Настоящее изобретение относится к способу конструирования и получения мутантной трансглутаминазы на основе пространственной структуры трансглутаминазы, полученной из Streptoverticillium mobaraense (здесь и далее обозначаемой как "MTG"), определенной с помощью методики рентгеноструктурного анализа, и полученным таким образом мутантным MTG. MTG широко применяется для переработки продуктов питания или тому подобное, так как она формирует гелеобразное вещество путем образования поперечных сшивок между белками. Мутантная MTG с усиленной трансглутаминазной активностью и термостабильностью, призвана снизить ее требуемое количество, а также мутантная MTG с модифицированными субстратной специфичностью и оптимальным значением pH позволит применять фермент в новой области.
Трансглутаминаза представляет собой фермент, катализирующий реакцию переацетилирования γ-карбоксамидной группы, содержащейся в пептидной последовательности белка. При взаимодействии белка с указанным ферментом может происходить реакция образования поперечных сшивок типа ε-(γ-Glu)-Lys и реакция замещения Gln на Glu путем дезамидирования.
Пока известными являются трансглутаминазы животных и микроорганизмов. Первые являются Ca2+-зависимыми ферментами и широко представлены в 20 внутренних органах, коже и крови животных. Они включают, например, трансглутаминазу печени морских свинок [K. Ikura et al., Biochemistry, Vol.27, p.2898 (1988)], трансглутаминазу эпидермальных кератиноцитов человека [M. A. Phillips et al., Proc. Natl. Acad. Sci. USA, Vol.87, p.9333 (1990)] и фактор XIII системы свертывания крови человека [A. Ichinose et al., Biochemistry, Vol.25, p.6900 (1990)]. Что касается последних, то Ca2+-независимые ферменты были обнаружены у бактерий класса Streptoverticillium, таких как Streptoverticillium griseocarneum IFO 12776, Streptoverticillium cinnamoneum подвида cinnamoneum IFO 12852 и Streptoverticillium mobaraense IFO 13819. Среди указанных ферментов трансглутаминаза, обнаруженная в культуральном супернатанте варианта Streptoverticillium mobaraense, обозначается как MTG (Микробная Трансглутаминаза). Ca2+-независимая трансглутаминаза была также обнаружена у Streptomyces lydicus NRRL B-3446 (JP-Kokai No.10-504721).
MTG представляет собой состоящий из 331 аминокислоты мономерный белок с молекулярной массой, равной приблизительно 38000 [Journal of Biological Chemistry, vol.268, p.11565 (1993)]. Ранее сообщалось о способе получения активной MTG путем экспрессии секреции с применением E.coli или дрожжей (JP-Kokai No.Hei 5-199883). Также сообщалось о способе получения активной MTG, в котором MTG экспрессируется в E.coli в виде белкового тельца включения, затем тельце включения солюбилизируется с денатурирующим белки агентом, а затем регенерируется путем удаления денатурирующего агента с получением активной MTG (JP-Kokai No.6-30771).
В отличие от трансглутаминаз животных трансглутаминазы микроорганизмов, такие как MTG, являются Ca2+-независимыми. Поэтому они применяются для производства гелеобразных продуктов питания, таких как желе, йогуртов, сыров или косметических средств в гелевой форме, а также для улучшения качества мясных продуктов (JP-Kokai No.64-27471). Они широко применяются в промышленности в качестве материалов для производства термостабильных микрокапсул или в качестве носителей для иммобилизированных ферментов.
Что касается условий протекания ферментативной реакции, то, например, студнеобразные продукты питания не могут быть превращены в гелевую форму, если время протекания ферментативной реакции является коротким, и наоборот, применение в качестве коммерческих продуктов затруднено, если время протекания ферментативной реакции является слишком большим. Поэтому, если MTG применяется для производства гелеобразных продуктов питания, таких как желе, йогуртов, сыров или косметических средств в гелевой форме, или для улучшения качества мясных продуктов, для получения предполагаемого продукта должны соответствующим образом регулироваться концентрации субстрата и фермента, температура реакции и время протекания реакции. Однако в связи с различием получаемых с применением MTG продуктов питания и реагентов случается, что для получения предполагаемых продуктов в некоторых случаях бывает недостаточно регулировать только концентрацию, температуру, время и тому подобное. Поэтому появляется необходимость в модификации ферментативной активности MTG.
Для модификации ферментативной активности MTG необходимо получить мутантную MTG и для выбора хорошего мутанта определить его активность, субстратную специфичность и тому подобное. Для получения мутантов необходимо воздействовать на гены дикого типа, и, соответственно, это является предпосылкой возможного получения рекомбинантного белка. Для MTG была установлена высокопродуктивная экспрессирующая система с применением E.coli (JP-Kokai No.6-30771). Однако MTG, экспрессированная подобным образом в большом количестве в E.coli, накапливается в микробных клетках в виде нерастворимых белковых телец включения. После их солюбилизации денатурированная MTG в белковых тельцах включения должна быть активирована путем ее повторной укладки. Таким образом, для получения одной мутантной MTG требуется от 2 до 3 недель. Поэтому требовалось разработать способ приемлемой модификации MTG с применением некой методики, отличной от методик случайно индуцированных мутаций в целой молекуле.
Краткое описание изобретения
Целью настоящего изобретения являлся способ усовершенствования MTG.
Другой целью настоящего изобретения являлась трансглутаминаза, обладающая усиленной реакционной способностью по отношению к субстратам.
В результате интенсивных исследований, выполненных с целью решения описанных выше задач, авторы изобретения обнаружили, что трансглутаминаза, обладающая усиленной реакционной способностью по отношению к субстрату, может быть получена на основе пространственной структуры. Настоящее изобретение было совершено на основе этого открытия.
А именно, краткое содержание настоящего изобретения представляет собой следующее:
(1) Способ конструирования и получения мутантной трансглутаминазы путем предсказания сайта связывания MTG с субстратом на основе пространственной структуры трансглутаминазы, определенной с помощью рентгеноструктурного анализа, и замены, вставки или делеции аминокислотных остатков, расположенных в предсказанном субстратсвязывающем сайте трансглутаминазы.
(2) Обладающая модифицированной субстратной специфичностью мутантная MTG, в которой, по крайней мере, одна из следующих позиций (остатки, расположенные в пределах 20 Å от активного остатка Cys64 и находящиеся на поверхности предположительного сайта связывания с акцептором ацильной группы) в аминокислотных последовательностях MTG, обозначенных как SEQ ID NO: 2, является модифицированной: 1-5, 26, 28, 58-59, 62, 69, 74-75, 77, 79, 235-236, 238-244, 248-250, 252-254, 277-278, 282-287, 289, 291, 296-297 и 300-304.
(3) Обладающая трансглутаминазной активностью и модифицированной субстратной специфичностью мутантная трансглутаминаза, в которой при сравнении аминокислотной последовательности указанной трансглутаминазы с аминокислотной последовательностью MTG по SEQ ID NO: 2, по крайней мере, одна из позиций, соответствующая следующим позициям (остатки, расположенные в пределах 20 Å от активного остатка Cys64 и находящиеся на поверхности предположительного сайта связывания с акцептором ацильной группы) в аминокислотных последовательностях MTG по SEQ ID NO: 2, является модифицированной: 1-5, 26, 28, 58-59, 62, 69, 74-75, 77, 79, 235-236, 238-244, 248-250, 252-254, 277-278, 282-287, 289, 291, 296-297 и 300-304.
(4) Обладающая трансглутаминазной активностью и модифицированной субстратной специфичностью мутантная трансглутаминаза, в которой при сравнении методом тридинга аминокислотной последовательности указанной трансглутаминазы с аминокислотной последовательностью MTG по SEQ ID NO: 2, по крайней мере, одна из позиций, соответствующая следующим позициям (остатки, расположенные в пределах 20 Å от активного остатка Cys64 и находящиеся на поверхности предположительного сайта связывания с акцептором ацильной группы) в аминокислотных последовательностях MTG по SEQ ID NO: 2, является модифицированной: 1-5, 26, 28, 58-59, 62, 69, 74-75, 77, 79. 235-236, 238-244, 248-250, 252-254, 277-278, 282-287, 289, 291, 296-297 и 300-304.
(5) Мутантная MTG или мутантная трансглутаминаза, обладающая модифицированной субстратной специфичностью вследствие снижения отрицательной заряженности путем модификации кислых аминокислотных остатков, расположенных в позициях, определенных любым из описанных выше пп.(2)-(4), или аминокислотных остатков, прилегающих к упомянутым кислым аминокислотным остаткам.
(6) Ген, кодирующий мутантную MTG или мутантную трансглутаминазу, определенную любым из описанных выше пп.(2)-(5).
(7) Рекомбинантная ДНК, содержащая ген, определенный в описанном выше п.(6).
(8) Микроорганизм, несущий рекомбинантную ДНК, определенную в описанном выше п.(7).
(9) Способ получения мутантной MTG или мутантной трансглутаминазы, включающий культивирование описанных в п.(8) микроорганизмов и сбор мутантной MTG или мутантной трансглутаминазы.
(10) Моноклинный кристалл MTG, принадлежащий к спейсерной группе P21.
А именно, настоящее изобретение относится к способу рационального конструирования и получения мутантной MTG на основе пространственной структуры MTG и полученной таким образом мутантной MTG.
Краткое описание фигур
Фигура 1 представляет собой ленточную модель, отображающую кристаллическую структуру MTG.
Фигура 2 отображает поверхностную неровность кристаллической структуры MTG, наблюдаемую в том же направлении, что и на Фигуре 1.
Фигура 3 отображает сравнения аминокислотных последовательностей MTG и трансглутаминазы из Streptoverticillium cinnamoneum.
Фигура 4 отображает сравнения аминокислотных последовательностей MTG и трансглутаминазы из Streptoverticillium lydicus.
Фигура 5 отображает наборы праймеров, применяемые для сайтспецифического мутагенеза S2Y·S2R·S2D мутантов.
Фигура 6 отображает наборы праймеров, применяемые для сайтспецифического мутагенеза del 1-2 и del 1-3 мутантов.
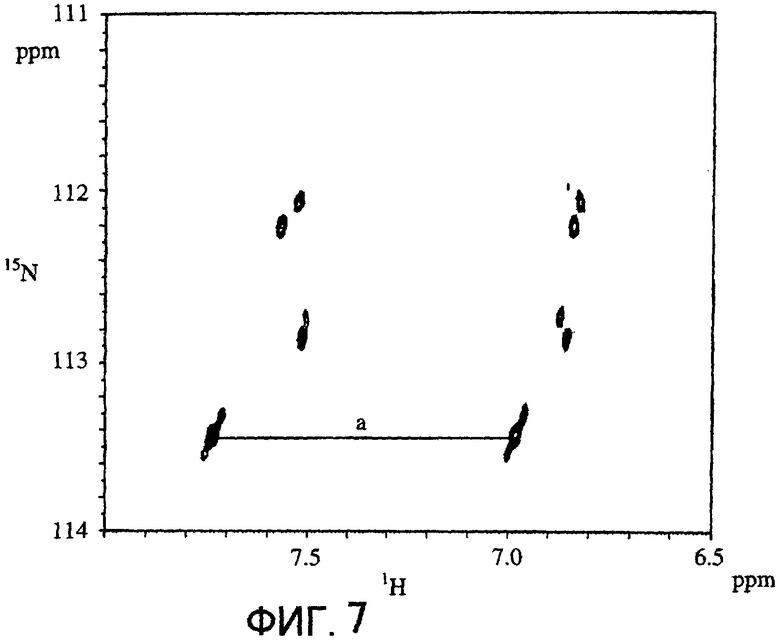
Фигура 7 отображает 1H-15N HSQC спектр овальбумина, вступившего во взаимодействие с диким типом в присутствии 15NH4Cl.
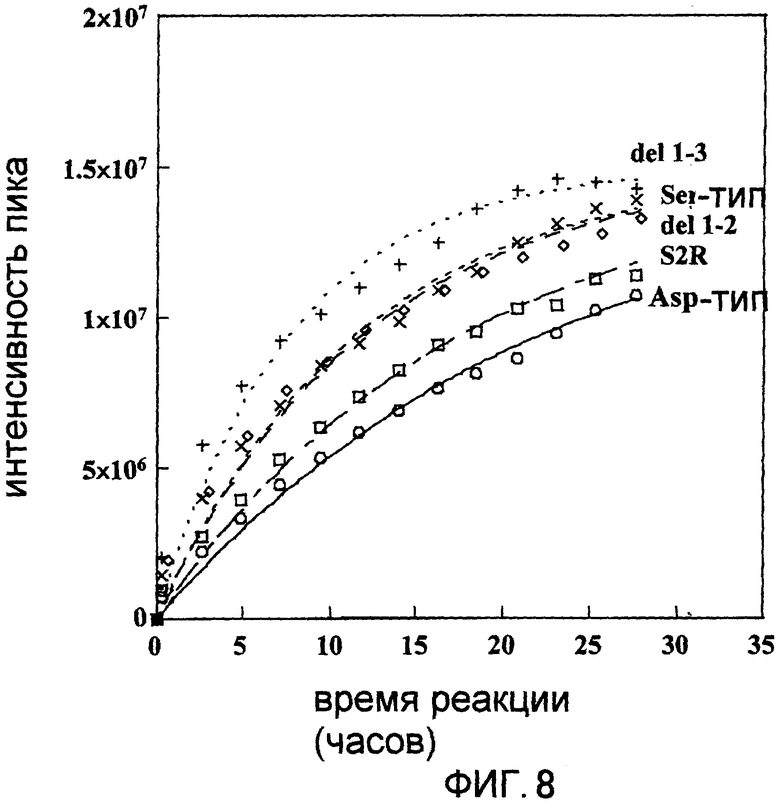
Фигура 8 отображает величину интенсивности пика сигнала "a" на Фигуре 7 в глутаминовых остатках овальбумина, вступившего во взаимодействие с диким типом, Ser-типом, S2R, del 1-2 или del 1-3 мутантами в присутствии 15NH4Cl, расположенную относительно времени реакции.
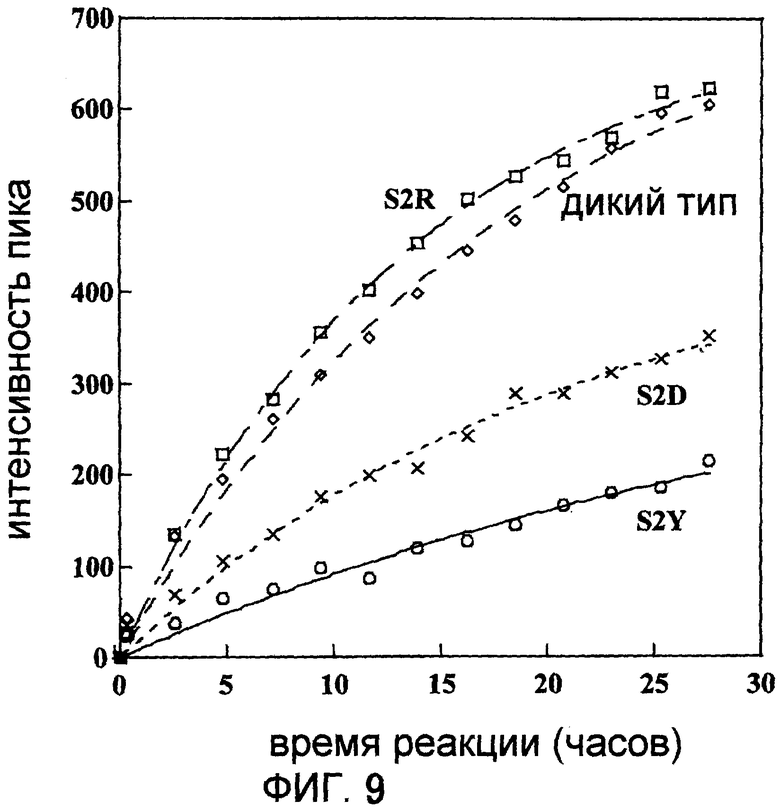
Фигура 9 отображает величину интенсивности пика сигнала "a" на Фигуре 7 в глутаминовых остатках овальбумина, вступившего во взаимодействие с диким типом, S2Y, S2R или S2D мутантами в присутствии 15NH4Cl, расположенную относительно времени реакции.
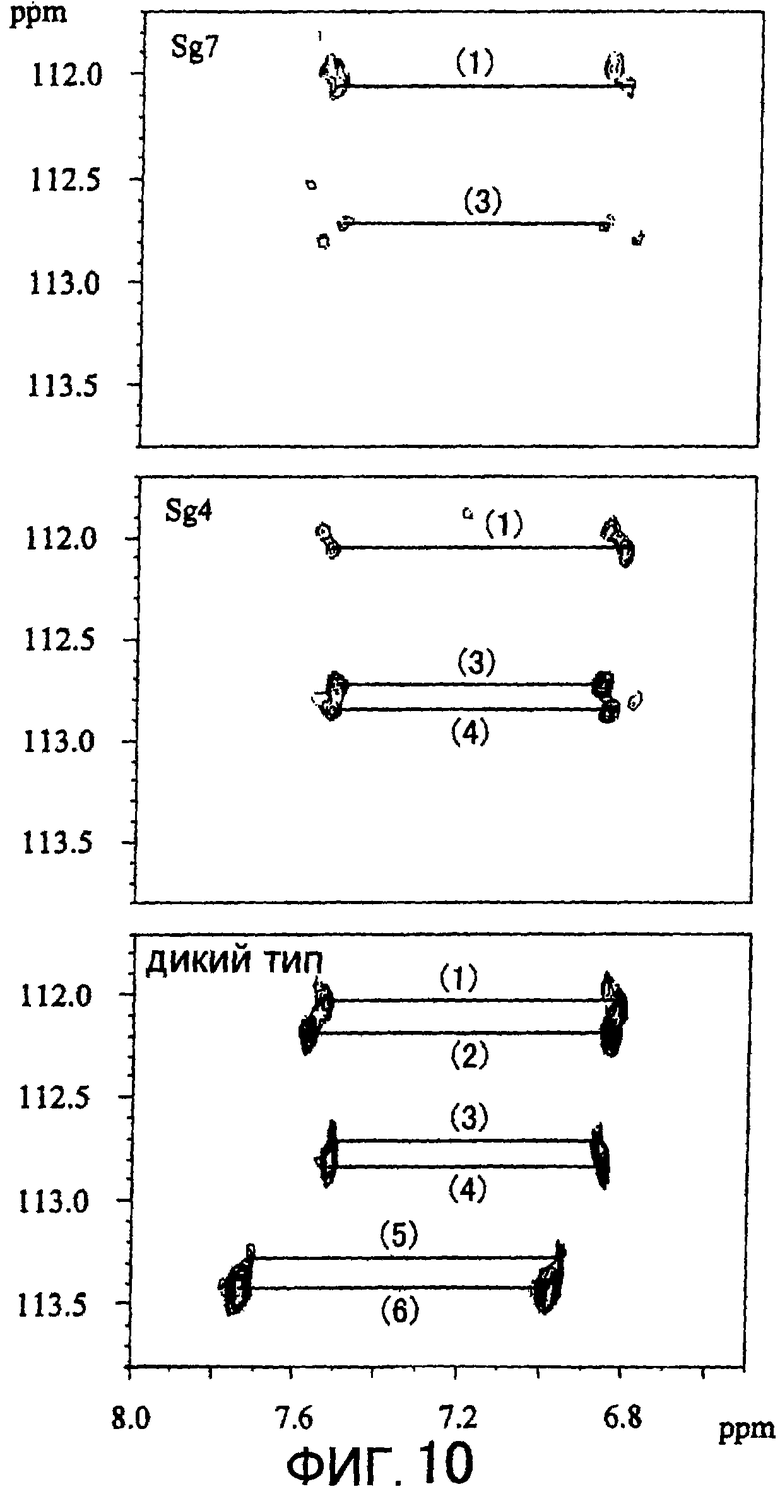
Фигура 10 отображает 1H-15N HSQC спектр овальбумина, вступившего во взаимодействие с диким типом, Sg4 или Sg7 мутантами в присутствии 15NH4Cl.
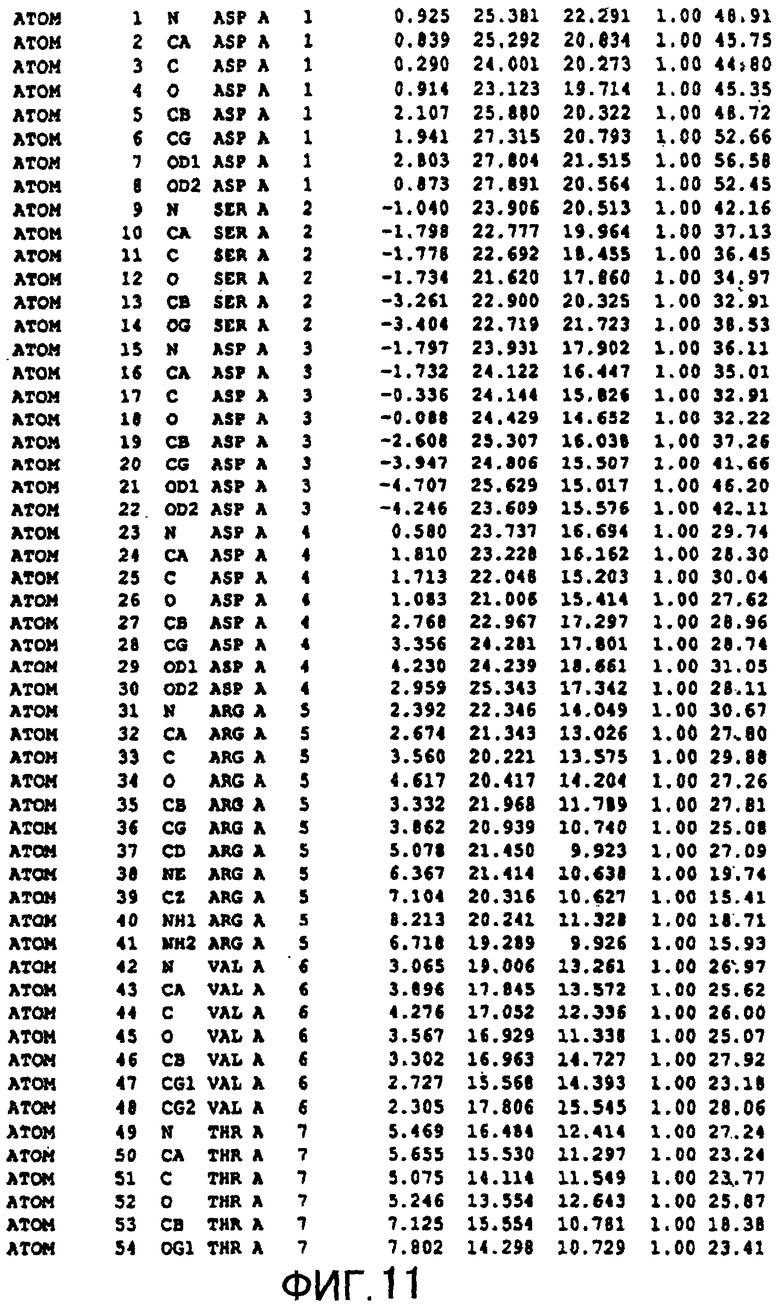
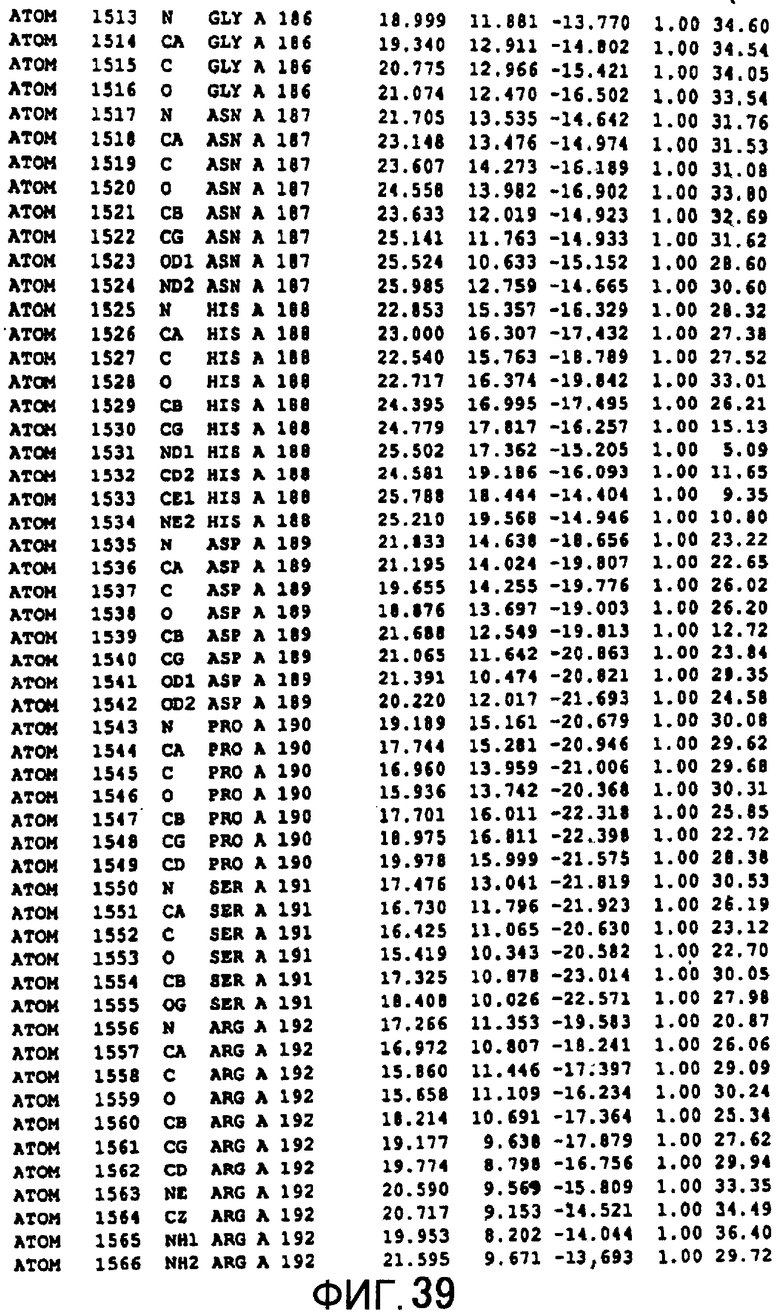
Фигура 11 отображает атомные координаты (1) MTG.
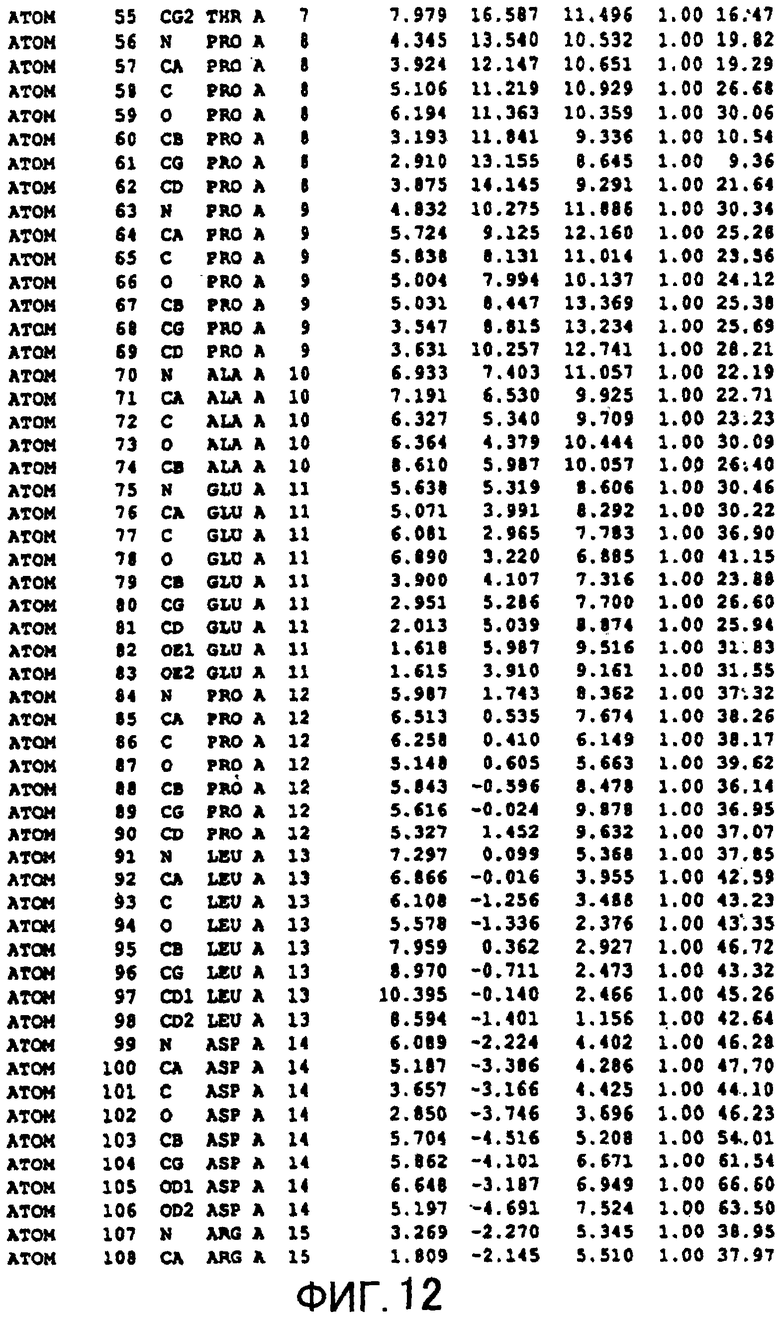
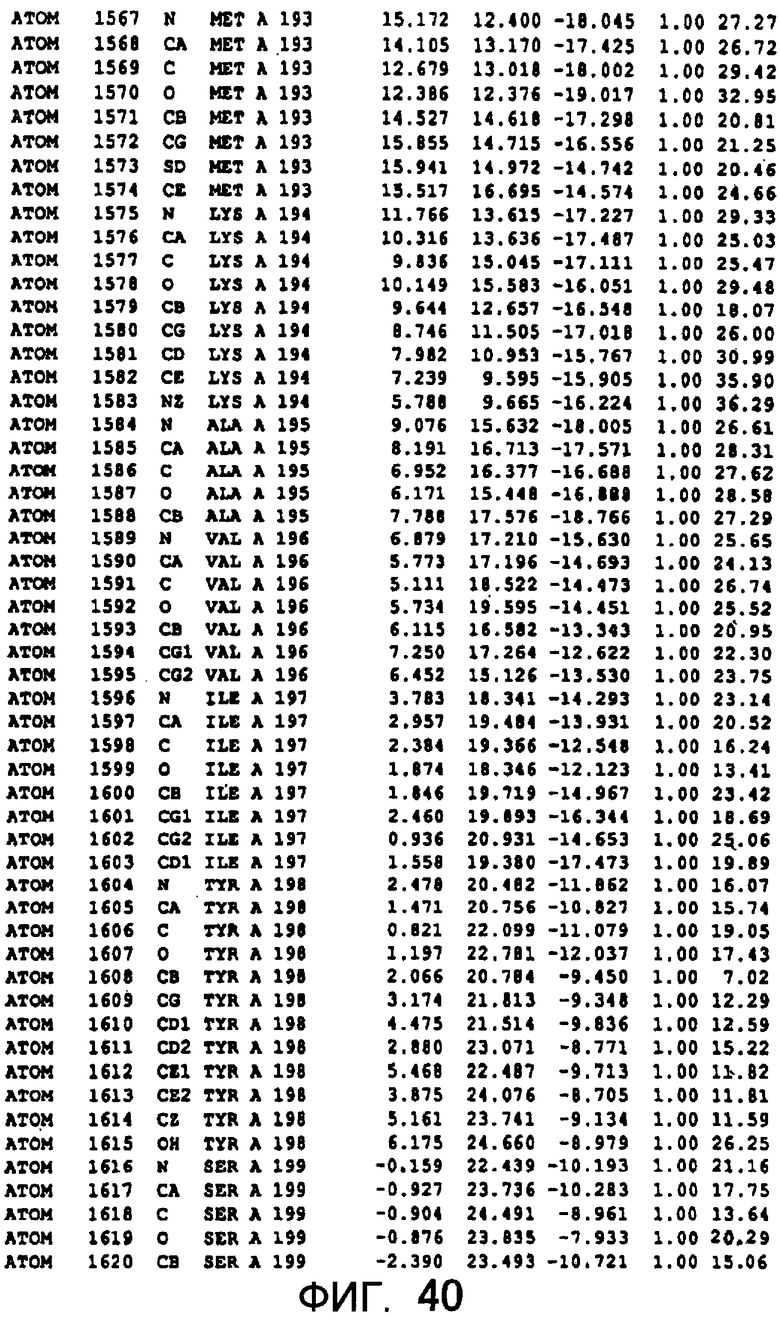
Фигура 12 отображает атомные координаты (2) MTG.
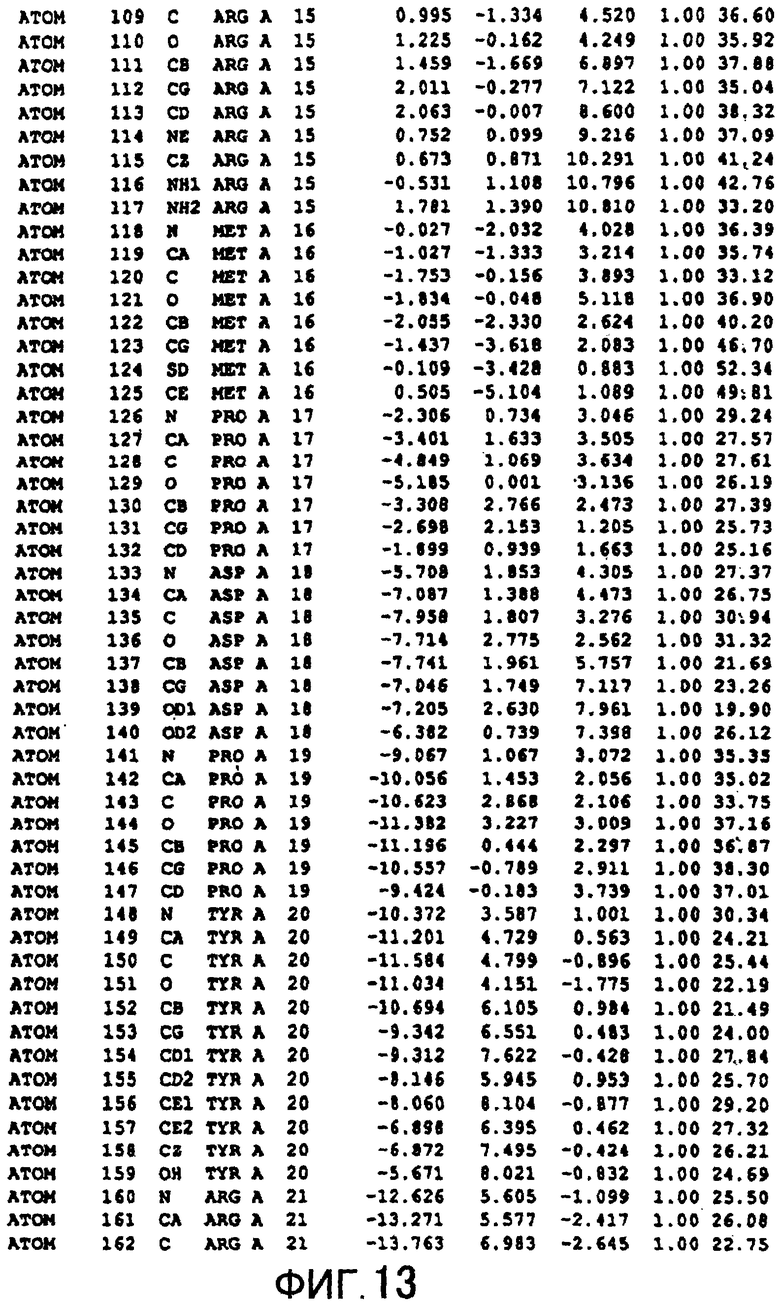
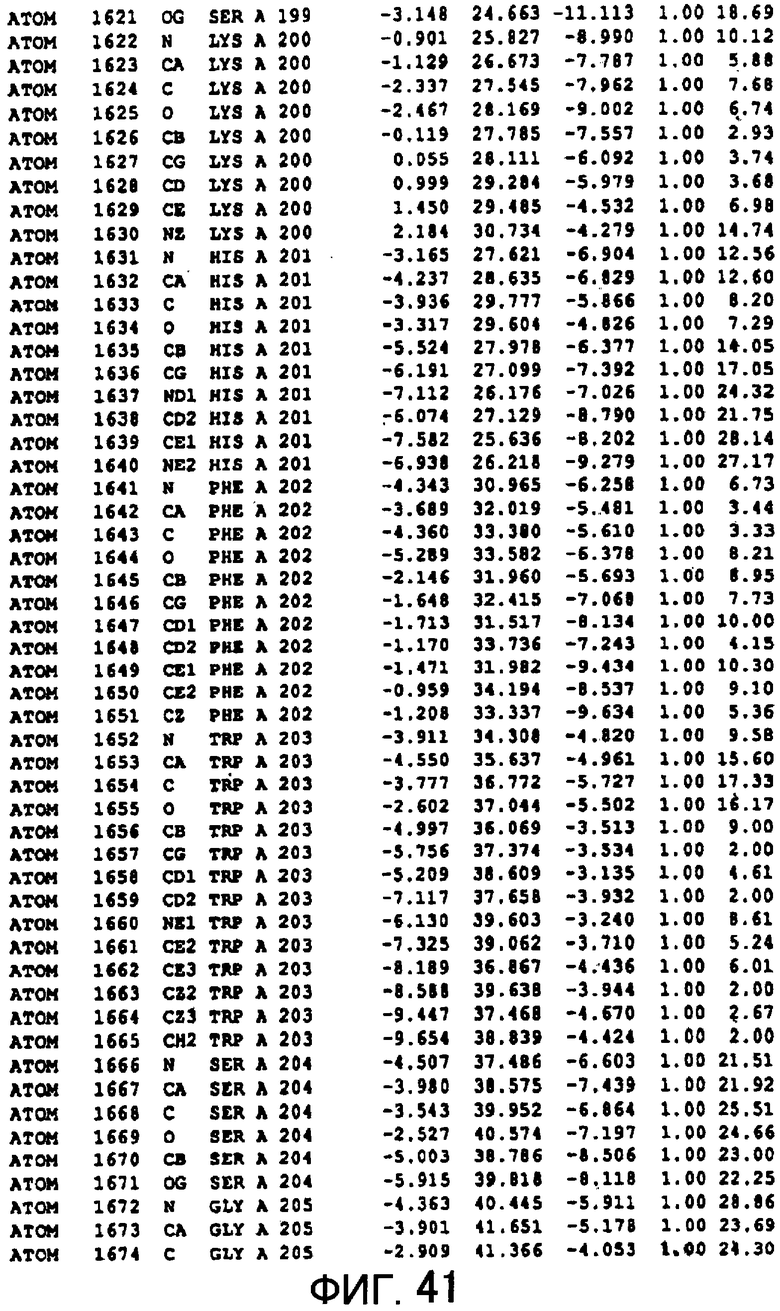
Фигура 13 отображает атомные координаты (3) MTG.
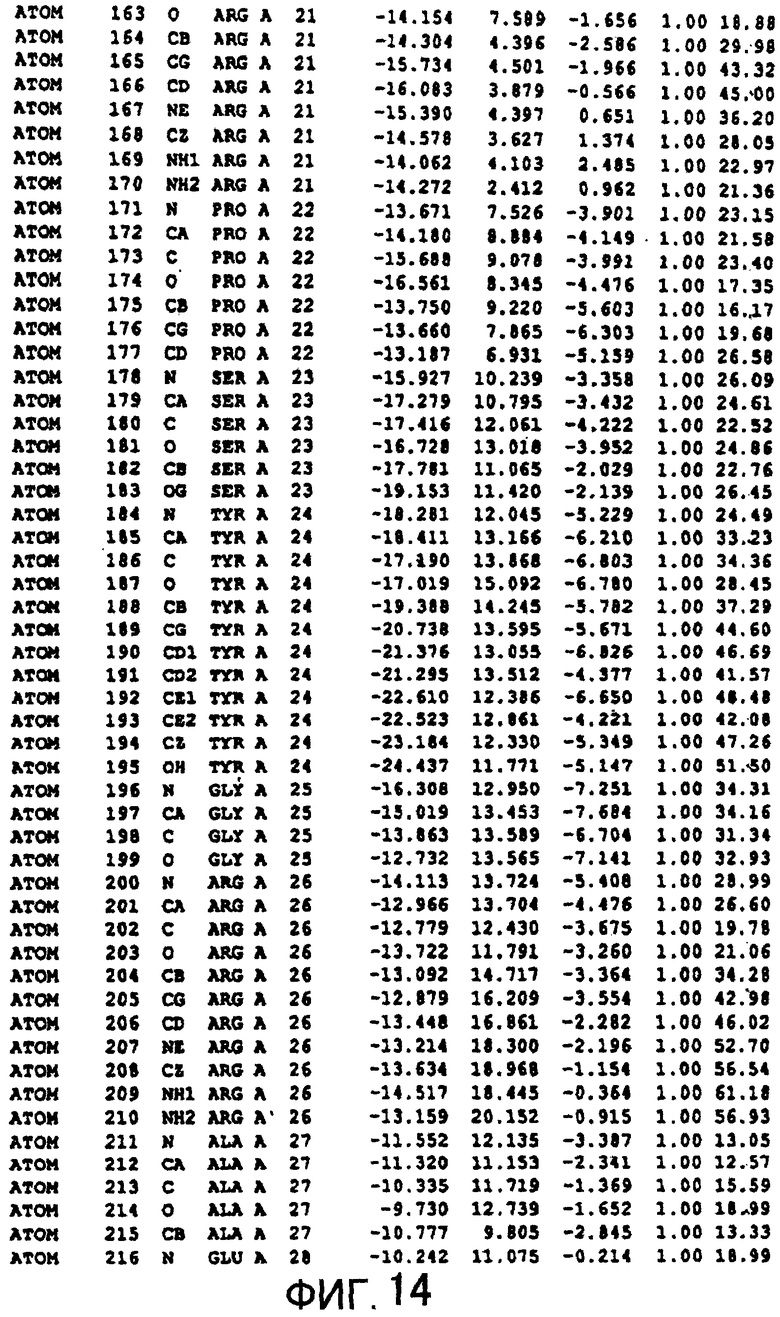
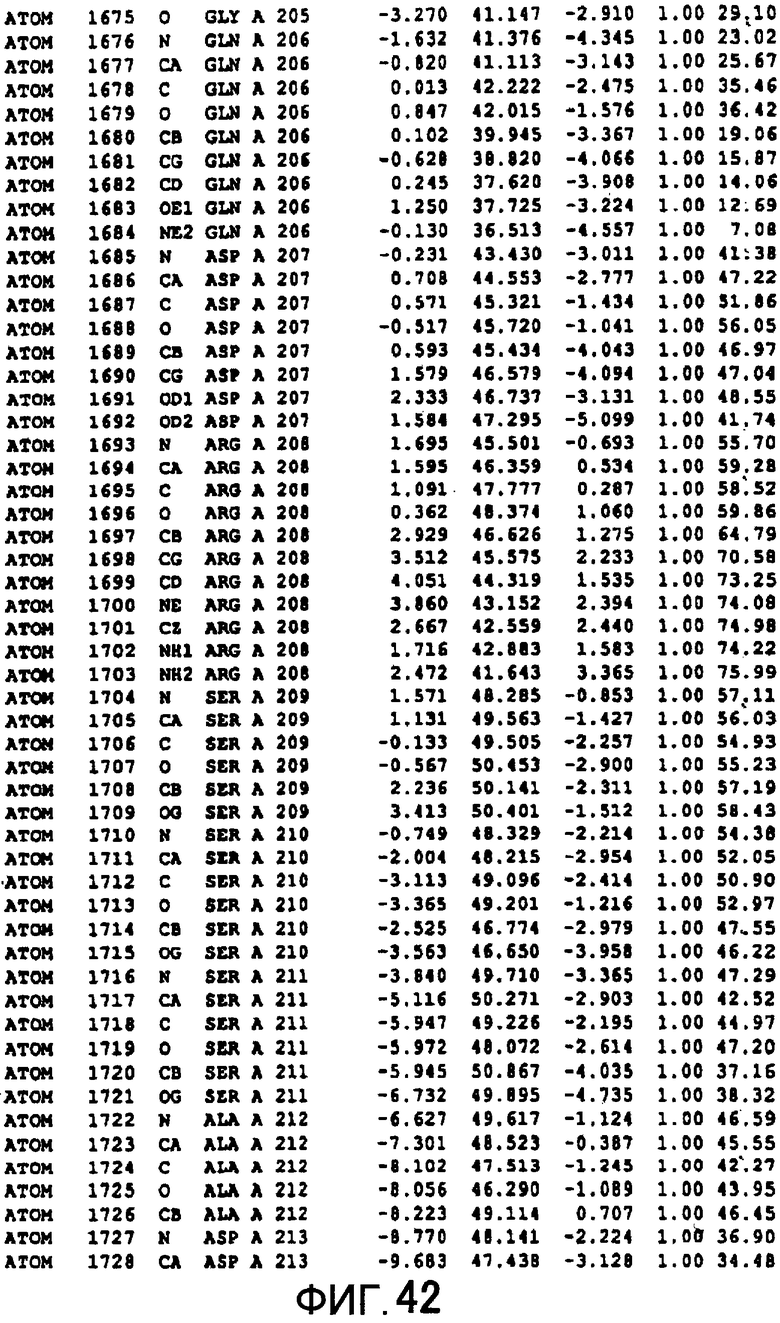
Фигура 14 отображает атомные координаты (4) MTG.
Фигура 15 отображает атомные координаты (5) MTG.
Фигура 16 отображает атомные координаты (6) MTG.
Фигура 17 отображает атомные координаты (7) MTG.
Фигура 18 отображает атомные координаты (8) MTG.
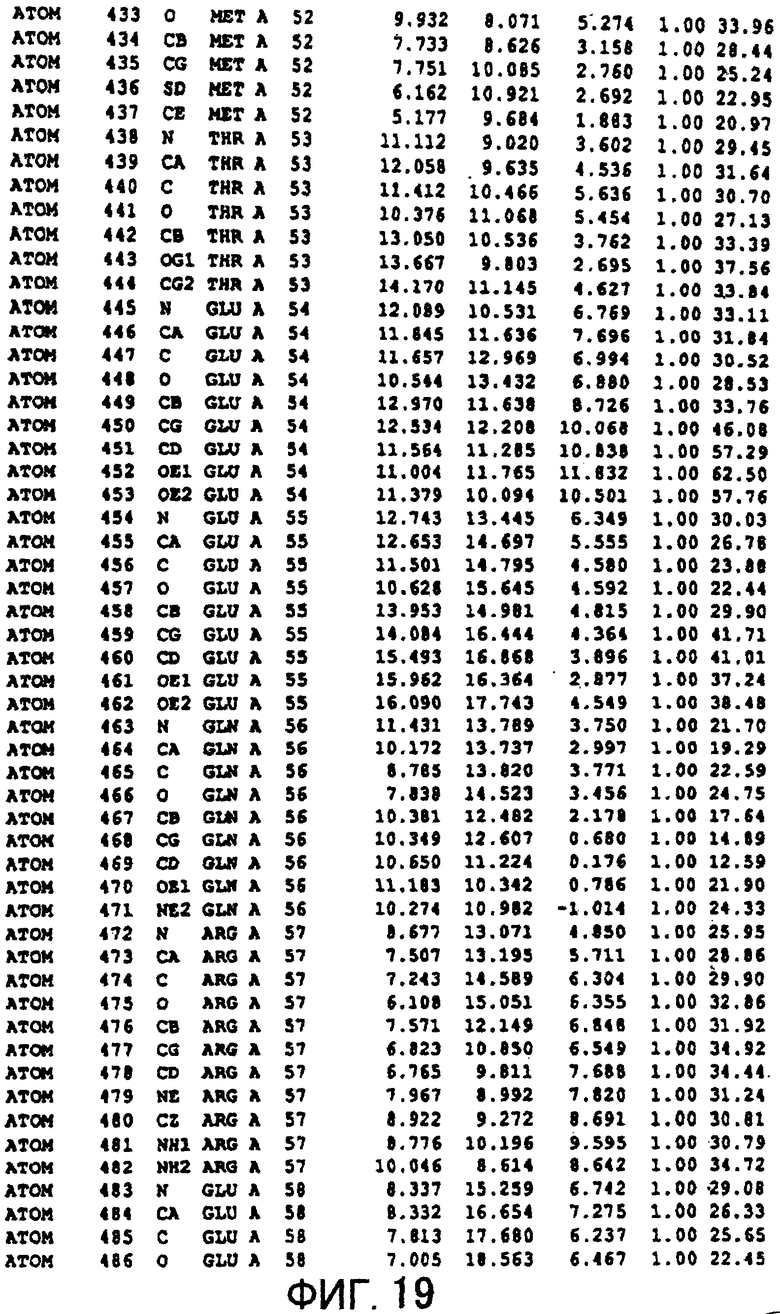
Фигура 19 отображает атомные координаты (9) MTG.
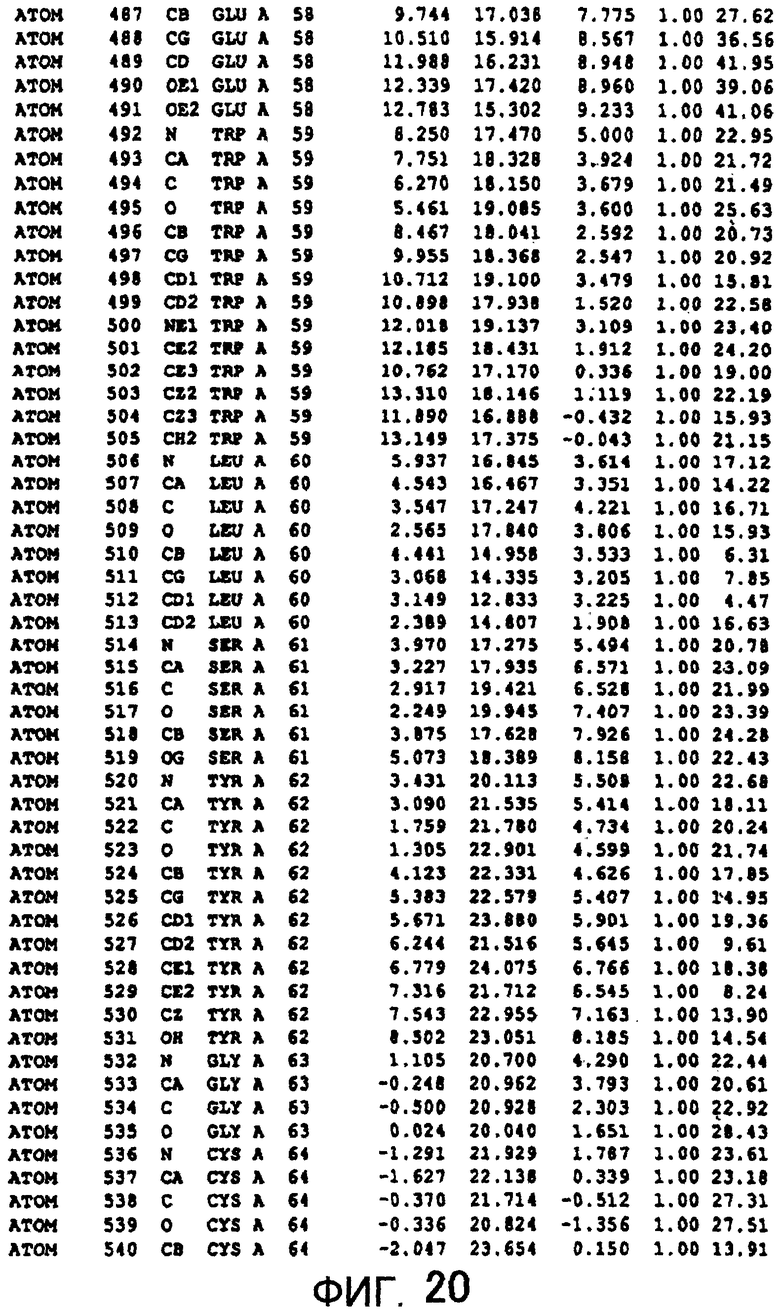
Фигура 20 отображает атомные координаты (10) MTG.
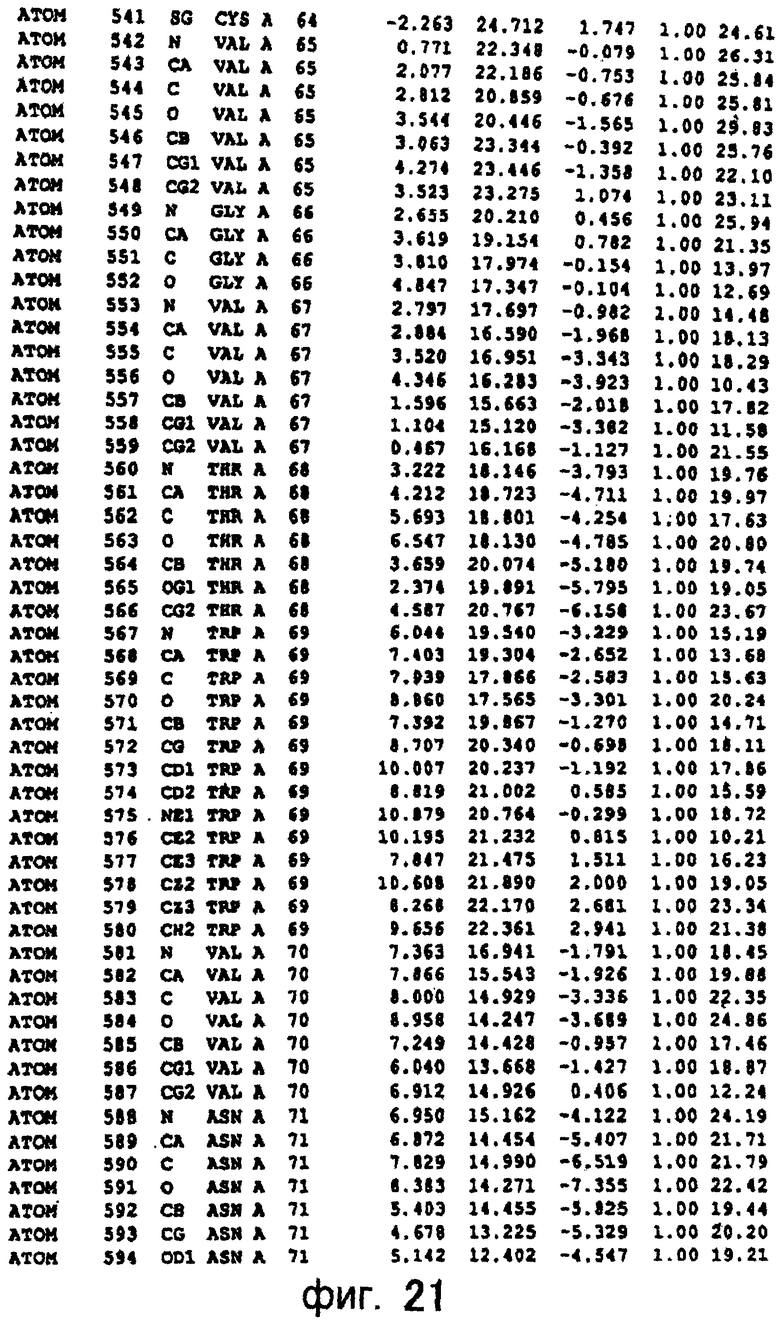
Фигура 21 отображает атомные координаты (11) MTG.
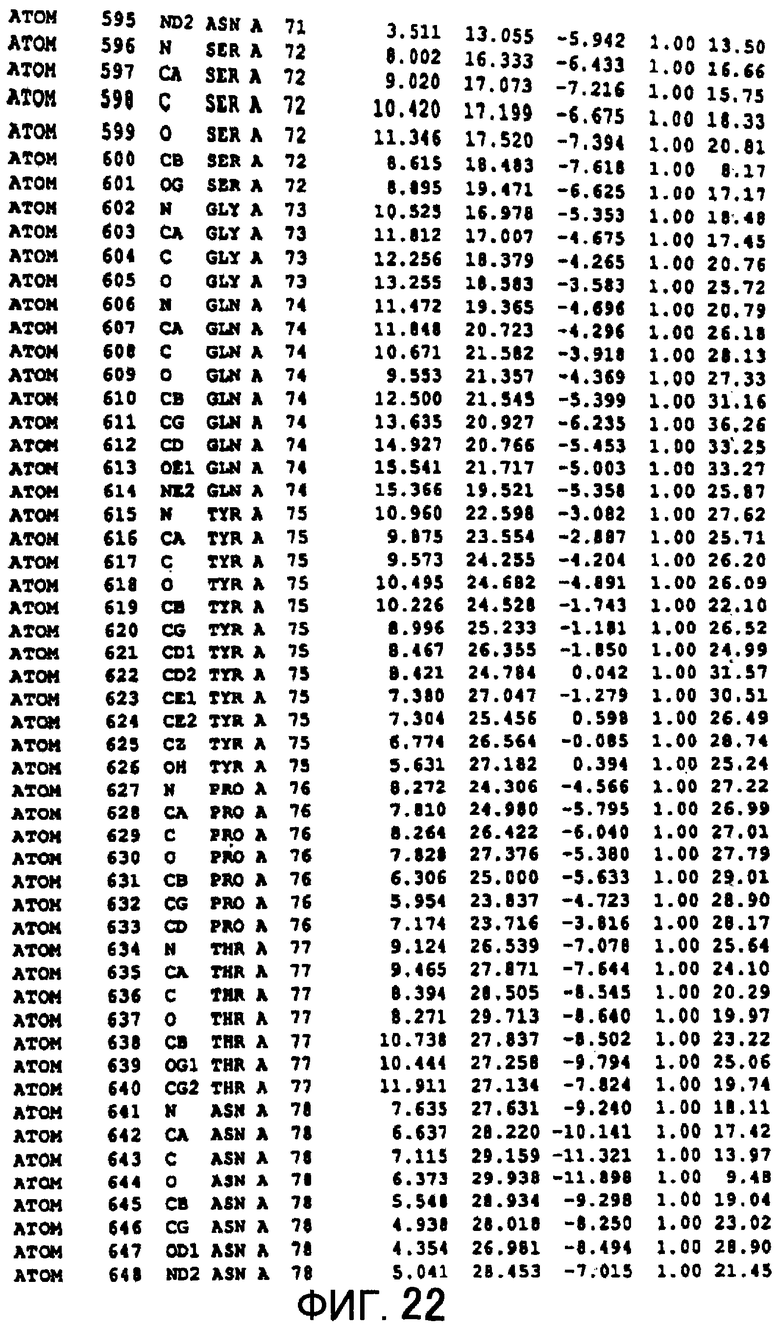
Фигура 22 отображает атомные координаты (12) MTG.
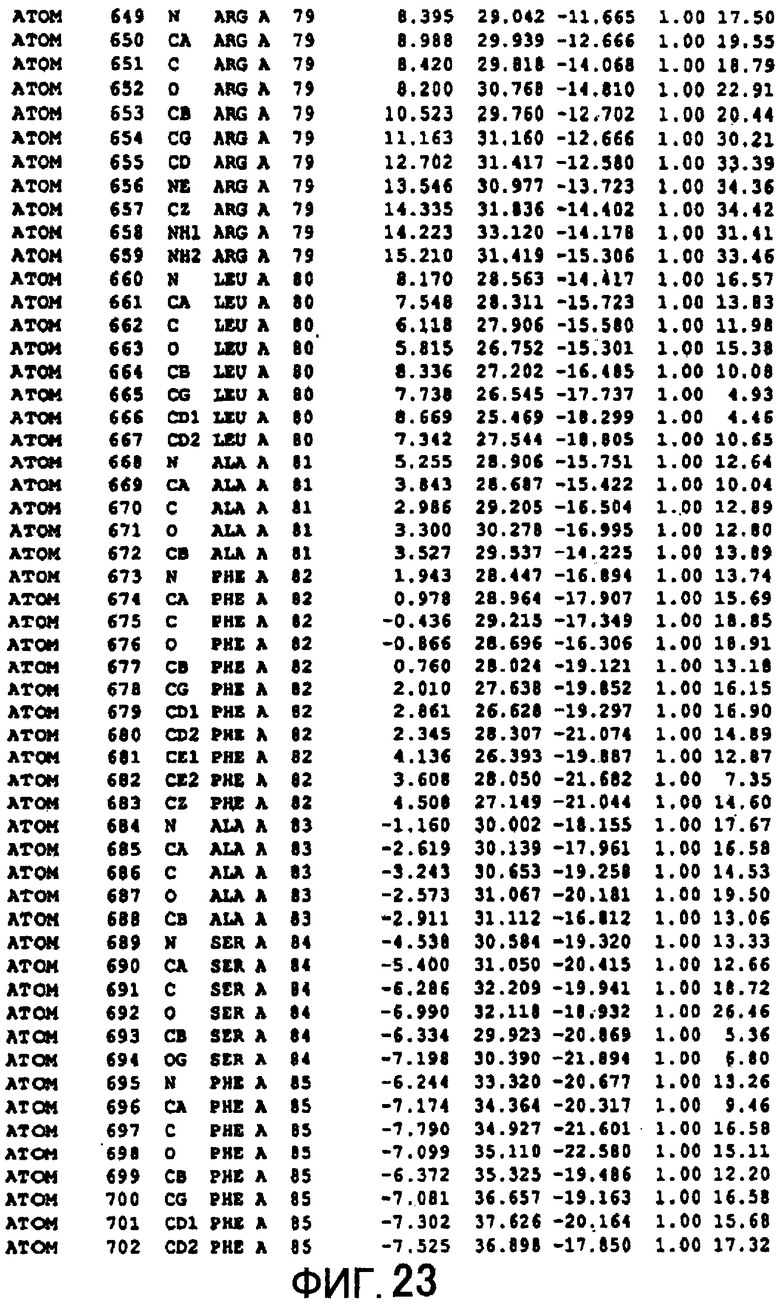
Фигура 23 отображает атомные координаты (13) MTG.
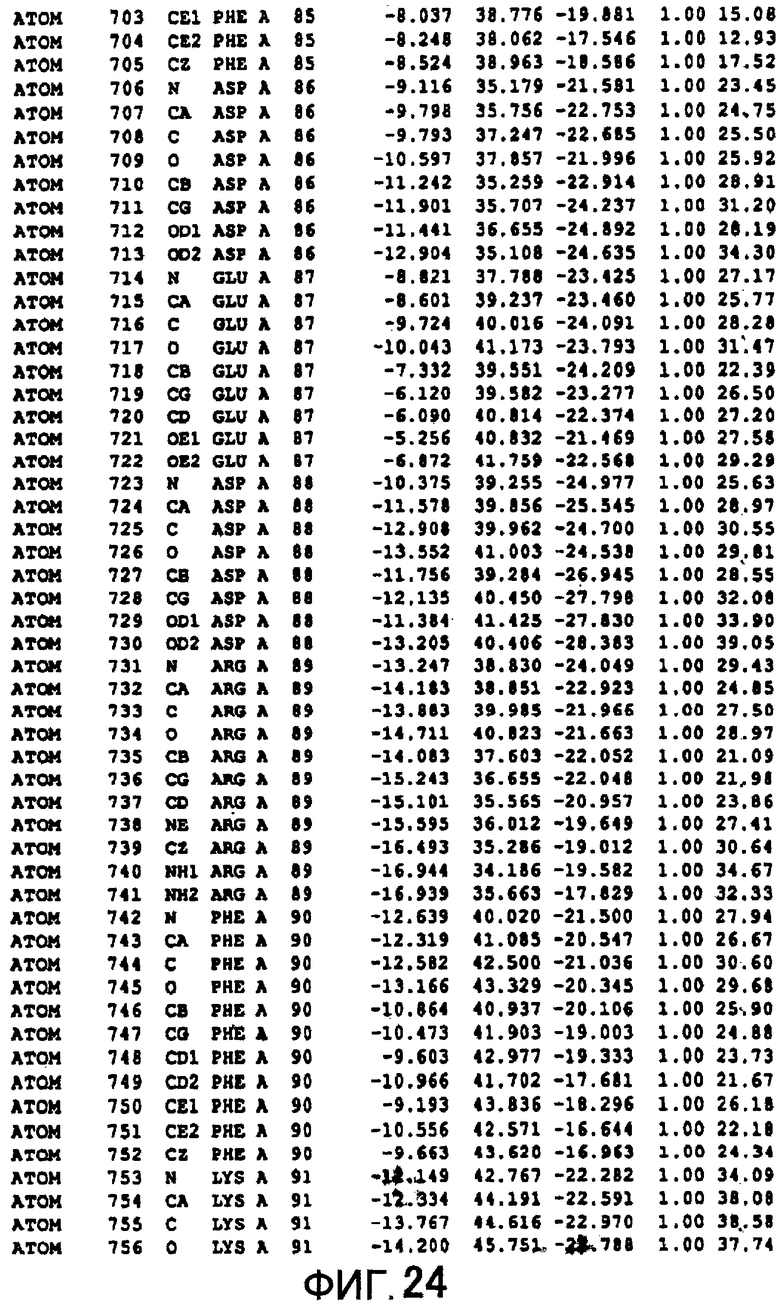
Фигура 24 отображает атомные координаты (14) MTG.
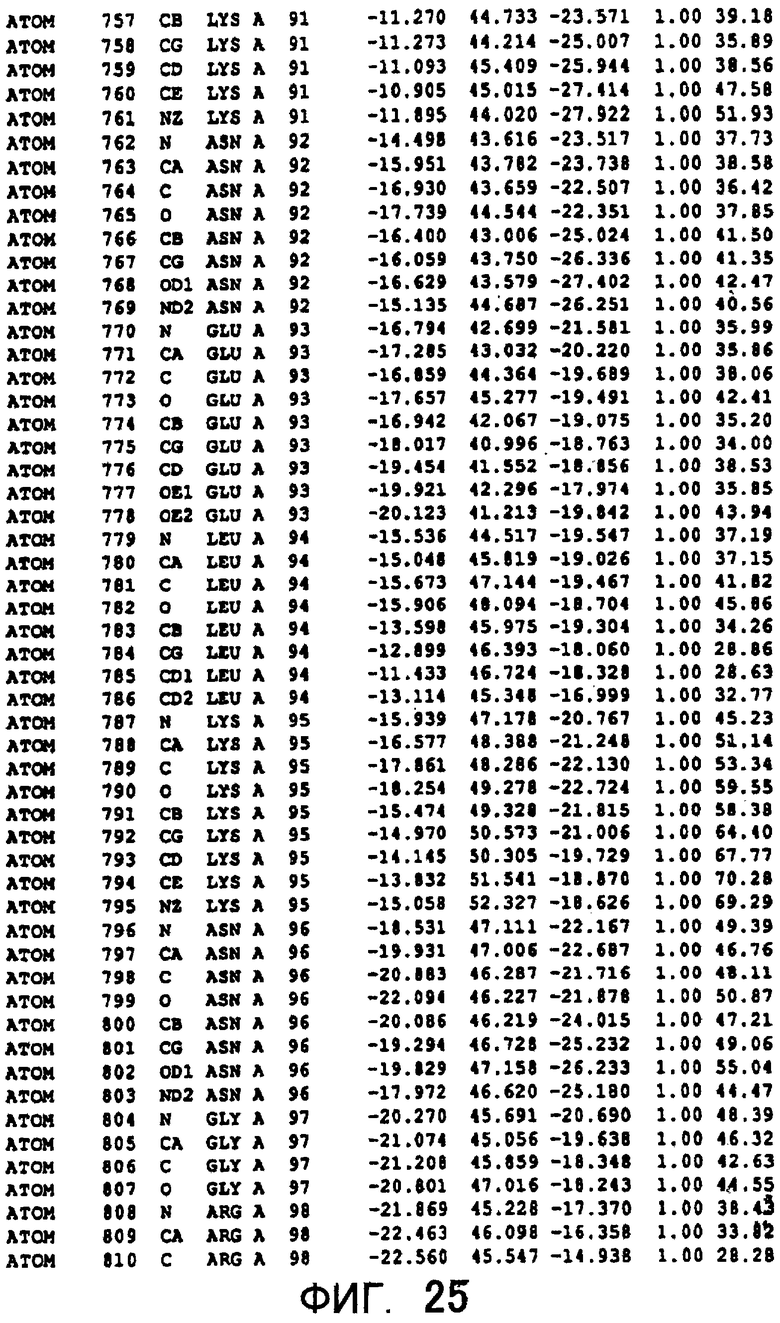
Фигура 25 отображает атомные координаты (15) MTG.
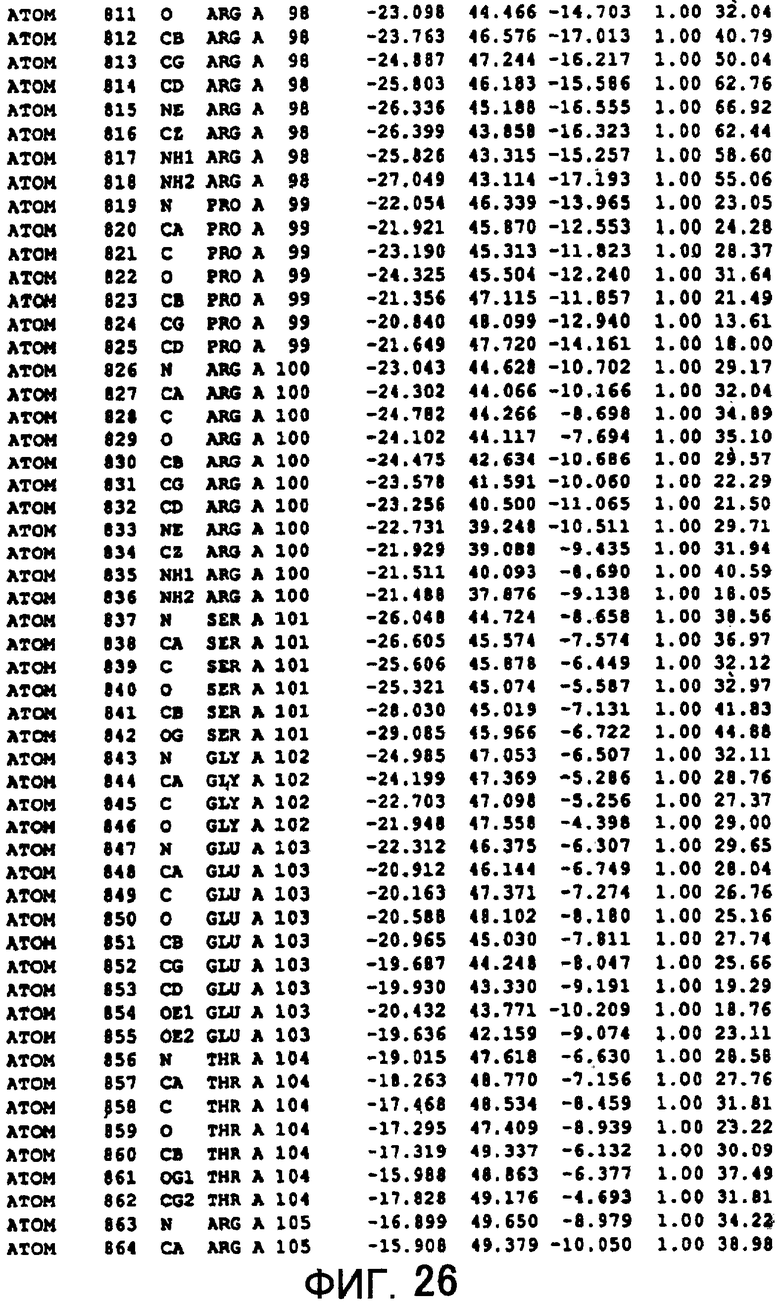
Фигура 26 отображает атомные координаты (16) MTG.
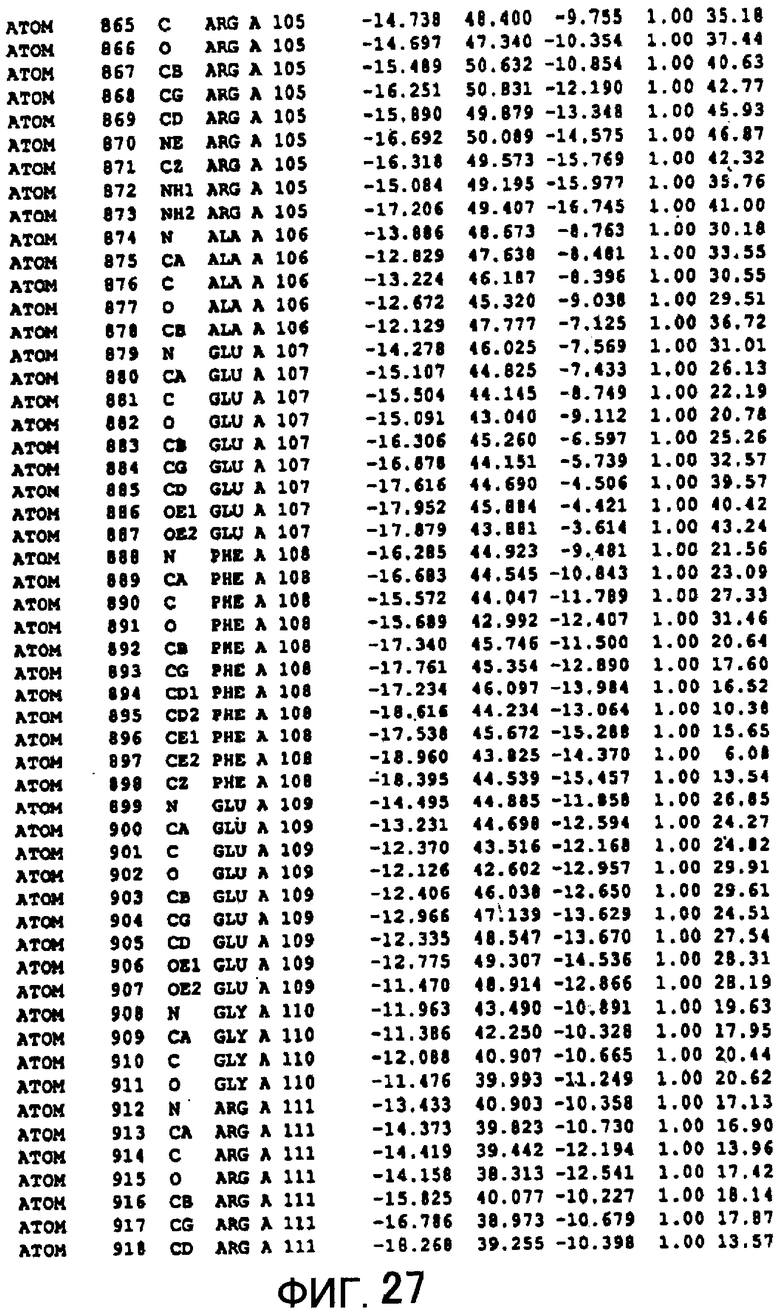
Фигура 27 отображает атомные координаты (17) MTG.
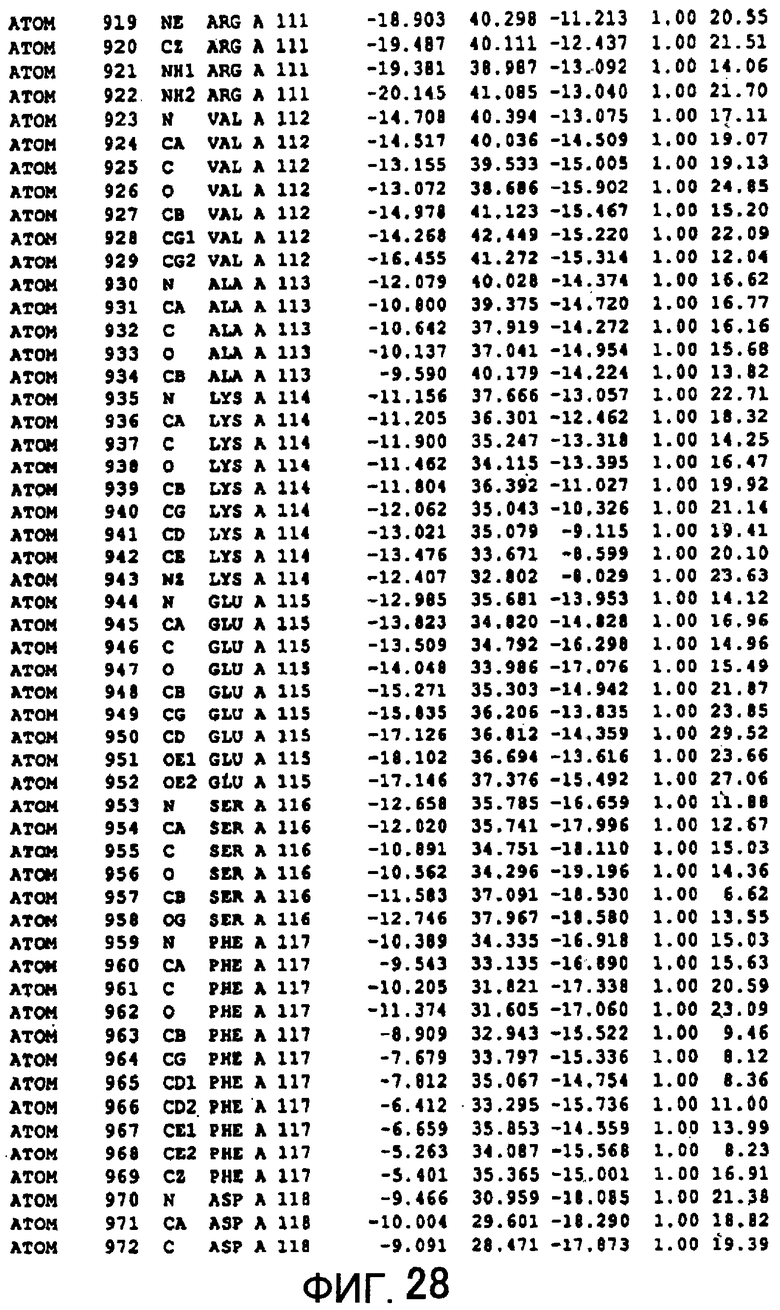
Фигура 28 отображает атомные координаты (18) MTG.
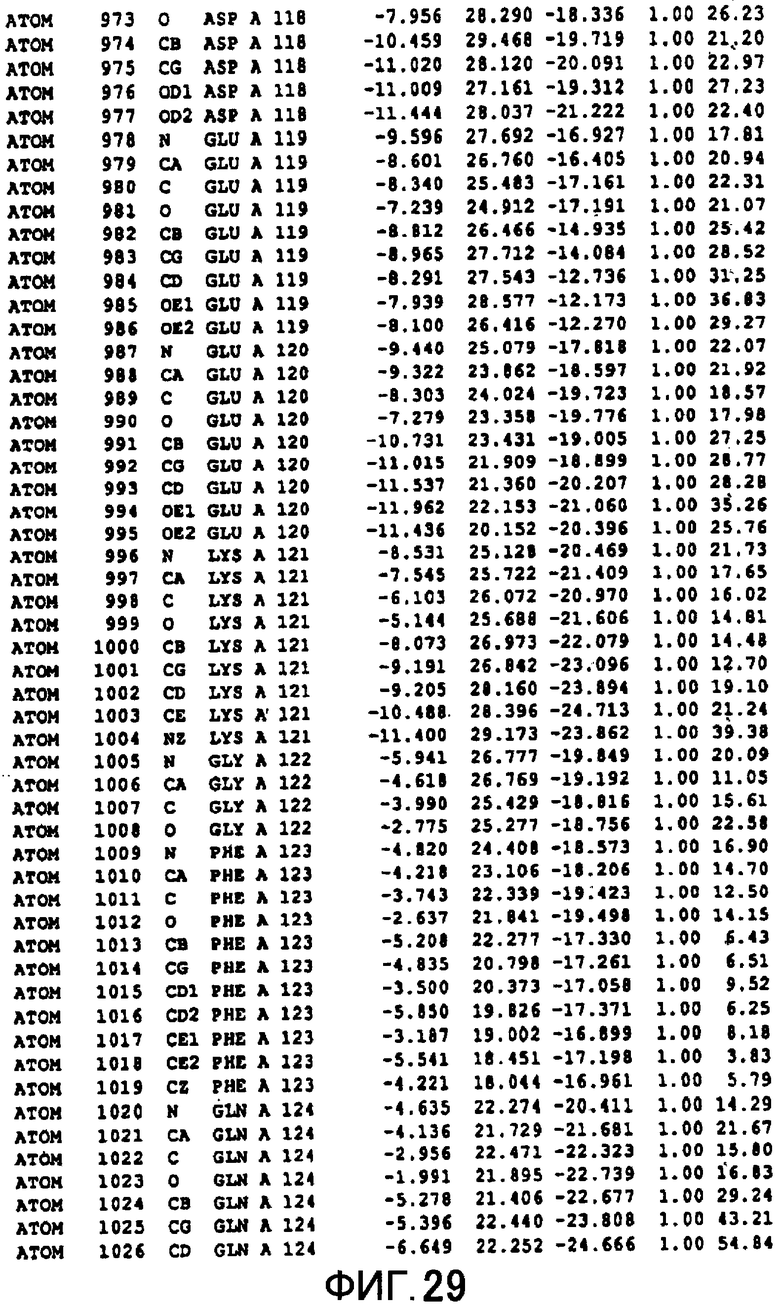
Фигура 29 отображает атомные координаты (19) MTG.
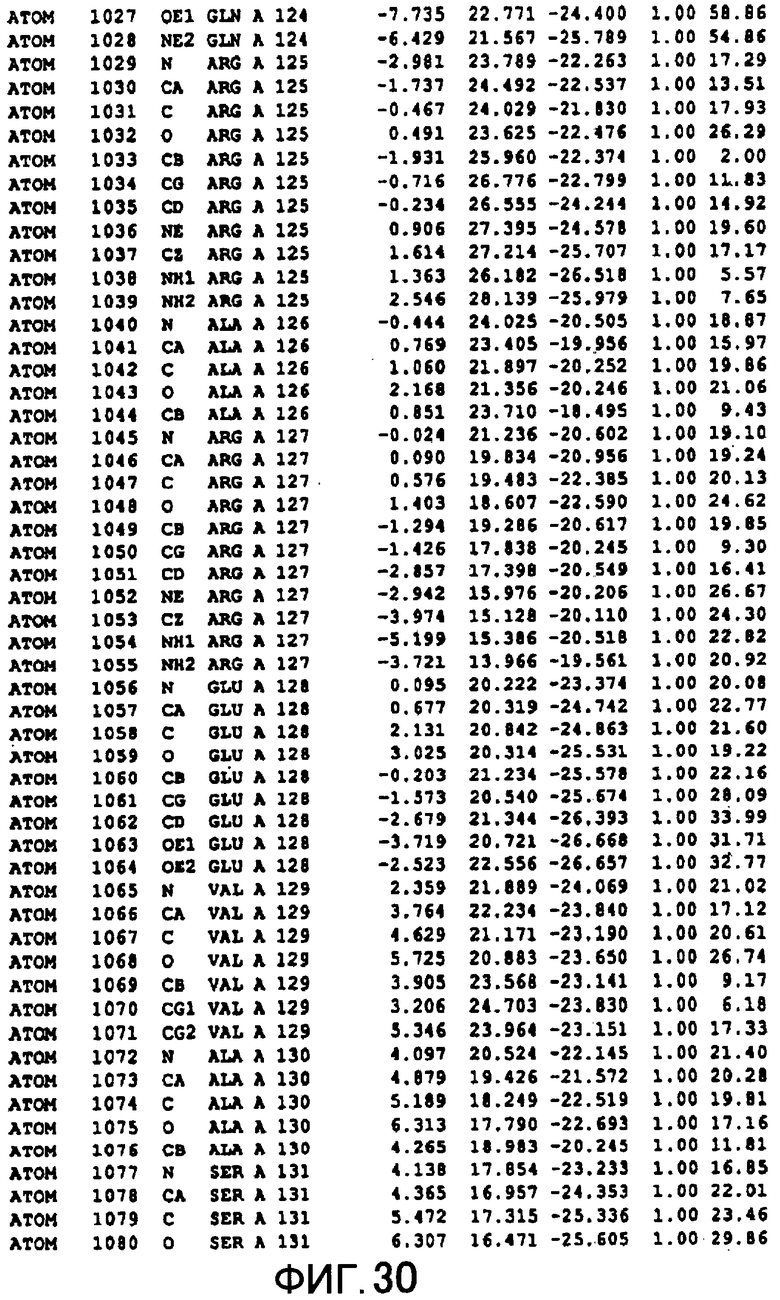
Фигура 30 отображает атомные координаты (20) MTG.
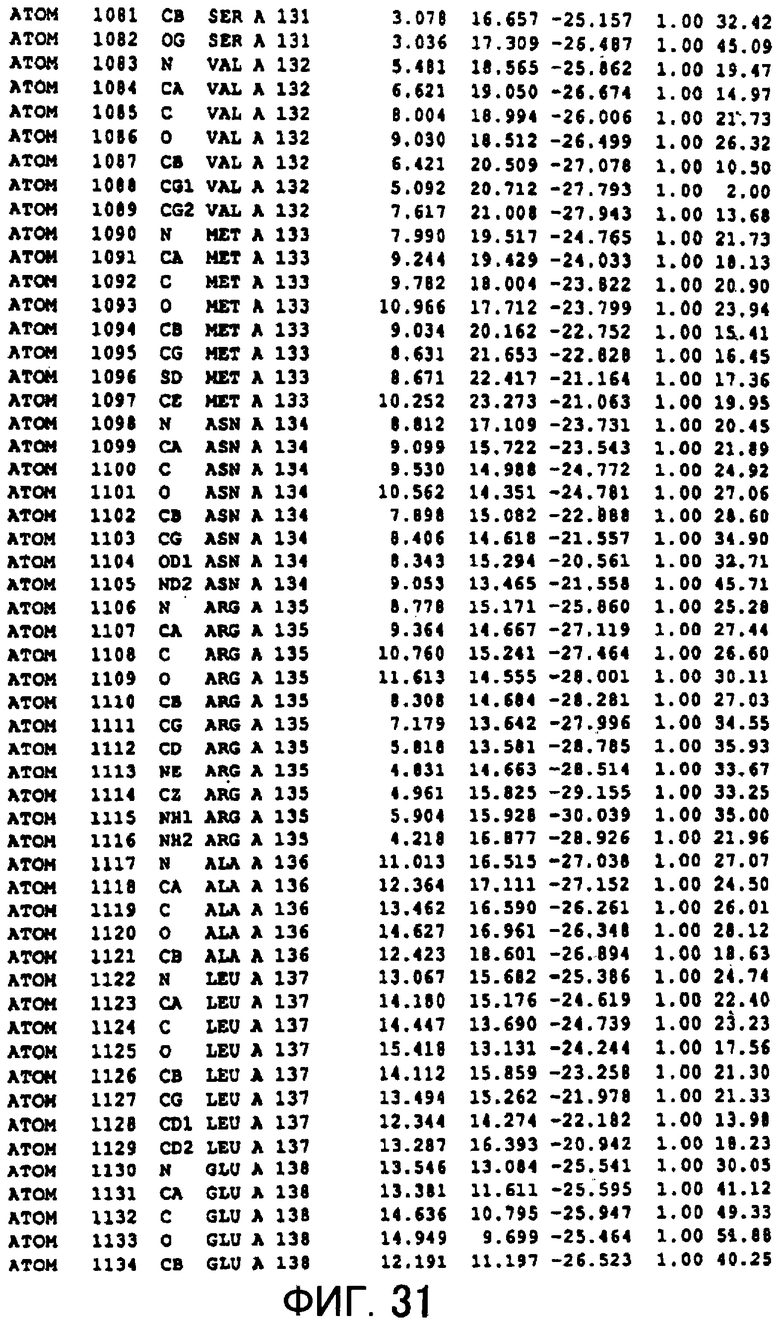
Фигура 31 отображает атомные координаты (21) MTG.
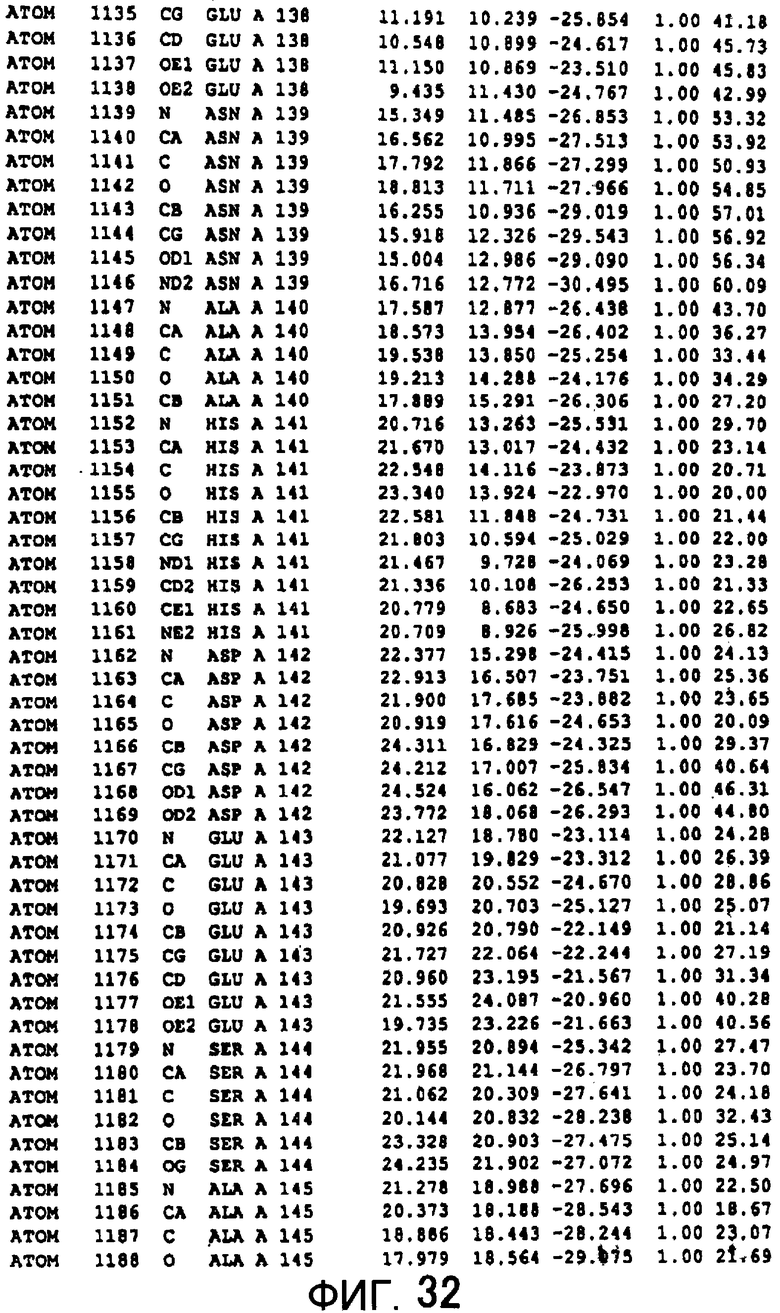
Фигура 32 отображает атомные координаты (22) MTG.
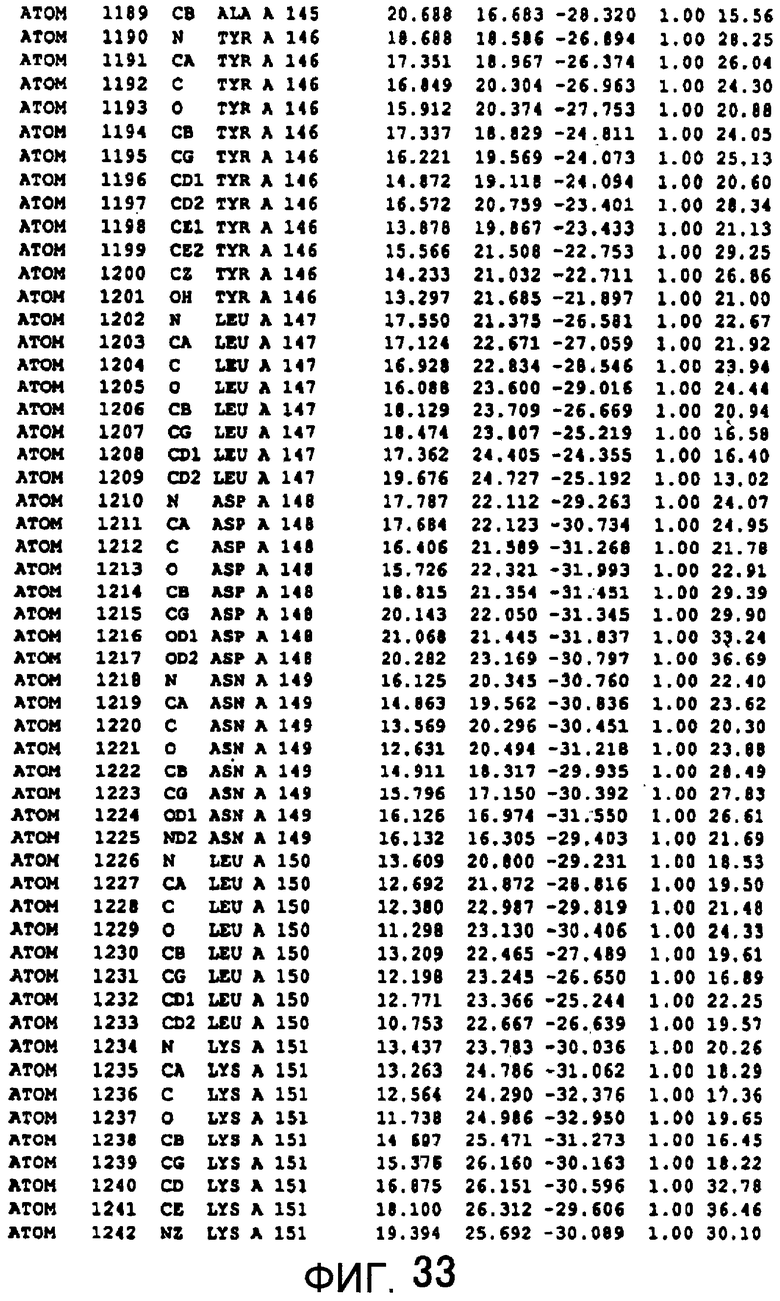
Фигура 33 отображает атомные координаты (23) MTG.
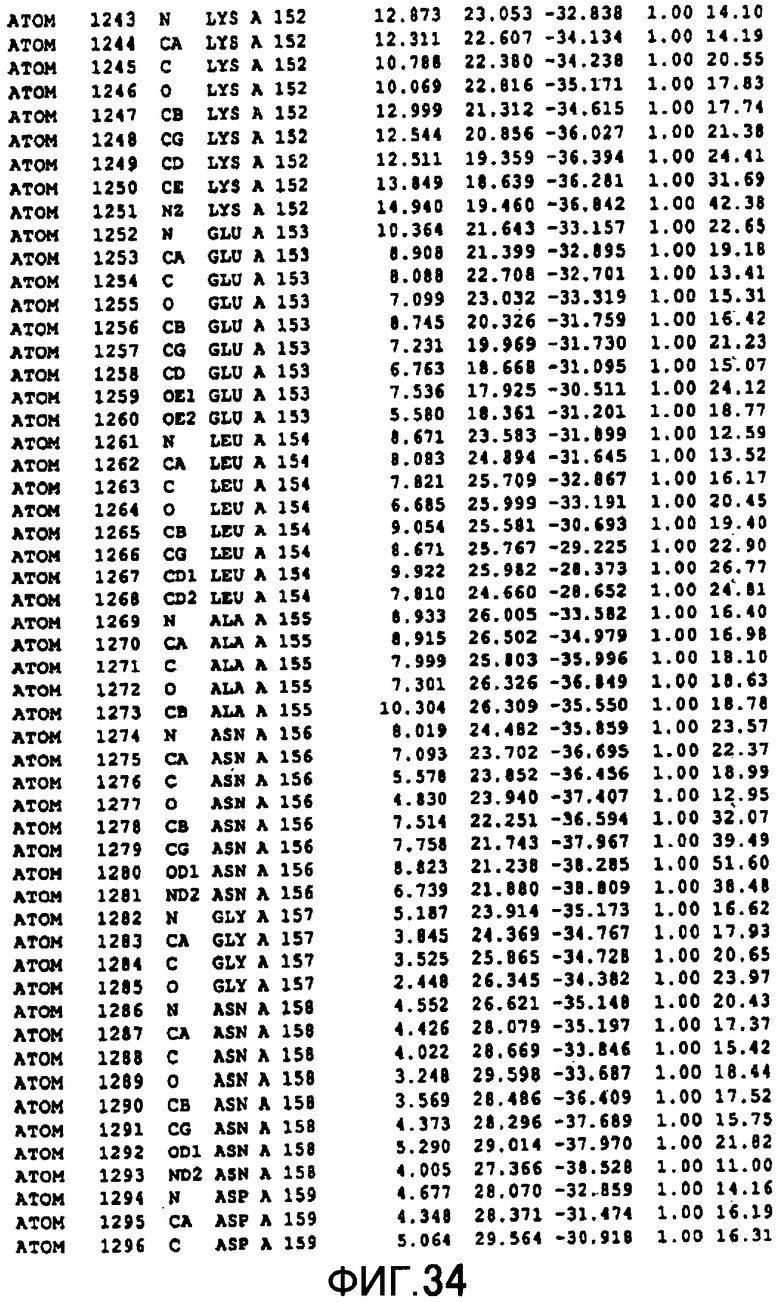
Фигура 34 отображает атомные координаты (24) MTG.
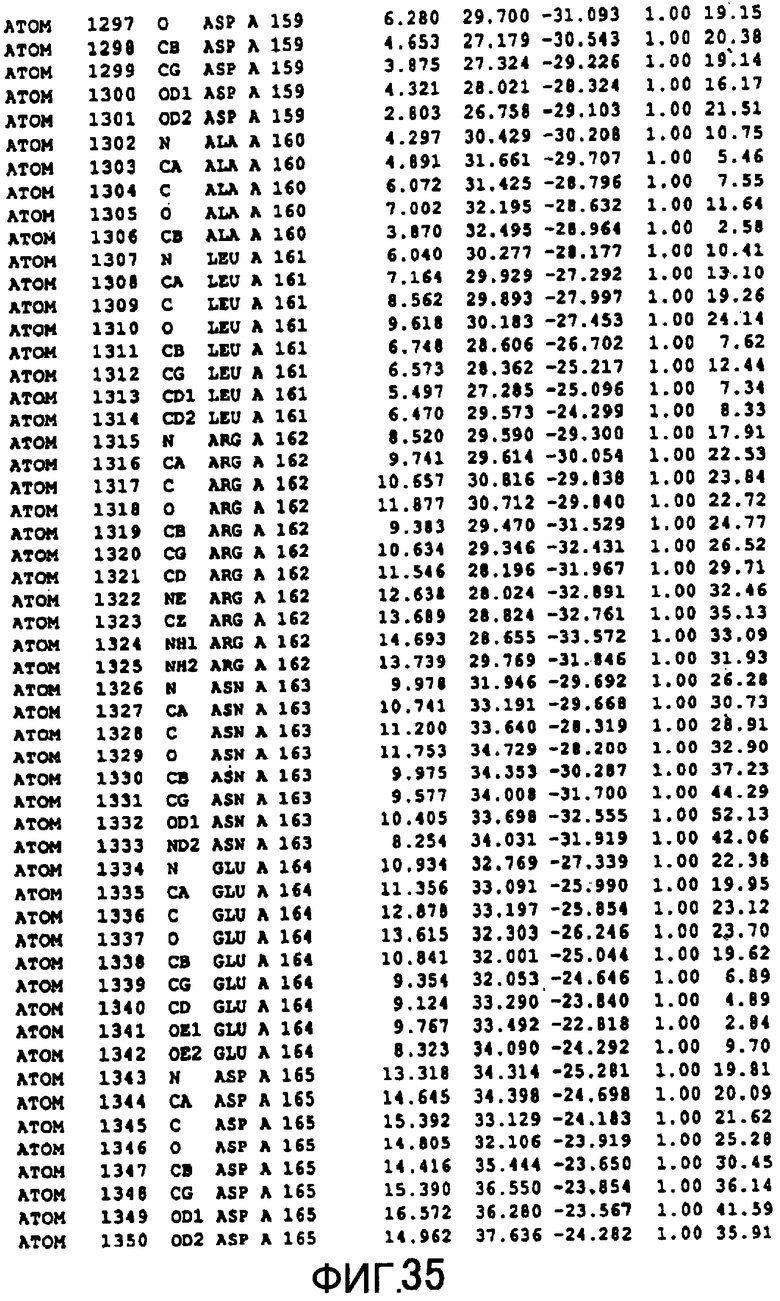
Фигура 35 отображает атомные координаты (25) MTG.
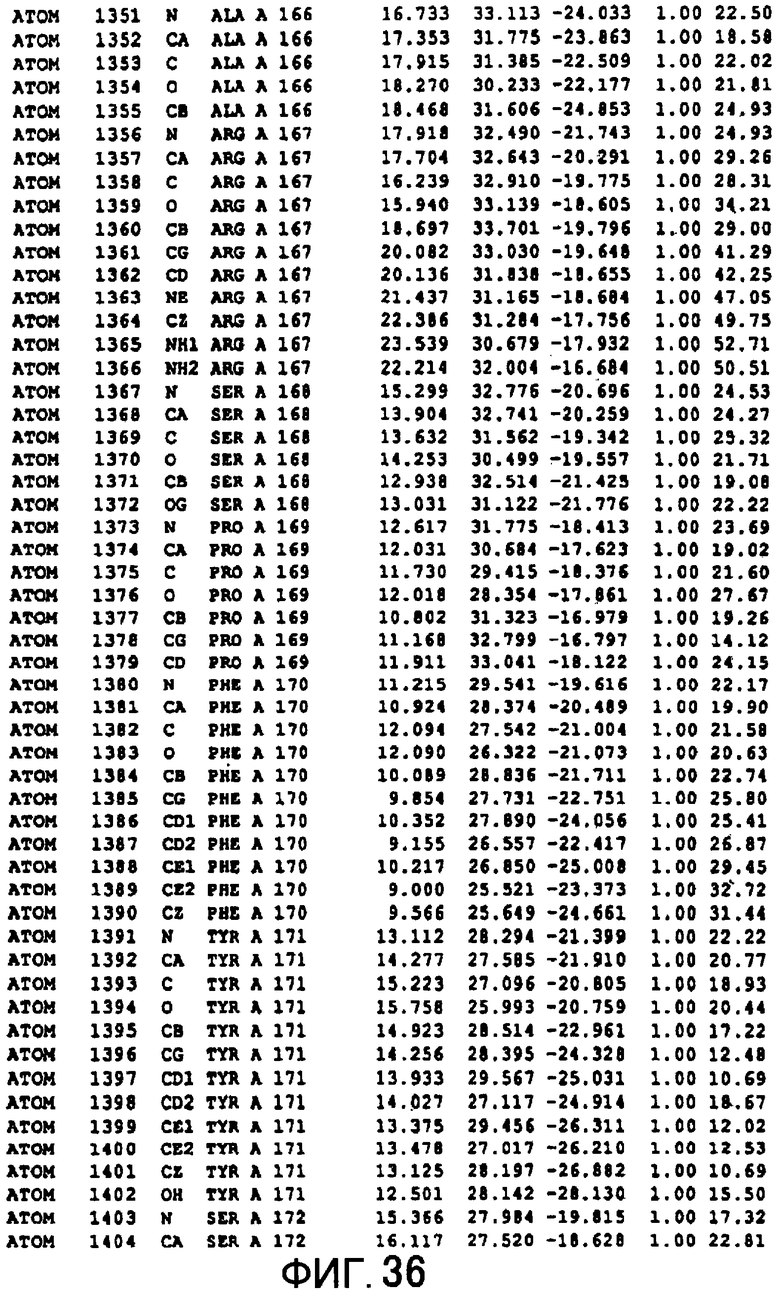
Фигура 36 отображает атомные координаты (26) MTG.
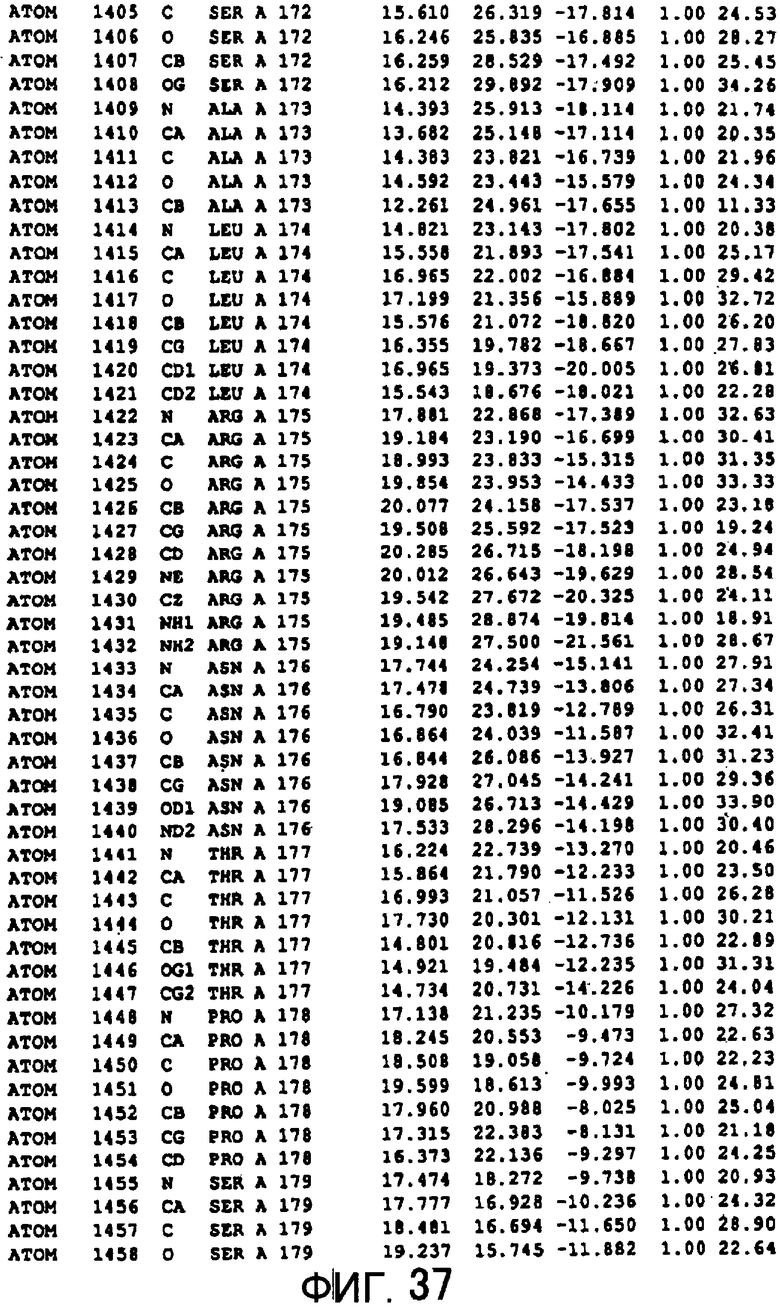
Фигура 37 отображает атомные координаты (27) MTG.
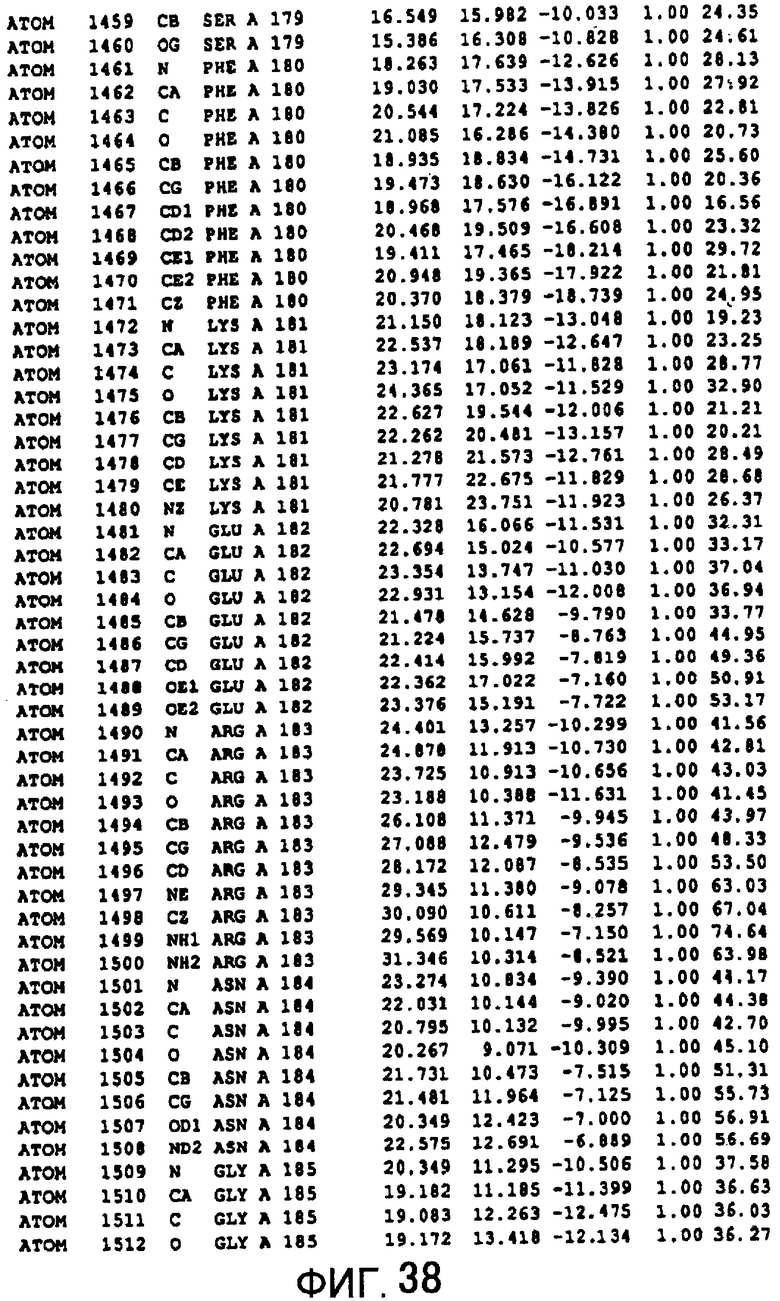
Фигура 38 отображает атомные координаты (28) MTG.
Фигура 39 отображает атомные координаты (29) MTG.
Фигура 40 отображает атомные координаты (30) MTG.
Фигура 41 отображает атомные координаты (31) MTG.
Фигура 42 отображает атомные координаты (32) MTG.
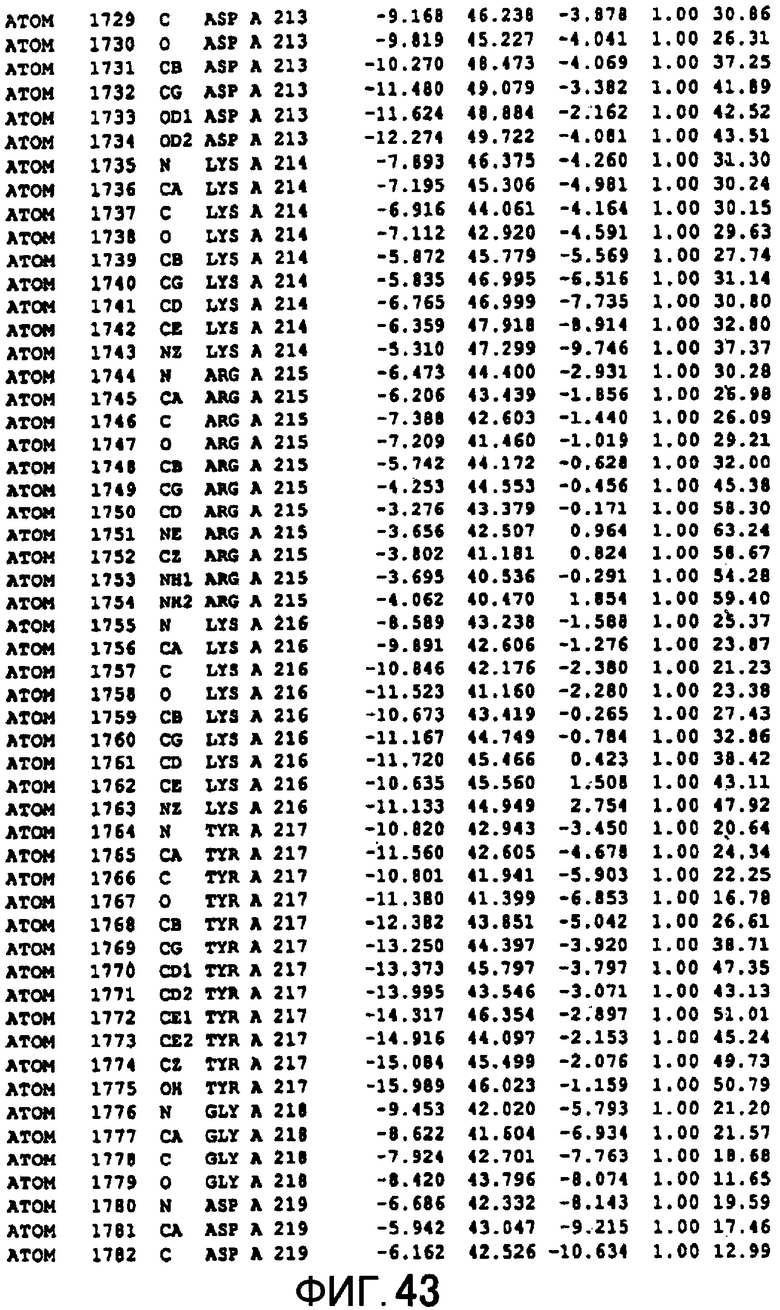
Фигура 43 отображает атомные координаты (33) MTG.
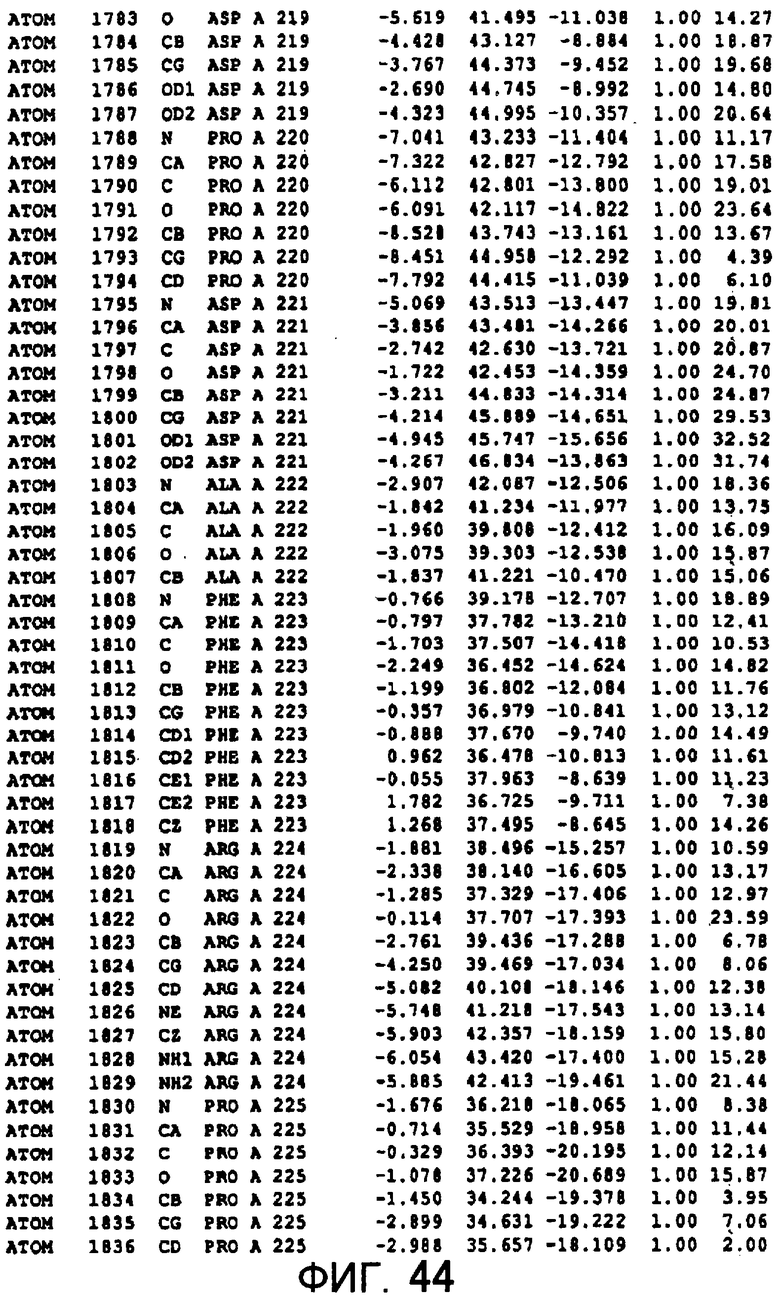
Фигура 44 отображает атомные координаты (34) MTG.
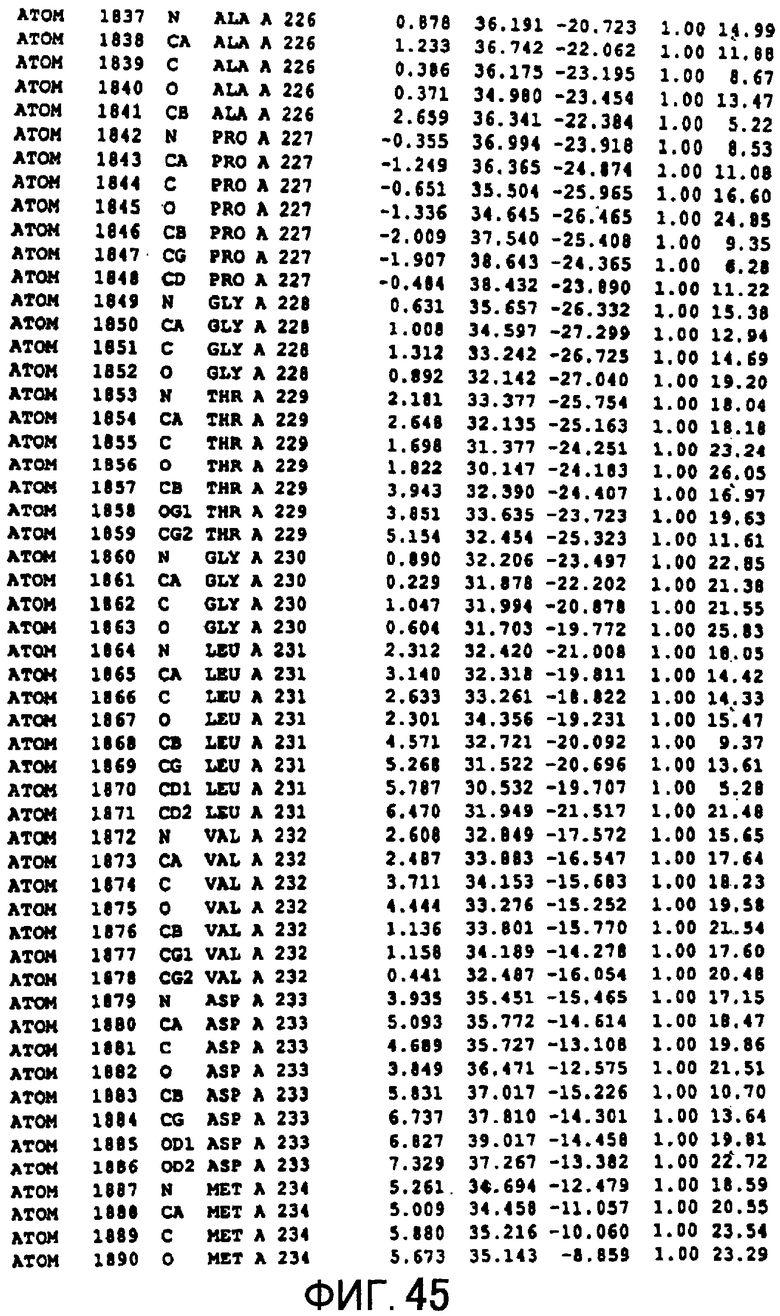
Фигура 45 отображает атомные координаты (35) MTG.
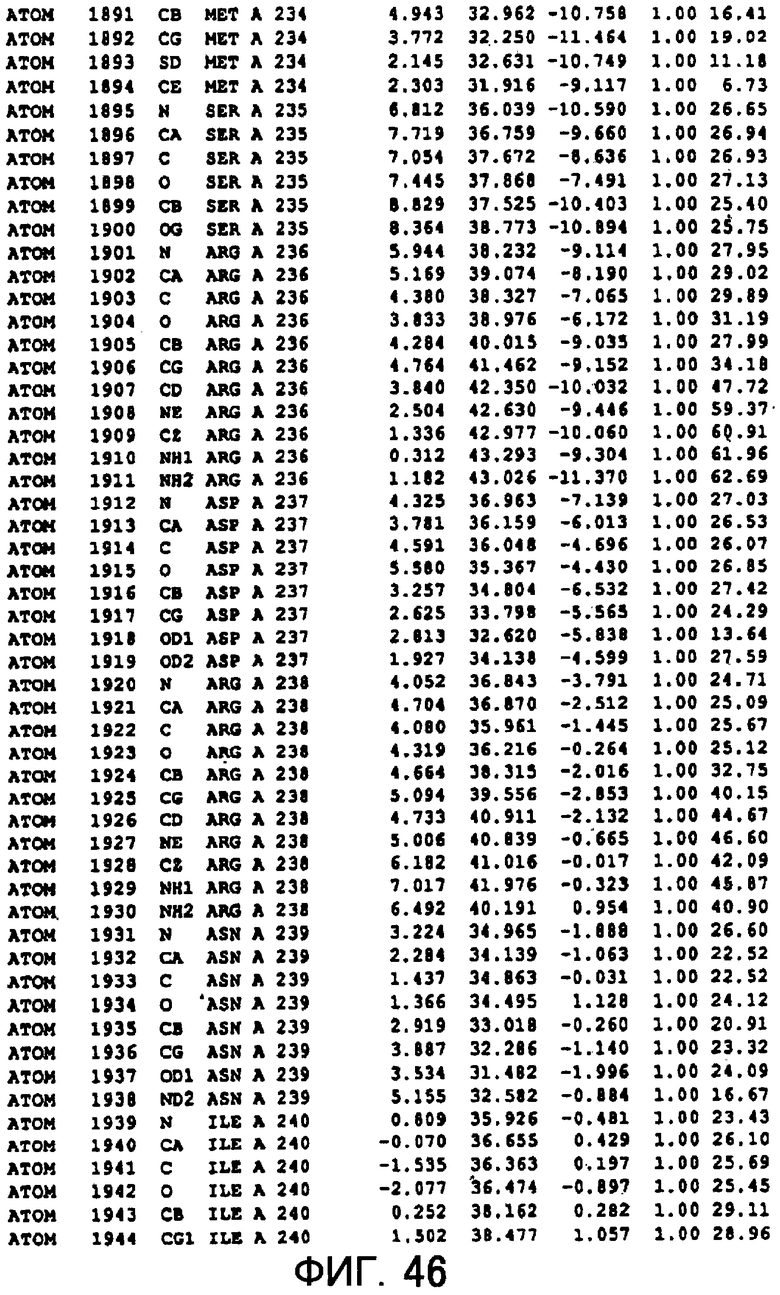
Фигура 46 отображает атомные координаты (36) MTG.
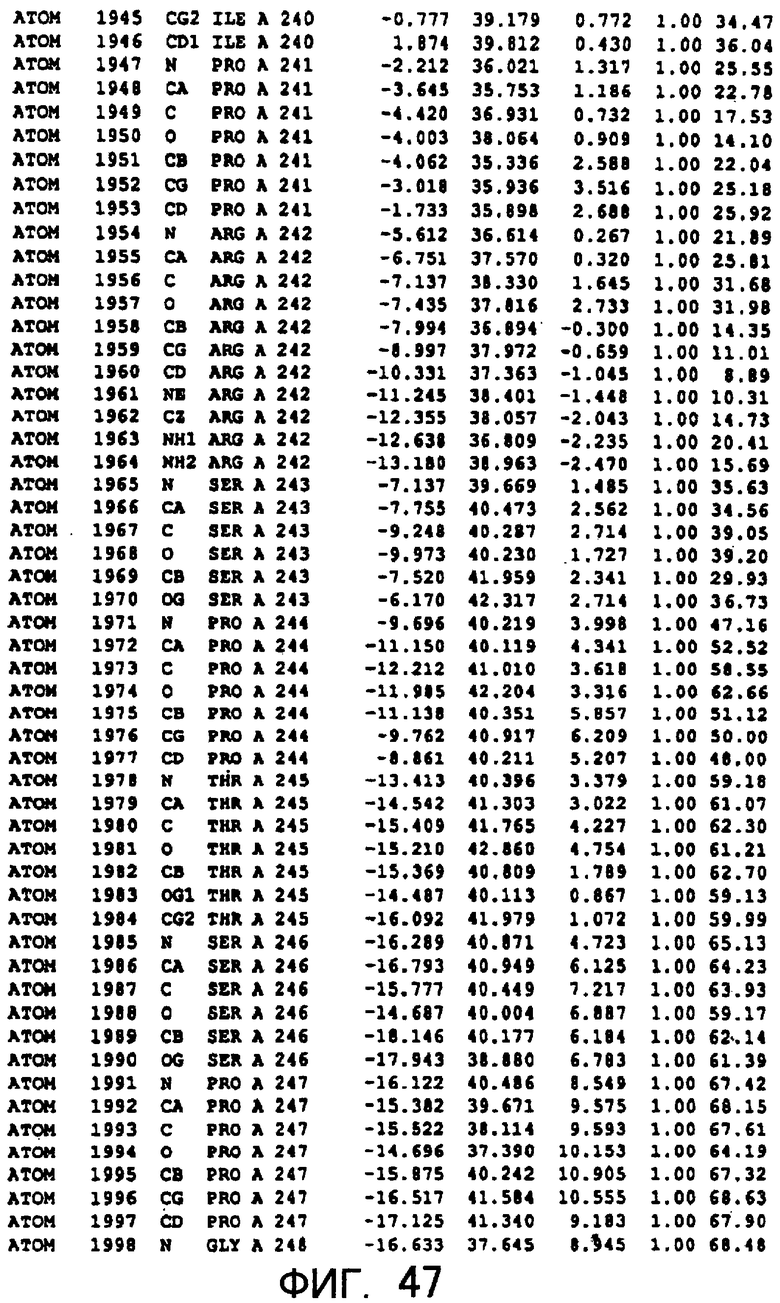
Фигура 47 отображает атомные координаты (37) MTG.
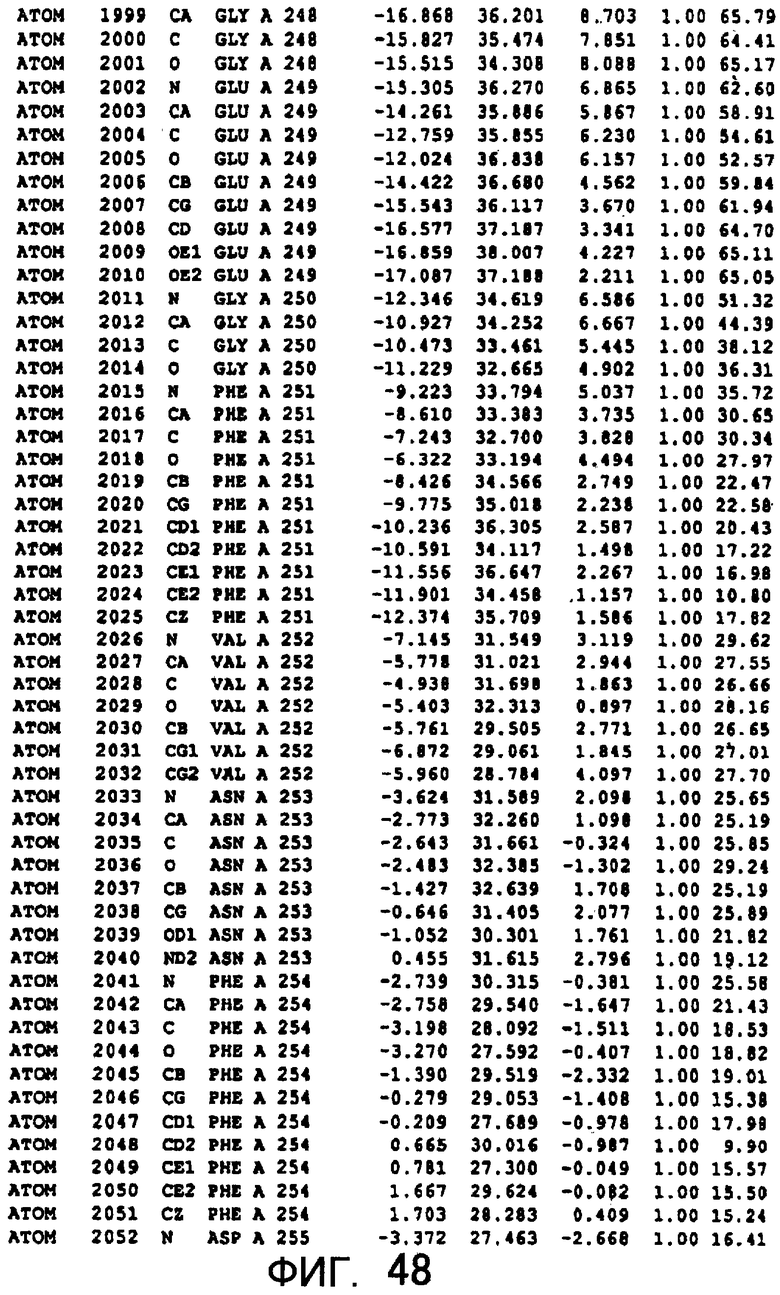
Фигура 48 отображает атомные координаты (38) MTG.
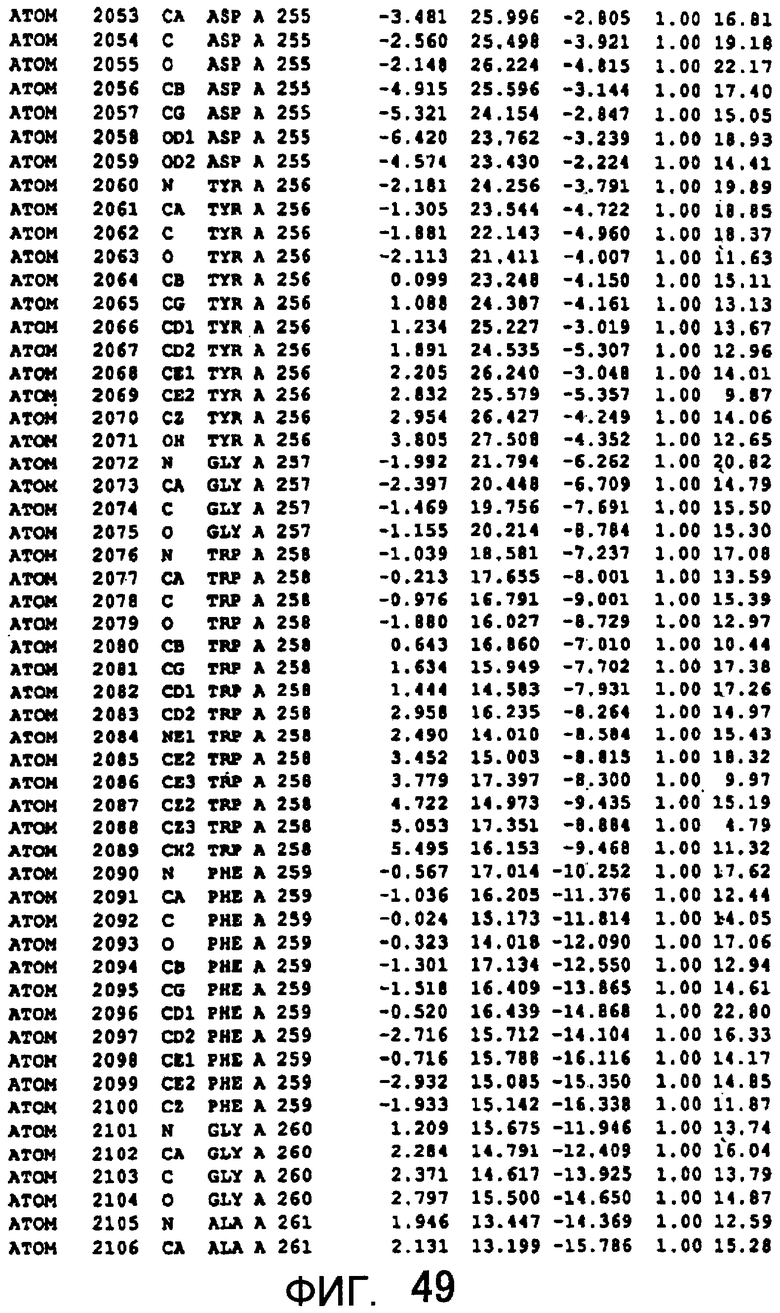
Фигура 49 отображает атомные координаты (39) MTG.
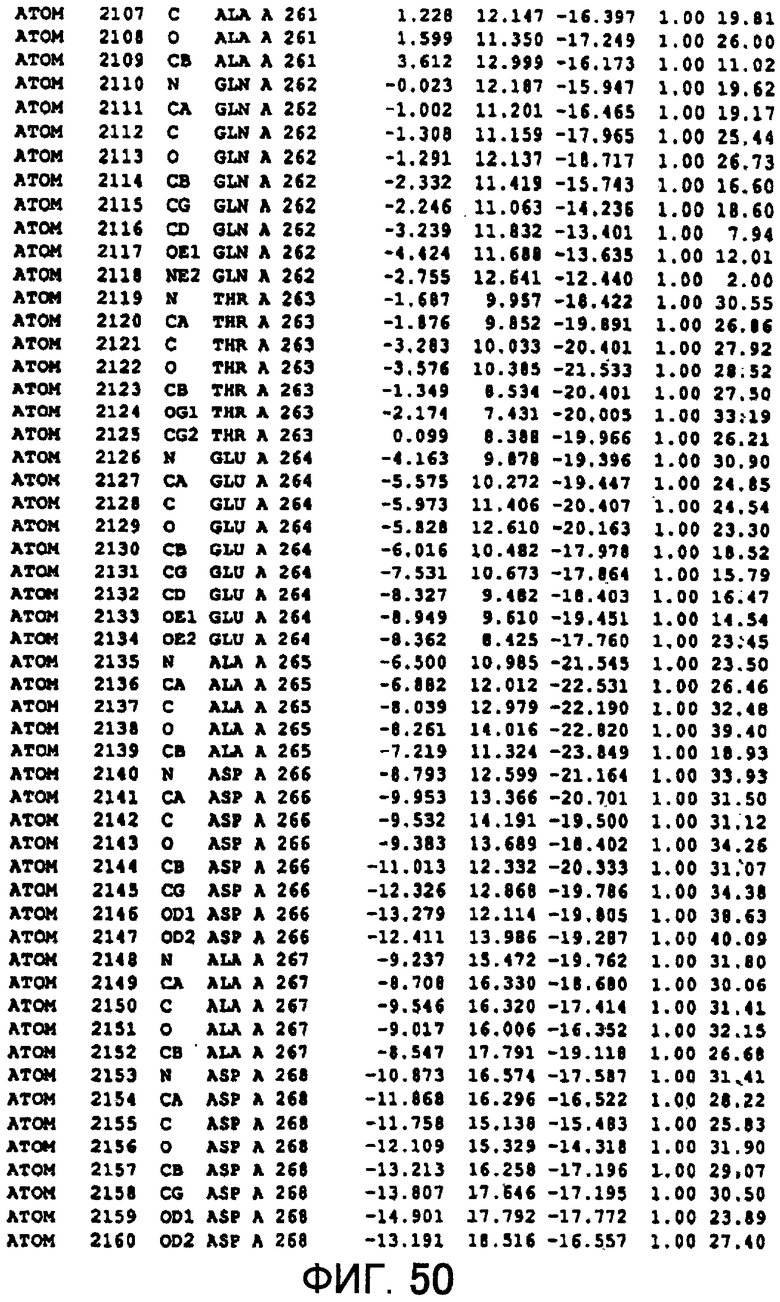
Фигура 50 отображает атомные координаты (40) MTG.
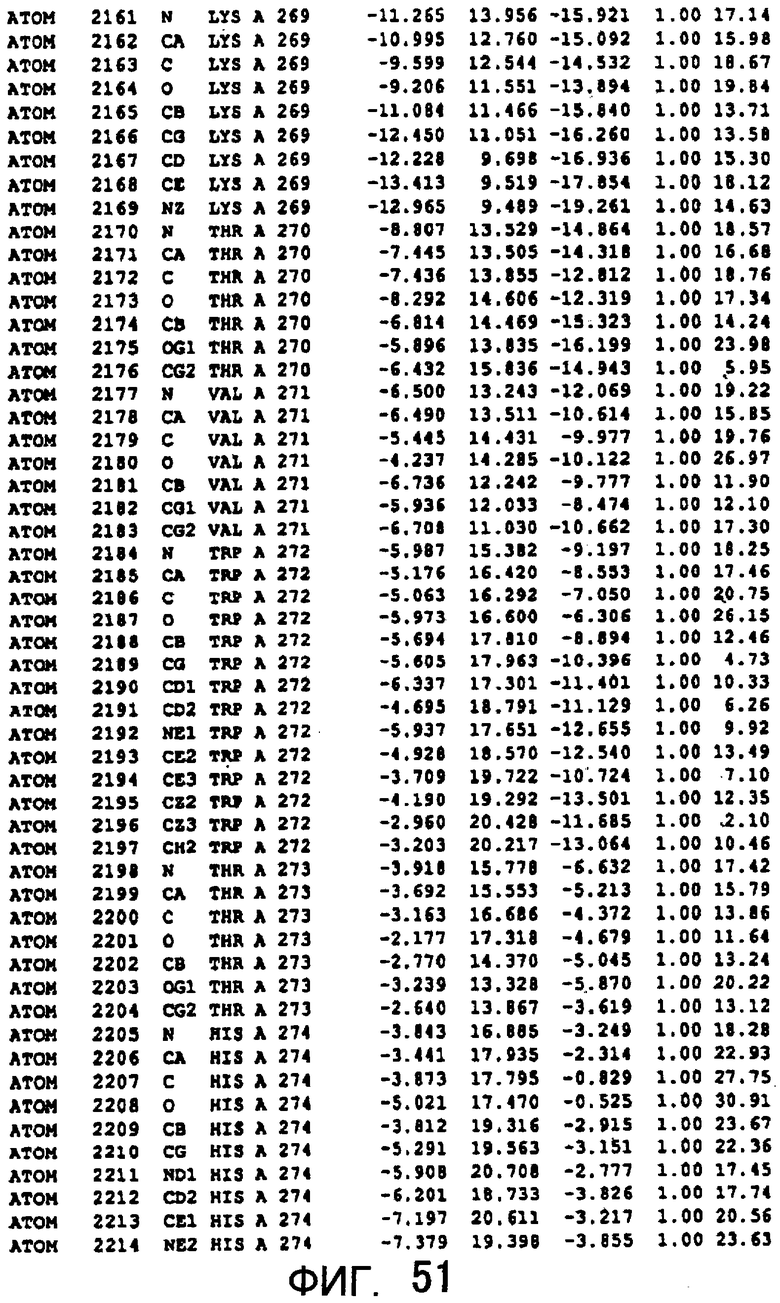
Фигура 51 отображает атомные координаты (41) MTG.
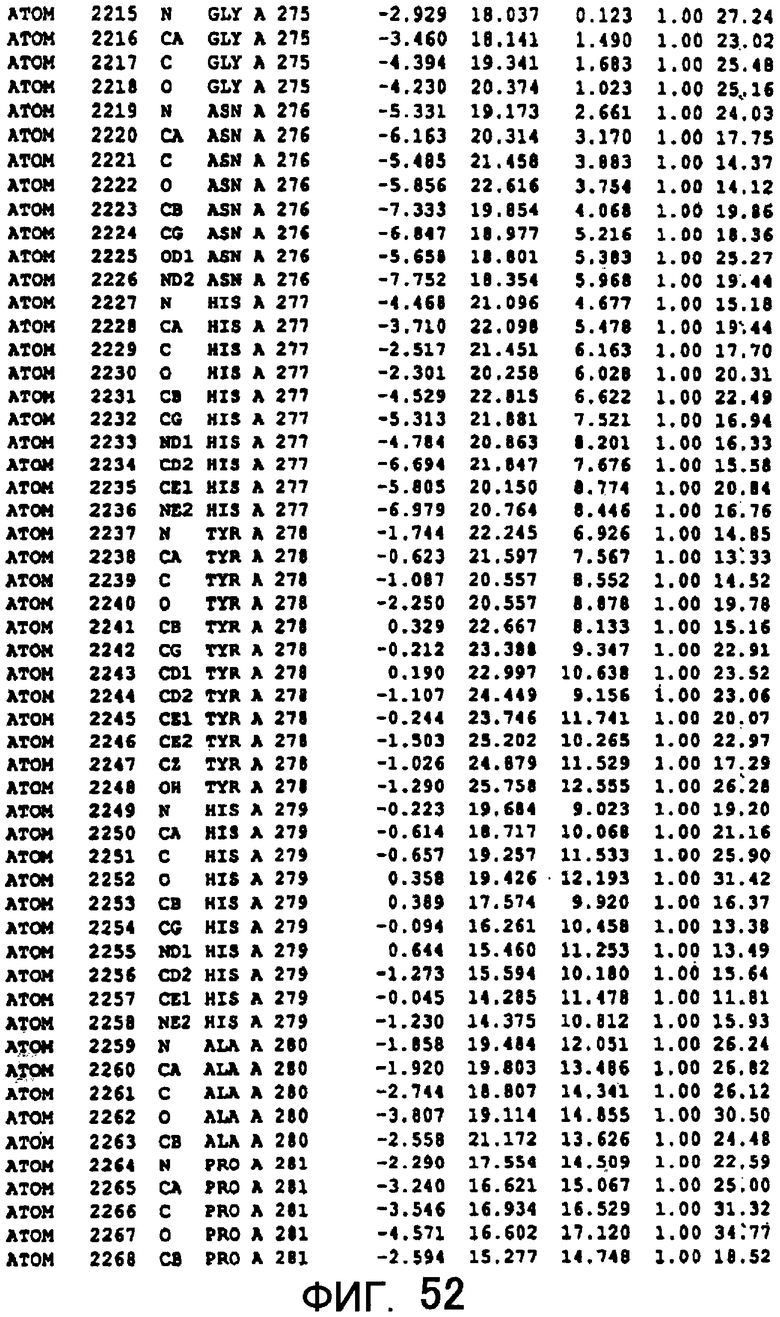
Фигура 52 отображает атомные координаты (42) MTG.
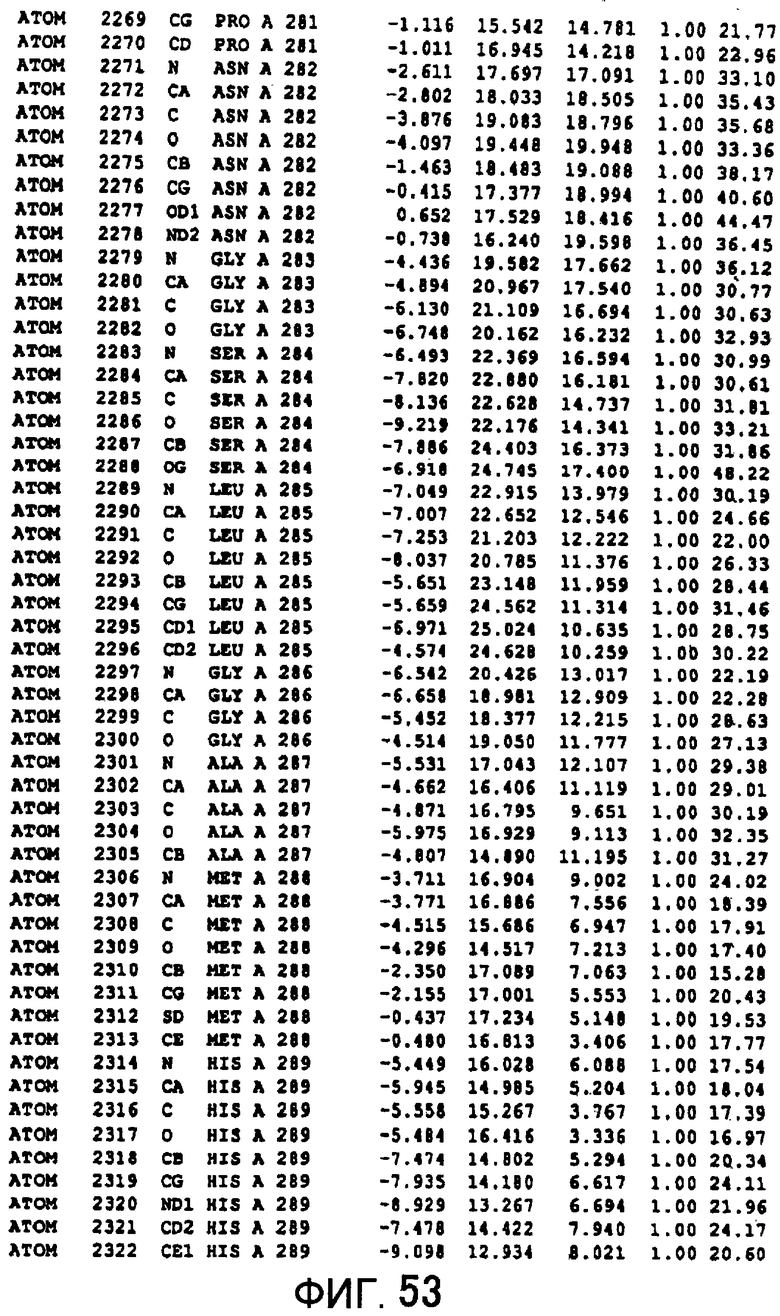
Фигура 53 отображает атомные координаты (43) MTG.
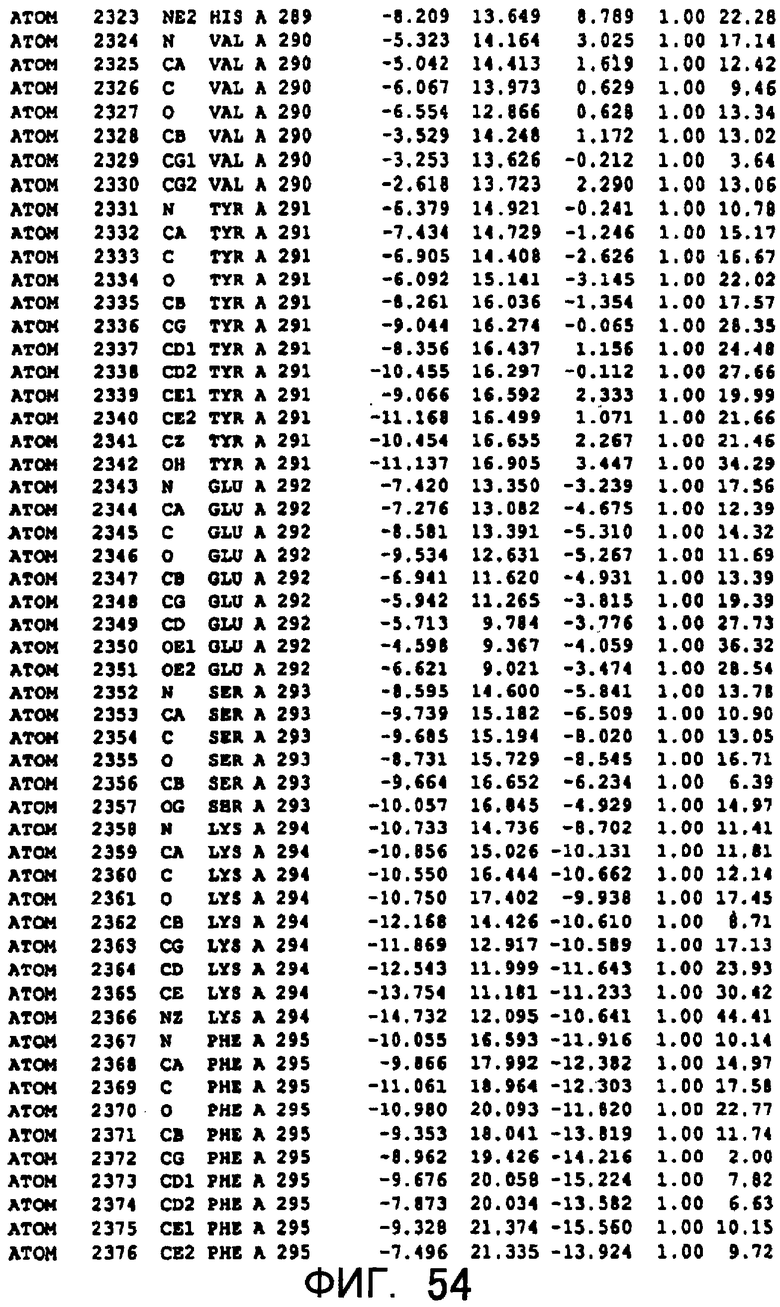
Фигура 54 отображает атомные координаты (44) MTG.
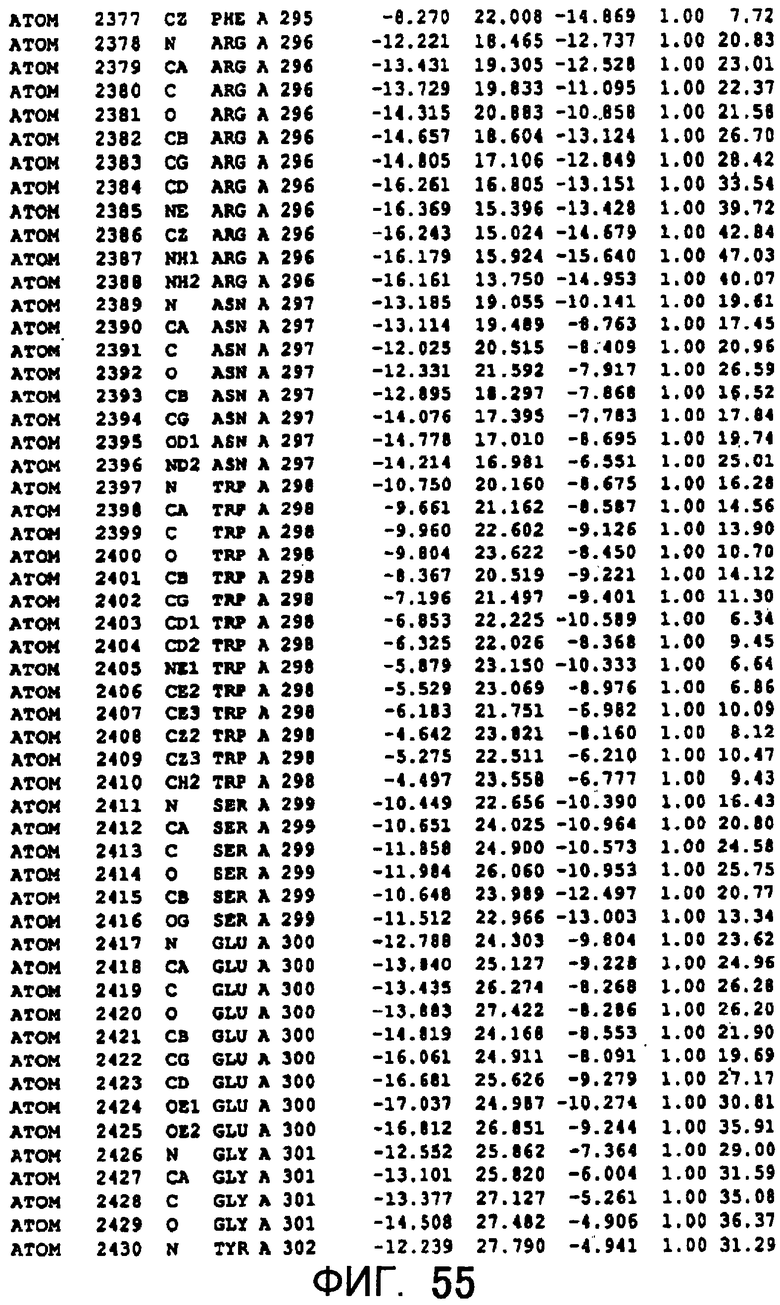
Фигура 55 отображает атомные координаты (45) MTG.
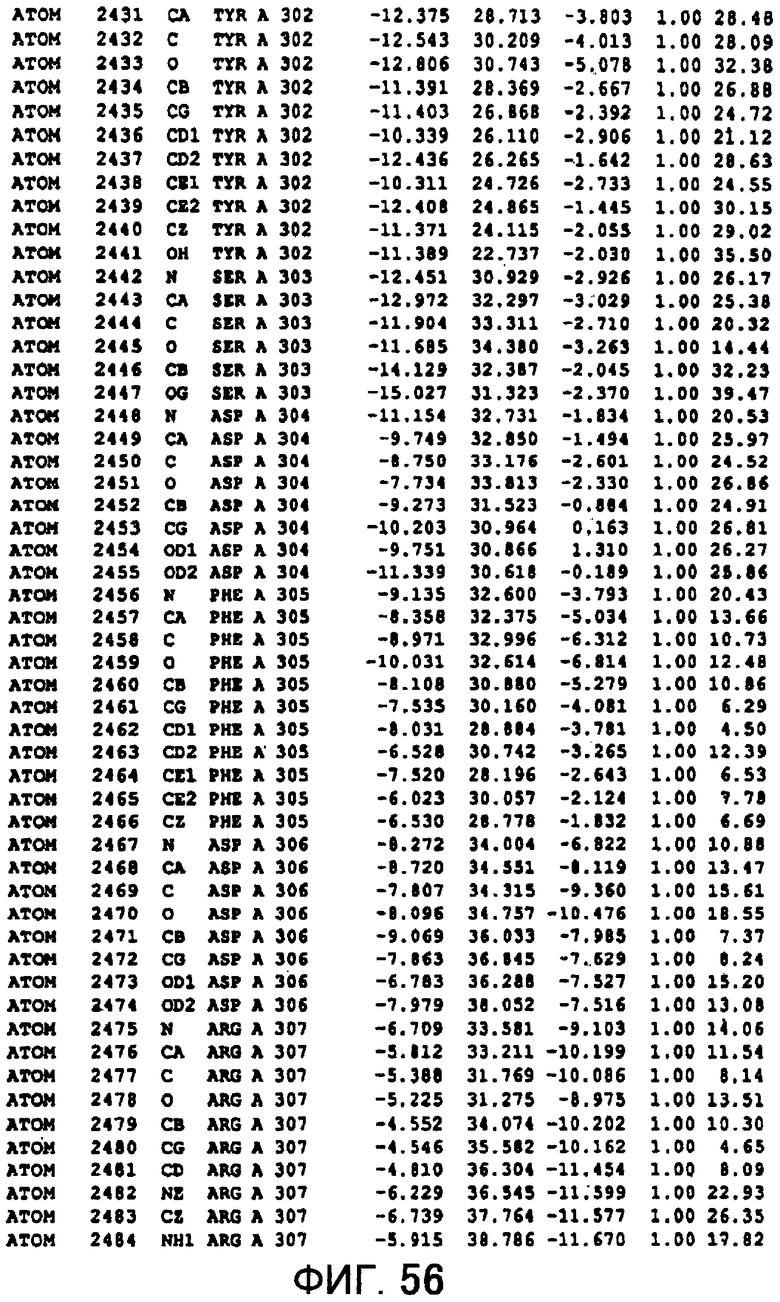
Фигура 56 отображает атомные координаты (46) MTG.
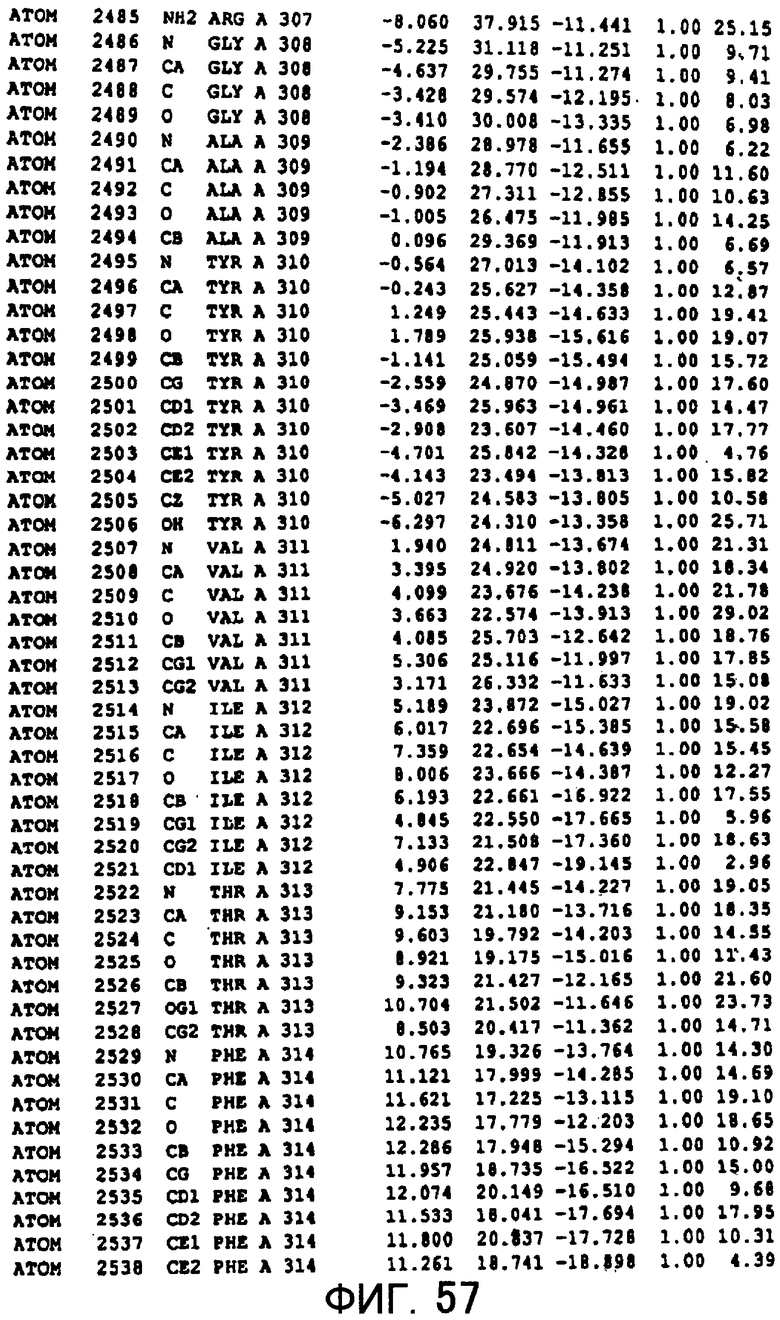
Фигура 57 отображает атомные координаты (47) MTG.
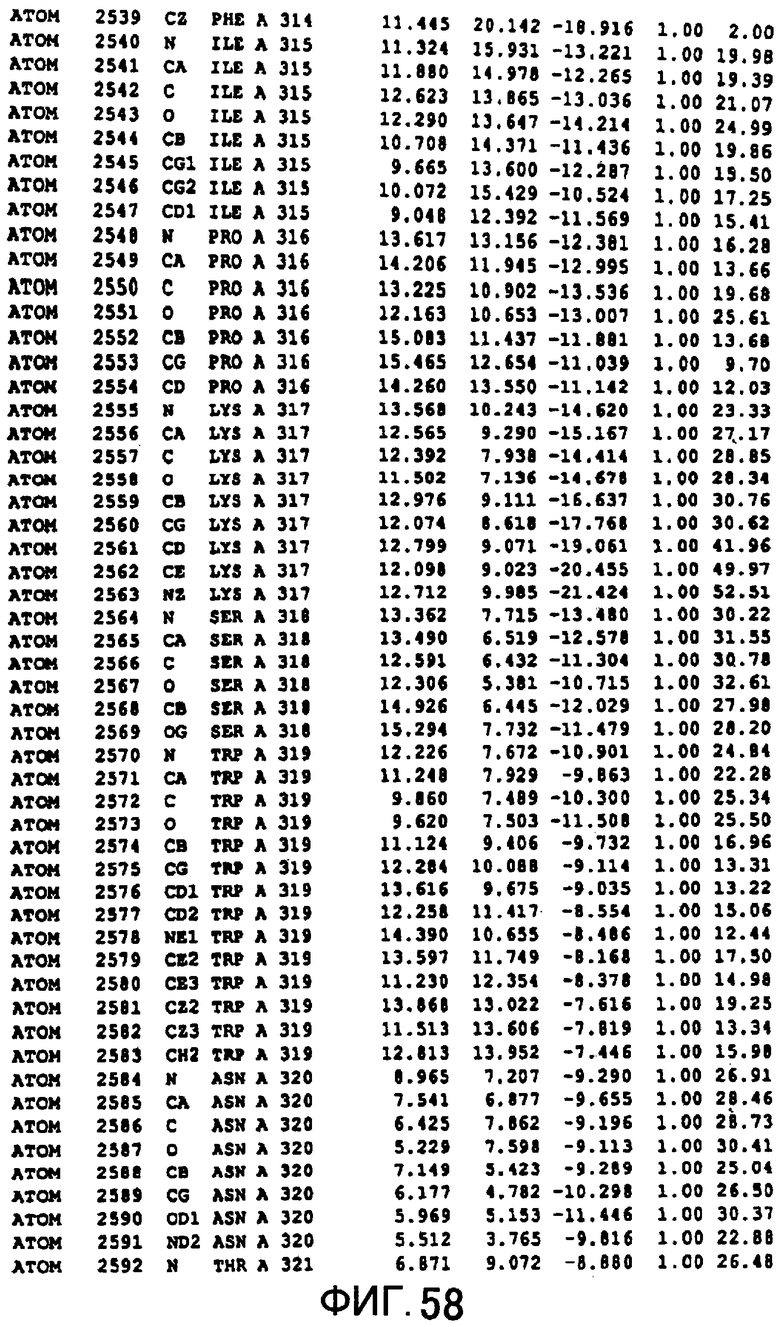
Фигура 58 отображает атомные координаты (48) MTG.
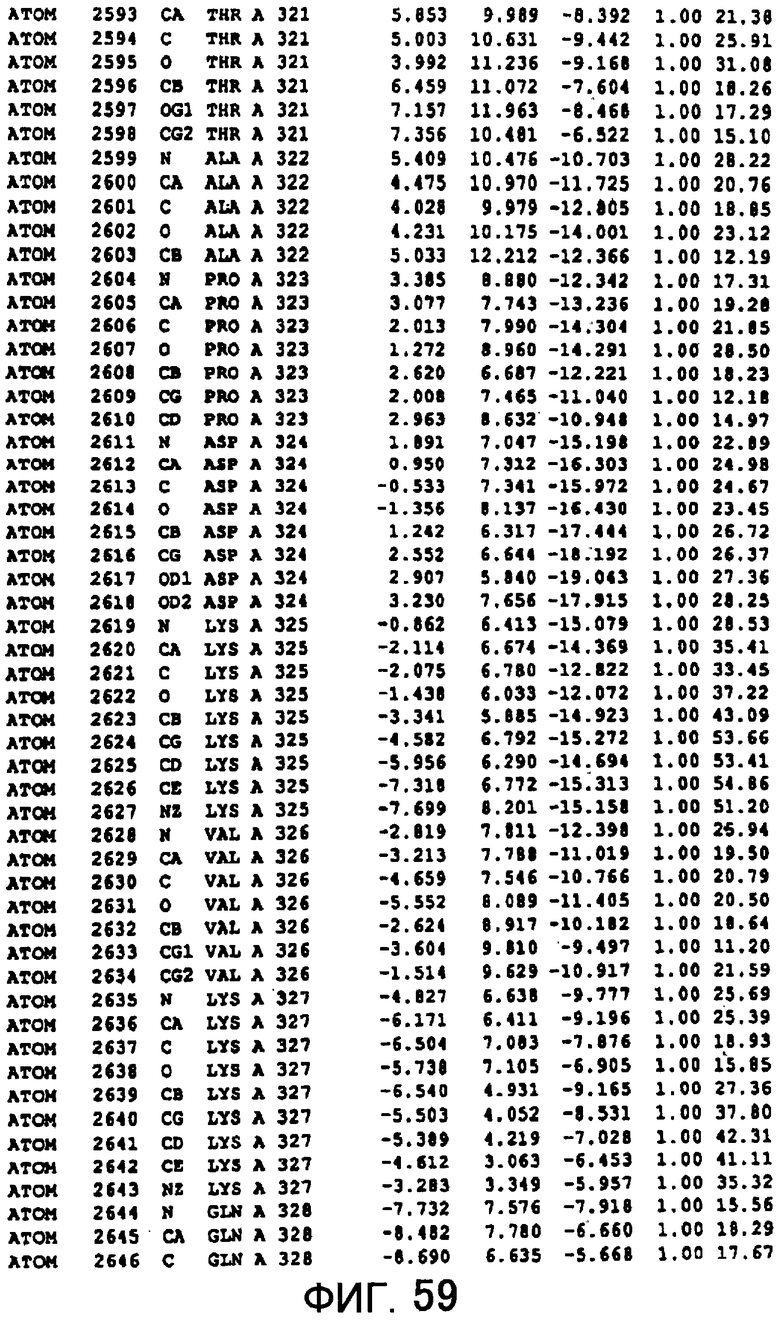
Фигура 59 отображает атомные координаты (49) MTG.
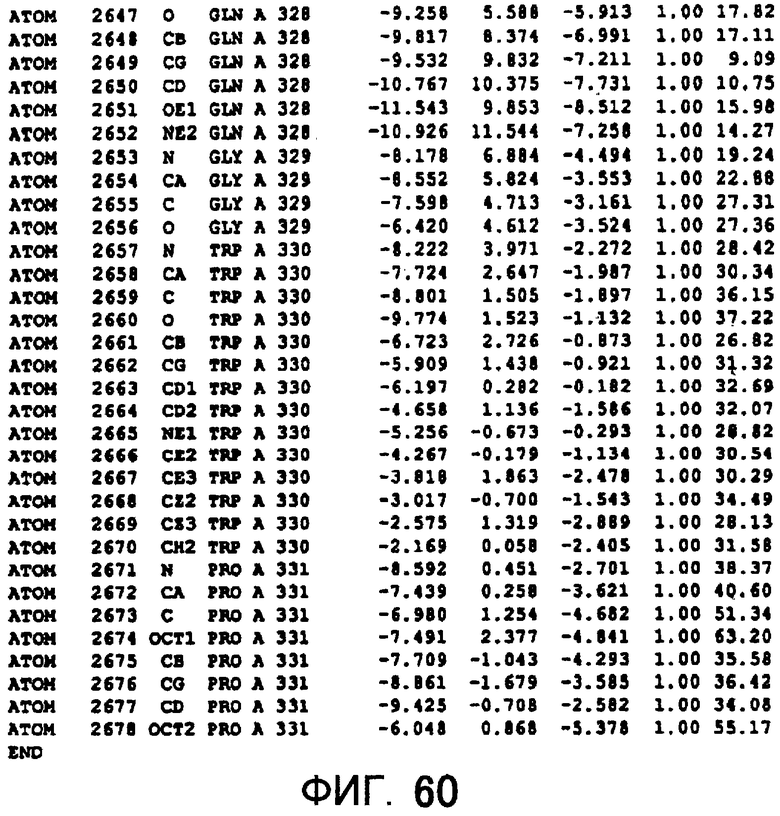
Фигура 60 отображает атомные координаты (50) MTG.
Описание предпочтительных воплощений
Настоящее изобретение будет подробно описано далее.
Трансглутаминаза широко применяется при производстве продуктов питания, таких как желатинов, сыров, йогуртов, тофу, вареных рыбных паштетов, ветчин, колбас и лапши, а также для улучшения качества мясных продуктов (JP-Kokai No.64-27471). Трансглутаминаза также применяется для различных промышленных целей, например, в производстве материалов для термостабильных микрокапсул и в производстве носителей для иммобилизированных ферментов. Трансглутаминаза катализирует реакцию переацетилирования γ-карбоксамидной группы глутаминового остатка, содержащегося в пептидной последовательности молекулы белка. Если в молекуле белка ε-аминогруппа остатка лизина действует в качестве акцептора ацильной группы, в молекуле белка и между молекулами образуется связь типа ε-(γ-Glu)-Lys.
Авторы изобретения обнаружили, что трансглутаминаза не взаимодействует со всеми глутаминовыми остатками, а именно возможность реакции, то есть реакционная способность трансглутаминазы, варьирует в зависимости от типов и расположения остатков, окружающих глутаминовые остатки. Поэтому, модификация реакционной способности трансглутаминазы по отношению к глутаминовым остаткам в определенных условиях окружающей среды приводит к изменению скорости гелеобразования и, как результат, к расширению диапазона дальнейшего применения трансглутаминазы.
В настоящем изобретении для модификации субстратной специфичности MTG с помощью метода рентгеноструктурного анализа определяли ее пространственную структуру, предсказывали способ связывания ее с субстратом и конструировали и осуществляли приемлемые для этой цели мутации. Применяемый здесь термин "субстратная специфичность" обозначает реакционную способность по отношению к каждому глутаминовому остатку в акцепторе ацильной группы.
Определение пространственной структуры белка методом рентгеноструктурного анализа может быть осуществлено в соответствие со следующей процедурой:
(1) Белок кристаллизуют. Кристаллизация является необходимой для определения пространственной структуры. В дополнение кристаллизацию применяют в промышленности для высококачественной очистки белка, а также в качестве способа стабильного хранения белка в высокой концентрации, причем данный способ обладает высокой устойчивостью к действию протеолитических ферментов.
(2) Полученные таким образом кристаллы подвергают воздействию рентгеновского излучения с регистрацией результатов дифракции. Во многих случаях кристаллы белка повреждаются воздействием рентгеновского излучения, что снижает их способность к дифракции. В подобных случаях в последнее время широко распространена методика измерения при низких температурах, когда кристаллы быстро охлаждают до приблизительно -173°C, и измеряют дифракцию в данных условиях. В заключение, для получения данных с высоким разрешением, применяемых для установления пространственной структуры, используется высокоинтенсивное синхротронное излучение.
(3) Для анализа кристаллической структуры в дополнение к данным дифракции требуется фазовая информация. Структура MTG не может быть определена методом молекулярного замещения, так как кристаллические структуры аналогичных белков до сих пор неизвестны. Поэтому фазовая проблема может быть решена с помощью метода изоморфного замещения тяжелым атомом. Данный метод включает введение в состав кристалла атома металла с большой атомной массой, такого как ртуть или платина, и получение фазовой информации благодаря вкладу атома металла, обладающего высокой способностью рассеивать рентгеновское излучение, в данные дифракции рентгеновского излучения. Определенное таким образом значение фазы может быть улучшено за счет сглаживания электронной плотности в контактирующей с раствором области кристалла. В связи с тем, что электронная плотность молекул воды в контактирующей с раствором области кристалла едва уловима вследствие их высоких флуктуаций, значение электронной плотности, близко совпадающее с истинным значением электронной плотности, может быть получено при приравнивании значения электронной плотности в данной области к 0, что улучшает фазу. В том случае когда множество молекул собраны в асимметричный элемент, значение фазы может быть дополнительно значительно улучшено за счет усреднения электронной плотности данных молекул. Модель белка пригодна для расчета карты электронной плотности с помощью улучшенной фазы. Данный расчет выполняют в виде компьютерной графики с помощью программы, такой как QUANTA (MSI Co., U.S.A.). Затем структуру уточняют с применением программы, такой как X-PLOR (MSI Co., U.S.A.), для завершения анализа структуры.
Авторы изобретения кристаллизовали очищенную обычную MTG из Streptoverticillium mobaraense и рекомбинантную MTG (Met-типа) с дополнительным Met остатком от инициирующего кодона N-конца, но не смогли получить пригодный для анализа кристалл. Тем не менее, исходя из того, что полученные кристаллы обычной MTG были лучше, чем кристаллы MTG Met-типа, авторы изобретения получили MTG Asp-типа (Пример 1) после сделанного предположения о том, что начинающаяся с Asp1 рекомбинантная MTG (MTG Asp-типа) подобно обычной, полученной путем удаления Met остатка в N-конце, может обладать способностью к образованию кристаллов, пригодных для анализа. В результате из MTG Asp-типа был получен хороший кристалл (Пример 2) и успешно была получена пространственная структура (Пример 3). Атомные координаты пространственной структуры MTG представлены на Фигурах 11-60.
Считается, что механизм реакции трансглутаминазы является следующим.
Содержащий Gln донор ацильной группы взаимодействует с трансглутаминазой с образованием промежуточного продукта реакции. Затем содержащий Lys акцептор ацильной группы атакует промежуточный продукт с образованием связи типа ε-(γ-Glu)-Lys между донором ацильной группы и акцептором ацильной группы. Для применения белка в качестве донора ацильной группы факт содержания им Gln не является значимым, но при этом необходимо, чтобы участок вокруг Gln мог соответствующим образом связываться с субстратсвязывающим сайтом MTG. Наоборот, в большинстве случаев бывает достаточно, чтобы белок содержал первичный амин в качестве акцептора ацильной группы. Поэтому модификация субстратной специфичности MTG означает модификацию строения сайтов связывания с донором ацильной группы. Если MTG может быть модифицирована таким образом, чтобы уменьшить ее субстратную специфичность или, другими словами, чтобы связываться с большим количеством доноров ацильной группы, увеличивается частота случаев, когда объединение белков становится возможным с применением меньшего количества MTG, а потому ожидается снижение ее применяемого количества. Дополнительно, если становится возможным объединение белков, в которых образование поперечных сшивок изначально было невозможно, то становится возможным применение MTG в новой области. Пространственная структура MTG необходима для предсказания способа связывания между MTG и донором ацильной группы.
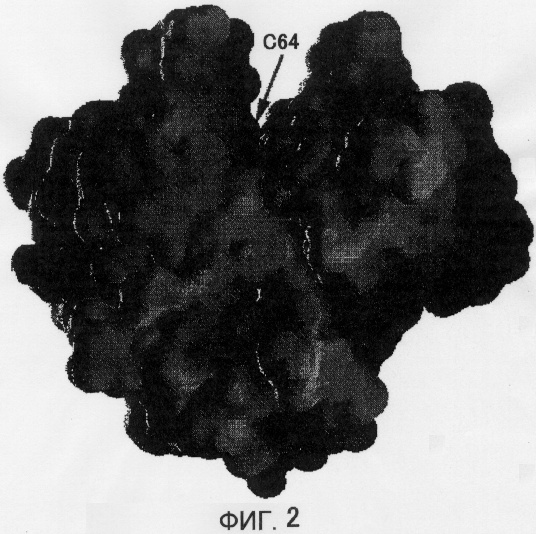
Установленная в настоящем изобретении молекула MTG имеет форму диска с размерами 65×59×41 Å (смотри Фиг.1 и 2). А именно, на Фигуре 1 показана ленточная модель кристаллической структуры MTG. α-Спирали и β-пластины показаны с помощью спиралей и стрелок соответственно. Активный остаток, Cys64, показан с помощью шарово-палочной модели (верхне-центральная часть).
Фигура 2 отражает вогнуто-выпуклую поверхность кристаллической структуры MTG в том же направлении, что на Фигуре 1. Обладающие электрическим зарядом участки обозначены темным светом, а не несущие электрического заряда участки обозначены светлым цветом. При рассмотрении диска сверху обнаруживается клиновидная щель. Во внутренней части щели расположен активный остаток Cys64. Тот факт, что остаток Asp255 расположен по соседству с активным остатком Cys64, наводит на мысль о том, что реакцию обуславливает SH-группа остатка Cys64, которая может дегидрироваться остатком Asp255. Боковая цепь Cys64 обращена к растворителю, и она может достигать донора ацильной группы и акцептора ацильной группы. В Ca2+-зависимом факторе XIII системы свертывания крови человека активный остаток Cys не обращен к растворителю. Поэтому для связывания с донором ацильной группы фактор XIII системы свертывания крови человека должен претерпевать значительные структурные изменения для связывания активного остатка с донором ацильной группы. С другой стороны, считается, что MTG способна связываться с донором ацильной группы без изменения своей структуры. Поэтому высказано предположение, что MTG может использовать больше остатков Gln в качестве субстрата по сравнению с фактором XIII системы свертывания крови человека. Кроме того, MTG обладает тем достоинством, что легко может быть сконструирован ее мутант, обладающий сниженной субстратной специфичностью, так как структурное изменение, необходимое для связывания с донором ацильной группы, может не приниматься во внимание. Более того, молекулярная масса MTG составляет менее 38000, в то время как для фактора XIII системы свертывания крови человека она равна 166000, что делает определение сайта связывания MTG с донором ацильной группы более простым. При рассмотрении структуры MTG становится понятным, что субстрат, скорее всего, связывается в клиновидной щели, направленной от раствора к Cys64.
В щели расположено множество гидрофобных аминокислотных остатков и ароматических аминокислотных остатков, направленных к активному остатку Cys64, и поэтому считается, что их совместное действие с гидрофобными остатками или ароматическими остатками донора ацильной группы является одним из ключей для определения субстратной специфичности. Полярные остатки, включая Asp, также разбросаны на поверхности щели, что наводит на мысль о том, что они играют важную роль в распознавании донора ацильной группы. Обладающая модифицированной субстратной специфичностью мутантная трансглутаминаза может быть получена путем замены, вставки или делеции расположенных в щели аминокислотных остатков, которые предположительно являются участком связывания донора ацильной группы, предпочтительно замещением полярных остатков (Asp, Glu, Lys и Arg) или ароматических остатков (Phe, Tyr и Trp) другими аминокислотами, замещением их полярными остатками или ароматическими остатками, делецией полярных остатков или ароматических остатков или вставкой полярных остатков или ароматических остатков. Необходимо, тем не менее, отметить, что в связи с предположительной утратой активности вследствие модификации остатков Cys64 и Asp255, играющих важную роль в активности, при получении мутантов указанные остатки должны быть исключены.
Как показано в представленных ниже Примерах, в субстратных белках к реакционному сайту прилегают кислые аминокислоты. Во многих случаях взаимодействующие с MTG остатки глутамина расположены на поверхности субстратного белка и считается, что легко контактирующие с растворителем кислые аминокислоты, скорее всего, расположены вокруг них. Если кислые аминокислотные остатки содержатся в реакционном сайте субстратного белка MTG, то отрицательные заряды в MTG и субстратном белке взаимоотталкиваются со снижением скорости ферментативной реакции. Отрицательный заряд может быть устранен путем модификации кислых аминокислотных остатков, представленных на поверхности щели MTG, содержащей остаток Cys64,который предположительно связывается с донором ацильной группы, или, альтернативно, отрицательный заряд может быть уменьшен путем превращения аминокислотных остатков, последовательно или пространственно прилегающих к кислым аминокислотным остаткам, в основные аминокислоты с получением обладающей модифицированной субстратной специфичностью трансглутаминазы. Подобные кислые аминокислоты включают остатки MTG с №№1, 3, 4, 28, 58, 249, 300 и 304, описанные в SEQ ID NO: 2.
Аминокислоты, прилегающие к кислым аминокислотам, представляют собой аминокислотные остатки, располагающиеся в пределах 5 A от карбоксильных групп кислых аминокислот, таких как аспарагиновая кислота или глутаминовая кислота, и предпочтительно представляют собой аминокислотные остатки со степенью взаимодействия с растворителем, по меньшей мере, равной 10%. Они включают остатки MTG с №№2, 5, 59, 248-250, 252, 278, 283-285, 289, 291, 296-297 и 302-303, описанные в SEQ ID NO: 2.
Аминокислотная последовательность MTG представлена на SEQ ID NO: 2, а нуклеотидная последовательность кодирующего MTG гена представлена на SEQ ID NO. 1.
Как показано в Примере 5, было подтверждено, что в молекуле белка скорость реакции на конкретном глутаминовом остатке может быть повышена путем удаления остатков Asp1 или Asp3 или путем замещения остатка Ser2. В связи с тем, что расстояние между активным остатком Cys64 и остатком Asp1 в N-конце составляет 20 Å, предполагается существование возможности модификации субстратной специфичности путем модификации аминокислотных остатков, расположенных в пределах 20 Å от остатка Cys64. В дополнение, в связи с подтверждением субстратной специфичности, по крайней мере, для петлевых участков (233-253 и 276-288), сделано предположение о возможности модификации субстратной специфичности при помощи модификации данных участков. Возможность модификации субстратной специфичности дополнительно возрастает при помощи модификации остатков, расположенных на поверхности расщелины, содержащей остаток Cys64, предположительно связывающийся с донором ацильной группы. Для определения того, обращены ли аминокислотные остатки в сторону растворителя или нет, с применением программы, такой как QUANTA, может быть рассчитана доступность растворителя. Доступность растворителя является выражаемой в процентах величиной, равной частному доступной для растворителя площади остатка белка и доступной для растворителя площади аминокислоты в своей свободной форме. Возможность модификации субстратной специфичности MTG может быть высока в случае модификации остатков, располагающихся в пределах 20 Å от остатка Cys64 и имеющих значение доступности растворителя, равное, по меньшей мере, 10%. Данные остатки соответствуют остаткам MTG с №№1-5, 26, 28, 58-59, 62, 69, 74-75, 77, 79, 235-236, 238-244, 248-250, 252-254, 277-278, 282-287, 289, 291, 296-297 и 300-304, описанным в SEQ ID NO. 2.
При конструировании и получении мутантной трансглутаминазы кодон в кодирующем трансглутаминазу гене, соответствующий аминокислотным остаткам в описанном выше субстратсвязывающем сайте, может быть модифицирован путем сайтспецифического мутагенеза, полученный ген может быть вставлен в подходящий вектор и перенесен реципиенту с последующим культивированием трансформанта. Требуемая мутантная трансглутаминаза может быть выделена из трансформированных культивируемых микробных клеток и оценена.
Методы сайт-специфического мутагенеза для проведения требуемой мутации в требуемых сайтах ДНК включают, например, описанный в Примерах метод PCR [Higuchi, R., 61, in PCR technology; Stockton Press, edited by Eriich, H. A. (1989); Carter P., Meth. In Enzymol., Vol.154, p.3382 (1987)]; и метод с применением фага [Kramer W. and Frits H.J., Methods in Enzymology, Vol.54, p.350 (1987), Kunkel, T A. et al. Methods in Enzymology, Vol.154, p.367 (1987)]. Векторы для введения кодирующих трансглутаминазу генов конкретно не ограничиваются настолько, насколько они могут реплицироваться в реципиенте. В случае применения Escherichia coli в качестве реципиента могут применяться плазмиды, способные автономно реплицироваться в данной бактерии. Например, могут применяться плазмиды pUC19, pET или pGEMEX.
Предпочтительными реципиентами являются, например, линии Escherichia coli, однако любая бактерия может быть использована в качестве реципиента настолько, насколько репликатор сконструированной рекомбинантной ДНК и мутантной трансглутаминазы может функционировать, мутантная ДНК может реплицироваться, а гены мутантной трансглутаминазы могут экспрессироваться в бактерии. Примером предпочтительного реципиента является Escherichia coli BL21(DE3)pLysS при использовании промотора T7 или Escherichia coli JM109 в других случаях.
Для введения реципиенту рекомбинантной ДНК, полученной путем сшивания вектора с фрагментом ДНК, содержащим кодирующий мутантную трансглутаминазу ген, может применяться обычный метод, без какого-либо конкретного ограничения. В случае использования в качестве реципиента Escherichia coli может быть применен метод хлорида кальция [J. Mol. Biol., Vol.53, p.159 (1970)], метод Hanahan [J. Mol. Biol, Vol.166, p.557 (1983)], метод SEM [Gene, Vol.96, p.23 (1990)] и метод по Chung et al. [Proceedings of the National Academy of Sciences of the USA, Vol.86, p.2172 (1989)].
MTG может быть экспрессирована в бактериальной клетке в большой степени путем культивирования трансформанта, несущего рекомбинантную ДНК, содержащую кодирующий мутантную трансглутаминазу ген, в подходящей среде, содержащей источник углерода, источник азота, неорганическое железо и, если это необходимо, дополнительно содержащей источник питания органической природы. Для экстракции мутантной трансглутаминазы из культивируемых клеток после культивации клетки собирают, суспендируют в буферном растворе, подвергают воздействию лизоцима, замораживания-оттаивания, разрушению ультразвуком и тому подобное, а затем разрушают и центрифугируют для разделения суспензии на супернатант и осадок.
В связи с тем, что мутантная трансглутаминаза продуцируется в виде белкового тельца включения и выделяется в виде осадка, она может быть солюбилизирована денатурирующим агентом или тому подобное, а затем денатурирующий агент может быть удален для выделения и очистки белка. Денатурирующими агентами для солюбилизации продуцируемого белкового тельца включения могут быть, например, мочевина (например, 8 M) и гуанидинхлорид (например, 6 M). Обладающий трансглутаминазной активностью белок регенерируют путем снижения концентрации денатурирующего агента при помощи разбавления или тому подобное. Применяемые для разбавления растворы могут включать фосфатные буферы и Трис-буферы.
После регенерации активности действующий белок может быть выделен и очищен при помощи подходящей комбинации известного способа выделения и способа очистки. Примерами таких способов являются высаливание, диализ, ультрафильтрация, гель-фильтрация, ионообменная хроматография, аффинная хроматография и высокоэффективная жидкостная хроматография с обращенной фазой.
Термин "субстратная специфичность" в контексте настоящего изобретения обозначает реакционную способность каждого остатка глутамина в молекуле субстратного белка по отношению к трансглутаминазе. Изменение субстратной специфичности трансглутаминазы означает изменение реакционной способности, по крайней мере, одного остатка глутамина. В том случае когда обладающая измененной субстратной специфичностью трансглутаминаза взаимодействует с белком, степень поперечного сшивания белка, а именно степень гелеобразования, также изменяется по сравнению с результатами, полученными при взаимодействии с обычной трансглутаминазой.
Способы сравнения субстратной специфичности трансглутаминаз включают, например, способ, при котором определяется скорость гелеобразования казеина, способ, при котором полученный в результате трансглутаминазной реакции поперечно-сшитый полимер анализируют при помощи электрофореза, и способ, при котором продукт реакции разделяют на фрагменты и анализируют. Также возможно, что с применением трансглутаминазы проводят взаимодействие субстратного белка с 15N-меченным ионом аммония, а затем определяют меченые остатки глутамина методом ЯМР (заявка на выдачу патента Японии №2000-141152).
Анализ субстратной специфичности при помощи последнего способа представляет собой способ сравнения реакционной способности индивидуального остатка глутамина в белке, основываясь на том выгодном факте, что азот карбоксамидной группы в действующем в качестве субстрата остатке глутамина является 15N-меченным при помощи трансглутаминазы. Субстратные специфичности трансглутаминаз могут сравниваться между собой, основываясь на том выгодном факте, что действующий в качестве субстрата для трансглутаминазы остаток глутамина является специфически 15N-меченным, и что скорость мечения варьирует в зависимости от реакционной способности трансглутаминазы по отношению к остатку глутамина.
Единицу активности трансглутаминазы определяют и обозначают следующим образом.
Проводят взаимодействие бензилоксикарбонил-L-глутаминилглицина и гидроксиламина в качестве субстратов, полученную гидроксамовую кислоту преобразовывают в ее комплекс с железом в присутствии трихлоруксусной кислоты и затем определяют ее количество по поглощению при 525 нм. Таким образом, получают калибровочную кривую, основанную на количестве гидроксамовой кислоты. Количество фермента, продуцирующего 1 мкМ гидроксамата за 1 минуту, обозначают как единицу трансглутаминазной активности, одна единица. Детали данного способа определения сообщались ранее (например, JP-Kokai No.64-27471).
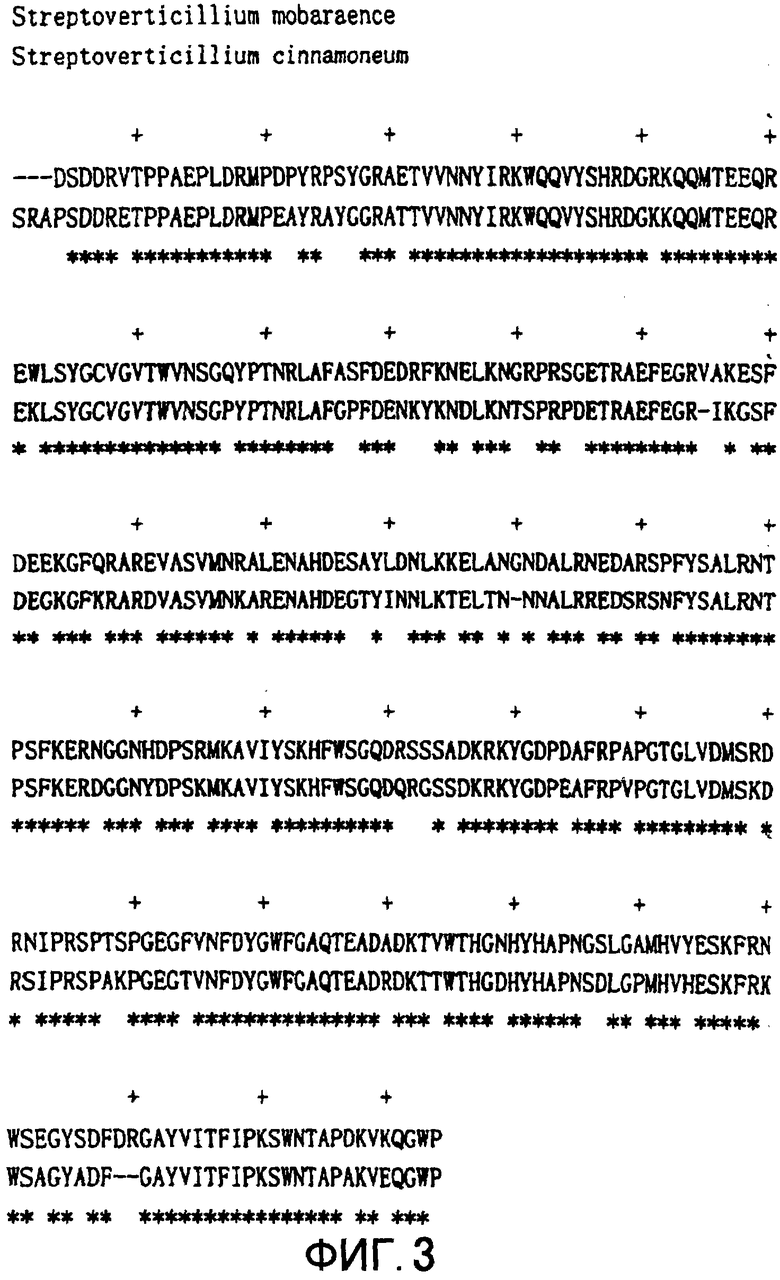
Кроме трансглутаминазы из Streptoverticillium mobaraence (MTG), на основе пространственной структуры MTG также возможно модифицировать фермент, обладающий трансглутаминазной активностью и гомологичной MTG аминокислотной последовательностью, или фермент, обладающий трансглутаминазной активностью и предположительно обладающий сходной с MTG пространственной структурой. Замена аминокислот, эффективная для модификации субстратной специфичности MTG или тому подобное, также эффективна для аналогичных ферментов, таких как ферменты из Streptoverticillium cinnamoneum и Streptoverticillium lydicus (JP-Kokai No.10-504721). Аминокислотная последовательность трансглутаминазы из Streptoverticillium cinnamoneum представлена на SEQ ID NO: 6, нуклеотидная последовательность кодирующего аминокислотную последовательность гена представлена на SEQ ID NO: 5, аминокислотная последовательность трансглутаминазы из Streptomyces lydicus представлена на SEQ ID NO: 8, и нуклеотидная последовательность кодирующего аминокислотную последовательность гена представлена на SEQ ID NO: 7. В этой связи гомологичность аминокислотной последовательности MTG и трансглутаминазы из Streptoverticillium cinnamoneum составляет 78% (Фигура 3), а гомологичность аминокислотной последовательности MTG и трансглутаминазы из Streptomyces lydicus составляет 79% (Фигура 4). А именно, на Фигуре 3 представлено сравнение аминокислотной последовательности MTG и трансглутаминазы из Streptoverticillium cinnamoneum. Сохраненные остатки аминокислот обозначены "*". На Фигуре 4 представлено сравнение аминокислотной последовательности MTG и трансглутаминазы из Streptoverticillium lydicus. Сохраненные остатки аминокислот обозначены "*".
Задание координат остатков аминокислот двух разных белков может быть сделано на основе сравнения аминокислотных последовательностей (Сравнение Сиквенса), если гомологичность их аминокислотных последовательностей превышает приблизительно 20%, и оно может быть сделано на основе пространственных структур и сравнения аминокислотных последовательностей (Тридинг), если гомологичность не превышает приблизительно 20%. Первый может быть выполнен с применением программы, такой как BLAST, а последний может быть выполнен с применением программы, такой INSIGHT II. BLAST может быть получен с помощью ftp в виде адаптированного к применяемому компьютеру файла, среди файлов, представленных в директории "/blast/executable" Интернет-сайта http://ncbi.nlm.nih.gov. Инструкции по применению детально описаны на сайте http://genome.nhgri.nih.gov/blastall/blast install. INSIGHT II является коммерчески доступным (MSI Co.).
Обладающая модифицированной субстратной специфичностью мутантная трансглутаминаза применяется для гелеобразных продуктов питания, таких как желе, йогурты, сыры и гелеобразная косметика; для улучшения качества мясных продуктов; для производства материалов для термостабильных микрокапсул и носителей иммобилизированных ферментов; для ферментативного мечения белков. Если белок является 15N-меченным, то становится возможным проведение структурного анализа при помощи ЯМР (заявка на выдачу патента Японии №2000-141151), а если он помечен первичным амином, то возможно улучшение его растворимости и стабильности (заявка на выдачу патента Японии №2000-141152). Применяемые в качестве субстратов для трансглутаминазы белки могут варьировать. Они включают составляющие мышц, такие как актин и миозин; компоненты плазмы крови человека, такие как альбумин, иммуноглобулин, факторы системы свертывания крови; ферменты, такие как протеазы и трансферазы; гормоны, такие как гормон роста и эритропоэтин; факторы роста клеток для пролиферации или ингибирования клеток; модуляторы иммунных реакций для дифференцировки, индукции и стимуляции клеток; продуцируемые клетками биологическиактивные белки, такие как монокины, цитокины и лимфокины. Источники данных белков конкретно не ограничиваются, и белки могут быть получены от животных, растений и микроорганизмов. Кроме того, ими могут являться белки, экспрессируемые Escherichia coli, дрожжами и клетками животных при введении им генов данных белков, или белки, экспрессируемые с применением бесклеточных систем синтеза белка.
Взаимодействие подобного белка в присутствии трансглутаминазы проводят в условиях, при которых трансглутаминаза может функционировать. Например, субстратный белок и трансглутаминазу инкубируют в диапазоне значений pH от приблизительно 5,0 до 9,0, предпочтительно от приблизительно 6,0 до 8,0, и в диапазоне температур от приблизительно 4 до 55°C, предпочтительно от приблизительно 25 до 40°C, в водном растворителе. В данной реакции концентрация субстратного белка предпочтительно находится в диапазоне от 1 мкМ до 1 M, и количество трансглутаминазы желательно находится в диапазоне от приблизительно 10 нМ до 100 мкМ, что соответствует от приблизительно 0,01 до 20 единиц на 1 М белка. Однако условия проведения реакции не ограничиваются описанными выше.
Диапазон применения MTG в промышленности может дополнительно быть расширен за счет возможной модификации термостабильности и оптимума pH в дополнение к модификации субстратной специфичности. В случае улучшения термостабильности становится возможным гелеобразование белка при более высоких температурах. При производстве упакованного тофу (соевого творога), обычно продаваемого в супермаркетах, MTG добавляют в соевое молоко, затем полученную смесь тщательно упаковывают и затем нагревают. Поэтому температура легко достигает точки денатурации MTG. В случае улучшения термостабильности MTG количество инактивированного денатурированием белка может быть снижено, а соответственно и снижена цена. Термостабильность может быть улучшена, например, путем введения остатка пролина, путем замещения остатков, образующих спиральную структуру с движением против часовой стрелки, остатками глицина [Protein Engineering, Vol.6, pp.85-91 (1993)] или путем заполнения полостей в белке [Biochemistry, Vol.32, pp.6171-6178 (1993)]. Кроме того, если оптимум pH может быть сдвинут в щелочную область, эффективность гелеобразования реагирующего в щелочных реакционных условиях вещества может быть повышена, как в случае с udon, получаемым путем объединения клейковины. Модификация оптимума pH может быть достигнута путем модификации значения pKa активных остатков [Protein Engineering, Vol.11, pp.383-388 (1998)].
ПРИМЕРЫ
Следующие примеры дополнительно иллюстрируют настоящее изобретение, что никоим образом не ограничивает изобретение.
Пример 1 Получение рекомбинантной MTG нативного типа Asp-типа
Экспрессирующая MTG плазмида pETMTGXa-01, которая будет описана ниже, была сконструирована с целью продуцирования MTG, содержащей N-концевую аминокислотную последовательность, идентичную содержащейся в диком типе Escherichia coli. pETMTGXa-01 представляет собой плазмиду, способную экспрессировать слитый белок, содержащий последовательность распознавания (IEGR) фактора Xa между аминоконцевой областью белка гена 10 T7 и MTG. Методика конструирования плазмиды pETMTGXa-01 описана далее. Сначала синтезировали праймер pGEXМТGF01 (SEQ ID NO: 9 в перечне последовательностей), содержащий кодирующий сайт расщепления BamHI, находящийся справа и выше по отношению к гену MTG сайт распознавания фактора Xa, а также содержащий сайт расщепления Sac I праймер pGEXМТGR01 (SEQ ID NO: 10 в перечне последовательностей). PCR проводили с применением этих плазмид и генов MTG в экспрессирующей MTG плазмиде pUCTRPMTG-02(+) (JP Kokai No.11-75876) в качестве матрицы, и клонировали амплифицированный фрагмент в pGEM-T Easy Vector (Promega). PCR проводили в виде 25 циклов в режиме "96°C в течение 30 секунд, 50°C в течение 15 секунд и 60°C в течение 1 минуты". Клонирование амплифицированного фрагмента проводили в соответствие с рекомендациями поставщика. Выбирали клон, в котором амплифицированные фрагменты были введены в направлении, обратном по отношению к гену lacZ. Анализировали нуклеотидные последовательности плазмид в клонах и присваивали плазмиде, соответствующей требуемой последовательности, имя "pGEMMTGXa". Затем вектор pET5a (Takara Shuzo Co., Ltd.) расщепляли при помощи EcoRI, дефосфорилировали концы и расщепляли при помощи BamHI. Плазмиду pGEMMTGXa расщепляли при помощи SalI, дефосфорилировали концы и расщепляли при помощи BamHI, и субклонировали полученный фрагмент, содержащий ген MTG, в описанном выше ферментативно-обработанном векторе pET5a для конструирования плазмиды pETMTGXa-01.
Штамм Escherichia coli BL21(DE3)pLysS (Promega) с интродуцированной плазмидой pETMTGXa-01 засевали в 50 мл среды L (10 г/л триптона, 5 г/л экстракта дрожжей, 5 г/л NaCl и 1 г/л глюкозы; pH 7,2), содержащей 200 мкг/мл ампициллина, и предынкубировали при 37°C в течение 6 часов. Затем 8 мл предкультуры пересевали на 4 часа при 37°C в казаминокислотную среду M9 (8 г/л казаминокислот, 5 г/л хлорида аммония, 0,2 г/л экстракта дрожжей, 2 мг/л гидрохлорида витамина B1, 14,5 мг/л двухводного хлорида кальция, 15,1 г/л двенадцативодного динатрийгидрофосфата, 3 г/л калийдигидрофосфата, 0,5 г/л пятиводного сульфата магния и 5 г/л глюкозы; где пятиводный сульфат магния и глюкозу стерилизовали по отдельности), содержащей 200 мкг/мл ампициллина. При достижении измеренной при 660 нм оптической плотностью (D660) значения, равного приблизительно 2,0, в культуру добавляли 0,8 мл 1M IPTG и дополнительно инкубировали культуру при 37°C в течение 14 часов.
Клетки собирали из бульона центрифугированием и суспендировали в 200 мл смеси 20 мМ Трис-HCl, 30 мМ NaCl и 5 мМ ЭДТА, pH 7,5. К полученной суспензии добавляли 1 мг/мл лизоцима. После инкубации полученной смеси при 4°C в течение 1 часа клетки разрушали обработкой ультразвуком. Содержащие денатурированную MTG белковые тельца включения собирали центрифугированием. Белковые тельца включения тщательно суспендировали в небольшом количестве воды. Для растворения белковых телец включения к полученной суспензии добавляли реагенты и воду таким образом, чтобы 30 мл суспензии содержали 8 M мочевины, 20 мМ фосфата натрия, 1 мМ ЭДТА и 20 мМ DTT. После инкубации при 37°C в течение 2 часов, pH снижали до значения, равного 4, путем покапельного добавления 1 M соляной кислоты, и удаляли осадок центрифугированием. Определяли концентрацию солюбилизированной MTG, оказавшуюся равной приблизительно 20 мг/мл. 20 мл описанного выше раствора солюбилизированной MTG медленно добавляли по каплям к 1000 мл 20 мМ ацетата натрия, pH 4,0, содержащего 2 мМ DTT, и оставляли смесь выдерживаться в течение 2 часов. Для завершения процесса повторной укладки спустя 2 часа к смеси по каплям добавляли 4 M гидроксид натрия для повышения pH до значения, равного 6. Определенная концентрация MTG составила приблизительно 0,14 мг/мл. Процедуру повторной укладки полностью проводили в холодной комнате при 4°C.
Полученный таким образом полный раствор концентрировали до объема, равного 200 мл, ультрафильтрацией (Hydrosart, молекулярное сито 10 кДа; Sartocon Slice Casette, Sartorius) и замещали растворитель 20 мМ Трис-HCl (pH 7,5) с применением Сефадекса G25(M) (Pharmacia Aktiebolag). К 250 мл полученного раствора добавляли 5 мг бычьего фактора Xa (Hematologic Technology Inc.) и оставляли смесь выдерживаться в течение ночи при 5°C. Определенная концентрация MTG составила приблизительно 0,36 мг/мл. Расщепленную фактором Xa N-концевую аминокислотную последовательность MTG анализировали на секвенаторе белка (Shimadzu Corporation) для подтверждения того факта, что прилегающая к N-концу последовательность была полностью расщеплена фактором Xa и что только рекомбинантная MTG содержит N-концевую аминокислотную последовательность зрелого типа.
После расщеплением фактором Xa pH раствора доводили до значения, равного 5,8, добавлением 1 M соляной кислоты, а затем пятикратно разбавляли раствор буферным раствором (20 мМ ацетат натрия, pH 5,8), применяемым для уравновешивания в катион-обменной хроматографии. Полученный раствор наносили на катионную колонку (CM Sepharose FF, 2,6ϕ×10 cm; Amersham Pharmacia Biotech), в достаточной мере уравновешенную буферным раствором. После повторного уравновешивания тем же самым буферным раствором фракцию белка (46 мл), элюированную линейным градиентом концентраций NaCl 0-0,4 M, выделяли с применением УФ-спектроскопии при 280 нм. Выделенную фракцию десятикратно разбавляли буферным раствором (20 мМ ацетат натрия, pH 5,5), применяемым для уравновешивания в катионобменной хроматографии. Треть разбавленного раствора наносили на катионную колонку (6 мл Resource S; Amersham Pharmacia Biotech), в достаточной мере уравновешенную тем же самым буферным раствором. После повторного уравновешивания тем же самым буферным раствором фракцию белка, элюированную линейным градиентом концентраций NaCl 0-0,5 M, разделяли на фракции с применением УФ-спектроскопии при 280 нм. Фракцию (2 мл), без определяемых методом ВЭЖХ с обращенной фазой примесей и свободную от форм MTG с низким значенем pI, выделяли из фронтальной части верхушки пика. Сходную очистку проводили дважды, и подвергали повторному хроматографическому разделению фракцию, побочную выделяемой, с получением в результате 8 мл (40 мг) фракции. Затем растворитель замещали 20 мМ фосфорной кислоты (pH 6,0) с применением Сефадекса G25 (M). Выделенную фракцию анализировали с применением изоэлектрического фокусирования для подтверждения того факта, что она представляет собой высокоочищенную рекомбинантную MTG, практически свободную от имеющих различное значение pI аналогов. Все хроматографические процедуры проводили при комнатной температуре.
Пример 2 Кристаллизация
Кристаллизацию проводили с применением метода диффузии пара в варианте "висячей капли". Одинаковые количества (2 мкл) раствора рекомбинантной MTG Asp-типа (концентрация: 15 мг/мл) и 83 мМ буферного раствора (pH 5,0) какодилата натрия, содержащего 25% полиэтиленгликоля 1000 и 25 мМ хлорида кальция, наносили по каплям на силиконизированное покровное стекло и смешивали. Покровное стекло помещали в буферный раствор таким образом, что капли смешанной жидкости нависали над лункой, заполненной 500 мкл буферного раствора, и оставляли выдерживаться при 20°C. Кристаллы формировались спустя неделю. Спустя 10 суток кристаллы вырастали в пластинчатые кристаллы достаточного для измерения размера (приблизительно 0,5×0,3×0,1 мм). Данный кристалл MTG анализировали при низкой температуре, так как кристалл MTG должен был повреждаться рентгеновским излучением, что приводит к значимому снижению разрешения при проведении анализа при обычной температуре. Кристалл переносили в 117 мМ буферный раствор (pH 5,0) какодилата натрия, содержащий 35% полиэтиленгликоля 1000 и 35 мМ хлорида кальция, и быстро охлаждали продувкой азотом при -173°C.
Результаты дифракции рентгеновских лучей с разрешением 2,7 Å получали на рентгеновском дифрактометре R-AXIS IIc (Rigaku Co., Ltd.) и определяли кристаллографические параметры. Спейсерной группой являлась P21, параметры кристаллической решетки составляли: a=78,4 Å, b=117,1 Å, c=85,7 Å и p=112,9°. Принимая в расчет, что асимметричная единица состоит из 4 молекул (молекулярная масса: 38,000), рассчитывали содержание воды в кристаллах, равное 50%.
Пример 3 Определение пространственной структуры
Кристаллы погружали в раствор солей тяжелых металлов и проводили скрининг производных тяжелых металлов. Результаты дифракции производных кристаллов с тяжелыми атомами получали с применением SMART6000 (Bruker) и оборудования для синхротронного облучения BL-6B (National Laboratory for High Energy Physics). Исходя из различия диаграммы Патерсона с обычными данными было показано, что хорошие изоморфнозамещенные тяжелыми атомами кристаллы получаются с применением EMTS, K2OSCl6 и K2IrCl6. Основные сайты связывания тяжелых металлов определяли исходя из дифференциальных диаграмм Патерсона соответственных изоморфнозамещенных тяжелыми атомами кристаллов, а затем с применением дифференциальных диаграмм Фурье определяли другие второстепенные сайты связывания тяжелых металлов. Координаты данных сайтов связывания тяжелых металлов уточняли с помощью программы определения фаз "MLPHARE" и рассчитывали фазы. С применением программы DM фазу усиливали путем solvent flattering и сглаживания гистограмм, а затем получали карты электронной плотности. В карте электронной плотности в составе асимметричной единицы были установлены 4 молекулы MTG. Для значительного улучшения качества электронной плотности соответствующая 4 молекулам электронная плотность была усреднена с помощью программы DM. Полученные с разрешением 2,7 Å карты электронной плотности оказались очень хорошими, и все аминокислотные остатки, от N-концевого до C-концевого, смогли быть вписаны в электронную плотность.
Первую модель сконструировали с помощью компьютерной графической программы QUANTA и уточнили структуру с помощью программы X-PLOR. Окончательные модели (Фигуры 1 и 2), уточненные с помощью данных рентгеновской дифракции с разрешением 2,4 Å на оборудовании для синхротронного облучения BL-6B (National Laboratory for High Energy Physics, Tsukuba), содержали все 331 аминокислотный остаток и 400 молекул воды. Кристаллографический фактор достоверности (R фактор), определенный с применением отражения при разрешении от 10 до 2,4 Å, составил 19,6%. На полученном с помощью программы PROCHECK графике Рамачандрана показано, что 82% отличных от глицина остатков расположены в наиболее предпочтительном участке, а 15% - во втором предпочтительном участке.
Пример 4 Получение сайт-специфического мутанта
В связи с тем, что N-концевой Asp1 расположен на входе в щель, предполагается, что он обладает некой способностью к распознаванию субстрата (Фигуры 1 и 2). Боковая цепь Asp1 не взаимодействует с другими образующими фермент остатками и поэтому считается, что даже в случае модификации остатка не существует риска искажения пространственной структуры белка. Эффекты Asp1 на субстратную специфичность исследовали путем получения мутанта, в котором данный остаток был удален (здесь и далее называемый "мутантом Ser-типа", так как он начинается с Ser2), и сравнения его субстратной специфичности с диким типом. В связи с тем, что Escherichia coli, полученная путем трансформирования плазмиды, содержащей кодирующий мутант Ser-типа ген, уже была сконструирована (JP-Kokai No.11-075876), для получения мутанта Ser-типа мы использовали эту плазмиду.
Также было сделано предположение, что субстратная специфичность может быть изменена путем замещения расположенного на входе в канал Ser2 другой аминокислотой. Мы сконструировали трех мутантов, то есть S2Y, полученного замещением Ser2 на Tyr с целью усилить гидрофобное взаимодействие, и S2R и S2D, полученных замещением Ser2 на Arg и Asp соответственно с целью усилить электростатическое взаимодействие. Данные мутанты вводили в MTG, содержащую дополнительный Met, соответствующий инициирующему кодону N-конца (Met-тип).
Дополнительно A-цепь инсулина (Sigma) с окисленным остатком цистеина была взята в качестве субстратного белка, и были исследованы глутаминовые остатки, которые могут являться субстратом для MTG. A-цепь инсулина представляет собой белок (SEQ ID NO: 23), состоящий из 21 аминокислотных остатков с глутаминовыми остатками в 5м и 15м положении относительно N-конца. Причиной выбора A-цепи инсулина для изучения структурных характеристик субстратного белка является тот факт, что она содержит множество глутаминовых остатков и позиции, взаимодействующих с MTG глутаминовых остатков могут быть легко определены в связи с ее малой молекулярной массой.
200 мкл 1 M 15NH4Cl и 2 мкл 1 мМ MTG добавляли к 1 мг A-цепи инсулина с окисленными остатками цистеина, перемешивали и проводили масс-спектрометрический анализ. Масс-спектрометрический анализ проводили во время фрагментирования индивидуальных аминокислотных остатков с целью выявления 15N-меченного остатка, которым, как было доказано, оказался глутаминовый остаток в 15м положении относительно N-конца. В результате было показано, что из двух глутаминовых остатков A-цепи инсулина глутаминовый остаток в 15м положении являлся субстратом для MTG.
Глутаминовая кислота присутствует в 17м положении A-цепи инсулина. Данный остаток глутаминовой кислоты прилегает к глутаминовому остатку в 15м положении с одним вставленным между ними остатком. В связи с тем, что боковые цепи аминокислот в полипептидной цепи склонны быть направленными в одном направлении, если между ними вставлен один остаток, считается, что глутаминовый остаток в 15 м положении и остаток глутаминовой кислоты в 17м положении не только близки друг другу в последовательности, но и пространственно. В связи с тем, что кислый аминокислотный остаток расположен рядом с глутаминовым остатком в 15м положении, не является предпочтительным, чтобы обладающая отрицательным электрическим зарядом аминокислота присутствовала в субстратсвязывающем сайте MTG. В большинстве случаев реакционноспособные по отношению к MTG глутаминовые остатки расположены на поверхности субстратного белка. Считается, что кислые аминокислоты, которые могут легко контактировать с растворителем, могут окружать глутаминовый остаток. Это также применимо и для отличных от инсулина субстратных белков.
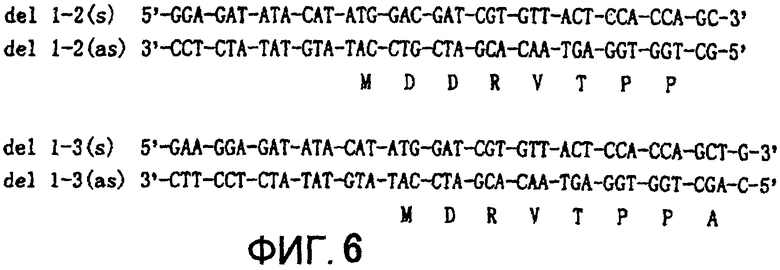
Соответственно, высказано предположение, что субстратная специфичность может быть усилена в том случае, когда отрицательный заряд понижается путем модификации аминокислот или путем преобразования окружающих аминокислот в основные аминокислоты с получением мутантной трансглутаминазы, обладающей измененной субстратной специфичностью. Мутантная MTG Ser-типа, где была удалена аспарагиновая кислота в положении SEQ ID No: 2, и S2R-мутант, в котором для понижения отрицательного заряда в окружении кислых аминокислот были расположены основные аминокислоты, соответствуют указанному выше критерию. Они удовлетворяют описанным ранее условиям. Дополнительно мутанты del 1-2 получали путем удаления остатков с №№1 и 2, а мутанты del 1-3 получали путем удаления остатков с №№1-3 для устранения отрицательного заряда Asp1 и Asp3, соответственно.
В связи с тем, что окружающие активный остаток Cys64 две петли (233-253 и 276-288) расположены таким образом, что субстрат вставляется между ними, то скорее всего именно они определяют субстратную специфичность. Однако петли не могут быть слепо удалены, так как они играют важную роль в укладке MTG. Поэтому на основе пространственной структуры MTG проводился поиск сайтов, удовлетворяющих следующим условиям (1) и (2): (1) сайт прочно реагирует с отличными от петель сайтами и не влияет на укладку, и (2) расстояние между N-концом и C-концом удаляемого сайта (остатка) не превышает 7 Å, и он может быть замещен 3 остатками глицина. 3 остатка глицина упоминаются только в той связи, что с их участием может быть образована измененная структура, но не в ограничительном контексте. Например, в связи с тем, что остатки с №№241-252 практически не взаимодействуют с другими сайтами, а расстояние между N-концом остатка №241 и C-концом остатка №252 составляет приблизительно 6 Å, остатки с №№241-252 могут быть замещены 3 остатками глицина. В качестве мутантов, удовлетворяющих таким условиям, когда окружающие активный остаток Cys64 две петли удаляются для выявления их вклада, была сделана попытка получить мутант Sg4 путем замещения остатков с №№241-252 3 остатками глицина и мутант Sg7 путем замещения остатков с №№278-287 3 остатками глицина.
(1) Получение MTG Ser-типа
Штамм Escherichia coli JM109, несущий плазмиду pUCTRPMTG-02(+), содержащую ген MTG Ser-типа (JP-Kokai No.11-075876), засевали в 50 мл среды L, содержащей 200 мкг/мл ампициллина, и предынкубировали при 30°C в течение 7 часов. Затем 16 мл предкультуры переносили в 800мл среды M9 и проводили дополнительную инкубацию при 37°C в течение 24 часов. Клетки собирали из бульона центрифугированием и суспендировали в 12,5 мл 20 мМ Трис и 30 мМ NaCl. К полученной суспензии добавляли 1 мг/мл лизоцима и 0,5 М ЭДТА и оставляли выдерживаться при 4°C в течение 1 часа. Затем клетки разрушали обработкой ультразвуком в течение 20 минут. Содержащие денатурированную MTG белковые тельца включения собирали центрифугированием. Белковые тельца включения тщательно суспендировали в небольшом количестве воды и затем для растворения белковых телец включения к полученной суспензии добавляли реагенты и воду таким образом, чтобы 5 мл суспензии содержали 8M мочевины, 20мМ фосфата натрия, 1 мМ ЭДТА и 20 мМ DTT.
После инкубации при 37°C в течение 2 часов, pH снижали до значения, равного 4, и удаляли осадок центрифугированием. Раствор солюбилизированной MTG медленно добавляли по каплям к 250 мл 20 мМ ацетата натрия (pH 4,0), содержащего 2 мМ DTT, и оставляли смесь выдерживаться в течение 2 часов. Спустя 2 часа pH повышали до значения, равного 5 и удаляли нерастворимые вещества центрифугированием. Для завершения процесса повторной укладки повышали pH до значения, равного 6. Процедуру повторной укладки полностью проводили в холодной комнате при температуре 4°C.
После замещения 250 мл раствора 20 мМ раствором ацетата натрия (pH 5,8) с сефадексом G25(M) (Pharmacia) выделяли белковую фракцию (55 мл), которую элюировали из ионообменной смолы (CM Sepharose FF, Pharmacia) линейным градиентом концентраций NaCl 0-0,4 M в 20 мМ растворе ацетата натрия (pH 5,8). Затем раствор замещали 20 мМ раствором фосфата натрия (pH 6,0) с сефадексом G25(M).
Если следующей за N-концевым Met аминокислотой является Ser и N-концевой Met может быть удален метионинаминопептидазой Escherichia coli, для получения мутанта Ser-типа, содержащего Ser в N-конце, соответствующий Ser кодон может быть расположен после инициирующего кодона.
(2) Получение мутанта замещением Ser2
Для продуцирования трех мутантных MTG (S2Y, S2R и S2D) Escherichia coli экспрессирующую MTG плазмиду pGEMMTG3 (кодирующая аминокислотную последовательность в SEQ ID No: 4 и содержащая последовательность оснований SEQ ID No: 3) конструировали и применяли в качестве матрицы для сайтспецифического мутагенеза с применением PCR. pGEMMTG3 представляет собой плазмиду, в высокой степени экспрессирующую MTG с индукцией промотором T7 и сконструированную путем удаления гена белка гена 10 T7 из плазмиды pGEM15TG(Xa), экспрессирующего слитый белок аминоконцевого участка белка гена 10 T7 и MTG [Biosci. Biotech. Biochem., Vol.61 (5), pp.830-835 (1997)] и введения синтетического гена, сконструированного таким образом, что становилась возможной экспрессии Escherichia coli MTG в высокой степени, как описано в JP-Kokai No.11-75876.
Процедуры конструирования pGEMMTG3 описаны ниже. Сначала синтезировали праймер pGEMTGF01 (SEQ ID No: 11), сконструированный путем удаления 42 остатков из белка гена 10 T7 и содержащий сайт расщепления PstI в своем 5'-конце, и праймер pGEMTGR01 (SEQ ID No: 12), содержащий сайт расщепления EcoRI в гене MTG. PCR проводили с применением этих праймеров и плазмиды pGEM15TG(Xa) в качестве матрицы и клонировали амплифицированные фрагменты в pUC19 (Takara Shuzo Co., Ltd.). PCR проводили в виде 25 циклов в режиме "96°C в течение 30 секунд, 50°C в течение 15 секунд и 60°C в течение 1 минуты". Анализировали нуклеотидные последовательности плазмид в клонах и присваивали плазмиде, соответствующей требуемой последовательности, имя "pUCTGN". В связи с тем, что плазмида pGEM15TG(Xa) содержит два сайта расщепления Ndel, фрагмент Ndel/EcoRI плазмиды pUCTGN замещали фрагментом Ndel/EcoRI плазмиды pGEM15TG(Xa) путем последовательности описанных ниже событий. А именно, небольшой фрагмент, полученный путем расщепления плазмиды pGEM15TG(Xa) с помощью Smal и EcoRI, субклонировали в векторе pBluescript (Toyobo Co., Ltd.), расщепленном с помощью Smal и EcoRI с получением плазмиды "pBS15TG(SE)". Затем фрагмент, содержащий ген трансглутаминазы, полученный путем расщепления плазмиды pUCTGN с помощью Ndel и EcoRI, сшивали с несодержащим ген трансглутаминазы сегментом, полученным путем расщепления плазмиды pBS15TG(SE) с помощью Ndel и EcoRI, с получением плазмиды "pBSTG(SE)". Затем большой фрагмент, полученный путем расщепления pGEM15TG(Xa) с помощью Smal и EcoRI, сшивали с содержащим ген трансглутаминазы сегментом, полученным путем расщепления pBSTG(SE) с помощью Ndel и EcoRI, получением "pGEMTG". В заключение для конструирования плазмиды pGEMMTG сегмент гена MTG (фрагмент расщепления PvuII - HindIII) плазмиды pGEMTG замещали геном MTG (фрагмент расщепления PvuII - HindIII) плазмиды pUCTRPMTG-02(+) (описанный в JP-Kokai No.11-75876).
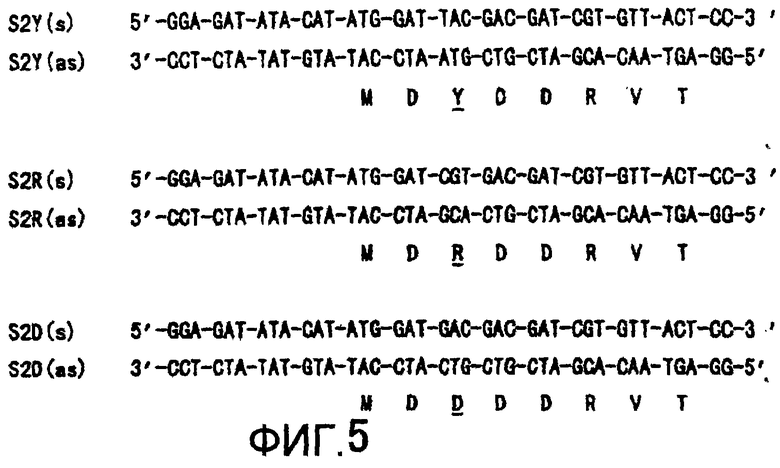
Мутацию проводили с применением набора для быстрообменного (зарегистрированная торговая марка) сайтспецифического мутагенеза (Stratagene) с применением праймера (Фигура 5, SEQ ID NOS: 13-18), соответствующего любому мутантному ферменту в соответствии с протоколом производителя. А именно, на Фигуре 5 представлены участки праймера, применяемые при сайтспецифическом мутагенезе мутанта S2Y·S2·PS2D. Escherichia coli JM109 (Takara Shuzo Co., Ltd.) трансформировали PCR-продуктом. Трансформированные клетки помещали в плашки с L-агаровой средой, содержащей 100 мкл/мл ампициллина, и инкубировали при 37°C в течение 16 часов. Полученные таким образом колонии собирали и культивировали в L-среде, содержащей 100 мкл/мл ампициллина, в течение ночи при встряхивании. Клетки отделяли от бульона путем центрифугирования, и экстрагировали плазмиду с применением Automatic DNA Isolation System PI-50 (KURABO) в соответствие с протоколом производителя. Кодирующая каждую мутантную MTG нуклеотидная последовательность была подтверждена при помощи ДНК-секвенирования. Плазмиду трансформировали в Escherichia coli BL21 (Promega).
Escherichia coli BL21(DE3) с интродуцированным геном MTG засевали в 50 мл среды L (10 г/л триптона, 5 г/л экстракта дрожжей, 5 г/л NaCl и 1 г/л глюкозы; pH 7,2), содержащей 200 мкг/мл ампициллина, и предынкубировали при 37°C в течение 6 часов. Затем 8 мл предкультуры пересевали в казаминокислотную среду M9 (8 г/л казаминокислот, 5 г/л хлорида аммония, 0,2 г/л экстракта дрожжей, 2 мг/л гидрохлорида витамина B1, 14,5 мг/л двухводного хлорида кальция, 15,1 г/л двенадцативодного динатрийгидрофосфата, 3 г/л калийдигидрофосфата, 0,5 г/л пятиводного сульфата магния и 5 г/л глюкозы; pH 7,0), содержащей 200 мкг/мл ампициллина, и дополнительно культивировали при 37°C в течение 4 часов. При достижении измеренной при 660 нм оптической плотностью (D660) значения, равного приблизительно 0,6, в культуру добавляли 0,8 мл 1M IPTG и дополнительно инкубировали культуру в течение 14 часов при 37°C. Воздействие на клетки после культивирования, солюбилизацию белковых телец включения, повторную укладку белка и очистку белка проводили, как описано в (1).
(3) Получение мутантов с модифицированными кислыми аминокислотными остатками
Экспрессирующую MTG плазмиду pGEMMTG3 применяли в качестве матрицы для сайтспецифического мутагенеза, а соответствующую каждому мутантному ферменту синтетическую ДНК (SEQ ID NOS: 19-22 в Фигуре 6) применяли в качестве праймера. Конструирование мутантной плазмиды, культивирование, воздействие на клетки, солюбилизацию белковых телец включения, повторную укладку белка и очистку белка проводили, как описано в Примере 4 (2).
(4) Получение мутантов Sg4 и Sg7 с удаленными петлями
Для генов Sg4 вырезали гены, соответствующие остаткам с №№1-240 и 253-331, а для гена Sg7 вырезали гены, соответствующие остаткам с №№1-277 и 288-331, а затем сшивали два фрагмента, соответственно. Способы конструирования экспрессирующей Sg4 плазмиды и экспрессирующей Sg7 плазмиды описаны ниже. PCR проводили с использованием праймера (SEQ ID NO: 24), содержащего сайт расщепления Smal в 5'-конце, праймеров (SEQ ID NOS: 25 и 26), содержащих фрагмент, кодирующий 3 глициновых остатка в 3'-конце, и pGEMМТП3 в качестве матрицы. PCR проводили сходным образом с использованием праймеров (SEQ ID NOS: 27 и 28), содержащих фрагмент, кодирующий 3 глициновых остатка в 5'-конце, праймера (SEQ ID NO: 29), содержащего сайт расщепления HindIII в 3'-конце, и pGEMMTG3 в качестве матрицы. Для получения амплифицированного гена Sg4 и гена Sg7 проводили PCR с использованием праймера (SEQ ID NO: 24), содержащего сайт расщепления Smal в 5'-конце, и праймера (SEQ ID NO: 29), содержащего сайт расщепления HindIII в 3'-конце, вместе с двумя амплифицированными фрагментами ДНК в качестве матрицы. Для гена Sg7 фрагмент расщепляли Smal и HindIII и клонировали в pGEMMTG между сайтом Smal и сайтом HindIII. С другой стороны, для гена Sg7 фрагмент расщепляли EcoRI и HindIII, и клонировали в pGEMMTG между сайтом EcoRI и сайтом HindIII. Затем для окончательного конструирования экспрессирующей Sg4 плазмиды и экспрессирующей Sg7 плазмиды подтверждали нуклеотидные последовательности вставок полученных плазмид.
Культивирование, воздействие на клетки, солюбилизацию белковых телец включения, повторную укладку белка и очистку белка проводили, как описано в Примере 4 (2).
Пример 5 Сравнение субстратной специфичности
Для сравнения субстратной специфичности в качестве субстратного белка применяли овальбумин. Овальбумин содержит 12 глутаминовых остатков, и реакционная способность каждого из них была отмечена с применением ЯМР. Для применения белка в продуктах питания, лекарственных средствах и тому подобное, например, должна быть увеличена скорость образования поперечных сшивок белков. Поэтому для некоторых мутантов было проведено сравнение времени, требующегося для гелеобразования казеина в соответственных случаях.
(1) Анализ субстратной специфичности с применением ЯМР
(1-1)Сравнение между диким типом и Ser-типом
Овальбумин, 15NH4Cl и трансглутаминазу смешивали таким образом, что конечная концентрация составляла 100 мг/мл для овальбумина, 200 мМ для 15NH4Cl и 1 мкМ для трансглутаминазы. Первое 1Н-15N HSQC-определение проводили спустя 20 минут, а затем повторяли 1Н-15N HSQC-определение 13 раз с интервалом в 2 часа 15 минут.Пример полученного после взаимодействия дикого типа в течение 3 часов измерения 1Н-15N HSQC представлен на Фигуре 7. И для дикого типа, и для Ser-типа интенсивность пика сигнала (ось ординат), показанную как "а" на Фигуре 7, откладывали относительно времени взаимодействия (в часах; горизонтальная ось) (Фигура 8). В результате было показано, что глутаминовый остаток, генерирующий сигнал, обозначенный как "а" на Фигуре 7, наиболее быстро метился 15N при его взаимодействии с мутантом Ser-типа по сравнению с взаимодействием с диким типом.
(1-2) Сравнение дикого типа с мутантом S2Y·S2·PS2D
Овальбумин, 15NH4Cl и трансглутаминазу смешивали таким образом, что конечная концентрация составляла 100 мг/мл для овальбумина, 200 мМ для 15NH4Cl и 1 мкМ для трансглутаминазы. Первое 1Н-15N HSQC-определение проводили спустя 20 минут, а затем повторяли 1Н-15N HSQC-определение 13 раз с интервалом в 2 часа 15 минут. Интенсивность пика (ось ординат) спектра, полученного при взаимодействии дикого типа, S2Y или S2D с глутаминовым остатком, дающего сигнал "а" на Фигуре 7, была начерчена, соответственно (Фигура 9). В результате было показано, что для глутаминового остатка, генерирующего сигнал, обозначенный как "а" на Фигуре 7, порядок скорости мечения 15N представлял собой вид S2Y<S2D<дикий тип<S2R.
(1-3) Сравнение с мутантами, содержащими модифицированные кислые аминокислотные остатки
Овальбумин, 15NH4Cl и трансглутаминазу смешивали таким образом, что конечная концентрация составляла 100 мг/мл для овальбумина, 200 мМ для 15NH4Cl и 1 мкМ для трансглутаминазы. Первое 1Н-15N HSQC-определение проводили спустя 20 минут, а затем повторяли 1Н-15N HSQC-определение 13 раз с интервалом в 2 часа 15 минут. Интенсивность пика (ось ординат) спектра, полученного при взаимодействии дикого типа, Ser-тип, S2R, del 1-2 или del 1-3 с глутаминовым остатком, дающего сигнал "а" на Фигуре 7, была начерчена (Фигура 8), соответственно, таким же образом, как и в Примере 5. В результате было показано, что реакционная способность и del 1-2, и del 1-3 была усилена по сравнению с диким типом. Более того, следует понимать, что реакционная способность del 1-3 была во много раз выше по сравнению с мутантом Ser-типа или S2R.
Поэтому образование поперечных сшивок между белками может быть получено с использованием меньшего количества трансглутаминазы по сравнению с используемым в настоящее время количеством путем создания обладающей модифицированной субстратной специфичностью мутантной трансглутаминазы, путем снижения отрицательной заряженности кислых аминокислот их модификацией или путем преобразования остатков аминокислот, прилегающих к остаткам данных кислых аминокислот, в основные, что приводит к значительному сокращению расходов.
(1-4) Сравнение с мутантами, в которых удален участок петли
Овальбумин, 15NH4Cl и трансглутаминазу смешивали таким образом, что конечная концентрация составляла 100 мг/мл для овальбумина, 200 мМ для 15NH4Cl и 1 мкМ для трансглутаминазы. Спустя 6 часовпроводили 1Н-15N HSQC-определение и в случае применения Sg4 и Sg7 сигнал был обнаружен только у 3 групп и 2 групп соответственно в то время как в случае применения дикого типа сигнал был обнаружен у 6 групп (Фигура 10). Из экспериментов без участия трансглутаминазы известно, что среди наблюдаемых на Фигуре 10 сигналов непронумерованные не относятся к глутаминовым остаткам, помеченным трансглутаминазной реакцией. Для спектра, помеченного с применением Sg7 овальбумина, представлена фигура, полученная при низкой концентрации, так как для снижения метящей эффективности реакционная способность Sg7 была снижена. Следует отметить, что по этой причине непронумерованные сигналы были слегка подчеркнуты.
При использовании Sg4 метились только соответственные (1), (3) и (4) глутаминовые остатки, в то время как диким типом метились глутаминовые остатки, соответственные (1)-(6). Поэтому было показано, что при использовании Sg4 количество глутаминовых остатков, которые могли бы служить субстратом, снижалось, а именно их субстратная специфичность была выше по сравнению со случаем использования дикого типа. Также было показано, что при использовании Sg7 субстратом могут служить только глутаминовые остатки, соответственные (1) и (3), хотя реакционная способность была снижена. Был сделан вывод, что область связывания субстрата была уменьшена путем удаления петли, и что количество типов субстратов, с которыми может взаимодействовать мутант, было сокращено. В результате было высказано предположение, что субстратная специфичность MTG может быть модифицирована путем получения ее мутантов, основанных на пространственной структуре. Путем использования обладающей модифицированной субстратной специфичностью трансглутаминазы возможно ее применение по отношению к объектам, отличных от описанных ранее, и ожидается новое развитие в отраслях промышленности.
(2) Сравнение времени гелеобразования желатина
57,6 мкг дикого типа, мутанта Ser-типа, S2Y, S2R или S2D соответственно добавляли к 2,5 г 8% (по массе) раствору казеина, и в каждом случае проводили сравнение времени, требуемого для гелеобразования желатина. Температура реакции равнялась 40°C. Требуемое для гелеобразования время составляло 90 минут для дикого типа, 85 минут для мутанта Ser-типа, 100 минут для мутанта S2Y, 85 минут для мутанта S2R и 95 минут для мутанта S2D. Таким образом, было показано, что гелеобразование происходит за короткий промежуток времени для мутанта Ser-типа или мутанта S2R, а для мутантов S2Y и S2D гелеобразование происходит медленно.
В соответствие с настоящим изобретением облегчен процесс усовершенствования MTG на основе пространственной структуры. Конкретно с помощью настоящего изобретения может быть получена трансглутаминаза, обладающая усиленной реакционной способностью по отношению к субстрату. Кроме того, с применением трансглутаминазы, обладающей усиленной реакционной способностью по отношению к субстрату, могут быть получены новые продукты и новые методики.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПРОДУЦИРОВАНИЯ МИКРОБНОЙ ТРАНСГЛУТАМИНАЗЫ | 2004 |
|
RU2316594C2 |
| СПОСОБ СЕКРЕТОРНОЙ ПРОДУКЦИИ БЕЛКА | 2002 |
|
RU2264463C2 |
| ГЕН ФОСФАТАЗЫ ФОСФАТИДНОЙ КИСЛОТЫ | 2012 |
|
RU2625025C2 |
| ГЕНЕТИЧЕСКИ СКОНСТРУИРОВАННАЯ ЗАВИСИМАЯ ОТ ПИРРОЛОХИНОЛИНХИНОНА ГЛЮКОЗОДЕГИДРОГЕНАЗА, СОДЕРЖАЩАЯ ИНСЕРЦИЮ АМИНОКИСЛОТЫ | 2005 |
|
RU2336306C1 |
| Рекомбинантный продуцент омега-амидазы человека Nit2 | 2021 |
|
RU2778559C1 |
| УЛУЧШЕННАЯ НИТРИЛГИДРАТАЗА | 2015 |
|
RU2689606C2 |
| ДОМЕН СВЯЗЫВАНИЯ БАКТЕРИАЛЬНОЙ АТР-СИНТАЗЫ | 2005 |
|
RU2418001C2 |
| РЕКОМБИНАНТНЫЕ АТТЕНУИРОВАННЫЕ МИКРООРГАНИЗМЫ CLOSTRIDIUM И ВАКЦИНА | 2007 |
|
RU2445364C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ СЕМЕЙСТВА ГЛУТАМАТА С ИСПОЛЬЗОВАНИЕМ КОРИНЕФОРМНОЙ БАКТЕРИИ | 2011 |
|
RU2496867C2 |
| КОНЪЮГАТ АНТИТЕЛА И ЛЕКАРСТВЕННОГО СРЕДСТВА, КОМПОЗИЦИИ НА ЕГО ОСНОВЕ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2015 |
|
RU2728235C2 |

Настоящее изобретение относится к биотехнологии, а именно к способу конструирования и получения мутантных трансглутаминаз на основе пространственной структуры трансглутаминазы, полученной из Streptoverticillium mobaraense (MTG), и полученной таким образом мутантной MTG. Настоящее изобретение относится также к способу модификации MTG на основе пространственной структуры и трансглутаминазе, обладающей усиленной этим способом реакционной способностью по отношению к субстрату. Данное изобретение позволяет предсказать сайт связывания MTG с субстратом на основе пространственной структуры, определенной с помощью рентгеноструктурного анализа кристаллов MTG, и сконструировать мутантные трансглутаминазы путем замены, вставки или делеции аминокислотных остатков, расположенных в субстратсвязывающем сайте трансглутаминазы. 5 з. п.ф-лы, 60 ил.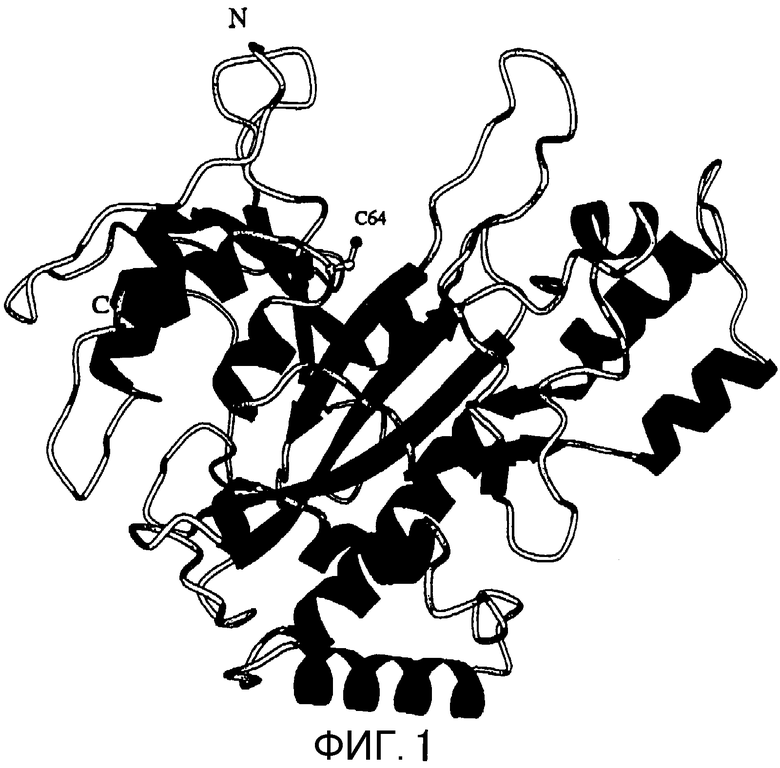
| Регулируемый подшипник | 1940 |
|
SU59654A1 |
| ПОЛИПЕПТИД, ОБЛАДАЮЩИЙ УРАТОКСИДАЗНОЙ АКТИВНОСТЬЮ, ФРАГМЕНТ ДНК, ОБЕСПЕЧИВАЮЩИЙ ЭКСПРЕССИЮ ПОЛИПЕПТИДА С УРАТОКСИДАЗНОЙ АКТИВНОСТЬЮ (ВАРИАНТЫ), ВЕКТОР ЭКСПРЕССИИ, СОДЕРЖАЩИЙ ФРАГМЕНТ ДНК, ОБЕСПЕЧИВАЮЩИЙ ЭКСПРЕССИЮ ПОЛИПЕПТИДА С УРАТОКСИДАЗНОЙ АКТИВНОСТЬЮ (ВАРИАНТЫ), ШТАММ, ЭКСПРЕССИРУЮЩИЙ ПОЛИПЕПТИД, ОБЛАДАЮЩИЙ УРАТОКСИДАЗНОЙ АКТИВНОСТЬЮ (ВАРИАНТЫ), СПОСОБ ПОЛУЧЕНИЯ ПОЛИПЕПТИДА, ОБЛАДАЮЩЕГО УРАТОКСИДАЗНОЙ АКТИВНОСТЬЮ | 1991 |
|
RU2105812C1 |

