Изобретение относится к биотехнологии, конкретно к генетической инженерии, и представляет собой плазмиду pcDNA-TCI, содержащую искусственный ген TCI, кодирующий белок-иммуноген, состоящий из множественных цитотоксических Т-клеточных эпитопов ВИЧ-1, и обеспечивающую его экспрессию в клетках эукариот, а также рекомбинантный аттенуированный штамм бактерий Salmonella enteritidis E-23/pcDNA-TCI, являющийся вектором доставки и представления выше указанной плазмиды иммунокомпетентным клеткам организма млекопитающих. Предложенный рекомбинантный штамм микроорганизмов является кандидатом для конструирования живой ДНК-вакцины против ВИЧ-1.
Среди новых направлений в создании вакцин против ВИЧ весьма перспективными считаются ДНК-вакцины [1]. К настоящему времени уже получен ряд кандидатных ДНК-вакцин против ВИЧ и показана способность таких вакцин индуцировать ВИЧ-специфический иммунный ответ на лабораторных животных [2, 3]. В то же время эксперименты по ДНК-вакцинации обозначили определенные проблемы. Одна из основных проблем ДНК-вакцинации связана с доставкой плазмиды в организм хозяина, поскольку при традиционном парентеральном введении ДНК-вакцины для достижения достаточно высокого иммунного ответа приходится вводить большие количества плазмидной ДНК, что может привести к появлению ауто-антител на собственную ДНК организма. Наиболее перспективным способом представления ДНК-вакцин иммунокомпетентным клеткам могут быть аттенуированные штаммы сальмонелл [4, 5, 6, 7].
В настоящее время создан ряд различных аттенуированных штаммов сальмонелл. Эти штаммы способны инвазировать лимфоидную ткань кишечного тракта, но обнаруживают пониженную патогенность даже при больших дозах [8, 9].
Известен штамм бактерий Salmonella typhimurium SL 7207, который несет ДНК-вакцину (плазмиду pRc/CMV-env, кодирующую gp120 ВИЧ-1) [10]. Штамм вводили мышам per os. В результате у мышей регистрировали выраженный ВИЧ-1 специфический CD8+ иммунный ответ, который выявляли как в мукозальной, так и в системной лимфоидной ткани. При внутримышечной иммунизации вышеуказанной плазмидной ДНК-вакциной регистрировали только системный CD8+ ответ, который был ниже, чем при использовании сальмонелл в качестве вектора доставки. Однако следует отметить, что существенным недостатком данного аналога является то, что не показана индукция специфических анти-ВИЧ-1 антител, а также не определена вируснейтрализующая активность сывороток иммунизированных животных.
Известен рекомбинантный аттенуированный штамм Salmonella enteritidis E-23 ВМС120, который несет ДНК-вакцину в виде плазмиды рВМС120, обеспечивающей синтез белка gp 120 вируса иммунодефицита человека в клетках млекопитающих [11, прототип]. Штамм обладает высокой способностью вызывать выраженную и действенную иммунизацию к ВИЧ-1. Однако есть существенная проблема, препятствующая получению эффективной вакцины против ВИЧ на основе только одного белка gp120, связанная с его поразительной антигенной изменчивостью [12]. В результате вирус изменяет свою антигенную структуру быстрее, чем иммунная система успевает переключиться на новые антигенные варианты возбудителя.
Подходящим кандидатом в белки-иммуногены является искусственный полиэпитопный белок TCI (Т cell immunogen) [13]. При проектировании TCI-иммуногена в рассмотрение принимались эпитопы, которые удовлетворяют определенным критериям. В частности, были отобраны эпитопы, которые в совокупности были ограничены, по крайней мере, десятью различными антигенами главного комплекса гистосовместимости (МНС) I класса, которые полностью покрывают генетическое разнообразие HLA-молекул I класса в популяциях Caucasian и Oriental (Mills et al. 1994). Длина белка TCI составляет 392 аминокислотных остатка. Белок содержит более восьмидесяти эпитопов (как CD8+ CTL, так и CD4+ Th), многие из которых перекрываются [13, 14].
Известна рекомбинантная плазмида pET-TCI, содержащая искусственный ген TCI, кодирующий целевой белок-иммуноген TCI [13]. Однако данная плазмидная конструкция предназначена для экспрессии указанного гена в клетках прокариот.
Технической задачей изобретения является создание рекомбинантного штамма сальмонелл, обеспечивающего доставку ДНК-вакцины в виде рекомбинантной плазмидной ДНК, кодирующей искусственный белок-иммуноген, TCI, содержащий набор большого числа основных антигенов ВИЧ-1, и вызывающего индукцию как специфического цитотоксического клеточного ответа, так и нейтрализующих ВИЧ-1 антител при введении животным, т.е. являющегося кандидатом в живую вакцину против ВИЧ-1.
Поставленная задача решается путем конструирования рекомбинантной плазмидной ДНК pcDNA-TCI, несущей ген искусственного полиэпитопного белка TCI под контролем промотора CMV, обеспечивающего экспрессию последнего в клетках млекопитающих, которая имеет размер 6657 п.н. и состоит из следующих фрагментов (фиг.1):
- NheI - HindIII - векторного фрагмента ДНК плазмиды pcDNA 3.1-mycchislacZ (-) [Invitrogen, США] размером 5423 п.н., содержащего промотор CMV и последовательность BGH poly А, обеспечивающие экспрессию гена TCI в клетках млекопитающих; ген устойчивости к ампициллину (bа) и pMBI ori, обеспечивающие селекцию и размножение целевой плазмиды в клетках бактерий Escherichia и Salmonella;
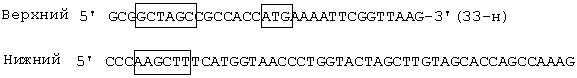
- NheI - HindIII - фрагмента размером 1234 п.н., содержащего ген TCI и последовательность Козак с инициирующим кодоном АТGразмером, полученного путем ПЦР плазмиды pET-TCI с использованием олигонуклеотидных праймеров, структура которых приведена ниже:
(49-н)
и обработки рестриктазами NheI - HindIII;
- генетические маркеры: ген устойчивости к ампициллину (blа);
- уникальные сайты рестрикции: NruI-209, NheI-896, BstEII-2089, ВаmHI 2112, ЈcoRI-2121, НindIII-2130, ApaI-2140, PstI- 3544, NarI-3491, SmaI-3304.
Молекулярная масса целевой рекомбинантной плазмидной ДНК равна 4,3×103 кДа.
Положение генов в плазмиде pcDNA-TCI: TCI - начало (896) - конец (2129); Neo-r - начало (3362) - конец (4156); Bla - начало (5661) - конец (6521); BGH polyA - начало (2250) - конец (2477); pMBI - начало (4843) - конец (5516); эукариотический промотор CMV pr - начало (209) - конец (863).
После трансформации клеток E.coli JM103 и рестрикционного отбора ДНК клонов была получена целевая плазмида pcDNA-TCI, несущая искусственный ген TCI и обеспечивающая его экспрессию в клетках эукариот. Полученная плазмида pcDNA-TCI обеспечивает синтез белка TCI молекулярной массой 44 кДа.
Известен аттенуированный штамм Salmonella enteritidis E-23 (cya, CRP), обладающий пониженной вирулентностью [15]. На мышах, телятах и обезьянах показано, что вирулентность этого штамма снижена примерно в 1000 раз по сравнению с родительским штаммом, при сохранении инвазивной активности. Этот штамм по ряду основных свойств может быть использован для решения поставленной задачи, т.е. для создания на его основе вакцины против ВИЧ-1.
Поставленная задача решается генетической трансформацией штамма Salmonella enteritidis E-23 (cya, CRP) плазмидой pcDNA-TCI, кодирующей белок TCI под контролем цитомегаловирусного промотора и обеспечивающей наработку этого белка в клетках млекопитающих. При введении полученного штамма лабораторным животным происходит индукция специфических анти-ВИЧ антител и специфического клеточного ответа.
Сконструированный штамм получил обозначение Salmonella enteritidis E-23/ pcDNA-TCI и депонирован под номером В-883 в коллекции НИИ ККМ ГНЦ ВБ "Вектор".
Культурально-морфологические признаки
Морфология: грамотрицательные палочки, средних размеров, расположены хаотично.
Культуральные свойства: растет на мясо-пептонном агаре, глюкозоминимальном агаре Девиса, средах Мак-Конки, Левина, Кауфмана, образует колонии в S-форме мелких размеров через 18 часов роста при 37°С. В мясо-пептонном бульоне, желчном бульоне, жидкой глюкозоминимальной среде А (Девиса) - диффузное помутнение через 18 часов роста. В цитратминимальной среде (Девиса) - отсутствие роста.
Резистентность к антибиотикам: на всех вышеперечисленных средах растет в присутствии ампициллина (100 мкг /мл).
Антигенная структура:
O9,12 Hg,m
Биохимические свойства:
Не утилизирует цитрат на агаре Симмонса, не продуцирует индол, не расщепляет мочевину, не обладает фенилаланилтрансаминазной активностью, реакция Фогес-Проскауэра - отрицательная; продуцирует сероводород в течение 48 часов, обладает лизин- и орнитилкарбоксилазными активностями и аргининдегидролазной активностью; ферментирует глюкозу, не ферментирует лактозу, сахарозу, мальтозу, сорбит, инозит, дульцит, адонит; в течение 48 часов ферментирует рамнозу, манит, трегалозу. Желатину не разжижает.
Чувствительность к бактериофагам:
Чувствителен к бактериофагу Р22.
Плазмидный профиль:
Плазмида pcDNA-TCI с молекулярной массой 4,3×103 кДа обеспечивает синтез белка TCI в клетках млекопитающих.
Изобретение иллюстрируется следующими графическими материалами.
Фиг.1. Рекомбинантная плазмидная ДНК pcDNA-TCI и схема ее конструирования.
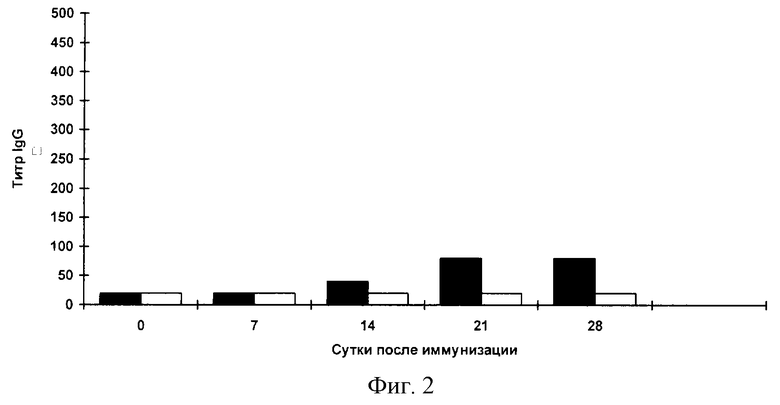
Фиг.2. Анализ системного гуморального ответа к очищенному вирусу ВИЧ-1 сывороток мышей, иммунизированных штаммом Salmonella enteritidis E-23/ pcDNA-TCI по данным ИФА.
 S.enteritidis E-23/ pcDNA-TCI
S.enteritidis E-23/ pcDNA-TCI
 S. enteritidis E-23
S. enteritidis E-23
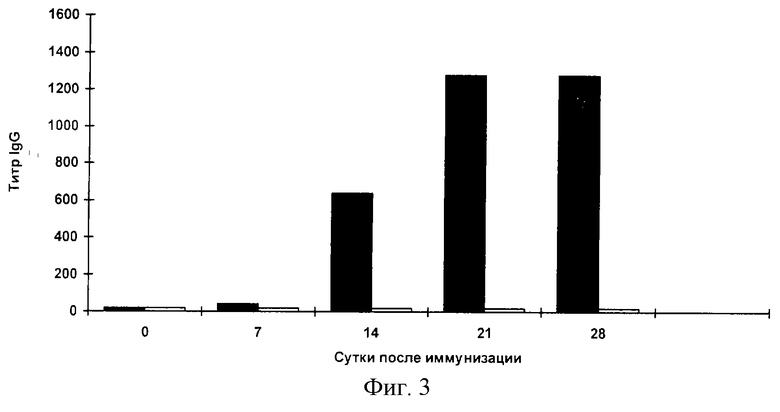
Фиг.3. Анализ системного гуморального ответа к очищенному белку TCI сывороток мышей, иммунизированных штаммом Salmonella enteritidis E-23/ pcDNA-TCI по данным ИФА.
 S. enteritidis E-23/ pcDNA-TCI
S. enteritidis E-23/ pcDNA-TCI
 S. enteritidis E-23
S. enteritidis E-23
Для лучшего понимания сущности изобретения ниже следуют примеры его осуществления.
Пример 1. Конструирование рекомбинантной плазмидной ДНК pcDNA-TCI.
Для конструирования целевой плазмиды pcDNA-TCI используют плазмиду pcDNA 3.1-mycchislacZ (-) [Invitrogen, США]. Фрагмент ДНК, содержащий ген TCI, получают путем ПЦР плазмиды pET-TCI [13], с использованием олигонуклеотидных праймеров (фиг.1). ПЦР проводят в 0,5 ml пробирках (BioTip Germany) с использованием следующих стоковых растворов:
dNТР - 10 mМ (СибЭнзим Россия)
Буфер - 10-кратный (СибЭнзим Россия)
Праймеры - 13 рМ/μ1
Thermus Aquaticus DNA Polymerase (Taq pol) - 5 U/μ1 (СибЭнзим Россия)
MgCl2 - 20 mM
Состав реакционного буфера (1-кратного): 60 mM Tris-HCl (pH 8,5 при 25°С), 25 mM KCl, 1,5 mM MgCl2, 10 mM 2-меркапоэтанол, 0,1% Тритон Х-100. Реакцию проводят на амплификаторе БИС-109 по следующей программе:
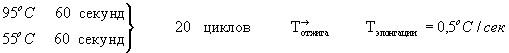
Последний цикл такой же. за исключением времени элонгации при 72°С - 4 мин. 0,5 мкг ПЦР фрагмента гена TCI обрабатывают рестриктазами NheI - НindIII и лигируют в стандартных условиях с 0,1 мкг плазмиды pcDNA 3.1-mycchislacZ (-), гидролизованной рестриктазами NheI - HindIII. Лигазной смесью трансформируют компетентные клетки E.coli JM103 и из клонов, выросших на среде с ампициллином, выделяют плазмидную ДНК и подвергают рестрикционному анализу с помощью ферментов NheI - HindIII, BglII и PvuII. Соответствие последовательности нуклеотидов встройки в отобранной рекомбинантной плазмиде проверяют секвенированием.
Пример 2. Получение рекомбинантного штамма Salmonella enteritidis E-23/ pcDNA-TCI.
Трансформация.
Электропорация. Культуру клеток Salmonella enteritidis E-23 выращивают в среде YT до ОП=0,5, охлаждают, 1 мл культуральной жидкости центрифугируют 2 мин при 6000 об/мин на центрифуге Eppendorf. Осадок клеток ресуспендируют в 800 мкл охлажденной, бидистилированной воды и осаждают центрифугированием 1 мин при 6000 об/мин. Процедуру промывки повторяют 5 раз. Затем осадок ресуспендируют в 200 мкл 10% глицерина, центрифугируют 1 мин при 6000 об/мин и ресуспендируют в 40 мкл 10% глицерина.
К суспензии клеток добавляют 0,5 мкг плазмидной ДНК. Образец помещают в кювету для электропорации (время импульса составляет 4 с при напряжении 2,5 В). Для электропорации используют электропоратор фирмы BIORAD. Затем добавляют но 300 мкл YT среды и инкубируют 1 час при 37°С. Клетки рассевают на StI-агар с ампициллином (100 мкг/мл). Выросшие на агаре с ампициллином колонии обозначают как Salmonella Enteritidis E-23/ pcDNA-TCI. Все манипуляции с плазмидной ДНК проводят согласно описанным методикам [16].
Пример 3. Исследование способности целевого штамма индуцировать гуморальный иммунный ответ.
Культивирование
Для иммунизации клетки S.enteritidis E-23/ pcDNA-TCI выращивают следующим образом: отдельную колонию сальмонеллы засевают в 50 мл среды YT, содержащей ампициллин (100 мкг/мл), и инкубируют 16 ч при 37°С; клетки осаждают центрифугированием и ресуспендируют в 1 мл физиологического раствора. Концентрацию клеток определяют титрованием.
Иммунизация. Для иммунизации используют мышей линии BALB/c. весом 12-15 грамм, которые были получены из вивария ГНЦ ВБ "Вектор". Животные содержались на стандартном рационе.
Иммунизацию животных полученным штаммом проводят однократно, перректально, введением 108 клеток S. enteritidis E-23/ pcDNA-TCI на мышь. Забор крови производят на 0, 7, 14, 21, 28, 35 сутки после введения сальмонелл.
Иммуноферментный анализ. Специфичность образующихся антител в сыворотках мышей анализируют с помощью ИФА. Для выявления антител против ВИЧ-1 используют очищенный инактивированный вирус ВИЧ-1 и очищенный белок TCI. В каждом эксперименте берут образцы крови от трех животных. После получения сыворотки образцы смешивают, после чего определяют титр специфических антител. Титр определяют как превышение титра сывороток мышей, иммунизированных S. enteritidis E-23/ pcDNA-TCI над титром сывороток мышей, иммунизированных исходным штаммом S.enteritidis E-23. На каждую точку делают два повтора. Результаты представлены на фиг.2,3. Начиная с третьей недели после иммунизации мышей клетками S.enteritidis E-23/ pcDNA-TCI обнаруживается продукция IgG к ВИЧ-1, причем продукция антител остается на высоком уровне на протяжении всего срока наблюдения.
Пример 4. Исследование способности целевого штамма индуцировать клеточный иммунный ответ.
Реакция бласттрансформации. Реакцию бласттрансформации спленоцитов проводят па 0, 7, 14, 21 сутки после введения сальмонелл по ранее описанной методике [17]. В качестве специфических антигенов используют: белок TCI в концентрации 2 мкг/мл; пептид N15 (DRVIEVVQGAYRAIR), пептид N16 (KQIINMWQEVGKAMYA). Последовательности пептидов N15, и N16 взяты из Los Alamos HIV Molecular Immunology Database (http://hiv-web.lanl.gov/). Они были специально включены в структуру белка TCI, так как представляются иммунной системе (СTL лимфоцитам) в комплексе с МНС I класса H-2d, что позволяет тестировать CTL ответы на эти пептиды на BALB/c мышах [13]. В качестве неспецифического антигена (отрицательный контроль) используют антиген ЕНЕС (неинфекционный штамм E.coli О:H157) - в концентрации 2 мкг/мл. В лунку планшета вносят по 100 мкл каждого компонента. В качестве митогена используют конканавалин А (производства "Sigma", США) в концентрации 5 мкг/мл. Стимулированные лимфоциты культивируют в течение 76 ч. За 18 ч до окончания культивирования добавляют МТТ (3-[4,5-dimethylthiazol-2-yl]2,5-diphenyltetrazolium bromide, производства "Sigma", США) в концентрации 2 мкг на лунку. По окончании срока культививирования в лунки вносят по 100 мкл изопропанола с добавлением 0.2 М НСl. Планшет выдерживают 18 ч в темноте при комнатной температуре, измеряют оптическую плотность в лунках при длине волны 570 нм и длине референс-волны 630 нм на многоканальном вертикальном автоматическом спектрофотометре Bio-Tec L-808U. Обработку результатов проводят с помощью программы КС-4 Bio-Tec.
Пролиферативную активность спленоцитов оценивают по индексу стимуляции, который рассчитывают как отношение средних значений абсолютных чисел DeltaOD стимулированных к среднему значению абсолютных чисел DeltaOD не стимулированных спленоцитов. Достоверным увеличением пролиферативной активности является 50% увеличение индекса стимуляции по отношению к аналогичному показателю в интактном контроле (таблица 1).
Результаты, приведенные в таблице 1, свидетельствуют о том, что иммунизация клетками S.enteritidis E-23/ pcDNA-TCI вызывает формирование клона клеток памяти, поскольку индекс стимуляции клеток in vitro специфическими белками ВИЧ достоверно увеличивается (Р<0.01) на 14 сутки по сравнению с показателями на 0 сутки (день проведения иммунизации). Этот показатель сохраняется на высоком уровне на всем сроке наблюдения.
Определение количества CTL лимфоцитов на основе реакции ELISPOT
При постановке реакции ELISPOT на первом этапе проводят сорбцию анти-INF-γ MAT с концентрацией 5 мкг/мл на лунку 96-луночного планшета ImmunoSpot M200. После инкубации в течение 12 ч при 4°C каждую лунку дважды промывают раствором PBS и блокируют средой RPMI 1640, содержащей 10% фетальную бычью сыворотку, в течение 2 ч. В качестве клеток-эффекторов используют спленоциты иммунизированных животных в концентрации 106/мл. Для стимуляции продукции INF-γ суспензией клеток используют белок TCI (1 мкг/мл) и два пептида: N15 (DRVIEVVQGAYRAIR), N16 (KQIINMWQEVGKAMYA), для оценки неспецифической продукции используют пептид ЕНЕС (отрицательный контроль). Клетки культивируют в присутствии 5% СО2 при 37°С в течение 24 ч. INF-γ-секретирующие клетки визуализируют, используя 0,5 мкг/мл биотинилированнных анти-INF-γ антитела и 0,25 мкг/мл конъюгата Avidin-HRP. Окрашивание производят добавлением субстратов для пероксидазы (4-хлор-1-нафтол, диаминобензидин фосфат). Реакцию останавливают удалением реагентов, лунки промывают 3 раза дистиллированной водой. Подсчет количества INF-γ-продуцирующих клеток осуществляют с помощью микроскопа. Результаты экспериментов сведены в таблицу 2.
Пример 5. Оценка вируснейтрализующей активности сывороток.
Вируснейтрализующую активность сывороток определяют методом учета ингибирования репродукции вируса ВИЧ-1 по снижению его инфекционного титра. Для этого используют перевиваемые лимфобластоидные клетки человека МТ-4. Клетки культивируют в концентрации 3.0-5.0×105 клеток в 1 мл среды RPMI 1640 с 10% сыворотки эмбрионов коров, 100 мкг/мл гентамицина. В реакции вируснейтрализации используют штамм ВИЧ-1899А (тип Х4, субтип В), полученный из коллекции штаммов вирусов иммунодефицита человека НИИ вирусологии им. Д.И.Ивановского РАМН. Множественность заражения составляет 100 ТЦД50. Сыворотки инактивируют на водяной бане при температуре 56°С в течение 30 мин. В лунки пластиковой 96-луночной панели (“Costar”, США) вносят по 50 мкл следующих разведений сывороток: 1:10, 1:20, 1:40, 1:80, 1:160, 1:320, 1:640, 1:1280, т.д и добавляют 50 мкл суспензии вируса. Смесь инкубируют в течение 1 ч при 37°С. После инкубации вносят по 100 мкл взвеси чувствительных клеток МТ-4 в количестве 90-100×103 кл/лунку. Панель инкубируют при 37°С в атмосфере с 5% СО2 и 98% влажности в течение 5-7 суток до момента учета результатов. В качестве контролей используют: “контроль вируса” - вирус с добавлением среды без сыворотки, “контроль клеток” - клетки в питательной среде, “контроль цитотоксичности сывороток” - клетки + сыворотки. Учет результатов проводят на 5-7 день после начала эксперимента. Ингибирование репродукции вируса оценивают по снижению его инфекционного титра, который определяют по ТЦД50 (50% тканевая цитопатическая доза). Данные по определению нейтрализующих титров антител в сыворотках крови иммунных животных сведены в таблицу 3.
Данные, представленные в таблице 3, свидетельствуют о том, что образующиеся после введения рекомбинантного штамма антитела обладают вируснейтрализующеи активностью.
Таким образом, новый сконструированный штамм Salmonella enteritidis E-23/ pcDNA-TCI при введении животным индуцирует выраженный специфический гуморальный и клеточный иммунный ответ к ВИЧ-1. Предложенный рекомбинантный штамм может быть использован для конструирования живой ДНК- вакцины против ВИЧ-1.
Литература
1. Bowers M. DNA and other technologies for HIV vaccines. BETA. 1998, 9: 12-28.
2. Hanke Т., Neumann V.C., Blanchard T.J. et al. Effective induction of HIV-specific CTL by multi-epitope using gene gun in a combined vaccination regime. Vaccine. 1999, 17: 589-596.
3. Robinson H.L, Montefiori D.C., Johnson R.P. et al. Neutralizing antibody-independent containment of immunodeficiency virus challenges by DNA priming and recombinant pox virus booster immunizations. Nat. Med. 1998, 5: 526-534.
4. Darji A., Guzman С.A., Gerstel В. et al. Oral somatic transgene vaccination using attenuated S.typhimurium. Cell. 1998, 91: 765-775.
5. Paglia P., Medina E., Arioli I. et al. Gene transfer in dendritic cells, induced by oral DNA vaccination with Salmonella typhimurium, results in protective immunity against a murine fibrosarcoma. Blood. 1998. 92: 3172-3176.
6. Sizemore D.R., Branstrom A.A., Sadoff J.C. Attenuated Shigella as a DNA delivery vehicle for DNA-mediated immunization. Science. 1995, 270: 299-302.
7. Xiang R,, Lode H.N., Chao Т.Н. et al. An autologous oral DNA vaccine protects against murine melanoma. Proc. Natl. Acad. Sci. 2000, 97: 5492-5497.
8. Levine M.M., Dougan G. Optimism over vaccines administered via mucosal surfaces. Lancet. 1998, 351: 1375-1376.
9. Mascola J., Nadel G. Vaccines for the prevention of HIV-1 disease. Current Opinion in Immunology. 2001, 13: 489-494.
10. Shata M.T, Reitz M.S Jr, DeVico A.L. et al. Mucosal and systemic HIV-1 Env-specific CD8(+) T-cells develop after intragastric vaccination with a Salmonella Env DNA vaccine vector. Vaccine. 2001, 20: 623-629.
11. Патент РФ №2192277, кл. А 61 К 39/12, опубл. БИ № 31 от 10.11.2002 г.
12. Letvin N.L. Strategies for an HIV vaccine. J. Clin. Invest. 2002, 110: 15-20.
13. Заявка на патент РФ №2002110800/13 от 22.04.2002 г., кл. С 12 N 15/49.
14. Bazhan S.I., Ilyichev A.A., Pokrovsky A.G. et al. The current approaches to development of an effective and safe vaccine against HIV-1. IV ISTC scientific advisory committee seminar on basic science in ISTC activities. Akademgorodok, Novosibirsk. 23-27 April, 2001. P. 19.
15. Бойченко М.Н., Воробьев А.А., Тымчук С.Н. Перспектива получения мутантов суа и crp сальмонелл для использования в качестве вакцинных штаммов. Вестн. РАМН. 1995, 10: 37-39.
16. Маниатис Т., Фрич Э., Сэмбрук Дж. Молекулярное клонирование. М.: Мир. - 1984.
17. Хоробрых В.В., Пронин А.В., Киркин А.Ф. и др. Методы постановки реакции бласт-трансформации в микромодификации. Иммунология. 1983, 3: 76-79.
Изобретение относится к области биотехнологии, конкретно к генетической инженерии. Предложена плазмидная рекомбинантная ДНК pcDNA-TCI. Рекомбинантная плазмидная ДНК характеризуется массой 4,3×103 кДа и имеет размер 6657 п.н. Предложенная рекомбинантная плазмидная ДНК обеспечивает экспрессию искусственного гена TCI, содержащего цитотоксические Т-клеточные эпитопы ВИЧ-1, в клетках эукариот. Кроме того, предложен рекомбинантный аттенуированный штамм бактерий Salmonella enteritidis E-23 pcDNA-TCI. Предложенный штамм получают путем генетической трансформации штамма Salmonella enteritidis E-23 плазмидой pcDNA-TCI. Штамм обеспечивает доставку в клетки эукариот ДНК-вакцины в виде рекомбинантной плазмидной ДНК pcDNA-TCI. Предложенная группа изобретений позволяет индуцировать в организме выраженный специфический гуморальный и клеточный иммунный ответ к ВИЧ-1. Предложенная группа изобретений может быть использована в медицине и вирусологии для конструирования живой ДНК-вакцины против ВИЧ-1. 2 н.п. ф-лы, 3 ил., 3 табл.
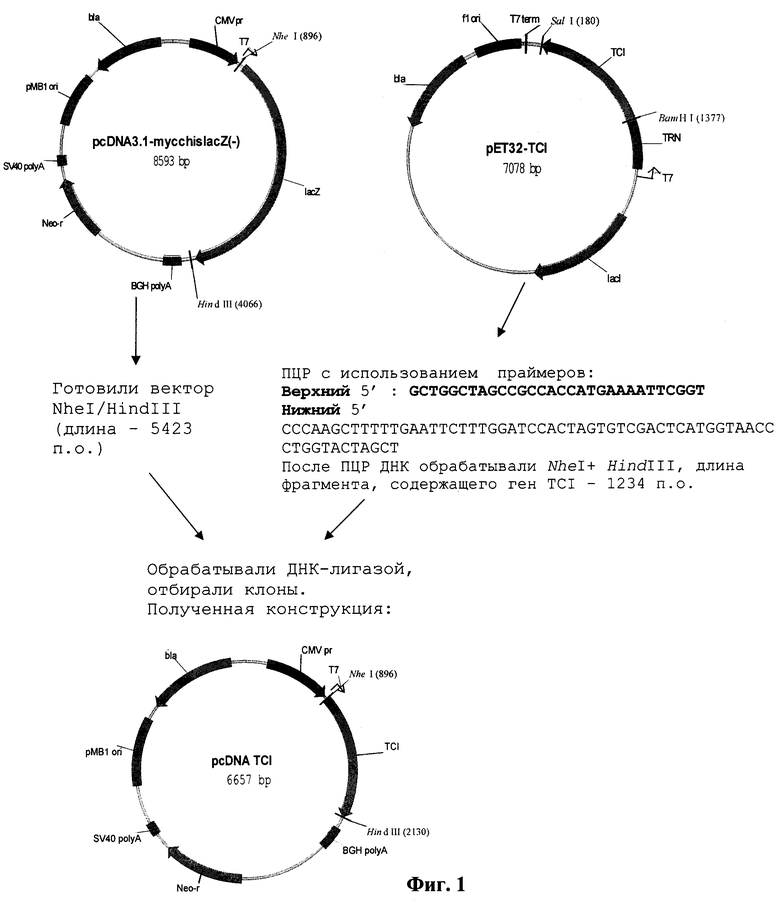
Nhel - HindIII - векторного фрагмента ДНК плазмиды pcDNA 3.1-mycchislacZ (-) [Invitrogen, США] размером 5423 п.н., содержащего промотор CMV и последовательность BGH poly А, обеспечивающих экспрессию гена TCI в клетках млекопитающих; ген устойчивости к ампициллину (bla) и pMBI ori, обеспечивающих селекцию и размножение целевой плазмиды в клетках бактерий Escherichia и Salmonella;
Nhel - HindIII - фрагмента размером 1234 п.н., содержащего искусственный ген TCI и последовательность Козак с инициирующим кодоном ATG, полученного путем ПЦР плазмиды рЕТ- TCI с использованием олигонуклеотидных праймеров:
Верхний
5'GCGGCTAGCCGCCACCATGAAAATTCGGTTAAG-3'(33-н)
Нижний
5'CCCAAGCTTTCATGGTAACCCTGGTACTAGCTTGTAGCACCAG CCAAAG
(49-н)
и обработкой рестриктазами Nhel - HindIII;
генетические маркеры: ген устойчивости к ампициллину (blа);
уникальные сайты рестрикции: Nrul-209, Nhel-896, BstEII-2089, BamHI-2112, Ecorl-2121, HindIII-2130, Apal-2140, Pstl-3544, Narl-3491, Smal-3304.
гены, имеющие положение в плазмиде pcDNA-TCI:
TCI - начало (896) - конец (2129); Neo-r - начало (3362) - конец (4156); Bla - начало (5661) - конец (6521); BGH polyA - начало (2250) - конец (2477); pMBI - начало (4843) - конец (5516); эукариотический промотор CMV pr - начало (209) - конец (863).
| ШТАММ БАКТЕРИЙ SALMONELLA ENTERIDITIS E-23 BMC120 ДЛЯ ИММУНИЗАЦИИ ПРОТИВ ВИРУСА ИММУНОДЕФИЦИТА ЧЕЛОВЕКА | 2001 |
|
RU2192277C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PUE 41, КОДИРУЮЩАЯ ФРАГМЕНТЫ БЕЛКОВ ОБОЛОЧКИ GP 120 И GP 41 ВИРУСА ИММУНОДЕФИЦИТА ЧЕЛОВЕКА ТИПА I И ШТАММ БАКТЕРИЙ ESCHERICHIA COLI - ПРОДУЦЕНТ ФРАГМЕНТОВ БЕЛКОВ ОБОЛОЧКИ GP 120 И GP 41 ВИРУСА ИММУНОДЕФИЦИТА ЧЕЛОВЕКА ТИПА I | 1990 |
|
SU1766070A1 |
| Рекомбинатная плазмидная ДНК pGp 120 - 428, кодирующая гибридный белок с антигенными свойствами белка @ р 120 ВИЧ-1 | 1991 |
|
SU1789562A1 |

