В соответствии с настоящим изобретением предложены анализы для скрининга, или для определения активности (включая определение эффективности препарата) агониста гена активации лимфоцитов 3 (LAG-3). В соответствии с настоящим изобретением также предложены наборы для осуществления указанных анализов.
Стимуляция функции Т-клеток начинается после взаимодействия Т-клеточного рецептора (TCR) с короткими пептидами, презентированными молекулами ГКГС I или II класса (ГКГС I для CD8 Т-клеток, ГКГС II для CD4 Т-клеток) на поверхности антигенпрезентирующих клеток (АПК). В первичных Т-клетках TCR сам по себе не способен активировать последующие сигнальные пути, чтобы запустить активацию Т-клеток. Для этого также требуются корецепторы, такие как CD4 для хелперных Т-клеток и CD8 для цитотоксических Т-клеток. Данные корецепторы связываются с соответствующими молекулами ГКГС и стабилизируют взаимодействие Т-клеток и АПК. Дополнительно к связыванию TCR с нагруженными антигеном ГКГС, как для хелперных Т-клеток, так и для цитотоксических Т-клеток требуется множество вторичных сигналов, чтобы они полностью активировались и начали ответ. В случае хелперных Т-клеток, первый такой сигнал обеспечивается CD28. Этот белок является рецептором для двух молекул, экспрессированных на АПК (CD80 и CD86), и вызывает пролиферацию Т-клеток, приводя к экспансии клонов Т-клеток, специфичных к антигену. Активация цитотоксических Т-клеток в меньшей степени зависит от CD28, но им требуются сигналы от других костимулирующих молекул, таких как CD70 и CD137.
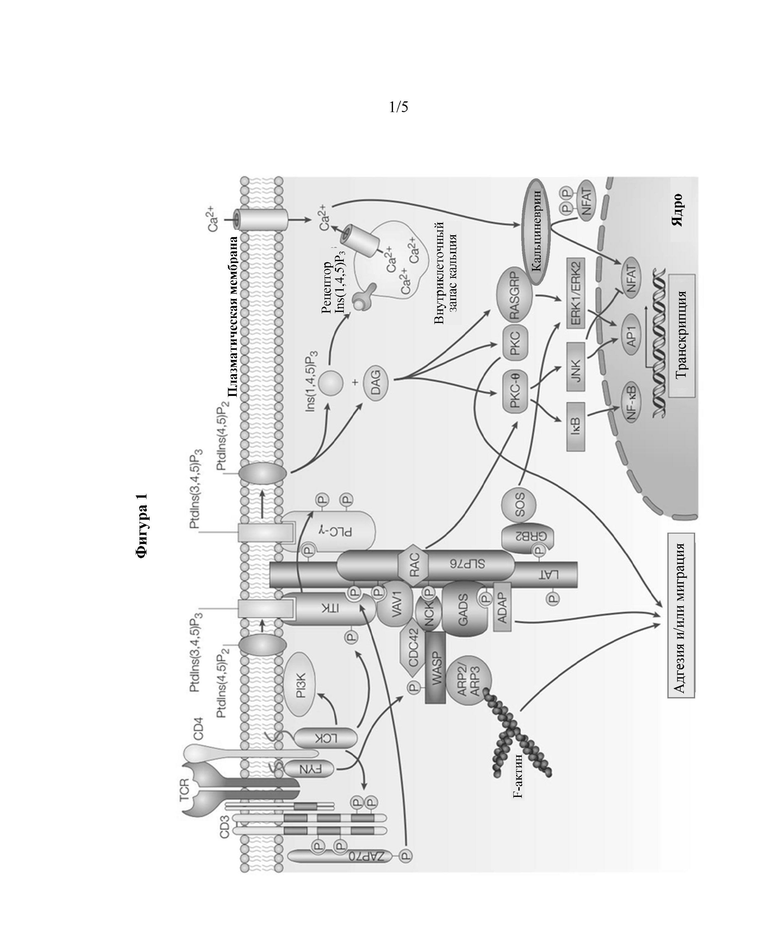
TCR расположен в непосредственной близости от комплекса сигнальных молекул, которые опосредуют активацию Т-клеток посредством множества сигнальных каскадов (см. обзор сигнальных путей TCR на Фигуре 1). Данные сигнальные молекулы включают семейство белков CD3. После надлежащего связывания TCR с комплексом пептид-ГКГС, вызываются конформационные изменения в связанных с ним цепях CD3, что приводит к их фосфорилированию и связыванию с последующими белками в сигнальном пути. Цепь дзета TCR также фосфорилируется после связывания TCR. Эти молекулы фосфорилируются Src-киназами - лейкоцитепецифичной тирозинкиназой (LCK) и Fyn, - по их С-концевым активирующим мотивам иммунорецептора на основе тирозина (ITAM).
Фосфорилированные CD3 ITAM привлекают и активируют киназу семейства Syk - дзета-активируемый белок 70 кДа (ZAP70). ZAP70 затем фосфорилирует связанный с мембраной каркасный белок, названный линкером для активации Т-клеток (LAT). LAT, в свою очередь, привлекает второй молекулярный каркас - содержащий домен SH2 лейкоцитарный белок с молекулярной массой 76 кДа (Slp-76). ZAP70 затем фосфорилирует Slp-76, и полученный в результате этого комплекс LAT-Slp-76 действует как каркас для привлечения сигнальных эффекторных молекул. Индуцируемая интерлейкином-2 тирозинкиназа (ITK) затем взаимодействует с комплексом LAT-Slp-76 и активируется посредством аутофосфорилирования. Это вызывает фосфорилирование эффекторной молекулы фосфолипазы С-гамма (PLC-γ1). PLC-γ1 передает сигналы TCR путем расщепления фосфатидилинозитолтрифосфата (PIP2) в плазматической мембране с образованием вторичных мессенджеров диацилглицерина (DAG) и инозитолтрифосфата (IP3).
DAG, связанный с мембраной липид, активирует множество последующих сигнальных белков, включая различные изоформы протеинкиназы С (PKC) и RAS-белок, высвобождающий гуаниловый нуклеотид (RasGRP). После активации посредством DAG, PKC-тета участвует в активации пути NF-κВ, тогда как RasGRP является ключевым активатором сигнальных путей МАРК. IP3 стимулирует отток Са2+ из эндоплазматического ретикулума в цитоплазму. Повышенные уровни Са2+ вызывают активацию протеинфосфатазы кальциневрина, которая затем дефосфорилирует фактор транскрипции Т-клеток - ядерный фактор активированных Т-клеток (NFAT). Дефосфорилированный NFAT затем мигрирует в ядро, чтобы присоединиться к другим факторам транскрипции, чтобы вызвать транскрипцию определенных генов.
Неконтролируемые иммунные ответы на патогены или собственные антигены могут вызывать воспалительное повреждение ткани и аутоиммунные заболевания. Для того чтобы это предотвратить, иммунные ответы регулируются равновесием между костимулирующими и ингибиторными сигналами, в совокупности называемыми иммунными контрольными точками, которые необходимы для поддержания аутотолерантности и защиты хозяина от повреждения ткани. Активированные Т-клетки экспрессируют множество коингибиторных рецепторов, таких как ген активации лимфоцитов 3 (LAG-3), белок запрограммированной клеточной смерти 1 (PD-1), ассоциированный с цитотоксическими Т-лимфоцитами белок 4 (CTLA-4) и Т-клеточный иммунорецептор, содержащий домены иммуноглобулина и ингибирующего мотива иммунорецептора на основе тирозина [ITIM] (TIGIT). Было показано, что ингибиторные рецепторы иммунных контрольных точек модулируют ответы Т-клеток на собственные белки, а также на хронические инфекции и антигены опухолей. Ингибиторные рецепторы иммунных контрольных точек являются мишенями для иммунотерапии рака благодаря их потенциалу для применения при множестве типов рака.
LAG-3 представляет собой гомологичный CD4 мембранный белок I типа с четырьмя внеклеточными доменами суперсемейства Ig. Аналогично CD4, LAG-3 олигомеризуется на поверхностях Т-клеток и связывается с молекулами ГКГС II класса на АПК, но со значительно более высокой аффинностью, чем CD4. LAG-3 экспрессируется на активированных CD4-положительных и CD8-положительных Т-лимфоцитах, где он связывается с комплексом CD3-TCR на поверхности клетки и отрицательно регулирует передачу сигналов. Вследствие этого, он отрицательно регулирует пролиферацию, функцию и гомеостаз Т-клеток. Когда происходит распознавание комплекса ГКГС II класса с пептидом специфичным TCR, внутриклеточные сигналы в Т-клетке передаются посредством TCR и в АПК посредством молекул ГКГС II класса. Отрицательная регуляторная роль передачи сигнала LAG-3 в Т-клетки реализуется в ответах первичных CD4+ и CD8+ Т-клеток человека (Macon-Lemaitre, et al., Immunology. 2005 Jun; 115 (2): 170-178). Тем не менее, молекулярный механизм, посредством которого LAG-3 отрицательно регулирует передачу сигнала в Т-клетках, неизвестен. Для ингибиторной функции LAG-3 требуется его внутриклеточная (ВК) область, но данная область не содержит обычный сигнальный мотив с известным механизмом передачи сигнала. Maeda et al. (J. Biol. Chem. 2019, RA119.007455) недавно сообщили, что LAG-3 передает два независимых ингибиторных сигнала посредством мотива FxxL в области вблизи мембраны и С-концевого повтора EX. Тем не менее, ранее не сообщали о таких мотивах в ингибиторных корецепторах, и молекулярный механизм ингибиторного сигнала, который передает LAG-3, остается неясным.
В WO 2017/037203 описаны антитела (такие как гуманизированное моноклональное антитело IMP761), и их антигенсвязывающие фрагменты, которые являются агонистами LAG-3, и их применение для лечения состояний, связанных с пролиферацией и/или активацией CD4+ и/или CD8+ Т-клеток, в частности, воспалительных и аутоиммунных расстройств.
Обычные способы, используемые для измерения активности антитела и других лекарственных средств, разработанных для нацеливания на рецепторы иммунных контрольных точек, основаны на первичных клетках человека и измерении функциональных конечных точек, таких как пролиферация клеток, экспрессия маркеров на поверхности клеток и продукция цитокинов. Такие анализы трудоемки и высоковариабельны вследствие того, что в их основе лежат первичные клетки донора, сложные протоколы анализа и непроверенные на соответствие требованиям реагенты для анализа. В результате, данные анализы трудно наладить в условиях разработки лекарственных средств с контролем качества. Для того чтобы решить данные проблемы, компания Promega Corporation разработала клеточные биоанализы с использованием биолюминесцентного репортера для отдельных мишеней иммунотерапии - иммунных контрольных точек, - и их комбинаций (Cheng et al., June 2016, Promega, «Quantitative Cell-Based Bioassays for Indivisual and Combination Immune Checkpoint Immunotherapy Targets»).
Биоанализы, разработанные Promega, включают биоанализы блокирования PD-1, CTLA-4, LAG-3 и TIGIT. В основе данных анализов лежит применение Т-клеток Jurkat, которые подвергли генетической инженерии, чтобы они экспрессировали на поверхности интересующий коингибиторный рецептор, и которые содержат репортерный ген люциферазы светлячка под контролем элемента отклика NFAT (NFAT-RE). Эти клетки экспрессируют эндогенные рецепторы TCR, CD3 и CD28. Когда указанные клетки связываются с соответствующим лигандом, TCR передает внутриклеточные сигналы, приводящие к повышенной опосредованной NFAT-RE люминесценции. Биолюминесцентный сигнал детектируют и проводят количественный анализ, используя субстрат люциферазы и стандартный люминометр. В указанных анализах также используют искусственные антигенпрезентирующие клетки (иАПК), экспрессирующие сконструированный белок поверхности клетки, разработанный таким образом, чтобы он активировал TCR независимым от антигена способом, и экспрессирующие на своей поверхности природный лиганд для коингибиторного рецептора (для биоанализа блокирования PD-1, CTLA-4 и TIGIT), или клетки Raji в присутствии суперантигена стафилококкового энтеротоксина Е (SEE) (для биоанализа блокирования LAG-3 - клетки Raji с естественной экспрессией ГКГС II класса - лиганд LAG-3). Связывание TCR с клетками Jurkat вызывает активность люциферазы. Совместное связывание коингибиторного рецептора с его природным лигандом ингибирует активность люциферазы. Опосредованное антителами блокирование связывания коингибиторного рецептора с его природным лигандом восстанавливает активность люциферазы.
Указанные биоанализы демонстрируют эффективность, необходимую для применения для скрининга для выявления антагонистического антитела, тестирования эффективности и исследований стабильности. Тем не менее, они не подходят для тестирования агонистических антител, таких как агонистическое антитело LAG-3 (например, IMP761, описанное в WO 2017/037203). В частности, присутствие природного лиганда для LAG-3 в биоанализе блокирования (такого как ГКГС II класса, экспрессированный клетками Raji) будет препятствовать тестированию агониста. Следовательно, есть потребность в анализах in vitro для идентификации и тестирования агонистов LAG-3.
В рамках настоящей заявки неожиданно было обнаружено, что IMP761, посредством агонизма LAG-3, ингибирует передачу сигнала TCR, в частности, регулируемую NFAT экспрессию генов, в LAG-3 положительных Т-клетках. Автор настоящего изобретения понял, что это может стать основой для биоанализов in vitro для определения активности агониста LAG-3, или получения агониста LAG-3, и для идентификации новых агонистов LAG-3.
В соответствии с настоящим изобретением предложен анализ (способ анализа) in vitro для определения активности агониста гена активации лимфоцитов 3 (LAG-3), который включает:
обеспечение множества эффекторных Т-клеток, каждая эффекторная Т-клетка экспрессирует на своей поверхности LAG-3 и Т-клеточный рецептор (TCR) и содержит репортерный ген, кодирующий репортер, причем экспрессия репортера регулируется опосредованным LAG-3 ингибированием передачи сигнала TCR в эффекторных Т-клетках; и
определение активности агониста по степени, до которой изменяется экспрессия репортера (т.е., повышается или снижается) в присутствии агониста, по сравнению с экспрессией репортера в отсутствие агониста.
Анализы в соответствии с настоящим изобретением для определения активности агониста включают анализы для определения эффективности препарата агониста, например, в рамках этапа контроля качества при получении агониста (в частности, такого как клеточный анализ эффективности, необходимый для выпуска продукта в соответствии с надлежащей производственной практикой, GMP), или для проверки стабильности препарата агониста, например, после некоторого периода хранения, или в качестве анализа для определения характеристик продукта.
В соответствии с настоящим изобретением также предложен анализ in vitro для скрининга в отношении агониста LAG-3, который включает:
обеспечение множества эффекторных Т-клеток, каждая эффекторная Т-клетка экспрессирует на своей поверхности LAG-3 и Т-клеточный рецептор (TCR) и содержит репортерный ген, кодирующий репортер, причем экспрессия репортера регулируется опосредованным LAG-3 ингибированием передачи сигнала TCR в эффекторных Т-клетках; и
определение того, является ли агонист-кандидат агонистом LAG-3, путем определения степени, до которой изменяется экспрессия репортера (т.е., повышается или снижается) в присутствии агониста-кандидата, по сравнению с экспрессией репортера в отсутствие агониста-кандидата.
Анализы в соответствии с настоящим изобретением для скрининга в отношении агониста LAG-3 можно применять, чтобы идентифицировать агонист LAG-3, например, из библиотеки агонистов-кандидатов. Такими агонистами-кандидатами могут быть лекарственные средства (например, синтетические низкомолекулярные соединения), или биологические агенты, такие как рекомбинантные или природные белки, антитела или их фрагменты или производные.
Репортер может экспрессироваться в эффекторных Т-клетках в отсутствие агониста, или агониста-кандидата.
Необязательно экспрессия репортера снижается в присутствии агониста, или агониста-кандидата, по сравнению с экспрессией репортера в отсутствие агониста, или агониста-кандидата.
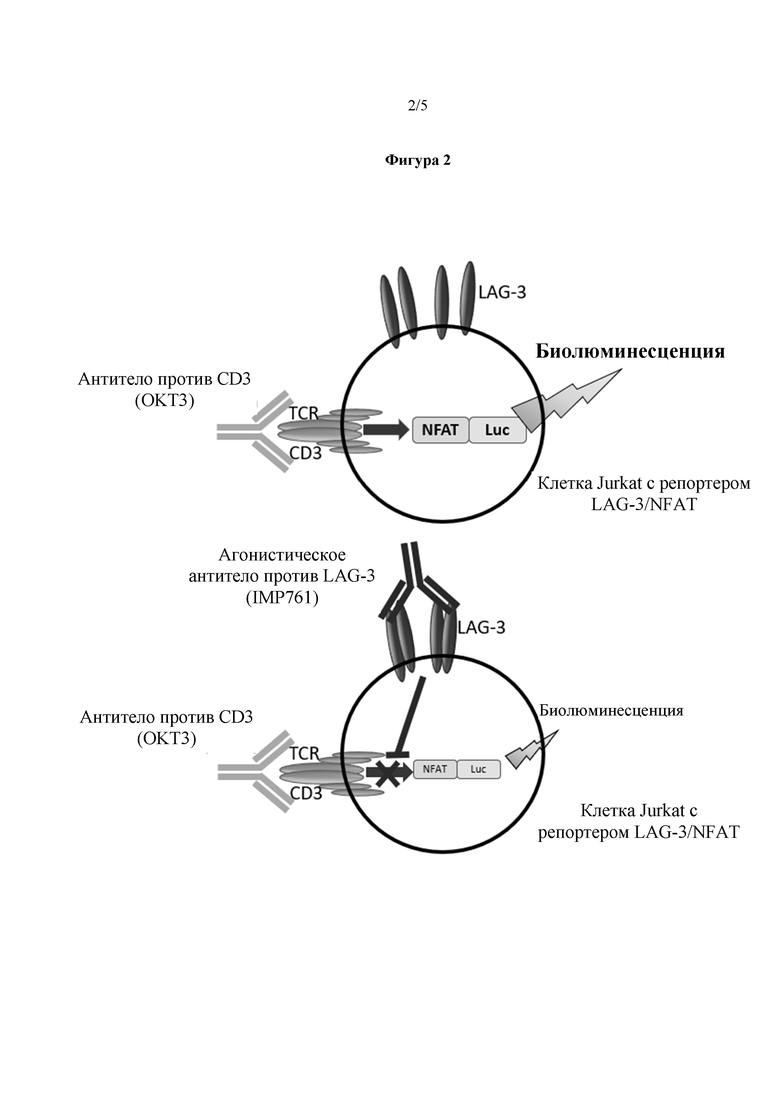
Например, опосредованное LAG-3 ингибирование передачи сигнала TCR в эффекторных Т-клетках может вызывать снижение базального уровня экспрессии репортера в эффекторных Т-клетках (см. Фигуру 2(a)). Таким образом, активность агониста LAG-3 (включая эффективность препарата агониста LAG-3) можно определить, или агонист LAG-3 можно идентифицировать, путем определения степени, до которой снижается базальный уровень экспрессии репортера в присутствии агониста, или агониста-кандидата, по сравнению с базальным уровнем экспрессии репортера в отсутствие агониста, или агониста-кандидата.
Формулировку «опосредованное LAG-3 ингибирование передачи сигнала TCR в эффекторных Т-клетках» используют в данной заявке в том смысле, что агонизм LAG-3, экспрессированного на поверхности эффекторных Т-клеток, ингибирует опосредованную TCR передачу сигнала в эффекторных Т-клетках с последующим изменением (т.е., повышением или снижением) экспрессии репортера. Можно ингибировать любой опосредованный TCR путь передачи сигналов в эффекторных Т-клетках. Например, экспрессия репортерного гена может находиться под контролем промотора или элемента отклика (RE), который отвечает на связывание фактора транскрипции, который является участником сигнального пути. Например, можно ингибировать сигнальный путь кальциневрина/NFAT в эффекторных Т-клетках. В частности, экспрессия репортерного гена может находиться под контролем элемента отклика NFAT, так что снижение передачи сигнала кальциневрина/NFAT, происходящее в результате агонизма LAG-3, вызывает снижение экспрессии репортера с репортерного гена.
Экспрессия репортера может измениться (т.е., повыситься или снизиться) в каждой эффекторной Т-клетке в ответ на активацию эффекторной Т-клетки посредством TCR. Необязательно анализы в соответствии с настоящим изобретением дополнительно включают: активацию эффекторных Т-клеток путем независимой от антигена, независимой от ГКГС II класса, опосредованной TCR активации Т-клеток в присутствии и отсутствие агониста, или агониста-кандидата; и определение активности (включая эффективность препарата) агониста, или агониста-кандидата, по степени, до которой изменяется экспрессия репортера в ответ на активацию эффекторных Т-клеток в присутствии агониста, или агониста-кандидата, по сравнению с экспрессией репортера в ответ на активацию эффекторных Т-клеток в отсутствие агониста, или агониста-кандидата.
Таким образом, в соответствии с настоящим изобретением предложен анализ in vitro для определения активности агониста гена активации лимфоцитов 3 (LAG-3), который включает:
обеспечение множества эффекторных Т-клеток, каждая эффекторная Т-клетка экспрессирует на своей поверхности LAG-3 и Т-клеточный рецептор (TCR) и содержит репортерный ген, кодирующий репортер, экспрессия которого изменяется в ответ на активацию эффекторной Т-клетки посредством TCR, причем экспрессия репортера регулируется опосредованным LAG-3 ингибированием передачи сигнала TCR в эффекторных Т-клетках;
активацию эффекторных Т-клеток путем независимой от антигена, независимой от ГКГС II класса, опосредованной TCR активации Т-клеток в присутствии и отсутствие агониста; и
определение активности агониста по степени, до которой изменяется экспрессия репортера в ответ на активацию эффекторных Т-клеток в присутствии агониста, по сравнению с экспрессией репортера в ответ на активацию эффекторных Т-клеток в отсутствие агониста.
В соответствии с настоящим изобретением также предложен анализ in vitro для скрининга в отношении агониста LAG-3, который включает:
обеспечение множества эффекторных Т-клеток, каждая эффекторная Т-клетка экспрессирует на своей поверхности LAG-3 и Т-клеточный рецептор (TCR) и содержит репортерный ген, кодирующий репортер, экспрессия которого изменяется в ответ на активацию эффекторной Т-клетки посредством TCR, причем экспрессия репортера регулируется опосредованным LAG-3 ингибированием передачи сигнала TCR в эффекторных Т-клетках;
активацию эффекторных Т-клеток путем независимой от антигена, независимой от ГКГС II класса, опосредованной TCR активации Т-клеток в присутствии и отсутствие агониста-кандидата; и
определение активности агониста-кандидата по степени, до которой изменяется экспрессия репортера в ответ на активацию эффекторных Т-клеток в присутствии агониста-кандидата, по сравнению с экспрессией репортера в ответ на активацию эффекторных Т-клеток в отсутствие агониста-кандидата.
Необязательно экспрессия репортера повышается в каждой эффекторной Т-клетке в ответ на активацию эффекторной Т-клетки посредством TCR, и снижается в присутствии агониста, или агониста-кандидата, в результате опосредованного LAG-3 ингибирования передачи сигнала TCR в эффекторной Т-клетке, и при этом активность агониста, или агониста-кандидата, определяют по степени, до которой снижается экспрессия репортера в ответ на активацию эффекторных Т-клеток в присутствии агониста, или агониста-кандидата, по сравнению с экспрессией репортера в ответ на активацию эффекторных Т-клеток в отсутствие агониста, или агониста-кандидата.
Чем больше степень, до которой изменяется экспрессия репортера в ответ на активацию эффекторных Т-клеток в присутствии агониста, или агониста-кандидата, по сравнению с экспрессией репортера в ответ на активацию эффекторных Т-клеток в отсутствие агониста, или агониста-кандидата, тем больше активность (или эффективность препарата) агониста, или агониста-кандидата, по отношению к LAG-3.
Необязательно эффекторные Т-клетки активируют путем бесклеточной активации Т-клеток.
Термин «бесклеточная активация Т-клеток» в данной заявке используют по отношению к активации эффекторных Т-клеток путем применения одного или более бесклеточных активаторов Т-клеток в отсутствие любых клеток, отличных от эффекторных Т-клеток. В частности, бесклеточная активация Т-клеток происходит без использования антигенпрезентирующей клетки (АПК), искусственной АПК (иАПК) или любой другой клетки, экспрессирующей ГКГС I класса или ГКГС II класса, такой как клетка Raji.
Бесклеточная активация Т-клеток предпочтительна, так как для клеточных реагентов требуются особые условия хранения. Клетки обычно хранят в замороженном состоянии, и их затем необходимо разморозить перед применением.
Формулировка «активация эффекторной Т-клетки посредством TCR», используемая в данной заявке, означает, что после связывания TCR активатором Т-клеток передается сигнал посредством TCR в эффекторной Т-клетке, приводящий к изменению (т.е., повышению или снижению) экспрессии репортера в эффекторной Т-клетке. Например, экспрессия с репортерного гена может находиться под контролем промотора или элемента отклика (RE), который отвечает на сигнал активации эффекторных Т-клеток. Например, с элементом отклика может связаться один или более факторов транскрипции после активации эффекторной Т-клетки посредством TCR.
Необязательно эффекторные Т-клетки активируют путем приведения в контакт эффекторных Т-клеток с независимым от антигена, независимым от ГКГС II класса активатором Т-клеток при условиях, подходящих для независимой от антигена, независимой от ГКГС II класса, опосредованной TCR активации эффекторных Т-клеток активатором Т-клеток.
В соответствии с настоящим изобретением также предложен набор для проведения анализа in vitro для определения активности, или для скрининга в отношении агониста LAG-3, который содержит: множество эффекторных Т-клеток, каждая эффекторная Т-клетка экспрессирует на своей поверхности LAG-3 и Т-клеточный рецептор (TCR) и содержит репортерный ген, кодирующий репортер, причем экспрессия репортера регулируется опосредованным LAG-3 ингибированием передачи сигнала TCR в эффекторных Т-клетках, причем экспрессия репортера изменяется (т.е., повышается или снижается) в каждой эффекторной Т-клетке в ответ на активацию эффекторной Т-клетки посредством TCR; и активатор Т-клеток, способный к независимой от антигена, независимой от ГКГС II класса, опосредованной TCR активации эффекторных Т-клеток.
Независимые от антигена, независимые от ГКГС II класса активаторы Т-клеток отличаются от с упер антигенов, таких как стафилококковые энтеротоксины (SE). SE содержат участки связывания с ГКГС II класса и TCR. Они связываются сначала с ГКГС II класса, затем с вариабельной областью цепи альфа или бета TCR. Взаимодействие с Т-клеточным рецептор позволяет SE действовать как «клин» между TCR и ГКГС. Это приводит к вытеснению любого антигенного пептида прочь от TCR и позволяет обойти нормальный механизм активации Т-клеток. SE, следовательно, представляют собой зависимые от ГКГС II класса активаторы Т-клеток.
Необязательно эффекторные Т-клетки приводят в контакт с концентрацией активатора Т-клеток, при которой происходит максимальное ингибирование экспрессии репортера в присутствии избытка агониста LAG-3. Использование такой концентрации активатора Т-клеток оптимизирует точность анализа.
Автор настоящего изобретения обнаружил, что концентрация активатора Т-клеток, при которой происходит максимальное ингибирование экспрессии репортера в присутствии избытка агониста LAG-3, представляет собой субоптимальную концентрацию активатора Т-клеток (т.е., концентрацию, которая меньше, чем концентрация активатора Т-клеток, при которой наблюдается наибольшая экспрессия репортера в ответ на активацию эффекторных Т-клеток активатором Т-клеток в отсутствие агониста, или агониста-кандидата).
Необязательно активатор Т-клеток приводят в контакт с эффекторными Т-клетками при концентрации, которая меньше, чем концентрация активатора Т-клеток, при которой наблюдается наибольшая экспрессия репортера в ответ на активацию эффекторных Т-клеток активатором Т-клеток в отсутствие агониста, или агониста-кандидата.
Должно быть понятно, что зависимая от антигена стимуляция распространяется только на антигенспецифичные Т-клетки, тогда как независимая от антигена стимуляция распространяется на до 100% эффекторных Т-клеток.
Примером активатора Т-клеток, способного к независимой от антигена, независимой от ГКГС II класса, опосредованной TCR активации эффекторных Т-клеток, является антитело против CD3. Антитело против CD3 связывается с CD3 и активирует комплекс TCR без антигенного пептида от АПК (т.е., независимая от антигена, независимая от ГКГС II класса, опосредованная TCR стимуляция эффекторных Т-клеток) (см. Фигуру 2(b)). Антитела против CD3 могут активировать до 100% эффекторных Т-клеток.
Необязательно активатор Т-клеток включает антитело против CD3, или его фрагмент или производное, у которого сохраняется способность к независимой от антигена, независимой от ГКГС II класса, опосредованной TCR активации эффекторных Т-клеток. Подходящие примеры антитела против CD3 включают OKT3 и UCHT1. Необязательно антитело против CD3 представляет собой OKT3.
Необязательно антитело против CD3, или его фрагмент или производное, приводят в контакт с эффекторными Т-клетками в концентрации ~6-30×10-12 М (1-4 нг/мл для полноразмерного антитела, или молярный эквивалент для его фрагмента или производного).
Необязательно набор в соответствии с настоящим изобретением содержит антитело против CD3, или его фрагмент или производное, у которого сохраняется способность к независимой от антигена, независимой от ГКГС II класса, опосредованной TCR активации эффекторных Т-клеток, при концентрации, позволяющей его применение в анализе в концентрации ~6-30×10-12 М (1-4 нг/мл для полноразмерного антитела). Например, набор может содержать одну или более аликвот антитела против CD3, или его фрагмента или производного, в концентрации ~6-30×10-12 М (1-4 нг/мл для полноразмерного антитела).
Необязательно эффекторные Т-клетки приводят в контакт с несколькими различными концентрациями агониста или агониста-кандидата. Например, можно провести параллельно множество различных анализов, в которых эффекторные Т-клетки для каждого отличного анализа приводят в контакт с отличной концентрацией агониста или агониста-кандидата.
Необязательно определяли значение IC50 агониста-кандидата или агониста для ингибирования экспрессии репортера. Это можно осуществить, например, по кривой доза-ответ, построенной по результатам, полученным путем приведения в контакт эффекторных Т-клеток с несколькими различными концентрациями агониста-кандидата или агониста.
Необязательно анализ в соответствии с настоящим изобретением дополнительно включает проведение анализа отрицательного контроля (например, параллельно с указанным анализом). Например, эффекторные Т-клетки можно привести в контакт с активатором Т-клеток при условиях, подходящих для независимой от антигена активации эффекторных Т-клеток активатором Т-клеток в отсутствие агониста, или агониста-кандидата, но в присутствии молекулы того же типа, что и агонист или агонист-кандидат, но у которой, как известно, отсутствует агонистическая активность по отношению к LAG-3 (т.е., в качестве отрицательного контроля).
Например, если агонист или агонист-кандидат представляет собой антитело, необязательно молекула для применения в качестве отрицательного контроля также представляет собой антитело. Предпочтительно антитело отрицательного контроля представляет собой антитело того же изотипа, что и агонистическое или агонистическое антитело-кандидат.
Необязательно активатор Т-клеток представляет собой бесклеточный активатор Т-клеток.
Необязательно набор в соответствии с настоящим изобретением не содержит клетку, экспрессирующую молекулы ГКГС II класса.
Необязательно набор в соответствии с настоящим изобретением не содержит клетки Raji.
Необязательно набор в соответствии с настоящим изобретением не содержит АПК, иАПК или любые другие клетки, экспрессирующие ГКГС I класса или ГКГС II класса, такие как клетки Raji.
Необязательно единственными клетками в наборе в соответствии с настоящим изобретением являются эффекторные Т-клетки.
Необязательно эффекторные Т-клетки в наборе в соответствии с настоящим изобретением представляют собой замороженные клетки, например, клетки "Разморозь и используй".
В соответствии с настоящим изобретением также предложен набор в соответствии с настоящим изобретением для проведения анализа в соответствии с настоящим изобретением.
Необязательно репортерный ген представляет собой гетерологичный репортерный ген.
Генетические репортеры широко применяют в качестве индикаторов для исследования экспрессии генов и клеточных событий, связанных с экспрессией генов, для фармацевтических и биомедицинских исследований. Обычно, репортерный ген, кодирующий репортер, клонируют в вектор экспрессии, который затем переносят в клетки. После переноса анализируют присутствие репортера в клетках путем непосредственного измерения самого белка репортера или ферментативной активности белка репортера. Предпочтительными репортерами являются такие, которые можно легко обнаружить, и провести их количественный анализ, когда они экспрессируются в эффекторных Т-клетках. Множество подходящих примеров известно специалистам в данной области, включая флуоресцентные и люминесцентные репортеры. Необязательно репортер представляет собой биолюминесцентный репортер, такой как люцифераза.
Биолюминесценция представляет собой особую форму хемилюминесценции, обнаруженной в живых организмах. Данный класс хемилюминесценции является процессом, катализируемым ферментом, при котором высокая эффективность испускания протона возникла в результате естественной эволюции. Данные ферменты называют люциферазами, а испускающие фотон субстраты называют люциферинами. Биолюминесцентные химические вещества эволюционировали из множества независимых источников и включают множество различных молекулярных структур. Из множества природных форм, две широко применяют в генетических репортерных анализах: люциферазу светлячка и люцифераза Renilla (Fan, and Wood (2007) Bioluminescent assays for high-throughput screening. Assay Drug Dev. Technol. 5, 127-36).
Анализы с биолюминесцентными репортерными генами имеют явное преимущество над флуоресцентными анализами, такими как анализ с использованием зеленого флуоресцентного белка (GFP), состоящее в том, что чувствительность таких анализов может быть в 10-1000 раз выше. Как при флуоресценции, так и при люминесценции вырабатываются фотоны вследствие энергетических переходов с молекулярных орбиталей в возбужденном состоянии на орбитали с меньшей энергией. Тем не менее, они отличаются тем, как образуются орбитали с возбужденным состоянием. При люминесценции возбужденные состояния возникают в результате экзотермических химических реакций, тогда как при флуоресценции возбужденные состояния возникают в результате поглощения света. В репортерных анализах предпочтительна биолюминесценция, так как не требуются фотоны для создания возбужденного состояния. Следовательно, они не создают характерный фон при измерении испускания фотонов образцом. В результате, низкий уровень фона позволяет точно измерить небольшие изменения излученного света от четырех до восьми порядков степени в линейном диапазоне анализа.
Технология люциферазного репортера основана на взаимодействии фермента люциферазы с люминесцентным субстратом люциферином, который высвобождает свет в процессе биолюминесценции. Биолюминесценция обнаружена во множестве различных организмов; тем не менее, именно люцифераза светлячка (Photinus pyralis), несомненно, является наиболее часто применяемым биолюминесцентным репортером благодаря как чувствительности, так и удобству анализа фермента, а также сильной взаимосвязи синтеза белка с активностью фермента. Данный мономерный фермент с молекулярной массой 61 кДа катализирует двухстадийную реакцию окисления с образованием света, обычно в области спектра от зеленого до желтого, обычно 550-570 нм. Ген, кодирующий люциферазу светлячка (luc), представляет собой кДНК, и ему не требуются какие-либо посттрансляционные модификации. Это означает, что люцифераза доступна в виде зрелого фермента сразу после трансляции с ее мРНК.
Подходящие репортерные гены, кодирующие люциферазу, доступны для приобретения у нескольких компаний, включая Promega.
После создания взаимосвязи функциональных регуляторных элементов с экспрессией гена люциферазы, обычно путем размещения регуляторного элемента непосредственно против хода транскрипции от гена, кодирующего люциферазу, активацию эффекторных Т-клеток можно легко детектировать по люминесцентному сигналу. Обычно репортерные гены располагают по ходу транскрипции от клонированных элементов отклика.
Необязательно репортерный ген находится под контролем промотора или элемента отклика. Подходящий элемент отклика включает элемент отклика NFAT (ядерного фактора активированных Т-клеток) (NFAT-RE).
Выше объясняли, что стимуляция TCR вызывает внутриклеточное высвобождение кальция и активацию кальциневрина, который дефосфорилирует цитоплазматический ядерный фактор активированных Т-клеток (NFAT). Дефосфорилированный NFAT перемещается в ядро и связывается с NFAT-RE, вызывая транскрипцию репортерного гена.
Необязательно эффекторные Т-клетки содержат гетерологичную нуклеиновую кислоту, содержащую репортерный ген.
Необязательно эффекторные Т-клетки содержат гетерологичную нуклеиновую кислоту, кодирующую LAG-3.
Термин "гетерологичный" репортерный ген или нуклеиновая кислота используют в данной заявке как означающий репортерный ген или нуклеиновую кислоту, которые в природе не присутствуют в эффекторных Т-клетках, но которые внедрили в указанные эффекторные Т-клетки или в эффекторные Т-клетки, из которых происходят указанные эффекторные Т-клетки, например, с помощью клонирования, рекомбинантной технологии или трансфекции (методик, которые хорошо известны квалифицированному специалисту).
Необязательно осуществляют двойную трансфекцию эффекторных Т-клеток гетерологичной нуклеиновой кислотой, содержащей репортерный ген (необязательно при этом репортерный ген функционально связан с промотором или элементом отклика, чтобы управлять экспрессией репортерного гена в эффекторной Т-клетке) и гетерологичную нуклеиновую кислоту, кодирующую LAG-3.
Необязательно набор в соответствии с настоящим изобретением дополнительно содержит молекулу того же типа, что и агонист или агонист-кандидат, но у которой, как известно, отсутствует агонистическая активность по отношению к LAG-3, для применения в качестве отрицательного контроля.
Например, если агонист или агонист-кандидат представляет собой антитело, необязательно молекула для применения в качестве отрицательного контроля также представляет собой антитело. Предпочтительно антитело отрицательного контроля представляет собой антитело того же изотипа, что и агонистическое или агонистическое антитело-кандидат.
Необязательно набор в соответствии с настоящим изобретением дополнительно содержит известный агонист LAG-3 для применения в качестве положительного контроля. Например, известный агонист может представлять собой агонистическое антитело, или его фрагмент или производное, у которого сохраняется агонистическая активность.
Должно быть понятно, что анализ в соответствии с настоящим изобретением следует проводить в отсутствие природного лиганда для LAG-3, поскольку природный лиганд, в противном случае, будет искажать результаты анализа.
Необязательно набор в соответствии с настоящим изобретением не содержит природный лиганд для LAG-3.
Эффекторные Т-клетки включают несколько типов Т-клеток, которые активно отвечают на стимул, такой как костимуляция. Они включают CD4+, CD8+ и регуляторные Т-клетки. Подходящим примером является клетка Jurkat. Клетки Jurkat представляют собой иммортализованные Т-лимфоциты, изначально полученные из периферической крови ребенка с Т-клеточным лейкозом (Schneider et al., 1977, Int J Cancer 19 (5): 621-6).
Необязательно агонист представляет собой агонистическое антитело против LAG-3, или его фрагмент или производное, у которого сохраняется агонистическая активность по отношению к LAG-3.
Необязательно эффекторные Т-клетки включают линию Т-клеток, которую подвергли двойной трансфекции нуклеиновой кислотой, кодирующей LAG-3, и репортерным геном под контролем промотора или элемента отклика, например, линию клеток Jurkat, которую подвергли двойной трансфекции нуклеиновой кислотой, кодирующей LAG-3, и репортерным геном NFAT/люцифераза (клетки Jurkat LAG-3+/NFAT-luc). Клетки Jurkat LAG3+/NFAT-luc2 доступны для приобретения у Promega (номер в каталоге CS194801). Клетки Jurkat LAG3+/NFAT-luc доступны для приобретения у BPS Bioscience (номер в каталоге 71278).
Необязательно агонист представляет собой агонистическое антитело против LAG-3, или его фрагмент или производное, у которого сохраняется агонистическая активность по отношению к LAG-3, и эффекторные Т-клетки включают клетки Jurkat LAG-3+/NFAT-luc2.
Необязательно агонистическое антитело против LAG-3 представляет собой агонистическое антитело против LAG-3, описанное в WO 2017/037203. Агонистические антитела против LAG-3, описанные в WO 2017/037203, включают моноклональное антитело мыши 13Е2 и гуманизированное антитело 13E2-Fc IgG4 человека (обозначенное IMP761). Антитела 13Е2 и IMP761 содержат последовательности CDR1-3 VH и CDR1-3 VL, представленные в Таблице 1 ниже:
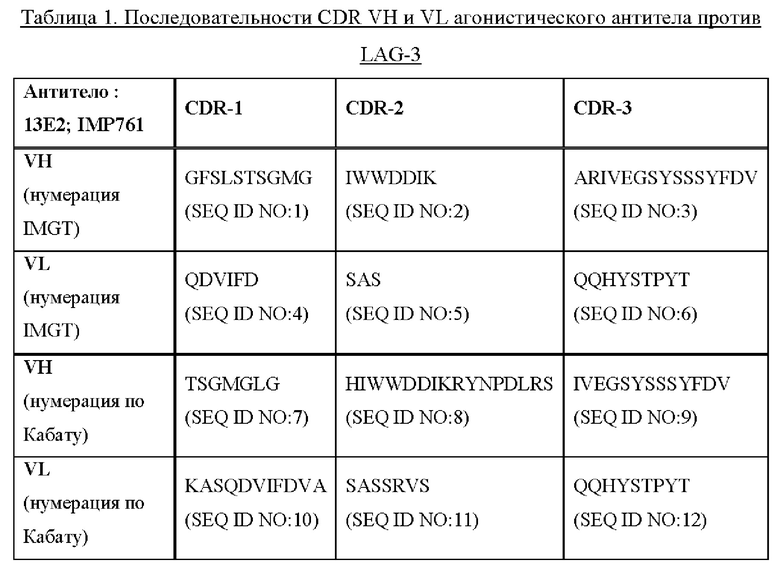
Необязательно агонистическое антитело против LAG-3, или его фрагмент или производное, содержит последовательности CDR1-3 VH и CDR1-3 VL, представленные в SEQ ID NO: 1-6, соответственно, или последовательности CDR1-3 VH и CDR1-3 VL, представленные в SEQ ID NO: 7-12, соответственно:
Необязательно агонистическое антитело против LAG-3 представляет собой IMP761.
Варианты реализации настоящего изобретения описаны ниже, исключительно в качестве примера, с отсылкой к сопроводительным фигурам, описанным далее.
На Фигуре 1 показано схематическое представление передачи сигнала TCR (из Belikov, Aleksey. (2016). The role of reactive oxygen species and mitochondria in T-cell activation. 10.13140/RG. 2.1.2916.0568);
На Фигуре 2 показан механизм действия анализа эффективности IMP761 в соответствии с вариантом реализации настоящего изобретения. В присутствии антитела против CD3, антитело против CD3 связывается с CD3 на поверхности репортерной клетки Jurkat, приводя к повышенной экспрессии люциферазы с репортерного гена NFAT/Luc вследствие опосредованной TCR передачи сигнала (верхняя часть). Связывание IMP761 с LAG-3 на поверхности репортерной клетки Jurkat ингибирует опосредованную TCR передачу сигнала, приводя к снижению экспрессии люциферазы с репортерного гена NFAT/Luc (нижняя часть);
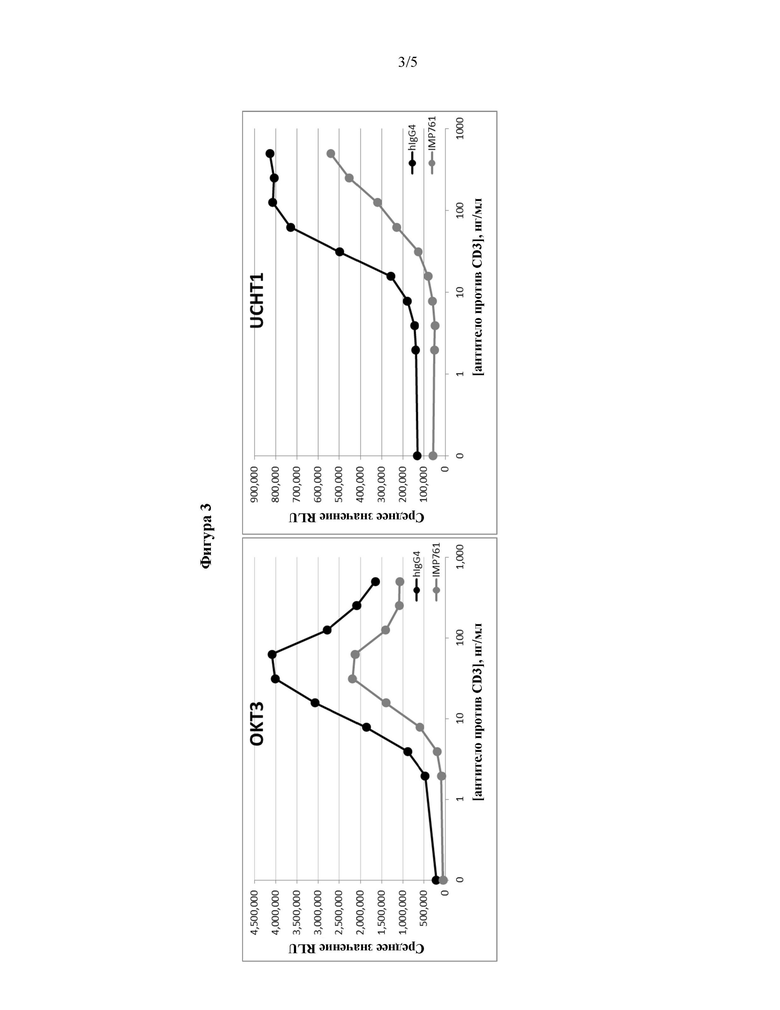
На Фигуре 3 показано влияние стимуляции клеток Jurkat LAG-3+/NFAT-luc2 различными концентрациями антител против CD3 OKT3 и UCHT1 в присутствии 300 нг/мл IMP761 или антитела IgG4 человека отрицательного контроля; и
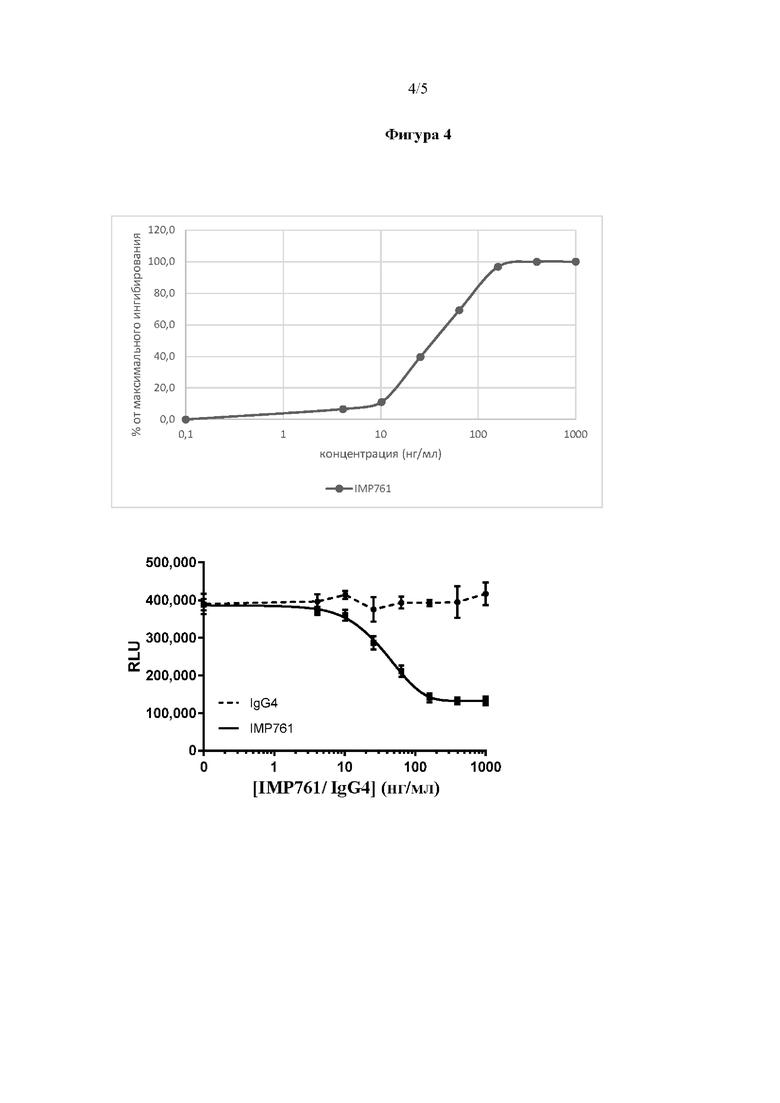
На Фигуре 4 показан пример результатов анализа эффективности IMP761 по сравнению с отрицательным контролем IgG4, в соответствии с вариантом реализации настоящего изобретения; и
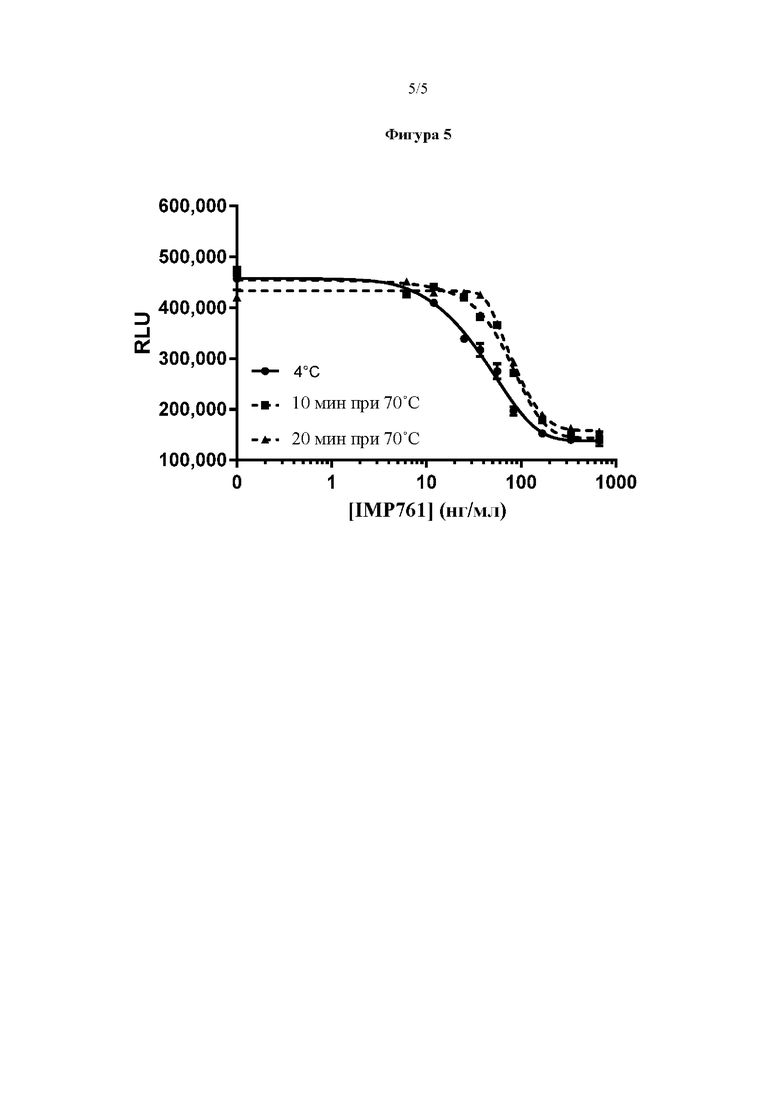
На Фигуре 5 показан пример результатов анализа эффективности с референсным препаратом IMP761 (4°С) по сравнению с препаратом IMP761 после денатурирующего температурного стресса (10 или 20 минут при 70°С), в соответствии с вариантом реализации настоящего изобретения.
Пример 1
Оптимизация протокола анализа эффективности IMP761
Эффекторные клетки Jurkat Lag-3+/NFAT-luc2 были изначально разработаны в Promega для определения активности антагонистического антитела против LAG-3 после активации TCR посредством стимуляции супер антигеном, презентированным молекулами ГКГС II. Блокирование антагонистическим антителом против LAG-3 взаимодействия LAG-3/ГКГС II приводит к повышению активации TCR и активности люциферазы. Для применения эффекторных клеток Jurkat Lag-3+/NFAT-luc2 для определения активности агонистического антитела против LAG-3 требуется совсем другая постановка эксперимента, что объяснено ниже.
Антитело против CD3 в качестве стимулятора клеток Jurkat.
В биоанализе Promega эффекторные клетки Jurkat Lag-3+/NFAT-luc2 активируют, применяя клетки Raji, в присутствии стафилококкового энтеротоксина Е или D (SEE или SED). Клетки Raji экспрессируют эндогенный ГКГС II класса -лиганд LAG-3. Важно проверять блокирующую активность антагонистического антитела против LAG-3 на взаимодействии LAG-3/ГКГС II. Тем не менее, так как не требуется взаимодействие LAG-3/ГКГС II для тестирования эффективности агонистического антитела против LAG-3, то не требуются ни клетки Raji, ни стафилококковый энтеротоксин. Использовали тип анализа на отдельных клетках с антителами против CD3, чтобы активировать эффекторные клетки Jurkat Lag-3+/NFAT-luc2 посредством передачи сигнала TCR.
Концентрация антитела против CD3, и связанное с LAG-3 ингибирование.
Исследовали влияние на анализ эффективности клеток двух различных антител против CD3 (OKT3 и UCHT1) при различных концентрациях антитела в диапазоне от 1 до 500 нг/мл.
Клетки Jurkat Lag-3+/NFAT-luc2 инкубировали с 300 нг/мл IMP761 или IgG4 человека (в качестве отрицательного контроля) в присутствии различных концентраций OKT3 или UCHT1 в течение 24 часов. Средние значения RLU, полученные для различных концентраций антитела против CD3, показаны ниже на Фигуре 3 и в Таблице 2.
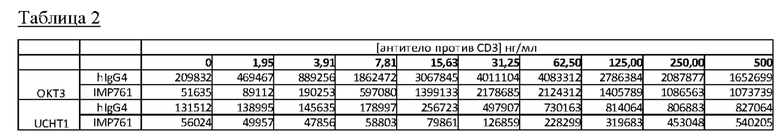
Люциферазная активность ингибируется IMP761 во всем диапазоне различных концентраций каждого антитела против CD3.
В линии клеток Jurkat без стимуляции присутствует базальный уровень экспрессии репортера, так что активность люциферазы и ингибирование активности люциферазы посредством IMP761 также исследовали в отсутствие антитела против CD3. Тем не менее, стимуляция клеток Jurkat антителом против CD3 приводит к более высоким значениям RLU, так что ингибиторное действие IMP761 более явно в присутствии антитела против CD3 (в частности, низкой концентрации антитела против CD3). Процент ингибирования активности люциферазы для антитела OKT3 показан ниже в Таблице 3.
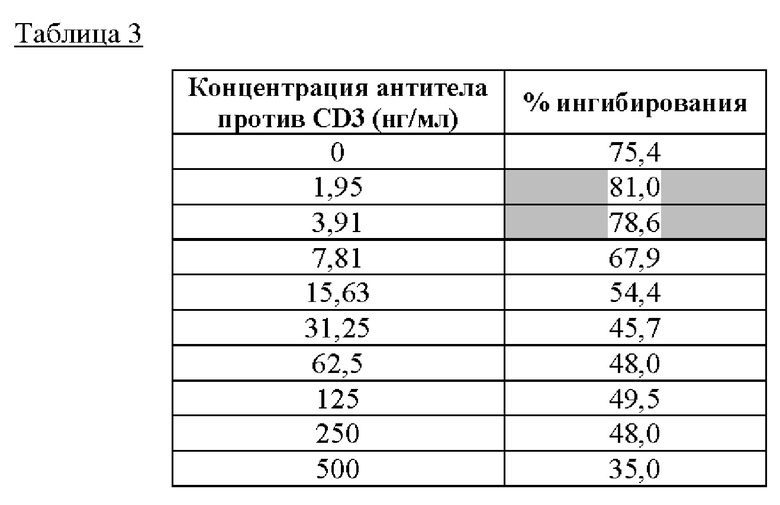
Пришли к заключению, что максимальный эффект IMP761 (ингибирование приблизительно на 80%) наблюдали при применении низкой концентрации антитела OKT3 (от 1 до 4 нг/мл). Таким образом, оптимальный анализ эффективности включает стимуляцию клеток Jurkat низкими концентрациями антитела против CD3 (например, антитела OKT3).
Пример 2
Анализ эффективности IMP761
В данном примере описан анализ эффективности в соответствии с вариантом реализации настоящего изобретения для измерения активности моноклонального антитела IMP761 in vitro. Указанный способ основан на способности IMP761 снижать активацию линии эффекторных клеток с LAG-3, индуцированную низкой дозой антитела против CD3 (клон OKT3, 3 нг/мл), имитируя стимуляцию антигеном Т-клеток. Линия эффекторных клеток с LAG-3 представляет собой линию Т-клеток Jurkat, экспрессирующих на своей поверхности LAG-3 и содержащих ген люциферазы под контролем элемента отклика NFAT (ядерного фактора активированных Т-клеток) (эффекторные клетки Jurkat Lag-3+/NFAT-luc2 от Promega). После связывания со своей мишенью, IMP761 запускает понижающую регуляцию индуцированной TCR регулируемой NFAT экспрессии (схематически проиллюстрировано на Фигуре 2, нижняя часть). Активность люциферазы в линии клеток использовали для измерения запускаемой TCR активации клеток, которая угнеталась в присутствии активности IMP761. Таким образом, в этом анализе измеряют эффективность IMP761 в отношении его способности ингибировать передачу сигнала TCR.
Реагенты
Эффекторные клетки Jurkat LAG-3+/NFAT-luc2 (Promega, номер в каталоге CS194801)
RPMI 1640 (GIBCO, номер в каталоге 31870-025)
L-глутамин (200 мМ) (GIBCO, номер в каталоге 25030-024)
HEPES (1 М) (GIBCO, номер в каталоге 15630-080)
Эмбриональная телячья сыворотка (ЭТС, GIBCO, номер в каталоге 10270106)
IMP761 (2,06 мг/мл) (IMMUTEP, партия: 270416)
IgG4 человека, контроль (Biolegend, номер в каталоге 403402)
Антитело против CD3 (OKT3) (eBioscience, номер в каталоге 16-0037-85)
Реагент Bio-Glo (PROMEGA, номер в каталоге G7941)
Твердые белые 96-луночные микропланшеты с плоским дном лунок (COSTAR, номер в каталоге 3917)
Аналитическая среда: RPMI 1640, L-глутамин (2 мМ), Hepes (10 мМ), ЭТС 1%
Концентрация клеток: 1,33×106 клеток/мл
Протокол
1. Пересеять эффекторные клетки Jurkat LAG-3+/NFAT-luc2 в -1 день перед проведением анализа, чтобы получить плотность клеток приблизительно 1 миллион/мл (от 0,8 до 1,2 миллиона/мл) в 0 день.
2. Приготовить аналитическую среду, при необходимости, и заранее нагреть среду в течение 30 минут при 37°С:
3. Приготовить, при необходимости, исходный раствор IMP761 для контроля качества (КК):
Приготовить исходный раствор 24000 нг/мл; например:
10 мкл исходного раствора IMP761 (2,06 мг/мл)+848,3 мкл аналитической среды.
Хранить в виде аликвот по 25 мкл в морозильной камере при -80°С.
4. Приготовить 3Х растворы IMP761 (партия 270416, в концентрации 2,06 мг/мл), отрицательного контроля IgG4 человека (или любого другого антитела).
Осуществить этапы предварительного разведения в зависимости от исходной концентрации антитела: объем аналитической среды (в мкл), который необходимо добавить, чтобы разбавить 2 мкл исходного раствора с получением предварительного разведения с концентрацией 100 мкг/мл, определяют следующим образом:
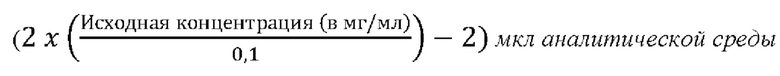

5. Приготовить 3Х растворы IMP761 для контроля качества (КК)
Разморозить аликвоту исходного раствора IMP761 КК [24000 нг/мл] и сделать серийные разведения

6. Приготовить реагент OKT3
3,6 мкл исходного раствора + 4 мл аналитической среды = 0,9 мкг/мл
150 мкл [0,9 мкг/мл] + 14850 мкл аналитической среды = 9 нг/мл (3Х)
7. Приготовить эффекторные клетки Jurkat LAG-3/NFAT-luc2
На 0 день посчитать клетки, применяя окрашивание трипановым синим.
Центрифугировать клетки при 1200 об/мин в течение 5 минут.
Удалить среду путем аспирации и ресуспендировать клетки при концентрации 3,75×106/мл в аналитической среде.
8. Распределить 3Х растворы по 96-луночному планшету.
Не использовать наружные лунки по причине краевого эффекта
Распределить по 40 мкл 3Х растворов IMP761/КК/аналитической среды (0) по соответствующим лункам аналитического планшета в двух повторах.
Распределить по 40 мкл/лунку 3Х раствора антитела против CD3/аналитической среды (нестимулированный контроль: нестим) в каждую лунку аналитического планшета в двух повторах. Конечная концентрация: 3 нг/мл
Распределить по 40 мкл на лунку (0,15×106/лунку)
Пример матрицы планшета (референсная партия/неизвестно)
9. Инкубировать планшет(ы) при 37°С во влажном термостате с 5% СО2 в течение 24 часов.
10. Приготовить реагент BioGlo
a) Поместить замороженный реагент BioGlo на комнатную температуру (КТ) на 3-6 часов перед применением, чтобы позволить ему разморозиться.
b) Перенести буфер (10 мл) в колбу с субстратом, пермешать и хранить при КТ в темноте до момента применения.
11. Уравновесить планшет(ы) при КТ в течение 15 минут.
12. Добавить 120 мкл/лунку BioGlo, избегать образования пузырьков/удалять их.
13. Инкубировать при КТ в течение от 5 до 15 минут.
14. Измерить люминесценцию (RLU), время накопления сигнала = 0,5 сек/лунку, применяя устройство для прочтения планшетов 2103 Envision Multilabel от PerkinElmer. Провести по 3 измерения для каждой лунки с интервалами по 45 секунд, рассчитать среднее значение по 3 измерениям с получением "Среднего значения RLU".
Результаты
На Фигуре 4 показан пример результатов, полученных при анализе эффективности, с применением антитела IgG4 в качестве отрицательного контроля. Максимальную активацию зарегистрировали как активацию, наблюдаемую, когда не присутствовало IMP761. Максимальное ингибирование зарегистрировали как активацию, наблюдаемую, когда концентрация IMP761 составляла 1000 нг/мл. С помощью пятипараметрической модели нелинейной регрессии определили, что IC50 IMP761 составляла 37 нг/мл.
Пример 3
Способность анализа эффективности IMP761 оценить денатурированное антитело IMP761.
Способность анализа эффективности, описанного в Примере 2, оценить пониженную эффективность денатурированного антитела IMP761 исследовали путем сравнения IC50 референсного IMP761, которое хранили при 4°С, с IC50 IMP761 из той же партии после термического стресса (10 минут при 70°С и 20 минут при 70°С). Результаты представлены на Фигуре 5 и в Таблице 4 ниже:
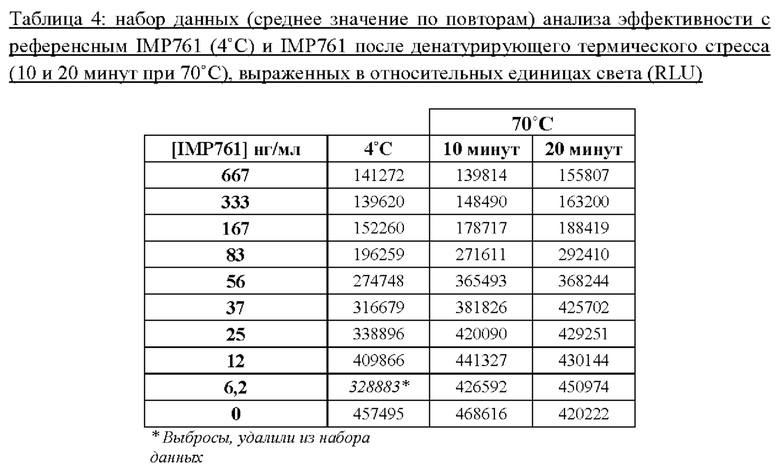
С помощью пятипараметрической модели нелинейной регрессии определили, что IC50 денатурированных антител IMP761 (74 нг/мл и 81,5 нг/мл после 10 минут и 20 минут при 70°С, соответственно) была выше, чем IC50 (41 нг/мл) референсного антитела IMP761, которое хранили при 4°С.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Immutep S.A.S.
<120> АНАЛИЗЫ
<130> P/80025.WO01
<140> PCT/EP2020/062206
<141> 2020-05-01
<150> GB 1906127.4
<151> 2019-05-01
<160> 12
<170> PatentIn версии 3.5
<210> 1
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность CDR
<400> 1
Gly Phe Ser Leu Ser Thr Ser Gly Met Gly
1 5 10
<210> 2
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность CDR
<400> 2
Ile Trp Trp Asp Asp Ile Lys
1 5
<210> 3
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность CDR
<400> 3
Ala Arg Ile Val Glu Gly Ser Tyr Ser Ser Ser Tyr Phe Asp Val
1 5 10 15
<210> 4
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность CDR
<400> 4
Gln Asp Val Ile Phe Asp
1 5
<210> 5
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность CDR
<400> 5
Ser Ala Ser
1
<210> 6
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность CDR
<400> 6
Gln Gln His Tyr Ser Thr Pro Tyr Thr
1 5
<210> 7
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность CDR
<400> 7
Thr Ser Gly Met Gly Leu Gly
1 5
<210> 8
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность CDR
<400> 8
His Ile Trp Trp Asp Asp Ile Lys Arg Tyr Asn Pro Asp Leu Arg Ser
1 5 10 15
<210> 9
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность CDR
<400> 9
Ile Val Glu Gly Ser Tyr Ser Ser Ser Tyr Phe Asp Val
1 5 10
<210> 10
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность CDR
<400> 10
Lys Ala Ser Gln Asp Val Ile Phe Asp Val Ala
1 5 10
<210> 11
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность CDR
<400> 11
Ser Ala Ser Ser Arg Val Ser
1 5
<210> 12
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Последовательность CDR
<400> 12
Gln Gln His Tyr Ser Thr Pro Tyr Thr
1 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ТЕСТ-СИСТЕМА ДЛЯ АНАЛИЗА ФУНКЦИОНАЛЬНОЙ АКТИВНОСТИ АНТИТЕЛ ПРОТИВ PD-1 И АНТИТЕЛ ПРОТИВ PD-L1 | 2019 |
|
RU2731896C1 |
| БИФУНКЦИОНАЛЬНЫЕ СВЯЗЫВАЮЩИЕ ПОЛИПЕПТИДЫ | 2019 |
|
RU2806747C2 |
| ОПРЕДЕЛЯЮЩИЕ КОМПЛЕМЕНТАРНОСТЬ УЧАСТКИ ДЛЯ СВЯЗЫВАНИЯ CD3 И СОДЕРЖАЩАЯ ИХ БИСПЕЦИФИЧЕСКАЯ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА | 2019 |
|
RU2738802C1 |
| УЛУЧШЕННЫЕ ИММУННЫЕ КЛЕТКИ С ДВОЙНОЙ кшРНК И КОМПОЗИЦИИ, СОДЕРЖАЩИЕ ИХ | 2019 |
|
RU2793922C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ С МУТИРОВАННЫМИ КОСТИМУЛЯТОРНЫМИ ДОМЕНАМИ CD28 | 2018 |
|
RU2800922C2 |
| ТРАНСГЕННОЕ ЖИВОТНОЕ, ОТЛИЧНОЕ ОТ ЧЕЛОВЕКА, И ЕГО ПРИМЕНЕНИЯ | 2010 |
|
RU2579701C2 |
| ЛЕЧЕНИЕ РАКА С ПОМОЩЬЮ ХИМЕРНОГО АНТИГЕННОГО РЕЦЕПТОРА К CD33 | 2015 |
|
RU2747384C2 |
| ИНГИБИТОРЫ Т-КЛЕТОЧНОЙ АКТИВАЦИИ | 2012 |
|
RU2657440C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К OX40 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2709742C2 |
| ЛИГАНД GITR И СВЯЗАННЫЕ С ЛИГАНДОМ GITR МОЛЕКУЛЫ И АНТИТЕЛА И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2004 |
|
RU2369636C2 |
Изобретение относится к области молекулярной биологии. Описаны способы анализа in vitro для определения активности агониста гена активации лимфоцитов 3 (LAG-3). Способ включает обеспечение множества эффекторных T-клеток, каждая эффекторная T-клетка экспрессирует на своей поверхности LAG-3 и T-клеточный рецептор (TCR) и содержит репортерный ген, кодирующий репортер, причем экспрессия указанного репортера регулируется опосредованным LAG-3 ингибированием передачи сигнала TCR в указанных эффекторных T-клетках. Далее следует определение активности указанного агониста по степени, до которой изменяется экспрессия указанного репортера в присутствии указанного агониста по сравнению с экспрессией указанного репортера в отсутствие указанного агониста, причем указанный способ осуществляют в отсутствие природного лиганда LAG-3. Также описаны наборы для осуществления указанного способа и его применения. Технический результат заключается в свойстве IMP761, посредством агонизма LAG-3, ингибировать передачу сигнала TCR, в частности регулируемую NFAT экспрессию генов, в LAG-3 положительных Т-клетках. 6 н. и 44 з.п. ф-лы, 5 ил., 4 табл., 3 пр.
1. Способ in vitro для определения активности агониста гена активации лимфоцитов 3 (LAG-3), включающий:
обеспечение множества эффекторных T-клеток, каждая эффекторная T-клетка экспрессирует на своей поверхности LAG-3 и T-клеточный рецептор (TCR) и содержит репортерный ген, кодирующий репортер, причем экспрессия указанного репортера регулируется опосредованным LAG-3 ингибированием передачи сигнала TCR в указанных эффекторных T-клетках; и
определение активности указанного агониста по степени, до которой изменяется экспрессия указанного репортера в присутствии указанного агониста по сравнению с экспрессией указанного репортера в отсутствие указанного агониста;
причем указанный способ осуществляют в отсутствие природного лиганда LAG-3.
2. Способ по п. 1 для определения эффективности препарата агониста LAG-3.
3. Способ in vitro для скрининга в отношении агониста LAG-3, включающий:
обеспечение множества эффекторных T-клеток, каждая эффекторная T-клетка экспрессирует на своей поверхности LAG-3 и T-клеточный рецептор (TCR) и содержит репортерный ген, кодирующий репортер, причем экспрессия указанного репортера регулируется опосредованным LAG-3 ингибированием передачи сигнала TCR в указанных эффекторных T-клетках; и
определение того, является ли агонист-кандидат агонистом LAG-3, путем определения степени, до которой изменяется экспрессия указанного репортера в присутствии указанного агониста-кандидата по сравнению с экспрессией указанного репортера в отсутствие указанного агониста-кандидата;
причем указанный способ осуществляют в отсутствие природного лиганда LAG-3.
4. Способ по п. 1 или 2, отличающийся тем, что указанный репортер экспрессируется на базальном уровне в указанных эффекторных T-клетках в отсутствие указанного агониста, или способ по п. 3, отличающийся тем, что указанный репортер экспрессируется на базальном уровне в указанных эффекторных T-клетках в отсутствие указанного агониста-кандидата.
5. Способ по любому из предшествующих пунктов, отличающийся тем, что экспрессия указанного репортера снижена в присутствии указанного агониста или агониста-кандидата по сравнению с экспрессией указанного репортера в отсутствие указанного агониста или агониста-кандидата.
6. Способ по любому из предшествующих пунктов, отличающийся тем, что экспрессия указанного репортера изменяется в каждой эффекторной T-клетке в ответ на активацию указанной эффекторной T-клетки посредством TCR, и который дополнительно включает:
активацию указанных эффекторных T-клеток путем независимой от антигена, независимой от ГКГС II класса, опосредованной TCR активации T-клеток в присутствии и отсутствие указанного агониста или агониста-кандидата; и
определение активности указанного агониста или агониста-кандидата, по степени, до которой изменяется экспрессия указанного репортера в ответ на активацию указанных эффекторных T-клеток в присутствии указанного агониста или агониста-кандидата по сравнению с экспрессией указанного репортера в ответ на активацию указанных эффекторных T-клеток в отсутствие указанного агониста или агониста-кандидата.
7. Способ по п. 6, отличающийся тем, что экспрессия указанного репортера повышается в каждой эффекторной T-клетке в ответ на активацию указанной эффекторной T-клетки посредством TCR и снижается в присутствии указанного агониста или агониста-кандидата в результате опосредованного LAG-3 ингибирования передачи сигнала TCR в указанной эффекторной T-клетке, и при этом активность указанного агониста или агониста-кандидата определяют по степени, до которой снижается экспрессия указанного репортера в ответ на активацию указанных эффекторных T-клеток в присутствии указанного агониста или агониста-кандидата по сравнению с экспрессией указанного репортера в ответ на активацию указанных эффекторных T-клеток в отсутствие указанного агониста или агониста-кандидата.
8. Способ по п. 6 или 7, отличающийся тем, что указанные эффекторные T-клетки активируют путем приведения в контакт указанных эффекторных T-клеток с независимым от антигена, независимым от ГКГС II класса активатором T-клеток при условиях, подходящих для независимой от антигена, независимой от ГКГС II класса, опосредованной TCR активации эффекторных T-клеток указанным активатором T-клеток.
9. Способ по п. 8, отличающийся тем, что указанные эффекторные T-клетки приводят в контакт с указанным активатором T-клеток при концентрации указанного активатора T-клеток, при которой происходит максимальное ингибирование экспрессии указанного репортера в присутствии избытка агониста LAG-3.
10. Способ по п. 8 или 9, отличающийся тем, что указанные эффекторные T-клетки приводят в контакт с указанным активатором T-клеток при концентрации указанного активатора T-клеток, которая меньше, чем концентрация указанного активатора T-клеток, при которой наблюдается наибольшая экспрессия указанного репортера в ответ на активацию указанных эффекторных T-клеток указанным активатором T-клеток в отсутствие указанного агониста или агониста-кандидата.
11. Способ по любому из пп. 8-10, отличающийся тем, что указанный активатор T-клеток содержит или состоит из антитела против CD3, или его фрагмента, или производного, у которых сохраняется способность к независимой от антигена, независимой от ГКГС II класса, опосредованной TCR активации эффекторных T-клеток.
12. Способ по п. 11, отличающийся тем, что указанное антитело против CD3 представляет собой OKT3.
13. Способ по п. 11 или 12, отличающийся тем, что указанное антитело против CD3, или его фрагмент, или производное приводят в контакт с указанными эффекторными T-клетками в концентрации ~6-30×10-12 М, предпочтительно в концентрации 1-4 нг/мл для полноразмерного антитела, или молярный эквивалент для его фрагмента или производного.
14. Способ по любому из предшествующих пунктов, отличающийся тем, что указанные эффекторные T-клетки активируют путем бесклеточной, независимой от антигена, независимой от ГКГС II класса, опосредованной TCR активации T-клеток.
15. Способ по любому из предшествующих пунктов, отличающийся тем, что указанные эффекторные T-клетки приводят в контакт с несколькими различными концентрациями указанного агониста или агониста-кандидата.
16. Способ по п. 15, который дополнительно включает определение значения IC50 указанного агониста или агониста-кандидата для ингибирования экспрессии указанного репортера.
17. Способ по любому из предшествующих пунктов, отличающийся тем, что указанные эффекторные T-клетки содержат гетерологичную нуклеиновую кислоту, содержащую указанный репортерный ген.
18. Способ по любому из предшествующих пунктов, отличающийся тем, что указанный репортерный ген находится под контролем промотора или элемента отклика.
19. Способ по п. 18, отличающийся тем, что указанный элемент отклика содержит элемент отклика NFAT (ядерного фактора активированных T-клеток) (NFAT-RE).
20. Способ по любому из предшествующих пунктов, отличающийся тем, что указанный репортер содержит биолюминесцентный репортер, такой как люцифераза.
21. Способ по любому из предшествующих пунктов, отличающийся тем, что указанные эффекторные T-клетки содержат гетерологичную нуклеиновую кислоту, кодирующую LAG-3.
22. Способ по любому из предшествующих пунктов, который дополнительно включает проведение анализа отрицательного контроля, в котором указанные эффекторные T-клетки активируют в отсутствие указанного агониста или агониста-кандидата, но в присутствии молекулы того же типа, что и указанный агонист или агонист-кандидат, но про которую известно, что у нее отсутствует агонистическая активность по отношению к LAG-3.
23. Способ по любому из предшествующих пунктов, отличающийся тем, что указанный агонист или агонист-кандидат представляет собой антитело против LAG-3, или его фрагмент, или производное, у которых сохраняется агонистическая активность по отношению к LAG-3.
24. Способ по любому из предшествующих пунктов, отличающийся тем, что указанные эффекторные T-клетки содержат клетки, происходящие из Jurkat.
25. Способ по любому из предшествующих пунктов, отличающийся тем, что указанный агонист представляет собой агонистическое антитело против LAG-3, или его фрагмент, или производное, у которых сохраняется агонистическая активность по отношению к LAG-3, и указанные эффекторные T-клетки содержат клетки Jurkat LAG-3+/NFAT-luc2.
26. Способ по любому из предшествующих пунктов, отличающийся тем, что указанное агонистическое антитело против LAG-3, или его фрагмент, или производное содержат последовательности CDR1-3 VH и последовательности CDR1-3 VL, представленные в SEQ ID NO: 1-6 соответственно или в SEQ ID NO: 7-12 соответственно.
27. Способ по любому из предшествующих пунктов, отличающийся тем, что указанное агонистическое антитело против LAG-3 представляет собой IMP761.
28. Набор для осуществления способа in vitro для определения активности агониста LAG-3 или для скрининга в отношении агониста LAG-3, причем указанный набор содержит:
множество эффекторных T-клеток, каждая эффекторная T-клетка экспрессирует на своей поверхности LAG-3 и T-клеточный рецептор (TCR) и содержит репортерный ген, кодирующий репортер, причем экспрессия указанного репортера регулируется опосредованным LAG-3 ингибированием передачи сигнала TCR в указанных эффекторных T-клетках, и причем экспрессия указанного репортера изменяется в каждой эффекторной T-клетке в ответ на активацию указанной эффекторной T-клетки посредством TCR; и
активатор T-клеток, способный к независимой от антигена, независимой от ГКГС II класса, опосредованной TCR активации указанных эффекторных T-клеток;
причем указанный набор не содержит природный лиганд LAG-3.
29. Набор согласно п. 28, отличающийся тем, что указанный набор не содержит клетку, экспрессирующую молекулы ГКГС II класса.
30. Набор согласно п. 28 или 29, отличающийся тем, что экспрессия указанного репортера повышается в ответ на активацию указанных эффекторных T-клеток посредством TCR.
31. Набор по любому из пп. 28-30, отличающийся тем, что указанные эффекторные T-клетки содержат гетерологичную нуклеиновую кислоту, содержащую указанный репортерный ген.
32. Набор по любому из пп. 28-31, отличающийся тем, что указанный репортерный ген находится под контролем промотора или элемента отклика.
33. Набор согласно п. 32, отличающийся тем, что указанный элемент отклика содержит элемент отклика NFAT (ядерного фактора активированных T-клеток) (NFAT-RE).
34. Набор по любому из пп. 28-33, отличающийся тем, что указанный репортер содержит биолюминесцентный репортер, такой как люцифераза.
35. Набор по любому из пп. 28-34, отличающийся тем, что указанные эффекторные T-клетки содержат гетерологичную нуклеиновую кислоту, кодирующую LAG-3.
36. Набор по любому из пп. 28-35, отличающийся тем, что указанный активатор T-клеток представляет собой бесклеточный активатор T-клеток.
37. Набор по любому из пп. 28-36, отличающийся тем, что указанный активатор T-клеток содержит антитело против CD3, или его фрагмент, или производное, у которых сохраняется способность к независимой от антигена, независимой от ГКГС II класса, опосредованной TCR активации эффекторных T-клеток.
38. Набор согласно п. 37, отличающийся тем, что указанное антитело против CD3 представляет собой OKT3.
39. Набор согласно п. 37 или 38, отличающийся тем, что указанное антитело против CD3, или его фрагмент, или производное присутствуют при концентрации, позволяющей их применение в указанном способе в концентрации ~6-30×10-12 М, предпочтительно в концентрации 1-4 нг/мл для полноразмерного антитела, или молярный эквивалент для его фрагмента или производного.
40. Набор по любому из пп. 28-39, который дополнительно содержит молекулу того же типа, что и указанный агонист, но про которую известно, что у нее отсутствует агонистическая активность по отношению к LAG-3, для применения в качестве отрицательного контроля.
41. Набор по любому из пп. 28-40, который дополнительно содержит известный агонист LAG-3 для применения в качестве положительного контроля.
42. Набор по любому из пп. 28-41, отличающийся тем, что указанные эффекторные T-клетки содержат клетки, происходящие из Jurkat.
43. Набор по любому из пп. 28-42, отличающийся тем, что указанные эффекторные T-клетки содержат клетки Jurkat LAG-3+/NFAT-luc2.
44. Набор по любому из пп. 28-43, отличающийся тем, что указанный набор не содержит АПК, иАПК или любые другие клетки, экспрессирующие ГКГС I класса или ГКГС II класса.
45. Набор по любому из пп. 28-44, отличающийся тем, что единственными клетками в указанном наборе являются указанные эффекторные T-клетки.
46. Набор по любому из пп. 28-45 для определения эффективности препарата агониста LAG-3.
47. Набор по любому из пп. 28-46 для осуществления способа по любому из пп. 1-27.
48. Применение набора по любому из пп. 28-45 для определения активности агониста LAG-3.
49. Применение набора по любому из пп. 28-45 для определения эффективности препарата агониста LAG-3.
50. Применение набора по любому из пп. 28-45 для скрининга в отношении агониста LAG-3.
| US 2017097333 A1, 06.04.2017 | |||
| Anonymous, "LAG3 / IL-2 Reporter - Jurkat Recombinant Cell Line", 31.12.2016, найдено в сети Интернет 28.09.2023 https://bpsbioscience.com/lag3-il-2-reporter-jurkat-recombinant-cell-line-79813 | |||
| РАСТВОРИМЫЕ ПОЛИПЕПТИДНЫЕ ФРАКЦИИ ПРОТЕИНА LAG-3, СПОСОБ ПОЛУЧЕНИЯ, ТЕРАПЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, АНТИТЕЛО | 1995 |
|
RU2178306C2 |

