Перекрестная ссылка на родственные заявки
Настоящая международная патентная заявка согласно 35 U.S.C. §119 (e) заявляет приоритет предварительной заявки США, серийный № 61/203605, зарегистрированной 24 декабря 2008 года, абандондонированной в настоящее время, и предварительной заявки, серийный номер 61/188112, зарегистрированной 6 августа 2008 года, абандондонированной в настоящее время, и содержание обеих из них включены полностью в настоящее изобретение.
УРОВЕНЬ ИЗОБРЕТЕНИЯ
Область изобретения
Настоящее изобретение относится в общем к областям медицины, микробиологии патогенов и технологиям визуализации. Более конкретно, настоящее изобретение относится к соединениям и репортерам, полезным для обнаружения и локализации бактериальных патогенов в ходе визуализации in vivo у субъекта.
Описание родственного уровня техники
Многочисленные бактериальные инфекции вызывают значительную заболеваемость и смертность во всем мире, и многие из наиболее важных видов бактерий являются положительными по бета-лактамазе, что делает их устойчивыми к стандартным подобным пенициллину антибиотикам. Диагностика многих из этих инфекций и определение резистентности к пенициллину часто затруднительны, и для этого перед определением восприимчивости необходимо продолжительное диагностическое лабораторное культивирование.
Например, в настоящее время почти одна треть населения в мире поражена туберкулезом, который остается острой угрозой общественного здоровья. Эта озабоченность значительно усиливается с учетом длительного существования во всем мире множественной лекарственной резистентности и распространенности штаммов с чрезвычайной лекарственной резистентностью, которые трудно поддаются лечению. Современные способы количественного определения и оценки жизнеспособности туберкулеза в лабораторных условиях, клетках тканевой культуры и при инфекции в моделях животных и человеческих моделях ограничены определением колониеобразующих единиц (КОЕ) и/или микроскопией тканей и слюны. Эти способы занимают много времени, часто трудны для интерпретации и относительно нечувствительны. Для большинства способов необходимы инвазивные процедуры, которые у животных и людей должны осуществляться посмертно. Эти несоответствия мешают отслеживать развитие болезни, эффективность вакцины и терапевтический результат как в моделях животных, так и у больных людей. Способы оптической визуализации позволят непосредственно в режиме реального времени неинвазивным образом с использованием живых животных определять жизнеспособность туберкулеза при инфекции, эффективность лечения и локализацию бактерий во время болезни.
Таким образом, в данной области техники существует признанная потребность в улучшенных способах визуализации бактериальной болезни. Более конкретно, предшествующий уровень техники имеет недостаток в чувствительных и специфичных способах оптической визуализации в режиме реального времени бета-лактамазоположительных бактерий, в способах, которые можно применять in vitro и у живых субъектов для диагностики и локализации бактериальной инфекции, для быстрого скрининга новых терапевтических средств и идентификации новых мишеней для лекарственного средства. Настоящее изобретение соответствует этой давно существующей потребности и задачам в данной области техники.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к способу обнаружения патогенных бактерий в режиме реального времени у субъекта. Способ включает введение субъекту или во взятый у субъекта биологический образец флуоресцентного, люминесцентного или колориметрического субстрата для бета-лактамазы патогенных бактерий и визуализацию субъекта или образца на наличие продукта бета-лактамазной активности на субстрате. Получают сигналы на длине волны, испускаемой бета-лактамазным продуктом, и таким образом выявляют патогенные бактерии у субъекта. Настоящее изобретение относится к родственному способу, дополнительно включающему получение трехмерной реконструкции испускаемого сигнала для определения локализации патогенных бактерий у субъекта. Настоящее изобретение относится к другому родственному способу, дополнительно включающему диагностику в режиме реального времени патофизиологического состояния, связанного с патогенными бактериями, на основе интенсивности испускаемого сигнала, превышающей измеренный контрольный сигнал.
Настоящее изобретение также относится к способу диагностики патофизиологического состояния у субъекта, связанного с патогенными бактериями. Способ включает введение субъекту флуорогенного субстрата для бета-лактамазы патогенных бактерий или приведение биологического образца, полученного от субъекта, в контакт с указанным субстратом, и визуализацию субъекта на наличие продукта бета-лактамазной активности на субстрате. Интенсивность флуоресцентного, люминесцентного или колориметрического сигнала измеряют в режиме реального времени на длине волны, испускаемой продуктом, таким образом, что интенсивность флуоресцентного, люминесцентного или колориметрического сигнала превышает интенсивность измеренного контрольного сигнала, и это коррелирует с диагнозом патофизиологического состояния. Настоящее изобретение относится к родственному способу, дополнительно включающему получение трехмерной реконструкции сигнала для определения локализации микробного патогена. Настоящее изобретение относится к другому родственному способу, дополнительно включающему введение одного или более терапевтических соединений, эффективных для лечения патофизиологического состояния. Настоящее изобретение относится к дополнительному родственному способу, включающему повторное введение субъекту флуорогенного субстрата и повторную визуализацию субъекта или приведение биологического образца, взятого у субъекта, в контакт с указанным флуорогенным субстратом; и визуализацию субъекта или указанного биологического образца для мониторинга эффективности терапевтического соединения, таким образом, что понижение испускаемого сигнала по сравнению с сигналом при диагнозе указывает на терапевтическое воздействие на патофизиологическое состояние.
Настоящее изобретение дополнительно относится к способу скрининга терапевтических соединений, эффективных для лечения у субъекта патофизиологического состояния, связанного с наличием патогенных бактерий у субъекта. Способ включает выбор потенциального терапевтического соединения для патогенных бактерий, приведение бактериальных клеток в контакт с флуоресцентным, люминесцентным или колориметрическим агентом обнаружения и приведение бактериальных клеток в контакт с потенциальным терапевтическим соединением. Флуоресцентный, люминесцентный или колориметрический сигнал, продуцируемый бактериальными клетками, измеряют в присутствии и отсутствии потенциального терапевтического соединения, и, таким образом, понижение сигнала в присутствии терапевтического соединения по сравнению с сигналом в его отсутствие указывает на терапевтическое воздействие этого соединения на патофизиологическое состояние.
Настоящее изобретение дополнительно относится к способу визуализации патогенных бактерий. Способ включает контакт патогенных бактерий с флуорогенным субстратом для их бета-лактамазного фермента, воздействие на патогенные бактерии длиной волны возбуждения на продукт бета-лактамазной активности на субстрате и получение флуоресцентных, люминесцентных или колориметрических сигналов, испускаемых продуктом. Таким образом, для визуализации патогенных бактерий создается трехмерная реконструкция полученных сигналов.
Настоящее изобретение дополнительно относится к флуорогенному субстрату CNIR-7 или CNIR7-ТАТ для бактериальной бета-лактамазы.
Настоящее изобретение дополнительно относится к способу обнаружения патогенных бактерий в режиме реального времени у субъекта. Способ включает введение субъекту субстрата, радиоактивно меченного изотопом, связанным с гамма-излучением, при этом субстрат предназначен для бета-лактамазы или другого фермента или белка, специфичных для патогенных бактерий. Во время активности радиоактивно меченного субстрата у субъекта проводят обнаружение гамма-излучения из указанного субстрата и получают сигналы, генерируемые испускаемыми гамма-лучами. Таким образом, на основе интенсивности сигналов, генерируемых гамма-лучами, обнаруживают патогенные бактерии с помощью созданной трехмерной реконструкции концентрации радиоактивной метки у субъекта. Настоящее изобретение относится к родственному способу, дополнительно включающему диагностику в режиме реального времени патофизиологического состояния, связанного с патогенными бактериями, который основан на их обнаружении. Настоящее изобретение относится к другому родственному способу, дополнительно включающему введение одного или более терапевтических соединений, которые эффективны для лечения этого патофизиологического состояния. Настоящее изобретение относится еще к одному родственному способу, дополнительно включающему повторное введение радиоактивно меченного субстрата субъекту и повторную визуализацию субъекта для мониторинга эффективности терапевтического соединения; при этом понижение гамма-излучения по сравнению с гамма-излучением при диагнозе указывает на терапевтическое воздействие на патофизиологическое состояние.
Настоящее изобретение дополнительно относится к радиоактивно меченному субстрату для бактериальной бета-лактамазы, подходящему для визуализации с помощью позитронно-эмиссионной томографии (ПЭТ) или однофотонной эмиссионной компьютерной томографии (ОФЭКТ), как описано в настоящем изобретении.
Другие и дополнительные объекты, признаки и преимущества станут очевидными из последующего описания представленных предпочтительных вариантов осуществления настоящего изобретения, которые приведены в целях раскрытия.
КРАТКОЕ ОПИСАНИЕ ФИГУР
Для достижения и возможности детального понимания аспектов вышеупомянутых признаков, преимуществ и целей изобретения, а также других раскрываемых аспектов, более конкретные описания изобретения, коротко сформулированные выше, могут быть приведены со ссылкой на определенные варианты их осуществления, которые показаны в прилагаемых фигурах. Указанные фигуры представляют собой часть описания. Вместе с тем, нужно отметить, что прилагаемые фигуры показывают предпочтительные варианты осуществления изобретения и поэтому не должны рассматриваться в качестве ограничения объема вариантов изобретения.
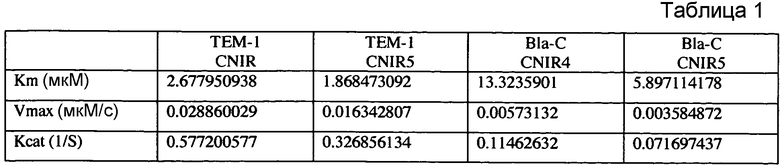
Фиг. 1A-1C показывают структуры CC1 и CC2 (фиг. 1A), CHPQ (фиг. 1B) и CR2 (фиг. 1C) перед гидролизом и после гидролиза с помощью бета-лактамазы.
Фиг. 2A-2B показывают структуры CNIR1, CNIR2, CNIR3 и CNIR4 и их гидролиз бета-лактамазой (фиг. 2A) и структуры CNIR9 и CNIR10 (фиг. 2B).
Фиг. 3A-3C показывают схему синтеза для приготовления субстстрата CNIR5, близкого к инфракрасному (фиг. 3A), флуоресцентную интенсивность против длины волны CNIR5 в присутствии и в отсутствие бета-лактамазы (фиг. 3B) и структуру CNIR5-QSY22 (фиг. 3C).
Фиг. 4A-4D показывают структуры QSY21 (фиг. 4A), дисульфоната QSY21 (фиг. 4B) и дисульфоната QSY22 (фиг. 4C), и химический синтез дисульфоната QSY22 (фиг. 4D).
Фиг. 5A-5B показывают структуру CNIR7 (фиг. 5A) и ее химический синтез (фиг. 5B).
Фиг. 6A-6B показывают химический синтез Bluco (фиг. 6A) и использование Bluco для последовательного биолюминесцентного репортерного анализа (SREL) для визуализации бета-лактамазы (фиг. 6B).
Фиг. 7 показывает обнаружение активности Bla в E. coli с CNIR5. Контроль содержит среду LB и CNIR5 без трансформированной E. coli.
Фиг. 8A-8D показывают спектры эмиссии для CNIR4 (фиг. 8A), CNIR5 (фиг. 8B), CNIR9 (фиг. 8C) и CNIR10 (фиг. 8D) перед и после расщепления Bla в течение 10 мин.
Фиг. 9A-9B показывают кинетические данные бета-лактамазы TEM-1 E. coli и бета-лактамазы Bla-C Mycobacterium tuberculosis с субстратами CNIR4 (фиг. 9A) и CNIR5 (фиг. 9B).
Фиг. 10A-10Н показывают кинетические данные флуоресцентных включений и коэффициенты распределения в них (фиг. 10E-10Н) бактерий Mycobacterium tuberculosis, единственных, в среде с CNIR4 (фиг. 10A, 10E), CNIR5 (фиг. 10B, 10F), с CNIR9 (фиг. 10C, 10G) и с CNIR10 (фиг. 10D, 10Н).
Фиг. 11A-11H показывают кинетические данные флуоресцентных включений (фиг. 11A-11D) и коэффициенты распределения в них (фиг. 11E-11H) макрофагов, инфицированных бактериями Mycobacterium tuberculosis, с CNIR4 (фиг. 11А, 11Е), CNIR5 (фиг. 11В, 11F), CNIR9 (фиг. 11C, 11G) и CNIR10 (фиг. 11D, 11Н).
Фиг. 12 демонстрирует изображения флуоресцентной конфокальной микроскопии, показывая внутриклеточное включение CNIR4 в макрофаги, инфицированные Mycobacterium tuberculosis. Окрашивание DAPI (синим) выявляет ядра инфицированных клеток, зеленая флуоресценция обусловлена меченной GFP M. tuberculosis, и красная флуоресценция обусловлена расщеплением CNIR4.
Фиг. 13A-13E показывают флуоресценцию у мышей, инфицированных Mycobacterium tuberculosis путем подкожной инокуляции CNIR4 (фиг. 13A), CNIR5 (фиг. 13B), CNIR9 (фиг. 13C) и CNIR10 (фиг. 13D) при разных концентрациях от 108 (нижняя область слева у каждой мыши), 107 (верхняя область слева), 106 (верхняя область справа). Фиг. 13E показывает сравнение сигнала с фоновым сигналом для каждого соединения при каждой концентрации бактерий, используемой для инфицирования.
Фиг. 14A-14E представляют собой изображения флуоресценции у мышей, которые были инфицированы Mycobacterium tuberculosis в легкие путем аэрозольной инокуляции, и показывают сигнал флуоресценции, измеренный для CNIR4 (фиг. 14A), CNIR5 (фиг. 14B), CNIR9 (фиг. 14C) и CNIR10 (фиг. 14D). В каждой из фигур 14A-14D мышь слева в каждой колонке не инфицирована, вторая мышь слева инфицирована M. tuberculosis, которая несет мутацию в гене blaC, и две мыши с правой стороны в каждой группе инфицированы M. tuberculosis дикого типа. Трем правым мышам в каждой группе за 24 часа перед визуализацией внутривенно (в/в) вводили CNIR4, CNIR5, CNIR9 или CNIR10. Фиг.14E представляет собой график сигнала в сравнении с фоновым сигналом для каждого соединения в легочной области при получении дорсального изображения.
Фиг. 15A-15F показывают флуоресценцию у мышей, инфицированных аэрозольным путем M. Tuberculosis, которую визуализируют с использованием субстрата CNIR5 в точках времени 1 час (фиг. 15A), 18 часов (фиг. 15B), 24 часа (фиг. 15C) и 48 часов (фиг. 15D). В каждой группе дорсального, вентрального изображения или боковых правых и левых видов неинфицированная мышь показана слева и инфицированная мышь показана справа. Всем мышам перед визуализацией в/в вводили CNIR5 в вышеуказанные точки времени. Фиг. 15F представляет собой график, количественно определяющий флуоресцентный сигнал, полученный из рассматриваемой области, которая обведена кругом в верхней группе фиг. 15A.
Фиг. 16A-16B показывают визуализацию флуоресценции мышей, инфицированных аэрозольным путем M. Tuberculosis (фиг. 16A) или неинфицированных (фиг. 16B), и получение изображения с использованием трансиллюминации, в отличие от отражения, для уменьшения фонового сигнала.
Фиг. 17A-17D показывают визуализацию экспрессии Bla с CNIR5 (7 нмоль) у нагой мыши с ксенотрансплантатом опухоли C6 дикого типа в левом плече и опухоли C6 с устойчивой трансфекцией cmv-bla в правом плече. Фиг. 17A показывает наложенную флуоресценцию и изображения в светлом поле в указанные точки времени. Фиг. 17B показывает график - кривую средней интенсивности каждой опухоли в зависимости от времени. Фиг. 17C показывает изображения удаленных опухолей и органов. Фиг. 17D показывает результаты CC1 анализа на Bla в экстрактах обеих опухолей.
Фиг. 18A-18C показывает визуализацию экспрессии Bla с CNIR6 (7 нмоль) у нагой мыши с ксенотрансплантатом опухоли C6 дикого типа в левом плече и опухоли C6 с устойчивой трансфекцией cmv-bla в правом плече. Фиг. 18A отображает химическую структуру CNIR6. Фиг. 18B показывает наложенную флуоресценцию и изображения в светлом поле в указанные точки времени. Фиг. 18C показывает кривую средней интенсивности каждой опухоли в зависимости от времени.
Фиг. 19A-19B демонстрируют биораспределение CNIR5 в количестве 7,5 нмоль в разных тканях через 4 часа (фиг. 19A) и через 24 часа (фиг. 19B).
Фиг. 20A-20B представляют собой изображения мыши in vivo, инфицированной M. Tuberculosis (фиг. 20A) и неинфицированной контрольной мыши (фиг. 20B), с использованием в качестве агента визуализации CNIR5 внутривенно.
Фиг. 21A-21C показывают порог обнаружения для SREL с использованием зонда CNIR. Фиг. 21A представляет собой гистограмму, показывающую возможность обнаружения менее 100 бактерий с использованием бета-лактамазного зонда CNIR с визуализацией SREL. Фиг. 21B-21C являются изображениями in vivo живых мышей, неинфицированных (фиг. 21B) или инфицированных M. Tuberculosis (фиг. 21C).
Фиг. 22 изображает структуры флуорофоров серии IRDye800.
Фиг. 23 изображает структуры цефоперазона и предлагаемого зонда CNIR.
Фиг. 24 представляют собой схему конструирования маленькой направленной библиотеки из субстратов Bluco.
Фиг. 25 показывает структуры новых зондов, содержащих аллильную связь в 3'-положении.
Фиг. 26 показывает структуры новых зондов, содержащих карбаматную связь в 3'-положении.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Используемые в настоящем описании формы единственного числа могут означать один или более. Используемые в формуле (формулах) изобретения слова в единственном числе в сочетании со словом "содержит" могут означать один или более чем один. Используемые слова "другой" или "другие" могут подразумевать по меньшей мере второй или дальнейший в отношении одного и того же элемента, или отличного элемента, или его компонентов, из формулы изобретения. Кроме того, если иначе не требуется по контексту, термины в единственном числе должны включать множественное число, и термины во множественном числе должны включать единственное число.
Используемый в формуле изобретения термин "или" означает "и/или", если явно не обозначено, что он относится только к альтернативам, или альтернативы являются взаимоисключающими, при этом в раскрытии употребляется определение, которое относится только к альтернативам и "и/или".
Используемый в настоящем изобретении термин "контактирует" относится к любому подходящему способу достижения контакта флуорогенного соединения, флуоресцентного, люминесцентного или колориметрического белка или радиоактивно меченого субстрата, подходящих для визуализации посредством ПЭТ или ОФЭКТ, с патогенными бактериями, например, без ограничения, Mycobacterium tuberculosis (Mbt) и Mycobacterium bovis (M. bovis). В условиях in vitro или ex vivo этот контакт достигается путем воздействия флуорогенного соединения или флуоресцентного, люминесцентного или колориметрического белка на одну или более бактериальных клеток в подходящей среде. Бактериальные клетки находятся в образцах, полученных из субъекта, и указанные образцы могут включать без ограничения плевральную жидкость и другие субстанции организма, включающие кровь, слюну, мочу и кал, в которых могут находиться бактерии. Для применений in vivo подходят любые известные способы введения флуорогенного соединения, флуоресцентного, люминесцентного или колориметрического белка, или радиоактивно меченного субстрата, согласно настоящему описанию.
Используемое в настоящем изобретении понятие "флуорогенный субстрат" относится к соединению или белку, или пептиду, или к другой биологически активной молекуле, которая в присутствии подходящего фермента приводит к получению продукта, который испускает или генерирует флуоресцентный, люминесцентный или колориметрический сигнал после возбуждения волной соответствующей длины. Например, без какого-либо ограничения, флуорогенный субстрат может продуцировать флуоресцентный, люминесцентный или колориметрический продукт в присутствии бета-лактамазы или люциферазы.
Используемое в настоящем изобретении понятие "радиоактивно меченный субстрат" относится к соединению или белку, или пептиду, или к другой биологически активной молекуле, присоединенной к короткоживущему радиоизотопу или связанной с ним, или иным образом включенной в короткоживущий радиоизотоп, который испускает позитроны для позитронно-эмиссионной томографии (ПЭТ) или гамма-лучи для однофотонной эмиссионной компьютерной томографии (ОФЭКТ).
Используемое в изобретении понятие "бета-лактамазоположительные бактерии" относится к патогенным бактериям, которые в природе секретируют фермент бета-лактамазу или приобретают бету-лактамазу в процессе патогенеза.
Используемый в настоящем изобретении термин "субъект" относится к любой цели лечения. Предпочтительно субъектом является млекопитающее, более предпочтительно субъектом является или представитель рогатого скота, или человек.
В одном варианте осуществления настоящего изобретения представлен способ обнаружения патогенных бактерий у субъекта в режиме реального времени, и указанный способ включает введение субъекту или в биологический образец, взятый у субъекта, флуоресцентного, люминесцентного или колориметрического субстрата для бета-лактамазы патогенных бактерий; визуализацию субъекта или образца на длине волны возбуждения на наличие продукта бета-лактамазной активности на субстрате; и получение сигналов на длине волны, испускаемых бета-лактамазным продуктом; и, таким образом, обнаружение патогенных бактерий у субъекта.
В дополнение к этому варианту осуществления способ включает получение трехмерной реконструкции испускаемого сигнала для определения локализации патогенных бактерий у субъекта. В другом дополнительном варианте осуществления способ включает диагностику в режиме реального времени патофизиологического состояния, связанного с патогенными бактериями, которая основана на интенсивности испускаемого сигнала, превышающей измеренный контрольный сигнал. Примером патофизиологического состояния является туберкулез.
В некоторых вариантах осуществления настоящего изобретения флуоресцентный субстрат может представлять собой флуорогенный субстрат. Примерами флуорогенных субстратов являются CNIR2, CNIR3, CNIR4, CNIR5, CNIR5-QSY22, CNIR7, CNIR9, CNIR10, CNIR7-ТАТ, каркасный люциферин, колориметрический реагент или их производные. Дополнительно в некоторых вариантах осуществления получаемая при визуализации длина волны составляет от приблизительно 540 нм до приблизительно 730 нм. Также диапазон испускаемых сигналов может быть от приблизительно 300 нм до приблизительно 900 нм. Во всех вариантах осуществления длина обнаруживаемой волны составляет от приблизительно 300 нм до приблизительно 900 нм. В некоторых вариантах осуществления колориметрические показатели можно визуально идентифицировать человеческим глазом по изменению цвета или измерить с помощью устройства для определения заданной числовой величины. Кроме того, патогенные бактерии могут включать виды бактерий Bacteroides, Clostridium, Streptococcus, Staphylococcus, Pseudomonas, Haemophilus, Legionella, Mycobacterium, Escherichia, Salmonella, Shigella или Listeria.
В другом варианте осуществления настоящего изобретения представлен способ диагностики у субъекта патофизиологического состояния, связанного с патогенными бактериями, который включает введение субъекту флуорогенного субстрата для бета-лактамазы патогенных бактерий; визуализацию субъекта с длиной волны возбуждения на наличие продукта бета-лактамазной активности на субстрате; и измерение в режиме реального времени интенсивности флуоресцентного, люминесцентного или колориметрического сигнала по длине волны, испускаемой продуктом; при этом интенсивность флуоресцентного, люминесцентного или колориметрического сигнала, превышающая интенсивность измеренного контрольного сигнала, коррелирует с диагнозом патофизиологического состояния.
В дополнение к этому варианту осуществления способ включает создание трехмерной реконструкции сигнала для определения локализации микробного патогена. В другом дополнительном варианте осуществления способ включает введение одного или более терапевтических соединений, эффективных для лечения патофизиологического состояния. Более того, способ включает повторное введение флуорогенного субстрата субъекту; и повторную визуализацию субъекта для мониторинга эффективности терапевтического соединения; при этом понижение испускаемого сигнала по сравнению с сигналом при диагнозе указывает на терапевтическое воздействие на патофизиологическое состояние. Во всех вариантах осуществления патофизиологическое состояние, патогенные бактерии, флуорогенный субстрат и показатели длины волны возбуждения и эмиссии соответствуют вышеприведенному описанию.
В другом варианте осуществления настоящего изобретения представлен способ диагностики у субъекта патофизиологического состояния, связанного с патогенными бактериями, который включает приведение образца, полученного из указанного субъекта, в контакт с колориметрическим субстратом для бета-лактамазы патогенных бактерий; при этом продукт бета-лактамазной активности на субстрате вызывает изменение цвета, видимого невооруженным глазом, таким образом, указывая на диагноз. Субстрат можно наносить на полоску, ватную палочку, основу или другие видимые индикаторы. Изменение цвета можно наблюдать невооруженным глазом и распознавать без какого-либо оборудования или возбуждения с помощью внешнего источника энергии.
Еще в одном варианте осуществления настоящего изобретения представлен способ скрининга терапевтических соединений, эффективных для лечения у субъекта патофизиологического состояния, связанного с наличием патогенных бактерий у субъекта, который включает выбор потенциального терапевтического соединения для патогенных бактерий; приведение бактериальных клеток в контакт с флуоресцентным, люминесцентным или колориметрическим агентом обнаружения; приведение бактериальных клеток в контакт с потенциальным терапевтическим соединением; и измерение флуоресцентного, люминесцентного или колориметрического сигнала, продуцируемого бактериальными клетками в присутствии потенциального терапевтического соединения и в его отсутствие; при этом понижение сигнала в присутствии терапевтического соединения по сравнению с сигналом в его отсутствие указывает на терапевтический эффект соединения против патогенных бактерий. В этом варианте осуществления патофизиологическое состояние и патогенные бактерии соответствуют описанию выше.
В одном аспекте этого варианта осуществления патогенные бактерии могут быть рекомбинантными бактериями, у которых стадия приведения бактерий в контакт с флуоресцентным, люминесцентным или колориметрическим агентом обнаружения включает трансформирование бактерий дикого типа вектором экспрессии, содержащим флуоресцентный, люминесцентный или колориметрический агент обнаружения. В этом аспекте флуоресцентный, люминесцентный или колориметрический агент обнаружения может содержать флуоресцентный белок. Примерами флуоресцентного белка являются mPlum, mKeima, Mcherry или tdTomato. Также в этом аспекте вектор экспрессии может содержать ген бета-галактозидазы, при этом способ дополнительно включает контакт рекомбинантных бактериальных клеток с флуорофором, который обладает эффектом эмиссии флуоресцентного сигнала в присутствии фермента бета-галактозидазы. Примерами флуорофоров являются C2FDG, C12RG или DDAOG. Дополнительно в этом аспекте вектор экспрессии может содержать ген люциферазы, при этом способ дополнительно включает контакт рекомбинантных бактериальных клеток с D-люциферином. Примерами люциферазы являются люцифераза светлячка, жука-щелкуна красного или rLuc δ.
В другом аспекте этого варианта осуществления флуоресцентный агент обнаружения может представлять собой флуорогенный субстрат бактериальной бета-лактамазы. В одном примере патогенные бактерии могут контактировать с флуорогенными субстратами CNIR2, CNIR3, CNIR4, CNIR5, CNIR5-QSY22, CNIR7, CNIR9, CNIR10, CNIR-ТАТ, каркасным люциферином, колориметрическим реагентом или их производными in vivo. В другом примере патогенные бактерии могут контактировать с флуорогенным субстратом CC1, CC2, CHPQ, CR2, CNIR1 или CNIR6 in vitro.
Еще в одном варианте осуществления настоящего изобретения представлен способ визуализации патогенных бактерий, включающий контакт патогенных бактерий с флуорогенным субстратом для их фермента бета-лактамазы; воздействие на патогенные бактерии длиной волны возбуждения на наличие продукта бета-лактамазной активности на субстрате; получение флуоресцентных, люминесцентных или колориметрических сигналов, испускаемых продуктом; и создание трехмерной реконструкции полученных сигналов, и, таким образом, визуализацию патогенных бактерий. В аспектах этого варианта осуществления патогенные бактерии могут контактировать in vivo или in vitro с флуорогенными или люминесцентными субстратами, как описано выше. Кроме того, во всех аспектах этого варианта осуществления патогенные бактерии и показатели длины волны возбуждения и эмиссии соответствуют описанию выше.
Еще в одном варианте осуществления настоящего изобретения представлен флуорогенный субстрат для бактериальной бета-лактамазы, который представляет собой CNIR-7 или CNIR7-ТАТ.
Еще в одном варианте осуществления настоящего изобретения представлен способ обнаружения у субъекта патогенных бактерий в режиме реального времени, который включает введение субъекту субстрата, радиоактивно меченного изотопом, связанным с гамма-излучением; при этом субстрат предназначен для бета-лактамазы или другого фермента или белка, специфичных для патогенных бактерий; обнаружение у субъекта гамма-излучения из радиоактивно меченного субстрата в процессе его активности; регистрацию сигналов, генерируемых испускаемыми гамма-лучами; и создание трехмерной реконструкции концентрации у субъекта с радиоактивной меткой на основе интенсивности сигналов, генерируемых гамма-излучением; и, таким образом, обнаружение патогенных бактерий.
В дополнение к этому варианту осуществления способ включает диагностику в режиме реального времени патофизиологического состояния, связанного с патогенными бактериями, которая основана на их обнаружении. В другом дополнительном варианте осуществления способ включает введение одного или более терапевтических соединений, эффективных для лечения патофизиологического состояния. Еще в одном дополнительном варианте осуществления способ включает повторное введение субъекту радиоактивно меченого субстрата; и повторную визуализацию субъекта для мониторинга эффективности терапевтического соединения; при этом понижение гамма-излучения по сравнению с гамма-излучением при диагнозе указывает на терапевтическое воздействие на патофизиологическое состояние. В этих дополнительных вариантах осуществления патофизиологическое состояние может представлять собой туберкулез.
В одном аспекте всех упомянутых вариантов осуществления радиоактивной меткой может быть позитронно-активный изотоп, и визуализацию можно осуществлять посредством позитронно-эмиссионной томографии (ПЭТ). В другом аспекте радиоактивная метка может представлять собой изотоп, непосредственно испускающий гамма-лучи, и визуализацию можно осуществлять посредством однофотонной эмиссионной компьютерной томографии (ОФЭКТ). Во всех аспектах упомянутых вариантов осуществления другой фермент или белок могут быть бета-лактамазоподобным ферментом или связывающим пенициллин белком. При этом во всех вариантах осуществления виды бактерий могут быть такими, как описанные выше.
Еще в одном варианте осуществления настоящего изобретения представлен радиоактивно меченный субстрат для бактериальной бета-лактамазы, подходящей для визуализации с помощью ПЭТ или ОФЭКТ. В этом варианте осуществления радиоактивной меткой может быть фтор-18, азот-13, кислород-18, углерод-11, технеций-99m, йод-123 или индий-111.
Настоящее изобретение относится к системам и способам оптической визуализации бактериальной болезни и/или инфекции. Эти системы и способы являются чрезвычайно чувствительными инструментами для определения количества и локализации бактерий во время болезни и для анализа in vivo антибактериальной активности препарата в режиме реального времени. Считается, что эти системы эффективны для обнаружения бактериальных патогенов на уровне одиночной клетки. Упомянутые системы и способы визуализации in vivo (IVI) можно применять непосредственно у пациентов в клинических условиях.
Системы и способы по настоящему изобретению применимы к видам бактерий, которые в природе обладают бета-лактамазной активностью или приобрели такую активность. Неограничивающие примеры бета-лактамазоположительных видов бактерий представляют собой Bacteroides, Clostridium, Streptococcus, Staphylococcus, Pseudomonas, Legionella, Mycobacterium, Haemophilus, Escherichia, Salmonella, Shigella или Listeria. В частности, рассматривается диагностика, локализация и количественное определение Mycobacterium, например, Mycobacterium tuberculosis и Mycobacterium bovis. Преимущество описанных в настоящем изобретении систем и способов состоит в том, что они не требуют создания бактериального штамма для обнаружения, но вместе с тем предполагается, что способы улучшения экспрессии, активности и/или секреции бета-лактамазы повышают чувствительность обнаружения. В этой связи считается, что бета-лактамаза бактериальных видов может быть обнаружена путем введения бета-лактамазы в бактерии или штамм любого рассматриваемого вида любым применимым способом, при котором осуществляется экспрессия и секреция бета-лактамазы на достаточном уровне, позволяющем проводить его чувствительное обнаружение. Это может быть достигнуто в условиях in vitro или in vivo с использованием известных и стандартных способов доставки, включающих использование фага, который является подходящим носителем для доставки, применяемым у млекопитающих.
Системы визуализации настоящего изобретения способны обнаруживать in vivo флуоресцентный, люминесцентный или колориметрический сигнал, генерируемый соединением или репортером, которые служат субстратом для бета-лактамазной активности. Системы визуализации известны в данной области техники и коммерчески доступны. Например, система последовательной репортер-ферментной флуоресценции (SREF), система последовательной репортер-ферментной люминесценции (SREL) или система биолюминесценции могут применяться для обнаружения продуктов бета-лактамазной активности. Кроме того, полученные сигналы можно использовать для создания трехмерного отображения, полезного для определения локализации бактериального патогена. Рядовой специалист в данной области техники сможет легко выбрать длину волны возбуждения и эмиссии для этих систем, на основе используемого соединения и/или репортера и типа сигнала, который необходимо обнаружить. Сигнал возбуждения, в качестве примера, может иметь длину волны в диапазоне от приблизительно 540 нм до приблизительно 730 нм, и примером сигнала эмиссии может быть сигнал в диапазоне от приблизительно 650 нм до приблизительно 800 нм. Также предполагается, что системы визуализации по настоящему изобретению могут также обнаруживать in vivo другие сигналы, например, генерируемые излучением, или любые обнаружимые или читаемые сигналы, продуцируемые действием бета-лактамаз на подходящем субстрате, или другие агенты обнаружения.
Бета-лактамазные субстраты по настоящему изобретению могут представлять собой химические субстраты или субстраты квантовых точек. Например, субстратами для визуализации с применением SREL или SREF могут быть флуорофор, каркасный люциферин или другое флуоресцентное, люминесцентное или колориметрическое соединение, репортер или другие реагенты для обнаружения, которые создают наилучший сигнал для требуемого применения. Субстрат является нетоксичным или имеет очень низкую токсичность на уровнях, которые позволяют хорошо проникать в любую ткань, и высокое соотношение сигнала к шумовому сигналу. Сигнал может представлять собой близкий к инфракрасному сигнал, инфракрасный или красный световой сигнал, например, от приблизительно 650 нм до приблизительно 800 нм.
Например, субстратами могут быть флуорогенные субстраты или субстраты квантовых точек, которые генерируют сигнал при расщеплении бета-лактамазой в условиях in vitro или in vivo. Флуорогенные субстраты могут содержать донор FRET (резонансный перенос энергии флуоресценции), такой как индоцианиновый краситель, например, Cy5, Cy5.5 или Cy7, и гаситель FRET, такой как гасящие группы QSY21, дисульфонат QSY21, QSY22 или дисульфонат QSY22. Дополнительно флуорогенные субстраты могут содержать перацетилированный D-глюкозамин для улучшения клеточной пенетрации и/или могут быть связаны с маленьким пептидом, таким как ТАТ, но не ограниченный им. Дополнительно субстрат можно модифицировать для повышения интенсивности его сигнала, улучшения способности к тканевой пенетрации, специфичности или способности к хорошему распределению во всех тканях. Кроме того, предполагается, что другие способы мечения ткани, клеток или других соединений в комбинации с упомянутыми субстратами являются полезными для улучшения чувствительности и обнаружения бактериальных патогенов.
В частности, флуорогенные субстраты могут обнаруживать активность бета-лактамазы в бактериальной клеточной культуре или в культуре однородных бактериальных клеток in vitro. Примерами химических флуорогенных субстратов являются CC1, CC2, CHPQ, CR2, CNIR1 или CNIR6. Альтернативно флуорогенным субстратом, применяемым для визуализации in vivo, может быть CNIR2, CNIR3, CNIR4, CNIR5, CNIR5-QSY22, CNIR7, CNIR9, CNIR10 или CNIR-ТАТ. Указанные флуорогенные субстраты являются полезными в системе последовательной репортер-ферментной флуоресценции (SREF). Предполагается, что бета-лактамазные субстраты эффективны для обнаружения однородных бактериальных клеток в условиях in vitro или in vivo.
Другим примером флуорогенного субстрата для обнаружения бета-лактамазы in vivo является каркасный люциферин, например, Bluco, Bluco2 или Bluco3, но не ограниченный вышеперечисленным. Этот субстрат содержит D-люциферин, основу люциферазы светлячка (Fluc) и бета-лактам, основу бета-лактамазы. Расщепление бета-лактама посредством фермента высвобождает D-люциферин, который люминесцирует при окислении Fluc. Каркасные люциферины полезны в системе последовательной репортер-ферментной люминесценции (SREL) или в других биолюминесцентных системах визуализации.
Флуоресцентные белки также могут быть полезными для обнаружения бактериальных патогенов в условиях in vitro и in vivo. Флуоресцентные белки (FP), такие как mPlum, mKeima, Mcherry и tdTomato, клонируют в векторы экспрессии. Рассматриваемый бактериальный патоген, такой как M. tuberculosis, трансформируют с конструкцией FP. Экспрессия флуоресцентного белка бактериями дает сигнал, обнаруживаемый при визуализации. В других системах визуализации можно задействовать рекомбинантные бактерии, которые трансформированы для секреции других ферментов, таких как бета-галактозидаза, которая в присутствии флуорофоров, например, C2FDG, C12RG или DDAOG, дает флуоресцентный сигнал. Также системы визуализации используют другие рекомбинантные системы, экспрессирующие другие люциферазы, такие как люцифераза жука-щелкуна красного или rLuc8, которые продуцируют сигнал в присутствии D-люциферина.
Альтернативно можно применять системы визуализации позитронно-эмиссионную томографию (ПЭТ) или однофотонную эмиссионную компьютерную томографию (ОФЭКТ). Образцы могут содержать субстраты бета-лактамазы, бета-лактамазоподобного фермента или другого подобного фермента или белка патогенных бактерий, описанных в настоящем изобретении. Способы визуализации ПЭТ и ОФЭКТ известны в данной области техники. Для визуализации ПЭТ образцы субстрата могут быть меченными позитрон-активным радиоизотопом, таким как, без ограничения, фтор-18, кислород-18, углерод-11 или азот-13. Для визуализации ОФЭКТ образцы субстрата могут быть меченными гамма-активным радиоизотопом, таким как, без ограничения, технеций-99m, йод-123 или индий-111. Образцы для ПЭТ и ОФЭКТ можно синтезировать и метить с использованием стандартных и известных способов химического и радиохимического синтеза.
Предполагается, что конструкцию и специфичность зондов можно максимизировать с использованием малых молекул, таких как цефероперазон, для моделирования бета-лактамазного кармана фермента. Таким образом, используя указанную высокопропускную систему малых молекул, в диагностических целях можно разрабатывать субстраты с наибольшей чувствительностью, подходящие для создания сигнала, который будет эффективно проникать из глубоких тканей, будет обнаружим с помощью современного оборудования для визуализации и будет предотвращать перекрестную реагентность с другими видами бактерий. Также, такие чувствительные и специфичные образцы субстрата эффективны на уровне единственной бактерии и могут количественно усиливать полученный от нее сигнал от 100 до 1000-кратного увеличения. Также предполагается, что для повышения специфичности исследования можно разработать бета-лактамазоподобные ферменты и связывающие пенициллин белки, отличные от бета-лактамазы M. tuberculosis.
Системы и способы, описанные в настоящем изобретении, являются эффективными для обнаружения, определения локализации, количества и жизнеспособности бактериального патогена в режиме реального времени. Визуализацию можно осуществлять in vitro с клеточной культурой или культурой однородных клеток, или ex vivo с клиническим образцом или экземпляром, используя системы SREL или SREF, или in vivo внутри субъекта, применяя любую из раскрытых систем визуализации. Образцы, используемые in vitro, могут включать без ограничения биоптаты, плевральную жидкость и другие субстанции организма, включающие кровь, слюну, мочу и кал, в которых может находиться бактериальный патоген. Таким образом, системы и способы согласно настоящему изобретению являются эффективными для диагностики патофизиологического состояния, такого как болезнь или инфекция, связанного с бактериальным патогеном. Поскольку существует возможность обнаружения на очень низком уровне, включающем единственную бактерию, можно проводить немедленную диагностику и в более ранний период инфекции, чем существующие в настоящее время способы диагностики. Системы и способы, описанные в настоящем изобретении, можно применять для тестирования и регулярной проверки работников здравоохранения, которые могут подвергаться риску бактериальной инфекции. Дополнительно, эти системы и способы могут также использоваться для проверки и обнаружения загрязнения на инструментах, посуде, оборудовании, рабочих поверхностях, одежде и на людях. Поскольку до 40% работников здравоохранения инфицированы высокорезистентным к лекарствам туберкулезом (XDR-TB) и основными областями локализации инфекции являются носовые проходы и трещины на руках, вызванные частым мытьем, настоящее изобретение является полезным в качестве способа скрининга на бактериальные патогены в учреждениях здравоохранения и среди работников здравоохранения. Упомянутые системы и способы можно использовать применительно к сельскому хозяйству и зоологии для обнаружения бета-лактамазы, при необходимости.
Также в технологии визуализации по настоящему изобретению существует устойчивая корреляция мощности сигнала и количества бактерий. Таким образом, эффективность соединений, лекарственных средств, фармацевтических соединений или других терапевтических агентов можно контролировать в режиме реального времени. Системы и способы, описанные таким образом в настоящем изобретении, обеспечивают высокопропускную систему скрининга антибактериальных веществ. Поскольку для обнаружения бета-лактамазы необходимо, чтобы бактерии были жизнеспособными, уровни фермента в присутствии одного или более терапевтических агентов служат мерой антибактериальной активности. Использование субстратов, подходящих для конкретных бактерий, позволяет быстро измерять изменения содержания бета-лактамазы и почти немедленно определять эффективность терапевтического агента. Высокопропускные системы являются полезными в случае от единственного зонда и до тысяч зондов в микропланшетах.
Следующие примеры приведены с целью демонстрации разных вариантов осуществления изобретения и не предназначены для какого-либо ограничения объема настоящего изобретения.
ПРИМЕР 1
Обнаружение Bla M. tuberculosis в культуре
Проводили сравнения возможных флуорогенных соединений и известных соединений, включающих нитроцефин (Calbiochem), субстрат CENTA Bla (Calbiochem), флуороциллин зеленый (Molecular Probes), CCF2-АМ (Invitrogen) и CCF4-АМ (Invitrogen), для обнаружения Bla в Mtb с использованием целых клеток и лизатов целых клеток, выращенных до ранней фазы логарифмического роста. Для всех этих образцов оценивали разведения для определения минимального числа бактерий или количества лизата, которое вызывало выраженный сигнал. Проводили титрование для определения количества фактически используемых КОЕ, перед анализом с интактными клетками и после анализа, и перед лизисом в случае лизатов. И чувствительность, и воспроизводимость оценивали четырехкратно с помощью спектрофотометрии, используя 96-луночные планшеты, которые инкубировали при 37°C в бактериальной культуральной среде в течение от 15 до 120 мин. Вначале использовали соединения в концентрациях, рекомендуемых изготовителем, для CNIR5 она составляет 2 нмоль, и указанную концентрацию применяли для визуализации in vivo. Проводили оценку разных концентраций наиболее чувствительных и репродуцируемых соединений в культуральной среде, чтобы определить минимальные концентрации, необходимые для максимального сигнала. Контроли в указанных экспериментах включают положительный контроль M. smegmatis и коммерчески доступный Bla (Sigma) и отрицательный контроль мутантным blaC Mtb (PM638, который предоставлен д-ром M. Pavelka, University of Rochester), в котором отсутствует Bla [1]. Также оценивали выработку Bla в BCG, поскольку в некоторых случаях BCG используют для визуализации in vivo (IVI) в BL2, где легко доступен более широкий диапазон оборудования для визуализации.
Оценка рекомбинантных конструкций Bla в мутантном blaC и туберкулезе дикого типа
Два многокопийных и два однокопийных вектора использовали для экспрессии Bla в Mtb. Основой многокопийных векторов является pJDC89, несущий промотор hsp60 (Phsp60) из pMV262, который показал умеренный уровень экспрессии генов. Указанный вектор также несет устойчивость к гигромицину, полилинкер ниже Phsp60, точку начала репликации E. coli и точку начала репликации микобактериального pAL5000. Чтобы увеличить экспрессию из указанного вектора, Phsp60 замещают на промотор L5 (PL5), который экспрессирует гены на уровне в 50-100 раз выше, чем Phsp60. Оба промотора являются относительно конститутивными и должны экспрессироваться в большинстве условий in vivo. Клонирования, если иначе не указано, в большинстве случаев осуществляют с использованием ПЦР-системы клонирования In-Fusion 2.0 PCR (Clontech), которая позволяет прямое клонирование фрагментов в любую линеаризованную конструкцию, используя минимальные области гомологии в 15 пар оснований (п.о.) на праймерах, используемых для полимеразной цепной реакции (ПЦР) рассматриваемой области.
Два сконструированных вектора модифицировали в донорные векторы Gateway (Invitrogen) путем клонирования фрагмента ПЦР, несущего ген ccdB и обе левую и правую рекомбинированные последовательности Gateway, расположенные ниже каждого промотора. Векторы, несущие указанную область, должны сохраняться в штамм ccdB Survivor, который позволяет сохранять эту область; при этом в другом штамме E. coli эта область будет летальной и ее используют, чтобы предотвратить сохранение нерекомбинантного вектора во время клонирования. Эти промоторы и ассоциированные области Gateway клонировали в pYUB412, который несет устойчивость к гигромицину, точку начала репликации E. coli, точку начала репликации фага L5 (attP) и рекомбиназу L5, то есть их объединяли в сайте attB в микобактериальной хромосоме и сохраняли с помощью устойчивости микобактерии в одной копии. В каждый из этих векторов клонировали Bla Mtb путем ПЦР с использованием праймеров, которые несут рекомбинированные последовательности Gateway, посредством реакции Gateway BP (Invitrogen). Эти векторы трансформировали в мутантные Mtb и blaC путем электропорации, согласно описанию [2]. Полученные штаммы Mtb оценивали для обнаружения, используя анализы in vitro, описанные для анализа эндогенного Bla, и показатели интенсивности сигнала по сравнению с показателями мутантного blaC в качестве отрицательного контроля и дикого типа с подходящим векторным скелетом единственным.
Хотя CNIR5 представляет собой высокопроницаемую мембрану, мощность сигнала можно увеличивать с помощью нацеливания Bla на мембрану клетки-хозяина, которая имеет большую площадь поверхности для репортера, чем единственная бактерия, и улучшает доступ к соединению. Поскольку фагосома микобактерии не является статической, взаимодействуя с несколькими путями рециклинга липидов и рецепторов, а также имеет несколько маркеров, присутствующих в эндосомах из рециклинга, нацеленные должным образом белки должны достигать плазматической мембраны клетки-хозяина через микобактериальную фагосому. Секреция Mtb Bla из бактерий происходит посредством ТАТ-сигнала, расположенного на его аминоконце, что делает идеальной метку карбоксиконца, нацеливающую этот белок на плазматическую мембрану. Белки, заякоренные гликозилфосфатидилинозитолом (GPI), такие как CD14, которые хорошо экспрессируются на поверхности макрофагов, локализуются на плазматической мембране посредством карбоксиконцевой сигнальной последовательности.
Создавали конструкцию слитого (Bla::GPI) с карбокси-концевыми 24 сигнальными последовательностями аминокислот GPI-заякоренного белка от CD14 и Bla от Mtb. Затем этот слитый белок помещали во все четыре системы экспрессии для Mtb с использованием системы Gateway и трансформировали и в дикий тип Mtb, и в мутантный blaC. Полученные штаммы, экспрессирующие Bla::GPI, мутантный blaC в качестве отрицательного контроля и оригинальный Bla, оценивали на содержание Bla на поверхности инфицированных макрофагов, используя внутриклеточные анализы. Исследовали как интактные инфицированные макрофаги, так и макрофаги, лизированные 0,1% тритоном.
Флуоресцентные спектры субстратов перед гидролизом и после гидролиза
Спектры возбуждения и эмиссии получали в 1 мл раствора ФБР в концентрации 1 мкмоль. К указанному раствору добавляли 10 нмоль очищенного Bla и снова отмечали спектры эмиссии и возбуждения, до выявления какого-либо дополнительного изменения. Увеличение сигнала флуоресценции в образцах после гидролиза Bla оценивали путем сравнения интенсивности эмиссии при значении 690 нм, которое является длиной волны пиковой эмиссии.
Ферментативная кинетика in vitro зондов в качестве субстратов Bla
Скорость увеличения (v) интенсивности флуоресценции при длине волны приблизительно 690 нм использовали как меру скорости гидролиза образца. Скорость (v) измеряли при разных концентрациях 5, 10, 20, 50, 80 мкмоль при концентрации Mtb Bla 1 нмоль. Для оценки значений влияния kcat и Km зонда на гидролиз Bla использовали график двойной-обратной зависимости скорости гидролиза субстрата (1/v) от концентрации субстрата (1/[образец]).
Биоустойчивость субстрата
Скорость спонтанного гидролиза субстрата при физиологических условиях также можно оценивать по скорости увеличения интенсивности флуоресценции при ~690 нм. Таким образом, устойчивость субстрата в водном буфере и в сыворотке можно легко оценить путем количественного определения флуоресценции после инкубации в течение 1 часа при комнатной температуре.
Визуализация экспрессии Bla в культивированных клетках
Субстрат тестировали с клеточными линиями, трансфицированными Bla (cmv-bla) и диким типом Jurkat и клеточной линией глиомы C6, и получали изображение с флуоресцентным микроскопом, используя известные условия визуализации [3].
Линейная корреляция между уровнями мРНК и сигналами NIRF
Клетки дикого типа и cmv-bla Jurkat смешивали в разных отношениях (10%, 20%, 40%, 60% и 80% клеток cmv-bla) с плотностью клеток 5×105/мл. После инкубации в течение 30 минут 5 мкмоль субстрата в каждой смеси клеток каждый образец промывали холодным фосфатно-буферным раствором (ФБР), центрифугировали и подвергали лизису. Измерения флуоресценции проводили на конечных супернатантах. Содержание мРНК и фермента Bla количественно определяли с помощью нозерн-блоттинга. Кривая зависимости концентрации мРНК от интенсивности флуоресценции Cy5.5 показывает, существует ли между ними линейная зависимость.
Локализация и регуляция бета-лактамазы M. tuberculosis в культуре
Транскрипцию Bla анализировали с помощью qRT-ПЦР (ПЦР с обратной транскрипцией) по ходу кривой роста Mtb, инокулированных при оптической плотности O.D. 0,05 и выращенных до стационарной фазы (O.D.=2). Оценку уровня транскрипции проводили путем ежедневного выделения РНК из аликвотных количеств этой же культуры, и все культуры выращивали в трех экземплярах. Выделение РНК [4] и qRT-ПЦР с использованием SYBR Green [5] проводили, как описано ранее. Содержание РНК подтверждали нозерн-блоттингом в одной или двух ключевых точках на кривой роста и все измерения стандартизировали по 16S рРНК. Данные сравнивали с измерением активности Bla с нитроцефином при тех же условиях, используя лизаты целых бактерий и целых клеток.
Изучали способность бета-лактамов индуцировать blaC. Транскрипты РНК анализировали в присутствии и в отсутствие бета-лактамов тем же образом по всей кривой роста. Инкубировали 50, 250 и 500 мкг/мл карбенициллина, который убивает Bla-отрицательные Mtb, совместно с Mtb, которые выращивали до ранней фазы логарифмического роста в течение двух часов, и определяли уровни транскриптов blaC вместе с активностью Bla в целых клетках и в лизатах целых клеток. Количественно определяли содержание Bla с помощью стандартной кривой, полученной с использованием коммерчески доступного Bla (Sigma), и выращенный сходным образом мутантный Mtb blaC включали в исследование в качестве отрицательного контроля активности BIa.
Обнаружение Bla в макрофагах
В общем, клетки J774A.1 высевали в 96-луночные плоскодонные планшеты с плотностью 1×104 клеток/на лунку и инкубировали в течение ночи при 37°C. Суспензии однородных клеток Mtb, выращенных до ранней фазы логарифмического роста, добавляли в разных вариантах множественного заражения от 1000 до 0,001 бактерий на клетку и инкубировали при 37°C в течение 30 мин. Затем лунки дважды промывали ФБР и свежей средой с добавлением 200 мкг/мл амикацина и инкубировали в течение 2 часов при 37°C, чтобы убить внеклеточные бактерии. После этого лунки промывали ФБР и инкубировали в свежей среде вместе с разными концентрациями тестового соединения в течение периода от 60 до 180 минут перед спектрофотометрическим измерением сигнала. Для оценки значения проницаемости клетки-хозяина в полученных показателях перед добавлением соединений 0,1% тритоном X-100 лизировали двойные лунки.
Во всех точках времени использовали четыре необработанные лунки для определения количества КОЕ, ассоциированных с клетками. Локализацию сигнала подтверждали с помощью флуоресцентной микроскопии для соединений, подтвердивших наибольшую эффективность. Микроскопические исследования проводили подобным образом, но с использованием восьмилуночного предметного стекла для определения локализации сигнала, определения процента бактерий с положительным сигналом и для оценки интенсивности локализованного сигнала.
Биопробы и фармакокинетика
После анестезии мышей умерщвляли смещением шейных позвонков в разные временные интервалы (30 минут, 240 минут, 12 часов, 24 часа, 48 часов и 72 часа) после инъекции (по три мыши в каждой точке времени). Забор образцов крови осуществляли пункцией сердца и быстро забирали ткани (опухолей, сердца, почки, печени, мочевого пузыря, желудка, мозга, поджелудочной железы, тонкой и толстой кишки, легких и селезенки) для измерения близкой к инфракрасной флуоресценции с помощью флуорометра. Данные выражали как единицы флуоресценции (ЕФ) на грамм ткани [ЕФ/(г ткани)].
Анализ активности бета-лактамазы
Измеряли уровень фермента Bla в ксенотрансплантированных опухолях с использованием следующего протокола: после забора опухоль дважды промывали холодным ФБР; добавляли лизирующий буфер фирмы Promega (4 мл/г ткани) и гомогенизировали раствор ткани; три раза замораживали и оттаивали гомогенат и центрифугированием собирали супернатант; оценивали активность Bla, используя флуорогенный субстрат CC1. Подтверждали наличие мРНК Bla в опухолях cmv-bla с помощью последующей экстракции РНК согласно протоколу Qiagen Inc. и проведения анализа RT-ПЦР. Эти измерения подтверждают корреляцию активности Bla с наблюдаемым близким к инфракрасному сигналу в опухолях с трансфекцией cmv-bla.
Определение экспрессии Bla РНК in vivo
Экстрагировали экспрессируемую in vivo РНК Bla с использованием стандартного протокола экстракции РНК для туберкулеза [6] и проводили qRT-ПЦР относительно конститутивного контрольного гена рРНК. Эти измерения обеспечивают способ оценки уровней экспрессии Bla во всех тканях по сравнению с уровнями наблюдаемых IVI сигналов. Если полученные уровни РНК ниже уровней, обнаруживаемых с помощью RT-ПЦР, при этом в тканях присутствуют КОЕ в количестве, которое можно подсчитать, проводят амплификацию кДНК перед RT-ПЦР, используя полимеразу phi29 (Fidelity Systems), которая способна линейно амплифицировать ДНК с высокой точностью, позволяя достоверно количественно определить уровни матрицы после амплификации.
Экспрессия, устойчивость и вирулентность штаммов Bla in vivo
Восемь групп по четыре мыши Balb/c аэрозольным путем инфицировали в диапазоне от 100 до 1000 КОЕ/легкое. Бактериальные штаммы оттаивали из хранившихся при -80°C, пропускали дважды через шприцевую иглу 27G, чтобы получить суспензии однородных клеток и использовать их для аэрозольного инфицирования. Аэрозольное инфицирование осуществляли с помощью «Мэдисонской» камеры, сконструированной в Висконсинском университете, которая разработана для прямой доставки капельной взвеси клеточных ядер в просвет альвеол х7ъ10). Инфицирование Mtb проводили с сертифицированным оборудованием ABSL3, разработанном для обращения с вирулентными штаммами туберкулеза. Инфицированные мыши находились в изолированном режиме в ABSL3 в Центре сравнительной медицины (Center for Comparative Medicine) до вскрытия трупов. Одну группу из четырех мышей для каждого бактериального штамма (blaC и WT) подвергали вскрытию во всех точках времени (1, 14, 28 и 72 дня) для определения КОЕ, содержания РНК для активности blaC и Bla в легких и селезенке. Определение уровней транскрипции РНК и активности Bla с использованием нитроцефина проводили согласно описанию в настоящем изобретении.
Исследовали устойчивость и влияние на вирулентность экспрессии рекомбинантного Bla in vivo для двух рекомбинантных штаммов, которые показали перспективность использования для IVI. Двенадцать групп из четырех мышей Balb/c инфицировали аэрозольным путем в диапазоне от 100 до 1000 КОЕ/на легкое, как описано выше. В одной группе из четырех мышей для каждого бактериального штамма (дикий тип, конструкция 1 и конструкция 2) проводили вскрытие во всех точках времени (1, 14, 28 и 72 дня) для определения КОЕ, проведения гистопатологии, определения присутствия соответствующей конструкции и активности Bla в легких и селезенке. Определяли процент бактериальной популяции, несущей конструкцию, с помощью анализов Bla, проводимых, по меньшей мере, на 20 отдельных колониях с титровальных планшетов КОЕ. Исследование активности Bla проводили на гомогенизированных тканях для оценки общего уровня оставшегося Bla. Оценку активности Bla проводили с использованием нитроцефина согласно настоящему описанию.
ПРИМЕР 2
Прижизненная визуализация с помощью микроскопии с использованием модели клеточной трансплантации
Универсальный донор Tr, CD8+T-клетки, моноциты, макрофаги и дендритные клетки трансплантировали сингенным мышам, инфицированным BCG, и наблюдали распределение указанных клеток в течение времени с помощью биолюминесцентной визуализации in vivo (BLI) и прижизненной микроскопии с визуальным контролем (IVM). Линия трансгенных мышей, у которых посредством промотора бета-актина вырабатывается люцифераза, служила источником тканей и клеток, которые излучают свечение у нетрансгенных животных [11-12]. У мышей указанной линии (L2G85) наблюдали яркую биолюминесценцию от люциферазы светлячка (Fluc), но слабую флуоресценцию от GFP, поэтому ее совмещали с отдельной линией, проявляющей сильную экспрессию GFP и флуоресценцию в лимфоцитах. Таким образом, можно отслеживать с помощью BLI пространственное распределение универсальных донорных стволовых клеток и других клеток у реципиента, по мере их размножения, перераспределения или выведения, и обнаруженные клетки можно впоследствии визуализировать с помощью FVM, используя GFP.
Мышиные L2G85 конструировали в основу FVB, таким образом, задействовали дикий тип мышей FVB/NJ (Jackson Labs) в качестве реципиентов клеток от L2G85, с профилактикой отторжения трансплантированных клеток. В общей сложности 80 мышей FVB/NJ интраназально инфицировали BCG в количестве 104 КОЕ в 20 мкл солевого раствора. Четырех мышей умерщвляли через 24 часа после инфицирования для определения исходного показателя КОЕ в легких. Через 14 дней после инфицирования дополнительно четыре мыши умерщвляли для гистопатологии и определения КОЕ в легких и селезенке. Также через 14 дней оставшихся 72 мыши разделяли на 4 группы и вводили в/в в хвостовую вену L2G85 Tr, клетки CD8T, моноциты, макрофаги, дендритные клетки или не вводили клетки (контроль). На 28, 42 и 56 день исследовали шесть групп по четыре мыши (включая контрольных мышей) согласно описанию [12] в присутствии D-люциферина.
После получения визуального изображения проводили более подробное исследование очевидных поражений путем прижизненной микроскопии (IVM) с использованием оптоволоконного конфокального флуоресцентного микроскопа (Cell-viZio, Mauna-Kea). Для IVM применяли гибкий минизонд, состоящий из десятков тысяч оптических волокон. Проводили общее обезболивание и обследуемую область изучали через небольшой быстро заживающий разрез, что предотвращает необходимость умерщвления животных после хирургического вмешательства и позволяет проводить визуализацию на клеточном уровне.
Контрольных мышей умерщвляли после визуализации, чтобы определить показатели КОЕ в легких и других органах, где обнаружили сигнал у мышей, которым вводили клетки. Получали дорсальное, вентральное и два боковых изображения, чтобы лучше определить источник фотонной эмиссии. Дополнительное подтверждение получали от подгруппы животных путем анализа тканей, инкубацией свежих тканей в D-люциферине и их осмотром без вышележащих тканей. Во всех достоверно инфицированных тканях проводили подробную гистопатологию для флуоресцентной микроскопии, чтобы обнаружить экспрессирующие GFP трансплантированные клетки, и проводили быстрое окрашивание гематоксилином и эозином и кислотой для идентификации бактерий и клеток в тканях.
Визуализация in vivo отдельных бактерий и иммунных клеток во время образования гранулемы
Применяя модель трансплантации, выбирали два типа трасплантируемых клеток, которые лучше всего позволяют визуализировать образование гранулемы, чтобы использовать их для визуализации и бактерий и клеток-хозяев у живых мышей. Выбирали три точки времени, когда поражения только становятся заметными, когда они хорошо сформированы, и последнюю точку времени, когда можно наблюдать сигнал от трансплантированных клеток. В общей сложности 32 мыши FVB/NJ инфицировали интраназально в количестве 104 КОЕ BCG, экспрессирующей IVI репортер, например tdTomato, в 20 мкл солевого раствора. Дополнительную группу из четырех контрольных мышей оставляли неинфицированными. Четырех экспериментальных мышей умерщвляли через 24 часа, чтобы определить исходные показатели КОЕ в легких после инфицирования. Через 14 дней после инфицирования дополнительно четырех экспериментальных мышей умерщвляли для гистопатологии и для определения КОЕ в легких и селезенке. Также через 14 дней оставшихся 24 мышей разделяли на 4 группы, 12 мышам из этих мышей вводили в/в в хвостовую вену клетки L2G85, и 12 мышам в качестве контроля клетки не вводили. В трех точках времени в двух группах из четырех мышей (группа введения по сравнению с группой без введения клеток) проводили визуализацию согласно описанию (12) в присутствии D-люциферина.
После получения визуального изображения проводили более подробное исследование очевидных поражений путем прижизненной микроскопии (IVM) с использованием оптоволоконного конфокального флуоресцентного микроскопа (Cell-viZio, Mauna-Kea). Проводили общее обезболивание и обследуемую область изучали через небольшой быстро заживающий разрез. Контрольных мышей умерщвляли после визуализации, чтобы определить показатели КОЕ в легких и других органах, где обнаружили сигнал у мышей, которым вводили клетки. Получали дорсальное, вентральное и два боковых изображения, чтобы лучше определить источник фотонной эмиссии. Дополнительное подтверждение получали от подгруппы животных путем анализа тканей, инкубацией свежих тканей в D-люциферине и их осмотром без вышележащих тканей. Применяли наборы фильтров как для трансплантируемых клеток, так и для сигнала бактериального репортера в иссеченных тканях. Для всех достоверно инфицированных тканей проводили подробную гистопатологию для флуоресцентной микроскопии, чтобы обнаружить экспрессирующие GFP трансплантированные клетки, а также сигнал бактериального репортера, и проводили быстрое окрашивание гематоксилином и эозином и кислотой для идентификации бактерий и клеток в тканях.
Анализ визуализации
Полученные изображения обрабатывали на компьютере с использованием коммерчески доступного программного обеспечения Living Image от компании Xenogen Inc. Рассматриваемые области (РО) оттеняли опухоли на флуоресцентных изображениях целого тела. Одной из основных характеристик системы визуализации IVIS является ее стандартизация по источнику прослеживаемой спектральной энергетической яркости Национального института стандартов и технологии (NIST). Эта калибровка обеспечивает преобразование импульсов ПЗС-камеры в излучение на рассматриваемой поверхности с учетом потерь через оптические устройства и апертуры (f/stop) и принимает во внимание время визуализации и биннинг. Полученное изображение, таким образом, представляют в физических единицах поверхностного излучения (sr) (фотон/сек/см2/sr). Интегрированный сигнал от РО (в единицах фотон/секунды) от инфицированных мышей, контрольных мышей и из нормальных тканей сравнивали у разных мышей (в соотношении инфицированные:контрольные:нормальные ткани). Статистический анализ проводили с использованием программного обеспечения Prism GraphPad 3.0 (GraphPad, Сан-Диего, Калифорния). Уровень значимости устанавливали как P<0,05.
ПРИМЕР 3
Флуорогенный субстрат для обнаружения бета-лактамаз
CC1, CC2, CHPQ и CR2
Флуорогенные соединения CC1, CC2, CHPQ и CR2 эффективны для обнаружения активности Bla in vitro и в культуре однородных клеток. Эти образцы не являются флуоресцентными перед гидролизом с помощью Bla и становятся флуоресцентными после реакции Bla (фиг. 1A-1B). Можно выбирать диапазон разных флуоресцентных эмиссий при необходимости обнаружения Bla: от синего с CC1 и CC2, зеленого с CHPQ до красного CR2. Эти новые флуорогенные субстраты меньше чем CCF2, легки для изготовления, просты в использовании, являются высокочувствительными для обнаружения активности Bla и облегчают обнаружение активности Bla в разнообразных биологических образцах.
Вставка олефиновой группы между 3'-углеродом лактама и уходящей группой помогает улучшить кинетическую эффективность гидролиза посредством Bla. Например, для CC1, значение kcat составляет 174 s-1, при этом значение kcat его аналога без вставки двойной связи составляет только 35 s-1. Наблюдается примерно 5-кратное увеличение каталитической эффективности. Предполагается, что эта разработка может служить общей стратегией создания широкого разнообразия флуорогенных субстратов для Bla, включая близкие к инфракрасным субстраты для флуоресцентной визуализации целого организма животных.
Также предполагается, что образцы можно улучшить новым гасителем QC-1 и близким к инфракрасному флуорофором IRDye 800CW. Дополнительно, образцы на основе IRDye можно модифицировать добавлением сульфонатных групп.
CNIR1, CNIR2, CNIR3, CNIR4, CNIR5, CNIR9 и CNIR10
Для отображения экспрессии Bla у живых животных с помощью флуоресцентной визуализации целого организма преимущество имеет близкий к инфракрасному/инфракрасный флуорогенный субстрат, поскольку инфракрасный/близкий к инфракрасному свет обладает лучшей проницаемостью в ткани и меньшим рассеиванием света, чем видимый свет, и намного меньшую абсорбцию гемоглобином [13]. Соединения CNIR1, CNIR2, CNIR3, CNIR4, CNIR5, CNIR9 и CNIR10 представляют собой ряд близких к инфракрасному флуорогенных субстратов для визуализации экспрессии Bla в культивируемых клетках (фиг. 2A-2B). Эти соединения полезны в качестве основы для создания проницаемого для клеток близкого к инфракрасному флуорогенного субстрата для Bla и могут применяться, чтобы изучить влияние необходимости доступности зонда для бактерий на внутриклеточном уровне или у животных.
Информация об активности Bla основана на резонансном переносе энергии флуоресценции (FRET). Образцы содержат донор FRET и гаситель FRET. Для визуализации в условиях in vivo флуорофор в идеале должен иметь эмиссию больше чем 650 нм и низкую токсичность. Индоцианиновые красители (Cy5, Cy5.5 и Cy7) имеют эмиссию от 650 до 800 нм и применяются у десятков тысяч пациентов при малом количестве зарегистрированных побочных эффектов. В этой связи в качестве донора FRET был выбран Cy5. Было показано, что гасящая группа QSY21, не флуоресцентная сама по себе, имеет широкий спектр поглощения от 540 до 730 нм, достигающий максимума при 660 нм, и является эффективным гасителем для эмиссии Cy5.
CNIR1 является чрезвычайно нефлуоресцентным, но вырабатывает высокофлуоресцентный продукт с 57-кратным увеличением интенсивности эмиссии при длине волны 660 нм после обработки с Bla [14]. Вместе с тем, сам CNIR1 не является проницаемым для клеток и, таким образом, не может давать отображение Bla in vivo. Для улучшения проницаемости мембран CNIR1 осуществляли конъюгацию CNIR1 с перацетилированным D-глюкозамином, CNIR3, который обладает хорошей клеточной проницаемостью и способностью давать отображение экспрессии Bla в живых однородных клетках. Добавление двух групп сульфоната на QSY21 для улучшения растворимости приводит к образованию CNIR4.
CNIR5 и CNIR6
Все соединения от CNIR1 до CNIR4 основаны на Cy5. Для визуализации у животных in vivo более предпочтителен Cy5.5, имеющий большую длину волны эмиссии. Таким образом, Cy5 заменяли на Cy5.5 и синтезировали CNIR5 (фиг. 3A). Конечный продукт очищали высокоэффективной жидкостной хроматографией (ВЭЖХ) и определяли показатели масс-спектрометрии (вычисленная масса для C122H123N11O39S10: 2687,98; наблюдаемая ионизация лазерной десорбцией с использованием матрицы - масс-спектометрия MALDI-MS [M+H]+: 2687,68). Непосредственно CNIR5 излучает слабую флуоресценцию при длине волны 690 нм при возбуждении, но после обработки Bla интенсивность возрастает больше чем в 9 раз (фиг. 3B). Кинетику его гидролиза посредством Bla измеряли в фосфатно-буферном растворе (ФБР) при уровне pH 7,1: каталитическая константа kcat=0,62±0,2 s-1, и константа Михаэлиса Km=4,6±1,2 мкмоль (значения были получены путем подбора методикой наименьшего квадрата по двойной-обратной зависимости скорости гидролиза от концентрации субстрата). Каталитическая эффективность CNIR5 составляет (kcat/km) 1,36×105 М-1s-1. CNIR5 проявлял высокую устойчивость в ФБР при скорости спонтанного гидролиза 1,75×10-7 s-1 и также в мышиной сыворотке, то есть наблюдали небольшое увеличение флуоресценции даже после инкубации в течение 12 часов. CNIR6 представляет собой аналог CNIR5 без перацетилированного D-глюкозамина и полезен в качестве контроля.
Также CNIR5 можно синтезировать путем замены QSY21 на QSY22 (фиг. 3C). Указанный синтез очень сходен с синтезом CNIR5 и не представляет трудностей. Синтез QSY22 рассмотрен ниже.
CNIR7
CNIR7 представляет собой модификацию CNIR5, который улучшает его чувствительность для визуализации Bla in vivo. Гасящая группа QSY21 дисульфонат, используемая в CNIR5, имеет максимальное поглощение при 675 нм, но максимальная эмиссия Cy5.5 при 690 нм. Поэтому, как в случае CNIR5, гасящая эффективность составляет только 90%, что в значительной степени способствует наблюдаемой фоновой флуоресценции. В FRET-паре QSY21 и Cy5 (CNIR1), в силу лучшего спектрального наложения между QSY21 и Cy5, гасящая эффективность составляла более 98%. Таким образом, гасящая группа, которая способна к абсорбции при 690 нм, будет лучше гасить Cy5.5 и может уменьшить фоновый сигнал. Применительно к QSY21 известно, что при замене индолина на тетрагидрохинолин максимум поглощения смещался в красную область на 14 нм.
Таким образом, синтезировали новую структуру дисульфонат QSY22 (фиг. 4A-4D) путем замены индолиновых групп в QSY21 на тетрагидрохинолины, которая сходным образом должна смещать максимум поглощения в красную область на 14 нм. Поскольку единственное структурное различие между этими двумя соединениями состоит в том, что в QSY22 задействован тетрагидрохинолин, который содержит шестичленное слитое кольцо, а в QSY21 используется пятичленный индолин, применяли реакцию сульфонирования, и предполагалось то же самое положение сульфонирования (пара) на бензольном кольце. Таким образом, дисульфонат QSY22 должен более эффективно гасить Cy5.5 и вызывать более слабый фоновый сигнал.
Во-вторых, значение kcat для CNIR5 составляет приблизительно 0,6 s-1, что намного меньше, чем показывает CC1 и CCF2. Также между гасителем и Cy5.5 вставлена двойная связь, следствием чего должно быть увеличение kcat. В-третьих, уменьшено расстояние между донором FRET, Cy5.5, и гасителем, дисульфонатом QSY22, чтобы улучшить эффективность переноса энергии. CNIR5 имеет длинную линкерную группу, содержащую цистеин, для включения в транспортер. В новом соединении CNIR7 транспортер связан с другим участком связывания на Cy5.5, поэтому больше нет необходимости включать в него длинный линкер. Кроме того, 4-аминотиофенол в CNIR5 замещен на 2-аминотиофенол, что должно дополнительно сократить расстояние между Cy5.5 и гасителем. Заключительное строение субстрата NIR, CNIR7, и его химический синтез показан на фиг. 5A-5B. Его синтез можно завершать еще более коротким путем, и он должен быть легче, чем синтез CNIR5.
CNIR7 также может содержать короткий катионоактивный пептид, такой как ТАТ-последовательность, для замещения ацетилированного D-глюкозамина. Чтобы избежать гидролиза пептидазами, вместо L-аминокислот используются D-аминокислоты. Было показано, что короткие катионоактивные пептиды, такие как третья спираль гомеодомена белка Antennapedia (15-16), основные домены белка HIV-I Rev и белка HTLV-1 Rex, основные домены белка HIV-1 Tat, способны проникать через плазматическую мембрану клеток.
Каркасный субстрат Bla для визуализации Bla при туберкулезе
Структура каркасного субстрата для Bla (Bluco) (фиг. 6A) содержит D-люциферин, основу люциферазы светлячка (Fluc) и бета-лактам, основу Bla. Фенольная группа D-люциферина является важной для его окисления посредством Fluc. Если указанная фенольная группа непосредственно присоединена к цефалоспорину в 3'-положении посредством эфирной связи, образуемый конъюгат должен быть плохим субстратом для Fluc, но оставаться субстратом для Bla. Открытие бета-лактамного кольца посредством Bla будет запускать спонтанную фрагментацию, что приведет к разрушению эфирной связи в 3'-положении и высвобождению свободного D-люциферина, который в этом случае может окисляться посредством Fluc в реакции образования света.
Для повышения устойчивости конъюгата выполняли окисление сульфида до сульфоксида на цефалоспорине, получая конечную структуру Bluco. Приготовление Bluco можно осуществлять посредством многоэтапного органического синтеза (фиг. 6B). Поскольку размер Bluco намного меньше, чем размер зондов ряда CNIR, Bluco способен лучше проникать через клеточную стенку M. tuberculosis. В нем можно просто использовать идентифицированную замену в 7-аминоположении, чтобы создать TB-специфичный каркасный люминесцентный субстрат для SREL-визуализации Bla в TB. Bluco также можно синтезировать с улучшенным Kcat путем вставки двойной связи (Bluco2) и с использованием карбаматного присоединения (Bluco3).
ПРИМЕР 4
FRET и кинетика флуоресцентных включений для CNIR4, CNIR5, CNIR9 и CNIR10
FRET in vitro
Определение активности Bla у E. coli с помощью CNIR5
Проводили предварительный эксперимент для проверки способности CNIR5 обнаруживать активность Bla у живых бактерий. E. coli трансформировали плазмидой с резистентностью к ампициллину и выращивали в течение ночи при 30°C. Клетки собирали и дважды промывали средой LB перед добавлением 500 нмоль CNIR5. Брали спектры флуоресценции с промежутками (Ex: 640 нм), данные показаны в фиг. 7. В конце измерения (t=160 минута) добавляли раствор очищенного Bla для подтверждения завершения гидролиза CNIR5. Результат указывает, что CNIR5 способен обнаруживать Bla у E. coli. Для сравнения, когда при тех же условиях использовали флуорогенный субстрат CCF2/AM от компании Invitrogen Inc., Bla у живых E. coli в среде LB не были обнаружены.
Спектры FRET
Фиг. 8B-8E показывают спектры эмиссии FRET для каждого из зондов CNIR4, CNIR5, CNIR9 и CNIR10 перед и после расщеплением с Bla в течение 10 мин.
Кинетика TEM-I E. coli и Bla-C M. tuberculosis с субстратами CNIR4 и CNIR5
Таблица 1 сравнивает кинетику ферментов TEM-1 E. coli и бета-лактамазы Bla-C M. tuberculosis с CNIR4 и CNIR5 в качестве субстратов (фиг. 9A-9B).
Проводили определение кинетики флуоресцентного включения в M. tuberculosis, используя указанные образцы CNIR. Показано включение и распределение зондов CNIR4 и CNIR5, использованных в качестве субстрата в среде только с M. tuberculosis (фиг. 10A-10Н) и в M. tuberculosis, инфицированных макрофагами (фиг. 11А-11Н).
Включение CNIR4 в M. tuberculosis
С помощью флуоресцентной конфокальной микроскопии показано, что CNIR4 внедряется внутриклеточно в инфицированные M. tuberculosis макрофаги (фиг. 12). Окрашивание DAPI (синим) обозначает ядра инфицированных клеток, клетки, меченные GFP M. Tuberculosis, проявляют зеленую флуоресценцию, и клетки, расщепленные с CNIR4, проявляют красную флуоресценцию. Отмечено, что флуоресценция с CNIR4 происходит от инфицированных клеток, при этом неинфицированные клетки не проявляют флуоресценции.
Обнаружение флуоресцентного сигнала от зондов CNIR in vivo
Мышей инфицировали подкожно M. tuberculosis в разных концентрациях. В нижний левый квадрант вводили 108 бактерий, в верхний левый квадрант вводили 107 бактерий и верхний правый квадрант вводили 106 бактерий. Флуоресценцию измеряли в присутствии каждого из зондов CNIR4, CNIR5, CNIR9 и CNIR10 (фиг. 13A-13D). У зонда CNIR5 выявлен наибольший флуоресцентный сигнал и его увеличение по мере повышения концентрации инокулята, и далее у зондов CNIR10 и CNIR9. CNIR4 не показал увеличение флуоресценции. Флуоресценцию от зондов CNIR4, CNIR5, CNIR9 и CNIR10 также измеряли у мышей, которых инфицировали в легкие аэрозольным путем M. tuberculosis дикого типа или M. tuberculosis, имевших мутацию в гене blaC в (фиг. 14A-14D). Наиболее высокая общая интенсивность флуоресценции выявлена у CNIR10 и далее у зондов CNIR9, CNIR5 и CNIR4 (фиг. 14E).
CNIR5 использовали в качестве субстрата для визуализации флуоресцентных включений и графического изображения их кинетики в течение времени у контрольных мышей и мышей, инфицированных M. tuberculosis аэрозольным путем, визуализацию которых проводили с использованием субстрата CNIR5. Изображения контрольных и инфицированных мышей получали в точки времени 1, 18, 24, 48 и 96 часов (фиг. 15A-15E). Пиковое включение CNIR5 происходило через 48 часов после аэрозольного инфицирования (фиг. 15E). Фиг. 16A-16B показывают изображения флуоресценции неинфицированных мышей или мышей, аэрозольно инфицированных M. tuberculosis, соответственно, визуализацию которых осуществляли с помощью трансиллюминации, а не отражения, для уменьшения фонового сигнала.
ПРИМЕР 5
Визуализация in vivo с CNIR5
CNIR5 в модели опухоли мыши
Клетки крысиной глиомы C6 в количестве приблизительно 1×106 вводили в левую лапу нагой мыши и такое же количество клеток крысиной глиомы C6, которые были устойчиво трансфицированы cmv-bla, вводили в правую лапу этой же нагой мыши. Когда размер опухолей достигал приблизительно 6 мм, мышам в хвостовую вену под анестезией вводили 7,0 нмоль CNIR5. Мышь сканировали в устройстве для визуализации IVIS200 с набором фильтров Cy5.5 (длина волны возбуждения: 615-665 нм; эмиссии: 695-770 нм), время исследования 1 секунда, в разное время после инъекции.
Фиг. 17A показывает ряд репрезентативных изображений, полученных перед инъекцией и через 2, 4, 12, 24, 48 и 72 часа после инъекции. Сразу через 2 часа после инъекции опухоли cmv-bla показывали более высокую интенсивность флуоресценции, чем опухоли C6 дикого типа (wt). Разница достигала наибольшего значения 1,6 через 24 часа и затем начинала уменьшаться примерно до 1,3 через 48 часов и 72 часа (фиг. 17B). В конце визуализации мышей умерщвляли, чтобы забрать органы и опухоли для визуализации ex vivo и для анализов биораспределения, с целью подтверждени данных визуализации. Фиг. 17C представляет собой изображение флуоресценции опухолей и органов, забранных у мертвой мыши через 24 часа после инъекции CNIR5, что согласуется с данными визуализации in vivo, при которой выявлена более интенсивная эмиссия Cy5.5 из иссеченой опухоли cmv-bla, чем из опухоли C6 wt. Для подтверждения экспрессии Bla в опухолях cmv-bla проводили анализ CC1 иссеченных опухолей от мышей, которым вводили CNIR5 (фиг. 17D); результат указывал на наличие высокого уровня экспрессии фермента у cmv-bla опухолей, тогда как в опухолях дикого типа наблюдалась небольшая активность Bla.
Чтобы дополнительно показать, что наблюдаемая разница обусловлена активацией CNIR5 посредством Bla, экспрессируемого опухолями, в качестве контроля был изготовлен CNIR6, который представляет собой аналог CNIR5, но без перацетилированного D-глюкозамина (фиг. 18A). CNIR6 может подвергаться гидролизу in vitro с помощью Bla так же эффективно, как CNIR5, но он не проницаем для клеток, и, таким образом, CNIR6 не может обладать способностью визуализации Bla in vivo. В фиг. 8B-18C показано отсутствие какой-либо значительно разницы между опухолями cmv-bla и контрольными опухолями на протяжении всего периода визуализации. Это отчетливо указывает на внедрение в клетки-мишени CNIR5 и его активацию посредством Bla. Данный результат также продемонстрировал значение группы D-глюкозамина для CNIR5 в целях визуализации Bla in vivo.
Биораспределение и фармакокинетика CNIR5 у мышей после в/в инокуляции
Мышам Balb/c в/в вводили CNIR5. Группы мышей умерщвляли для забора и обработки органов. Присутствие CNIR5 оценивали по интенсивности флуоресценции в каждом органе в течение времени. Фиг. 19A-19B показывают наличие сигнала CNIR5 как через 4 часа, так и через 24 часа после инъекций, соответственно. Устойчивый сигнал наблюдали от всех тканей, что предполагает системное распределение CNIR5 после 24 часов, при этом за это время не выявлено значительного ослабления сигнала.
Визуализация in vivo для локализации инфекции M. tuberculosis у мышей с Bla
Шесть групп из четырех мышей Balb/c в каждой группе аэрозольным путем инфицировали в диапазоне от 100 до 1000 КОЕ/на легкое, как описано в примере 1. Одну группу из четырех мышей использовали для визуализации во всех точках времени, и в каждой точке времени другую группу из четырех мышей умерщвляли и вскрывали для проведения гистопатологии и определения КОЕ в легких и селезенке. Через 24 часа, 7, 14, 28 и 72 дня проводили визуализацию на том же комплексе ABSL3, используя оборудование для визуализации Xenogen IVIS200. В группе контроля визуализации задействовали четырех неинфицированных бактериями животных, которым вводили реагент обнаружения с целью контроля фоновой флуоресценции от нерасщепленного соединения. Животных обезболивали изофлуораном в светонепроницаемой камере и проводили визуализацию при длине волны возбуждения 640 нм и c фиксацией изображений при 690 нм. В хвостовую вену внутривенно вводили CNIR5 в количестве 5 нмоль, которое, как было показано, являлось достаточным для IVI. Изображения получали перед инъекцией соединения и через 1, 2 и 4 часа после инъекции. При выявлении сигнала в любой из указанных точек времени впоследствии проводили визуализацию животных через 24, 48 и 72 часа, чтобы следить за рассеиванием сигнала.
Показаны изображения in vivo мыши, которая была инфицирована M. tuberculosis дикого типа (фиг. 20A) и контрольной мыши (фиг. 20B). Обеим мышам в/в вводили CNIR5 перед визуализацией. На указанных фигурах показано, что сигнал у инфицированной мыши поступал из легких. Трехмерная реконструкция сигнала показывает, что в среднем локализация сигнала находилась между легкими. Поскольку сигнал был усреднен и у мышей имеются два легких, предполагается, что указанная локализация является точкой максимального источника. Таким образом, можно применять соединение CNIR5 для определения локализации M. tuberculosis у живых млекопитающих. Для получения этого изображения использовали систему визуализации Xenogen/Caliper IVIS Spectrum.
Определение порога обнаружения M. tuberculosis у мышей с Bla
Зонд бета-лактамазы CNIR может обнаружить 100 или менее бактерий M. tuberculosis с помощью SREL - визуализации мышей в режиме реального времени (фиг. 21A). SREL-визуализацию проводили у живых неинфицированных мышей в качестве контроля (фиг. 21B) или у мышей, инфицированных M. tuberculosis (фиг. 21C). Цветная полоса показывает уровни эмиссии при 680 нм после возбуждения волной 620 нм. Цвет указывает на присутствие мощного сигнала, источником которого являются легкие, инфицированные Mtb, и показывает специфическую локализацию инфекции. Также могут быть определены пороги обнаружения для Pseudomonas, Staphylococcus и Legionella.
Визуализация in vivo инфекции M. tuberculosis у морских свинок с Bla
Инфицировали шесть групп из четырех морских свинок и проводили визуализацию тем же образом, как описано для мышей, со следующими исключениями. Во-первых, анализы проводили в точки времени только до 28 дней после инфицирования, поскольку предполагалось, что морские свинки начнут проявлять значительную смертность в более поздние точки времени. Во-вторых, для морских свинок были необходимы реагенты обнаружения в количестве в 20 раз больше (приблизительно 100 нмоль для CNIR5), чтобы достигнуть того же уровня содержания в сыворотке, которое требовалось у мышей, и соединение вводили через латеральную метатарзальную вену. Морских свинок инфицировали аэрозолем с помощью оборудования ABSL3 и до визуализации содержали в изоляции. Визуализацию проводили с комплектом ABSL3, используя оборудование для визуализации IVIS200, через 24 часа, 7, 14 и 28 дней после инфицирования. Четырех неинфицированных бактериями животных, которым вводили реагент для обнаружения, задействовали в качестве контрольной группы для визуализации, для отслеживания фоновой флуоресценции от нерасщепленного соединения.
Перед визуализацией в хвостовую вену внутривенно вводили CNIR5 в количестве 100 нмоль, которое оказалось достаточным для IVI. Изображения получали перед инъекцией соединения и через 1, 2 и 4 часа после инъекции. Если сигнал наблюдали в какой-либо из указанных точек времени, то через 24, 48 и 72 часа после инъекции проводили визуализацию животных, чтобы следить за рассеиванием сигнала.
ПРИМЕР 6
Визуализация in vivo с CNIR7
Биораспределение CNIR7 в тканях мыши
Анализировали биораспределение CNIR7 в тканях мыши перед визуализацией in vivo. Трем мышам внутривенно вводили CNIR7 (в дозе 10 нмоль в 100 мкл солевого буфера). После анестезии мышей умерщвляли смещением шейных позвонков в разные временные интервалы (30 минут, 240 минут, 12 часов, 24 часа, 48 часов и 72 часа) после инъекции (по три мыши в каждой точке времени). Забор образцов крови осуществляли пункцией сердца и быстро забирали ткани (сердца, почки, печени, мочевого пузыря, желудка, мозга, поджелудочной железы, тонкой и толстой кишки, легких и селезенки) для измерения близкой к инфракрасной флуоресценции с помощью флуорометра. Данные выражали как единицы флуоресценции (ЕФ) на грамм ткани [ЕФ/(г ткани)].
Визуализация in vivo мышиной модели с CNIR7
Ксенотрансплантат опухолевых клеток глиомы С6 использовали у нагих мышей для визуализации CNIR7. Мышам проводили ингаляционную анестезию 2% изофлураном в 100% кислороде с низкой скоростью потока 1 л/мин. В латеральную вену хвоста вводили 10 нмоль CNIR7 в 100 мкл буфера ФБР. Изображения трех мышей получали с помощью системы флуоресцентной визуализации мелких животных in vivo, используя оптическую систему CCD IVIS200 (Xenogen Inc.). Эта система подходит для визуализации in vivo путем биолюминесценции и флуоресценции и способна быстро сканировать мелких грызунов в единственной проекции, то есть скорость сканирования для флуоресцентной визуализации составляет всего 1 секунду. С этой системой также доступен полный набор программного обеспечения для визуализации. Для визуализации NIRF с Cy5.5 использовали набор фильтров с фильтром возбуждения (640±25 нм) и фильтром эмиссии (695-770 нм). Отображения флуоресценции получали с монохромной камерой CCD с высокой чувствительностью к красному свету, оборудованному линзой с C-креплением. Мышей умерщвляли для анализа биораспределения. Часть образцов опухолевой ткани использовали для оценки активности Bla.
ПРИМЕР 7
Флуоресцентные белки
Оценка потенциала флуоресцентных белков для IVI
Флуоресцентный белок (FP) mPlum имеет самую большую длину волны 649 нм и очень хороший стоксов сдвиг 59 нм, что означает и очень хорошую проницаемость в тканях и хорошее соотношение сигнала к шуму. Хотя указанный белок не настолько яркий, как EGFP, он обладает сходной фотостабильностью, и его показатели длины волны и стоксова сдвига должны более чем восполнить это различие во время IVI, но вместе с тем, возможно, что in vitro его поведение отличается. Второй FP с большой длиной волны (620 нм) представляет собой белок mKeima, который обладает лучшими показателями стоксова сдвига, чем mPlum, но при 180 нм возникает обеспокоенность, что на длину волны возбуждения должен накладываться фон. Вместе с тем, mKeima обладает яркостью, сходной с mPlum, и становится неясно, какой из FP будет лучше вести себя во время IVI. Другим FP с относительно большой длиной волны (610 нм) является mCherry, и его яркость в четыре раза больше, чем яркость mPlum или mKeima. Стоксов сдвиг белка mCherry составляет только 23 нм, таким образом, соотношение сигнала к шуму может оставаться проблемой, несмотря на большую яркость. Белок FP tdTomato имеет самую короткую длину волны (581 нм), но является при этом самым ярким, а именно в 20 раз более ярким, чем mPlum и mKeima.
Четыре FP, mPlum, mKeima, mCherry и tdTomato клонировали в векторы экспрессии с помощью ПЦР-клонирования Gateway. Каждую из этих конструкций трансформировали в Mtb и анализировали in vitro с использованием 96-луночных планшетов. Их анализировали в культуральной среде при стандартных условиях роста и с помощью внутриклеточных анализов роста. Все конструкции оценивали спектрофотометрически и микроскопически, используя восьмилуночное предметное стекло. В спектрофотометрических исследованиях оценивали оптимальную длину волны возбуждения, а также оптимальную длину волны эмиссии для каждой конструкции. В качестве отрицательного контроля использовали EGFP для эмиссии при больших значениях длины волны и вектор единственный использовали для оценки влияния аутофлуоресценции непосредственно от бактерий и макрофагов. Микроскопия позволяет оценить любую вариабельность мощности сигнала и устойчивость разных векторов после выращивания в культуральной среде посредством подсчета процента флуоресценции в бактериальной популяции.
Группа оценки in vitro для FP
Конструкции FP анализировали по показателям устойчивости в культуре, эффективности транскрипции и трансляции, порога обнаружения и сигнала во время/после введения изониазида. Вначале для оценки были выбраны, по меньшей мере, два трансформанта с каждой конструкцией FP, поскольку в отдельных трансформантах FP ранее наблюдали вариабельность интенсивности сигнала и устойчивости конструкций. Затем был выбран единственный оптимальный штамм для исследований in vivo каждого FP.
Устойчивость в культуре оценивали по росту каждого штамма в отсутствие выбора и определяли процент бактерий, которые остаются флуоресцентными после роста в течение 30 дней и при наличии указанного выбора и определения. Это подтверждали путем помещения растворов в чашки Петри в присутствии и в отсутствие соответствующего антибиотика, для оценки процентного содержания в культуре бактерий, несущих выбираемый из плазмиды маркер. По анализам эффективности транскрипции и трансляции становится понятно, функционирует ли промотор в каждой конструкции должным образом и влияет ли использование кодона на трасляцию с той точки зрения, что это может влиять на интенсивность сигнала. Анализы проводили путем RT-ПЦР от Mtb, несущих каждую конструкцию FP, чтобы сравнить прирост индукции, используя разные промоторы и однокопийные или многокопийные векторы для корреляции этой индукции с конструкциями, экспрессирующими другие репортеры. Эти отношения должны быть сопоставимыми независимо от экспрессируемого репортера.
Проводили измерения интенсивности флуоресценции и содержания белка и сравнивали их для каждого штамма, используя спектрофотометрические исследования и вестерн-блоттинг, соответственно. Соотношения белка к РНК к флуоресцентному сигналу должны быть сопоставимыми, независимо от экспрессируемого репортера или уровня экспрессии транскрипта РНК. При неэффективной трансляции некоторых репортеров предполагается, что их соотношение белка к транскриптам РНК будет уменьшаться с увеличением уровня экспрессии РНК. Такое наблюдение интерпретируется как необходимость коррекции использования кодонов для указанных FP в целях улучшения эффективности трансляции. Вместе с тем, также возможно, что это является результатом неустойчивости белка или секвестрации во внедренных элементах при сверхэкспрессии.
Порог обнаружения определяли с помощью оценки флуоресценции в предельных разведениях из параллельно приготовленных культур. Эти данные оценивали по КОЕ и количественному определению с помощью флуоресцентной микроскопии, для подтверждения того, что данные флуоресценции непосредственно связаны с жизнеспособностью бактерий. Влияние обработки изониазидом (INH) оценивали путем добавления 1 мкг/мл изониазида в культуры, которые уже проанализировали на КОЕ и флуоресценцию в 96-луночном планшете. Исследования КОЕ и флуоресценции проводили в режиме реального времени, используя спектрофотометр с камерой для инкубации, установленной на 37°C, путем внесения на планшет аликвотных количеств для определения КОЕ немедленно после добавления и в разные точки времени до 48 часов после добавления INH. Таким образом, обеспечивается понимание мощности сигнала, устойчивости и продолжительности сигнала после введения антибиотика в каждую конструкцию.
Устойчивость и влияние на вирулентность выбранных рекомбинантных FP
В исследованиях вирулентности все штаммы параллельно сравнивали с диким типом. Двадцать групп из четырех мышей Balb/c инфицировали аэрозольным путем в количестве от 100 до 1000 КОЕ/на легкое, как описано в примере 1. В одной группе из четырех мышей для каждого бактериального штамма (дикий тип, FP1, FP2, FP3, FP4) проводили вскрытие во всех точках времени (1, 14, 28 и 72 дня), чтобы определить КОЕ, провести гистопатологическое исследование, определить присутствие соответствующей конструкции и уровень флуоресценции в легких и селезенке. Процент популяции бактерий, которые несут конструкцию, определяли с помощью флуоресцентной микроскопии, которую проводили на титровальных планшетах, по меньшей мере, на 20 отдельных колониях по КОЕ. Уровни флуоресценции измеряли в гомогенизированных тканях для оценки общего содержания остаточного FP.
Флуоресцентные белки у мышей после аэрозольного инфицирования
Шесть групп мышей Balb/c по четыре мыши на группу инфицировали аэрозольным путем в количестве от 100 до 1000 КОЕ/на легкое каждым бактериальным штаммом, несущим конструкции mPlum, mKeima, mCherry и tdTomato и векторный скелет единственный (в общей сложности 30 групп). Бактериальные штаммы оттаивали для аэрозольного инфицирования, как описано в примере 1. Пять групп из четырех мышей, по одной группе с каждым FP и одну группу только с вектором единственным, использовали для визуализации во всех точках времени, и в каждой точке времени другие пять групп из четырех мышей умерщвляли и вскрывали для проведения гистопатологии и определения КОЕ в легких и селезенке. Через 24 часа, 7, 14, 28 и 72 дня проводили визуализацию с тем же комплектом ABSL3, используя оборудование для визуализации Xenogen IVIS200 с оптимальными фильтрами возбуждения и эмиссии для каждого FP. Если для FP необходимо использовать другой набор фильтров в устройстве IVIS, вектор единственный также подвергали визуализации в каждой группе животных, с использованием того же набора фильтров для контроля аутофлуоресценции. Таким образом подтверждали приемлемость каждого FP для IVI, а также чувствительность этой системы, поскольку бактериальная нагрузка варьировала в течение эксперимента от очень низкой (100 КОЕ/на легкое) до очень высокой (>105 КОЕ/на легкое) в более поздних точках времени после инфицирования. Использование вектора единственного контролирует и аутофлуоресценцию и потенциальную разницу вирулентности, обусловленные присутствием FP.
ПРИМЕР 8
Жук-щелкун красный (CBR) для обнаружения туберкулеза в культуральной среде
Ген CBR клонировали во все из четырех конструкций, описанных для Bla, с использованием уже внедренных сайтов рекомбинации Gateway. Эти плазмиды позволяют экспрессию из обоих промоторов L5 и hsp60. Способность каждого штамма продуцировать свечение в присутствии D-люциферина сравнивали в ростовой среде с использованием 96-луночных планшетов в многорежимном микропланшетном ридере со способностью обнаружения люминесценции и инжекторами, позволяющими измерять мгновенную эмиссию во время добавления D-люциферина, а также кинетику постоянного ослабления сигнала. Все анализы проводили по четыре раза с предельным разведением бактерий и определением КОЕ, чтобы сопоставить количество жизнеспособных бактерий с производимым сигналом. Устойчивость конструкций неселективно анализировали по их росту в течение 7 дней, с последующими спектрофотометрическим исследованием и флуоресцентной микроскопией. Эти данные сопоставляли с КОЕ, чтобы определить отношение сигнала к жизнеспособности бактерий, и с помощью микроскопии вычисляли процент бактерий, продуцирующих положительный сигнал. В этих анализах оценивали влияние конструкций на жизнеспособность бактерий по динамике роста бактерий, несущих эту конструкцию, по сравнению с бактериями, несущими только вектор единственный.
Оценка экспрессии, устойчивости и вирулентности CBR у мышей
Устойчивость рекомбинантного CBR и его влияние на вирулентность исследовали для двух штаммов с предполагаемой перспективностью для IVI. В исследованиях на вирулентность все штаммы сравнивали параллельно с диким типом. Двенадцать групп из четырех мышей Balb/c аэрозольно инфицировали в количестве от 100 до 1000 КОЕ/на легкое, как описано в примере 1.
В одной группе из четырех мышей для каждого штамма бактерий (дикий тип, CBR1 и CBR2) проводили вскрытие во всех точках времени (1, 14, 28 и 72 дня) для определения КОЕ, гистопатологического исследования, определения присутствия соответствующей конструкции и уровня люминесценции в легких и селезенке. Процент популяции бактерий, несущих конструкцию, определяли с помощью флуоресцентной микроскопии, проводимой, по меньшей мере, на 20 отдельных колониях с титровальных планшетов КОЕ. Уровни люминесценции также измеряли в гомогенизированных тканях, чтобы оценить общее содержание остаточного CBR.
Визуализация у мышей штаммов туберкулеза и BCG, экспрессирующих CBR
Шесть групп из четырех мышей Balb/c в каждой группе инфицировали аэрозольным путем каждым штаммом бактерий, несущих RLuc8, и векторным каркасом единственным в количестве от 100 до 1000 КОЕ/на легкое (в общей сложности двенадцать групп), как описано в примере 1. Две группы из четырех мышей, одна из которых с RLuc8 и другая с вектором единственным, использовали для визуализации во всех точках времени и еще две группы из четырех мышей в каждой точке времени умерщвляли и вскрывали для определения КОЕ в легких и селезенке. Визуализацию выполняли через 24 часа, 7, 14, 28 и 72 дня с тем же комплектом ABSL3, используя оборудование для визуализации Xenogen IVIS200. Перед визуализацией в хвостовую вену внутривенно вводили от 1 до 5 мкмоль D-люциферина, что, как показано, было достаточно для IVI.
Изображения получали перед инъекцией соединения и через 1, 2 и 4 часа после инъекции. Если в любой из указанных точек времени был выявлен сигнал, проводили последующую визуализацию животных через 24, 48 и 72 часа, чтобы следить за рассеиванием сигнала. Животным проводили изофлурановую анестезию путем введения 2% изофлурана в 100% кислороде, используя систему Matrix (Xenogen) в светонепроницаемой камере, и проводили визуализацию с временем интегрирования от 3 до 5 минут, биннинг 10 пикселей. Это позволяет подтвердить полезность CBR для IVI, а также чувствительность этой системы, поскольку бактериальная нагрузка варьирует в течение эксперимента от очень низкой (100 КОЕ/на легкое) до очень высокой (>105 КОЕ/на легкое) в более поздние точки времени после инфицирования. Использование только вектора единственного контролирует и аутофлуоресценцию и потенциальную разницу в вирулентности, обусловленную присутствием гена CBR.
ПРИМЕР 9
Оценка потенциала других люциферазных систем для IVI
Ген RLuc8 люциферазы клонировали в описанную микобактериальную систему экспрессии с помощью ПЦР клонирования Gateway. Конструкции вводили в Mtb и изучали ими продукцию свечения в бактериальной культуральной среде с использованием целых клеток. Если интактные бактерии продуцировали свет, сопоставимый с CBR, то можно было проводить сравнение внутриклеточной бактериальной системы с системой CBR у мышей. Обе бактериальные люциферазные системы, грамположительные и грамотрицательные, имеют преимущество в том, что они продуцируют свой собственный субстрат. Оба оперона клонировали в системы экспрессии с помощью рестрикционного расщепления, чтобы удалить их из рассматриваемого вектора, с последующими лигированием в адаптеры Gateway и рекомбинантным клонированием Gateway. Конструкции изучали в отношении светового излучения из Mtb в бактериальной среде. Все исследования на биолюминесценцию проводили в 96-луночных планшетах, как описано для системы Bla, за исключением того, что световое излучение измеряли на люминесцентной установке для спектрофотометра. Чувствительность оценивали в предельных разведениях и определение КОЕ проводили параллельно во всех образцах с тем, чтобы световое излучение можно было рассчитать по отношению к КОЕ.
Обнаружение туберкулеза в макрофагах с использованием люциферазы
Исследовали влияние секреции и нацеливания на мембраны люциферазной активности в макрофагах. Секреция из микобактерий достигалась путем присоединения аминоконцевой сигнальной последовательности ТАТ из Mtb BlaC (BlaSS) и размещением указанного слияния в ту же конструкцию, которую оптимально экспрессирует CBR в Mtb. Секрецию подтверждали анализом культуры фильтратов и целых клеток из штаммов Mtb, экспрессирующих CBR, BlaSS::CBR и вектор единственный, которые выращивали до ранней фазы логарифмического роста. В культуре фильтратов из этого штамма должна выявляться намного большая продукция света, чем из штамма, экспрессирующего CBR, и продукция света из целых клеток от BlaSS::CBR должна быть такой же или более низкой, чем у CBR Mtb. Карбоксиконцевой заякоренный GPI от CD14, используемый для Bla, присоединяли к BlaSS::CBR, чтобы получить слитый белок BlaSS::CBR::GPI.
Продукцию света Mtb, экспрессирующей BlaSS::CBR::GPI, изучали с использованием анализа внутриклеточных макрофагов, по сравнению со штаммами, экспрессирующими CBR и BlaSS::RLuc8. Макрофаги J774A.1 находились в 96-луночных планшетах, чтобы была возможность титровать бактерии и изучать разные концентрации соединений. Все анализы выполняли четыре раза тем же образом, как описано для Bla. Для оценки значения проницаемости клетки-хозяина в полученных показателях перед добавлением D-люциферина 0,1% тритоном X-100 лизировали двойные лунки. Во всех точках времени четыре лунки без обработки использовали для определения количества КОЕ, связанного с клетками. На внутриклеточное обнаружение CBR может влиять проницаемость для D-люциферина эукариотических клеток и микобактериальных вакуолей, таким образом, чрезвычайно важной является оценка его чувствительности для бактерий внутри макрофагов. Вместе с тем, внутриклеточный рост бактерий вряд ли будет значительно влиять на бактериальные люциферазные системы.
Продукцию света в каждой из бактериальных люциферазных систем и RLuc8 подтверждали с помощью внутриклеточных анализов. Двойные лунки лизировали 0,1% тритоном X-100 перед добавлением коэлентеразина для оценки значения проницаемости клетки-хозяина в полученных для RLucS показателях. Локализацию сигнала подтверждали для тех конструкций, которые показали максимальную эффективность. Эти анализы проводили сходным образом, но с использованием восьмилуночного предметного стекла. Микроскопия позволяет определять локализацию, процент бактерий с положительным сигналом и оценивать интенсивность локализованного сигнала.
ПРИМЕР 10
Обнаружение Bgal с помощью соединений для IVI
Описанный ранее ген Bgal без промотора (17) клонировали в микобактериальные векторы экспрессии посредством фермента рестрикции и лигированием на адаптеры Gateway. Эти векторы переносили в Mtb для оценки в бактериальной культуральной среде с использованием микобактериального проницаемого флуоресцентного реагента 5-ацетиламино-флуорексин-ди-бета-D-галактопиранозида (C2FDG) в 96-луночные планшеты, как описано выше [18]. Это соединение не является флуоресцентным до его расщепления посредством Bgal, с длиной волны возбуждения 460 нм и эмиссии 520 нм. Вектор, который излучает наиболее мощный флуоресцентный сигнал, использовали для конструирования дополнительных слияний, с которыми возможна секреция Bgal и локализация клеток-хозяев.
Секреция Bgal является важной, поскольку способствует определению значения проницаемости микобактерий для способности разных соединений обнаруживать Bgal. Для секреции Bgal присоединяют аминоконцевую сигнальную последовательность ТАТ от Mtb BlaC (BlaSS), и это слияние помещают в ту же конструкцию, которая оптимально экспрессирует Bgal в Mtb. Секрецию подтверждают путем анализа культуры фильтратов и целых клеток из штаммов Mtb, экспрессирующих Bgal, BlaSS::Bgal и вектор единственный, которые выращивают до фазы раннего логарифмического роста. Тот же карбоксиконцевой заякоренный GPI от CD14, используемый для Bla, присоединяют к BlaSS::Bgal, чтобы создать слитый белок BlaSS::Bgal::GPI.
Все конструкции Bgal оценивали на чувствительность обнаружения флуоресценции с помощью C2FDG, 5-додеканоиламинорезоруфин-ди-бета-D-галактопиранозида (C12RG) и 9H-(1,3-дихлор-9,9-диметилакридин-2-он-7-ил)-бета-D-галактопиранозида (DDAOG). Все соединения являются коммерчески доступными от фирмы Molecular Probes, подразделения Invitrogen. Известно, что C2FDG эффективно внедряется и обнаруживает Bgal в Mtb, указанное соединение обеспечивает положительный контроль, хотя его длина волны эмиссии не имеет преимущества для IVI. C12RG хорошо внедряется в эукариотические клетки и имеет более длинную волну эмиссии (590 нм), но сходное соединение C12FDG плохо обнаруживает Bgal в Mtb, и предполагается, что оно плохо проходит через мембрану бактерий.
Чтобы подтвердить влияние проницаемости и локализации на излучаемый сигнал, измеряли активность Bgal в интактных клетках и лизатах целых клеток для всех штаммов и соединений. Показано, что соединение DDAOG хорошо работает для IVI, поскольку хорошо проходит через мембраны эукариот и имеет наибольшую длину волны эмиссии после расщепления посредством Bgal (660 нм). Считается, что DDAOG будет наилучшим соединением для дальнейших исследований, если оно будет хорошо обнаруживать активность Bgal.
Экспрессия Bgal, устойчивость и вирулентность у мышей
Изучали устойчивость и влияние на вирулентность рекомбинантного Bgal для двух штаммов, которые показали перспективность использования для IVI. В исследованиях на вирулентность все штаммы сравнивали параллельно с диким типом. Двенадцать групп из четырех мышей Balb/c каждая инфицировали аэрозольным путем в диапазоне от 100 до 1000 КОЕ/на легкое, как описано выше. В одной группе из четырех мышей для каждого бактериального штамма (дикий тип, Bgal 1 и Bgal 2) проводили вскрытие во всех точках времени (1, 14, 28 и 72 дня) для определения КОЕ, проведения гистопатологии, определяли присутствие соответствующей конструкции и активность Bla в легких и селезенке с C2FDG. Определяли процент бактериальной популяции, несущей конструкцию, с помощью анализов Bla, проводимых, по меньшей мере, на 20 отдельных колониях с титровальных планшетов КОЕ. Уровни Bla измеряли в гомогенизированных тканях для оценки общего содержания остаточного Bgal в каждой точке времени.
Визуализация у мышей штамма микобактерии туберкулеза, экспрессирующего Bgal
Поскольку все клетки от мышей L2G85 экспрессируют Fluc из промотора ACTB, макрофаги костномозгового происхождения от мышей L2G85 инфицировали штаммом Mtb, экспрессирующим Bgal, и сравнивали с тем же штаммом, несущим вектор единственный. Инфицирование макрофагов осуществляли макрофагами костномозгового происхождения от мышей L2G85, инфицированных тем же образом, как мыши для других анализов внутриклеточного роста в макрофагах J774A.1. Двойные лунки лизировали 0,1% тритоном X-100 перед добавлением Lugal, чтобы оценить роль проницаемости клетки-хозяина в полученных данных. Во всех точках времени использовали четыре необработанные лунки, чтобы определить количество КОЕ, связанного с клетками. Локализацию сигнала подтверждали микроскопией для конструкций, показавших максимальную эффективность. Это исследование проводили сходным образом, но с использованием восьмилуночного предметного стекла. Микроскопия позволяет определить локализацию, процент бактерий с положительным сигналом и оценить интенсивность локализованного сигнала. Исследования IVI проводили на мышах согласно тем же протоколам, как описано для CBR, за исключением применения Lugal вместо люциферина для детекции.
ПРИМЕР 11
Конструкция зонда на основе кристаллической структуры моделей бета-лактамаз и других белков
Моделирование кармана фермента BlaC
Карман фермента бета-лактамазы M. tuberculosis (BIaC) моделировали с использованием малых молекул, чтобы улучшить конструкцию и специфичность зонда. Применяли высокопропускной скрининг маленьких молекул, такой как библиотеки малых молекул, для идентификации соединений, которые связывают активный щелевидный сайт BlaC, и получали его кристаллическую структуру. Образцы-кандидаты исследовали, синтезировали и тестировали in vitro.
Бета-лактамазоподобные ферменты и связывающие пенициллин белки
Два первичных бета-лактамазоподобных белка (BlaX) и два первичных связывающих пенициллин белка (PBP) в M. tuberculosis подвергали клонированию, сверхэкспрессии и очистке. Определяли Кm и константы связывания для BlaX и PBP с цефероперазоном, пенициллином и ципрофлоксацином. Кристаллическую структуру для возможных белков-кандидатов изучали и использовали для конструирования специфичных зондов с улучшенной активностью зонда.
Отношения активности структуры между ферментами Mtb и бета-лактамазой E. coli TEM-1
Изучали кристаллические структуры BlaC и TEM-1 с цефоперазоном. Моделировали образцы на основе цефероперазона, конструировали и синтезировали их. Образцы-кандидаты использовали для определения Кm для BlaC и TEM-1.
ПРИМЕР 12
Улучшение REF-чувствительности посредством новых гасителей и красителей
Предшествующие субстраты, используемые для REF-визуализации (резонансный перенос энергии флуоресценции), успешно отображали легочные инфекции туберкулеза у мышей, и эта стратегия предполагала большие перспективы. Вместе с тем, поскольку порог обнаружения в легких составляет >10000 бактерий, будет иметь преимущество улучшение обнаружимости посредством повышения REF-чувствительности зондов. В последнее время в LiCor были разработаны новые красители и гасители, которые работают в диапазоне 800 нм, и это предвещает значительное улучшение соединений по изобретению. Указанный новый краситель с наименованием IRDye800CW обладает примерно в 10 раз большей яркостью, чем Cy5.5, и в силу большого значения длины волны должен проникать через ткани млекопитающих намного лучше, чем Cy5.5. Были разработаны соединения на основе этого красителя и соответствующий гаситель под наименованием QC-1. Соединение на основе указанного красителя и гаситель позволяют значительно улучшить существующую REF-систему. Также исследовали два красителя IRDye800, IRDye800RS и IRDye800CW (фиг. 22), в качестве донора FRET для применения визуализации in vivo. Оба из них имеют одинаковые спектры флуоресценции при длине волны возбуждения в 780 нм и эмиссии в 820 нм, но они отличаются по содержанию сульфонатных групп, которых в IRDye800CW больше, чем в IRDye800RS. Это различие может быть результатом разного биораспределения in vivo, и в этой связи исследовали оба указанных красителя. В качестве реципиента FRET в анализе флуорогенности используется IRDye QC-1, который представляет собой соответствующий краситель с высокой гасящей эффективностью для IRDye800 (фиг. 22). Включение в зонд указанных молекул аналогично процедуре синтеза, применяемой для изготовления CNIR5, с той же химической реакцией соединения между сложным эфиром NHS и амином, как описано выше. Вначале кинетика гидролиза указанных зондов CNIR, образованных из красителя IRDye800, отличается наличием и TEM-1 Bla и Mtb BlaC, и образцы визуализируют на наличие Mtb in vivo при подкожном и легочном инфицировании.
Соединения на основе IRDye800CW вначале исследуют in vitro, с последующими внутриклеточными анализами и работой на животных моделях, с целью подтверждения при подкожном и легочном инфицировании. Визуализируют флуоресцентные включения на участке инфекции, используя систему визуализации IVIS на уровне целого организма животных, и подтверждают на клеточном уровне в гомогенатах тканей во флуорометре, с использованием тканевых срезов и флуоресцентной конфокальной микроскопии, и путем прижизненной микроскопии инфицированных тканей. Комбинацию этих способов применяют ко всем зондам, которые исследуют на мышиной модели инфекции, что позволяет подробно характеризовать параметры мечения инфицированных тканей и параметры внедрения зонда в инфицированные клетки-хозяева.
Улучшение SREL и REF-чувствительности посредством структурной модификации зондов
Существующие образцы способны обнаруживать и визуализировать активность Mtb BlaC, но вместе с тем их действие для Mtb BlaC не является оптимальным. Зонд, улучшающий кинетику фермента с Mtb BlaC, будет обеспечивать повышенную чувствительность как для обнаружения, так и для визуализации. Кристаллическая структура Mtb BlaC показывает основное отличие от другого класса А бета-лактамаз, которое состоит в том, что Mtb BlaC имеет больший активный участок кармана. Это структурное различие предлагает возможность конструирования зонда с улучшенной кинетикой для Mtb BlaC. Три основных подхода используют для улучшения структуры модификации зондов BlaC, на основе цефоперазона, скрининга ограниченной библиотеки соединений и модификации уходящих групп. Идентифицированные соответствующие соединения дополнительно изучают в анализах in vitro с Mtb внутриклеточными бактериями и инфицированием мышей подкожным и легочным путями.
Рациональный подход, основанный на структуре цефоперазона.
Кинетика CNIR5 посредством TEM-1 Bla и Mtb BlaC:
для TEM-1 Bla, kcat=0,33 s-1, KM=1,9 мкмоль, kcat/KM=1,74×105 s-1M-1;
для Mtb BIaC, kcat=0,07 s-1, KM=5,9 мкмоль, kcat/KM=1,2×104 s-1М-1.
Эти кинетические данные указывают, что CNIR5 является предпочтительным субстратом для TEM-1 Bla, но не для Mtb BlaC. Чтобы идентифицировать структурные элементы, необходимые для специфической активности для Mtb BlaC, кинетику ряда антибиотиков группы лактама цефалоспорина (цефоперазон, цефалотин, цефазолин, цефтазидим, цефокситин, цефамандол, цефотаксим и цефалексин) измеряли с TEM-1 Bla и Mtb BlaC. Результаты показали, что цефоперазон (фиг. 23) является предпочтительным субстратом для Mtb BlaC по сравнению с TEM-1 Bla.
для TEM-1 Bla, k cat=0,26 s-1, KM=262 мкмоль, kcat/KM=1×103 s-1M-1;
для Mtb BlaC, kcat=2,01 s-1, KM=76 мкмоль, kcat/KM=2,6×104 s-1М-1.
Его значение kcat/KM для Mtb BlaC (2,6×104 s-1М-1) лучше, чем это значение у CNIR5 (1,2×104 s-1M-1), но его значение kcat/KM для TEM-1 Bla (1×103 s-1M-1) в 100 раз меньше, чем значение у CNIR5 (1,74×105 s-1М-1). Существует предположение, что основная структурная группа, отвечающая за селективность, происходит из большой группы, связанной с 7 амином в цефоперазоне, которое подтверждается обнаружением на рентгеновской структуре Mtb BlaC-BlaC наличия большого субстрата связывающего кармана на 7 участке.
Таким образом, группу в 7 положении цефоперазона включают в CNIR5 с целью создания зонда Mtb BlaC, который должен проявлять улучшенную кинетику фермента (фиг. 23). Указанный зонд анализировали in vitro, во-первых, 1) на его устойчивость в буферах и в мышиной сыворотке, 2) на его кинетику в присутствии очищенного Mtb и внутриклеточного Mtb, и 3) на его кинетику в присутствии очищенного TEM-1 Bla. Затем параметры его проходимости через мембраны сравнивали с CNIR5, чтобы оценить, проявляет ли он сопоставимые или улучшенные показатели мембранного транспорта и удерживания к показателям предшествующих зондов по изобретению. Затем проводили исследования на животных путем подкожного и аэрозольного инфицирования с последующей визуализацией Mtb.
Чтобы лучше понять взаимоотношения структуры и активности (SAR) в зонде цефоперазон-CNIR, проводили компьютерное моделирование его связывания с BlaC. Параллельно зонд кристаллизовали вместе с BlaC, чтобы решить вопрос сложной структуры. Полученную информацию о структуре применяли для рационального конструирования улучшенного зонда.
Быстрый анализ ограниченной библиотеки структур для идентификации зондов с улучшенной чувствительностью
После синтезирования и тестирования зонда цефоперазон-CNIR применили подход с библиотекой для улучшения селективности параллельно с усовершенствованием SAR с помощью рентгеноструктурного анализа.
Поскольку зонды Bluco намного легче в изготовлении, чем зонды CNIR, использовали субстраты на основе Bluco, чтобы обеспечить простое и быстрое считывание для кинетики фермента. Bluco использовали в качестве матрицы для конструирования малой направленной библиотеки аналогов цефоперазона. Чтобы построить такую библиотеку и создать многообразие, использовали 8-замещенный пиперазин-2,3-дионы (A) и 6-замещенные фенилглицилметилэфиры (B), все из которых коммерчески доступны. В результате получали 48 элементов.
Затем библиотека реагировала с предшественником Bluco (C) для создания конечных 48 аналогов Bluco. Библиотеку изготовляли на твердой подложке посредством карбоксилатной группы на D-люциферине. Перед включением всех указанных соединений в создаваемую библиотеку по изобретению проводили компьютерное исследование модели каждого элемента на основе доступной рентгеновской структуры BlaC, чтобы подтвердить потенциальное соответствие всех элементов активному участку кармана BlaC.
Скрининг библиотеки проводили в анализах с высокой пропускной способностью с помощью микропланшетного ридера люминесценции. Перед скринингом кинетики на первом этапе проводили скрининг устойчивости соединений в буферах. Кинетику оценивали путем сравнения уровня люминесценции оригинального субстрата Bluco по изобретению и люциферина в качестве положительного контроля в ранних точках времени при совместной инкубации. Соединения с полезной кинетикой проявляли быстрый гидролиз и высвобождение люциферина, что приводило к высокому уровню люминесценции через несколько минут после добавления субстрата; при этом оригинальные молекулы Bluco согласно изобретению показывали максимальные уровни люминесценции через несколько часов совместной инкубации. Эти исследования обеспечивают новые соединения, которые можно использовать для оптической визуализации в качестве основы для субстратов CNIR и Bluco, проявляющих улучшенную кинетику с BlaC и более высокую чувствительность.
Модифицированные уходящие группы для улучшенной кинетики
Аллильная связь в 3'-положении
Ранее было показано, что вставка двойной связи между фенольным эфиром значительно увеличивает кинетику высвобождения фенольной группы [JACS, 2003, 125, 11146-11147]. Например, kcat был увеличен в 5 раз до 54 s -1 для фенольной уходящей группы. На основе этого наблюдения была вставлена двойная связь в зонд CNIR. Например, для структуры, показанной на фиг. 22, соответствующий зонд показан на фиг. 25. В предыдущих примерах двойная связь имела цисконфигурацию, но в этом случае предполагается трансконфигурация по причине намного более крупной аллильной группы. Аналогично, вставка двойной связи в Bluco приводит к образованию Bluco2, у которого предполагается более хорошая кинетика, чем у Bluco (фиг. 24).
Карбаматная связь в 3'-положении
Второй тип связи в 3'-положении предлагает более быструю фрагментацию после гидролиза и, таким образом, улучшение чувствительности. В этой конструкции используется карбаматная связь и аминоаналог D-люциферина, амино-D-люциферин. Карбаматная связь широко использовалась в разработке пролекарства в качестве превосходной уходящей группы. Расщепление Bla высвобождает карбамат, который впоследствии разлагается на углекислый газ и свободный амино-D-люциферин (фиг. 26), субстрат для люциферазы. Аналогично указанную связь применяли также в зонде CNIR (фиг. 26).
Улучшение SREL и REF-чувствительности посредством оценки распределения в тканях
Исследования тканевого распределения проводили с использованием флуоресценции субстратов CNIR, чтобы определить имеющиеся концентрации. Поскольку расщепление увеличивает флуоресценцию, определяли распределение нерасщепленного субстрата посредством инкубации в присутствии BlaC и измерением флуоресценции, и по анализу прямой флуоресценции определяли концентрации субстрата. Указанный способ определяет присутствие субстрата в тканях приблизительно и не является точным, поскольку образцам ткани свойственна аутофлуоресценция, но вместе с тем присутствие потенциальных ингибиторов и спонтанный гидролиз субстрата могут влиять на полученные данные. Более подробные данные распределения ткани получают с помощью анализа распределения зонда с радиоактивной меткой. CNIR5 метили радиоактивным йодом, например, I125, таким образом, можно легко отслеживать распределение зонда in vivo. Ароматические группы в CNIR5 подвергают йодированию сходным образом с использованием протокола мечения тирозина в белках. Меченый зонд вводят мышам и проводят динамическую визуализацию ОФЭКТ. В разные интервалы времени мышей умерщвляют для забора органов с целью подсчета радиоактивности. Параллельно проводят непосредственный анализ свободной фракции зонда с помощью ВЭЖХ с использованием растворимых фракций, полученных после вскрытия. Гомогенаты тканей (общие и растворимые) анализировали с помощью флуоресценции, используя холодный зонд, и растворимые гомогенаты анализировали ВЭЖХ с последующим выявлением сцинтилляции фракций, с горячим зондом. Тот же эксперимент будет проведен с новыми Mtb-специфичными зондами после их разработки и подтверждения того, что обеспечено понимание их потенциала для улучшения распределения в тканях.
Улучшение чувствительности SREL и REF посредством использования бета-галактозидазы
Поскольку возможно наличие у различных ферментных систем SREL/REF значительных преимуществ перед BlaC по причине лучшей кинетики ферментов или доступности субстратов, использовали бета-галактозидазу (lacZ) с флуоресцентным (DDAOG) или люминесцентным субстратом (Lugal) для SREL/REF с Mtb. Для визуализации in vitro успешно использовались и DDAOG и Lugal, и для визуализации подкожной инфекции у мышей применяли Lugal. Хотя DDAOG показал многообещающие результаты in vitro, авторы не проводили его анализов in vivo. Для авторов будет важно определить, является ли DDAOG таким же чувствительным, как Lugal, in vivo, поскольку использование флуоресцентного субстрата будет иметь некоторое преимущество перед люминесцентным субстратом, для которого необходимо доставка люциферазы вместе с субстратом. Подобные вопросы возникают для указанной системы в отношении субстратов для Bluco. DDAOG или модифицированные соединения, которые улучшены на основе DDAOG, могут, в конечном счете, оказаться одной из наиболее чувствительных систем, и существует ряд колориметрических репортерных систем, уже применяемых многочисленными исследователями, которые могут сделать эту систему непосредственно ценной в обществе с проблемой туберкулеза и которые должны успешно применяться для визуализации инфекции туберкулеза у живых животных, больных туберкулезом.
ПРИМЕР 13
Улучшение специфичности SREL- и REF-зондов с использованием больших лактамов
Применяли стратегию, подобную стратегии для разработки зондов с улучшенной чувствительностью, чтобы разработать зонды, селективные для Mtb BlaC по бета-лактамазам, присутствующим в других видах бактерий. Лучшими характеристиками указанных ферментов бета-лактамаз обладают E. coli TEM-1, которые используются для ряда анализов кинетики и в качестве полезного репортера в эукариотических системах. Различие в используемом подходе по сравнению с подходом к улучшению чувствительности, в первую очередь, является фокус на соединения, обладающие наибольшей разницей в кинетике между Mtb BlaC и E. coli TEM-1. Хотя кинетика большинства бета-лактамов лучше, чем кинетика фермента TEM-1, были идентифицированы три бета-лактама, которые проявляют более хорошую кинетику с Mtb BlaC, чем с TEM-1. Такими бета-лактамами являются цефоперазон, цефотаксим и цефокситин. Эти соединения значительно отличаются по своей кинетике, но цефоперазон от 10 до 100 раз быстрее по кинетике с ферментом Mtb, чем с ферментом TEM-1, и предполагается, что он является хорошим кандидатом для разработки зондов, специфичных для этого фермента. Соединение CNIR создано на основе цефоперазона, и его специфичность анализировали с помощью определения кинетических параметров его фермента с использованием очищенного BlaC и TEM-1 в 96-луночном планшете с флуоресценцией для считывания результатов.
Улучшение специфичности зондов SREL и REF с использованием ограниченной структурной библиотеки
Для улучшения специфичности зондов SREL и REF согласно изобретению можно также использовать библиотеку соединений, разработанных в примере 12, но с применением модифицированного высокопропускного скрининга, сфокусированного на специфичности, а не на кинетике фермента. В основном каждое соединение синтезируют как субстрат на основе Bluco, как описано выше, и соединения анализируют в присутствии очищенного BlaC и TEM-1 в высокопропускном анализе люминесцентности согласно изобретению. Проводили скрининг всех соединений с BlaC, чтобы идентифицировать наилучшие, и с TEM-1 Bla, чтобы идентифицировать соединения, которые являются плохими субстратами для других ферментов. Дополнительно проводили предварительный скрининг всех соединений на устойчивость при 37°C в воде, чтобы гарантировать использование в дальнейших исследованиях только устойчивых соединений. Анализы проводили параллельно и все результаты выражали как отношение люминесценции BlaC к TEM-1. Вначале авторы установили свой порог для молекул, которые через 30 минут реакции показали более чем 10-кратное ускорение кинетики с ферментом BlaC. Была создана компьютерная модель каждого соединения к кристаллической структуре ферментов BlaC и TEM-1, чтобы установить устойчивые соотношения активности и структуры (SAR). Предположение о возможности применения полученных результатов к субстратам CNIR, используемым для REF, было сначала подтверждено путем сравнения активности зондов цефоперазона, которые созданы на основе CNIR и Bluco. Согласно полученным данным лактамы с идентифицированной высокой специфичностью в дальнейшем использовали при создании зондов REF и анализировали их способность к обнаружению целых клеток Mtb in vitro, при выращивании их внутриклеточно в макрофагах и во время инфекции у мышей после подкожной и аэрозольной инокуляции.
ПРИМЕР 14
Оценка CBR для визуализации у живых мышей
Начальные исследования изобретения показали, что люцифераза жука-щелкуна красного (CBR) очень хорошо действует как репортер для Mtb in vitro и в клетках культуры ткани. Было обнаружено, что CBR сопоставим с люциферазой светлячка (FFlux) в плане продуцируемого сигнала и порога обнаружения in vitro. Вместе с тем, при подкожном и легочном инфицировании мышей порог обнаружения для CBR был значительно лучше, чем для FFlux. Это предварительное наблюдение могло быть обусловлено различиями в материале инокуляции, влиянием на метаболизм бактерий in vivo или кинетикой люминесценции. Исследовали каждый из этих параметров путем тщательного анализа полезности CBR в качестве репортера жизнеспособности Mtb при легочном и подкожном инфицировании. Кинетику люминесценции оценивали и сравнивали непосредственно с FFlux на тех же животных, используя подкожную инокуляцию на разных участках и в комбинации, используя спектральное разделение биолюминесцентного сигнала, чтобы показать ответственный репортер. Легочные инфекции анализировали отдельно в парах мышей, инфицированных параллельно с сопоставимым количеством бактерий. Достигнуто понимание сути потенциальной чувствительности CBR при гипоксических поражениях путем исследования влияния на интенсивность сигнала in vitro при состояниях низкой оксигенации. Изучали другие состояния стресса, которые можно встретить in vivo, такие как низкий уровень pH и присутствие ROS и RNS.
Анализ визуализации CBR для терапевтической оценки
Поскольку сигнал CBR люциферазы зависит от присутствия АТФ, система визуализации по изобретению предлагает уникальную возможность быстро оценивать влияние лечения на жизнеспособность бактерий. Некоторые из основных вопросов относительно этой системы заключаются в скорости получения измеримого различия в сигнале и насколько точно ее можно использовать для определения минимальной ингибирующей дозы MIC. Определение MIC проводят для Mtb, используя этот анализ для изониазида и рифампицина. Полученные экспериментально данные MIC сравнивали с полученными данными в анализах OD и КОЕ. Кинетику потери сигнала оценивали в присутствии антибиотика в количестве 0,5x, 1x и 5x MIC с использованием анализа целых Mtb и внутриклеточных Mtb в макрофагах. После определения кинетики in vitro и сравнения с разницей в жизнеспособности КОЕ исследовали способность роста бактерии после лечения и определяли наличие высокой корреляции между КОЕ и люминесценцией. После определения корреляции между КОЕ и люминесценцией для выращенных in vitro бактерий исследовали кинетику влияния на люминесценцию при лечении подкожной и легочных инфекций у мышей. Применяли оба пути инокуляции, поскольку предполагались различия в доступности бактерий в легочной среде и в подкожной среде, что также делало вероятным различия в кинетике потери сигнала. Эти исследования обеспечивают понимание полезности CBR для быстрой оценки лечения у мышей. Эксперименты нацелены на острую фазу инфекции и позволяют быстро получать результаты, но существует необходимость проведения последующих экспериментов во время фазы хронической инфекции у мышей, с целью установления полезности указанной системы также для оценки лекарственных препаратов, когда возможно отсутствие репликации бактерий с высокой скоростью.
Разработка двойной системы оптической визуализации CBR-REF
Система CBR имеет преимущества, поскольку она должна позволить быстро получать данные о жизнеспособности бактерий, но в некоторых случаях этот тип системы, возможно, не является оптимальным. В ситуациях, когда скорость метаболизма бактерий недостаточна для максимальной продукции света, системы на основе люциферазы, возможно, не являются настолько чувствительными, как при оптимальных состояниях метаболизма. Влияние лекарственных препаратов оценивали с использованием CBR, проводили количественное определение бактерий в разных тканях и применяли REF для определения их локализации в клетке. Для понимания потенциальной полезности этих двух систем для оценки количества бактерий в разной окружающей среде исследовали кинетику потери сигналов и CBR и REF у мышей после инфицирования легочным и подкожным путем. Люминесценцию немедленно уменьшали после доставки антибиотика, и для выявления потери сигнала REF требовалось целых 24 часа. Разница в чувствительности CBR и REF к метаболической активности обеспечивает возможность оценки количества бактерий в реальном времени в сочетании с состоянием метаболизма. Эта система имеет важное значение для разработки, поскольку остается неясным состояние метаболизма всех бактерий во время инфекции Mtb у животных. Эта система визуализации обеспечивает, в первую очередь, способ прямого наблюдения перехода к разным средам у живых животных, по присутствию или отсутствию каждого сигнала в режиме реального времени. Предполагается, что эта способность имеет особую важность для оценки лекарственных препаратов, потому что препараты могут иметь бактерицидное действие в некоторых окружающих средах и не иметь такого действия в других средах, что является важнейшим соображением для продолжения доклинических исследований.
В настоящем изобретении цитированы следующие ссылки:

Любые патенты или публикации, упомянутые в настоящем описании, являются показательными для уровней специалистов в области техники настоящего изобретения. Эти патенты и публикации включены в изобретение в тех же пределах, как если бы каждая отдельная публикация была конкретно и отдельно включена в него путем ссылки.
Специалист в данной области техники сможет легко оценить, что настоящее изобретение хорошо адаптировано для осуществления задач и получения упомянутых результатов и преимуществ, а также свойственных ему результатов и преимуществ. Специалистам в данной области техники будет очевидно, что при осуществлении настоящего изобретения можно делать различные модификации и изменения, не отступая от сущности или объема изобретения. Специалисты в данной области техники будут осуществлять изменения в изобретении и другие применения, которые охватываются сущностью изобретения, как определено объемом формулы изобретения.
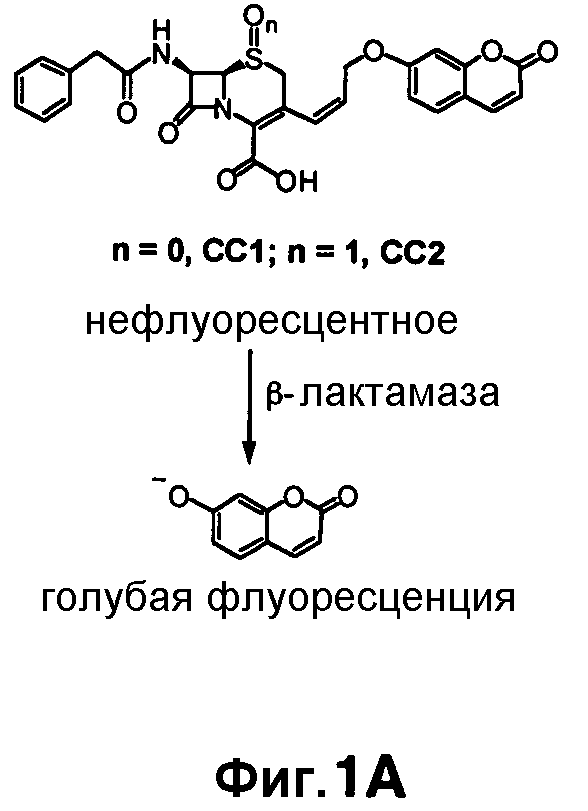
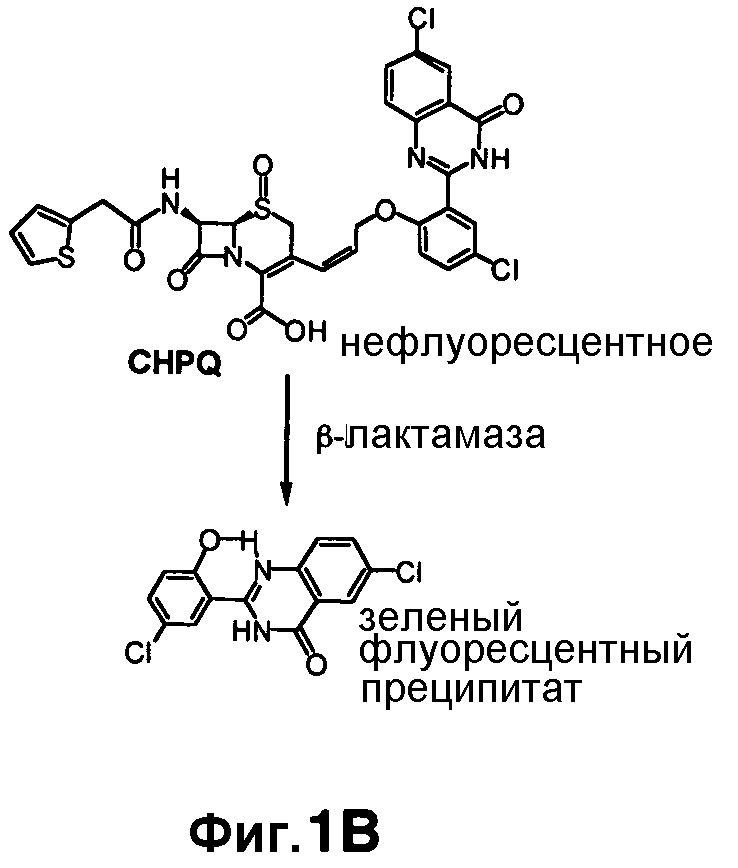
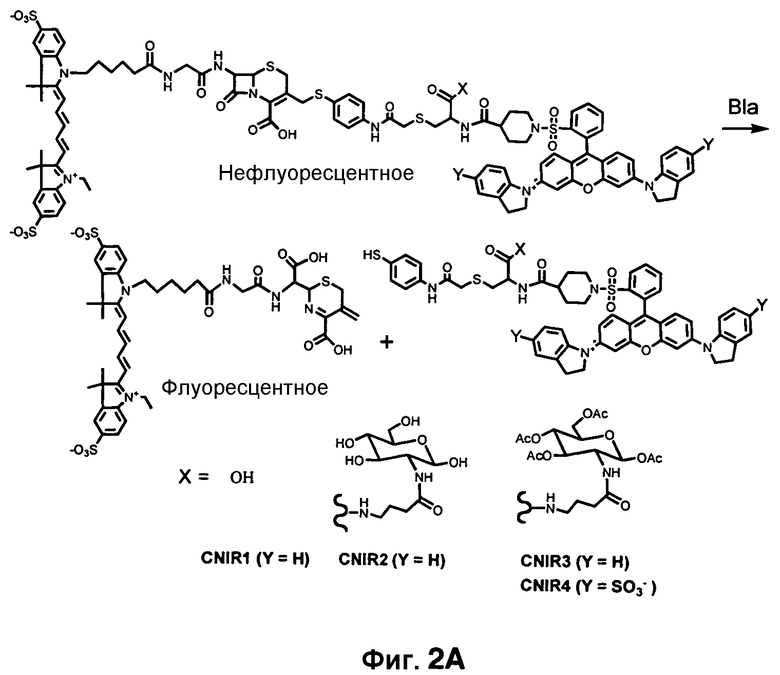
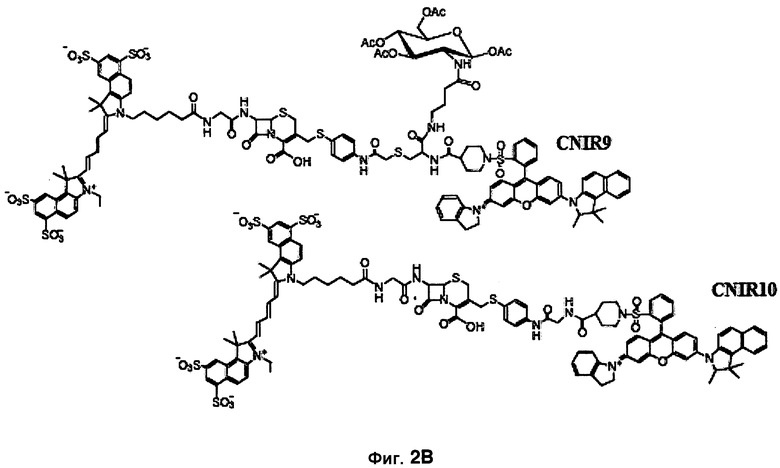
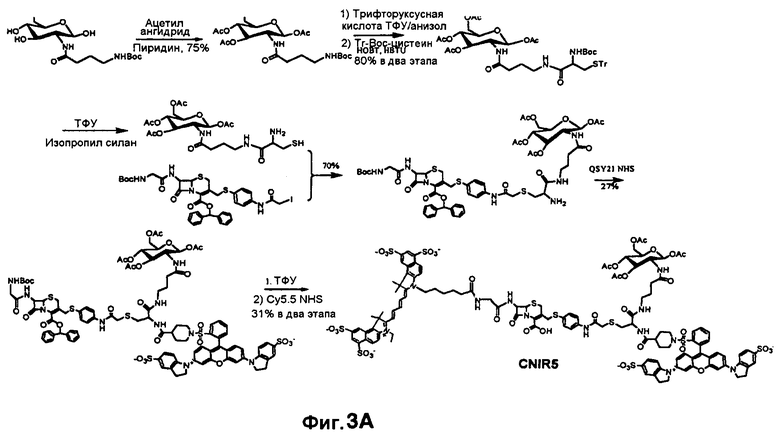
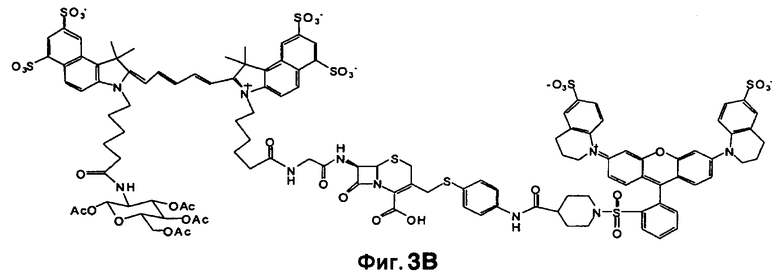
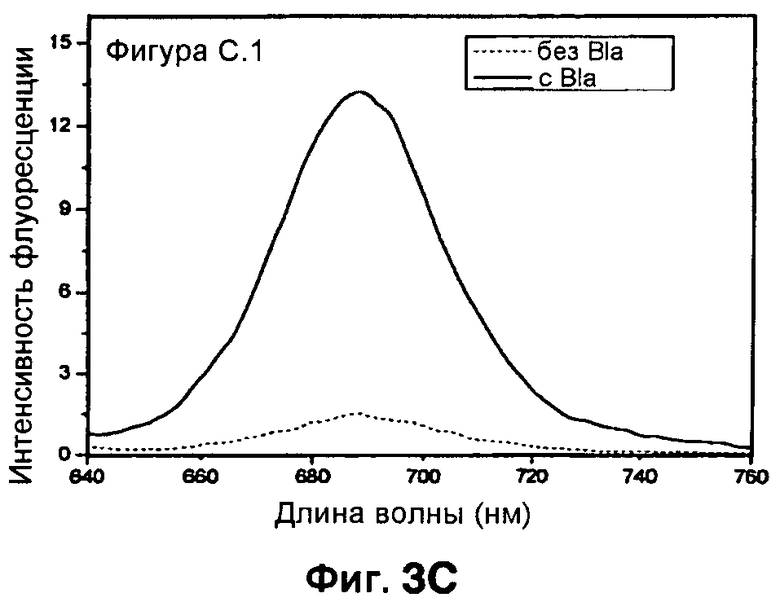
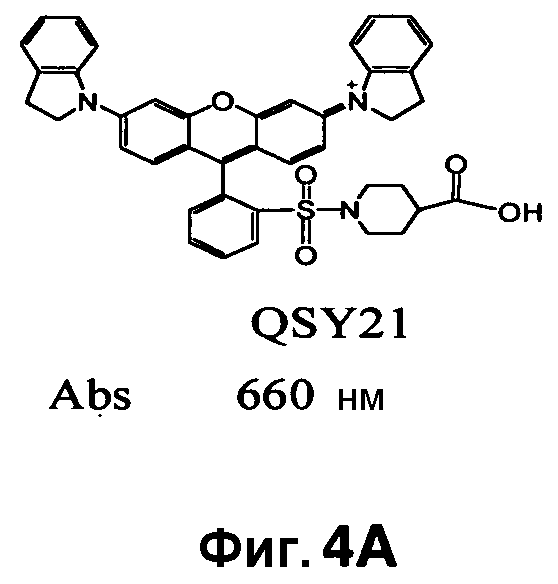
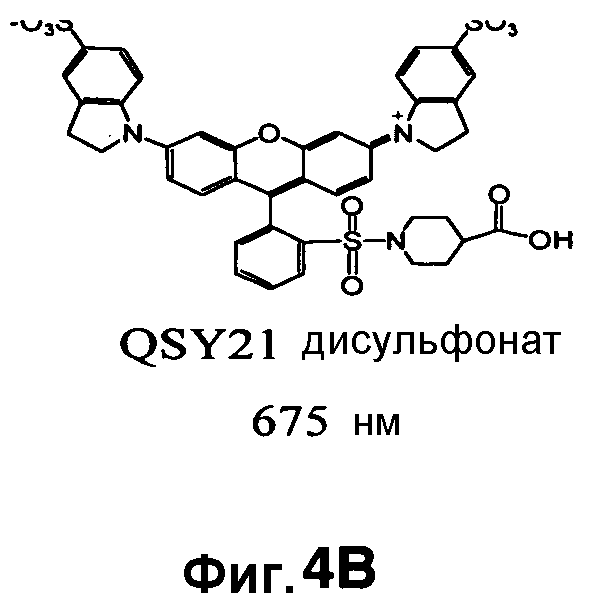
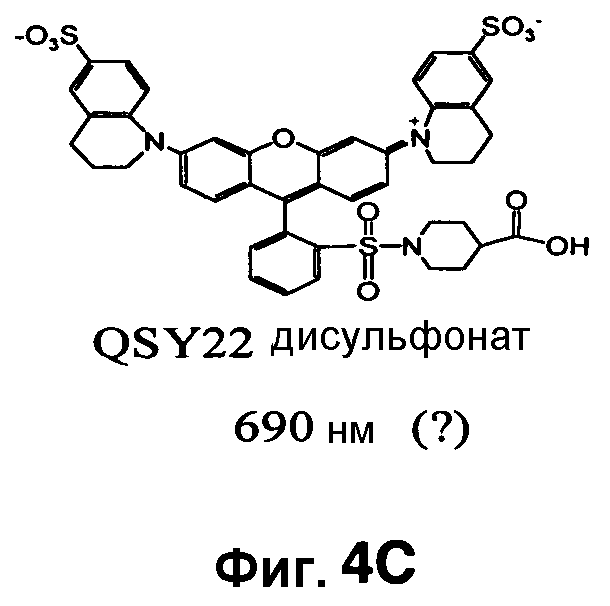
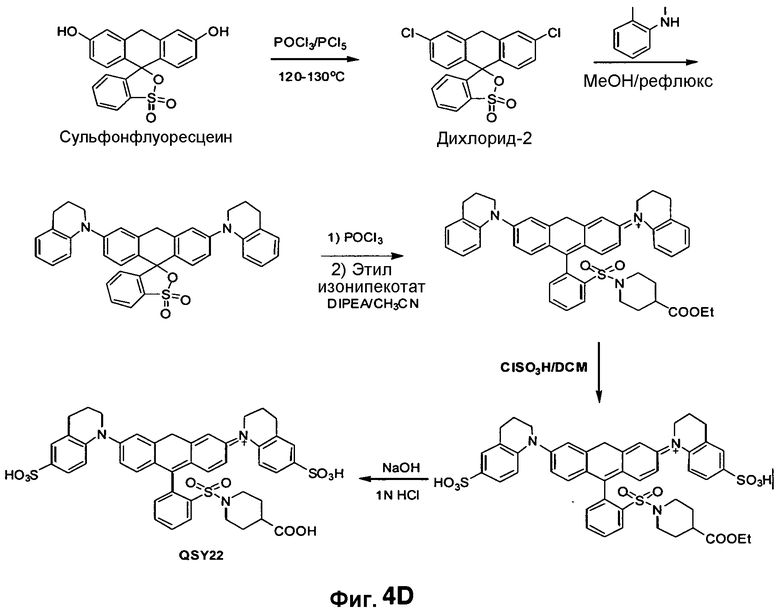
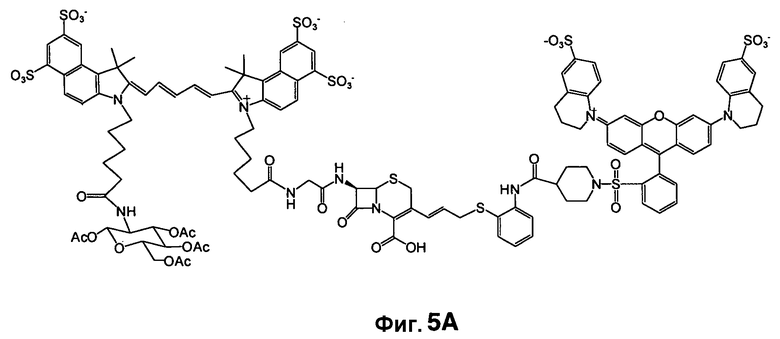
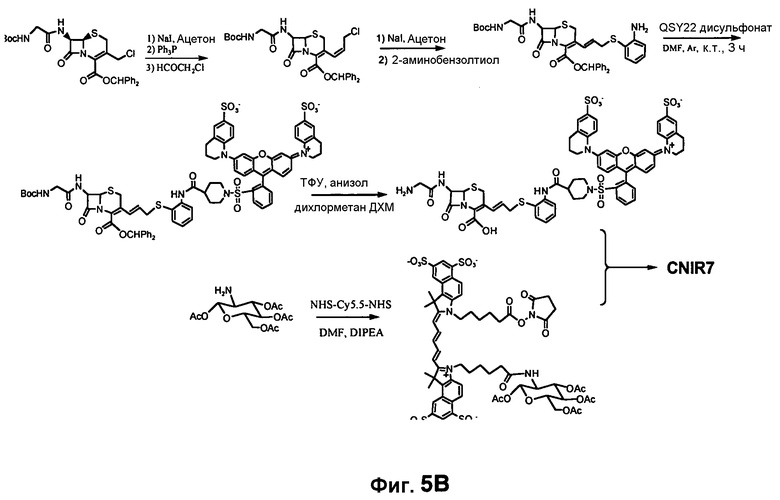
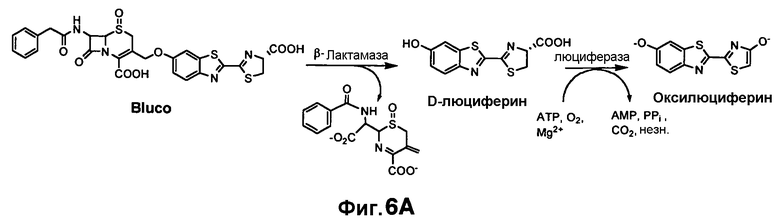
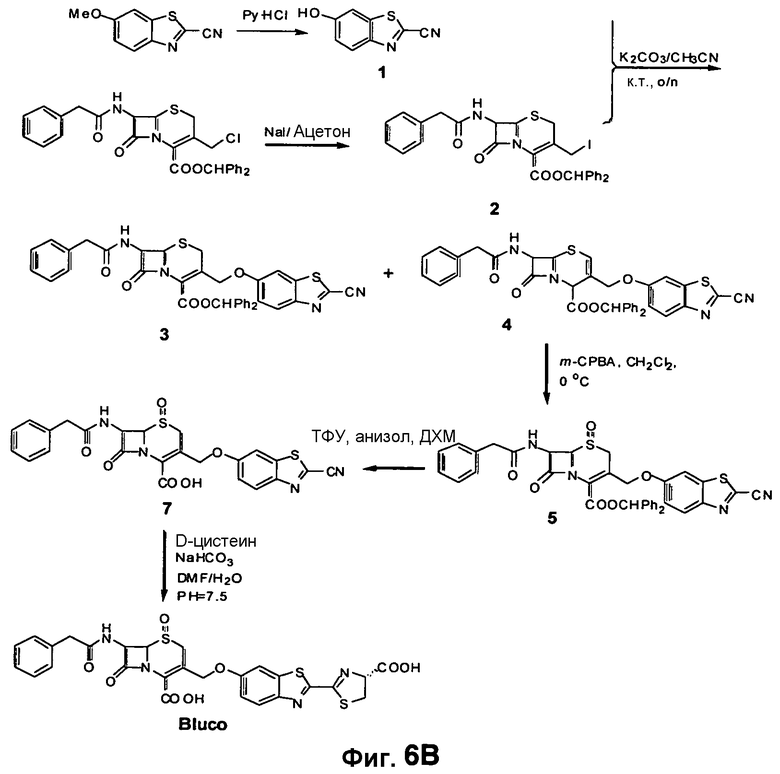
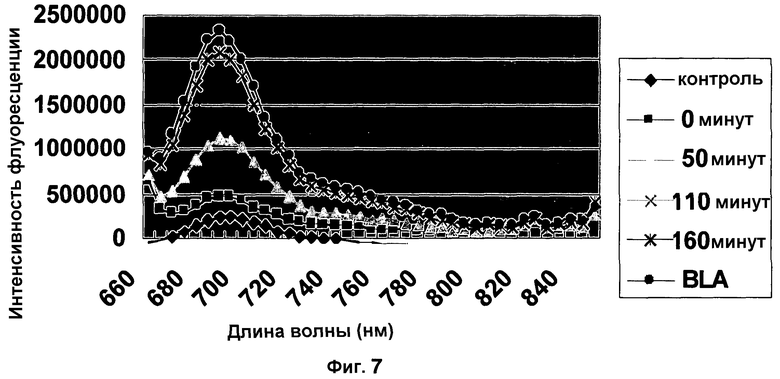
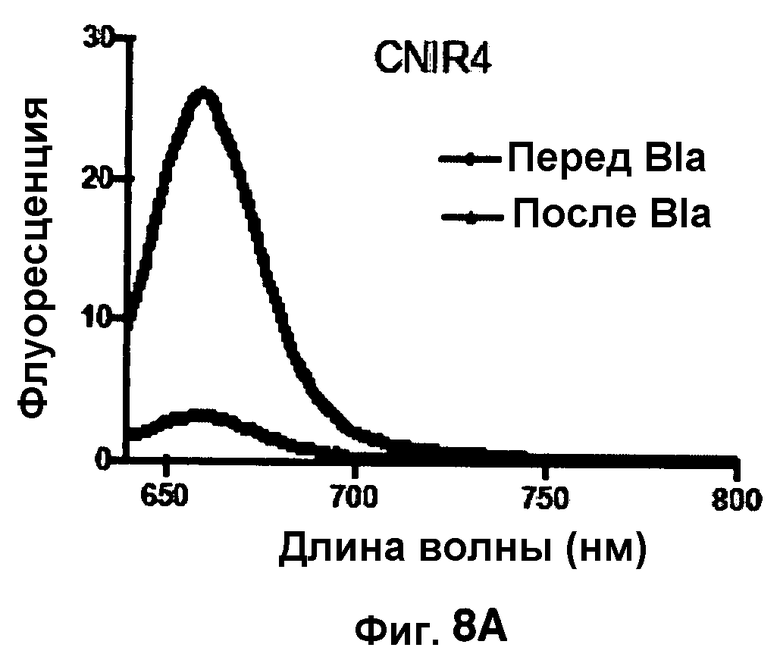
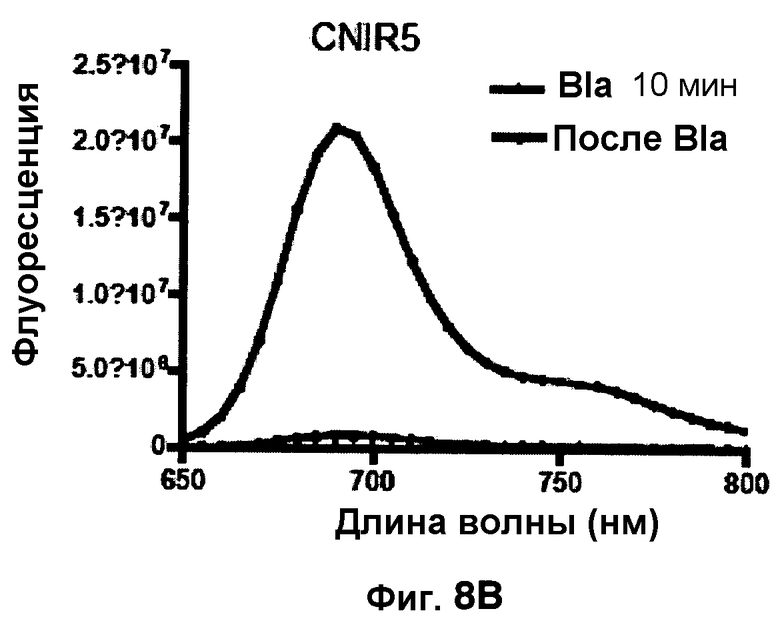
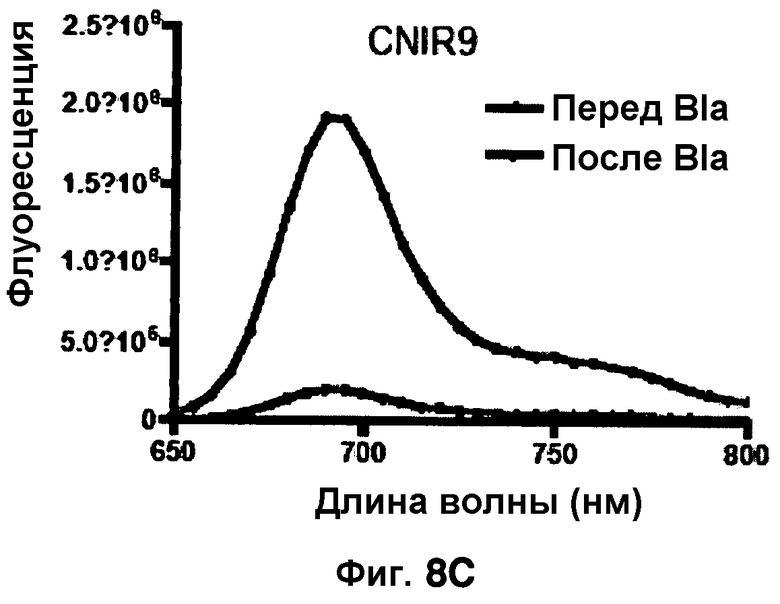
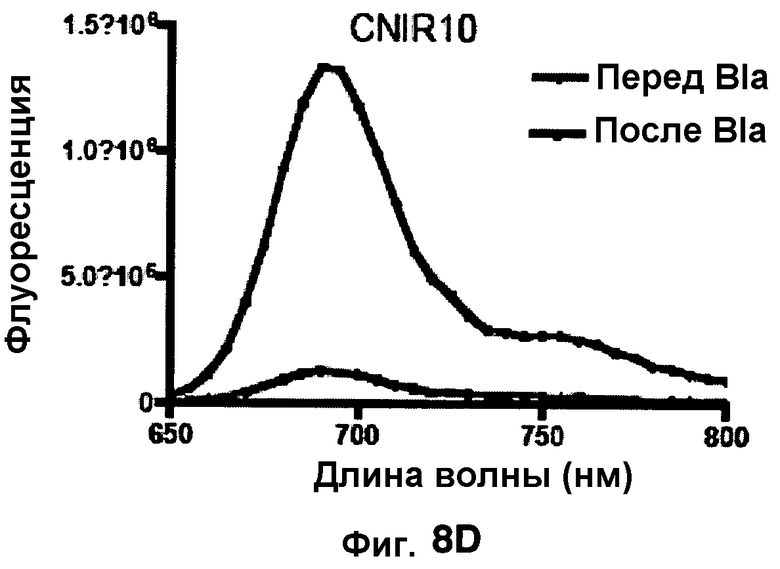
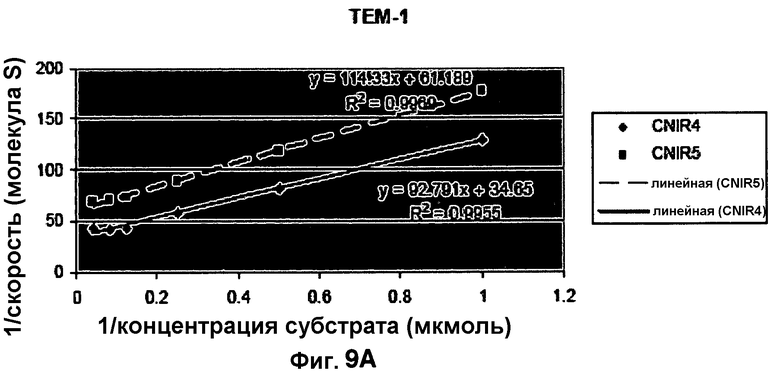
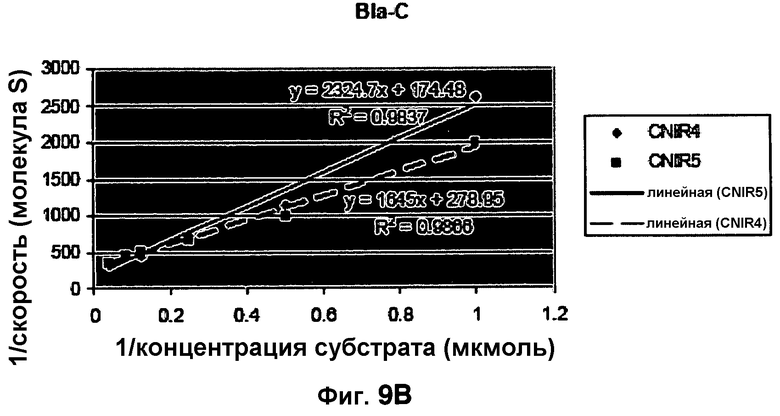
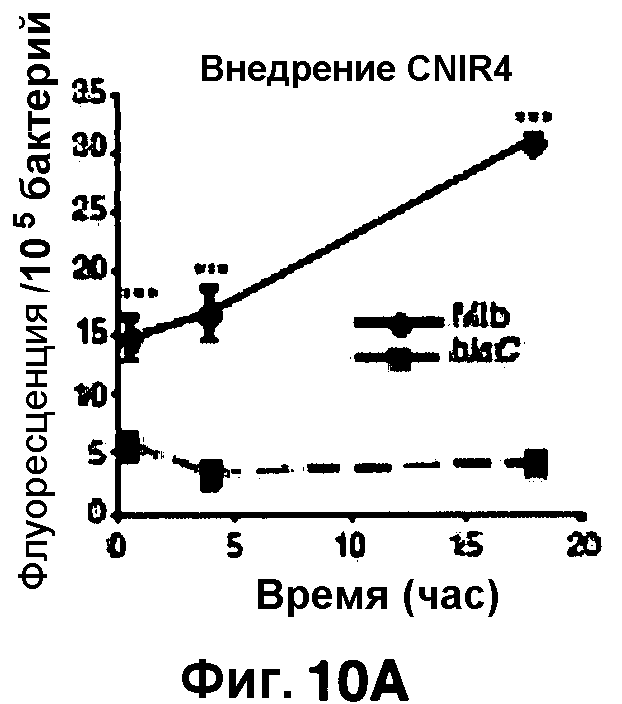
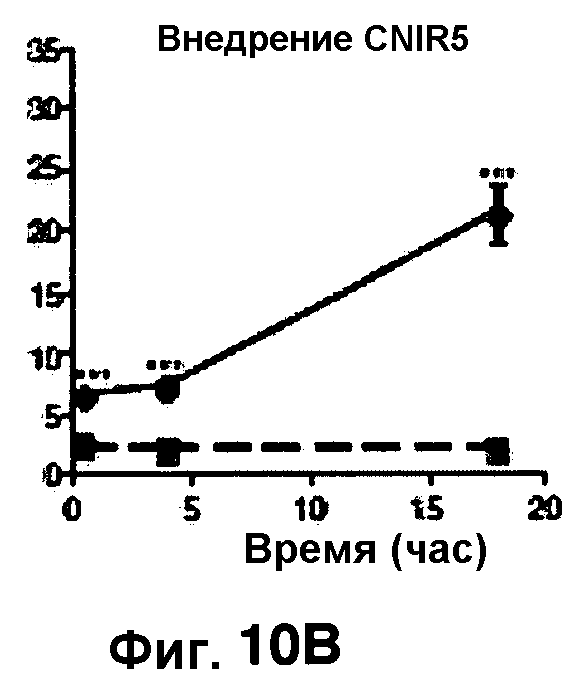
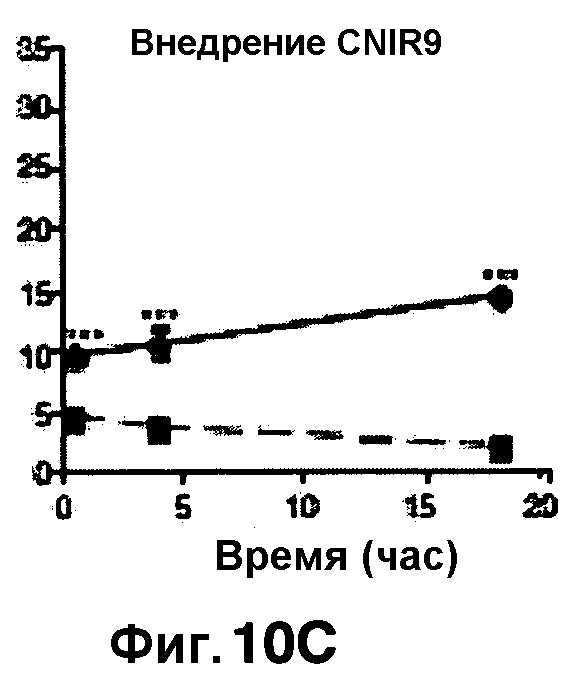
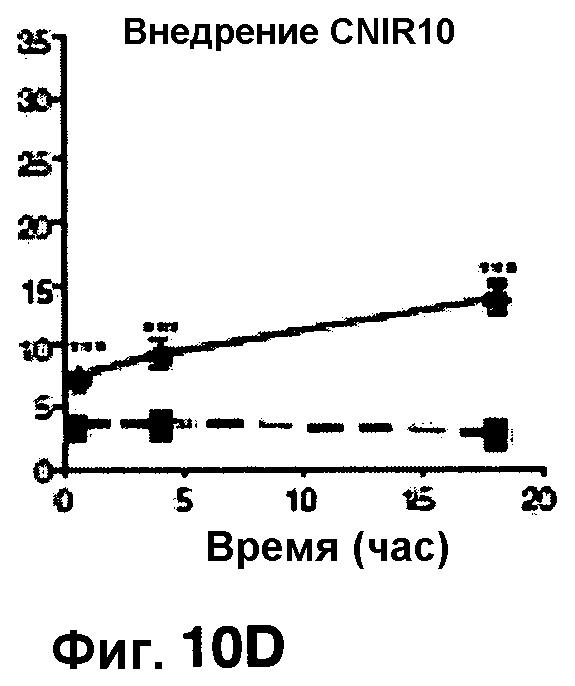
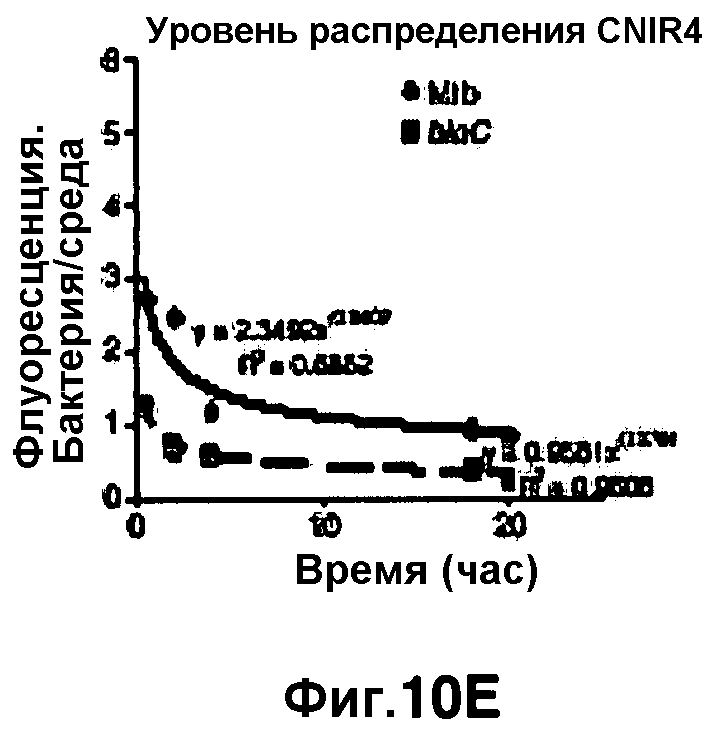
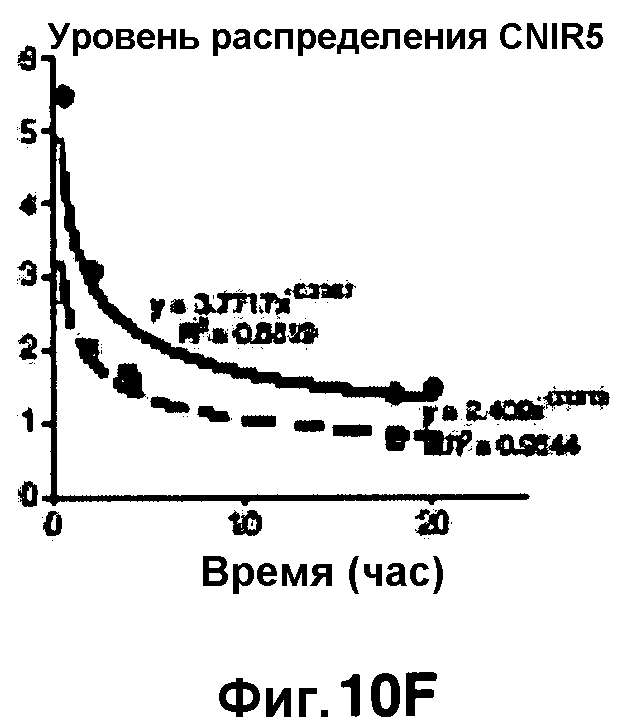
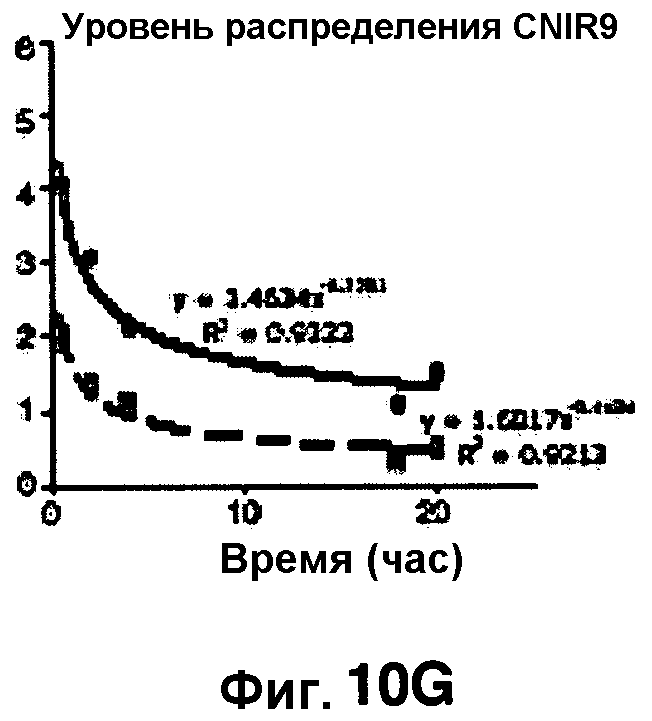
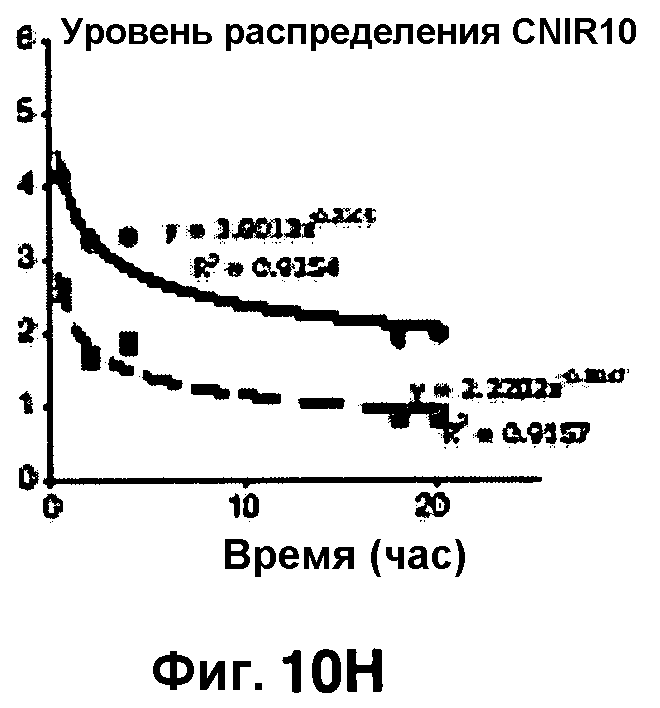
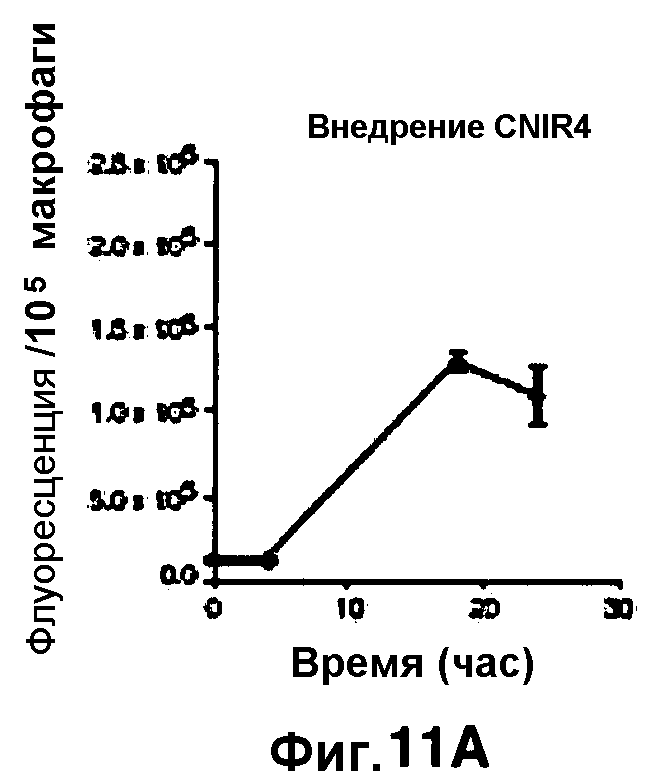
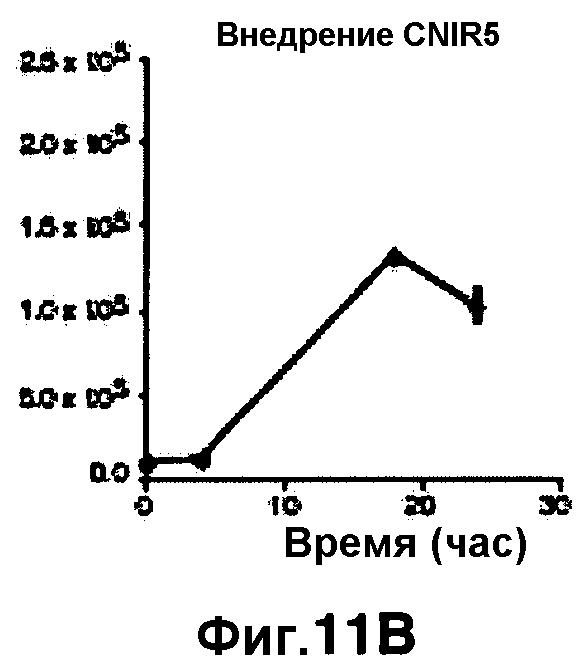
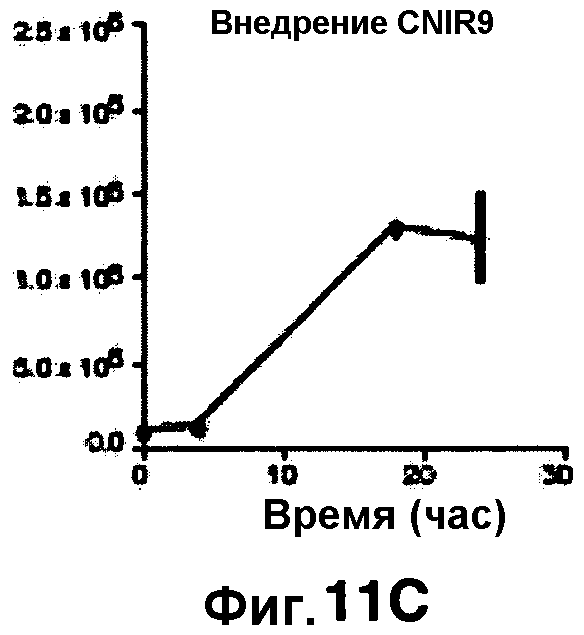
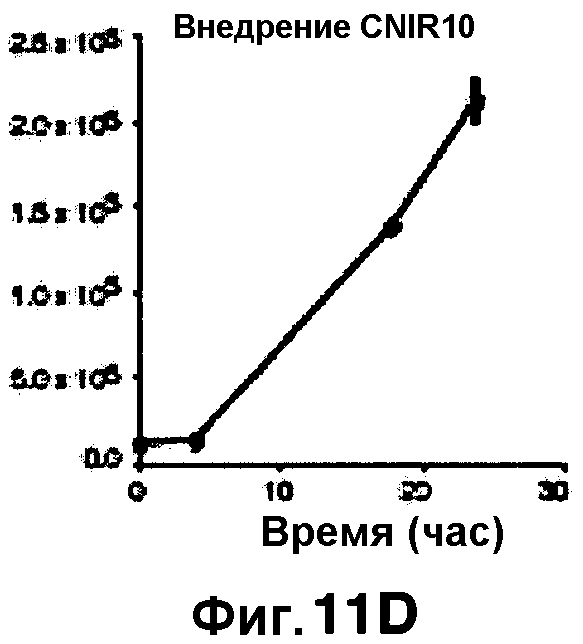
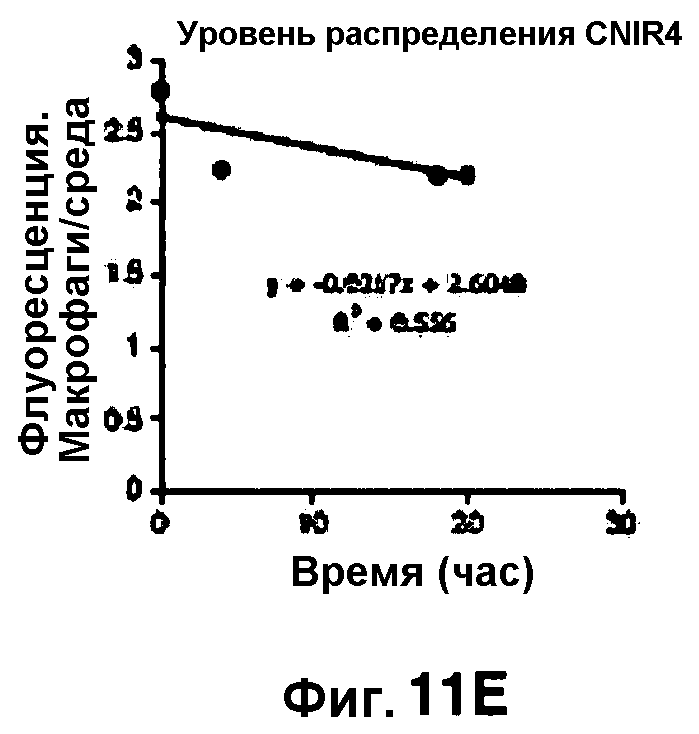
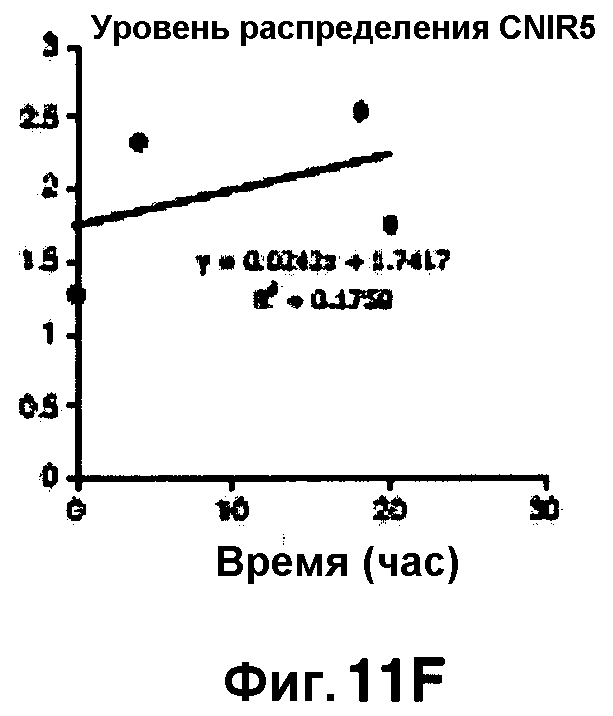
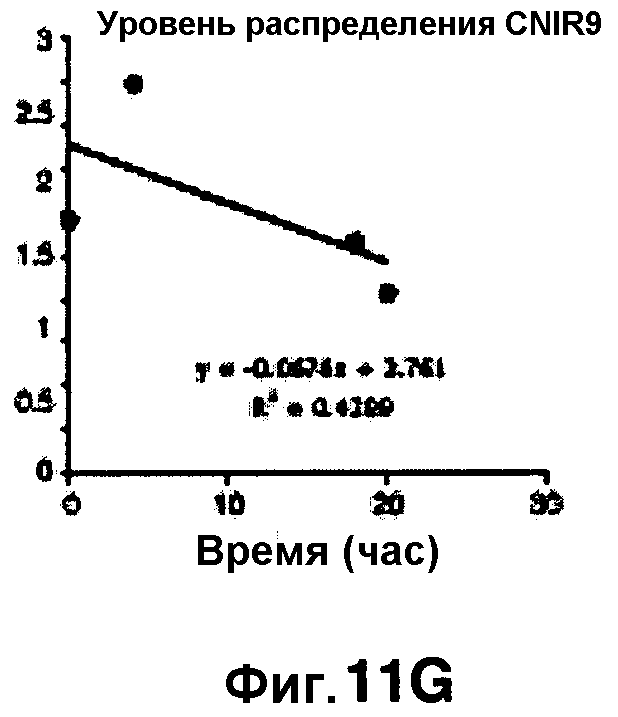
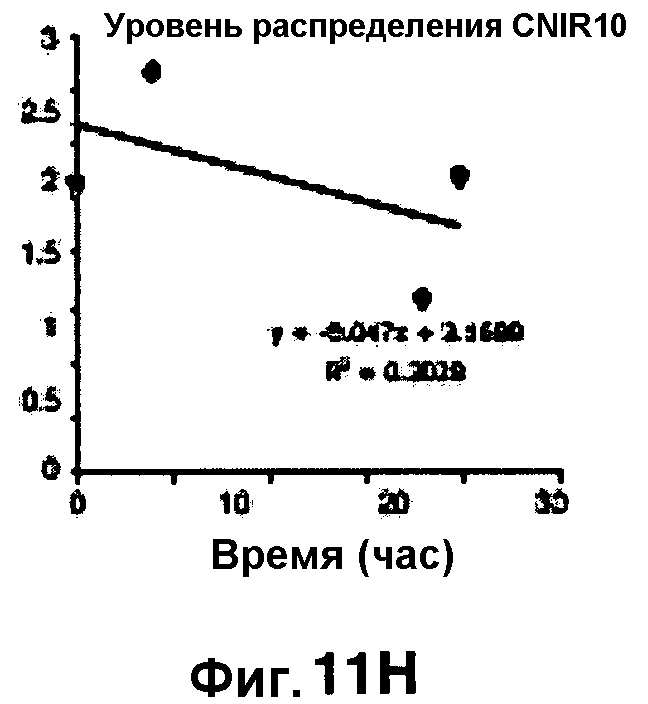
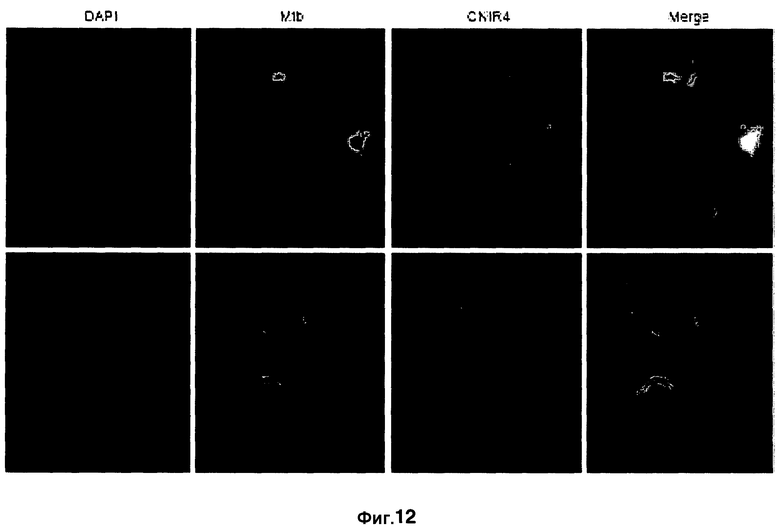



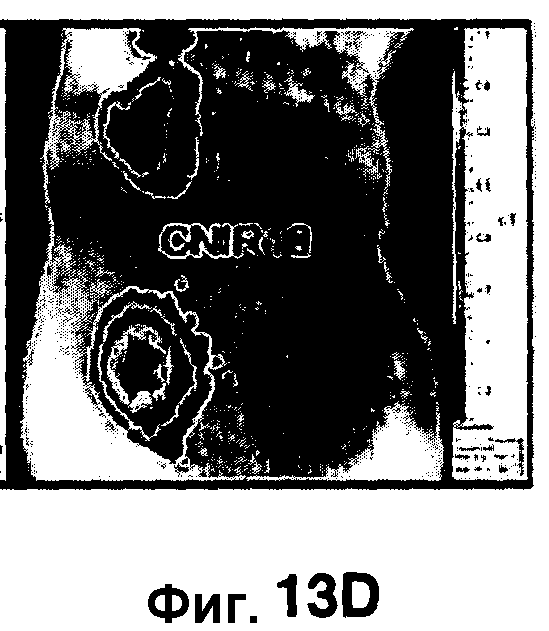
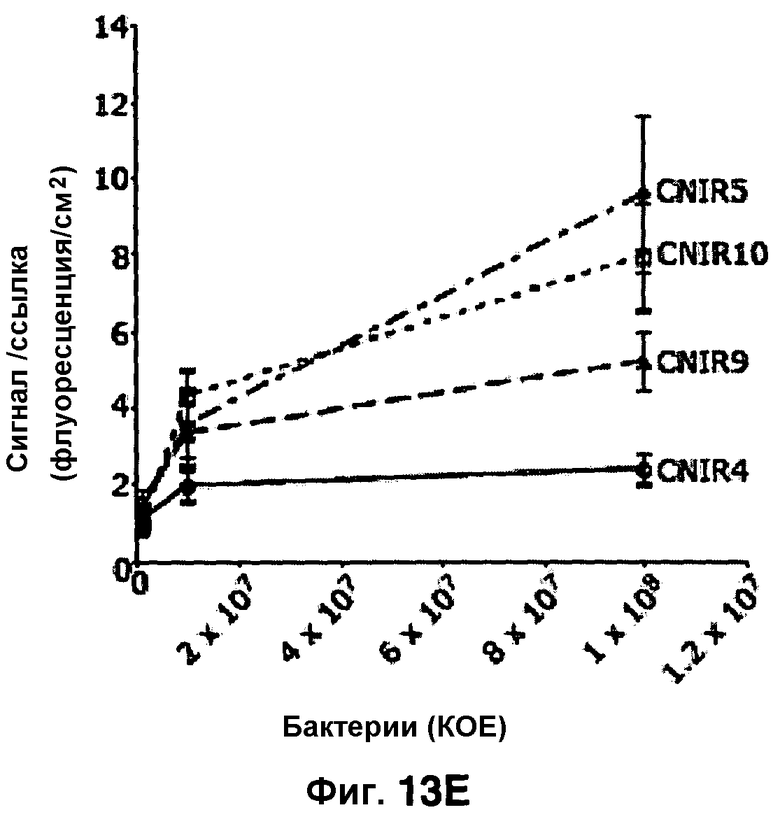

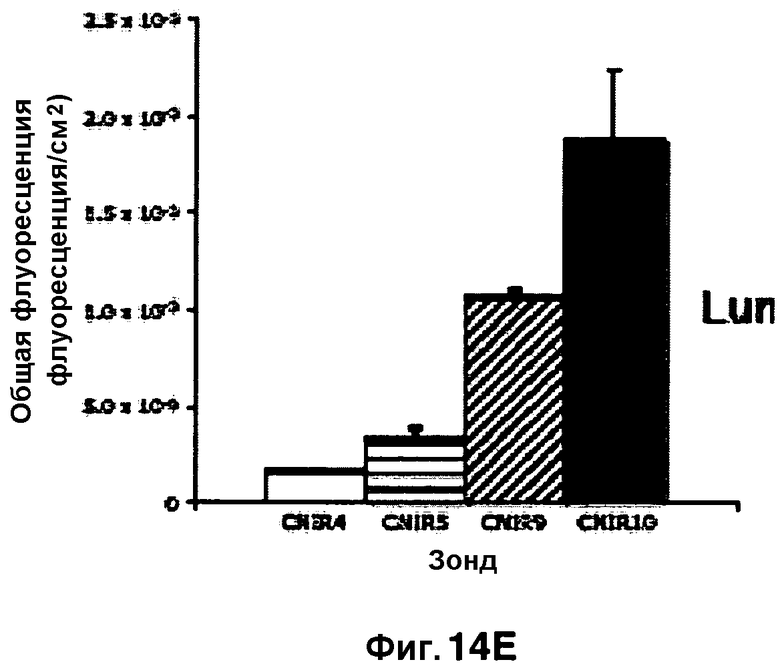
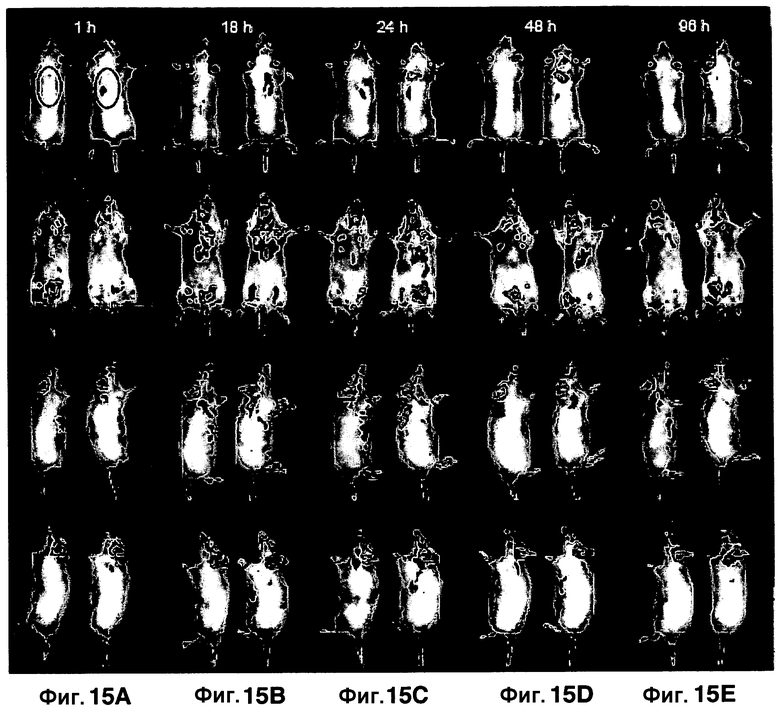
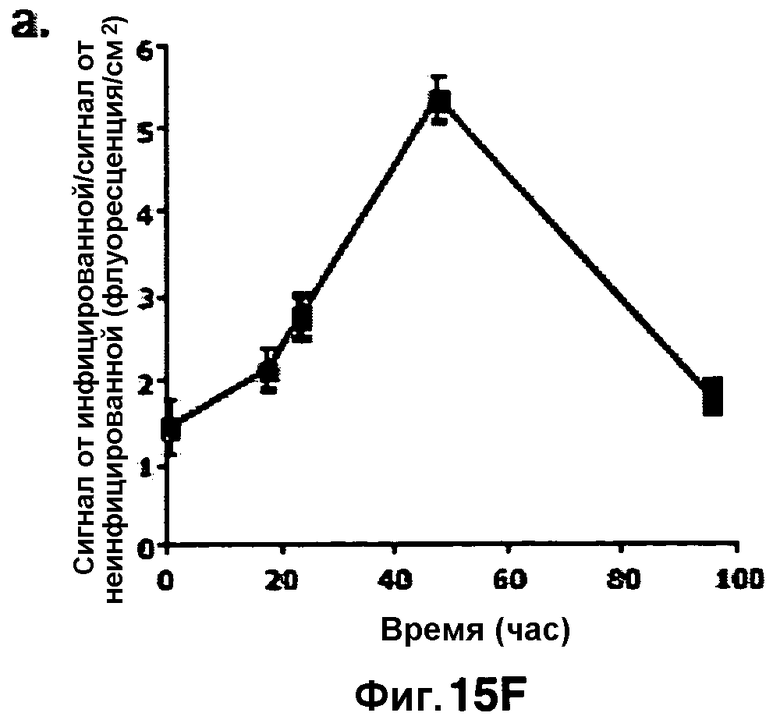
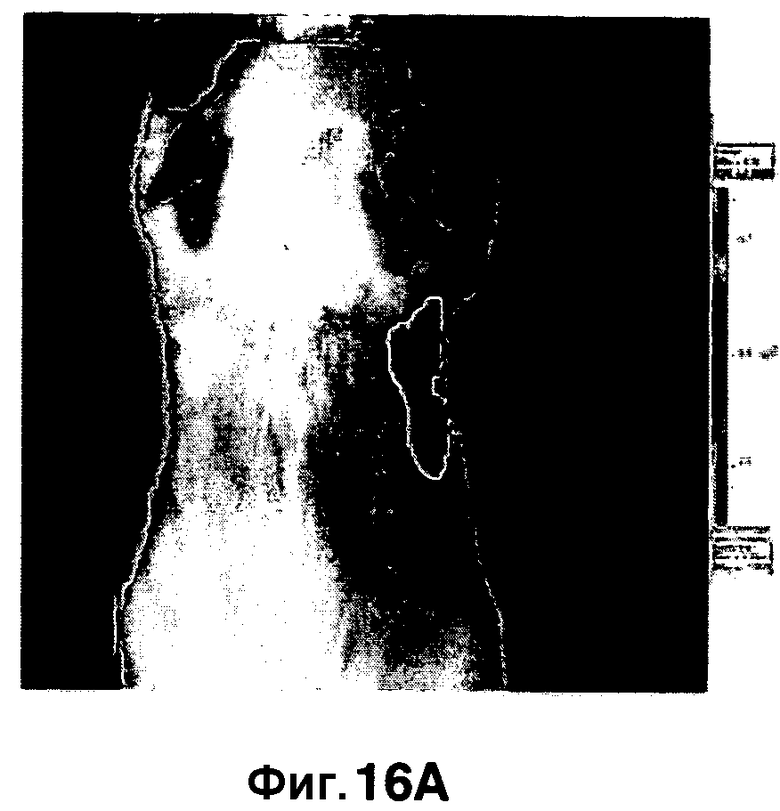
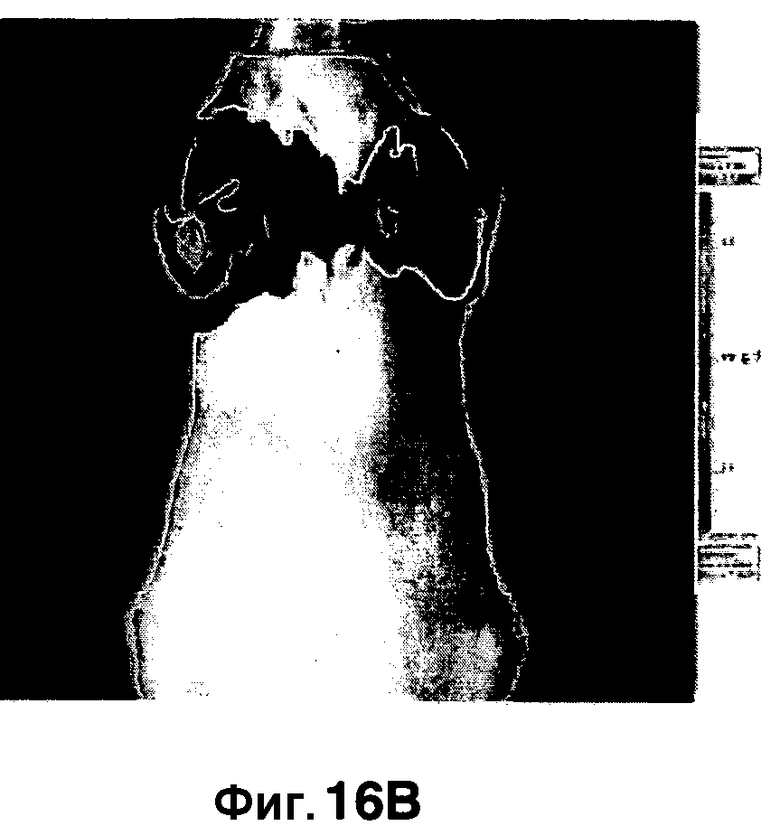
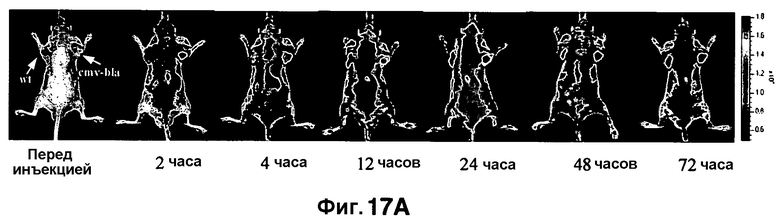
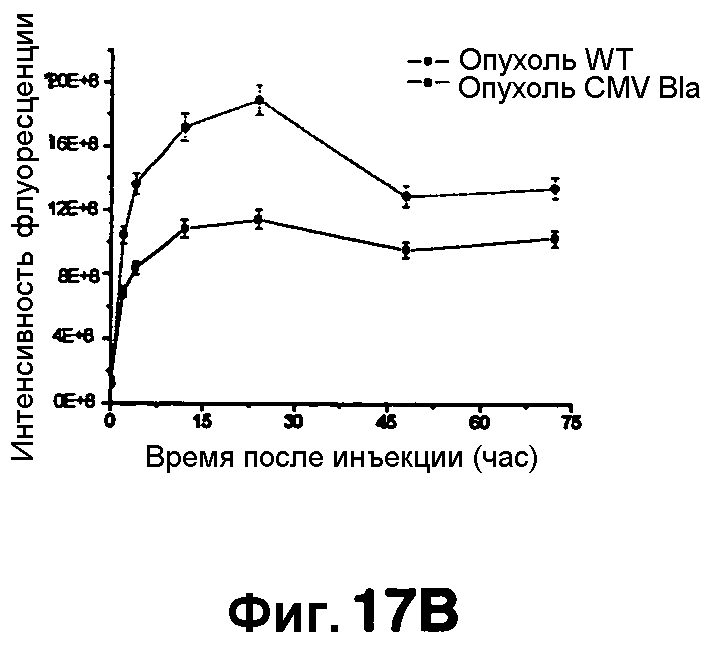

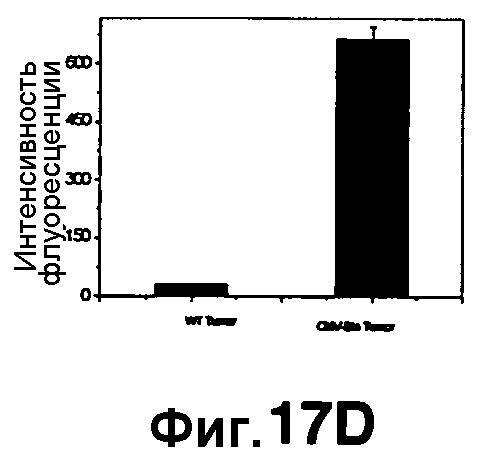
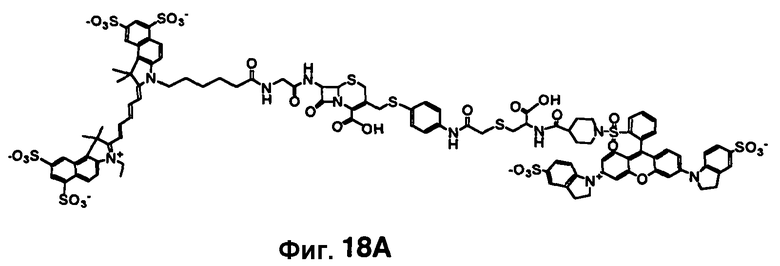
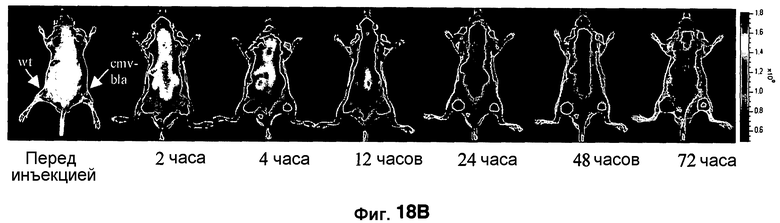
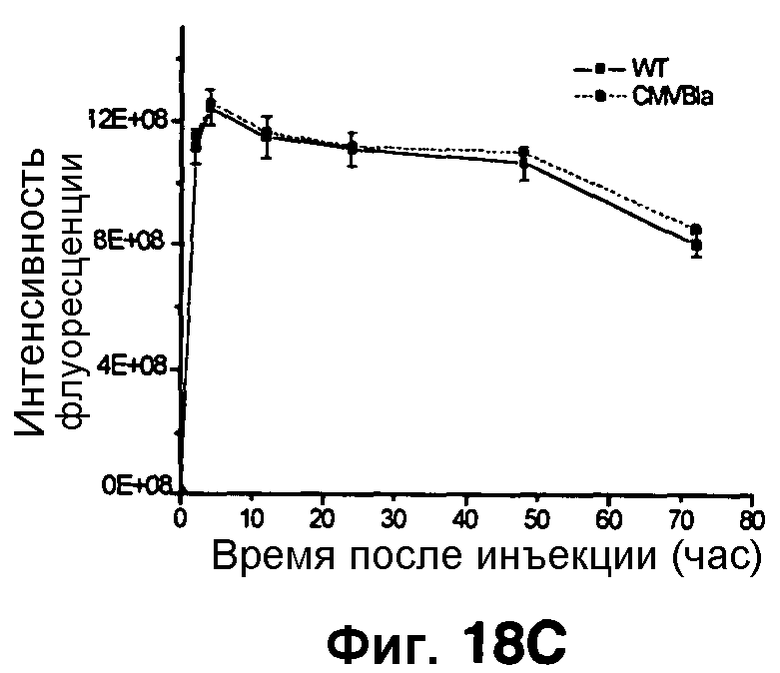
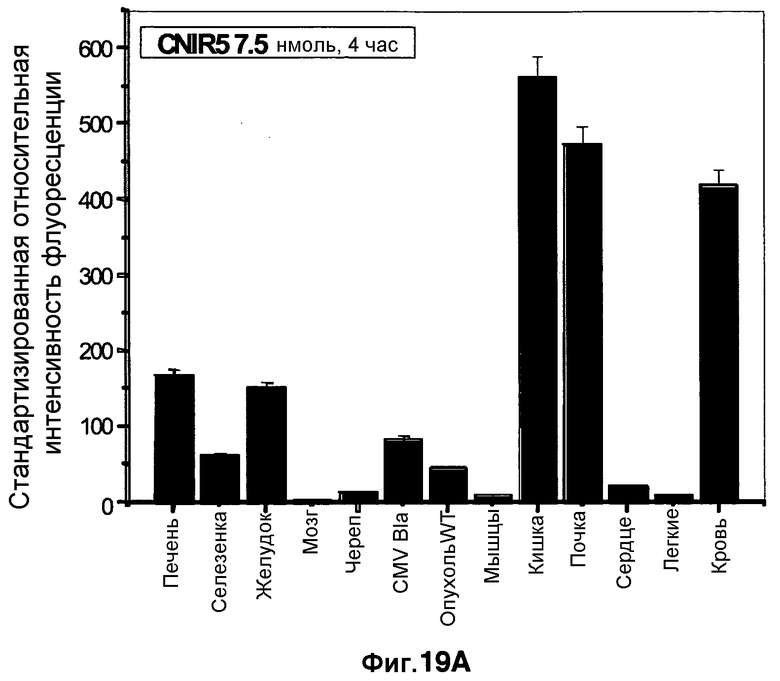
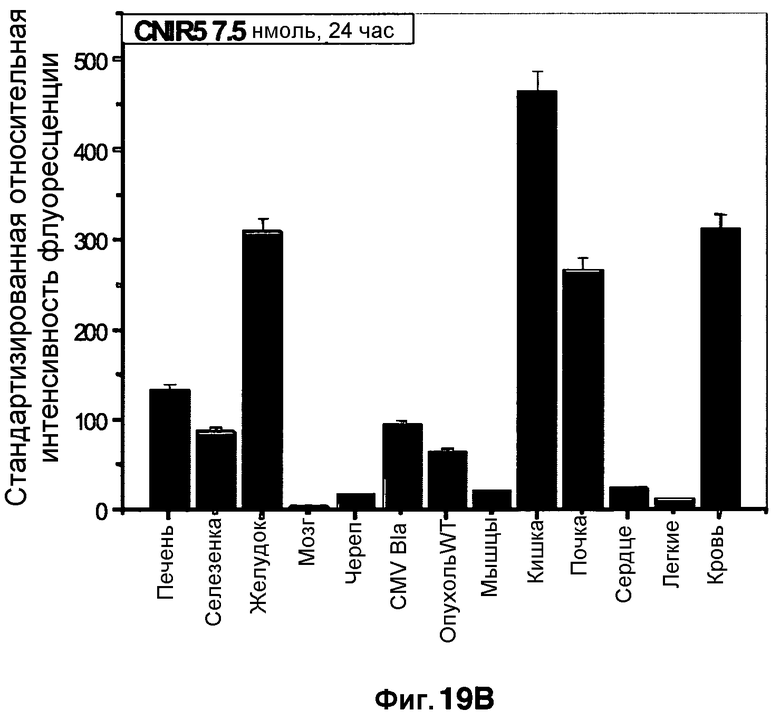
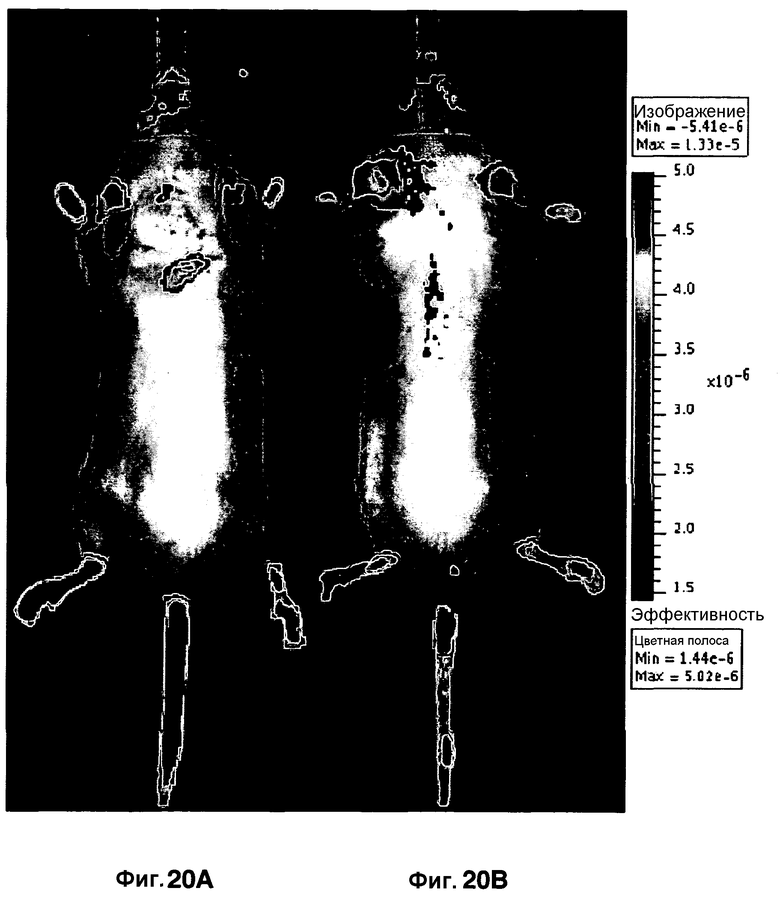
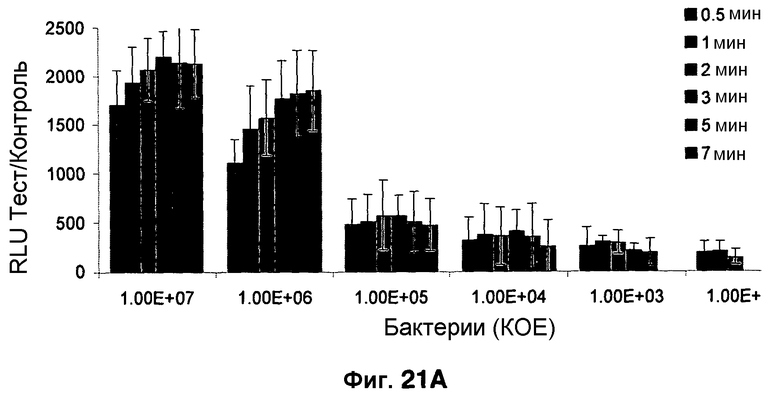
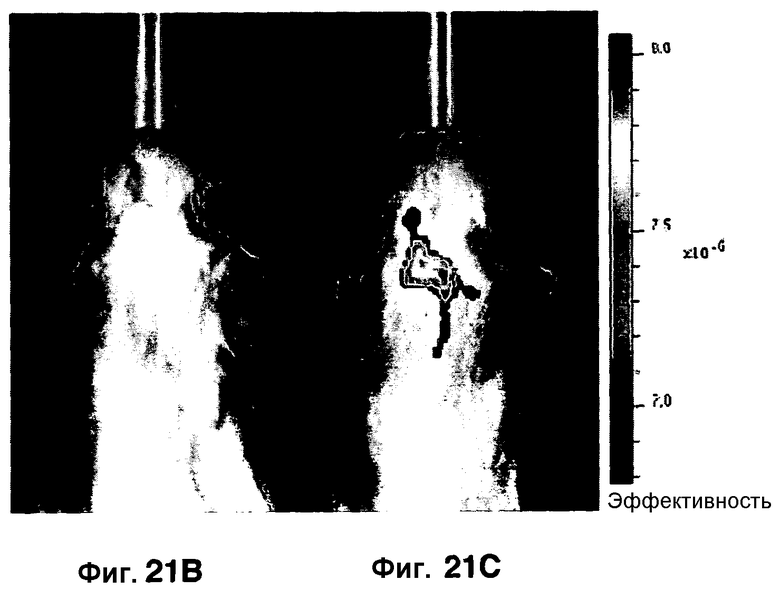
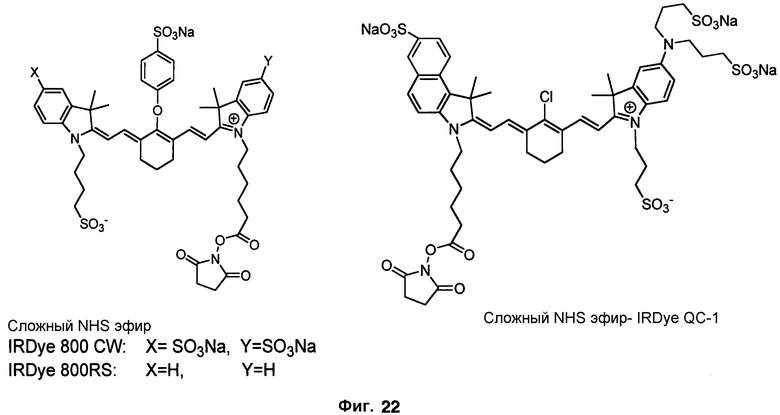
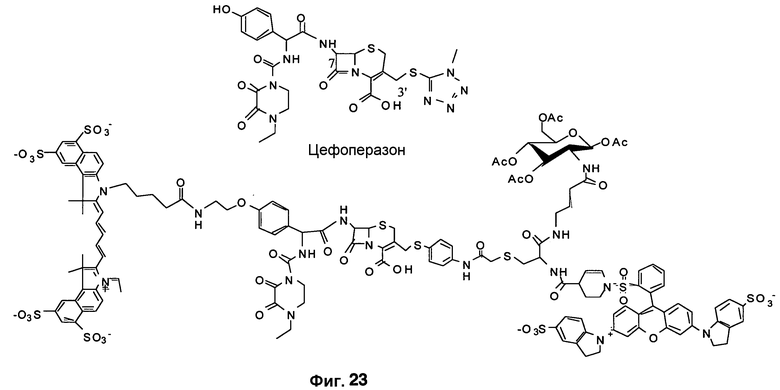
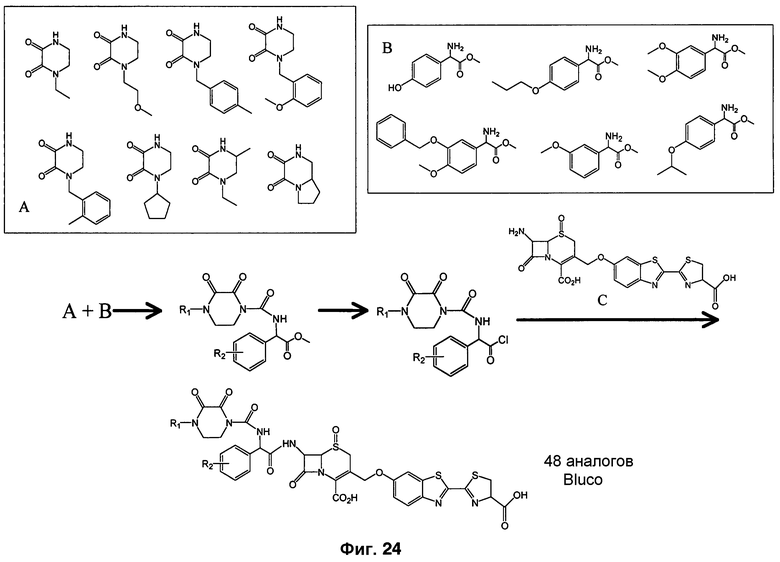
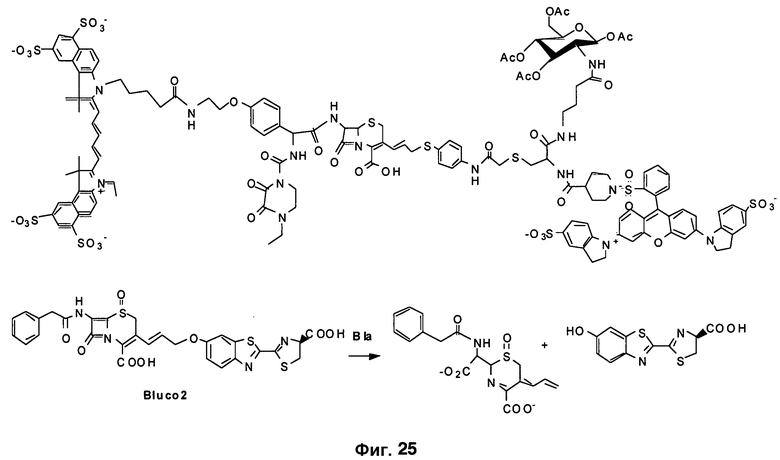
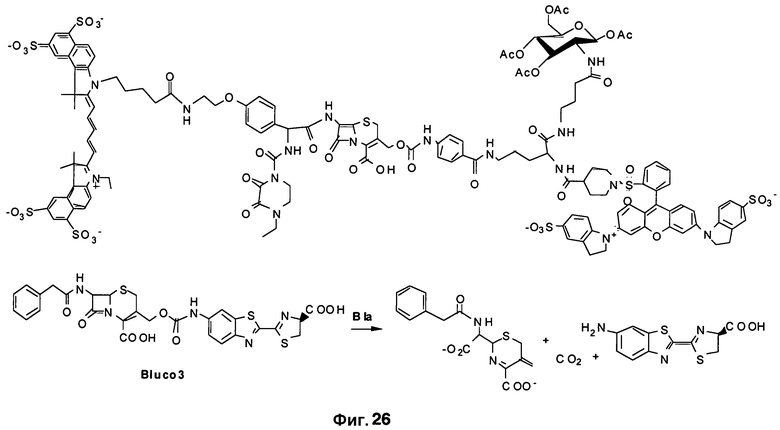
Изобретение относится к области микробиологии, а именно к использованию бактериальной бета-лактамазы для диагностики in vitro и визуализации, диагностики и лечения in vivo. Способ обнаружения патогенных бактерий в режиме реального времени у субъекта заключается в том, что осуществляют введение субъекту или взятие у субъекта образца флуорогенного субстрата для бета-лактамазы патогенных бактерий; визуализацию субъекта или образца на наличие флуоресцентного продукта бета-лактамазной активности на субстрате; получение сигналов на длине волны, испускаемой флуоресцентным бета-лактамазным продуктом, и обнаружение патогенных бактерий у субъекта в режиме реального времени. При этом флуорогенный субстрат представляет собой CNIR5, CNIR5-QSY22, CNIR7, CNIR7-TAT, CNIR9 или CNIR10. Способ мониторинга развития патофизиологического состояния, связанного с патогенными бактериями у субъекта. Способ скрининга соединений, обладающих терапевтическим эффектом против патогенной Mycobacterium у субъекта. Способ визуализации патогенных бактерий c флуорогенным субстратом для бактериальной бета-лактамазы. Использование заявленного изобретения позволяет улучшить визуализацию патогенных бактерий и мониторинг эффективности терапевтических соединений. 5 н. и 15 з.п. ф-лы, 26 ил., 1 табл., 14 пр.
1. Способ обнаружения патогенных бактерий в режиме реального времени у субъекта, включающий:
введение субъекту или во взятый у субъекта образец флуорогенного субстрата для бета-лактамазы патогенных бактерий;
визуализацию субъекта или образца на наличие флуоресцентного продукта бета-лактамазной активности на субстрате; и
получение сигналов на длине волны, испускаемой флуоресцентным бета-лактамазным продуктом; и обнаружение таким образом патогенных бактерий у субъекта в режиме реального времени, где указанный флуорогенный субстрат имеет химическую структуру, представляющую собой
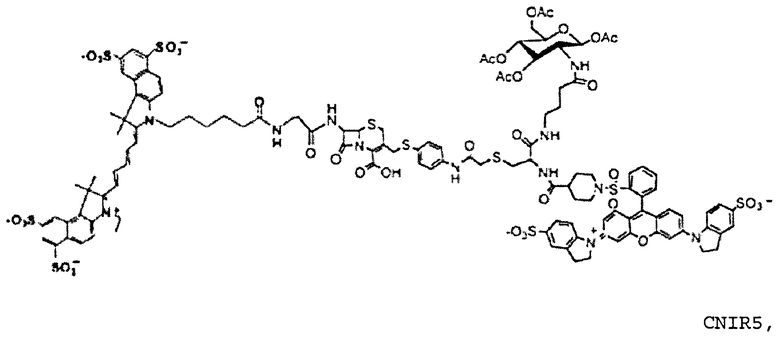
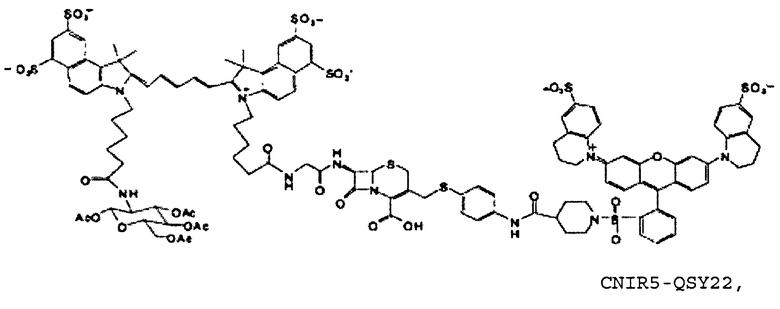
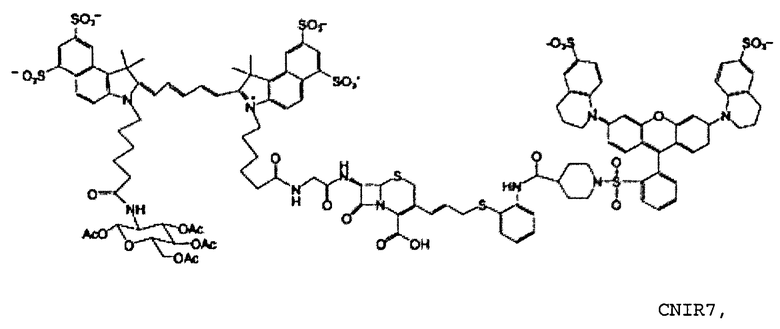
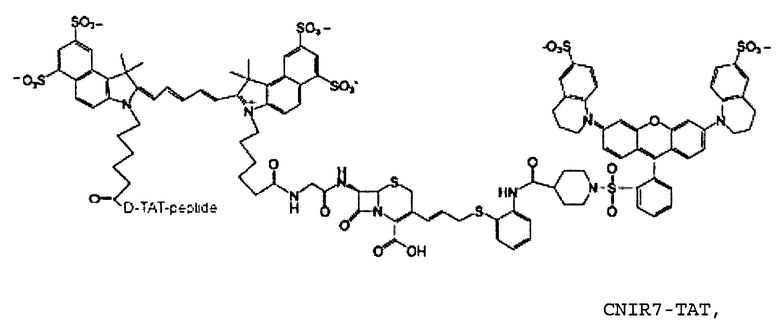
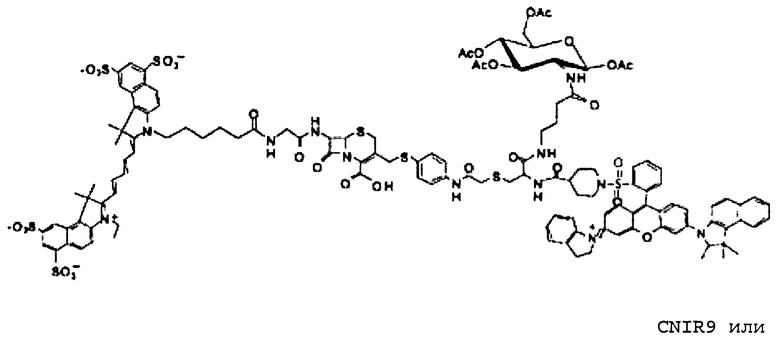
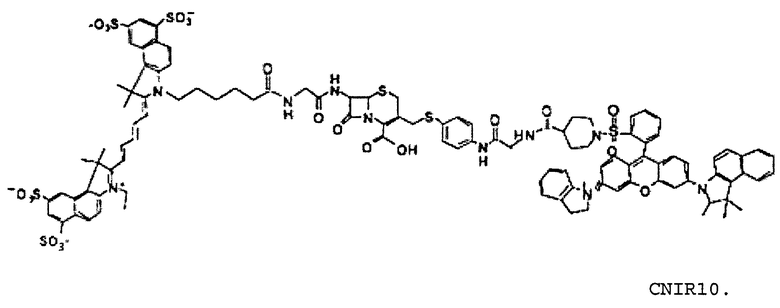
2. Способ по п.1, дополнительно включающий создание трехмерной реконструкции испускаемого сигнала для определения локализации патогенных бактерий у субъекта.
3. Способ по п.1, в котором патогенные бактерии включают виды бактерий Bacteroides, Clostridium, Streptococcus, Staphylococcus, Pseudomonas, Haemophilus, Legionella, Mycobacterium, Escherichia, Salmonella, Shigella или Listeria.
4. Способ по п.1, в котором длина волны визуализации составляет от приблизительно 300 нм до приблизительно 900 нм.
5. Способ по п.1, в котором длина волны испускаемых сигналов составляет от приблизительно 300 нм до приблизительно 900 нм.
6. Способ мониторинга развития патофизиологического состояния, связанного с патогенными бактериями у субъекта, включающий:
введение субъекту флуорогенного субстрата для бета-лактамазы патогенных бактерий или приведение биологического образца, полученного у субъекта, в контакт с флуорогенным субстратом для бета-лактамазы патогенных бактерий, который представляет собой CNIR5, CNIR5-QSY22, CNIR7, CNIR7-TAT, CNIR9 или CNIR10;
визуализацию указанного субъекта или указанного биологического образца на наличие флуоресцентного продукта бета-лактамазной активности на субстрате; и
измерение в режиме реального времени интенсивности флуоресцентного сигнала, полученного на длине волны, испускаемой флуоресцентным продуктом; и
сравнение измеренной в режиме реального времени интенсивности флуоресцентного сигнала с интенсивностью флуоресцентного сигнала флуоресцентного продукта, предварительно измеренной у субъекта, где повышение интенсивности флуоресцентного сигнала, измеренной в реальном времени, по сравнению с предварительно измеренной интенсивностью флуоресцентного сигнала коррелирует с развитием патофизиологического состояния у субъекта.
7. Способ по п.6, дополнительно включающий создание трехмерной реконструкции сигнала для определения локализации микробного патогена.
8. Способ по п.6, дополнительно включающий введение одного или более терапевтических соединений, эффективных для лечения патофизиологического состояния.
9. Способ по п.8, дополнительно включающий:
повторное введение флуорогенного субстрата субъекту или приведение биологического образца, полученного от субъекта, в контакт с указанным флуорогенным субстратом; и
визуализацию субъекта или указанного биологического образца для мониторинга эффективности терапевтического соединения; где понижение испускаемого сигнала после повторного введения флуорогенного субстрата указывает на терапевтическое воздействие на патофизиологическое состояние.
10. Способ по п.6, в котором патофизиологическое состояние представляет собой туберкулез.
11. Способ по п.6, в котором патогенные бактерии включают виды бактерий Bacteroides, Clostridium, Streptococcus, Staphylococcus, Pseudomonas, Haemophilus, Legionella, Mycobacterium, Escherichia, Salmonella, Shigella или Listeria.
12. Способ по п.6, в котором длина волны визуализации составляет от приблизительно 300 нм до приблизительно 900 нм.
13. Способ по п.6, в котором длина волны испускаемых сигналов составляет от приблизительно 300 нм до приблизительно 900 нм.
14. Способ скрининга соединений, обладающих терапевтическим эффектом против патогенной Mycobacterium у субъекта, который включает:
выбор потенциального терапевтического соединения для патогенной Mycobacterium;
приведение клеток Mycobacterium в контакт с флуорогенным субстратом для бета-лактамазы патогенной Mycobacterium, который представляет собой CNIR5, CNIR5-QSY22, CNIR7, CNIR7-TAT, CNIR9 или CNIR10;
приведение бактериальных клеток в контакт с потенциальным терапевтическим соединением; и
измерение флуоресцентного сигнала, продуцируемого бактериальными клетками в присутствии потенциального терапевтического соединения и в его отсутствие; где понижение сигнала в присутствии терапевтического соединения по сравнению с сигналом в его отсутствие указывает на терапевтический эффект соединения против патогенных бактерий.
15. Способ по п.14, в котором патофизиологическое состояние представляет собой туберкулез.
16. Способ визуализации патогенных бактерий, включающий:
приведение патогенных бактерий в контакт с флуорогенным субстратом для их фермента бета-лактамазы, который представляет собой CNIR5, CNIR5-QSY22, CNIR7, CNIR7-TAT, CNIR9 или CNIR10;
воздействие на патогенные бактерии длиной волны возбуждения, для флуоресцентного продукта бета-лактамазной активности на субстрате;
получение флуоресцентных сигналов, излучаемых флуоресцентным продуктом; и
получение трехмерной реконструкции полученных сигналов, за счет чего осуществляется визуализация патогенных бактерий.
17. Способ по п.16, в котором патогенные бактерии включают виды бактерий Bacteroides, Clostridium, Streptococcus, Staphylococcus, Pseudomonas, Haemophilus, Legionella, Mycobacterium, Escherichia, Salmonella, Shigella или Listeria.
18. Способ по п.16, в котором длина волны возбуждения составляет от приблизительно 540 нм до приблизительно 730 нм.
19. Способ по п.16, в котором длина волны испускаемых сигналов составляет от приблизительно 650 нм до приблизительно 800 нм.
20. Флуорогенный субстрат для бактериальной бета-лактамазы, представляющий собой CNIR5, CNIR5-QSY22, CNIR7, CNIR7-TAT, CNIR9 или CNIR10.
| XING B | |||
| et al Cell-permeable near-infrared fluorogenic substrates for imaging beta-lactamase activity | |||
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| ZANG Y | |||
| et al Beta-Lactamase inhibitors and the inducibility of the Beta-lactamase of Mycobacterium tuberculosis | |||
| Am Rev Respir Dis | |||
| Пуговица для прикрепления ее к материи без пришивки | 1921 |
|
SU1992A1 |
| YAO H | |||
| et al A bioluminogenic substrate for | |||

