Данное изобретение относится к способу идентификации новой мишени для применения при разработке терапевтических форм и лекарственных средств, эффективных против туберкулеза. Конкретно, данное изобретение относится к способу идентификации новой и ранее не известной мишени при разработке терапевтических форм, включая противотуберкулезные лекарственные средства, особенно имеющие отношение к существованию персистенции микобактерий при гипоксии.
Туберкулез является ведущей причиной смерти от одного инфекционного агента и ежегодно убивает более 3 миллионов человек по всему миру. В 1998 году число случаев туберкулеза в Индии, по оценкам, составляло 2078076, из которых 935134 случая, вероятно, являлись инфекционными. Последствия инфекции, очевидно, представляют собой итог длительного взаимодействия между патогенным микроорганизмом и иммунной защитой хозяина. В большинстве случаев у инфицированного субъекта развивается эффективный иммунный ответ, кульминацией которого является образование гранулемы вокруг участков инфицирования и остановка прогрессирования заболевания. Условия внутри гранулемы, как предполагается, представляют собой гипоксию. Клинические исследования показывают, что бациллы внутри таких гранулем не погибают, но вместо этого остаются в состоянии покоя. Это называется латентной инфекцией. Примерно в 10% случаев латентная инфекция реактивируется, что приводит к активному инфекционному заболеванию спустя месяцы или годы после первоначального инфицирования. Большое число латентно инфицированных субъектов сильно препятствует снижению распространения туберкулеза и скорости переноса M.tuberculosis. Адаптация M.tuberculosis в течение спектра инфекции и заболевания, вероятно, осуществляется через конкретные генетические пути, которые модулируются специфичными физиологическими условиями и условиями среды в тканях хозяина.
Имеется срочная необходимость в понимании данных путей для того, чтобы разработать новые и более направленные стратегии профилактики, контроля и лечения туберкулеза. Общепринятые лекарственные средства направлены на пути, требуемые для бактериального роста и деления, такие как пути биосинтеза клеточной стенки и репликации ДНК. Полагают, что их низкая активность в отношении медленнорастущих или нерастущих бактерий является важной причиной, по которой применяемые в настоящее время режимы требуют так много времени для эрадикации инфекции.
Гены devR-devS, соответственно обозначенные как Rv333133c и Rv3132c в аннотированном геноме M.tuberculosis, как предсказано, кодируют регулятор ответа DevR и сенсор гистидинкиназы DevS, соответственно. Данная генетическая система была ранее идентифицирована в лаборатории авторов изобретения путем вычитающей гибридизации с использованием РНК из вирулентных и авирулентных штаммов M.tuberculosis (ссылка 1). Авторы изобретения описывают способ идентификации данной системы в качестве новой и не известной ранее мишени для разработки терапевтических форм, включая противотуберкулезные лекарственные средства, особенно имеющие отношение к существованию персистенции микобактерий при гипоксии.
Поэтому главной целью изобретения является идентификация мишени, которая ответственна за рецидив/реактивацию у пациента данного заболевания, или той мишени, что позволяет данному организму адаптироваться к гипоксии.
Другой целью данного изобретения является идентификация мишени, ответственной за рецидив/реактивацию у пациента данного заболевания, или той мишени, что позволяет данному организму адаптироваться к гипоксии, и разработка терапевтических форм и противотуберкулезных лекарственных средств.
По данному изобретению предоставляется способ идентификации новой мишени для применения при разработке терапевтических форм и лекарственных средств, эффективных против туберкулеза, включающий:
I. разрушение гена devR (Rv3133c), расположенного во вставке EcoRI-HindIII размером ˜3,3 т.п.н. плазмиды pJT53.34, кассетой устойчивости к канамицину (KmR),
II. конструирование pJQ200SkdevR::kan из разрушенного гена devR,
III. введение указанной плазмиды в M.tuberculosis H37Rv путем электропорации,
IV. селекцию трансформантов с одним кроссинговером, указывающим на интеграцию плазмиды, на чашках с агаром middle brook 7H10, содержащих 20 мкг/мл канамицина,
V. их анализ путем полимеразной цепной реакции (PCR) на наличие последовательностей генов devR, KmR и гена устойчивости к сахарозе (SacB),
VI. проведение Саузерн-анализа в отношении данных последовательностей с зондом devR, зондом devS, зондом гена устойчивости к канамицину, так что выявляется M.tuberculosis Dup devR, содержащая копию дикого типа и разрушенную копию локуса devR,
VII. выращивание M.tuberculosis Dup devR в среде middle brook 7H9, содержащей 20 мкг/мл канамицина и 2% сахарозы, в течение 7 суток,
VIII. помещение указанного выращенного штамма M.tuberculosis Dup devR на множество чашек со средой middle brook 7H10, содержащей 20 мкг/мл канамицина и 2% сахарозы, с получением трансформантов, устойчивых к канамицину,
IX. проведение Саузерн-гибридизации в отношении указанной выращенной M.tuberculosis devR с последующей процедурой полимеразной цепной реакции для подтверждения указанного аллельного обмена,
X. проведение анализа путем полимеразной цепной реакции в отношении указанных трансформантов на предмет разрушенного гена devR::kan,
XI. проведение вестерн-блоттинга и иммунной электронной микроскопии в отношении указанного разрушенного гена devR::kan для подтверждения функционального разрушения указанного гена,
XII. оценку способности к росту мутантного по devR штамма M.tuberculosis в условиях ограничения количества кислорода при экспрессии генов devR и devS,
XIII. оценку роста и жизнеспособности указанного мутантного по devR штамма M.tuberculosis в условиях ограничения количества кислорода в аэробных условиях при экспрессии генов devR и devS,
XIV. проведение анализа RT-PCR в отношении указанного выращенного штамма на предмет транскриптов, полученных из оперона Rv3134c-devR-devS,
XV. сканирование указанных транскриптов путем использования системы документации гелей Ultra-Violet Product и проведение денситометрического анализа по отношению к данным транскриптам путем использования компьютерного программного обеспечения,
XVI. тестирование вирулентности мутантного по devR штамма M.tuberculosis на морских свинках.
Способ идентификации новой мишени для применения при разработке терапевтических форм и лекарственных средств, эффективных против туберкулеза, описан здесь подробно с помощью сопровождающих чертежей, где:
на фиг. 1 показано конструирование мутантного штамма M.tuberculosis devR;
a) анализ путем гибридизации по Саузерну рекомбинантных штаммов M.tuberculosis,
b) PCR-анализ репрезентативного мутантного по devR клона.
На фиг. 2 показаны:
a) вестерн-блот-анализ мутантной по devR M.tuberculosis,
b) иммунная электронная микроскопия мутантной по devR M.tuberculosis,
с) морфологический анализ in vitro мутантного по devR штамма и штамма H37Rv M.tuberculosis.
На фиг. 3 показан экспрессионный анализ оперона Rv3134c-devR-devS на штамме дикого типа и мутантном штамме M.tuberculosis.
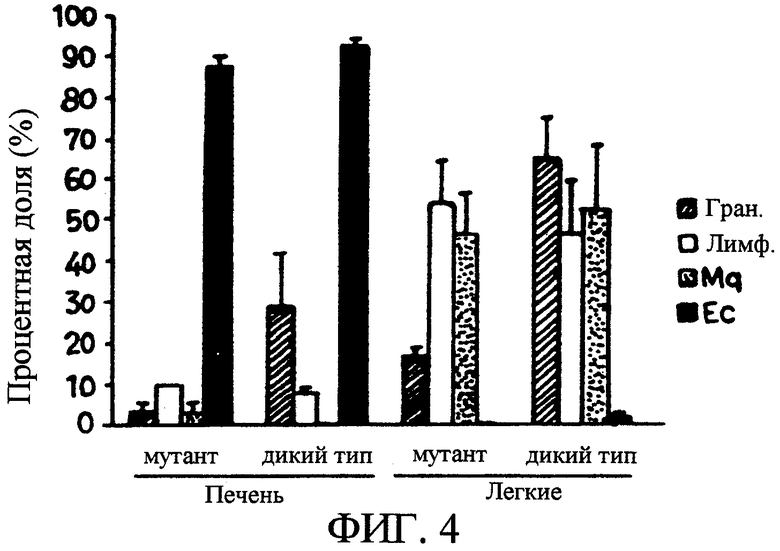
На фиг. 4 показаны характеристики гранулемы печени и легких у морских свинок, инфицированных мутантным по devR штаммом и штаммом H37RV M.tuberculosis.
Согласно способу данного изобретения ген devR M.tuberculosis разрушают посредством аллельного обмена с использованием стандартных технологий. В кратком изложении ген devR, расположенный во вставке EcoRI-HindIII массой ˜3,3 т.п.н. плазмиды pJT53.34, разрушают геном устойчивости к канамицину (KmR) в уникальном участке PpuMI. Разрушенный аллель devR вырезают в виде фрагмента ApaI-BamHI и клонируют в плазмиду pJQ200SK, конструируя pJQ200SkdevR::kan. Данную плазмиду вводят в M.tuberculosis H37Rv путем электропорации. Трансформанты с одним кроссинговером, которые показательны в смысле интеграции плазмиды, отбирают на чашках с агаром middle brook 7H10, содержащих 20 мкг/мл канамицина, и анализируют путем полимеразной цепной реакции (PCR) на предмет наличия последовательностей devR, KmR и гена сахарозы (sacB). Анализируют двадцать пять колоний KmR, и только 6 являются позитивными на предмет KmR по PCR, что указывает на высокую частоту спонтанного KmR (˜75%). Все 6 колоний были позитивны по PCR на предмет sacB, что указывает на наличие одного события кроссинговера при образовании колоний KmR. Три (из 6) при PCR являются позитивными на предмет devR дикого типа (513 н.п) и на предмет продуктов devR::kan (1,8 т.п.н.), указывая на то, что в хромосому M.tuberculosis произошла интеграция копии devR::kan плазмидного происхождения. PCR-анализы с использованием фланкирующих хромосому праймеров вместе с праймерами, специфичными для гена KmR, позволили предполагать, что интеграция плазмиды произошла вблизи локуса devR или devS в одном из клонов. После гибридизации по Саузерну с зондом devR получают два сигнала размером 3,8 т.п.н. и ˜9,5 т.п.н. в отличие от единственного сигнала гибридизации размером 3,8 т.п.н., полученного для исходного штамма. Идентичные результаты получают с использованием в качестве зонда гена devS (фиг. 1А). При гибридизации с зондом на основе гена KmR высвечивался фрагмент размером ˜9,5 т.п.н., содержащий остов вектора, полученный в результате события кроссинговерной рекомбинации слева от гена KmR в локусе devR или devS. Меродиплоидный штамм, содержащий копию дикого типа и разрушенную копию локуса devR, обозначают M.tuberculosis Dup devR (фиг. 1А).
Событие аллельного обмена подвергали селекции на второй стадии путем выращивания M.tuberculosis Dup devR в среде middle brook 7H9, содержащей канамицин (20 мкг/мл) и 2% сахарозы, в течение 7 суток. Серийные разведения высевали на среду 7H10, содержащую канамицин (20 мкг/мл) и 2% сахарозы. Всего получали 87 трансформантов, характеризующихся KmR и устойчивых к сахарозе (SucR); 31 из этих трансформантов являются при PCR негативными в плане гена devR дикого типа, из них 18 являются позитивными в плане разрушенного гена devR::kan. Устойчивость к сахарозе появлялась в результате одного из двух событий; ген sacB или терялся в результате разделения на части (64%), или накапливал мутацию(-и), приводящую(-ие) к потере функции (36%). Аллельный обмен подтверждают Саузерн-гибридизацией. В кратком изложении 16 (из 18) KmR-SucR-трансформантов обрабатывают зондами на основе devR и devS. Событие двойного кроссинговера происходило в 15 клонах, что приводило к присутствию копии devR::kan (сигнал гибридизации, равный 5,1 т.п.н.) вместо гена devR дикого типа (сигнал, равный 3,8 т.п.н.), причем мутанты характеризовались приращением размера на 1,3 т.п.н., что соответствовало кассете KmR, вставленной в локус devR. Гибридизация с геном KmR подтверждала сохранение в хромосоме, разрушенной кассетой KmR, копии гена devR. Отсутствие гибридизации с зондом sacB подтверждало то, что данный ген был потерян в результате разделения на части за счет тандемной дупликации (не показано). Репрезентативный клон показан на фиг. 1А. Один нокаут-клон devR случайным образом выбран для дальнейшей характеристики. Перестановку генов также подтверждали путем PCR (фиг. 1В).
Функциональное разрушение гена devR в мутанте подтверждают путем вестерн-блоттинга и иммунной электронной микроскопии с использованием стандартных процедур. Обработанные ультразвуком культуры M.tuberculosis H37Rv, мутанта по devR и E. coli, полученные в логарифмической фазе, подвергали денатурирующему электрофорезу в полиакриламидном геле, белки переносили на нитроцеллюлозную мембрану и обрабатывали поликлональным антителом против DevR в разведении 1:1000, которое было получено от кроликов против белка DevR M.tuberculosis. Иммунореактивность оценивали с использованием конъюгированного с пероксидазой хрена иммуноглобулина G осла против кроличьих антител и субстрата 3,3'-диаминобензидина. Белок DevR визуализируют в M.tuberculosis H37Rv и в рекомбинантной E.coli, избыточно экспрессирующей белок DevR, но не в мутатном штамме. Мечение DevR происходило на поверхности и в цитозоле организмов дикого типа (25±2,4 гранул золота на бациллу), но не в мутантном штамме (2±0,88 гранул золота на бациллу, фиг. 2В).
Морфологию мутантных и исходных бацилл сравнивают путем сканирующей электронной микроскопии. Мутантные бациллы логарифмической фазы были длиннее по размеру (в среднем 4-6 мкм) по сравнению с бациллой дикого типа (в среднем 2-4 мкм, фиг. 2С). Сравнивают кривые роста мутантного по devR и исходного штаммов, культивированных в трех параллелях в аэробных условиях в 7H9, содержащей 10% альбумин-декстрозы и 0,05% Tween 80. Среднее A590 возрастало от 0,054 на 1 сутки до 1,41 на 28 сутки для культур дикого типа и от 0,048 на 1 сутки до 1,9 на 28 сутки для мутантных культур. Различия в значении A590 в разные моменты времени для мутантного и исходного штаммов не является статистически значимым (по тесту ранговых сумм Вилкоксона), и это указывает на то, что мутация devR не имеет значимого влияния на рост M.tuberculosis в аэробных условиях.
Оценивали эффект ограничения кислорода на экспрессию генов M.tuberculosis devR и devS, которые, как было показано ранее, совместно транскрибируются в M.tuberculosis. Использовали модель покоя M.tuberculosis in vitro, в которой культуры выращивались на Dubos Tween-альбумин без перемешивания и медленно оседали на дно культурального сосуда, при которой рост ограничивался низкой концентрацией кислорода. Через ˜30 суток бактерии вступали в стационарную (нереплицирующуюся) фазу роста. В течение этого периода бактерии дикого типа нарастали в ˜18 раз. Организмы дикого типа адаптировались к постепенно нарастающему кислородному голоданию без детектируемой утраты жизнеспособности. Мутантные бактерии нарастали только в ˜5 раз за период продолжительностью в 30 суток. В конце эксперимента жизнеспособность мутантного штамма составляла ˜25% от таковой для штамма дикого типа. Аэробные шейкерные культуры исходного и мутантного штаммов M.tuberculosis выращивали одновременно до логарифмической фазы (A590˜0,4). РНК выделяют с использованием набора Rneasy Mini (Qiagen, Германия) из 30-суточной и 40-суточной культур логарифмической фазы, полученных без перемешивания, и проводят RT-PCR-анализ для транскриптов, происходящих из оперона Rv3134c-devR-devS (Rv3134c является первым геном в данном опероне). Гели сканируют с использованием системы документации гелей UVP и проводят денситометрический анализ с использованием аналитического программного обеспечения LabworksTM (Ultra-Violet product, США). В культурах дикого типа экспрессия генов devR, devS и Rv3134c в условиях гипоксии положительно регулируется примерно в 3-4 раза. Положительная регуляция также наблюдается в мутантном штамме за исключением того, что базовый уровень экспрессии гена devS и Rv3134c примерно в 2,5 раза ниже, чем наблюдаемая в штамме дикого типа. Как и ожидалось, в мутантном штамме не было обнаружено транскриптов гена devR (фиг. 3). Экспрессия и положительная регуляция гена devS в мутантном штамме, как полагают, имеют место вследствие инициации транскрипции, происходящей выше по цепи, поскольку экспрессия кассеты KmR (внутри гена devR) также подлежит положительной регуляции в сходных условиях (данные не показаны).
Способ идентификации новой мишени для применения при разработке терапевтических форм и противотуберкулезных лекарственных средств подробно описан здесь.
Действие мутации devR на рост in vivo и способность вызывать заболевание у морских свинок оценивают, как описано. Нелинейных морских свинок-альбиносов (пять животных на группу) подвергали подкожной инъекции 0,1 мл раствора жизнеспособных бацилл в фосфатно-солевом буфере (M.tuberculosis H37RvЧ106 CFU и devR-мутант 3,2Ч107 CFU). Морских свинок забивали через 47 суток после инфекции. Одно из животных (группа H37Rv), которое умерло не от туберкулеза до даты забоя, отбрасывали из анализа. Количество видимых случаев туберкулеза внутренних органов подсчитывали немедленно после забоя, как описано. У морских свинок, инфицированных M.tuberculosis H37Rv, отмечена тяжелая форма повреждения легких, печени, селезенки и лимфатических узлов. Визуальные коэффициенты для морских свинок, инфицированных исходным и мутантным штаммами, варьировали от 43 до 93 (в среднем 77) и от 23 до 48 (в среднем 38,4), соответственно, причем различие является значимым. Печень является наиболее затронутым органом, и у морских свинок, инфицированных исходным штаммом, наблюдается тяжелое вмешательство с наличием множественных крупных туберкул и областей некроза. Как показано, селезенка и легкие затронуты умеренно с наличием множественных мелких туберкул. Заметно меньшее количество видимых повреждений наблюдается в органах морской свинки, инфицированной мутантным штаммом (таблица 1). Селезенки гомогенизируют и серийные разведения высевают на косые агары LJ. Всего из селезенок животных, инфицированных исходным штаммом, выделяют 7,09±0,83 log10 cfu против 4,4±1,21 log10 cfu, полученных из животных, инфицированных мутантным штаммом, причем различие является значимым.
Серийные секции размером 5 мкм из образцов аутопсии печени и легких подвергали полуколичественной экспертизе гистологических признаков (архитектура органа, процент области секции, занятой гранулемой, и процент главных клеточных компонентов внутри гранулемы), как описано выше. Секции печени трех из пяти морских свинок, инфицированных мутантным штаммом, характеризовались нормальной архитектурой и не содержали гранулемы или воспалительных клеточных инфильтратов. У оставшихся двух животных, инфицированных мутантным штаммом, наблюдалась минимальная, хорошо организованная, не некротическая эпителиоидная клеточная гранулема.
В секциях печени от всех четырех морских свинок, инфицированных исходным штаммом, показано наличие хорошо оформленных гранулем, которые состояли из эпителиоидных клеток и лимфоцитов. Клетки других типов отсутствовали. У одного из животных гранулема была выраженной (65%) и сопровождалась частичным разрушением архитектуры органа (фиг. 4). По сравнению с печенью отмечалось гораздо более интенсивное вовлечение легких. Архитектура легких всех морских свинок, инфицированных мутантным штаммов, является нормальной. Хотя было показано присутствие гранулемы у всех животных, она была минимальной и состояла как из лимфоцитов, так и из макрофагов. Гигантские клетки и другие клетки некроза не были обнаружены ни в одном из легких. Для животных, инфицированных исходным штаммом, легкое одной из морских свинок является полностью разрушенным, и оно частично повреждено у оставшихся трех. Гранулема изменялась от 40% до 85% и варьировалась от преимущественно лимфоцитарной до гистиоцитарной (фиг. 4).
Через 7 недель после инфекции у морских свинок, инфицированных мутантным штаммом, по сравнению с инфицированными исходным штаммом, наблюдалась менее выраженная патология органов, и из них была получена примерно в тысячу раз меньшая бактериальная загрузка. Преобладание эпителиоидных клеток над макрофагами и лимфоцитами в печени по сравнению с легким указывает на хорошо выраженный иммунный ответ и более развернутое разрешение гранулемы в последнем.
Таким образом, очевидно, что двухкомпонентная система DevR-DevS участвует в вирулентности M.tuberculosis и действительно может быть ключевым регуляторным связующим звеном между ограничением кислорода и инициацией и поддержанием адаптивного ответа на гипоксию. Адаптация микробактерий к анаэробному микроокружению, как полагают, обеспечивает бациллам туберкулеза способ неограниченно долго оставаться в персистирующем состоянии покоя/стационарной фазы внутри воспалительных и некротических участков повреждения, таких как гранулема. Поэтому данная генетическая система может служить важной мишенью для разработки новых и неизвестных лекарственных средств для лечения туберкулеза, особенно состояния персистирования.
Изобретение относится к биотехнологии и может быть использовано в медицине и фармацевтической промышленности. Способ предусматривает следующие стадии: разрушение гена devR плазмиды pJT53.34; конструирование из разрушенного гена плазмиды pJQ2000KdevR::kan и введение ее в M.tuberculosis H37Rv с последующей селекцией трансформантов на среде Миддлбрука 7Н10 с канамицином. Далее трансформанты проверяют на наличие последовательностей генов devR, KmR и гена устойчивости к сахарозе (SacB), затем выявляют M.tuberculosis Dup devR, содержащую копию дикого типа и разрушенную копию локуса devR. После проведенных анализов эти микобактерии выращивают на той же среде в присутствии канамицина и сахарозы с последующим размножением микобактерий в тех же условиях выращивания. У полученного мутантного в отношении devR штамма M.tuberculosis осуществляют проверку на подтверждение аллельного обмена, разрушения гена devR::kan, в т.ч. и его функционального разрушения, а также оценивают его ростовые свойства. Далее проводят анализ на наличие транскриптов, полученных из оперона Rv 3134с-devR-devS, их сканирование и тестирование вирулентности полученного мутантного по devR штамма на морских свинках, включающий гистопатологический анализ инфицированных органов морских свинок, выделение идентифицированных мутантных микобактерий из селезенок инфицированных животных и количественный анализ бактериальной загрузки. Использование изобретения позволяет идентифицировать как мишень, ответственную за рецидив/реактивацию у пациента данного заболевания, так и мишень, которая позволяет организму адаптироваться к гипоксии, и соответственно разработать терапевтические формы и лекарственные средства для эффективного лечения туберкулеза. 5 з.п. ф-лы. 4 ил.
I. разрушение гена devR (Rv3133c), расположенного во вставке EcoRI-HindIII размером ˜3,3 т.п.н. плазмиды pJT53.34,
II. конструирование pJQ200SkdevR: : kan из разрушенного гена devR,
III. введение указанной плазмиды в M.tuberculosis H37Rv,
IV. селекцию трансформантов с одним кроссинговером, указывающим на интеграцию плазмиды, на чашках с агаром Миддлбрука 7Н10, содержащих канамицин,
V. их анализ путем полимеразной цепной реакции (PCR) на наличие последовательностей генов devR, KmR и гена устойчивости к сахарозе (SacB),
VI. проведение Саузерн-анализа в отношении указанных последовательностей с зондом devR, зондом devS, зондом гена устойчивости к канамицину, так что выявляется M.tuberculosis Dup devR, содержащая копию дикого типа и разрушенную копию локуса devR,
VII. выращивание M.tuberculosis Dup devR в среде Миддлбрука 7Н9, содержащей канамицин и сахарозу,
VIII. помещение указанного выращенного штамма M.tuberculosis Dup devR во множество чашек со средой 7Н10, содержащей канамицин и сахарозу, с получением трансформантов, устойчивых к канамицину, получая таким образом мутантный в отношении devR штамм M.tuberculosis,
IX. проведение Саузерн-гибридизации в отношении указанной выращенной M.tuberculosis devR с последующей процедурой полимеразной цепной реакции для подтверждения указанного аллельного обмена,
X. проведение анализа путем полимеразной цепной реакции в отношении указанных трансформантов на предмет разрушенного гена devR: : kan,
XI. проведение вестерн-блоттинга и иммунной электронной микроскопии в отношении указанного разрушенного гена devR: : kan для подтверждения функционального разрушения указанного гена,
XII. оценку способности к росту мутантного по devR штамма M.tuberculosis в условиях ограничения количества кислорода при экспрессии генов devR и devS,
XIII. оценку способности к росту указанного мутантного по devR штамма M.tuberculosis в условиях ограничения количества кислорода в аэробных условиях при экспрессии генов devR и devS,
XIV. проведение анализа RT-PCR в отношении указанного выращенного штамма на предмет транскриптов, полученных из оперона Rv3134c-devR-devS,
XV. сканирование указанных транскриптов путем использования системы документации гелей Ultra-Violet Product и проведение денситометрического анализа по отношению к данным транскриптам стадии путем использования компьютерного программного обеспечения,
XVI. тестирование вирулентности мутантного по devR штамма M.tuberculosis на морских свинках, включающее (а) гистопатологический анализ инфицированных органов (легких и печени) морских свинок, инфицированных мутантным по devR штаммом и штаммом дикого типа M.tuberculosis, и (b) выделение M.tuberculosis из селезенок инфицированных животных и количественный анализ бактериальной загрузки.
| CN 1309703 А, 22.08.2001 | |||
| DASGUPTA, N | |||
| et al | |||
| Characterization of two-component system, devR-desS, of Mycobacterium tuberculosis | |||
| Tubercle and Lung Disease, 2000, vol.80, №3, р.141-159 | |||
| 0 |
|
SU184437A1 | |

