Изобретение относится к антагонистам фактора роста клеток сосудистого эндотелия (ФРЭККС), к терапевтическому составу этих антагонистов, а также к методам использования антагонистов в диагностических и терапевтических целях.
Предпосылки изобретения
Двумя основными клеточными компонентами кровеносных сосудов являются эндотелиальные клетки и гладкие мышечные клетки. Эндотелиальные клетки образуют выстилку внутренней поверхности всех кровеносных сосудов и представляют собой барьер между кровью и тканями, препятствующий образованию тромбов. Помимо того, эндотелиальные клетки являются важным компонентом в формировании новых капилляров и кровеносных сосудов. Таким образом, эндотелиальные клетки пролиферируют во время ангиогенеза, или неоваскуляризации, связанного с ростом и метастазированием опухолей и с другими заболеваниями или нарушениями, иными чем неопластические.
По имеющимся данным пролиферация эндотелиальных клеток может вызываться различными полипептидами, которые естественным образом встречаются в природе. К числу таких полипептидов относятся основные и кислотные факторы роста фибробластов (ФРФ) (Берджесс и Масиаг, "Ежег. биохим.обозрение" (Animal Rev. Biochem. ) 58, 575 (1989)), фактор роста эндотелиальных клеток, образующийся из тромбоцитов (ФРЭКТ) (Исикава и др., "Природа" (Nature), 338, 557 (1989)), и фактор роста эндотелиальных клеток кровеносных сосудов (ФРЭККС) (Леунг и др. "Наука" (Science) 246, 1306 (1989), Феррара и Ген-цель, "Научн.сообщ.по биохим. и биофиз." (Biochem. Biophys. Res. Commun), 161, 851 (1989), Тишер и др. , "Научн. сообщ. по биохим. и биофиз." (Biochem. Biophys. Res. Commun), 165, 1198 (1989), Феррара и др. PCT Pat. Publ. N WO 90/13649 (опубликовано 15 ноября 1990 г.); Феррара и др. Заявка на патент США (U. S. Pat. App.) N 07/360229).
ФРЭККС был впервые определен в среде, кондиционированной фолликулярными или фолликулостеллярными клетками бычьего гипофиза. Биохимический анализ показывает, что бычий ФРЭККС представляет собой димерный белок с примерным молекулярным весом около 45000 дальтон и с вероятной митогенной специфичностью для эндотелиальных клеток кровеносных сосудов. ДНК, кодирующая бычий ФРЭККС, была выделена посредством скрининга библиотеки кДНК, приготовленной из таких клеток с использованием в качестве пробы гибридизации олегонуклеотидов, основанных на последовательности аминокислот аминного конца молекулы белка.
ФРЭККС человека был приготовлен сначала посредством скрининга библиотеки кДНК, приготовленной из таких клеток человека, с использованием кДНК бычьего ФРЭККС в качестве пробы для гибридизации. Одна из кДНК, выделенных таким способом, кодирует белок, состоящий из 165 аминокислот и имеющий более чем 95% гомологию с бычьим ФРЭККС. Данный белок получил название человеческого ФРЭККС (ФРЭККС). Митогенная активность человеческого ФРЭККС была подтверждена путем экспрессии кДНК человеческого ФРЭККС в клетках млекопитающих. Среда, кондиционированная клетками, которые были трансфецированы кДНК человеческого ФРЭККС, поддерживала пролиферацию капиллярных эндотелиальных клеток, в то время как контрольные клетки не проявляли такой активности [Леунг и др., "Наука" (Science), 246: 1306 (1989)].
Помимо ФРЭККС в библиотеках кДНК человека были определены еще несколько дополнительных кДНК, которые кодируют изоформы ФРЭККС, состоящие из 121, 189, и 206 аминокислот (совместно называемые белками, близкими к ФРЭККС). Белок, состоящий из 121 аминокислоты, отличается от ФРЭККС наличием делеции 44 аминокислот между позициями аминокислотных остатков 116 и 159 в последовательности ФРЭККС. Белок, состоящий из 189 аминокислот, отличается от ФРЭККС наличием вставки добавочных 24 аминокислот после позиции аминокислотного остатка 116, и, по всей видимости, этот белок идентичен человеческому фактору проницаемости кровеносных сосудов (чФПКС). Белок, состоящий из 206 аминокислот, отличается от ФРЭККС наличием вставки добавочной 41 аминокислоты после позиции аминокислотного остатка 116 [Хоук и др. "Молекулярная эндокринология" (Mol. Endocrinol.), 5: 1806 (1991); Феррара и др. "Журнал клеточной биохимии" (J. Cell Biochem.), 47: 211 (1991); Феррара и др. "Эндокринол. обозрение" (Endrocrine Reviews), 13: 18 (1992); Кек и др. "Наука" (Science), 246: 1309 (1989); Коннолли и др. "Журнал биологической химии" (J. Biol. Chem. ), 264: 20017 (1989); Кек и др. ЕРО. Pat. Publ. N 0370989 (опубликовано 30 мая 1990 г.).
ФРЭККС не только стимулирует пролиферацию эндотелиальных клеток кровеносных сосудов, но также индуцирует проницаемость кровеносных сосудов и ангиогенез. Ангиогенез, который включает образование новых кровеносных сосудов из уже существующего эндотелия, является важным компонентом многих заболеваний и нарушений, в том числе таких, как рост и метастазирование опухолей, ревматический артрит, псориаз, атеросклероз, диабетическая ретинопатия, ретролентальная фиброплазия, неоваскулярная глаукома, гемангиомы, иммунное отторжение трансплантированной роговицы и других тканей, а также хронические воспаления.
В случае роста опухолей ангиогенез имеет особенно важное значение при переходе от гиперплазии к неоплазии, а также для обеспечения питания растущей твердой опухоли [Фолкман и др. "Природа" (Nature), 339: 58 (1989)]. Ангиогенез также позволяет опухолям находиться в контакте с кровеносной системой хозяина, в результате чего могут быть определены направления метастазирования для клеток опухоли. Данные, подтверждающие роль ангиогенеза в метастазировании клеток опухоли, были получены, в частности, в результате исследований, показавших корреляцию между количеством и плотностью микрососудов на гистологических срезах инвазивной карциномы груди человека и фактическим наличием метастазов, распространившихся на значительное расстояние [Вейднер и др."Медицинск, журнал Новой Англии" (New Eng. J. Med.), 324: 1 (1991)].
В свете значения роста эндотелиальных клеток кровеносных сосудов и ангиогенеза, а также роли этих процессов в различных заболеваниях и нарушениях желательно было бы иметь средства для уменьшения или остановки одного или нескольких из биологических эффектов, оказываемых ФРЭККС.
Желательно также иметь средства для обнаружения присутствия ФРЭККС в нормальных и патологических условиях, и особенно в случае наличия рака.
Краткое описание изобретения
В настоящем изобретении представлены вещества-антагонисты ФРЭКСС, включая (а) антитела и их варианты, способные к специфическому связыванию ФРЭККС, рецептора ФРЭККС или же комплекса ФРЭККС и рецептора ФРЭККС; (б) рецептор ФРЭККС и его варианты и (в) варианты ФРЭККС. Подобные антагонисты ингибируют митогенную, ангиогенную и прочую биологическую активность ФРЭККС и, следовательно, могут быть применены для лечения заболеваний или нарушений, характеризующихся избыточной неоваскуляризацией, в том числе, например, опухолей (в особенности твердых злокачественных опухолей), ревматоидного артрита, псориаза, атеросклероза, диабетической и другой ретинопатии, ретролентальной фиброплазии, неоваскулярной глаукомы, гемангиомы, гиперплазии щитовидной железы (включая болезнь Грейва), иммунного отторжения трансплантировенной роговицы и других тканей, а также хронических воспалений. Антагонисты могут быть применены также для лечения заболеваний или нарушений, характеризующихся нежелательной избыточной проницаемостью кровеносных сосудов, таких как отеки, связанные с опухолью мозга, асциты, связанные со злокачественными опухолями, синдром Мейгса, воспаление легких, нефротический синдром, перикардиальный выпот (например, связанный с перикардитом) и плевральный выпот.
Помимо того, антагонисты ФРЭККС являются полиспецифическими моноклональными антителами, которые способны связываться: (а) с иными эпитопами, чем эпитоп ФРЭККС, например эпитопами белка, участвующего в процессах тромбогенеза или тромболиза, или антигеном поверхности опухолевой клетки и (б) с ФРЭККС, рецептором ФРЭККС или же с комплексом ФРЭККС и рецептора ФРЭККС.
Помимо того, антагонисты ФРЭККС могут быть присоединены к цитотоксическому остатку.
Кроме того, данное изобретение относится к выделению нуклеиновых кислот, кодирующих моноклональные антитела согласно вышеописанному, а также к линиям клеток гибридомы, продуцирующим подобные моноклональные антитела.
Помимо того, данное изобретение относится к фармацевтическим составам, включающим антагонист ФРЭККС в количестве, достаточном для снижения или остановки митогенной или ангиогенной активности ФРЭККС млекопитающих.
В другом плане данное изобретение относится к методам лечения, состоящим в применении к млекопитающему, желательно к пациенту-человеку, нуждающемуся в подобном лечении, физиологически эффективного количества антагониста ФРЭККС. При желании, совместно с антагонистом ФРЭККС могут быть применены (одновременно или последовательно) другие антагонисты ФРЭККС или антиопухолевые или антиангиогенные вещества (одно или более).
Кроме того, данное изобретение относится к методу обнаружения ФРЭККС в пробах посредством приведения пробы в контакт с антителом, способным специфически связываться с ФРЭККС, и определения степени такого связывания.
Краткое описание иллюстраций
Фиг. 1 показывает эффект моноклональных антител к ФРЭККС (A4.6.1 или В2.6.2), а также постороннего антитела к фактору роста гепатоцитов (анти-ФРГ) на связывание моноклональных антител к ФРЭККС с ФРЭККС.
Фиг. 2 показывает эффект моноклональных антител к ФРЭККС (A4.6.1 или B2.6.2), а также постороннего анти-ФРГ на биологическую активность ФРЭККС в культурах бычьих клеток капиллярного эндотелия коры надпочечников (КЭКН).
Фиг. 3 показывает эффект моноклональных антител к ФРЭККС (A4.6.1, B2.6.2 или A2.6.1) на связывание ФРЭККС с бычьими клетками КЭКН.
Фиг. 4 показывает эффект применения моноклонального антитела к ФРЭККС (A4.6.1) на скорость роста мышиных опухолей NEG55.
Фиг. 5 показывает эффект применения моноклонального антитела к ФРЭККС (A4.6.1) на величину мышиных опухолей NEG55 после применения в течение пяти недель.
Фиг. 6 показывает эффект применения моноклонального антитела к ФРЭККС A4.6.1 (Ат ФРЭККС) на рост мышиных опухолей SK-LMS-1.
Фиг. 7 показывает эффект применения различных по величине доз моноклонального антитела к ФРЭККС A4.6.1 (Ат ФРЭККС) на рост мышиных опухолей A673.
Фиг. 8 показывает эффект моноклонального антитела к ФРЭККС A4.6.1 на рост и выживаемость культуры клеток глиобластомы NEG55 (G55).
Фиг. 9 показывает эффект моноклонального антитела к ФРЭККС A4.6.1 на рост и выживаемость культуры клеток рабдомиосаркомы A673.
Фиг. 10 показывает эффект моноклонального антитела к ФРЭККС A4.6.1 на хемотаксис человеческих эндотелиальных клеток, вызванный человеческой синовиальной жидкостью.
Подробное описание изобретения
Термин "ФРЭККС" используется здесь и далее по отношению к человеческому фактору роста эндотелиальных клеток кровеносных сосудов, состоящему из 165 аминокислот, а также по отношению к близким человеческим факторам роста эндотелиальных клеток кровеносных сосудов, состоящим из 121, 189 и 206 аминокислот, согласно описанию Леунг и др. "Наука" (Science), 246: 1306 (1989) и Хоук и др., "Молекулярная эндокринология" (Mol. Endocrin.), 5: 1806 (1991), а также по отношению к встречающимся в природе аллельным или модифицированным формам таких факторов роста.
Настоящее изобретение включает вещества-антагонисты ФРЭККС, которые способны ингибировать одну или более из биологических активностей ФРЭККС, например его митогенную или ангиогенную активность. Действие антагонистов ФРЭККС заключается в препятствовании связыванию ФРЭККС с клеточным рецептором, в остановке жизнедеятельности или уничтожении клеток, которые были активированы ФРЭККС, или в препятствовании активации эндотелиальных клеток кровеносных сосудов после того, как ФРЭККС связывается с клеточным рецептором. Для целей данного изобретения все подобные моменты воздействия антагонистов ФРЭККС будут считаться эквивалентными. Таким образом, в состав данного изобретения включаются антитела, предпочтительно моноклональные, или же фрагменты таковых, которые связываются с ФРЭККС, рецептором ФРЭККС или же с комплексом ФРЭККС и рецептора ФРЭККС. Помимо того, в состав данного изобретения включаются фрагменты ФРЭККС и варианты аминокислотной последовательности ФРЭККС, которые связываются с рецептором ФРЭККС, но не проявляют биологической активности нативного ФРЭККС. Помимо того, в состав данного изобретения включаются рецептор ФРЭККС, а также его фрагменты и варианты его аминокислотной последовательности, которые способны связывать ФРЭККС.
Термин "рецептор ФРЭККС" (рФРЭККС) используется здесь и далее по отношению к клеточному, как правило, поверхностному рецептору, находящемуся на эндотелиальных клетках кровеносных сосудов человека, а также к его вариантам, которые сохраняют способность связывать ФРЭККС. Как правило, те рецепторы ФРЭККС и их варианты, которые являются антагонистами ФРЭККС, будут фигурировать в изолированной форме, в отличие от ситуации в естественных условиях, где они могут быть интегрированы в состав клеточной мембраны или прикреплены к поверхности клетки. Одним из примеров рецептора ФРЭККС является fms-подобная тирозинкиназа (flt), трансмембранный рецептор из группы тирозинкиназ [ДеФриз и др. "Наука" (Science), 255: 989 (1992); Шибуя и др. "Онкоген" (Oncogene), 5: 519 (1990)]. Рецептор flt состоит из внеклеточного домена, трансмембранного домена и внутриклеточного домена, имеющего тирозинкиназную активность. Внеклеточный домен участвует в связывании ФРЭККС, в то время как внутриклеточный домен участвует в передаче сигнала.
Другим примером рецептора ФРЭККС является рецептор flk-1 (называемый также KDR) [Мэтьюс и др. "Известия национальн. Акад. Наук" (Proc. Nat. Acad. Sci. ), 88: 9026 (1991); Терман и др. "Онкоген" (Oncogene), 6: 1677 (1991); Терман и др. "Научн. сообщ. по биохим. и биофиз". (Biochem. Biophys. Res. Commun.), 187: 1579 (1992)].
Связывание ФРЭККС с рецептором flt приводит к образованию по меньшей мере двух комплексов с высоким молекулярным весом от 205000 до 300000 дальтон. Комплекс с весом 300000 дальтон, по-видимому, является димером, включающим две молекулы рецептора, связанные с одной молекулой ФРЭККС.
В состав данного изобретения включены также варианты рФРЭККС. Примеры таких вариантов включают усеченные формы рецепторов, где отсутствуют трансмембранный и цитоплазматический домены, а также слитые белки, в которых полимеры или полипептиды, отличные от рФРЭККС, присоединены к рФРЭККС или же, предпочтительно, к его усеченным формам. Примером такого полипептида, отличного от рФРЭККС, является иммуноглобулин. В этом случае, например, внеклеточный домен рФРЭККС замещен Fv-доменом легкой или (предпочтительно) тяжелой цепи иммуноглобулина, причем C-конец внеклеточного домена рецептора ковалентно соединен с аминоконцом фрагмента тяжелой цепи (CH-1, сочленения, CH-2, или другого фрагмента). Подобные варианты построены таким же образом, как известные иммуноадгезоны, см., например: Гаскойн и др. "Известия национальн. Акад. Наук" (Proc. Nat. Acad. Sci.), 84: 2936 (1987); Капон и др. "Природа" (Nature), 337: 525 (1989); Аруффо и др. "Клетка" (Cell), 61: 1303 (1990); Ашкенази и др. "Известия национальн. Акад. Наук" (Proc. Nat. Acad. Sci.), 88: 10535 (1991); Беннетт и др. "Журн. биол. химии" (J. Biol. Chem.), 266: 23060 (1991). В других случаях рФРЭККС может быть присоединен к небелковому полимеру, такому как полиэтиленгликоль (ПЭГ) [см. Дэвис и др. Патент США (U. S. Patent) N 4179337; Гудсон и др. "Биотехнология" (Biotechnology), 8: 343-346 (1990); Абуховски и др. "Журн. биол. химии" (J. Biol. Chem.), 252: 3578 (1977); Абуховски и др. "Журн. биол. химии" (J. Biol. Chem.), 252: 3582 (1977)] или углеводы [см., например: Маршалл и др. "Архив биохимии и биофизики" (Arch. Biochem. Biophys. ), 167: 77 (1975)]. Посредством этого достигается увеличение срока биологического полураспада рФРЭККС и уменьшается вероятность того, что рецептор окажет иммуногенное влияние на млекопитающее, к которому этот рецептор будет применен. Использование рФРЭККС аналогично использованию антител к ФРЭККС, если принять во внимание сродство антагониста и число мест его связывания с ФРЭККС.
Внеклеточный домен рецептора ФРЭККС сам по себе или же слитый с иммуноглобулинным полипептидом или другим полипептидом-носителем в особенности полезен для использования в качестве антагониста ФРЭККС ввиду того, что внеклеточный домен может связывать ФРЭККС клетки-хозяина, не связанный с рФРЭККС на поверхности клетки.
Кроме того, рФРЭККС и его варианты могут использоваться в скрининговых анализах для выявления агонистов и антагонистов ФРЭККС. Например, клетки хозяина, трансфецированные ДНК, кодирующей рФРЭККС (например, ДНК flt или flk-1), проявляют избыточную экспрессию рецепторного полипептида на поверхности клетки, в результате чего такие рекомбинантные клетки хозяина могут идеально служить для анализа того, как тестируемое вещество (например, небольшая молекула, линейный или циклический пептид или полипептид) связывается с рФРЭККС. Подобным же образом могут быть использованы белки, в которых рФРЭККС слит с другим белком, например слитый белок рФРЭККС-ИгГ. К примеру, слитый белок может быть присоединен к иммобилизованному основанию, после чего определяется способность тестируемого вещества удалять ФРЭККС, помеченный радиоактивным изотопом, из домена слитого белка, включающего рФРЭККС.
Термин "рекомбинантный", применяемый в отношении к ФРЭККС, рецептору ФРЭККС, моноклональным антителам или прочим белкам, обозначает белки, которые производятся путем экспрессии рекомбинантной ДНК в клетках хозяина. Клетка-хозяин может быть прокариотической (например, бактериальная клетка, такая как Е. coli) или эукариотической (например, клетка дрожжей или млекопитающих).
Моноклональные антитела-антагонисты
Термин "моноклональное антитело" используется здесь и далее для обозначения антитела, полученного из популяции достаточно однородных антител, т. е. индивидуальные антитела, составляющие популяцию, идентичны в своей специфичности и сродству, за исключением возможных, естественно встречающихся мутаций, которые могут присутствовать в незначительных количествах. Необходимо обратить внимание на то, что в результате подобных, естественно встречающихся, мутаций состав моноклональных антител в данном изобретении, которое в большинстве своем содержит антитела, способные специфически связывать ФРЭККС, рецептор ФРЭККС или же комплекс ФРЭККС и рецептора ФРЭККС, может также включать незначительные количества прочих антител.
Таким образом, термин "моноклональное" указывает на характер антитела, происходящего из достаточно однородной популяции антител, но здесь не имеется в виду, что антитела должны производиться каким-либо определенным путем. Например, моноклональные антитела, описанные в данном изобретении, могут быть получены гибридомным методом [Колер и Милстейн."Природа" (Nature), 256: 495 (1975)] или с применением методов, использующих рекомбинантную ДНК [Кабилли и др. Патент США (U. S. Patent) N 4816567].
При получении моноклональных антител гибридомным методом мышь или другое подходящее животное-хозяин иммунизируется антигеном посредством подкожной, внутриперитонеальной или внутримышечной инъекции с целью выявить лимфоциты, которые производят или же способны производить антитела, специфически связывающиеся с белком(ами), использованным(и) для иммунизации. В качестве альтернативы, лимфоциты могут быть иммунизированы in vitro. Затем лимфоциты сливают с клетками миеломы с использованием соответствующего агента, например такого, как полиэтиленгликоль, чтобы создать гибридомную клетку [Годинг, "Моноклональные антитела: Принципы и практика" (Monoclonal Antibodies: Principles and Practice, с. 59-103 (Academic Press, 1986)].
Таким антигеном могут являться ФРЭККС, рФРЭККС или же комплекс ФР ЭККС и рФРЭККС. Антиген может представлять собой фрагмент или часть ФРЭККС или рФРЭККС, обладающие одним или несколькими аминокислотными остатками, которые участвуют в связывании ФРЭККС с одним из его рецепторов. Например, иммунизация внеклеточным доменом рФРЭККС (т.е. усеченным полипептидом рФРЭККС, в котором отсутствуют трансмембранный и внутриклеточный домены) будет особенно пригодна для получения антител, являющихся антагонистами ФРЭККС, поскольку именно внеклеточный домен участвует в связывании ФРЭККС.
Также могут быть использованы моноклональные антитела, способные к связыванию комплекса ФРЭККС и рФРЭККС, в особенности если они при этом не связываются с неассоциированными (не находящимся в составе комплекса) ФРЭККС и рФРЭККС. Подобные антитела, таким образом, связываются только с клетками, которые находятся в данный момент в состоянии активации, вызванном ФРЭККС, и, соответственно, не изолированы свободными ФРЭККС и рФРЭККС, что является обычной ситуацией в клетках млекопитающих. Такие антитела обычно связывают эпитоп, который занимает одно или несколько мест контакта между рецептором и ФРЭККС. Такие антитела были получены и для других лигандных рецепторных комплексов и могут быть аналогичным образом получены и в данном случае. Подобные антитела, независимо от наличия или отсутствия способности связываться с неассоциированными ФРЭККС и рФРЭККС, не обязательно должны нейтрализовать или ингибировать биологическую активность неассоциированных ФРЭККС и рФРЭККС.
Приготовленные таким образом гибридомные клетки высеваются и выращиваются в подходящей культуральной среде, которая предпочтительно должна содержать одно или несколько веществ, которые ингибируют рост или выживание неслитых, родительских клеток миеломы. Например, в случае, если в родительских клетках миеломы отсутствует фермент гипоксантингуанинфосфорибозилтрансфераза (ГГФРТ или ГФРТ), культуральная среда для гибридом обычно будет содержать гипоксантин, аминоптерин и тимидин (среда HAT), каковые вещества препятствуют росту клеток, не обладающих ГГФРТ.
Предпочтительно выбирать такие клетки миеломы, которые эффективно сливаются, поддерживают стабильный высокий уровень экспрессии антител в отобранных клетках, производящих антитела, и являются чувствительными к средам, таким, например, как среда HAT. Среди таких клеток предпочтительными клеточными линиями являются: мышиные линии миеломы, такие как линии, происходящие от мышиных опухолей МОРС-21 и МРС-11, которые можно получить из Центра распределения клеток Института им. Солка в Сан-Диего (Калифорния, США); клетки SP-2, которые можно получить из Американской коллекции типовых культур в Роквилле (Мэриленд, США); и клетки P3X63Ag8U.1, описанные Йелтоном и др. ("Текущие вопросы в микробиологии и иммунологии" (Curr. Top. Microbiol. Immunol.), 81: 1 (1978). Кроме того, были описаны клеточные линии человеческой миеломы и человеческо-мышиной гетеромиеломы, способные производить человеческие моноклональные антитела (Козбор, "Журн. иммунологии" (J.Immunol.), 133: 3001 (1984); Бродер и пр. "Получение моноклональных антител: методы и их применение" (Monoclonal Antibody Production Techniques and Applications), c. 51-63 (Marcel Dekker Inc., New York, 1987).
Культурная среда, в которой выращиваются клетки гибридомы, подвергается анализу для производства моноклональных антител, направленных против соответствующего антигена. Предпочтительно, чтобы специфичность связывания моноклональных антител, производимых клетками гибридомы, определялась путем иммунопреципитации или анализа связывания in vitro, например радиоиммуноанализом (RIA) или твердофазным иммуноферментным анализом (ELISA). Данное изобретение относится к тем моноклональным антителам, которые предпочтительно образуют иммунопреципитат с ФРЭККС, рФРЭККС или комплекс ФРЭККС и рФРЭККС, или же предпочтительно связывают по меньшей мере один из этих антигенов при анализе связывания и способны ингибировать биологическую активность ФРЭККС.
После определения клеток гибридомы, которые производят антагонистические антитела желаемой специфичности, сродства и активности, клоны могут быть субклонированы методом ограниченных разбавлений и выращены стандартными методами [Годинг, "Моноклональные антитела: Принципы и практика" (Monoclonal Antibodies: Principles and Practice, с. 59-104 (Academic Press, 1986)]. К подходящим культуральным средам относятся, например, среда Игла, модифицированная Дулбекко (СИМД), или среда RPMI-1640. Кроме того, клетки гибридомы могут выращиваться in vivo в животных как асцитные опухоли.
Моноклональные антитела, продуцированные субклонами, отделяются от культуральной среды, асцитной жидкости или плазмы путем использования обычных методов иммуноглобулинной очистки, таких как, например, белок A-Сефароза, гидроксилапатитная хроматография, электрофорез в гелях, диализ или аффинная хроматография.
ДНК, кодирующая моноклональные антитела, описанные в данном изобретении, может быть с легкостью выделена и секвенирована обычными методами (например, с использованием олигонуклеотидных проб, способных связываться специфически с генами, кодирующими тяжелую и легкую цепу мышиных антител). В качестве предпочтительного источника ДНК служат клетки гибридомы, описанные в данном изобретении. После выделения ДНК может быть помещена в векторы экспрессии, которые затем трансфецируются в клетки хозяина, такие как обезьяньи клетки линии COS, клетки яичника китайского хомячка (СНО) или клетки миеломы, которые в иной ситуации не продуцируют иммуноглобулинный белок, для того чтобы достигнуть синтеза моноклональных антител в рекомбинантных клетках хозяина.
ДНК может быть модифицирована по выбору для того, чтобы изменить характер иммуноглобулина, продуцируемого экспрессией этой ДНК. Так, например, гуманизированные формы мышиных антител могут быть получены путем замещения комплементарной определяющей области (CDR) вариабельного домена мышиного антитела на соответствующую область человеческого антитела. В некоторых вариантах отдельные аминокислоты из базовой области (FR) мышиного антитела также замещаются на соответствующие аминокислотные остатки человеческого антитела [Картер и др. "Известия национальн. Акад. Наук" (Proc. Nat. Acad. Sci.), 89: 4285 (1992); Картер и др. "Биотехнология" (Biotechnology), 10: 163 (1992)]. Химерные формы мышиных антител могут быть получены также путем замещения гомологичных мышиных последовательностей ДНК на последовательность, кодирующую отдельные области человеческих постоянных цепей иммуноглобулина (тяжелой и легкой) [Кабилли и др., Патент США (U. S. Patent) N 4816567; Моррисон и пр. "Известия национальн. Акад. Наук" (Proc. Nat. Acad. Sci.), 81: 6851 (1984)].
Антитела, описанные в данном изобретении, включают варианты антител, такие как химерные (в том числе "гуманизированные") антитела и гибридные антитела, представляющие собой иммуноглобулинные цепи, способные связывать ФРЭККС, рецептор ФРЭККС, комплекс ФРЭККС и рецептора ФРЭККС, или же эпитоп, не принадлежащий к ФРЭККС.
В состав таких антител включаются все виды, классы (например, IgA, IgD, lgE, IgG и IgM) и подклассы иммуноглобулинов, а также и фрагменты антител (например, Fab, F(ab')2 и Fv), пока они обладают способностью связывать ФРЭККС, рецептор ФРЭККС или комплекс ФРЭККС и рецептора ФРЭККС и пока они проявляют антагонизм по отношению к биологической активности ФРЭККС.
В случае предпочитаемого варианта данного изобретения моноклональные антитела будут проявлять сродство к иммунизирующему антигену в размере по меньшей мере 109 литров/моль, как определяется, например, анализом Скатчарда [Мансон и Поллард, "Аналит. биохимия" (Anal. Biochem), 107: 220 (1980)]. Кроме того, моноклональные антитела обычно будут ингибировать митогенную или ангиогенную активность ФРЭККС по крайней мере на 50%; предпочтительно на 80% и наиболее предпочтительно на 90%, как определяется, например, анализом выживания или пролиферации клеток in vitro, подобно описанному в Примере 2.
Для некоторых терапевтических и диагностических применений желательно, чтобы моноклональные антитела реагировали не со всеми различными молекулярными формами ФРЭККС. Например, может быть желательно получить моноклональное антитело, которое способно специфически связываться только с полипептидом ФРЭККС, имеющим последовательность из 165 аминокислот, но не с полипептидами ФРЭККС, имеющими последовательности из 121 или 189 аминокислот. Такие антитела легко определяются путем сравнения анализов ELISA или путем сравнения иммуноприпитации различных полипептидов ФРЭККС.
Конъюгаты с цитотоксическими остатками
В некоторых вариантах желательно иметь цитотоксический остаток в виде конъюгата с моноклональным антителом, специфическим для ФРЭККС, или же в виде конъюгата с рФРЭККС. В подобных вариантах цитотоксин служит для приостановки жизнедеятельности или для уничтожения клеток, которые проявляют экспрессию ФРЭККС или рецептора ФРЭККС или же связывают эти белки. Конъюгат нацеливается на клетку-мишень через посредство домена, который способен связываться с ФРЭККС, рФРЭККС или комплексом ФРЭККС и рФРЭККС.
Таким образом, моноклональные антитела, способные связываться с ФРЭККС, рФРЭККС или комплексом ФРЭККС и рФРЭККС, конъюгируются с цитотоксинами. Подобным же образом конъюгируется с цитотоксином ФРЭККС. В то время, как в оптимальном случае моноклональные антитела способны нейтрализовать активность ФРЭККС и без участия цитотоксина, в случае данного варианта не является необходимым, чтобы моноклональное антитело или рецептор были способны к какой-либо другой активности, кроме связывания с ФРЭККС, рФРЭККС или комплексом ФРЭККС и рФРЭККС.
В обычном случае в качестве цитотоксина выступает белковый цитотоксин, например токсин возбудителя дифтерии, рицин или токсин Pseudomonas, хотя в ряде случаев с некоторыми классами иммуноглобулинов домен Fc самого моноклонального антитела может служить в качестве цитотоксина [например, в случае антител IgG2, которые способны фиксировать комплемент и участвовать в антителозависимой клеточной цитотоксичности (АТКЦ)]. Однако цитотоксин не обязательно должен иметь белковую природу и может включать химиотерапевтические вещества, до настоящего времени употреблявшиеся, например, для лечения опухолей.
Обычно цитотоксин присоединяется к моноклональному антителу или же фрагменту такого антитела через амидную связь в составе основной цепи молекулы антитела в ее домене Fc (или же замещая этот домен или его часть). В случае, когда рФРЭККС обеспечивает функцию нацеливания на клетку, цитотоксический остаток присоединяется к любому домену рецептора, который не участвует в связывании ФРЭККС. Предпочтительно, чтобы остаток замещал трансмембранный или цитоплазматический домены рецептора или присоединялся к ним. Оптимальный участок замещения должен быть определен путем стандартных экспериментов, не требующих дополнительных навыков.
Конъюгаты, являющиеся слитыми белками, легко производятся в рекомбинантной культуре клеток путем экспрессии гена, который кодирует данный конъюгат. В альтернативном случае конъюгаты производятся путем ковалентного перекрестного сшивания цитотоксического остатка с боковой цепью или с C-терминальным карбоксилом антитела рецептора с использованием методик, которые известны сами по себе как дисульфидный обмен или сшивание посредством тиоэфирной связи, например, с использованием иминотиолата и метил-4-меркаптобутиримадата.
Конъюгаты с другими остатками
Моноклональные антитела и рФРЭККС, являющиеся антагонистами ФРЭККС, также могут конъюгироваться с веществами, которые сами по себе не могут быть непосредственно отнесены к цитотоксинам, но которые повышают активность описанных составов. Например, моноклональные антитела, способные связываться с ФРЭККС, рФРЭККС или комплексом ФРЭККС и рФРЭККС, сливают с гетерологичными полипептидами, такими как последовательности вирусов, клеточные рецепторы, цитокины (такие как ТНФ), интерфероны или интерлейкины, с полипептидами, имеющими прокоагулянтную активность, и с прочими биологически или иммунологически активными полипептидами. Подобные слитые конструкции легко изготовляются с помощью рекомбинантных методов. Обычно такие неиммуноглобулинные полипептиды замещаются на постоянный домен (или домены) антитела к ФРЭККС или к комплексу ФРЭККС и рФРЭККС или на трансмембранный и/или внутриклеточный домены рФРЭККС. В альтернативном случае они замещаются на вариабельный домен одного антигенсвязывающего участка описанного антитела к ФРЭККС.
Предпочтительными являются те варианты, в которых такие неиммуноглобулинные полипептиды соединены с или заменяют постоянные домены описанного антитела [Беннетт и др. "Журнал биологической химии" (J. Biol. Chem.), 266: 23060 (1991)]. В альтернативном случае они замещаются на Fv описанного антитела для создания химерного поливалентного антитела, включающего по крайней мере один остающийся антигенсвязывающий участок, специфичный для ФРЭККС, рФРЭККС или комплекса ФРЭККС и рФРЭККС, и суррогатный антигенсвязывающий участок, имеющий иную специфичность, чем исходное антитело.
Гетероспецифические антитела
Моноклональные антитела, способные связываться с ФРЭККС, рФРЭККС или комплексом ФРЭККС и рФРЭККС, должны содержать только один связывающий участок для данных эпитопов, обычно одиночный комплекс тяжелой и легкой цепей или фрагмент такого комплекса. Однако подобные антитела могут также иметь антигенсвязывающие домены, способные связывать иные эпитопы, нежели эпитопы ФРЭККС, рФРЭККС или комплекса ФРЭККС и рФРЭККС. Например, в результате замещения соответствующей аминокислотной последовательности или аминокислотных остатков нативных антител к ФРЭККС, рФРЭККС или к комплексу ФРЭККС и рФРЭККС на остатки, определяющие комплементарность, и, в случае необходимости, остовные остатки антитела, специфического по отношению к антигену, иному чем ФРЭККС, рФРЭККС или комплекс ФРЭККС и рФРЭККС, может быть создано полиспецифическое антитело, включающее один антигенсвязывающий участок, специфический для ФРЭККС, рФРЭККС или комплекса ФРЭККС и рФРЭККС, и другой антигенсвязывающий участок, специфический для антигена, иного чем ФРЭККС, рФРЭККС или комплекс ФРЭККС и рФРЭККС. Такие антитела являются по меньшей мере бивалентными, но могут быть и поливалентными, в зависимости от числа антигенсвязывающих участков, которым обладают антитела выбранного класса. Например, антитела класса IgM будут поливалентными.
Предпочтительными являются те варианты, в которых такие антитела способны связывать эпитопы ФРЭККС или рФРЭККС, а также (а) полипептид, активный при коагиляции крови, такой как белок C или тканевый фактор, или (б) цитотоксический белок, такой как фактор некроза опухолей (ФНО), или (в) рецептор клеточной поверхности, отличный от рФРЭККС, такой как рецепторы CD4 или HER-2 [Мэддон и др., "Клетка" (Cell), 42: 93 (1985); Куссенс и др., "Наука" (Science), 230: 1137 (1985)]. Гетероспецифические, мультивалентные антитела могут быть получены путем котрансформации клетки-хозяина ДНК, кодирующей тяжелую и легкую цепи обоих антител, с последующим сохранением, путем имунноаффинной хроматографии или подобным методом, той части экспрессированных антител, которые обладают желаемыми антигенсвязывающими свойствами. В альтернативном случае подобные антитела получаются путем рекомбинации моноспецифических антител in vitro.
Моновалентные антитела
Моновалентные антитела, способные связываться с ФРЭККС или комплексом ФРЭККС и рФРЭККС, особенно полезны в качестве антагонистов ФРЭККС. Данное изобретение не ограничено описанием какого-либо отдельного механизма биологической активности; предполагается, что активация клеточных рецепторов ФРЭККС следует механизму, в котором связывание ФРЭККС и клеточных рецепторов ФРЭККС индуцирует агрегацию рецепторов, что в свою очередь активирует киназную активность внутриклеточного рецептора. Поскольку моновалентные антитела к рецептору ФРЭККС не способны индуцировать подобную агрегацию и вследствие этого активировать ФРЭККС описанным способом, они являются идеальными антагонистами ФРЭККС.
Необходимо, однако, заметить, что такие антитела должны быть направлены на ФРЭККС-связывающий участок рецептора или же должны быть иным образом способны интерферировать со связыванием ФРЭККС и рецептора ФРЭККС, скажем, пространственным образом преграждать доступ ФРЭККС к рецептору. Однако, как описано в данном изобретении, те антитела к ФРЭККС, которые не способны интерферировать со связыванием ФРЭККС, могут быть использованы в виде конъюгата с неиммуноглобулинными остатками, например с цитотоксинами.
Методы приготовления моновалентных антител относятся к числу широко известных в данной области. Например, один из методов состоит в рекомбинантной экспрессии легкой цепи и модифицированной тяжелой цепи иммуноглобулина. Тяжелая цепь усекается, в принципе, в любом месте участка Fc так, чтобы предотвратить сшивку тяжелой цепи. В альтернативном случае подходящие остатки цистеина замещаются другим аминокислотным остатком или вырезаются так, чтобы предотвратить сшивку. Для приготовления моновалентных антител используются также методы in vitro. Например, фрагменты Fab получаются путем ферменативного расщепления интактных антител.
Использование в диагностике
Для использования в диагностике антитела к ФРЭККС, описанные в данном изобретении, обычно должны быть помечены остатком, который легко обнаружить. Это может быть любой остаток, который, прямо или косвенно, может продуцировать обнаруживаемый сигнал. Например, это могут быть радиоизотопы, такие как 3H, 14C, 32P, 35S, 125I; флюоресцентное или хемилюминесцентное соединение, такое как изотиоцианат флюоресцеина, родамин или люциферин; метки, помеченные радиоизотопами, такими как, например, 125I, 32P, 14C или 3H, или ферменты, такие как щелочная фосфатаза, бетагалактозидаза или пероксидаза хрена.
Здесь может применяться любой известный в данной области метод для конъюгирования отдельных антител к обнаруживаемым остаткам, включая описанные методы [Хантер и др. "Природа" (Nature), 144: 945 (1962); Дэвид и др. "Биохимия" (Biochemistry), 13: 1014 (1974); Пейн и др. "Методы иммунол." (J. Immunol. Meth.), 40: 219 (1981), и Нигрен, "Журн. гистохим. и цитохим." (J. Histochem. and Cytochem.), 30: 407 (1982)].
Антитела и рецепторы данного изобретения могут быть использованы в любом известном методе анализа, например в анализе конкурентного связывания, анализе методом прямых и косвенных иммуносэндвичей и анализе методом иммунопреципитации (Зола, "Моноклональные антитела: практическое пособие" (Monoclonal Antibodies: A Manual of Techniques), с. 147-158 (CRC Press Inc. 1987).
Анализ конкурентного связывания основан на способности помеченного стандарта (это может быть ФРЭККС или же его иммунологически активная часть) конкурировать за ограниченное количество антитела с образцом ФРЭККС. Количество ФРЭККС в образце обратно пропорционально количеству стандарта, который связывается с антителами или рецепторами. Чтобы способствовать определению количества стандарта, которое переходит в связанное состояние, антитела или рецепты обычно переводятся в нерастворимую форму перед анализом или после него таким образом, что стандарт и образец, связанные с антителами или рецепторами, могут быть легко отделены от стандарта и образца, которые находятся в свободном состоянии.
Анализ методом иммуносэндвичей включает использование двух антител или рецепторов, каждый из которых способен связываться с различной иммуногенной областью или эпитопом белка, подлежащего обнаружению. При анализе методом иммуносэндвичей тестируемый образец связывается с первым антителом или рецептором, иммобилизованным на твердой основе, а затем второе антитело или рецептор связывается с образцом, образуя таким образом нерастворимый тройной комплекс [Дэвид и Грин, Патент США N 4376110]. Второе антитело или рецептор может быть помечено обнаруживаемым остатком (прямой метод иммуносэндвичей) или же может быть определено с использованием антитела к иммуноглобулину, помеченного обнаруживаемым остатком (косвенный метод иммуносэндвичей). Например, один из типов сэндвичей - это твердофазный иммуноферментный анализ (ELISA), и в этом случае обнаруживаемым остатком является фермент.
Антитела рецептора также используются для выявления in vivo. когда антитело или рФРЭККС, помеченные обнаруживаемым остатком, вводится пациенту, предпочтительно в кровеносную систему, и анализируется присутствие и местонахождение меченого антитела или рецептора в организме пациента. Такая визуализация может использоваться, например, при определении стадии заболевания и при лечении неоплазм. Антитело или рФРЭККС метятся любым остатком, обнаружимым в организме млекопитающих, известными в этой области методами, например ядерным магнитным резонансом, радиологическим методом и т.д.
Антагонисты-варианты ФРЭККС
В дополнение к описанным антителам могут быть использованы другие применяемые антагонисты ФРЭККС, а именно фрагменты и аминокислотные варианты нативного ФРЭККС, которые связываются с рецептором ФРЭККС, но не проявляют биологической активности, свойственной нативному ФРЭККС. Например, к таким антагонистам относятся фрагменты и аминокислотные варианты, которые составляют домен нативного ФРЭККС, связывающийся с рецептором, но не включают в себя домен, который проявляет биологическую активность, или имеют иные недостатки, так что они не могут активировать клеточный рецептор ФРЭККС, как, например, в случае с аминокислотным фрагментом, лишенным способности индуцировать агрегацию или активацию клеточных рецепторов ФРЭККС. Термин "домен, связывающийся с рецептором", относится к аминокислотным последовательностям ФРЭККС, которые участвуют в связывании с рецептором ФРЭККС. Термин "биологически активный домен" или "домен, обладающий биологической активностью", относится к аминокислотным последовательностям ФРЭККС, которые отвечают за определенную биологическую активность фактора, например его митогенную или ангиогенную активность.
ФРЭККС, по-видимому, способен образовывать комплекс с двумя или более молекулами рФРЭККС на поверхности клетки; это предполагает наличие у ФРЭККС по меньшей мере двух отдельных участков для связывания рФРЭККС; представляется также, что ФРЭККС связывается с такими клеточными рецепторами последовательным образом, сначала с одним участком и потом с другим, прежде чем происходит активация, как это делают гормоны роста - пролактин и подобные ему [см. Каннингем и др. "Наука" (Science), 254: 821 (1991); деВос и др. "Наука" (Science), 255: 306 (1992); Фу и др. "Наука" (Science), 256: 1677 (1992)] . Соответственно выбираются антагонисты-варианты ФРЭККС, в которых один участок ФРЭККС для связывания рецептора (обычно участок, который участвует в первоначальном связывании ФРЭККС и рФРЭККС) остается без изменения (или, в случае модификации, варьируется для того, чтобы усилить связывание), в то время как второй участок ФРЭККС, связывающий рецептор, обычно модифицируется путем субституции(й) или делеции(й) неконсервативных аминокислотных остатков с целью лишить этот участок его связывающих свойств.
Домены ФРЭККС, связывающие рецептор, и домены рФРЭККС, связывающие ФРЭККС, определяются любыми известными в этой области методами, в том числе рентгенографией, анализом мутаций или анализом связывания антител. Анализ мутаций включает в себя методы неспецифического насыщающего мутагенеза в совокупности с отбором утекающих мутантов и инсерционным мутагенезом. Другая стратегия, пригодная для определения доменов, которые связываются с рецептором, в составе лигандов известна как аланин (Ала)-сканирующий мутагенез [Каннингем и др. "Наука" (Science), 244: 1081-1985 (1989)]. Этот метод включает в себя определение областей, содержащих заряженные боковые цепи аминокислот. Заряженные остатки определяются на (например, Арг, Асп, Гис, Лиз и Глю) и замещаются на Ала (на одной области на каждую мутантную молекулу), и тестируется связывание полученных лигандов с рецептором, чтобы оценить значение отдельных областей в связывании с рецептором. Еще более мощным методом для локализации доменов, которые связываются с рецептором, является использование нейтрализующих антител к ФРЭККС [Ким и др. "Факторы роста" (Growth Factors), 7: 53 (1992)]. Обычно для локализации доменов, которые связываются с рецептором, используется комбинация таких методов и подобных им.
Термин "аминокислотный вариант", используемый по отношению к ФРЭККС, относится к полипептидам, аминокислотные последовательности которых до некоторой степени отличаются от аминокислотных последовательностей нативных форм ФРЭККС. Как правило, антагонистические варианты аминокислотных последовательностей будут обладать не менее чем 70% гомологией c по меньшей мере одним доменом нативного ФРЭККС, который связывается с рецептором; предпочтительно, чтобы гомология была около 80%; еще более предпочтительно, чтобы гомология с доменом нативного ФРЭККС, который связывается с рецептором, была около 90%. Варианты аминокислотных последовательностей обладают субституциями, делециями и/или инсерциями в определенных позициях последовательности нативного ФРЭККС таким образом, что варианты сохраняют способность связываться с рецептором ФРЭККС (и таким образом конкурировать с нативным ФРЭККС за связывание с рецептором ФРЭККС), но лишены способности индуцировать один или более биологических эффектов ФРЭККС, таких как пролиферация эндотелиальных клеток, ангиогенез или проницаемость сосудов.
"Гомология" определяется как процент остатков в аминокислотной последовательности, которые идентичны остаткам аминокислотной последовательности домена нативного ФРЭККС, который связывается с рецептором после того, как эти последовательности были сопоставлены и учтены пропуски в них, если это необходимо для достижения максимальной процентной гомологии. Для такого сопоставления в данной области исследований существуют соответствующие методы и компьютерные программы. Одна из таких компьютерных программ, "Align 2", была создана компанией "Генентек, Инк." (Genentech, Inc.), и документация к ней была занесена в архивы Управления по Авторским Правам США (United States Copyright Office, Washington, DC 20559) 10 декабря 1991 года. Субституционными вариантами являются варианты, в которых из нативной последовательности удален хотя бы один аминокислотный остаток и в той же позиции на его место вставлена другая аминокислота. Субституции могут быть одиночными или множественными, когда в одной и той же молекуле замещены две или более аминокислоты.
Инсерционными вариантами являются варианты, в которых одна или более аминокислот вставлены непосредственно по соседству с аминокислотой в определенной позиции нативной последовательности. "Непосредственно по соседству с аминокислотой" означает, что аминокислоты присоединяются либо к α-карбоксильной функциональной группе, либо к α-аминогруппе.
Делеционными вариантами являются варианты, в которых одна или более аминокислот удалены из нативной последовательности. Обычно делеционные варианты обладают делецией одной или более аминокислот в определенном участке молекулы. Фрагменты и аминокислотные варианты ФРЭККС легко могут быть получены известными в данной области методами, в том числе сайт-направленным мутагенезом ДНК, кодирующей нативный фактор. Мутировавшая ДНК вставляется в cоответствующий вектор экспрессии, и клетки-хозяева затем трансфецируются рекомбинантным вектором. Рекомбинантные клетки-хозяева выращиваются в соответствующей культуральной среде, и желаемый фрагмент или аминокислотный вариант ФРЭККС, выраженный в клетках-хозяевах, выделяется затем из культуры рекомбинантных клеток хроматографией или другими методами очистки.
В альтернативном случае фрагменты и аминокислотные варианты ФРЭККС приготовляются in vitro, например, путем протеолиза нативного ФРЭККС или синтезом с использованием стандартных методов твердофазного синтеза пептидов, соответственно описанию Меррифилда ["Журн. Амер. хим. об-ва" (J. Am. Chem. Soc.), 85: 2149 (1963)], хотя могут быть использованы и другие эквивалентные химические синтезы, известные в данной области. Твердофазный синтез инициируется с C-конца пептида путем присоединения защищенной аминокислоты к соответствующей смоле. Аминокислоты присоединяются к пептидной цепи с использованием стандартных методов для создания пептидных связей.
Терапевтическое использование
Для использования в терапевтической практике антагонисты, описанные в данном изобретении, вводятся млекопитающему, предпочтительно человеку, в фармацевтически приемлемой форме, включая введение внутривенно в виде шарика или постоянного вливания в течение некоторого периода времени, а также следующими путями: внутримышечной, интраперитонеальным, интрацереброспинальным, подкожным, внутрисуставным, внуртисиновиальным, внутриоболочечным, оральным, локальным или ингаляционным. Антагонисты также могут вводиться внутриопухолевым, околоопухолевым, внутриочаговым и околоочаговым путями для обеспечения локального действия наряду с системным терапевтическим действием. Ожидается, что особенно пригодным должно быть интраперитонеальное введение, например, для лечения опухолей яичников.
Подобные формы введения включают фармацевтически приемлемые носители, которые по своей природе не обладают ни токсическим, ни терапевтическим действием. Примерами таких носителей являются ионообменные вещества, квасцы, стеарат алюминия, лецитин, белки плазмы (такие, как белок плазмы человека), буферные вещества, такие как фосфаты, глицин, сорбиновая кислота, сорбат калия, частичные глицеридные смеси насыщенных овощных жирных кислот, вода, соли или электролиты, такие как сульфат протамина, гидрофосфат натрия, гидрофосфат калия, хлорид натрия, соли цинка, коллоидная окись кремния, трисиликат магния, поливинилпирролидон, вещества с целлюлозной основой и полиэтиленгликоль.
Носители для локальной или основанной на геле форм антагонистов включают полисахариды, такие как натриевая соль карбоксиметилцеллюлозы или метилцеллюлозы, поливинилпирролидон, полиакрилаты, полимеры полиоксиэтилен-полиоксипропиленового блока, полиэтиленгликоль и спирты. Для введения во всех случаях используются обычные лекарственные формы, получаемые со складов. К таким формам относятся, например, микрокапсулы, нанокапсулы, липосомы, пластыри, ингаляционные препараты, аэрозоли, подъязычные таблетки и препараты с постоянным высвобождением вещества. Антагонист в таких препаратах будет обычно содержаться в концентрации примерно от 0,1 мг/мл до 100 мг/мл.
Подходящие примеры препаратов с постоянным высвобождением вещества включают полупроницаемые матрицы твердых гидрофобных полимеров, содержащих антагонист; подобные матрицы имеют определенную форму, например это могут быть пленки или микрокапсулы. К примерам матриц с постоянным высвобождением относятся полиэфиры, гидрогели [например, поли(2-гидроксиэтил-метакрилат)] , описанные Лангером и др., "Ж. иссл. биомед. материалов" (J. Biomed. Mater. Res.), 15: 167 (1981) и Лангером, "Химич. технол." (Chem. Tech.), 12: 98-105 (1982), или поли(винилалкоголь), полилактиды (Патент США N 3773919), сополимеры L-глутаминовой кислоты и гаммаэтил-L-глутамата (Сидман и др. "Биополимеры" (Biopolymers), 22: 547 (1983), недеградируемый этиленвинилацетат (Лангер и др. см. выше), деградируемые сополимеры молочной и гликолевой кислот, такие как Lupron DepotТМ (инъецируемые микросферы, состоящие из полимеров молочной и гликолевой кислот и ацетата лейпролида), и поли-D-(-)- 3-гидроксибутировой кислоты. В то время, как такие полимеры, как этиленвинилацетат и cополимер молочной и гликолевой кислот, способны к постоянному высвобождению молекул в течение более 100 дней, определенные гидрогели высвобождают белки за более короткие периоды времени. Когда инкапсулированные полипептидные антагонисты остаются в организме на долгое время, они могут денатурировать или агрегироваться в результате воздействия влаги при температуре 37oC, что ведет к потере биологической активности и возможным изменениям в иммуногенности. С целью стабилизации могут быть разработаны разумные стратегии, в зависимости от действующего механизма. Например, если обнаружен механизм агрегации, выражающийся в формировании межмолекулярной S-S-связи посредством тиодисульфидного обмена, стабилизация может быть достигнута путем модификации сульфгидрильных остатков, лиофилизации с целью удаления кислых растворов, контролирования влажности, использования соответствующих добавок и разработки специфических полимерных матричных составов.
Антагонистические составы с постоянным высвобождением ФРЭККС включают также антагонистические антитела и рФРЭККС, заключенные в липосомах. Липосомы, содержащие антагонисты, могут быть получены известными в данной области методами, например, описанными Эпстейном и др., "Известия национальн. Акад. Наук" (Proc. Nat. Acad. Sci.), 82: 3688 (1985); Хуанг и др. "Известия национальн. Акад. Наук" (Proсс. Nat. Acad. Sci.), 77: 4030 (1980); Патент США N 4485045 и Патент США N 4544545. Липосомы, как правило, имеют небольшую величину (величиной около 200-800 ангстрем) и принадлежат к однослойному типу, в котором содержание липидов выше, чем 30 мол.% холестерина; выбранное соотношение может изменяться для подбора оптимальных условий терапии. Липосомы с продолжительным сроком циркуляции покрываются Патентом США N 5013556.
Еще одним путем использования данного изобретения является инкорпорирование антагониста ФРЭККС внутрь изделий, имеющих определенную форму. Такие изделия могут быть использованы для модулирования роста клеток эндотелия и ангиогенеза. Кроме того, такие изделия могут быть использованы для модулирования инвазии опухолей и метастазов.
При профилактике или лечении заболевания необходимая доза антагониста будет зависеть от типа заболевания, от его степени серьезности и протекания, от того, вводятся ли антитела с профилактической или терапевтической целью, от предыдущей терапии, от истории болезни пациента и его реакции на антагонист и от указаний лечащего врача. Антагонист может вводиться пациенту различными способами, единовременно или в качестве серии назначений.
Антагонисты ФРЭККС могут быть использованы для лечения различных неопластических и ненеопластических заболеваний и нарушений. Неоплазмы и близкие состояния, которые поддаются такому лечению, включают карциному груди, карциному легких, карциному желудка, карциному пищевода, колоректальную карциному, карциному печени, карциному яичников, текому, арренобластому, карциному шейки матки, карциному эндометрия, гиперплазию эндометрия, эндометриоз, фибросаркомы, хориосаркомы, рак головы и шеи, носоглоточную карциному, карциному гортани, гепатобластому, саркому Капоши, меланому, карциномы кожи, гемангиому, кавернозную гемангиому, гемангиобластому, карциному поджелудочной железы, ретинобластомы, астроцитому, глиобластому, шванному, олигодендроглиому, медуллобластому, нейробластому, рабдомиосаркому, остеогенную саркому, лейомиосаркому, карциномы мочеполового тракта, карциномы щитовидной железы, опухоль Вильмса, карциному клеток почки, карциному предстательной железы, аномальную пролиферацию сосудов, связанную с факоматозами, отеки (такие как отеки, связанные с опухолями мозга) и синдром Мейгса.
Ненеопластические заболевания, которые поддаются лечению, включают такие, как ревматоидный артрит, псориаз, атеросклероз, диабетические и другие ретинопатии, ретролентальная фиброплазия, неоваскулярнуя глаукома, тироидные гиперплазии (в том числе болезнь Граве), трансплантация роговицы и других тканей, хронические воспаления, воспаление легких, нефротический синдром, асцит, преэклампсия, перикардиальный выпот (например, связанный с перикардитом) и плевральный выпот.
В зависимости от типа заболевания и от степени его серьезности первоначальная доза для введения пациенту будет составлять от 1 мкг/кг до 15 мг/кг и может вводиться путем одного или многих отдельных введений или путем постоянного вливания. Обычная дневная доза может варьировать примерно от 1 мкг/кг до 100 мг/кг и более, в зависимости от вышеупомянутых факторов. Для повторного введения в течение нескольких дней и более, в зависимости от условий, лечение повторяется, пока не достигается желаемое подавление симптомов болезни. Однако могут использоваться и другие режимы дозировки. Успех лечения легко определяется обычными методами и анализами, например радиографической визуализацией опухолей.
В соответствии с другим применением изобретения эффективность антагониста в предотвращении или лечении болезней может быть улучшена путем введения антагониста серийно или же в комбинации с другим веществом, эффективным для данной цели, таким как фактор некроза опухоли (ФНО); антитело, способное ингибировать или нейтрализовать ангиогенную активность кислого или основного фактора роста фибробластов (ФРФ) или фактора роста гепатоцитов (ФРГ); антитело, способное ингибировать или нейтрализовать коагулянтную активность тканевого фактора, белка C или белка S (см. Эсмон и др. РСТ Патентная публикация N WO 91/01753, опубл. 21 февраля 1991 г.); или одно или более обычных терапевтических веществ, таких как, например, алкилирующие соединения, антагонисты фолиевой кислоты, антиметаболиты метаболизма нуклеиновых кислот, антибиотики, аналоги пиримидинов, 5-флюороурацил, пуриновые нуклеозиды, амины, аминокислоты, триазольные нуклеозиды или кортикостероиды. Подобные вещества могут присутствовать во вводимом составе или могут вводиться отдельно. Кроме того, антагонист может вводиться серийно или же в комбинации с радиологическим лечением, которое может включать как иррадиацию, так и введение радиоактивных веществ.
В соответствии с одним из применений изобретения при комбинированной терапии подвергается атаке васкуляризация опухоли. Один или более антагонист ФРЭККС вводятся пациенту с опухолью в терапевтически эффективных дозах, определенных, например, при наблюдении некроза опухоли или ее метастазных фокусов, если они имеются. Такая терапия продолжается до тех пор, пока перестает наблюдаться дальнейшее улучшение или клиническое обследование показывает, что опухоль или ее метастазы исчезли. Затем вводится ФНО, отдельно или в комбинации с дополнительным веществом, таким как альфа-, бета- или гамма-интерферон, антитело к HER2, херегулин, антитело к херегулину, D-фактор, интерлейкин-1 (IL-1), интерлейкин-2 (IL-2), фактор стимуляции колонии гранулоцитов-макрофагов (ФСК-ГМ) или вещества, которые способствуют микрососудистой когуляции в опухолях, таких как антитела к белкам C или S или связывающий белок C4b (см. Эсмон и др. PCT Патентная публикация N WO 91/01753, опубл. 21 февраля 1991 г.), или тепло и радиация.
Поскольку эффективность дополнительных веществ будет варьировать, желательно сравнить их влияние на опухоль путем стандартного матричного скрининга. Производится повторное введение антагониста ФРЭККС и ФНО, пока не будет достигнут желаемый клинический эффект. В альтернативном случае антагонист(ы) ФРЭККС и ФНО вводятся совместно и, при желании, вместе с дополнительными веществами. В случае, когда обнаруживаются твердые опухоли в суставах или других местах, более изолированных от общей циркуляции, описанные здесь терапевтические агенты вводятся в изолированную опухоль или орган. В других случаях совместно с антагонистом ФРЭККС пациенту вводятся антагонист ФРФ или тромбоцитный фактор роста (ТФР), такой как нейтрализующее антитело к ФРФ или ТФР. Применение антагонистов ФРЭККС при оптимальных обстоятельствах может быть приостановлено во время залечивания раны или желательной неоваскуляризации.
Прочие применения
Антитела к ФРЭККС, описываемые в данном изобретении, могут служить в качестве агентов для аффинной очистки. В этом процессе антитела к ФРЭККС иммобилизуются на подходящем носителе, таком как Сефадекс или фильтровальная бумага, с использованием известных методов. Иммобилизованное антитело затем приводится в контакт с образцом, содержащим ФРЭККС, который необходимо очистить; далее база промывается соответствующим растворителем, таким как глициновый буфер, pH 5,0, который освобождает ФРЭККС от антитела.
Нижеследующие примеры предлагаются только в качестве иллюстрации и не должны восприниматься как в чем-либо ограничивающие изобретение.
ПРИМЕР 1
Приготовление моноклональных антител к ФРЭККС
Для получения ФРЭККС, конъюгированного с гемоцианином морского блюдечка (ГМБ) для иммунизации, рекомбинантный ФРЭККС [165 аминокислот; Леунг и др. "Наука" (Science), 246: 1306 (1989)] смешивали с ГМБ в отношении 4:1 в присутствии 0,05% глютаральдегида и смесь инкубировали при комнатной температуре в течение 3 часов, слегка помешивая. Смесь была затем диализована против фосфатного буферного физиологического раствора (ФБРФ) при температуре 4oC в течение ночи.
Мыши линии Balb/c были иммунизованы четыре раза в течение каждых двух недель путем интраперитонеальных инъекций пятью мкг ФРЭККС, конъюгированными с 20 мкг ГМБ, и реиммунизированы той же дозой ФРЭККС, конъюгированного с ГМБ, за четыре дня до слияния клеток.
Клетки селезенки от иммунизованной мыши были слиты с клетками миеломы линии P3X63Ag8U.1. (Йелтон и др. "Текущие вопросы в микробиологии и иммунологии" (Curr. Top. Microbiol. Immunol.), 81: 1 (1978) с использованием 35% полиэтиленгликоля (ПЭГ). (Ярмуш и др. "Известия национальн. Акад. Наук" (Proc. Nat. Acad. Sci.), 77: 2899 (1980). Гибридомы отбирались в среде HAT.
Супернатанты культур гибридомных клеток подвергались скринингу на продукцию антитела к ФРЭККС; применялся твердофазный иммуносорбентный анализ ELISA с микротитровочными пластинками, покрытыми ФРЭККС. Антитело, которое привязывалось к ФРЭККС в каждой из лунок, определялось при помощи козьего антимышиного иммуноглобулина IgG, конъюгированного со щелочной фосфатазой, и хромогенного субстрата п-нитрофенилфосфата [Харлоу и Лейн, "Антитела: Лабораторное руководство" (Antibodies: A Laboratory Manual), с. 597 (Cold Spring Harbor Laboratory, 1988)]. Клетки гибридомы, которые, как было установлено этим методом, производят антитела к ФРЭККС, были субклонированы путем лимитирующего растворения, и два из этих клонов, названные A4.6.1 и B2.6.2, были выбраны для дальнейших исследований.
ПРИМЕР 2
Характеризация моноклональных антител к ФРЭККС
А. Специфичность по отношению к антигену.
Связывающие специфичности моноклональных антител к ФРЭККС, произведенных гибридомами A4.6.1 и B2.6.2, были определены методом ELISA. Моноклональные антитела были добавлены в лунки микротитровочных пластинок, которые до этого были покрыты ФРЭККС, ФРФ, ФРГ или эпидермальным фактором роста (ЭФР). Связанное антитело было обнаружено при помощи козьих антимышиных иммуноглобулинов IgG, конъюгированных с пероксидазой. Результаты данного анализа подтверждают, что моноклональные антитела, продуцированные гибридомами A4.6.1 и B2.6.2, связываются с ФРЭККС, но связывания их с прочими белковыми факторами роста не наблюдается.
Б. Картирование эпитопа.
Анализ конкурентного связывания ELISA использовался для того, чтобы определить, связываются ли моноклональные антитела, продуцированные гибридомами A4.6.1 и B2.6.2, к тем же или к различным эпитопам (участкам) в пределах ФРЭККС [Ким и др. "Инфекц. иммунол. " (Infect. Immunol), 57: 944 (1989)]. Индивидуальные немеченые моноклональные антитела к ФРЭККС (A4.6.1 и B2.6.2) или постороннее антитело к ФРГ (изотип IgG1) были добавлены в лунки микротитровочных пластинок, до этого покрытых только ФРЭККС. Затем были добавлены биотинилированные моноклональные антитела (ВIO-A4.6.1 и BIO-В2.6.2). Отношение биотинилированных моноклональных антител к немеченым моноклональным антителам было около 1:1000. Связывание биотинилированных моноклональных антител визуализировалось добавлением авидинконъюгированной пероксидазы, а затем о-фенилен-диаминдигидрохлорида и перекиси водорода. Цветная реакция, которая показывает, какое количество биотинилированных моноклональных антител связалось с эпитопом, требует измерения оптической плотности (O.D) на длине волны 495 нанометров.
Как показано на фиг. 1, в каждом случае связывание биотинилированного антитела к ФРЭККС было ингибировано соответствующим немеченым антителом к ФРЭККС или антителом к ФРГ. Эти результаты показывают, что моноклональные антитела, продуцированные гибридомами A4.6.1 и B2.6.2, связываются с различными эпитопами в пределах ФРЭККС
В. Изотипы.
Анализ ELISA использовался для того, чтобы определить моноклональные антитела, продуцированные гибридомами A4.6.1 и B2.6.2. Образцы культуральной среды (супернатанта), в которой росла каждая из гибридом, добавлялись в лунки микротитровочных пластинок, до этого покрытых ФРЭККС. Добытые моноклональные антитела к ФРЭККС инкубировались с различными изотипспецифичными козьими антимышиными иммуноглобулинами IgG, конъюгированного со щелочной фосфатазой, а связывание конъюгированных моноклональных антител к ФРЭККС было определено путем добавления n-нитрофенилфосфата. Цветная реакция измерялась при длине волны 405 нанометров с помощью счетчика на пластинке для анализа ELISA.
В результате применения этого метода был определен изотип (IgG1) моноклональных антител, продуцированных гибридомами A4.6.1 и B2.6.2.
Г. Связывающее сродство.
Сродство для моноклональных антител к ФРЭККС, продуцированных гибридомами A4.6.1 и В2.6.2, было определено путем анализа конкурентного связывания. Заранее известная субоптимальная концентрация моноклонального антитела была добавлена к образцам, содержащим 20000 - 40000 импульсов в минуту помеченного радиоактивным иодом 125I-ФРЭККС (1-2 нанограмма) и различные, известные количества немеченого ФРЭККС (1 - 1000 нг). После инкубации в течение часа при комнатной температуре было добавлено 100 мкл козьей антимышиной антисыворотки Ig (Пел-Фриз, Роджерс, Аризона, США), и смеси инкубировались в течение еще одного часа при комнатной температуре. Комплексы антитела и связанного белка (иммунные комплексы) были осаждены добавлением 500 мкл 6% полиэтиленгликоля (ПЭГ, мол. вес 8000) при 4oC, а затем центрифугированы при 2000 х G 20 мин 4oC. Количество связанного с моноклональным антителом 125I-ФРЭККС в каждом образце определялось подсчетом осадка на счетчике гамма-излучения.
Из этих данных были определены константы сродства путем Скатчард-анализа. Сродство моноклонального антитела к ФРЭККС, продуцированного гибридомой A4.6.1, было определено как 1,2 х 109 л/моль. Сродство моноклонального антитела к ФРЭККС, продуцированного гибридомой B2.6.2, было определено как 2,5 х 109 л/моль.
Д. Ингибирование митогенной активности ФРЭККС.
Бычьи клетки капиллярного эндотелия коры надпочечников (КЭКН) [Феррара и др. , "Известия национальн. Акад. Наук" (Proc. Nat. Acad. Sci.), 84: 5773 (1987)] были высеяны с плотностью 104 клеток/мл в 12-луночных пластинках. В каждую лунку было добавлено 2,5 нг/мл ФРЭККС в присутствии или в отсутствие различных концентраций моноклональных антител к ФРЭККС, продуцированных гибридомами A4.6.1 и В2.6.2, а также постороннего моноклонального антитела к ФРГ. После роста культуры в течение 5 дней клетки в каждой лунке были подсчитаны с помощью счетчика Коултера. В качестве контроля КЭКН выращивались в отсутствие ФРЭККС.
Как показано на фиг. 2, оба моноклональных антитела к ФРЭККС ингибировали способность добавленного ФРЭККС поддерживать рост и выживание бычьих КЭКН. Моноклональное антитело к ФРЭККС, продуцированное гибридомой A4.6.1, полностью ингибировало митогенную активность ФРЭККС (на более чем 90%), тогда как моноклональное антитело, продуцированное гибридомой В2.6.2, лишь частично ингибировало митогенную активность ФРЭККС.
Е. Ингибирование связывания ФРЭККС.
Бычьи клетки капиллярного эндотелия коры надпочечников (КЭКН) были высеяны с плотностью 2,5 • 104 клеток/0,5 мл/лунку в 24-луночных микротитровочных пластинках в СИМД (среда Игла, модифицированная Дулбекко), содержащей 10% плазмы теленка, 2 мМ глютамина и 1 нг/мл основного фактора роста фибробластов. После роста культуры в течение ночи клетки были один раз промыты связывающим буфером (равные объемы CИМД и среды F12 плюс 25 мМ HEPES и 1% альбумина бычьей плазмы) при 4oC.
12000 импульсов в минуту (ивм) 125I-ФРЭККС (около 5 • 104 ивм/нг/мл) были преинкубированы в течение 30 мин с 5 мкг моноклональных антител к ФРЭККС, продуцированных гибридомами A4.6.1, В2.6.2 или A2.6.1 (общий объем 250 мкл), после чего смеси были добавлены к КЭКН на микротитровочных пластинках. После инкубации клеток 3 часа при 4oC клетки были промыты 3 раза связывающим буфером при 4oC, переведены в раствор добавлением 0,5 мл 0,2 н. NaOH, и сигнал был подсчитан с помощью гамма-счетчика. Как показано на фиг. 3 (вверху), моноклональное антитело, продуцированное гибридомой
A2.6.1 и B2.6.2, прекращало прикрепление ФРЭККС к КЭКН. В противоположном случае моноклональные антитела к ФРЭККС, продуцированные гибридомами A4.6.1, не влияют на прикрепление ФРЭККС к КЭКН. Это согласуется с результатами, полученными путем анализа проницаемости клеток, описанной выше. Моноклональные антитела, продуцированные гибридомами A4.6.1, сильнее препятствовали прикреплению ФРЭККС, чем моноклональные антитела, продуцированные гибридомами B2.6.2.
Как показано на фиг. 3 (внизу), моноклональное антитело, продуцированное гибридомой A4.6.1, полностью ингибировало связывание ФРЭККС с КЭКН при соотношении ФРЭККС к антителу 1:250.
Ж. Перекрестняя реакция с другими изоформами ФРЭККС.
Для определения того, реагирует ли моноклональное антитело, продуцированное гибридомой A4.6.1, с 121- и 189-аминокислотными формами ФРЭККС, это антитело было проанализировано на предмет способности иммунопреципитировать подобные полипептиды.
Человеческие клетки 293 были трансфецированы векторами, включающими нуклеотидные кодрующие последовательности 121-й 189-аминокислотных полипептидов ФРЭККС, описанных Леунг и др., "Наука" (Science), 246: 1306 (1989). Спустя два дня после трансфекции, клетки были переведены в среду, лишенную цистеина и метионина. Клетки инкубировались в этой среде 30 минут, а затем были добавлены 35S-метионин и 35S-цистеин, 100 мкКюри/мл каждого. После мечения клетки перевели в среду, лишенную плазмы, и инкубировали три часа. Культуральная среда была собрана, и клетки были лизированы инкубацией 30 мин в лизирующем буфере (150 мМ NaCl, 1% NP40, 0,5% дезоксихолат, 0,1% додецилсульфат натрия (SDS), 50 мМ Трис, pH 8,0). Клеточные остатки были удалены из лизатов путем центрифугирования при 200 G в течение 30 мин.
Аликвоты 500 мкл культуральной среды и клеточных лизатов инкубировали вместе с 2 мкл моноклонального антитела, продуцированного гибридомой A4.6.1 (2,4 мг/мл) в течение 1 часа при 4oC, а затем инкубировали с 5 мкл антимышиного иммуноглобулина IgG кролика 1 час при 4oC. Иммунные комплексы меченого 35S ФРЭККС и моноклонального антитела к ФРЭККС были преципитированы на Сефарозе с белком А (Фармация), затем подверглись электрофорезу в 12% полиакриламидном геле с SDS в восстанавливающих условиях. Гель был экспонирован на рентгенограмму для анализа иммунопреципитированных белков, меченых радиоизотопами, путем авторадиографии.
Результаты этого анализа показали, что моноклональное антитело к ФРЭККС, продуцированное гибридомой A4.6.1, перекрестно реагировало с 121- и 189-аминокислотными формами ФРЭККС.
ПРИМЕР 3
Приготовление слитого белка "рецептор ФРЭККС-IgG".
Нуклеотидные и аминокислотные кодирующие последовательности рецептора ФРЭККС, flt, описаны Шибуя и др., "Онкоген" (Oncogene), 5: 519-524 (1990). Кодирующая последовательность внеклеточного домена рецептора ФРЭККС, flt, была слита с кодирующей последовательностью тяжелой цепи человеческого IgG1 в два приема.
Сайт-направленный мутагенез применялся для того, чтобы включить рестрикционный сайт BstBI в ДНК, кодирующую flt, в 5'-сайте к кодону для аминокислоты 759 flt и перевести единственный BstEII рестрикционный сайт в плазмиде pBSSKFC в сайт BstBI [Беннетт и др. "Журн. биол. химии" (J. Biol. Chem. ), 266: 23060-23067 (1991)]. Модифицированная плазмида была обработана EcoRI и BstBI, и получившийся большой фрагмент плазмидной ДНК был соединен с EcoRI-BstBI фрагментом ДНК flt, кодирующей внеклеточный домен (аминокикслоты 1-758) рецептора ФРЭККС, flt.
Полученная конструкция была обработана ClaI и NotI; был получен фрагмент длиной примерно 3,3 kb, который был путем лигирования вставлен в множественный клонирующий сайт вектора экспрессии млекопитающих pHЕВO2 [Леунг и др. "Нейрон" (Neuron), 8: 1045 (1992)]. Концы фрагмента длиной 3,3 kb могут быть модифицированы, например, посредством добавления линкеров, чтобы вставка фрагмента в вектор могла быть ориентирована в правильном направлении для экспрессии.
Клетки-хозяева млекопитающих, например клетки CEN4 (Леунг и др. см. выше), трансфецировались плазмидой pHЕВO2, содержащей вставку flt, путем электропорации. Трансфецированные клетки выращивались в среде, содержащей около 10% эмбриональной бычьей плазмы, 2 мМ глютамина и антибиотики; около 75% клеток были перенесены в среду, свободную от плазмы. Среда кондиционировалась 3-4 дня до того, как собирались клетки, и слитый белок "flt-IgG" был выделен из кондиционированной среды хроматографией на аффинной матрице с белком А, в целом согласно описанию Беннетта и др. "Журнал биологической химии" (J. Biol. Chem.), 266: 23060-23067 (1991).
ПРИМЕР 4
Ингибирование роста опухоли антагонистами ФРЭККС.
Различные человеческие линии опухолевых клеток анализировались методом ELISA на продукцию ФРЭККС. Было обнаружено, что ФРЭККС производится в клетках опухолей яичников, легких, прямой кишки, желудка, груди и мозга. Три линии опухолевых клеток продуцировали ФРЭККС: NEG55 (G55) (линия клеток глиомы человека получена от Д-ра М. Вестфал, Отделение нейрохирургии, Университ, Госпиталь Эппендор, Гамбург, Германия), А-673 (линия клеток рабдомиосаркомы человека получена из Американской коллекции типовых культур (АКТК), Роквилл, Мэриленд, США 20852, номер линии CRL 1598) и SK-LMS-1 (линия клеток лиомиосаркомы человека получена из АКТК, номер линии НТВ 88); эти линии были использованы для дальнейших исследований.
Самкам мышей (Beige/nude) возрастом от 6 до 10 недель (полученные из лаборатории Чарльз Ривер, Уилмингтон, Массачусеттс, США) были подкожно введены 1-5 • 106 опухолевых клеток в 100-200 мкл физиологического раствора с фосфатным буфером (ФРФБ). В различное время после того, как установился рост опухоли, мышам вводили, интраперитонеально, один или два раза в неделю различные дозы моноклонального антитела к ФРЭККС, продуцированного гибридомой A4.6.1, постороннего моноклонального антитела к gp120 (5В6) или ПБС. Величина опухоли измерялась каждую неделю, и по завершении исследования опухоли были вырезаны и взвешены.
Эффект различных доз моноклонального антитела к ФРЭККС, продуцированного гибридомой A4.6.1, на рост опухолей NEG55 показан у мышей на фиг. 4 и 5. Фиг. 4 показывает, что у тех мышей, которым было введено от 25 до 100 мкг моноклонального антитела к ФРЭККС, продуцированного гибридомой A4.6.1, начиная с одной недели после инокуляции клетками NEG55, скорость роста опухолей была значительно более медленной, чем у мышей, которым вводили постороннее моноклональное антитело или ФРФБ. Фиг.5 показывает, что к тому моменту, когда прошло пять недель после инокуляции клетками NEG55, величина опухоли у мышей, которым было введено моноклональное антитело к ФРЭККС, продуцированное гибридомой A4.6.1, была на 50% (в случае, когда мыши получали 25 мкг антитела) или на 85% (в случае, когда мыши получали 100 мкг антитела) меньше, нежели у мышей, которым вводили постороннее моноклональное антитело или ФРФБ.
Эффект моноклонального антитела к ФРЭККС, продуцированного гибридомой A4.6.1, на рост опухолей SK-LMS-1 у мышей показан на фиг. 6. К тому моменту, когда прошло пять недель после инокуляции клетками SK-LMS-1, средняя величина опухоли у мышей, которым было введено моноклональное антитело к ФРЭККС, продуцированное гибридомой A4.6.1, была на 75% меньше, нежели у мышей, которым вводили постороннее моноклональное антитело или ФРФБ.
Эффект моноклонального антитела к ФРЭККС, продуцированного гибридомой A4.6.1, на рост опухолей A673 у мышей показан на фиг.7. К тому моменту, когда прошло четыре недели после инокуляции клетками A673, средняя величина опухоли у мышей, которым было введено моноклональное антитело к ФРЭККС, продуцированное гибридомой A4.6.1, была на 60% (в случае, когда мыши получали 10 мкг антитела) до более чем на 90% (в случае, когда мыши получали 50 - 400 мкг антитела) меньше, нежели у мышей, которым вводили постороннее моноклональное антитело или ФРФБ.
ПРИМЕР 5
Анализ непосредственного эффекта антитела к ФРЭККС на рост клеток опухоли в культуре.
Клетки глиобластомы человека NEG55 или клетки рабдомиосаркомы человека А-673 были высеяны с плотностью 7 • 103 клеток/лунку в многолуночных пластинках (12 лунок на пластинку) в среде F12/СИМД, содержащей 10% эмбриональной плазмы теленка, 2 мМ глютамина и антибиотики. К культурам клеток было добавлено моноклональное антитело к ФРЭККС, продуцированное гибридомой A4.6.1; его конечная концентрация была от 0 до 20,0 мкг/мл. По истечении 5 дней клетки, растущие в лунках, были диссоциированы с помощью трипсина и подсчитаны с помощью счетчика Коултера.
Результаты этого анализа показаны на фиг. 8 и 9. Очевидно, что моноклональное антитело к ФРЭККС, продуцированное гибридомой A4.6.1, не имело сколько-нибудь значительного эффекта на рост клеток NEG55 или A-673 в культуре. Эти результаты свидетельствуют, что моноклональное антитело к ФРЭККС, продуцированное гибридомой A4.6.1, не имеет цитотоксических свойств, и с уверенностью подтверждают, что наблюдаемый эффект антитела на рост опухолей совершается за счет ингибирования неоваскуляризации, опосредованной ФРЭККС.
ПРИМЕР 6
Эффект антитела к ФРЭККС на хемотаксис эндотелиальных клеток.
Хемотаксис эндотелиальных клеток, а также иных клеток, включая моноциты и лимфоциты, играет важную роль в патогенезе ревматоидного артрита. Миграция эндотелиальных клеток и их пролиферация сопровождают ангиогенез, происходящий в ревматоидной синовиальной оболочке. Ткань, насыщенная кровеносными сосудами (паннус), вторгается в хрящ сустава и разрушает его.
Чтобы определить, интерферируют ли антагонисты ФРЭККС с этим процессом, проанализировали эффект антитела к ФРЭККС, продуцированного гибридомой A4.6.1, на хемотаксис эндотелиальных клеток, стимулированный синовиальной жидкостью, взятой у пациентов с ревматоидным артритом. В качестве контроля проанализировали эффект антитела к ФРЭККС, продуцированного гибридомой А4.6.1, на хемотаксис эндотелиальных клеток, стимулированный синовиальной жидкостью, взятой у пациентов с остеоартритом (ангиогенез, происходящий при ревматоидном артрите, не имеет места при остеоартрите).
Хемотаксис эндотелиальных клеток анализировался в модифицированных камерах Бойдена согласно стандартным методам [Томпсон и др."Раковые исследования" (Cancer Res.), 51: 2670 (1991); Филлипс и др. "Изв. эскпер. биол. и мед. "(Proc. Exper. Biol. Med.), 197: 458 (1991)]. Около 104 эндотелиальных клеток пуповины человека были нанесены на фильтры, покрытые желатином (размер пор 0,8 микрон) в многолуночных (48 лунок) микрокамерах в среде, содержащей 0,1% эмбриональной плазмы теленка. По истечении двух часов камеры были перевернуты, и были добавлены образцы (синовиальная жидкость при ревматоидном артрите, синовиальная жидкость при остеоартрите, основной ФРФ (оФРФ) (до конечной концентрации 1 мкг/мл) или ФРФБ и моноклональное антитело к ФРЭККС, продуцированное гибридомой A4.6.1 (до конечной концентрации 10 мкг/мл). По истечении еще двух-трех часов все мигрировавшие клетки были окрашены и подсчитаны.
Результаты этого исследования показаны на фиг. 10. Значения, показанные в столбце, обозначенном "син. жидк.", а также данные внизу страницы показывают среднее количество эндотелиальных клеток, которые мигрировали в присутствии синовиальной жидкости, оФРФ или только ФРФБ. Значения, показанные в столбце, обозначенном "син. жидк. + а-т ФРЭККС", показывают среднее количество эндотелиальных клеток, которые мигрировали в присутствии синовиальной жидкости и добавленного антитела к ФРЭККС, продуцированного гибридомой A4.6.1. Значения, показанные в столбце, обозначенном "% подавления", отражают процентное сокращение миграции эндотелиальных клеток, вызванной присутствием синовиальной жидкости, при добавлении антитела к ФРЭККС. Как видно из этих данных, антитело к ФРЭККС существенно ингибировало свойства синовиальной жидкости при ревматоидном артрите (средний процент ингибирования 53.40), но не остеоартритной синовиальной жидкости (средний процент ингибирования 13.64) вызывать миграции эндотелиальных клеток.
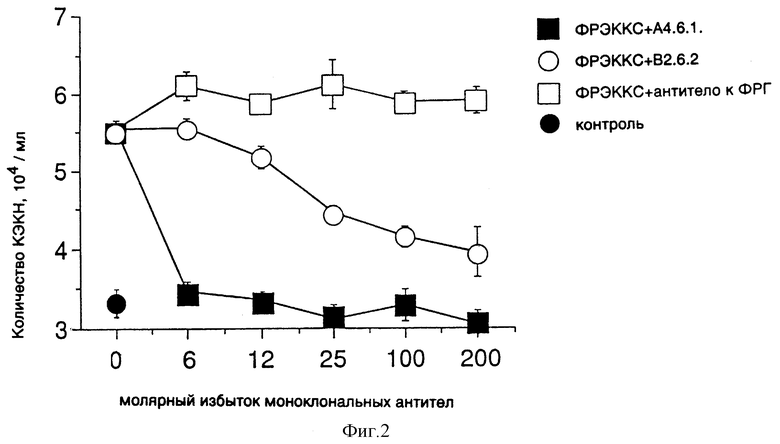
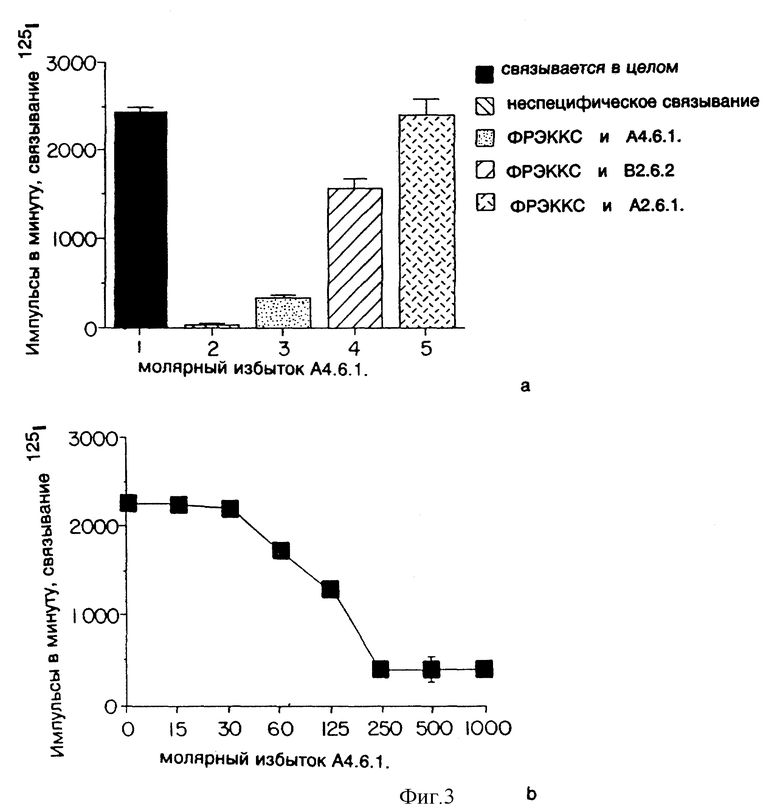
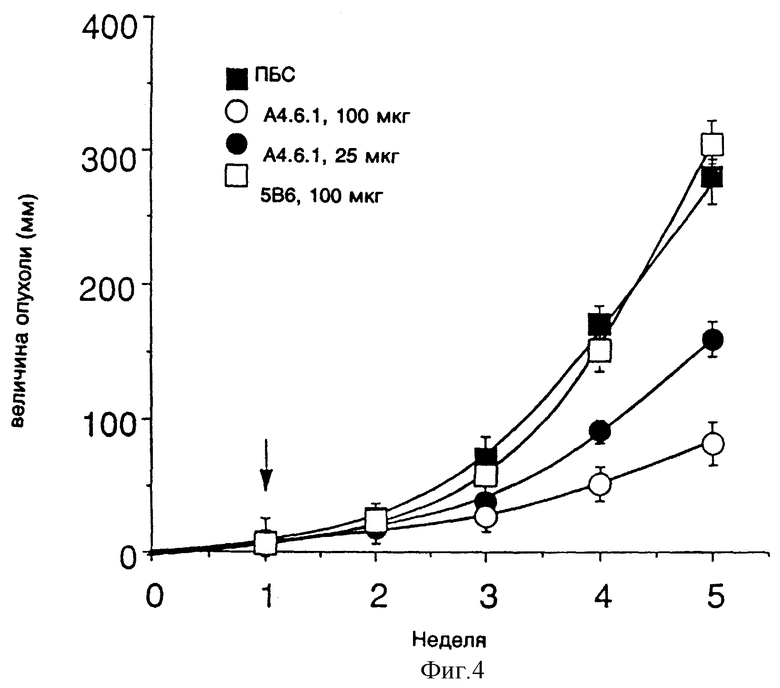
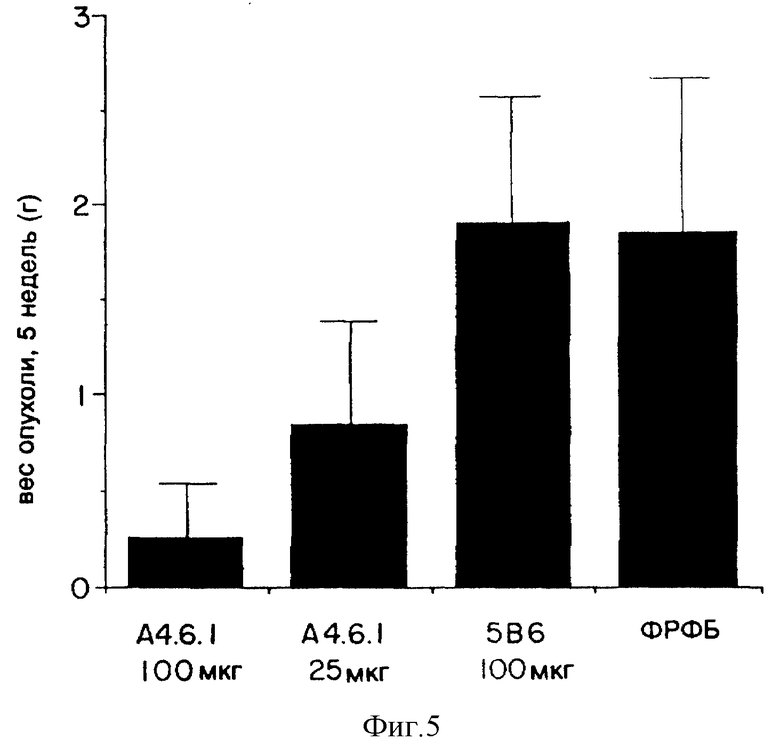
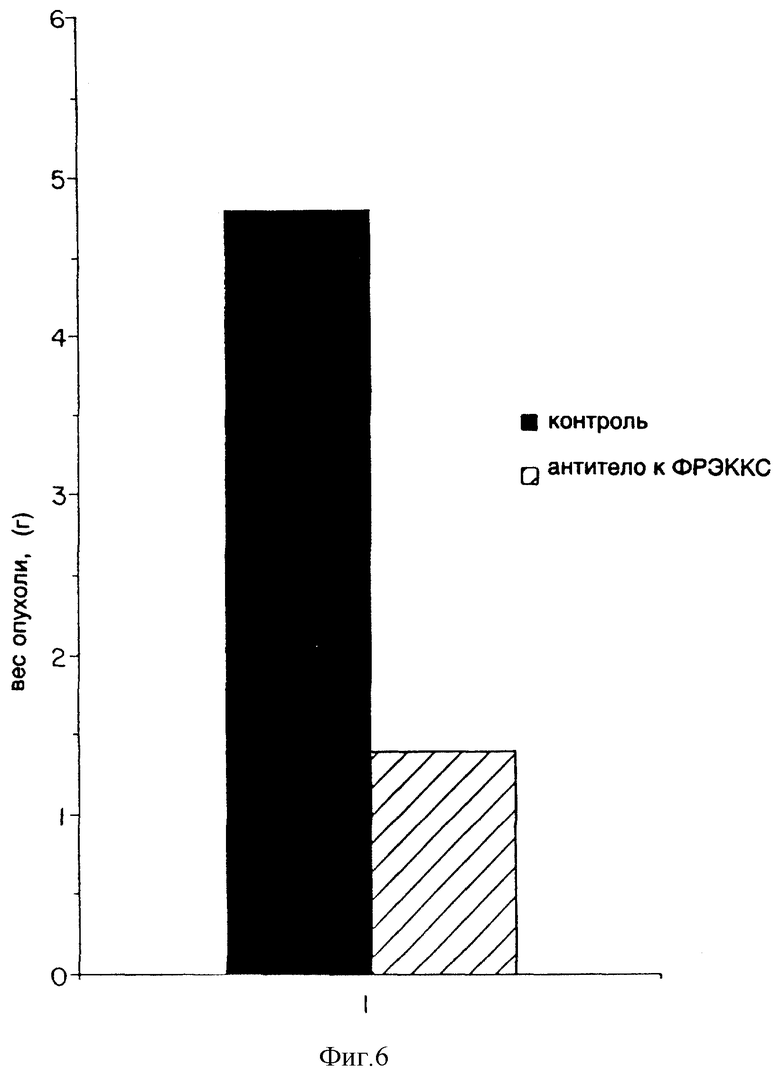
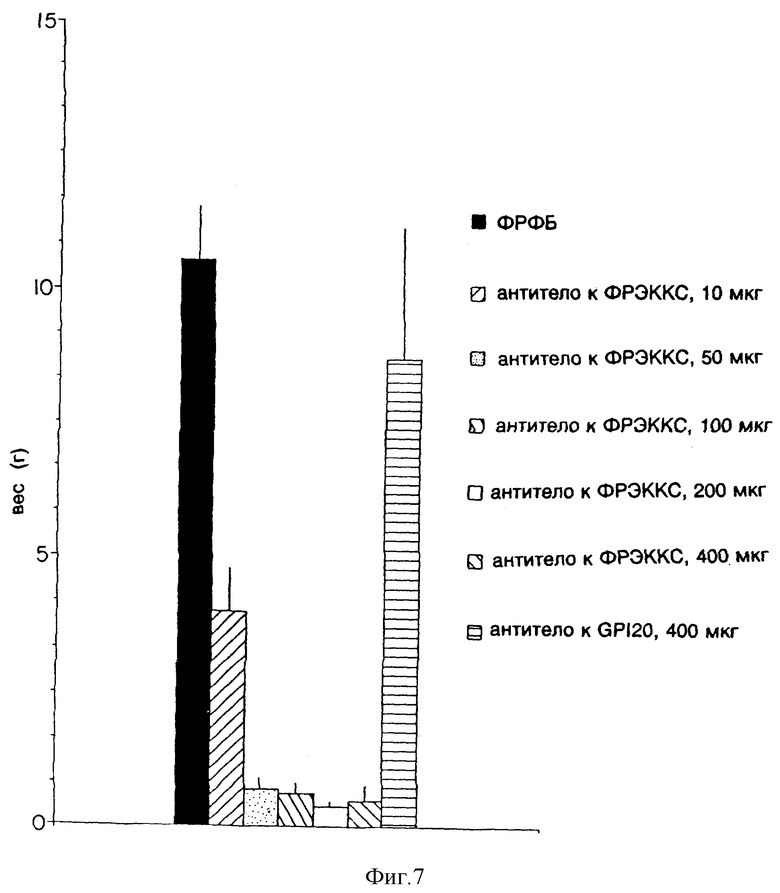
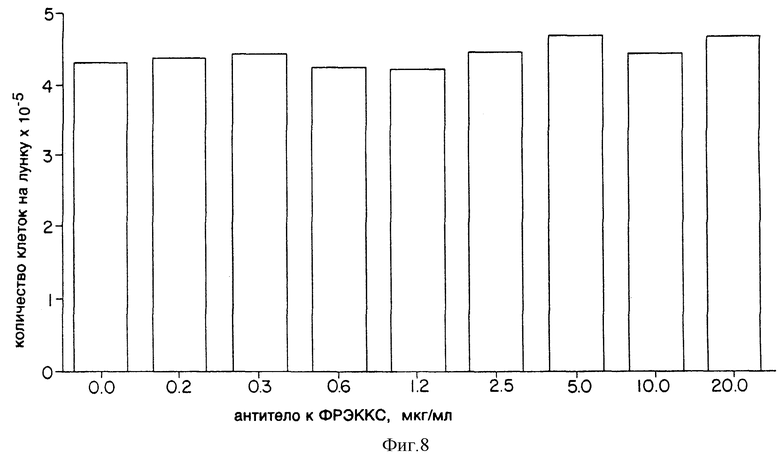
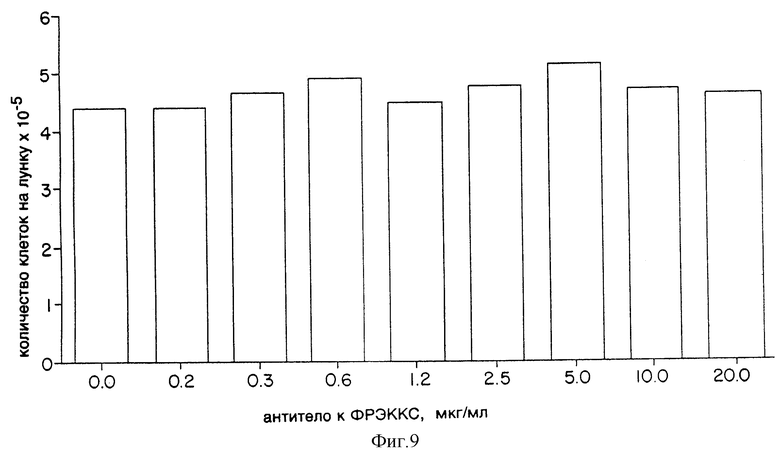
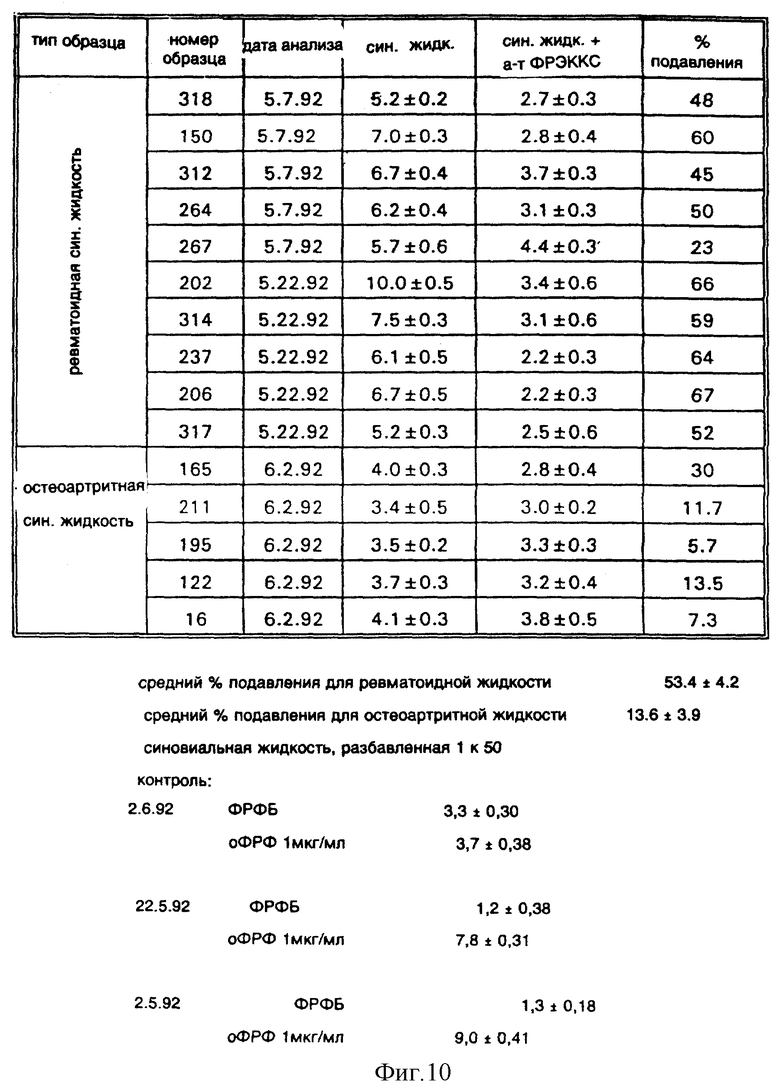
Изобретение относится к области медицины и касается композиции для ингибирования ангиогенеза, моноклонального антитела, полипептида и способа ингибирования роста опухоли. Изобретение включает антагонисты фактора роста эндотелиальных клеток кровеносных сосудов человека (hVEGFr), в том числе моноклональные антитела, рецепторы вышеуказанных антагонистов, варианты этих антагонистов, ингибирующих ангиогенную и митогенную активности антагонистов hVEGFr. Антагонисты могут использоваться для ингибирования роста опухоли, а также процессов, связанных с избыточной пролиферацией и неоваскуляризацией. Преимущество изобретения заключается в разработке новых препаратов для решения проблем, связанных с избыточной пролиферацией и неоваскуляризацией. 5 с. и 16 з.п.ф-лы, 10 ил.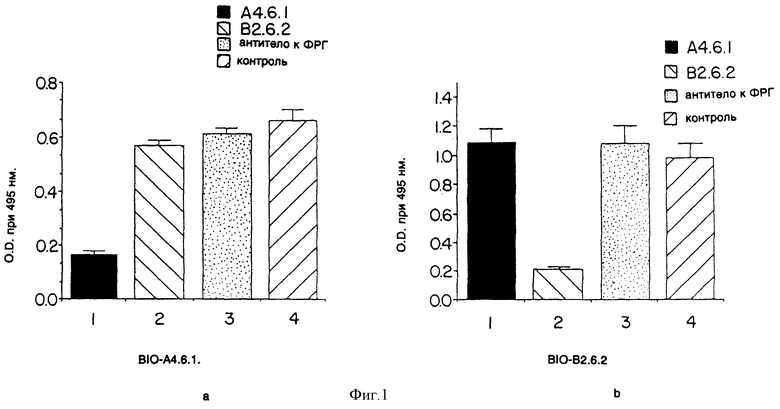
| Штамм гибридных культивируемых клеток животных MUS мUSсULUS L. - продуцент моноклонального антитела к антигену мембран жировых глобул женского молока | 1989 |
|
SU1698287A1 |
| МАРТИН D.S | |||
| et al., Role of Murine tumor model in cancer treatment research, Cancer Research, 1986, v.46, pp.2189 - 2192 | |||
| McKenzie S.J | |||
| Generation and characterization of monoclonal antibodyes specific for the human new oncogene product, Oncogene | |||
| Механизм для сообщения поршню рабочего цилиндра возвратно-поступательного движения | 1918 |
|
SU1989A1 |
