Изобретение относится к области иммунологии.
Изобретение может быть использовано в иммунологических, серологических и вирусологических исследованиях по изучению и оценке защитных свойств препаратов нового поколения против вирусных инфекционных заболеваний, а также для разработки методов биологической защиты.
Достижения молекулярной биологии, генетической инженерии, иммунологии и биотехнологии открыли принципиально новые перспективные подходы конструирования различных типов вакцин. Практическую реализацию получили два направления: создание искусственных вакцин путем химического синтеза пептидов, имитирующих протективные антигены патогенов, и путем генно-инженерной технологии. Причем последнее направление получило наибольшее развитие и практическую реализацию. К вакцинам, полученным с помощью генно-инженерных технологий, относятся: векторные рекомбинантные вакцины, ДНК-вакцины, химерные вакцины, трансгенные вакцины.
В начале 90-х годов был предложен принципиально новый подход конструирования вакцин, при котором в качестве вакцинного препарата используется плазмида, содержащая ген(ы), кодирующий чужеродный целевой белок (протективные антигены патогена), так называемая полинуклеотидная вакцина, или ДНК-вакцина [1]. При создании ДНК-вакцин чаще всего используются плазмиды E.coli pUC типа, поскольку последние обеспечивают высокий выход и чистоту ДНК. Для достижения максимальной экспрессии клонированных генов в клетках млекопитающих используются вирусные промоторы, чаще всего цитомегаловирусный.
В настоящее время сконструированы и апробированы на различных экспериментальных моделях полинуклеотидные вакцины против ряда вирусных, бактериальных и паразитарных инфекций [2].
Интенсивная работа ведется по конструированию ДНК-вакцин против флавивирусных инфекций, включая лихорадку Денге и энцефалиты (клещевой, японский, Западного Нила, Сент Луис, долины Муррей) [3, 4].
В рамках международного сотрудничества по борьбе с малярией проводится работа по созданию ДНК-вакцины против малярии, вызываемой Plasmodium falciparum [5].
В ряде экспериментов доказана возможность успешного применения полинуклеотидных вакцин для лечения аллергических болезней [6].
Недавно обнаружено, что полинуклеотидные вакцины могут индуцировать толерантность к экспрессируемым аутоантигенам и, следовательно, использоваться для лечения аутоиммунных заболеваний, в частности диабета [7, 8].
Полинуклеотидные вакцины могут оказаться весьма эффективными не только как средства профилактики, но и в качестве лечебных препаратов при терапии хронических персистентных инфекций, так как они индуцируют сильный клеточноопосредованный иммунный ответ (цитотоксические Т-лимфоциты, Th-1-клетки), который приводит к элиминации патогенов.
Параллельно развитию методов конструирования вакцинных препаратов появляются новые данные о механизмах презентации антигенов иммунной системе хозяина. Так, в настоящее время сформировалось мнение о том, что клеточные органеллы животных - протеасомы - участвуют в процессинге антигенов, представляемых молекулами первого класса главного комплекса гистосовместимости. Реализуется данный процесс в непосредственной связи с убиквитинконъюгирующей системой [9]. Совместно с убиквитином потенциальной специфичностью к протеасомному комплексу обладает и ряд других конститутивных пептидов млекопитающих, что связано с наличием в их последовательностях сочетаний некоторых аминокислотных остатков, являющихся сильными сигналами деградации. К числу последних относится фермент орнитиндекарбоксилаза (ODC).
К основным клеточным процессам, регулируемым протеасомной системой, относятся: клеточный цикл и деление, дифференцировка, эмбриогенез, апоптоз, сигнальная трансдукция, репарация ДНК, трансмембранный и везикулярный транспорт, реакция на стрессовые воздействия, в том числе на патогены. Многообразие процессов, регулируемых протеасомной системой, позволяет сделать вывод о ее главенствующей роли в такой кооперативной реакции организма, как иммунный ответ [10]. Использование протеасомоузнающих сигналов, обуславливающих антигенный процессинг и ускоренную презентацию иммунокомпетентным клеткам при конструировании вакцин нового поколения, может позволить получать высокоэффективные защитные препараты с повышенной иммуногенностью.
На сегодняшний день в научной литературе имеются данные об экспериментальной оценке пептидных вакцин, полученных при использовании протеасомной деградации in vitro, против ряда инфекционных заболеваний вирусной и бактериальной этиологии [11, 12]. Однако использование технологии ДНК-вакцинации при реализации механизма направленного протеасомоопосредованного процессинга антигенов не описано.
Способ ДНК-вакцинации при реализации механизма направленного протеасомоопосредованного процессинга антигенов не имеет аналогов в Российской Федерации.
В качестве модельного возбудителя инфекционного заболевания, против которого разрабатываются профилактические средства, выбирают хорошо изученный возбудитель.
Вирус Венесуэльского энцефаломиелита лошадей (ВЭЛ) является одним из наиболее полно изученных как структурно, так и с точки зрения иммуно- и патогенеза возбудителей вирусных инфекций. Структурно возбудитель содержит одну молекулу РНК, ковалентно связанную с белком капсида и окруженную димером гликопротеинов оболочки Е1 и Е2, однако в условиях высокой множественности заражения возможно формирование полиплоидных вирионов, содержащих под одним суперкапсидом несколько нуклеокапсидов.
Оба гликопротеина оболочки (Е1 и Е2) имеют значение для сохранения инфекционности и патогенности вируса, однако если Е1 в большей степени определяет этап связывания вируса с рецепторами клеточной оболочки и последующий виропексис, то Е2 преимущественно индуцирует иммунный ответ организма на вирусную инфекцию, что показано в исследованиях с сыворотками реконвалесцентов.
При клонировании фрагментов структурных пептидов вирусов, имеющих антигенные детерминанты, необходимым условием является сохранение наиболее актуальных эпитопов. К настоящему времени антигенная структура гликопротеина Е2 (фиг.1) достаточно хорошо изучена, а эпитопы на его поверхности картированы как американскими, так и отечественными специалистами [13]. Иммунодоминантное положение занимает высоко консервативный участок, в состав которого входят фрагменты гена белка Е2 - М вируса ВЭЛ (фиг.1), расположенный в центральной зоне белка Е2 на небольшом удалении от сайта гликозилирования.
Целью настоящего изобретения является разработка способа ДНК-вакцинации, основанного на протеасомоопосредованной технологии (на примере вируса ВЭЛ) с использованием рекомбинантных плазмид pCI-neo-ODC-nsP1, pCI-neo-ODC-E2-L, pCI-neo-ODC-E2-M.
Сущность изобретения состоит в том, что впервые в Российской Федерации в результате молекулярно-биологических и иммунологических исследований разработан способ ДНК-вакцинации, основанный на протеасомоопосредованной технологии, с использованием рекомбинантных плазмид pCI-neo-ODC-nsP I, pCI-neo-ODC-E2-L, pCI-neo-ODC-E2-M. Рекомбинантные плазмиды pCI-neo-ODC-nsP1, pCI-neo-ODC-E2-L, pCI-neo-ODC-E2-M получены в результате последовательного клонирования ген-эквивалентов неструктурного белка nsP1, фрагментов L, М структурного белка Е2 вируса ВЭЛ соответственно в составе гена орнитиндекарбоксилазы сначала в прокариотический плазмидный вектор рЕТ-23b, а затем в эукариотический плазмидный вектор pCI-neo.
В качестве контроля используются плазмиды pCI-neo-ODC и pCI-neo-Е2-М, полученные в результате последовательного клонирования гена ODC и фрагмента М гена Е2 вируса ВЭЛ соответственно сначала в прокариотический плазмидный вектор рЕТ-23b, а затем в эукариотический плазмидный вектор pCI-neo. Рекомбинантные плазмиды pCI-neo-ODC-nsP1, pCI-neo-ODC-E2-L, pCI-neo-ODC-E2-M, pCI-neo-ODC и pCI-neo-Е2-М депонированы в Специализированную коллекцию рекомбинантных плазмид Вирусологического центра НИИ микробиологии Министерства обороны Российской Федерации за №№02/1, 02/2, 02/3, 02/4 и 02/5 соответственно.
Характеристика и молекулярно-биологические свойства рекомбинантных плазмид pCI-neo-ODC-nsP1, pCI-neo-ODC-E2-L, pCI-neo-ODC-E2-M, pCI-neo-ODC и рС1-neo-Е2-М представлены в таблицах 1, 2.
Краткое описание чертежей
На фиг.1 представлена диаграмма конформационных эпитопов белка Е2 вируса ВЭЛ, проецированных на клонируемые фрагменты пептида в составе векторных плазмид рЕТ-23b и pCI-neo.
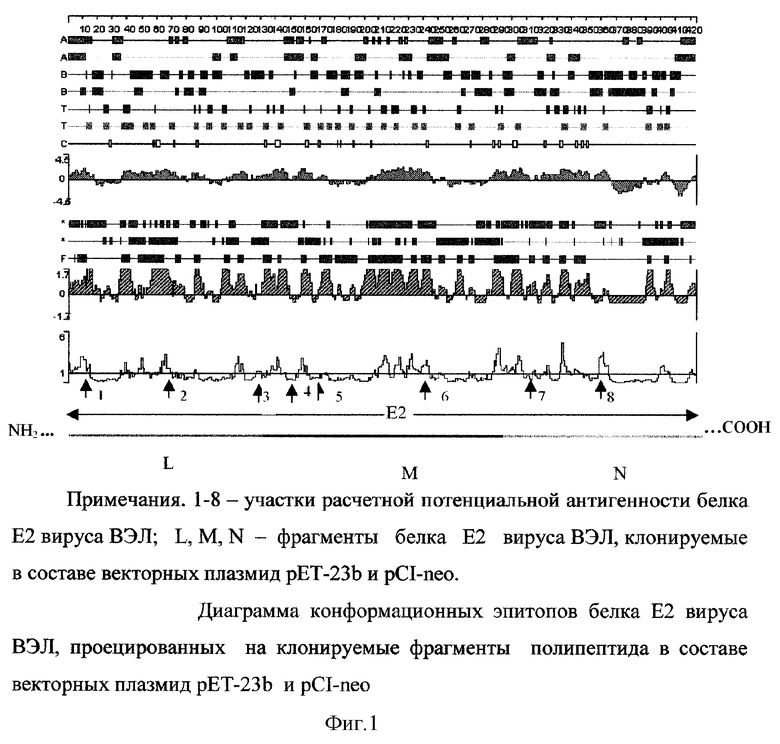
На фиг.2 показана схема ДНК-иммунизации внутримышечно белых мышей-самцов линии BALB/c (8-10 г) рекомбинантными плазмидами pCI-neo-ODC-nsP1, pCI-neo-ODC-E2-L, pCI-neo-ODC-E2-M и их комбинацией.
Пример выполнения
Пробирки с культурами Е.coli, содержащими рекомбинантные плазмиды pCI-neo-ODC-nsP1, pCI-neo-ODC-E2-L, pCI-neo-ODC-E2-M, pCI-neo-ODC и pCI-neo-Е2-М, извлекают из холодильника, температура хранения минус (20,0±2,0)°С. Проводят культивирование на среде LB с добавлением 25 мкг/мл ампициллина. Продолжительность культивирования составляет от 12 до 18 часов при температуре (37,0±0,5)°С.
Лизис бактериальных клеток осуществляют лизоцимом (10 мг/мл в 0,25 М Трис-HCl, рН 8,0) в присутствии 10%-ного SDS. Хромосомную ДНК удаляют высокоскоростным центрифугированием с последующей двойной экстракцией смесью фенол/хлороформ и одной экстракцией хлороформом. Остатки РНК удаляют методом гель-фильтрации на колонках с сефарозой 2В и осаждают двойным объемом перегнанного холодного 70%-ного этилового спирта. Концентрацию выделенной плазмидной ДНК определяют спектрофотометрически, учитывая тот факт, что одна оптическая единица при 260 нм соответствует 50 мкг ДНК в 1 мл [14]. После осаждения центрифугированием осадок плазмидной ДНК растворяют в буфере ТЕ до концентрации 1 мг/мл.
Выделенные и очищенные препараты оценивают затем электрофоретически в 0,8% агарозном геле. Электрофорез проводят при температуре (20,0±2,0)°С в течение 18 часов при напряженности электрического поля 1-2 В/см. Размер пластины 20×20 см2.
Полученные препараты представляют собой очищенные препараты ДНК рекомбинантных плазмид pCI-neo-ODC-nsP1, pCI-neo-ODC-E2-L, pCI-neo-ODC-E2-M, pCI-neo-ODC и pCI-neo-Е2-М, которые используются в качестве ДНК-вакцин против вируса Венесуэльского энцефаломиелита лошадей.
Для иммунизации (фиг.2) используют белых мышей BALB/c массой 7-8 г. Водные растворы плазмидных ДНК (концентрация ДНК 1 мг/мл) вводят однократно в мышцу бедра задней лапы белой мыши (-100 мкг ДНК/мышь).
В качестве положительного контроля служат белые мыши, иммунизированные подкожно в холку суспензией желточных мешков, инфицированных вакцинным штаммом вируса ВЭЛ (иммунизирующая доза - 3,7 lg ED50/мышь). Отрицательными контрольными группами при ДНК-вакцинации являются животные, иммунизированные плазмидами, содержащими ген-эквиваленты ODC и пептида М, а также интактные мыши. Использование этих плазмид необходимо для оценки возможного неспецифического действия орнитиндекарбоксилазы и специфического фрагмента белка Е2 на иммунитет лабораторных животных.
Иммунизацию животных проводят трехкратно. Бустерные иммунизации проводят через 4 и 3 недели соответственно. У 50% мышей на 14 сутки отбирают кровь для анализа методом ТФИФА. Остальных животных инфицируют смертельной дозой вирулентного штамма вируса ВЭЛ (100 LD50/мышь). После заражения срок наблюдения за грызунами составляет 14 суток.
С целью изучения иммуногенного и протективного эффекта комбинированной вакцинации лабораторным животным одновременно вводят смесь изучаемых препаратов плазмидной ДНК с ген-эквивалентами фрагмента белка Е2-L и М (pCI-neo-ODC-E2-L, pCI-neo-ODC-E2-M).
Иммуногенную активность рекомбинантных плазмид, несущих фрагменты генома вируса ВЭЛ, инъецированных как по отдельности, так и в комбинации, определяют по титру гуморальных антител в сыворотке крови иммунизированных белых мышей с помощью метода ТФИФА. Оцениваемые препараты, их комбинация, группы животных и титры антител представлены в таблице 3.
У всех животных, иммунизированных вакцинным штаммом вируса ВЭЛ (положительный контроль), выявляются специфические антитела к вирусу (в разведении не меньшем, чем 1:100). В то же время у большей части мышей, иммунизированных смесью плазмид pCI-neo-ODC-E2-L+pCI-neo-ODC-E2-M (83%) и плазмидой pCI-neo-ODC-nsP1 (78%) также отмечается положительная сероконверсия (таблица 4).
Протективную эффективность рекомбинантных плазмид рассчитывают, определяя разницу величины процента выживших (после заражения вирулентным вирусом ВЭЛ) мышей в группе, иммунизированной вакцинным штаммом вируса ВЭЛ, и в группе животных, инъецированных рекомбинантной плазмидой или смесью плазмид (при 100% гибели неиммунизированных мышей).
Комбинированная внутримышечная иммунизация плазмидной ДНК со встроенными ген-эквивалентами пептидов L и М белка Е2 вируса ВЭЛ индуцирует у большей части белых мышей выработку специфических антител и защиту от гибели при заражении летальной дозой вирулентного штамма возбудителя (таблица 4).
 ±δх) в сыворотках интактных животных равна 84,9±9,6;
±δх) в сыворотках интактных животных равна 84,9±9,6;
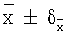 ) определялись по таблице B.C.Генеса [15];
) определялись по таблице B.C.Генеса [15];
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
1. Donnelly J.J., Liu M.A., Ulmer J.B. Antigen presentation and DNA vaccines // Amer. J.Respir. Crit Care Med. - 2000. - Vol.162. - P.190-193.
2. Robinson H.L., Boyle C.A., Feltquate D.M. et al. DNA immunization for influenza virus: studies using hemagglutinin - and nucleoprotein - expressing DNAs vaccine // J. Inf. Dis. - 1997. - Vol.176. - P.50-55.
3. Chang G.J., Davis B.S., Hunt A.R. et al. Flavivirus DNA vaccines: current status and potential // Arm. N.-Y. Acad. Sci. - 2001. - Vol.951. - P.272-285.
4. Kinney R.M., Huang C.Y.H. Development of new vaccines against dengue fever and Japanese encephalitis // Intervirology. - 2001. - Vol.44, №2/3. -P.176-197.
5. Kumar S., Epstein J.E., Richie T.L. et al. A multilateral effort to develop DNA vaccines against falciparum malaria // Trends Parasitol. - 2002. - Vol.18, №3. - P.129-135.
6. Von Herrath M.G., Whitton J.L. DNA vaccination of treatment autoimmune diabetes // Ann. Med. - 2000. - Vol.32, №5. - P.285-292.
7. Hsu C.H., Chua K.Y., Tao M.N. et al. Immunoprophylaxis of allergen-induced immunoglobulin E and airway hyperresponsiveness in vivo by genetic immunization // Nat. Med. - 1996. - Vol.2. - P.540-544.
8. Robinson W.H., Garren H., Utz P.J. et al. Proteomics for the development of DNA tolerising vaccines to treatment to immune disease // Clin. Immunol - 2002. - Vol.103, №1. - P.7-12.
9. Hochshtrasser M. Ubiquitin-dependent protein degradation // J. Annu. Rev. Genetics. - 1996. - Vol.30. - P.405-439.
10. Tanaka К. Molecular biology of proteasomes // Biochem. Biophys. Res. Commun. - 1998. - Vol.247. - P.537-541.
11. Yang Y., 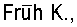 Ahn К., Petrson P.A. In vivo assembly of the proteasomal complexes, implications for antigen processing // J.Biol. Chem. - 1995. - Vol.270. - P.27687-27694.
Ahn К., Petrson P.A. In vivo assembly of the proteasomal complexes, implications for antigen processing // J.Biol. Chem. - 1995. - Vol.270. - P.27687-27694.
12. Salter R.D., Howell D.N., Cresswell P. Genes regulating HLA class I antigen expression in T-B Lymphoblastoid hybrids // Immunogenetics. - 1995. - Vol.21. - P.235-246.
13. Fields Virology. Edited by B.N. Fields, D.M. Knipe, P.M. Howley et al. - Philadelphia, Lippincott-Raven Publishers. - 1996. - 480 p.
14. Маниатис Т., Фрич Э., Сэмбрук ДЖ. Методы генетической инженерии. Молекулярное клонирование / Пер. с англ. под ред. А.А.Баева, К.Г.Скрябина. - М.: Мир, 1984. - 479 с.
15. Генес B.C. Некоторые простые методы кибернетической обработки данных диагностических и физиологических исследований. - М.: Наука, 1998. - 208 с.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pCI-neo-ODC-nsP1 И ЕЕ ВАРИАНТЫ pCI-neo-ODC-E2-L И pCI-neo-ODC-E2-M, КОДИРУЮЩИЕ БЕЛКИ ВИРУСА ВЕНЕСУЭЛЬСКОГО ЭНЦЕФАЛОМИЕЛИТА ЛОШАДЕЙ И БЕЛОК ОРНИТИНДЕКАРБОКСИЛАЗЫ, ПРЕДНАЗНАЧЕННЫЕ ДЛЯ РАЗРАБОТКИ СРЕДСТВ ПРОФИЛАКТИКИ ВИРУСНЫХ ИНФЕКЦИОННЫХ ЗАБОЛЕВАНИЙ (НА ПРИМЕРЕ ВИРУСА ВЭЛ) | 2004 |
|
RU2280691C2 |
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОГЕННЫХ ПРЕПАРАТОВ С ПОМОЩЬЮ ПРОТЕАСОМНОЙ СИСТЕМЫ, ПРЕДНАЗНАЧЕННЫЙ ДЛЯ РАЗРАБОТКИ СРЕДСТВ ПРОФИЛАКТИКИ ВИРУСНЫХ ИНФЕКЦИОННЫХ ЗАБОЛЕВАНИЙ | 2004 |
|
RU2279290C2 |
| ШТАММ РЕКОМБИНАНТНОГО ВИРУСА ОСПОВАКЦИНЫ, ЭКСРЕССИРУЮЩИЙ СТРУКТУРНЫЕ БЕЛКИ ВИРУСА ВЕНЕСУЭЛЬСКОГО ЭНЦЕФАЛОМИЕЛИТА ЛОШАДЕЙ И ПРИГОДНЫЙ ДЛЯ ПРОИЗВОДСТВА ИММУНОБИОЛОГИЧЕСКИХ ПРЕПАРАТОВ И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 1993 |
|
RU2091489C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PC-NS3, ОБЕСПЕЧИВАЮЩАЯ ИНТЕГРАЦИЮ КОМПЛЕКСА ГЕНОВ C, PRM, E, NS1, NS2A, NS2B, NS3 ВИРУСА КЛЕЩЕВОГО ЭНЦЕФАЛИТА (ВКЭ) В ГЕНОМ ВИРУСА ОСПОВАКЦИНЫ (ВОВ) И РЕКОМБИНАНТНЫЙ ШТАММ ВИРУСА ОСПОВАКЦИНЫ, ЭКСПРЕССИРУЮЩИЙ В КЛЕТКАХ ИММУНИЗИРОВАННОГО ОРГАНИЗМА КОМПЛЕКС ГЕНОВ C, PRM, E, NS1, NS2A, NS2B, NS3 ВИРУСА КЛЕЩЕВОГО ЭНЦЕФАЛИТА | 1995 |
|
RU2112038C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PCDNA-TCI, ОБЕСПЕЧИВАЮЩАЯ ЭКСПРЕССИЮ ИСКУССТВЕННОГО ГЕНА TCI В КЛЕТКАХ ЭУКАРИОТ, И РЕКОМБИНАНТНЫЙ АТТЕНУИРОВАННЫЙ ШТАММ БАКТЕРИЙ SALMONELLA ENTERITIDIS E-23/PCDNA-TCI КАК КАНДИДАТ ДЛЯ КОНСТРУИРОВАНИЯ ЖИВОЙ ДНК-ВАКЦИНЫ ПРОТИВ ВИРУСА ИММУНОДЕФИЦИТА ЧЕЛОВЕКА | 2003 |
|
RU2248396C2 |
| Генетическая конструкция на основе оптимизированного гена консенсусного гликопротеина вируса бешенства для профилактики бешенства | 2018 |
|
RU2707544C1 |
| ВАКЦИННЫЙ СОСТАВ, ПОТЕНЦИРОВАННЫЙ КОМБИНАЦИЕЙ ДНК И АНТИГЕНА | 2002 |
|
RU2294212C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pTBI-HBsAg, СОДЕРЖАЩАЯ ХИМЕРНЫЙ ГЕН TBI-HBsAg ПОД КОНТРОЛЕМ ПРОМОТОРА p7.5K ВИРУСА ОСПОВАКЦИНЫ, И ШТАММ РЕКОМБИНАНТНОГО ВИРУСА ОСПОВАКЦИНЫ, ВЫЗЫВАЮЩИЙ ИММУННЫЙ ОТВЕТ ПРОТИВ ВИЧ И ГЕПАТИТА B ЧЕЛОВЕКА В ОРГАНИЗМЕ ЖИВОТНЫХ | 2001 |
|
RU2194075C2 |
| ВАКЦИНА ПРОТИВ ВИЧ/СПИД | 2010 |
|
RU2475264C2 |
| РЕКОМБИНАНТНЫЙ ГЕН L1HPV16, РЕКОМБИНАНТНАЯ ПЛАЗМИДА pQE-L1/16, БЕЛОК L1HPV16 И ИХ ПРИМЕНЕНИЕ | 2016 |
|
RU2628693C1 |
Изобретение относится к области иммунологии, а именно к способу презентации вирусных антигенов иммунной системе хозяина, основанный на протеасомоопосредованной технологии, предназначенному для разработки методов биологической защиты. Сущность изобретения состоит в том, что впервые в Российской Федерации в результате молекулярно-биологических и иммунологических исследований разработан способ ДНК-вакцинации, основанный на протеасомоопосредованной технологии, с использованием рекомбинантных плазмид pCI-neo-ODC-nsP1, pCI-neo-ODC-E2-L, pCI-neo-ODC-E2-M. Рекомбинантные плазмиды pCI-neo-ODC-nsP1, pCI-neo-ODC-E2-L, pCI-neo-ODC-E2-M получены в результате последовательного клонирования ген-эквивалентов неструктурного белка nsP1, фрагментов L, М структурного белка Е2 вируса ВЭЛ соответственно в составе гена орнитиндекарбоксилазы сначала в прокариотический плазмидный вектор рЕТ-23b, а затем в эукариотический плазмидный вектор pCI-neo. Преимущество изобретения заключается в разработке полинуклеотидных вакцин, индуцирующих клеточноопосредованный иммунный ответ. 4 табл., 2 ил.
Способ презентации антигенов вируса Венесуэльского энцефаломиелита лошадей иммунной системе хозяина, основанный на протеасомоопосредованной технологии, характеризующийся тем, что ДНК-вакцинацию осуществляют с использованием рекомбинантных плазмид, содержащих ген орнитиндекарбоксилазы и ген-эквивалент неструктурного белка nsP1, или ген-эквивалент фрагмента L, или ген-эквивалент фрагмента М гликопротеина оболочки Е2 вируса ВЭЛ, выбранных из группы: соответственно pCI-neo-ODC-nsP1, pCI-neo-ODC-E2-L, pCI-neo-ODC-E2-M, имеющих номера №№02/1, 02/2, 02/3, зарегистрированных в коллекции Вирусологического центра НИИ микробиологии Министерства обороны Российской Федерации или сочетанием плазмид pCI-neo-ООС-Е2-L, pCI-neo-ODC-E2-М.
| УСТРОЙСТВО ДЛЯ ОБРАБОТКИ КРУП | 2007 |
|
RU2337755C1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| ШТАММ РЕКОМБИНАНТНОГО ВИРУСА ОСПОВАКЦИНЫ, ЭКСРЕССИРУЮЩИЙ СТРУКТУРНЫЕ БЕЛКИ ВИРУСА ВЕНЕСУЭЛЬСКОГО ЭНЦЕФАЛОМИЕЛИТА ЛОШАДЕЙ И ПРИГОДНЫЙ ДЛЯ ПРОИЗВОДСТВА ИММУНОБИОЛОГИЧЕСКИХ ПРЕПАРАТОВ И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 1993 |
|
RU2091489C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК P UHBС, КОДИРУЮЩАЯ ГЕН БЕЛКА КОРОВОГО АНТИГЕНА ВИРУСА ГЕПАТИТА В, ПОЗВОЛЯЮЩАЯ ЭКСПОНИРОВАТЬ ЧУЖЕРОДНЫЕ ЭПИТОПЫ НА ПОВЕРХНОСТИ КОРА. | 1995 |
|
RU2121504C1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| Электромагнитный насос | 1923 |
|
SU811A1 |

