ёОбласть техники, к которой относится изобретение
Изобретение относится к новой протеазе, которая эффективно отщепляет проструктурную часть протрансглутаминазы с преобразованием ее в активную форму трансглутаминазы, и к кодирующей ее нуклеиновой кислоте, причем указанная протрансглутаминаза продуцируется актиномицетами.
Настоящее изобретение также относится к способу продуцирования микробной трансглутаминазы в ее активной форме с использованием указанной протеазы. Кроме того, настоящее изобретение относится к способу продуцирования нейтральной металлопротеазы.
Уровень техники изобретения
Трансглутаминаза представляет собой фермент, который катализирует реакцию ацильного переноса γ-карбоксиламидных групп на пептидную цепь белка. Когда данный фермент взаимодействует с белком, могут происходить образование перекрестной связи ε-γ-Glu)-Lys и замена Gln на Glu путем дезамидирования. Трансглутаминазы используют для производства гелеобразных пищевых продуктов, таких как желе, йогурт, сыр или гелеобразных косметических средств и других, для улучшения качества мяса и т.д. (публикация заявки на выдачу патента Японии № 1-50382, прошедшая экспертизу) (JP-Kokoku). Более того, трансглутаминаза представляет собой фермент, имеющий высокий уровень применения в промышленности в том плане, что он используется для производства материалов для термостабильных микрокапсул, носителя для иммобилизованных ферментов и т.д.
Ранее были известны трансглутаминазы из животных, которые являются зависимыми от кальция в плане проявления своей активности, и трансглутаминазы из микроорганизмов (микробная(-е) трансглутаминаза(-ы)), которая(-е) здесь и далее также указываются как «MTG»), которые являются не зависимыми от кальция в плане проявления своей активности. В ряду MTG была обнаружена трансглутаминаза из бактерии, принадлежащей к роду Streptoverticillium. Такие бактерии Streptoverticillium включают в себя, например, Streptoverticillium griseocarneum IFO 12776, Streptoverticillium cinnamoneum subsp. cinnamoneum IFO 12852, Streptoverticillium mobaraense (здесь и далее может быть сокращенно обозначена как S. mobaraense) IFO 13819 и др. (публикация неоцененной заявки на выдачу патента Японии (JP-Kokai) № 64-27471).
Поскольку данные трансглутаминазы, однако, продуцировались путем очистки из культур, таких как культуры описанных выше микроорганизмов, имели место проблемы в плане количества и эффективности, и тому подобного. Затем, в качестве способа эффективной секреции гетерологичных белков, был разработан способ, в котором в качестве хозяина выбрана коринеформная бактерия, где белок слияния, содержащий трансглутаминазу, присоединяли ниже домена сигнального пептида коринеформной бактерии, и трансглутаминаза эффективно секретировалась с получением ее высокого выхода (WO 01/23591). В данном исследовании описан способ, в котором MTG секретируется в неактивной форме как протрансглутаминаза (здесь и далее указана как «про-MTG»), в которой проструктурная часть присоединена к MTG, и затем проструктурная часть про-MTG отщепляется протеазой с преобразованием ее в трансглутаминазу, имеющую активность, а также способ, в котором активная трансглутаминаза продуцируется непосредственно в культуральную среду в требуемом и достаточном количестве путем совместной экспрессии с SAM-P45, представляющей собой сериновую протеазу, происходящую из актиномицетов, в коринеформной бактерии, продуцирующей про-MTG.
Хотя способ, в котором активная трансглутаминаза непосредственно продуцируется путем совместной экспрессии про-MTG и протеазы, которая позволяет создать проструктурную часть про-MTG в коринеформной бактерии, считается исключительно эффективным способом продуцирования трансглутаминазы, субстратная специфичность SAM-P45 не так строга, и она может расщеплять и приводить к деградации не только проструктурной части про-MTG, но также в некоторой степени и самой трансглутаминазы, поэтому обращение с SAM-P45 может быть непростым. В случаях, когда используется SAM-P45, способ продуцирования трансглутаминазы должен быть строго контролируемым, иначе не будет происходить расщепления продуцируемой трансглутаминазы в культуральной среде.
Таким образом, еще оставалась потребность в протеазе, которая может избирательно отщеплять только проструктурную часть про-MTG и вызывать как можно меньшее избыточное расщепление самой трансглутаминазы для осуществления преимущественного продуцирования активной формы трансглутаминазы.
Известным ферментом, который отщепляет проструктурную часть про-MTG, кроме SAM-P45, является диспаза из Bacillus polymyxa (Eur. J. Biochem., vol. 257, p. 570-576 (1998)). Однако, для отщепления проструктурной части требуется большое количество фермента, и имеется риск избыточного расщепления самой трансглутаминазы. Кроме того, диспаза представляет собой реагент для культуры клеток, поскольку для индустриального применения данный фермент является дорогим.
Сущность изобретения
Еще остается необходимость в протеазе, которая может избирательно отщеплять только проструктурную часть про-MTG, и вызывать как можно меньшее избыточное расщепление самой трансглутаминазы для осуществления преимущественного продуцирования активной формы трансглутаминазы, как указывалось выше. Кроме того, если могли использоваться протеазы, способные избирательно отщеплять только проструктурную часть про-MTG и вызывать как можно меньшее избыточное расщепление самой трансглутаминазы, их полагали предпочтительными для продуцирования активной трансглутаминазы. Более того, если протеазы, подходящие для продуцирования трансглутаминазы, которые могут избирательно отщеплять проструктурную часть про-MTG, могли секретироваться из клетки, полагали, что они являются более предпочтительными, поскольку активная трансглутаминаза могла непосредственно продуцироваться в культуральную среду за счет совместной экспрессии протеаз с про-MTG.
Таким образом, целью настоящего изобретения является предоставление протеазы, которая может использоваться для продуцирования трансглутаминазы и которая избирательно отщепляет проструктурную часть про-MTG.
В частности, целью изобретения является предоставление протеазы, которая избирательно отщепляет проструктурную часть про-MTG, где указанная протеаза может продуцироваться за счет использования коринеформной бактерии в качестве хозяина и может секретироваться из клетки.
Целью изобретения также является предоставление молекулы нуклеиновой кислоты, кодирующей указанную протеазу.
Другой целью изобретения является предоставление способа эффективного продуцирования MTG с использованием указанной протеазы.
Более того, целью изобретения является предоставление способа продуцирования указанной протеазы.
Авторами настоящего изобретения найдена протеаза, которая будет избирательно отщеплять проструктурную часть про-MTG, но будет вызывать как можно меньшее избыточное расщепление самой трансглутаминазы, и затем они смогли выделить и очистить нейтральную металлопротеазу, имеющую такое свойство. Авторами изобретения также получена ДНК, кодирующая указанную протеазу, которую вводили в коринеформную бактерию и затем успешно осуществляли ее секреторную экспрессию с использованием коринеформной бактерии в качестве хозяина. Кроме того, настоящий фермент действительно взаимодействовал с про-MTG с расщеплением проструктурной части и затем получали активную трансглутаминазу. Авторы изобретения осуществляли настоящее изобретение путем идентификации нейтральных металлопротеаз, происходящих из микроорганизмов из других источников, имеющих эквивалентную функцию, которые, как было подтверждено, также могут использоваться в качестве активных MTG.
То есть настоящее изобретение относится к нейтральной металлопротеазе из актиномицетов, имеющей высокую избирательность при отщеплении проструктурной части про-MTG, и к кодирующей ее молекуле нуклеиновой кислоты.
Настоящее изобретение также относится к способу продуцирования активной MTG, включающему отщепление проструктурной части про-MTG нейтральной металлопротеазой.
Настоящее изобретение также относится к способу продуцирования указанной металлопротеазы, включающему введение молекулы нуклеиновой кислоты, кодирующей указанную нейтральную металлопротеазу в коринеформную бактерию, культивирование коринеформной бактерии, в которую была введена указанная молекула нуклеиновой кислоты, с обеспечением за счет этого экспрессии указанной нейтральной металлопротеазы и получение указанной металлопротеазы, которая секретируется из клетки.
Более конкретно, настоящее изобретение относится к нейтральной металлопротеазе SVP35 из актиномицетов, имеющей следующие свойства:
1) Молекулярная масса: около 35000 (измерена путем SDS-PAGE)
2) Оптимальное значение pH: 6,0-8,0, более конкретно 6,5-7,5, в частности, примерно 7,0
3) pH-стабильность: pH 4-10
4) Оптимальная температура: около 45°C
5) Температурная стабильность: стабильна при температуре ниже, чем примерно 50°C
6) Она сильно ингибируется этилендиаминотетрауксусной кислотой, 1,10-фенантролином и фосфорамидоном, которые являются ингибиторами металлопротеаз, и ингибитором субтилизина Streptomyces (SSI) из актиномицетов.
Настоящее изобретение также относится к нейтральной металлопротеазе SVP70, имеющей следующие свойства:
1) Молекулярная масса: около 71000 (измерена путем SDS-PAGE)
2) Оптимальное значение pH: 6,0-8,0, более конкретно 6,5-7,5, в частности, примерно 7,0
3) pH-стабильность: pH 5-10
4) Оптимальная температура: интервал около 50°C-55°С, в частности, примерно 55°С
5) Она испытывает сильное ингибирующее действие этилендиаминотетрауксусной кислоты, 1,10-фенантролина и фосфорамидона, которые являются ингибиторами металлопротеаз, дитиотреитола, который является SH-восстановителем, и ингибитора субтилизина Streptomyces (SSI), происходящего из актиномицетов.
Настоящее изобретение также относится к молекуле нуклеиновой кислоты, кодирующей указанную SVP35 или SVP70.
Настоящее изобретение также относится к способу продуцирования активной MTG, включающему отщепление проструктурной части про-MTG указанной SVP35 или SVP70.
Более того, настоящее изобретение относится к способу продуцирования SVP35 или SVP70, включающему введение молекулы нуклеиновой кислоты, кодирующей указанную SVP35 или SVP70 в коринеформную бактерию, культивирование коринеформной бактерии, в которую введена указанная молекула нуклеиновой кислоты, и получение SVP35 или SVP70, секретируемых из клеток.
Краткое описание чертежей
Фиг.1 представляет собой график, на котором показана зависимость активности SVP35 и SVP70 от рН.
Фиг.2 представляет собой график, на котором показана pH-стабильность SVP35 и SVP70.
Фиг.3 представляет собой график, на котором показана зависимость активности SVP35 и SVP70 от температуры.
Фиг.4 представляет собой график, на котором показана температурная стабильность SVP35.
На фиг.5 отображены ингибирующая активность различных соединений в отношении активности SVP35 и SVP70.
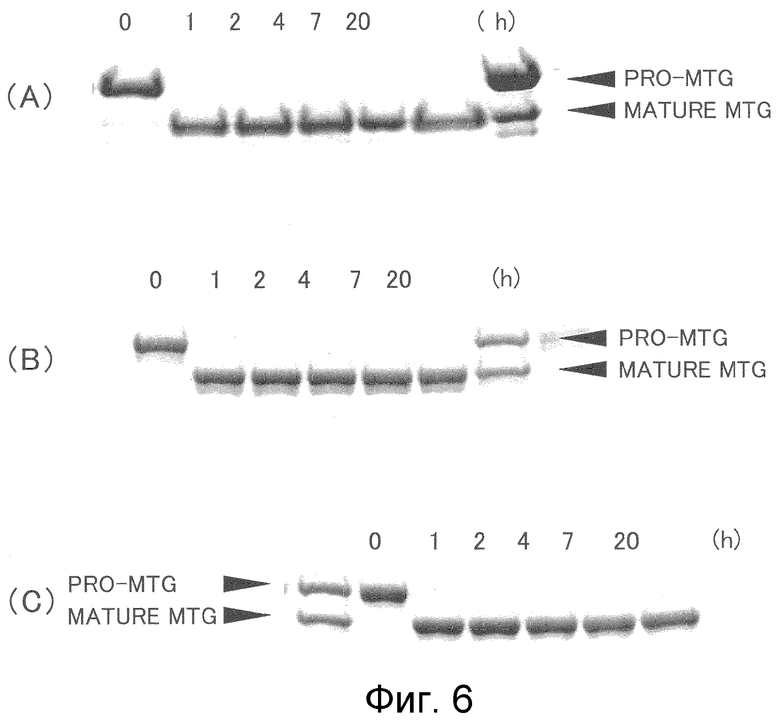
На фиг.6 отображены последовательные изменения преобразования про-MTG в активную форму MTG, осуществляемые за счет SVP35 и SVP70, в виде относительного изменения количества белка.
Фиг.7 (A) и (B) представляют собой графики, на которых изображено изменение активности трансглутаминазы во времени при взаимодействии про-MTG с SVP70 и SAM-P45, соответственно. (A): добавление SVP70,  : добавленное количество 1/200 относительно субстрата,
: добавленное количество 1/200 относительно субстрата,  : добавленное количество 1/500 относительно субстрата; (B): добавление SAM-P45,
: добавленное количество 1/500 относительно субстрата; (B): добавление SAM-P45,  : добавленное количество 1/10 относительно субстрата,
: добавленное количество 1/10 относительно субстрата,  : добавленное количество 1/50 относительно субстрата.
: добавленное количество 1/50 относительно субстрата.
Фиг.8 (A) и (B) представляют собой графики, на которых отображено изменение количества белка MTG во времени при взаимодействии про-MTG с SVP70 и SAM-P45, соответственно. (A): добавление SVP70, : добавленное количество 1/200 относительно субстрата, : добавленное количество 1/500 относительно субстрата; (B): добавление SAM-P45, : добавленное количество 1/10 относительно субстрата, : добавленное количество 1/50 относительно субстрата.
Описание предпочтительных вариантов осуществления
В общем известно, что секреторный белок транслируется в виде препептида или препропептида, и после этого его сигнальный пептид («пре-часть») отщепляется, приводя к преобразованию в зрелый пептид или пропептид, пропептид расщепляется в домене, называемом проструктурой, становясь зрелым пептидом. При использовании здесь данного термина проструктурная часть секреторного белка может просто называться «проструктурой». Кроме того, используемая здесь «сигнальная последовательность» относится к последовательности, которая расположена с N-конца предшественника секреторного белка и она не присутствует во встречающемся в природе зрелом белке, и «сигнальный пептид» относится к пептиду, который отщепляется от такого белка-предшественника. В общем сигнальная последовательность отщепляется протеазой после секреции из клетки.
При использовании здесь данного термина белок, который не содержит сигнальный пептид, но содержит проструктурную часть может называться «пробелок», например «протрансглутаминаза» или «про-MTG». При использовании здесь данного термина проструктурная часть секреторного белка может просто называться «проструктурой» или «проструктурной частью», и данные термины могут использоваться здесь взаимозаменяемо.
Среди протеаз, которые, как считается, могут легко экспрессироваться в коринеформной бактерии, авторы изобретения вначале провели поиск протеазы, имеющей высокую специфичность и избирательность в отношении интересующего субстрата, т.е. протеазы, которая избирательно отщепляет проструктурную часть про-MTG и вызывает как можно меньшее избыточное расщепление самой трансглутаминазы.
Когда MTG секретируется актиномицетами внеклеточно, полагают, что она вначале секретируется в виде про-MTG, с последующим отщеплением проструктурной части про-MTG, что приводит к образованию активной формы MTG (Eur. J. Biochem., vol. 257, p. 570-576 (1998)). В соответствии с этим авторы изобретения ожидали, что в актиномицетах, продуцирующих MTG, существует протеаза, которая отщепляет проструктурную часть про-MTG. Поскольку данная протеаза исходно представляет собой протеазу, которая отщепляет проструктурную часть, ожидается, что данная протеаза имеет высокую избирательность в отношении субстратов и расщепляет только проструктурную часть, при этом она действует на саму MTG в меньшей степени.
Кроме того, структурный ген про-MTG актиномицетов и структурный ген протеазы SAM-P45 могут эффективно экспрессироваться в коринеформной бактерии, и они могут секретироваться внеклеточно. Основываясь на данной информации, было выполнено значимое исследование, направленное на поиск интересующей протеазы из продуцирующей MTG бактерии, которая представляет собой актиномицет, и в результате было выявлено, что продуцирующий MTG штамм Streptoverticillium mobaraense характеризуется высокой избирательностью расщепление проструктурной части про-MTG и продуцирует новые нейтральные металлопротеазы, которые могут использоваться для продуцирования активной MTG. Авторами настоящего изобретения выделены и очищены данные нейтральные металлопротеазы и продемонстрированы их энзимологические характеристики. Более того, авторами изобретения определены аминокислотные последовательности N-концевых частей данных металлопротеаз и получены гены, кодирующие данные металлопротеазы.
Кроме того, авторы изобретения вводили ген данного фермента в коринеформной бактерии, обеспечивая его экспрессию в системе, где применяется коринеформная бактерия в качестве хозяина, и в результате фермент секреции. Более того, фермент действительно взаимодейстовал на про-MTG с проструктурной частью, что приводило к отщеплению проструктурной части с выходом активной трансглутаминазы. Также обнаружили нейтральные металлопротеиназы из микроорганизмов других источников, имеющие сходные функции, которые, как было выявлено, также могут использоваться для продуцирования активной формы MTG.
Здесь и далее будут иллюстрироваться более конкретные варианты осуществления настоящего изобретения.
Нейтральные металлопротеазы по настоящему изобретению могут быть получены с поверхностей культивируемых актиномицетов или из культуральной надосадочной жидкости актиномицетов, включая Streptoverticillium mobaraense, Streptomyces griseus, Streptomyces coelicolor и т.д.
В следующей части вначале описываются впервые обнаруженные нейтральные металлопротеазы Streptoverticillium mobaraense IFO13819.
Культивирование бактерии для получения нейтральной металлопротеазы по настоящему изобретению, например актиномицета, описанного выше, может проводиться по способам, обычно применяемым для культивирования актиномицетов. То есть в качестве среды для культуры может использоваться обычная среда, содержащая обычные источники углерода, источники азота, неорганические ионы и др. Глюкоза, крахмал и другое может использоваться в качестве источников углерода. Пептон, дрожжевой экстракт, мясной экстракт, солодовый экстракт, соль аммония и другое может необязательно использоваться в качестве источника азота, если это необходимо. Культивирование может проводиться в аэробных условиях, которые надлежащим образом контролируются в пределах интервала рН от pH 5,0 до 8,5 и в температурном интервале от 15 до 37°C. Для продуцирования нейтральных металлопротеаз по настоящему изобретению культивирование предпочтительно продлевают до достижения максимального количества требуемой нейтральной металлопротеазы, и затем его можно прекратить. Хотя подходящий период культивирования зависит от температуры, pH и типа среды, обычно данный период предпочтительно составляет примерно от 1 до 12 суток. После периода культивирования культуру можно разделить на клетки и культуральную надосадочную жидкость путем центрифугирования или подобного способа.
Новые нейтральные металлопротеазы по настоящему изобретению могут быть получены из культуральной надосадочной жидкости, а также из выделенных клеток, в частности с поверхности клеток. Для очистки фермента могут адаптироваться любые способы, которые обычно применяют для очистки фермента, например способ высаливания сульфатом аммония, способ гель-фильтрации, ионообменная хроматография, гидрофобная хроматография и тому подобное, протеаза может быть очищена более эффективно с использованием высокоэффективной жидкостной хроматографии (ВЭЖХ) и т.д. Измерение ферментативной активности нейтральной металлопротеазы, полученной таким путем, может осуществляться путем взаимодействия фермента с пептидом, который содержит область, связывающую про-часть протрансглутаминазы и зрелую трансглутаминазу, например, с синтетическим пептидом Glu-Pro-Ser-Phe-Arg-Ala-Pro-Asp-Ser (SEQ ID NO: 11) (Peptide Institute), в качестве субстрата, и расчета снижения количества субстрата.
Как указано выше, нейтральная металлопротеаза по изобретению, очищенная из выделенных клеток, в частности с поверхности клеток, или из надосадочной жидкости культуры, может анализироваться на предмет N-концевой аминокислотной последовательности на газофазовом аминокислотном секвенаторе для определения частичной аминокислотной последовательности. Более того, могут оцениваться ферментативные свойства (оптимальный pH, pH-стабильность, оптимальная температура, действие ингибитора и т.д.) выделенной и очищенной нейтральной металлопротеазы.
В одном из вариантов осуществления настоящего изобретения нейтральную металлопротеазу, названную SVP35, получали с поверхности клеток Streptoverticillium mobaraense, а нейтральную металлопротеазу, названную SVP70, можно получить из культуральной надосадочной жидкости Streptoverticillium mobaraense.
В одном из осуществлений настоящего изобретения нейтральная металлопротеаза по изобретению представляет собой нейтральную металлопротеазу SVP35, имеющую следующие свойства:
1) Молекулярная масса: около 35000 (измерена путем SDS-PAGE)
2) Оптимальное значение pH: 6,0-8,0, более конкретно 6,5-7,5, в частности, примерно 7,0
3) pH-стабильность: pH 4-10
4) Оптимальная температура: около 45°C
5) Температурная стабильность: стабильна при температуре ниже чем примерно 50°C
6) Ингибиторы: она сильно ингибируется этилендиаминотетрауксусной кислотой, 1,10-фенантролином и фосфорамидоном, которые являются ингибиторами металлопротеаз, и ингибитором субтилизина Streptomyces (SSI), происходящим из актиномицетов.
В другом осуществлении настоящего изобретения нейтральная металлопротеаза по изобретению представляет собой нейтральную металлопротеазу SVP70, имеющую следующие свойства:
1) Молекулярная масса: около 71000 (измерена путем SDS-PAGE)
2) Оптимальное значение pH: 6,0-8,0, более конкретно 6,5-7,5, в частности, примерно 7,0
3) pH-стабильность: pH 5-10
4) Оптимальная температура: интервал около 50°C-55°С, в частности, примерно 55°С
5) Ингибиторы: она сильно ингибируется этилендиаминотетрауксусной кислотой, 1,10-фенантролином и фосфорамидоном, которые являются ингибиторами металлопротеаз, дитиотреитолом, который является SH-восстановителем, и ингибитором субтилизина Streptomyces (SSI), происходящего из актиномицетов.
Когда SVP35 или SVP70 взаимодействует с про-MTG, обе они характеризуются высокой избирательной активностью расщепления в отношении проструктурной части MTG. То есть, поскольку оба фермента характеризуются эффективным преобразованием про-MTG в активную MTG, в то время как активность в отношении деградации, полученной в результате активной MTG, является низкой, оба они являются подходящими ферментами для продуцирования активной MTG с использованием про-MTG в качестве сырого материала. N-концевые аминокислотные последовательности двух новых нейтральных металлопротеаз показаны в SEQ ID NO: 1 для SVP35, и в SEQ ID NO: 2 для SVP70, что выявляет гомологию между данными последовательностями. Поэтому проводился поиск белков, имеющих гомологию в отношении данных протеаз в их N-концевых аминокислотных последовательностей, обнаружили металлопротеазу SGMP II (J. Biochem. Vol. 110, p. 339-344 (1991)) из Streptomyces griseus, а также обнаружили три металлопротеазы (CAB76000, CAB76001, CAB69762 GenBank/EMBL/DDBJ) из Streptomyces coelicolor и т. п. Данные протеазы могут также использоваться тем же способом, что и SVP35 и SVP70, для избирательного расщепления проструктурной части про-MTG, и они могут использоваться для продуцирования активной MTG с использованием про-MTG а качестве сырого материала.
Далее описан способ продуцирования нейтральной металлопротеазы по настоящему изобретению по технологии рекомбинантной ДНК.
Известны некоторые примеры продуцирования полезных белков, включающих в себя ферменты, физиологически активные вещества и тому подобное, с использованием технологии рекомбинантной ДНК. Преимущество применения способа рекомбинантной ДНК представляет собой возможность массового продуцирования полезных белков, которые в природе встречаются в минимальном количестве.
Для продуцирования нейтральной металлопротеазы по настоящему изобретению, с использованием технологии рекомбинантной ДНК, вначале получали генетическую конструкцию, которая содержит промотор, последовательность, кодирующую надлежащий сигнальный пептид, фрагмент нуклеиновой кислоты, кодирующий нейтральную металлопротеазу по изобретению, и регуляторную последовательность (оператор или терминатор, и т.д.), которая необходима для экспрессии гена нейтральной металлопротеазы в коринеформной бактерии, в надлежащем положении, так что они могли бы функционировать. Нейтральная металлопротеаза по изобретению может иметь проструктурную часть на N-конце. Векторы, которые могут использоваться для данной конструкции, конкретно не ограничены и включают в себя любой вектор, который может функционировать в коринеформной бактерии, и они могут автономно реплицироваться, как плазмиды, или интегрироваться в хромосому бактерии. Когда в качестве хозяина используется коринеформная бактерия, в качестве вектора особенно предпочтительны плазмиды, происходящие из коринеформных бактерий. Они включают в себя, например, pHM1519 (Agric. Biol. Chem., 48, 2901-2903 (1984))) pAM330 (Agric. Biol. Chem., 48, 2901-2903 (1984), и модифицированные плазмиды, которые обладают генами лекарственной резистентности.
Примеры Corynebacterium, которые могут использоваться в качестве бактерий-хозяев по настоящему изобретению, включают в себя мутантные штаммы, происходящие из штаммов дикого типа, включающих в себя Brevibacterium saccharolyticum ATCC14066, Brevibacterium immariophilum ATCC14068, Brevibacterium lactofermentum (Corynebacterium glutamicum) ATCC13869, Brevibacterium roseum ATCC13825, Brevibacterium flavum (Corynebacterium glutamicum) ATCC14067, Corynebacterium acetoacidophilum ATCC13870, Corynebacterium glutamicum ATCC13032, Corynebacterium lilium (Corynebacterium glutamicum) ATCC15990, Brevibacterium ammoniagenes (Corynebacterium ammoniagenes) ATCC6871 и тому подобное, или мутантные штаммы, происходящие от мутантных штаммов данных диких типов.
Мутантные штаммы, которые используются по настоящему изобретению, включают в себя, например, мутантные штаммы, дефективные по способности продуцировать глутамат, штаммы, мутантные по продуцированию аминокислот, таких как лизин и тому подобное, и мутантные штаммы, модифицированные в плане продуцирования других веществ, таких как нуклеиновые кислоты, например инозин. Такие мутантные штаммы могут быть получены путем обработки ультрафиолетовым облучением или химическим мутагеном, таким как N-метил-N'-нитрозогуанидин и тому подобное, и затем выбора штаммов с увеличенной способностью к продуцированию-секреции белков.
В частности, Corynebacterium glutamicum AJ1203 (FERM BP-734) (исходное размещение в банке 26 марта 1984 г.) (в настоящее время National Institute of Advanced Industrial Science and Technology, International Patent Organism Depositary, Tsukuba Central 6, 1-1, Higashi 1-Chome, Tsukuba-shi, Ibaraki-ken, 305-8566, Япония), которые выделяли из Corynebacterium glutamicum (C. glutamicum) дикого типа ATCC13869, в виде мутантного штамма, устойчивого к стрептомицину, как ожидается, имеет мутацию в функциональном гене, ассоциированном с секрецией белков, и его способность секретировать-продуцировать гетерологичные белки исключительно высока, составляя по накопленному количеству в 2 или в 3 раза больше по сравнению с родительским штаммом (штаммом дикого типа), в оптимальных условиях культивирования, так что он подходит в качестве бактерии хозяина (см. WO 02/081694). Кроме того, предпочтительно использовать в качестве хозяина штамм, который получали путем модификации данного штамма, так что данный штамм не будет более продуцировать белок клеточной поверхности, поскольку очистка гетерологичных белков, секретируемых в среду, станет проще, что особенно предпочтительно. Такая модификация может проводиться путем введения мутации в ген белка клеточной поверхности на хромосоме или в область регуляции его экспрессии путем мутагенеза или способов рекомбинации генов.
Примеры промоторов из коринеформных бактерий включают в себя промоторы генов белков клеточной поверхности PS1, PS2, и SlpA, промоторы генов биосинтетических систем различных аминокислот, например гена глутаминсинтетазы, гена аспартокиназы в системе биосинтеза лизина, и т.п.
Сигнальный пептид, который применяется по настоящему изобретению, представляет собой сигнальный пептид секреторного белка из коринеформной бактерии, хозяина, и предпочтительно представляет собой сигнальный пептид белка клеточной поверхности из коринеформной бактерии. Белки клеточной поверхности коринеформной бактерии включают в себя PS1 и PS2 из C. glutamicum (JP-Kokai № 6-502548) и SlpA из C. ammoniagenes (JP-Kokai № 10-108675).
Для продуцирования нейтральной металлопротеазы с сильной активностью в плане избирательного расщепления проструктуры про-MTG с использованием технологии рекомбинантной ДНК требуется ДНК, кодирующая такую нейтральную металлопротеиназу.
В одном из осуществлений настоящего изобретения нейтральную металлопротеазу SVP35 продуцируют с использованием технологии рекомбинантной ДНК. ДНК, кодирующая SVP35, может быть получена следующим образом.
Вначале определяют аминокислотную последовательность очищенной SVP35. Для определения такой аминокислотной последовательности может использоваться способ Эдмана (Edman, P., Acta Chem. Scand. 4, 227 (1950)). Для определения аминокислотной последовательности может использоваться газофазовый белковый секвенатор от Shimadzu Co. Ltd. Co. Ltd. и т.п.
Для нейтральной металлопротеазы SVC35 по изобретению путем секвенирования 20 аминокислотных остатков с N-конца обнаружена последовательность, показанная в SEQ ID NO: 1.
Данная информация может использоваться для синтеза подходящего праймера для ПЦР и получения зонда для получения нейтральной металлопротеиназы по настоящему изобретению. Например ген протеазы из актиномицетов, который, как ожидается, характеризуется гомологией на основе результатов поиска гомологии с N-концевой аминокислотной последовательностью, например ген металлопротеазы (GenBank/EMBL/DDBJ CAB76001) из Streptomyces coelicolor, может подвергаться ПЦР с использованием ДНК актиномицетов, полученной по способу Saito and Miura [Biochem. Biophys. Acta, 72, 619 (1963)], в качестве матрицы, для амплификации фрагмента гена, кодирующего данную протеазу. Амплифицированный фрагмент может использоваться в качестве зонда.
Затем ДНК актиномицетов, полученную способом Saito and Miura, например хромосомную ДНК Streptoverticillium mobaraense IFO13819, расщепляют различными подходящими ферментами рестрикции, например различными ферментами рестрикции, которые распознают последовательности из 6 оснований. Расщепленную хромосомную ДНК актиномицетов можно анализировать способами, хорошо известными специалистам в данной области, например способом блот-гибридизации по Саузерну, описанным в Molecular Cloning 2nd edition [J. Sambrook E. F. Fritsch and T. Maniatis, Cold Spring Harbor Laboratory Press, p9. 31 (1989)] и т.п., с использованием меченного 32P ПЦР-продукта, полученного путем описанной выше ПЦР. Например, молекула нуклеиновой кислоты, кодирующая нейтральную металлопротеазу по настоящему изобретению или ее часть, может клонироваться путем получения фрагмента, гомологию которого в отношении использованного зонда подтверждали путем блоттинга по Саузерну, и клонирования его в подходящий вектор. Способы, необходимые для такого генного клонирования хорошо известны в данной области (см., например, J. Sambrook, E. F. Fritsch and T. Maniatis, Cold Spring Harbor Laboratory Press, p1. 90 (1989)).
В одном из вариантов осуществления настоящего изобретения ПЦР проводят с использованием хромосомной ДНК Streptomyces coelicolor A3(2) в качестве матрицы для получения зонда. Далее одну полосу размером примерно 8 тыс.п.н., которая гибридизуется с меченными 32P зондами, выявляют в продукте расщепления SphI хромосомной ДНК Streptoverticillium mobaraense IFO13819. Таким образом, хромосомную ДНК Streptoverticillium mobaraense IFO13819, полученную описанным выше способом, расщепляют с помощью SphI, фрагмент, примерно равный 8 тыс.п.н., получают посредством электрофореза в агарозном геле, полученный фрагмент вводят в участок SphI в pUC18, и затем его вводят в компетентные клетки Escherichia coli JM109 с образованием библиотеки. Интересующие клоны могут быть получены путем скрининга образованной библиотеки с использованием синтетического олигонуклеотида в качестве зонда по способам гибридизации колоний, описанным в Molecular Cloning 2nd edition (выше), и путем отбора штамма, который несет плазмиду, содержащую генный фрагмент SVP35, клонированный в плазмиду. Плазмиду, полученную из данного штамма, обозначают здесь как pVSV1. Анализируют последовательность нуклеотидов фрагмента, клонированного в pVSV1, предсказывают первичную аминокислотную структуру для подтверждения того, что данный фрагмент кодирует ранее установленную N-концевую аминокислотную последовательность. Таким образом, подтверждают, что полученный ген представляет собой ген, кодирующий SVP35.
Затем можно конструировать рекомбинантную молекулу нуклеиновой кислоты для экспрессии нейтральной металлопротеазы по настоящему изобретению путем лигирования генетической конструкции, содержащей ДНК, кодирующую полученную металлопротеазу, с подходящим вектором в зависимости от свойств используемого хозяина. Клетки коринеформной бактерии-хозяина трансформируют рекомбинантной молекулой нуклеиновой кислоты. Трансформированные клетки можно культивировать в подходящей среде с получением нейтральной металлопротеазы по настоящему изобретению, секретируемой или накапливаемой в среде и/или в клетке.
Следующим будет описан способ продуцирования активной MTG из про-MTG с использованием нейтральной металлопротеазы.
Нейтральная металлопротеаза, использованная в продуцировании активной MTG, может взаимодействовать с про-MTG в виде фракции, содержащей нейтральную металлопротеазу, полученную из культуральной среды бактерии, продуцирующей нейтральную металлопротеиназу. Также она может использоваться в виде более высокоочищенной нейтральной металлопротеазы с высокой специфичной активностью. Далее, как описаной ниже, может также использоваться нейтральная металлопротеаза, которая может быть получена путем культивирования клетки, трансформированной рекомбинантной молекулой нуклеиновой кислоты, которая может быть получена путем соединения ДНК, кодирующей нейтральной металлопротеазы, имеющей высокую избирательную активность отщепления проструктурной части про-MTG.
Про-MTG, используемая для продуцирования MTG, может быть фракцией, содержащей про-MTG, полученной из культуральной среды продуцирующей про-MTG бактерии. Также может использоваться более высокоочищенная про-MTG. Взаимодействие можно проводить в условиях, в которых количество добавленной металлопротеазы, добавленной к про-MTG, составляет от 1/10 до 1/500 по массе, в которых температура взаимодействия подобрана в интервале от 15 до 50°C и интервал pH составляет от pH 5,0 до 9.
Кроме того, генетическая конструкция, которая получена как описано выше и которая содержит ДНК, кодирующую нейтральную металлопротеазу по настоящему изобретению, может вводиться в микроорганизм, содержащий генетическую конструкцию, кодирующую про-MTG, в частности в коринеформную бактерию, для продуцирования в одной бактериальной клетке и про-MTG, и нейтральной металлопротеазы по настоящему изобретению, притом, что про-MTG может преобразовываться в зрелую MTG в указанных выше условиях. Более подробный способ эффективного продуцирования про-MTG клетках коринеформной бактерии, генетическая конструкция, используемая для такого способа, и коринеформная бактерия, в которую вводят данную генетическую конструкцию, описаны, например, в WO 01/23591. Более конкретно, например, коринеформная бактерия, которая может эффективно осуществлять внеклеточную секрецию белка про-MTG, может быть получена введением генетической конструкции в коринеформную бактерию, где генетическая конструкция получена путем соединения последовательности, содержащей последовательность, кодирующую про-MTG, которая расположена ниже (в положении даунстрим) последовательности, кодирующей домен сигнального пептида коринеформной бактерии, в частности домен сигнального пептида белка клеточной поверхности, с соответствующим промотором ниже его. Сигнальный пептид, промотор и хозяин, которые могут использоваться для данной цели, могут быть выбраны из сигнальных пептидов, промоторов и хозяев, подходящих для экспрессии нейтральной металлопротеазы по настоящему изобретению, как указано выше. Сочетание векторов, которые совместимы в одной клетке, также известно специалистам в данной области. Таким образом, зрелую MTG можно получить путем введения подходящей генетической экспрессирующей конструкции, содержащей ДНК, кодирующую нейтральную металлопротеазу по настоящему изобретению, указанную выше, в коринеформную бактерию, продуцирующую про-MTG, или наоборот, путем введения подходящей генетической экспрессирующей конструкции, кодирующей про-MTG, в коринеформную бактерию, продуцирующую нейтральную металлопротеазу по настоящему изобретению, позволяя таким путем генетическим конструкциям, которые могут экспрессировать про-MTG и нейтральную металлопротеазу по настоящему изобретению, сосуществовать в одной и той же бактерии и путем поддержания культуры в подходящих условиях, таких, что нейтральная металлопротеаза по настоящему изобретению может быть активной.
Трансглутаминаза, продуцируемая способом по настоящему способу может выделяться и очищаться из реакционной смеси по способам, хорошо известным специалистам в данной области. Например, трансглутаминаза может выделяться и очищаться путем удаления клеток из смеси центрифугированием и т.д. и затем с использованием подходящих способов, таких как высаливание, преципитация этанолом, ультрафильтрация, хроматография путем гель-фильтрации, ионообменная колоночная хроматография, аффинная хроматография, жидкостная хроматография с высоким давлением среды, обращенно-фазовая хроматография, гидрофобная хроматография или их комбинация.
Настоящее изобретение далее описано в следующих примерах, которые никоим образом не предназначаются для ограничесния настоящего изобретения.
ПРИМЕРЫ
Пример 1. Нейтральная металлопротеаза, продуцируемая Streptoverticillium mobaraense IFO13819
(1) Очистка нейтральной металлопротеазы (SVP70), продуцированной Streptoverticillium mobaraense IFO13819
В колбу Sakaguchi объемом 5 л (колба для перемешивания) помещали 800 мл культуральной среды ISP2 (4 г дрожжевого экстракта, 10 г солодового экстракта, 4 г глюкозы на литр воды, значение рН доведено до 7,3), инокулировали Streptoverticillium mobaraense IFO13819 с чашки и культивировали при встряхивании при 30°C в течение 9 суток при 120 об/мин. Культуральную среду центрифугировали и собирали надосадочную жидкость культуры. Ее фильтровали с использованием фильтра Depth (размер пор 3 мкм, Saltorius Co. Ltd.) с последующим концентрированием с использованием мембраны Sartocon Slice с размером пор 10000 Да (Saltorius Co. Ltd.). Концентрат разбавляли в 10 раз буфером Tris-HCl/5 мМ хлорид кальция (pH 7,5), наносили на колонку с DEAE-Sepharose FF (2,6 ø × 10 см, Amersham Pharmacia Co. Ltd.), уравновешенную тем же буфером, используя FPLC (Amersham Pharmacia Co. Ltd.), и элюировали с использованием линейного градиента концентрации 0-0,5 M хлорида натрия. Собирали фракцию, содержащую активный ингредиент, наносили на колонку с фенил-Sepharose HP (1,6 ø × 10 см, Amersham Pharmacia Co. Ltd.), уравновешенную раствором 1,5 M сульфат аммония/20 мМ буфер MES/5 мМ хлорид кальция (pH 6,0), элюировали с использованием линейного градиента концентрации 1,5-0 M сульфата аммония и собирали активную фракцию. Полученную в результате активную фракцию диализовали против раствора 20 мМ буфер MES/5 мМ хлорид кальция (pH 6,0) при 4°C в течение ночи с получением очищенного раствора фермента.
Измерение ферментативной активности на каждой стадии проводили следующим образом.
Раствор фермента добавляли к 20 мМ натрий-фосфатному буферу, содержащему пептид GPSFRAPDS (Peptide Institute) (SEQ ID NO: 11) с получением 170 мкл общего объема жидкости, и проводили взаимодействие в течение 10 минут при 30°C с последующим нагреванием до 95°C в течение 5 минут для окончания взаимодействия. 80 мкл данного раствора анализировали посредством ВЭЖХ в следующих условиях и ее активность измеряли, основываясь на сниженном количестве субстрата:
Прибор: система ВЭЖХ L-6300 (Hitachi Co. Ltd)
Колонка: YMC-PACK ODS 120A 4,6 × 150 мм (YMC)
Элюент: (A) 0,1% TFA (B) 80% ацетонитрил / 0,1% TFA
Условия элюции: линейный градиент концентрации 12-16% ацетонитрила (в течение 15 мин)
Скорость потока: 1,0 мл/мин
Длина волны детекции: 220 нм
С использованием данных условий пептид GPSFRAPDS элюировали со временем задержки от 13 до 14 мин, а продукт деградации FRAPDS элюировали со временем задержки от 7,5 до 8,5 мин.
Количество фермента, которое катализирует расщепление одного (1) нмоль про-MTG в минуту, определяли как одну (1) единицу ферментативной активности.
(2) Очистка нейтральной металлопротеазы (SVP35), продуцированной Streptoverticillium mobaraense IFO13819
В колбу Sakaguchi объемом 5 л (колба для перемешивания) помещали 800 мл культуральной среды ISP2, инокулировали Streptoverticillium mobaraense IFO13819 с чашки и культивировали при встряхивании при 30°C в течение 48 часов при 120 об/мин. Культуральную среду центрифугировали, и надосадочную жидкость культуры удаляли для сбора клеток. Клетки ресупендировали в 20 мМ буфере Tris-HCl/30 мМ хлорид натрия (рН 7,5), встряхивали на льду в течение 4 часов и затем надосадочную жидкость собирали центрифугированием. Полученную надосадочную жидкость фильтровали с использованием фильтра Depth (размер пор 0,22 мкм, сделано Saltorius Co. Ltd.), затем ее наносили на колонку с СМ-Sepharose FF (1,6 ø × 10 см, Amersham Pharmacia Co. Ltd.), уравновешенную 20 мМ буфера Tris-HCl (рН 7,5), содержащего 5 мМ хлорида кальция и 0,01 мМ хлорида цинка, используя FPLC (Amersham Pharmacia Co. Ltd.), элюировали тем же буфером с использованием линейного градиента концентрации 0-0,5 M хлорида натрия. Собирали фракцию содержащую активный ингредиент, наносили на колонку с фенил-Sepharose HP (1 мл, Amersham Pharmacia Co. Ltd.), уравновешенную 20 мМ буфера Tris-HCl, содержащего 1,5 M сульфата аммония, 5 мМ хлорида кальция и 0,01 мМ хлорида цинка, и элюировали с использованием линейного градиента концентрации 1,5-0 M сульфата аммония. Собирали активную фракцию, и деминерализовали посредством 20 мМ буфера Tris-HCl (рН 7,5), содержащего 5 мМ хлорида кальция и 0,01 мМ хлорида цинка, с использованием колонки PD-10 (Amersham Pharmacia) с получением частично очищенного раствора фермента.
Ферментативную активность на каждой стадии измеряли с использованием пептида GPSFRAPDS в качестве субстрата тем же способом, что в (1).
(3) Оценка свойств нейтральной металлопротеазы (SVP35), продуцированной Streptoverticillium mobaraense IFO13819
i) Субстратная специфичность
1 мг/мл раствора инсулина B и раствора про-MTG, полученных в 20 мМ буфере Tris-HCl (pH 7,5), содержащем 5 мМ хлорида кальция и 0,01 мМ хлорида цинка, использовали в качестве субстрата, и раствор фермента добавляли к раствору для взаимодействия при 30°C в течение 2 часов, а затем пептидные фрагменты отделяли путем ВЭЖХ в следующих условиях:
Прибор: L-7100 / 7200 / 7405 / D-7600 (Hitachi Co. Ltd.)
Колонка: VYDAC C18 4,6 мм I.D. × 250 мм (VYDAC)
Элюент: (A) 0,1% TFA (B) 80% ацетонитрил/ 0,1% TFA
Условия элюции: линейный градиент концентрации 4-44% ацетонитрила
Скорость потока: 0,5 мл/мин
Длина волны детекции: УФ 220 нм
Аминокислотные последовательности полученных пептидных фрагментов анализировали с использованием PPSQ-10 (Shimadzu Co. Ltd.) для характеристики последовательностей точек расщепления SVP35. В результате подтверждали, что пептид расщеплялся главным образом перед (с N-конца) Phe, часто перед Leu, иногда перед Tyr, Trp, Ile, Val, и что SVP распознавала ароматические аминокислоты и гидрофобные аминокислоты с объемными боковыми цепями, расположенные в P'1 от участка расщепления.
ii) Оптимум pH
В 0,15 M буфере GTA (уравновешенном 3,3-диметилглутаровой кислотой, трис-(гидроксиметил)аминометаном, 2-амино-2-метил-1,3-пропандиолом) от pH 3 до pH 10, SVP35 позволяли взаимодействовать с Gly-Pro-Ser-Phe-Arg-Ala-Pro-Asp-Ser в качестве субстрата при 30°C в течение 10 минут. В результате было выявлено, что оптимум pH для SVP35 составлял около 7,0, и что если активность при pH 7,0 определяли как 100%, SVP35 имела активность 70% или более при pH 6,0-8,0 и активность 80% или более при pH 6,5-7,5 (см. фиг.1).
iii) pH-стабильность
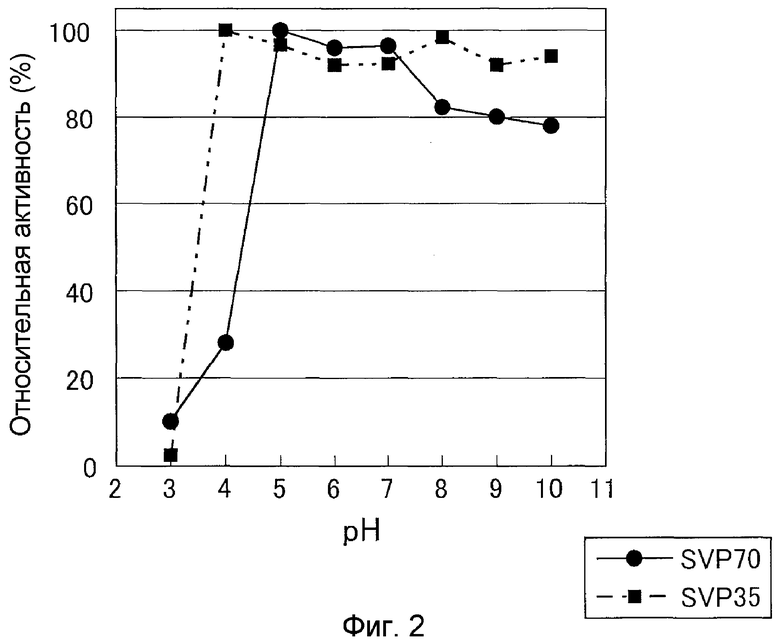
К 10 мкл очищенного раствора фермента SVP35 добавляли 40 мкл 0,15 M буфера GTA каждого pH от pH 3 до pH 10, оставляли при 4°C в течение ночи с последующим добавлением 0,1 M фосфата натрия (pH 7,0) для доведения объема жидкости до 400 мкл и доводили объем до pH 7,0. В растворы данных ферментов добавляли в качестве субстрата Gly-Pro-Ser-Phe-Arg-Ala-Pro-Asp-Ser, обеспечивали взаимодействие при pH 7,0 при 30°C в течение 10 минут. В результате показали, что SVP35 была стабильна в интервале от pH 4 до pH 10 (если активность при pH 4,0 определяли как 100%, она имела активность 90% или более при pH 4-10) (см. фиг.2).
iv) Оптимум температуры
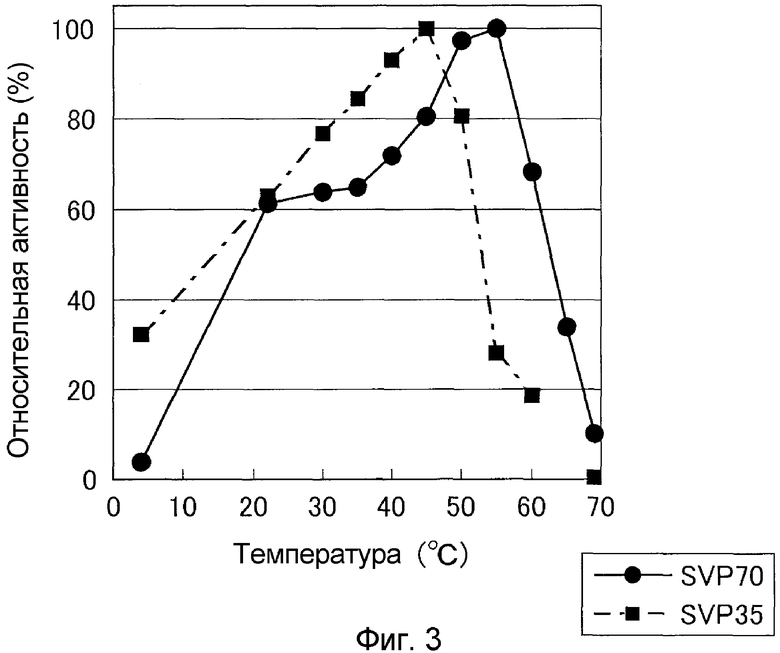
К очищенному раствору фермента, разбавленного 20 мМ буфером Tris-HCl (pH 7,5), содержащим 5 мМ хлорида кальция, и 0,01 мМ хлорида цинка, добавляли Gly-Pro-Ser-Phe-Arg-Ala-Pro-Asp-Ser и обеспечивали взаимодействие при pH 7,0, от 5 до 65°C в течение 10 минут. В результате показали, что оптимальная температура для SVP35 составляла около 45°C, и она имела высокую активность в интервале от 40 до 50°C (она имела активность 80% или более по сравнению с таковой при 45°C) (см. фиг.3).
v) Температурная стабильность
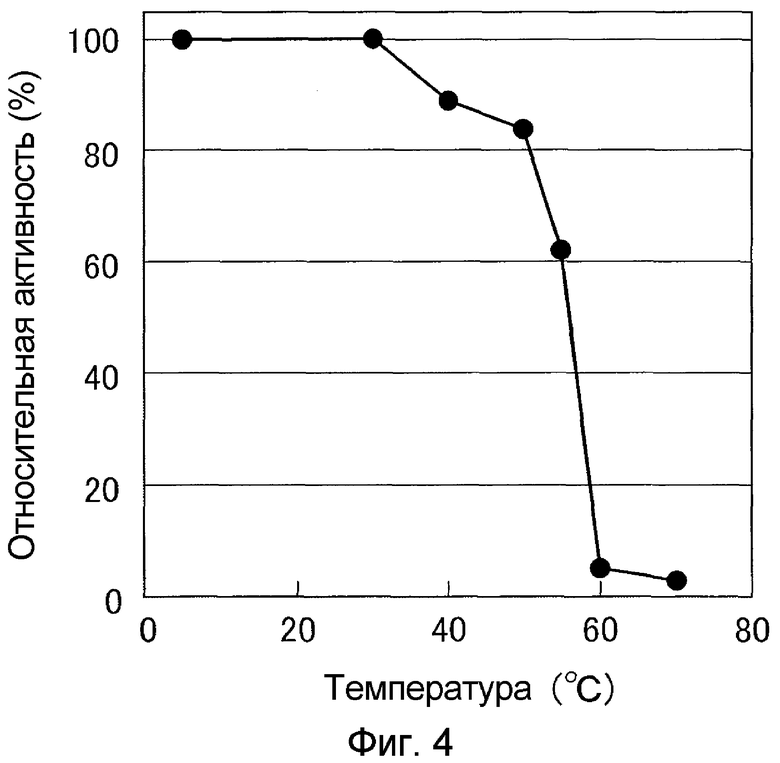
К 10 мкл очищенного раствора фермента добавляли 40 мкл 20 мМ буфера Tris-HCl (pH 7,5), содержащего 5 мМ хлорида кальция и 0,01 мМ хлорида цинка, для обработки при 4°C или при 30-70°C в течение 15 минут, затем охлаждали на льду, добавляли 250 мкл 20 мМ натрий-фосфатного буфера (pH 7,0). К раствору данного фермента добавляли Gly-Pro-Ser-Phe-Arg-Ala-Pro-Asp-Ser в качестве субстрата и обеспечивали взаимодействие при 30°C в течение 5 минут. Если активность фермента, обработанного при 4°C, определяли как 100%, рассчитывали оставшуюся активность при каждой температуре. В результате было показано, что SVP35 сохраняет 80% активности при 50°C, но она теряет свою активность при 60°C (см. фиг.4).
vi) Ингибиторы
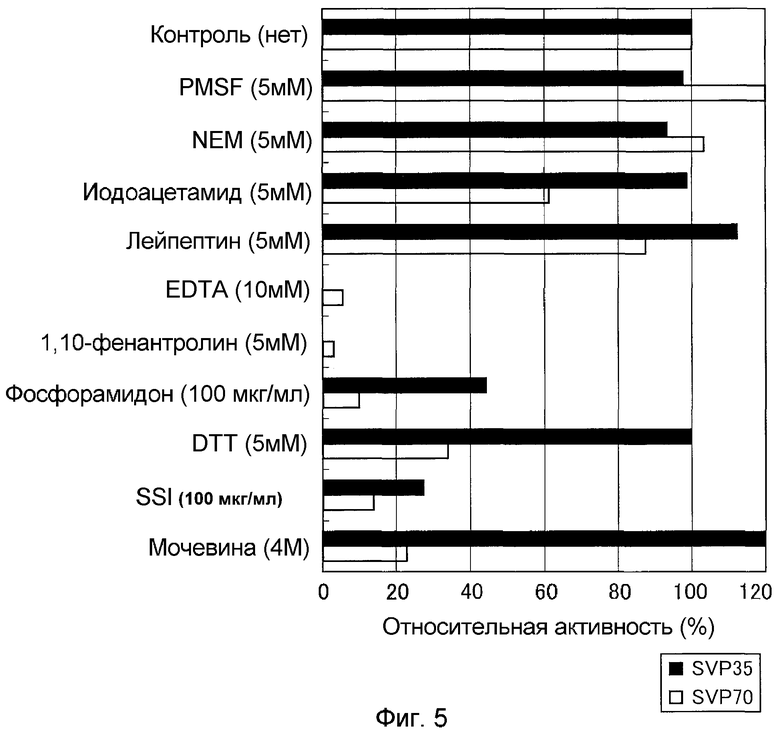
К 20 мМ натрий-фосфатного буфера (pH 7,0), содержащего различные соединения в концентрациях, показанных на фиг.5, добавляли раствор очищенного фермента и оставляли на 60 минут при комнатной температуре. Затем добавляли в качестве субстрата Gly-Pro-Ser-Phe-Arg-Ala-Pro-Asp-Ser и позволяли взаимодействовать в течение 10 минут при 30°C. Относительную активность добавления каждого соединения рассчитывали, считая активность расщепления Gly-Pro-Ser-Phe-Arg-Ala-Pro-Asp-Ser в отсутствии соединений 100%-ной. В результате было показано, что SVP35 сильно ингибируется этилендиаминотетрауксусной кислотой, 1,10-фенантролином и фосфорамидоном, которые представляют собой ингибиторы металлопротеаз, и ингибитором субтилизина Streptomyces (SSI), происходящим из актиномицетов (см. фиг.5).
(4) Характеристика свойств нейтральной металлопротеазы (SVP70), продуцированной Streptoverticillium mobaraense IFO13819
i) Субстратная специфичность
Субстратную специфичность оценивали, как описано в (3)-i). В результате выявлено, что субстрат расщепляется главным образом перед (с N-концевой стороны) Phe, часто Leu, иногда Tyr, Trp, Ile, Val и что SVP70 распознает ароматические аминокислоты и гидрофобные аминокислоты с объемными боковыми цепями, расположенные в P'1 от участка расщепления.
ii) Оптимум pH
Оптимум pH для SVP70 оценивали сходно с (3)-ii). В результате было выявлено, что оптимум pH SVP70 составлял примерно 7,0 и что если активность при pH 7,0 определить как 100%, SVP70 имеет активность 90% или более при pH 6,0-8,0 и активность 95% или более при pH 6,5-7,5 (см. фиг.1).
iii) pH-стабильность
pH-стабильность оценивали сходно с (3)-iii). В результате было показано, что SVP70 была стабильна при pH от 5 до 10, но она была менее стабильна, чем SVP35, в слабощелочных условиях (см. фиг.2). Конкретно, если активность при pH 5,0 определить как 100%, она имела активность 90% или более в интервале от pH 5 до pH 7 и активность около 80% или более даже в интервале от pH 7 до pH 10.
iv) Оптимум температуры
Оптимум температуры SVP70 оценивали сходно с (3)-iv). В результате было показано, что оптимум температуры для SVP70 находился в интервале примерно от 50 до 55°C, особенно, около 55°C (см. фиг.3).
v) Ингибиторы
Ингрибиторную активность различных соединений в отношении SVP70 оценивали по аналогии с (3)-vi). В результате SVP70 подвергалась сильному ингибирующему действию этилендиаминотетрауксусной кислоты, 1,10-фенантролина и фосфорамидона, которые представляют собой ингибиторы металлопротеаз, и восстановителя дитиотреитола, мочевины, и ингибитора субтилизина Streptomyces (SSI), происходящего из актиномицетов (см. фиг.5).
(5) Секвенирование N-концевой аминокислотной последовательности SVP35 и SVP70
Очищенные ферменты SVP35 и SVP70, полученные выше в (1) и (2), переносили на мембрану из поливинилидендифторида (PVDF) с использованием картриджа для мембран (Perkin Elmer Co. Ltd.) и анализировали N-концевую аминокислотную последовательность с помощью газофазового белкового секвенатора PPSQ-10 (Shimadzu Co. Ltd.). Аминокислотная последовательность SVP35 показана в SEQ ID NO: 1, и аминокислотная последовательность SVP70 показана в SEQ ID NO: 2. В этих последовательностях можно видеть гомологию.
В соответствии с этим искали белки, которые характеризуются какой-либо гомологией в отношении данных N-концевых аминокислотных последовательностей, затем обнаружили металлопротеазу SGMP II (J. Biochem., Vol. 110, p. 339-344, 1991) из Streptomyces griseus и три металлопротеазы (GenBank/EMBL/DDBJ CAB76000, то же CAB76001, и то же CAB69762), и т.д., из Streptomyces coelicolor. Данные протеазы также могут использоваться для избирательного отщепления проструктурной части про-MTG, и затем они могут использоваться для продуцирования активной формы MTG по настоящему изобретению.
(6) Клонирование гена SVP35 и его секреторной экспрессии в коринеформных бактериях
Хромосомную ДНК Streptomyces coelicolor A3(2) получали с использованием способа Saito and Miura [Biochem. Biohhys. Acta, 72, 619 (1963)]. Праймеры, показанные в SEQ ID NO: 3 и SEQ ID NO: 4, синтезировали с использованием последовательности гена металлопротеазы (GenBank/EMBL/DDBJ CAB76001) из Streptomyces coelicolor, которые имеют гомологию в отношении N-концевой аминокислотной последовательности. Праймеры, показанные в SEQ ID NO: 3 и SEQ ID NO: 4, использовали для выполнения ПЦР с использованием хромосомной ДНК Streptomyces coelicolor A3(2) в качестве матрицы и амплифицировали генную область гена металлопротеазы. Для реакции ПЦР использовали ДНК-полимеразу Pyrobest (Takarasyuzo Co. LTD.) и по условиям реакции следовали протоколу, рекомендованному производителем. Хромосомную ДНК Streptoverticillium mobaraense IFO13819, полученную способом Saito and Miura, расщепляли различными ферментами рестрикции, которые распознают последовательность длиной 6 оснований, расщепленный образец анализировали гибридизацией путем блоттинга по Саузерну, как описано в Molecular Cloning 2nd edition [J. Sambrook E. F. Fritsch and T. Maniatis, Cold Spring Harbor Laboratory Press, p9. 31 (1989)], с использованием в качестве зонда меченного 32P продукта ПЦР, и при расщеплении SphI выявляли единичную полосу размером примерно 8 тыс.п.н. Соответственно, хромосомную ДНК Streptoverticillium mobaraense IFO13819, которую получали следующим способом, расщепляли SphI, и фрагмент размером примерно 8 тыс.п.н. получали посредством электрофореза в агарозном геле с использованием EASYTRAP Ver. 2 (Takarasyuzo Co. LTD.). Полученный фрагмент встраивали в участок SphI на pUC18, которую вводили в компетентные клетки Escherichia coli JM109 (Takarasyuzo Co. LTD.) с образованием библиотеки. Библиотеку подвергали скринингу на бактериальный штамм, который содержит плазмиду, в которую клонировали генный фрагмент SVP35, путем гибридизации колоний, как описано в Molecular Cloning 2nd edition [J. Sambrook E. F. Fritsch and T. Maniatis, Cold Spring Harbor Laboratory Press, p1. 90 (1989)] с использованием синтетического нуклеотида в качестве зонда.
Плазмиду выделяли из штамма, полученного выше, и обозначали как pVSV1. Определяли нуклеотидную последовательность фрагмента, клонированного в pVSV1. Нуклеотидная последовательность данного клонированного фрагмента показана в SEQ ID NO: 5. Предсказывали первичную аминокислотную структуру, кодируемую данным геном, что позволяло определять целую первичную аминокислотную последовательность SVP35, содержащую сигнальную последовательность SVP35, включающую в себя аминокислотную последовательность ранее определенной N-концевой части, и область, признанную проструктурной частью. Полная аминокислотная последовательность SVP35 показана в SEQ ID NO: 6. Предполагается, что аминокислоты № 1-36 аминокислотной последовательности, описанной в SEQ ID NO: 6, относятся к сигнальной последовательности, аминокислоты № 37-216 относятся к проструктурной части и аминокислоты № 217-537 соответствуют зрелой SVP35.
Праймеры, показанные в SEQ ID NO: 7 и SEQ ID NO: 8, синтезированы с использованием pVSV1 в качестве матрицы при ссылке на последовательность SEQ ID NO: 5, и генную область, содержащую проструктурную часть SVP35 и зрелую SVP35 амплифицировали способом ПЦР. Для ПЦР использовали ДНК-полимеразу Pyrobest (Takarasyuzo Co. Ltd.) и по условиям реакции следовали протоколу, рекомендованному производителем.
Далее с использованием pPKSPTG1, описанной в WO 01/23591, в качестве матрицы, область, включающую в себя 5'-вышележащую область (в положении апстрим), содержащую промоторную область гена PS2, который представляет собой белок клеточной поверхности C. glutamicum, и сигнальную последовательность SlpA белка клеточной поверхности C.ammoniagenes, амплифицировали способом ПЦР с использованием комбинации олигонуклеотидов SEQ ID NO: 9 и SEQ ID NO: 10. Праймер, показанный в SEQ ID NO: 10, содержит последовательность, кодирующую N-концевые аминокислоты SVP35, содержащие проструктурную часть.
Затем ген гетерологичного пре-про-SVP35 генного фрагмента слияния, который был присоединен к 5'-вышележащей области (в положении апстрим), содержащей промоторную область гена PS2 и сигнальную последовательность SlpA, белка клеточной поверхности C.ammoniagenes, амплифицировали проведением перекрестной ПЦР с SEQ ID NO: 8 и SEQ ID NO: 9, используя смесь по 1 мкл каждого из амплифицированных растворов ПЦР. Амплифицированный фрагмент размером примерно 2,3 тыс.п.н. выявляли путем электрофореза в агарозном геле. ПЦР-продукт подвергали электрофорезу в агарозном геле для получения фрагмента размером примерно 2,3 тыс.п.н., после затупления его концов с использованием набора для затупления ДНК (Takarasyuzo Co. Ltd.) фрагмент встраивали в SmaI-участок pCV7, как описано в JP-Kokai № 9-070291 с получением pVSV1. Нуклеотидную последовательность встроенного фрагмента определяли общепринятым способом для подтверждения того, что сконструированный ген слияния соответствует ожиданиям.
C. glutamicum ATCC13869 трансформировали конструированной pVSV1 и отбирали штаммы, выращенные на агаровой среде CM2S, включающей 5 мг/л хлорамфеникола (10 г дрожжевого экстракта, 10 г триптона, 5 г сахарозы, 5 г NaCl, 15 г агара на литр дистиллированной воды). Затем подвергшиеся селекции C. glutamicum ATCC13869, несущие pVSV1, культивировали в культуральной среде MMTG (60 г глюкозы, 0,4 г гептагидрата сульфата магния, 30 г сульфата аммония, 1 г однозамещенного фосфата калия, 0,01 г гептагидрата сульфата железа-II, 0,01 г пентагидрата сульфата марганца, 450 мкг гидрохлорида тиамина, 450 мкг биотина, 0,15 г DL-метионина, 50 г карбоната кальция на литр дистиллированной воды, pH доведен до 7,5), содержащей 5 мг/л хлорамфеникола, при 30°C в течение 30 часов. 1 мл культуральной надосадочной жидкости центрифугировали для разделения надосадочной жидкости культуры и бактерий. В культуральной надосадочной жидкости выявляли активность SVP35 и в результате после электрофореза SDS-PAGE (Nature, 227, 380-685 (1970)) по способу Лэммли подтверждали секрецию-экспрессию примерно 200 мг/л SVP35.
Пример 2. Преобразование предшественника трансглутаминазы (про-MTG), продуцированного Streptoverticillium mobaraense IFO13819, в активную форму
С применением в качестве очищенного субстрата про-MTG (1 мг/мл), экспрессированную в Corynebacterium glutamicum нейтральную протеазу (SVP35, SVP70) из Streptoverticillium mobaraense или нейтральную металлопротеазу SGMP II из Streptomyces griseus смешивали в соотношении субстрат:фермент = 200:1, смеси позволяли взаимодействовать при 30°C. Через 0, 1, 2, 4, 7, 20 часов реакционную смесь последовательно отбирали, аликвоты реакционной смеси смешивали с буфером для образца SDS-PAGE, нагревали при 95°C в течение 3 минут и затем подвергали SDS-PAGE по способу Лэммли (Nature, 227, 680-685 (1970)). Результат показан на фиг.6. Как можно видеть на фиг.6, при действии данных протеаз про-MTG преобразовывалась в зрелую форму, и количество продуцированной MTG не снижалось даже при долгосрочном взаимодействии. Трансглутаминазную активность (TG) отобранных фракций измеряли гидроксаматным способом и подтверждали достаточную активность. Кроме того, SGMP II очищали от актиназы (Kakenseiyaku Co. Ltd.) по способу сравнения (J. Biochem., Vol. 110, p. 339-344, 1991).
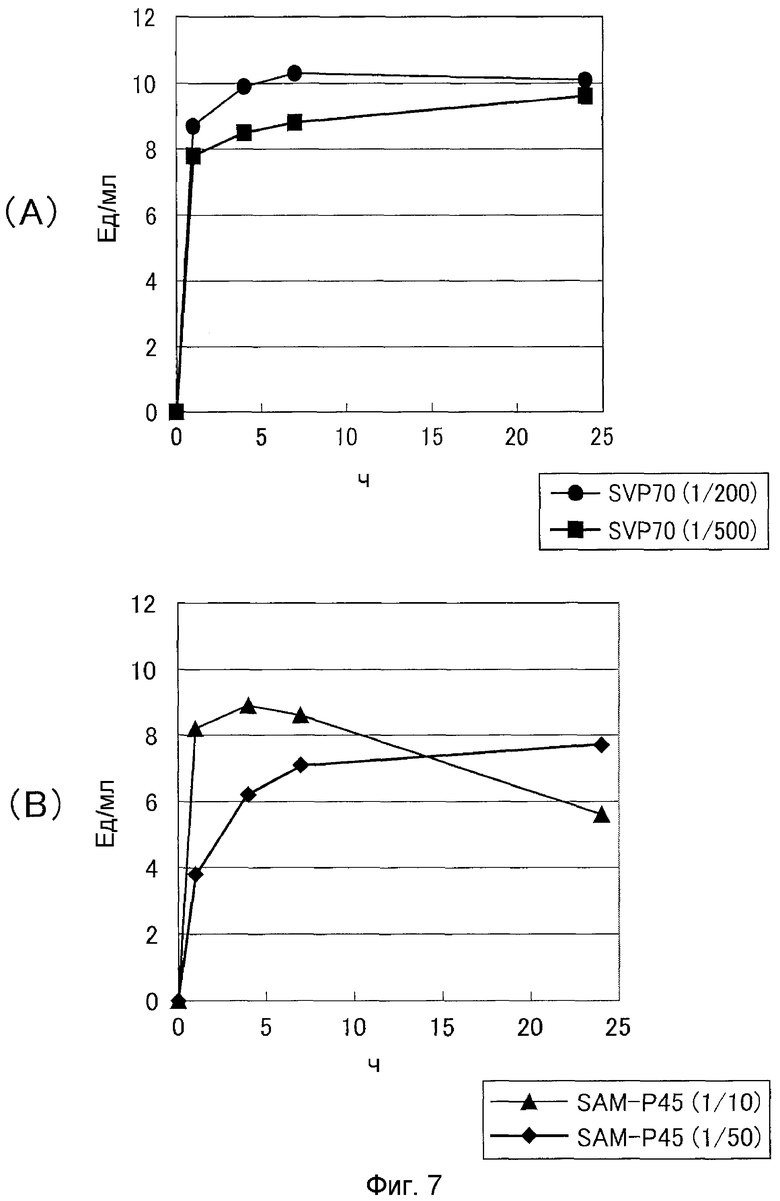
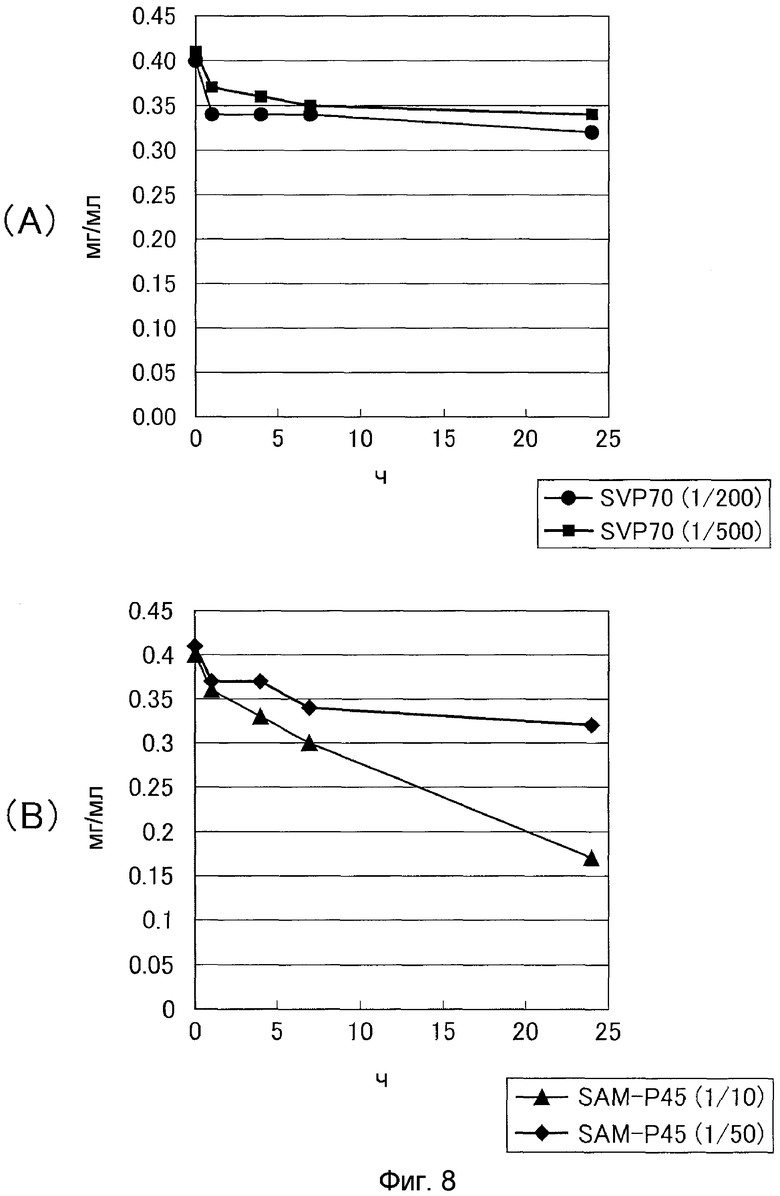
Затем нейтральную металлопротеазу SVP70 из Streptoverticillium mobaraense и сериновую протеазу SAM-P45 (Streptomyces albogriseolus) в качестве контроля добавляли к про-MTG с постепенно увеличивающимся количеством данных ферментов и позволяли взаимодействовать при 30°C и pH 7,0. Через 1, 4, 7 и 24 часа реакционную смесь последовательно отбирали для определения активности TG гидроксаматным способом (см. фиг.7). Концентрацию белка TG измеряли путем обращенно-фазовой хроматографии (см. фиг.8). В результате было показано, что SVP может преобразовывать про-MTG в активную MTG в таком малом количестве, как 1/500 по отношению к субстрату. Было показано, что SAM-P45 генерирует только недостаточную трансглутаминазную активность даже в количестве 1/50 по отношению к субстрату и что полное преобразование в активную форму не наблюдается. С другой стороны, когда SAM-P45 добавляли в количестве 1/10 в отношении субстрата, наблюдали преобразование в активную MTG, но при снижении количества и активности белка MTG. Это указывает на то, что за счет SAM-P45 происходило расщепление зрелой MTG.
Настоящее изобретение относится к новой протеазе из актиномицета Streptoverticillium mobaraense, которая специфично расщепляет проструктурную часть предшественника трансглутаминазы с ее активацией, и к гену данной протеазы. Новая протеаза по настоящему изобретению может экспрессироваться в большом количестве коринеформной бактерией, и, таким образом, изобретение относится к способу эффективного продуцирования трансглутаминазы из микроорганизмов.
Преимуществом применения нейтральной металлопротеазы из актиномицетов по настоящему изобретению для продуцирования активной MTG состоит в том, что данные ферменты имеют сильную активность по избирательному отщеплению проструктурной части про-MTG и что данные ферменты могут экстрацеллюлярно экспрессироваться коринеформной бактерией.
Поскольку показано, что про-MTG из актиномицетов может эффективно экспрессироваться и секретироваться коринеформной бактерией, возможно эффективное продуцирование активной MTG одной бактериальной клеткой путем совместной экспрессии и секреции про-MTG и нейтральной металлопротеазы. В данном случае достаточно экспрессировать нейтральную металлопротеазу только в количестве, необходимом и достаточном для отщепления проструктурной части про-MTG.
Литература
1. JP-Kokoku No. 1-50382.
2. JP-Kokai No. 64-27471.
3. WO publication No. 01/2351.
4. JP-Kokai No. 6-502548.
5. JP-Kokai No. 10-108675.
6. Eur. J. Biochem., Vol. 257, pages 570-576, 1998.
7. J. Biochem., Vol. 110, pages 339-244, 1991.








| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ГЕТЕРОЛОГИЧНОГО СЕКРЕТОРНОГО БЕЛКА | 2000 |
|
RU2224796C2 |
| СПОСОБ СЕКРЕТОРНОЙ ПРОДУКЦИИ БЕЛКА | 2002 |
|
RU2264463C2 |
| СПОСОБ МОДИФИЦИРОВАНИЯ ТРАНСГЛУТАМИНАЗ МИКРООРГАНИЗМОВ | 2001 |
|
RU2260051C2 |
| СПОСОБ СЕКРЕТОРНОЙ ПРОДУКЦИИ БЕЛКА | 2012 |
|
RU2588439C2 |
| ПРОДУЦИРОВАНИЕ ТЕРМОЛИЗИНА И ЕГО ВАРИАНТОВ И ЕГО ИСПОЛЬЗОВАНИЕ В ЖИДКИХ ДЕТЕРГЕНТАХ | 2008 |
|
RU2536255C2 |
| ПРОДУЦИРОВАНИЕ ТЕРМОЛИЗИНА И ЕГО ВАРИАНТОВ И ЕГО ИСПОЛЬЗОВАНИЕ В ЖИДКИХ ДЕТЕРГЕНТАХ | 2014 |
|
RU2733541C2 |
| СПОСОБЫ УЛУЧШЕНИЯ ЭФФЕКТИВНОСТИ БЕЛКОВ | 2008 |
|
RU2569106C2 |
| СПОСОБ АМПЛИФИЦИРОВАНИЯ ЛОКУСОВ В БАКТЕРИАЛЬНОЙ КЛЕТКЕ | 2009 |
|
RU2577985C2 |
| НОВЫЕ ГЕНЫ, КОДИРУЮЩИЕ НОВЫЕ ПРОТЕОЛИТИЧЕСКИЕ ФЕРМЕНТЫ | 2002 |
|
RU2423525C2 |
| ПРОТЕАЗА Streptomyces | 2008 |
|
RU2486244C2 |
Изобретение относится к биотехнологии. Культивируют коринеформную бактерию, в которую введен ген, кодирующий нейтральную металлопротеазу из Streptoverticillium. Отщепляют проструктурную часть протрансглутаминазы нейтральной металлопротеазой, которую продуцирует данная бактерия. Характеристики нейтральной металлопротеазы приведены в тексте. Изобретение также относится к молекуле нуклеиновой килоты, кодирующей нейтральную металлопротеазу. Настоящее изобретение позволяет получать протеазу, которую можно использовать для продуцирования трансглутаминазы и которая избирательно отщепляет проструктурную часть протрансглутаминазы. 5 н. и 3 з.п. ф-лы, 8 ил.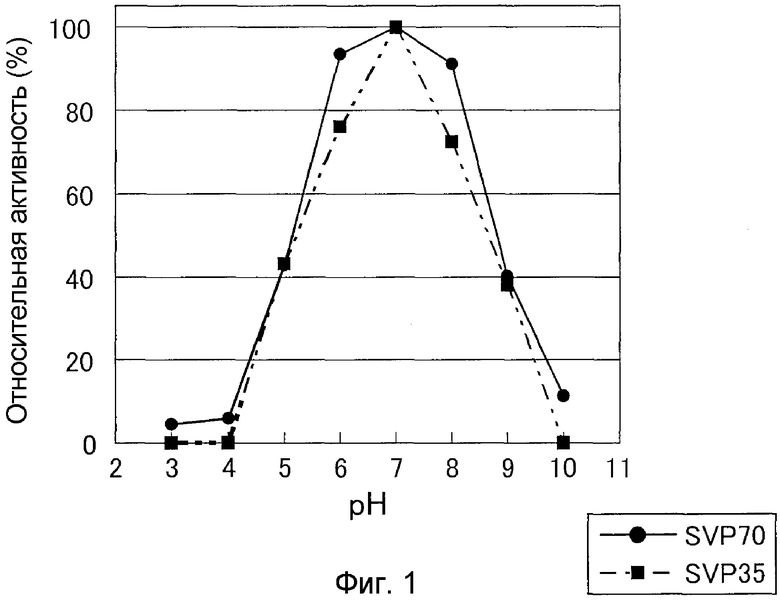
1) она имеет молекулярную массу, примерно равную 35000;
2) она имеет оптимум рН, равный рН 7,0;
3) она стабильна при рН 4-10;
4) она имеет оптимальную температуру, примерно равную 45°С;
5) она стабильна при температуре ниже, чем примерно 50°С; и
6) она сильно ингибируется этилендиаминотетрауксусной кислотой, 1,10-фенантролином и фосфорамидоном, которые являются ингибиторами металлопротеаз, и ингибитором субтилизина Streptomyces (SSI) из актиномицетов.
1) она имеет молекулярную массу, примерно равную 71000;
2) она имеет оптимум рН, равный 7,0;
3) она стабильна при рН 5-10;
4) она имеет оптимальную температуру, примерно равную 55°С; и
5) она сильно ингибируется этилендиаминотетрауксусной кислотой, 1,10-фенантролином и фосфорамидоном, которые являются ингибиторами металлопротеаз, дитиотреитолом, который представляет собой восстановитель SH-группы, и ингибитором субтилизина Streptomyces (SSI), происходящим из актиномицетов.
1) она имеет молекулярную массу, примерно равную 35000;
2) она имеет оптимум рН, равный рН 7,0;
3) она стабильна при рН 4-10;
4) она имеет оптимальную температуру, примерно равную 45°С;
5) она стабильна при температуре ниже, чем примерно 50°С; и
6) она сильно ингибируется этилендиаминотетрауксусной кислотой, 1,10-фенантролином и фосфорамидоном, которые являются ингибиторами металлопротеаз, и ингибитором субтилизина Streptomyces (SSI) из актиномицетов.
1) она имеет молекулярную массу, примерно равную 71000;
2) она имеет оптимум рН, равный 7,0;
3) она стабильна при рН 5-10;
4) она имеет оптимальную температуру, примерно равную 55°С; и
5) она сильно ингибируется этилендиаминотетрауксусной кислотой, 1,10-фенантролином и фосфорамидоном, которые являются ингибиторами металлопротеаз, дитиотреитолом, который представляет собой восстановитель SH-группы, и ингибитором субтилизина Streptomyces (SSI), происходящим из актиномицетов.
| US 6100053 А 08.08.2000 | |||
| US 2002187525 A 12.12.2002 | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| Походная разборная печь для варки пищи и печения хлеба | 1920 |
|
SU11A1 |
| СПОСОБ ПОЛУЧЕНИЯ ГЕТЕРОЛОГИЧНОГО СЕКРЕТОРНОГО БЕЛКА | 2000 |
|
RU2224796C2 |
| US 2003045438 A 06.03.2003 | |||
| СПОСОБ СТАБИЛИЗАЦИИ ВОДНОГО РАСТВОРА ИЛИ СУСПЕНЗИИ МЕТАЛЛОПРОТЕАЗЫ И СТАБИЛИЗИРОВАННЫЙ ВОДНЫЙ РАСТВОР ИЛИ СУСПЕНЗИЯ МЕТАЛЛОПРОТЕАЗЫ | 1994 |
|
RU2167937C2 |
| RU 2001113066 A 27.02.2003. | |||

