Изобретение относится к микробиологической и медицинской промышленности, генной и белковой инженерии, конкретно - к получению корового антигена вируса гепатита В (HBcAg), несущего иммуногенный эпитоп района preS1 поверхностного белка вируса гепатита В (27-37 а.о.)
Вирусные капсиды являются благодарным объектом для конструирования антигенов с заданными свойствами, поскольку жесткая фиксация и многократное повторение антигенных детерминант на поверхности вирусных капсидов обеспечивают их высокую антигенность и иммуногенность [1]. Конструирование вирусных капсидов, экспонирующих чужеродные эпитопы, обещает стать перспективным направлением в создании эффективных вакцин. Одним из наиболее многообещающих векторов экспонирования чужеродных детерминант является нуклеокапсидный белок вируса гепатита В человека. Он представляет собой капсид, состоящий из 180 одинаковых белковых субъединиц, способных к самоорганизации. Способность субъединиц к самосборке сохраняется как в эукариотических, так и в прокариотических системах экспрессии [2].
Химерные капсиды на основе HBcAg могут быть использованы как компоненты диагностикумов и вакцин. Последнее тем более перспективно, что HBcAg при иммунизации не только обеспечивает высокий уровень синтеза специфических антител, но и обладает ярко выраженным свойством стимулировать Т-клеточный иммунный ответ [3].
В настоящее время в литературе описаны способы получения химерных частиц HBcAg, несущих эпитопы различных вирусов - вируса ящура (FMDV), риновируса человека (HRV2) [4], вируса гепатита В, вируса гепатита С и других [5]. Химерные частицы были способны индуцировать иммунный ответ на встроенный пептид при иммунизации лабораторных животных и человека [6].
Известны конструкции HBcAg, несущие детерминанты районов preS поверхностного белка вируса гепатита В, в которых эпитопы встраивались в С-конец или в N-конец кор-антигена вируса гепатита В [7, 8]. Такие химерные белки хорошо самоорганизовывались в частицы, подобные нативному кору, однако иммунный ответ на встроенную детерминанту был невысок (на три порядка ниже, чем на HBcAg).
Более перспективным местом для встроек, по-видимому, является область петли Е1, соответствующая 71-89 аминокислотным остаткам HBcAg. Авторами работы [9] было показано, что этот район кора поверхностно экспонирован и является основной антигенной детерминантой HBcAg. Эпитоп района preS1 (31-36 а.о.), встроенный в основную антигенную детерминанту HbcAg, стимулировал высокий титр антител к данной вставке у иммунизированных животных [10, прототип]; данная конструкция является наиболее близким аналогом, и она выбрана в качестве прототипа. К недостаткам описанной конструкции можно отнести то, что она содержит лишь небольшой фрагмент последовательности района preS1 HBsAg-эпитоп, узнающийся моноклональными антителами МА 18/7. В то же время исследования с использованием панелей моноклональных антител показывают, что иммуногенный участок в данном районе шире. В работе Coursaget P. с соавторами [11] установлено, что этот район состоит из 29-36 аминокислотных остатков, в другой работе, проведенной Kuroki К с соавторами [12], описан район 27-36 аминокислотных остатков. Также результаты работы, посвященной определению данной детерминанты с помощью бактериального дисплея [13], показали, что присутствуют клоны, взаимодействующие с сыворотками животных, иммунизированных белком preS1, имитирующие последовательность 31-34 аминокислотного остатка, но большая часть полученных клонов узнает район 28-31 аминокислотных остатков. Таким образом, можно сделать вывод, о том, что иммунноактивный эпитоп данного района шире, чем предполагалось ранее, и конструкции, основанные на HBcAg, включающем последовательность района preS1 31-34 аминокислотных остатков, не смогут в полной мере стимулировать иммунный ответ к иммуноактивной области района preS1 поверхностного белка вируса гепатита В.
Технической задачей изобретения является создание химерных частиц HBcAg, содержащих последовательность уточненного, более широкого эпитопа района preS1 (27-37 а.о.) поверхностного белка вируса гепатита В (HBsAg), и бактериального штамма E.coli - продуцента белка НВс со встройкой эпитопа района preS1 (27-37 а.о.) поверхностного белка вируса гепатита В (HBsAg).
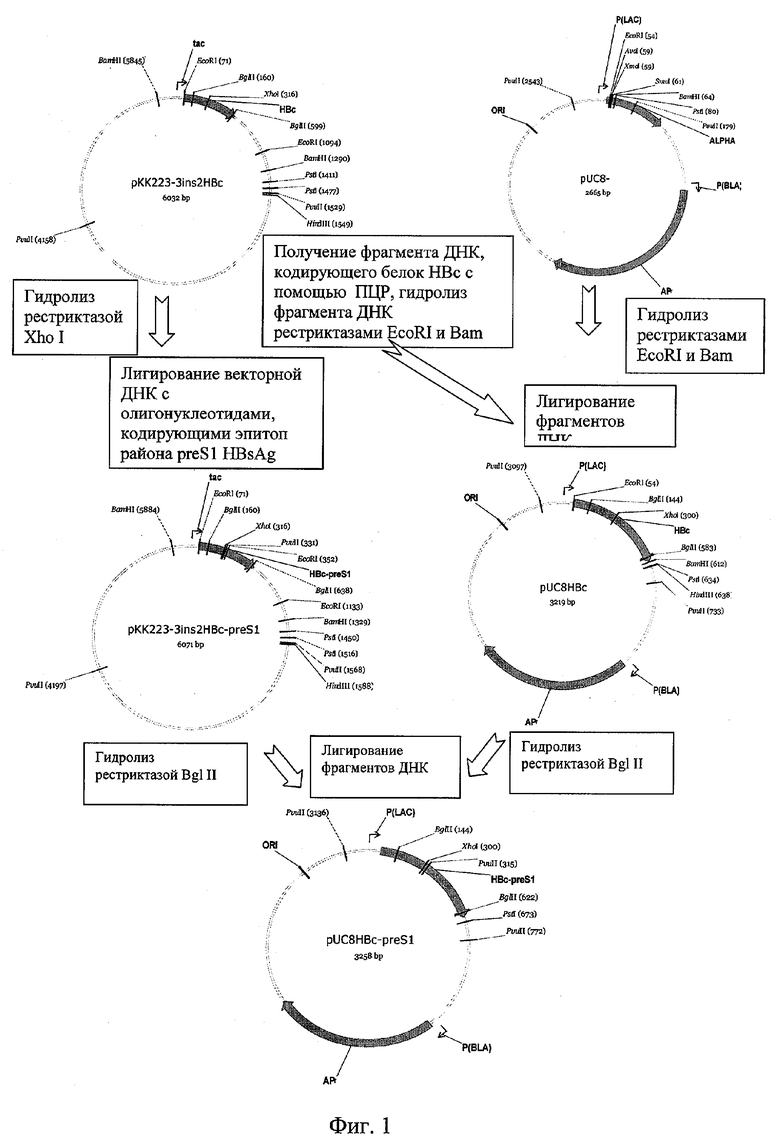
Поставленная задача решается путем введения с помощью клонирования олигонуклеотидов, кодирующих эпитоп (27-37 а.о.) района preS1 поверхностного белка вируса гепатита В в плазмиду, содержащую ген HBcAg. Все генно-инженерные манипуляции проводились согласно руководству Маниатис Т. и другие [14]. Сконструированная плазмида pUCHBc-preS1 содержит плазмидный вектор pUC8 [15] и ген химерного белка HBc-preS1. Ген химерного белка получен путем клонирования олигонуклеотидов
5'-TCG AGC GAT CAC CAG CTG GAC CCG GCG TTC AGA GCG AAT
CG СТА GTG GTC GAC CTG GGC CGC AAG TCT CGC TTA AGC T-5',
кодирующих эпитоп района preS1 в плазмиде рКНВс [16], содержащей ген НВс, в район, соответствующий 81 аминокислотному остатку НВс белка (с использованием ранее введенного XhoI сайта рестрикции). Фрагмент ДНК, кодирующий ген НВс, получен методом ПЦР с использованием плазмиды рКНВс (в качестве матрицы) и олигонуклеотидов:
Прямой: 5'-ACGAATTCTATGGACATTGATCCTT
Обратный: 5'-CAAGGGATCCTAACATTGAGATTCC
Полученный фрагмент гидролизуют ферментами рестрикции EcoRI и BamHI и лигируют с векторм pUC8, гидролизованным теми же ферментами. Для получения плазмидного вектора pUC8HBc-preS1, несущего ген химерного белка HBc-preS1, фрагмент гена HBc-preS1 (с использованием рестриктазы BglII) из плазмиды рКНВс клонируют в плазмиде pUC8HBc.
Полученная целевая плазмида pUC8HBc-preS1 (фиг.1) с молекулярной массой 2,11 МД имеет размер 3258 п.о. и состоит из следующих элементов:
- EcoRI - BamHI-фрагмента векторной плазмиды pUC18 размером 2686 п.о., содержащего ген b-лактамазы, сайт инициации репликации плазмиды pBR322, часть lac-гена из М13mp18, промотор Plac, полилинкерную последовательность из М13mp18; уникальные сайты рестрикции с координатами: Hind III (677), Pst I (673), Afl III (2956), Cfr10 I (1983), Ssp I (1263),
- BamHI - EcoRI-фрагмента ДНК плазмиды р7,5С [17] размером 890 п.о., содержащего ген химерного корового белка вируса гепатита В, несущего эпитоп района preS1 (27-37 а.о.) поверхностного белка вируса гепатита В в районе, соответствующем 81-82 аминокислотным остаткам HBcAg;
содержит:
- в качестве генетического маркера bla-ген ампициллин резистентности (ген b-лактамазы), определяющий устойчивость к ампициллину при трансформации клеток E.coli;
- уникальные сайты рестрикции и их координаты: Hind III (677), Pst I (673), Afl III (2956), Cfr10 I (1983), Ssp I (1263).
Для получения бактериального штамм-продуцента гибридного белка кор-антигена HBc-preS1 компетентные клетки бактерий Escherichia coli DH5αF' трансформируют сконструированной целевой плазмидой pUC8HBc-preS1. Полученный штамм бактерий характеризуется следующими признаками:
Морфологические признаки:
Клетки мелкие утолщенной палочковидной формы, граммотрицательные, неспороносные.
Культуральные признаки:
Клетки хорошо растут на простых питательных средах. При росте на агаре "Дифко"-колонии круглые, гладкие, прижатые, мутные, блестящие серые, край ровный. При росте в жидких средах (например, LB-бульон) образуют интенсивную ровную муть. Клетки растут при 37°С при оптимуме рН от 6,8 до 7,0.
Устойчивость к антибиотикам:
Клетки проявляют устойчивость к ампицилину (100 мкг/мл), обусловленную наличием плазмиды.
Штамм E.coli DH5αF'/pUC8HBc-preS1 обеспечивает синтез гибридного белка HBc-preS1 с уровнем продукции 2% от суммарного клеточного белка.
Полученный штамм депонирован в НИИ "Коллекция культур микроорганизмов " Государственного научного центра вирусологии и биотехнологии "Вектор", регистрационный номер В-1055.
Сущность изобретения заключается в том, что для экспрессии гена гибридного корового белка вируса гепатита В, несущего встройку эпитопа района preS1 (27-37 а.о.) поверхностного белка вируса гепатита В, олигонуклеотиды, кодирующие эпитоп района preS1 поверхностного белка вируса гепатита В, встраивают в район гена корового белка, соответствующий 81-82 аминокислотному остатку корового белка. Полученный гибридный ген клонируют в экспрессирующий плазмидный вектор pUC8HBc с целью повысить продукцию целевого белка. В результате получают плазмиду pUC8HBc-preS1, кодирующую ген гибридного белка HBc-preS1.
Экспрессию гибридного гена осуществляют в клетках E.coli DH5αF'. Индикацию экспрессии осуществляют с помощью белкового электрофореза и иммуноблоттинга. Рекомбинантный гибридный белок узнается моноклональными антителами к N-концу НВс белка. Уровень экспрессии определяют с помощью денситометрии полиакриламидного геля, окрашенного Кумаси. Уровень экспрессии составляет 2% общего белка биомассы.
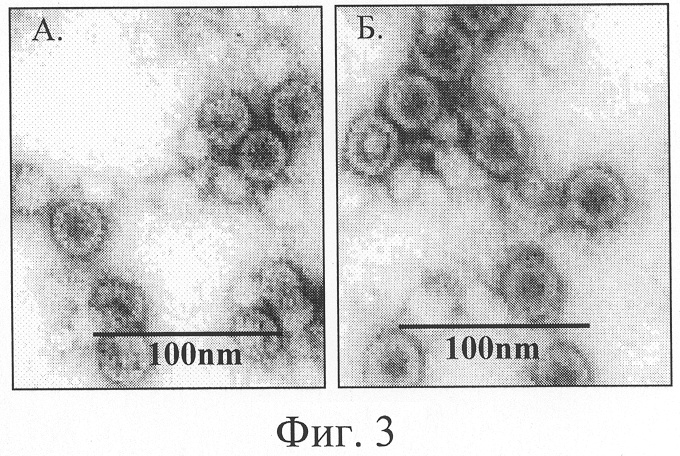
С помощью электронной микроскопии показано, что гибридный белок собирается в частицы, по форме и морфологии подобные природным коровым частицам вируса гепатита В.
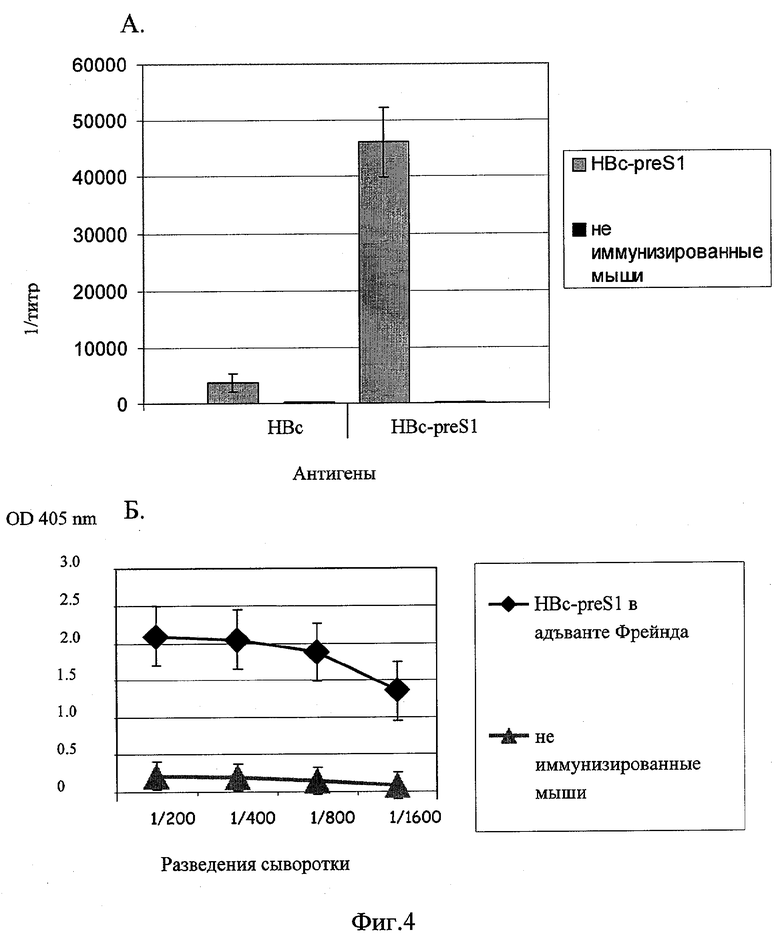
Иммуногенность гибридного рекомбинантного белка HBc-preS1 определяют внутримышечной иммунизацией лабораторных животных раствором данного белка. Лабораторных мышей линии Balb/с (12-14 г) иммунизируют внутримышечно трехкратно дозой 5 мкг белка, с использованием адъюванта Фрейнда. Оценку напряженности иммунного ответа проводят методом иммуноферментного анализа (ИФА). В качестве антигена для ИФА используют рекомбинантный белок HBc-preS1 или пептид с последовательностью эпитопа района preS1 поверхностного белка. Иммунизация полученным химерным гибридным белком HBc-preS1 стимулирует синтез специфических антител как к целому белку HBc-preS1, так и к отдельному эпитопу района preS1 HBsAg.
Таким образом, впервые получены плазмидная ДНК и штамм-продуцент, обеспечивающие экспрессию HBcAg, несущего эпитоп района preS1 HBsAg, вызывающего напряженный иммунный ответ к эпитопу района preS1 HBsAg.
Изобретение иллюстрируется следующими фигурами:
Фиг.1. Физическая карта плазмиды pUC8HBc-preS1 и схема ее конструирования.
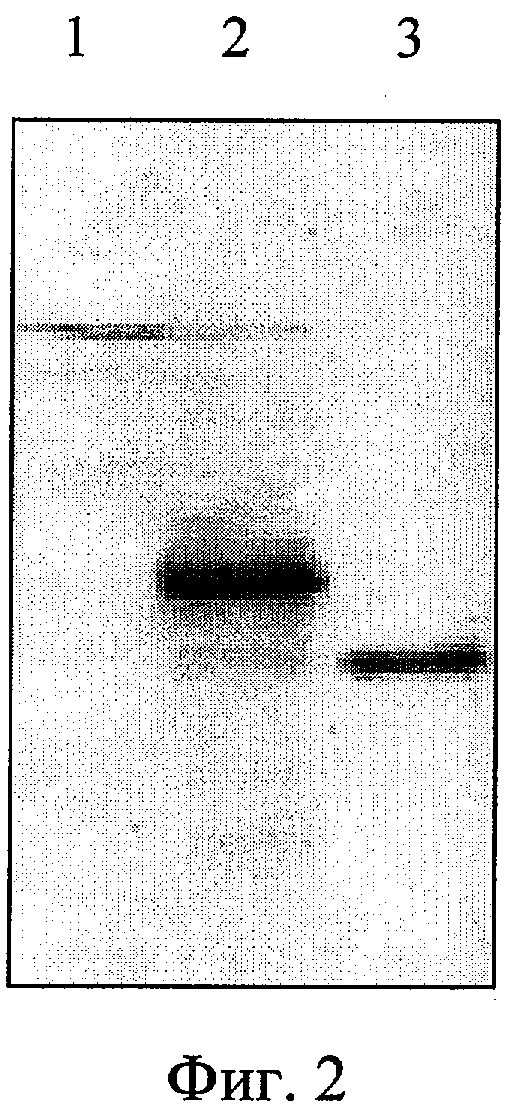
Фиг.2. Электрофоретический анализ в 15% полиакриламидном геле с последующим иммуноблотингом клеточного лизата штамма E.coli DH5αF'/pUC8HBc-preS1. Дорожки: 1 - лизат клеток E.coli DH5αF', 2 - лизат клеток E.coli DH5αF'/pUC8HBc-preS1, 3 - очищенный белок НВс.
Фиг.3. Электронная фотография рекомбинантных белков HBcAg (Фиг.3А) и HBcAg-preS1 (Фиг.3Б). Препараты HBcAg готовят методом негативного контрастирования 2%-ным раствором фосфорно-вольфрамовой кислоты и исследуют в электронном микроскопе "JEM 100C".
Фиг.4. Данные ИФА сывороток животных, иммунизированных гибридным белком HBcAg-preS1. А. Результаты ИФА с использованием в качестве антигенов целых белков. Б. Результаты ИФА с использованием в качестве антигена пептида DHQLDPAFRAN.
Для лучшего понимания сущности изобретения ниже следуют примеры его конкретного выполнения.
Пример 1. Конструирование плазмиды pUC8HBc-preS1.
Конструирование генов, гибридных НВс белков, содержащих эпитопы поверхностного белка вируса гепатита В.
1 мкг плазмидной ДНК рКНВс [16] гидролизуют с помощью фермента XhoI, в реакционной смеси, содержащей 5 ед. XhoI, 50 mM трис-HCl рН 7,6, 10 mM MgCl2, 1 mM DTT. Реакцию ведут 1 ч при 37°С. Олигонуклеотидный дуплекс получают, смешивая олигонуклеотиды по 20 рМ каждый в 10 мкл раствора, содержащего 20 mM DTT. Пробирку со смесью нагревают до 75°С и медленно остужают до комнатной температуры. 1 рМ гидролизованной ДНК смешивают с 20 рМ олигонуклеотидного дуплекса и лигазой бактериофага Т4 (2 ед.а.) в смеси, содержащей также 50 mM Tris-HCl (pH 7.8 при 25°С); 10 mM MgCl2; 10 mM DTT; 1 mM rATP; 25 мкг/мл BSA при 14°С в течение 16 часов. Инкубируют в течение 16 часов при 14°С. Полученной лигазной смесью трансформируют клетки E.coli DH5αF'. Клоны, содержащие плазмиду с гибридным геном, отбирают с помощью рестрикционного анализа. Правильность нуклеотидной последовательности встроенных фрагментов ДНК подтверждают секвенированием по методу Сэнгера.
Конструирование плазмиды pUC8HBc-preS1.
Фрагмент ДНК, кодирующий ген гибридного белка НВс, получают методом ПЦР. 1 нг плазмидной ДНК рКНВс смешивают с 20 рМ прямого и обратного олигонуклеотидов в растворе, содержащем 0,1 М смесь трифосфатов, 60 mM Tris-HCl (pH 8.5 при 25°С); 1.5 mM MgCl2; 25 mM KCl; 10 mM 2-меркаптоэтанол; 0.1% Тритон Х-100, 10 ед.а. Taq-полимеразы. Реакцию полимеризации проводят при 55°С, 30 циклов. Далее полученный фрагмент ДНК гидролизуют ферментами рестрикции BamHI и EcoRI в буфере, содержащем 100 mM Tris-HCl (pH 7.6 при 25°С); 10 mM MgCl2; 50 mM NaCl; 1 mM DTT). Векторную плазмиду pUC8 также гидролизуют ферментами BamHI и EcoRI. Полученные фрагменты ДНК лигируют с помощью лигазы бактериофага Т4 (2 ед.а.) в буфере, содержащем 50 mM Tris-HCl (pH 7.8 при 25°С); 10 mM MgCl2; 10 mM DTT; 1 mM rATP; 25 мкг/мл BSA при 14°С в течение 16 часов. Полученной лигазной смесью трансформируют клетки E.coli DH5αF'. Клоны, содержащие плазмиду с гибридным геном, отбирают с помощью рестрикционного анализа. Правильность нуклеотидной последовательности встроенных фрагментов ДНК подтверждают рестрикционным анализом.
Клонирование гена гибридного белка HBc-preS1 в плазмиде pUC8HBc.
Плазмиды pUC8HBc и pKHBc-preS1 гидролизуют рестриктазой Bgl II, фрагменты ДНК векторной плазмиды pUC8HBc и фрагмент гена HBc-preS1 лигируют с помощью лигазы бактериофага Т4 (2 ед.а.) в смеси, содержащей также 50 mM Tris-HCl (рН 7.8 при 25°С); 10 mM MgCl2; 10 mM DTT; 1 mM rATP; 25 мкг/мл BSA при 14°С в течение 16 часов. Полученной лигазной смесью трансформируют клетки E.coli DH5αF'. Клоны, содержащие целевую плазмиду с гибридным геном, отбирают с помощью рестрикционного анализа.
Пример 2. Характеризация рекомбинантного белка HBc-preS1.
Клетки E.coli DH5αF'/pUC8HBc-preS1 растят на среде LB в течение 16 часов при 37°С. Электрофоретическое разделение белков проводят в 13%-ном полиакриламидном геле в присутствии 0,1% ДСН по Лэмли [18]. Перенос белков из геля на фильтр осуществляли при напряжении электрического поля 10 В/см в течение 2-х часов. Фильтры помещали в 3%-ный раствор БСА в 0,01 М трис-HCl, рН 7,5, 0,05% твин-20 на 1 час, после чего инкубировали 1 час с раствором с моноклональными антителами к HBcAg, разведенным в буфере 0,01 М трис-HCl, рН 7,5, 0,15М NaCl, 0,05% твин-20, 0,05% NaN3, 0,001 M ФМСФ. Этот же раствор использовали для последующей отмывки фильтров и для инкубации с конъюгатом антител с щелочной фосфатазой (фиг.2).
Гибридный белок Hbc-preS1 выделяют с помощью иммуноафинной хроматографии с использованием сефарозы, конъюгированной с антителами из сыворотки кроликов, иммунизированных HbcAg. Выделенные частицы гибридного белка исследуют с помощью электронной микроскопии. В полученных препаратах обнаружены частицы, по форме и морфологии аналогичные природному кору (фиг.3).
Пример 3. Иммунизация животных гибридным белком HBc-preS1.
Для иммунизации были использованы мыши линии BALB/c, весом 12-15 грамм, полученные из вивария ГНЦ ВБ "Вектор". Животные содержались на стандартном рационе.
Иммунизацию проводят трехкратно, внутримышечно, дозой 5 мкг гибридного белка HBc-preS1 в физиологическом растворе на животное. При первом введении раствор гибридного белка смешивают с полным адъювантом Фрейнда, через 14 суток проводят вторую иммунизацию той же дозой препарата, раствор белка смешивают с неполным адъювантом Фрейнда. Последнюю иммунизацию проводят через 14 суток раствором белка в той же дозе. Забор крови животных для определения титра специфических антител осуществлялся через 14 дней после последней иммунизации. В каждой группе было по 10 животных.
Иммуноферментный анализ.
Специфичность образующихся антител в сыворотках мышей анализируют с помощью ИФА. Для выявления антител против HBc-preS1 используют очищенные рекомбинантный белок HBcAg-preS1 и синтетические пептиды с аминокислотной последовательностью эпитопа preS1 (27-37). Рекомбинантный белок (10 нг на лунку) сорбируют на планшеты в течение ночи в растворе 15 mM Na2СО3, 35 mM NaHCO3, рН 9,2 при 8°С. Для разведения сывороток и для промывки планшетов используют буфер, содержащий 147 mM NaCl, 3 mM KCl, 10 mM трис-HCl, 0,05% твин-20 (рН 7,2). Инкубацию с сыворотками мышей проводят в течение часа при 37°С.Для выявления антител используют антивидовой конъюгат антител со щелочной фосфатазой. Титр определяют как разведение сыворотки, при котором оптическая плотность двукратно превышает фоновое значение. Используя данные титров сывороток мышей одной группы, вычисляют среднее арифметическое и стандартное отклонение. Результаты иммуноферментного анализа представлены на фиг.4А.
Для выявления антител, специфичных эпитопу preS1 (27-37) биотинилируют синтетические пептиды с аминокислотной последовательностью эпитопа preS1 (27-37 DHQLDPAFRAN). Стрептовидин (10 нг на лунку) сорбируют на планшеты в течение ночи в растворе 15 mM Na2СО3, 35 mM NaHCO3, рН 9,2 при 8°С. Для связывания биотинилированных пептидов и разведения сывороток используют буфер 147 mM NaCl, 3 mM KCl, 10 mM трис-HCl, 0,1% казеин (рН 7,2). Для промывки планшетов используют 147 mM NaCl, 3 mM KCl, 10 mM трис-HCl, 0,05% твин-20 (рН 7,2). Пептиды сорбируют в течение часа при 37°С (100 нг/лунку). Инкубацию с сыворотками мышей проводят в течение часа при 37°С. Для выявления антител используют антивидовой конъюгат антител со щелочной фосфатазой и краситель SIGMA FAST p-nitrophenyl phosphat. Результаты оценивают на фотометре для микропланшет BioRad модель 680 на длине волны 405 нм. Титр определяют как разведение сыворотки, при котором оптическая плотность двукратно превышает фоновое значение. Используя данные титров сывороток мышей одной группы, вычисляют среднее арифметическое и стандартное отклонение. Результаты иммуноферментного анализа представлены на фиг.4Б.
Таким образом, получен рекомбинантный штамм Е.coli DH5αF'/pUC8HBc-preS1, продуцирующий белок HbcAg, содержащий иммуноактивный эпитоп района preS1 поверхностного белка вируса гепатита В. Показано, что иммунизация данным белком, очищенным из клеток Е.coli, стимулирует синтез антител, специфичных району 27-37 аминокислотных остатков района preS1 поверхностного белка вируса гепатита В. Полученный белок HBc-preS1 может быть использован в качестве компонента вакцин против вируса гепатита В.
Литература
1. Clarke B.E., Newton S.E., Carrol A.R., Francis M.J., Appleyard G., Syred A.D., Highfild P.E., Rowlands D.J., Brown F. // Improved immunogenicity of a peptide epitope after fusion to hepatitis В core protein. // Nature - 1987. - V.330. - P.381-384.
2. Francis M.J., Hastings G.Z., Brown A.L., Grace K.G., Rowlands D.J., Brown F., Clarke B.E. Immunological properties of hepatitis В core antigen fusion proteins. // Proc. Natl. Acad. Sci. USA. - 1990. - V.87. - P.2545-2549.
3. Milich D.R., McLachlan A.// The nucleocapsid of hepatitis В virus is both a T cell-independent and a T-cell-dependent antigen. // Science. - 1986. - V.12. - P.1398-401.
4. Brown A.L., Francis M.J., Hastings G.Z., Parry N.R., Barnett P.V., Rowlands D.J., Clarke B.E. // Foreign epitopes in immunodominant regions of hepatitis В core particles are highly immunogenic and conformationally restricted. // Vaccine. - 1991. - V.9(8). - P.595-601.
5. Clarke B.E., Brown A.L., Grace K.G., Hastings G.Z., Brown F., Rowlands D.J., Francis M.J. // Presentation and immunogenicity of viral epitopes on the surface of hybrid hepatitis В virus core particles produced in bacteria. // J.Gen. Virol. - 1990. - V.71. - P.1109-17.
6. Pumpens P., Grens E. // HBV core particles as a carrier for В cell / T cell epitopes. // Intervirology. - 2001. - V.44. - P.98-114.
7. Schodel F., Milich D.R., Will H. // Hepatitis В virus nucleocapsid / pre-S2 fusion proteins expressed in atenuated Salmonella for oral vaccination. // J.Immunol. - 1990. - V.145. - №12. - P.4317-4321.
8. Pumpens P., Borisova G.P., Crowther R.A., Grens E. // Hepatitis В virus core particles as epitope carriers. // Intervirology. - 1995. - V.38. - P.63-74.
9. Argos P., Fuller S.D. // A model for the hepatitis В virus core protein: prediction of antigenic sites and relationship to RNA virus capsid. // EMBO J. - 1988. - V.7. - P.819-824.
10. Макеева Н.В., Калинина Т.Н., Худяков Ю.Е., Самошин В.В., Смирнова Е.А., Семилетов Ю.А., Павлюченкова Р.Н., Кадошников Ю.Н., Смирнов В.Д. // Гетерологичные эпитопы в центральной части core-белка вируса гепатита В. // Мол. биология. - 1995. - Т.29. - С.211-224.
11. Coursaget P., Lesage G., Le-Cann P., Mayelo V., Bourdil С. // Mapping of linear B-cell epitopes of hepatitis В surface antigen. // Res. Virol. - 1991. - V.142(6). - P.461-467.
12. Kuroki К, Floreani M, Mimms LT, Ganem D. // Epitope mapping of the PreS1 domain of the hepatitis В virus large surface protein // Virology. - 1990. - V.176(2). - P.620-624.
13. Xin Z.T., Liu C., Dong В., Gao Y.P., Shao N.S., Liu W., Zhang J., Dong J., Ling S.G., Xue Y.N. // A subtractive fluorescence-activated cell-sorting strategy to identify mimotopes of HBV-preS protein from bacterially displayed peptide library. // J Immunol Methods. - 2004. - V.293(1-2). - P.13-21.
14. Маниатис Т., Фрич Э., Сэмбрук Дж. // Методы генетической инженерии. Молекулярное клонирование. - М.: Мир. - 1984.
15. Hanna Z., Fregau C., Prefontaine G., Brousseau R. // Construction of a family of universal expression plasmid vectors. // Gene. - 1984. - V.30. - P.247-250.
16. Патент РФ №2112039, кл. С 12 N 15/51, опубл. БИ №15 от 27.05.98.
17. Патент РФ №1651555, кл. С 12 N 15/51, опубл. БИ №10 от 30.05.94.
18. Laemmly W.K. // Cleavage of structural proteins during the assembly of the heard of bacteriophage T4. // Nature. - 1970. - V.227. - P.680-685.
Изобретение относится к микробиологической и медицинской промышленности, генной и белковой инженерии. Предложена рекомбинантная плазмидная ДНК pUC8HBc-preS1. Плазмида кодирует ген химерного белка корового антигена вируса гепатита В, несущего эпитоп района preS1 (27-37 а.о.) поверхностного белка вируса гепатита В. Предложен также штамм бактерий Escherichia coli DH5αF'/pUC8HBc-preS1, несущий такую плазмиду и продуцирующий гибридный белок HBc-preS1. Изобретение может быть использовано в медицине. 2 н.п. ф-лы, 4 ил.
плазмидный вектор pUC18 размером 2686 п.о., раскрытый по сайтам EcoRI BamHI и содержащий ген ампициллин резистентности, сайт инициации репликации плазмиды pBR322, часть lac-гена из М13mp18, промотор Plac, полилинкерную последовательность из М13mp18, уникальные сайты рестрикции с координатами Hind III (677), Pst I (673), Afl III (2956), Cfr10 I (1983), Ssp I (1263);
BamHI EcoRI - фрагмент ДНК плазмиды р7,5С размером 890 п.о., содержащий ген химерного корового белка вируса гепатита В, несущего эпитоп района preS1 (27-37 а.о.) поверхностного белка вируса гепатита В в районе, соответствующем 81-82 аминокислотным остаткам HBcAg;
генетические маркеры:
bla-ген ампициллин резистентности (ген b-лактамазы), определяющий устойчивость к ампициллину при трансформации клеток Е.coli;
| МАКЕЕВА Н.В | |||
| и др., "Гетерологичные эпитопы в центральной части core-белка вируса гепатита В", Мол | |||
| биология, 1995, т.29, с.211-224 | |||
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PKHBC-33, СОДЕРЖАЩАЯ ГЕН КОРОВОГО АНТИГЕНА ВИРУСА ГЕПАТИТА B, ВКЛЮЧАЮЩЕГО ПОСЛЕДОВАТЕЛЬНОСТЬ ГЛАВНОГО НЕЙТРАЛИЗУЮЩЕГО ЭПИТОПА ПОВЕРХНОСТНОГО АНТИГЕНА ВИРУСА ГЕПАТИТА B (137 - 147), И ШТАММ БАКТЕРИЙ ESCHERICHIA COLI - ПРОДУЦЕНТ КОРОВОГО АНТИГЕНА ВИРУСА ГЕПАТИТА B С ЭКСПОНИРОВАННЫМ НА ЕГО ПОВЕРХНОСТИ ГЛАВНЫМ НЕЙТРАЛИЗУЮЩИМ ЭПИТОПОМ ПОВЕРХНОСТНОГО АНТИГЕНА ВИРУСА ГЕПАТИТА B (137 - 147) | 1995 |
|
RU2112039C1 |

