Изобретение относится к области молекулярной биологии, биотехнологии и лабораторной техники и может быть использовано для нахождения неизвестных гидрофобных лигандов липидпереносящих белков (ЛПБ), установления липидсвязывающей и липидпереносящей активности белков по отношению к определенному гидрофобному лиганду и изучения липид-белковых взаимодействий in vitro. Способ может быть применен в практике лабораторных работ для поиска неизвестных липидпереносящих белков и их природных лигандов, а также для анализа их биохимических свойств; в медицине для установления и исследования отклонений в липидном обмене в патологически измененных тканях и органах; в сельскохозяйственной биотехнологии для поиска новых липидпереносящих белков и гидрофобных веществ, важных в сельскохозяйственной практике. Найденные белки могут в дальнейшем быть изменены с использованием методов генетической инженерии и молекулярной биологии с целью повышения в продуценте (как животном, так и растительном) содержания жиров и жирорастворимых витаминов.
Известно, что динамика обмена липидов, включающая их спонтанное перемещение в мембране и переход в состав новой мембраны при индуцируемом переносе под действием специальных липидпереносящих белков (ЛПБ), играет важную роль в жизнедеятельности клетки. ЛПБ определяют формирование мембранных везикул и соответственно контролируют внутриклеточный транспорт и секрецию [Геннис Р. Биомембраны: Молекулярная структура и функции: Пер. с англ. М.: Мир, 1997; стр.9-48].
Изучение функций ЛПБ является одной из важных задач современной биологии и медицины. Интерес к этой проблеме с одной стороны объясняется тем, что различные липиды являются одним из основных «строительных блоков» клетки и энергозапасающим депо организма, а с другой - гидрофобные лиганды играют важную роль в передаче клеточных сигналов, развитии некоторых патологий, связанных с нарушением распределения гидрофобных лигандов, и изменении статуса клеток и тканей (апоптоз, клеточная дифференцировка и т.д.). Так, пациенты, имеющие дефицит по белку PI-TPα, могут демонстрировать генерализованное вырождение нейронов спинного столба, продолговатого мозга и рогов спинного мозга, которое ведет к развитию признаков, подобным тем, которые характерны для прогрессирующих нейродегенеративных заболеваний [Hamilton, В.A.; Smith, D.J.; Mueller, К.L.; Kerrebrock, A.W.; Bronson, R.Т.; van Berkel, V.; Daly, M.J.; Kruglyak, L.; Reeve, M.P.; Nemhauser, J.L.; Hawkins, T.L.; Rubin, E.M. and Lander, E.S. Neuron 18, 711-722; 1997]. CRALBP-дефицитные пациенты демонстрируют аутосомальную рецессивную депигментацию сетчатки, вызванную нарушениями в транспорте 11-cis-ретинальдегида и 11-cis -ретинола [Maw, M.A.; Kennedy, В.; Knight, A.; Bridges, R.; Roth, K.E.; Mani, E.J.; Mukkadan, J.K.; Nancarrow, D.; Crabb, J.W. and Denton, M. J. Nat. Genet. 17, 198-200; 1997]. У пациентов, страдающих расстройством координации движения (атаксией), вызванной дефицитом витамина E (AVED), наличествуют мутации в гене, кодирующем белок α-ТТР. Эти мутации приводят к атаксии, потере нейронов, атрофии сетчатки, массовому накоплению липофусцина («пигмента старения») и пигментному ретиниту [Yokota, Т.; Uchihara, Т.; Kumagai, J.; Shiojiri, Т.; Pang, J.J.; Arita, M.; Arai, H.; Hayashi, M.; Kiyosawa, M.; Okeda, R. and Mizusawa, H.J. Neurol. Neurosurg. Psychiatry 68,521-525; 2000].
Важный этап установления функции ЛПБ - нахождение их специфического природного липидного лиганда. Некоторые предположения в этом отношении можно выдвинуть на основе анализа первичной структуры белка, идентифицировав характерные липидсвязывающие домены. Однако решающее значение имеет эксперимент, демонстрирующий взаимодействие ЛПБ с предполагаемым лигандом и свидетельствующий о наличии у исследуемого полипептида транспортной активности.
В настоящее время определение неизвестного природного гидрофобного лиганда ЛПБ чаще проводится по следующей схеме:
1. Выделение изучаемого белка из природного источника или получение рекомбинантного аналога этого белка. Его очистка и идентификация.
2. Проведение экспериментов по связыванию исследуемого белка с набором различных заранее известных липидных лигандов, как правило, нанесенных на твердую подложку (нитроцеллюлозу). Выбор из оттестированных лигандов тех, которые проявили наибольшее сродство к белку.
3. Создание модельной системы in vitro, в которой возможно зарегистрировать липидпереносящую активность исследуемого белка. В большинстве случаев для этой цели создаются донорные и акцепторные искусственные липосомы. В состав донорной липосомы вводят переносимый лиганд, акцепторная липосома его не содержит. Донорные и акцепторные липосомы соединяют в общем объеме, а в водную фазу вводят исследуемый белок. При наличии у белка липидпереносящей активности по отношению к лиганду белок за время совместной инкубации с липосомами индуцирует перенос липида от донорной липосомы к акцепторной.
4. В дальнейшем донорные и акцепторные липосомы разделяют (чаще - ультрацентрифугированием) и тем или иным способом устанавливают наличие исследуемого липида во фракции акцепторных липосом. На основе этого делают вывод о том, что белок является переносчиком определенного липида [Rogers D.P, Bankaitis VA. Int Rev Cytol. 2000; 197, 35-81].
Известен способ определения переносящей активности белка SPF (супернатантного белкового фактора) из печени крысы в отношении сквалена доказательства того, что белок SPF (супернатантный белковый фактор) из печени крысы является переносчиком сквалена, заключающийся в том, что создаются искусственные донорные липосомы, состоящие из фосфатидилхолина желтка куриного яйца (2,5 мкМ), дицетилфосфата (0,25 мкМ), меченного тритием сквалена (2,0×106 расп./мин) и меченного изотопом углерода 14С триглицеро[карбоксил-14С]олеата (1,0×106 расп./мин). Последний используется в качестве маркера неспецифического переноса сквалена, который происходит за счет слипания донорных и акцепторных мембран. В этом способе в качестве акцептора используется неочищенная фракция «тяжелых» клеточных мембран, полученных при разрушении тканей печени крысы. Подготовленные таким образом липосомы инкубируют при 37°С в течение 30 мин с фракцией «тяжелых» мембран. Объем инкубационной смеси составляет 1 мл (1 мл буферного раствора SET (0.25 M сахароза, 1 мМ EDTA и 10 мМ Трис-HCl, рН 7.4) + 0,05 мг SPF). После инкубации смесь центрифугируется при ускорении 15 тыс. g и отбирается 0,8 мл надосадочной жидкости, в которой и содержатся неосевшие при центрифугировании донорные липосомы. После этого рассчитывают количество сквалена, перешедшего под действием белка во фракцию тяжелых мембран. Это значение получают как разность между исходным количеством сквалена в донорных липосомах и остаточным количеством после инкубации. Введя поправку на процент неспецифического слипания донорных и акцепторных мембран, и, как следствие, - попадания в акцепторную фракцию сквалена без помощи белка, устанавливают, что за время эксперимента под действием SPF до 35% скалена перешло в акцепторную фракцию тяжелых мембран. Это доказывает, что белок SPF обладает липидтранспортной активностью в отношении сквалена и является его гидрофобным лигандом [Shibata N., Arita M., Misaki Y., Dohmae N., Takio K., Ono Т., Inoue K. and Arai H. PNAS 2001. 98(5), 2244-2249].
К недостаткам этого метода можно отнести то, что из-за нахождения донорных и акцепторных мембран в одном инкубационном объеме возникает необходимость использовать для детекции переноса липидов их радиоактивно меченные производные, а для коррекции расчетов необходимо вводить в состав донорной мембраны второй радиоактивный липид с другим типом метки.
Описанный способ приводит к тому, что весь лиганд, специфически доставленный белком в акцепторные мебраны, фактически находится в осадке. О его присутствии в акцепторе можно судить только по возникшей недостаче лиганда в донорной мембране. Выделить же индивидуальный перенесенный лиганд из осадка неочищенной фракции «тяжелых» мембран практически невозможно, так как его количество там очень мало по сравнению с другими (неизвестными) липидами, формирующими эту фракцию.
Известен наиболее близкий по технической сущности к предлагаемому изобретению способ определения липидпереносящей активности белка из печени крупного рогатого скота и установления его гидрофобного лиганда, заключающийся в том, что на поверхности водного раствора тестируемого белка в контролируемых условиях формируется липидный монослой, включающий предполагаемый гидрофобный лиганд. Способ применяют для доказательства того, что белок из печени крупного рогатого скота индуцирует обмен фосфатидилхолина между искусственно созданным липидным монослоем и водной фазой. С использованием данного способа также показывают, что этот белок индуцирует обмен фосфатидилхолина между липидным монослоем и искусственными липосомами.
При реализации способа осуществляют обмен фосфатидилхолина между монослоем, состоящим из 16:0/18:1[14C-метилхолин]фосфатидилхолина и белком, растворенным в нижнем фазовом водном слое. 20,6 нМ фосфатидилхолина распределяют монослоем на поверхности водного раствора белка. Буфер содержит 10-2 М Трис-ацетата при рН 7,4. Обменивающий белок вводится под липидный слой в водную фазу. Равновесие достигается при переходе 49,8% 14C-меченного фосфатидилхолина в жидкость. Учитывая количество веществ, использованных в эксперименте, делается вывод, что с одной молекулой белка взаимодействует одна молекула липида.
Для контроля поверхностного натяжения при формировании монослоя и изменения поверхностного натяжения при переходе липида в водный раствор (в составе водорастворимого комплекса с белком) было необходимо использовать умножитель сигналов, выпрямитель и самописец. Параллельно было необходимо контролировать падение радиоактивности поверхностного слоя липида и нарастание радиоактивности водной фазы. Для этих целей в составе устройства предусмотрен радиометр. Равновесие при переходе липида в раствор достигается примерно через три часа его инкубации с белком [Demel R.A., Wirtz K.W.A., Kamp H.H., van Kessel G.W.S.M. and van Deenen L.L.M. Nature New Biology 1973. 246, 102-105].
Однако относительная сложность технической реализации метода и использование радиоактивных липидных производных ограничивает его применение. Данный метод не позволяет изолировать и идентифицировать неизвестный гидрофобный лиганд из сложной смеси липидов и служит только для демонстрации липидсвязывающей (липидпереносящей) активности определенного белка по отношению к конкретному, заранее известному, липиду, который и вводится в формируемый липидный монослой.
Задачей предлагаемого изобретения является разработка нового упрощенного способа определения липидпереносящей способности белков.
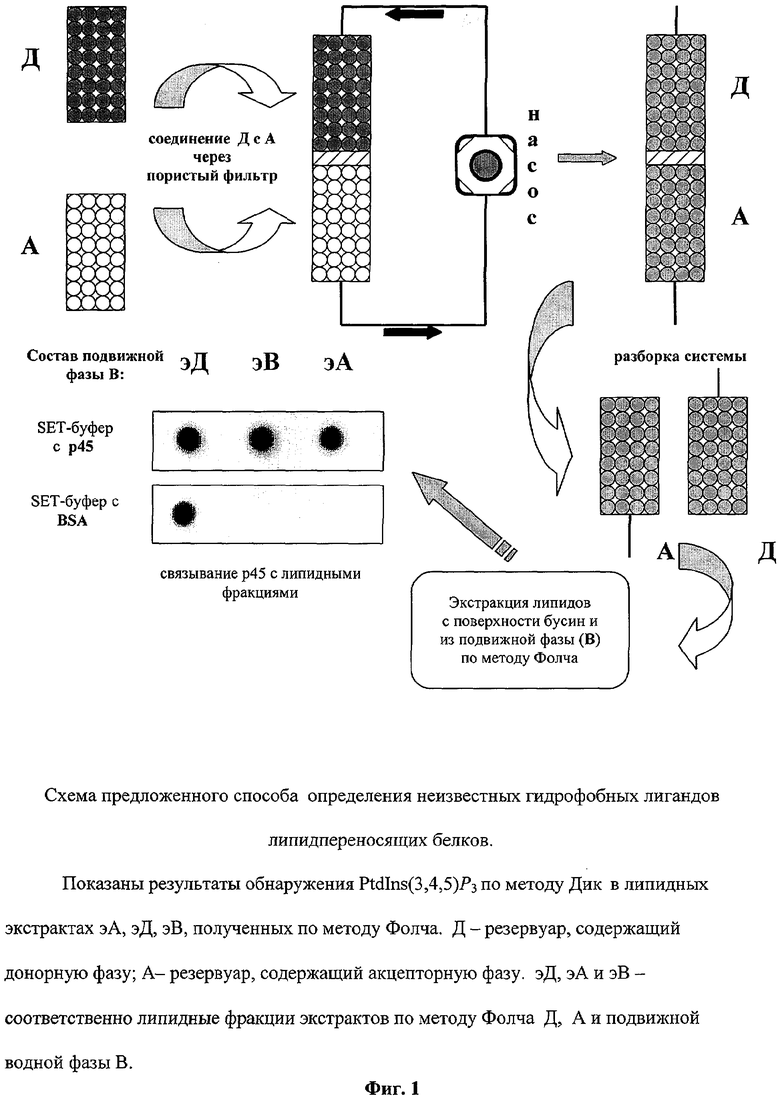
Поставленная задача решается за счет того, что способ определения липидпереносящей способности белков состоит в том, что предварительно готовят донорную и акцепторную фазы, для чего наполнитель в форме стеклянных шариков, покрытых гидрофильным сополимером, помещают в две равные емкости, одну из которых для формирования акцепторной фазы заливают раствором базового липида в смеси смешивающегося и не смешивающегося с водой органических растворителей в объемном соотношении 1:4, для формирования донорной фазы вторую емкость заливают вышеуказанным раствором, дополнительно содержащим переносимый липид, систему выдерживают в течение 10-15 минут, сливают растворы и промывают наполнитель 10-кратньм количеством SET-буфера, сформированные донорные и акцепторные фазы, находящиеся в равных емкостях, соединяют между собой пористой перегородкой и в направлении от донорной к ацепторной фазе пропускают раствор исследуемого белка в SET-буфере по замкнутому контуру в течение 45-60 минут с последующей экстракцией акцепторной фазы и раствора белка смесью равных частей хлороформа, метанола и серного эфира, а определение переносимого липида проводят известным методом.
Техническая реализация способа предусматривает наличие двух емкостей, содержащих наполнитель - стеклянные шарики в качестве твердой подложки для полимерного слоя. На их поверхности создают слой гидрофильного сополимера. Подготовленными таким образом шариками заполняются емкости. Оставшееся свободным от шариков пространство в каждой емкости заполняют специальным раствором липидов в смеси органических растворителей, один из которых смешивается с водой, а другой - не смешивается. Содержимое одной из емкостей, содержащей наполнитель - покрытые сополимером шарики (твердой подложкой), обрабатывают смесью органических растворителей, содержащей базовый липид, формирующий липидный слой (фосфатидилхолин (PtdCho) и тестируемый на способность переноситься белком липид (фосфатидилинозит-3,4,5-трифосфат (PtdIns(3,4,5)P3). Другую емкость обрабатывают подобным образом, за исключением того, что в смеси органических растворителей присутствует только PtdCho. Первая из описанных емкостей, таким образом, будет содержать «донорную фазу («Д») - источник предполагаемого переносимого лиганда, а вторая - «акцепторную» фазу («А»), куда под действием белка будет переноситься лиганд.
Липидный слой формируется на границе раздела фаз воды, связанной с гидрофильным сополимером и несмешивающегося с водой органического растворителя. После формирования на подложке липидных слоев каждую емкость независимо промывают 10 объемами SET-буфера (0.25 М сахароза, 1 мМ EDTA и 10 мМ Трис-HCl, рН 7.4). Затем обе емкости герметично соединяют через пористую перегородку, предотвращающую непосредственный контакт шариков. Через собранную систему пропускают раствор водорастворимого липидсвязывающего белка молекулярной массой 45 кДа из обонятельного эпителия крысы («р45») в SET-буфере. Этот раствор представляет собой подвижную фазу «В». Направление циркуляции подвижной фазы показано на фиг.1 черными стрелками. Через 45-60 мин водную фазу сливают и систему разбирают. Шарики, заполнявшие емкости А и Д, а также собранную водную фазу В экстрагируют согласно методу Фолча [Folch J., Less M. and Sloane G.J. Biol. Chem. 1957, 226, 497-509]. Полученные таким образом липидные фракции (обозначенные далее эА, эД и эВ) исследуют по методу Дик [Deak M., Csamayor A., Currie R.A., Downes C.P. and Alessia, D.R, FEBS Lett. 1999, 451, 220-226], используя для детекции переносимого лиганда связывающийся с ним р45. Обнаружение р45 на нитроцеллюлозных фильтрах осуществляют с помощью моноклональных антител к этому белку и фотолюминесцентной системы (ECL «Amersham Biosciences» Великобритания).
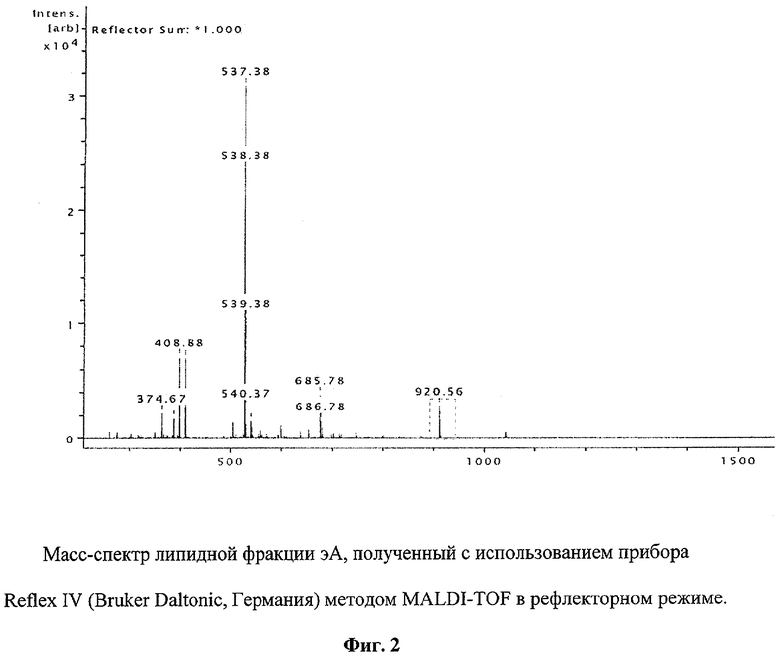
Точную масс-спектрометрическую идентификацию микроколичеств переносимого лиганда во фракциях эВ и эА проводят с использованием прибора Reflex IV (Bruker Daltonic, Германия) методом MALDI-TOF в рефлекторном режиме.
Техническим результатом заявленного способа является возможность отказаться от использования радиоактивных аналогов липидов, используемых для контроля неспецифического (не обусловленного белком) переноса, так как донорный и акцепторный слои пространственно разнесены и не контактируют непосредственно. Это приводит к тому, что исследуемый гидрофобный лиганд в ходе эксперимента может включиться в состав акцепторного липидного слоя, только предварительно соединившись с белком, перейдя в виде этого водорастворимого комплекса в водную фазу и попав с ней в объем, содержащий акцепторный липидный слой.
Кроме того, контролируя липидный состав фаз, экстрагируя после эксперимента липиды акцепторной фазы и липиды, входящие в водную фазу в виде белок-липидного комплекса, можно вычленить индивидуальный липид, являющийся лигандом липидпереносящего белка. Способ дает возможность использовать при формировании донорной фазы сложной смесью липидов неизвестного состава (к примеру - тотальный липидный экстракт ткани). Тогда в ходе эксперимента в состав акцепторного слоя будет включен только определенный липид, отобранный (отселектированный) используемым в эксперименте белком. Очевидно, что в этом случае при анализе липидного состава акцепторного слоя любой вновь появившейся липид, не входивший в состав акцепторной фазы при ее формировании до эксперимента, будет искомым природным лигандом белка.
Другим важным преимуществом заявляемого технического решения является очевидная возможность изучать влияние на процесс переноса липида белком различных агентов, вводимых в подвижную фазу, и физико-химических условий (малых регуляторных молекул, других белков, рН среды, температуры и т.д.).
В дополнение к вышеперечисленному, введя в донорный слой конкретный известный липид, можно проводить тестирование сложной белковой фракции (например - тотального белкового экстракта ткани) на наличие липидпереносящей активности по отношению к этому липиду. Затем, проведя стандартными хроматографическими методами разделение белковой смеси на индивидуальные компоненты, можно вычленить конкретный белок, ответственный за липидтранспортную активность по отношению к изучаемому липиду.
Изобретение иллюстрируется следующими примерами.
Пример 1. Для реализации способа две емкости заполняют стеклянными шариками (диаметром 2,5 мм), которые служат твердой подложкой для полимерного слоя (фиг.1). В качестве емкостей применяют пустые микроколонки объемом 5 мл (кат. № # А809В) от системы выделения ДНК Wizard® Plus Minipreps («Promega», США). Для создания стабильного слоя связанной воды на поверхность шариков наносят гидрофильный сополимер, представляющий собой 10% полиакриламидный гель с сополимеризованным полиэтиленглем 40000 (10% от массы акриламида) в SET-буфере состава: 0.25 М сахароза, 1 мМ EDTA и 10 мМ Трис-HCl, рН 7.4. Подготовленными таким образом шариками заполняют обе емкости. Затем емкости заполняют индивидуальным для каждого резервуара раствором липидов в смеси органических растворителей, один из которых смешивается с водой, а другой - не смешивается (метанол-хлороформ в объемном соотношении 1:4). Раствор для емкости Д содержит фосфатидилхолин (PtdCho) - липид, формирующий базовый гидрофильный слой, и фосфатидилинозит-3,4,5-трифосфат (PtdIns(3,4,5)P3, предполагаемый гидрофобный лиганд липидсвязывающего белка р45, в соотношении 10:1 (соответственно 100 мкг:10 мкг). Раствор для емкости А содержит только PtdCho (100 мкг). Липидный слой формируется на границе раздела фаз связанной с сополимером воды и гидрофобного органического растворителя. После нанесения липидов и формирования на поверхности сополимера гидрофобных слоев каждую емкость независимо промывают 10 объемами SET-буфера. Затем оба резервуара герметично соединяют через пористую перегородку, предотвращающую непосредственный контакт шариков. В качестве пористой перегородки используют полиамидный диффузионный диск диаметром 8 мм и толщиной 1,5 мм от адаптера хроматографической колонки «IEC» фирмы «Whatman» (США).
Через собранную систему пропускают 50 нМ раствор р45 в SET-буфере. Этот раствор представляет собой подвижную фазу В. Скорость протока ≈5 свободных объемов системы в час (˜10 мл/ч при 30°С). Направление циркуляции подвижной фазы показано на фиг.1 черными стрелками.
Через 1 ч водную фазу сливают и систему разбирают. Шарики, заполнявшие емкости А и Д, а также собранную водную фазу В экстрагируют согласно методу Фолча. Для этого извлеченные из емкостей шарики помещают в смесь равных частей хлороформа, метанола и серного эфира. Объем экстрагирующей смеси равняется пяти объемам донорной и акцепторной фаз (по 25 мл смеси на каждую фазу в отдельности). Интенсивно встряхивают в течение 10 мин. Дают отстояться и фильтруют через бумажный фильтр. Отфильтрованную смесь, содержащую экстрагированные липиды, переносят в емкости для упаривания и сушат в вакуумном экстракторе. Пленку липидного осадка с внутренней поверхности этих емкостей количественно смывают смесью метанола и хлороформа в объемном соотношении 2:1. Полученные таким образом липидные фракции (обозначенные далее эА, эД и эВ), исследуют по методике Дик [Deak М., Csamayor A., Currie R.A., Downes C.P. and Alessia, D.R. FEBS Lett. 1999, 451, 220-226], используя для детекции перенесенного лиганда связывающийся с ним р45. Для этого полученные по методике Фолча липидные экстракты фаз Д, А и В наносят на нитроцеллюлозный фильтр Hybond-C Extra (Echelon, США) в смеси хлороформ : метанол : вода в объемных отношениях 1:2:0,8. Образцы подсушивают на воздухе при комнатной температуре в течение 1 ч. Для предотвращения неспецифической сорбции белка мембрану обрабатывают блокирующим буферным раствором «ТБС + Твин-20» (50 мМ Tris-HCl-буфер, рН 7,0-8,0, с 150 мМ NaCl и 0,5% Твин-20), содержащим 3% бычьего сывороточного альбумина (BSA), свободного от жирных кислот. Обработка производится в течение 2 ч при комнатной температуре. Затем мембрану переносят в свежий буферный раствор того же состава, содержащий р45 в концентрации 30 нМ, и инкубируют в течение часа при комнатной температуре с перемешиванием. После этого мембрану промывают «ТБС + Твин-20» (3 раза по 10 мин) и обрабатывают при 37°С первичными поликлональными антителами к р45 в разведении антител 1:750. Затем мембрану трижды отмывают в «ТБС + Твин-20» и обрабатывают в течение 1 ч при 37°С вторичными антикроличьими антителами, конъюгированными с пероксидазой хрена в разведении 1:5000. Проявление связавшегося в зонах нанесения экстрактов р45 осуществляют методом хемолюминесцентной детекции (ECL-kit, Amersham, Великобритания). Сорбция р45 в точках нанесения липидных экстрактов после эксперимента свидетельствует о наличии в этих фракциях PtdIns(3,4,5)P3. Таким образом, применение способа демонстрирует, что во время эксперимента липидсвязывающийся белок р45 индуцирует перераспределение PtdIns(3,4,5)P3 между резервуарами Д и А в системе (на фиг.1 это отражено появлением серого окрашивания резервуара А и уменьшением интенсивности серого цвета в резервуаре Д). Это позволяет сделать вывод о том, что липидпереносящая способность белка р45 в отношении PtdIns(3,4,5)P3 определена и доказана.
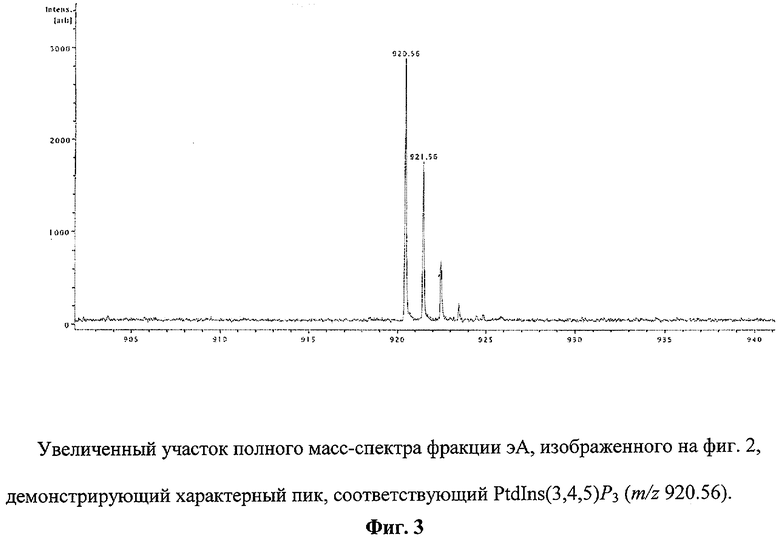
Пример 2. Точную масс-спектрометрическую идентификацию переносимого лиганда во фракциях эВ и эА проводят с использованием прибора Reflex IV (Bruker Daltonic, Германия) методом MALDI-TOF в рефлекторном режиме. Во фракции, полученной при экстракции липидов с поверхности бусин из резервуара А, как и ожидалось, наряду с пиком, соответствующим PtdCho (m/z 537.38), обнаруживается пик, соответствующий PtdIns(3,4,5)P3 (m/z 920.56) (фиг.2). На фиг.3 этот пик показан крупно. Присутствие в масс-спектре фракции эВ только уникального пика с m/z 920.56, соответствующего молекулярной массе переносимого лиганда, указывает на наличие в этой фракции PtdIns(3,4,5)P3, связываемого р45.
Пример 3. В условиях, аналогичных Примеру 1, отличающихся тем, что в состав водной фазы В вместо р45 входил бычий сывороточный альбумин (BSA) (в концентрации 50 нМ), было определено, что этот белок не приводит к перераспределению PtdIns(3,4,5)P3 между фазами Д и А и, следовательно, не проявляет липидпереносящей активности в отношении этого липида (фиг.1).
| название | год | авторы | номер документа |
|---|---|---|---|
| Липопротеиновые комплексы и их получение и применения | 2012 |
|
RU2627173C2 |
| Способ получения водонерастворимых биологически активных соединений | 1977 |
|
SU689200A1 |
| СПОСОБ КОНСТРУИРОВАНИЯ, ПРОВЕРКИ И ПРИМЕНЕНИЯ АССОЦИАТОВ МАКРОМОЛЕКУЛ И КОМПЛЕКСНЫХ АГРЕГАТОВ С ПОВЫШЕННОЙ ПОЛЕЗНОЙ НАГРУЗКОЙ И КОНТРОЛИРУЕМОЙ СТЕПЕНЬЮ АССОЦИАЦИИ/ДИССОЦИАЦИИ | 1998 |
|
RU2211027C2 |
| ПЕГИЛИРОВАННЫЕ ЛИПОСОМЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2796539C2 |
| ЦЕЛЕВЫЕ ВЕЗИКУЛЯРНЫЕ КОНСТРУКЦИИ ДЛЯ ЗАЩИТЫ КЛЕТКИ И ДЛЯ ЛЕЧЕНИЯ ИНФЕКЦИЙ H. PYLORI | 2000 |
|
RU2207113C2 |
| НОВЫЕ КОМПОЗИЦИИ ЛИПОСОМ | 2006 |
|
RU2454229C2 |
| Способ доставки гранзима Б в клетки млекопитающих | 2021 |
|
RU2796685C2 |
| ЛИПОСОМАЛЬНЫЕ КОМПОЗИЦИИ И СОДЕРЖАЩИЕ ИХ ТВЕРДЫЕ ПЕРОРАЛЬНЫЕ ЛЕКАРСТВЕННЫЕ ФОРМЫ | 2018 |
|
RU2751192C2 |
| ЛЕКАРСТВЕННАЯ ФОРМА СПЕЦИФИЧЕСКОГО ИММУНОБИОЛОГИЧЕСКОГО ЛЕКАРСТВЕННОГО СРЕДСТВА ДЛЯ ЛЕЧЕНИЯ И ПРОФИЛАКТИКИ ВИЧ ИНФЕКЦИИ И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2014 |
|
RU2600031C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ АДЪЮВАНТНЫХ ВИРОСОМ И АДЪЮВАНТНЫЕ ВИРОСОМЫ, ПОЛУЧАЕМЫЕ УКАЗАННЫМИ СПОСОБАМИ | 2014 |
|
RU2694367C2 |
Изобретение относится к области молекулярной биологии и может быть использовано для определения гидрофобных лигандов липидпереносящих белков, установления липидсвязывающей и липидпереносящей активности белков и изучения липид-белковых взаимодействий in vitro. Способ включает приготовление донорных и акцепторных фаз. Сформированные донорные и акцепторные фазы соединяют между собой пористой перегородкой и пропускают раствор исследуемого белка в SET-буфере с последующей экстракцией акцепторной фазы и раствора белка смесью равных частей хлороформа, метанола и серного эфира. Преимущество предложенного способа состоит в том, что он является упрощенным и менее трудоемким по сравнению с известными способами. 3 ил.
Способ определения липидпереносящей способности белков, состоящий в том, что предварительно готовят донорную и акцепторную фазы, для чего наполнитель в форме стеклянных шариков, покрытых гидрофильным сополимером, помещают в две равные емкости, одну из которых для формирования акцепторной фазы заливают раствором базового липида в смеси смешивающегося и не смешивающегося с водой органических растворителей в объемном соотношении 1:4, для формирования донорной фазы вторую емкость заливают вышеуказанным раствором, дополнительно содержащим переносимый липид, систему выдерживают в течение 10-15 мин, сливают растворы и промывают наполнитель 10-кратным количеством SET-буфера, сформированные донорные и акцепторные фазы, находящиеся в равных емкостях, соединяют между собой пористой перегородкой и в направлении от донорной к акцепторной фазе пропускают раствор исследуемого белка в SET-буфере по замкнутому контуру в течение 45-60 мин с последующей экстракцией акцепторной фазы и раствора белка смесью равных частей хлороформа, метанола и серного эфира.
| DEMEL R.A | |||
| et al | |||
| Phosphatidylcholine exchange protein from beef liver | |||
| NATURE NEW BIOLOGY | |||
| Приспособление для склейки фанер в стыках | 1924 |
|
SU1973A1 |
| ROGERS D.P | |||
| et al | |||
| Phospholipid transfer proteins and physiological functions | |||
| Int | |||
| Rev | |||
| Cytol | |||
| ЩИТОВОЙ ДЛЯ ВОДОЕМОВ ЗАТВОР | 1922 |
|
SU2000A1 |
| WO 03036259, 01.05.2003. | |||

