Перекрестная ссылка на родственные заявки
В настоящей заявке испрашивается приоритет по предварительной заявке на патент США No 60/701855, поданной 21 июля 2005, которая во всей своей полноте включена в настоящее описание посредством ссылки.
Предшествующий уровень техники
Настоящее изобретение относится к области молекулярной биологии, в частности к области экспрессии рекомбинантных белков, а также к экспрессии и процессингу, включая, в частности, посттрансляционный процессинг рекомбинантных полипротеинов или белков-предшественников.
В последние годы антитела, используемые в качестве диагностических и терапевтических средств, находят все большее применение. Моноклональное антитело ОКТ3 (Johnson and Johnson), на применение которого было получено разрешение Управления по контролю за качеством пищевых продуктов, медикаментов и косметических средств (FDA), впервые было использовано для лечения пациентов с отторжением почечного трансплантата. Герцептин (торговый знак Genentech Inc., South San Francisco, CA), то есть гуманизованное моноклональное антитело, используемое для лечения пациентов с метастазами рака молочной железы, был разрешен к применению в 1998. Множество методов лечения, разработанных на основе антител, дали многообещающие результаты на различных стадиях клинических испытаний. Одним из ограничений для широкого клинического применения технологии с использованием антител является то, что для достижения терапевтической эффективности обычно требуются большие количества антител, но продуцирование антител в достаточном количестве связано со значительными затратами. В качестве клеточных линий млекопитающих для промышленного продуцирования гликозилированных человеческих белков, таких как антитела и другие биотерапевтические средства, чаще всего используются клетки яичника китайского хомячка (CHO) и клетки миеломы NSO (Humphreys and Glover 2001. Curr. Opin. Drug Discov. Devel. 4:172-85). В процессе культивирования в течение 5-7 дней в ферментере периодического действия клеточные линии млекопитающих обычно продуцируются в количестве от 50 до 250 мг/л, а в процессе культивирования в течение 7-12 дней в ферментере периодического действия с подпиткой эти клетки продуцируются в количестве 300-600 мг/л. Негликозилированные белки иммуноглобулина могут быть успешно продуцированы в дрожжах или в E.coli (см., например, Humphreys DP, et al., 2000, Protein Expr Purif. 20(2):252-64), однако наилучшее продуцирование в бактериальных системах экспрессии достигалось с использованием фрагментов антител. (Humphreys, D.P. 2003. Curr. Opin. Drug Discov. Devel. 2003 6:188-96).
Важным этапом в области экспрессии множества генных сегментов было открытие интеинов (см., например, Hirata, R et al., 1990, J. Biol. Chem. 265:6726-6733; Kane, PM et al., 1990, Science 250:, 651 -657; Xu, M-Q and Perler, FB, 1996, EMBO Journal 15(19):5146-5153). Интеины представляют собой белковые эквиваленты интронов генных последовательностей и облегчают сплайсинг белка. Как указывается в патенте США 7026526, Snell K., сплайсинг белка представляет собой процесс, в котором внутренняя область белка-предшественника (интеина) вырезается, а фланкирующие области белка (экстеины) лигируются с образованием зрелого белка. Этот процесс наблюдается в различных белках прокариотов и эукариотов (Perler, F. B., Xu, M. Q., Paulus, H. Current Opinion in Chemical Biology 1997, 1, 292-299; Perler, F. B. Nucleic Acids Research 1999, 27, 346-347). Интеин содержит компоненты, необходимые для катализа сплайсинга белка, и часто имеет эндонуклеазный домен, который участвует в транспорте интеина (Perler, F. B., et al., Nucleic Acids Research 1994, 22, 1127-1127).
Главным назначением систем на основе интеина является разработка технологий очистки и получение новых гибридных белков в результате экспрессии генных сегментов, так, например, в патенте США 7026526 были описаны конструкции ДНК с модифицированными интеинами, используемые для экспрессии множества генных продуктов в виде отдельных белков, сообщающих множество различных признаков растениям. Однако в этом патенте не приводится каких-либо указаний на то, что эти системы могут быть с успехом использованы для экспрессии отдельных белков, которые подвергаются сборке с образованием функциональных мультимерных белков; белков, подвергаемых внеклеточной секреции; белков млекопитающих или белков, продуцируемых в эукариотических клетках-хозяевах. Примечательно, что иммуноглобулины подпадают под все эти категории.
Трудности применения метода на основе модифицированных интеинов, описанного в патенте США 7026526, к другим генам или в других целях осложняется тем, что в данном случае необходимо определить степень потенциальной важности влияния нужных экстеиновых сегментов генной последовательности на присутствующую интеиновую систему. В раболе Паулюса (Paulus) сообщается: “Действительно, на сплайсинг белка, даже если он полностью катализируется интеином, могут заметно влиять последовательности экстеинов. Такое влияние подтверждается тем фактом, что экспрессия систем сплайсинга химерного белка, в которые между чужеродными кодирующими последовательностями в той же рамке считывания были встроены последовательности интеина, часто приводит к серьезным побочным реакциям, таким как расщепление, происходящее выше или ниже границ сплайсинга (Xu M-Q, et al., 1993, Cell 75:1371-77; и Shingledecker K, et al., 1998, Gene 207:187-95). Это позволяет предположить, что способность интеинов принимать конфигурацию, оптимальную для сплайсинга белка, не вызывая при этом побочных реакций, зависит от конкретных экстеинов”. См., Paulus H, 2000, Protein splicing and related forms of protein autoproces-sing, Annu. Rev. Biochem. 69:447-96. Другой автор утверждает, что “Хотя сообщение белкам нужных свойств и активностей может быть осуществлено с применением рациональных методов конструирования, однако небольшие изменения, необходимые для эффективного и практического продуцирования сконструированного продукта, часто лежат за пределами наших возможностей (Shao, Z. and Arnold, F.H. 1996. Curr. Opin. Struct. Biol. 6, 513-518).... Тем не менее, было обнаружено, что области, непосредственно фланкирующие интеины, влияют на эффективность сплайсинга (Chong, S. et al., 1998, Nucleic Acids Res. 26, 5109-5115; Southworth, M.W. et al., 199, Biotechniques 27, 110-114) и некоторые белки-хозяева могут быть не совместимы с активностью интеинов. И хотя высокий уровень экспрессии и чистота продукта являются важными факторам, однако они не имеют существенного значения, если конечный продукт является неактивным.” См. Amitai G. and Pietrokovski, 1999, Nature Biotechnology 17:854-855.
Поэтому в модифицированной интеиновой системе, где предпочтительным результатом является расщепление без последующего лигирования, присутствие чужеродного экстеина по отношению к данной последовательности интеина может фактически влиять на эффективную комбинацию таких признаков, как точное расщепление, отсутствие повторного лигирования и отсутствие побочных реакций. Очевидно, что адаптация метода модифицированных интеинов для рекомбинантного продуцирования некоторых белков, конечные продукты которых, например иммуноглобулины и другие биотерапевтические средства, обладают функциональной активностью, представляет значительные трудности для осуществления каких-либо нововведений.
В настоящем изобретении такая проблема рассматривалась не только в связи с применением интеиновых систем, но также и с точки зрения возможности применения доменов hedgehog. Белки семейства hedgehog представляют собой межклеточные сигнальные молекулы, имеющие важное значение для формирования структур в эмбрионах млекопитающих. См., например, Mann, R. K. and Beachy, P.A. (2000) Biochim. Biophys. Acta. 1529, 188-202; Beachy, PA, (1997) Cold Spring Harb Symp Quant Biol 62: 191-204. Нативные белки-предшественники hedgehog расшепляются с образованием C-концевых (Hh-C) и N-концевых фрагментов (Hh-N) посредством реакции аутопроцессинга, которая аналогична сплайсингу белков. Система hedgehog предоставляет пока еще не исследованные возможности для творческой разработки систем, включающих модифицированные варианты, подходящие для экспрессии множества отдельных фрагментов белка.
Предпринимавшиеся ранее попытки экспрессии полноразмерной молекулы антитела/иммуглобулина методами рекомбинантных ДНК с использованием одного вектора были недостаточно успешными и обычно давали в значительной степени отличающиеся уровни экспрессии тяжелой и легкой цепей молекулы антитела/иммуноглобулина, а конкретнее, более низкий уровень экспрессии второго гена. Для других целей могут потребоваться относительно более высокие уровни экспрессии одной цепи, чем это необходимо для оптимального продуцирования мультимерных антител или их функциональных фрагментов с соответствующей укладкой. В связи с этим может возникнуть проблема, связанная с субоптимальной стехиометрией экспрессии тяжелой и легкой цепей в данной клетке, которая приводит к общему низкому выходу подвергнутого сборке мультимерного антитела. Фанг (Fang) и др. показали, что для экспрессии высоких уровней полноразмерного биологически функционального антитела из одного вектора требуется эквимолярная экспрессия тяжелой и легкой цепей (см. Fang et al., 2005, Nature Biotechnology 23:584-590; публикация заявки на патент США 2004/0265955A1). Кроме того, стандартные системы экспрессии на основе векторов, которые независимо экспрессируют множество полипептидов, в значительной степени зависят от таких факторов, как взаимодействие промоторов (например, интерференция промоторов). Такие взаимодействия могут негативно влиять на эффективность экспрессии генов и/или на сборку экспрессированных цепей, или для их осуществления может потребоваться использование более чем одного вектора (см., например, патент США 6331415, Cabilly et al.). Такая потребность в использовании нескольких векторов является серьезным недостатком, поскольку она может быть связана с некоторыми осложнениями, такими как потеря одного или нескольких отдельных векторов, а обычно - с необходимостью проведения дополнительных манипуляций.
Другим фактором, ограничивающим возможность экспрессировать две или несколько кодирующих последовательностей в одном векторе, является упаковывающая способность самого вектора. Так, например, при рассмотрении соответствующей последовательности вектора/кодирующей последовательности такими факторами являются упаковывающая способность вектора (например, приблизительно 4500 п.н. для адено-ассоциировнного вируса, AAV); продолжительность in vitro/in vivo экспрессии рекомбинантного белка в трансфицированной вектором клетке или органе (например, непродолжительное время экспрессии в аденовирусных векторах); тип клеток, способных к эффективному поддерживанию инфекции, индуцированной вектором, если используется вирусный вектор; и нужный уровень экспрессии генного(ых) продукта(ов). Требование регулируемой экспрессии двух или более генных продуктов в сочетании с ограниченной упаковывающей способностью вирусных векторов, таких как аденовирус и AAV, ограничивает выбор векторной конструкции и систем для экспрессии определенных генов, таких как гены иммуноглобулина или его фрагментов.
В других методах экспрессии двух или более последовательностей белков или полипептидных последовательностей в одном векторе для экспрессии отдельных кодирующих последовательностей используют два или более промотора или один промотор и внутренний сайт связывания с рибосомой (IRES), расположенный между двумя представляющими интерес кодирующими последовательностями. Использование двух промоторов в одном векторе может приводить к снижению уровня экспрессии белка, вызываемому интерференцией промоторов. Если две кодирующие последовательности разделены последовательностью IRES, то уровень трансляции второй кодирующей последовательности часто бывает значительно ниже уровня трансляции первой последовательности (Furler et al. 2001. Gene Therapy 8:864-873). В публикации заявки на патент США 2004/0241821 описаны флавивирусные векторы, в которых гетерологичная кодирующая последовательность расположена за последовательностью, кодирующей вирусный полипротеин, и эти последовательности отделены друг от друга последовательностью IRES. Стратегия заякоривания вектора в ядре, применяемая в целях экспрессии рекомбинантного гена, включая гибридные белки, в которых сегменты разделены сайтами узнавания протеазой, описана в публикации заявки на патент США 2005/0026137.
Связывание белков в форме полипротеинов в одной открытой рамке считывания (sORF) представляет собой механизм, наблюдаемый при репликации многих природных вирусов, включая пикорнавирусы. После трансляции кодируемые вирусом протеиназы опосредуют быстрое внутримолекулярное (цис)расщепление полипротеина с образованием дискретных продуктов зрелого белка. Вирусы ящура (FMDV) принадлежат к группе пикорнавирусов и экспрессируют одну длинную открытую рамку считывания, кодирующую полипротеин размером приблизительно 225 кД. Полноразмерный продукт трансляции подвергается быстрому внутримолекулярному (цис)расщеплению у С-конца области 2A, которое происходит между предшественником капсидного белка (Р1-2А) и репликативными доменами полипротеина 2BC и P3, и это расщепление опосредуется самой областью 2A по механизму прерывания действия рибосомы (Ryan et al. 1991. J. Gen. Virol. 72:2727-2732); Vakharia et al. 1987. J. Virol. 61:3199-3207). Были идентифицированы остатки незаменимых аминокислот, участвующих в экспрессии протеазной активности области 2A вируса FMDV. Были также охарактеризованы область 2A и аналогичные домены афтовирусов и кардиовирусов, принадлежащих семейству пикорнавирусов (Donnelly et al. 1997. J. Gen. Virol. 78:13-21).
Другие попытки применения методов протеолитического процессинга были описаны ранее для продуцирования рекомбинантного инсулина, например, в EP055945 (Genentech); и EP037723 (“Регенты Университета Калифорнии”). Однако применение этих методов для осуществления рекомбинантной экспрессии гораздо более крупных и более сложных функциональных белков, таких как иммуноглобулины, потребует резкого изменения технологии их получения. Примерами функциональных молекул антител могут служить гетеромультимеры, необходимые для сборки четырех или более цепей (например, две тяжелые цепи и две легкие цепи иммуноглобулина).
В связи с этим возникает необходимость получения альтернативных и/или улучшенных систем для генерирования рекомбинантных белков. Особенно острой проблемой является, в частности, разработка методов эффективной и/или “правильной” экспрессии полноразмерных иммуноглобулинов и их антигенсвязывающих фрагментов, которые имели бы преимущества по сравнению с используемыми современными технологиями. Настоящее изобретение направлено на решение этой проблемы путем получения одной векторной конструкции с применением ряда стратегий, таких как использование интеинов, hedgehog-сегментов аутопроцессинга, автокаталитических вирусных протеаз и их вариантов соответственно. Независимо от этого проблема, связанная с необходимостью разработки метода эффективной сборки мультимерных продуктов (например, иммуноглобулина), может быть решена путем регуляции стехиометрического соотношения субъединиц (например, тяжелой и легкой цепей или их фрагментов). В некоторых вариантах конструкции в одной ORF (sORF) кодируют пептидный компонент аутопроцессинга, который может быть использован для широкомасштабной экспрессии полипептида или для экспрессии биологически функционального полипептида, такого как фермент, иммуноглобулин, цитокин, хемокин, рецептор, гормон, компоненты двухгибридной системы или других представляющих интерес мультисубъединичных белков.
Краткое описание сущности изобретения
Настоящее изобретение относится к экспрессионным кластерам, векторам, рекомбинантным клеткам-хозяевам и к способам рекомбинантной экспрессии и процессинга, включая посттрансляционный процессинг рекомбинантных полипротеинов и белков-предшественников.
В одном из вариантов настоящее изобретение относится к экспрессирующему вектору для генерирования одного или нескольких рекомбинантных белковых продуктов, содержащему sORF-вставку, где указанная sORF-вставка включает в себя первую последовательность нуклеиновой кислоты, кодирующую первый полипептид; промежуточную последовательность нуклеиновой кислоты, кодирующую первый сайт расщепления белка; и вторую последовательность нуклеиновой кислоты, кодирующую второй полипептид, где указанная промежуточная последовательность нуклеиновой кислоты, кодирующая указанный первый сайт расщепления белка, соответствующим образом расположена между указанной первой последовательностью нуклеиновой кислоты и указанной второй последовательностью нуклеиновой кислоты и где указанный экспрессирующий вектор способен экспрессировать полипептид, кодируемый sORF и расщепляемый в указанном первом сайте расщепления белка. В одном из вариантов изобретения указанный первый сайт расщепления белка включает в себя сайт расщепления посредством аутопроцессинга. В одном своем варианте изобретения указанный сайт аутопроцессинга включает в себя сегмент интеина или модифицированный сегмент интеина, где указанный модифицированный (или немодифицированный) сегмент интеина обеспечивает расщепление полипептидов без полного лигирования экспрессированных первых полипептидов с экспрессированными вторыми полипептидами. В одном из вариантов изобретения указанный сайт аутопроцессинга включает в себя сегмент hedgehog или модифицированный сегмент hedgehog, где указанный модифицированный (или немодифицированный) сегмент hedgehog позволяет расщеплять экспрессированные первые полипептиды и экспрессированные вторые полипептиды. В одном из вариантов изобретения экспрессируется множество отдельных белков (например, первые полипептиды, вторые полипептиды, третьи полипептиды и т.п.). В одном из вариантов изобретения указанные первый полипептид и второй полипептид обладают способностью к мультимерной сборке. В одном из вариантов изобретения по меньшей мере один из указанных первых и вторых полипептидов обладает способностью к внеклеточной секреции. В одном из вариантов изобретения по меньшей мере один из указанных первых и вторых полипептидов происходит от млекопитающего. В одном из своих вариантов настоящее изобретение относится к векторам и способам генерирования антител, подвергнутых сборке.
В некоторых вариантах настоящее изобретение относится к конструкциям и к способам рекомбинантной экспрессии множества отдельных белков. В конкретных вариантах изобретения указанные белки обладают способностью к внеклеточной секреции. В конкретных вариантах изобретения указанные белки происходят от млекопитающего. В конкретных вариантах изобретения указанные белки обладают способностью к мультимерной сборке. В конкретных вариантах изобретения указанными белками являются иммуноглобулины.
В одном из вариантов изобретения рассматривается введение сайта узнавания протеазой отщепляемого сигнального пептида или последовательность аутопроцессинга полипептида (включая интеин, С-концевой hedgehog-домен аутопроцессинга, происходящий от дрозофилы, мыши, человека и животных других видов (Dassa et al, Trends in Genetics, Vol. 20 No. 11 Nov, 2004, 538-542; Ibrahim et al, Biochimica et Biophysics Acta 1760 (2006) 347-355). Авторы настоящего изобретения отмечают, что в некоторых случаях последовательность аутопроцессинга полипептида, если она упоминается в связи с протеолитическим процессингом, может называться протеолитическим сайтом. Обычно используются C-концевые домены аутопроцессинга генов warthog, groundhog и других hog-содержащих генов, происходящих от нематод, таких как Caenorhabditis elegans (Snell EA et al, Proc. R. Soc. B (2006) 273, 401-407; Aspock et al, Genome Research, 1999, 9:909-923); и домен аутопроцессинга Hoglet-C, происходящий от колониальных жгутиковых (Сhoanoflagellatа) (Aspock et al., Genome Research, 1999, 9:909-923). Бактериальные интеин-подобные (BIL) домены A-типа, такие как домены, происходящие от бактерий, таких как Clostridium thermocellum, и домены BIL B-типа, происходящие от бактерий, таких как Rhodobacter sphaeroides (Dassa et al., Journal of Biological Chemistry, Vol. 279, No. 31, July 30, 32001-32007) (дикого типа, в усеченной форме или в других модифицированных формах), и введенные в последовательность рекомбинантного белка-предшественника обеспечивают эффективную экспрессию и расщепление про-белка, в результате этого высвобождаются биологически активная часть или нужные белки, экспрессированные в полипротеине. Осуществление такого процесса позволяет избежать необходимости ко-экспрессии природных протеолитических ферментов, осуществляющих процессинг про-белков. Альтернативно, протеаза, которая является когнатной для конкретного сайта узнавания, может экстенсивно экспрессироваться вместе с последовательностью про-белка, при этом сайт узнавания протеазой находится между ними, в результате чего указанная протеаза высвобождается посредством протеолиза, а затем высвобождается отщепляемая часть белка-предшественника посредством последующего протеолитического расщепления, что приводит к высвобождению активной части белка-предшественника. В другом варианте изобретения пептидная последовательность аутопротеолитического процессинга 2А может быть встроена в белок-предшественник между частью зрелого (биоактивного) белка и отщепляющейся частью белка-предшественника, так чтобы после экспрессии сконструированного рекомбинантного белка осуществлялся его аутопроцессинг.
В другом своем варианте настоящее изобретение относится к способу эффективной экспрессии рекомбинантных молекул иммуноглобулина, включающему рекомбинантную экспрессию полипротеина, содержащего по меньшей мере одну область тяжелой цепи и по меньшей мере одну область легкой цепи, где указанные области отделены друг от друга одним или несколькими сайтами узнавания протеазой, сигнальными пептидами, последовательностями интеина, которые опосредуют расщепление, но не присоединение полипептидов; последовательностью hedgehog, другими интеин-подобными или hedgehog-подобными последовательностями аутопроцессинга или их вариантами или такими последовательностями, как пептид 2A, который разделяет фланкирующие пептиды в процессе трансляции. В другом варианте изобретения протеаза может экспрессироваться как часть полипротеина, отделенная от остального полипротеина сайтами узнавания протеазой, где каждый сайт узнавания протеазой является когнатным для ко-экспрессируемой протеазы. Затем действие протеолитической или сигнальной пептидазы приводит к высвобождению протеазы и других отдельных белков из первичного продукта трансляции. Вышеописанные способы разделения белковых субъединиц в полипротеине могут также применяться в комбинации друг с другом в целях достижения нужного протеолиза и получения продуктов экспрессии белка.
В варианте изобретения, относящемся к экспрессии иммуноглобулина, дупликация области, кодирующей легкую цепь, позволяет повышать эффективность сборки и/или экспрессии полноразмерной молекулы иммуноглобулина в том случае, если области, кодирующие легкую цепь, присутствуют в экспрессионном кластере и/или в экспрессирующем векторе вместе с областью, кодирующей тяжелую цепь, в отношении 1:1. В контексте настоящего изобретения белки тяжелой и легкой цепей могут представлять собой функциональные фрагменты природных тяжелых и легких цепей (функциональный фрагмент сохраняет способность связываться с цепью антитела-аналога, а также способность связываться с когнатным антигеном, как хорошо известно специалистам). Так, например, настоящее изобретение относится к конструкциям и к способам, в которых отношение области, кодирующей компонент легкой цепи, к области, кодирующей компонент тяжелой цепи, составляет 1:1 или более. Так, например, в одном из вариантов изобретения отношение L:H составляет 2:1 или более, а в других вариантах изобретения это отношение составляет 3:1, 3:2, 4:1 или более.
В предпочтительном аспекте изобретения последовательность, кодирующая легкую цепь иммуноглобулина или ее фрагмент, дуплицируется в последовательности, кодирующей полипротеин, а последовательности, кодирующие тяжелую и легкую цепи иммуноглобулина, присутствуют в молярном отношении “легкая цепь: тяжелая цепь”, составляющем примерно 2:1, и экспрессируются в отношении “легкая цепь: тяжелая цепь”, составляющем более чем 1:1. Последовательности легкой и тяжелой цепей в указанном полипротеине присоединены посредством сайтов расщепления протеазой, сигнальных (или лидерных) пептидов, интеинов или сайтов аутопроцессинга.
Протеазами (эндопротеазами), сигнальными пептидазами и аминокислотными последовательностями их сайтов узнавания, которые могут быть использованы для разделения компонентов биологически активного белка в продукте трансляции полипротеина, и их последовательностями узнавания протеазами являются, но не ограничиваются ими, сайты узнавания: фурином, RXR/K-R (SEQ ID NO:1); VP4 IPNV, S/TXA-S/AG (SEQ ID NO:2); протеазой вируса гравировки табака (TEV), EXXYXQ-G(SEQ ID NO:3); протеазой 3C риновируса, LEVLFQ-GP (SEQ ID NO:4); протеазой PC5/6; протеазой PACE, протеазой LPC/PC7; энтерокиназой, DDDDK-X (SEQ ID NO:5); фактором Xa, IE/DGR-X (SEQ ID NO:6); тромбином, LVPR-GS (SEQ ID NO:7); гененазой I, PGAAH-Y(SEQ ID NO:8); протеазой MMP; ядерным белком включения a(N1a) потивируса мозаики турнепса; NS2B/NS3 флавивирусов денге типа 4 (DEN4), протеазой NS3 вируса желтой лихорадки (YFV); ORF V вируса мозаики цветной капусты; и протеазой KEX2, MYKR-EAD (SEQ ID). Другим вариантом внутреннего сайта расщепления является CB2. Положение в последовательности узнавания протеазами, в которой происходит расщепление, показано дефисом.
В одном из вариантов изобретения используемыми сигнальными последовательностями являются последовательности дикого типа, мутированные последовательности или неспецифически мутированные последовательности, и эти последовательности были выбраны посредством скрининга с применением методов, известных специалистам.
В объем настоящего изобретения, описанный выше, также входят экспрессионный кластер, в котором конкретная последовательность, кодирующая полипротеин или белок-предшественник (про-белок, полипротеин), функционально присоединена к последовательностям регуляции транскрипции; экспрессирующие векторы; и рекомбинантные клетки-хозяева, содержащие указанный экспрессирующий вектор или экспрессионный кластер.
Настоящее изобретение относится к системе экспрессии полноразмерного иммуноглобулина или его фрагмента, основанной на экспрессии последовательностей, кодирующих тяжелую и легкую цепь под транскрипционным контролем одного промотора, где разделение тяжелой и легкой цепей опосредуется интеинами или модифицированными интеинами (которые обеспечивают расщепление, но не лигирование высвобождаемых молекул белков, либо такое лигированием белков может быть предотвращено антителами или другими фланкирующими последовательностями белков, которые могут быть соответствующим образом модифицированы), либо С-концевым hedgehog-доменом ацтопроцессинга, происходящим от дрозофилы, мыши, человека и животных других видов, либо C-концевыми доменами аутопроцессинга генов warthog, groundhog и других hog-содержащих генов, происходящих от нематод, таких как Caenorhabditis elegans, либо доменом аутопроцессинга Hoglet-C, происходящим от колониальных жгутиковых (Сhoanoflagellatа), либо бактериальными интеин-подобными (BIL) доменами A-типа, такими как домены, происходящие от бактерий, таких как Clostridium thermocellum, либо доменами BIL B-типа, происходящими от бактерий, таких как Rhodobacter sphaeroides. Интеинами, которые могут быть использованы в настоящем изобретении, являются, но не ограничиваются ими, интеины VMA Saccharomyces cerevisiae, Pyrococcus, Synechocystis и другие интеины, известные специалистам. Разделение тяжелой и легкой цепей может также опосредоваться сайтом аутопроцессинга, например, последовательностью 2А или 2А-подобной последовательностью.
В одном из своих аспектов настоящее изобретение относится к вектору для экспрессии рекомбинантного иммуноглобулина, который включает в себя промотор, функционально присоединенный к последовательности, кодирующей первую цепь молекулы иммуноглобулина или ее фрагмент; к последовательности, кодирующей сайт аутопроцессинга; и к последовательности, кодирующей вторую цепь молекулы иммуноглобулина или ее фрагмент, где указанная последовательность, кодирующая сайт аутопроцессинга, встроена между последовательностью, кодирующей первую цепь молекулы иммуноглобулина, и последовательностью, кодирующей вторую цепь молекулы иммуноглобулина. Первой или второй цепью молекулы иммуноглобулина может быть тяжелая цепь или легкая цепь, а последовательностью, кодирующей рекомбинантный иммуноглобулин, может быть полноразмерная кодирующая последовательность или ее фрагмент. Вторая область, соответствующая легкой цепи, отделена от смежной области сайтом узнавания протеазой, сигнальным пептидом или сайтом аутопроцессинга, таким как сайт 2А. При этом могут присутствовать две копии последовательности L-цепи и одна копия последовательности Н-цепи (или множество копий каждой цепи), при условии что каждый компонент цепи антитела имеет соответствующий сайт процессинга или последовательность, ассоциированную с этим сайтом, которые могли бы обеспечивать продуцирование “правильно” процессированных цепей антител.
Вектором может быть любой рекомбинантный вектор, способный экспрессировать полноразмерный полипептид, например молекулу иммуноглобулина или ее фрагмент, и таким вектором является, например, плазмидный вектор, в частности вектор, подходящий для экспрессии генов в клетках млекопитающих; бакуловирусный вектор для экспрессии в клетках насекомых; вектор на основе аденоассоциированного вируса (AAV); лентивирусный вектор; ретровирусный вектор; компетентный по репликации аденовирусный вектор; дефектный по репликации аденовирусный вектор; аттенюированный аденовирусный вектор; вектор на основе герпесвируса или невирусный (плазмидный) вектор и т.п.
Сайты аутопроцессинга включают в себя пептидную последовательность 2A, например последовательность 2А, происходящую от вируса ящура (FMDV). В другом предпочтительном аспекте изобретения такой вектор содержит последовательность, кодирующую дополнительный сайт протеолитического расщепления, расположенный между последовательностью, кодирующей первую цепь молекулы иммуноглобулина или ее фрагмент, и последовательностью, кодирующей вторую цепь молекулы иммуноглобулина или ее фрагмент (то есть последовательность, смежную с последовательностью сайта аутопроцессинга, такого как сайт 2А), а также последовательностью, смежной со второй последовательностью легкой цепи. В одном из репрезентативных подходов дополнительным сайтом протеолитического расщепления является сайт расщепления фурином с консенсусной последовательностью RXK/R-R (SEQ ID NO:1). Вектор для экспрессии рекомбинантного иммуноглобулина, содержащий пептид аутопроцессинга, может включать любой из ряда промоторов, где указанный промотор является конститутивным, регуляторным или индуцибельным, а также клеткоспецифическим, тканеспецифическим или видоспецифическим. Такой вектор может также включать последовательность, кодирующую сигнальную последовательность для одной или нескольких последовательностей, кодирующих цепи иммуноглобулина, белки-предшественники или т.п.
Настоящее изобретение также относится к клеткам-хозяевам или к стабильным клонам клеток-хозяев, инфицированных вектором, который содержит последовательность, кодирующую тяжелую и легкую цепи иммуноглобулина (то есть антитела); последовательность, кодирующую сайт аутопроцессинга; и может также содержать дополнительную последовательность, кодирующую дополнительный сайт протеолитического расщепления; и, необязательно, область, кодирующую протеазу, которая аналогичным образом отделена от остальной(ых) кодирующей(их) последовательности(ей) сайтом аутопроцессинга или сайтом узнавания протеазой. Использование таких клеток или клонов для генерирования полноразмерных рекомбинантных иммуноглобулинов или их фрагментов также входит в объем настоящего изобретения. Подходящими клетками-хозяевами являются, но не ограничиваются ими, культивированные клетки насекомых, таких как Spodoptera frugiperda, клетки микробов, включая бактерии; клетки дрожжей, таких как Saccharomyces cerevisiae или Pichia pastoris; клетки грибов, таких как Trichoderma reesei, Aspergillus, Aureobasidum и Penicillium species, а также клетки млекопитающих, такие как клетки яичника китайского хомячка (например, CHO-K1, ATCC CCL 61; CHO DG44, Chasin et al. 1986, Som. Cell. Molec. Genet. 12:555), клетки почек детеныша хомячка (BHK-21, BHK- 570, ATCC CRL 8544, ATCC CRL 10314), клетки COS, клетки мышиных эмбрионов (NIH-3T3, ATCC CRL 1658), клетки Verо (клетки почек африканской зеленой мартышки, депонированные в ATCC CRL 1587), клетки почек собак (например, MDCK, ATCC CCL 34), клетки крысиного гипофиза (GH1, ATCC CCL 34), клетки некоторых человеческих клеточных линий, включая клетки почек человеческого эмбриона (например, HEK293, ATCC CRL 1573), а также могут быть использованы различные системы трансгенных животных, включая, но не ограничиваясь ими, свиней, мышей, крыс, овец, коз и коров. Известно, что для экспрессии в яичном белке используются куры, а для экспрессии в молоке используются трансгенные овцы, козы и коровы, и т.п. В качестве клеток-хозяев могут быть также использованы клетки растений.
В своем родственном аспекте настоящее изобретение относится к рекомбинантной молекуле иммуноглобулина или ее фрагменту, продуцируемым такими клетками или клонами, где указанный иммуноглобулин включает в себя аминокислоты, кодируемые генами сайта аутопроцессинга, сигнального пептида и интеина, hog-содержащими генами С-концевого домена аутопроцессинга, генами бактериальных интеин-подобных доменов (BIL) или генами сайтов узнавания протеазой; а также к способам их продуцирования. В случае использования интеина предпочтительным является модифицированный интеин, при этом указанный интеин должен быть таким, чтобы две цепи антитела не подвергались совместному сплайсингу с образованием одной полипептидной цепи, либо концы полипептидов антител должны быть такими, чтобы они не могли подвергаться совместному сплайсингу, опосредуемому этим интеином. Интеин вводят между N-экстеином и С-экстеином с сохранением рамки считывания, например, между тяжелой цепью и легкой цепью иммуноглобулина, при условии что этот интеин и/или ближайшая к области стыка аминокислотная последовательность первичного продукта трансляции полипротеина будут обеспечивать расщепление с высвобождением экстеинов, но при этом не будет происходить лигирования этих экстеиновых белков.
Настоящее изобретение также относится к стратегии посттрансляционного процессинга белка с использованием hedgehog- домена процессинга белка, расположенного между первой частью экспрессированного белка и второй частью белка. Hedgehog-домен процессинга белка (Hh-C) может быть, но необязательно, усечен в целях делеции холестерин-переносящей части для того, чтобы происходило только расщепление белка. В случае если не происходит полного вырезания Hh-C, то включение домена сигнального пептида у N-конца второй части белка может приводить к протеолитическому отщеплению зрелого второго белка от Hh-C/части первого белка. Кроме того, в объем данного аспекта изобретения входят неприродные рекомбинантные молекулы ДНК, содержащие последовательность, кодирующую полипротеин, который включает в себя hedgehog-домен процессинга белка, расположенный между последовательностью, кодирующей первую часть экспрессированного белка, и последовательностью, кодирующей вторую часть экспрессированного белка, так что трансляция полипротеина происходит из одного транскрипта.
В другом своем аспекте настоящее изобретение относится к модифицированному фурину, отличающемуся тем, что он имеет дополнительную пептидную область, которая нацеливает новосинтезированный белок фурин в просвет эндоплазматического ретикулума. Настоящее изобретение также включает в себя описанную здесь стратегию получения интеина или модифицированного интеина.
В другом своем аспекте настоящее изобретение относится к применению стратегии расщепления полипротеина/аутопроцессинга, процессинга интеина, отщепления сигнального пептида или протеолитического расщепления в технологии получения двухгибридной и трехгибридной системы (и их вариантов). Первый и второй белки или первый, второй и третий белки экспрессируются в виде полипротеина из одного транскрипта в подходящей клетке-хозяине, а последовательности, кодирующие эти белки, разделены сайтом аутопроцессинга (например, 2А), интеином, сигнальным пептидом или сайтами узнавания протеазой. Такая стратегия позволяет избежать необходимости ко-трансфекции более чем одним вектором или осуществления экспрессии каждого белка из отдельного транскрипта, как это обычно происходит в стандартных методах, а поэтому стратегия, применяемая в настоящем изобретении, является более экономичной и, кроме того, позволяет повысить эффективность и уровень экспрессии, а близкое расположение потенциальных пар связывания по отношению друг к другу будет, очевидно, способствовать повышению вероятности присоединения друг к другу партнеров по связыванию. В конкретном варианте изобретения указанный полипротеин включает в себя белок-“приманку”, сайт аутопроцессинга, интеин, сигнальный пептид или последовательность узнавания протеазой и встроенные последовательности кДНК, представляющие собой один или несколько потенциальных белков-“жертв”, которые взаимодействуют с представляющим интерес белком-“приманкой”. Такая стратегия клонирования и экспрессии схематически проиллюстрирована на фиг. 8 и 9.
В одном из своих вариантов настоящее изобретение относится к конструкциям ДНК для экспрессии множества генных продуктов в клетке, включащим один промотор у 5'-конца данной конструкции; интеин-содержащий элемент, включающий в себя две или более последовательности экстеинов, кодирующих отдельные белки, и одну или несколько последовательностей интеина, присоединенные к карбокси-концевой кодирующей части каждой экстеиновой последовательности, за исключением последней экспрессируемой экстеиновой последовательности; и 3'-концевую последовательность, включающую сигнал полиаденилирования, расположенный за последней последовательностью, кодирующей белок экстеин, где указанный интеин-содержащий элемент экспрессируется в виде белка-предшественника, содержащего по меньшей мере один интеин, фланкированный кодируемыми экстеином белками, и по меньшей мере один из указанных интеинов может катализировать вырезание экстеинов; и где в указанном интеин-содержащем элементе предпочтительно по меньшей мере один аминокислотный остаток был заменен или добавлен так, чтобы вырезанные экстеины не подвергались лигированию посредством интеина. В конкретном варианте изобретения указанные конструкции имеют такую конфигурацию, при которой по меньшей мере две последовательности экстеина после их экспрессии в виде белков обладают способностью подвергаться мультимерной сборке. В одном из вариантов изобретения по меньшей мере две последовательности экстеина могут кодировать молекулу иммуноглобулина или другую молекулу, распознающую антиген. В одном из вариантов изобретения по меньшей мере одна последовательность экстеина после ее экспрессии в виде белка обладает способностью к внеклеточной секреции. В одном из вариантов изобретения по меньшей мере одна последовательность экстеина представляет собой ген млекопитающего.
В некоторых вариантах настоящее изобретение относится к конструкциям и способам экспрессии иммуноглобулина с использованием модифицированного или немодифицированного интеина, где экспрессированные сегменты иммуноглобулина не подвергаются повторному лигированию/присоединению, что позволяет продуцировать подвергнутое сборке антитело из множества субъединиц. В конкретном варианте изобретения модифицированный интеин имеет модификацию аминокислотного остатка, локализованного в первом положении С-экстеина. В конкретном варианте изобретения такая модификация присутствует в положении предпоследней аминокислоты интеинового сегмента.
В некоторых вариантах настоящее изобретение относится к конструкциям и к способам экспрессии любого гена или комбинации генов. В конкретном варианте изобретения С-экстеин является модифицированным. В другом конкретном варианте изобретения указанный С-экстеин модифицирован с использованием сигнальной последовательности. В другом конкретном варианте изобретения С-концевой экстеиновый компонент отсутствует.
В некоторых вариантах настоящее изобретение относится к конструкциям и к способам экспрессии генов антител с использованием модифицированного сигнального пептида для второй цепи иммуноглобулина (тяжелой или легкой цепи) и третьей цепи, если она присутствует, где указанные гены находятся за интеином или hedgehog-доменом аутопроцессинга. В одном из вариантов изобретения сегменты расположены в следующем порядке: первая цепь - первый интеин или hedgehog - первый модифицированный сигнальный пептид - вторая цепь - второй модифицированный сигнальный пептид - третья цепь (в случае, если этот вариант состоит из двух цепей, то, например, третья цепь или сегмент “второй модифицированный сигнальный пептид - третья цепь” отсутствуют). В другом варианте изобретения второй интеин или сегмент hedgehog расположен за второй цепью. В конкретном варианте изобретения использование такого модифицированного сигнального пептида позволяет повышать уровень секреции антитела. В одном из вариантов изобретения используемый сигнальный пептид модифицируют для снижения гидрофобности. В одном из вариантов изобретения такой сигнальный пептид является немодифицированным.
В некоторых вариантах настоящее изобретение относится к sORF-векторам для временной экспрессии. В другом варианте изобретения настоящее изобретение относится к sORF-векторам для экспрессии в стабильных экспрессионных системах. В одном из вариантов изобретения стабильные клетки-хозяева получают известными методами, например, путем трансфекции, и другими методами.
Хотя в настоящей заявке конкретно описано множество репрезентативных конструкций для экспрессии антитела, специфичного к фактору некроза опухоли α (альфа), однако следует отметить, что с применением аналогичных стратегий с заменой последовательностей, кодирующих другие белки, могут быть легко получены и другие конструкции. Конкретными примерами могут служить другие молекулы иммуноглобулинов и биотерапевтические молекулы. Другими конкретными примерами являются антитела, специфичные к селектину E/L, интерлейкину-12, интерлейкину-18 или рецептору эритропоэтина или любое другое антитело с нужной специфичностью, аминокислотная последовательность и/или кодирующая последовательность которого известны специалистам.
В одном из своих вариантов настоящее изобретение относится к экспрессирующему вектору, используемому для продуцирования одного или нескольких рекомбинантнных белковых продуктов и содержащему sORF-вставку, где указанная sORF-вставка включает в себя первую последовательность нуклеиновой кислоты, кодирующую первый полипептид; первую промежуточную последовательность нуклеиновой кислоты, кодирующую сайт расщепления первого белка, и вторую последовательность нуклеиновой кислоты, кодирующую второй полипептид; где указанная промежуточная последовательность нуклеиновой кислоты, кодирующая указанный сайт расщепления первого белка, соответствующим образом локализована между указанной первой последовательностью нуклеиновой кислоты и указанной второй последовательностью нуклеиновой кислоты и где указанный экспрессирующий вектор способен экспрессировать sORF-кодируемый полипептид, расщепляемый в указанном первом сайте расщепления белка. В одном из вариантов изобретения указанный первый сайт расщепления белка содержит сайт аутопроцессинга.
В одном из вариантов изобретения указанный сайт аутопроцессинга содержит интеиновый сегмент или модифицированный интеиновый сегмент, где указанный модифицированный интеиновый сегмент обеспечивает расщепление, но не лигирование указанного первого полипептида с указанным вторым полипептидом. В одном из вариантов изобретения указанный сайт аутопроцессинга содержит сегмент hedgehog или модифицированный сегмент hedgehog, где указанный модифицированный сегмент hedgehog обеспечивает отщепление указанного первого полипептида от указанного второго полипептида. В одном из вариантов изобретения указанные первый и второй полипептиды обладают способностью к мультимерной сборке. В одном из вариантов изобретения по меньшей мере один из указанных первых и вторых полипептидов обладает способностью к внеклеточной секреции. В одном из вариантов изобретения по меньшей мере один из указанных первых и вторых полипептидов происходит от млекопитающего.
В одном из вариантов изобретения по меньшей мере один из указанных первых и вторых полипептидов содержит тяжелую цепь иммуноглобулина или ее функциональный фрагмент. В одном из вариантов изобретения по меньшей мере один из указанных первых и вторых полипептидов содержит легкую цепь иммуноглобулина или ее функциональный фрагмент. В одном из вариантов изобретения указанный первый полипептид содержит тяжелую цепь иммуноглобулина или ее функциональный фрагмент, а указанный второй полипептид содержит легкую цепь иммуноглобулина или ее функциональный фрагмент, где указанные первый и второй полипептиды расположены в любом порядке. В одном из вариантов изобретения указанные первый и второй полипептиды, взятые вместе, обладают способностью к мультмерной сборке с образованием функционального антитела или другой молекулы, распознающей антиген.
В одном из вариантов изобретения указанный первый полипептид расположен выше указанного второго полипептида. В одном из вариантов изобретения указанный второй полипептид расположен выше указанного первого полипептида.
В одном из вариантов изобретения экспрессирующий вектор также содержит третью последовательность нуклеиновой кислоты, кодирующую третий полипептид, где указанная третья последовательность нуклеиновой кислоты соответствующим образом раположена за указанной второй последовательностью нуклеиновой кислоты и где, независимо от этого, указанная третья последовательность может быть такой же, как указанная первая или вторая последовательность нуклеиновой кислоты, либо она может отличаться от этих последовательностей. В одном из вариантов изобретения по меньшей мере два из указанных первого, второго и третьего полипептидов, взятые вместе, обладают способностью к сборке мультимерного белка.
В одном из вариантов изобретения указанный экспрессирующий вектор также содержит вторую промежуточную последовательность нуклеиновой кислоты, кодирующую второй сайт расщепления белка, где указанная вторая промежуточная последовательность нуклеиновой кислоты соответствующим образом расположена за указанной первой и второй последовательностью нуклеиновой кислоты и где указанная вторая промежуточная последовательность может быть такой же, как указанная первая промежуточная последовательность нуклеиновой кислоты, либо она может отличаться от такой последовательности. В одном из вариантов изобретения экспрессирующий вектор также содержит третью последовательность нуклеиновой кислоты, кодирующую третий полипептид, и вторую промежуточную последовательность нуклеиновой кислоты, кодирующую второй сайт расщепления белка; где указанная вторая промежуточная последовательность нуклеиновой кислоты и третья последовательность нуклеиновой кислоты, расположенные в таком порядке, соответствующим образом локализованы за указанной второй последовательностью нуклеиновой кислоты. В одном из вариантов изобретения указанная третья последовательность нуклеиновой кислоты кодирует тяжелую цепь иммуноглобулина, легкую цепь иммуноглобулина или их функциональные фрагменты соответственно. В одном из вариантов изобретения указанная третья последовательность нуклеиновой кислоты кодирует легкую цепь иммуноглобулина или ее функциональный фрагмент. В одном из вариантов изобретения указанная третья последовательность нуклеиновой кислоты кодирует тяжелую цепь иммуноглобулина или ее функциональный фрагмент.
В одном варианте экспрессирующего вектора указанная первая промежуточная последовательность нуклеиновой кислоты, кодирующая первый сайт расщепления белка, содержит нуклеиновую кислоту сигнального пептида, кодирующую сайт отщепления сигнального пептида или модифицированную последовательность сайта отщепления сигнального пептида. В одном из вариантов изобретения указанный экспрессирующий вектор также содержит последовательность нуклеиновой кислоты сигнального пептида, кодирующую сайт отщепления сигнального пептида и соответствующим образом расположенную перед указанной первой последовательностью нуклеиновой кислоты или указанной второй последовательностью нуклеиновой кислоты.
В одном из вариантов изобретения экспрессирующий вектор также содержит две последовательности нуклеиновой кислоты сигнального пептида, каждая из которых независимо кодирует сайт отщепления сигнального пептида, где одна последовательность нуклеиновой кислоты сигнального пептида соответствующим образом расположена перед указанной первой нуклеиновой кислотой, кодирующей первый полипептид, а другая последовательность нуклеиновой кислоты сигнального пептида соответствующим образом расположена перед указанной второй нуклеиновой кислотой, кодирующей указанный второй полипептид. В некоторых вариантах изобретения указанные две последовательности сигнального пептида являются одинаковыми или различными.
В одном из вариантов изобретения последовательность нуклеиновой кислоты сигнального пептида кодирует сайт отщепления сигнального пептида легкой цепи иммуноглобулина или модифицированный сайт отщепления сигнального пептида легкой цепи иммуноглобулина. В одном из вариантов изобретения последовательность нуклеиновой кислоты сигнального пептида кодирует модифицированный или немодифицированный сайт отщепления сигнального пептида легкой цепи иммуноглобулина, где указанный модифицированный сайт обладает способностью расщеплять и повышать секрецию по меньшей мере одного из указанных первого полипептида, второго полипептида и подвергнутых сборке молекул указанных первого и второго полипептидов и где уровень секреции в присутствии указанного сайта сигнального пептида примерно в 10-100 раз превышает уровень секреции в отсутствие указанного сайта сигнального пептида.
В одном из вариантов изобретения промежуточная последовательность нуклеиновой кислоты, кодирующая первый сайт расщепления белка, содержит последовательность интеина или модифицированную последовательность интеина, выбранные из группы, состоящей из: последовательности Pho Pol I Pyrococcus horikoshii, последовательности VMA Saccharomyces cerevisiae, Synechocystis spp, последовательности DnaE штамма РСС6803, последовательности GyrA Mycobacterium xenopi, последовательности ДНК-полимеразы GB-D Pyrococcus spp., последовательности бактериального интеин-подобного домена (BIL) А-типа и последовательности BIL B-типа.
В одном из вариантов изобретения промежуточная последовательность нуклеиновой кислоты, кодирующая первый сайт расщепления белка, содержит С-концевой домен аутопроцессинга, являющийся членом семейства hedgehog, где указанный член семейства hedgehog происходит от дрозофилы, мыши, человека или других насекомых или животных. В одном из вариантов изобретения промежуточная последовательность нуклеиновой кислоты, кодирующая первый сайт расщепления белка, содержит С-концевой домен аутопроцессинга, происходящий от гена warthog, groundhog или другого hog-содержащего гена нематоды, или домен Hoglet, происходящий от колониальных жгутиковых.
В одном из вариантов изобретения указанные первый и указанный второй полипептиды содержат функциональное антитело или другую молекулу, распознающую антиген, где указанное, антитело обладает специфичностью связывания с антигеном, выбранным из группы, состоящей из: фактора некроза опухоли - α, рецептора эритропоэтина, RSV, EL/селектина, интерлейкина-1, интерлейкина-12, интерлейкина-13, интерлейкина-18, интерлейкина-23, CXCL-13, GLP-1R и амилоида-бета. В одном из вариантов изобретения указанный первый и второй полипептиды содержат пару иммуноглобулиновых цепей антитела D2E7, ABT-007, ABT-325, EL246 или ABT-874. В одном из вариантов изобретения каждый из указанных первого и второго полипептидов независимо выбран из сегментов тяжелой цепи или легкой цепи иммуноглобулина, происходящих от аналогичного сегмента антител D2E7, ABT-007, ABT-325, EL246, ABT-874 или других антител.
В одном из вариантов изобретения вектор дополнительно содержит промоторный регуляторный элемент для указанной sORF-вставки. В одном из вариантов изобретения указанный промоторный регуляторный элемент является индуцибельным или конститутивным. В одном из вариантов изобретения указанный промоторный регуляторный элемент является тканеспецифическим. В одном из вариантов изобретения указанный промотор содержит основной поздний промотор аденовируса.
В одном из вариантов изобретения вектор также содержит нуклеиновую кислоту, кодирующую протеазу, способную отщеплять указанный первый сайт расщепления белка. В одном из вариантов изобретения указанная нуклеиновая кислота, кодирующая протеазу, соответствующим образом расположена в указанной sORF-вставке, где указанный экспрессирующий вектор также включает в себя дополнительную нуклеиновую кислоту, кодирующую второй сайт расщепления, расположенный между указанной нуклеиновой кислотой, кодирующей протеазу, и по меньшей мере одной из указанных первой нуклеиновой кислоты и второй нуклеиновой кислоты.
В одном из своих вариантов настоящее изобретение относится к клетке-хозяину, содержащей описанный здесь вектор. В одном из вариантов изобретения такой клеткой-хозяином является прокариотическая клетка. В одном из вариантов изобретения указанной клеткой-хозяином является Escherichia coli. В одном из вариантов изобретения указанной клеткой-хозяином является эукариотическая клетка. В одном из вариантов изобретения указанная эукариотическая клетка выбрана из группы, состоящей из клеток одноклеточных организмов, клеток животных, клеток растений и клеток грибов. В одном из вариантов изобретения указанной эукариотической клеткой является клетка животного, выбранная из группы, состоящей из клеток млекопитающего, клеток птиц и клеток насекомых. В предпочтительном варианте изобретения указанной клеткой-хозяином является клетка CHO или дефицитная по дигидрофолат-редуктазе клетка CHO. В одном из вариантов изобретения указанной клеткой-хозяином является клетка COS. В одном из вариантов изобретения указанной клеткой-хозяином является дрожжевая клетка. В одном из вариантов изобретения указанной клеткой-хозяином является Saccharomyces cerevisiae. В одном из вариантов изобретения указанной клеткой-хозяином является клетка Sf9 насекомого Spodoptera frugiperda. В одном из вариантов изобретения указанной клеткой-хозяином является клетка почки человеческого эмбриона.
В одном из своих вариантов настоящее изобретение относится к способу продуцирования рекомбинантного полипротеина или множества белков, включающему культивирование клетки-хозяина в культуральной среде в условиях, благоприятных для осуществления экспрессии белка в векторе. В одном из вариантов изобретения указанный способ дополнительно включает в себя выделение и/или очистку указанного векторного белка. В одном из вариантов изобретения указанное множество белков может подвергаться мультимерной сборке. В одном из вариантов изобретения рекомбинантный полипротеин или указанное множество белков являются биологически функциональными и/или обладают терапевтическими свойствами.
В одном из своих вариантов настоящее изобретение относится к способу продуцирования белка иммуноглобулина или его функционального фрагмента, подвергнутого сборке антитела или другой молекулы распознавания антигена, где указанный способ включает в себя культивирование клетки-хозяина по п.26 формулы изобретения в культуральной среде в условиях, благоприятных для продуцирования белка иммуноглобулина или его функционального фрагмента, подвергнутого сборке антитела или другой молекулы распознавания антигена.
В одном из своих вариантов настоящее изобретение относится к белку или полипротеину, продуцированному способом согласно изобретению. В одном из своих вариантов настоящее изобретение относится к подвергнутому сборке иммуноглобулину; подвергнутой сборке другой молекуле распознавания антигена или к отдельной цепи иммуноглобулина или его функционального фрагмента, продуцированным способами согласно изобретению. В одном из вариантов изобретения указанный иммуноглобулин, другая молекула распознавания антигена или отдельная цепь иммуноглобулина или его функциональный фрагмент обладают способностью повышать уровень связывания или стимулировать антигенспецифическое связывание с фактором некроза опухоли - α, рецептором эритропоэтина, интерлейкином-18, EL/селектином или интерлейкином-12. В одном из вариантов изобретения указанным иммуноглобулином является D2E7, а его функциональным фрагментом является фрагмент D2E7.
В одном из своих вариантов настоящее изобретение относится к фармацевтической композиции или к лекарственному средству, содержащим белок и фармацевтически приемлемый носитель. Наполнители и носители для фармацевтических препаратов могут быть выбраны по усмотрению специалиста в данной области.
В одном из своих вариантов настоящее изобретение относится к экспрессирующему вектору, в котором первый сайт расщепления белка включает в себя сайт расщепления клеточной протеазой или сайт расщепления вирусной протеазой. В одном из вариантов изобретения указанный первый сайт расщепления белка содержит сайт, узнаваемый фурином; VP4 IPNV; протеазой вируса гравировки табака (TEV); протеазой 3С риновируса; протеазой РС5/6; протеазой РАСЕ, протеазой LPC/PC7; энтерокиназой; протеазой фактора Ха; тромбином; гененазой I; протеазой ММР; ядерным белком включения a(N1a) потивируса мозаики турнепса; NS2B/NS3 флавивирусов денге типа 4 (DEN4); протеазой NS3 вируса желтой лихорадки (YFV); ORF V вируса мозаики цветной капусты; протеазой КЕХ2; СВ2; или сайт 2А. В одном из вариантов изобретения указанный первый сайт расщепления белка представляет собой сайт отщепления внутреннего вирусного сигнального пептида. В одном из вариантов изобретения указанный сайт отщепления внутреннего вирусного сигнального пептида включает в себя сайт, происходящий от вируса гриппа С, вируса гепатита С, хантавируса, флавивируса или вируса коревой краснухи.
В одном из своих вариантов настоящее изобретение относится к способу экспрессии белков двухгибридной системы, где указанная двухгибридная система содержит белок-"приманку" и кандидат на белок-"жертву", где указанный способ включает в себя стадии: получения клетки-хозяина, в которую был введен экспрессирующий вектор, кодирующий полипротеин, содержащий часть белка-"приманки", и часть кандидата на белок-"жертву", где указанные части разделены последовательностью аутопроцессинга, последовательностью сигнального пептида или сайтом расщепления протеазой; и культивирования данной клетки-хозяина в условиях, благоприятствующих экспрессии и аутопроцессингу полипротеина или расщеплению указанного полипротеина протеазой. В одном из вариантов изобретения указанный полипротеин также включает в себя отщепляемый компонент трехгибридной системы.
В одном из вариантов изобретения экспрессирующий вектор не содержит последовательности 2А. В одном из своих вариантов настоящее изобретение относится к экспрессирующему вектору, в котором указанный первый сайт расщепления белка содержит последовательность 2А FMDV и 2А-подобный домен, происходящий от других пикорнавирусов, вируса насекомых, ротавируса типа С, трипоносомы или Thermakoga maritime.
В одном из своих вариантов настоящее изобретение относится к экспрессирующему вектору, экспрессирующему рекомбинантный белок и содержащему последовательность, кодирующую полипротеин, где указанный полипротеин содержит по меньшей мере первый и второй сегмент белка, где указанные сегменты белка разделены сайтом расщепления белка, расположенным между ними; а указанный сайт расщепления белка содержит последовательность пептида аутопроцессинга, последовательность отщепления сигнального пептида или последовательность расщепления протеазой; и где указанная кодирующая последовательность экспрессируется в клетке-хозяине и расщепляется в этой клетке-хозяине.
В одном из своих вариантов настоящее изобретение относится к экспрессирующему вектору, в котором встроенная последовательность нуклеиновой кислоты дополнительно кодирует метку.
Другие аспекты, признаки и преимущества настоящего изобретения будут очевидны из нижеследующего описания изобретения, которое вместе с прилагаемым к нему графическим материалом приводится в целях раскрытия сущности настоящего изобретения.
В общих чертах используемые здесь термины и выражения имеют свои общепринятые значения, которые встречаются в научных монографиях, реферативных журналах и в других научных материалах, доступных специалистам. Для лучшего понимания конкретного употребления этих терминов в контексте описания настоящего изобретения ниже приводятся определения этих терминов.
Не ограничиваясь какой-либо конкретной теорией, можно лишь отметить, что в настоящем описании могут обсуждаться мнения или позиции авторов относительно основных принципов или механизмов, лежащих в основе настоящего изобретения. Следует также отметить, что независимо от окончательной точности какого-либо объяснения или гипотезы данный вариант осуществления изобретения может, тем не менее, иметь юридическую силу и практическое применение.
Краткое описание графического материала
На фигуре 1 проиллюстрирована предпочтительная конструкция экспрессирующего вектора для стабильной экспрессии sORF.
На фигуре 2 проиллюстрирована предпочтительная конструкция экспрессирующего вектора для стабильной экспрессии sORF, также содержащая дополнительную (вторую) промежуточную последовательность нуклеиновой кислоты, кодирующую второй сайт расщепления белка (который может представлять собой сайт аутопроцессинга) и третью последовательность нуклеиновой кислоты, кодирующую третий полипептид. Указанный вектор способен экспрессировать более чем два полипептида.
На фигуре 3 проиллюстрирована предпочтительная конструкция экспрессирующего вектора для временной экспрессии sORF (например, рТТ3-HC-Ssp-GA-int-LC-0aa).
На фигуре 4 проиллюстрирован экспрессирующий вектор, имеющий сегмент для экспрессии двухгибридной системы. Указанный векторный экспрессионный кластер имеет структуру, позволяющую сначала транслировать белок-"приманку" в виде гибридного белка "GAL4::bait::2А", который после трансляции пептида 2А подвергается аутопроцессингу. Второй открытой рамкой считывания (ORF) является гибридный белок "NFkappaB::библиотека".
На фигуре 5 представлена в линейном виде экспрессионная область плазмиды, показанная на фигуре 4 (2-компонентная гибридная система с пептидом расщепления 2А).
На фигуре 6 проиллюстрированы интеин-содержащие sORF-векторы, предназначенные для экспрессии иммуноглобулина.
На фигуре 7 проиллюстрировано несколько sORF-конструкций с выбранными точковыми мутациями, предназначенные для экспрессии подвергаемых сборке мультимерных молекул, таких как антитела.
На фигуре 8 проиллюстрированы sORF-конструкции с модифицированными сигнальными пептидами, например с модифицированными сигнальными пептидами легкой цепи иммуноглобулина.
На фигуре 9 проиллюстрированы sORF-конструкции, полученные с использованием hedgehog-доменов аутопроцессинга.
Подробное описание изобретения
Для лучшего понимания настоящего изобретения ниже приводится его подробное описание и неограничивающие примеры его осуществления.
Настоящее изобретение относится к системам, например, к конструкциям и способам, предназначенным для экспрессии структурных и биологически активных белков, таких как фермент, гормон (например, инсулин), цитокин, хемокин, рецептор, антитело или другая молекула. Предпочтительным белком является иммуномодуляторный белок, такой как интерлейкин, полноразмерный иммуноглобулин или его фрагмент, другая известная антиген-узнающая молекула или другая биотерапевтическая молекула. Общее описание таких систем приводится в конкретном обсуждении молекулы иммуноглобулина, рекомбинантное продуцирование которой осуществляют посредством экспрессии последовательностей, кодирующих тяжелую и легкую цепи, под транскрипционным контролем одного промотора, где превращение одного продукта трансляции (полипротеина) в отдельные тяжелые и легкие цепи опосредуется интеинами, hog-содержащими доменами аутопроцессинга, последовательностью 2A или 2A-подобной последовательностью, которые разделяют фланкирующие пептиды в процессе трансляции рибосомами или является результатом протеолитического процессинга в одном или нескольких сайтах узнавания протеазой, локализованных между двумя цепями зрелого биологически активного белка.
Межгенный сайт (родственный интеину сегмент, или hog-домен, 2A или 2A-подобный домен, или сайт узнавания протеазой; или их варианты) может называться сайтом расщепления. В случае экспрессии множества из трех или более белковых сегментов такой сайт расщепления может быть локализован между любыми по меньшей мере двумя сегментами из этого множества сегментов, либо этот сайт расщепления может быть расположен позади каждого сегмента, но позади последнего сегмента этот сайт необязательно, но предпочтительно отсутствует. Если используется множество сайтов расщепления, то эти сайты могут быть, независимо, одинаковыми или различными.
В одном из своих аспектов настоящее изобретение относится к к вектору для экспрессии рекомбинантного иммуноглобулина, включающему промотор, функционально присоединенный к последовательности, кодирующей первую цепь молекулы иммуноглобулина или ее фрагмент; последовательность, кодирующую сайт аутопроцессинга или другой сайт протеолитического расщепления; и последовательность, кодирующую вторую цепь молекулы иммуноглобулина или его фрагмент, где последовательность, кодирующая сайт аутопроцессинга или другой сайт протеолитического расщепления, встроена между последовательностью, кодирующей первую цепь молекулы иммуноглобулина, и последовательностью, кодирующую вторую цепь молекулы иммуноглобулина; и третью область, кодирующую легкую цепь иммуноглобулина, также отделенную от основной части полипротеина сайтом аутопроцессинга или другим сайтом протеолитического расщепления.
В одном из вариантов изобретения первой или второй цепью молекулы иммуноглобулинового полипротеина могут быть тяжелая цепь или легкая цепь. Последовательность, кодирующая рекомбинантный сегмент иммуноглобулина, может представлять собой полноразмерную кодирующую последовательность или ее фрагмент. В конкретном варианте изобретения вторая последовательность, кодирующая легкую цепь, должна быть частью последовательности, кодирующей процессируемый полипротеин согласно изобретению, то есть в целом, присутствуют три сегмента, содержащие две легких цепи и одну тяжелую цепь, расположенные в любом порядке. В конкретных вариантах изобретения конструкции имели конфигурацию, где указанные компоненты были расположены в следующем порядке: a) IgH-IgL; b) IgL-IgH; c) IgH-IgL-IgL; d) IgL-IgH-IgL; e) IgL-IgL-IgH; f) IgH-IgH-IgL; g) IgH-IgL-IgH; и/или h) IgL-IgH-IgH. В одном из вариантов изобретения “дефис” может означать положение, в котором находится последовательность сайта расщепления.
Альтернативно, последовательности, кодирующие тяжелую и легкую цепи иммуноглобулина, были лигированы в той же рамке считывания с расположенной между ними последовательностью, кодирующей интеин; при этом указанный интеин был модифицирован так, что он не обладает активностью сплайсинга, или концы тяжелой и легкой цепей были сконструированы так, чтобы сплайсинг предпочтительно не осуществлялся, или так, чтобы сплайсинг осуществлялся с низкой эффективностью с преобладанием несплайсированных молекул антител. Кроме того, модифицированный интеин может быть также дополнительно модифицирован так, чтобы в нем отсуствовала эндонуклеазная область (если эта эндонуклеазная область присутствовала в нем ранее), при условии что при этом будет сохраняться сайт-специфическая активность протеолитического расщепления, которая будет приводить к освобождению полипептидов легкой и тяжелой цепей антитела от промежуточной интеиновой части первичного продукта трансляции. Полипептидом легкой или тяжелой цепи антитела может быть N-экстеин или С-экстеин.
Указанным вектором может быть рекомбинантный вектор, способный экспрессировать полноразмерный полипротеин, например вектор на основе аденоассоциированного вируса (AAV); лентивирусный вектор; ретровирусный вектор; компетентный по репликации аденовирусный вектор; дефектный по репликации аденовирусный вектор и аттенюированный аденовирусный вектор; вектор на основе герпесвируса или невирусный (плазмидный) вектор или любой другой вектор, известный специалистам, причем выбор вектора, подходящего для экспрессии, зависит от клетки-хозяина, в которой экспрессируется иммуноглобулин или другой(ие) белок(белки). Векторами, подходящими для экспрессии генов в клетках насекомых, являются бакуловирусные векторы. Специалистам известно множество векторов, и многие из них являются коммерчески доступными или могут быть легко получены самим специалистом.
Сайты расщепления
Предпочтительными ресстрикционными сайтами аутопроцессинга являются последовательность интеина; модифицированный интеин; последовательность hedgehog; другая последовательность семейства "hog"; последовательность 2А, например последовательность 2А, происходящая от вируса ящура (FMDV); и их варианты.
Протеазами, последовательности узнавания которых могут быть заменены последовательностью 2А, являются, но не ограничиваются ими, фурин; модифицированный фурин, нацеленный не на транс-сеть Гольджи, а на эндоплазматический ретикулум; VP4 IPNV; протеаза TEV; протеаза TEV, дефицитная по сигналу ядерной локализации (TEV Nls); протеаза 3C риновируса; протеаза PC5/6; протеаза PACE, протеаза LPC/PC7; энтерокиназа; протеаза Xa; тромбин; гененаза I; и протеаза MMP, обсуждаемые выше. Другими эндопротеазами, используемыми для осуществления настоящего изобретения, являются, но не ограничиваются ими, такие протеазы, как ядерный белок включения a(N1a) потивируса мозаики турнепса; (Kim et al. 1996. Virology 221:245-249); протеазы флавивирусов NS2B/NS3 типа вируса денге 4 (DEN4) (Falgout et al. 1993. J. Virol. 67:2034-2042; Lai et al. 1994. Arch. Virol. Suppl. 9:359-368); протеаза NS3 вируса желтой лихорадки (YFV) (Chambers et al. 1991. J. Virol. 65:6042-6050); ORF V вируса мозаики цветной капусты (Torruella et al. 1989. EMBO Journal 8:2819-2825); интеины, примером которых является интеин Psp-GBD Pol (Xu, M.Q. 1996. EMBO 15: 5146-5153); внутренний отщепляемый сигнальный пептид, примером которого является внутренний отщепляемый сигнальный пептид вируса гриппа С (Pekosz A. 1992. Proc. Natl. Acad. Sci. USA 95: 3233-13238); и протеаза KEX2, MYKR-EAD (SEQ ID NO:9); KEX2 и модифицированная KEX2, которая нацелена на ЭР (см., Chaudhuri et al. 1992. Eur. J. Biochem. 210:811-822). Модифицированная KEX2, которая направлена только на ЭР, имеет кодирующую и аминокислотную последовательности, представленные в таблицах 7А и 7В соответственно, и называется KEX2-sol-KDEL. Первичная аминокислотная последовательность KEX2, происходящая от Saccharomyces cerevisiae, была модифицирована в целях удаления мембраносвязанного домена и присоединения последовательности KDEL, нацеленной на ЭР, к С-концу белка. Другими человеческими протеазами, подходящими для расщепления полипротеинов, содержащих соответствующие сайты узнавания, являются протеазы, описанные в публикации заявки на патент США 2005/0112565. Белок sonic hedgehog выделенный из Drosophila melanogaster, в частности, из домена его процессинга, может также служить для высвобождения белков из первичного продукта трансляции полипротеина.
В объем настоящего изобретения входит модифицированная протеаза фурин, нацеленная на эндоплазматический ретикулум (ЭP), а не на транс-сеть Гольджи (TGN), как природная протеаза фурин. В работе Vorhees et al. 1995. EMBO Journal 14:4961-4975 описана часть EEDE (SEQ ID NO:10) фурина (аминокислоты 775-778), участвующая в направленной доставке данной протеазы к TGN (Nakayama et al. 1997. Biochem. Journal 327:625-635). В работе Zerangue et al. 2001. Proc. Natl. Acad. Sci. USA 98:2431-2436 описана передача сигналов в ЭР, включая последовательность KKXX у С-конца белка. Таким образом, модифицированный фурин был получен и использован в целях нацеливания фуриновой протеолитической активности не на компартмент TGN или не только на компартмент TGN и более дальние компартменты, а на в ЭP.
В другом аспекте указанный вектор включает в себя последовательность, кодирующую дополнительный сайт расщепления, расположенный между последовательностью, кодирующей первую цепь молекулы иммуноглобулина или ее фрагмента, и последовательностью, кодирующей вторую и/или третью цепь (например, дубликат первой или второй цепи) молекулы иммуноглобулина или ее фрагмента (то есть последовательностью, смежной с последовательностью сайта расщепления, которым может быть сайт расщепления 2А). В одном из репрезентативных подходов дополнительным сайтом протеолитического расщепления является сайт расщепления фурином, содержащий консенсусную последовательность RXK(R)R (SEQ ID NO:1 ).
Регуляторные последовательности, включающие промоторы; клетки-хозяева
Вектор для экспрессии рекомбинантного иммуноглобулина или другого белка может включать любой из ряда известных промоторов, где таким промотором является конститутивный, регулируемый или индуцибельный промотор, а также клеткоспецифический, тканеспецифический или видоспецифический промотор. Другими конкретными примерами являются промоторы, восприимчивые к тетрациклину (Gossen M, Bujard H, Proc Natl Acad Sci U S A. 1992, 15;89(12):5547-51). Указанный вектор представляет собой репликон, адаптированный к данной клетке-хозяину, в которой может быть экспрессирован данный химерный ген, и указанный вектор предпочтительно также представляет собой репликон, который является функциональным в бактериальных клетках, преимущественно в Esсherichia coli, то есть в клетках, удобных для проведения манипуляций, обычно применяемых в молекулярной биологии.
Клетками-хозяевами для экспрессии гена могут быть, но не ограничиваются ими, клетки животного, в частности клетки млекопитающего, либо такими клетками могут быть микробные клетки (клетки бактерий, дрожжей, грибов, а предпочтительно эукариотические клетки) или клетки растений. Особенно подходящими клетками-хозяевами являются культивированные клетки насекомых, такие как клетки Spodoptera frugiperda, дрожжевые клетки, такие как Saccharomyces cerevisiae или Pichia pastoris, клетки грибов, таких как Trichoderma reesei, Aspergillus, Aureobasidum и Penicillium spр., а также могут быть использованы клетки млекопитающих, такие как CHO (клетки яичника китайского хомячка), ВНК (клетки почек детеныша хомячка), клетки COS, клетки 293, клетки 3T3 (мышиные), клетки Verо (клетки почек африканской зеленой мартышки) и клетки различных трансгенных животных, включая, но не ограничиваясь ими, свиней, мышей, крыс, овец, коз и коров. Для экспрессии в яичном белке используются куры, а для экспрессии в молоке используются трансгенные овцы, козы и коровы, и т.п. В соответствии с настоящим изобретением для экспрессии и расщепления антител в одной ORF, например для экспрессии sORF под регуляторным контролем промотора полиэдрина или другого сильного промотора в клеточной линии насекомых может быть использован бакуловирус, в частности векторы на основе AcNPV, и такие векторы и клеточные линии хорошо известны специалистам и являются коммерчески доступными. Промоторы, используемые в клетках млекопитающих, могут быть конститутивными (промотор TK герпесвируса, McKnight, Cell 31:355, 1982; ранний промотор SV40, Benoist et al. Nature 290:304, 1981, промотор вируса саркомы Рауса, Gorman et al. Proc. Natl. Acad. Sci. USA 79:6777, 1982; промотор цитомегаловируса, Foecking et al. Gene 45:101, 1980; промотор вируса мышиной опухоли молочной железы; в общих чертах см. Etcheverry in Protein Engineering: Principles and Practice, Cleland et al., eds, pp.162-181, Wiley & Sons, 1996) или регулируемыми (промотор металлотионеина, см., например, Hamer et al. J. Molec. Appl. Genet. 1:273, 1982). Векторы могут быть получены на основе вирусов, инфицирующих конкретные клетки млекопитающих, в частности ретровирусов, вирусов коровьей оспы и аденовирусов, и их производных, которые известны специалистам и являются коммерчески доступными. Такими промоторами являются, но не ограничиваются ими, промотор цитомегаловируса, поздний промотор аденовируса и промотор вируса коровьей оспы 7.5K. Могут быть также использованы векторы на основе дрожжей и грибов (см., например, Van den Handel, C. et al. (1991), Bennett, J.W. and Lasure, LL (eds.), More Gene Manipulations in Fungi, Academy Press, Inc., New York, 397-428), а также их промоторы, хорошо известные и широко используемые специалистами. Хорошо известным конститутивным дрожжевым промотором является промотор энолазы, а хорошо известным регулируемым промотором является промотор алкоголь-дегидрогеназы.
Выбор конкретных промоторов, последовательностей терминации транскрипции и других необязательных последовательностей, таких как последовательности, кодирующие тканеспецифические последовательности, в значительной степени зависит от типа клеток, в которых желательно осуществление экспрессии. Такими клетками могут быть клетки бактерий, дрожжей, грибов, млекопитающих, насекомых, кур или клетки других животных.
Сигнальные последовательности
Кодирующая последовательность белка, подвергаемого расщеплению, протеолитическому процессингу или аутопроцессингу, которая встроена в данный вектор, может также содержать одну или несколько последовательностей, кодирующих одну или несколько сигнальных последовательностей. Эти кодируемые сигнальные последовательности могут быть связаны с одним или несколькими зрелыми сегментами в полипротеине. Так, например, за последовательностью, кодирующей лидерную последовательность тяжелой цепи иммуноглобулина, может следовать последовательность, кодирующая тяжелую цепь, которая функционально присоединена с сохранением рамки считывания к остальной части последовательности, кодирующей данный полипротеин. Аналогичным образом последовательность, кодирующая лидерный пептид легкой цепи, или последовательность, кодирующая другой лидерный пептид, могут быть присоединены с сохранением рамки считывания к одной или обеим последовательностям, кодирующим легкую цепь иммуноглобулина, где указанная цепь лидерной последовательности отделена от сайта аутопроцессинга (такого как 2А) смежной цепью или последовательностью, кодирующей последовательность узнавания протеазой, с сохранением соответствующей рамки считывания.
Стехиометрия тяжелой и легкой цепей иммуноглобулина
Во многих описанных здесь вариантах изобретения легкая цепь (IgL) и тяжелая цепь (IgH) иммуноглобулина/антитела присутствуют в векторе или экспрессируются в клетке-хозяине в отношении примерно 1:1 (IgL:IgH). Хотя рекомбинантные методы, описанные здесь и в других работах, разработаны исходя из эквимолярной экспрессии тяжелой и легкой цепей (см., например, публикацию заявки на патент США 2005/0003482A1 или Международной патентной заявки WO2004/113493), однако в других вариантах настоящее изобретение относится к способам и к экспрессионным кластерам и векторам, разработанным на основе последовательностей, кодирующих легкую и тяжелую цепи в отношении 2:1, и ко-экспрессии указанных цепей, посредством аутопроцессинга или протеолитического процессинга, если первичным продуктом трансляции является полипротеин. В некоторых вариантах изобретения это отношение превышает 1:1 и составляет, например, приблизительно 2:1 или более. В конкретном варианте изобретения последовательности, кодирующие легкую и тяжелую цепь, используются в отношении, превышающем 1:1 (IgL:IgH). В конкретном варианте изобретения такое отношение IgL:IgH составляет 2:1.
Настоящее изобретение относится к клеткам-хозяевам или к стабильным клонам клеток-хозяев, трансформированным или инфицированным вектором, который содержит: последовательность, кодирующую тяжелую цепь и одну или по меньшей мере две легких цепи иммуноглобулина (то есть антитела); последовательности, кодирующие сайты расщепления, такие как сайты аутопроцессинга, сайты узнавания протеазой или сигнальные пептиды, расположенные между ними; и может дополнительно содержать последовательность или последовательности, кодирующие дополнительный сайт протеолитического расщепления. В объем настоящего изобретения также входит применение таких клеток или клонов в целях генерирования полноразмерных рекомбинантных иммуноглобулинов или их фрагментов или других биологически активных белков, состоящих из множества субъединиц (например, молекул, состоящих из двух цепей или множества цепей, или молекул, которые по своей природе продуцируются в виде про-белка и расщепляются или процессируются с высвобождением пре-белка и активной части белка). Неограничивающими примерами являются инсулин, интерлейкин-18, интерлейкин-1, белок остеогенеза 4, белок морфогенеза кости 2, любые другие белки морфогенеза кости, состоящие из двух цепей, фактор роста нервных клеток, ренин, химотрипсин, трансформирующий фактор роста β и интерлейкин 1β.
В своем родственном аспекте настоящее изобретение относится к рекомбинантной молекуле иммуноглобулина или к ее фрагменту или к другому белку, продуцируемому такими клетками или клонами, где указанный иммуноглобулин содержит аминокислоты, происходящие от сайта аутопроцессинга (такого как домен интеина или домен hedgehog), сайта расщепления или сайта отщепления сигнального пептида; а также к способам, векторам и к клеткам-хозяевам, применяемым для продуцирования таких молекул. В некоторых вариантах настоящее изобретение относится к клеткам-хозяевам, содержащим одну или несколько описанных здесь конструкций.
Настоящее изобретение относится к конструкциям, состоящим из одного вектора, экспрессирующего молекулу иммуноглобулина или ее фрагмент, и к способам их применения in vitro или in vivo. Такие векторы содержат последовательности аутопроцессинга или другие последовательности узнавания протеазой, расположенные между первой и второй, а также между второй и третьей кодирующими последовательностями иммуноглобулина, что позволяет экспрессировать функциональную молекулу антитела с использованием одного промотора и транскрипта. Репрезентативные векторные конструкции содержат последовательность, кодирующую сайт аутопроцессинга, расположенный между открытыми рамками считывания, и могут также содержать дополнительный сайт протеолитического расщепления, который является смежным с сайтом аутопроцессинга и обеспечивает удаление аминокислот, составляющих сайт аутопроцессинга после расщепления. Указанные векторные конструкции могут быть использованы в способах повышения уровня продуцирования полноразмерных биологически активных иммуноглобулинов или их фрагментов in vitro и in vivo. Другие биологически активные белки, содержащие по меньшей мере две различных цепи, могут быть получены с применением такой же стратегии, хотя очевидно, что отношение последовательностей, кодирующих эти цепи, к последовательностям, кодирующим другие цепи, необязательно должно превышать 1.
Хотя в настоящем изобретении описаны конкретные композиции и способы, однако следует отметить, что для осуществления настоящего изобретения могут быть использованы любые различные альтернативные композиции и способы. Следует также отметить, что выбор кластеров и векторов для экспрессии полипротеинов, клеток-хозяев и способов согласно изобретению может быть осуществлен в соответствии с процедурами, известными специалистам. Для осуществления настоящего изобретения могут быть применены, если это не оговорено особо, стандартные методы биологии клетки, молекулярной биологии (включая методы рекомбинантных ДНК), микробиологии, биохимии и иммунологии, которые известны специалистам в данной области. Такие методы подробно описаны в литературе, например, в таких руководствах, как Molecular Cloning: A Laboratory Manual, second edition (Sambrook et al., 1989); Oligonucleotide Synthesis (M. J. Gait, ed., 1984); Animal Cell Culture (R. I. Freshney, ed., 1987); Methods in Enzymology (Academic Press, Inc.); Handbook of Experimental Immunology (D. M. Weir & C. C. Blackwell, eds.); Gene Transfer Vectors for Mammalian Cells (J. M. Miller & M. P. Calos, eds., 1987); Current Protocols in Molecular Biology (F. M. Ausubel et al., eds., 1993); PCR: The Polymerase Chain Reaction, (Mullis et al., eds., 1994); и Current Protocols in Immunology (J. E. Coligan et al., eds., 1991), каждое из которых во всей своей полноте вводится в настоящее описание посредством ссылки.
Если это не оговорено особо, то все используемые здесь термины имеют общепринятые значения, известные специалистам, и, если это не оговорено особо, то настоящее изобретение осуществляют с применением стандартных методов микробиологии и техники рекомбинантных ДНК, известных специалистам.
Термин “модифицированный", используемый в контексте настоящего изобретения, относится к сегменту белка, в котором по сравнению с исходной молекулой по меньшей мере один аминокислотный остаток был заменен, делетирован или добавлен. Аналогичным образом, термин “нуклеиновая кислота”, используемый в контексте настоящего изобретения, относится к сегменту, в котором по сравнению с исходной молекулой по меньшей мере одна субъединица нуклеиновой кислоты была заменена, делетирована или добавлена.
Используемый здесь термин “интеин" обычно означает внутренний сегмент белка, который облегчает его собственное удаление и осуществляет объединение фланкирующих сегментов, известных как экстеины. Известно множество примеров интеинов в организмах различных типах, которые в некоторых случаях имеют общие структурные и/или функциональные признаки. В настоящем изобретении могут быть использованы интеины широкого ряда и их варианты, которые уже существуют и будут в дальнейшем выявлены или открыты. См., например, Gogarten JP et al., 2002, Annu Rev Microbiol. 2002;56:263-87; Perler, F. B. (2002), InBase, the Intein Database. Nucleic Acids Res. 30, 383-384 (а также веб-сайт в Интернете New England Biolabs, Inc., Ipswich, MA; http://www.neb.com/neb/inteins.html; Amitai G, et al., Mol Microbiol. 2003, 47(1):61-73; Gorbalenya AE, Nucleic Acids Res. 1998; 26(7): 1741-1748. неканонические интеины). Термины “интеин-содержащий элемент” или “элемент сплайсинга интеина”, используемые по отношению к белкам, могут охватывать части фланкирующих экстеинов, структурные признаки которых могут играть определенную роль в реакциях расщепления, лигирования и т.п. Этот термин может также означать категорию, к которой относится система, полученная на основе интеинов и включающая компонент “модифицированный интеин”.
Используемый здесь термин “модифицированный интеин” означает синтезированный интеин или природный интеин, где в “элементе сплайсинга интеина” был заменен, делетирован или добавлен по меньшей мере, один аминокислотный остаток, так чтобы в результате такой модификации, расщепленные или вырезанные экстеины не подвергались полному лигированию, опосредуемому интеином.
Используемый здесь термин hedgehog означает семейство генов (и соответствующих сегментов белков), имеющих структуру, обладающую аутопротеолитической функцией. Членами этого семейства являются, например, аналоги, происходящие от дрозофилы, мыши, человека и других видов. Кроме того, термин “hedgehog-сегмент" включают в себя не только члены этого семейства, но также имеет более широкое значение и относится к доменам аутопроцессинга генов warthog, groundhog и других hog-содержащих генов, происходящих от нематод, таких как Caenorhabditis elegans, и к домену аутопроцессинга Hoglet-C, происходящему от колониальных жгутиковых. См., например, Perler FB. Protein splicing of inteins and hedgehog autoproteolysis: structure, function, and evolution, Cell. 1998, 92(1 ):1 -4; Koonin, EV et al., (1995) A protein splice-junction motif in hedgehog family proteins. Trends Biochem Sci. 20(4): 141-2; Hall TM et al., (1997) Crystal structure of a hedgehog autoprocessing domain: homology between hedgehog and self-splicing proteins. Cell 91(1): 85-97; Snell EA et al, Proc. R. Soc. B (2006) 273, 401-407; Aspock et al, Genome Research, 1999, 9:909-923. Конкретным примером hedgehog-сегмента является sonic hedgehog, происходящий от Drosophila melanogaster. Этот термин может также означать категорию, к которой относится система, полученная на основе hedgehog и содержащая “модифицированный компонент hedgehog”.
Термин “модифицированный сегмент hedgehog” может означать синтетический или природный сегмент hedgehog, где в элементе сплайсинга hedgehog по меньшей мере один аминокислотный остаток был заменен, делетирован или добавлен так, чтобы в результате такой модификации расщепленные или вырезанные экстеины не подвергались полному лигированию.
Используемый здесь термин “вектор” означает молекулу ДНК или РНК, такую как плазмида, вирус или другой носитель, который содержит одну или несколько гетерологичных или рекомбинантных последовательностей ДНК и который предназначен для переноса этих последовательностей в другие клетки-хозяева. Термин “экспрессирующий вектор ” и “вектор для генотерапии” означает любой вектор, который является эффективным для встраивания и введения гетерологичных ДНК-фрагментов в клетке. Клонирующий или экспрессирующий вектор может включать дополнительные элементы, например экспрессирующий вектор может иметь две системы репликации, которые позволяют ему функционировать в двух организмах, например сохранять способность к экспрессии в человеческих клетках и способность в клонированию и амплификации в клетках прокариотического хозяина. В этих целях может быть использован любой подходящий вектор, который является эффективным для встраивания нуклеиновых кислот в клетки, так чтобы это приводило к экспрессии белка или полипептида, например вирусный или невирусный плазмидный вектор. Для осуществления настоящего изобретения могут быть использованы любые клетки, способные к эффективной экспрессии, например клетки насекомых и эукариотические клетки, такие как клетки дрожжей или млекопитающих.
Термины “гетерологичная ДНК” и “гетерологичная РНК” означают нуклеотиды, которые не являются эндогенными (то есть нативными) для данной клетки или не являются частью генома или вектора, в котором они присутствуют. Обычно гетерологичные ДНК или РНК добавляют в клетку путем трансдукции, инфецирования, трансфекции, трансформации, электропорации, биобаллистической трансформации или т.п. Такими нуклеотидами обычно являются по меньшей мере одна кодирующая последовательность, и эта кодирующая последовательность необязательно должна быть экспрессированной. Термин “гетерологичная ДНК” может означать “гетерологичную кодирующую последовательность” или “трансген”.
Используемые здесь термины “белок” и “полипептид” являются взаимозаменяемыми и обычно означают представляющие интерес “белки” и “полипептиды”, которые могут быть экспрессированы с использованием векторов согласно изобретению, содержащих сайт аутопроцессинга. Такими “белками” и “полипептидами” могут быть любые белки и полипептиды, которые обычно используются для диагностических или терапевтических исследований, более подробно описанных ниже. Используемый здесь термин “полипротеин” означает белок, подвергаемый процессингу в целях продуцирования двух или более полипептидных продуктов.
Используемый здесь термин “мультимер” означает белок, состоящий из двух или нескольких полипептидных цепей (иногда называемых “субъединицами”), которые после сборки образуют функциональный белок. Мультимеры могут состоять из двух (димеры), трех (триметы), четырех (тетрамеры) или более (пентамеры и так далее) пептидных цепей. Мультимеры могут образовываться в результате самосборки, либо для их сборки может потребоваться определенный компонент, такой как катализатор. Мультимеры могут состоять лишь из одних идентичных пептидных цепей (гомомультимеры) или из двух или нескольких различных пептидных цепей (гетеромультимеры). Такие мультимеры могут быть структурно или химически функциональными. Многие мультимеры известны и широко используются специалистами, включая, но не ограничиваясь ими, ферменты, гормоны, антитела, цитокины, хемокины и рецепторы. Такие мультимеры могут иметь биологическое применение (например, в фармацевтике) и промышленное применение (например, в биопроцессинге/биосинтезе).
Используемый здесь термин “метка" означает пептид, который может быть введен в экспрессирующий вектор, в котором он способен обеспечивать детектирование и/или очистку одного или нескольких продуктов экспрессии векторных вставок. Такие метки хорошо известны специалистам и могут включать радиоактивно меченные аминокислоты, либо они могут быть присоединены к полипептиду, состоящему из биотинилированных частей, которые могут быть детектированы по меченому авидину (например, стрептавидину, содержащему флуоресцентный маркер или активный фермент, который может быть детектирован методами оптической микроскопии или колориметрии). В системах экспрессии и очистки белка широко используются аффинные метки, такие как FLAG, глутатион-S-трансфераза; белок, связывающийся с мальтозой; домен, связывающийся с целлюлозой, тиоредоксин, NusA, мистин, домен, связывающийся с хитином; кутиназа, AGT, GFP и др. Другими неограничивающимися примерами меток для полипептидов являются, но не ограничиваются ими, гистидиновые метки, радиоизотопы или радионуклиды (например, 3Н, 14С, 35S, 90Y, 99Tc, 111In, 125I, 131I, 177Lu, 166Ho или 153Sm); флуоресцентные метки (например, ФИТЦ, родамин, комплексы “лантанид-фосфор”), ферментные метки (например, пероксидаза хрена, β-галактозидаза, люцифераза, щелочная фосфатаза), хемилюминесцентные маркеры, биотинильные группы, предварительно определенные полипептидные эпитопы, распознаваемые вторичной репортерной молекулой (например, двухкомпонентные последовательности “лейциновой молнии”, сайты связывания для “вторых” антител, металлосвязывающие домены, эпитопные метки) и магнитные вещества, такие как гадолиниевые хелатные комплексы.
Используемый здесь термин “дефектный по репликации”, относящийся к вирусному вектору для генотерапии согласно изобретению, означает, что вирусный вектор не может также независимо реплицироваться и упаковываться в его геноме. Так, например, если клетка индивидуума инфицирована вирионами rAAV, то гетерологичный ген экспрессируется в инфицированных клетках, однако поскольку инфицированные клетки не содержат генов rep и cap AAV и вспомогательных функциональных генов, то rAAV не способен реплицироваться.
Используемый здесь термин “ретровирусный вектор для переноса генов” означает экспрессирующий вектор, содержащий нуклеотидную последовательность, кодирующую трансген, а также нуклеотидные последовательности, необходимые для упаковки вектора. Предпочтительно ретровирусный вектор для переноса генов также содержит последовательности, необходимые для экспрессии трансгена в клетках.
Используемый здесь термин “система упаковки” означает ряд вирусных конструкций, содержащих гены, которые кодируют вирусные белки, участвующие в упаковке рекомбинантного вируса. Такие конструкции системы упаковки обычно вводят в упаковывающую клетку.
Используемый здесь термин система лентивирусных векторов “второй генерации” означает лентивирусную систему упаковки, не содержащую функциональных вспомогательных генов, таких как гены, в которых были делетированы или инактивированы вспомогательные гены vif, vpr, vpu и nef. См., например, Zufferey et al. 1997. Nat. Biotechnol. 15:871-875.
Используемый здесь термин система лентивирусных векторов “третьей генерации” означает лентивирусную систему упаковки, которая обладает свойствами векторной системы второй генерации и в которой отсутствует функциональный ген tat, такую как система, в которой ген tat был делетирован или инактивирован. Ген, кодирующий rev, обычно присутствует на отдельной экспрессирующей конструкции. См., например, Dull et al. 1998. J. Virol. 72:8463-8471.
Используемый здесь термин "псевдотипированный", относящийся к вирусу или к вирусному вектору, означает замену нативного белка вирусной оболочки гетерологичным или функционально модифицированным белком вирусной оболочки.
Используемый здесь термин “функционально присоединенный”, относящийся к рекомбинантной ДНК-конструкции или к рекомбинантному ДНК-вектору, означает что нуклеотидные компоненты рекомбинантной ДНК-конструкции или рекомбинантного ДНК-вектора обычно ковалентно присоединены друг к другу. Вообще говоря, “функционально присоединенные” последовательности ДНК являются непрерывными, а в случае секреторной лидерной последовательности они являются непрерывными и находятся в одной и той же рамке считывания. Однако энхансеры не должны быть обязательно смежными с последовательностями, экспрессию которых они активируют. Этот термин является синонимом термина “правильно локализованный”.
Энхансерные последовательности влияют на зависимую от промотора экспрессию гена и могут быть локализованы в 5'- или 3'-областях нативного гена. “Энхансеры” представляют собой цис-действующие элементы, которые стимулируют или ингибируют транскрипцию смежных генов. Энхансер, который ингибирует транскрипцию, также называется “сайленсером”. Энхансеры могут функционировать (то есть они могут быть связаны с кодирующей последовательностью) в любой ориентации на расстоянии нескольких тысяч пар (т.п.н.) от кодирующей последовательности и от положения, находящегося за транскрибируемой областью. Кроме того, для повышения уровня транскрипции стабильно интегрированных генных кластеров могут быть использованы инсуляторы или последовательности расплетания хроматина, такие как области связывания с матриксом (Chung, Cell, 1993, Aug 13;74(3):505-14, Frisch et al, Genome Research, 2001, 12:349-354, Kim et al, J. Biotech 107, 2004, 95-105).
Используемый здесь термин "ген" или "кодирующая последовательность" означает последовательность нуклеиновой кислоты, которая транскрибируется (ДНК) и транслируется (мРНК) в полипептид in vitro или in vivo, в том случае если она функционально присоединена к соответствующим регуляторным последовательностям. Ген может включать, а может и не включать области, находящиеся перед кодирующей областью и за кодирующей областью, например, 5'-нетранслируемую последовательность (5'-UTR) или "лидерную" последовательность и 3'-UTR-последовательности или "трейлерные" последовательности, а также промежуточные последовательности (интроны), расположенные между отдельными кодирующими сегментами (экзонами).
Термин "промотор" означает последовательность ДНК, которая регулирует связывание РНК-полимеразы и тем самым стимулирует синтез РНК, то есть минимальную последовательность, достаточную для регуляции транскрипции. Промоторы и соответствующая экспрессия белка или полипептида могут быть клеткоспецифическими, тканеспецифическими или видоспецифическими. Конструкции или векторы, полученные на основе нуклеиновых кислот согласно изобретению, также включают в себя энхансерные последовательности, которые могут быть, а могут и не быть смежными с промоторной последовательностью.
Термины "последовательности регуляции транскрипции" или "последовательности регуляции экспрессии", используемые здесь в широком смысле, включают в себя промоторную последовательность и физически ассоциированные с ней последовательности, которые модулируют или регулируют транскрипцию ассоциированной кодирующей последовательности часто в ответ на сигналы, передаваемые питательными элементами или факторами окружающей среды. Такие ассоциированные последовательности могут определять уровень ткане- или клеткоспецифической экспрессии, уровень ответа на внешний сигнал, уровень связывания белка, который повышает или снижает уровень транскрипции, и т.п. “Регуляторный промотор” представляет собой промотор, на активность которого влияет цис- или транс-действующий фактор (например, индуцибельный промотор, который активируется внешним сигналом или агентом).
"Конститутивный промотор” представляет собой любой промотор, который регулирует уровень продуцирования РНК во многих или во всех тканях/клетках почти во все периоды времени, так, например, таким промотором может быть область предраннего энхансера/промотора человеческого CMV, которая стимулирует конститутивную экспрессию клонированных ДНК-вставок в клетках млекопитающих.
Используемые здесь термины “белок регуляции транскрипции”, фактор регуляции транскрипции” и “фактор транскрипции” являются взаимозаменяемыми и означают ядерный белок, который связывается с элементом ДНК-ответа, и тем самым, регулирует экспрессию ассоциированного гена или генов на уровне транскрипции. Белки регуляции транскрипции обычно непосредственно связываются с элементом ДНК-ответа, однако в некоторых случаях связывание с ДНК может опосредоваться связыванием с другим белком, который, в свою очередь, связывается с элементом ДНК-ответа или является связанным с элементом ДНК-ответа.
Используемый здесь термин "сайт связывания с рибосомой", или "IRES", означает элемент, который непосредственно стимулирует присоединение внутреннего сайта связывания рибосомы с инициирующим кодоном, таким как ATG, цистрона (белок-кодирующей области), что, тем самым, приводит к кэп-независимой трансляции гена. См., например, Jackson R J. et al. 1990. Trends Biochem Sci 15:477-83 и Jackson R. J.and Kaminski, A. 1995. RNA 1:985-1000. В приведенных здесь примерах описано использование любого элемента IRES, способного стимулировать прямое присоединение внутреннего сайта связывания с рибосомой к инициирующему кодону цистрона. Используемый здесь термин "под транскрипционным контролем IRES" означает, что трансляция ассоциируется с IRES и осуществляется по кэп-независимому механизму. Так, например, последовательности, кодирующие тяжелую цепь и две легкие цепи, могут транслироваться посредством IRES, разделяющего отдельные кодирующие последовательности, при этом для отделения двух цепей друг от друга не требуется осуществления протеолитического процессинга или аутопроцессинга.
Термины “сайт аутопроцессинга" или “последовательность аутопроцессинга” определены здесь как последовательность сайта посттрансляционного или ко-трансляционного расщепления посредством процессинга. Такой термин “сайт аутопроцессинга” или “последовательность аутопроцессинга” означает последовательность ДНК или аминокислотную последовательность, представленные здесь как сайтом, последовательностью или доменом 2А или 2А-подобным сайтом, последовательностью или доменом. Используемый здесь термин “пептид аутопроцессинга” определен как продукт экспрессии пептида из последовательности ДНК, кодирующей сайт или последовательность аутопроцессинга, где указанный пептид после его трансляции опосредует быстрое внутримолекулярное (цис) расщепление белка или полипептида, содержащего сайт аутопроцессинга, с образованием дискретных зрелых белковых или полипептидных продуктов.
Используемый здесь термин "дополнительный сайт протеолитического расщепления" означает последовательность, которую вводят в экспрессионную конструкцию согласно изобретению и которая является смежной с сайтом аутопроцессинга, таким как последовательность 2А или 2А-подобная последовательность, и обеспечивает удаление дополнительных аминокислот, оставшихся после отщепления последовательности посредством аутопроцессинга. Репрезентативными “дополнительными сайтами протеолитического расщепления”, описанными в настоящей заявке, являются, но не ограничиваются ими, сайты расщепления фурином, содержащие консенсусную последовательность RXK/R-R. Такие сайты расщепления фурином могут отщепляться под действием эндогенных субтилизин-подобных протеаз, таких как фурин, и других подобных сериновых протеаз на пути секреции белка.
Используемые здесь термины "иммуноглобулин" и "антитело" означают интактные молекулы, а также их фрагменты, такие как Fa, F(ab')2 и Fv, способные связываться с представляющей интерес антигенной детерминантой. Такие молекулы, как "иммуноглобулин" и "антитело", состоят из двух одинаковых легких полипептидных цепей, молекулярная масса которых составляет 23000 дальтон, и двух одинаковых тяжелых цепей, молекулярная масса которых составляет 53000-70000. Эти четыре цепи связаны дисульфидными связями в конфигурации “Y”. Тяжелые цепи подразделяются на гамма (IgG), мю (IgM), альфа (IgA), дельта (IgD) или эпсилон (IgE) и являются основой для присвоения иммуноглобулину того или иного класса, который определяет эффекторную функцию данного антитела. Легкие цепи подразделяются на цепи каппа или лямбда. Что касается терминов “иммуноглобулина или его фрагментов”, то следует отметить, что такой термин, как “его фрагмент”, означает иммунологически функциональный фрагмент иммуноглобулина, в частности фрагмент, который связывается со своим когнатным лигандом с аффинностью связывания, составляющей по меньшей мере 10% от аффинности связывания интактного иммуноглобулина.
Fab-фрагмент антитела представляет собой одновалентный антиген-связывающий фрагмент молекулы антитела. Fv-фрагмент представляет собой генетически сконструированный фрагмент, содержащий вариабельную область легкой цепи и вариабельные области тяжелой цепи, экспрессируемые в виде двух цепей.
Термин “гуманизованное антитело” означает молекулу антитела, в которой одна или несколько аминокислот заменены в областях, не связывающихся с антигеном, так чтобы они сохраняли наибольшее сходство с аминокислотами человеческого антитела, а также связывающую активность исходного антитела. См., например, патент США № 6602503.
Используемый здесь термин “антигенная детерминанта” означает фрагмент молекулы (то есть эпитоп), с которым взаимодействует конкретное антитело. Различные области белка или пептида либо гликопептида белка или гликопротеина могут индуцировать вырабатывание антител, которые специфически связываются с данной областью или с трехмерной структурой на данном белке. Такие области или структуры называются антигенными детерминантами или эпитопами. Антигенная детерминанта может конкурировать с интактным антигеном (то есть с иммуногеном, используемым для вырабатывания иммунного ответа) за связывание с антителом.
Термин "фрагмент", если он относится к рекомбинантному белку или полипептиду согласно изобретению, означает пептид или полипептид, имеющий аминокислотную последовательность, которая является частью аминокислотной последовательности, но не всей последовательности, соответствующей полноразмерному белку или полипептиду, и которая сохраняет по меньшей мере одну функцию или активность соответствующего полноразмерного белка или полипептида. Такой фрагмент предпочтительно включает в себя по меньшей мере 20-100 смежных аминокислотных остатков указанного полноразмерного белка или полипептида.
Используемые здесь термины “введение" или “доставка" означают введение белка (включая иммуноглобулин) человеку или животному, нуждающемуся в этом, любым способом, известным специалистам. Фармацевтические носители и препараты или композиции также хорошо известны специалистам. Способами введения могут быть внутривенное, внутримышечное, интрадермальное, подкожное и трансдермальное введение, а также введение в слизистую, внутрь опухоли или через слизистую. Альтернативно, эти термины могут также означать доставку вектора для рекомбинантной экспрессии белка в клетку или клеточную культуру и/или в клетки или в органы индивидуума. Такое введение или такая доставка могут быть осуществлены in vivo, in vitro или ex vivo. Вектор для экспрессии рекомбинантного белка или полипептида может быть введен в клетку путем трансфекции, которая обычно означает введение гетерологичной ДНК в клетку физическими методами (например, трансфекция, опосредуемая фосфатом кальция, электропорация, микроинжекция или липофекция); путем инфицирования, которое обычно означает введение посредством инфекционного агента, то есть вируса; или путем трансдукции, которая обычно означает стабильное инфицирование клетки вирусом или перенос генетического материала из одного микроорганизма в другой микроорганизм посредством вирусного агента (например, бактериофага).
Термин "трансформация" обычно относится к бактериям, содержащим гетерологичную ДНК или клетки, которые экспрессируют онкоген, в результате чего они трансформируются в непрерывно растущие клетки, например в опухолевые клетки. Вектором, используемым для “трансформации” клетки, может быть плазмида, вирус или другой носитель.
Обычно клетка называется “трансдуцированной”, “инфицированной”, “трансфицированной” или “трансформированной” в зависимости от способа введения, доставки или встраивания гетерологичной ДНК (то есть вектора) в клетку. Используемые здесь термины “трансдуцированный”, “трансфицированный” и “трансформированный” могут быть взаимозаменяемыми независимо от способа введения гетерологичной ДНК.
Используемые здесь термины “стабильно трансформированный", “стабильно трансфицированный” и “трансгенный” относятся к клеткам, которые имеют неприродную (гетерологичную) последовательность нуклеиновой кислоты, интегрированную в геном. Стабильная трансфекция характеризуется устойчивостью клеточных линий или клонов, состоящих из популяции дочерних клеток, содержащих трансфицированную ДНК, стабильно реплицирующуюся посредством интеграции в их геномы или в виде эписомного элемента. В некоторых случаях “трансфекция” не является стабильной, то есть она является временной. В случае временной трансфекции экзогенная или гетерологичная ДНК экспрессируется в клетке-хозяине, но при этом введенная последовательность не интегрируется в геном данной клетки-хозяина, либо эта клетка не способна реплицироваться.
Используемый здесь термин “введение ex vivo” означает процесс, при котором у индивидуума берут первичные клетки, а затем в эти клетки вводят вектор для продуцирования трансдуцированных, инфицированных или трансфицированных рекомбинантных клеток, после чего такие рекомбинантные клетки снова вводят тому же самому или другому индивидууму.
Термин "мультицистронный транскрипт" означает молекулу мРНК, содержащую более чем одну белок-кодирующую область или цистрон. мРНК, включающая две кодирующие области, называется “бицистронным транскриптом”. “5'-проксимальная” кодирующая область или цистрон представляют собой кодирующую область, в которой кодон инициации трансляции (обычно AUG) наиболее близко расположен к 5'-концу мультицистронной молекулы мРНК. “5'-дистальная” кодирующая область или цистрон представляют собой область или цистрон, в которых кодон инициации трансляции (обычно AUG) находится на значительном расстоянии от 5'-конца данной мРНК.
Термины “5'-дистальный” и “нижерасположенный” являются синонимами и означают кодирующие области, не смежные с 5'-концом молекулы мРНК.
Используемый здесь термин "ко-транскрибируемый" означает, что две (или более) открытых рамки считывания или кодирующие области или два (или более) полинуклеотида находятся под транскрипционным контролем одного элемента регуляции транскрипции или регуляторного элемента, содержащего промотор.
Используемый здесь термин "клетка-хозяин" означает клетку, которая была трансдуцирована, инфицирована, трансфицирована или трансформирована вектором. Таким вектором может быть плазмида, вирусная частица, фаг и т.п. Условиями культивирования, такими как температура, рН и т.п., являются условия, ранее используемые для культивирования клеток-хозяев, выбранных для экспрессии, и такие условия хорошо известны специалистам. Следует также отметить, что термин “клетка-хозяин” также включает в себя природную трансдуцированную, инфицированную, трансфицированную или трансформированную клетку и ее потомство.
Используемые здесь термины "биологическая активность" и “биологически активный” относятся к активности, которой обладает конкретный белок в клеточной линии в культуре или в бесклеточной системе, такой как ELISA-планшеты, в анализе на связывание лиганд-рецептор. Термин “биологическая активность иммуноглобулина, антитела или его фрагмента” означает его способность связываться с антигенной детерминантой и, тем самым, стимулировать иммунологическую функцию. “Биологическая активность” гормона или интерлейкина известна специалистам.
Используемые здесь термины "опухоль” и "рак" относятся к клеткам, у которых отсутствует, по меньшей мере частично, регуляция ее роста по сравнению с клетками с нормальным ростом и/или развитием. Так, например, в большинстве случаев опухолевые или раковые клетки обычно не подвержены контактному ингибированию и могут обладать инвазивными свойствами и/или способностью к метастазированию.
Антителами являются белки иммуноглобулины, которые представляют собой гетеродимеры, состоящие из тяжелой и легкой цепи. Типичным антителом является мультимерное антитело с двумя тяжелыми цепями и с двумя легкими цепями (или с их функциональными фрагментами), которые связаны друг с другом. Антитела могут иметь и другую упорядоченную полимерную структуру, то есть они могут быть димерными, тримерными, тетрамерными, пентамерными и т.п., в основном в зависимости от изотипа. Было подтверждено, что такие антитела крайне трудно поддаются экспрессии в полноразмерной форме из одного вектора или из двух векторов в системах экспрессии клеточной культуры млекопитающих. В настоящее время для продуцирования антител применяются несколько методов, таких как in vivo иммунизация животных для продуцирования “поликлональных” антител; культивирование клеток В-клеточной гибридомы in vitro для продуцирования моноклональных антител (см. публикацию Kohler, et al. 1988. Eur. J. Immunol. 6:511 ; Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, 1988; которая вводится в настоящее описание посредством ссылки) и метод рекомбинантных ДНК (описанный, например, Cabilly et al., в патенте США No. 6331415, который вводится в настоящее описание посредством ссылки).
Хорошо известно, что основная молекулярная структура иммуноглобулиновых полипептидов включает в себя две идентичных легких цепи с молекулярной массой приблизительно 23000 дальтон и две одинаковые тяжелые цепи с молекулярной массой 53000-75000, где указанные четыре цепи связаны дисульфидными связями в конфигурации “Y”. Аминокислотная последовательность простирается от N-конца верхней части Y до С-конца нижней части каждой цепи. У N-конца расположена вариабельная область (длиной приблизительно 100 аминокислот), обладающая специфичностью связывания с антигеном.
Настоящее изобретение относится к усовершенствованным способам продуцирования иммуноглобулинов всех типов, включая, но не ограничиваясь ими, полноразмерные антитела и фрагменты антител, имеющих нативную последовательность (то есть последовательность, продуцируемую в ответ на стимуляцию антигеном), одноцепочечные антитела, которые представляют собой комбинацию антигенсвязывающей вариабельной области тяжелой и легкой цепей в одной полипептидной цепи со стабильной укладкой; одновалентные антитела (содержащие димер тяжелой цепи/легкой цепи, связанный с Fc-областью второй тяжелой цепи); “Fab-фрагменты”, которые включают в себя полноразмерную область “Y” молекулы иммуноглобулина, то есть ветви “Y”, либо одну легкую цепь или одну тяжелую цепь или их части (то есть агрегаты, состоящие из одной тяжелой цепи и одной легкой цепи, обычно известные как “Fab”); “гибридные иммуноглобулины”, которые обладают специфичностью к двум или более различным антигенам (например, квадромы или биспецифические антитела, описанные, например, в патенте США № 6623940); "композиционные иммуноглобулины", в которых тяжелая и легкая цепи имитируют цепи от различных видов или цепи с различными специфичностями; и “химерные антитела”, в которых части каждой из аминокислотных последовательностей тяжелой и легкой цепи происходят от более чем одного вида (то есть вариабельная область происходит от одного источника, такого как мышиное антитело, а константная область происходит от другого источника, такого как человеческое антитело).
Композиции и способы согласно изобретению могут быть применены для продуцирования иммуноглобулинов или их фрагментов, где указанная тяжелая цепь или легкая цепь является цепью “млекопитающего”, “химерной цепью” или цепью, модифицированной для повышения ее эффективности. Модифицированными антителами являются варианты аминокислотных последовательностей и варианты, кодируемые последовательностью нуклеиновой кислоты, которые сохраняют биологическую активность немодифицировнной формы, а также варианты, имеющие измененную активность, то есть имеют модификации в константной области, которые улучшают фиксацию комплемента, взаимодействие с мембранами и другие эффекторные функции, или модификации в вариабельной области, которые улучшают антигенсвязывающие свойства. Композиции и способы согласно изобретению могут также включать каталитические иммуноглобулины или их фрагменты.
Нуклеотидная последовательность, кодирующая “вариант” иммуноглобулина, может кодировать аминокислотную последовательность “варианта” иммуноглобулина, в которой по сравнению с эталонной полипептидной последовательностью модифицированы одна или несколько аминокислот. Также обсуждаются варианты модификаций других представляющих интерес биологически активных последовательностей белка (и их кодирующих последовательностей). Вариант полинуклеотидной последовательности может кодировать вариант аминокислотной последовательности, содержащий “консервативные” замены, где замененная аминокислота обладает структурными или химическими свойствами, аналогичными свойствам исходной аминокислоты. Следует отметить, что такой вариант представляющего интерес белка может быть получен путем замены аминокислотной последовательности на последовательность, которая, по существу, идентична (по меньшей мере, примерно на 80-99%, включая все целые промежуточные значения в данном интервале) природной аминокислотной последовательности, где указанный вариант образует функциональный эквивалент, который имеет трехмерную структуру и сохраняет биологическую активность природного белка. Специалистам в области биологии хорошо известно, что в последовательностях белка могут быть сделаны некоторые аминокислотные замены, не влияющие на функцию данного белка. Консервативные аминокислотные замены или замены на аналогичные аминокислоты обычно не влияют на функцию белка. Аналогичными аминокислотами могут быть аминокислоты, которые имеют аналогичный размер и/или заряд, так, например, такими парами аналогичных аминокислот являются аспартат и глутамат, и изолейцин и валин.
Замены одной аминокислоты на другую аминокислоту допускается в том случае, если это не приводит к нарушению образования нативной вторичной и третичной структуры. Сходство между парами аминокислот оценивается специалистами с применением различных методов. См. например, публикцию Dayhoff et al., Atlas of Protein Sequence and Structure, 1978. Volume 5, Supplement 3, Chapter 22, pages 345-352, которая вводится в настоящее описание посредством ссылки и в которой приводятся таблицы частоты встречаемости аминокислотных замен, которые могут быть использованы для оценки сходства аминокислот. Таблицы частоты встречаемости Dayhoff и др. были составлены на основе сравнения аминокислотных последовательностей белков, обладающих такими же функциями, как белки, происходящие от ряда эволюционно отдаленных источников.
Мутантные варианты, имеющие замены, инсерции и делеции в описанных нуклеотидных (и аминокислотных) последовательностях, могут быть легко получены методами, хорошо известными специалистам. Эти варианты могут иметь такое же применение, как конкретно описанные последовательности, при условии что такие варианты имеют последовательность, по существу, идентичную конкретно описанной последовательности согласно изобретению и сохраняют желательные функциональные свойства.
Используемый здесь термин “значительная идентичность последовательностей” означает гомологию (или идентичность), которая является достаточной для сохранения функций мутантного полинуклеотида или белка, аналогичных функциям полинуклеотида или белка, от которых происходит данный вариант. При этом предпочтительно, чтобы идентичность последовательностей превышала 70% или 80%, более предпочтительно 85% или 90%, а альтернативно 95%, включая все целые значения в интервале 70-100%. Специалист в данной области может самостоятельно вносить модификации, а именно замены, инсерции и делеции, которые позволяли бы получить последовательности с эквивалентными функциями или с улучшенными функциями или с какими-либо другими технологическими преимущественными свойствами. Следует отметить, что в объем настоящего изобретения не входят аспекты/варианты, которые могут относится к любым природным белкам или которые относятся к способу оценки уже известного продукта. Специалистам хорошо известно, что полинуклеотидные последовательности согласно изобретению могут быть усеченными или как-либо иначе модифицированы, так чтобы некоторые из полученных фрагментов и/или мутантов исходной полноразмерной последовательности сохраняли нужные свойства полноразмерной последовательности. Специалистам известны различные рестрикционные ферменты, подходящие для получения фрагментов от более крупных молекул нуклеиновых кислот. Кроме того, известно, что экзонуклеаза Вal31 может быть с успехом использована для времяспецифической рестрикции ДНК. См., например, руководство Maniatis et al. 1982. Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, New York, pages 135-139, которое вводится в настоящее описание посредством ссылки. См. также Wei et al. 1983. J. Biol. Chem. 258:13006-13512. Посредством процедур с применением экзонуклеазы Bal31 (обычно называемым процедурами на основе системы “erase-a-base”) средний специалист в данной области может самостоятельно удалить нуклеотиды из одного или с обоих концов рассматриваемых нуклеиновых кислот в целях получения широкого спектра фрагментов, которые являются функционально эквивалентыми рассматриваемым нуклеотидным последовательностям. С применением этого метода средний специалист в данной области может получить сотни фрагментов с регулируемыми варьирующимися длинами, которые могут охватить любой участок вдоль всей длины исходной кодирующей последовательности. Средний специалист в данной области путем рутинного экспериментирования может проанализировать или скринировать полученные фрагменты на их свойства и определить описанные здесь функции этих фрагментов. Также хорошо известно, что мутантные последовательности полноразмерной последовательности или их фрагменты могут быть легко получены посредством сайт-направленного мутагенеза. См., например, публикации Larionov, O.A. and Nikiforov, V.G. 1982. Genetika 18:349-59; Shortle et al. (1981) Annu. Rev. Genet. 15:265-94; которые вводится в настоящее описание посредством ссылки. Специалистом в данной области путем рутинного экспериментирования могут быть введены мутации типа делеций, инсерций или замен, а затем могут быть идентифицированы полученные мутанты, обладающие желательными свойствами полноразмерной последовательности дикого типа или ее фрагментов, например мутанты, обладающие гормональной, цитокиновой, антигенсвязывающей или другой биологической активностью.
Дополнительно или альтернативно, модифицированная полинуклеотидная последовательность может кодировать модифицированную аминокислотную последовательность, содержащую “неконсервативные” замены, где указанная замененная аминокислота обладает структурными или химическими свойствами, отличающимися от свойств исходных аминокислот. Модифицированные полинуклеотиды, кодирующие иммуноглобулин, могут также кодировать модифицированные аминокислотные последовательности, содержащие аминокислотные инсерции или делеции либо то и другое. Кроме того, модифицированный полинуклеотид, кодирующий иммуноглобулин, может кодировать такой же полипептид, как и эталонная полинуклеотидная последовательность, но из-за вырожденности генетического кода он может иметь полинуклеотидную последовательность, отличающуюся от эталонной полинуклеотидной последовательности одним или несколькими нуклеотидами.
Термин “фрагмент”, если он относится к рекомбинантному иммуноглобулину согласно изобретению, означает полипептид, имеющий аминокислотную последовательность, которая является частью аминокислотной последовательности, но не всей, последовательностью соответствующего полноразмерного белка иммуноглобулина и которая, по существу, сохраняет одну биологическую функцию или активность соответствующего полноразмерного белка или по меньшей мере одну из функций или активностей соответствующего полноразмерного белка. Такой фрагмент предпочтительно включает в себя по меньшей мере 20-100 смежных аминокислотных остатков указанного полноразмерного иммуноглобулина, а предпочтительно сохраняет способность связываться с таким же антигеном, как и полноразмерное антитело.
Используемый здесь термин “идентичность последовательностей” означает идентичность последовательностей нуклеиновой кислоты или аминокислотных последовательностей в двух или более последовательностях, сопоставляемых путем их выравнивания с использованием компьютерной программы для сравнения последовательностей. Используемый здесь термин “% гомологии” является синонимом термина “% идентичности" и означает уровень идентичности двух или более последовательностей нуклеиновых кислот или аминокислотных последовательностей, выравниваемых с помощью программы для сравнения последовательностей. Так, например, используемый здесь термин “80%-ная гомология” означает 80%-ную идентичность последовательностей, установленную с помощью определенного алгоритма, в соответствии с которым гомолог данной последовательности имеет более чем 80%-ную идентичность с последовательностью по всей его длине.
Оптимальное выравнивание последовательностей, проводимое в целях их сравнения, может быть осуществлено, например, с помощью алгоритма Смита и Уотермана для определения локальной гомологии. (Smith and Waterman. 1981. Adv. Appl. Math. 2:482), с помощью алгоритма выравнивания Нидлмана и Вунша для определения гомологии. (Needleman and Wunsch 1970. J. Mol. Biol. 48:443), с помощью программы поиска сходства методом Пирсона и Липмана (Pearson and Lipman 1988. Proc. Natl. Acad. Sci. USA 85:2444), с помощью компьютерной программы реализации этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в пакете программ Wisconsin Genetics software Package, Genetics Computer Group, Madison, Wis.); с помощью алгоритма BLAST (Altschul et al. 1990. J Mol. Biol. 215:403-410), с помощью общедоступных компьютерных программ, имеющихся на веб-сайте Национального Центра Биотехнологической Информации (National Center for Biotechnology Information) (см., nlm.nih.gov/) или путем визуальной оценки (в общих чертах, см. ниже Ausubel et al.). В целях осуществления настоящего изобретения оптимальное выравнивание последовательностей для их сравнения наиболее предпочтительно проводить с помощью алгоритма Смита и Уотермана для определения локальной гомологии (Smith and Waterman. 1981. Adv. Appl. Math. 2:482). См. также Altschul et al. 1990 и Altschul et al. 1997.
Термины “идентичный” или “процент идентичности”, используемые для двух или более последовательностей нуклеиновых кислот или белков, относятся к двум или более последовательностям или подпоследовательностям, являющимся идентичными или имеющими определенный процент идентичности аминокислотных остатков или нуклеотидов при их сравнении и сопоставлении на максимальное соответствие, которое определяют с помощью одного из описанных здесь алгоритмов сравнения последовательностей, например алгоритма Смита-Уотермана, и других известных алгоритмов, например BLAST, или путем визуальной оценки.
В соответствии с этим настоящее изобретение также охватывает варианты последовательностей, кодирующих полипептиды, расщепляемые посредством аутопроцессинга, а также сами полипептиды, которые на 80, 85, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99% или более (включая все целые значения в пределах величин 80-100) идентичны нативной последовательности. Настоящее изобретение также охватывает аминокислотные фрагменты полипептидов, которые представляют собой непрерывную цепь, состоящую по меньшей мере из 5, по меньшей мере из 10 или по меньшей мере из 15 остатков; фрагменты, гомологичные этим фрагментам в соответствии с описанными критериями идентичности, и фрагменты последовательностей нуклеиновых кислот, которые представляют собой непрерывную цепь по меньшей мере из 15, по меньшей мере из 30 или по меньшей мере из 45 остатков.
Считается, что последовательность нуклеиновой кислоты “селективно гибридизуется” с рассматриваемой последовательностью нуклеиновой кислоты, если эти две последовательности специфически гибридизуются друг с другом в условиях гибридизации и промывки умеренной или высокой жесткости. Условиями гибридизации являются температура плавления (Tm) комплекса или зонда, связывающегося с нуклеиновой кислотой. Так, например, под условиями “максимальной жесткости” обычно подразумевается, что Tm составляет Tm-5°С (на 5°С ниже Tm зонда); под условиями “высокой жесткости” подразумевается, что Tm примерно на 5-10° ниже Tm зонда; под условиями “умеренной жесткости” подразумевается, что Tm примерно на 10-20°С ниже Tm зонда, а под условиями “низкой жесткости” подразумевается, что Tm примерно на 20-25° ниже Tm зонда. По своим функциям условия максимальной жесткости могут быть использованы для идентификации последовательностей, абсолютно идентичных или почти идентичных последовательностям гибридизационного зонда, а условия высокой жесткости используются для идентификации последовательностей, которые примерно на 80% или более идентичны последовательностям данного зонда.
Условия гибридизации умеренной и высокой жесткости хорошо известны специалистам (см., например, Sambrook, et al, 1989, Главы 9 и 11, и Ausubel, F. M., et al., 1993). Одним из примеров условий “высокой жесткости” является гибридизация примерно при 42°С в 50% формамиде, 5× SSC, 5× растворе Денхардта, 0,5% ДСН и 100 мкг/мл денатурированной ДНК-носителя, с последующей двукратной промывкой в 2× SSC и 0,5% ДСН при комнатной температуре и с двумя дополнительными промывками в 0,1× SSC и 0,5% ДСН при 42°С. Варианты последовательности 2А, кодирующие полипептид, обладающий такой же биологической активностью, как и представляющий интерес природный белок, и гибридизующийся в условиях гибридизации умеренной или высокой жесткости, входят в объем настоящего изобретения.
Из-за вырожденности генетического кода может быть продуцирован ряд последовательностей, кодирующих последовательность полипептида 2А или 2А-подобного полипептида или другую последовательность расщепления протеазой или сигнальной пептидазой. Так, например, триплет CGT кодирует аминокислоту аргинин. Аргинин альтернативно кодируется кодонами CGA, CGC, CGG, AGA и AGG. В связи с этим следует отметить, что такие замены синонимичных кодонов в кодирующей области подпадают под определение вариантов последовательностей, которые входят в объем настоящего изобретения.
Кроме того, следует отметить, что такие варианты последовательностей могут гибридизоваться, а могут и не гибридизоваться с родительской последовательностью в условиях высокой жесткости. Это может происходить, например, если данный вариант последовательности включает в себя другой кодон для каждой из аминокислот, кодируемых родительским нуклеотидом. Тем не менее, такие варианты конкретно рассматриваются в настоящем изобретении и входят в его объем.
В настоящее время применение антител в качестве терапевтических средств ограничено продуктивной емкостью и стоимостью современных технологий. Использование одного улучшенного вирусного или невирусного вектора для экспрессии иммуноглобулина (или другого белка) облегчает экспрессию и доставку двух или более кодирующих последовательностей, то есть кодирующих последовательностей иммуноглобулинов или других белков, обладающих би- или мультиспецифичностью, из одного вектора. Настоящее изобретение направлено на устранение указанных ограничений и может быть применимо к любому иммуноглобулину (то есть антителу) или его фрагменту или к другому многокомпонентному белку или к паре связывающихся белков, подробно описанных в настоящей заявке, включая сконструированные антитела, такие как одноцепочечные антитела, полноразмерные антитела или их фрагменты, двухцепочечные гормоны, двухцепочечные цитокины, двухцепочечные хемокины, двухцепочечные рецепторы и т.п.
IRES
Элементы внутренних сайтов связывания с рибосомами (IRES) были впервые обнаружены в мРНК пикорнавирусов (Jackson et al. 1990. Trends Biochem. Sci. 15:477-83 и Jackson and Kaminski. 1995. RNA 1:985-1000). Примерами IRES, обычно используемых специалистами, являются IRES, представленные в таблице 1, а также IRES, описанные в патенте США No. 6692736. Примерами известных “IRES" являются, но не ограничиваются ими, IRES, выделяемые из пикорнавируса (Jackson et al., 1990), IRES, выделяемые из вирусных или клеточных мРНК-источников, таких как, например, белок, связывающийся с тяжелой цепью иммуноглобулина (BiP), васкулярный эндотелиальный фактор роста (VEGF) (Huez et al. 1998. Mol. Cell. Biol. 18:6178-6190), фактор роста фибробластов 2 (FGF-2), инсулиноподобный фактор роста (IGFII), фактор инициации трансляции elF4G, дрожжевые факторы транскрипции TFIID и HAP4, IRES вируса энцефаломиокардита (EMCV), поставляемого фирмой Novagen (Duke et al. 1992. J. Virol 66:1602-9); и IRES VEGF (Huez et al. 1998. Mol. Cell. Biol. 18:6178-90). Было обнаружено, что IRES присутствует в различных вирусах, таких как кардиовирус, риновирус, афтовирус, HCV, вирус мышиного лейкоза Фрейнда (FrMLV) и вирус мышиного лейкоза Молони (MoMLV). Используемый здесь термин “IRES" включает в себя функциональные варианты последовательностей IRES, при условии что такие варианты обладают способностью стимулировать непосредственное присоединение внутреннего сайта связывания с рибосомой к инициирующему кодону цистрона. IRES может происходить от млекопитающих, вирусов или простейших.
IRES стимулирует непосредственное присоединение внутренней рибосомы к инициирующему кодону нижерасположенного цистрона, что приводит к кэп-независимой трансляции. Таким образом, продукт нижерасположенного цистрона может экспрессироваться из бицистронной (или мультицистронной) мРНК, и при этом не требуется ни расщепления полипротеина, ни продуцирования моноцистронной мРНК. Внутренние сайты связывания с рибосомой имеют длину примерно 450 нуклеотидов и характеризуются умеренной консервативностью первичной последовательности и жесткой консервативностью вторичной структуры. Наиболее важным отличительным признаком первичной последовательности IRES является богатый пиримидином сайт, начало которого находится приблизительно на 25 нуклеотидов выше от 3'-конца IRES. см. Jackson et al. (1990).
Были идентифицированы и охарактеризованы IRES трех основных классов пикорнавирусов, а именно: кардио- и афтовирусов (например, вируса энцефаломиокардита, Jang et al. 1990. Gene Dev 4:1560-1572); энтеровирусов и риновирусов (например, полиовирусов, Borman et al. 1994. EMBO J. 13:3149-3157); и вируса гепатита А (HAV), Glass et al. 1993. Virol 193:842-852). Для первых двух классов применимы два общих принципа. Во-первых, большинство последовательностей IRES, состоящих из 450 нуклеотидов, сохраняют конкретные вторичные и третичные структуры, стимулирующие связывание с рибосомой и инициацию трансляции. Во-вторых, сайт связывания с рибосомой представляет собой триплет AUG, локализованный у 3'-конца IRES, то есть расположенный приблизительно на 25 нуклеотидов ниже консервативного олигопиримидинового пути. Инициация трансляции может происходить либо в сайте связывания с рибосомой (кардовирусов), либо в следующем расположенном ниже AUG (энтеро/риновируса). Инициация происходит в обоих сайтах афтовирусов. HCV и пестивирусы, такие как вирус коровьей диареи (BVDV) или вирус классической лихорадки свиней (CSFV), имеют длинные 5'-UTR длиной в 341 и 370 нуклеотидов соответственно. Эти 5'-UTR фрагмент образуют аналогичные вторичные структуры РНК и могут обладать достаточно эффективной функцией IRES (Tsukiyama-Kohara et al. 1992. J. Virol. 66:1476-1483; Frolov et al. 1998. RNA 4:1418-1435). Недавние исследования показали, что последовательности 5'-UTR вируса мышиного лейкоза Фрейнда (MLV) и последовательности элемента 30S, подобного ретротранспозону крысиного вируса (VL30), содержат IRES-структуру, происходящую от ретровируса (Torrent et al. 1996. Hum. Gene Ther 7:603-612).
В эукариотических клетках трансляция обычно инициируется посредством сканирования рибосомой последовательности от 5'-конца кэпированной мРНК под контролем факторов инициации. Однако было обнаружено, что некоторые клеточные мРНК имеют IRES-структуру, опосредующую кэп-независимую трансляцию (van der Velde, et al. 1999. lnt J Biochem Cell Biol. 31:87-106). Примерами элементов IRES являются, но не ограничиваются ими, белок, связывающийся с тяжелой цепью иммуноглобулина (BiP) (Macejak et al. 1991. Nature 353:90-94), мРНК усиков дрозофил (Oh et al. 1992. Gene and Dev 6: 1643-1653), фактор роста фибробластов-2 (FGF-2) (Vagner et al. 1995. Mol. Cell. Biol. 15:35-44), тромбоцитарный фактор роста В (PDGF-B) (Bernstein et al. 1997. J. Biol. Chem. 272:9356-9362), инсулиноподобный фактор роста II (Teerink et al. (1995) Biochim. Biophys. Acta 1264:403-408) и фактор инициации трансляции elF4G (Gan et al. 1996. J. Biol. Chem. 271:623-626). Недавно было также обнаружено, что васкулярный эндотелиальный фактор роста (VEGF) имеет элемент IRES (Stein et al. 1998. Mol. Cell. Biol. 18:3112-3119; Huez et al. 1998. Mol. Cell. Biol.18:6178-6190). Другими примерами последовательностей IRES являются последовательности пикорнавируса (HAV) (Glass et al. 1993. Virology 193:842-852); EMCV (Jang and Wimmer. 1990. Gene Dev. 4:1560-1572); полиовируса (Borman et al. 1994. EMBO J. 13:3149-3157); HCV (Tsukiyama-Kohara et al. 1992. J. Virol. 66:1476-1483); пестивируса BVDV (Frolov et al. 1998. RNA. 4:1418-1435); LRV-1 Leishmania (Maga et al. 1995. Mol. Cell. Biol. 15:4884-4889); ретровирусов: MoMLV (Torrent et al. 1996. Hum. Gene Ther. 7:603-612), VL30 и вируса мышиной саркомы Харви, REV (Lopez-Lastra et al. 1997, Hum. Gene Ther. 8:1855-1865). IRES может быть получен стандартными методами рекомбинантных ДНК, а также методами синтеза, известными специалистам. Для удобства клонирования рестрикционные сайты могут быть присоединены по концам используемых фрагментов IRES.
Для экспрессии двух или более белков из одного транскрипта, определяемого вирусным или невирусным вектором, последовательность внутреннего сайта связывания с рибосомой (IRES) обычно используется в целях инициации экспрессии второй, третьей и четвертой кодирующей последовательности и т.п. Если две кодирующих последовательности присоединены посредством IRES, то уровень трансляционной экспрессии второй кодирующей последовательности часто значительно снижается (Furler et al. 2001, Gene Therapy 8:864-873). Действительно, использование IRES в целях регуляции транскрипции двух или более кодирующих последовательностей, функционально присоединенных к тому же самому промотору, может приводить к снижению уровня экспрессии второй, третьей и т.п. кодирующих последовательностей по сравнению с кодирующей последовательностью, смежной с промотором. Кроме того, последовательность IRES может быть достаточно длинной для обеспечения полной упаковки вектора, например IRES eCMV имеет длину 507 пар нуклеотидов.
Экспрессия белков в форме полипротеинов (в виде первичного продукта трансляции) представляет собой стратегию, адаптированную для репликации многих вирусов, включая, но не ограничиваясь ими, пикорнавирусы. После трансляции кодируемые вирусом пептиды аутопроцессинга опосредуют быстрое внутримолекулярное (цис) расщепление полипротеина с получением дискретных (зрелых) белковых продуктов. По сравнению с использованием IRES преимущество настоящего изобретения заключается в том, что оно относится к получению вектора для экспрессии рекомбинантного белка или полипептида, содержащего пептидную последовательность аутопроцессинга (представленную здесь как последовательность пептида 2A) или другие сайты расщепления протеазой, которые облегчают экспрессию двух или более последовательностей, кодирующих белок или полипептид, с использованием одного промотора, где указанные два или более белка или полипептида экспрессируются в преимущественном молярном отношении. Что касается иммуноглобулинов, то такой полипротеин кодируется последовательностью, кодирующей одну тяжелую цепь, и последовательностями, кодирующими одну или две легких цепи, причем между каждыми из этих последовательностей находится кодируемый сайт аутопроцессинга или сайт узнавания протеазой.
В интеин-содержащей конструкции может присутствовать только один из сегментов тяжелой и легкой цепей, экспрессируемых в виде гибридного полипептида с сохранением рамки считывания вместе с интеином, находящимся между двумя цепями иммуноглобулинов, причем их соответствующие признаки позволяют осуществлять расщепление в стыках цепи “интеин-иммуноглобулин” без повторного лигирования двух иммуноглобулиновых белков. В другой интеин-содержащей конструкции присутствуют один или несколько дополнительных сегментов иммуноглобулина, необязательно отделенных от первого и/или второго сегмента сайтом расщепления. Так, например, метод с использованием интеина применяется для экспрессии одного сегмента тяжелой цепи и одного сегмента легкой цепи или для экспрессии одной тяжелой цепи и двух легких цепей и т.п.
Термины “сайт аутопроцессинга” или “последовательность аутопроцессинга”, определенные выше, означают кодирующую ДНК или аминокислотную последовательность, где после трансляции происходит быстрое внутримолекулярное (цис) расщепление полипептида, содержащего сайт аутопроцессинга, с образованием дискретных зрелых белковых продуктов. Такой “сайт аутопроцессинга” может также называться сайтом ко-трансляционного или посттрансляционного процессинга, например, используемым здесь сайтом, последовательностью или доменом 2A или интеином. Трансляционный эффект сайта, последовательности или домена 2A был продемонстрирован путем модификации активности рибосомы, проводимой в целях стимуляции гидролиза сложноэфирных связей, и тем самым, высвобождения полипептида из трансляционного комплекса в соответствии с механизмом, который позволяет синтезировать дискретный нижерасположенный продукт трансляции (Donnelly, 2001). Альтернативно, сайт или домен 2A осуществляют “аутопротеолиз” или “вырезание” посредством “отщепления” их собственных С-концов по цис-механизму с продуцированием первичных продуктов расщепления (Furler and Palmenberg. 1990. Ann. Rev. Microbiol. 44:603-623). Указанный сайт аутопроцессинга может быть заменен другими последовательностями узнавания протеазой, включая сайты расщепления сигнальной пептидазой. В полипротеинах также используются интеины.
Интеины
Используемый здесь “интеин” представляет собой сегмент в экспрессированном белке, связанный с N-концом первичного продукта экспрессии посредством N-экстеина и с С-концом первичного продукта экспрессии посредством С-экстеина. Природные интеины опосредуют вырезание интеинов и повторное присоединение (лигирование белка) N- и C-экстеинов. Однако в продуктах экспрессии согласно изобретению первичная последовательность интеина или фланкирующая аминокислотная последовательность экстеина должны быть такими, чтобы расщепление белкового остова происходило без лигирования экстеинов либо с пониженным или минимальным уровнем такого лигирования, в результате чего белки экстеины будут высвобождаться из первичного продукта трансляции (полипротеина), но не будут лигироваться с образованием гибридного белка. Интеиновая часть первичного продукта экспрессии (белок, синтезированный мРНК перед каким-либо протеолитическим расщеплением) опосредует протеолитическое расщепление на стыках N-экстеина/интеина и интеина/С-экстеина. Вообще говоря, природные интеины также опосредуют совместный сплайсинг (присоединение в результате образования пептидной связи) N-экстеина и С-экстеина. Однако целью настоящего изобретения является экспрессия двух полипептидов (конкретно представленных тяжелой и легкой цепями молекулы антитела), при которой предпочтительно не происходит лигирования белка. Это может быть достигнуто путем введения интеина, который по своей природе или в результате мутации не обладает лигирующей активностью. Альтернативно сплайсинг может быть предотвращен путем мутации, внесенной в целях замены аминокислоты (аминокислот) в сайте сплайсинга или в положении рядом с таким сайтом сплайсинга для предупреждения лигирования высвобождаемых белков. См., Xu and Perler, 1996, EMBO J. 15:5146-5153; при этом Ser, Thr или Cys обычно присутствуют в начале С-экстеина.
Интеины представляют собой класс белков, гены которых находятся только в генах других белков. Вместе с фланкирующими их генами хозяина, называемыми экстеинами, интеины транскрибируются в виде одной мРНК и транслируются в виде одного полипептида. После трансляции интеины инициируют событие автокатализа, в результате чего они вырезаются, а фланкирующие их сегменты белков хозяина лигируются с образованием новой полипептидной связи. Эта реакция катализируется только интеином и не требует участия каких-либо других клеточных белков, ко-факторов или ATФ. Интеины присутствуют в различных одноклеточных организмах и имеют различные размеры. Многие интеины содержат эндонуклеазный домен, которые придает им мобильность в геномах.
Опосредуемые интеинами реакции применяются в биотехнологии, в частности, для проведения процедур in vitro, таких как очистка и конструирования белковых чипов, а также в методах селекции видов растений (Perler, F.B. 2005. IUBMB Life 57(7):469-76). В нуклеотидные последовательности нативных интеинов были внесены мутации, и сообщалось, что некоторые из полученных мутантов имели другие свойства (Xu and Perler, 1996. EMBO J. 15(9), 5146-5153). Также известно, что в катализе посттрансляционного аутопроцессинга, осуществляемого по аналогичным механизмам, участвуют помимо интеинов бактериальные интеин-подобные домены (BIL) и hedgehog-домены (Hog) аутопроцессинга, то есть другие 2 члена суперсемейства Hog/интеинов (HINT) (Dassa et.al. 2004. J. Biol. Chem. 279(31):32001-32007).
Интеины присутствуют в конкретных белках хозяев в виде вставок с сохранением рамки считывания. В реакции самосплайсинга сами интеины вырезаются из белка-предшественника, а фланкирующие области, экстеины, соединяются с восстановлением функции гена хозяина. Такие элементы также имеют эндонуклеазную функцию, ответственную за их мобильность в геномах. Интеины имеют различные размеры (134-1650 аминокислот) и были идентифицированы в геномах эубактерий, эукариотов и архебактерий. Эксперименты, проводимые с использованием систем с моделью сплайсинга/репортера, показали, что домены, обладающие эндонуклеазной функцией, функцией расщепления белка и функцией сплайсинга белка, могут быть разделены (Xu and Perler. 1996. EMBO J. 15:5146-5153). В примерах, описанных ниже, для создания гибридного белка с последовательностями, происходящими от тяжелой и легкой цепи антитела, используется интеин Pho Pol I Pyrococcus horikoshii, VMA Saccharomyces cerevisiae и Synechocystis spp. Мутации, введенные в интеин для предотвращения сплайсинга интеинов, приводят к саморасщеплению одного полипептида с продуцированием “правильно” кодируемых тяжелой и легкой цепей антитела. Эта стратегия может быть также применена для экспрессии других многоцепьевых белков, гормонов или цитокинов и адаптирована для процессинга белков-предшественников (про-белков) с образованием зрелых биологически активных форм. Хотя в настоящем описании в качестве примера приводятся интеины Pho Pol I Pyrococcus horikoshii, VMA S. cerevisiae и Synechocystis spp., однако в векторах для экспрессии полипротеинов и в способах согласно изобретению могут быть использованы и другие известные интеины.
Помимо интеинов Pho Pol I Pyrococcus horikoshii, VMA S. cerevisiae и Synechocystis spp. специалистам известны и многие другие интеины (см., например, Perler, F.B. 2002, InBase, the Intein Database, Nucl. Acids Res. 30(1):383-384, базу данных и реестр интеинов (Intein Database and Registry), доступные на веб-сайте New England Biolabs website, например, на http://tools.neb.com/inbase/). Интеины были идентифицированы в организмах широкого ряда, таких как дрожжи, микобактерии и архебактерии, обладающие максимальной термофильностью. Некоторые интеины обладают эндонуклеазной активностью, а также активностью, направленной на сайт-специфическое расщепление и сплайсинг белка. Эндонуклеазная активность не является необходимой для осуществления настоящего изобретения, и область, кодирующая эндонуклеазу, может быть делетирована при условии сохранения активности расщепления белка.
Механизм сплайсинга белка был хорошо изучен (Chong et al. 1996. J. Biol. Chem. 271: 22159-22168; Xu and Perler. 1996. EMBO J 15: 5146-5153) и было обнаружено, что консервативные аминокислоты присутствуют в положениях сплайсинга интеинов и экстеинов (Xu et al. 1994. EMBO J 13:5517-5522). Описанные здесь конструкции содержат последовательность интеина, присоединенную к 5'-концу первой кодирующей последовательности, а вторая кодирующая последовательность присоединена к С-концу интеина с сохранением рамки считывания. Подходящие последовательности интеина могут быть выделены из любых белков, которые, как известно, содержат элементы сплайсинга белка. База данных, содержащая все известные интеины, имеется в Интернете (Perler, F. B. 1999. Nucl. Acids Res. 27: 346-347). Интеин-кодирующую последовательность присоединяют (в той же рамке считывания) к 3'- 5'-концу второй кодирующей последовательности. Для доставки этого белка в определенную органеллу соответствующий пептидный сигнал может быть присоединен к кодирующей последовательности белка.
За второй экстеин-кодирующей последовательностью могут снова находиться интеин-кодирующая последовательность - экстеин-кодирующая последовательность, и число таких повторов зависит от нужного числа белков, экспрессируемых в этой клетке. Для создания конструкций, содержащих множество интеинов, может оказаться полезным использование интеиновых элементов от различных источников. После экспрессии последовательности последнего гена может оказаться желательным встраивание последовательности терминации транскрипции, включая предпочтительно последовательность полиаденилирования. Порядок расположения последовательности полиаденилирования и последовательности терминации может быть определен специалистом в данной области. В одном из вариантов изобретения за последовательностью полиаденилирования может следовать последовательность терминации.
Модифицированные элементы сплайсинга интеина были сконструированы так, чтобы представляющий интерес модифицированный интеин мог катализировать отщепление экстеинов от интеинов, но не мог катализировать лигирование этих экстеинов (см., например, патент США 7026526 и публикацию заявки на патент США 20020129400). Мутагенез С-концевого стыка экстеина в ДНК-полимеразе GB-D Pyrococcus sp. приводит к продуцированию модифицированного элемента сплайсинга, который индуцирует отщепление экстеинов от интеинов, но предотвращает последующее лигирование экстеинов (Xu and Perler. 1996. EMBO J 15: 5146-5153). Мутация с заменой серина 538 на аланин или глицин (Ser → Ala или GIy) индуцирует отщепление, но предотвращает лигирование. В таком положении замена Ser на Met или Ser на Thr также используется для достижения экспрессии полипротеина, который расщепляется на отдельные сегменты и не подвергается, по меньшей мере частично, последующему лигированию. Мутация эквивалентных остатков в других элементах сплайсинга интеина может также предотвращать лигирование сегментов экстеина, что обусловлено относительной консервативностью аминокислот у С-концевого стыка экстеин/интеин. В случае низкой консервативности/гомологии, когда, например, первые несколько остатков, например приблизительно пять остатков С-экстеина и/или последние несколько остатков сегмента интеина, систематически варьируются, данные сегменты экстеина, в частности сегменты экстеина, описанные в настоящей заявке и известные специалистам, скринируют на их способность к отщеплению, но не к сплайсингу. Существуют интеины, которые не содержат эндонуклеазного домена, и такими интеинами являются интеин dnaE Synechocystis spp и белок GyrA Mycobacterium xenopi (Magnasco et al, Biochemistry, 2004, 43, 10265-10276; Telenti et al. 1997. J. Bacterid. 179: 6378-6382). Другие интеины были либо обнаружены в природе, либо были созданы искусственно путем удаления доменов, кодирующих эндонуклеазу, из последовательностей, кодирующих содержащие эндонуклеазу интеины (Chong et al. 1997. J. Biol. Chem. 272: 15587-15590). При необходимости выбирают интеин, который, по своей природе, состоит из минимального числа аминокислот, необходимых для осуществления функции сплайсинга, а именно интеин белка GyrA Mycobacterium xenopi (Telenti et al. 1997, см. выше). В альтернативном варианте изобретения выбирают интеин, не обладающий эндонуклеазной активностью, такой как интеин белка GyrA Mycobacterium xenopi, или интеин VMA Saccharomyces cerevisiae, который был модифицирован для удаления эндонуклеазных доменов (Chong et al. 1997, см. выше).
Дополнительная модификация элемента сплайсинга интеина может приводить к изменению скорости реакции расщепления, что позволяет регулировать количество белка путем простой модификации генной последовательности элемента сплайсинга.
В одном из вариантов изобретения было показано, что если С-концевой экстеин был конструирован так, что его первым остатком является глицин или аланин, то такая модификация предотвращает лигирование экстеина с ДНК-полимеразой GB-D Pyrococcus sp. (Xu and Perler. 1996. EMBO J 15: 5146-5153). В этом варианте предпочтительные С-концевые белки экстеины обычно содержат глициновый или аланиновый остаток, который расположен за N-концевым метионином в нативной аминокислотной последовательности. Присоединение глицина или аланина данного экстеина к С-концу интеина приводит к образованию нативной аминокислотной последовательности после процессинга полипротеина. В другом варианте изобретения в С-конец экстеина искусственно вводят глицин или аланин либо путем модификации нативной последовательности, либо путем добавления дополнительного аминокислотного остатка к N-концу данной нативной последовательности. В этом варианте изобретения нативная аминокислотная последовательность белка может быть модифицирована в одной аминокислоте после процессинга полипротеина. В других вариантах изобретения могут быть использованы другие модификации, описанные в патенте США 7026526.
ДНК-последовательность интеина ДНК-полимеразного GB-D Pyrococcus sp. представлена в SEQ ID NO:1 в патенте США No. 7026526. Положение N-концевой точки стыка с экстеином представляет собой последовательность "aac" (нуклеотиды 1-3 SEQ ID NO:1) и кодирует аспарагиновый остаток. Сайты сплайсинга нативного белка-предшественника ДНК-полимеразы GB-D находятся на участке от нуклеотида 3 до нуклеотиде 1614 в SEQ ID NO:1. Положение С-концевого стыка экстеина представляет собой последовательность "agс" (нуклеотиды 1615-1617 SEQ ID NO:1) и кодирует сериновый остаток. Мутация С-концевого экстеина с заменой серина на аланин или на глицин приводит к образованию модифицированного элемента сплайсинга интеина, который способен стимулировать вырезание полипротеина без лигирования экстеиновых элементов.
ДНК-последовательность минимального интеина GyrA Mycobacterium xenopi представлена в SEQ ID NO:2 в патенте США 7026526. Положение N-концевого стыка экстеина представляет собой последовательность "tac" (нуклеотиды 1-3 SEQ ID NO:2) и кодирует тирозиновый остаток. Сайты сплайсинга белка-предшественника находятся на участке от нуклеотида 3 до нуклеотида 597 в SEQ ID NO:2. Положение С-концевого стыка экстеина представляет собой последовательность "aсс" (нуклеотиды 598-600 SEQ ID NO:2) и кодирует треониновый остаток. Мутация С-концевого экстеина с заменой треонина на аланин или глицин приводит к образованию модифицированного элемента сплайсинга интеина, который способен стимулировать вырезание полипротеина без лигирования экстеиновых элементов.
Системы 2A
Возращаясь к варианту изобретения, относящемуся к процессингу под действием протеазы 2А, следует отметить, что активность домена 2A может приводить к удалению рибосом между кодонами, что препятствует образованию пептидных связей (de Felipe et al. 2000. Human Gene Therapy 11:1921-1931; Donnelly et al. 2001. J. Gen. Virol. 82:1013-1025), хотя считается, что такой домен действует подобно аутолитическому ферменту (Ryan et al. 1989. Virology 173:35-45). Исследования, в которых область, кодирующую последовательность 2А вируса ящура (FMDV), клонировали в экспрессирующие векторы и трансфицировали в клетки-мишени, показали, что 2А FMDV-опосредуемое отщепление искусственных репортерных полипротеинов является эффективным в гетерологичных экспрессионных системах широкого ряда (в лизате из семян пшеницы и трансгенные растения табака (Halpin et al., патент США No 5846767 (1998) и Halpin et al. 1999. The Plant Journal 17:453-459); в клеточной линии человеческой глиомы Hs 683 (de Felipe et al. 1999. Gene Therapy 6:198-208), обозначаемая далее “de Felipe II"); в лизате кроличьих ретикулоцитов и человеческих клеток HTK-143 (Ryan et al. 1994. EMBO J. 13:928-933); и в клетках насекомых (Roosien et al. 1990. J. Gen. Virol. 71:1703-1711). 2A-FMDV-опосредованное расщепление гетерологичного полипротеина с образованием биологически релевантной молекулы было продемонстрировано для IL-12 (гетеродимер p40/p35; Chaplin et al. 1999. J. Interferon Cytokine Res. 19:235-241). В трансфицированных клетках COS-7, 2A FMDV опосредует расщепление полипротеина p40-2A-p35 с образованием биологически функциональных субъединиц p40 и p35, обладающих активностью, ассоциированной с IL-12.
Последовательность 2A FMDV была введена в экспрессирующие векторы отдельно или в комбинации с различными последовательностями IRES в целях конструирования бицистронных, трицистронных и тетрацистронных векторов. Эффективность 2A-опосредуемой экспрессии генов у животных была продемонстрирована Furler (2001) с использованием рекомбинантных векторов на основе аденоассоциированного вируса (AAV), кодирующих α-синуклеин и EGFP или Cu/Zn-супероксид-дисмутазу (SOD-1) и EGFP, присоединенные посредством последовательности 2А FMDV. EGFP и α-синуклеин экспрессировались в векторах, которые включали последовательность 2А, на значительно более высоких уровнях, чем в соответствующих векторах на основе IRES, а SOD-1 экспрессировалась на сравнительных или немного более высоких уровнях.
Репрезентативными последовательностями ДНК, кодирующих сайт аутопроцессинга, являются вирусные последовательности, происходящие от пикорнавирусов, включая, но не ограничиваясь ими, энтеровирусы, риновирусы, кардиовирусы, афтовирусы или вирус ящура (FMDV). В предпочтительном варианте изобретения последовательность, кодирующая сайт аутопроцессинга, происходит от FMDV. Сайтами аутопроцессинга являются, но не ограничиваются ими, домены 2А и 2А-подобные домены (см. публикацию Donnelly et al. 2001. J. Gen. Virol. 82:1027-1041, которая во всей своей полноте вводится в настоящее описание посредством ссылки).
Альтернативно, сайт аутопроцессинга может быть заменен сайтом узнавания протеазой. Подходящими сайтами узнавания протезой и когнатными сайтами узнавания являются, но не ограничиваются ими, сайты узнавания: фурином, RXR/K-R (SEQ ID NO:1); VP4 IPNV, S/TXA-S/AG (SEQ ID NO:2); протеазой вируса гравировки табака (TEV), EXXYXQ-G (SEQ ID NO:3); протеазой 3C риновируса, LEVLFQ-GP (SEQ ID NO:4); протеазой PC5/6; протеазой PACE, протеазой LPC/PC7; энтерокиназой, DDDDK-X (SEQ ID NO:5); протеазой Xa, IE/DGR-X (SEQ ID NO:6); тромбином, LVPR-GS (SEQ ID NO:7); гененазой I, PGAAH-Y(SEQ ID NO:8); и протеазой MMP; и внутренний отщепляемый сигнальный пептид, примером которого является внутренний отщепляемый сигнальный пептид вируса гриппа С (Pekosz A. 1998. Proc. Natl. Acad. Sci. USA 95: 113233-13238) (MGRMAMKWLVVIICFSITSQPASA, SEQ ID NO:11). Протеаза может продуцироваться в транс- или в цис-форме как часть полипротеина, и в этом случае она будет находиться в том же участке транскрипции и будет отделена от остального первичного продукта трансляции, например, сайтом аутопроцессинга или сайтом узнавания протеазой.
Поскольку в настоящее время участилась выдача разрешений на клиническое применение все большего числа антител, используемых в качестве терапевтических средств, то за последние 20 лет, методы производства этих терапевтических белков еще более усовершенствуются (Wurm, FM, 2004, "Production of recombinant protein therapeutics in cultivated mammalian cells," Nat. Biotechnol. 22(11): 1393). Однако для промышленного продуцирования антител желательно разработать еще более эффективные и надежные способы. Некоторыми желательными отличительными особенностями такого продуцирования являются более высокие уровни секреции антител в культуральную среду, повышенная генетическая стабильность клеточных линий-продуцентов и более высокая скорость продуцирования клеточных линий.
При поиске более эффективных способов продуцирования терапевтических антител авторами настоящего изобретения были разработаны способы экспрессии тяжелой и легкой цепей антител из одной открытой рамки считывания. В одном из таких способов последовательность, кодирующая интеин, была использована для разделения генов тяжелой и легкой цепей антител в одной открытой рамке считывания (sORF). Преимуществами такого метода экспрессии антител в sORF являются возможность изменять отношения генов тяжелой и легкой цепей, близкое расположение друг от друга полипептидов тяжелой и легкой цепей, которое благоприятствует сборке множества субъединиц в ЭР, и возможность осуществления высокоэффективной секреции белка.
Другой метод экспрессии моноклональных антител в клетках млекопитающих включает в себя введение генов тяжелой и легкой цепей в двух отдельных ORF, где каждый из этих генов имеет собственный промотор и регуляторные последовательности. При применении такого способа возникает проблема, связанная с интерференцией промоторов. Альтернативным способом введения последовательностей, кодирующих тяжелую и легкую цепь антитела, в экспрессионные клеточные линии является использование внутреннего сайта связывания с рибосомой (IRES) для разделения последовательностей, кодирующих тяжелую и легкую цепь антитела. Этот способ не нашел широкого применения из-за низкой эффективности трансляции кодирующей последовательности, расположенной за последовательностью IRES. Недавно был описан способ, в котором для разделения последовательностей, кодирующих тяжелую и легкую цепи антитела, используется последовательность, кодирующая пептид вируса ящура (пептид 2A) (Fang et.al. 2005. Nat. Biotechnol. 23(5):584-90). В этом способе тяжелая и легкая цепи антитела и пептид 2А транскрибируются в виде одной мРНК. Однако полипептиды тяжелой и легкой цепей антитела расщепляются еще до их внедрения в эндоплазматический ретикулум (ЭР). Кроме того, после расщепления/разделения тяжелой и легкой цепей у С-конца тяжелой цепи остаются две неприродных аминокислоты. Система экспрессии интеинов согласно изобретению имеет фундаментальное отличие. Эта система отличается от 2А-метода тем, что в ней полипептиды тяжелой и легкой цепей транслируются и транспортируются в ЭР в виде одного полипротеина. Преимущество способа согласно изобретению заключается в отсутствии необходимости включения неприродных аминокислот в зрелые молекулы антител.
Ниже приводится полное описание всех антитело-продуцирующих векторов, содержащих экспрессионные кластеры: промотор - сигнал секреции - тяжелая цепь - интеин дикого типа, такой как интеин Pol I Р. horikoshii - сигнал секреции - легкая цепь - polyA; промотор - сигнал секреции - тяжелая цепь - модифицированный интеин, такой как интеин Pol I Р. horikoshii - легкая цепь - polyA; промотор - сигнал секреции - тяжелая цепь - модифицированный интеин Pol, такой как интеин Pol I Р. horikoshii - сигнал секреции - легкая цепь - модифицированный интеин Pol, такой как интеин Pol I Р. horikoshii - сигнал секреции - легкая цепь - polyA; промотор - сигнал секреции - тяжелая цепь - интеин дикого типа или модифицированный интеин, такой как интеин Pol I Р. horikoshii - модифицированный сигнал секреции - легкая цепь - polyA; промотор - сигнал секреции - легкая цепь - интеин дикого типа или модифицированный интеин, такой как интеин Pol I Р. horikoshii - модифицированный сигнал секреции - тяжелая цепь - polyA; промотор - сигнал секреции - тяжелая цепь - интеин дикого типа или модифицированный интеин, такой как интеин Pol I Р. horikoshii - модифицированный сигнал секреции - легкая цепь - интеин дикого типа или модифицированный интеин, такой как интеин Pol I Р. horikoshii - модифицированный сигнал секреции - легкая цепь - polyA; промотор - сигнал секреции - тяжелая цепь - сайт расщепления фурином - модифицированный интеин, такой как интеин Pol I Р. horikoshii - сайт расщепления фурином - сигнал секреции - легкая цепь - polyA; и промотор - тяжелая цепь - сайт расщепления фурином - модифицированный интеин, такой как интеин Pol I Р. horikoshii - сайт расщепления фурином - легкая цепь - сайт расщепления фурином - модифицированный интеин, такой как интеин Pol I Р. horikoshii - сайт расщепления фурином - легкая цепь - polyA. В других конструкциях используется модифицированный интеин Pol Psp-GBD.
Конкретным примером описанного здесь полипротеина является интеин Pol I Р. horikoshii, который был присоединен с сохранением рамки считывания перед тяжелой и легкой цепями D2E7 и за ними соответственно. Аминокислота, присутствующая в положении -1, представляет собой лизин, а аминокислота, присутствующая в положении +1, представляет собой метионин, то есть первую аминокислоту сигнального пептида легкой цепи. Использование метионина в положении +1 позволяет предотвращать сплайсинг, то есть присоединение тяжелой и легкой цепей, как было продемонстрировано авторами настоящего изобретения в последних разделах данного описания, при этом следует отметить, что для сплайсинга в положении +1 необходимо присутствие нуклеофильного аминокислотного остатка, такого как серин, цистеин или треонин. Помимо интеинов дикого типа могут быть использованы и мутанты, полученные путем замены последней аминокислоты аспарагина и предпоследней аминокислоты гистидина, поскольку такие мутации обычно приводят к блокированию сплайсинга и сохранению способности к расщеплению в стыке при N-концевом сплайсинге (Mills, 2004; Xu, 1996, Chong, 1997). Альтернативно, могут быть также введены мутации с заменой 1-й аминокислоты интеина, поскольку такие мутации обычно приводят к блокированию сплайсинга, к сохранению способности к расщеплению в точке N-концевого сплайсинга и либо к блокированию расщепления, либо к аттенюированию такого расщепления в точке N-концевого сплайсинга (Nichols, 2004; Evans, 1999, и Xu, 1996). Так, например, это было продемонстрировано на примере “полного блокирования сплайсинга и ингибирования образования разветвленного промежуточного продукта, приводящего к расщеплению в обоих стыках при сплайсинге” (Xu, M.Q., EMBO vol. 15:5146-5153).
В альтернативном варианте полипептида включение сайта рсщепления фурином позволяет модифицировать последовательность стыка с последующим ее вырезанием посредством расщепления фурином в процесс секреции. Последовательность интеина дикого типа приводится в таблице 9. В ДНК-полимеразе I GB-D Pyrococcus spp., последовательности стыков расщепления/сплайсинга представляют собой RQRAIKILAN/S (SEQ ID NO: 138) (N-концевые) и HN/SYYGYYGYAK (SEQ ID NO: 139) (C-концевые). При этом желательно, чтобы область, кодирующая эндонуклеазу, вырезалась посредством рестрикции ферментом Hindlll. Такое расщепление, сплайсинг и эндонуклеазные функции не зависят друг от друга, и указанная эндонуклеазная область может быть заменена небольшим линкером с получением мини-интеинов, которые еще обладают способностью к расщеплению и сплайсингу (Telenti et al. 1997. J. Bacteriol. 179:6378-6382). Было отмечено, что по меньшей мере один дрожжевой интеин функционирует в клетках млекопитающих (Mootz et al. 2003. J. Am. Chem. Soc. 125:10561-10569). В таблицах 8A и 8B приводятся кодирующие последовательности и аминокислотные последовательности интеиновой конструкции D2E7 (иммуноглобулина); а в таблице 8C приводится полная нуклеотидная последовательность экспрессирующей векторной конструкции интеина D2E7. Описана также гибридная конструкция, кодирующая тяжелую цепь D2E7 (Humira - зарегистрированный торговый знак для адалимумаба), присоединенную к модифицированному интеину Рol1 Psp, который сам присоединен к области, кодирующей легкую цепь D2E7. Последовательность легкой цепи может быть дуплицирована, причем в этой последовательности интеин, сигнальный пептид или сайт(ы) расщепления протеазой отделяют ее от остальной части полипротеина. В этом варианте изобретения за сигналом секреции тяжелой цепи следует зрелая тяжелая цепь. Этот интеин был модифицирован, как описано выше, то есть серин 1 был заменен треонином, а внутренний HindIII-фрагмент был вырезан для блокирования эндонуклеазной активности. Такой интеин был присоединен к зрелой области легкой цепи D2E7 с сохранением рамки считывания. Альтернативный вариант включает в себя сигнал секреции легкой цепи, расположенный со стороны 5'-конца от зрелой легкой цепи. На фигурах 10 и 11 схематически представлена конструкция интеина D2E7 и вектор для ее экспрессии, а в таблицах 8A-8C представлены нуклеотидные последовательности данной экспрессирующей конструкции, полноразмерный экспрессирующий вектор и аминокислотная последовательность конструкции интеина D2E7.
Сигнальные пептиды и сигнальные пептидазы
Гипотеза сигнальной последовательности, которая утверждает, что белок в своих аминокислотных последовательностях содержит информацию о его доставке в мембрану, была выдвинута более чем тридцать лет назад. Милстейн (Milstein) и сотрудники обнаружили, что легкая цепь IgG миеломных клеток синтезируется в высокомолекулярной форме и превращается в зрелую форму при внедрении везикул эндплазматического ретикулума (микросом) в систему трансляции, и на основании этих данных была предложена модель, в которой микросомы содержат протеазу, превращающую белок-предшественник в зрелую форму посредством удаления амино-концевого пептидного сегмента. Гипотеза сигнальной последовательности вскоре была расширена путем включения различных нацеливающих последовательностей в белки, локализованные в различных внутриклеточных мембранах, таких как митохондрии и хлоропласт. Позднее было обнаружено, что эти различные нацеливающие последовательности отщепляются от экспортируемого белка специфическими сигнальными пептидазами (SP-азами).
Существуют по меньшей мере три различные SP-азы, участвующие в отщеплении сигнальных пептидов в бактериях. SP-аза I может осуществлять процессинг нелипопротеиновых субстратов, которые экспортируются по SecYEG-пути или по пути транслокации двух аргининов (Tat). Липопротеины, которые экспортируются по Sec-пути, расщепляются SP-азой II. SP-аза IV расщепляет препилины типа IV и препилин-подобные белки, которые являются компонентами аппарата секреции типа II.
В эукариотах транспорт белков в мембрану эндоплазматического ретикулума (ЭР) опосредуется сигнальными пептидами, которые доставляют эти белки либо в процессе трансляции, либо после трансляции по механизму транслокации Sec61. ЭР-сигнальные пептиды имеют признаки, аналогичные признакам пептидов бактериальных аналогов. ЭР-сигнальные пептиды отщепляются от экспортируемого белка после его транспорта в просвет ЭР под действием комплекса сигнальных пептидаз (SPC). Сигнальные пептиды, которые направляют белки в различные участки эукариотических клеток, отличаются друг от друга, поскольку эти клетки содержат множество различных мембранных и водных компартментов. Белки, которые направляются в ЭP, часто содержат отщепляемые сигнальные последовательности. Интересно отметить, что многие искусственные пептиды могут функционировать как сигналы транслокации. Самым важным ключевым признаком, очевидно, является их гидрофобность, уровень которой выше определенной пороговой величины. ЭP-сигнальные пептиды имеют более высокое содержание лейциновых остатков, чем бактериальные сигнальные пептиды. Частица, узнающая сигнал (SRP), связывается с отщепляемыми сигнальными пептидами после их высвобождения из рибосомы. SRP необходим для нацеливания образовавшегося белка в мембрану ЭP. После транслокации этого белка в просвет ЭP экспортированный белок подвергается процессингу под действием SPC. В другом варианте используются ферменты, осуществляющие процессинг сигнального пептида (лидерной последовательности), которые обычно присутствуют в эукариотических клетках. В эукариотах нацеливание белков в мембрану эндоплазматического ретикулума (ЭP) опосредуется сигнальными пептидами, которые направляют данный белок в процессе трансляции или после трансляции по механизму транслокации Sec61. ЭP-сигнальные пептиды отщепляются от экспортируемого белка после его транспорта в просвет ЭP под действием комплекса сигнальных пептидаз (SPC). Большинство известных ЭP-сигнальных пептидов отщепляются либо у N-конца или во внутренней части последовательности белка, либо вообще не отщепляются. Недавно было обнаружено, что ряд вирусных полипротеинов, таких как полипротеины, присутствующие в вирусе гепатита С, хантавирусе, флавивирусе, вирусе коревой краснухи и вирусе гриппа С, содержат внутренние сигнальные пептиды, которые, по всей вероятности, отщепляется под действием SPC ЭP. Такие исследования по созреванию вирусных полипротеинов показали, что SPC может расщеплять не только амино-концевые сигнальные пептиды, но также и пептиды, находящиеся за внутренними сигнальными пептидами.
Аспарагиновая протеаза типа презенилина, которая является пептидазой сигнального пептида (SPP), расщепляет сигнальные пептиды в их трансмембранной области. SPP играет главную роль в генерировании происходящих от сигнального пептида эпитопов HLA-E у человека. Недавно было обнаружено, что ряд вирусных полипротеинов, таких как полипротеины, присутствующие в вирусе гепатита С, хантавирусе, флавивирусе, вирусе коревой краснухи и вирусе гриппа С, содержат внутренние сигнальные пептиды, которые, по всей вероятности, отщепляются под действием SPC ЭP. Мутагенез предполагаемых элементов, специфичных к субстрату сигнальной пептидазы, может также блокировать инфекционность вируса. Такие исследования по созреванию полипротеинов также представляют особый интерес, поскольку они показали, что SPC может расщеплять не только амино-концевые сигнальные пептиды, но также и пептиды, находящиеся за внутренними сигнальными пептидами. Сигнальные пептидазы хорошо известны специалистам. См., например, Paetzel M. 2002. Chem. Rev. 102(12): 4549; Pekosz A. 1998. Proc. Natl. Acad. Sci. USA. 95:13233-13238; Marius K. 2002. Molecular Cell 10: 735-744; Okamoto K. 2004. J. Virol. 78:6370-6380, Vol. 78; Martoglio B. 2003. Human Molecular Genetics 12: R201 -R206; и Xia W. 2003. J. Cell Sci. 116:2839-2844.
Направление белков в мембрану эндоплазматического ретикулума (ЭP) опосредуется сигнальными пептидами, которые направляют данный белок в процессе его трансляции или после трансляции по механизму транслокации Sec61. ЭP-сигнальные пептиды отщепляются от экспортируемого белка после его транспорта в просвет ЭP под действием комплекса сигнальных пептидаз (SPC). Большинство известных ЭP-сигнальных пептидов отщепляются либо у N-конца, либо вообще не отщепляются или отщепляются во внутренней части последовательности белка. Недавно было обнаружено, что ряд вирусных полипротеинов, таких как полипротеины, присутствующие в вирусе гепатита С, хантавирусе, флавивирусе, вирусе коревой краснухи и вирусе гриппа С, содержат внутренние сигнальные пептиды, которые, по всей вероятности, отщепляются под действием SPC ЭP. Такие исследования по созреванию вирусных полипротеинов показали, что SPC может расщеплять не только амино-концевые сигнальные пептиды, но также и пептиды, находящиеся за внутренними сигнальными пептидами.
В настоящем изобретении используются внутренние отщепляемые сигнальные пептиды для экспрессии полипептида в одном транскрипте. Затем один транскрибируемый полипептид отщепляется под действием SPC, в результате чего образуются отдельные пептиды, которые расположены отдельно друг от друга, либо отдельные пептиды, которые подвергаются сборке в белок. Способы согласно изобретению могут быть применимы к экспрессии тяжелой и легкой цепей иммуноглобулина в одном транскрибируемом полипептиде с последующим их расщеплением и сборкой в зрелый иммуноглобулин. Такая технология может быть применена к полипептидным цитокинам, факторам роста или к ряду других белков, например IL-12p40 и IL-12p35, которые экспрессируются в одном транскрибируемом полипептиде, а затем подвергаются сборке в IL-12, либо IL-12p40 и IL-23p19, которые экспрессируются в одном транскрибируемом полипептиде, а затем подвергаются сборке в IL-23.
Метод сигнальных пептидаз может быть применим к экспрессируюшим векторам млекопитающих, которые могут обеспечивать экспрессию функционального антитела или другого процессированного продукта, процессированного из предшественника или полипротеина. В случае антитела это антитело продуцируется из вектора в виде полипротеина, содержащего тяжелую и легкую цепи, при этом промежуточная последовательность, присутствующая между тяжелой и легкой цепями, представляет собой внутренний отщепляемый сигнальный пептид. Этот внутренний отщепляемый сигнальный пептид может быть вырезан под действием протеаз, присутствующих в ЭР, главным образом, сигнальных протеаз, презенилина или презенилин-подобных протеаз, что будет приводить к фолдингу и сборке тяжелой и легкой цепей и образованию функциональной молекулы, а желательно, секретируемой молекулы. Помимо внутреннего отщепляемого сигнального пептида, происходящего от вируса гепатита С, могут быть использованы и другие внутренние отщепляемые последовательности, которые могут вырезаться под действием протеаз, присутствующих в ЭР. Аналогичным образом настоящее изобретение не ограничивается клетками-хозяевами, в которых расщепление осуществляется сигнальными пептидазами, а также включает в себя клетки, в которых действуют другие протеазы, включая, но не ограничиваясь ими, презенилин, презенилин-подобную протеазу и другие протеазы для процессинга полипептидов. Такие протеазы описаны в цитируемых здесь статьях и т.п.
Кроме того, настоящее изобретение не ограничивается экспрессией тяжелой и легкой цепей иммуноглобулина, но также включает в себя и другие полипептиды и полипротеины, экспрессируемые в одном транскрипте с последующим отщеплением внутреннего сигнального пептида и высвобождением каждого отдельного пептида или белка. Эти белки могут быть, а могут и не быть подвергнуты сборке с образованием зрелого продукта.
В объем настоящего изобретения входят также экспрессионные конструкции, в которых отдельные полипептиды следуют друг за другом, то есть расположены в следующем порядке: “пептид 1 - внутренний отщепляемый сигнальный пептид - пептид 2” или “пептид 2 - внутренний отщепляемый сигнальный пептид - пептид 1”. Настоящее изобретение также включает в себя экспрессию более чем двух пептидов, присоединенных посредством внутренних отщепляемых сигнальных пептидов, таких как “пептид 1 - внутренний отщепляемый сигнальный пептид - пептид 2 - внутренний отщепляемый сигнальный пептид - пептид 3” и т.п.
Кроме того, настоящее изобретение относится к экспрессии обоих трансмембранных белков типа I и типа II, а также других сайтов расщепления протеазой, окружающих экспрессионные конструкции. Одним из примеров может служить введение сайта расщепления фурином или сайта расщепления PC5/6 за тяжелой цепью иммуноглобулина в целях облегчения отщепления дополнительных аминокислотных остатков у карбокси-конца пептида тяжелой цепи, например, с получением конструкции “тяжелая цепь - сайт расщепления фурином - внутренний отщепляемый сигнальный пептид - легкая цепь". Настоящее изобретение также включает в себя более чем один внутренний отщепляемый сигнальный пептид, расположенный отдельно или в тандеме, например, в виде конструкции “тяжелая цепь - сайт расщепления фурином - внутренний отщепляемый сигнальный пептид - внутренний отщепляемый сигнальный пептид - легкая цепь". Кроме того, настоящее изобретение включает в себя варианты, в которых сохраняются или удаляются сами сигнальные пептиды тяжелой и легкой цепи, такие как “сигнальный пептид HC - тяжелая цепь - сайт расщепления фурином - внутренний отщепляемый сигнальный пептид - сигнальный пептид LC - легкая цепь”.
Ниже приводится описание антитело-продуцирующих векторов, некоторые из которых уже упоминались в настоящем описании. Векторными конструкциями являются, но не ограничиваются ими, конструкции, представленные ниже.
Конкретным примером может служить гибридная конструкция, кодирующая тяжелую цепь D2E7 (Humira/адалимумаб), присоединенную к внутреннему отщепляемому сигнальному пептиду, который сам присоединен к области, кодирующей легкую цепь D2E7. В этом варианте изобретения за сигналом секреции тяжелой цепи следует зрелая тяжелая цепь. Последовательность внутреннего отщепляемого сигнального пептида происходит от вируса гриппа С. Сайт расщепления фурином находится у карбокси-конца тяжелой цепи. Для минимизации влияния на зрелое антитело третий от конца аминокислотный остаток тяжелой цепи подвергают мутации путем замены пролина на аргинин с получением сайта расщепления фурином. Альтернативный вариант также включает в себя сигнал секреции легкой цепи, расположенный со стороны 5'-конца от зрелой легкой цепи. См. таблицы 9A-9C. В этом примере используется минимальная последовательность внутреннего отщепляемого сигнального пептида, происходящего от вируса гриппа С (MGRMAMKWLVVIICFSITSQPASA, SEQ ID NO:11). Для повышения эффективности отщепления может быть также использована более длинная последовательность. См. GenBank рег. № AB126196. Могут быть также использованы различные нуклеотидные последовательности, кодирующие аналогичную аминокислотную последовательность.
В настоящем изобретении могут быть также использованы внутренние отщепляемые сигнальные пептиды для получения одного или нескольких зрелых полипептидов в полипротеине, кодируемом в одном транскрипте. Затем один транскрибированный полипептид отщепляется под действием SPC, в результате чего образуются отдельные пептиды, которые расположены отдельно друг от друга, либо отдельные пептиды, которые подвергаются сборке в белок. Настоящее изобретение может быть применено к экспрессии тяжелой и легкой цепей иммуноглобулина в одном транскрибируемом полипептиде с последующей их сборкой в зрелый иммуноглобулин. Настоящее изобретение может быть применено для экспрессии полипептидных цитокинов, факторов роста или ряда других белков, например для экспрессии IL-12p40 и IL-12p35, в одном транскрибируемом полипептиде с последующей сборкой в IL-12, либо для экспрессии IL-12p40 и IL-23p19 в одном транскрибируемом полипептиде с последующей сборкой в IL-23.
Позиционное субклонирование последовательности 2A или другой протеазы или сайта расщепления (узнавания) сигнальной пептидазой, расположенных между двумя или более гетерологичными последовательностями ДНК, в целях получения векторной конструкции согласно изобретению позволяет обеспечивать доставку двух или более генов в один экспрессирующий вектор и экспрессию этих генов в данном векторе. Предпочтительно сайты аутопроцессинга, такие как последовательности 2А FMDV или последовательности узнавания протеазой, являются уникальными средствами для экспрессии в одном вирусном векторе и для доставки из одного вирусного вектора двух или множества белков, полипептидов или пептидов, которые могут представлять собой отдельные части, например, антитела, гетеродимерного рецептора или гетеродимерного белка.
Последовательность 2A FMDV представляет собой область полипротеина, функция которой в геноме FMDV направлена на одно отщепление у его С-конца, и действует, тем самым, по цис-механизму. Сообщалось, что домен 2А FMDV обычно имеет длину примерно в девятнадцать аминокислот (LLNFDLLKLAGDVESNPGP, SEQ ID NO:12; TLNFDLLKLAGDVESNPGP, SEQ ID NO: 13; Ryan et al. 1991. J. Gen. Virol. 72:2727-2732), однако было показано, что олигопептиды, имеющие по меньшей мере четырнадцать аминокислотных остатков (LLKLAGDVESNPGP, SEQ ID NO: 14), опосредуют расщепление у С-конца 2А по механизму, аналогичному механизму процессинга нативного полипротеина FMDV.
Варианты последовательности 2A были проанализированы на их способность опосредовать эффективный процессинг полипротеинов (Donnelly et al. 2001). В объем настоящего изобретения входят гомологи и варианты последовательности 2A, и такими последовательностями являются, но не ограничиваются ими, следующие последовательности:
QLLNFDLLKLAGDVESNPGP, SEQ ID NO:15;
NFDLLKLAGDVESNPGPFF, SEQ ID NO:16;
LLKLAGDVESNPGP, SEQ ID NO:17;
NFDLLKLAGDVESNPGP, SEQ ID NO:18;
APVKQTLNFDLLKLAGDVESNPGP, SEQ ID NO:19;
VTELLYRMKRAETYCPRPLLAIHPTEARHKQKIVAPVKQTLNFDLLKLAGDVESNPGP, SEQ ID NO:20;
LLAIHPTEARHKQKIVAPVKQTLNFDLLKLAGDVESNPGP, SEQ ID NO:141; и
EARHKQKIVAPVKQTLNFDLLKLAGDVESNPGP, SEQ ID NO:142.
Последовательности 2A и их варианты могут быть использованы для получения векторов, экспрессирующих полипротеины аутопроцессинга, включая любой вектор (плазмидный или вирусный), который содержит последовательности, кодирующие белки или полипептиды, присоединенные посредством сайтов аутопроцессинга или других сайтов расщепления протеазой, так что после расщепления полипротеина, обусловленного присутствием сайта аутопроцессинга или другого сайта расщепления, отдельные белки экспрессируются в соответствующих молярных отношениях и/или количествах. Эти белки могут быть гетерологичными по отношению к самому вектору, по отношению друг к другу или по отношению к сайту аутопроцессинга, например сайту FMDV, а поэтому сайты аутопроцессинга, присутствующие в гетерологичных белках и кодирующих последовательностях и используемые в настоящем изобретении, не отличаются по своей способности осуществлять или опосредовать расщепление от соответствующих сайтов аутопроцессинга, происходящих от того же источника.
В одном из вариантов изобретения последовательность 2А FMDV, присутствующая в векторе согласно изобретению, кодирует аминокислотные остатки, составляющие последовательность LLNFDLLKLAGDVESNPGP (SEQ ID NO: 12). Альтернативно, вектор согласно изобретению может кодировать аминокислотные остатки других 2А-подобных областей, обсуждаемых в работе Donnelly et al. 2001. J. Gen. Virol. 82:1027-1041, включая, но не ограничиваясь ими, 2А-подобный домен, происходящий от пикорнавируса, вируса насекомых, ротавируса типа С, повторяющихся последовательностей трипаносом или бактерии Thermatoga maritima.
В настоящем изобретении рассматривается использование вариантов последовательностей нуклеиновой кислоты, кодирующих пептидную последовательность 2А или 2А-подобную пептидную последовательность, таких как последовательность нуклеиновой кислоты, кодирующая полипептид 2А или 2А-подобный полипептид и имеюшая кодон для одной или нескольких аминокислот, отличающийся от кодона родительского нуклеотида. Такие варианты конкретно рассматриваются в настоящем изобретении и входят в его объем. В объем настоящего изобретения также входят варианты последовательностей пептидов и полипептидов 2А. Аналогичным образом протеазы, находящиеся в цис- или транс-положениях, могут опосредовать протеолитический процессинг с помощью когнатных сайтов узнавания (расщепления) протеазой, расположенных между областями полипротеина.
В других экспериментах, проводимых с использованием экспрессионных конструкций “интеин - антитело”, авторами настоящего изобретения было продемонстрировано, что реакция сплайсинга белка, опосредуемая интеином Pol I Pyrococcus horikoshii, может происходить в клетках млекопитающих (293E), в ЭP и в аминокислотных последовательностях тяжелой и легкой цепей антитела (D2E7). Путем проведения реакции этого типа в формате экспрессии антитела в одной открытой рамке считывания (sORF) авторами настоящего изобретения было продемонстрировано, что такая реакция может быть осуществлена в клетках млекопитающих (293E), в ЭP и в аминокислотных последовательностях тяжелой и легкой цепей антитела с использованием двух конструкций, pTT3-HcintLC1aa-p.hori и pTT3- HcintL.C3aa-p.hori. См. таблицы 11A и 12A.
Указанные конструкции были получены на основе вектора PTT3. Этот вектор имеет ориджин репликации вируса Эпштейна-Барра (EBV), что позволяет осуществлять его эписомную амплификацию в трансфицированных клетках 293E (в клетках, которые экспрессируют ядерный антиген 1 вируса Эпштейна-Барра) в суспензионной культуре (Durocher, 2002, “High level and high-throughput recombinant protein production by transient transfection of suspension-growing human 293-EBNA1 cells”, Nucleic Acids Research 30(2):E9). Каждый вектор имеет одну ORF, транскрипционно экспрессируемую под регуляторным контролем промотора CMV. В ORF интеин Poll P. horikoshii был встроен с сохранением рамки считывания между тяжелой и легкой цепями антитела D2E7, каждая из которых имеет сигнальный пептид (SP). Конструкции pTT3-HcintLC1aa-p.hori и pTT3-HcintLC3aa-p.hori имеют 1 аминокислоту нативного экстеина или 3 аминокислоты нативного экстеина на любой стороне интеина, отделяющие последовательности тяжелой и легкой цепей антитела D2E7 от последовательности интеина. Эти конструкции были введены в клетки 293E посредством временной трансфекции. Были проанализированы образцы супернатанта культуры и клеточного осадка.
Образцы клеточного осадка подвергали лизису в условиях, благоприятствующих разделению цитозольных и внутриклеточных мембранных фракций. Обе эти фракции анализировали с помощью вестерн-блот-анализа (WB) с использованием антител против тяжелой цепи или антител против легкой цепи каппа. На этих блотах авторы настоящего изобретения наблюдали экспрессию 4 молекул белка, соответствующих трехкомпонентной форме, наблюдаемой в ORF данных конструкций (130 кДа), то есть гибрида H и L, который был образован в результате сплайсинга (80 кДа) и содержал тяжелую цепь антитела (50 кДа) и легкую цепь антитела (25 кДа). Первые 2 молекулы белка были детектированы с использованием антител против тяжелой цепи и антител против легкой цепи, при этом тяжелая цепь детектировалась только антителом против тяжелой цепи, а легкая цепь детектировалась только антителом против легкой цепи. Присутствие в обеих конструкциях молекул белка размером в 80 кДа, которые были детектированы с использованием антител против тяжелой и легкой цепи, указывало на событие сплайсинга белка. Кроме того, все четыре молекулы белка присутствовали преимущественно в субклеточной мембранной фракции, которая содержала эндоплазматический ретикулум (ЭP). Это свидетельствовало о том, что сигнальный пептид тяжелой цепи (кодируемый в начале ORF) направляет весь полипептид в ЭP, где происходит реакция сплайсинга. Не ограничиваясь какой-либо конкретной теорией, можно лишь отметить, что свободные полипептиды тяжелой и легкой цепей, вероятно, образуются в результате расщепления на границах N-концевого и С-концевого неполного сплайсинга.
Образцы клеточного осадка были также использованы для экстракции всей РНК и проведения Нозерн-блот-анализа с помощью зонда для тяжелой цепи антитела и зонда для легкой цепи антитела. Нозерн-блот-анализ выявил присутствие в этих sORF-конструкциях трехкомпонентной мРНК (3,4 т.п.н.), которая была гибридизована с зондом тяжелой цепи и с зондом легкой цепи, но не выявил отдельных мРНК для тяжелой цепи или легкой цепи. В противоположность этому, в образцах клеточного осадка, в которых была осуществлена экспрессия антитела D2E7 с применением стандартного метода, а именно путем введения тяжелой и легкой цепей антитела от двух отдельных ORF, присутствующих в двух векторах pTT3, мРНК тяжелой цепи (1,4 т.п.н.) и легкой цепи (0,7 т.п.н.), были детектированы с использованием зондов для тяжелой цепи или легкой цепи соответственно. В контрольных образцах клеточного осадка какой-либо трехкомпонентой мРНК не обнаруживалось.
Вышеописанные данные продемонстрировали, что с использованием конструкций, содержащих одну ORF (тяжелая цепь D2E7 - интеин Р. horikoshi - легкая цепь D2E7), была транскрибирована одна мРНК, содержащая все 3 белка. Этот трехкомпонентный транскрипт был транслирован в трехкомпонентый полипептид, который в процессе котрансляции транспортировался в ЭP под действием сигнального пептида тяжелой цепи, присутствующего у N-конца трехкомпонентного полипротеина. Благодаря этой конструкции опосредуемая интеином реакция сплайсинга белка происходит внутри ЭP. Это дает основание предположить, что опосредуемые интеином реакции могут быть использованы для экспрессии антител, а также других мульти-субъединичных секретируемых белков, то есть белков, которые для их укладки и соответствующей посттрансляционной модификации должны поступать на секреторный путь.
Были также проанализированы супернатанты культур. Вестерн-блот-анализ и ELISA-анализ позволяют детектировать антитело, секретируемое в результате экспрессии конструкции pTT3-HcintLC1 aa-p.hori. Эти исследования более подробно обсуждаются ниже, при этом уровень экспрессии секретируемого антитела повышался благодаря точковым мутациям и мутациям в последовательности, кодирующей сигнальный пептид легкой цепи.
Мутации были введены в целях ингибирования опосредуемого интеином лигирования, но осуществление реакций расщепления на границах N-концевого и С-концевого сплайсинга приводило к повышению уровней секреции антитела.
Для повышения эффективности секреции антител были сконструированы и протестированы три типа точковых мутаций. Мутация первого типа была введена в кодон первого серинового остатка С-концевого экстеина, и эти конструкции имели замену Ser→Met (S→M) (конструкция pTT3-HcintLC-p.hori, конструкция E и конструкция A). Мутация второго типа была введена в положение, кодирующее первый сериновый остаток интеина, и такая конструкция имела замену Ser→Thr (S→T) (конструкция E). Мутация третьего типа была введена в кодон гистидинового остатка, который представляет собой вторую от конца (предпоследнюю) аминокислоту интеина, и такие конструкции имели мутацию с заменой His на Ala (H→A) (конструкция A и конструкция B). Эти мутации были введены либо отдельно, либо в комбинации. Все мутантные конструкции были сконструированы в целях сохранения способности к расщеплению на границах N-концевого и С-концевого сплайсинга и в целях снижения уровня сплайсинга высвобождаемых экстеинов или того и другого в соответствии с механизмами реакции, описанными в литературе. Как будет описано ниже, секреция антитела D2E7 достигается с использованием ряда этих конструкций.
В одном из экспериментов данные конструкции были введены в клетки 293E посредством временной трансфекции, и через 7 дней супернатанты культур анализировали на титры антител IgG с помощью ELISA. Титры антител для конструкций pTT3-HcintLC3aa-p.hori, pTT3-HcintLC1aa-p.hori, pTT3-HcintLC-p.hori, E, A и B составляли 17,0+0,6, 113,8+2,6, 225,8+10,0, 9,3+0,5, 161,7+4,4 и 48,2+1,0 нг/мл (среднее + ср.кв.от.) соответственно.
Эти образцы супернатанта были также проанализированы с помощью электрофореза в ДСН-ПААГ в денатурирующих условиях, а затем блот-анализа с использованием антитела против тяжелой цепи человеческого IgG и антитела против человеческой легкой цепи каппа. На этих вестерн-блотах тяжелая цепь антитела (~50 кДа) и легкая цепь антитела (~25 кДа) визуально наблюдались в супернатантах, полученных для конструкций pTT3-HcintLC-p.hori и A, что соответствовало ранговому порядку уровней IgG, измеренных с помощью ELISA.
Образцы клеточного осадка, взятые от этих трансфектантов, были также охарактеризованы с помощью вестерн-блот-анализа. Трехкомпонентный полипептид (-130 кДа), а также полосы тяжелой цепи антитела (~50 кДа) и легкой цепи антитела (~25 кДа) наблюдались в клеточных осадках, содержащих все вышеописанные конструкции. Из этих конструкций конструкция pTT3-HcintLC-p.hori и конструкция A давали наиболее яркие полосы тяжелой цепи и легкой цепи, что позволяет сделать вывод о корреляции между уровнем внутриклеточных свободных тяжелых и легких цепей и уровнем антител, подвергнутых сборке и секреции антител. Продукт сплайсинга (~80 кДа), который представляет собой гибрид тяжелой и легкой цепей антитела, присутствовал в клеточных осадках, полученных с использованием конструкции pTT3- HcintLC3aa-p.hori и, в меньшей степени, в клеточных осадках, полученных с использованием конструкции pTT3-HcintLC1aa-p.hori, при этом в конструкциях pTT3-HcintLC-p.hori и в конструкциях A, B и E эти продукты отсутствовали. Это указывает на то, что уровень сплайсинга белка был обратно пропорционален эффективности секреции антитела, что совпадает с предположением о том, что объединение тяжелой цепи и легкой цепей антитела должно приводить к нарушению укладки, если исходить из общих данных о структуре антитела, и такое нарушение укладки должно приводить к предотвращению секреции, опосредуемому клеточными механизмами деградации белков с неправильной укладкой. Другим видом белка, присутствующего на этих блотах, был гибрид “интеин - легкая цепь” (80 кДа, распознаваемый антителом против легкой цепи, но не антителом против тяжелой цепи), и этот гибрид был образован в результате расщепления на границах N-концевого сплайсинга в отсутствие любых других реакций расщепления. Указанная полоса присуствовала в конструкциях A, B, E, pTT3-HcintL.C3aa-p.hori, pTT3-HcintLC1aa-p.hori, но обычно отсутствовала в конструкциях pTT3-HcintLC-p.hori и H, описанных выше. Поэтому количество белка такого типа было также обратно пропорционально уровню секреции антитела. И наконец, полоса интеина в этих клеточных лизатах была также детектирована с использованием кроличьей сыворотки, содержащей поликлональное антитело против пептида P. horikoshii, конъюгированное с KLH.
Авторами настоящего изобретения было продемонстрировано, что антитело D2E7, секретированное с использованием sORF-конструкции pTT3-HcintLC-p.hori, имело “правильные” N-концевые последовательности тяжелой и легкой цепей, и соответствующие молекулярные массы тяжелой и легкой цепей и интактного антитела.
Антитело D2E7, секретируемое с использованием одной sORF-конструкции pTT3-HcintLC-p.hori, очищали посредством аффинной хроматографии на белке А и анализировали N-концевые последовательности их тяжелых и легких цепей. Полученные однозначные результаты показали, что N-концевой пептидной последовательностью тяжелой цепи является EVQLVESGGG (SEQ ID NO:21), а N-концевой пептидной последовательностью легкой цепи является DIQMTQSPSS (SEQ ID NO:22). Таким образом, в данной конструкции сайты расщепления, используемые сигнальной пептидазой, имеющей последовательность DIQMTQSPSS, являются такими же, как и сайты, используемые в стандартном методе, в котором для экспрессии антитела DE27 использовались две ORF/два вектора.
Эти данные имеют важное научное значение для создания конструкций следующего поколения, то есть конструкций, в которых пептидаза ЭP млекопитающих может узнавать и точно отщеплять сигнальный пептид в новосинтезированном полипротеине, даже если имеются некоторые очевидные условия для его презентации (см. ниже).
Это очищенное антитело, а также антитело D2E7, продуцированное стандартным промышленным методом, анализировали с помощью масс-спектрометрии. В денатурирующих условиях легкая цепь D2E7, полученная из конструкции pTT3-HcintLC-p.hori, давала один единственный пик на масс-спектре, а ее молекулярная масса (MW) составляла 23408,8, тогда как молекулярная масса (MW) легкой цепи D2E7, продуцированной стандартным промышленным методом, составляла 23409,7, то есть имела близкое сходство с предыдущим значением. Кроме того, в денатурирующих условиях тяжелая цепь D2E7, полученная из конструкции pTT3-HcintLC-p.hori давала на масс-спектре один главный пик и 2 слабых пика с молекулярными массами (MW) 50640,6, 50768,2 и 50802,4 соответственно, а молекулярные массы (MW) тяжелой цепи D2E7, продуцированной стандартным промышленным методом, составляли 50641,7, 50768,6 и 50804,1 соответственно, и эти значения также имели близкое сходство со стандартными значениями. Эти 3 пика соответствовали стандартным вариантам тяжелой цепи D2E7.
Молекулярные массы (MW) интактного антитела D2E7, продуцированного в природных условиях из конструкции pTT3-HcintLC-p.hori, а также антитела D2E7, продуцированного промышленным методом, были также определены с помощью масс-спектрометрии. Антитело, продуцированное из конструкции pTT3-HcintLC-p.hori, имело 3 пика с MW 148097,6, 148246,9 и 148413,1 соответственно, а антитело D2E7, продуцированное промышленным методом, также имело 3 пика с MW 148096,0, 148252,3 и 148412,8 соответственно.
Эти данные со всей очевидностью продемонстрировали, что антитело D2E7, продуцированное из конструкции pTT3-HcintLC-p.hori, имело размер, идентичный размеру антитела, продуцированного стандартным промышленным методом, как в денатурирующих, так и в природных условиях. Одним из преимуществ системы экспрессии антител согласно изобретению по сравнению со стандартным промышленным методом является ее способность продуцировать антитела с полностью аутентичными аминокислотными последовательностями. Антитело, продуцированное с использованием системы 2А, описанной, например, Fang et.al. Nature Biotechnology, 2005, имело 2 дополнительных неприродных аминокислоты у С-конца тяжелой цепи, чего невозможно было избежать из-за природного характера расщепления.
Авторами настоящего изобретения было продемонстрировано, что антитело D2E7, продуцированное из sORF-конструкции pTT3-HcintLC-p.hori, имело аффинность связывания с TNF, аналогичную аффинности связывания антитела D2E7, продуцированного промышленным методом. Взаимодейстие путем связывания между антагонистами rhTNFa, нанесенными на биосенсорный чип посредством иммобилизованного козьего антитела против человеческих IgG, и уровень растворимого rhTNFa измеряли в реальном времени на приборе Biacore 3000 (Pharmacia LKB Biotechnology, Uppsala, Sweden) в соответствии с инструкциями производителя и стандартными процедурами. Вкратце, аликвоты rhTNFa разводили в буфере HBS-EP (Biacore) и 150 мкл-аликвоты инъецировали в матрицы с иммобилизованным белком при скорости потока 25 мл/мин. Эквивалентную концентрацию аналита одновременно инъецировали на необработанную эталонную поверхность, которая служила в качестве контрольной сенсорограммы для вычитания фонового значения общего показателя преломления. В интервалах между циклами поверхность сенсорного чипа восстанавливали двумя 5-минутными инъекциями 10 мM глицина при скорости потока 25 мл/мин. Затем полученные экспериментальные сенсорограммы связывания оценивали с помощью компьютерной программы BIA Еvaluation 4.0.1 в целях определения кинетических параметров. Данные для каждого антагониста соответствовали Лангмюровской модели 1:1. Для проведения этих исследований данные по связыванию и диссоциации анализировали в соответствии с протоколом общего анализа путем подбора кривой, где такой подбор кривой проводили локально для максимальной способности связывания аналита (RU) или для параметра Rmax. В этом случае программа вычисляла одну константу диссоциации (kd), константу ассоциации (ka) и константу аффинности (Kd). Константа равновесной диссоциации (Kd) = kd/ka. Кинетическую скорость ассоциации, кинетическую скорость диссоциации и общую аффинность определяли с использованием различных концентраций TNFα в интервале 1-100 нМ. Кинетическая скорость ассоциации, кинетическая скорость диссоциации и общая аффинность для антитела D2E7, продуцированного из конструкции pTT3-HcintLC-p.hori, составляли 1,616 (M-1с-1), 5,69-5 (с-1) и 3,54-11 (M) соответственно, а кинетическая скорость ассоциации, кинетическая скорость диссоциации и общая аффинность для антитела D2E7, продуцированного промышленным методом, составляли 1,736 (M-1с-1), 6,72-5 (с-1) и 3,89-11 (M) соответственно. Анализ Biacore показал, что антитело D2E7, продуцированное с использованием этой sORF-конструкции, имело такую же аффинность связывания с TNFα, как и антитело D2E7, продуцированное стандартным промышленным методом.
Модификация сигнального пептида
Авторами настоящего изобретения было продемонстрировано, что в модели sORF-конструкции “тяжелая цепь - интеин - легкая цепь” уровень секреции антитела был примерно в 10 раз выше в том случае, когда гидрофобность последовательности сигнального пептида легкой цепи была уменьшена в результате сайт-направленного мутагенеза.
Авторами настоящего изобретения была создана конструкция H, в которой последовательность сигнального пептида легкой цепи, то есть “MDMRVPAQLLGLLLLWFPGSRC" (SEQ ID NO:23), расположенная за последовательностью интеина P. horikoshi, была заменена последовательностью "MDMRVPAQLLGDEWFPGSRC" (SEQ ID NO:24). В эксперименте по трансфекции аналогичного типа, описанном выше, супернатант клеток, которые экспрессируют эту конструкцию, содержал 2047+116 нг/мл антитела, как было измерено с помощью ELISA-анализа. Этот уровень секреции антитела аналогичен уровню секреции антитела, полученного с применением 2A-технологии (1,6 мкг/мл). Вестерн-блот-анализ этого супернатанта показал яркие полосы, соответствующие тяжелой и легкой цепи антитела.
В контрольном эксперименте в вектор, экспрессирующий это антитело, была введена такая же мутация сигнального пептида легкой цепи с применением стандартного метода (то есть экспрессии тяжелой и легкой цепей антитела из двух отдельных открытых рамок считывания в двух отдельных векторах). В этой конструкции замена SEQ ID NO:23 на SEQ ID NO:24 приводила к предотвращению секреции антитела, как это и предполагалось, поскольку в конструкции, полученной стандартным способом, гидрофобная область играет важную роль в доставке комплекса частиц узнавания сигнала (SRP) в ЭP и в направлении его транспорта в транслокон. Было установлено, что в экспериментальной sORF-конструкции функция нацеливания сигнального пептида легкой цепи не имеет важного значения, даже если он может узнаваться и отщепляться сигнальной пептидазой в ЭP, и это подтверждает гипотезу, согласно которой вся ORF транспортируется в ЭP под действием сигнального пептида тяжелой цепи, присутствующего в начале ORF.
Антитело D2E7, секретированное с использованием sORF-конструкции H, очищали с помощью аффинной хроматографии на белке А и анализировали N-концевую последовательность легкой цепи. N-концевой пептидной последовательностью легкой цепи является последовательность MDMRVPAQLL (SEQ ID NO:26) (однозначно), которая представляет собой неотщепленный сигнальный пептид. Хотя в литературе высказывается предположение, что функция области H сигнального пептида ЭP млекопитающего состоит, главным образом, в нацеливании на комплекс (SRP) и в направлении его транспорта посредством транслокона, однако данные, полученные авторами настоящего изобретения, позволяют предположить, что гидрофобная (Н) область сигнального пептида также играет определенную роль в узнавании и расщеплении сигнальной пептидазой.
Авторами настоящего изобретения было продемонстрировано, что антитела D2E7, секретируемые с использованием конструкции pTT3-HcintLC-p.hori и конструкции H, являются биологически активными в клеточных анализах. Антитело D2E7, продуцированное с использованием конструкции pTT3-HcintLC-p.hori и конструкции Н, очищали и тестировали на его способность нейтрализовать TNFα-индуцированную цитотоксичность в клетках L929. Этот анализ проводили, в основном, как описано в патенте США 6090382 (см. пример 4 в этом патенте). Человеческий рекомбинантный TNFα является цитотоксичным в мышиных клетках L929 и был использован в данном анализе. Анти-TNFα антитело, как и антитело D2E7, может нейтрализовать такую цитотоксичность, а поэтому анализ с использованием клеток L929 является одним из клеточных анализов, который может быть применен для оценки биологической активности конкретного препарата антитела D2E7. Этот анализ показал, что антитело D2E7, продуцированное с помощью конструкции pTT3-HcintLC-p.hori и конструкции Н, нейтрализовало TNFα-индуцированную цитотоксичность. Величины IC50 для этого антитела были аналогичны соответствующим величинам для антитела D2E7, продуцированного стандартным промышленным методом.
Авторами настоящего изобретения были проведены исследования дополнительных конструкций с различными структурами в области сигнального пептида легкой цепи. Для идентификации оптимальной структуры sORF-конструкции, которая обеспечивала бы высокоэффективную секрецию антитела, авторами настоящего изобретения было создано несколько дополнительных конструкций, которые отличались своими участками в области, расположенной рядом с сайтом С-концевого сплайсинга и следующего за ним сигнального пептида. Конструкция J вместо последовательности “MDMRVPAQLLGDEWFPGSRC" (SEQ ID NO:24), присутствующей в конструкции Н, имела последовательность “MDMRVPAQWFPGSRC" (SEQ ID NO:25), следующую за последним остатком N интеина, причем внутри этого сигнального пептида гидрофобная область затем удалялась, тогда как С-концевая область, а также сайт расщепления сигнальной пептидазой сохранялись. Конструкция K направляла экспрессию зрелой последовательности легкой цепи, следующей непосредственно за последним остатком N интеина. Конструкция L вместо последовательности “MDMRVPAQLLGLLLLWFPGSRC" (SEQ ID NO:23), присутствующей в конструкции pTT3-HcintLC-p.hori, направляла экспрессию последовательности “MDMRVPAQLLGLLLLWFPGSGG" (SEQ ID NO:27), следующей за последним остатком N интеина, в которой перед сайтом расщепления сигнальной пептидазой были заменены аминокислоты в положении -1 и -2.
В данном эксперименте эти конструкции были введены в клетки 293E посредством временных трансфекций, и через 7 дней, культивированные супернатанты анализировали на титры антител IgG с помощью ELISA. Титры антител для конструкций H, J, K и L составляли 2328,5+79,9, 1289,7+129,6, 139,3+4,7, и 625,0+20,6 нг/мл (среднее + ср.кв.от.) соответственно.
Образцы клеточного осадка после таких трансфекций также анализировали с помощью вестерн-блот-анализа. Все конструкции имели полосу трехкомпонентного полипептида (~130 кДа), полосу тяжелой цепи (~50 кДа) и полосу легкой цепи (~25 кДа), описанные выше, но ни одна из них не обнаруживала детектируемого продукта сплайсинга (80 кДа и продукта, распознаваемого антителом против тяжелой цепи и легкой цепи). В этой группе конструкций конструкция К продуцировала наиболее характерный паттерн вестерн-блота (WB) и отличалась тем, что она продуцировала лишь очень небольшое количество внутриклеточных легких цепей, а вместо этого продуцировала молекулы белка, соответствующего гибриду “интеин - легкая цепь”, то есть продукту одного события расщепления на границе N-концевого сплайсинга. Белок такого типа отсутствовал в других конструкциях этой группы. Конструкция K отличалась от других конструкций по двум признакам: она не имела сайта расщепления сигнальной пептидазой и имела аспарагиновую кислоту вместо метионина или серина в положении 1-го аминокислотного остатка С-концевого экстеина. Оба этих признака или любой из них могут способствовать предотвращению расщепления в области между интеином и легкой цепью антитела, что может приводить к снижению уровня секреции антитела.
Антитело D2E7, секретированное с использованием sORF-конструкций J и L, очищали с помощью аффинной хроматографии на белке А и анализировали на N-концевые последовательности их легких цепей. Этот анализ показал, что такой N-концевой пептидной последовательностью легкой цепи, продуцированной с использованием конструкции J, была последовательность MDMRVPAQLL, которая представляет собой неотщепленный сигнальный пептид, а N-концевой пептидной последовательностью легкой цепи, продуцированной с использованием конструкции L, была последовательность DIQMTQSPSS, которая представляет собой зрелую легкую цепь, образованную в результате “правильного” отщепления сигнального пептида. Поэтому по сравнению с конструкцией pTT3-HcintLC-p.hori конструкция L имеет структуру, которая способствует повышению уровня секреции антитела (0,6 - 1 мкг/мл при различных временных трансфекциях), а его легкая цепь также имеет нужную N-концевую последовательность.
Авторами настоящего изобретения были разработаны механизмы экспрессии подвергнутого сборке антитела из sORF-конструкций с использованием интеинов и с применением методов дополнительного увеличения уровней секреции антител. Внутриклеточные образцы, взятые из клеток, трансфицированных большинством описанных sORF-конструкций, содержали две молекулы легкой цепи антитела, соответствующие непроцессированным и процессированным легким цепям. В клетке, трансфицированной любой из конструкций позитивного контроля или конструкцией pTT3-HcintLC-p.hori, секретировалась только процессированная легкая цепь, что указывало на то, что непроцессированные легкие цепи, которые имели связанные сигнальные пептиды легкой цепи дикого типа, не могли подвергаться сборке и секреции. В противоположность этому непроцессированные легкие цепи конструкций H и J обладали способностью к сборке и секреции, и обе они имели мутированные сигнальные пептиды. Степень процессинга сигнального пептида легкой цепи, определяемая по распределению внутриклеточного полипептида легкой цепи в непроцессированных и процессированных формах, варьировалась в зависимости от конкретной конструкции. По сравнению с конструкцией pTT3-HcintLC-p.hori конструкция L имела повышенное количество процессированных легких цепей, что приводило к увеличению уровня секреции антител.
На основании вышеописанных экспериментальных данных можно утверждать, что одним из способов увеличения уровня секреции антитела с использованием sORF-конструкций является повышение эффективности процессинга сигнального пептида легкой цепи. Это может быть достигнуто путем систематического анализа мутаций в гидрофобной области, а также в области, находящейся вблизи от сайта расщепления, и путем анализа сигнальных пептидов различной длины. Это может быть также осуществлено путем скрининга дрожжей на пептидные последовательности, которые могут эффективно расщепляться при таком представлении, и путем аналогичного скрининга в клетках CHO.
Другим методом, который может быть применен для увеличения уровней секреции антител с использованием sORF-конструкций, является анализ различных 5'- и 3'-нетранслируемых областей (UTR) на повышение стабильности трехкомпонентной мРНК, поскольку такие мРНК являются более крупными, чем традиционные мРНК, кодирующие тяжелую и легкую цепи антител по отдельности.
Другим методом повышения уровней секреции антител с использованием sORF-конструкций является генерирование и отбор стабильных клеточных линий CHO или NSO и амплификация рекомбинантных генов с использованием DHFR или GS для увеличения числа копий этих генов. Уровень секреции антител независимо увеличивается при изменении эписомной (временной) локализации рекомбинантных генов на геномную (стабильную) локализацию. Такой уровень секреции также увеличивается с увеличением числа копий и/или в результате модификаций в 5' и 3' UTR в промоторных и в энхансерных последовательностях. Векторы, экспрессирующие дигидрофолат-редуктазу (dhfr), трансфицировали в dhfr-дефицитные клеточные линии. Клеточные линии с большим числом копий векторов отбирали с использованием метотрексата, конкурентного ингибитора dhfr (Kaufman, R.J. & Sharp, P.A. J Mol. Biol. (1982) 159:601-621). В качестве другой независимой альтернативы для увеличения уровня экспрессии используются экспрессирующие векторы, несущие энхансер промотора цитомегаловируса в комбинации с селективным маркером глутамин-синтетазой (Bebbington, CR. (1991) Methods 2:138-145). Помимо увеличения числа копий рекомбинантных генов этот способ также предусматривает отбор клеточных линий дифференцировки, которые являются особенно подходящими для осуществления процессинга с использованием моделей sORF-конструкций.
Использование модифицированных интеинов, содержащих вставки
В целях прослеживания дальнейшего пути внутриклеточных белков интеинов, которые были отделены от полипептидов тяжелой и легкой цепи D2E7, авторами настоящего изобретения были получены 4 конструкции, в которые была введена гистидиновая метка в положения аминокислотной последовательности FRKVR!RGRG (! означает положения инсерции, - HT1) и EGKR!IPEF (-HT2), в обеих конструкциях pTT3-LcintHC-p.hori и в конструкции H. Была выдвинута гипотеза, что эти 2 положения в интеине P.horikoshi представляют собой петли, которые могут быть толерантными к вставкам и при этом сохранять свою 3-мерную структуру, а следовательно и свои функции. В одном эксперименте после инкубирования в течение 4 дней, проводимого после трансфекции клеток 293E, супернатанты культуры анализировали на титры антител IgG с помощью ELISA. Титры антител для конструкций pTT3-LcintHC-p.hori-HT1, pTT3-LcintHC-p.hori-HT2, конструкции H-HT1, конструкции H-HT2 и конструкции H составляли 78,3+3,2, 67,3+0,6, 663,0+15,5, 402,7+5,5, 747,0+22,5 нг/мл (среднее + ср.кв.от.) соответственно. Использование интеина P.horikoshii, имеющего вставки в обоих 2 положениях, позволяет секретировать подвергнутое сборке антитело. В частности, использование интеина, имеющего внутренюю метку, введенную в положение 1, давало такой же уровень секреции антитела, как и использование интеина, не содержащего каких-либо вставок.
Представленные выше данные показали, что разработка sORF-конструкций настоящего изобретения включает в себя использование модифицированных интеинов, содержащих внутреннюю метку. Специалистам известны различные метки. Метками согласно изобретению являются, но не ограничиваются ими, флуоресцентные метки и хемилюминесцентные метки. С использованием таких конструкций может быть проведен мониторинг количества экспрессируемого полипротеина с помощью флуоресцентной детекции в отдельных клетках. Кроме того, эти клетки могут быть отсортированы по уровню экспрессии белка с помощью FACS. Использование таких меток является особенно подходящим для получении генераций стабильных клеточных линий, поскольку оно позволяет проводить отбор клеток или клеточных линий с высокой продуцирующей способностью с помощью FACS-анализа. Как описано в настоящем изобретении, полноразмерные интеины наблюдались в клеточном лизате после их самоотщепления от фланкирующих тяжелой и легкой цепей антитела. На основе этого могут быть детектированы флуоресцентно меченные интеины, которые могут быть использованы для продуцирования стабильной клеточной линии. Метки могут быть также использованы для очистки белков.
Исходя из описанных выше данных авторами настоящего изобретения был сделан вывод, что реакция сплайсинга белка, опосредуемая интеином Pol I P. horikoshi, может происходить в клетках 293E, в ЭP и в аминокислотных последовательностях тяжелой и легкой цепей антитела (в частности, антитела D2E7, взятого в качестве примера). Мутации типа точковых замен, такие как S → M, в положении первой аминокислоты С-концевого экстеина, и Н → А в положении предпоследней аминокислоты интеина приводят к увеличению уровней секреции антител. Снижение гидрофобности области H сигнального пептида легкой цепи, например, в конструкциях H и J приводит к еще большему увеличению уровней секреции антител. Уровень секреции антител в конструкции, не содержащей сигнального пептида легкой цепи, является относительно низким, что, вероятно, обусловлено менее эффективным расщеплением на границе С-концевого сплайсинга. Для повышения эффективности такого расщепления применяются два метода. Первый метод включает в себя введение аминокислоты, не являющейся аспарагиновой кислотой, в положение +1. В некоторых описанных здесь конструкциях в положение +1 был также введен метионин, что приводило к эффективному расщеплению на границе С-концевого сплайсинга. Вторым методом повышения эффективности такого расщепления является изменение пространства между С-концевым сайтом расщепления и глобулярной структурой легкой цепи с помощью линкера, за которым находится, но необязательно, сайт расщепления другого типа, например сайт, описанный в настоящей заявке.
Хотя были описаны и протестированы различные конструкции, содержащие интеин P. horikoshii и антитело DE27, однако в sORF-конструкциях согласно изобретению могут быть использованы и другие интеины и интеин-подобные белки (включая белки семейства hedgehog и родственного семейства), например встроенные между тяжелой и легкой цепями антитела. Белки тяжелой и легкой цепей антитела могут быть также заменены другими мультисубъединичными белками (включая белки, состоящие из двух субъединиц, и белки, имеющие более чем две субъединицы).
Помимо конструкций с интеином Poll P. horikoshii, описанных выше, авторами настоящего изобретения были сконструированы аналогичные конструкции с использованием интеина Sce.VMA и мини-интеина dnaE Ssp.: pTT3-Hc-VMAint-LC-0aa, pTT3-Hc-VMAint-LC-1aa, pTT3-Hc-VMAint-LC-3aa, pTT3-Hc-Ssp-GA-int-LC-0aa, pTT3-Hc-Ssp-GA-int-LC-1aa и pTT3-Hc-Ssp-GA-int-LC-3aa. Эти конструкции были перенесены в клетки 293E, а затем были проанализированы супернатанты и образцы клеточного осадка.
В одном из экспериментов после 7-дневного инкубирования, проводимого после трансфекции клеток 293E, супернатанты культур анализировали на титры антител IgG с помощью ELISA-анализа. Титры антител для конструкций pTT3-Hc-VMAint-LC-0aa, pTT3-Hc-VMAint-LC-1aa, pTT3-Hc-VMAint-LC-3aa, pTT3-HC-Ssp-GA-int-LC-0aa, pTT3-HC-Ssp-GA-int-LC-1aa и pTT3-HC-Ssp-GA-int-LC-3aa составляли 9,0 ± 3,5, 12,0 ± 0,0, 39,7 ± 1,2, 90,0 ± 2,0, 38,7 ± 1,5 и 32 ± 2,6 нг/мл (среднее ± ср. кв.от.) соответственно.
Образцы клеточного осадка после этих трансфекций также анализировали с помощью вестерн-блот-анализа. Во всех этих образцах наблюдались трехкомпонентные полипептиды. Кроме того, полипептид тяжелой цепи наблюдался в конструкциях pTT3-Hc-VMAint-LC-0aa, pTT3-HC-Ssp-GA-int-LC-0aa, pTT3-HC-Ssp-GA-int-LC-1aa и pTT3-HC-Ssp-GA-int-LC-3aa, а полипептид легкой цепи наблюдался в pTT3-HC-Ssp-GA-int-LC-0aa, pTT3-HC-Ssp-GA-int-LC-1aa и pTT3-HC-Ssp-GA- int-LC-3aa.
Результаты этих экспериментов показали, что интеины как класс белков могут быть с успехом использованы в стратегиях экспрессии белка sORF, как уже указывалось авторами настоящего изобретения. Кроме того, в конструкциях, аналогичных описанным здесь конструкциям, помимо интеина могут быть также использованы бактериальные интеин-подобные (BIL) домены и hedgehog-домены (Hog) аутопроцессинга, то есть другие 2 члена суперсемейства Hog/интеинов (HINT).
Кроме того, поскольку эндонуклеазные области, которые присутствуют во многих интеинах, включая интеин Poll P. horikoshii и интеин Sce.VMA, не используются в стратегии экспрессии генов согласно изобретению, то такой эндонуклеазный домен может быть делетирован и заменен небольшим линкером с получением “мини-интеинов”.
Эти сконструированные мини-интеины могут быть также использованы для получения описанных конструкций, и они имеют то преимущество, что кодирующая область интеина является значительно меньшей по размеру и позволяет использовать более крупную последовательность, кодирующую представляющие интерес полипептиды, и/или значительно облегчать манипуляции с рекомбинантными молекулами ДНК.
Одной из проблем, связанных с использованием пептидов аутопроцессинга, таких как последовательности 2А или 2А-подобные последовательности или последовательности узнавания протеазой, является то, что C- или N-концы одной или нескольких цепей полипептида содержат аминокислоты, происходящие от пептида аутопроцессинга, то есть аминокислотные остатки, происходящие от 2А или от последовательности узнавания протеазой, в зависимости от положения расщепления и соответствующего положения конкретной цепи в первичном продукте трансляции. Эти аминокислотные остатки являются “чужеродными” для хозяина и могут вырабатывать иммунный ответ при экспрессии или доставке in vivo рекомбинантного белка (то есть при его экспрессии из вирусного или невирусного вектора, используемого в генотерапии, или при его введении в качестве in vitro продуцируемого рекомбинантного белка). Кроме того, аминокислотные остатки, происходящие от 2А или от последовательности узнавания протеазой, если они не удаляются, могут препятствовать секреции белка в клетках-продуцентах и/или изменять конформацию белка, что будет приводить к продуцированию неоптимального уровня экспрессии и/или к снижению биологической активности рекомбинантного белка.
Экспрессионные генные конструкции сконструированы так, чтобы они содержали дополнительный сайт протеолитического расщепления между полипептид-кодирующей последовательностью и сайтом аутопроцессинга (то есть 2А-последовательность) или другим сайтом расщепления протеазой, где указанный дополнительный сайт протеолитического расщепления может быть использован в настоящем изобретении в качестве средства для удаления аминокислотных остатков сайта аутопроцессинга, оставшихся после расщепления.
Примерами дополнительных сайтов протеолитического расщепления являются сайт расщепления фурином с консенсусной последовательностью RXK(R)R (SEQ ID NO:1), которая может быть отщеплена эндогенными субтилизин-подобными протеазами, такими как фурин и другие сериновые протеазы, действующие на пути секреции белка. В публикации заявки на патент США 2005/0042721 указывается, что остатки 2A у N-конца первого белка могут быть эффективно удалены путем введения сайта расщепления фурином RAKR между первым полипептидом и последовательностью 2A. Кроме того, было показано, что использование плазмиды, содержащей последовательность 2A и сайт расщепления фурином, смежный с сайтом 2A, приводит к увеличению уровня экспрессии белка по сравнению с плазмидой, содержащей лишь одну последовательность 2А. Такое увеличение уровня экспрессии имеет то преимущество, что благодаря удалению остатков 2А из N-конца данного белка могут быть использованы более длинные последовательности 2А или 2А-подобные последовательности или другие последовательности аутопроцессинга. Эти более длинные последовательности аутопроцессинга, такие как последовательности 2A или 2A-подобные последовательности, могут еще больше облегчать эквимолярную экспрессию двух или более полипептидов под контролем одного промотора. Еще большее увеличение экспрессии иммуноглобулина достигается, в том случае если в данном полипротеине присутствуют две кодируемые последовательности легкой цепи иммуноглобулина и только одна кодируемая последовательность тяжелой цепи иммуноглобулина.
При этом предпочтительно использовать антитела или их аналоги со всеми свойствами человеческих антител. Использование этих реагентов позволяет избежать продуцирования нежелательных иммунных ответов, индуцируемых антителами или их аналогами, происходящими от нечеловеческих антител. Во избежание продуцирования иммунных ответов хозяина на аминокислотные остатки, происходящие от пептидов аутопроцессинга, последовательность, кодирующая сайт протеолитического расщепления, может быть встроена (с применением стандартной технологии, известной специалистам) между последовательностью, кодирующей первый белок, и последовательностью, кодирующей пептид аутопроцессинга, так чтобы пептидная последовательность аутопроцессинга была удалена из экспрессированного полипептида, то есть из антитела. Такая стратегия является особенно подходящей при использовании терапевтических и диагностических антител in vivo.
Для осуществления настоящего изобретения может быть использован любой известный дополнительный сайт протеолитического расщепления, который может быть экспрессирован с помощью рекомбинантных ДНК-векторов. Примерами дополнительных сайтов протеолитического расщепления, которые могут быть встроены между последовательностью, кодирующей полипептид или белок, и последовательностью аутопроцессинга (такой как последовательность 2А), являются, но не ограничиваются ими, сайт расщепления фурином, RXK(R)R (SEQ ID NO:1); сайт расщепления фактором Xa, IE(D)GR (SEQ ID NO:6); сайт расщепления сигнальной пептидазой I, например, LAGFATVAQA (SEQ ID NO:28); и сайт расщепления тромбином, LVPRGS (SEQ ID NO:7).
В настоящем изобретении в качестве альтернативных методов, в которых для экспрессии более чем одного зрелого белка из одной открытой рамки считывания используются IRES, фурин, 2А и интеин, также рассматривается процессинг белка с использованием домена белка hedgehog, расположенного в полипротеине между частями первого и второго белков. Авторами настоящем изобретении была сконструирована одна открытая рамка считывания для осуществления экспрессии тяжелой и легкой цепи антитела с hedgehog-доменом аутопроцессинга в целях разделения генов тяжелой и легкой цепей антитела. В клетках, несущих такую ORF, была транскрибирована одна мРНК, которая состоит по меньшей мере из одной тяжелой цепи антитела, одной легкой цепи антитела и одного hedgehog-домена аутопроцессинга, и такая мРНК была использована для продуцирования соответствующего полипротеина. После трансляции hedgehog-домен аутопроцессинга опосредует разделение тяжелой и легкой цепей антитела.
Семейство белков hedgehog содержит консервативные сигнальные молекулы, которые действуют как морфогены в различных системах развития и ответственны за развитие у человека заболеваний широкого ряда (Kalderon, D. 2005. Biochem Soc Trans. Dec;33(Pt6):1509-12). Белки hedgehog имеют 2 структурных домена, N-концевой домен (Hh-N), который участвует в передаче сигналов в клетках, и C-концевой домен (Hh-C), который катализирует событие посттрансляционного аутопроцессинга, приводящее к отщеплению белка между этими двумя доменами, к присоединению холестериновой молекулы к С-концу N-концевого домена и, тем самым, к активации молекулы, участвующей в передаче сигнала (Traci et al. 1997. Cell, 91, 85-97).
Преимуществами предлагаемой технологии экспрессии sORF-антитела является возможность изменения отношения количества генов тяжелой и легкой цепей, близкая локализация полипептидов тяжелой и легкой цепей, способствующая их сборке в мульти-субъединицы в ЭP, и возможность осуществления высокоэффективной секреции белка.
Hh-C-домены белка могут быть использованы для катализа реакции аутопроцессинга в ЭP, которая приводит к посттрансляционному расщеплению в положении между полипептидом тяжелой цепи и полипептидом Hh-C антитела в описанной ниже экспериментальной конструкции с одной открытой рамкой считывания.
Белки семейства hedgehog имеют N-концевой сигнальный домен и C-концевой домен аутопроцессинга. С-концевые домены аутопроцессинга этих белков сами отщепляются от N-концевых доменов и присоединяют к своим С-концам холестериновую молекулу посредством 2-стадийной реакции (Porter et al. 1996. Science. 274(5285):255-9). Помимо холестерина аутопроцессинг также стимулируют и другие нуклеофилы, такие как DTT или глутатион (Lee et al. 1994. Science, 266, 1528-1537). Поскольку реакция расщепления катализируется С-концевым доменом аутопроцессинга, то аналогичная реакция расщепления происходит также в том случае, когда N-концевой сигнальный домен белка hedgehog заменяют полипептидом тяжелой или легкой цепи антитела. Такая реакция может быть использована для разделения тяжелой и легкой цепей антитела, присутствующих в полипротеине, кодируемом одной открытой рамкой считывания.
Сначала проводят тест на экспрессию антитела в системе временной экспрессии и для этой цели создают конструкцию на основе вектора PTT3. Этот вектор имеет ориджин репликации EBV, который позволяет осуществлять эписомную амплификацию этого вектора в трансфицированных клетках 293E (в клетках, которые экспрессируют ядерный антиген 1 вируса Эпштейна-Барра) в суспензионной культуре (Durocher et al. 2002). Каждый вектор имеет одну открытую рамку считывания под контролем промотора CMV. В одном из экспериментов по созданию конструкции pTT3-HC-Hh-C25-LC весь С-концевой домен белка sonic hedgehog Drosophila melanogaster встраивали с сохранением рамки считывания в положение между тяжелой и легкой цепями антитела D2E7, каждая из которых имеет сигнальный пептид (SP). Эти конструкции вводили в клетки 293E посредством временной трансфекции. Затем были проанализированы супернатанты культуры и образец клеточного осадка.
Образцы клеточного осадка лизировали в условиях, благоприятствующих разделению цитозольных и внутриклеточных мембранных фракций. Обе эти фракции анализировали методами иммуноблоттинга с использованием антител против тяжелой цепи или антител против легкой цепи каппа. На этих блотах наблюдались молекулы белка, а именно полипротеин (HC-Hh-C25-LC), Hh-C25-LC и отдельные тяжелые цепи (HC) и легкие цепи (LC). Присутствие последних 3 молекул белка указывало на реакцию аутопроцессинга. Свободная тяжелая цепь образовывалась в результате расщепления, катализируемого Hh-C-доменом белка, а свободные полипептиды легкой цепи образовывалась в результате расщепления сигнальной пептидазой. Сегрегация молекул белка в субклеточной мембранной фракции, которая содержала эндоплазматический ретикулум (ЭP), дает основание предположить, что сигнальный пептид тяжелой цепи в начале ORF согласно изобретению направляет всю ORF в ЭP, где происходит реакция расщепления.
Из образцов клеточного осадка была также экстрагирована вся РНК, после чего проводили Нозерн-блот-анализ с помощью зонда для тяжелой цепи антитела и зонда для легкой цепи антитела. На этих Нозерн-блотах наблюдалась трехкомпонентная мРНК, которая была гибридизована с зондом тяжелой цепи и с зондом легкой цепи, что подтвержадет sORF-природу этой экспериментальной конструкции. В противоположность этому, в образцах клеточного осадка, в которых была осуществлена экспрессия антитела D2E7 с применением стандартного метода, а именно путем введения тяжелой и легкой цепей антитела от двух отдельных ORF, присутствующих в двух векторах pTT3, мРНК тяжелой цепи (1,4 т.п.н.) и легкой цепи (0,7 т.п.н.) были детектированы с использованием зондов для тяжелой цепи или для легкой цепи соответственно.
Эти эксперименты продемонстрировали, что использование конструкций, содержащих одну ORF (тяжелая цепь D2E7 - Hh-C25- легкая цепь D2E7), приводит к транскрипции одной мРНК, содержащей все 3 белка. Этот трехкомпонентный транскрипт был транслирован в трехкомпонентый полипептид, который в процессе котрансляции транспортировался в ЭP под действием сигнального пептида тяжелой цепи, присутствующего в начале ORF. Это указывает на то, что Hh-C-домен белка может быть использован для экспрессии антител, а также других мульти-субъединичных секретируемых белков и/или других белков, которые для их укладки и соответствующей посттрансляционной модификации должны направляться по секреторному пути.
Помимо клеточных осадков были проанализированы супернатанты культур с помощью Вестерн-блот-анализа и ELISA на секретированные антитела, обсуждаемые в настоящей заявке. Конструкции, полученные с использованием делетированного hh-C25, могут быть протестированы для сравнения эффективности процессинга полипротеинов и уровня секреции антитела.
Было показано, что делеция C-концевой аминокислоты 63 в Hh-C25-домене белка приводит к образованию Hh-C17-домена белка, который может катализировать процессинг белка, но не присоединение холестерина. Hh-C17 также экспрессировался как рекомбинантный белок, и была определена его кристаллическая структура (Traci et al. 1997. см. выше). Поэтому в другой эспериментальной конструкции, pTT3-HC-C17-LC, такой усеченный домен белка был встроен между тяжелой и легкой цепями антитела D2E7.
При выравнивания в целях определения гомологии белков hedgehog и интеинов, которые были проанализированы авторами настоящего изобретения в аналогичных экспериментальных конструкциях, подробно описанных в настоящей заявке, предсказанную концевую β-складчатую вторичную структуру удлиняли на 8 аминокислот, которые могут оказывать, а могут и не оказывать, влияние на эффективность аутопроцессинга. Поэтому также была проанализирована дополнительная конструкция pTT3-HC-C17sc-LC.
Эти конструкции вводили в клетки 293E посредством временной трансфекции, и через 7 дней супернатанты культуры могут быть проанализированы на титры антител IgG с помощью ELISA. Титры антител для pTT3-HC-C25-LC, pTT3-HC-C17-LC, pTT3-HC-C17sc-LC и pTT3-HC-C17hn-LC составляли 0,038, 0,042, 0,040 и 0,046 мкг/мл соответственно.
Образцы этих супернатантов также анализировали с помощью электрофореза в ДСН-ПААГ (в денатурирующих условиях) и подвергали блоттингу с использованием антитела, специфичного для тяжелой цепи человеческого IgG, и антитела, специфичного для человеческой легкой цепи каппа. На этих вестерн-блотах могут наблюдаться белки тяжелой цепи антитела (~50 кДа) и легкой цепи антитела (~25 кДа), уровни которых коррелируют с уровнями IgG, измеренными с помощью ELISA.
Образцы клеточного осадка после указанных трансфекций также анализировали с помощью вестерн-блот-анализа. Присутствие и относительная плотность описанных белков четырех видов для различных конструкций могут быть также подвергнуты сравнению в целях определения эффективности процессинга белка в каждой из экспериментальных конструкций.
В белках, принадлежащих к другому классу и подвергающихся аутопроцессингу, то есть в интеинах, последними двумя аминокислотами часто являются HisAsn. В процессе сплайсинга белка, катализируемого интеинами, Asn подвергается циклизации с участием His, что приводит к расщеплению пептидной связи между интеином и его С-концевым фланкирующим полипептидом. В отличие от интеинов самопроцессирующиеся белки hedgehog по своей природе не имеют С-концевого фланкирующего полипептида и не имеют консервативного Asn в этом положении полипептида. В одной из экспериментальных конструкций pTT3-HC-C17hn-LC в это положение вместо Ser-Cys авторами был введен His-Asn. Не ограничиваясь какой-либо теорией, следует лишь отметить, что сайт расщепления, созданный в данном положении, позволяет осуществлять более эффективное разделение самопроцессирующегося белка hedgehog и легкой цепи антитела в этой конкретной экспериментальной конструкции. Эффективность секреции антитела оценивали, как описано выше.
Были охарактеризованы антитела, продуцируемые с использованием sORF-конструкций, содержащих самопроцессирующийся белок hedgehog. Антитело D2E7, секретированное с использованием вышеупомянутой sORF-конструкции, очищали с помощью аффинной хроматографии на белке А и анализировали на N-концевые последовательности тяжелой и легкой цепей. Эти очищенные антитела, а также антитело D2E7, полученное стандартным промышленным методом, анализировали с помощью масс-спектрометрии, как описано выше, в денатурирующих условиях. С помощью масс-спектрометрии в нативных условиях определяли молекулярную массу (MW) интактного антитела D2E7, продуцированного с использованием указанных конструкций, а также антитела D2E7, продуцированного промышленным методом.
Уровень связывания антитела D2E7 с человеческим TNFα анализировали на приборе Biacore, как описано выше. Кинетическую скорость ассоциации, кинетическую скорость диссоциации и общую аффинность определяли с использованием различных концентраций TNFα в пределах 1-100 нМ.
В настоящем изобретении рассматривается применение любых векторов, используемых для введения конструкций, содержащих последовательность, кодирующую два или более полипептида или белка, и сайт аутопроцессинга в клетки. Специалистам известно множество примеров векторов для экспрессии генов, которые могут иметь вирусное или невирусное происхождение. Векторами в методах доставки невирусных генов, которые могут быть использованы для осуществления настоящего изобретения, могут служить, но не ограничиваются ими, плазмиды, липосомы, комплексы “нуклеиновая кислота/липосома”, катионные липиды и т.п.
Вирусные векторы
Вирусные и другие векторы могут эффективно трансдуцировать клетки и вводить свою собственную ДНК в клетку-хозяина. При генерировании рекомбинантных вирусных векторов гены, не играющие важной роли, заменяют экспрессируемыми последовательностями, кодирующими представляющие интерес белки или полипептиды. Репрезентативными векторами являются, но не ограничиваются ими, вирусные и невирусные векторы, такие как ретровирусный вектор (включая лентивирусные векторы), аденовирусные векторы (Ad), включая компетентные по репликации вирусы, дефектные по репликации вирусы и их аттенюированные формы; векторы на основе аденоассоциированных вирусов (AAV), векторы на основе обезьяньего вируса 40 (SV-40), векторы на основе бычьего папиломавируса, векторы на основе вируса Эпштейна-Барра, герпесвирусные векторы, векторы на основе вируса коровьей оспы, векторы на основе вируса мышиного лейкоза Молони, векторы на основе вируса мышиной саркомы Харви, векторы на основе вируса опухоли мышиной молочной железы, векторы на основе вируса саркомы Рауса и невирусные плазмиды. Бакуловирусные векторы хорошо известны специалистам и являются подходящими для экспрессии в клетках насекомых. Специалистам также хорошо известно множество векторов, подходящих для экспрессии в клетках млекопитающих или в других эукариотических клетках, и многие из них являются коммерчески доступными. Коммерческими источниками таких векторов являются, но не ограничиваются ими, Stratagene, La JoIIa, CA; Invitrogen, Carlsbad, CA; Promega, Madison, Wl и Sigma-Aldrich, St. Louis, MO. Последовательности многих векторов могут быть взяты из Genbank, а дополнительную информацию об этих векторах можно найти в Интернете в Riken BioSource Center.
Вектор обычно содержит ориджин репликации, и такой вектор может дополнительно содержать, а может и не содержать “маркер” или “селективный маркер”, с помощью которого данный вектор может быть идентифицирован и отобран. Хотя может быть использован любой селективный маркер, однако в рекомбинантных векторах обычно используются известные селективные маркеры, и выбор селективных маркеров зависит от клетки-хозяина. Примерами генов селективных маркеров, кодирующих белки, сообщающие резистентность к антибиотикам или к другим токсинам, являются, но не ограничиваются ими, гены резистентности к ампициллину, метотрексату, тетрациклину, неомицину (Southern et al. 1982. J Mol Appl Genet. 1:327-41), микофеноловой кислоте (Mulligan et al. 1980. Science 209:1422-7), пуромицину, зеомицину, гигромицину (Sugden et al. 1985. Mol Cell Biol. 5:410-3), дигидрофолат-редуктазе, глутамин-синтетазе и G418. Как известно специалистам, экспрессирующие векторы обычно включают в себя ориджин репликации, промотор, функционально присоединенный к кодирующей последовательности или к экспрессируемым последовательностям, а также сайты связывания с рибосомой, сайты РНК-сплайсинга, сайт полиаденилирования и последовательности терминации транскрипции, если это необходимо для экспрессии кодирующей(их) последовательности(ей).
Если вектор или другие последовательности ДНК называются здесь “рекомбинантными”, то это означает лишь то, что эти последовательности ДНК представляют собой функционально присоединенные последовательности, которые в своей изолированной форме или в природной форме обычно не являются функционально присоединенными. Регуляторные (экспрессионные и/или регулирующие) последовательности функционально присоединены к кодирующей последовательности нуклеиновой кислоты, если эти экспрессионные и/или регуляторные последовательности регулируют транскрипцию и, при необходимости, трансляцию последовательности нуклеиновой кислоты. Такие экспрессионные и/или регуляторные последовательности могут включать промоторы, энхансеры, терминаторы транскрипции, старт-кодон (то есть ATG), расположенный со стороны 5'-конца от кодирующей последовательности, сигналы сплайсинга с вырезанием интронов и стоп-кодоны.
Известно, что аденовирусные векторы, используемые в генотерапии, обладают способностью к высокоэффективной временной экспрессии, имеют превосходные титры и обладают способностью трансдуцировать делящиеся и неделящиеся клетки in vivo (Hitt et al. 2000. Adv in Virus Res 55:479-505). Рекомбинантные Ad-векторы согласно изобретению содержат сайт упаковки, позволяющий вектору проникать в дефектные по репликации вирионы Ad; последовательность, кодирующую два или более представляющих интерес полипептидов или белков, например тяжелые и легкие цепи представляющего интерес иммуноглобулина; и последовательность, кодирующую сайт аутопроцессинга отдельно или в комбинации с дополнительным сайтом протеолитического расщепления. Другими элементами, необходимыми или полезными для включения в инфекционные вирионы, являются 5'- и 3'-ITR Ad; гены E2, части гена E4 и необязательно ген E3.
Дефектные по репликации Ad-вирионы, инкапсулирующие рекомбинантные Ad-векторы, получают стандартными методами, известными специалистам, с использованием Ad-упаковывающих клеток и с применением технологии упаковки. Примеры таких методов можно найти, например, в патенте США No 5872005. Последовательность, кодирующую два или более представляющих интерес полипептидов или белков, обычно встраивают в аденовирус в делетированную область E3 вирусного генома. Предпочтительные аденовирусные векторы, используемые для осуществления настоящего изобретения, не экспрессируют один или более генных продуктов Ad дикого типа, например E1a, E1b, E2, E3 и E4. Предпочтительными вариантами являются вирионы, которые обычно используются вместе с упаковывающими клеточными линиями, комплементирующими функции генных областей E1, E2A, E4 и, необязательно, E3. См., например, патенты США №№ 5872005, 5994106, 6133028 и 6127175.
Таким образом, используемые здесь термины “аденовирус” и “аденовирусная частица” означают сам вирус или его производные и включают в себя все его серотипы и подтипы, а также все природные и рекомбинантные формы, если это не оговорено особо. Такими аденовирусами могут быть аденовирусы дикого типа либо аденовирусы, которые могут быть модифицированы различными методами, известными специалистам или описанными в настоящей заявке. Такими модификациями являются модификации в геноме аденовируса, который упакован в частице, образующей инфекционный вирус. Такими модификациями являются делеции, известные специалистам, такие как делеции в одной или в кодирующих нескольких областях E1a, E1b, E2a, E2b, E3 или E4. Примерами упаковывающих клеток и клеток-продуцентов являются клетки, происходящие от клеток 293, A549 или HeLa. Аденовирусные векторы очищают и на их основе получают препараты стандартными методами, известными специалистам.
Аденоассоциированный вирус (AAV) представляет собой зависящий от вируса-помощника человеческий парвовирус, способный к латентному инфицированию клетки посредством его интеграции в хромосому клетки. Благодаря своей способности интегрироваться в хромосому и непатогенной природе AAV может быть с успехом использован в качестве вектора для генотерапии человека. Вирионы rAAV, используемые для осуществления настоящего изобретения, могут быть продуцированы стандартным методом, известным специалистам, и могут быть сконструированы так, чтобы они содержали в качестве функционально присоединенных компоненов по ходу транскрипции регуляторные последовательности, включая последовательности инициации и терминации транскрипции, и представляющую(ие) интерес кодирующую(ие) последовательность(и). Более конкретно, рекомбинантные AAV-векторы согласно изобретению содержат: сайт упаковки, позволяющий вектору проникать в дефектные по репликации вирионы AAV; последовательность, кодирующую два или более представляющих интерес полипептидов или белков, например тяжелую и легкую цепи представляющего интерес иммуноглобулина; последовательность, кодирующую сайт аутопроцессинга, локализованный отдельно или в комбинации с одним или несколькими дополнительными сайтами протеолитического расщепления. AAV-векторы, используемые для осуществления настоящего изобретения, конструируют так, чтобы они в качестве функционально присоединенных компонентов также содержали по ходу транскрипции регуляторные последовательности, включая последовательности инициации и терминации транскрипции. Эти компоненты фланкируются у 5'- и 3'-концов функциональными последовательностями ITR AAV. Термин “функциональные последовательности ITR AAV” означает, что последовательности ITR выполняют функцию “спасения”, репликации и упаковки вириона AAV.
Рекомбинантные AAV-векторы были также охарактеризованы на их способность регулировать экспрессию и продуцирование выбранных рекомбинантных полипептидных или белковых продуктов в клетках-мишенях. Таким образом, рекомбинантные векторы содержат по меньшей мере все последовательности AAV, играющие главную роль в инкапсидировании и в образовании физических структур для инфицирования рекомбинантных вирионов AAV (rAAV). Следовательно, ITR AAV, используемые в экспрессирующих векторах, необязательно должны иметь нуклеотидную последовательность дикого типа (например, как описано в работе Kotin, 1994. Hum. Gene Ther. 5:793-801), и они могут быть модифицированы путем инсерции, делеции или замены нуклеотидов, либо ITR AAV могут быть получены из любых различных серотипов AAV. Вообще говоря, AAV-вектором может быть любой вектор, происходящий от серотипа аденоассоциированного вируса, известного специалистам.
Обычно экспрессионный AAV-вектор вводят в клетку-продуцент с последующим введением конструкции вируса-помощника AAV, где указанная конструкция-помощник включает в себя кодирующие области AAV, способные экспрессироваться в указанной клетке-продуценте и комплементировать функции вируса-помощника AAV, отсутствующие в AAV-векторе. Такая конструкция-помощник может быть создана в целях ингибирования экспрессии крупных белков Rep (Rep78 и Rep68), обычно путем введения мутации в старт-кодон после p5, а именно замены кодона ATG на ACG, как описано в патенте США № 6548286, который вводится в настоящее описание посредством ссылки. После этого в клетку-продуцент вводят вирус-помощник и/или дополнительные векторы, где указанные вирус-помощник и/или дополнительные векторы обладают дополнительными функциями, способными поддерживать эффективное продуцирование вируса rAAV. Затем клетки-продуценты культивируют с продуцированием rAAV. Эти стадии осуществляют в соответствии со стандартной методикой. Дефектные по репликации вирионы AAV, инкапсулирующие рекомбинантные AAV-векторы согласно изобретению, получают стандартными методами, известными специалистам, с использованием AAV-упаковывающих клеток и с применением технологии упаковки. Примеры таких методов можно найти, например, в патентах США №№ 5436146; 5753500, 6040183, 6093570 и 6548286, которые во всей своей полноте вводятся в настоящее описание посредством ссылки. Другими композициями и методами упаковки являются композиции и методы, описанные в работе Wang et al. (в публикации заявки на патент США 2002/0168342), которая также во всей своей полноте вводится в настоящее описание посредством ссылки, и методы, известные специалистам.
Клетками-хозяевами, подходящими для осуществления настоящего изобретения и продуцирующими rAAV или другие вирионы в экспрессирующих векторах, являются клетки млекопитающих, клетки насекомых, микроорганизмы и дрожжи. Клетками-хозяевами могут быть также упаковывающие клетки, в которых обеспечивается стабильное сохранение генов rep и cap (или других генов) AAV, или клетки-продуценты, в которых обеспечивается стабильное сохранение и упаковка генома AAV-вектора. Репрезентативными упаковывающими клетками и клетками-продуцентами являются клетки, происходящие от клеток 293, A549 или HeLa. AAV-векторы очищают и на их основе приготавливают препараты в соответствии со стандартной технологией, известной специалистам. Другими подходящими клетками-хозяевами (в зависимости от вектора) являются клетки яичника китайского хомячка (CHO), дефицитные по дигидрофолат-редуктазе варианты клеток CHO, такие как клетки CHO DX B11 или CHO DG44 (см., например, Urlaub and Chasin. 1980. Proc. Natl. Acad. Sci. 77:4216-4220), клетки PerC.6 (Jones et al. 2003. Biotechnol. Prog. 19:163-168) или клетки мышиной миеломы Sp/20 (Coney et al. 1994. Cancer Res. 54:2448-2455).
Ретровирусные векторы
Стандартным средством для доставки генов также являются ретровирусные векторы (Miller, 1992. Nature 357: 455-460). Для осуществления настоящего изобретения могут быть использованы ретровирусные векторы, а более конкретно, лентивирусные векторы. В соответствии с этим используемый здесь термин “ретровирус” или “ретровирусный вектор” включает в себя “лентивирус” и “лентивирусные векторы” соответственно. Были проанализированы ретровирусные векторы, и было обнаружено, что они являются подходящими носителями для стабильного введения представляющих интерес генов в геном клеток-мишеней широкого ряда. Способность ретровирусных векторов вводить неаранжированные однокопийные трансгены в клетки делает эти ретровирусные векторы наиболее подходящими для переноса генов в клетки. Кроме того, ретровирусы проникают в клетки-хозяева посредством связывания гликопротеинов ретровирусной оболочки со специфическими рецепторами клеточной поверхности, находящимися на этих клетках-хозяевах. Следовательно, псевдотипированные ретровирусные векторы, в которых кодируемый нативный белок оболочки заменен гетерологичным белком оболочки, обладающим клеточной специфичностью, отличающейся от специфичности нативного белка оболочки (например, способностью связываться с другим рецептором клеточной поверхности по сравнению с нативным белком оболочки), также могут быть использованы для осуществления настоящего изобретения. Способность ретровирусных векторов, включающих одну или несколько последовательностей, кодирующих белок-мишень, к направленной доставке этих последовательностей в конкретные клетки-мишени является желательным фактором для осуществления настоящего изобретения.
Настоящее изобретение относится к ретровирусным векторам, которыми являются, например, трансфицирующие ретровирусные векторы, содержащие одну или несколько трансгенных последовательностей, и упаковывающие ретровирусные векторы, содержащие один или несколько упаковывающих элементов. В частности, настоящее изобретение относится к псевдотипированным ретровирусным векторам, которые кодируют гетерологичный или функционально модифицированный белок оболочки и используются для продуцирования псевдотипированных ретровирусов.
Коровая последовательность ретровирусных векторов согласно изобретению может быть легко выделена из ретровирусов широкого ряда, включая, например, ретровирусы типа B, C и D, а также спумавирусы и лентивирусы (см. RNA Tumor Viruses, Second Edition, Cold Spring Harbor Laboratory, 1985). Примером ретровируса, подходящего для использования в композициях и способах согласно изобретению, является, но не ограничивается им, лентивирус. Другими ретровирусами, подходящими для использования в композициях и способах согласно изобретению, являются, но не ограничиваются ими, вирус птичьего лейкоза; вирус бычьего лейкоза; вирус мышиного лейкоза; вирус норки, индуцирующий образование фокусов выживающих клеток; вирус мышиной саркомы, вирус ретикулоэндотелиоза и вирус саркомы Рауса. Особенно предпочтительными вирусами мышиного лейкоза являются вирусы 4070A и 1504A (Hartley and Rowe. 1976. J. Virol. 19:19-25), вирус Абельсона (ATCC No. VR-999), вирус Фрейнда (ATCC No VR-245), вирус Граффи и Гросса (ATCC No VR-590), вирус саркомы Кирстени-Харви и Раушера (ATCC No VR-998) и вирус мышиного лейкоза Молони (ATCC No VR-190). Эти ретровирусы могут быть легко получены из депозитариев или из коллекций, таких как Американская коллекция типовых культур (ATCC; Manassas, VA), либо они могут быть выделены из известных источников стандартными методами. Существуют и другие коммерчески доступные источники.
Последовательность ретровирусного вектора согласно изобретению может быть получена из лентивирусов. Предпочтительным лентвирусом является вирус иммунодефицита человека, например, типа 1 или 2 (то есть ВИЧ-1 или ВИЧ-2, причем ВИЧ-1 ранее назывался ассоциированным с лимфаденопатией вирусом 3 (HTLV-III) и вирусом, ассоциированным с синдромом приобретенного иммунодефицита (СПИД'а) (ARV)), либо другой идентифицированный вирус, родственный вирусам ВИЧ-1 или ВИЧ-2, и ассоциированный со СПИД'ом или СПИД-подобным заболеванием. Другими лентивирусами являются овечий вирус висны/мэди, вирус кошачьего иммунодефицита (FIV), бычий лентивирус, вирус обезьяньего иммунодефицита (SIV), вирус инфекционной анемии лошадей (EIAV) и вирус козьего артрита-энцефалита (CAEV).
Ретровирусы подходящих родов и штаммов хорошо известны специалистам (см., например, руководство Fields Virology, Third Edition, edited by B. N. Fields et al. 1996. Lippincott-Raven Publishers, см., например, Chapter 58, Retroviridae: The Viruses and Their Replication, Classification, pages 1768-1771, включая таблицу 1, которое вводится в настоящее описание посредством ссылки). Системы упаковки ретровирусов, используемые для генерирования клеток-продуцентов и клеточных линий, продуцирующих ретровирусы, и методы получения таких систем упаковки также известны специалистам.
Типичные системы упаковки включают в себя по меньшей мере два упаковывающего вектора, а именно первый упаковывающий вектор, который содержит первую нуклеотидную последовательность, включающую гены, gag, pol или gag и pol; и второй упаковывающий вектор, который содержит вторую нуклеотидную последовательность, включающую гетерологичный или функционально модифицированный ген оболочки. Ретровирусные элементы могут происходить от лентивируса, такого как ВИЧ. Такие векторы могут не содержать функционального гена tat и/или функциональных вспомогательных генов (vif, vpr, vpu, vpx, nef). Эта система может также содержать третий упаковывающий вектор с нуклеотидной последовательностью, содержащей ген rev. Такая упаковывающая система может быть получена в виде упаковывающих клеток, содержащих первую, вторую и, необязательно, третью нуклеотидную последовательность.
Настоящее изобретение может быть применено к ряду экспрессионных систем, в частности систем эукариотических клеток, а предпочтительно клеток млекопитающих. Если нативные белки являются гликозилированными, то предпочтительно, чтобы такой экспрессионной системой была система, обеспечивающая гликозилирование экспрессированных белков аналогично природному гликозилированию.
Лентивирусы имеют несколько общих структурных белков вириона, включая гликопротеины оболочки SU (gp120) и TM (gp41), которые кодируются геном env; CA (p24), MA (p17), и NC (p7-11), которые кодируются геном gag; и RT, PR и IN, кодируемые геном pol. ВИЧ-1 и ВИЧ-2 содержат вспомогательные и другие белки, участвующие в регуляции синтеза и процессинга вирусной РНК и других репликативных функций. Эти вспомогательные белки, кодируемые генами vif, vpr, vpu/vpx и nef, могут отсутствовать в рекомбинантной системе (либо они могут быть инактивированы). Кроме того, гены tat и rev могут отсутствовать, либо они могут быть инактивированы, например, посредством мутации или делеции.
Системы упаковки лентивирусных векторов первой генерации позволяют получить отдельные упаковывающие конструкции для gag/pol и env, в которых из-за соображений безопасности обычно используется гетерологичный или функционально модифицированный белок оболочки. В системах лентивирусных векторов второй генерации вспомогательные гены, vif, vpr, vpu и nef делетированы или инактивированы. Системами лентивирусных векторов третьей генерации являются системы, в которых ген tat был делетирован или каким-либо иным способом инактивирован (например, посредством мутации).
Недостаток регуляции транскрипции, которая в нормальных условиях обеспечивается геном tat, может быть скомпенсирована благодаря использованию сильного конститутивного промотора, такого как предранний энхансер/промотор (HCAAV-IE) человеческого цитомегаловируса. Как очевидно для каждого специалиста, могут быть выбраны и другие промоторы/энхансеры исходя из их способности к конститутивной промоторной активности, специфичности по отношению к ткани-мишени (например, печень-специфический промотор) или других факторов, относящихся к желательной регуляции экспрессии. Так, например, в некоторых вариантах изобретения для достижения регулируемой экспрессии желательно использовать индуцибельный промотор, такой как промотор гена tet. Ген, кодирующий белок rev, может присутствовать в отдельной экспрессирующей конструкции, в результате чего типичная система лентивирусных векторов третьей генерации будет включать четыре плазмиды, то есть одну плазмиду для gagpol, одну - для rev, одну - для гена оболочки и одну - для вектора переноса. Независимо от генерации используемой упаковывающей системы гены gag и pol могут присутствовать в одной конструкции или в отдельных конструкциях.
Обычно упаковывающие векторы вводят в упаковывающую клетку, и такое введение в клетку осуществляют путем трансфекции, трансдукции или инфицирования. Методы трансфекции, трансдукции или инфицирования хорошо известны специалистам. Ретровирусный трансфицирующий вектор согласно изобретению может быть введен в упаковывающую клеточную линию путем трансфекции, трансдукции или инфицирования с получением клеточной линии или клетки-продуцента. Упаковывающие векторы согласно изобретению могут быть введены в человеческие клетки или клеточные линии стандартными методами, включая, например, опосредуемую фосфатом кальция трансфекцию, липофекцию или электропорацию. В некоторых вариантах изобретения упаковывающие векторы вводят в клетки вместе с доминантным селективным маркером, таким как neo, дигидрофолат-редуктаза (DHFR), глутамин-синтетаза или ADA, с последующим отбором в присутствии соответствующего лекарственного средства и выделением клонов. Селективный маркерный ген может быть физически сцеплен с генами, присутствующими в упаковывающем векторе.
Стабильные клеточные линии, в которых функциональные элементы упаковки имеют конфигурацию, обеспечивающую их экспрессию в подходящей упаковывающей клетке, известны специалистам. Например, см. патент США No 5686279; и Ory et al. 1996. Proc. Natl. Acad. Sci. 93:11400-11406, где описаны упаковывающие клетки. Дополнительное описание продуцирования стабильных клеточных линий можно найти в публикациях Dull et al. 1998. J. Virol. 72(11):8463-8471; и Zufferey et al. 1998. J. Virol. 72:9873-9880.
В работе Zufferey et al. 1997. Nat. Biotechnol. 15:871-75 описана плазмида для упаковки лентивируса, в которой последовательности, расположенные со стороны 3'-конца гена pol, включая ген оболочки ВИЧ-1, были делетированы. Такая конструкция содержит последовательности tat и rev, а 3'-LTR заменен последовательностями poly A. Последовательности 5'-LTR и psi были заменены другим промотором, таким как индуцибельный промотор. Так, например, может быть использован промотор CMV или его производное.
Для усиления экспрессии лентивирусных белков и повышения безопасности в упаковывающих векторах могут быть также изменены упаковывающие функции. Так, например, могут быть удалены все последовательности ВИЧ, расположенные выше от гена gag. Также могут быть удалены последовательности, расположенные ниже от гена оболочки. Кроме того, для повышения уровня сплайсинга и трансляции РНК могут быть проведены стадии модификации данного вектора.
При этом может быть использована, но необязательно, кондиционированная система упаковки, описанная Dull et al. 1998, см. выше. Также желательно использовать самоинактивирующийся вектор (SIN), который повышает биологическую безопасность вектора благодаря делеции длинного концевого повтора (LTR) ВИЧ-1, как описано, например, Zufferey et al. 1998. J. Virol. 72:9873- 9880. Могут быть также использованы индуцибельные векторы, такие как индуцируемые тетрациклином LTR.
Промоторы
Векторы согласно изобретению обычно включают в себя гетерологичные регуляторные последовательности, которыми являются, но не ограничиваются ими, конститутивные промоторы, такие как предранний промотор цитомегаловируса (CMV), LTR RSV, LTR MOMLV и промотор PGK; ткане- или клеткоспецифические промоторы, включая промоторы mTTR, TK, HBV, hAAT, регулируемые или индуцибельные промоторы, энхансеры и т.п.
Подходящими промоторами являются промотор LSP (III et al. 1997. Blood Coagul. Fibrinolysis 8S2:23-30) и промотор EF1-альфа (Kim et al. 1990. Gene 91(2):217-23) и Guo et al. 1996. Gene Ther. 3(9):802-10). Наиболее предпочтительными промоторами являются промотор фактора элонгации 1-альфа (EF1a), промотор фосфоглицерат-киназы-1 (PGK), промотор предраннего гена цитомегаловируса (CMV), химерные печень-специфические промоторы (LSP), энхансер цитомегаловируса/промотор куриного бета-актина (CAG), промотор чувствительности к тетрациклину (TRE), промотор транстиретина (TTR), промотор обезьяньего вируса 40 (SV40) и промотор CK6. Предпочтительным промотором, используемым в целях осуществления настоящего изобретения, является главный поздний промотор аденовируса (Berkner and Sharp. 1985. Nucl. Acids Res. 13:841-857). Конкретный пример последовательности экспрессирующего вектора, в котором используется главный поздний промотор аденовируса, приводится ниже. Последовательности этих и различных других промоторов известны специалистам. Подходящие последовательности могут быть легко получены из общедоступных баз данных и введены в векторы, используемые для осуществления настоящего изобретения.
Особенно предпочтительным промотором, используемым для осуществления настоящего изобретения, является главный поздний промотор аденовируса. Экспрессионный кластер может содержать в направлении 5' → 3' главный поздний промотор аденовируса; треткомпонентную лидерную последовательность, функционально присоединенную к первой последовательности, кодирующей представляющий интерес белок или цепь представляющего интерес белка; последовательность, кодирующую последовательность аутопроцессинга или последовательность расщепления протеазой; вторую последовательность, кодирующую представляющий интерес белок или цепь представляющего интерес белка; и, необязательно, последовательность, кодирующую сайт аутопроцессинга или сайт расщепления протеазой, за которой следует третья последовательность, кодирующая представляющий интерес белок или цепь представляющего интерес белка. Все эти кодирующие последовательности ковалентно присоединены в одной и той же рамке считывания, так что в полипротеин-кодирующей последовательности не происходит терминации трансляции. В процессе синтеза белка или после завершения синтеза полипептида происходит аутопроцессинг или протеолитический процессинг, проиводящий к расщеплению полипротеина с образованием соответствующих цепей белка или белков. В случае синтеза иммуноглобулина последовательность, кодирующая легкую цепь, присутствует в полипротеин-кодирующей последовательности в двух копиях. При этом предпочтительно области, кодирующие лидерную последовательность, могут быть связаны с последовательностями белков или цепей белков; при этом процессинг, происходящий под действием сигнальных пептидаз, имеет дополнительное преимущество, заключающееся в удалении некоторых аминокислотных остатков на N-концах белков, расположенных ниже сайтов процессинга. Компонентами для тяжелой цепи иммуноглобулина являются Met, то есть аминокислота метионин, кодируемая инициирующим кодоном; HC, тяжелая цепь; LC, легкая цепь, SPPC, сайт аутопроцессинга или сайт расщепления протеазой. Экспрессионные конструкции для синтеза иммуноглобулина могут включать указанные компоненты в следующем порядке: Met - протеаза SPPC - лидерная последовательность HC - HC - SPPC- лидерная последовательность LC - LC - SPPC - лидерная последовательность LC - LC; Met - протеаза - SPPC- лидерная последовательность LC - LC - SPPC- лидерная последовательность LC - LC - SPPC- лидерная последовательность HC - HC; Met-протеаза - SPPC - лидерная последовательность LC - LC - SPPC- лидерная последовательность HC - HC - SPPC - лидерная последовательность LC - LC; лидерная последовательность HC - HC -SPPC-лидерная последовательность LC - LC - SPPC- лидерная последовательность LC - LC; лидерная последовательность LC - LC-SPPC - лидерная последовательность HC - HC - SPPC - лидерная последовательность LC - LC; лидерная последовательность LC - LC-SPPC-лидерная последовательность LC - LC - SPPC- лидерная последовательность HC -H C; Met-протеаза - SPPC - лидерная последовательность HC - HC - SPPC - лидерная последовательность LC - LC.
Конкретный пример полипротеин-кодирующей последовательности (продукт: Met - лидерная последовательность НС - HC - сконструированный сайт расщепления фурином - сайт расщепления протеазой TEV - протеаза TEV Nia - сайт расщепления протеазой TEV - лидерная последовательность LC - LC) схематически показан на фиг.1, а на фиг. 2 схематически представлен вектор для экспрессии этой конструкции. Анти-TNFα антитело (D2E7) представляет собой репрезентативное антитело с его соответствующими последовательностями HC и LC. Для продуцирования терапевтического антитела может не потребоваться лидерная последовательность LC. SPPS представляет собой сайт узнавания протеазой TEV, а сайт расщепления фурином находится со стороны 5'-конца от TEV-сайта. Расщепление фурином после расщепления протеазой TEV приводит к восстановлению “правильного” С-концевого лизинового остатка в тяжелой цепи. Полноразмерная последовательность ДНК экспрессирующего вектора D2E7-TEV представлена в таблице 1.
Была сконструирована конкретно проиллюстрированная конструкция для экспрессии полипротеина D2E7 (D2E7-LС-LC-HC), кодирующая тандемный повтор LC и расщепляемая благодаря присутствию протеазной последовательности 2A, используемой в качестве сайта расщепления. С-конец легкой цепи D2E7 был модифицирован с добавлением сайтов расщепления фурином. Это приводило к замене Glu на Arg в положении (обычно) предпоследней аминокислоты и к присоединению лизина у С-конца. Благодаря введению двух последовательностей LC со стороны 5'-конца от HC две копии LC сохраняли ту же самую аминокислотную последовательность. Полная нуклеотидная последовательность экспрессирующего вектора представлена в таблице 6C, а аминокислотная последовательность и последовательность, кодирующая полипротеин, представлены в таблицах 6B и 6A соответственно. См. также SEQ ID №№:29-31. Схематическая карта экспрессирующего вектора представлена на фиг. 7.
Другим конкретно проиллюстрированным полипротеином (и его кодирующей последовательностью) является полипротеин ABT-007-TEV; см. таблицы 2B и 2A соответственно. См. также SEQ ID N№: 33 и 32. Это рекомбинантное антитело специфически связывается с рецептором эритропоэтина (EpoR). Полноразмерная последовательность экспрессирующего вектора, кодирующего сконструированный полипротеин ABT-007-TEV, представлена в таблице 2C (SEQ ID NO:35). См. также SEQ ID NO:34. Такой вектор схематически представлен на фиг. 3.
Другим конкретно проиллюстрированным полипротеином и его кодирующей последовательностью является полипротеин ABT-874-TEV; см. таблицы 3B и 3A соответственно. Это антитело специфически связывается с интерлейкином-12. Экспрессирующий вектор схематически представлен на фиг 4. См. также SEQ ID N№: 35-37.
Еще одним конкретно проиллюстрированным полипротеином (и его кодирующей последовательностью) является полипротеин EL246-GG-TEV; см. таблицы 4B и 4A. Антитело, кодируемое этим вектором, специфически связывается с E/L-селектином. Экспрессирующий вектор схематически представлен на фиг 5. См. также SEQ ID N№:38-40.
ABT-325-TEV представляет собой сконструированное антитело, которое специфически связывается с интерлейкином-18. Последовательности, кодирующие полипротеин, и его аминокислотные последовательности представлены в таблицах 5A и 5B соответственно, а полноразмерная последовательность экспрессирующего вектора представлена в таблице 5C. Экспрессирующий вектор для синтеза полипротеина представлен на фиг. 6. См. также SEQ ID №№:41-43.
Настоящее изобретение также относится к протеазе TEV, из которой был удален сигнал локализации в ядре (NLS) (TEV NLS-). Протеаза TEV или TEV(NLS-) может также временно или стабильно экспрессироваться в клетках как часть отдельного вектора или отдельного транскрипта. Белок TEV(NLS-), который ранее локализовался в NLS-части, может заякориваться в ЭP или в рибосоме путем включения ЭP-заякоривающей последовательности или путем присоединения к небольшому белку, связывающемуся с рибосомой, соответственно.
Хотя в настоящей заявке обсуждается протеолитическое расщепление белков-предшественников и полипротеинов в процессе их синтеза или в клетках после синтеза, однако при этом следует отметить, что указанные полипротеины и белки-предшественники (про-протеины) могут быть получены после сбора этих белков с использованием соответствующей(их) протеазы(протеаз) in vitro.
В объем настоящего изобретения входят конкретные экспрессированные антитела (иммуноглобулины), которыми могут быть, inter alia, антитело, специфически связывающееся с фактором некроза опухоли (сконструированное антитело, соответствующее и/или происходящее от HUMIRA/D2E7, торговый знак адалимумаба Abbott Biotechnology Ltd., Hamilton, Bermuda); антитело, специфически связывающееся с интерлейкином-12 (сконструированное антитело, происходящее от ABT-874); антитело, специфически связывающееся с интерлейкином-18 (сконструированное антитело, происходящее от ABT-325); антитело, специфически связывающееся с рекомбинантным рецептором эритропоэтина (сконструированное антитело, происходящее от ABT-007); или антитело, специфически связывающееся с E/L-селектином (сконструированное антитело, происходящее от EL246-GG). Последовательности, кодирующие сконструированные полипротеины, и аминокислотные последовательности таких полипротеинов представлены в таблицах 1-5. Другими антителами, подходящими для использования в настоящем изобретении, являются, например, Remicade (инфликсимаб); Rituxan/Mabthera (ритуксимаб); Herceptin (трастузумаб); Avastin (бевацизумаб); Synagis (паливизумаб); Erbitux (цетуксимаб); Reopro (абциксимаб); Orthoclone OKT3 (муромонаб-CD3); Zenapax (даклизумаб); Simulect (базиликсимаб); Mylotarg (гемтузумаб); Campath (алемтузумаб); Zevalin (ибритумомаб); Xolair (омализумаб); Bexxar (тозитумомаб) и Raptiva (эфализумаб), где обычно после торгового знака/названия торговой марки в скобках приводится их соответствующее родовое название. Другими подходящими белками являются, например, один или несколько из нижеследующих белков, таких как эпоэтин-альфа, эпоэтин-бета, этанерцепт, дарбепоэтин-альфа, филграстим, интерферон-бета 1a, инетрферон-бета 1b, интерферон-альфа 2b, инсулин гларгин, соматотропин, терипаратид, фоллитропин-альфа, дорназа, фактор VIII, фактор VII, фактор IX, имиглюцераза, незиритид, ленограстим и фактор фон Виллебранда, где каждое одно или несколько родовых названий может соответствовать продуктам, имеющим одно или несколько торговых знаков/названий торговой марки. Специалистам должны быть известны и другие антитела и белки, подходящие для их использования в настоящем изобретении.
В настоящем изобретении также рассматривается регулируемая экспрессия последовательности, кодирующей два или более представляющих интерес полипептидов, белков или про-протеинов. Системы регуляции генов могут быть использованы для модуляции экспрессии конкретного(ых) гена(ов). В одном из репрезентативных подходов система регуляции или переключения генов включает в себя химерный фактор транскрипции, содержащий лиганд-связывающий домен, домен активации транскрипции и ДНК-связывающий домен. Такие домены могут быть выделены фактически из любого источника и могут быть объединены любым из существующих методов с получением нового белка. Система регуляции генов также включает в себя ДНК элемента ответа, которая взаимодействует с химерным фактором транскрипции. Этот элемент регуляции транскрипции является смежным с регулируемым геном.
Репрезентативными системами регуляции транскрипции, которые могут быть использованы для осуществления настоящего изобретения, являются, например, система экдизонов Drosophila (Yao et al. 1996. Proc. Natl. Acad. Sci. 93:3346), система экдизонов Bombyx (Suhr et al. 1998. Proc. Natl. Acad. Sci. 95:7999), система рецепторов синтетического прогестерона GeneSwitch (торговый знак Valentis, The Woodlands, TX), в которой в качестве индуктора используется RU486 (Osterwalder et al. 2001. Proc. Natl. Acad. Sci. USA 98(22): 12596-601); системы Tet и RevTet (регулируемые тетрациклином экспрессионные системы, торговые знаки BD Biosciences Clontech, Mountain View, CA), в которых для регуляции (“включения” или “отключения”) транскрипции мишени используются небольшие молекулы, такие как тетрациклин (Tc) или его аналоги, например доксициклин (Knott et al. 2002. Biotechniques 32(4):796, 798, 800); регуляторная система ARIAD Regulation Technology (Ariad, Cambridge, MA), которая была получена с использованием небольшой молекулы вместе с двумя внутриклеточными молекулами, каждая из которых присоединена либо к активатору транскрипции, либо к ДНК-связывающему белку. Совместное присутствие этих компонентов приводит к активации транскрипции представляющего интерес гена. Ariad представляет собой систему, основанную на гомодимеризации, и систему, основанную на гетеродимеризации (Rivera et al. 1996. Nature Med. 2(9):1028- 1032; Ye et al. 2000. Science 283:88-91).
Экспрессирующие векторные конструкции согласно изобретению, включающие последовательности нуклеиновой кислоты, кодирующие антитела или их фрагменты или другие гетерологичные белки или про-протеины в виде рекомбинантных полипептидов, расщепляемых посредством аутопроцессинга или под действием протеазы, могут быть введены в клетки in vitro, ex vivo или in vivo для доставки чужеродных, терапевтических или трансгенных белков в клетки, например в соматические клетки, или для продуцирования рекомбинантных полипептидов в клетках, трансдуцированных вектором.
Клетки-хозяева и доставки векторов
Векторные конструкции согласно изобретению могут быть введены в подходящие клетки in vitro или ex vivo стандартными методами, известными специалистам. Такими методами являются, например, трансфекция с использованием фосфата кальция, микроинжекция в культивированные клетки (Capecchi, 1980, Cell 22:479-488), электропорация (Shigekawa et al. 1988. BioTechnology 6:742-751), опосредуемый липосомами перенос генов (Mannino et al. 1988. BioTechnology 6:682-690), опосредуемая липидами трансдукция (Feigner et al. 1987. Proc. Natl. Acad. Sci. USA 84:7413-7417) и доставка нуклеиновых кислот методами высокоскоростной микробаллистики (Klein et al. 1987. Nature 327:70-73).
Для in vitro и ex vivo экспрессии может быть использована любая клетка, эффективная для экспрессии функционального белкового продукта. Специалистам известны различные примеры клеток и клеточных линий, используемых для экспрессии белка. Так, например, для экспрессии могут быть использованы прокариотические клетки и клетки насекомых. Кроме того, могут быть использованы эукариотические микроорганизмы, такие как дрожжи. Методы экспрессии рекомбинантных белков в прокариотических системах, в системах насекомых и в дрожжевых системах, по существу, известны специалистам и могут быть адаптированы для экспрессии антител или других белков с применением композиций и способов согласно изобретению.
Примерами клеток, используемых для экспрессии, также являются клетки млекопитающих, такие как фибробласты; клетки млекопитающих, не являющихся человеком, таких как овцы, свиньи, мыши и коровы; клетки насекомых и т.п. Конкретными примерами клеток млекопитающих являются, но не ограничиваются ими, клетки COS, клетки VERO, клетки НеLa, клетки яичника китайского хомячка (CHO), клетки CHO DX B11, клетки CHO DG44, клетки PerC.6, клетки Sp2/0, клетки 293, клетки NSO, фибробласты 3T3, клетки W138, клетки BHK, клетки HEPG2 и клетки MDCK.
Клетки-хозяева культивируют в стандартной питательной среде, модифицированной, если это необходимо, для индуцирования промоторов, отбора трансформантов или амплификации генов, кодирующих нужные последовательности. Клетки-хозяева млекопитающих могут быть культивированы в различных средах. Наиболее подходящими для культивирования клеток-хозяев являются коммерчески доступные среды, такие как среда Хэмса F10 (Sigma), минимальная питательная среда (MEM)(Sigma), RPMI 1640 (Sigma) и модифицированная по способу Дульбекко среда Игла (DMEM) (Sigma). В указанные среды обычно добавляют, если это необходимо, гормоны и/или другие факторы роста (такие как инсулин, трансферрин или эпидермальный фактор роста), соли (такие как хлорид натрия и фосфат кальция и магния), буферы (такие как HEPES), нуклеозиды (такие как аденозин и тимидин), антибиотики, микроэлементы и глюкозу или эквивалентный источник энергии. Как хорошо известно специалистам, могут быть также включены и другие необходимые добавки в соответствующих концентрациях. Соответствующие условия культивирования конкретных клеточных линий, такие как температура, рН и т.п., по существу, хорошо известны специалистам, при этом предполагаемые условия культивирования многочисленных клеточных линий описаны, например, в каталоге ATCC (доступном на сайте Интернета "atcc.org/SearchCatalogs/AllCollections.cfm") или в прилагаемых инструкциях поставщиков.
Для доставки множества генов, сцепленных с последовательностью аутопроцессинга, в целях экспрессии двух или более белков или полипептидов у животных-молелей или у человека экспрессирующие векторы могут быть введены in vivo различными способами (например, внутрикожно, внутривенно, внутрь опухоли, в головной мозг, через воротную вену, внутрибрюшинно, внутримышечно, в мочевой пузырь и т.п.). В зависимости от способа введения терапевтические белки вырабатывают местный ответ (в головном мозге или в мочевом пузыре) или системный ответ (при других способах введения). Использование тканеспецифических промоторов, расположенных со стороны 5'-конца по отношению к открытой(ым) рамке(ам) считывания, приводит к тканеспецифической экспрессии белков или полипептидов, кодируемых целой открытой рамкой считывания.
Различные методы введения рекомбинантного экспрессирующего вектора, несущего трансген, в клетки-мишени in vitro, ex vivo или in vivo были ранее описаны и хорошо известны специалистам. Настоящее изобретение относится к терапевтическим способам, к вакцинам и к противораковой терапии, проводимой путем инфицирования клеток-мишеней рекомбинантными векторами, содержащими последовательность, кодирующую два или более представляющих интерес белков или полипептидов, и экспрессии указанных белков или полипептидов в клетках-мишенях.
Так, например, рекомбинантные векторы согласно изобретению могут быть введены in vivo в различные органы широкого ряда, включая, но не ограничиваясь ими, головной мозг, печень, кровеносные сосуды, мышцы, сердце, легкие и кожу.
В случае переноса гена ex vivo, клетки-мишени хозяина выделяют и генетически модифицируют в лаборатории с использованием рекомбинантных векторов согласно изобретению и в соответствии с методами, хорошо известными специалистам.
Рекомбинантные векторы согласно изобретению могут быть введены стандартными способами введения, включая, но не ограничиваясь ими, способы, описанные выше. Рекомбинантные векторы согласно изобретению могут присутствовать в различных препаратах, которыми являются, но не ограничиваются ими, жидкие растворы и суспензии, микровезикулы, липосомы и растворы для инъекций или вливаний. Предпочтительная форма препарата зависит от способа введения и целей терапевтического применения.
Преимуществами рекомбинантных экспрессирующих векторных конструкций согласно изобретению, предназначенных для продуцирования иммуноглобулина или другого биологически активного белка in vivo, являются: возможность введения одного вектора для длительной и пролонгированной экспрессии антитела у пациентов; возможность in vivo экспрессии антитела или его фрагмента (или другого биологически активного белка), обладающего всеми биологическими активностями; и возможность создания природных посттрансляционных модификаций антитела, продуцируемого в человеческих клетках. При этом экспрессируемый белок предпочтительно идентичен или достаточно идентичен природному белку, но в большинстве случаев не стимулирует вырабатывание иммунных ответов при его введении пациенту или при его непрерывной экспрессии у пациента, нуждающегося в этом.
Рекомбинантные векторные конструкции согласно изобретению могут быть также использованы для in vitro продуцирования рекомбинантных антител и других биологически активных белков в целях их применения в терапии или в научных исследованиях. Методы продуцирования рекомбинантного белка хорошо известны специалистам и могут быть применены для экспрессии рекомбинантных антител, осуществляемой с использованием описанных здесь векторных конструкций, содержащих сайт аутопроцессинга или другой сайт расщепления протеазой.
В одном из своих аспектов настоящее изобретение относится к способам продуцирования рекомбинантного иммуноглобулин а или его фрагмента путем введения экспрессирующего вектора, такого как вышеописанный вектор, в клетку с получением трансфицированной клетки, где указанный вектор включает в себя в направлении 5'→3': промотор, функционально присоединенный к последовательностям, кодирующим тяжелую цепь и две легкие цепи иммуноглобулина или его фрагмента; последовательность аутопроцессинга, такую как последовательность 2А или 2А-подобную последовательность, или сайт расщепления протеазой, расположенные между указанными цепями. Следует отметить, что последовательность, кодирующая тяжелую цепь иммуноглобулина, или последовательность, кодирующая легкую цепь иммуноглобулина, могут быть локализованы со стороны 5'-конца по отношению к последовательности 2А (то есть первой последовательности) в данной векторной конструкции. Альтернативно, протеаза, когнатная сайту расщепления протеазой, может экспрессироваться как часть полипротеина, то есть она может самопроцессироваться из остальной части полипротеина или протеолитически отщепляться отдельной (или той же самой) протеазой. Белки с множеством других цепей или другие белки (такие как белки в друхгибридных или трехгибридных системах) могут экспрессироваться в процессированной активной форме благодаря замене релевантных кодирующих последовательностей, перемежающихся с сайтами аутопроцессинга или сайтами узнавания протеазой, также имеющими нужных размер, с продуцированием отдельных белков.
Двухгибридная (и другая) система была использована для скрининга библиотек кДНК на ранее не идентифицированные партнеры по связыванию, которые, как известно, представляют собой лиганд или субъединицу белкового комплекса. При введении в эту систему соответствующих изменений могут быть также идентифицированы белки или субъединицы, которые ингибируют связывание, конкурируют за связывание или блокируют связывание с известным комплексом. Хотя двухгибридные (или другие) системы были применены в ряде научных исследований, однако эти системы могут оказаться неэффективными из-за высокой частоты ложноположительных или ложноотрицательных результатов. Такие ложные сигналы были получены по меньшей мере в нескольких случаях, что может быть обусловлено дисбалансом между относительной экспрессией белка-"приманки" и кандидатов на белки-партнеры по связыванию или кандидатами на белки-дезинтеграторы. Дополнительным преимуществом такой стратегии согласно изобретению является то, что клетка-хозяин трансфицируется или трансформируюется только одной плазмидой, и то, что для отбора этой плазмиды вместо двух процедур отбора, как это предусматривается в двухгибридных схемах с использованием бинарных векторов, требуется только одна процедура отбора. Этот способ может быть также адаптирован для его применения к трехгибридным системам. Обсуждение двухгибридных систем приводится в публикациях Toby & Golemis. 2001. Methods 24:201-217; Vidal and Legrain. 1999, Nucl. Acids Res. 27:919-929; Drees, B. 1999. Curr. Op. Chem. Biol. 3:64-70; и Fields and Song. 1989, Nature 340:245-246. На фиг.9 схематически представлен полипротеин и проиллюстрирована стратегия экспрессии посредством аутопроцессинга или расщепления протеазой для белка-“приманки” и белка-“жертвы” (или кандидата на белок-“жертву”), а на фиг.8 проиллюстрирован вектор, содержащий экспрессионный кластер для продуцирования белка-“приманки” и белка-“жертвы” сначала как гибридный белок GAL4::bait::2A, который подвергается самопроцессингу после трансляции пептида 2A. Второй открытой рамкой считывания (ORF) является гибридный белок NFkappaB::библиотека. Для конструирования белка-“приманки” в MCS1 необходима трансляция в той же рамке считывания с образованием самопроцессирующейся пептидной последовательности 2A. Конструирование экспрессионной библиотеки в нижерасположен-ной MCS2 имеет менее важное значение.
Описанная здесь стратегия может быть также адаптирована для экспрессии белков, которые экспрессируются в форме про-белков, процессирующихся в зрелую активную форму путем протеолитического расщепления, что позволяет получить композиции для рекомбинантной экспрессии и разработать методы такой экспрессии. Примерами таких белков являются, но не ограничиваются ими, интерлейкины 1 и 18 (IL-1 и IL-18), исулин и т.п. IL-1 и IL-18 продуцируются в цитоплазме воспалительных клеток. В этих молекулах отсутствует традиционный сигнал секреции, и такие молекулы должны расщепляться протеазой с последующей секрецией в биологически активной форме. IL-1 процессируется в зрелую форму под действием интерлейкин-конвертирующего фермента (ICE). Про-IL-18 превращается в зрелый IL-18 под действием каспаз. Продуцирование этих молекул в рекомбинантной форме является трудной задачей, поскольку эти клетки, которые часто используются в качестве хозяев, не экспрессируют протеазы, необходимые для продуцирования биологически активных зрелых форм этих белков. Экспрессия этих цитокинов без про-доменов приводит к инактивации молекул и/или к снижению уровней их продуцирования. Настоящее изобретение относится к первичным продуктам трансляции, которые содержат между про-доменом и аминокислотой зрелого полипептида сконструированный сайт аутопроцессинга (например, последовательность 2A) или встроенный сайт расщепления протеазой, и в данном случае не требуется экспрессии потенциально токсичной протеазы одновременно с представляющим интерес белком.
В своем родственном аспекте настоящее изобретение относится к способу продуцирования рекомбинантного иммуноглобулина или его фрагмента путем введения экспрессирующего вектора, такого как вышеописанный вектор, в клетку, где указанный вектор также включает в себя дополнительный сайт протеолитического расщепления, расположенный между первой и второй кодирующими последовательностями иммуноглобулина. Предпочтительным дополнительным сайтом протеолитического расщепления является сайт расщепления пурином с консенсусной последовательностью RXK/R-R (SEQ ID NO:1). Обсуждение приводится в публикации заявки на патент США 2005/0003482A1.
В одном из репрезентативных аспектов изобретения встраивание или введение вектора в клетку осуществляют в одну или несколько стадий, таких как культивирование трансфицированной клетки в условиях, подходящих для отбора клетки и экспрессии полипротеина или про-белка; измерение уровня экспрессии иммуноглобулина или его фрагмента или другого(их) белка(ов); и сбор иммуноглобулина или его фрагмента или другого(их) белка(ов).
В другом своем аспекте настоящее изобретение относится к клеткам, экспрессирующим рекомбинантный иммуноглобулин или его фрагмент или другой(ие) белок(и) или представляющий интерес белок, где указанная клетка содержит экспрессирующий вектор для экспрессии двух или более цепей иммуноглобулина или его фрагментов или другого(их) про-белка(ов) или белка(ов); промотор, функционально присоединенный к первой кодирующей последовательности иммуноглобулина или другой цепи или ее фрагмента; последовательность, кодирующую сайт аутопроцессинга или другой сайт расщепления, такую как последовательность 2А или 2А-подобную последовательность, или сайт узнавания протеазой, и вторую кодирующую последовательность иммуноглобулина или его другой цепи или фрагмента, где указанная последовательность, кодирующая сайт аутопроцессинга или сайт узнавания протеазой, встроена между первой и второй кодирующими последовательностями. В родственном аспекте изобретения указанная клетка содержит экспрессирующий вектор, описанный выше, где указанный экспрессирующий вектор также содержит дополнительный сайт протеолитического расщепления между первой и второй кодирующими последовательностями представляющего интерес иммуноглобулина. Предпочтительным дополнительным сайтом протеолитического расщепления является сайт расщепления фурином с консенсусной последовательностью RXR/K-R (SEQ ID NO:1).
Используемый здесь термин “кодирующая последовательность для первой цепи молекулы иммуноглобулина или его фрагмента” означает последовательность нуклеиновой кислоты, кодирующую молекулу белка, включая, но не ограничиваясь ими, легкую цепь или тяжелую цепь антитела или иммуноглобулина или его фрагмента.
Используемый здесь термин “кодирующая последовательность для второй цепи молекулы иммуноглобулина или его фрагмента” означает последовательность нуклеиновой кислоты, кодирующую молекулу белка, включая, но не ограничиваясь ими, легкую цепь или тяжелую цепь антитела или иммуноглобулина или его фрагмента. При этом следует отметить, что в одном из аспектов изобретения повышенный уровень экспрессии наблюдается в том случае, если присутствуют две копии кодирующей последовательности легкой цепи иммуноглобулина на одну копию кодирующей последовательности тяжелой цепи иммуноглобулина.
Последовательность, кодирующая первую и вторую цепь антитела или иммуноглобулина или его фрагмента, включает в себя тяжелую цепь или ее фрагмент, происходящий от IgG, IgM, IgD, IgE или IgA. В более широком смысле последовательность, кодирующая цепь антитела или иммуноглобулина или его фрагмента, также включает в себя легкую цепь или ее фрагмент, происходящий от IgG, IgM, IgD, IgE или IgA. Генами всех молекул антитела, а также их модифицированных или производных форм являются, например, гены других фрагментов молекул узнавания антитена, таких как Fab, одноцепочечный Fv (scFv) и F(ab')2. Антитела и их фрагменты могут происходить от животного или могут представлять собой химерное антитело “человек - мышь”, гуманизованное антитело; антитело, модифицированное путем деиммунизации (Biovation Ltd); антитело, модифицированное в целях изменения аффинности связывания с Fc-рецепторами, или полностью человеческое антитело. При этом желательно, чтобы антитело или другой рекомбинантный белок не вырабатывали иммунного ответа у человека или животного, которым они были введены.
Антитела могут быть биспецифическими, и такими антителами являются, но не ограничиваются ими, диантитела, квадромы, мини-антитела, антитела ScBs и антитела с укладкой типа “выступ в углублении” (“knobs-into-holes”).
Продуцирование и выделение самих антител может быть осуществлено различными методами, хорошо известными специалистам (Harlow et al. 1988. Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory). Другие представляющие интерес белки собирают и/или очищают и/или используют в соответствии с методами, хорошо известными специалистам.
Для практической реализации настоящего изобретения продуцирование антитела или его варианта (аналога) методами рекомбинантных ДНК может быть осуществлено путем культивирования модифицированной клетки-хозяина в условиях, благоприятствующих росту клеток-хозяев и экспрессии кодирующих последовательностей. Для подтверждения успешной экспрессии может быть проведен мониторинг уровней антитела против данного антигена в соответствии со стандартными методами, такими как ELISA, РИА и т.п. Антитела выделяют из супернатанта культуры стандартными методами, известными специалистам. Разумеется, что очищенные формы этих антител могут быть легко получены стандартными методами очистки, включая, но не ограничиваясь ими, аффинную хроматографию на колонках с белком А, G-белком или белком L, или методами в зависимости от конкретного антигена или даже от нужной специфичности конкретного эпитопа антигена. Антитела могут быть также очищены с помощью стандартной хроматографии, такой как колоночная ионообменная хроматография или эксклюзионная хроматография, в комбинации с другими технологиями, такими как преципитация сульфатом аммония и фильтрация на мембране с соответствующим размером пор. Если данные экспрессионные системы сконструированы так, чтобы они включали сигнальные пептиды, то полученные антитела секретируются в культуральную среду или супернатант, однако возможно также и внутриклеточное продуцирование.
Продуцирование и отбор антигенспецифических полностью человеческих моноклональных антител у мышей, которым были введены локусы человеческих Ig, были описаны в литературе (Jakobovits et al. 1998. Advanced Drug Delivery Reviews 31:33-42; Mendez et al. 1997. Nature Genetics 15: 146-156; Jakobovits et al. 1995. Curr Opin Biotechnol 6: 561-566; Green et al. 1994. Nature Genetics Vol. 7:13-21).
Высокий уровень экспрессии терапевтических моноклональных антител был достигнут в молоке трансгенных коз, и было показано, что их уровни связывания с антигеном были эквиваленты уровням связывания моноклональных антител, продуцированных стандартным методом культивирования клеток. Этот метод основан на получении человеческих терапевтических белков в молоке трансгенных животных, несущих генетическую информацию, которая позволяет осуществлять экспрессию человеческих терапевтических белков в молоке этих животных. После продуцирования таких рекомбинантных белков эти белки могут быть успешно выделены из молока с помощью стандартной технологии. См., например, Pollock et al. 1999. J. Immunol. Meth. 231:147-157 and Young et al. 1998. Res Immunol. 149(6): 609-610. Было продемонстрировано, что молоко, яичный белок, кровь, моча и зародышевая плазма трансгенных животных, а также трансгенные коконы шелкопряда являются потенциальными источниками для продуцирования рекомбинантных белков в промышленном масштабе (Houdebine L M. 2002. Curr Opin Biotechnol 13:625-629; Little et al. 2000. Immunol Today, 21 (8):364-70; and Gura T. 2002. Nature, 417:584-5860). В настоящем изобретении также рассматривается применение экспрессионных систем для экспрессии у трансгенных животных рекомбинантного антитела или его варианта (аналога) или другого(их) представляющего(их) интерес белка(ов), где в указанных системах используются векторы согласно изобретению, кодирующие сайт аутопроцессинга и/или сайт узнавания протеазой.
Было также успешно продемонстрировано продуцирование рекомбинантных белков в растениях, включая, но не ограничиваясь ими, картофель, томаты, табак, рис и другие растения, трансформированные посредством инфицирования агробактерией (Agrobacterium), или посредством биобаллистической трансформации, трансформации протопластов и т.п. Была продемонстрирована экспрессия рекомбинантного человеческого GM-CSF в семенах трансгенных растений табака, а также экспрессия антител, включая одноцепочечные антитела в растениях. См., например, Streaffield and Howard. 2003. Int. J. Parasitol. 33:479-93; Schillberg et al. 2003. Cell Mol Life Sci. 60:433A5; Pogue et al. 2002. Annu. Rev. Phytopathol. 40:45-74; and McCormick et al. 2003. J Immunological Methods, 278:95-104. В настоящем изобретении рассматривается применение экспрессионных систем для экспрессии в трансгенных растениях рекомбинантного иммуноглобулина или его фрагмента или другого(их) представляющего(их) интерес белка(ов), где в указанных системах используются векторы согласно изобретению, кодирующие сайт расщепления протеазой и/или сайт аутопроцессинга.
Экспрессионные системы на основе бакуловирусных векторов, используемые в комбинации с клетками насекомых, находят все большее применение как основа для практической реализации продуцирования рекомбинантного белка. Сообщалось, что применение бакуловирусных экспрессионных систем для клеточных культур млекопитающих имеет то преимущество, что оно облегчает культивирование клеток и обеспечивает более высокие уровни экспрессии. См., например, Ghosh et al. 2002. Mol Ther. 6:5-11 и lkonomou et al. 2003. Appl Microbiol Biotechnol. 62:1-20. В настоящем изобретении рассматривается применение экспрессионных систем на основе бакуловирусных векторов для экспрессии в трансгенных растениях рекомбинантного иммуноглобулина или его фрагмента, где в указанных системах используются векторы согласно изобретению, кодирующие сайт аутопроцессинга. Бакуловирусные векторы и подходящие клетки-хозяева хорошо известны специалистам и являются коммерчески доступными.
Экспрессионные дрожжевые системы, включая двух- или трехгибридные системы, могут быть также использованы для экспрессии рекомбинантного иммуноглобулина или его фрагмента или другого(их) представляющего(их) интерес белка(ов), где в указанных системах используются векторы согласно изобретению, кодирующие сайт аутопроцессинга. См., например, патент США №5643745, который вводится в настоящее описание посредством ссылки.
Следует отметить, что экспрессионные кластеры, векторы и рекомбинантные клетки-хозяева согласно изобретению, содержащие кодирующие последовательности для самопроцессирующегося пептида, присутствующего отдельно или в комбинации с дополнительными кодирующими последовательностями для сайта протеолитического расщепления, находят применение в любой системе экспрессии белка, обеспечивающей экспрессию рекомбинантных иммуноглобулинов или их фрагментов, про-белков, биологически активных белков и белковых компонентов двухгибридных и трехгибридных систем, некоторые из которых известны специалистам и примеры которых приводятся в настоящей заявке. Векторы согласно изобретению могут быть легко адаптированы для их использования в любой системе экспрессии белка самим специалистом в данной области.
Если в настоящей заявке заявлены соединение, конструкция или композиция, то в этой связи следует отметить, что соединения, конструкции или композиции, известные специалистам, включая соединения, конструкции или композиции, упоминаемые в ссылочных материалах, не входят в объем настоящего изобретения. Если в настоящем изобретении используется группа Маркуша или другие группы, включая все отдельные члены этой группы и все возможные комбинации и субкомбинации, объединяемые в эту группу, то, очевидно, что такая группа, любой ее отдельный член или подгруппа также входят в объем настоящего изобретения.
Пример 1. Экспрессия иммуноглобулинов с применением опосредуемого интеином процессинга
Стратегия эффективной экспрессии молекул антител основана на экспрессии полипротеинов с использованием конструкций, в которых интеин локализован между тяжелой и легкой цепями, причем последовательность интеина и/или последовательности стыка должны быть модифицированы так, чтобы происходило высвобождение белковых компонентов без лигирования N-концевых и C-концевых белков. В таких конструкциях может присутствовать одна копия каждой из соответствующих тяжелой и легкой цепей или легкая цепь может присутствовать в двух копиях, либо может присутствовать множество копий тяжелой и легкой цепей, при условии что функциональная последовательность расщепления будет обеспечивать разделение каждого иммуноглобулинового белка в данном полипротеине. Такая стратегия с использованием интеина может быть применена более чем один раз, либо по меньшей мере на одном конце иммуноглобулинового белка могут присутствовать другие последовательности протеолитического процессинга или последовательности расщепления ферментами.
Интеин, происходящий от Pyrococcus horikoshii, был введен в конструкцию методом, вкратце описанным выше, и было показано, что такое введение позволяет успешно продуцировать “правильно” процессированное и полностью функциональное антитело D2E7. Дополнительными протестированными интеинами являются интеины от Saccharomyces cerevisiae и штамма PCC6803 Synechocystis spp., и ELISA-анализ показал, что эти интеины способствуют продуцированию секретированного антитела.
ПЦР-амплификация и субклонирование интеина Pho Pol I Pyrococcus horikoshii:
Для амплификации интеина Pho Pol I Pyrococcus horikoshii (NCBI/белок, рег. #059610, GenBank, рег.# для полноразмерной последовательности ДНК-полимеразы I - BA000001.2:1686361..1690068, взятые из полноразмерной геномной последовательности P. horikoshii), проводимой с использованием геномной ДНК в качестве матрицы и с использованием смеси ДНК-полимераз (Platinum Taq Hi Fidelity DNA Polymerase Supermix) (Invitrogen, Carlsbad, CA), были получены нижеследующие олигонуклеотиды. Геномную ДНК закупали в ATCC.
P. horikoshii int-5' AGCATTTTACCAGATGAATGGCTCCC (SEQ ID NO:52)
P. horikoshii int-3' AACGAGGAAGTTCTCATTATCCTCAAC (SEQ ID NO:53)
ПЦР проводили в соответствии со следующей программой:
ПЦР-продукт субклонировали в pCR2.1-TOPO (Invitrogen), а затем вставку секвенировали и подтверждали ее правильность. На этой стадии было выявлено, что последовательность у 3'-конца интеина отсутствовала из-за ошибки при считывании. Отсутствующая последовательность была затем введена в процессе проведения последующих реакций ПЦР для связывания интеина с тяжелой и легкой цепью D2E7.
Для создания гибридной конструкции “тяжелая цепь D2E7 - интеин - легкая цепь D2E7” конструировали олигонуклеотидные праймеры. Праймеры были сконструированы так, чтобы ПЦР-продукт можно было использовать в последующих ПЦР-реакциях в качестве праймеров.

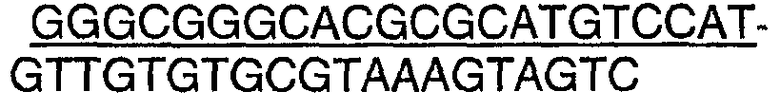

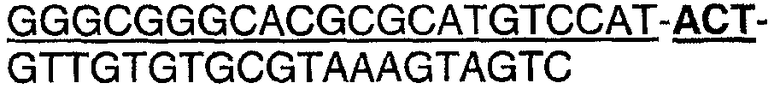
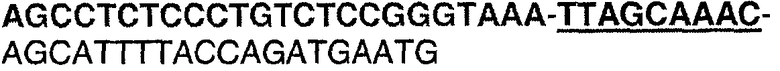

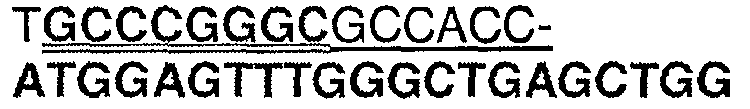
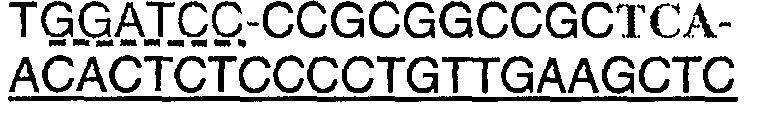
Маркировка шрифта для идентификации компонентов последовательностей:
Последовательность тяжелой цепи (жирный шрифт, [красный]) - Последовательность легкой цепи (подчеркнута)- Последовательность интеина P.horikoshi (обычный шрифт Arial) - последовательность экстеина P.horikoshi (жирный шрифт с подчеркиванием, [синий]) - последовательность узнавания ферментом Srfl, GCCCGGGC (двойное подчеркивание, [зеленый]) - последовательность узнавания ферментом BamHI, GGATСC (подчеркнуто пунктиром, [фиолетовый]); стоп-кодон, TCA (шрифт Times New Roman, [оливковый] цвет), - последовательность Козака. Kozak
ПЦР-амплификация и сборка гибрида “тяжелая цепь D2E7 - интеин- легкая цепь D2E7”:
ПЦР проводили с использованием в качестве матрицы клона интеина pCR2.1-TOPO-p.horikoshii, полученного как описано выше, а также с использованием праймеров P. horikoshii int-5' и модифицированного P.hori-3' для восстановления исходной последовательности со стороны 3'-конца интеина. Во избежание присоединения поли-А-хвоста, которое происходит под действием полимеразы Platinum Taq, использовали ДНК-полимеразу Pful.
ПЦР проводили в соответствии со следующей программой:
Продукт ПЦР-амплификации подвергали гель-очистке с использованием набора для гель-экстракции (Qiaquick Gel Extraction kit) (Qiagen, Valencia, CA). Этот продукт использовали в качестве матрицы в нижеследующей серии реакций.
Три серии реакций ПЦР проводили для генерирования кодирующих последовательностей интеина с варьирующимся числом экстеиновых остатков, фланкирующих 5'- и 3'-концы кодирующей последовательности интеина. Кодоны экстеинов происходят от природного гена ДНК-полимеразы в P. hohkoshii, природной частью которого является данный интеин. Были использованы следующие праймеры: праймер Set 1, который не вводит последовательность экстеина (с нулевой последовательностью экстеина) (HC-интеин-5' и модифицированная LC-интеин-3'), праймер Set 2, который вводит одну аминокислоту (3 пары оснований) у обоих концов интеина (HC-интеин (1а.к.)-5' и модифицированная LC-интеин (1а.к.)-3'), и праймер Set 3, который вводит три аминокислоты (9 пар оснований) у обоих концов интеина (HC-интеин (3а.к.)-5' и модифицированная LC-интеин (3а.к.)-3').
Программа проведения ПЦР была аналогична программе, описанной выше. PCR-продукты подвергали гель-очистке с использованием набора для гель-экстракции (Qiaquick Gel Extraction kit) (Qiagen). Эти продукты использовали в качестве праймеров в последующей серии реакций.
Три серии реакций ПЦР проводили для генерирования гибрида, включающего тяжелую цепь D2E7 и интеин, между которыми находятся 0, 1 или 3 аминокислоты экстеина. В качестве матрицы для этих реакций служила ДНК тяжелой цепи D2E7. Во всех реакциях в качестве 3'-праймеров были использованы ПЦР-продукты, описанные выше соответственно, а в качестве 5'-праймера были использована HC-Srfl-5'. В этих реакциях была использована ДНК-полимераза Pful.
ПЦР проводили в соответствии со следующей программой:
ПЦР-продукт подвергали гель-очистке с использованием набора для гель-экстракции (Qiaquick Gel Extraction kit) (Qiagen). Этот продукт использовали в качестве праймеров в последующей серии реакций.
Три серии реакций ПЦР проводили для генерирования гибрида, включающего тяжелую цепь D2E7 - интеин - легкую цепь D2E7, между которыми находятся 0, 1 или 3 аминокислоты экстеина. В качестве матрицы для этих реакций служила ДНК легкой цепи D2E7. Во всех реакциях в качестве 5'-праймеров были использованы ПЦР-продукты, непосредственно описанные выше соответственно, а в качестве 3'-праймера была использована LC-BamHI-3'. В этих реакциях была использована ДНК-полимераза Pful.
ПЦР проводили в соответствии со следующей программой:
При электрофорезе на геле полученный ПЦР-продукт был размазанным и неплотно рассеянным по гелю. Продукты этих реакций были непосредственно использованы в последнем раунде ПЦР в качестве матрицы, а HC-Srfl-5' и LC-BamHI-3' использовали в качестве праймеров. В этих реакциях была использована ДНК-полимераза Pful. Программа проведения ПЦР была аналогична программе, описанной выше. ПЦР-продукты подвергали гель-очистке с использованием набора для гель-экстракции (Qiaquick Gel Extraction kit) (Qiagen).
Очищенные ПЦР-продукты, описанные выше, субклонировали в pCR-Bluntll-TOPO (Invitrogen) с помощью набора для ПЦР-клонирования (Zero Blunt TOPO PCR Cloning Kit, Invitrogen). Затем клоны секвенировали для подтверждения того, что данные конструкции имеют предполагаемые последовательности нуклеиновой кислоты. Нужные клоны были обнаружены в продукте каждого типа. Кластер “тяжелая цепь D2E7 - интеин - легкая цепь D2E7” вырезали из pCR-Bluntll-TOPO с использованием Srfl и Notl, и затем субклонировали в вектор pTT3, рестриктированной теми же ферментами, и подвергали гель-очистке.
Были созданы три экспрессионные конструкции для гибрида “тяжелая цепь D2E7 - интеин - легкая цепь D2E7” с использованием интеина P. horikoshii, а именно: pTT3-HcintLC-p.hori (см. фиг.14, где дана плазмидная карта); pTT3-HcintLC1aa-p.hori; и pTT3-HcintLC3aa-p.hori.
Таблица 10A. Нуклеотидная последовательность pTT3-HcintLC-p.hori (SEQ ID NO:62)




Таблица 10В. Аминокислотная последовательность открытой рамки считывания в pTT3-HcintLC-p.hori (SEQ ID NO:63)
Вышеуказанным текстом/набором символов шрифта представлены последовательности:
Вектор pTT3 -тяжелая цепь - интеин - легкая цепь
Следующие 2 конструкции отличались от вышеуказанной конструкции только тем, что они включали нативные экстеиновые последовательности P. horikoshii (подчеркнута). Эти последовательности показаны от конца кодирующей области тяжелой цепи D2E7 (последние 9 пар оснований показаны красным цветом) до 5'-конца кодирующей области легкой цепи D2E7 (первые 9 пар оснований показаны [розовым] цветом на отдельной строке)
Таблица 11A. Неполная кодирующая последовательность рTT3-HcintLC1aa-p.hori (SEQ ID NO:64)

Таблица 11В. Неполная аминокислотная последовательность рTT3-HcintLC1aa-p.hori, в которой перед тяжелой цепью находятся 4 аминокислоты и четыре аминокислоты находятся позади интеина (SEQ ID NO:65)

3'-последовательность тяжелой цепи - интеин - экстеин- 5'- последовательность легкой цепи
Таблица 12A. Неполная кодирующая последовательность pTT3-HcintLC3aa-p.hori (SEQ ID NO:66)


Таблица 12В. Неполная аминокислотная последовательность рTT3-HcintLC3aa-p.hori, представляющая собой интеин и фланкирующие последовательности (SEQ ID NO:67)

3'-последовательность тяжелой цепи - интеин - экстеин - 5'- последовательность легкой цепи
Для получения конструкций A, B, E, H, I, J, K и L были использованы следующие праймеры:


Для получения конструкции A в качестве матрицы 2 использовали плазмиду pTT3 HC-int-LC P.hori, и перекрывающиеся ДНК-фрагменты амплифицировали с использованием праймеров для мутагенеза YKF1 и YKR3, а также праймеров для мутагенеза YKR1 и YKF3 соответственно. ДНК-фрагмент, связывающий вышеуказанные 2 фрагмента, получали посредством ПЦР-амплификации с использованием смеси вышеуказанных 2 ПЦР-фрагментов в качестве матрицы и праймеров YKF3 и YKR3. Затем этот ПЦР-фрагмент разрезали рестрикционными ферментами EcoR I и Not I и клонировали в конструкцию pTT3-HC-int-LC-P.hori, разрезанную теми же самыми рестрикционными ферментами.
Конструкцию B получали так же, как и конструкцию А, за исключением того, что вместо праймеров YKF1 и YKR1 использовали праймеры для мутагенеза YKF2 и YKR2 и в качестве ПЦР-матрицы вместо плазмиды pTT3-HC-int-LC-P.hori использовали плазмиду pTT3-HC-int-LC-1aa P.hori, а в качестве остова для клонирования использовали вектор pTT3-HC-int-LC-P.hori.
Для получения конструкции E, ДНК-фрагмент 2 амплифицировали с использованием плазмиды pTT3-HC-int-LC-1aa-P.hori в качестве матрицы и с использованием праймера YKF4 и праймера для мутагенеза YKR4. Этот ПЦР-фрагмент разрезали ферментами Sac II и Mfe II и клонировали в плазмиду pTT3-HC-int-LC-P.hori, разрезанную теми же самыми рестрикционными ферментами.
Для получения конструкции H в качестве матрицы использовали плазмиду pTT3-HC-int-LC-P.hori и перекрывающиеся фрагменты амплифицировали с использованием праймера для мутагенеза YKF5 и праймера YKR3 для одного фрагмента, а также праймера F3 и праймера для мутагенеза R5 для другого фрагмента. Второй раунд ПЦР-амплификации проводили с использованием вышеуказанных 2 фрагментов в качестве матриц и с использованием праймеров YKF3 и YKR3. Этот фрагмент гидролизовали рестрикционными ферментами EcoR I и Not I и клонировали в конструкцию pTT3-HC-int-LC-P.hori, разрезанную теми же самыми ферментами.
Для получения конструкции J в качестве матрицы 2 использовали плазмиду pTT3-HC-int-LC-P.hori и перекрывающиеся фрагменты амплифицировали с использованием праймера для мутагенеза YKF13 и праймера YKR3 для одного фрагмента, а также праймера F3 и праймера для мутагенеза R13 для другого фрагмента. Второй раунд ПЦР-амплификации проводили с использованием вышеуказанных 2 фрагментов в качестве матриц и с использованием праймеров YKF3 и YKR3. Этот фрагмент гидролизовали рестрикционными ферментами EcoR I и Not I и клонировали в конструкцию pTT3-HC-int-LC-P.hori, разрезанную теми же самыми ферментами.
Для получения конструкции K в качестве матрицы 2 служила pTT3-HC-int-LC-P.hori. Перекрывающиеся фрагменты амплифицировали с использованием праймера для мутагенеза YKF14 и праймера YKR3 для одного фрагмента, а также праймера F3 и праймера для мутагенеза R14 для другого фрагмента. Второй раунд ПЦР-амплификации проводили с использованием вышеуказанных 2 фрагментов в качестве матриц и с использованием праймеров YKF3 и YKR3. Этот фрагмент гидролизовали рестрикционными ферментами EcoR I и Not I и клонировали в конструкцию pTT3-HC-int-LC-P.hori, разрезанную теми же самыми ферментами.
Для получения конструкции L в качестве матрицы 2 использовали плазмиду pTT3-HC-int-LC-P.hori и перекрывающиеся фрагменты амплифицировали с использованием праймера для мутагенеза YKF15 и праймера YKR3 для одного фрагмента, а также праймера F3 и праймера для мутагенеза R15 для другого фрагмента. Второй раунд ПЦР-амплификации проводили с использованием вышеуказанных 2 фрагментов в качестве матриц и с использованием праймеров YKF3 и YKR3. Этот фрагмент гидролизовали рестрикционными ферментами EcoR I и Not I и клонировали в конструкцию pTT3-HC-int-LC-P.hori, разрезанную теми же самыми ферментами.
Были подтверждены нуклеотидные последовательности всех конструкций. Все конструкции имели такую же последовательность, как и pTT3-HC-int-LC-P.hori, за исключением последовательностей, находящихся между последними кодонами тяжелой цепи D2E7 (кодирующими PGK) и первыми кодонами зрелой последовательности легкой цепи D2E7 (кодирующими DIQ). Последовательности в этой области, которые включают в себя интеин дикого типа или мутантный интеин в комбинации с сигнальной последовательностью дикого типа или с сигнальной мутантной последовательностью легкой цепи, представлены ниже для всех конструкций.
Таблица 13A. Неполная кодирующая последовательность конструкции А (SEQ ID NO:98)


Таблица 13В. Неполная аминокислотная последовательность, представляющая собой интеин и фланкирующие последовательности в конструкции А (SEQ ID NO:99)

Таблица 14A. Неполная кодирующая последовательность в конструкции В (SEQ ID NO:100)

Таблица 14В. Неполная аминокислотная последовательность в конструкции В (SEQ ID NO:101)

Таблица 15A. Неполная кодирующая последовательность в конструкции Е (SEQ ID NO:102)

Таблица 15В. Неполная аминокислотная последовательность в конструкции Е (SEQ ID NO:103)

Таблица 16A. Неполная кодирующая последовательность в конструкции Н (SEQ ID NO:104)

Таблица 16В. Неполная аминокислотная последовательность в конструкции Н (SEQ ID NO:105)

Таблица 17A. Неполная кодирующая последовательность в конструкции J (SEQ ID NO:106)

Таблица 17В. Неполная аминокислотная последовательность в конструкции J (SEQ ID NO:107)

Таблица 18A. Неполная кодирующая последовательность в конструкции К (SEQ ID NO:108)

Таблица 18В. Неполная аминокислотная последовательность в конструкции К (SEQ ID NO:109)

Таблица 19A. Неполная кодирующая последовательность в конструкции L (SEQ ID NO:110)

Таблица 19В. Неполная аминокислотная последовательность в конструкции L (SEQ ID NO:111)

3'-последовательность тяжелой цепи - интеин + последовательность сигнального пептида легкой цепи - зрелая последовательность легкой цепи
Для амплификации интеина VMA Saccharomyces cerevisiae (GenBank accession #AB093499), проводимой с использованием геномной ДНК в качестве матрицы и с использованием ДНК-полимеразы Pfu-I Hi Fidelity (Stratagene), были получены нижеследующие олигонуклеотиды. Геномную ДНК получали из культуры Saccharomyces cerevisiae с помощью набора Yeast-Geno-DNA-Template kit (G Biosciences, cat. #786-134).
Интеин VMA Sсe 5': TGCTTTGCCAAGGGTACCAATGTTTT (SEQ ID NO:112)
Интеин VMA Sсe 3': ATTATGGACGACAACCTGGTTGGCAA (SEQ ID NO:113)
ПЦР проводили в соответствии со следующей программой:
Для продуцирования 0 а.к.-, 1 а.к.- или 3 а.к.-вариантов интеина, которые получали так же, как интеины для конструкций P. horikoshii, в качестве матрицы использовали ПЦР-продукт, а в качестве праймеров использовали нижеследующие пары праймеров. В этой реакции использовали ДНК-полимеразу Pfu-I Hi Fidelity (Stratagene).

ПЦР проводили с использованием программы, описанной выше. ПЦР-продукт реакции каждого типа субклонировали в pCR-Bluntll-TOPO (Invitrogen) и вставку каждого типа секвенировали и подтверждали ее правильность.
Для получения гибрида “тяжелая цепь D2E7 - интеин- легкая цепь D2E7” посредством гомологичной рекомбинации в конструкцию pTT3-HcintLC Р. horikoshii, введенную в E. coli, конструировали олигонуклеотидные праймеры. Для конструирования 40 пар оснований между ПЦР-генерированным вектором (содержащим вектор pTT3, области тяжелой и легкой цепей, но не содержащим интеин P. horikoshii) и вставкой интеина VMA, две ДНК могут быть смешаны и трансформированы в E.coli в условиях, не способствующих лигированию, что будет приводить к гомологичной рекомбинации в E.coli двух фрагментов в pTT3-HC-VMAint-LC в 0 а.к.-, 1 а.к.- и 3 а.к.-вариантах.
Праймеры для гомологичной рекомбинации интеина VMA:
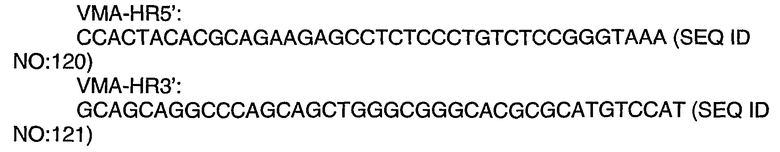
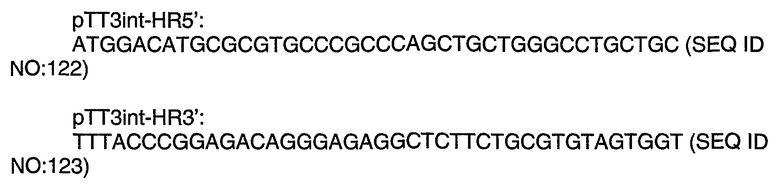
ПЦР для интеина проводили в соответствии со следующей программой, в которой использовали ДНК-полимеразу Pfu-I Hi Fidelity (Stratagene):
ПЦР для вектора проводили в соответствии со следующей программой, в которой использовали смесь ДНК-полимераз Platinum Taq Hi Fidelity Supermix (Invitrogen):
Для осуществления гомологичной рекомбинации интеина VMA в pTT3-HcintLC была использована нижеследующая стратегия. ПЦР-продукты подвергали гель-очистке и каждый продукт элюировали в 50 мкл элюирующего буфера с использованием набора для гель-экстракции Qiaquick Gel Extraction kit (Qiagen). 3 мкл векторного ПЦР-продукта перемешивали в пробирке Эппендорфа и добавляли 3 мкл нужного ПЦР-продукта интеина VMA (0-a.к.-, 1 а.к-. или 3 а.к.-варианта в отдельных пробирках). Клетки E.coli трансформировали каждой из этих смесей, а затем клетки высевали на планшеты с LB + ампициллином и инкубировали в течение ночи при 37оС. Колонии культивировали до получения 2 мл-культур, а затем получали плазмидную ДНК с помощью наборов Wizard Prep (Promega), после чего ее анализировали путем гидролиза рестрикционными эндонуклеазами и подвергали электрофорезу в агарозном геле. Клоны, обнаруживающие нужный характер рестрикции, анализировали на последовательность ДНК.
Были созданы три экспрессионные конструкции для гибрида “тяжелая цепь D2E7 - интеин - легкая цепь D2E7” с использованием интеина VMA S. cerevisiae: pTT3-Hc-VMAint-LC-0aa; pTT3-Hc-VMAint-LC-1aa; и pTT3-Hc-VMAint-LC-3aa. См. также фиг. 15 для плазмидной карты.
Таблица 20. Полноразмерная последовательность гибрида “плазмиды pTT3 - тяжелая цепь D2E7 - интеин - легкая цепь D2E7” (SEQ ID NO:124)



Вектор pTT3 -тяжелая цепь - интеин - легкая цепь
Нижеследующие конструкции отличаются от вышеуказанной конструкции только тем, что они включали нативные экстеиновые последовательности S. cerevisiae (показана [синим]). Эти последовательности показаны от конца кодирующей области тяжелой цепи D2E7 (последние 9 пар оснований показаны [красным] цветом) до 5'-конца кодирующей области легкой цепи D2E7 (первые 9 пар оснований показаны [розовым] цветом).
Таблица 21. Неполная кодирующая последовательность pTT3-HC-VMAint-LC-1aa (SEQ ID NO: 125)


3'-последовательность тяжелой цепи - интеин - экстеин - 5'-последовательность легкой цепи
Таблица 22. pTT3-HC-VMAint-LC-3aa (SEQ ID NO:126)

3'-последовательность тяжелой цепи - интеин - экстеин - 5'-последовательность легкой цепи
Интеин DnaE штамма PCC6803 Synechocystis spp.: синтез, ПЦР-амплификация и клонирование
Интеин DnaE штамма PCC6803 Senechocestis spp. представляет собой интеин, отщепляемый в природных условиях (NCBI accession # S76958 и S75328). Авторами настоящего изобретения были присоединены N-концевые и С-концевые половины этого интеина в виде одной открытой рамки считывания путем искусственного синтеза. Кодирующая последовательность нужной последовательности белка была оптимизирована по кодонам для экспрессии в клетках СНО (www.geneart.com). Полученная нуклеотидная последовательность представлена в таблице 23.
Таблица 23. Ssp-Di (кодирующая последовательность, оптимизированная для экспрессии в Cricetulus griseus) (См. также SEQ ID №№:127 и 128)

Для амплификации интеина DnaE штамма PCC6803 Synechocystis spp., проводимой с использованием вышеуказанной синтетической ДНК в качестве матрицы и с использованием смеси Platinum Taq Hi Fidelity Supermix (Invitrogen), были получены нижеследующие олигонуклеотиды. С помощью этих праймеров также вводили экстеиновые последовательности для генерирования 0 а.к.-, 1 a.к.- и 3 а.к.-вариантов, а также последовательности для гомологичной рекомбинации ПЦР-продукта в вектор pTT3-HcintLC, полученный на основе интеина VMA S. cerevisiae:

ПЦР проводили в соответствии со следующей программой:
Для осуществления гомологичной рекомбинации оптимизированного по кодонам интеина DnaE штамма PCC6803 Synechocystis spp. в pTT3-HcintLC была использована нижеследующая стратегия. ПЦР-продукты подвергали гель-очистке и каждый продукт элюировали в 50 мкл элюирующего буфера (с использованием набора для гель-экстракции Qiaquick Gel Extraction kit (Qiagen)). 2 мкл векторного ПЦР-продукта (такого же продукта, который был использован для гомологичной рекомбинации с помощью интеина VMA) перемешивали в пробирке Эппендорфа и добавляли 3 мкл нужного ПЦР-продукта интеина DnaE штамма PCC6803 Synechocystis spp. (0-a.к.-, 1 а.к-. или 3 а.к.-варианта в отдельных пробирках). Клетки E.coli трансформировали нуклеиновыми кислотами, а затем высевали на планшеты с LB + ампициллином и инкубировали в течение ночи при 37оС. Колонии культивировали до получения 2 мл-культур, а затем получали ДНК с помощью набора Wizard Prep (Promega), после чего ее анализировали путем гидролиза рестрикционными эндонуклеазами и подвергали электрофорезу в агарозном геле. Клоны, обнаруживающие нужный характер рестрикции, анализировали на последовательность ДНК для подтверждения присутствия нужных последовательностей.
Были созданы три экспрессионные конструкции для гибрида “тяжелая цепь D2E7 - интеин - легкая цепь D2E7” с использованием интеина DnaE штамма PCC6803 Synechocystis spp.: pTT3-Hc-Ssp-GA-int-LC-0aa (см. фиг. 16 для плазмидной карты); pTT3-Hc-Ssp-GA-int-LC-1aa; и pTT3-Hc- Ssp-GA-int-LC-3aa.
Таблица 24. Полноразмерная последовательность гибрида “плазмида pTT3 - тяжелая цепь D2E7 - Ssp - GA-интеин - легкая цепь D2E7 (SEQ ID NO:135)



Вектор pTT3 -тяжелая цепь - интеин - легкая цепь
Нижеследующие конструкции отличаются от вышеуказанной конструкции только тем, что они включали нативные экстеиновые последовательности штамма PCC6803 Synechocystis spp. (показаны синим). Эти последовательности показаны от конца кодирующей области тяжелой цепи D2E7 (последние 9 пар оснований показаны [красным] цветом) до 5'-конца кодирующей области легкой цепи D2E7 (первые 9 пар оснований показаны розовым цветом).
Таблица 25. pTT3-HC-Ssp-GA-int-LC-1aa, соответствующая часть кодирующей последовательности (SEQ ID NO:136)

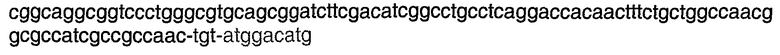
Вектор pTT3 - тяжелая цепь - интеин - легкая цепь
Таблица 26. pTT3-HC-Ssp-GA-int-LC-3aa, соответствующая часть кодирующей последовательности (SEQ ID NO:137)

Вектор pTT3 - тяжелая цепь - интеин - легкая цепь
Кроме того, в таблицах 8А-8С представлены соответствующие последовательности для гибридного белка “D2E7-интеин”, экспрессирующий вектор и кодирующая последовательность, полученная с использованием мутированного (с заменой серина на треонин) интеина Pol GBD Pyrococcus Ssp.
Таблица 8A. Кодирующая последовательность гибридного белка “D2E7-интеин” (SEQ ID NO:48)


Таблица 8В. Аминокислотная последовательность гибридной конструкции белка “D2E7-интеин” (SEQ ID NO:49)

Таблица 8С. Полноразмерная нуклеотидная последовательность экспрессирующего вектора для гибридной конструкции “D2E7-интеин” (SEQ ID NO:50)




Таблица 9. Аминокислотная последовательность нативного интеина Pol Psp-GBD с ограниченной информацией о фланкирующих последовательностях (NCBI, рег. № AAA67132.1) (SEQ ID NO:51)

“/” означает границу сплайсинга, подчеркнутые аминокислоты представляют собой последовательности интеина, а остальная последовательность представляет собой информацию о последовательности экстеина.
Пример 2. Конструирование полипротеиновых последовательностей иммуноглобулина и векторов с hedgehog-доменом аутопроцессинга Drosophila melanogaster, последовательностей C17 и C25
Другой стратегией эффективной экспрессии молекул антитела является экспрессия полипротеина, в котором hedgehog-домен расположен между тяжелой и легкой цепями и в котором последовательность hedgehog-домена и/или последовательности стыка имеют модификацию, приводящую к высвобождению белковых компонентов без присоединения холестерина к N-концевому белку. В таких конструкциях может присутствовать одна копия каждой из соответствующих тяжелой и легкой цепей, либо легкая цепь может дуплицироваться с образованием по меньшей мере двух легких цепей, либо может присутствовать множество копий тяжелой и легкой цепей, при условии что функциональная последовательность расщепления будет стимулировать разделение всех иммуноглобулиновых белков в данном полипротеине. Стратегия с использованием конкретного сайта расщепления (например, hedgehog-домена) может быть применена более чем один раз, либо может быть независимо использован каждый из множества сайтов расщепления. Таким образом, по меньшей мере на одном конце иммуноглобулина или белка, происходящего от иммуноглобулина, может присутствовать и другая последовательность протеолитического процессинга или последовательность расщепления ферментом.
Для амплификации С-концевого hedgehog-домена аутопроцессинга Drosophila melanogaster (Hh-C), последовательностей Hh-C17, усеченных последовательностей Hh-C17 (и одной последовательности с мутацией) и Hh-C25 (GenBank accession #L02793.1), проводимой с использованием геномной ДНК в качестве матрицы и смеси ДНК-полимераз Platinum Taq Hi Fidelity PCR Supermix (Invitrogen), конструировали нижеследующие олигонуклеотиды. Геномую ДНК получали из клеток D.Mel-2 дрозофилы, находящихся в замороженном сосуде (Invitrogen, cat. #10831-014).

ПЦР проводили в соответствии со следующей программой:
Для получения гибрида “тяжелая цепь D2E7 - Hh-C - легкая цепь D2E7” посредством гомологичной рекомбинации в конструкцию pTT3-HcintLC Р. horikoshii, введенную в E. coli, конструировали олигонуклеотидные праймеры. Для конструирования 40 пар оснований между ПЦР-генерированным вектором (содержащим вектор pTT3, области тяжелой и легкой цепей, но не содержащим интеин P. horikoshii) и вставками Hh-C-домена два ДНК-фрагмента смешивали и трансформировали в E.coli в условиях, не способствующих лигированию, что приводило к гомологичной рекомбинации двух фрагментов в pTT3-HC-Hh-C-LC в E.coli (в различных вариантах в зависимости от исходных ПЦР-продуктов).
Праймеры для гомологичной рекомбинации Hh-C-доменов:

Праймеры для гомологичной рекомбинации pTT3-HcintLC:

ПЦР для Hh-C-домена проводили в соответствии со следующей программой, в которой использовали ДНК-полимеразу Pfu-I Hi Fidelity (Stratagene):
ПЦР для вектора проводили в соответствии со следующей программой, в которой использовали смесь ДНК-полимераз Platinum Taq Hi Fidelity Supermix (Invitrogen):
Для осуществления гомологичной рекомбинации Hh-C-доменов в pTT3-HcintLC была применена нижеследующая стратегия. ПЦР-продукты подвергали гель-очистке и каждый продукт элюировали в 50 мкл элюирующего буфера (с использованием набора для гель-экстракции Qiaquick Gel Extraction kit (Qiagen)). 3 мкл векторного ПЦР-продукта перемешивали в пробирке Эппендорфа и добавляли 3 мкл нужного ПЦР-продукта Hint-домена (различные варианты). Клетки E.coli трансформировали продуктами ПЦР-амплификации и высевали на планшеты с LB + ампициллином, а затем инкубировали в течение ночи при 37оС и колонии культивировали до получения 2 мл-культур, после чего плазмидную ДНК экстрагировали с помощью набора Wizard Prep (Promega) и образцы ДНК анализировали путем гидролиза рестрикционными эндонуклеазами и подвергали электрофорезу в агарозном геле. Клоны, обнаруживающие нужный характер рестрикции, анализировали на последовательность ДНК для подтверждения присутствия нужной последовательности.
Было создано пять экспрессионных конструкций для экспрессии гибрида “тяжелая цепь D2E7 - Hh-C - легкая цепь D2E7” с использованием С-концевого hedgehog-домена аутопроцессинга Drosophila melanogaster: pTT3-HC-Hh-C17-LC; pTT3 - HC - Hh-C17 - SC - LC; PTT3 - HC - Hh - C17 - HN - LC; и pTT3-HC-Hh-C25-LC.
Таблица 27. Полноразмерная последовательность гибрида “плазмида pTT3-тяжелая цепь D2E7-Hh-C17-легкая цепь D2E7” (SEQ ID NO:155)



Вектор pTT3 - тяжелая цепь - Hh-C17 - легкая цепь
Нижеследующие конструкции отличались от вышеуказанной конструкции только тем, что они включали усеченную область С17, и это приводило к предотвращению переноса холестериновой активности. Эти последовательности показаны от конца кодирующей области тяжелой цепи D2E7 (последние 9 пар оснований кодирующей последовательности НС, первая строка таблицы) до 5'-конца кодирующей области легкой цепи D2E7 (первые 9 пар оснований кодирующей последовательности LС, последняя строка таблицы).
Таблица 28. Неполная кодирующая последовательность плазмиды pTT3-HC-C17-sc-LC (SEQ ID NO: 156)

3'-последовательность тяжелой цепи-Hh-C17-5'-последовательность легкой цепи
Нижеследующая конструкция отличалась от вышеуказанной конструкции pTT3-HC-C17-sc-LC только тем, что она имела мутацию в двух последних аминокислотах в hedgehog-области С17, а именно замену SC на HN (подчеркнуты). Эти последовательности показаны от конца кодирующей области тяжелой цепи D2E7 (последние 9 пар оснований кодирующей последовательности НС, первая строка таблицы) до 5'-конца кодирующей области легкой цепи D2E7 (последняя строка таблицы).
Таблица 29. Неполная кодирующая последовательность плазмиды pTT3-HC-C17-hn-LC (SEQ ID NO:157)

3'-последовательность тяжелой цепи-Hh-C17-мутация-5'-последовательность легкой цепи
В нижеследующей конструкции вместо С17 присутствует полноразмерная область С25 Hint-домена. Эти последовательности показаны от конца кодирующей области тяжелой цепи D2E7 (последние 9 пар оснований кодирующей последовательности НС, первая строка таблицы) до 5'-конца кодирующей области легкой цепи D2E7 (последние 9 пар оснований кодирующей последовательности LС, последняя строка таблицы).
Таблица 29В. Неполная кодирующая последовательность плазмиды pTT3-HC-C25 - Hint-LC (SEQ ID NO:158)
[3'-последовательность тяжелой цепи-Hh-домен C25-5'-последовательность легкой цепи]
(SEQ ID NO: 140)
Аминокислотная последовательность Hh-C25 и родственных конструкций (стрелка вниз указывает на сайт расщепления;
Пример 3. Экспрессия антитела с последовательностью узнавания протеазой TEV для протеолитического процессинга
Были получены конструкции и экспрессирующие векторы для направления экспрессии антител, специфичных к фактору некроза опухоли -α, интерлейкину-12, интерлейкину-18 и рецептору эритропоэтина, где между сегментами последовательности тяжелой и легкой цепей иммуноглобулина, которые составляют представляющее интерес антитело, присутствуют последовательности узнавания протеазой TEV. При этом предпочтительно, чтобы такие конструкции включали экспрессирующие векторы, содержащие главный поздний промотор аденовируса и энхансер цитомегаловируса, регулирующие транскрипцию тяжелой цепи представляющего интерес антитела, находящейся за лидерной последовательностью в той же рамке считывания. Кодирующая последовательность тяжелой цепи присоединена с сохранением рамки считывания к сайту расщепления фурином и к последовательности узнавания протеазой TEV (E-P-V-Y-F-Q-G), за которыми следует кодирующая область для протеазы TEV с делетированной областью сигнала локализации в ядре (Ceriani et al. (1998) Plant Molec Biol. 36:239), а за этой областью следует вторая последовательность узнавания протеазой TEV. Вторая последовательность узнавания протеазой TEV присоединена с сохранением рамки считывания к лидерной последовательности легкой цепи антитела, связанной с кодирующей областью представляющей интерес легкой цепи антитела и стоп-кодоном. За кодирующей областью расположен сигнал полиаденилирования. Соответствующие последовательности представлены ниже.
Таблица 1. Полноразмерная последовательность ДНК TEV-экспрессирующего вектора D2E7 (Humira/адалимумаб) (SEQ ID NO:44)



Таблица 2A. TEV-конструкция ABT-007: кодирующая последовательность для полипротеина (SEQ ID NO:32)

Таблица 2В. Аминокислотная последовательность полипротеина TEV ABT-007 (SEQ ID NO:33)


Таблица 2С. Полноразмерная последовательность TEV-экспрессирующего вектора в конструкции ABT-007 (SEQ ID NO:34)



Таблица 3A. Кодирующая последовательность для полипротеина TEV ABT-874 (J695) (SEQ ID NO:35)

Таблица 3B. Аминокислотная последовательность полипротеина TEV ABT-874 (J695) (SEQ ID NO:36)

Таблица 3С. Полноразмерная нуклеотидная последовательность TEV-экспрессирующего вектора в конструкции ABT-874 (J695) (SEQ ID NO:37)



Таблица 4A. Последовательность нуклеиновой кислоты, кодирующая полипротеин TEV EL246 GG (антитело против селектина E/L) (SEQ ID NO:38)


Таблица 4В. Аминокислотная последовательность полипротеина TEV EL246 GG (антитело против селектина E/L) (SEQ ID NO:39)

Таблица 4С. Полноразмерная нуклеотидная последовательность для вектора, экспрессирующего полипротеин TEV EL246 GG (антитело против селектина E/L) (SEQ ID NO:40)



Таблица 5A. Кодирующая последовательность для полипротеина TEV ABT-325 (SEQ ID NO:41)


Таблица 5В. Аминокислотная последовательность полипротеина TEV ABT-325 (SEQ ID NO:42)

Таблица 5C. Полноразмерная нуклеотидная последовательность вектора, экспрессирующего полипротеин TEV ABT-325 (SEQ ID NO:43)



Таблица 6A. Кодирующая последовательность для полипротеиновой конструкции LC-LC-HC D2E7 (SEQ ID NO:29)


Таблица 6B. Аминокислотная последовательность полипротеина LC-LC-HC D2E7 (SEQ ID NO:30)

Таблица 6C. Полноразмерная нуклеотидная последовательность ДНК-вектора, экспрессирующего полипротеин LC-LC-HC D2E7 (SEQ ID NO:31)




Пример 4. Экспрессия антитела в виде полипротеина с конструкцией, содержащей внутренний отщепляемый сигнальный пептид
В других вариантах настоящее изобретение относится к получению кодирующих последовательностей и экспрессирующих векторов, а также к способам экспрессии антитела. Первичная экспрессионная конструкция содержит полипротеин с внутренним отщепляемым сигнальным пептидом, экспрессия и последующее отщепление которого приводит к образованию многоцепьевой молекулы антитела (например, молекулы, имеющей две цепи).
Таблица 7A. Кодирующая последовательность для конструкции, содержащей внутренний отщепляемый сигнальный пептид D2E7 (SEQ ID NO:45)

Таблица 7В. Аминокислотная последовательность полипротеина, содержащего внутренний отщепляемый сигнальный пептид D2E7 (SEQ ID NO:46)

Таблица 7С. Полноразмерная последовательность ДНК-вектора, экспрессирующего полипротеин, содержащий внутренний отщепляемый сигнальный пептид D2E7 (SEQ ID NO:47)



Материалы и методы:
Трансфекцию описанных конструкций в клетки 293-6E осуществляли следующим образом. Используемыми клетками являются клетки HEK293-6E в экспоненциальной фазе роста (0,8 - 1,5×106 клеток/мл), которые подвергали пересеву в культуре менее чем 30 раз, а затем полученные культуры инокулировали в свежую культуральную среду до концентрации 3×105 клеток/мл через каждые три или четыре дня. Культуральной средой является экспрессионная cреда FreeStyleTM 293 (GIBCOTM Cat. No. 12338-018, Invitrogen, Carlsbad, CA), в которую были добавлены 25 мкг/мл генетицина (G418)(GIBCOТМ Cat. No.10131-027) и 0,1% Pluronic F-68 (поверхностно-активное вещество, GIBCOТМ, Cat. No. 24040-032). Средой для трансфекции является экспрессионная среда FreeStyleТМ (GIBCOТМ Cat. No. 12338-018) с конечной концентрацией 10 мМ буферного раствора HEPES/мл среды (GIBCOТМ Cat. No. 15630-080). Для трансфекции добавляли выбранный ДНК-вектор до концентрации 1 мкг (тяжелая цепь + легкая цепь)/мл рассматриваемого антитела, в котором были сделаны замены исходя из результатов экспериментов по оптимизации. В качестве медиатора трансфекции добавляли ПЭИ (полиэтиленимин), линейный, 25 кДа, 1 мг/мл стерильного маточного раствора, pH 7,0 (Polysciences, Inc., Warrington, PA) в отношении ДНК:ПЭИ= 1:2. В качестве питательной среды использовали триптоновую среду N1 (порошок TN1 от Organotechnie France, Cat No. 19554, поставляемый TekniScience Inc. Tel# 1-800-267-9799). 5% мас./об. маточного раствора в среде для экспрессии клеток 293 FreeStyleTM добавляли до конечной концентрации 0,5%. Обычно эксперименты проводили на стандартном лабораторном оборудовании. Клетки подсчитывали на системе для подсчета клеток Cedex Cell Counting System (Innovatis, Bielefeld, Germany).
Каждую процедуру лабораторной трансфекции проводили в 125-миллилитровой колбе Эрленмейера следующим образом. Аликвоту 20 мл свежей культуральной среды инокулировали 1×106 жизнеспособных клеток/мл. (Примечание: для повышения объема культура должна составлять 20-25% от номинальной емкости сосуда, например, 100 мл культуры в 500 мл-колбе). Затем культуры помещали в инкубатор при 37оС в атмосфере с повышенной влажностью с 5% CO2 при скорости вращения 130 об/мин.
Препарат комплекса ДНК-ПЭИ получали путем нагревания среды для трансфекции до 37оС в водяной бане с последующим оттаиванием замороженного маточного раствора ПЭИ и растворов ДНК при комнатной температуре (и хранением при -20°С). Количество используемых ДНК и ПЭИ вычисляли исходя из общего объема трансфицируемой культуры. Для 20 мл культуры с 2,5 мл комплекса ДНК/ПЭИ и 2,5 мл Tn1 требуется всего 25 мкг ДНК и 50 мкг ПЭИ. Комплексы ДНК:ПЭИ (например, для десяти трансфекций) получали путем объединения 12,5 мл среды для трансфекции в пробирке А, в которую был добавлен раствор, содержащий выбранную векторную ДНК до конечной концентрации 10 мкг/мл, и 12,5 мл смеси среды для трансфекции и ПЭИ (20 мкг/мл, конечная концентрация). ПЭИ-смесь интенсивно перемешивали примерно 10 секунд, а затем смешивали с раствором ДНК. После объединения смесей ПЭИ и ДНК указанную комбинацию интенсивно перемешивали в течение 10 секунд. Затем эту смесь оставляли на 15 минут (но не более чем на 20 минут) при комнатной температуре. Затем добавляли 2,5 мл раствора комплекса ДНК:ПЭИ на 20 мл клеток HEK-6E. Примерно через 20-24 часа после трансфекции в каждую колбу добавляли 5% добавки TN1 до конечной концентрации 0,5%.
Плотность и жизнеспособность клеток определяли на 4-й и 7-й дни. Клеточные осадки (2 мл-аликвоты культуры) собирали на 4-й день для проведения Вестерн-анализа и Нозерн-блот-анализа. Осадки замораживали при -80°С и хранили до проведения анализов. Через 7 дней после трансфекции клетки собирали путем центрифугирования при 1000 об/мин (10 минут) и супернатанты фильтровали на бумаге для предварительной фильтрации и на системе 0,22 мкм-фильтров Corning CA. Образцы супернатантов также хранили при -80°С, после чего их анализировали, например, с помощью ELISA.
Для Нозерн-блот-анализа полноразмерную РНК выделяли из временно трансфицированных клеток 293-6E следующим образом. Замороженный клеточный осадок оттаивали на льду. РНК очищали с помощью мини-набора Qiagen Rneasy Mini Kit (Qiagen, cat. #74104) в соответствии с инструкциями производителя.
Формальдегидный/агарозный гель получали следующим образом. 2 грамма агарозы (Ambion, cat. #9040) кипятили в 161,3 мл дистиллированной воды. Затем добавляли 4 мл 1 M MOPS (морфолинопропансульфоновой кислоты), рН 7,0, 1 мл 1 M NaOAc, 0,4 мл 0,5 M EDTA и смесь охлаждали до 60°С. После этого добавляли 33,3 мл 37% формальдегида (J.T. Baker, cat #2106-01) и расплавленный агарозный раствор слегка перемешивали. Затем гель выливали и помещали в вытяжной шкаф для отверждения.
Буфер для электрофореза получали путем смешивания 30 мл 1 M MOPS, pH 7,0, 7 мл 1 M NaOAc, 3 мл 0,5M EDTA и ДЭПК-(диэтилпирокарбонат)-обработанной dH2O, рН до 1,5.
РНК-образцы получали путем смешивания 3 частей формальдегидного загрузочного красителя (Ambion, cat. #8552) с 1 частью РНК. На одну дорожку загружали 3-5 мкг РНК. В качестве маркеров молекулярной массы РНК использовали РНК-лэддер в 0,5-10 т.п.н. (Invitrogen, cat. #15623-200). Образцы нагревали при 65°С в течение 5 минут для денатурации и охлаждали на льду. Затем в каждый образец добавляли 0,5 мкл 10 мкг/мкл этидийбромида (Pierce, cat. #17898) Каждый образец быстро центрифугировали для осаждения жидкости.
Гель-электрофорез осуществляли следующим образом. Формальдегидный/агарозный гель покрывали буфером для электрофореза, а затем загружали образцы и проводили электрофорез при 150 В в течение 2 часов в вытяжном шкафу. Полосы наблюдали путем просвечивания ультрафиолетовым излучением и фотографировали в непрерывном режиме съемки.
Капиллярный перенос осуществляли путем смачивания геля ДЭПК-обработанной dH2O с несколькими заменами в течение пяти минут для удаления формальдегида. Затем гель погружали в 50 мM NaOH, 10 мM NaCl на 20 минут при комнатной температуре для дополнительной денатурации любой двухцепочечной РНК. Этот гель ополаскивали один раз ДЭПК-обработанной dH2O, а затем для нейтрализации погружали на 20 минут в 20×SSC (175,3 г NaCl; 88,2 г цитрата натрия; pH доводили до ~7,0 путем добавления 10M NaOH и объем доводили до 1 л) при комнатной температуре. Hybond-N+-мембрану (Amersham Biosciences, cat #RPN303B) погружали в ДЭПК-обработанную dH2O для смачивания и обрезали до размера геля. 3M фильтровальную бумагу (Whatman cat#3030917) обрезали до размера геля и мембраны. Систему для переноса создавали путем наложения слоя 3M бумаги на твердый носитель поверх резервуара с 20×SSC, так, чтобы эта бумага подобно фитилю пропускала 20×SSC через слои, находящиеся на поверхности. Гель помещали на этот фитиль, Hybond-N+-мембрану, 3 листа 3M бумаги, обрезанные по размеру, и толстую пачку бумаги для гель-блотов (Gel Blot Paper) (Schleicher & Schuell, cat. #10427920). Поверх этой пачки помещали плоский носитель и ставили груз (обычно литровую бутылку с водой), если это необходимо, для гарантии эффективного капиллярного переноса. Все открытые части резервуара покрывали пластиковой пленкой для предотвращения выпаривания. Эту систему оставляли на ночь при комнатной температуре для осуществления переноса. Затем систему для переноса разбирали и блот погружали в 6×SSC для удаления всей агарозы. Мембрану оставляли для сушки на воздухе и облучали ультрафиолетом для перекрестного сшивания блота.
ДНК-зонды, используемые в качестве матриц, представляют собой кодирующую область для тяжелой и легкой цепей D2E7. 100 нг нужной матрицы метили щелочной фосфатазой с использованием набора реагентов для прямого мечения (AlkPhos Direct Labeling Reagents kit)(система для прямого мечения щелочной фосфатазой, Amersham Biosciences, cat. #RPN3680) в соответствии с инструкциями производителя. Стадии предварительной гибридизации и гибридизации осуществляли с использованием того же самого набора для мечения (содержащего буфер для гибридизации). Мембраны подвергали предварительной гибридизации в течение по меньшей мере 1 часа при 65°С в печи для гибридизации, а затем зонд кипятили и добавляли непосредственно в буфер/блот для предварительной гибридизации. Гибридизацию проводили в течение ночи при 65°С в печи для гибридизации. Раствор для гибридизации декантировали и мембрану быстро промывали 2×SSC для удаления раствора для гибридизации, а затем два раза промывали 2×SSC, 0,1% ДСН при 65°С по 15 минут каждым и, наконец, два раза промывали 0,1×SSC, 0,1% ДСН при 65°С, по 15 минут каждым. Для визуализации полос на мембране применяли хемилюминесценцию. На блоты наносили детектирующий реагент CDP-Star (при этом наблюдалась зависимая от щелочной фосфатазы фотохимическая реакция, вызываемая 1,2-диоксетановым субстратом Amersham Biosciences, cat. #RPN3682) в течение 5 минут при комнатной температуре. Из блотов удаляли избыточное количество реагента, а затем их помещали в пластиковые изоляторы. Блоты экспонировали с пленкой Kodak Biomax MR (рентгеновской пленкой, Kodak, cat. #8952855) с начальной экспозицией 10 секунд, которую затем доводили до 10 минут. Пленки проявляли с использованием процессора Kodak M35A X-OMAT (проявителя для рентгеновских пленок/процессора для обработки рентгеновских пленок).
Образцы клеточного осадка для вестерн-блот-анализа получали следующим образом. Для анализа внутриклеточной экспрессии антитела клетки подвергали лизису в лизирующем буфере NP 40 (50 мM трис-HCl, pH 7,5, 150 нM NaCl, 1% NP40 (октилфенолполиэтиленгликолевый эфир), 5 мМ BME и смесь ингибиторов протеазы III) при инкубировании на льду в течение 10 минут. Фракции мембран и нерастворимых белков собирали путем центрифугирования при 16000 об/мин в течение 30 минут на микроцентрифуге. Для анализа геля использовали супернатант, взятый из растворимой внутриклеточной или цитозольной фракции, с добавлением загрузочного ДСН-буфера с ДТТ. Клеточный осадок суспендировали в равном объеме буфера для лизиса и добавляли ДТТ-содержащий буфер для загрузки в ДСН-гель. Образцы супернатантов культуры для проведения вестерн-блот-анализа получали следующим образом. Супернатанты культуры либо концентрировали на устройстве Centricon Ultra (устройство для ультрафильтрации, Millipore) с отсечкой MW 30000 дальтон либо непосредственно использовали для вестерн-блоттинга. Для иммуноблоттинга (вестерн-анализа) образцы разделяли на 4-12% бис-трис (полиакриламидных)-гелях NUPAGE и переносили на ПВДФ-мембрану стандартными методами. Эти мембраны инкубировали в течение 1 часа в блокирующем растворе (PBS с 0,05% твином 20 (полиоксиэтиленсорбитанмонолаурат) и 5% сухим молоком), после чего промывали, инкубировали с ПХ-конъюгированным поликлональным кроличьем антителом против человеческого IgG или ПХ-конъюгированным поликлональным кроличьем антителом против человеческой легкой цепи каппа, полученным от DakoCytomation (Denmark), при разведении 1:1000 в буфере PBST, а затем снова промывали при комнатной температуре в PBST с тремя заменами. Для детектирования использовали детектирующую систему ЭХЛ + вестерн-блот-анализ (хемилюминесцентная и хемифлуоресцентная детекция), поставляемую фирмой GE/Amersham Biosciences (Piscataway, NJ).
ELISA-анализы проводили стандартными методами с использованием козьего антитела против человеческих IgG, UNLB и ПХ-конъюгированного козьего антитела против человечесих IgG, поставляемых Southern Biotech (Birmingham, AL), 2% молока в PBS, используемом в качестве буфера для блоттинга, и К-синего (3,3', 5,5'-тетраметилбензидина и пероксида водорода (H2O2, Neogen, Lansing, МI) в качестве субстрата. Планшеты считывали на микропланшет-ридере Spectramax при начальной длине волны 650 нм и эталонной длине волны 490 нм.
Секретированное антитело подвергали аффинной очистке стандартными методами с использованием сфер с белком А-агарозой, поставляемых фирмой Invitrogen (Carlsbad, CA), иммунологически чистого (lmmuno Pure (A)) буфера для связывания с IgG, поставляемого Pierce, PBS, pH 7,4, в качестве промывочного буфера и 0,1 M уксусной кислоты/150 мМ NaCl, pH 3,5, в качестве элюирующего буфера (нейтрализованного с использованием 1 M трис, pH 9,5).
Определение молекулярной массы интактного антитела. Молекулярные массы образцов интактного D2E7, продуцированного с использованием конструкции pTT3-HC-int-LC-P.hori, анализировали с помощью ЖХ-МС. Для обессоливания и загрузки образцов на масс-спектрометр Q Star Pulsar i (Applied Biosystems, SN K1820202) использовали капиллярную ВЭЖХ-систему 1100 (Agilent SN DE 14900659) с белковой микроловушкой (Michrom Bioresources, Inc. cat. 004/25109/03). Образцы элюировали в градиенте буфера A (0,08% FA, 0,02% TFA в ВЭЖХ-воде) и буфера B (0,08% FA и 0,02% TFA в ацетонитриле) при скорости потока 50 мкл/мин в течение 15 минут.
Определение молекулярной массы легкой и тяжелой цепей. Образцы нативного D2E7, продуцированного с использованием конструкции pTT3-HC-int-LC-P.hori, анализировали с помощью ЖХ-МС. Восстановление дисульфидных связей, которые соединяют легкие и тяжелые цепи, проводили в 20 мМ ДTT при 37оС в течение 30 минут. Для отделения легких цепей от тяжелых цепей и для их загрузки на масс-спектрометр Q Star Pulsar i (Applied Biosystems, SN K1820202) использовали капиллярную ВЭЖХ-систему 1100 (Agilent SN DE 14900659) с колонкой PLRP-S (Michrom Bioresources, Inc., 8 мкм, 4000 A, 1,0Ч150 мм, P/N 901-00911-00). Колонку нагревали при 60°С. Элюирование образцов проводили в ВЭЖХ-градиенте буфера A (0,08% FA, 0,02% TFA в ВЭЖХ-воде) и буфера B (0,08% FA и 0,02% TFA в ацетонитриле) при скорости потока 50 мкл/мин в течение 60 минут.
Рестрикционные эндонуклеазы поставлялись компанией New England Biolabs (Beverly, MA). Для клонирования использовали специально приготовленные олигонуклеотиды, ДНК-полимеразы, ДНК-лигазы и штаммы E.coli, поставляемые компанией Invitrogen (Carlsbad, CA). Смесь ингибиторов протеазы III поставлялась Calbiochem (La JoIIa, CA). Для выделения и очистки ДНК использовали продукты компании Qiagen (Valencia, CA).
Заявления, относящиеся к ссылочному материалу и вариантам
Все ссылочные материалы, упомянутые в настоящей заявке, например патентные документы, включая опубликованные или выданные патенты или их эквиваленты; публикации патентных заявок; неопубликованные патентные заявки и непатентная литература или другие первоисточники во всей своей полноте вводятся в настоящее описание посредством ссылки, так, как если бы они были отдельно введены в настоящее описание посредством ссылки. В случае возникновения каких-либо разногласий между цитируемыми здесь работами и описанием настоящей заявки премущество отдается последнему. Некоторые упомянутые здесь работы вводится в настоящее описание посредством ссылки для получения, например, подробной информации об источниках исходных материалов, о дополнительных исходных материалах, о дополнительных реагентах, о дополнительных способах синтеза, о дополнительных способах анализа, о дополнительных биологических материалах, о дополнительных клетках и о дополнительных способах применения настоящего изобретения.
Все упомянутые здесь патенты и публикации является показателями уровня области техники, к которой относится настоящее изобретение. Цитируемые здесь работы указывают на состояние уровня техники на момент выхода данной публикации или подачи заявки, причем считается, что эта информация может быть использована в данной заявке, если это необходимо, для исключения конкретных вариантов, которые известны специалистам в данной области. Так, например, что касается заявленных и рассматриваемых здесь композиций, то следует отметить, что те соединения, которые были известны еще до появления настоящего изобретения и которые могут быть получены специалистами в данной области, включая соединения, описанные в цитируемых здесь работах, не включены в заявленные здесь композиции.
Любое приложение или приложения к настоящей заявке вводятся в настоящее описание посредством ссылки как часть данного описания и/или графического материала.
Используемые здесь термины “включать", “включает в себя", “включенный" или “ включающий в себя" могут быть интерпретированы как термины, определяющие присутствие установленных признаков, элементов, стадий или компонентов, но не исключают присутствия или добавления одного или нескольких других признаков, элементов, стадий, компонентов или их групп. Таким образом, эти используемые здесь термины являются синонимами терминов “содержащий”, “имеющий” или “характеризующийся” и имеют инклюзивный смысл или допускают возможность каких-либо других добавлений. Используемый здесь термин “состоящий из” следует понимать как исключающий присутствие каких-либо других элементов, стадий или ингредиентов и т.п., не определенных в формуле изобретения. Используемый здесь термин “состоящий, по существу, из” не исключает присутствия материалов или стадий, которые не оказывают значительного влияния на основные и новые свойства заявленного объекта (например, активного ингредиента). В каждом рассматриваемом случае любой из терминов “ включающий в себя”, “состоящий, по существу, из” и “состоящий из” может быть заменен любым из двух других терминов, и это означает, что определяемый ими объект представляет собой отдельные варианты осуществления изобретения и/или определяет отдельные объемы изобретения, которые необязательно должны быть одинаковыми. Настоящее изобретение, проиллюстрированное в настоящем описании, может быть практически успешно осуществлено в отсутствие какого-либо элемента или элементов либо ограничения или ограничений, конкретно не описанных в настоящей заявке.
Все описанные здесь интервалы, например интервалы температур, интервалы времени, интервалы количеств или концентраций или другие интервалы указанных величин и т.п., и кроме того, все промежуточные интервалы и субинтервалы, а также все отдельные величины, входящие в данные интервалы, входят в объем настоящего изобретения. Настоящее изобретение не ограничивается описанными здесь вариантами его осуществления, включая любые указанные в описании графические или репрезентативные материалы, которые приводятся в качестве иллюстративных и неограничивающих примеров. При этом следует отметить, что любые субинтервалы или отдельные величины, входящие в интервал или субинтервал, указанный в настоящем описании, могут быть исключены из соответствующего пункта формулы изобретения.
Настоящее изобретение было описано со ссылками на различные конкретные и/или предпочтительные варианты и способы осуществления изобретения. Однако следует отметить, что в него могут быть внесены различные изменения и модификации, не выходящие за рамки объема и существа изобретения. При этом очевидно, что для осуществления настоящего изобретения, описанного здесь в общих чертах, каждый специалист в данной области может использовать и другие композиции, способы, устройства, элементы устройства, материалы, процедуры и технологии, не прибегая к излишнему экспериментированию; и в настоящем изобретении могут быть применены помимо уже описанных, например, другие исходные материалы, биологические материалы, реагенты, методы синтеза, методы очистки, аналитические методы, методы оценки и биологические методы. Все известные функциональные эквиваленты ранее описанных здесь объектов (например, композиций, способов, устройств, элементов устройств, материалов, процедур и технологий и т.п.) также входят в объем настоящего изобретения. Используемые здесь термины и выражения являются описательными, но не ограничивающими терминами, и употребление этих терминов и выражений не исключает возможности включения любых эквивалентов проиллюстрированных и описанных здесь признаков или их частей, но при этом следует отметить, что в объем заявленного изобретения могут быть включены различные модификации. Таким образом, очевидно, что хотя в настоящем описании представлены примеры и предпочтительные варианты осуществления изобретения и его необязательные признаки, однако для специалиста в данной области очевидно, что в настоящее изобретение могут быть внесены модификации и изменения, не выходящие за рамки описанного здесь существа изобретения, и указанные модификации и изменения также входят в объем настоящего изобретения, определяемого прилагаемой формулы изобретения.
Дополнительные ссылки
US 6258562, US 6090382; US 6455275; EP1080206B1 ; WO 9960135; US 5912167; US 5162601; WO 199521249A1; US 5149783; US 5955072; US 5532142; US 20040224391; US 6537806; US 5846767; US 20030099932; WO 9958663; US 20030157641; US 2003048306A2; US 6114146; US 6060273; US 5925565; US 20040241821 ; WO 2003100021 A2; WO 2003100022A2; US 20040265955; US 20050003482; US 20050042721; WO 2005017149; WO 2004113493; US 20050136035; WO 2004108893; US 6692736; US 20050147962; US 6331415; US 6632637; US 20040063186; US 7026526; US 6365377; WO 2005123915; US 5665567; WO 9741241 A1; EP 0701616B1; US 20060010506; WO 2006048459; US 6852510; WO 2005072129; US 5648254; US 6908751; US 20050221429; WO 2005071088; WO 2005108585; WO 2005085456; US 7029876; US 6638762; US 6544780; US 5519164; WO 2003031630; US 6294353; WO 2005047512; US 7052905; US 7018833; US 20020034814; US 20040126883; US 20050002907; US 20050112095; US 20050214258; EP 0598029.
Mathys S. et al., 1999, Gene 231 (1-2): 1-13 “Characterization of a self- splicing mini-intein и its conversion into autocatalytic N- и C-terminal cleavage elements: facile production of protein building blocks for protein ligation”.
| название | год | авторы | номер документа |
|---|---|---|---|
| УСОВЕРШЕНСТВОВАННЫЕ ВЕКТОРЫ ЭКСПРЕССИИ МЛЕКОПИТАЮЩИХ И ИХ ПРИМЕНЕНИЕ | 2009 |
|
RU2502800C2 |
| ОПТИМИЗИРОВАННАЯ ЭКСПРЕССИОННАЯ КАССЕТА ДЛЯ ЭКСПРЕССИИ ПОЛИПЕПТИДА С ВЫСОКИМ ВЫХОДОМ | 2013 |
|
RU2682884C2 |
| УЛУЧШЕННЫЕ СПОСОБЫ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ПОЛИПЕПТИДА | 2014 |
|
RU2711322C1 |
| СЛИТЫЕ ПОЛИПЕПТИДЫ, СОДЕРЖАЩИЕ ДОМЕН WAP, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2639526C2 |
| СЛИТЫЕ СЕРПИНОВЫЕ ПОЛИПЕПТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2642310C2 |
| СИСТЕМА ЭКСПРЕССИИ И СЕКРЕЦИИ | 2013 |
|
RU2670491C2 |
| БАКТЕРИЯ Е.coli, ПРОДУЦИРУЮЩАЯ ГЕТЕРОЛОГИЧНЫЙ ПОЛИПЕПТИД, И СПОСОБ ПРОДУЦИРОВАНИЯ ПОЛИПЕПТИДА | 2001 |
|
RU2287574C2 |
| АНТИТЕЛА ПРОТИВ TNF-α И ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2595379C2 |
| СЛИТЫЕ СЕРПИНОВЫЕ ПОЛИПЕПТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2727452C1 |
| СПОСОБЫ И КОМПОЗИЦИЯ ДЛЯ СЕКРЕЦИИ ГЕТЕРОЛОГИЧНЫХ ПОЛИПЕПТИДОВ | 2010 |
|
RU2585488C2 |





















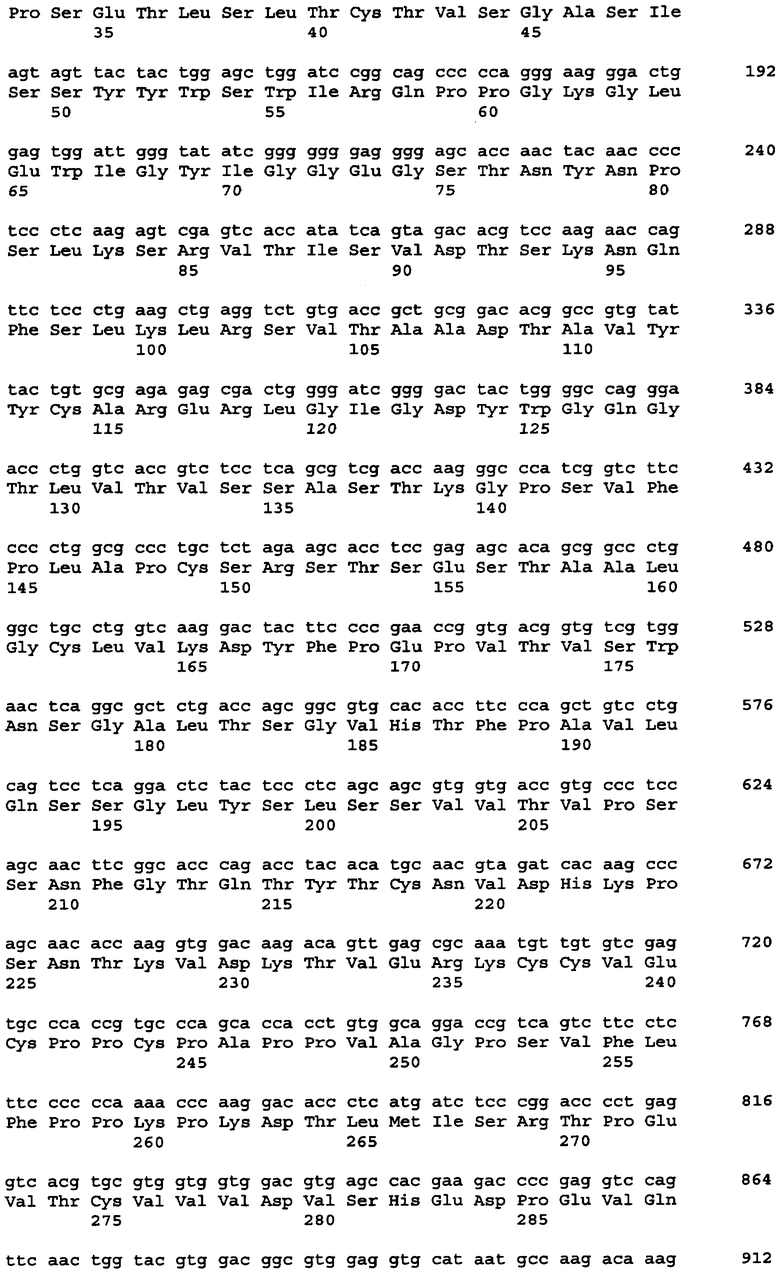
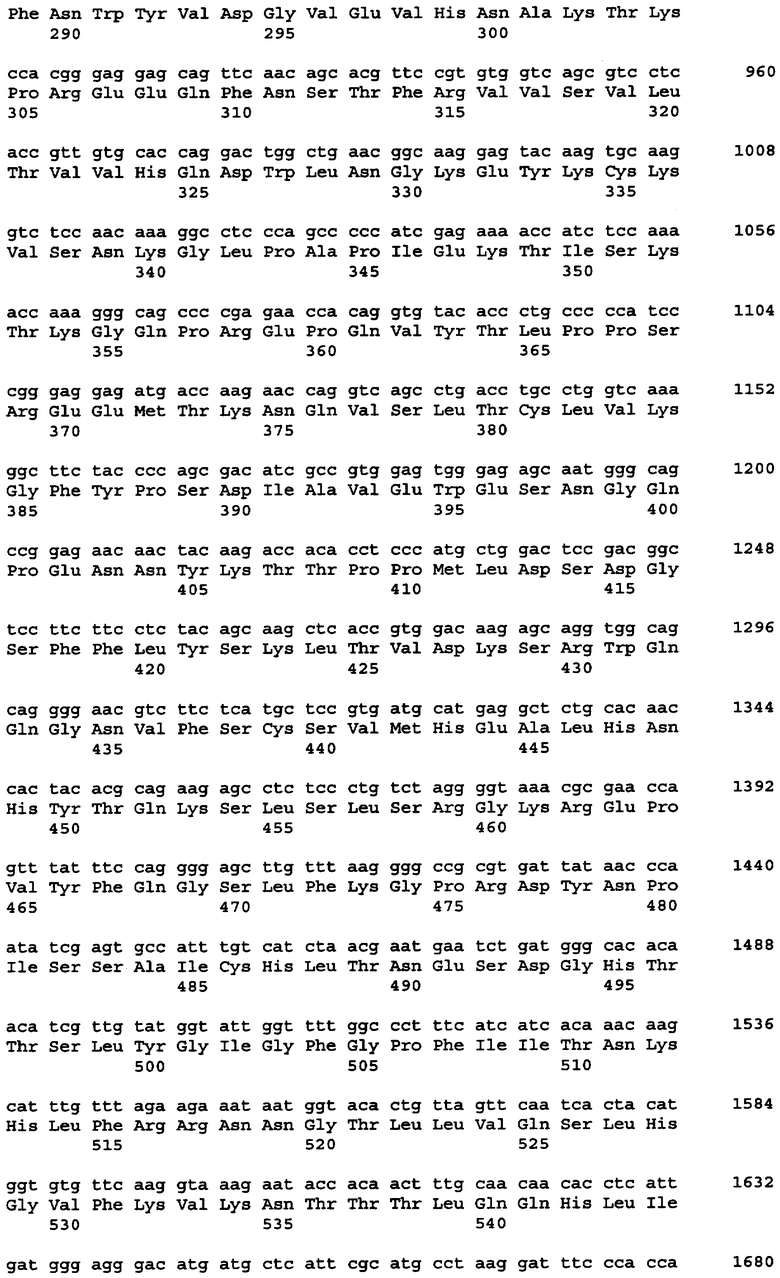
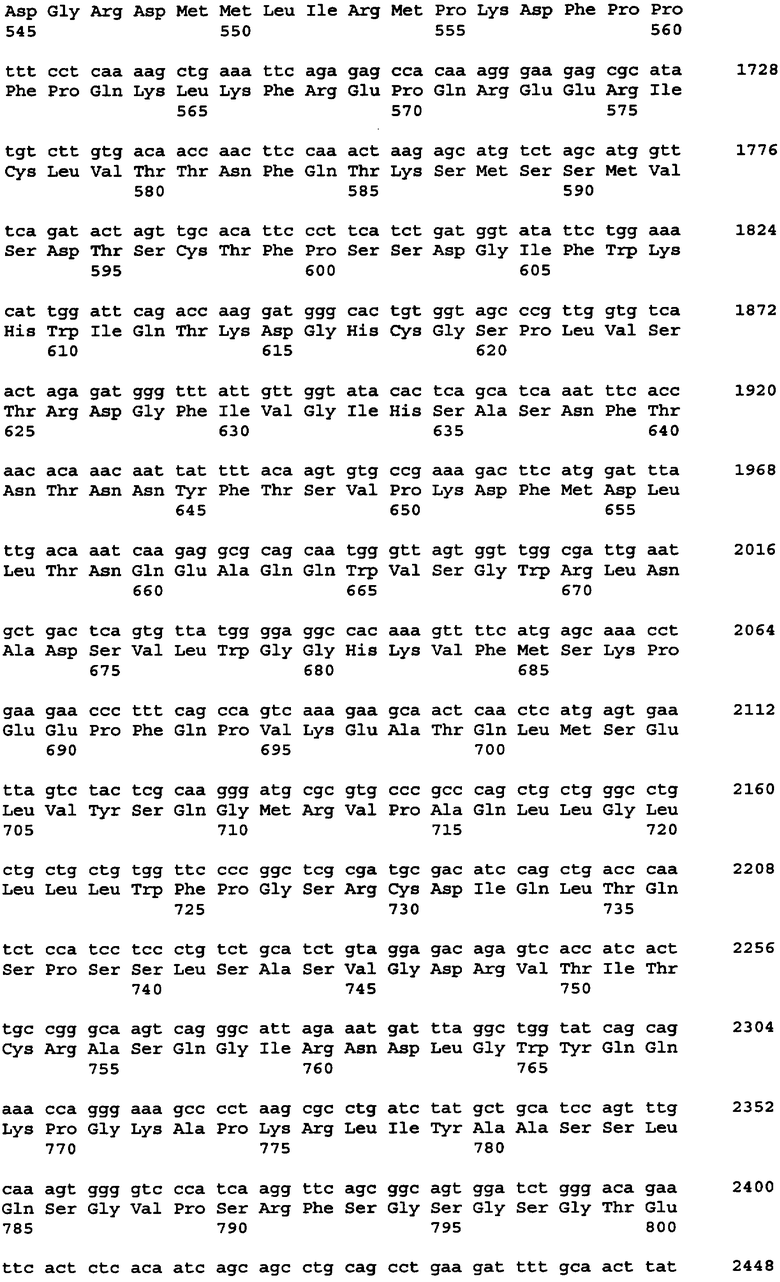











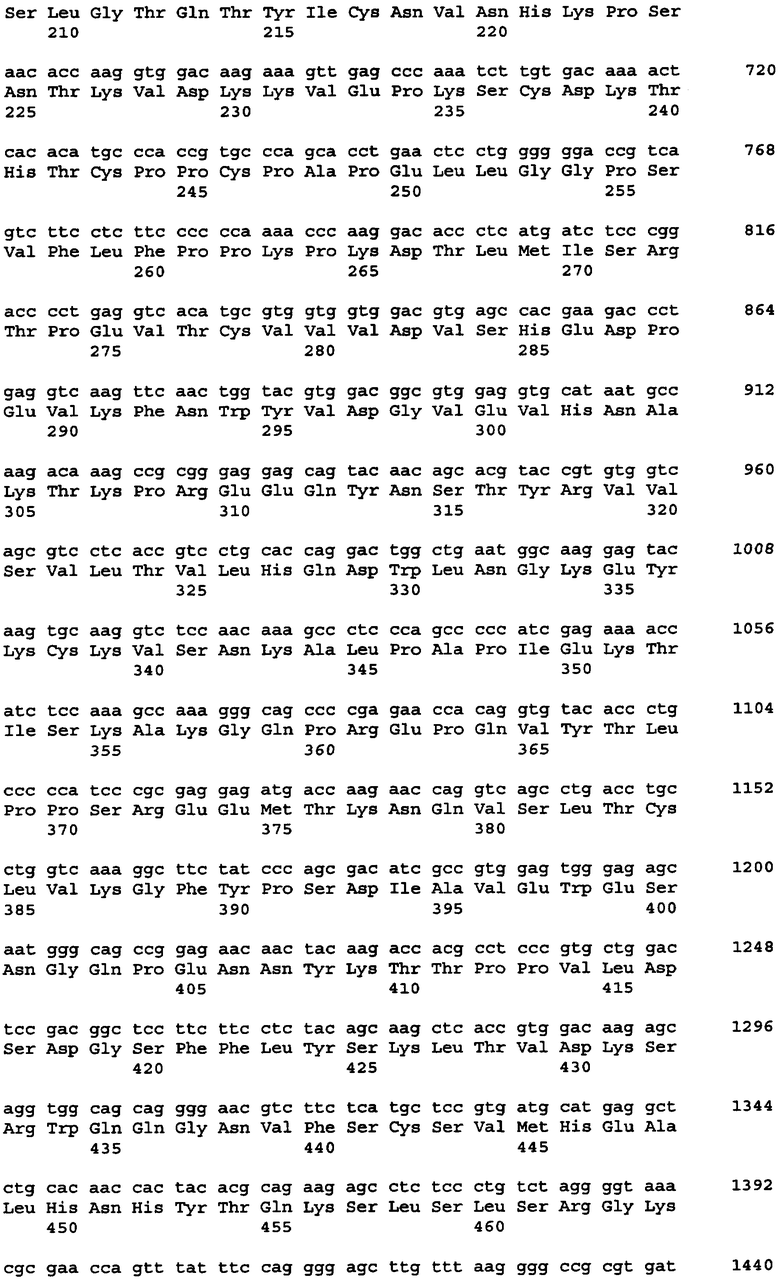
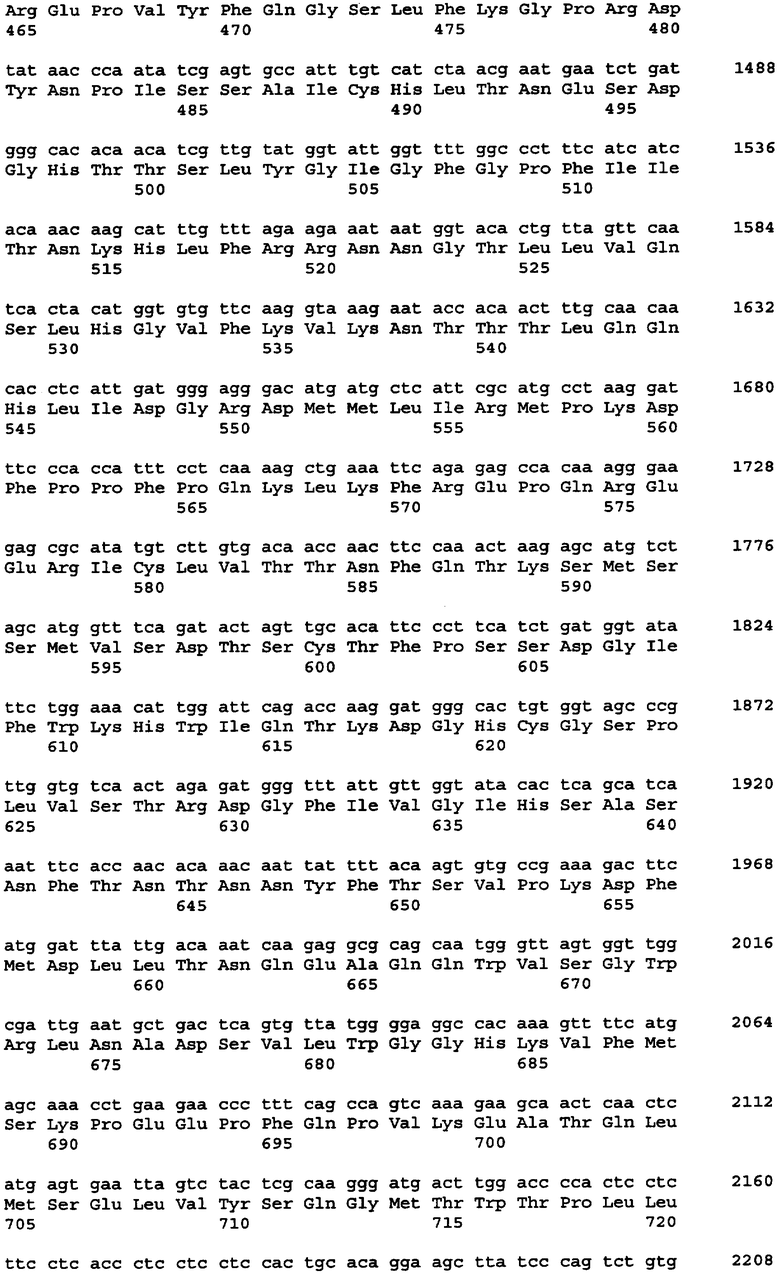
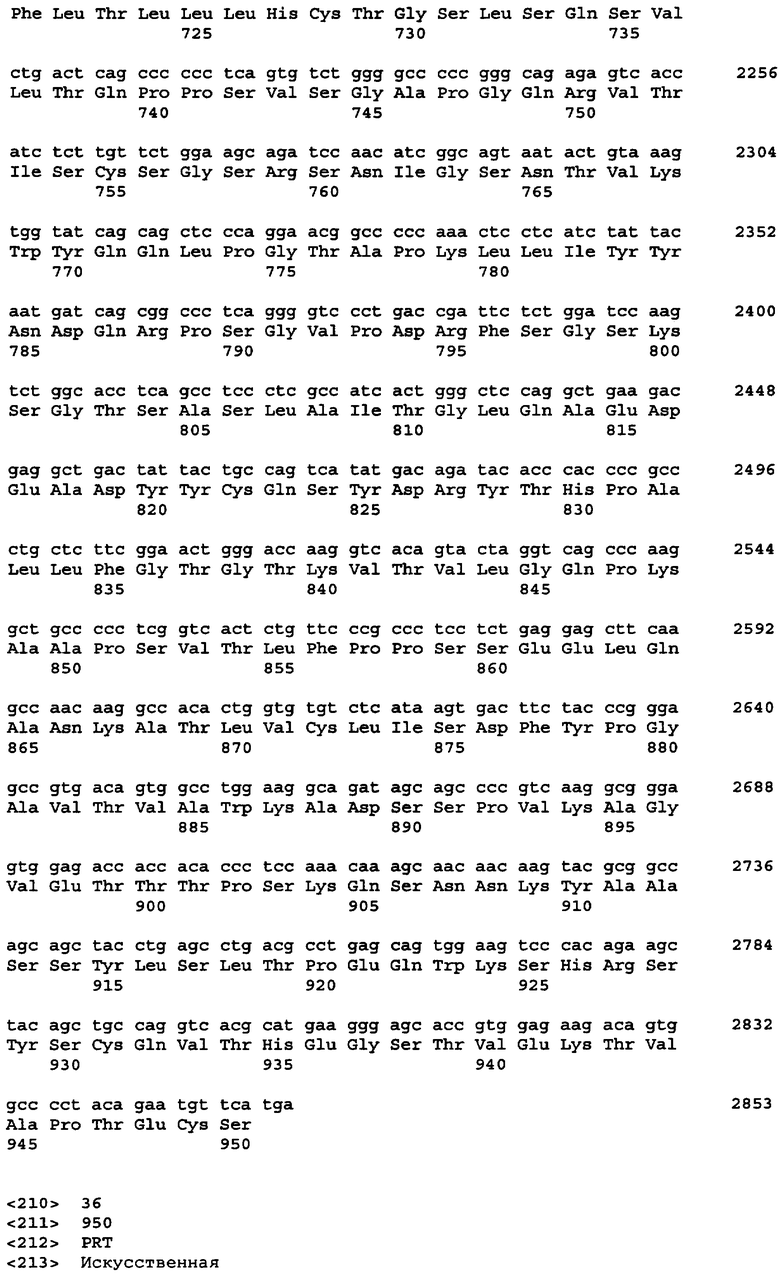










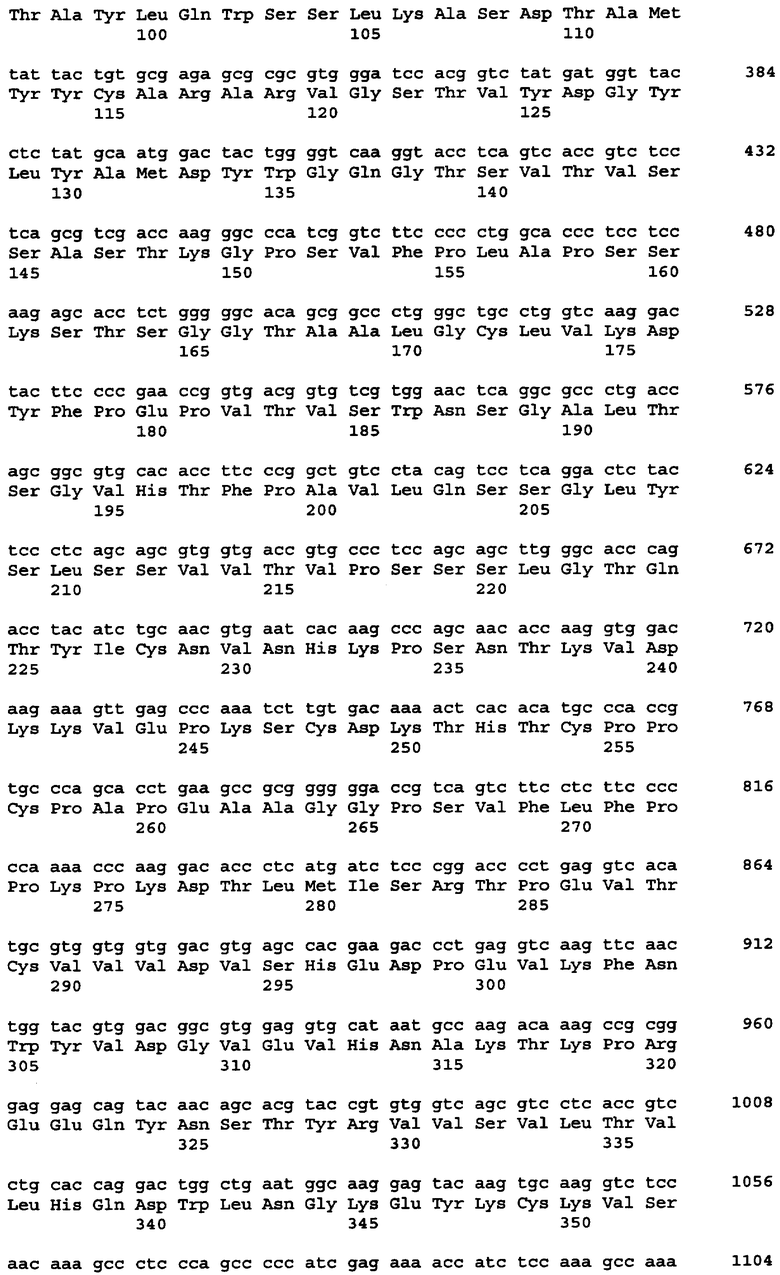













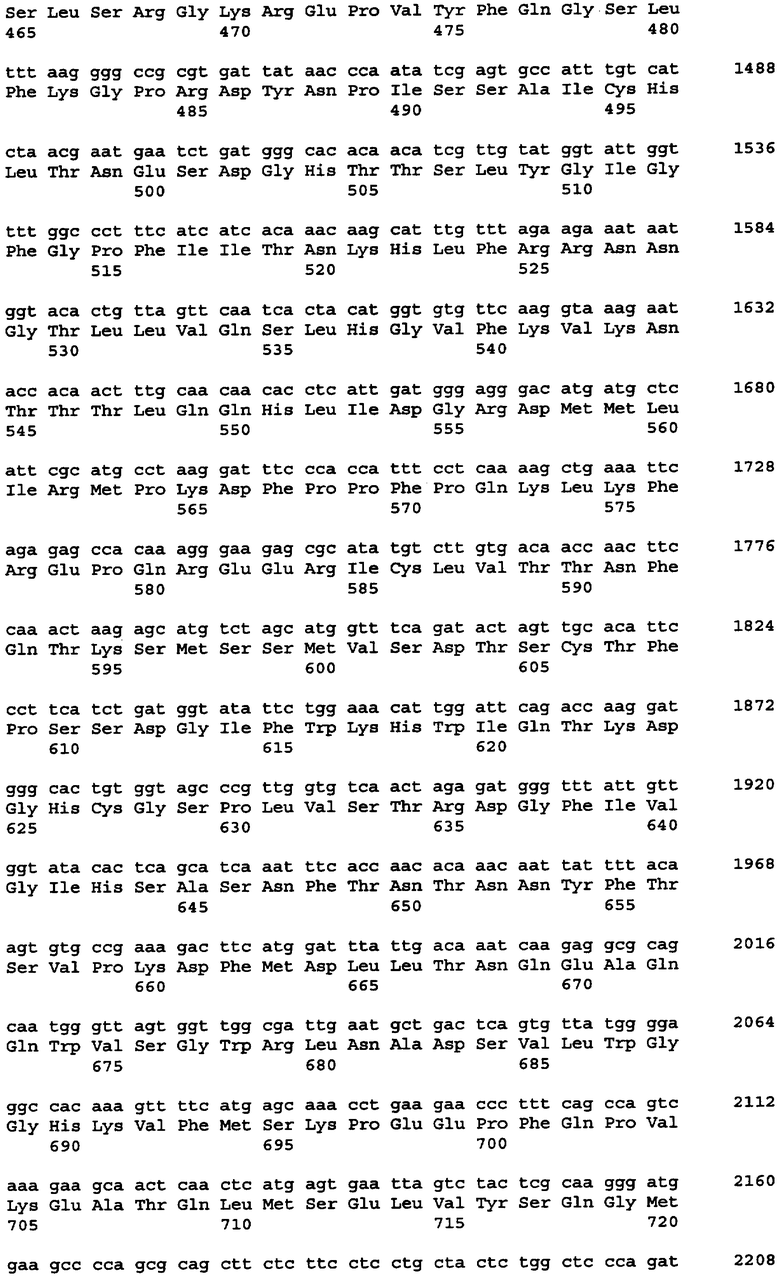
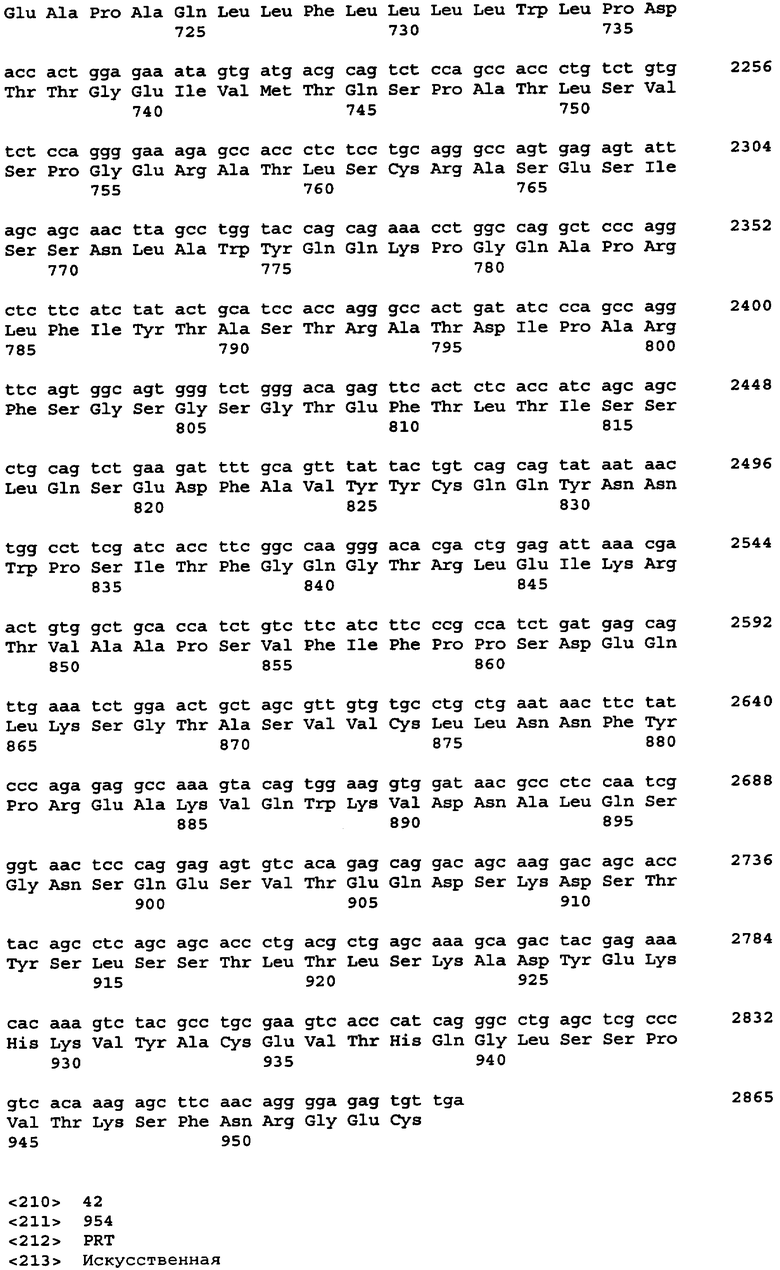
















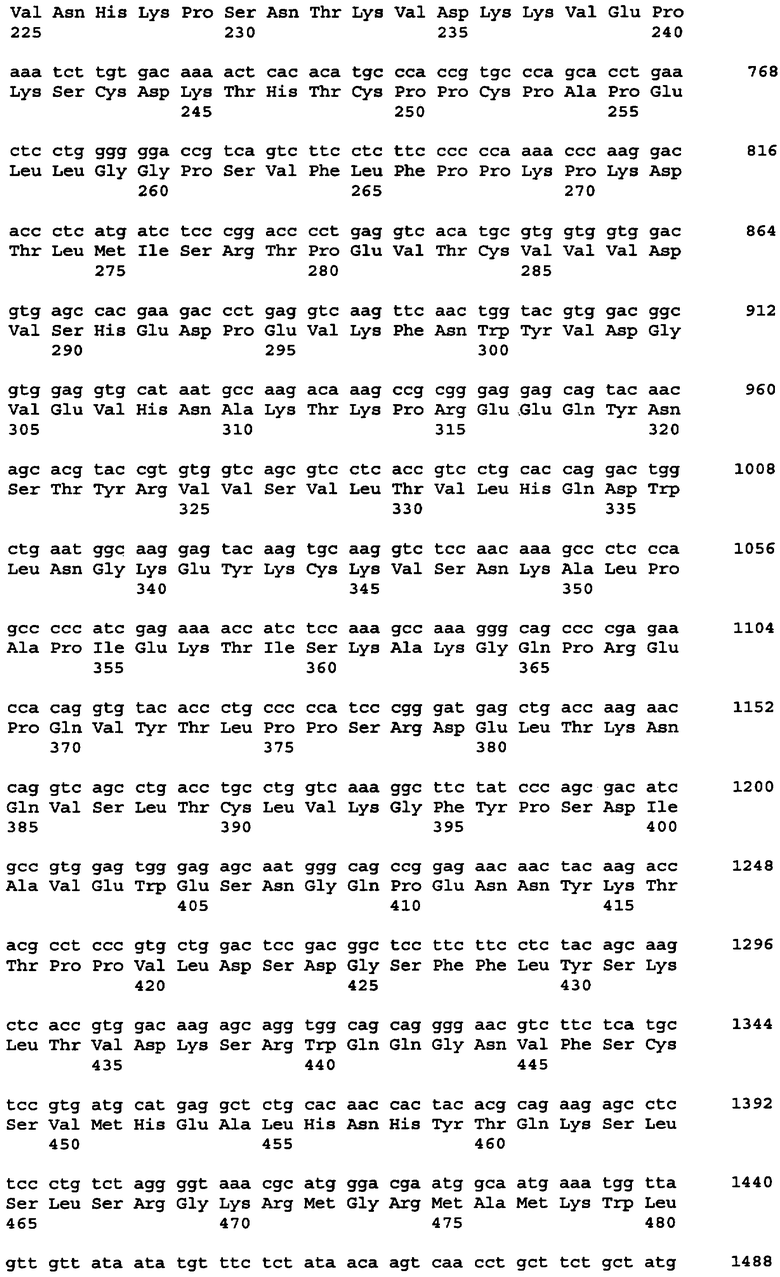












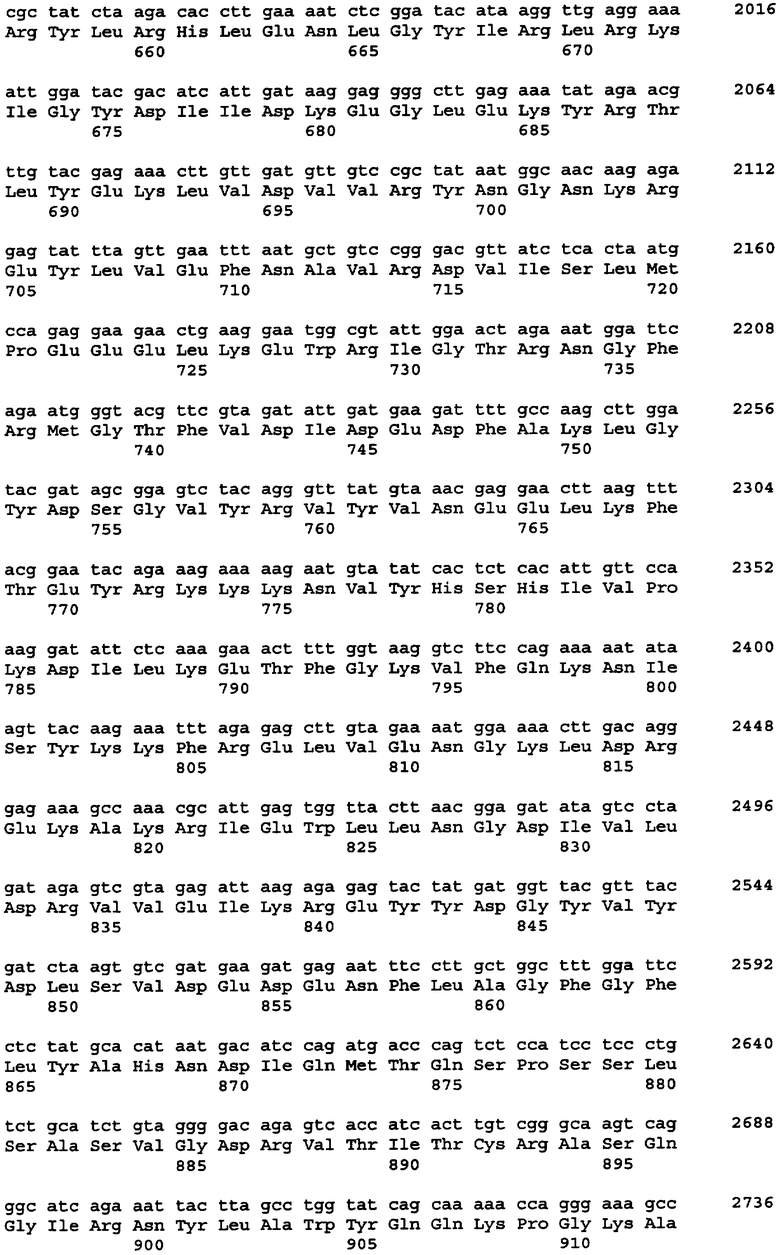























































































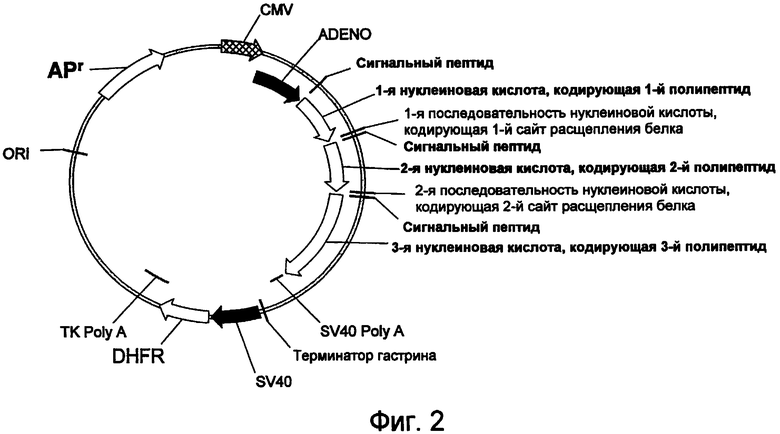
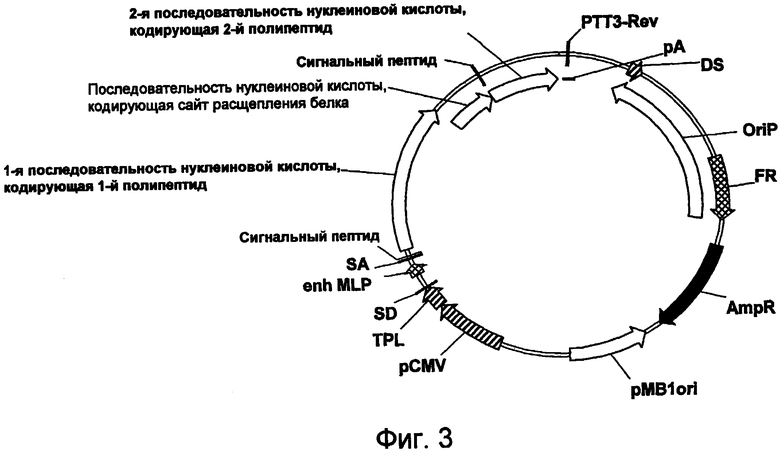
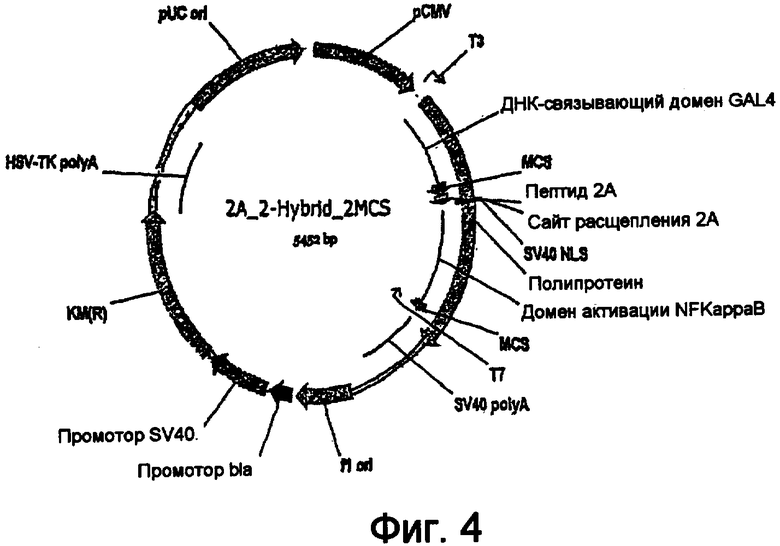
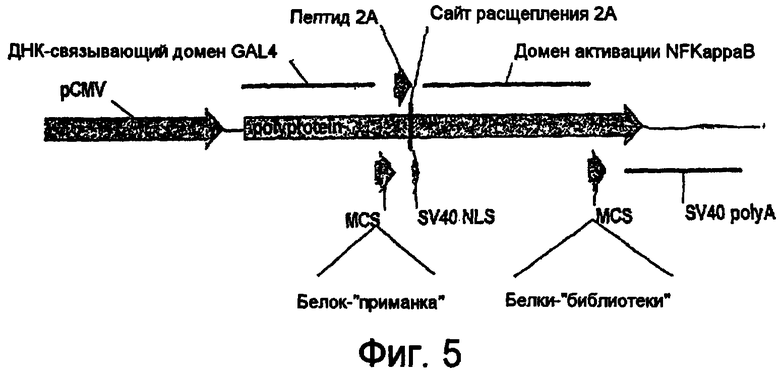
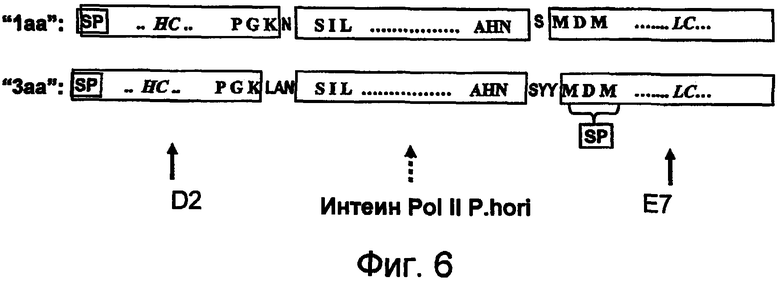
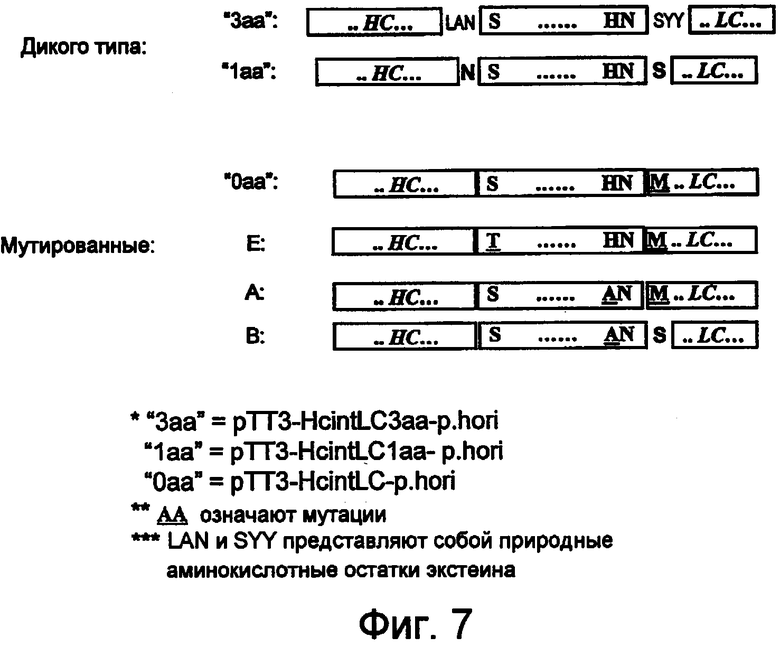

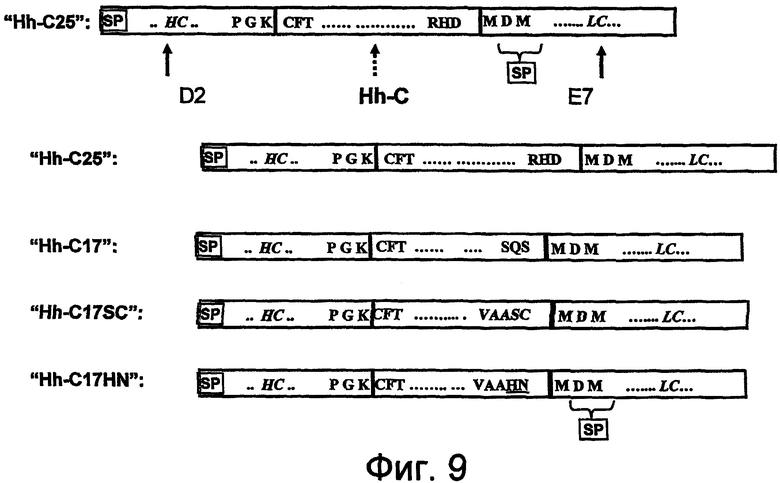
Изобретение относится к области биотехнологии, конкретно к экспрессионным конструкциям, и может быть использовано для экспрессии иммуноглобулина. Экспрессионный вектор содержит одну открытую рамку считывания (sORF)-вставку, которая включает первую последовательность нуклеиновой кислоты, кодирующую первый полипептид; первую промежуточную последовательность нуклеиновой кислоты, кодирующую первый сайт расщепления белка, включающий элемент аутопроцессинга с сегментом интеина, обеспечивающий протеолитическое расщепление полипептида sORF между первым полипептидом и сегментом интеина и вторым полипептидом, но не лигирование указанного первого полипептида с указанным вторым полипептидом; и вторую последовательность нуклеиновой кислоты, кодирующую второй полипептид. Экспрессионный вектор способен экспрессировать в клетке-хозяине полипептид млекопитающего, кодируемый sORF и расщепляемый в указанном первом сайте расщепления белка, состоящий из первого полипептида - тяжелой цепи иммуноглобулина и второго полипептида - легкой цепи иммуноглобулина, которые обладают способностью к сборке в мультимер. Изобретение обеспечивает продуцирование функционального антитела с "правильной" укладкой и сборкой. 4 н. и 36 з.п. ф-лы, 9 ил., 57 табл., 4 пр.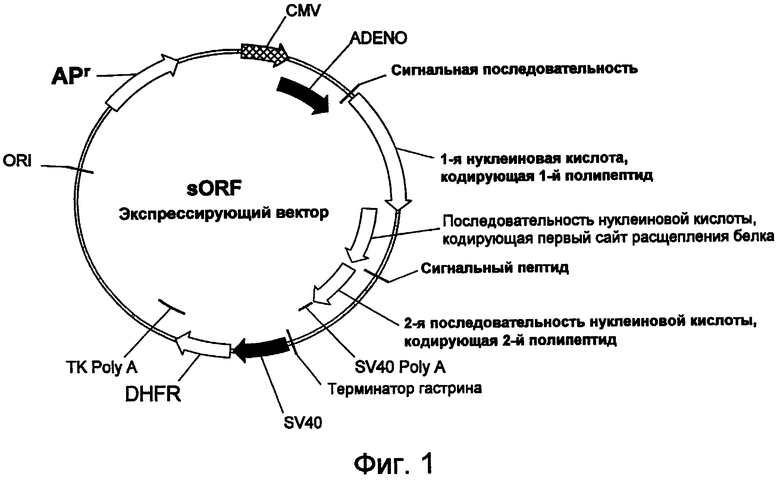
1. Экспрессионный вектор для генерирования одного или более рекомбинантных белковых продуктов, содержащий одну открытую рамку считывания (sORF)-вставку, где указанная sORF-вставка включает первую последовательность нуклеиновой кислоты, кодирующую первый полипептид; первую промежуточную последовательность нуклеиновой кислоты, кодирующую первый сайт расщепления белка; и вторую последовательность нуклеиновой кислоты, кодирующую второй полипептид, где указанная промежуточная последовательность нуклеиновой кислоты, кодирующая указанный первый сайт расщепления белка, соответствующим образом расположена между указанной первой последовательностью нуклеиновой кислоты и указанной второй последовательностью нуклеиновой кислоты, и где указанный экспрессионный вектор способен экспрессировать полипептид, кодируемый sORF и расщепляемый в указанном первом сайте расщепления белка; где указанный первый сайт расщепления белка включает элемент аутопроцессинга, включающий сегмент интеина или модифицированный сегмент интеина, где указанные сегмент интеина или модифицированный сегмент интеина позволяют протеолитическое расщепление полипептида sORF между первым полипептидом и сегментом интеина или модифицированного интеина и между сегментом интеина или модифицированного интеина и вторым полипептидом, но не лигирование указанного первого полипептида с указанным вторым полипептидом, где указанные первый и второй полипептиды происходят от млекопитающего, где указанный первый полипептид содержит тяжелую цепь иммуноглобулина или ее функциональный фрагмент, а указанный второй полипептид содержит легкую цепь иммуноглобулина или ее функциональный фрагмент, и где указанные первый и второй полипептиды расположены в любом порядке, и где указанные первый полипептид и второй полипептид обладают способностью к сборке в мультимер; где указанный сегмент интеина или модифицированный сегмент интеина содержат последовательность Pho Pol I Pyrococcus horikoshii, последовательность VMA Saccharomyces cerevisiae, последовательность DnaE штамма РСС6803 Synechocystis spp, последовательность GyrA Mycobacterium xenopi, последовательность ДНК-полимеразы GB-D Pyrococcus spp., последовательность бактериального интеин-подобного домена (BIL) А-типа или последовательность BIL В-типа.
2. Экспрессионный вектор по п.1, где указанный первый полипептид расположен выше от указанного второго полипептида.
3. Экспрессионный вектор по п.1, где указанный второй полипептид расположен выше от указанного первого полипептида.
4. Экспрессионный вектор по п.1, дополнительно содержащий третью последовательность нуклеиновой кислоты, кодирующую третий полипептид, где указанная третья последовательность нуклеиновой кислоты функционально расположена после указанной второй последовательности нуклеиновой кислоты; где указанная третья последовательность независимо может быть той же самой, что и указанная первая или вторая последовательности нуклеиновой кислоты, либо она может отличаться от указанных последовательностей.
5. Экспрессионный вектор по п.4, где, по меньшей мере, два из указанных первого, второго и третьего полипептидов, взятые вместе, обладают способностью к сборке в мультимер.
6. Экспрессионный вектор по п.1, дополнительно содержащий вторую промежуточную последовательность нуклеиновой кислоты, кодирующую второй сайт расщепления белка, где указанная вторая промежуточная последовательность нуклеиновой кислоты функционально расположена после указанной первой и указанной второй последовательностью нуклеиновой кислоты; где указанная вторая промежуточная последовательность может быть той же самой, что и указанная первая промежуточная последовательность нуклеиновой кислоты, либо она может отличаться от указанной первой промежуточной последовательности нуклеиновой кислоты.
7. Экспрессионный вектор по п.1, дополнительно содержащий третью последовательность нуклеиновой кислоты, кодирующую третий полипептид, и вторую промежуточную последовательность нуклеиновой кислоты, кодирующую второй сайт расщепления белка, где указанная вторая промежуточная последовательность нуклеиновой кислоты и третья последовательность нуклеиновой кислоты, в этом порядке, функционально расположены после указанной второй последовательности нуклеиновой кислоты.
8. Экспрессионный вектор по п.4, где указанная третья последовательность нуклеиновой кислоты кодирует тяжелую цепь или легкую цепь иммуноглобулина, или, соответственно, их функциональные фрагменты.
9. Экспрессионный вектор но п.4. где указанная третья последовательность нуклеиновой кислоты кодирует легкую цепь иммуноглобулина или ее функциональный фрагмент.
10. Экспрессионный вектор по п.4, где указанная третья последовательность нуклеиновой кислоты кодирует тяжелую цепь иммуноглобулина или ее функциональный фрагмент.
11. Экспрессионный вектор по п.1, где указанная первая промежуточная последовательность нуклеиновой кислоты, кодирующая первый сайт расщепления белка, включает нуклеиновую кислоту сигнального пептида, кодирующую сайт отщепления сигнального пептида или модифицированную последовательность сайта отщепления сигнального пептида.
12. Экспрессионный вектор по п.1, дополнительно содержащий последовательность нуклеиновой кислоты сигнального пептида, кодирующую сайт отщепления сигнального пептида, и функционально расположенную перед указанной первой последовательностью нуклеиновой кислоты или указанной второй последовательностью нуклеиновой кислоты.
13. Экспрессионный вектор по п.1, дополнительно содержащий две последовательности нуклеиновой кислоты сигнального пептида, каждая из которых независимо кодирует сайт отщепления сигнального пептида, где указанная одна последовательность нуклеиновой кислоты сигнального пептида функционально расположена перед указанной первой нуклеиновой кислотой, кодирующей указанный первый полипептид, а другая последовательность нуклеиновой кислоты сигнального пептида функционально расположена перед указанной второй нуклеиновой кислотой, кодирующей указанный второй полипептид.
14. Экспрессионный вектор по п.11, где указанная последовательность нуклеиновой кислоты сигнального пептида кодирует сайт отщепления сигнального пептида легкой цепи иммуноглобулина или модифицированный сайт отщепления сигнального пептида легкой цепи иммуноглобулина.
15. Экспрессионный вектор по п.2, где указанные функциональное антитело или другая молекула, распознающая антиген, имеют антигенную специфичность, направленную на связывание с антигеном, выбранным из группы, состоящей из фактора некроза опухоли а, рецептора эритропоэтина, RSV, E/L-селектина, интерлейкина-1, интерлейкина-12, интерлейкина-13, интерлейкина-18, интерлейкина-23, CXCL-13, GLP-1R и бета-амилоида.
16. Экспрессионный вектор по п.1, где указанный первый и второй полипептиды предоставляют пару иммуноглобулиновых цепей, происходящих из антитела адалимумаба (D2E7), аминокислотных последовательностей иммуноглобулина SEQ ID NO:33 (АВТ-007), аминокислотных последовательностей иммуноглобулина SEQ ID NO:42 (АВТ-325), аминокислотных последовательностей иммуноглобулина SEQ ID NO:39 (EL246) или аминокислотных последовательностей иммуноглобулина SEQ ID NO:36 (АВT-874).
17. Экспрессионный вектор по п.1, где каждый из указанных первого и второго полипептидов независимо выбран из сегментов тяжелой цепи иммуноглобулина или легкой цепи иммуноглобулина, происходящих из аналогичного сегмента адалимумаба (D2E7), аминокислотных последовательностей иммуноглобулина SEQ ID NO:33 (АВТ-007), аминокислотных последовательностей иммуноглобулина SEQ ID NO:42 (АВТ-325), аминокислотных последовательностей иммуноглобулина SEQ ID NO:39 (ЕL246) или аминокислотных последовательностей иммуноглобулина SEQ ID NO:36 (АВТ-874) или других антител.
18. Экспрессионный вектор по п.1, где указанный вектор дополнительно содержит промоторный регуляторный элемент для указанной sORF-вставки.
19. Экспрессионный вектор по п.18, где указанный промоторный регуляторный элемент является индуцибельным или конститутивным.
20. Экспрессионный вектор по п.18, где указанный промоторный регуляторный элемент является тканеспецифическим.
21. Экспрессионный вектор по п.18, где указанный промотор включает основной поздний промотор аденовируса.
22. Экспрессионный вектор по п.1, где указанный вектор дополнительно включает нуклеиновую кислоту, кодирующую протеазу, способную расщеплять указанный первый сайт расщепления белка.
23. Экспрессионный вектор по п.22, где указанная нуклеиновая кислота, кодирующая протеазу, оперативно расположена в указанной sORF-вставке, где указанный экспрессирующий вектор также включает в себя дополнительную нуклеиновую кислоту, кодирующую второй сайт расщепления, расположенный между указанной нуклеиновой кислотой, кодирующей протеазу, и, по меньшей мере, одной из указанных первой и второй нуклеиновых кислот.
24. Экспрессионный вектор по п.1, где указанный вектор не содержит последовательности 2А.
25. Экспрессионный вектор по п.1, где указанная промежуточная последовательность нуклеиновой кислоты дополнительно кодирует метку.
26. Клетка-хозяин, кроме эмбриональной клетки человека, для получения белков иммуноглобулина, содержащая вектор но любому из пп.1-25.
27. Клетка-хозяин по п.26, являющаяся прокариотической клеткой.
28. Клетка-хозяин по п.27, являющаяся клеткой Escherichia coli.
29. Клетка-хозяин по п.26, являющаяся эукариотической клеткой.
30. Клетка-хозяин по п.29, где указанная эукариотическая клетка выбрана из группы, состоящей из клеток одноклеточных организмов, клеток животных, клеток растений и клеток грибов.
31. Клетка-хозяин но п.30, где указанной эукариотической клеткой является клетка животного, выбранная из группы, состоящей из клеток млекопитающих, клеток птиц и клеток насекомых.
32. Клетка-хозяин по п.31, являющаяся клеткой яичника китайского хомячка (CНO) или дефицитной по дигидрофолат-редуктазе клеткой СНО.
33. Клетка-хозяин по п.32, являющаяся клеткой COS.
34. Клетка-хозяин по п.30, являющаяся дрожжевой клеткой.
35. Клетка-хозяин по п.34, где указанной дрожжевой клеткой является Saccharomyces cerevisiae.
36. Клетка-хозяин по п.31, являющаяся клеткой Sf9 насекомого Spodoptera frugiperda.
37. Способ получения белков иммуноглобулина, включающий культивирование клетки-хозяина по п.26 в культуральной среде в условиях, благоприятных для осуществления экспрессии белков иммуноглобулина.
38. Способ по п.37, дополнительно включающий выделение и/или очистку указанных белков.
39. Способ по п.37 или 38, где указанные белки являются терапевтическими белками.
40. Способ получения белка иммуноглобулина, где указанный способ включает культивирование клетки-хозяина, кроме эмбриональной клетки человека, содержащей экспрессионный вектор по любому из пп.1-25 в культуральной среде в условиях, благоприятных для продуцирования белка иммуноглобулина.
| FANG J | |||
| et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Nat | |||
| Biotechnol | |||
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| US 2004265966 A1, 30.12.2004 | |||
| US 2004235011 A1, 25.11.2004 | |||
| US 2004191786 A1, 30.09.2004 | |||
| US 2002129400 A1, 12.09.2002 | |||
| Устройство для определения места повреждения изоляции и степени замыкания на землю в цепи оперативного постоянного тока | 1957 |
|
SU112820A1 |
| СПОСОБ ИДЕНТИФИКАЦИИ И ХАРАКТЕРИЗАЦИИ ФУНКЦИЙ, ПОТЕНЦИАЛЬНО ИМЕЮЩИХСЯ В БИОЛОГИЧЕСКОМ ОБРАЗЦЕ, СОДЕРЖАЩЕМ НУКЛЕИНОВЫЕ КИСЛОТЫ | 1999 |
|
RU2249045C2 |

