Предпосылки для изобретения
Настоящее изобретение касается способа получения L-глутаминовой кислоты с помощью ферментации, сопряженной с ее осаждением. L-глутаминовая кислота широко используется в качестве различных добавок и т.д.
В целом, L-глутаминовая кислота вырабатывается в ходе ферментационного процесса с использованием т.н. коринебактерий, являющихся продуцентами L-глутаминовой кислоты и относящихся к родам Brevibacterium, Corynebacterium или Microbacterium или представляющих их мутантные штаммы («Amino Acid Fermentation», Gakkai Shuppan Center, 1986, pp.195-215). В качестве способов выработки L-глутаминовой кислоты путем ферментации с использованием других бактериальных штаммов известен способ, в котором используются микроорганизмы, относящиеся к родам Bacillus, Streptomyces, Penicillum или подобным (патент США № 3220929), способ, основанный на использовании микроорганизмов, относящихся к родам Pseudomonas, Arthrobacter, Serratia, Candida или подобным (патент США № 3563857), способ, в котором используются микроорганизмы, относящиеся к родам Bacillus, Pseudomonas, Serratia, Aerobacter aerogenes (в настоящее время определяемого как Enterobacter aerogenes) или подобным (японская патентная публикация (Ко Ко Ku) № 32-9393), способ, основанный на использовании мутантного штамма Escherichia coli (открытая японская патентная публикация (Ко Kai) № 5-244970 и другие. Кроме того, заявители настоящего изобретения представляют способ выработки L-глутаминовой кислоты с использованием микроорганизма, относящегося к родам Klebsiella, Erwinia или Pantoea (открытая японская патентная заявка № 2000-106869).
Кроме того, были заявлены различные способы повышения способности к выработке L-глутаминовой кислоты за счет усиления активностей ферментов, участвующих в биосинтезе L-глутаминовой кислоты за счет использования методологии рекомбинантной ДНК. Например, было сообщено, что внесение гена, кодирующего цитратсинтазу, происходящую от Escherichia coli или Corynebacterium glutamicum, было эффективным с точки зрения повышения способности вырабатывать L-глутаминовую кислоту бактериями родов Corynebacterium или Brevibacterium (японская патентная публикация № 7-121228). Кроме того, в открытой японской патентной заявке № 61-268185 заявляется клетка, несущая рекомбинантную ДНК, включающую ген глутаматдегидрогеназы, происходящий от коринебактерии Corynebacterium. Далее, открытая японская патентная заявка №63-214189 заявляет способ повышения способности вырабатывать L-глутаминовую кислоту путем амплификации гена глутаматдегидрогеназы, гена изоцитратдегидрогеназы, гена аконитатгидратазы и гена цитратсинтазы.
Хотя уровень выработки L-глутаминовой кислоты в существенной степени повышается в результате воспроизводства упоминавшихся выше микроорганизмов или улучшения способов выработки, тем не менее разработка способов более эффективной выработки L-глутаминовой кислоты при низких затратах является необходимой с точки зрения дальнейшего повышения потребности в будущем.
Известен способ, в соответствии с которым ферментация осуществляется так, что кристаллизующаяся L-аминовая кислота накапливается в культуре (открытая японская патентная заявка № 62-288). В этом способе концентрация L-аминокислоты в культуре поддерживается на уровне, ниже некоторого значения путем осаждения накопленной в культуре L-аминокислоты. В частности, L-триптофан, L-тирозин или L-лейцин осаждают в ходе ферментационного процесса путем доведения температуры и рН культуры или путем добавления поверхностно-активного агента к культуральной среде.
При том, что способ ферментации, сопряженный с осаждением L-аминокислоты, известен, на что было указано выше, а аминокислотами, пригодными для ферментации таким способом, являются аминокислоты, характеризующиеся относительно низкой растворимостью в воде, в то время как не было известно примеров применения данного способа в отношении хорошо растворимых в воде аминокислот, таких как L-глутаминовая кислота. Кроме того, культуральная среда для осаждения L-глутаминовой кислоты должна характеризоваться низким значением рН. Однако вырабатывающие L-глутаминовую кислоту бактерии, такие как те бактерии, которые упоминались выше, не могут расти в кислых условиях, а следовательно, ферментация L-глутаминовой кислоты должна проводиться в нейтральных условиях (патенты США №№ 3220929 и 3032474; K.C.Chao & J.W.Foster, 1959, J.Bacteriol., 77, pp.715-725). Таким образом, выработка L-глутаминовой кислоты в ферментационном процессе в сопряжении с осаждением была неизвестна. Более того, известно, что рост большинства ацидофильных бактерий подавляется в присутствии органических кислот, таких как уксусная кислота, молочная кислота и янтарная кислота («Extreme Environment Microorganism Handbook», ed. Y.Oshima, Sci. Forum, p.231; R.M.Borichewski, 1967, J.Bacteriol., 93, pp.597-599; и др.). Таким образом, предполагается, что большое число микроорганизмов является чувствительным к L-глутаминовой кислоте, которая также является органической кислотой, в кислых условиях; при этом отсутствуют сообщения о попытках поиска микроорганизмов, проявляющих способность вырабатывать L-глутаминовую кислоту в кислых условиях.
Резюме изобретения
В описанной выше современной ситуации объектом настоящего изобретения явился поиск и воспроизводство микроорганизма, который бы вырабатывал L-глутаминовую кислоту в условиях низких значений рН, а также представление способа выработки L-глутаминовой кислоты с использованием полученного микроорганизма путем ферментации, сопровождающейся осаждением L-глутаминовой кислоты.
Заявители настоящего изобретения в ходе анализа улучшения выработки L-глутаминовой кислоты в ферментационном процессе предположили, что подавление выработки L-глутаминовой кислоты, накопленной в культуральной среде при высокой концентрации, являлось одним из факторов, не позволяющих достичь повышения уровня выработки. Например, у клеток имеется экскреционная (выделительная) система и система поглощения для L-глутаминовой кислоты. Однако, если L-глутаминовая кислота, однажды уже выделившаяся в результате экскреции в среду, снова поглощается этой клеткой, то это приводит не только к существенному снижению эффективности выработки, но также и к подавлению реакций биосинтеза L-глутаминовой кислоты. С целью исключения подавления выработки таким высококонцентрированным накоплением L-глутаминовой кислоты заявители настоящего изобретения протестировали микроорганизмы, которые могут размножаться в кислых условиях и в присутствии высокой концентрации L-глутаминовой кислоты. В результате удалось успешно выделить из почвы микроорганизмы, для которых характерны такие свойства, что тем самым позволило завершить настоящее изобретение.
Таким образом, в настоящем изобретении представляется следующее.
(1) Микроорганизм, который может метаболизировать источник углерода при конкретной величине рН в жидкой культуральной среде, содержащей L-глутаминовую кислоту в концентрации насыщения и данный источник углерода, и который обладает способностью накапливать L-глутаминовую кислоту в количестве, превышающем то ее количество, которое соответствует концентрации насыщения в жидкой среде при данном рН.
(2) Микроорганизм в соответствии с п.(1), который может расти в жидкой культуральной среде.
(3) Микроорганизм в соответствии с пп.(1) или (2), при том, что рН не превышает 5,0.
(4) Микроорганизм в соответствии с любым из пп.(1)-(3), обладающий по крайней мере одним из следующих признаков:
(a) микроорганизм проявляет повышенную активность фермента, катализирующего реакцию биосинтеза L-глутаминовой кислоты; и
(b) микроорганизм характеризуется уменьшенной или отсутствующей активностью фермента, катализирующего реакцию, отклоняющуюся от биосинтетического пути L-глутаминовой кислоты и обеспечивающего выработку иного, нежели L-глутаминовая кислота, соединения.
(5) Микроорганизм в соответствии с п.(4), при том, что фермент, который катализирует реакцию биосинтеза L-глутаминовой кислоты, является по крайней мере одним из группы, включающей цитратсинтазу, фосфоенолпируваткарбоксилазу и глутаматдегидрогеназу.
(6) Микроорганизм в соответствии с пп.(4) или (5), при том, что ферментом, который катализирует реакцию, отклоняющуюся от биосинтетического пути L-глутаминовой кислоты и связанную с выработкой иного, нежели L-глутаминовой кислота, соединения, является α-кетоглутаратдегидрогеназа.
(7) Микроорганизм в соответствии с любым из пп.(1)-(6), при том, что этот микроорганизм относится к роду Enterobacter.
(8) Микроорганизм в соответствии с п.(7), являющийся видом Enterobacter agglomerans.
(9) Микроорганизм в соответствии с п.(8), несущий мутацию, которая обусловливает сниженную интенсивность внеклеточной секреции вязкого материала по сравнению со штаммом дикого типа в случае культивирования в среде, содержащей сахариды.
(10) Способ выработки L-глутаминовой кислоты с помощью ферментации, который включает культивирование микроорганизма, определенного в любом из пп.(1)-(9), в жидкой культуральной среде, рН которой доведена до величины, при которой происходит осаждение L-глутаминовой кислоты, с целью выработки и накопления L-глутаминовой кислоты и осаждения L-глутаминовой кислоты в культуральной среде.
(11) Способ скрининга микроорганизма, пригодного для выработки L-глутаминовой кислоты с помощью ферментации, сопряженной с осаждением L-глутаминовой кислоты в жидкой культуральной среде, который включает внесение содержащего микроорганизмы образца в кислую среду, содержащую L-глутаминовую кислоту в концентрации насыщения и источник углерода, и отбор штамма, который способен метаболизировать этот источник углерода.
(12) Способ в соответствии с п.(11), при том, что штамм, способный расти в данной культуральной среде, выбирается как штамм, который способен метаболизировать данный источник углерода.
(13) Способ в соответствии с пп.(11) или (12), при том, что рН среды не превышает 5,0.
В соответствии со способом по настоящему изобретению L-глутаминовая кислота может быть выработана с помощью ферментации, сопряженной с осаждением L-глутаминовой кислоты. В результате концентрация L-глутаминовой кислоты в среде поддерживается ниже определенной, и L-глутаминовая кислота может вырабатываться, не будучи подверженной ингибированию высокой концентрацией L-глутаминовой кислоты.
Краткое описание чертежей
На фигуре 1 показана рестрикционная карта фрагмента ДНК, производного от плазмиды pTWVEK101 Enterobacter agglomerans.
На фигуре 2 отображено сравнение аминокислотной последовательности, выведенной из нуклеотидной последовательности гена sucA энтеробактерии Enterobacter agglomerans и такого же гена кишечной палочки Escherichia coli. Верхняя последовательность - Enterobacter agglomerans; нижняя последовательность - Escherichia coli (такое взаиморасположение принято здесь и далее).
На фигуре 3 отображено сравнение аминокислотной последовательности, выведенной из нуклеотидной последовательности гена sucB Enterobacter agglomerans и такого же гена Escherichia coli.
На фигуре 4 отображено сравнение аминокислотной последовательности, выведенной из нуклеотидной последовательности гена sdhB Enterobacter agglomerans и такого же гена Escherichia coli.
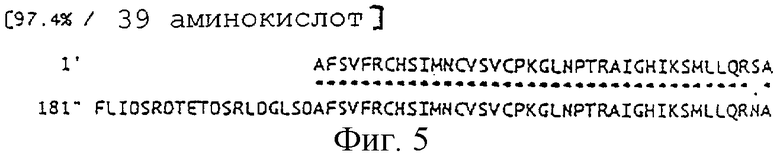
На фигуре 5 отображено сравнение аминокислотной последовательности, выведенной из нуклеотидной последовательности гена sucC Enterobacter agglomerans и такого же гена Escherichia coli.
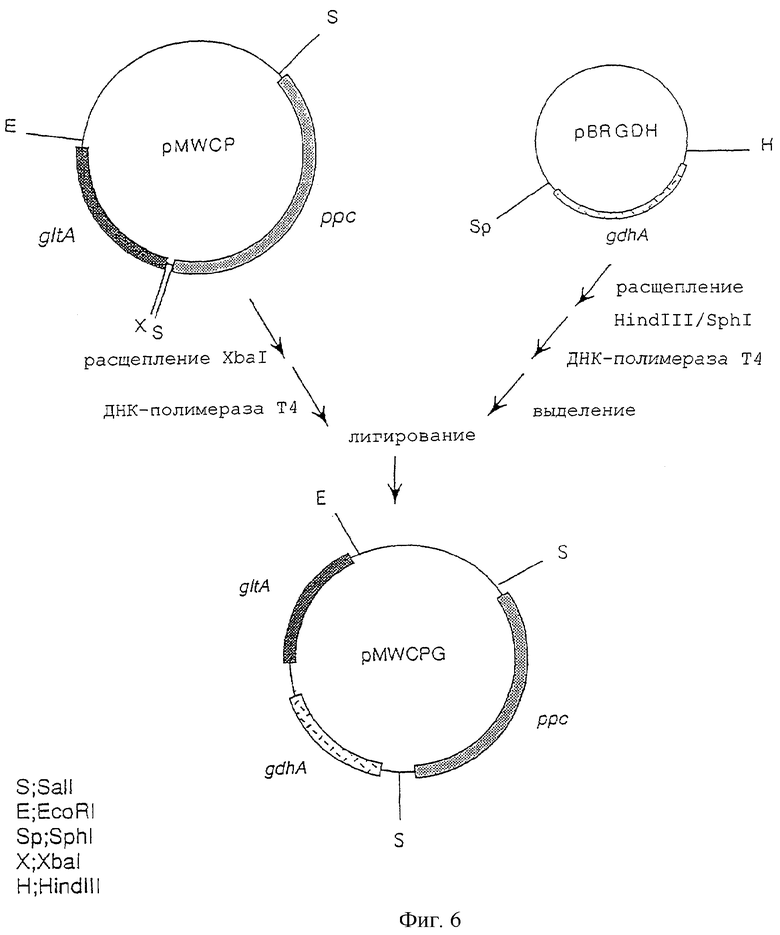
На фигуре 6 показана конструкция плазмиды pMWCPG, включающей ген gltA, ген ррс и ген gdhA.
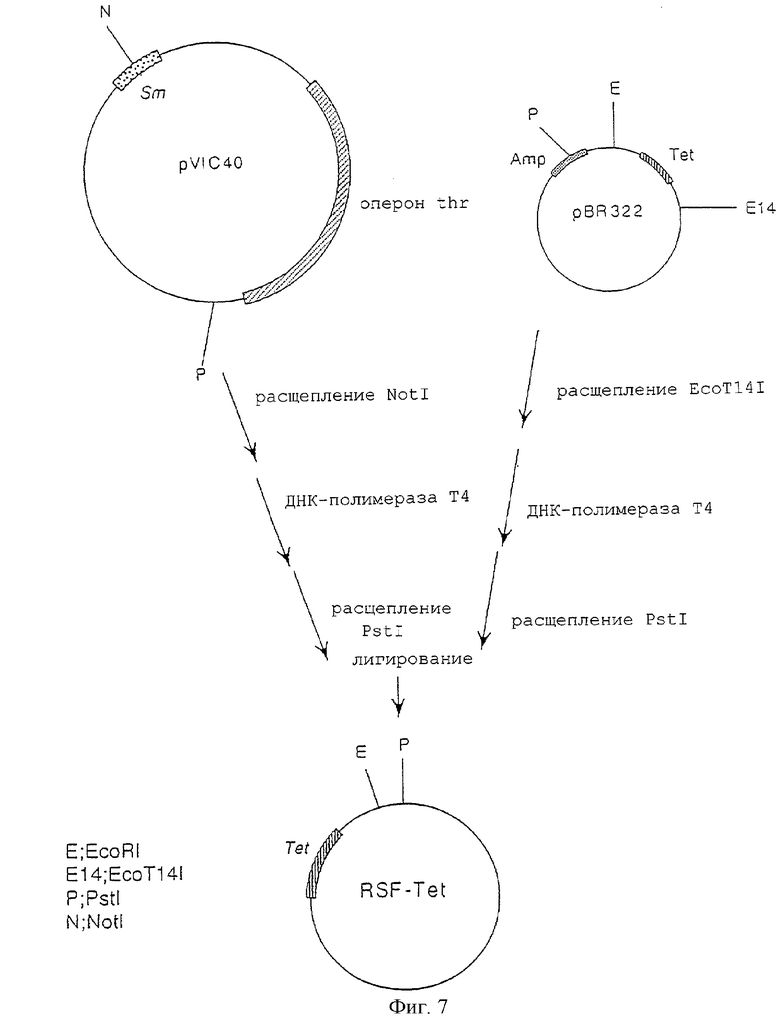
На фигуре 7 показана конструкция плазмиды RSF-Tet, включающей сайт начала репликации, производный от плазмиды RSF1010, характеризующейся сродством к широкому кругу хозяев, и ген резистентности к тетрациклину.
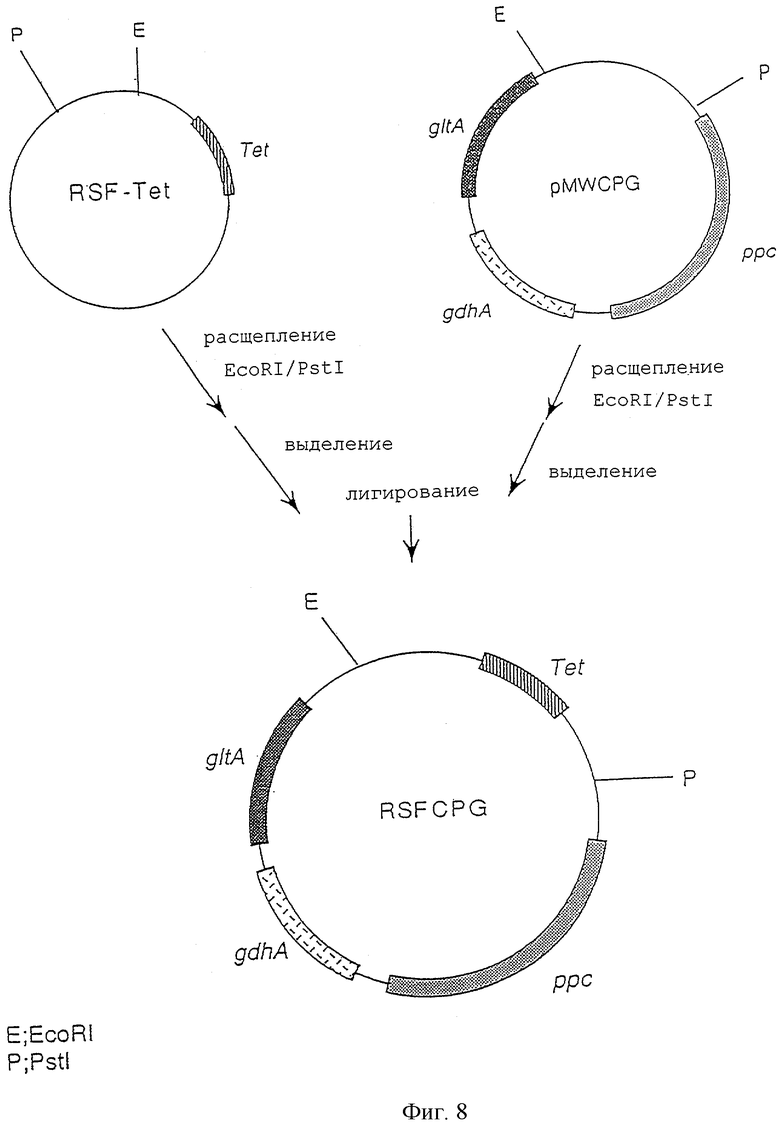
На фигуре 8 изображена конструкция плазмиды RSFCPG, включающей сайт начала репликации, производный от плазмиды RSF1010, характеризующейся сродством к широкому кругу хозяев, ген резистентности к тетрациклину, ген gltA, ген ррс и ген gdhA.
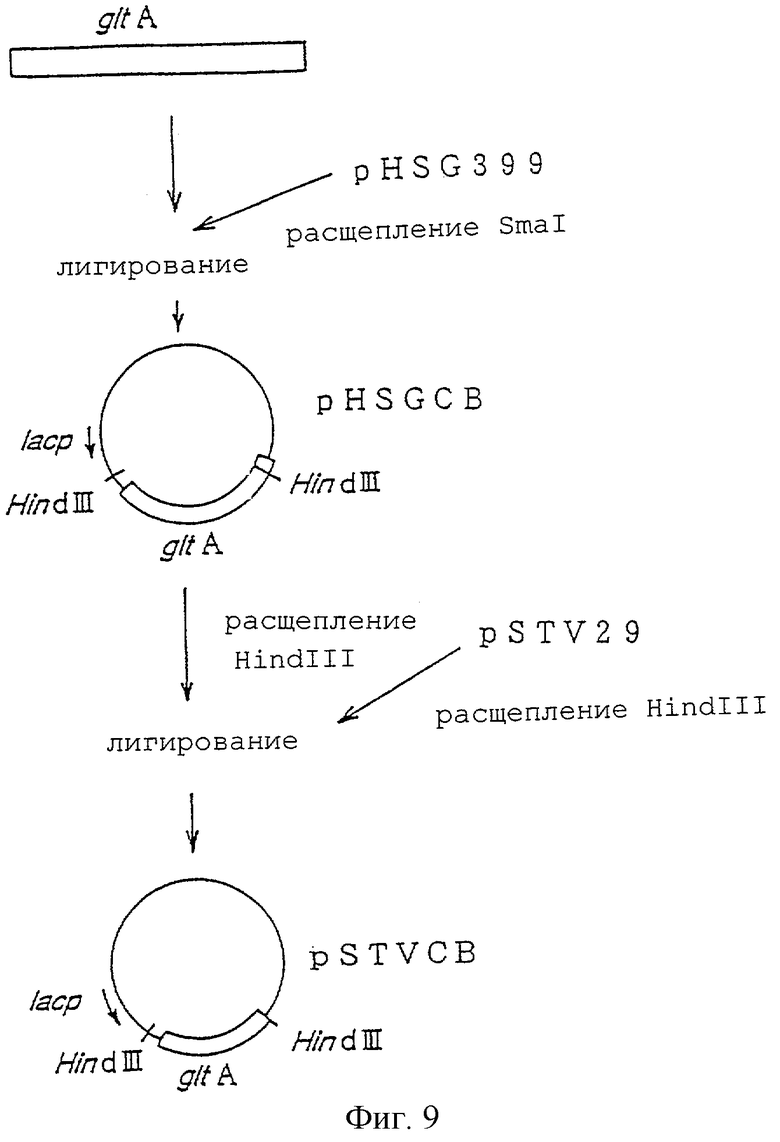
На фигуре 9 показана конструкция плазмиды pSTVCB, включающей ген gltA.
Подробное описание изобретения
Здесь и далее настоящее изобретение будет описано подробно.
Микроорганизмом по настоящему изобретению является микроорганизм, который: (1) способен метаболизировать источник углерода при конкретной величине рН в жидкой культуральной среде, содержащей L-глутаминовую кислоту в концентрации насыщения и данный источник углерода, и (2) обладает способностью накапливать L-глутаминовую кислоту в количестве, превышающем то количество, которое соответствует концентрации насыщения в данной культуральной среде при данном значении рН.
Термин «концентрация насыщения» обозначает концентрацию L-глутаминовой кислоты, растворенной в жидкой культуральной среде, при которой эта жидкая среда оказывается насыщенной по L-глутаминовой кислоте.
Здесь и далее будет описан способ скрининга микроорганизма, который способен метаболизировать источник углерода в жидкой среде, содержащей L-глутаминовую кислоту в концентрации насыщения и данный источник углерода при конкретной величине рН. Содержащий микроорганизмы образец вносят в жидкую культуральную среду, содержащую L-глутаминовую кислоту в концентрации насыщения и источник углерода, при конкретной величине рН: отбирают штамм, который способен метаболизировать этот источник углерода. Конкретное значение рН в принципе не ограничено, но обычно не должно превышать примерно 5,0, предпочтительно не должно превышать примерно 4,5 и более предпочтительно не должно превышать примерно 4,3. Микроорганизм по настоящему изобретению используется для выработки L-глутаминовой кислоты с помощью ферментации, сопряженной с осаждением L-глутаминовой кислоты. Если значение рН слишком высоко, то затруднительным становится обеспечить выработку микроорганизмом достаточного для осаждения количества L-глутаминовой кислоты. Следовательно, значение рН предпочтительно находится в указанном выше диапазоне.
Если величина рН водного раствора, содержащего L-глутаминовую кислоту, снижается, то растворимость L-глутаминовой кислоты в значительной степени уменьшается до величины, примерно равной рКа γ-карбоксильной группы (4,25 при 25°С). Растворимость становится наименьшей при достижении изоэлектрической точки (рН=3,2) - и L-глутаминовая кислота сверх количества, определяющего концентрацию насыщения, осаждается из раствора. С учетом зависимости от состава культуральной среды L-глутаминовую кислоту обычно растворяют в количестве 10-20 г/л при рН=3,2, 30-40 г/л при рН=4/0 и 50-60 г/л при рН=4,7 (при примерно 30°С). Обычно нет необходимости доводить рН до величин меньше 3,0, потому что эффект осаждения L-глутаминовой кислоты достигает плато-фазы при рН, меньшем некоторого значения. Однако величина рН может быть и меньше 3,0.
Кроме того, выражение «микроорганизм способен метаболизировать источник углерода» означает, что он способен размножаться или утилизировать источник углерода даже без способности к размножению: следовательно, это определяет то, что он катаболизирует источники углерода, такие как сахариды или органические кислоты. Более конкретно, например, если микроорганизм размножается в случае его культивирования в жидкой среде, содержащей L-глутаминовую кислоту в концентрации насыщения, при рН=4,0-5,0, предпочтительно при рН=4,0-4,5, более предпочтительно при рН=4,0-4,3, а наиболее предпочтительно рН=4,0, при подходящей температуре, например при 28°С, 37°С или 50°С, в течение 2-4 дней, то этот микроорганизм способен метаболизировать источник углерода в среде. Далее, например, даже если микроорганизм не размножается в случае его культивирования в жидкой культуральной среде, содержащей L-глутаминовую кислоту в концентрации насыщения, при рН=4,0-5,0, предпочтительно при рН=4,0-4,5, более предпочтительно при рН=4,0-4,3, а наиболее предпочтительно рН=4,0, при подходящей температуре, например при 28°С, 37°С или 50°С, в течение 2-4 дней, то этот микроорганизм, который утилизирует данный источник углерода в среде, является тем микроорганизмом, который способен метаболизировать источник углерода в среде.
Микроорганизм, который способен метаболизировать источник углерода, включает микроорганизмы, которые способны расти в жидкой культуральной среде.
Выражение микроорганизм «способен расти» означает то, что он способен размножаться или способен вырабатывать L-глутаминовую кислоту даже при отсутствии способности к размножению. Более конкретно, например, если микроорганизм размножается в случае его культивирования в жидкой культуральной среде, содержащей L-глутаминовую кислоту в концентрации насыщения, при рН=4,0-5,0, предпочтительно при рН=4,0-4,5, более предпочтительно при рН=4,0-4,3, а наиболее предпочтительно рН=4,0, при подходящей температуре, например при 28°С, 37°С или 50°С, в течение 2-4 дней, то этот микроорганизм способен расти в данной культуральной среде. Далее, например, даже если микроорганизм не размножается в случае его культивирования в жидкой синтетической среде, содержащей L-глутаминовую кислоту в концентрации насыщения, при рН=4,0-5,0, предпочтительно при рН=4,0-4,5, более предпочтительно при рН=4,0-4,3, а наиболее предпочтительно рН=4,0, при подходящей температуре, например при 28°С, 37°С или 50°С, в течение 2-4 дней, то этот микроорганизм, который обеспечивает повышение количества L-глутаминовой кислоты в культуральной среде, является тем микроорганизмом, который способен расти в данной культуральной среде.
Описанный выше отбор может быть повторен дважды или большее число раз при тех же самых условиях или при измененяющейся величине рН или концентрации L-глутаминовой кислоты. Исходный отбор может быть осуществлен в среде, содержащей L-глутаминовую кислоту в концентрации, меньшей концентрации насыщения, а после этого последующий отбор может быть проведен в среде, содержащей L-глутаминовую кислоту в концентрации насыщения. Далее могут быть отобраны штаммы, обладающие предпочтительными характеристиками, такими как лучшие параметры размножения.
В дополнение к свойству, описанному выше, микроорганизм по настоящему изобретению характеризуется способностью накапливать L-глутаминовую кислоту в количестве, превышающем то количество, которое соответствует концентрации насыщения L-глутаминовой кислоты в жидкой культуральной среде. Величина рН упомянутой выше жидкой культуральной среды предпочтительно такова же или близка к таковой у культуральной среды, использовавшейся для скрининга микроорганизма, обладающего описанным выше свойством (1). Обычно микроорганизм становится чувствительным к высокой концентрации L-глутаминовой кислоты тогда, когда величина рН снижается. Следовательно, предпочтительным является, чтобы рН не была слишком низкой с точки зрения резистентности к L-глутаминовой кислоте, но при этом низкое значение рН предпочтительно с точки зрения выработки L-глутаминовой кислоты, сопряженной с ее осаждением. Для удовлетворения таким условиям величина рН может находиться в диапазоне 3-5, предпочтительно в диапазоне 4-5, более предпочтительно в диапазоне 4,0-4,7, еще более предпочтительно в диапазоне 4,0-4,5 и конкретно предпочтительно в диапазоне 4,0-4,3.
В качестве микроорганизма по настоящему изобретению или его воспроизводительного материала можно упомянуть, например, микроорганизмы, относящиеся к родам Enterobacter, Klebsiella, Serratia, Pantoea, Erwinia, Escherichia, Corynebacterium, Alicyclobacillus, Bacillus, Saccharomyces и подобное. Среди них предпочтительными являются микроорганизмы, относящиеся к роду Enterobacter. Здесь и далее описание микроорганизмов по настоящему изобретению в основном будет относиться к микроорганизмам рода Enterobacter, хотя настоящее изобретение может быть применено к микроорганизму, относящемуся к другим родам, т.е. не ограничивается родом Enterobacter.
В качестве микроорганизма, относящегося к роду Enterobacter, можно, в частности, упомянуть вид Enterobacter agglomerans, предпочтительно Enterobacter agglomerans штамма AJ13355. Этот штамм был выделен из пробы почвы из Иваты (провинция Шицуока, Япония) в качестве штамма, который способен размножаться в культуральной среде, содержащей L-глутаминовую кислоту и источник углерода при низких значениях рН.
Физиологические характеристики штамма AJ13355 таковы:
(1) грамотрицательная бактерия;
(2) отношение к кислороду - факультативный анаэроб;
(3) позитивна по каталазе;
(4) негативна по оксидазе;
(5) способностью восстанавливать нитраты не обладает;
(6) тест Фогса-Проскауера: +;
(7) тест с метиловым красным: -;
(8) негативна по уреазе;
(9) выработка индола: +;
(10) обладает подвижностью;
(11) слабая активность по выработке сероводорода в среде TSI;
(12) позитивна по β-галактозидазе;
(13) свойства по ассимиляции сахаридов:
арабиноза - +;
сахароза - +;
лактоза - +;
ксилоза - +;
сорбитол - +;
инозитол - +;
трегалоза - +;
мальтоза - +;
глюкоза - +;
адонитол - -;
раффиноза - +;
салицин - -;
мелибиоза - +;
(14) ассимиляция глицерина - +;
(15) свойства по ассимиляции органических кислот:
лимонная кислота - +;
тартаровая кислота - -;
глюконовая кислота - +;
уксусная кислота - +;
малоновая кислота - -;
(16) негативна по аргининдегидратазе;
(17) негативна по орнитиндекарбоксилазе;
(18) негативна по лизиндекарбоксилазе;
(19) негативна по фенилаланиндеаминазе;
(20) образуемый пигмент - желтый;
(21) имеется способность разжижать желатин;
(22) диапазон рН для роста: может расти при рН=4, хорошо растет при рН=4,5-7;
(23) температура для роста: хорошо растет при 25°С, хорошо растет при 30°С, хорошо растет при 37°С, возможен рост при 42°С, рост при 45°С невозможен.
Основываясь на этих бактериологических параметрах, исследованный штамм АJ13355 был идентифицирован по принадлежности к виду Enterobacter agglomerans.
Штамм AJ13355 Enterobacter agglomerans был внесен в коллекцию Национального института биологии и биотехнологии человека Агентства по науке и технологии в промышленности Министерства международной торговли и промышленности Японии (по адресу 1-3 Higashi 1-chome, Tsukuba-shi, Ibaraki 305-8566, Japan) 19 февраля 1998 года с получением депозитарного №FERM Р-16644. Затем на основании Будапештского соглашения он был внесен в международный депозитарий 11 января 1999 года с получением депозитарного №FERM BP-6614.
Микроорганизм по настоящему изобретению может являться микроорганизмом, исходно обладающим способностью вырабатывать L-глутаминовую кислоту, или микроорганизмом, у которого способность вырабатывать L-глутаминовую кислоту придана или усилена с помощью внесения мутаций, с применением методов рекомбинантной ДНК или подобного.
Способность вырабатывать L-глутаминовую кислоту может быть придана или усилена, например, путем повышения активности фермента, который катализирует реакцию в биосинтезе L-глутаминовой кислоты. Способность вырабатывать L-глутаминовую кислоту также может быть усилена путем снижения активности фермента, который катализирует реакцию, отклоняющуюся от биосинтетического пути L-глутаминовой кислоты и приводящую к образованию иного, нежели L-глутаминовая кислота, соединения, или путем нарушения такой активности.
В качестве ферментов, катализирующих реакцию в биосинтезе L-глутаминовой кислоты, могут быть упомянуты глутаматдегидрогеназа (здесь и далее обозначаемая как GDH), глутаминсинтетаза, глутаматсинтаза, изоцитратдегидрогеназа, аконитатгидратаза, цитратсинтаза (здесь и далее обозначаемая как CS), фосфоенолпируваткарбоксилаза (здесь и далее обозначаемая как РЕРС), пируватдегидрогеназа, пируваткиназа, енолаза, фосфоглицеромутаза, фосфоглицераткиназа, глицеральдегид-3-фосфатдегидрогеназа, триозофосфатизомераза, фруктозобифосфатальдолаза, фосфофруктокиназа, глюкозофосфатизомераза и т.д. Среди этих ферментов предпочтительными являются один или два фермента из CS, РЕРС, GDH, или все три этих фермента. Далее, предпочтительным является, чтобы активность всех этих трех ферментов (CS, РЕРС и GDH) повышалась у микроорганизма по настоящему изобретению. В частности, предпочтительным является фермент CS Brevibacterium lactofermentum, поскольку он не подвержен ингибированию α-кетоглутаровой кислотой, L-глутаминовой кислотой и НАД.
С целью усиления активности ферментов CS, РЕРС или GDH, например ген, кодирующий CS, РЕРС или GDH, может быть клонирован в состав подходящей плазмиды, а микроорганизм-хозяин может быть трансформирован с использованием полученной плазмиды. В результате число копий гена, кодирующего фермент CS, РЕРС или GDH (здесь и далее эти гены обозначаются как «ген gltA», «ген ррс» и «ген gdhA» соответственно), в трансформированном штамме клеток увеличивается, что обусловливает повышение активности ферментов CS, РЕРС или GDH.
Клонированные ген gltA, ген ррс и ген gdhA вносят в упомянутый выше исходный родительский штамм по отдельности или в случайном сочетании двух из них, или все три вместе. Когда вносят два или три типа генов, то эти два или три типа генов могут быть клонированы в одну и ту же плазмиду и внесены в организм-хозяин, или же они могут быть клонированы по отдельности в два или три типа плазмид, которые могут сосуществовать и быть внесенными в данный организм-хозяин.
Могут быть внесены в один и тот же организм-хозяин два или большее число генов, кодирующих ферменты одного типа, но происходящие от различных видов микроорганизмов.
Тип упоминавшихся выше плазмид, в принципе, ни чем не ограничивается, помимо того, что они должны автономно реплицироваться в клетках микроорганизма, относящегося, например, к роду Enterobacter или подобному: при этом можно, например, упомянуть плазмиды pUC19, pUC18, pBR322, pHSG299, pHSG298, pHSG399, pHSG398, RSF1010, pMW119, pMW118, pMW219, pMW218, pACYC177, pACYC184 и т.д. Помимо этого также могут быть использованы векторы, основанные на фаговой ДНК.
Трансформация может быть осуществлена, например, с помощью метода Моррисона (D.M.Morrison, 1979, Meth. Enzymol., 68, 326), метода, в котором проницаемость ДНК повышается путем обработки клеток бактерий-реципиентов хлоридом кальция (M.Mandel & A.Higa, 1970, J. Mol. Biol., 53, 159), с помощью электропорации (J.H.Miller, 1992, «A Short Course in Bacterial Genetics», Cold Spring Harbor Lab. Press, США.) и подобного.
Активность ферментов CS, РЕРС или GDH также может быть повышена за счет интеграции множественных копий гена gltA, гена ррс или гена gdhA в хромосомную ДНК упоминавшегося выше исходного родительского штамма, взятого в качестве организма-хозяина. С целью внесения множественных копий гена gltA, гена ррс или гена gdhA в хромосомную ДНК микроорганизма, относящегося к роду Enterobacter или подобного, может быть использована нуклеотидная последовательность, которая в большом числе копий присутствует в хромосомной ДНК, такая как повторяющаяся ДНК или инвертированные повторы, имеющиеся на концах транспозонов. С другой стороны, множественные копии гена могут быть внесены в хромосомную ДНК путем переноса транспозона, включающего ген gltA, ген ррс или ген gdhA. В результате число копий гена gltA, гена ррс или гена gdhA в трансформированном клеточном штамме будет увеличено, а следовательно, повысится и активность ферментов CS, РЕРС или GDH.
В качестве организмов-источников гена gltA, гена ррс или гена gdhA, число копий которых предполагается увеличить, может быть использован любой организм, лишь бы он обладал активностями ферментов CS, РЕРС или GDH. Помимо прочего предпочтительными являются бактерии, являющиеся прокариотическими организмами, например те, которые относятся к родам Enterobacter, Klebsiella, Erwinia, Pantoea, Serratia, Escherichia, Corynebacterium, Brevibacterium и Bacillus. В качестве конкретных примеров могут быть упомянуты Escherichia coli, Brevibacterium lactofermentum и т.д. Ген gltA, ген ррс или ген gdhA могут быть получены из состава хромосомной ДНК описанных выше микроорганизмов.
Ген gltA, ген ррс или ген gdhA могут быть получены с использованием мутантного штамма, являющегося дефицитным по активности ферментов CS, РЕРС или GDH, с целью выделения фрагмента ДНК, который комплементирует ауксотрофность, обусловливаемую хромосомной ДНК упоминавшихся выше микроорганизмов. С учетом того, что нуклеотидные последовательности этих генов у кишечной палочки Escherichia и коринебактерии Corynebacterium ранее были уже определены (Biochemistry, 22, pp.5243-5249, 1983; J.Biochem., 95, pp.909-916, 1984; Gene, 27, pp.193-199, 1984, Microbiology, 140, pp.1817-1828, 1994; Mol. Gen. Genet., 218, pp.330-339, 1989; Mol. Microbiol., 6, pp.317-326, 1992), они также могут быть получены с применением ПЦР с использованием затравок, основанных на каждой из таких нуклеотидных последовательностей, и с использованием хромосомной ДНК в качестве матрицы.
Активность ферментов CS, РЕРС или GDH также может быть увеличена путем усиления экспрессии гена gltA, гена ррс или гена gdhA кроме упомянутой выше амплификации.
Например, экспрессия может быть усилена путем замены промотора гена gltA, гена ррс или гена gdhA на другие, более сильные промоторы. Например, в качестве сильных промоторов известны промотор lac, промотор trp, промотор tac, промоторы PR и PL λ-фага и другие сильные промотры. Ген gltA, ген ррс или ген gdhA, в составе которых заменяют промотор, клонируют в состав плазмиды и вносят в микроорганизм-хозяин или интегрируют в состав хромосомной ДНК микроорганизма-хозяина с помощью повторяющейся ДНК, инвертированных повторов, транспозона или подобного.
Активность ферментов CS, РЕРС или GDH также может быть усилена путем замены промотора в составе гена gltA, гена ррс или гена gdhA, находящихся в составе хромосомы, на более сильные промоторы (см. международную патентную заявку WO 87/03006 и открытую японскую патентную заявку №61-268183) или встраивания сильного промотора выше кодирующей последовательности каждого из этих генов (см. Gene, 29, pp.231-241, 1984). В частности, гомологичная рекомбинация может быть осуществлена между ДНК, включающей ген gltA, ген ррс или ген gdhA, в составе которых проведена замена на более сильный промотор или его фрагмент, и соответствующим геном в составе хромосомы.
Примерами ферментов, катализирующих реакцию, отклоняющуюся от биосинтетического пути L-глутаминовой кислоты и приводящую к образованию иного, нежели L-глутаминовая кислота, соединения, являются α-кетоглутаратдегидрогеназа (здесь и далее обозначается как α-KGDH), изоцитратлиаза, фосфатацетилтрансфераза, ацетаткиназа, синтаза ацетогидрооксокислот, ацетолактатсинтаза, форматацетилтрансфераза, лактатдегидрогеназа, глутаматдекарбоксилаза, 1-пирролиндегидрогеназа и т.д. Среди этих ферментов предпочтительным является α-KGDH.
С целью достижения снижения или нарушения активности упомянутого выше фермента у микроорганизма, относящегося к роду Enterobacter или подобного, мутация, обусловливающая снижение или блокировку внутриклеточной активности такого фермента, может быть внесена в последовательность гена, кодирующего упомянутый выше фермент с применением стандартного метода мутагенеза или генетической инженерии.
Примеры методов мутагенеза включают, например, методы, основанные на использовании рентгеновского или ультрафиолетового облучения, методы, связанные с обработкой мутагенным агентом, таким как N-метил-N'-нитро-N-нитрозогуанидином, и т.д. Сайт, по которому происходит возникновение мутации в данном гене, может приходиться на кодирующий сегмент, детерминирующий этот фермент, или на сегмент, регулирующий его экспрессию, такой, каковым является промотор.
Примеры методов генетической инженерии включают, например, методы, основанные на генной рекомбинации, трансдукции, слияния клеток и т.д. Например, ген резистентности к химическому соединению встраивают в состав клонированного гена-мишени с целью конструирования гена, который утрачивает свои исходные функции (дефектный ген). После этого такой дефектный ген вносят в клетку микроорганизма-хозяина, в результате чего ген-мишень из состава генома заменяется на упомянутый выше дефектный ген в результате прохождения гомологичной рекомбинации (этап «разрушения гена»).
Снижение или нарушение внутриклеточной активности фермента-мишени и степень снижения такой активности может быть определена путем измерения уровня активности этого фермента в клеточном экстракте или в его очищенной фракции, выделенных из анализируемого штамма с последующим сравнением со штаммом дикого типа. Например, активность фермента α-KGDH может быть измерена с применением метода Рида с соавт. (L.J.Reed & В.B.Mukherjee, 1969, Meth. Enzymol., 13, pp.55-61).
В зависимости от фермента-мишени являющийся мишенью мутантный штамм может быть выбран, исходя из фенотипа такого мутантного штамма. Например, мутантный штамм, дефицитный по активности α-KGDH или характеризующийся сниженной активностью α-KGDH, не может размножаться или характеризуется существенно сниженным пролиферационным индексом в минимальной среде, содержащей глюкозу, или в минимальной среде, содержащей уксусную кислоту или L-глутаминовую кислоту в качестве единственного источника углерода, в аэробных условиях. Однако нормальное размножение становится возможным даже в таких же условиях после добавления янтарной кислоты или лизина, метионина и диаминопимеловой кислоты к минимальной среде, содержащей глюкозу. С использованием этого эффекта в качестве индикаторного могут быть отобраны мутантные штаммы, характеризующиеся сниженной активностью α-KGDH или дефицитные по такой активности.
Способ получения дефицитного по α-KGDH штамма Brevibacterium lactofermentum с применением гомологичной рекомбинации подробно описан в международной патентной заявке WO 95/34672. Сходные способы могут быть применены в отношении и других микроорганизмов.
Далее, такие методы, как клонирование генов и расщепление и лигирование ДНК, трансформация и т.д. подробно описаны в руководстве «Molecular Cloning», 2d ed., Cold Spring Harbor Press (1989) и других.
В качестве конкретного примера мутантного штамма, дефицитного по активности фермента α-KGDH или характеризующегося сниженной активностью α-KGDH, полученного в соответствии с описанным выше, можно упомянуть штамм AJ13356 Enterobacter agglomerans. Штамм AJ13356 Enterobacter agglomerans был внесен в коллекцию Национального института биологии и биотехнологии человека Агентства по науке и технологии в промышленности Министерства международной торговли и промышленности Японии (по адресу 1-3 Higashi 1-chome, Tsukuba-shi, Ibaraki 305-8566, Japan) 19 февраля 1998 года с получением депозитарного № FERM Р-16645. Затем на основании Будапештского соглашения он был внесен в международный депозитарий 11 января 1999 года с получением депозитарного №FERM BP-6615. Штамм AJ13356 Enterobacter agglomerans дефицитен по активности фермента α-KGDH в результате разрушения гена, кодирующего Е1-субъединицу α-KGDH (ген sucA).
Когда Enterobacter agglomerans, как пример микроорганизма по настоящему изобретению, культивируют в среде, содержащей сахариды, то при этом во внеклеточное пространство секретируется вязкий материал, в результате чего снижается производственная эффективность. Следовательно, при использовании Enterobacter agglomerans, обладающей таким свойством секретировать вязкий материал, предпочтительным является использование такого мутантного штамма, который бы секретировал меньшее количество вязкого материала по сравнению со штаммом дикого типа. Примеры методов мутагенеза включают, например, методы, основанные на использовании рентгеновского или ультрафиолетового облучения, методы, связанные с обработкой мутагенным агентом, таким как N-метил-N'-нитро-N-нитрозогуанидином, и т.д. Мутантный штамм, обладающий сниженным уровнем секреции вязкого материала, может быть отобран путем инокуляции мутировавших бактериальных клеток в среде, содержащей сахариды, например, на пластине среды LB, содержащей 5 г/л глюкозы, с культивированием его при наклоне культуральной пластины на 45 градусов, отбирая колонию, которая не смывается с пластины жидкостью.
В настоящем изобретении способствование или усиление способности к выработке L-глутаминовой кислоты и придания других важных свойств, таких как мутирование в сторону секреции менее вязкого материала, описанного выше, могут быть осуществлены в произвольном порядке.
Путем культивирования микроорганизма по настоящему изобретению в жидкой культуральной среде, величина рН которой доведена до значения, при котором L-глутаминовая кислота осаждается, L-глутаминовая кислота может быть выработана и накоплена с осаждением в культуральной среде. L-глутаминовая кислота также может быть осаждена путем постановки данной культуры при нейтральном рН и затем продолжения и завершения культивирования при рН, при котором L-глутаминовая кислота осаждается.
Величина рН, при которой L-глутаминовая кислота осаждается, означает ту величину, при которой L-глутаминовая кислота осаждается тогда, когда микроорганизм вырабатывает и накапливает L-глутаминовую кислоту.
В качестве упоминавшейся выше культуральной среды обычная питательная среда, содержащая источник углерода, источник азота, минеральные соли и по необходимости следы органических компонентов, такие как аминокислоты и витамины, может быть использована так, лишь бы величина рН была доведена до того значения, при котором происходит осаждение L-глутаминовой кислоты. Может быть использована как синтетическая культуральная среда, так и естественная культуральная среда. Источник углерода и источник азота в составе культуральной среды могут быть любыми, если только они могут быть утилизированы культивируемым штаммом.
В качестве источника углерода используются сахариды, такие как глюкоза, глицерин, фруктоза, сахароза, мальтоза, манноза, галактоза, гидролизат крахмала и меласса. Кроме того, органические кислоты, такие как уксусная кислота и лимонная кислота, могут быть использованы как по отдельности, так и в сочетании с другим источником углерода.
В качестве источника азота используются аммиак, соли аммония, такие как сульфат аммония, карбонат аммония, хлорид аммония, фосфат аммония и ацетат аммония, нитраты и т.д.
В качестве следовых органических компонентов используются аминокислоты, витамины, жирные кислоты, нуклеиновые кислоты, вещества, содержащие эти соединения, такие как пептон, кас-аминокислоты, дрожжевые экстракты и продукты расщепления соевых белков. Когда используется штамм, являющийся ауксотрофным мутантом, который нуждается в аминокислоте и чем-либо подобном для своих метаболических процессов или роста, тогда добавление такого питательного компонента является обязательным.
В качестве минеральных солей используются фосфаты, соли магния, соли кальция, соли железа, соли марганца и т.д.
В качестве метода культивирования аэрационная культура обычно осуществляется при контроле температуры ферментации, которая составляет 20-42°С при рН=3-5, предпочтительно 4-5, более предпочтительно 4-4,7 и более предпочтительно 4-4,5. Таким образом, после культивирования в течение примерно от 10 часов до 4 дней в культуре накапливается существенное количество L-глутаминовой кислоты. Накопленная L-глутаминовая кислота, превышающая количество, соответствующее концентрации насыщения, осаждается в культуральной среде.
По завершении культивирования осажденная в культуре L-глутаминовая кислота может быть собрана с помощью центрифугирования, фильтрования и подобного. Растворенная в культуральной среде L-глутаминовая кислота может быть собрана в соответствии с известными методами. Например, L-глутаминовая кислота может быть выделена путем концентрирования культурального бульона до кристаллизации или выделена с применением ион-обменной хроматографии или подобного. Осажденная в культуральном бульоне L-глутаминовая кислота может быть выделена вместе с L-глутаминовой кислотой, которая была растворена в культуральной среде, после проведения кристаллизации.
В соответствии со способом по настоящему изобретению количество L-глутаминовой кислоты, превосходящее то ее количество, которое соответствует концентрации насыщения, осаждается, а концентрация L-глутаминовой кислоты, растворенной в культуральной среде, сохраняется на постоянном уровне. Следовательно влияние высокой концентрации L-глутаминовой кислоты на микроорганизмы может быть снижено. Соответственно становится возможным воспроизводить микроорганизм, обладающий улучшенными свойствами по выработке L-глутаминовой кислоты. Кроме того, поскольку L-глутаминовая кислота осаждается в виде кристаллов, то подкисление культурального бульона из-за накопления L-глутаминовой кислоты подавляется: следовательно, количество щелочи, используемой для поддержания рН данной культуры, может быть в существенной степени снижено.
Примеры
Здесь и далее настоящее изобретение будет более подробно описано путем отсылки к нижеследующим примерам.
Пример 1
Скрининг микроорганизма, характеризующегося резистентностью к L-глутаминовой кислоте в кислой среде
Скрининг микроорганизма, характеризующегося резистентностью к L-глутаминовой кислоте в кислой среде, осуществляли следующим образом. Каждый из примерно 500 природных образцов, включая пробы почвы, плодов растений, частей растений, речной воды в количестве 1 г каждая суспендировали в 5 мл стерилизованной воды, после чего 200 мкл наносили на 20 мл твердой культуральной среды, рН которой доводили до 4,0 с помощью соляной кислоты. Состав данной культуральной среды был следующим: 3 г/л глюкозы, 1 г/л (NH4)2SO4, 0,2 г/л MgSO4·7Н2O, 0,5 г/л КН2РО4, 0,2 г/л NaCl, 0,1 г/л CaCl2·7H2O, 0,01 г/л FeSO4·7H2O, 0,01 г/л MnSO4·4H2O, 0,72 мг/л ZnSO4·2H2O, 0,64 мг/л CuSO4·5H2O, 0,72 мг/л CoCl2·6H2O, 0,4 мг/л борной кислоты, 1,2 мг/л Na2MoO4·2H2O, 50 мкг/мл биотина, 50 мкг/л пантотената кальция, 50 мкг/л фолиевой кислоты, 50 мкг/л инозитола, 50 мкг/л ниацина, 50 мкг/л р-аминобензольной кислоты, 50 мкг/л пиридоксина гидрохлорида, 50 мкг/л рибофлавина, 50 мкг/л тиамина гидрохлорида, 50 мг/л циклогексимида и 20 г/л агара.
Культуральную среду, на которую высевали упомянутые выше образцы, инкубировали при 28°С, 37°С или 50°С в течение 2-4 дней: были получены 378 штаммов, каждый из которых образовывал колонии.
После этого каждый из этих штаммов, полученный в соответствии с описанным выше, вносили в тест-пробирку длиной 16,5 см и диаметром 14 мм, содержащую 3 мл жидкой культуральной среды (с доведением рН до 4,0 с помощью соляной кислоты), содержащей L-глутаминовую кислоту в концентрации насыщения, и культивировали при 28°С, 37°С или 50°С в течение от 24 часов до 3 дней при встряхивании. Затем отбирали выросшие штаммы. Состав упомянутой среды был следующим: 40 г/л глюкозы, 20 г/л (NH4)2SO4, 0,5 г/л MgSO4·7H2O, 2 г/л КН2PO4, 0,5 г/л NaCl, 0,25 г/л CaCl2·H2O, 0/02 г/л FeSO4·7H2O, 0,02 г/л MnSO4·4H2O, 0,72 мг/л ZnSO4·2H2O, 0,64 мг/л CuSO4·5H2O, 0,72 мг/л CoCl2·6H2O, 0,4 мг/л борной кислоты, 1,2 мг/л Na2MoO4·2H2O, 2 г/л дрожжевого экстракта.
В результате были получены 78 штаммов микроорганизмов, обладающих резистентностью к L-глутаминовой кислоте в кислой среде.
Пример 2
Отбор штаммов, обладающих лучшим ростом в кислой среде, среди микроорганизмов, характеризующихся резистентностью к L-глутаминовой кислоте
Различные микроорганизмы, обладающие резистентностью к L-глутаминовой кислоте в кислой среде, полученные в соответствии с описанным выше, были внесены в тест-пробирку длиной 16,5 см и диаметром 14 мм, содержащую 3 мл культуральной среды (с доведением рН до 4,0 с помощью соляной кислоты), полученной путем добавления 20 г/л глутаминовой кислоты и 2 г/л глюкозы к среде М9 (J.Sambrook, E.F.Fritsh & T.Maniatis, 1989, «Molecular Cloning», Cold Spring Harbor Lab. Press): мутность полученного раствора измеряли в зависимости от времени с целью выбора штаммов, обладающих лучшими показателями роста. В результате в качестве штамма, обладающего наилучшим темпом роста, был выделен штамм АJ13355, происходящий из почвы района Ивата (провинция Шицуока, Япония). Этот штамм был определен по принадлежности к виду Enterobacter agglomerans на основе его бактериологических параметров, описанных выше.
Пример 3
Получение штамма, характеризующегося меньшим уровнем секреции вязкого материала, из штамма AJ13355 Enterobacter agglomerans
Поскольку штамм AJ13355 Enterobacter agglomerans секретирует во внеклеточное пространство вязкий материал в случае его культивирования в среде, содержащей сахариды, производственная эффективность оказывается невысокой. Поэтому штамм, характеризующийся секрецией менее вязкого материала, был получен с помощью ультрафиолетового облучения (J.H.Miller et al., 1992, «A Short Course in Bacterial Genetics: Laboratory Manual», Cold Spring Harbor Lab. Press, p.150).
Штамм AJ13355 Enterobacter agglomerans был облучен ультрафиолетом в течение 2 минут при расстоянии в 60 см от 60-ваттной ультрафиолетовой лампы с последующим культивированием в среде LB в течение ночи с целью фиксации мутации. Мутировавший штамм разбавляли и вносили в среду LB, содержащую 5 г/л глюкозы и 20 г/л агара таким образом, что примерно 100 колоний на одной пластине могли образовываться и культивироваться при 30°С в течение ночи при том, что эту пластину наклоняли на 45°: затем отбирали 20 колоний, не проявлявших стекания вязкого материала.
Из отобранных, как отмечено выше, штаммов был выбран штамм SC17, который удовлетворял условиям отсутствия появления ревертантов даже после 5 пересевов на среду LB, содержащую 5 г/л глюкозы и 20 г/л агара и подтверждения параметров роста, ранее установленных для родительского штамма в среде LB, являющейся средой LB, содержащей 5 г/л глюкозы и среду М9 (J.Sambrook et al. 1989, «Molecular Cloning», Cold Spring Harbor Lab. Press), к которой добавляли 20 г/л L-глутаминовой кислоты и 2 г/л глюкозы и рН которой доводили до 4,5 с помощью соляной кислоты.
Пример 4
Конструирование вырабатывающей глутаминовую кислоту бактерии на материале штамма SC17 Enterobacterium agglomerans
(1) Получение дефицитного по α-KGDH штамма на материале штамма SC17 Enterobacterium agglomerans
Штамм, дефицитный по ферменту α-KGDH и характеризующийся усиленной системой биосинтеза L-глутаминовой кислоты, был получен на материале штамма SC17 Enterobacterium agglomerans.
(i) Клонирование гена α-KGDH (здесь и далее обозначаемого как sucAB) штамма AJ13355 Enterobacterium agglomerans
Ген sucAB бактерии Enterobacter agglomerans штамма AJ13355 был клонирован путем отбора фрагмента ДНК, комплементирующего свойство неспособности ассимилировать уксусную кислоту штамма кишечной палочки E.coli, дефицитного по гену E1-субъединицы фермента α-KGDH (здесь и далее обозначаемого как sucA), с помощью хромосомной ДНК штамма AJ13355 Enterobacter agglomerans.
Хромосомная ДНК штамма AJ13355 Enterobacter agglomerans была выделена с помощью метода, обычно применяемого при экстракции хромосомной ДНК из Escherichia coli («Text for Bioengineering Experiments», ed. Soc. Biosci. & Bioeng., Baifukan, Japan, 1992, pp.97-98). В качестве вектора использовали плазмиду pTWV228 (резистентной к ампициллину), доступную на коммерческой основе от Takara Shuzo Co. Ltd.
Хромосомную ДНК штамма AJ13355, расщепленную рестриктазой ЕсоТ221, и плазмиду pTWV228, расщепленную рестриктазой PstI, лигировали друг на друга с использованием лигазы фага Т4 и использовали для трансформации дефицитного по гену sucA штамма JRG465 Escherichia coli (J.Herbert et al., 1969, Mol. Gen. Genet., 105, 182). Среди полученных в соответствии с описанным выше трансформированных штаммов отбирали штамм, растущий в ацетатной минимальной среде, а из него экстрагировали плазмиду, которую обозначили как pTWVEK101. У клеток Escherichia coli штамма JRG465, несущих плазмиду pTWVEK101, восстанавливалась ауксотрофность по янтарной кислоте или L-лизину и L-метионину помимо свойства накапливать уксусную кислоту. Это предполагает, что в составе плазмиды pTWVEK101 имеется ген sucA энтеробактерии Enterobacter agglomerans.
На фиг.1 показана рестрикционная карта ДНК-фрагмента, производного от Enterobacter agglomerans, из состава плазмиды pTWVEK101. Полученная нуклеотидная последовательность участка, показанного на фиг.1, приведена как SEQ ID NO: 1. В составе этой последовательности были обнаружены нуклеотидные последовательности, предположительно представляющие две полноразмерные открытые рамки считывания (ORF), и две нуклеотидные последовательности, предположительно являющиеся частичными последовательностями обнаруженных ORF. В SEQ ID NOS: 2-5 показаны аминокислотные последовательности, которые могут кодироваться указанными открытыми рамками или их частичными последовательностями в направлении от их 5'-конца. В результате тестирования параметров их гомологии было установлено, что сегмент обнаруженной нуклеотидной последовательности, включая 3'-концевую частичную последовательность гена Fe,S-сукцинатдегидрогеназы (sghB), полноразмерные гены sucA и Е2-субъединицы α-KGDH (sucB), и 5' концевую частичную последовательность гена sucC, кодирующего β-субъединицу сукцинил-КоА-синтетазы. На фиг.2-5 показаны результаты сравнения аминокислотных последовательностей, выведенных из этих нуклеотидных последовательностей, с полипептидами кишечной палочки E.coli (Eur. J. Biochem., 141, pp.351-359, 1984; Eur. J. Biochem., 141, pp.361-374, 1984; Biochemistry, 24, pp.6245-6252, 1985). Таким образом, в каждом случае аминокислотные последовательности проявляют очень высокий уровень гомологии. Кроме того, было установлено, что в геноме Enterobacter agglomerans имеется такой же генный кластер sdhB-sucA-sucB-sucC, что и на хромосоме Escherichia coli (Eur. J. Biochem., 141, pp.351-359, 1984; Eur. J. Biochem., 141, pp.361-374, 1984; Biochemistry, 24, pp.6245-6252, 1985).
(ii) Получение дефицитного по ферменту α-KGDH штамма, производного от штамма SC17 Enterobacter agglomerans
Гомологичная рекомбинация была осуществлена с использованием гена sucAB Enterobacter agglomerans, полученного в соответствии с описанным выше, с целью получения дефицитного по ферменту α-KGDH штамма Enterobacter agglomerans.
После расщепления плазмиды pTWVEK101 рестриктазой SphI с целью вырезания фрагмента, включающего ген sucA, полученный фрагмент «затупляли» по концам с помощью фрагмента Кленова (Takara Shuzo Co. Ltd.) и с использованием ДНК-лигазы Т4 (Takara Shuzo Co. Ltd.) лигировали его в состав плазмиды pBR322, предварительно расщепленной рестриктазой EcoRI и «затупленной» по концам с помощью фрагмента Кленова. Полученную плазмиду расщепляли по сайту распознавания рестриктазой BglII, расположенному в самом центре гена sucA, с использованием именно этой рестриктазы, «затупляли» концы с помощью фрагмента Кленова и затем опять лигировали с использованием ДНК-лигазы Т4. Предполагалось, что ген sucA не функционировал из-за мутации типа «сдвига рамки», которая была внесена в ген sucA, находящийся в составе плазмиды, сконструированной в описанной выше процедуре.
Плазмиду, сконструированную в соответствии с описанным выше, расщепляли рестриктазой ApaLI и подвергали электрофорезу в агарозном геле с целью выделения фрагмента ДНК, включающего ген sucA, в составе которого имеется мутация «сдвига рамки», а также ген резистентности к тетрациклину, происходящий из плазмиды pBR322. Выделенный фрагмент ДНК опять лигировали с использованием ДНК-лигазы Т4 с получением плазмиды, предназначенной для разрушения гена α-KGDH.
Плазмиду, предназначенную для разрушения гена α-KGDH, полученную в соответствии с описанным выше, использовали для трансформации бактерии Enterobacter agglomerans штамма SC17 путем электропорации (J.H.Miller, 1992, «A Short Course in Bacterial Genetics: Handbook», Cold Spring Harbor Lab. Press, p.279): штамм, у которого ген sucA на его хромосоме оказывался заменен на мутантный тип гена в результате гомологичной рекомбинации с плазмидой, получали с использованием в качестве индикатора признака резистентности к тетрациклину. Полученный штамм обозначили как штамм SC17sucA.
С целью подтверждения того, что штамм SC17sucA дефицитен по активности фермента α-KGDH, активность этого фермента была определена по методу Рида с соавт. (L.J.Reed & B.B.Mukherjee, 1969, Meth. Enzymol., 13, pp.55-61) с использованием клеток этого штамма, культивируемых в среде LB до логарифмической фазы роста. В результате у штамма активность фермента OC-KGDH была определена на уровне 0,073 (Δ ABS/мин на 1 мг белка) для штамма SC17, в то время как у штамма SC17sucA активность α-KGDH полностью отсутствовала: следовательно, это подтверждает, что, как это и предполагалось, имеет место дефицитность по гену sucA.
(2) Усиление системы биосинтеза L-глутаминовой кислоты Enterobacter agglomerans штамма SC17sucA
В штамм SC17sucA последовательно вносили ген цитрат-синтазы, ген фосфоенолпируваткарбоксилазы и глутаматдегидрогеназы, происходящие от Escherichia coli.
(i) Получение плазмиды, несущей ген gltA, ген ррс и ген gdhA, производные от Escherichia coli
Процедуры получения плазмиды, несущей ген gltA, ген ррс и ген gdhA, будут описаны со ссылками на фиг.6 и 7.
Плазмиду, несущую ген gdhA, производный от Escherichia coli, - pBRGDH (открытая японская патентная заявка № 7-203980) - расщепляли рестриктазами HindIII и SphI; оба полученных конца «затупляли» с использованием ДНК-полимеразы Т4 и затем полученный фрагмент ДНК, содержащий ген gdhA, очищали и выделяли. Отдельно от этого плазмиду, включающую ген gltA и ген ррс, производные от Escherichia coli, - pMWCP (международная патентная заявка WO 97/08294) - расщепляли рестриктазой XbaI и затем оба конца «затупляли» с использованием ДНК-полимеразы Т4. Полученный фрагмент смешивали с очищенным ДНК-фрагментом, указанным выше, несущим ген gdhA, и лигировали с использованием лигазы Т4 с получением плазмиды pMWCPG, которая соответствует плазмиде pMWCP, дополнительно включающей ген gdhA (фиг.6).
В то же время плазмиду pVIC40 (открытая японская патентная заявка № 8-047397), включающую сайт начала репликации, активный в широком круге организмов-хозяев из состава плазмиды RSF101, расщепляли рестриктазой NotI, обрабатывали ДНК-полимеразой Т4 и расщепляли рестриктазой PstI. Плазмиду pBR322 расщепляли рестриктазой ЕсоТ14I, обрабатывали ДНК-полимеразой Т4 и расщепляли рестриктазой PstI. Оба полученных продукта смешивали друг с другом и лигировали друг на друга с использованием лигазы Т4 с получением плазмиды RSF-Tet, включающей сайт начала репликации, RSF1010, и ген резистентности к тетрациклину (фиг.7).
Далее плазмиду pMWCPG расщепляли рестриктазами EcoRI и PstI с последующим выделением и очисткой фрагмента ДНК, включающего ген gltA, ген ррс и ген gdhA. Плазмиду RSF-Tet сходным образом расщепляли рестриктазами EcoRI и PstI с последующим выделением и очисткой фрагмента ДНК, включающего сайт начала репликации, происходящий от RSF1010. Оба полученных продукта смешивали и лигировали друг на друга с использованием лигазы Т4 с получением плазмиды RSFCPG, которая соответствует плазмиде RSF-Tet, дополнительно включающей ген gltA, ген ррс и ген gdhA (фиг.8). То, что полученная плазмида RSFCPG экспрессирует ген gltA, ген ррс и ген gdhA, было подтверждено с применением теста на комплементирование ауксотрофности у дефицитного по генам gltA, ррс или gdhA штамма, производного от Escherichia coli, с последующим измерением активности каждого из соответствующих ферментов.
(ii) Получение плазмиды, включающей ген gltA, производный от Brevibacterium lactofermentum
Плазмида, включающая ген gltA, происходящий из генома Brevibacterium lactofermentum, была сконструирована следующим образом. ПЦР была осуществлена с использованием затравок ДНК, имеющих нуклеотидные последовательности, которые показаны в SEQ ID NO 6 и 7, которые были получены на основе параметров нуклеотидной последовательности гена gltA коринебактерии Corynebacterium glutamicum (Microbiology, 140, pp.1817-1828, 1994), используя хромосомную ДНК Brevibacterium lactofermentum штамма АТСС13869 в качестве матрицы, с целью получения фрагмента гена gltA длиной примерно 3 тысячи пар нуклеотидов. Этот фрагмент встраивали в плазмиду pHSG399 (приобретена у Takara Shuzo Co. Ltd.), расщепленную рестриктазой Smal, с получением плазмиды pHSGCB (фиг.9). После этого плазмиду pHSGCB расщепляли рестриктазой HindIII и вырезали фрагмент гена gltA длиной примерно 3 тысячи пар нуклеотидов, встроенный в плазмиду pSTV29 (приобретена у Takara Shuzo Co. Ltd.), расщепленную рестриктазой HindIII, с получением плазмиды pSTVCB (фиг.9). То, что полученная плазмида pSTVCB экспрессирует ген gltA, было подтверждено с помощью измерения активности соответствующего фермента у Enterobacter agglomerans штамма AJ13355.
(iii) Внесение плазмид RSFCPG и pSTVCB в штамм SC17sucA
Штамм SC17sucA Enterobacter agglomerans был трансформирован с использованием плазмиды RSFCPG с применением электропорации с получением трансформированного штамма SC17sucA/RSFCPG, характеризующегося резистентностью к тетрациклину. Далее, штамм SC17sucA/-RSFCPG был трансформирован с использованием плазмиды pSTVCB с применением электропорации с получением трансформированного штамма SC17sucA/RSFCPG+pSTVCB, характеризующегося резистентностью к хлорамфениколу.
Пример 5
Получение штамма, характеризующегося улучшенной резистентностью к L-глутаминовой кислоте в среде с низким значением рН
Штамм, характеризующийся улучшенной резистентностью к высокой концентрации L-глутаминовой кислоты в среде с низким значением рН (здесь и далее также обозначаемый как «резистентный штамм к высокой концентрации Glu при низком рН») был выделен на материале Enterobacter agglomerans штамма SC17sucA/RSFCPG+pSTVCB.
Штамм SC17sucA/RSFCPG+pSTVCB культивировали в течение ночи при 30°С в среде LBG (10 г/л триптона, 5 г/л дрожжевого экстракта, 10 г/л NaCl, 5 г/л глюкозы), затем клетки, промытые солевым раствором, подходящим образом разбавляли и высевали на пластину среды М9-Е (4 г/л глюкозы, 17 г/л Na2HPO4·12Н2O, 3 г/л КН2PO4, 0,5 г/л NaCl, 1 г/л NH4Cl, 10 мМ MgSO4, 10 мкМ CaCl2, 50 мг/л L-лизина, 50 мг/л L-метионина, 50 мг/л DL-диаминопимеловой кислоты, 25 мг/л тетрациклина, 25 мг/л хлорамфеникола, 30 г/л L-глутаминовой кислоты; рН доводят до 4,5 с помощью водного раствора аммиака). Колонию, появившуюся после культивирования при 32°С в течение 2 дней, выделяли как колонию, характеризующуюся резистентностью к высокой концентрации Glu при низком рН.
Для полученного штамма определяли темп роста в жидкой культуральной среде М9-Е и тестировали способность к выработке L-глутаминовой кислоты в больших тест-пробирках объемом 50 мл, содержащих по 5 мл среды для тестирования выработки L-глутаминовой кислоты (40 г/л глюкозы, 20 г/л (NH4)2SO4, 0,5 г/л MgSO4·7H2O, 2 г/л КН2PO4, 0,5 г/л NaCl, 0,25 г/л CaCl2·7H2O, 0,02 г/л FeSO4·7H2O, 0,02 г/л MnSO4·4H2O, 0,72 мг/л ZnSO4·2H2O, 0,64 мг/л CuSO4·5H2O, 0,72 мг/л CoCl2·6H2O, 0,4 мг/л борной кислоты, 1,2 мг/л Na2MoO4·2Н2O, 2 г/л дрожжевого экстракта, 200 мг/л L-лизина гидрохлорида, 200 мг/л L-метионина, 200 мг/л DL-α,ε-диаминопимеловой кислоты, 25 мг/л тетрациклина гидрохлорида, 25 мг/л хлорамфеникола). Штамм, который проявлял наилучший темп роста и такую же способность вырабатывать L-глутаминовую кислоту, что и родительский штамм - штамм SC17sucA/RSFCPG+pSTVCB, - был обозначен как штамм AJ13601 Enterobacter agglomerans. Штамм AJ13601 был внесен в коллекцию Национального института биологии и биотехнологии человека Агентства по науке и технологии в промышленности Министерства международной торговли и промышленности Японии (по адресу 1-3 Higashi 1-chome, Tsu-kuba-shi, Ibaraki 305-8566, Japan) 18 августа 1999 года с получением депозитарного №FERM Р-17516. Затем на основании Будапештского соглашения он был внесен в международный депозитарий 6 июля 2000 года с получением депозитарного №FERM BP-7207.
Пример 6
Культивирование Enterobacter agglomerans штамма AJ13601 для выработки L-глутаминовой кислоты
Штамм AJ13601 Enterobacter agglomerans высевали в ферментационные колбы 1-L, содержащие по 300 мл среды, включающей 40 г/л глюкозы, 20 г/л (NH4)2SO4, 0,5 г/л MgSO4·7Н2О, 2 г/л КН2PO4, 0,5 г/л NaCl, 0,25 г/л CaCl2·7H2O, 0,02 г/л FeSO4·7H2O, 0,02 г/л MnSO4·4H2O, 0,72 мг/л ZnSO4·2H2O, 0,64 мг/л CuSO4·5H2O, 0,72 мг/л CoCl2·6H2O, 0,4 мг/л борной кислоты, 1,2 мг/л Na2MoO4·2H2O, 2 г/л дрожжевого экстракта, 200 мг/л L-лизина гидрохлорида, 200 мг/л L-метионина, 200 мг/л DL-α,ε-ди-аминопимеловой кислоты, 25 мг/л тетрациклина гидрохлорида, 25 мг/л хлорамфеникола, и культивировали при 34°С в течение 14 часов при рН=6,0. Величину рН этой культуры контролировали путем внесения газообразного аммиака в культуральную среду.
Культуру, полученную в соответствии с описанным выше, центрифугировали при 5000 об/мин в течение 10 минут и собранные клетки высевали в ферментационные колбы 1-L, содержащие по 300 мл культуральной среды, включающей 40 г/л глюкозы, 5 г/л (NH4)2SO4, 1,5 г/л MgSO4·7H2O, 6 г/л КН2PO4, 1,5 г/л NaCl, 0,75 г/л CaCl2·7H2O, 0,06 г/л FeSO4·7H2O, 0,06 г/л MnSO4·4H2O, 2,16 мг/л ZnSO4·2H2O, 1,92 мг/л CuSO4·5H2O, 2,16 мг/л CoCl2·6H2O, 1,2 мг/л борной кислоты, 3,6 мг/л Na2MoO4·2H2O, 6 г/л дрожжевого экстракта, 600 мг/л L-лизина гидрохлорида, 600 мг/л L-метионина, 600 мг/л DL-α,ε-диаминопимеловой кислоты, 25 мг/л тетрациклина гидрохлорида, 25 мг/л хлорамфеникола, и культивировали при 34°С и при рН=4,5 с целью обеспечения выработки культурой L-глутаминовой кислоты. Величину рН этой культуры контролировали путем внесения газообразного аммиака в культуральную среду. После того, как исходно включаемая глюкоза истощалась, добавляли 600 г/л глюкозы.
В результате культивирования на выработку L-глутаминовой кислоты, осуществленного в течение 50 часов в соответствии с описанным выше, существенное количество кристаллов L-глутаминовой кислоты было осаждено в ферментационной колбе. В табл.1 показана концентрация L-глутаминовой кислоты, растворенной в культуральном бульоне в данный момент времени, и концентрация L-глутаминовой кислоты, определенной путем растворения кристаллов в 2 М едкого калия. Кристаллы L-глутаминовой кислоты собирали из культуры путем декантации после остановки процесса культивирования.
Пример 7
Культивирование Enterobacter agglomerans штамма AJ13601 для выработки L-глутаминовой кислоты
Следующий эксперимент был осуществлен с целью подтверждения того, что Enterobacter agglomerans штамма AJ13601 уже обладает способностью вырабатывать L-глутаминовую кислоту даже в таких условиях, когда присутствуют кристаллы L-глутаминовой кислоты.
Штамм AJ13601 Enterobacter agglomerans высевали в ферментационные колбы 1-L, содержащие по 300 мл среды, включающей 40 г/л глюкозы, 20 г/л (NH4)2SO4, 0,5 г/л MgSO4·7Н2O, 2 г/л КН2PO4, 0,5 г/л NaCl, 0,25 г/л CaCl2·7H2O, 0,02 г/л FeSO4·7H2O, 0,02 г/л MnSO4·4H2O, 0,72 мг/л ZnSO4·2H2O, 0,64 мг/л CuSO4·5H2O, 0,72 мг/л CoCl2·6Н2О, 0,4 мг/л борной кислоты, 1,2 мг/л Na2MoO4·2H2O, 2 г/л дрожжевого экстракта, 200 мг/л L-лизина гидрохлорида, 200 мг/л L-метионина, 200 мг/л DL-α,ε-диаминопимеловой кислоты, 25 мг/л тетрациклина гидрохлорида, 25 мг/л хлорамфеникола, и культивировали при 34°С в течение 14 часов при рН=6,0. Величину рН этой культуры контролировали путем барботажа культуральной среды газообразным аммиаком. Культуру, полученную в соответствии с описанным выше, центрифугировали при 5000 об/мин в течение 10 минут и затем отобранные клетки культивировали в среде, в которой L-глутаминовая кислота содержится в виде кристаллов. Использовавшаяся среда включала 40 г/л глюкозы, 5 г/л (NH4)2SO4, 1,5 г/л MgSO4·7H2O, 6 г/л КН2PO4, 1,5 г/л NaCl, 0,75 г/л CaCl2·7H2O, 0,06 г/л FeSO4·7H2O, 0,06 г/л MnSO4·4H2O, 2,16 мг/л ZnSO4·2H2O, 1,92 мг/л CuSO4·5H2O, 2,16 мг/л CoCl2·6H2O, 1,2 мг/л борной кислоты, 3,6 мг/л Na2MoO4·2H2O, 6 г/л дрожжевого экстракта, 600 мг/л L-лизина гидрохлорида, 600 мг/л L-метионина, 600 мг/л DL-α,ε-диаминопимеловой кислоты, 25 мг/л тетрациклина гидрохлорида, 25 мг/л хлорамфеникола с добавлением кристаллов L-глутаминовой кислоты в количестве 40 г/л. Клетки высевали в ферментационную колбу 1-L, содержащую 300 мл такой среды, и культивировали при 34°С и при рН=4,3 с целью обусловливания выработки этой культуры L-глутаминовой кислоты. Величину рН этой культуры контролировали путем подачи газообразного аммиака в культуральную среду. После того, как исходно включаемая глюкоза истощалась, добавляли 600 г/л глюкозы. В данной среде только 39 г/л добавленной L-глутаминовой кислоты растворялись при рН=4,3, а оставшиеся 1 г/л присутствовали в виде кристаллов.
В результате культивирования на выработку L-глутаминовой кислоты, осуществленного в течение 53 часов в соответствии с описанным выше, существенное количество кристаллов L-глутаминовой кислоты было осаждено в ферментационной колбе. В табл.2 показана концентрация L-глутаминовой кислоты, растворенной в культуральном бульоне, количество L-глутаминовой кислоты, присутствующей в виде кристаллов в тот же момент времени, и концентрация L-глутаминовой кислоты, определенной путем растворения кристаллов в 2 М КОН. Кристаллы L-глутаминовой кислоты собирали из культуры путем декантации после остановки процесса культивирования. Полученные результаты показали, что Enterobacter agglomerans штамма AJ13601 накапливает L-глутаминовую кислоту и осаждает ее в виде кристаллов даже в условиях присутствия кристаллов L-глутаминовой кислоты.
Пример 8
Культивирование штамма AJ13601 Enterobacter agglomerans для выработки L-глутаминовой кислоты (3)
Штамм AJ13601 Enterobacter agglomerans может расти не только при кислых значениях рН, но также и при нейтральном рН. Следовательно, было подтверждено, что кристаллы L-глутаминовой кислоты также могут быть осаждены при исходном культивировании при нейтральном значении рН и последующей выработкой L-глутаминовой кислоты в процессе культивирования в условиях постепенного самопроизвольного снижения рН.
Клетки на одной пластине (8,5 см в диаметре) штамма AJ13601 Enterobacter agglomerans, культивировавшегося в агарсодержащей среде LBG (10 г/л триптона, 5 г/л дрожжевого экстракта, 10 г/л NaCl, 5 г/л глюкозы, 15 г/л агара), включающей 25 мг/л тетрациклина гидрохлорида и 25 мг/л хлорамфеникола, при 30°С в течение 14 часов, высевали в ферментационные колбы 1-L, содержащие по 300 мл среды, включающей 40 г/л глюкозы, 5 г/л (NH4)2SO4, 1,5 г/л MgSO4·7H2O, 6 г/л КН2PO4, 1,5 г/л NaCl, 0,75 г/л CaCl2·7H2О, 0,06 г/л FeSO4·7Н2O, 0,06 г/л MnSO4·4H2O, 2,16 мг/л ZnSO4·2H2O, 1,92 мг/л CuSO4·5Н2O, 2,16 мг/л CoCl2·6H2O, 1,2 мг/л борной кислоты, 3,6 мг/л Na2MoO4·2H2O, 6 г/л дрожжевого экстракта, 600 мг/л L-лизина гидрохлорида, 600 мг/л L-метионина, 600 мг/л DL-α,ε-диаминопимеловой кислоты, 25 мг/л тетрациклина гидрохлорида, 25 мг/л хлорамфеникола, и исходно культивировали при 34°С и при рН=7,0. Величину рН этой культуры контролировали путем введения газообразного аммиака в культуральную среду. После того, как исходно включаемая глюкоза истощалась, добавляли 600 г/л глюкозы.
По мере накопления L-глутаминовой кислоты величина рН снижалась самопроизвольно. Количество добавляемого газообразного аммиака обеспечивали таким образом, чтобы величина рН постепенно снижалась с 7,0 до 4,5 в течение периода времени между 15 и 24 часами после начала культивирования, а через 24 часа после начала культивирования величина рН достигала бы 4,5. После этого культивирование продолжали еще в течение 12 часов.
В результате культивирования на выработку L-глутаминовой кислоты, осуществленного в течение 36 часов в соответствии с описанным выше, существенное количество кристаллов L-глутаминовой кислоты было осаждено в ферментационной колбе. В табл.3 показана концентрация L-глутаминовой кислоты, растворенной в культуральном бульоне, количество L-глутаминовой кислоты, присутствующей в виде кристаллов в тот же момент времени, и концентрация L-глутаминовой кислоты, определенной путем растворения кристаллов в 2 М КОН. Кристаллы L-глутаминовой кислоты собирали из культуры путем декантации после остановки процесса культивирования.











| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ L-ГЛУТАМИНОВОЙ КИСЛОТЫ | 2002 |
|
RU2282662C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ГЛУТАМИНОВОЙ КИСЛОТЫ | 2002 |
|
RU2288271C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ГЛУТАМИНОВОЙ КИСЛОТЫ | 1999 |
|
RU2188236C2 |
| ОРГАНИЧЕСКАЯ АЗОТСОДЕРЖАЩАЯ КОМПОЗИЦИЯ И УДОБРЕНИЕ, ВКЛЮЧАЮЩЕЕ ЕЕ | 2002 |
|
RU2291139C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ГЛУТАМИНОВОЙ КИСЛОТЫ | 1999 |
|
RU2194076C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2003 |
|
RU2307165C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ СЕМЕЙСТВА ГЛУТАМАТА ИЛИ L-ВАЛИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia | 2009 |
|
RU2418064C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ ESCHERICHIA | 2005 |
|
RU2311454C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ БАКТЕРИЙ, ПРИНАДЛЕЖАЩИХ К РОДУ ESCHERICHIA | 2005 |
|
RU2304615C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН rcsA | 2006 |
|
RU2359029C2 |



Изобретение относится к биотехнологии. Предложен штамм Enterobacter agglomerans/FERM BP-7207, который способен метаболизировать источник углерода при величине рН, при которой L-глутаминовая кислота осаждается в жидкой культуральной среде, содержащей L-глутаминовую кислоту в концентрации насыщения и источник углерода, а также обладает способностью накапливать L-глутаминовую кислоту в количестве, превосходящем то количество, которое соответствует ее концентрации насыщения в жидкой культуральной среде при данном рН; и способ получения L-глутаминовой кислоты с помощью ферментации, который включает культивирование данного микроорганизма в жидкой культуральной среде, величина рН которой доводится до такой величины, при которой L-глутаминовая кислота осаждается, с получением и накоплением L-глутаминовой кислоты и осаждением L-глутаминовой кислоты в данной культуральной среде. Указанный микроорганизм получают внесением содержащего микроорганизмы образца в кислую культуральную среду, содержащую L-глутаминовую кислоту в концентрации насыщения и источник углерода, и отбором штамма, который способен метаболизировать данный источник углерода. Данное изобретение позволяет получать L-глутаминовую кислоту с высокой степенью эффективности. 4 н. и 16 з.п. ф-лы, 9 ил., 3 табл.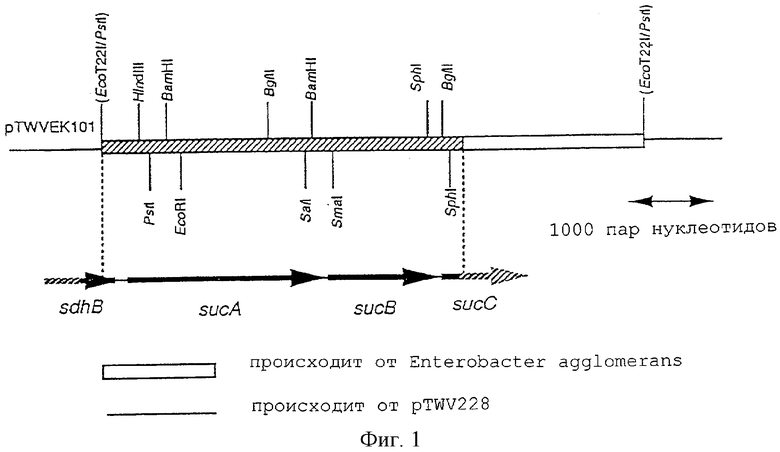
(a) микроорганизм характеризуется усиленной активностью фермента, который катализирует реакцию биосинтеза L-глутаминовой кислоты, и
(b) микроорганизм характеризуется пониженной или недостаточной активностью фермента, который катализирует реакцию, отклоняющуюся от пути биосинтеза L-глутаминовой кислоты и приводящую к образованию иного, нежели L-глутаминовая кислота, соединения.
(a) микроорганизм характеризуется усиленной активностью фермента, который катализирует реакцию биосинтеза L-глутаминовой кислоты, и
(b) микроорганизм характеризуется пониженной или недостаточной активностью фермента, который катализирует реакцию, отклоняющуюся от пути биосинтеза L-глутаминовой кислоты и приводящую к образованию иного, нежели L-глутаминовая кислота, соединения.
| US 3220929, 30.11.1965 | |||
| ШТАММ БАКТЕРИЙ CORYNEBACTERIUM GLUTAMICUM В-7198-ПРОДУЦЕНТ L-ГЛЮТАМИНОВОЙ КИСЛОТЫ | 1996 |
|
RU2107723C1 |
| Способ крашения тканей | 1922 |
|
SU62A1 |

