Настоящее изобретение относится к области лечения иммунодефицита и вирусных инфекций. Более конкретно, настоящее изобретение относится к использованию ингибиторов циклооксигеназы-2 (СОХ-2) или их производных в целях иммуномодуляции для лечения иммунодефицита и вирусных заболеваний, а в частности ВИЧ-инфекций и СПИДа и родственных состояний.
Простагландины играют важную роль в воспалительных процессах, и ингибирование образования простагландинов является главной целью при разработке противовоспалительных лекарственных средств. Нестероидные противовоспалительные средства (НСПВС) ингибируют циклооксигеназу (СОХ), которая представляет собой фермент, участвующий в биосинтезе промежуточных соединений простагландинов из арахидоновой кислоты. В клинической медицине используется несколько НСПВС, включая лекарственные средства, такие как индометацин, пироксикам, теноксикам, диклофенак, мелоксикам, тенидап, изоксикам, ацетилсалициловая кислота, дифлунизал, сулиндак, ибупрофен, напроксен и кетопрофен.
В настоящее время НСПВС являются широко распространенными лекарственными средствами, применяемыми во всем мире.
Эти НСПВС представляют собой клинически эффективные лекарственные средства и обладают жаропонижающим, противовоспалительным и антитромботическим действием. Главными показаниями для этого класса лекарственных средств являются артриты, включая остеоартрит и ревматоидный артрит, костно-мышечные боли и общие болезненные состояния. Однако эти лекарственные средства имеют серьезные побочные эффекты. Наиболее часто встречающимися побочными эффектами являются образование язв и кровотечения в области желудочно-кишечного тракта, ингибирование агрегации тромбоцитов и взаимодействие с другими лекарственными средствами.
В начале 1990-х годов была клонирована вторая изоформа фермента СОХ. В настоящее время эта новая изоформа СОХ известна как СОХ-2 (Vane et al., 1998, Ann. Rev. Pharmacol. Toxicol., 38, р.97-120).
В настоящее время хорошо известны две изоформы СОХ, СОХ-1 и СОХ-2 (недавно было также высказано предположение о существовании СОХ-3). СОХ-1 присутствует в большинстве тканей и может рассматриваться как фермент "домашнего хозяйства". Активность фермента СОХ-1 обеспечивает защиту, например, выстилки желудочно-кишечного тракта. Однако обычно СОХ-2 не присутствует в нормальных тканях, но она продуцируется в процессе воспаления. Некоторые из побочных эффектов НСПВС связаны с ингибированием фермента СОХ-1. НСПВС ингибируют как фермент СОХ-1, так и фермент СОХ-2 (см. таблицы 1-3):
Величины IC50 и отношение СОХ-2/СОХ-1 для различных НСПВС в макрофагах морской свинки, используемой в качестве модели (величины IC50 взяты у Engelhart et al., J. Inflammatory Res., 44, p.422-43, 1995)
(мкмоль/литр)
(мкмоль/литр)
Величины IC50 для различных НСПВС в штатных клетках, ингибирующих СОХ-1 (коровьи эндотелиальные клетки) и СОХ-2 (стимулированные макрофаги)(величины IC50 взяты у Taketo et al., J. National Cancer Institute, 90, p.1529-1536, 1998)
(мкмоль/литр)
(мкмоль/литр)
Ингибирование синтеза рекомбинантного PGH (СОХ-1 и СОХ-2) (величины IC50 взяты у Laneuvill et al., J. Fharm. Exp. Ther., 271, p.927-34, 1994)
(мкмоль/литр)
(мкмоль/литр)
За последние десять лет было идентифицировано несколько новых селективных ингибиторов COX-2 и так называемых "преимущественных" ингибиторов COX-2. Было разработано несколько таких ингибиторов COX-2, и небольшая их часть недавно поступила в продажу. Некоторые из этих новых ингибиторов COX-2 не обнаруживали ингибирования COX-1 в клинических дозах. Интенсивные клинические исследования и клиническая практика по использованию этих ингибиторов COX-2 выявили, что эти новые ингибиторы COX-2 обладают значительными преимуществами с точки зрения их безопасности по сравнению с неселективными НСПВС. Обзорное описание ингибиторов СОХ-2 можно найти, например, у Golden et al., 1999, Osteoarthritis. 25, p.359-379, Mitchel et al., 1999, Brit. J. Pharmacol., 128, p.1121-1132, Lipsky, 1999, Am. J. Med., 106 (5B), p.515-575, Taketo, 1998, J. National Cancer Inst., 90, p.1529-1537, Griswold et al., 1996, Med. Res. Rev., 16, p.181-206 и Reitz et al., 1995, Ann. Rep. Med. Chem., 30, p.179-188.
Другими представляющими интерес публикациями, относящимися к различным ингибиторам СОХ-2, являются, например: Lane, 1997, J. Rheumatol., 24 (suppl. 49), p.20-24, Mehlish et al., 1998, Clin. Pharmacol. Ther., 63, p.1-8, Zhao et al., 1997, Arthritis Rheum., 40 (suppl.), S88, Ehrich et al., 1997, Arthritis Rheum., 40 (suppl.), S93, Maziasz et al., 1997, Arthritis Rheum., 40 (suppl.), S195, Mengle-Gaw et al., 1997, Arthritis Rheum., 40 (suppl.), S93, Morrison, 1999, Clin. Ther., 21, p.943-953, Chan et al., 1995, J. Pharmacol. Exp. Ther., 274, p.1531-37, Riendeau et al., 1997, Br. J. Pharmacol., 121, p.105-117, Black et al., 1999, J. Med. Chem., 42, p.1274-81, Cuo et al., 1996, J. Biol. Chem., 271, p.19134-39, Geiss, 1999, Scand. J. Rheumatol., 109 (suppl.), p.31-37, Warner et al., 1999, PNAS USA, 96, p.7563-68, Bjarnson et al., 1977, Scand. J. Gastroenterol., 32, p.126-130, Danneberg et al., 1999, Semin. Oncol., 26, p.499-504, Mitchell et al., 1993, PNAS USA, 90, p.11693-97, Futaki et al., 1994, Prostaglandins, 47, p.55-9, Futaki et al., 1993, J. Pharm. Pharmacol., 46, p.753-5, Masferrer et al., 1994, PNAS USA, 91, p.3228-32, Klein et al., 1994, Biochem. Pharmacol., 48, p.1605-10, Reitz et al., 1994, J. Med. Chem., 37, p.3878-81, Seibert et al., 1994, PNAS USA, 91, p.12013-17, Klein et al., 1996, Biochem. Pharmacol., 51, p.285-90, Nantal et al., 1998, 9th Intern. Conference Inflamm. Res. Assoc., Nov 1-5, Pennig et al., 1997, J. Med. Chem. 40, p.1347-65 and Puig et al., 2000, J. Med. Chem., 43, p.214-223.
Ингибиторы СОХ-2 представляют собой относительно широкую группу соединений с различными структурами. Соединения, которые селективно ингибируют СОХ-2, за последнее десятилетие описаны во многих патентных документах. Некоторыми из них являются WO 94/26781, WO 94/20480, WO 94/13635, WO 95/00501, WO 94/27980, WO 94/15932, WO 95/21817, WO 95/15316, WO 96/06840, WO 96/03388, WO 96/03387, WO 96/03392, WO 96/25405, WO 96/24584, WO 96/03385, WO 96/16934, WO 98/41516, WO 98/43966, WO 99/12930, ЕРО 673366, WO 98/41511, WO 98/47871, WO 99/20110, WO 99/23087, WO 99/14194, WO 99/14195, WO 99/15513 и WO 99/15503, и патенты США №№ 5380738, 5344991, 5393790, 5434178, 5474995, 5475018 и 5510368.
В настоящее время были получены два соединения, рофекоксиб (4-(4-метилсульфонил)фенил-3-фенил-2(5Н)-фуранон) (I), выпускаемый под товарным знаком Vioxx® и селекоксиб 4-(5-(4-метилфенил)-3-(трифторметил)-1Н-пиразол-1-ил)бензолсульфонамид (II), выпускаемый под товарным знаком Celebra®:
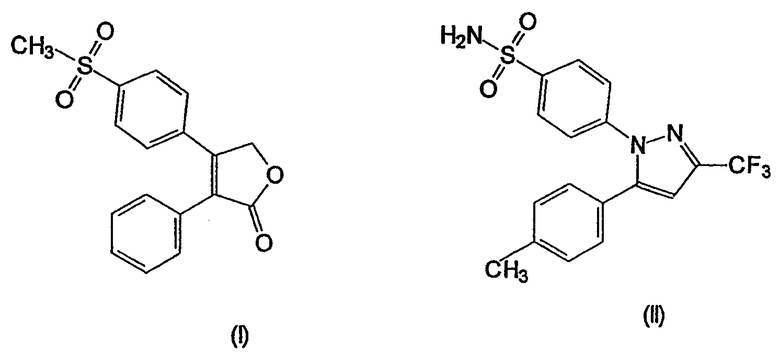
Рофекоксиб описан в WO 93/0500501, Merck Frosst Canada и кроме того, в работах Morrison, 1999, Clin. Ther., 21, p.943-953, Chan et al., 1995, J. Pharmacol. Exp. Ther., 274, p.1531-37 и Nantel et al., 1998, см.выше.
Целекоксиб описан в работах Geiss, 1999, Scand. J. Rheumatol., 109 (suppl.), p.31-37 и Penning et al., 1997, J. Med. Chem. 40, p.1347-65. Как указывалось, целекоксиб является в 375 раз более селективным по отношению к СОХ-2, чем по отношению к СОХ-1.
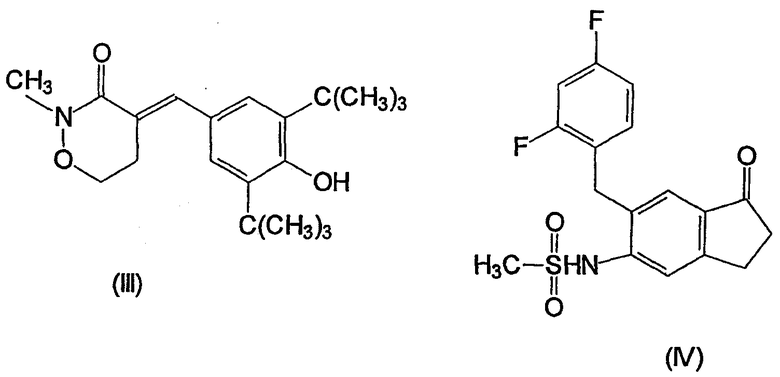
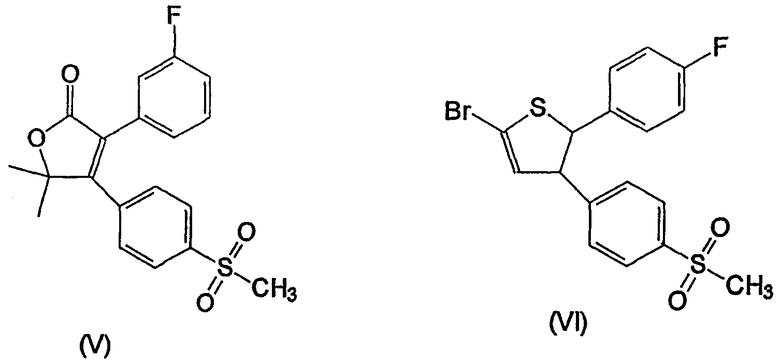
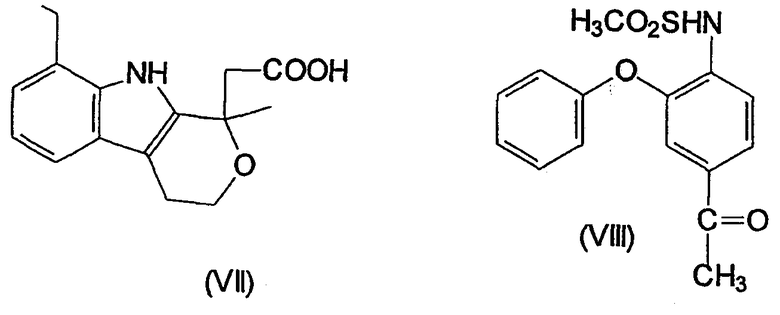
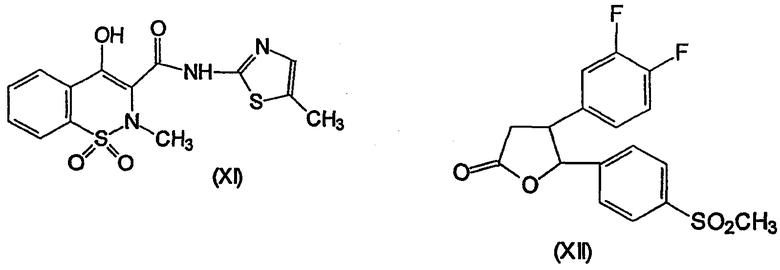
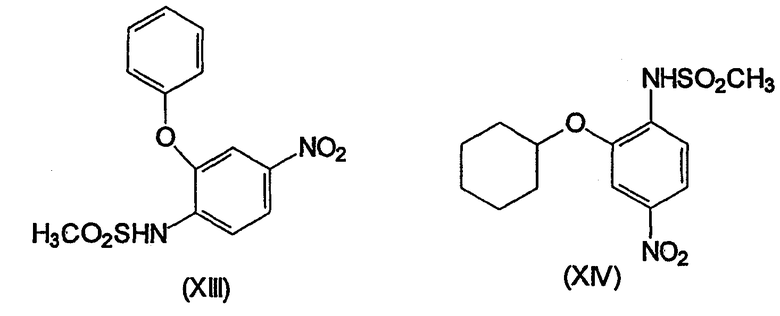
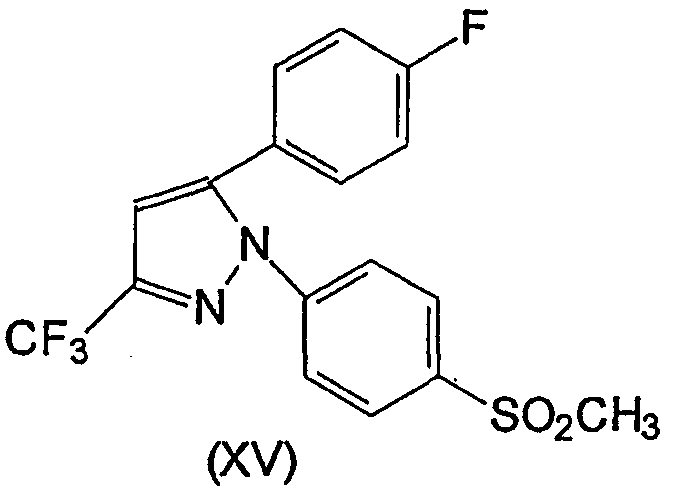
Несколько других ингибиторов СОХ-2 было оценено в биологических системах, и некоторыми из них являются BF 389 (III), CGP 28232 (IV), DFP, DFU (V), DuP 697 (VI), этодолак (VII), FK 3311 (VIII), флозулид (IX), L-745337 (X), мелоксикам (Mobic®, US 4233299, 4-гидрокси-2-метил-N-(5-метил-2-тиазолил)-1,1-диоксид-2Н-1,2-бензотиазин-3-карбоксамид) (XI), трициклический MF (XII), нимезулид (XIII), NS-398 (XIV) и SC-58125 (XV):

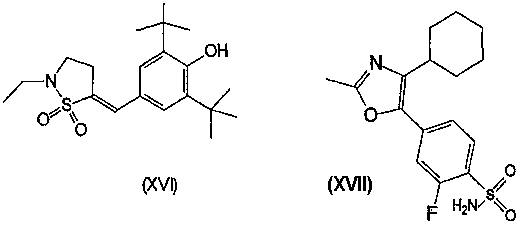
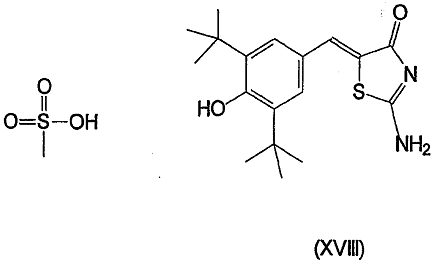
Другими описанными соединениями, ингибирующими СОХ-2, являются S-2474 (от Shionogi, EP 595546, 5(E)-(3,5-ди-трет-бутил-4-гидрокси)бензилиден-2-этил-1,2-изотиазолидин-1,1-диоксид) (XVI), JTE-522 или RWK-57504 (4-(4-циклогексил-2-метил-5-оксазолил)-2-фторбензолсульфонамид)(XVII), мезилат дарбуфелона (Pfizer, WO 94/03448, монометансульфонатная соль 2-амино-5-((3,5-бис(1,1-диметилэтил)-4-гидроксифенил)метилен-4(5Н)-тиазолон) (XVIII), 6089 (от Kotobuki Pharmaceutical)(XIX), валдекоксиб (Pharmacia, 4-(5-метил-3-фенил-4-изоксазол)бензолсульфонамид)(XX), паракоксиб-натрий (Pharmacia, натриевая соль N-((4-(5-метил-3-фенил-4-изоксазолил)фенил)сульфонил)пропанамид) (XXI), 4-(2-оксо-3-фенил-2,3-дигидрооксазол-4-ил)бензолсульфонамид (Almirall-Prodespharma)(XXII) и эторикоксиб (MK-633, Merck & Co.):

Вышеописанные соединения образуют предпочтительные ингибиторы СОХ-2, используемые в способах, описанных ниже.
Показаниями для применения ингибиторов СОХ-2 являются артрит, костно-мышечные боли и общие болезненные состояния, которые обычно устраняются классическими НСПВС, такими как индометацин, дихлорфенак и напроксен. Недавно было предложено использовать ингибиторы СОХ-2 в противораковой терапии, а также для предупреждения рака. Ингибиторы СОХ-2 могут быть также использованы при болезни Альцгеймера и других ассоциированных с деменцией процессах, протекающих в головном мозге.
Возможность клинического применения ингибиторов СОХ-2 обсуждается, например, в работах Nature, 367, p.215-216 (1994), in Drug News and Perspectives, 7, p.501-512 (1994), Annual Reports in Medicinal Chem., 30, p.179-188 (1995) и Oncogene, 18, p.7908-7916 (1999).
В этих работах не было высказано каких-либо конкретных предположений относительно возможности использования ингибиторов СОХ-2 в противовирусной терапии или, более конкретно, в терапии ВИЧ/СПИДа, и ингибиторы СОХ-2 не были тестированы на действие против ВИЧ. Кроме того, не было высказано каких-либо предположений относительно возможности использования ингибиторов СОХ-2 (или неселективных ингибиторов СОХ) в качестве иммуностимулирующих агентов для лечения иммунодефицита вирусного и невирусного происхождения.
ВИЧ-инфекция и СПИД являются главной угрозой для здоровья более чем 33 миллионов человек, инфицированных этим вирусом, во всем мире. Большинство инфицированных людей проживает в Африке (в регионе ниже Сахары) и в отдельных частях Азии. В настоящее время существует два класса соединений против СПИДа, которые используются в клинической практике; а именно, ингибиторы обратной транскриптазы ВИЧ и ингибиторы протеазы ВИЧ. Ингибиторы обратной транскриптазы ВИЧ могут быть подразделены на ненуклеозидные ингибиторы обратной транскриптазы (NNRTI) и нуклеозидные ингибиторы обратной транскриптазы (NRTI).
Наиболее часто используемыми NNRTI являются невирапин, делавиридин, эфавиренц, эмивирин и Т180. Наиболее часто используемыми NRTI являются зидовудин, диданозин, ставудин и зальцитабин. Ингибиторами протеазы ВИЧ, имеющими клиническое применение, являются инклинавир, палинавир и саквиравир.
Существующие методы лечения ВИЧ-инфекций и СПИДа основаны на использовании комбинации нескольких лекарственных средств, так называемой смеси ингибиторов обратной транскриптазы и ингибиторов протеазы. Эти комбинации, называемые HAART (высокоактивная антиретровирусная терапия), являются достаточно эффективными и могут снижать количество вируса в крови пациента до недетектируемых уровней. Однако HAART не приводит к полному излечению пациента, поскольку этот вирус еще присутствует в иммунных клетках и через некоторое время заболевание может возникнуть снова, а после прерывания лечения часто наблюдаются пики виремии и быстрое прогрессирование СПИДа. Кроме того, иммунодефицит и ВИЧ-специфическое нарушение Т-клеточной функции все еще наблюдаются в процессе HAART-терапии. Указанная терапия требует длительного применения и является весьма дорогостоящей. Стоимость лишь одного лекарственного средства часто превышает 15000 долларов США. Кроме того, существуют и некоторые другие проблемы, связанные с этой терапией, и именно, трудности, связанные с соблюдением пациентом режима лечения (сложные схемы введения лекарственного средства), продуцирование резистентных вирусов, неидеальная фармакокинетика и побочные эффекты, такие как, например, подавление функции головного мозга и длительных метаболических эффектов.
Описание терапии против ВИЧ можно найти в недавно опубликованных обзорах, см., например, Hilgegroth, 1998, Pharm. uns. Zeit., 1998, 27, p.22-25, Hilgegroth, 1998, Pharm. uns Zeit., 7, p.111-116, Stellbrink, 1997, Dk Ärztebl., 94, p.2497-2503, Rettle et al., 1998, Int. J. STD AIDS, 9, p.80-87, De-Clercq, 1998, Antiviral Res., 38, p.153-179, Gait et al., 1995, TIBTECH, 13, p.430-438 & Redshaw et al., in "Emerging Drugs: The Prospects of Improved Medicines", Chapter 6, p.127-154, 1997.
В заключение можно сделать вывод, что, хотя комбинации множества лекарственных средств, таких как HAART, значительно улучшают прогноз для пациентов, страдающих ВИЧ-инфекцией, однако, получение новых соединений для противовирусной терапии ВИЧ, а в частности, агентов, стимулирующих иммунную систему, является крайне необходимым для современной медицины. Настоящее изобретение направлено на получение таких соединений.
Экспрессия СОХ-2 обычно ограничивается головным мозгом/отростками головного мозга, синовиальной жидкостью при артритах и участками поражения тканей. СОХ-2 не был обнаружен в нормальных лимфатических узлах или в лимфоцитах. Однако было неожиданно обнаружено, что у мышей, инфицированных вирусом, вызывающим иммунодефицитное состояние MAID, в клетках лимфатических узлов экспрессируются высокие уровни СОХ-2. Кроме того, положительные CD4+ и CD8+-Т-клетки, а также В-клетки из лимфатических узлов мыши с MAID содержали высокие уровни СОХ-2 (см. пример 2). Было установлено, что указанный фермент СОХ-2 может служить мишенью для направленной доставки иммуностимулирующего средства в целях ослабления симптомов иммунодефицита, например ослабления нарушения Т-клеточной функции, например, посредством генерирования антиген-специфического иммунного ответа.
Не претендуя на какую-либо конкретную теорию, можно предположить, что активность СОХ-2 способствует увеличению продуцирования PGE2, который, в свою очередь повышает уровни сАМР, активирующего путь передачи сигнала РКА, что приводит к нарушению функции лимфоцитов. Исследования, проведенные на мышах с MAID in vivo, продемонстрировали, что ингибиторы СОХ-2 улучшают иммунные функции Т-клеток (см. пример 6).
Настоящее изобретение относится к новому способу лечения или предупреждения иммунодефицита, а в частности к способу лечения ВИЧ-инфекций и СПИДа, предусматривающему введение субъекту терапевтически эффективного количества ингибитора СОХ-2, или его производного, или его фармацевтически приемлемой соли.
Таким образом, в первом своем аспекте, настоящее изобретение относится к способу лечения или предупреждения расстройства, характеризующегося повышенной СОХ-2-активностью, такого как расстройство, характеризующееся пониженной иммунной функцией (например, повышенной экспрессией СОХ-2) у человека или другого животного, где указанному животному вводят терапевтически эффективное количество ингибитора СОХ-2 или его производного или его фармацевтически приемлемой соли.
Используемый здесь термин повышенная СОХ-2-активность означает повышенные уровни активности, обусловленные продуцированием повышенного числа молекул СОХ-2 (например, повышенной экспрессией) и/или продуцированием более активных молекул (например, образующихся путем превращения латентных форм в активные формы или путем предотвращения ингибирования активных форм). Предпочтительно указанное расстройство характеризуется ослаблением иммунной функции, то есть иммунодефицитным состоянием, выражающимся, например, нарушением функции лимфоцитов. Используемый термин "иммунодефицит" означает нарушение функции клеток, участвующих в нормальных иммунных ответах, а в частности В- и Т-клеток. Таким образом, описанные здесь соединения могут быть использованы для достижения иммуностимулирующего эффекта в целях увеличения иммунных ответов. Таким образом, предполагается, что ингибиторы СОХ-2 обладают иммуномодулирующим действием. Предпочтительными состояниями, которые могут быть подвергнуты лечению, являются вирус-индуцированные иммунодефицитные расстройства.
Таким образом, применением вышеуказанного способа может быть, но не ограничивается им, лечение ВИЧ- или СПИД-ассоциированных расстройств у субъектов. Так например, приблизительно у 50% пациентов с вариабельным неклассифицируемым иммунодефицитом наблюдается нарушение Т-клеточной функции, аналогичное нарушению Т-клеточной функции при ВИЧ-инфекции, и у таких пациентов благоприятный эффект может давать иммуностимулирующее лечение. В соответствии с настоящим изобретением любой ингибитор СОХ-2 может быть введен субъекту, нуждающемуся в лечении ВИЧ-инфенции/СПИДа. Таким образом, предпочтительными состояниями для лечения способами настоящего изобретения являются инфекции, вызываемые ретровирусами, а в частности ВИЧ (и инфекции, вызываемые родственными вирусами у других животных, например SIV, FIV, MAIDS), и развивающийся в результате этих инфекций СПИД, а также вариабельный неклассифицируемый иммунодефицит и состояния, родственные вышеупомянутым состояниям.
Субъектами, которые могут быть подвергнуты лечению, являются предпочтительно человек и домашние питомцы, или сельскохозяйственные животные, такие как собаки, кошки, обезьяны, лошади, овцы, козы, коровы, кролики, крысы и мыши.
Альтернативно, настоящее изобретение относится к ингибитору СОХ-2, или его производному, или фармацевтически приемлемой соли, предназначенному для лечения или предупреждения расстройства, характеризующегося повышенной СОХ-2-активностью, описанной выше, или к использованию указанного ингибитора СОХ-2, его производного или его фармацевтически приемлемой соли для получения лекарственного средства для лечения или предупреждения расстройства, характеризующегося повышенной СОХ-2-активностью, описанной выше. Используемый здесь термин "лечение" означает снижение или ослабление одного или нескольких симптомов указанного расстройства предпочтительно до нормальных уровней, например инфекционности вируса, или снижение или ослабление нарушения иммунной функции. Термин "предупреждение" означает полное предупреждение, то есть отсутствие детектируемого инфекционного агента, например вируса, и/или поддержание его нормальных уровней, по сравнению с уровнями, соответствующими конкретному симптому (например, СОХ-2-активности), или снижение, или ослабление степени или продолжительности (то есть замедление) начала появления данного симптома.
Фермент циклооксигеназа-2 представляет собой новую мишень для терапии ВИЧ-инфекций/СПИДа. Термин "ингибитор СОХ-2" означает соединение, способное ингибировать фермент циклооксигеназу 2, но не оказывающее при этом значительного ингибирующего действия на циклооксигеназу-1, при его введении в конкретной концентрации. Таким ингибитором предпочтительно являются соединения, обладающие селективностью в пользу ингибирования циклооксигеназы-2 по сравнению с циклооксигеназой-1 (например, как было определено по отношению СОХ-1:СОХ-2 при IC80 в тесте WHMA, см. ниже), где указанное отношение составляет, по крайней мере, 10, более предпочтительно, по крайней мере, 50, и даже более предпочтительно, по крайней мере, 100. (Отношение селективности для одного конкретного соединения может варьироваться в зависимости от биологического анализа и формы, в которой оно выражено (предпочтительно это отношение выражается как СОХ-1:СОХ-2 при IC50 или IC80, см. таблицы 1-4). Указанные здесь отношения означают данные, полученные в одном или нескольких подходящих хорошо известных анализах на СОХ, проводимых предпочтительно с использованием очищенных человеческих ферментов, например, отношение величин IC50, определенных, например, Engelhart et al., 1995, см. выше. Однако, предпочтительно, указанным анализом является тест WHMA.
Ряд анализов на относительную активность СОХ-1 и СОХ-2 было осуществлено с использованием аналитических систем широкого ряда, от выделенных очищенных ферментов до интактных клеток и клеточных моделей, полученных от различных видов. Однако в настоящее время наиболее широко используемой моделью является анализ цельной крови человека (WBA) и модифицированная версия Уильяма Харви модифицированного анализа цельной крови человека (WHMA), который является предпочтительным анализом. Эти анализы позволяют использовать для тестирования легко доступные человеческие клетки, которые являются предпочтительным при введении данных соединений человеку. Следует также учитывать связывание НСПВС с белками плазмы. Кроме того, оценку селективности предпочтительно проводят при IC80, а не при IC50, вследствие того что кривые концентрации для ингибирования СОХ-2 и СОХ-1 не являются параллельными, а также вследствие того что большинство соединений используются в дозах, дающих постоянные концентрации в плазме, близкие к 80%-ному ингибированию (Warner et al., 1999, PNAS USA, 96, p.7563-7568).
В анализе WBA на СОХ-1, кровь обрабатывают тестируемым агентом, а затем, через 60 минут, кальциевым ионофором и инкубируют в течение 30 минут, после чего плазму собирают. Для анализа на СОХ-2 кровь обрабатывают аспирином для ингибирования СОХ-1, а через 6 часов обрабатывают липополисахаридом и тестируемым агентом и инкубируют в течение 18 часов, после чего плазму собирают. Затем с помощью радиоиммуноанализа оценивают содержание тромбоксана В2 в плазме, как показателя активности СОХ.
В тесте WHMA, анализ на СОХ-1 проводят, как описано выше. Для анализа на СОХ-2, кровь обрабатывают кондиционированной средой, взятой от культур эпителиальных клеток дыхательных путей человека (А549), обработанных интерлейкином 1β в течение 24 часов, и инкубируют с этой средой вместе с тестируемым агентом в течение 60 минут, после чего добавляют кальциевый ионофор, а через 30 минут добавляют диклофенак для прекращения продуцирования прозаноидов. Затем плазму собирают и анализируют с помощью радиоиммуноанализа на содержание в ней простагландина Е2, как показателя активности СОХ-2. В этом последнем анализе периоды инкубирования, проводимого для оценки активности СОХ-1 и СОХ-2, являются аналогичными, что делает эти активности более сравнимыми, а поэтому анализ WHMA является предпочтительным.
Селективность, определенная исходя из СОХ-2/WHMA-СОХ-1 при IC80, с использованием указанного анализа, показана в таблице 4, где величины 0,2 и 0,02 представляют 5- и 50-кратное увеличение селективности для СОХ-2, соответственно.
(Отношение СОХ-2/СОХ-1 при IC80, определенное в соответствии с тестом WHMA и взятое у Warner et al., см. выше):
Поэтому в предпочтительном варианте осуществления изобретения отношение селективностей определяют в анализе WHMA при IC80, и соединения, имеющие отношение селективности СОХ-2:СОХ-1, составляющие менее чем 0,2, предпочтительно менее чем 0,05, например, менее чем 0,02, предпочтительно менее чем 0,01, например, <0,005, являются особенно предпочтительными для использования в способах настоящего изобретения. Альтернативно, предпочтительные соединения имеют отношение селективности СОХ-1:СОХ-2 (в соответствии с анализом WHMA при IC80), составляющее более чем 2, предпочтительно более чем 5, особенно предпочтительно более чем 50 или 100, как указывалось ранее.
Используемый здесь термин "ингибирование" означает снижение измеряемой активности циклооксигеназы-2. Это может быть достигнуто путем воздействия на транскрипцию, трансляцию, пост-трансляционную модификацию или активность СОХ-2. Однако предпочтительно такое ингибирование может быть достигнуто путем ингибирования ферментативной активности, то есть путем блокирования активного центра уже присутствующих активных молекул СОХ-2.
Предпочтительно ингибиторы СОХ-2, используемые для лечения иммунодефицита или вирусной инфекции, а в частности ВИЧ-инфекций и СПИДа, имеют IC50 для СОХ-2 примерно менее чем 0,5 мкмоль/литр, а более предпочтительно примерно менее чем 0,2 мкмоль/литр.
Описанный способ относится к использованию ингибиторов СОХ-2 или их производных для предупреждения и лечения различных состояний, включая иммунодефицит и вирусные инфекции, а в частности ВИЧ-инфекции и СПИД.
В одном из предпочтительных вариантов осуществления настоящего изобретения ингибитор СОХ-2, предназначенный для лечения в соответствии с настоящим изобретением, выбран из сульфонамидов кислоты.
В одном из предпочтительных вариантов осуществления настоящего изобретения ингибиторы СОХ-2, используемые в настоящем изобретении, выбраны из соединений нижеследующей общей формулы А, включая простые эфиры и тиоэфиры метансульфонамида

где Х представляет атом кислорода или серы или алкильную группу, а предпочтительно группу -СН2-;
R1 представляет циклоалкильную или арильную группу, которые могут быть, но необязательно, замещены одной или более группами или атомами, предпочтительно, одним или более атомами галогена, такого как фтор;
R2, R3, R4 и R5 независимо представляют атом водорода, нитро- или ацильную группу или алкильную группу, которая может быть, но необязательно, замещена одной или более группами (например, ацильной группой) или одним или более атомами, либо, альтернативно, R2 и R3, R3 и R4 или R4 и R5, взятые вместе с атомом углерода, с которым они связаны, образуют циклопентаноновую группу;
или их производного, или фармацевтически приемлемой соли.
В указанных соединениях Х предпочтительно представляет атом кислорода. В других предпочтительных соединениях R1 представляет арильную группу или арильную группу, замещенную одним или более атомами фтора, или циклоалкильную группу.
В других предпочтительных соединениях R2 и R3 представляют атомы водорода, а R4 представляет группу -NO2 или -СОСН3. Альтернативными предпочтительными соединениями являются соединения, в которых R2 представляет атом водорода, а R3 и R4, взятые вместе, образуют циклопентаноновую группу.
Особенно предпочтительными соединениями формулы А, используемыми в настоящем изобретении, являются описанные здесь соединения, такие как флозулид, NS-398, нимезулид, FK 3311 и L-745337.
В другом предпочтительном варианте осуществления настоящего изобретения ингибитор СОХ-2, используемый в настоящем изобретении, выбран из диарилгетероциклов.
Одним из примеров соединений семейства диарилгетероциклов, которые могут быть использованы в настоящем изобретении как ингибиторы СОХ-2, являются соединения нижеследующей общей формулы В
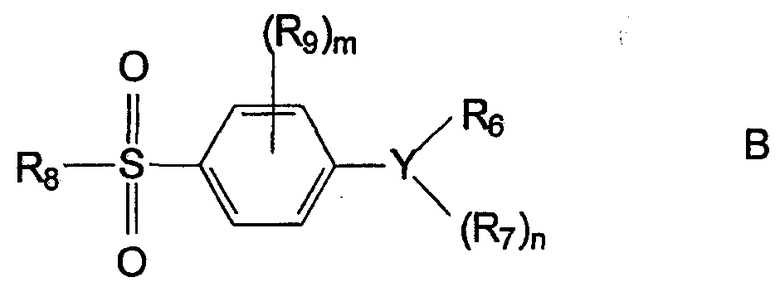
где Y представляет циклическую группу, предпочтительно выбранную из оксазолила, изоксазолила, тиенила, дигидрофурила, фурила, пирролила, пиразолила, тиазолила, имидазолила, изотиазолила, циклопентенила, фенила и пиридила;
n равно целому числу от 0 до 3;
m равно целому числу от 0 до 4;
R6 представляет кетоциклильную, циклоалкильную или арильную группу, которые могут быть, но необязательно, замещены одной или более группами или атомами, предпочтительно одним или более атомами галогена, такого как фтор;
R7 независимо представляет заместитель, которым может быть любая функциональная группа, предпочтительно атом водорода или атом галогена, предпочтительно фтора или брома, или алкильная группа (предпочтительно -СН3), которая может быть, но необязательно, замещена одной или более группами или атомами, предпочтительно одним или более атомами фтора, например -CF3;
R8 представляет алкильную группу, предпочтительно, -СН3 или NHR10, предпочтительно, -NH2;
R9 представляет атом галогена, предпочтительно фтор; и
R10 представляет атом водорода или алкильную группу, необязательно замещенную одной или более группами или атомами, предпочтительно ацильной группой;
или их производное, или фармацевтически приемлемая соль.
Соединения этого класса описаны в US 6025353 как антиангиогенные агенты и в US 6025353 также имеются дополнительное описание предпочтительных заместителей и соединений настоящего изобретения.
В указанных соединениях R8 предпочтительно представляет NH2- или -СН3. В других предпочтительных соединениях Y представляет пиразолильную, фурильную или тиенильную группу. R6 предпочтительно представляет арильную группу, необязательно замещенную одним или более атомами фтора. Предпочтительно n равно 1 или 2. R7 предпочтительно представляет атом брома, ацильную группу или замещенную алкильную группу, такую как CF3.
Особенно предпочтительными соединениями формулы В, используемыми в настоящем изобретении, являются описанные здесь соединения, такие как целекоксиб, рофекоксиб, DuP-697, SC-58125, DFP, DFU, CGP 28232 и трициклический MF.
Используемый здесь термин "алкил" означает любую длинноцепочечную или короткоцепочечную прямую, разветвленную или циклическую, алифатическую насыщенную или ненасыщенную углеводородную группу, необязательно моно- или полизамещенную гидроксигруппой, алкоксигруппой, ацилоксигруппой, нитрогруппой, алкоксикарбонилоксигруппой, аминогруппой, арильной группой, оксогруппой или галогеном, если это не оговорено особо. Ненасыщенные алкильные группы могут быть моно- или полиненасыщенными, и такими группами являются как алкенильные группы, так и алкинильные группы. Такие группы могут содержать вплоть до 40, предпочтительно от 1 до 10 атомов углерода.
Используемый здесь термин "циклические кольца" предпочтительно означает С5-7-кольцо, которое необязательно содержит один или несколько гетероатомов, выбранных из кислорода, азота и серы.
Используемый здесь термин "ацил" означает как карбоксилатную, так и карбонатную группы, так, например, ацилокси-замещенными алкильными группами является, например, алкилкарбонилоксиалкил. В таких группах, любая алкиленовая часть предпочтительно имеет число атомов углерода, определенное для вышеуказанных алкильных групп. Предпочтительными арильными группами являются фенильные и моноциклические 5-7-членные гетероароматические группы, а в частности, фенил и указанные группы сами могут быть, но необязательно, замещенными.
Характерными замещенными алкильными группами R1 являются алкоксиалкильная, гидроксиалкоксиалкильная, полигидроксиалкильная, гидроксиполиалкиленоксиалкильная группы и т.п., такие как алкоксиметильная, алкоксиэтильная и алкоксипропильная группы или ацилоксиметильная, ацилоксиэтильная и ацилоксипропильная группы, например, пивалоилоксиметил.
Используемые здесь замещенные группы могут быть моно- или полизамещенными гидроксигруппой, алкоксигруппой, ацилоксигруппой, нитрогруппой, алкоксикарбонилоксигруппой, аминогруппой, арильной группой, оксогруппой или галогеном, если не указано особо.
В другом предпочтительном варианте осуществления настоящего изобретения ингибитор СОХ-2 выбран из модифицированных классических НСПВС, например из их пролекарств, сложных эфиров или солей.
Новые селективные ингибиторы СОХ-2 были получены на основе химических структур классических НСПВС. Таким соединением может быть мелоксикам, которым является оксекам (СОХ-2-специфический аналог хорошо известного пироксикама) или производные уксусной кислоты, такие как этодолак (СОХ-2-специфический аналог диклофенака). Другими примерами некоторых наиболее предпочтительных соединений этого класса являются активные по отношению к СОХ-2 производные индометацина и зомепирак.
Дополнительный список семейств и подсемейств соединений настоящего изобретения приводится в патентах и в патентных заявках, относящихся к ингибиторам СОХ-2, например в патентных документах, на которые имеются ссылки в настоящем описании. В этих патентных документах также проиллюстрированы и перечислены конкретные соединения, которые также являются наиболее предпочтительными ингибиторами СОХ-2 настоящего изобретения.
Однако особенно предпочтительными соединениями являются диизопропилфторфосфат, L-745337, рофекоксиб, NS398, SС58125, этодолак, мелоксикам, целекоксиб, нимезулид.
Способы получения ингибиторов СОХ-2, используемых в настоящем изобретении, хорошо известны специалистам, и в частности, описаны в работах, упомянутых выше.
Ингибитор СОХ-2, предназначенный для лечения и предупреждения описанных здесь заболеваний, например, иммунодефицита и вирусных инфекций, а в частности, ВИЧ-инфекций/СПИДа в соответствии с настоящим изобретением, может иметь один или несколько асимметрических центров и/или одну или несколько двойных связей, то есть настоящее изобретение относится к использованию изомеров и рацематов описанных здесь соединений. Все такие возможные изомеры входят в объем настоящего изобретения. Ингибитор СОХ-2 может иметь форму изомерной смеси соединений или, более предпочтительно, форму очищенного изомера или его фармацевтически приемлемой соли.
Фармацевтическая композиция ингибитора(ов) СОХ-2 для лечения состояний в соответствии с настоящим изобретением, например иммунодефицита и вирусных инфекций, может быть получена в виде фармацевтически приемлемых солей и может также содержать фармацевтически приемлемые носители, хорошо известные специалистам.
Таким образом, настоящее изобретение также относится к фармацевтическим композициям, содержащим ингибитор СОХ-2 или его производное или фармацевтически приемлемую соль и фармацевтически приемлемый разбавитель, носитель или наполнитель. Термин "фармацевтически приемлемый" означает, что данный ингредиент должен быть совместимым с другими ингредиентами в указанной композиции, а также он должен быть физиологически приемлемым для реципиента.
В других вариантах своего осуществления, настоящее изобретение также относится к использованию таких композиций и к способам предупреждения/лечения с использованием указанных композиций, как описано выше.
Если ингибитор СОХ-2 является основным, то соли могут быть получены из фармацевтически приемлемых нетоксичных кислот, включая неорганические и органические кислоты. Особенно предпочтительными солями являются соли соляной, бромистоводородной, фосфорной, серной, лимонной, малеиновой и винной кислот.
Если ингибитор СОХ-2 является кислотным, то соли могут быть получены из фармацевтически приемлемых нетоксичных оснований, включая неорганические и органические основания. Особенно предпочтительными солями являются соли натрия, калия и меглумина.
Для лечения или предупреждения описанных здесь заболеваний, например, иммунодефицита или вирусных заболеваний, включая ВИЧ-инфекции/СПИД, ингибиторы СОХ-2 могут быть введены перорально, ректально, местно, буккально, путем ингаляции или парентерально (например, внутримышечно, подкожно, внутрибрюшинно или внутривенно) в форме инъекции или вливания. Предпочтительными способами введения являются пероральное введение, ректальное введение или введение путем инъекции или вливания. Наиболее предпочтительной формой введения является пероральное введение.
При всех способах введения ингибитор СОХ-2 вводят в виде разовых лекарственных форм, обычно содержащих хорошо известные фармацевтически приемлемые носители, адъюванты и наполнители. Так, например, активный ингредиент может быть введен, необязательно, вместе с другими активными веществами в виде комбинированного препарата с одним или несколькими стандартными носителями, разбавителями и/или наполнителями для получения стандартных галеновых препаратов, таких как таблетки, драже, порошки, пастилки, саше, облатки, эликсиры, суспензии, эмульсии, растворы, сиропы, аэрозоли (в твердой или в жидкой среде), мази, мягкие и твердые желатиновые капсулы, суппозитории, стерильные растворы для инъекций, стерильные порошки в упаковках и т.п. Для твердых имплантатов могут быть также использованы биологически разлагаемые полимеры (такие как полиэфиры, полиангидриды, полимолочная кислота или полигликолевая кислота). Указанные композиции могут быть стабилизированы сушкой вымораживанием, переохлаждением или с использованием пермазима.
Подходящими наполнителями, носителями или разбавителями являются лактоза, декстроза, сахароза, сорбит, маннит, крахмалы, аравийская камедь, фосфат кальция, альгинаты, трагакант, желатин, силикат кальция, микрокристаллическая целлюлоза, поливинилпирролидон, целлюлоза, водный сироп, вода, смесь вода/этанол, смесь вода/гликоль, смесь вода/полиэтилен, гликоль, пропиленгликоль, метилцеллюлоза, метилгидроксибензоаты, пропилгидроксибензоаты, тальк, стеарат магния, минеральное масло или жирные вещества, такие как твердый жир, или их подходящие смеси. Указанные композиции могут, кроме того, включать смазывающие агенты, смачивающие агенты, эмульгаторы, суспендирующие агенты, консерванты, подсластители, отдушки, усилители адсорбции, например, для назальной доставки (соли желчных кислот, лецитины, поверхностно-активные вещества, жирные кислоты, хелатообразующие агенты) и т.п. Композиции настоящего изобретения могут быть изготовлены хорошо известными методами так, чтобы обеспечивалось быстрое, замедленное или пролонгированное высвобождение активного ингредиента после введения этих композиций пациенту.
Активный ингредиент, предназначенный для введения, может быть соответствующим образом модифицирован для использования в фармацевтической композиции. Так активный ингредиент может быть стабилизирован, например, с использованием соответствующих добавок, таких как соли или вещества, не являющиеся электролитами, ацетат, ДСН, EDTA, цитратные или ацетатные буферы, маннит, глицин, HSA или полисорбат.
Для повышения липофильности, усиления клеточного транспорта, повышения растворимости или обеспечения точной доставки могут быть получены конъюгаты. Эти конъюгаты могут быть разлагаемыми, так чтобы данные конъюгаты действовали как пролекарство. Стабильность может быть также достигнута использованием соответствующих комплексов металла, например Zn, Са или Fe.
Активный ингредиент может быть включен в соответствующий носитель для доставки в конкретные клетки, органы или ткани или для нацеливания на эти клетки, органы или ткани. Так, например, фармацевтические композиции могут быть изготовлены в форме микроэмульсий, липосом, ниосом или наночастиц, которые могут абсорбировать, адсорбировать включать или связывать активный ингредиент. Это позволяет эффективно превращать данный продукт в нерастворимую форму.
Указанные частицы для увеличения времени их присутствия в кровотоке могут нести на своей поверхности соответствующие молекулы (например, сывороточные компоненты, поверхностно-активные вещества, полиоксамид 908, ПЭГ и т.п.) или молекулы для сайт-специфической направленной доставки, такие как лиганды для рецепторов, находящихся на конкретных клетках. Соответствующие методы доставки лекарственных средств и их нацеливания на конкретные участки хорошо известны специалистам, см., например, работы Kreuter, 1994, Eur.J. Drug Metab. Pharmacokinet., 3, p.253-256; Shen, 1997, J. Drug Targeting, 5(1), p.11-13; Mrsny, 1997, J. Drug Targeting, 5(1), p.5-9; Pettit & Gombotz, 1998, TIBTECH, 16, p.343-349; и Duncan, 1997, J. Drug Targeting, 5(1), p.1-4, касающиеся доставки лекарственных средств, и Simari & Nabel, 1996, работы Semin. Intervent. Cardiol., 1, p.77-83; Torchillin, 1998, J. Microencapsulation, 15(1), p.1-19; Klyashchitsky & Owen, 1998, J. Drug Targeting 5(6), p.443-458; Kreuter, 1996, J. Anat., 189, p.503-505; Fasano, 1998, TIBTECH, 16, p.152-157; Kataoka et al., 1993, 24, p.119-132; Anderson, 1998, Nature, 392(suppl), p.25-30; Langer, 1998, Nature, 392(suppl), p.5-10; Gregoriadis, 1995, TIBTECH, 13, p.527-536; Gregoriadis et. al., 1997, FEBS Lett., 402, p.107-110; Rolland, 1998, Critical Reviews in Therapeutics Drug Carrier Systems, 15(2), p.143-198; Hope et al., 1998, Molec. Memb. Biol., 15, p.1-14; и Scherman et al., 1998, Curr. Opinion Biotech., 9(5), p.480-485, касающиеся доставки молекул пептидов и нуклеиновых кислот. Так описание специфической сайт-направленной доставки можно найти, например, в работе Schafer et al., 1992, Pharm. Res., 9, p.541-546, где указывается, что наночастицы могут быть аккумулированы в ВИЧ-инфицированных макрофагах. Очевидно, что такие методы имеют конкретное применение в способах настоящего изобретения, описанных в данной заявке.
Указанные дериватизированные или конъюгированные активные ингредиенты подпадают под определение ингибирующих молекул, используемых в соответствии с настоящим изобретением.
Так, например, фармацевтическая композиция для перорального введения содержит активный ингредиент(ы) и физиологически приемлемые агенты, подходящие для изготовления таблеток, капсул, растворов, суспензий или других хорошо известных композиций для перорального введения. Такие композиции могут быть получены любым известным методом, применяемым для изготовления пероральных фармацевтических композиций. Указанные композиции могут содержать один или более биологически активных агентов или один или более агентов, выбранных из группы, состоящей из консервантов, инертных разбавителей, агентов, увеличивающих вязкость, красителей, подсластителей, гранулирующих агентов, дезинтегрирующих агентов, связующих агентов, осмотически активных агентов, смачивающих агентов, суспендирующих агентов, материалов для приготовления композиций замедленного высвобождения, масел и воды.
Фармацевтические композиции для других непероральных способов введения, например суппозитории для ректального введения или растворы для инъекций или вливаний, могут быть получены хорошо известными способами с использованием добавок, подходящих для таких композиций. Все композиции для инъекций и вливаний должны быть стерильными.
Активный ингредиент в таких композициях может составлять примерно от 0,01% до 99% от массы композиции, предпочтительно примерно от 0,1 до 50%, например 10%.
Для лечения расстройств, указанных в настоящем описании, например иммунодефицитов и вирусных инфекций, с использованием ингибиторов СОХ-2, суточные дозы этих ингибиторов составляют в пределах примерно 0,005-150 мг/кг массы тела. Эта доза в значительной степени зависит от выбора соединения ингибитора СОХ-2, от клинического состояния (типа вируса, статуса инфекции и состояния данного пациента), от возраста и веса пациента, способа введения и от использования пациентом общих лекарственных средств в процессе лечения, включая продолжительность курса лечения. Более предпочтительные дозы обычно составляют 0,01-50 мг/кг массы тела в день, и даже более предпочтительно, 0,05-20 мг/кг массы тела в день. Так, например, взрослому человеку могут быть ежедневно перорально введены 25 мг рофекобсиба или 200 мг целекоксиба.
Разовые лекарственные формы обычно включают 1-500 мг активного ингредиента.
В соответствии с одним из аспектов настоящего изобретения один ингибитор СОХ-2 может быть объединен с одним или несколькими другими ингибиторами СОХ-2 для лечения описанных здесь расстройств, например иммунодефицита или вирусной инфекции.
В соответствии с другим аспектом настоящего изобретения один ингибитор СОХ-2 может быть объединен с одним или несколькими другими ингибиторами СОХ-2 или с одним или несколькими лекарственными средствами, имеющими различные режимы действия и предназначенными для лечения таких расстройств, как иммунодефицит, ВИЧ-инфекция или СПИД. Примерами таких комбинаций могут служить комбинации ингибитора СОХ-2 с одним или несколькими NNRTI или комбинации с одним или несколькими NRTI, или комбинации с одним или несколькими ингибиторами протеазы ВИЧ, или комбинации одного или нескольких HAART с ингибитором СОХ-2.
В другом своем аспекте настоящее изобретение относится к способам и/или композициям, в которых один или несколько ингибиторов СОХ-2 объединены с соединениями, улучшающими переносимость активного ингредиента, а особенно во время продолжительного лечения. Типичными соединениями являются антигистамин и ингибиторы протонного насоса.
Таким образом, настоящее изобретение относится к композиции, содержащей ингибитор СОХ-2, описанный выше, вместе с одним или несколькими другими ингибиторами СОХ-2 и/или с одним или несколькими дополнительными активными ингредиентами. Кроме того, настоящее изобретение относится к использованию таких композиций и к способам использования таких композиций, описанных выше. Настоящее изобретение также относится к продукту, содержащему компоненты, описанные выше и используемые в виде комбинированного препарата для одновременного, раздельного или последовательного введения в целях лечения или предупреждения состояний или расстройств, описанных выше.
Настоящее изобретение подробно описано в нижеследующих и неограничивающих примерах со ссылками на нижеследующий графический материал, где
на фигуре 1 представлены уровни циклического АМР после MAIDS-инфицирования CD8+ (А), CD4+ (В) и В-клеток (С). Мононуклеарные клетки выделяли из лимфоузлов MAIDS-инфицированных мышей в различные периоды времени и разделяли на CD4+-, CD8+- и В-клетки путем негативного отбора с использованием клеточного сортера FACS. Уровни внеклеточного сАМР оценивали путем обработки ультразвуком и с помощью радиоиммуноанализа. Столбцы представляют собой средние величины ± стандартное отклонение (n=3 отдельных мышей);
на фигуре 2 представлены уровни MAIDS-сАМР в CD4+-, Thy-1.2-негативных и позитивных популяциях. Клетки лимфоузлов, взятые от трех инфицированных и трех контрольных мышей соответствующего возраста, разделяли на CD4+-, Thy-1.2+ (незаштрихованные столбцы) и CD4+-, Thy-1.2- (заштрихованные столбцы) популяции с использованием FACS-сортера, и оценивали уровни внутриклеточного сАМР, как показано на фигуре 1. Столбцы представляют собой средние величины ± стандартное среднеквадратичное отклонение (n=3);
на фигуре 3 представлены уровни активности протеинкиназы А у мышей с MAIDS по сравнению с мышами дикого типа. (А) Оценку киназных активностей с использованием кемптида в качестве субстрата в присутствии (общая активность, столбцы, заштрихованные косыми линиями) или в отсутствие (свободная активность, закрашенные черным столбцы) 5 мкМ cAMP проводили в солюбилизированных детергентом экстрактах клеток лимфоузлов, выделенных из мышиной селезенки. Для получения лишь РКА-специфической активности, фосфотрансферазную активность, которая не ингибировалась РКА-специфическим ингибитором протеинкиназы (PKI, 1 мкМ), вычитали. Активности у инфицированных мышей (MAIDS; n=4) были проиллюстрированы путем сравнения с активностью у инфицированных мышей дикого типа одного помета. (В) Связывание с [3Н-сАМР] измеряли в тех же самых экстрактах, что и в (А), и вычисляли молярные количества R-мономера;
на фигуре 4 показана иммунная локализация С-субъединицы РКА в клетках MAIDS-инфицированных мышей и мышей дикого типа. Мононуклеарные клетки от контрольных мышей (верхняя панель) и MAIDS-инфицированных мышей (две нижние панели) связывали с предметными стеклами с помощью цитоцентрифуги (400Чг), фиксировали и подвергали иммунному окрашиванию поликлональным антителом против РКА-С и ПХ-конъюгированным "вторым" антителом (бурая окраска). Контрастное окрашивание проводили гематоксилином (синяя окраска на хроматин);
на фигуре 5 проиллюстрировано воздействие антагониста РКА типа I, Rp-8-бром-сАМР-фосфортиоата (Rp-8-Br-cAMPS), на функцию Т-клеток у MAIDS-инфицированных мышей и у мышей дикого типа. TCR/CD3-стимулированную пролиферацию Т-клеток оценивали с помощью Т-клеток, выделенных у MAIDS-инфицированных мышей (А) и у неинфицированных контрольных мышей (В). Действие возрастающих концентраций агониста сАМР (8-СРТ-сАМР) на TCR/CD3-стимулированную пролиферацию CD3+-Т-клеток, выделенных у MAIDS-инфицированных мышей (незаштрихованные кружки, пунктирная линия) и у контрольных мышей (закрашенные кружки и сплошная линия), оценивали отдельно в том же самом эксперименте (С). Представлены средние значения для определений с тремя повторностями ±стандартные отклонения. Систематизированные данные представлены в таблице 4 (n=11). Примечание: масштабы на панелях А и В отличаются, а на панели С, TCR/CD3-индуцированная пролиферация в отсутствие агониста сАМР была нормализована на 100% для Т-клеток MAIDS-инфицированных мышей и контрольных мышей;
на фигуре 6 проиллюстрирована секреция PGE2 нормальными клетками и MAIDS-клетками лимфоузлов in vitro. Неотсортированные клетки лимфоузлов от MAIDS-инфицированных мышей (сплошные столбцы, n=9), взятых через 20 недель после инфицирования, и от контрольных мышей соответствующего возраста (затененные столбцы, n=4) культивировали в течение 48 ч в полной среде, после чего измеряли уровни секретированного PGE2 в супернатантах с помощью ELISA;
на фигуре 7 проиллюстрировано действие неселективного ингибитора СОХ на иммунную функцию Т-клеток у нормальных и у MAIDS-инфицированных мышей. Столбец 1 - контрольные мыши + антитело против CD3; столбец 2 - контрольные мыши + антитело против CD3 + индометацин; столбец 3 - MAIDS-инфицированные мыши + антитело против CD3; столбец 4 - MAIDS-инфицированные мыши + антитело против CD3 + индометацин. Т-клеточные пролиферативные ответы оценивали в смешанной популяции неотсортированных мононуклеарных клеток лимфоузлов по включению [3Н]-тимидина в отсутствие и в присутствии неселективного ингибитора СОХ, индометацина, (50 нг/мл). Активацию Т-клеток осуществляли путем перекрестного связывания анти-CD3 антитела (mAb 2С11; 4 мкг/мл). Столбцы представляют средние значения ± стандартное отклонение для контрольных мышей (n=3) и MAIDS-инфицированных мышей (n=5), дополнительные данные см. в таблице 5. Клетки культивировали 72 часа, и во время этого культивирования включение [3Н]-тимидина происходило в последние 4 ч;
на фигуре 8 проиллюстрирована экспрессия СОХ-2 различными субпопуляциями лимфоцитов лимфоузлов нормальных мышей (А) и MAIDS-инфицированных мышей MAIDS (В). CD4+-, CD8+-Т- и В-клетки сортировали на FACS-сортере путем позитивного отбора на основе экспрессии молекул CD4, CD8 и В220, соответственно. CD11b-клетки были отсортированы путем негативного отбора (исходя из отсутствия CD11b). Затем клетки от MAIDS-инфицированных и нормальных мышей лизировали и 10 мкг белка от каждого образца подвергали иммуноблот-анализу на экспрессию СОХ-2. Блоты одновременно реагировали с антителами против актина, используемыми в качестве контроля;
на фигуре 9 проиллюстрирована экспрессия CD11b-клеток лимфоузлов от MAIDS-мышей и мышей дикого типа. Показана экспрессия CD11b (определенная путем проточной цитометрии) различными субпопуляциями лимфоцитов лимфоузлов (CD4+-, CD8+-Т-клетками и В220+-В-клетками) от MAIDS-инфицированных и контрольных мышей. R1: высокий уровень экспрессии CD11b; R2: низкий уровень экспрессия CD11b; R3: экспрессия CD11b отсутствует;
на фигуре 10 представлены уровни экспрессии СОХ-2 в лимфоузлах MAIDS-инфицированных мышей и мышей дикого типа. Приготавливали замороженные срезы лимфоузлов и подвергали их иммуногистохимическому окрашиванию на СОХ-2 (бурая окраска). (а) Нормальный контрольный лимфоузел с зародышевым центром, окрашенным на СОХ-2. (b) Нормальный лимфоузел при более сильном увеличении. Клетки, дающие позитивную окраску на ПХ-цветную реакцию, представляют собой макрофаги, "окрашивающиеся тельца" с фаготитированным материалом (стрелки). (с) Лимфоузел MAIDS-инфицированных мышей (через 20 недель после инфицирования). Примечания: измененная морфология и архитектура. (d) Лимфоузел MAIDS-инфицированных мышей, окрашенный на СОХ-2, при более сильном увеличении. Примечание: ряд клеток давал бурое иммуноокрашивание в цитоплазме и многочисленные митотические фигуры;
на фигуре 11 проиллюстрировано влияние введения in vivo неселективного ингибитора СОХ на иммунную функцию Т-клеток у ВИЧ-инфицированных пациентов. Т-клеточные пролиферативные ответы оценивали по включению [3Н]-тимидина в CD3+-Т-клетках у 3 пациентов (пациенты 1-3), участвующих в клиническом испытании фазы II и принимавших, в дополнение к проводимой тройной комбинированной терапии, 25 мг индометацина перорально три раза в день в течение 14 дней. На верхних панелях проиллюстрирована иммунная функция Т-клеток на день 0, на 14-й день (после 2-недельного лечения) и на 28-й день (через 2 недели после окончания лечения), которая соответствует столбцам 1, 2 и 3. Активацию Т-клеток осуществляли путем перекрестного связывания с антителами против CD3 (mAb SрVT3b). А: базальная пролиферация после активации Т-клеток; В: пролиферация в присутствии Rp-8-Br-cAMPS (1 мМ). Примечание: различия в степени сАМР-опосредованного иммунодефицита очевидны при сравнении верхней и нижней панели. Столбцы представляют средние значения ± стандартное отклонение для определений с тремя повторностями. Клетки культивировали 72 часа, и во время этого культивирования включение [3Н]-тимидина происходило в последние 16 ч;
на фигуре 12 проиллюстрировано влияние введения in vivo неселективного ингибитора СОХ, индометацина, на пролиферацию Т-клеток у ВИЧ-инфицированных пациентов, как описано на фигуре 11, но в данном случае участвуют 7 пациентов, указанных как пациенты 1-7, соответственно, закрашенными кружками, незаштрихованными кружками, закрашенными треугольниками, незаштрихованными треугольниками, закрашенными квадратами, незаштрихованными квадратами и закрашенными ромбами. Строили кривые средних значений для определений с тремя повторностями, соединительные линии иллюстрируют улучшение состояния каждого пациента;
на фигуре 13 проиллюстрировано действие рофекоксиба, специфического ингибитора СОХ-2, на иммунную функцию Т-клеток у MAIDS-инфицированных мышей. Т-клеточные пролиферативные ответы оценивали в смешанной популяции неотсортированных мононуклеарных клеток лимфоузлов по включению [3Н]-тимидина в отсутствие и в присутствии возрастающих концентраций (1,9-500 нМ) специфического ингибитора СОХ-2, рофекоксиба. Активацию Т-клеток осуществляли путем перекрестного связывания антитела против CD3 (mAb 2С11, 4 мкг/мл). Представлены средние значения для определений с тремя повторностями, а также построена сигмоидальная кривая. Клетки культивировали 72 часа, и во время этого культивирования включение [3Н]-тимидина происходило в последние 4 часа;
на фигуре 14 проиллюстрировано действие целекоксиба, специфического ингибитора СОХ-2, на иммунную функцию Т-клеток у MAIDS-инфицированных мышей, как описано на фигуре 13 для рофекоксиба;
на фигуре 15 проиллюстрировано действие рофекоксиба и целекоксиба по сравнению с действием индометацина на секрецию PGE2 клетками лимфоузлов (ЛУ) ex vivo для контрольных мышей (1) или для MAIDS-инфицированных мышей (2). Неотсортированные клетки ЛУ культивировали в полной среде в отсутствие или в присутствии индуктора PGE2, липополисахарида (ЛПС; 4 мкг/мл), неспецифического ингибитора циклооксигеназы, индометацина (50 мг/мл), и специфических ингибиторов СОХ-2, рофекоксиба (0,125 мкМ) и целекоксиба (0,125 мкМ). Через 48 ч измеряли концентрацию PGE2 в супернатантах с помощью ИФА. Затем анализировали 3 отдельных инфицированных мыши (20-я неделя) и пул от 3 контрольных мышей соответствующего возраста. Указаны средние значения ± стандартные отклонения;
на фигуре 16 проиллюстрировано влияние in vivo-обработки MAIDS-инфицированных мышей рофекоксибом на иммунную функцию Т-клеток. MAIDS-инфицированные мыши оставляли необработанными (необработанные мыши 1-3), либо им, перорально (per os), в течение семи дней давали рофекоксиб (3 мг/кг/день, один раз в день, обработанные мыши 1 и 2), который вводили через трубку, вставленную в желудочек. Затем Т-клеточные пролиферативные ответы оценивали in vitro в смешанной популяции неотсортированных мононуклеарных клеток лимфоузлов обработанных и необработанных животных по включению [3Н]-тимидина в отсутствие (столбец А) и в присутствии Rp-8-Br-cAMPS (0,5 или 1,0 мМ, столбцы В и С, соответственно). Активацию Т-клеток осуществляли во всех образцах путем перекрестного связывания антитела против CD3 (mAb 2С11, 4 мкг/мл). Контроль был представлен пролиферацией Т-клеток у неинфицированных мышей. Представлены средние значения для определений с тремя повторностями. Клетки культивировали 72 часа, и во время этого культивирования включение [3Н]-тимидина происходило в последние 4 часа;
на фигуре 17 проиллюстрировано влияние in vivo-обработки MAIDS-инфицированных мышей рофекоксибом или целекоксибом на иммунную функцию Т-клеток. MAIDS-инфицированным мышам инъецировали наполнитель (интралипид), затем этих мышей обрабатывали рофекоксибом в интралипиде путем внутрибрюшинной инъекции (3 мг/кг/день, один раз в день, n=6) либо их обрабатывали целекоксибом путем внутрибрюшинной инъекции (20 мг/кг/день, один раз в день, n=5) в течение 18-20 дней. Затем Т-клеточные пролиферативные ответы оценивали in vitro, как описано на фигуре 16, но без Rp-8-Br-cAMPS. Контроль был представлен пролиферацией Т-клеток у неинфицированных мышей. Представлены средние значения для определений с тремя повторностями (черные кружки), а также 25% и 75% процентили (области в рамках) и медиана (линия в рамке). Столбцы представляют диапазон; и
на фигуре 18 проиллюстрировано действие in vivo-обработки MAIDS-инфицированных мышей мелоксикамом на иммунную функцию Т-клеток. Осмотические насосы (Alzet, 100 мкл) с мелоксикамом (скорость высвобождения 70 мкг/животное/день) или с забуференным фосфатом физиологическим раствором (PBS) подкожно имплантировали MAIDS-инфицированным мышам (через 14 недель после инфицирования) и здоровым мышам в течение 14 дней. (а) Затем оценивали in vitro Т-клеточные пролиферативные ответы, как описано на фигуре 17. Для каждой группы представлены средние значения ± стандартная ошибка среднего. Влияние обработки мелоксикамом на стимулированную анти-CD3 антителом пролиферацию клеток от MAIDS-инфицированных мышей (закрашенные столбцы) по сравнению с MAIDS-индуцированными мышами, получавшими PBS (незаштрихованные столбцы), было значимым (р<0,05). (b) В смешанные культуры клеток лимфоузлов от групп мышей (а), обработанных in vivo мелоксикамом или PBS, снова добавляли мелоксикам (2,5 мкг/мл) в клеточной культуре in vitro, и оценивали Т-клеточную пролиферацию, индуцированную антителом против CD3, как описано в (а), и эффект повторного добавления мелоксикама in vitro (незаштрихованные столбцы) сравнивали с ответом клеток, полученным без добавления in vitro (закрашенные столбцы) (р=0,005). (с) В in vitro культуры смешанных клеточных культур лимфоузлов от групп мышей (а), обработанных in vivo мелоксикамом или PBS, добавляли Rp-8-Br-cAMPS (0,5 мМ), затем оценивали Т-клеточную пролиферацию, индуцированную антителом против CD3, как описано в (а), и эффект добавления Rp-8-Br-cAMPS in vitro (незаштрихованные столбцы) выражали как число, определяющее во сколько раз это индуцирование превышало индуцирование клеток без in vitro добавления (заштрихованные столбцы). Статистический анализ проводили с помощью U-критерия Манна-Уитни для сравнения двух групп животных и с помощью парного критерия Уилкоксона для сравнения одной и той же группы с различными обработками.
ПРИМЕРЫ
Пример 1
Мыши с синдромом мышиного приобретенного иммунодефицита (MAIDS) имеют нарушение функции Т-клеток, индуцированное сАМР/РКА типа I
MAIDS(синдром мышиного приобретенного иммунодефицита)
Во многих исследованиях MAIDS рассматривается как возможная модель ВИЧ-инфекции человека. Этот синдром развивается после инфицирования дефектным по репликации ретровирусом, кодирующим вариантный полипротеин Pr60gag (Chattopadhyay et al., 1991, J. Virol., 65, p.4232-4241; Jolicoeur, 1991, FASEB, J. 5, p.2398-2405). Этот синдром ассоциируется с прогрессирующей пролиферацией лимфоцитов в селезенке и в лимфоузлах и вызывает серьезные иммунные дефекты. Хотя дефектный ретровирус, ответственный за MAIDS-инфекции, инфицирует большинство В-клеток (Aziz, 1989, Nature, 338, p.505-508), однако при этом наблюдаются серьезные нарушения функции CD4+-Т-клеток и отсутствие у них реакции на стимуляцию митогеном in vitro. Большая часть CD4+-Т-клеток (но не CD8+-Т-клеток) инфицированных мышей также характеризуется необычным Thy-1-негативным фенотипом (Holmes et al., 1990, Eur. J. Immunol., 20, p.2783-2787; Moutschen et al., 1994, Scand. J. Immunol., 39, p.216-224 (MAIDS)). У нормальных, неинфицированных мышей, CD4+-Thy-1--Т-клетки селективно обнаруживались в зародышевых центрах, где они соответствовали недавно появившимся антиген-специфическим мигрантам.
Механизм, по которому вариантный белок Pr60gag индуцирует аномалию Т-клеток, пока не выявлен. Было заявлено, что растворимые факторы, секретированные инфицированными клетками, влияют на функцию Т-клеток (Simard J. Virol., 68, p.1903-1912) на расстоянии, но природа таких медиаторов до сих пор не выяснена. Другие исследования позволяют предположить, что для индуцирования дефектов Т-клеток необходимы прямые когнатные взаимодействия между CD4+-Т-клетками и антиген-презентирующими клетками (Green, 2001, J. Virol., 70, p.2569-2575; de Leval, 1998, J. Virol., 72, p.5285-5290).
Путь "аденилатциклаза-сАМР-протеинкиназа А" играет важную роль в регуляции иммунных ответов (Kammer, 1991, Immunol. Today, 9, p.222-229). Известно, что возрастающие концентрации сАМР ингибируют пролиферативные ответы Т-клеток на различные стимуляторы, такие как антитело против CD3 (анти-CD3 mAb) и интерлейкин-2. Недавно было высказано предположение, что негативная регуляция тирозинкиназы JАК3 может представлять собой механизм, посредством которого сАМР ингибирует пролиферацию Т-клеток (Kolenko, 1999, Blood, 93, p.2308-2318). Циклический АМР может также индуцировать негативную регуляцию мембранных белков, поскольку мышиные тимоциты или клетки тимомы, обработанные сАМР-индуцирующими агентами, такими как норэпинефрин, ингибируют экспрессию Thy-1 в соответствии с механизмом, ответственным за дестабилизацию мРНК (Wajeman-Chao, J. Immunol., 161, p.4825-4833).
Простагландин E2 (PGE2), сильный индуктор сАМР, секретируется, главным образом, моноцитами, макрофагами и активированными Т-клетками. PGE2 дают сдвиг в равновесии Т-клеток-хелперов в направлении от Т-клеток-хелперов типа 1 к Т-клеткам-хелперам типа 2 путем ингибирования IL-2 и усиления продуцирования IL-4 (Betz & Fox, 1991, J. Immunol., 146, p.108-113; Meyaard, 1997, Blood, 89, p.570-576). Наблюдается также сдвиг в дифференцировке В-клеток в сторону продуцирования IgE (Fedyk & Phipps, 1996, PNAS USA, 93, p.10978-10983). Синтез простагландинов происходит в результате последовательного действия циклооксигеназы-1 и -2 (СОХ-1 и СОХ-2) и специфических ГФ-синтаз (глицерат-трифосфат-синтаз) (Smith & DeWitt, 1996, Adv. Immunol., 62, p.167-215). Экспрессия СОХ-1 является, главным образом, конститутивной и широко распространенной, тогда как СОХ-2 индуцируется только в некоторых типах клеток (макрофагах, фибробластах, клетках гладких мышц) NO и воспалительными цитокинами, такими как IL-1 и TNF-α.
Механизм, ответственный за нарушение функции Т-клеток при MAIDS, пока не выяснен. Предполагается, что в нем участвуют преимущественно CD4+-Т-клетки, а в некоторых работах высказывается предположение, что изменение CD8+-Т-клеток обусловлено лишь отсутствием адекватной помощи CD4+-Т-клеток. В противоположность этому, ингибирование В-клеточных ответов является важным фактором и не может объясняться лишь дефектом CD4+-лимфоцитов. Поэтому наблюдаемое авторами настоящего изобретения селективное увеличение сАМР в В-клетках и CD4+-Т-клетках, но не в CD8+-клетках, подтверждает участие сАМР в анергическом процессе, ассоциированном с MAIDS.
В соответствии с этими данными, авторы настоящего изобретения впервые продемонстрировали селективное увеличение продуцирования сАМР в определенной субпопуляции клеток в данной модели заболевания. Если растворимый фактор, такой как простагландин E2, действительно ответственен за индуцирование сАМР, то как можно объяснить селективность его действия в отношении определенной субпопуляции? В первом исследовании было проведено сравнение экспрессии различных простаноидных рецепторов на CD4+- и CD8+-Т-клетках и был сделан вывод об аналогичном характере экспрессии в обеих субпопуляциях. Нормальные CD8+-Т-клетки являются абсолютно чувствительными к сАМР-индуцирующему действию PGE2. Возможное объяснение может быть дано на пострецепторном уровне; Т-клетки памяти/активированные Т-клетки являются более чувствительными к PGE2, чем "необученные" Т-клетки. При MAIDS, где происходят процессы, зависимые от МНС класса II, CD4+-Т-клетки могут приобретать конкретное состояние активации, придающее им большую чувствительность к воздействию данной концентрации PGE2. Пострецепторная модуляция простаноидных эффектов опосредуется, главным образом, рецептор-G-киназами (GRK), которые способствуют отделению белка G от соответствующего мембранного рецептора. Воспалительные состояния, такие как ревматоидный артрит, характеризуются негативной регуляцией GRK, а поэтому повышенной чувствительностью лимфоцитов к сАМР-индуцирующим агентам, таким как катехоламины. Уровни активности GRK в CD4+- и CD8+-Т-клетках инфицированных мышей неизвестны.
Способы, используемые в примерах 1 и 2
Мыши и клеточная суспензия
Самцов мышей С57ВL/6 выращивали в лаборатории авторов настоящего изобретения. Мышам в возрасте 4 и 5 недель два раза внутрибрюшинно (i.p.) инъецировали 0,25 мл бесклеточного вирусного экстракта. Контрольным мышам соответствующего возраста два раза, i.p., инъецировали 0,25 мл забуференного фосфатом физиологического раствора (PBS). Через различные промежутки времени после инфицирования мышей умерщвляли посредством CO2-асфикции. Периферические лимфатические узлы (паховые, подмышечные и шейные) диссоциировали шприцем с получением моноклеточных суспензий и пропускали через нейлоновый препарат для окраски клеток, три раза промывали полной средой RPMI 1640 и подсчитывали на цитометре Тома после исключения трипанового синего.
Вирус
Вирусный экстракт приготавливали из лимфатических узлов мышей, которым за 2 месяца до этого инъецировали RadLV-Rs, как описано выше. Лимфатические узлы собирали, измельчали в PBS и центрифугировали при 1,5·104 g в течение 30 минут. Затем супернатант снова центрифугировали в течение 30 минут при 1,5·104g. Этот бесклеточный вирусный экстракт хранили в жидком азоте. Для количественной оценки вирусных частиц проводили анализ методом бляшек ХС. Этот вирусный препарат содержал 103 бляшкообразующих единиц (PFU) экотропного вируса/мл.
Антитела
Для Вестерн-блот-экспериментов были использованы следующие поликлональные антитела: "первое" антитело: поликлональное кроличье антитело против СОХ-1 или кроличье антитело против СОХ-2. (Santa Cruz Biotechnology); "второе антитело": конъюгированное с пероксидазой хрена антикроличье антитело, закупленное у Trancduction Laboratories (Trancduction Laboratories, UK). Для проточной цитометрии, были использованы следующие моноклональные антитела moAb: РЕ-конъюгированное антитело против CD4/L3Т4 (YTS.191.1), ФИТЦ-конъюгированное антитело против CD45R/В220 (RА3-6В2), ФИТЦ-конъюгированное антитело против CD11b/Мас-1 (М1/70), ФИТЦ-конъюгированное антитело против CD161/NK-1.1 (PK136), ФИТЦ-конъюгированное антитело против CD8a (Ly-2) и CD16/CD32 (рецептор FcγIII/II) (2.4G2) (все они были закуплены у Pharmingen: San Diego, CA, USA). Анти-СD3 moAb (145-2C11) очищали в лаборатории авторов настоящего изобретения. Конканавалин А (ConA) был закуплен у Boehringer Mannheim Biochemica, а фитогемаглютинин-М (РНА) был закуплен у Difco.
Проточная цитометрия и клеточный сортинг
Анализ проводили на проточном клеточном сортере FACStar-plus с программным обеспечением Cellquest (Becton Dickinson). Для отбора жизнеспособных лимфоцитов было использовано прямое и боковое рассеяние. Для двухцветного анализа, ФИТЦ (зеленый) и РЕ (оранжевый), возбуждение в голубом диапазоне при 488 нм обеспечивалось аргоновым ионным лазером (модель с воздушно-водным охлаждением - Air-to-Water cooled model, Spinnaker 1161; Spectra Physics, Mountain View, CA). Для сортировки клеток 60·106 клеток инкубировали с антителом против FcγRII (Fc Block) для предупреждения неспецифических взаимодействий, а затем подвергали мечению антителами, конъюгированными с флуорохромом, в течение 20 минут на льду. CD4+-Т-клетки негативно отбирали путем элиминации CD8+-, B220+-, CD11b+-клеток. Аналогичным образом CD8+-Т-клетки негативно отбирали путем элиминации CD4+-, B220+-, CD11b+-клеток, а В-клетки отбирали путем элиминации CD8+-, CD4+-, CD11b+-клеток. Для каждой сортировки отобранную фракцию повторно анализировали с помощью проточной цитометрии для оценки чистоты, которая всегда превышала 97%.
Количественная оценка циклического АМР
Моноклеточные суспензии клеток лимфоузлов получали как описано выше, два раза промывали RPMI 1640 и центрифугировали при 1500·g в течение 3 минут. Затем клетки разрушали ультразвуком для облегчения высвобождения внутриклеточного сАМР в раствор для экстракции (0,01 н. HCl, 95% этанол). Раствор, содержащий клеточный лизат, центрифугировали при 13·104 g в течение 15 минут, и супернатант переносили в свежую пробирку. Экстракт выпаривали в центрифужном концентраторе (Speed Vac) при 45°С, и осадок хранили при -20°С. Непосредственно перед использованием этот осадок ресуспендировали в аналитическом буфере, и уровни сАМР измеряли с помощью радиоиммуноанализа (РИА) с использованием аналитической системы с 125I-меченым сАМР (Amersham, England). Концентрацию сАМР в тестируемых образцах определяли путем сравнения с нелинейной стандартной кривой. Для позитивных и негативных контролей, клетки лимфоузлов (1·106) инкубировали, соответственно с 1 мМ дибутирил-сАМР и 0,5 мМ DDA (ингибитор аденилилциклазы) в течение 30 минут при 37°С в инкубаторе в атмосфере воздуха с 5% CO2 и с повышенной влажностью, а затем измеряли концентрацию сАМР.
Гомогенизация клеток и иммуноблоттинг
Клетки (50·106) гомогенизировали путем обработки ультразвуком (2·15 сек) на льду в буфере, содержащем 10 мМ фосфат калия, рН 7,1, 250 мМ сахарозу, 1 мМ EDTA, 0,1% тритон Х-100 и 10 мкг/мл каждого из ингибиторов протеазы: химостатина, лейпептина, пепстатина А и антипаина (Tasken et al., 1993, J. Biol. Chem., 268, p.21276-21283) и центрифугировали в течение 30 минут (15000 g) для удаления нерастворившегося материала. Концентрации белка определяли с помощью анализов Брэдфорда (BioRad). Для проведения иммуноблоттинга 40 мкг белка выделяли с помощью электрофореза в 10% ПААГ с ДСН, переносили на PVDF-мембраны и инкубировали с антителами в смеси TBS/Твин с 5%-ным обезжиренным сухим молоком и 0,1% BSA (Blotto). "Первые" антитела детектировали с помощью ПХ-конъюгированных "вторых" антител (Jackson Laboratories/Trancduction Laboratories) и ЭХЛ (Amersham).
Фосфотрансферазная активность РКА
Каталитическую активность РКА анализировали путем фосфорилирования РКА-специфического субстрата (Leu-Arg-Arg-Ala-Ser-Leu-Gly)(Kemp et al., 1976, PNAS USA, 73, p.1038-1042), кемптида (Peninsula Laboratories INC.) с использованием [γ-32Р]-ATP (удельная активность 0,25 Ки/мМоль, Amersham) в смеси для анализа, описанной R. Roskoski (Methods Enzymol., 1983, 99, p.36). Фосфотрансферазную активность измеряли в присутствии или в отсутствие сАМР (5 мкМ) и PKI (1 мкМ) и вычитали низкие уровни активности, которые не ингибировались PKI, для определения РКА-специфической активности.
Измерение связывания циклического АМР
Количественную оценку специфического связывания [3Н]-сАМР с солюбилизированными регуляторными субъединицами РКА осуществляли, как описано Cobb & Corbin (Methods in Enzymology, 159, p.202-208, 1988) в смеси, содержащей [2,8-2Н]-сАМР (2,25 мкМ; удельная активность 5 Ки/мМоль; Du Pont-New England Nuclear). Молярные отношения субъединиц R вычисляли исходя из двух сайтов связывания сАМР на каждом мономере регуляторной субъединицы.
Иммуноцитохимия
Контрольные и инфицированные лимфоциты лимфатических узлов фиксировали холодным ацетоном в течение 5 минут и дважды промывали в течение 5 минут в 0,1% сапонине в PBS. Эндогенную пероксидазу блокировали путем инкубирования с 0,3%-ной перекисью водорода в 0,1% сапонине/PBS в течение 15 минут. После промывки в сапонине/PBS предметные стекла инкубировали в течение 30 минут при комнатной температуре с блокирующим буфером (1,5%-ная нормальная козья сыворотка в 0,1% сапонине/PBS), а затем инкубировали в течение 60 минут с раствором "первого" антитела при комнатной температуре в камере с повышенной влажностью. Антитело против Са брали от Santa Cruz и разводили 1:1000 в PBS, содержащем 0,1% сапонина и 0,5% нормальной козьей сыворотки. Затем предметные стекла промывали, как описано выше, и инкубировали с биотинилированным козьим антикроличьим антителом. Это антитело детектировали с использованием АМС-комплекса (Novastain Super ABC Kit, Novocastra). Пероксидазу выделяли с использованием диаминобензидина (DAB)(Dako), который давал коричневый осадок в присутствии H2O2. Предметные стекла подвергали контрастному окрашиванию гематоксилином-эозином (Sigma). Специфичность тестировали путем инкубирования в цитоцентрифуге со специфическим пептидом против субъединицы РКА-Сα.
Иммуногистохимия
Иммуногистохимический анализ осуществляли на тонких гистологических срезах (толщиной 2 мкМ) тканей, фиксированных в 4%-параформальдегиде и залитых в полимер (JB4-JBPolysciences). Срезы пропитывали трипсином (0,24%) в течение 1 минуты при 37°С, а затем твином 20 (2%) в течение 30 минут при 37оС. Эндогенные пероксидазы гасили путем инкубирования с H2O2 (1%) в течение 30 минут при комнатной температуре. Неспецифические сайты насыщали нормальной козьей сывороткой (1,5%) в течение 1 ч при 37оС. После этого срезы инкубировали в течение ночи при 4°С с "первым" поликлональным кроличьим антителом против СОХ-1 или кроличьим антителом против СОХ-2 (Santa Cruz Biotechnology), а затем инкубировали с биотинилированным козьим антикроличьим антителом. Это антитело детектировали с использованием АМС-комплекса (Novastain Super ABC Kit, Novocastra). Пероксидазу выделяли с использованием диаминобензидина (DAB)(Dako), который давал коричневый осадок в присутствии H2O2. Срезы подвергали контрастному окрашиванию гематоксилином-эозином (Sigma). Специфичность тестировали путем инкубирования срезов с нормальной кроличьей сывороткой вместо "первого" антитела.
Анализы на пролиферацию клеток у MAIDS-мышей
Анализы на пролиферацию клеток осуществляли путем инкубирования 0,1·106 CD3+-Т-клеток/мл в объеме 100 мкл в плоскодонных 96-луночных микротитрационных планшетах. Активация достигалась путем добавления монодисперсных магнитных сфер, покрытых козьим антителом против мышиных IgG (Dynal, cat. № 110.02) в отношении клетка:сфера=1:1, с последующим добавлением антитела против CD3 (клон 2С11) при конечном разведении 4 мкг/мл для рассматриваемых экспериментов. Оптимальную концентрацию антитела осторожно титровали с установкой начальных параметров "setup" и всегда проводили параллельные эксперименты при некоторых других разведениях антитела. Пролиферацию анализировали путем инкубирования клеток в течение 72 часов, при этом включение [3Н]-тимидина (0,4 мкКи) наблюдалось в последние 4 часа и эти клетки собирали с использованием сборщика клеток (Skatron, Sterling, VA, USA) на стекловолоконные фильтры. Уровень включенного предшественника подсчитывали в сцинтилляционном анализаторе (Tri-Carb, Packard, Meriden, CT, USA). Аналоги cAMP, если они использовались, добавляли за 30 минут до активации, осуществляемой добавлением анти-CD3 антител. 8-СРТ-сАМР закупали у Sigma (St. Louis, MO), а Sp- и Rp-8-Br-cAMPS закупали у Biolog Life Science Company (Bremen, Germany) и все они были растворены до концентраций 4-10 мМ в PBS, и концентрации вычисляли с использованием коэффициентов экстинкции, указанных в инструкциях производителей. Индометацин растворяли в воде и использовали при концентрации 50 нг/мл.
Определение PGE2
500 мкл супернатанта от 48-часовой культуры клеток лимфоузлов, выделенных от контрольных и инфицированных мышей, вводили пипеткой в полипропиленовые 1,5 мл-пробирки, в которые затем добавляли 500 мкл смеси вода:этанол (1,4) и 10 мкл охлажденной льдом уксусной кислоты. Содержимое пробирок осторожно перемешивали и оставляли на 5 минут при комнатной температуре. Затем пробирки центрифугировали при 2500·g в течение 2 минут. Супернатанты собирали и пропускали через С18-мини-колонки Amprep, которые были примированы 2 колоночными объемами 10% этанола. После этого колонки промывали 1 объемом H2O и 1 колоночным объемом гексана. Затем PGE2 элюировали 2·0,75 мл этилацетата. Фракции собирали и выпаривали досуха в атмосфере азота. И, наконец, каждую фракцию снова разводили в 100 мкл аналитического буфера и PGE2 анализировали с помощью набора для ИФА Amersham в соответствии с рекомендациями производителей.
Статистические анализы
Для сравнения двух групп субъектов использовали U-критерий (двусторонний) Манна-Уитни. Коэффициенты корреляции (r) вычисляли с помощью рангового критерия Спирмана. Статистический анализ и анализ путем построения кривых по точкам осуществляли с использованием программных пакетов "Statistica" (Statsoft Inc., Tulsa, OK) и "Sigma Plot" (Jandel Corporation, Erkrath, Germany). Результаты представлены как медианы и 25% и 75% процентили, и если это не оговорено особо, то р-величины являются двусторонними и рассматриваются как значимые при <0,05.
Эксперименты
MAIDS-инфекции приводят к увеличению количества сАМР в CD4+-Т-клетках. Мышей, инокулированных смесью ретровирусов RadLV-Rs, которые, как известно, вызывают MAIDS, умерщвляли через различные промежутки времени после инфицирования и клетки лимфоузлов сортировали путем негативного отбора с использованием проточного цитометра/клеточного сортера для получения чистых В-клеток и CD4+- и CD8+-Т-клеток. После инфицирования оценивали внутриклеточные уровни сАМР в различных популяциях клеток. Как видно на фиг.1, уровни сАМР в CD4+-Т-клетках через несколько недель после инфицирования заметно увеличивались (более чем в 20 раз). На последних стадиях, уровни сАМР в В-клетках также увеличивались, тогда как в CD8+-Т-клетках наблюдались лишь незначительные изменения. Кроме того, при разделении CD4+-Т-клеток на Thy-1.2+- и Thy-1.2--клетки путем позитивного сортинга было очевидно, что в Thy-1.2--клетках наблюдалось значительное увеличение уровней сАМР (фигура 2, в 6 раз). Эта популяция, обычно содержащая низкие уровни сАМР, также обнаруживала более высокие базальные уровни сАМР по сравнению с уровнями сАМР в Thy-1.2+-клетках в случае, когда обе популяции были взяты от неинфицированных мышей.
Оценка фосфотрансферазной активности РКА в постнуклеарных супернатантах, взятых от солюбилизированных детергентом экстрактов, показала, что общие уровни сАМР-зависимой киназной активности были пониженными в клетках MAIDS-лимфоузлов, тогда как изменения активности, наблюдаемые в отсутствие сАМР, были незначительными (фигура 3А). Этот результат соответствует хронической активации и диссоциации РКА, приводящими либо к разложению субъединицы С, либо к траснлокации субъединицы С. Оценка связывания сАМР (фигура 3В) не выявила каких-либо изменений в общих уровнях субъединицы R РКА. Иммуноцитохимический анализ клеток лимфоузлов от MAIDS-инфицированных и контрольных мышей указывал на повышенные уровни иммунореактивной субъединицы С РКА в ядрах (фигура 4). Эти данные также соответствуют активации "сАМР-РКА"-пути при MAIDS.
Антагонист РКА типа I способствует усилению пролиферации Т-клеток MAIDS
Для оценки влияния повышенного уровня сАМР и активации РКА на ингибирование TCR/CD3-индуцированной пролиферации Т-клеток, авторами настоящего изобретения был использован замещенный серой сАМР-аналог (Rp-8-Br-cAMPS), действующий как полный антагонист для РКА типа I (Gjertsen, Mellgren, et al., 1995, 1665/id). На фигуре 5А показано, что в Т-клетках, взятых от MAIDS-инфицированных мышей, TCR/CD3-стимулированная пролиферация составляла менее чем 10% по сравнению с Т-клетками, взятыми от неинфицированных контрольных мышей (фигура 5В). Кроме того, при оценке действия антагониста РКА типа I в Т-клетках, инфицированных MAIDS, авторы наблюдали зависимое от концентрации увеличение TCR/CD3-индуцированной пролиферации, которая, при более высоких концентрациях увеличивалась более чем в 4 раза (фигура 5А), тогда как при обработке контрольных Т-клеток никакой стимуляции не наблюдалось (фигура 5В). Исследование одиннадцати MAIDS-инфицированных мышей показало, что все они имели значительно более низкую пролиферацию Т-клеток по сравнению с контролем (р<0,001), и у 10 из 11 мышей антагонист РКА типа I вызывал увеличение пролиферации Т-клеток (р<0,01; 2,2-кратное увеличение медианы, таблица 5). Стимулирующее действие антагониста сАМР не являлось насыщенным даже при использовании самых высоких концентраций (фигура 5А и аналогичные данные (не приводятся), полученные для всех мышей, представлены в таблице 5). Это указывает на то, что растворимость данного соединения, его аффинность или доступность для клеток может быть ограничивающим фактором для наблюдаемого эффекта. Таким образом, более проницаемый и сильный антагонист РКА типа I, если он доступен, может еще более увеличивать TCR/CD3-индуцированную пролиферацию MAIDS-инфицированных Т-клеток.
Кроме того, действие агониста сАМР (на TCR/CD3-индуцированную пролиферацию исследовали на пяти MAIDS-инфицированных мышах и на четырех контрольных мышах. Т-клетки от MAIDS-инфицированных мышей обнаруживали явный сдвиг чувствительности к ингибированию пролиферации клеток при экзогенном добавлении 8-СРТ-сАМР (фигура 5С и таблица 5). Более того, при максимальных уровнях пролиферации Т-клеток от MAIDS-инфицированных мышей и контрольных Т-клеток, нормализованных на 100% (фигура 5С, данные не приводятся), очевидно, что помимо сдвинутой влево кривой сАМР-ингибирования, наклоны кривых для Т-клеток от MAIDS-мышей значительно отличались (коэффициенты Хилла для клеток MAIDS-мышей составляли 0,6 (0,54-1,52), а для нормальных Т-клеток эти коэффициенты составляли 2,2 (1,9-2,5), таблица 5, р<0,05). Повышенная чувствительность к ингибированию сАМР-аналогом позволяет предположить, что повышенные уровни эндогенного сАМР влияют на примирование сАМР-связывающего сайта РКА типа I и приводят к последующему увеличению аффинности сайта А для экзогенно введенного сАМР-аналога. Сдвиг в наклоне кривой, соответствующей кооперативному двухлигандному сайту связывания, в сторону кривой, соответствующей явному некооперативному ингибированию аналогом 8-СРТ-сАМР, также указывает на занятость В-сайта повышенными уровнями эндогенного сАМР.
(кратность увеличения)
(IC50, мкМ)
(коэффициент Хилла)
процентили)
(3740-9986)
n=11
(1,9-3,7)
n=11
(0,08-0,52)
n=5
(0,54-1,52)
n=5
(56539-82038)
n=6
(1,0-1,3)
n=6
(0,33-0,46)
n=4
(1,93-2,47)
n=4
Пример 2
Индуцированное циклическим АМР нарушение функции MAIDS-Т-клеток обусловлено повышенным продуцированием PGE2 CD11b-позитивными клетками с повышенными уровнями СОХ-2
Повышенное продуцирование PGE2 MAIDS-клетками
Смешанные популяции клеток лимфоузлов выделяли у MAIDS-инфицированных и контрольных мышей и культивировали in vitro. Уровни секретированного PGE2 оценивали в супернатантах среды после 48-часового культивирования, и в результате этой оценки было выявлено, что MAIDS-инфицированные клетки секретировали в 7-8 раз большее количество PGE2, чем контрольные клетки.
Ингибирование продуцирования PGE2 приводит к восстановлению пролиферации Т-клеток при MAIDS
Затем смешанные клетки лимфоузлов активировали анти-CD3 антителами для индуцирования пролиферации Т-клеток, и через 72 часа оценивали включение [3Н]-тимидина. И в этом случае, пролиферация клеток у MAIDS-инфицированных мышей составляла лишь 10-20% от пролиферации Т-клеток у неинфицированных мышей. Однако добавление к этим культурам индометацина для ингибирования продуцирования PGE2 в смешанных культурах приводило к значительному увеличению пролиферации клеток у пяти MAIDS-инфицированных мышей до уровней, сравнимых с уровнями контрольных мышей (фигура 6). Исследование 10 дополнительных MAIDS-инфицированных мышей (таблица 6) показало, что воздействие индометацина на Т-клеточную пролиферацию смешанных культур лимфоцитов было в высокой степени значимым (р<0,01). В противоположность этому, обработка контрольных культур индометацином не приводила к изменению пролиферации.
СОХ-2 экспрессируется на высоких уровнях в лимфатических узлах MAIDS-инфицированных мышей
Конститутивно экспрессируемый СОХ-1 является нормальным источником циклооксигеназной активности, продуцирующей PGE2. Однако у MAIDS-инфицированных мышей, у которых должны наблюдаться повышенные уровни PGE2, какого-либо увеличения уровней СОХ-1 обнаружено не было (данные не приводятся). Экспрессия СОХ-2 обычно ограничивается головным мозгом/отростками головного мозга, артритной синовиальной жидкостью и участками поражения тканей. СОХ-2 не был обнаружен в нормальных лимфоузлах или в лимфоцитах, как показано, например, для контрольных лимфоцитов на фигуре 8 (верхняя панель). Однако авторами было неожиданно обнаружено, что у MAIDS-инфицированных мышей неочищенные клетки лимфоузлов экспрессировали высокие уровни СОХ-2 (фигура 8, нижняя панель). Кроме того, позитивно отобранные CD4+ и CD8+-Т-клетки, а также В-клетки от MAIDS-инфицированных лимфатических узлов содержали высокие уровни СОХ-2. В противоположность этому, негативно отобранные CD11b--клетки содержали лишь низкие уровни СОХ-2.
В результате исследования CD4+ и CD8+-Т-клеток, а также В-клеток (В220-маркер) от MAIDS-инфицированных и контрольных мышей посредством проточной цитометрии было выявлено, что CD11b-маркер не экспрессировался на Т- или В-клетках. Однако отдельная фракция CD4+-Т-клеток и В-клеток от MAIDS-инфицированных мышей представляла "яркие" CD11b-клетки (дискриминационное окно для меченного R1), а дополнительный пул CD4+-Т-клеток и В-клеток, а также CD8+-Т-клеток представлял "бледные" CD11b-клетки (дискриминационное окно для меченного R2), что свидетельствовало о том, что они имели значимые, но более низкие уровни экспрессии CD11b. Таким образом, субпопуляции MAIDS-инфицированных CD4+-Т- и CD8+-Т-клеток представляли собой "яркие" и "бледные" CD11b-клетки, соответственно, при этом большинство В-клеток были позитивными. Эти результаты, взятые с учетом того факта, что именно CD11b+-клетки, а не CD11b--клетки, экспрессируют СОХ-2, указывают на то, что как В-клетки, так и Т-клетки в лимфоузлах MAIDS-инфицированных мышей экспрессируют СОХ-2.
В результате исследования интактных лифоузлов у MAIDS-инфицированных мышей посредством иммуногистохимического анализа было выявлено, что у MAIDS-инфицированных мышей (через 19 недель после инфицирования), в отличие от контрольных мышей, общая архитектура изменяется с потерей зародышевых центров (фигура 10, с-а). При большем увеличении предметных стекол, иммуноокрашенных на СОХ-2, видно, что лимфоузлы от контрольных животных имеют лишь бурую ПХ-окраску в фагоцитированном материале в макрофагах (ложноположительные "окрашивающие тельца", фигура 10b), тогда как бтльшая часть MAIDS-инфицированных клеток лимфоузлов были позитивно окрашенными на СОХ-2 (фигура 10d).
антитело
анти-CD3 антитело
(839-3051)
n=10
(335-4317)
n=7
(3594-8019)
n=10
(8963-37037)
n=8
** означает р<0,001; н.о. означает не определены.
Пример 3
ВИЧ-пациенты обнаруживают маргинальные эффекты при лечении неселективным ингибитором СОХ in vivo
Методы
Негативный отбор CD3+-Т-клеток периферической крови, взятых у ВИЧ-инфицированных пациентов
CD3+-Т-клетки периферической крови очищали путем негативного отбора из лейкоцитарной пленки здоровых доноров (Ullevaal University Hospital Blood Center, Oslo, Norway). Вкратце, мононуклеарные клетки периферической крови выделяли путем центрифугирования в градиенте плотности (Lymphoprep, Nycomed, Oslo, Norway) с последующим негативным отбором с использованием монодисперсных магнитных сфер, непосредственно покрытых антителами против CD14 и CD19, и сфер с крысиными антимышиными антителами IgG, покрытых антителами против CD56, и магнита. Все магнитные сферы поставлялись от Dynal (Осло, Норвегия, кат. № 111.12, 111.04 и 110.11, соответственно), а антитела против CD56 поставлялись от Pharmingen (San Diego, CA, cat. no. 31660.d). Все стадии осуществляли при 4°С. Клеточные суспензии были проанализированы с помощью проточной цитометрии и было обнаружено, что они более чем на 90% состоят из CD3+-клеток.
Анализы на пролиферацию с использованием Т-клеток ВИЧ-инфицированного пациента
Анализы на пролиферацию клеток осуществляли путем инкубирования 0,75·106 CD3+-Т-клеток/мл в объеме 100 мкл в плоскодонных 96-луночных микротитрационных планшетах. Активация достигалась путем последовательного добавления монодиспергированных магнитных сфер, покрытых козьим антимышиным IgG (Dynal, cat. № 110.02) в отношении клетка:сфера=1:1, с последующим добавлением антитела против CD3 (клон SpvT3b) при конечном разведении 1:125000 для рассматриваемых экспериментов. Оптимальную концентрацию антитела осторожно титровали с установкой начальных параметров "setup" и всегда проводили параллельные эксперименты при некоторых других разведениях антитела. Пролиферацию анализировали путем инкубирования клеток в течение 72 часов, при этом, включение [3Н]-тимидина (0,4 мкКи) наблюдалось в последние 16 часов. Эти клетки промывали и собирали на стеклянных фильтрах, а затем анализировали в β-сцинтилляционном счетчике. Аналоги сАМР, если они использовались, добавляли за 30 минут до активации, осуществляемой добавлением анти-CD3 антител. 8-СРТ-сАМР брали от Sigma (St. Louis, MO).
Эксперименты
Целью продолжающихся клинических испытаний фазы II была оценка иммуномодулирующего эффекта кратковременного лечения неселективным ингибитором СОХ (индометацином) на "суррогатные" параметры Т-клеток у ВИЧ-инфицированных пациентов. В соответствии с одобренным протоколом, пациенты получали 50 мг индометацина три раза в день (полная доза составляла 150 мг/день) в течение 2 недель, при этом образцы брали на день 0, на 14-й день и на 28-й день (через 2 недели после прерывания лечения). Однако из-за побочных эффектов, таких как боли в эпигастральной области и диспепсия, и из-за прекращения исследований пациентов на начальном этапе, эта доза была уменьшена до 25 мг индометацина 3 раза в день (полная доза составляла 75 мг/день). На фигуре 11 проиллюстрирована иммунная функция Т-клеток (определенная по пролиферации Т-клеток после активации) у 3 пациентов (пациенты 1-3), которые полностью проходили исследование. На верхней панели показаны уровни пролиферации после активации Т-клеток вначале (0-й день), после завершения лечения индометацином (14-й день) и через 2 недели после завершения лечения (28-й день). Как можно видеть, у пациентов 1 и 2 не увеличивалась их иммунная функция при in vivo-введении неселективного антагониста СОХ. Однако у пациента 3 Т-клеточные ответы возрастали приблизительно в 2,5 раз и оставались на этом уровне в течение 2 недель после прекращения введения индометацина. На фигуре 11b на нижней панели проиллюстрирована пролиферация Т-клеток после инкубирования с РКА-I-селективным антагонистом cAMP Rp-8-Br-cAMPS in vitro в клеточных культурах. Различия в степени cAMP-опосредованного нарушения Т-клеточной функции очевидны исходя из изменения пролиферации, наблюдаемой при введении антагониста (ср. верхнюю и нижнюю панели, где во все интервалы времени у пациентов 1 и 3 наблюдалось увеличение пролиферации в 2 раза, тогда как у пациента 2 никакого эффекта не наблюдалось). Как проиллюстрировано на фиг.11, индометацин не обладает надежным действием, которое может быть объяснено отсутствием СОХ-2-селективности, а также дозозависимыми ограничениями, обусловленными побочными эффектами.
Пример 4
ВИЧ-пациенты обнаруживают маргинальные эффекты после введения неселективного ингибитора СОХ in vivo(продолжение экспериментов, описанных в примере 3)
Методы
Эти методы описаны в примере 3.
Эксперименты
Результаты продолжающихся клинических испытаний фазы II (продолжение примера 3), полученные для 7 пациентов, которым, в дополнение к проводимой тройной комбинированной терапии, перорально вводили 25 мг индометацина три раза в день в течение 14 дней, представлены на фигуре 12. Пациенты 1-3 соответствовали пациентам, описанным в примере 3. Из-за побочных эффектов, связанных с введением индометацина и описанных выше (пример 3), доза индометацина была ограничена до 25 мг три раза в день. При этой предельно допустимой дозе эффекты указанного неселективного ингибитора СОХ были маргинальными. Через 14 дней после лечения только два из семи пациентов обнаруживали явное увеличение иммунной функции Т-клеток, измеряемой по пролиферации Т-клеток после их активации, тогда как у одного пациента наблюдалось снижение иммунной функции, а у четырех пациентов наблюдались незначительные изменения. Через две недели после прекращения введения индометацина, у пяти из семи пациентов наблюдалось усиление иммунных ответов по сравнению с ответами на день 0. Однако только у двух пациентов наблюдалось более чем двукратное увеличение пролиферации Т-клеток.
Пример 5
Ингибиторы СОХ-2 улучшают иммунную функцию MAIDS-инфицированных Т-клеток in vitro
Методы
Методы, используемые в анализе на пролиферацию, описаны в примере 1. Анализ на PGE2 описан в примере 1.
Эксперименты
Анализ на пролиферацию
Смешанные клетки лимфоузлов выделяли у MAIDS-инфицированных мышей через 17 недель после инфицирования. Для индуцирования пролиферации Т-клеток клетки активировали антителами против CD3 и через 72 часа оценивали включение [3Н]-тимидина как показателя иммунной функции. И снова пролиферация клеток у MAIDS-инфицированных мышей составляла лишь 5-20% от пролиферации Т-клеток неинфицированных мышей (2000-12000 им/мин в MAIDS-клетках, а в клетках неинфицированных мышей - в среднем 55000 им/мин). Однако при введении в культуры рофекоксиба (фигура 13) или целекоксиба (фигура 14) наблюдалось зависимое от концентрации 2-3-кратное увеличение пролиферации клеток у MAIDS-инфицированных мышей. В противоположность этому, обработка контрольных культур неинфицированных мышей рофекоксибом или целекоксибом не приводила к увеличению пролиферации (0,8-1,0-кратное увеличение в присутствии ингибиторов СОХ-2, то есть увеличения не наблюдалось, данные не приводятся). В Т-клетках MAIDS-инфицированных мышей концентрация рофекоксиба и целекоксиба, которая давала полумаксимальный эффект (EC50), составляла приблизительно 0,01 мкМ для рофекоксиба и 0,03 мкМ для целекоксиба. Тот факт, что субмикромолярные концентрации являются эффективными, явно указывает на то, что наблюдаемое увеличение иммунного ответа опосредовано ингибированием СОХ-2, но не СОХ-1, который ингибировался лишь при микромолярных концентрациях рофекоксиба и целекоксиба (величины взяты у Warner et al., 1999, PNAS USA, 96, p.7563-7568). Таким образом, восстановление ингибированной иммунной функции Т-клеток рофекоксибом и целекоксибом приводило к уменьшению продуцирования PGE2 в смешанных культурах и, тем самым, к снижению уровней Т-клеточного сАМР посредством ингибирования СОХ-2.
Продуцирование PGE2
Было также проанализировано влияние ингибиторов СОХ-2, рофекоксиба и целекоксиба, на уровни PGE2. Как видно на фигуре 15, неочищенные клетки лимфоузлов, взятые у MAIDS-инфицированных мышей, секретировали в 5-6 раз большее количество PGE2, чем клетки лимфоузлов, взятые от здоровых мышей (см. также фиг.6). Кроме того, уровни PGE2, продуцированного в ответ на ЛПС, у инфицированных мышей увеличивались в 8-10 раз, а у неинфицированных мышей - приблизительно в 2 раза. При инкубировании клеток в присутствии ингибиторов СОХ-2, рофекоксиба и целекоксиба, секреция PGE2 в MAIDS-инфицированных клетках лимфоузлов была аналогична секреции в неинфицированных клетках. Действие индометацина (сравнительная пролиферация на фиг.7) рассматривалось как контроль.
Пример 6
Ингибитор СОХ-2 повышает иммунную функцию MAIDS-инфицированных Т-клеток in vivo
Методы и эксперименты
Инфицированным мышам (через 17 недель после инфицирования) один раз в неделю per os (то есть перорально) вводили дозу рофекоксиба, соответствующую дозе, рекомендованной для человека (с учетом того факта, что у грызунов выведение лекарственного средства происходит в 7 раз быстрее). У MAIDS-инфицированных мышей обычно развивается синдром иммунопролиферации с увеличением лимфоузлов и селезенки. В соответствии с этим, необработанные инфицированные животные имели среднюю массу селезенки 1,3 г и среднюю массу пула лимфоузлов 1,7 г. В противоположность этому, у MAIDS-инфицированных мышей, получавших рофекоксиб в течение 7 дней, средняя масса селезенки составляла 0,8 г и средняя масса пула лимфоузлов составляла 0,3 г, что указывало на отмену лимфопролиферации.
Результаты представлены на фигуре 16. Оценка иммунной функции Т-клеток для неочищенных клеток лимфоузлов от инфицированных обработанных и необработанных мышей явно показала, что необработанные инфицированные животные обнаруживали индуцированную анти-CD3 антителом пролиферацию в диапазоне 2000-10000 им/мин (в среднем 7300 им/мин), а инфицированные мыши, которым вводили рофекоксиб в течение одной недели, давали Т-клеточные ответы, которые в 2,7-5,6 раза выше, чем инфицированные необработанные мыши. Кроме того, хотя инфицированные необработанные мыши обнаруживали повышенную индуцированную анти-CD3 антителом пролиферацию Т-клеток в присутствии Rp-8-Br-cAMPS, однако этот эффект был в 2-3 раза ниже у мышей, обработанных рофекоксибом, что указывало на то, что обработка рофекоксибом in vivo приводила снижению уровней PGE2 и отмене cAMP-опосредованного ингибирования функции Т-клеток.
Пример 7
In vivo-обработка MAIDS-инфицированных мышей рофекоксибом и целекоксибом приводила к усилению Т-клеточных ответов на анти-CD3 антитело и иммунных ответов
Методы и эксперименты
Инфицированных мышей обрабатывали рофекоксибом и целекоксибом в соответствии с дозой, рекомендованной для человека (с учетом того факта, что у грызунов выведение лекарственного средства происходит в 7 раз быстрее, 3 и 20 мг/кг/день, соответственно). Парентеральное введение осуществляли путем внутрибрюшинной инъекции ингибиторов СОХ-2, приготовленных в интралипиде. Результаты представлены на фигуре 17.
Оценка иммунной функции Т-клеток в неочищенных клетках лимфоузлов от инфицированных обработанных и необработанных мышей через 18-20 дней после инфицирования явно показала, что необработанные инфицированные животные обнаруживали индуцированную анти-CD3 антителом пролиферацию приблизительно 10000 им/мин, а инфицированные мыши, которые получали рофекоксиб в течение 18-20 дней, давали Т-клеточные ответы на анти-CD3 антитело, которые приблизительно в два раза превышали ответы инфицированных необработанных мышей. Аналогичным образом, обработка целекоксибом приводила к повышению иммунного ответа в клетках у большинства животных, относящихся к группе инфицированных мышей, и этот ответ был почти в 3 раза выше, чем ответ у необработанных неинфицированных мышей.
Пример 8
In vivo-обработка MAIDS-инфицированных мышей мелоксикамом приводила к увеличению Т-клеточной иммунной функции
Методы и эксперименты
Инфицированных мышей обрабатывали дозой 2,8 мг/кг/день мелоксикама, которая соответствовала дозе, рекомендованной для человека с учетом того факта, что у грызунов выведение лекарственного средства происходит в 7 раз быстрее. Парентеральное введение осуществляли путем подкожной имплантации осмотических насосов, наполненных водорастворимым соединением для инъекции мелоксикама. Проводили оценку Т-клеточной функции, и полученные результаты представлены на фигуре 18.
Оценка иммунной функции Т-клеток в неочищенных клетках лимфоузлов от обработанных и контрольных (PBS-обработанных) инфицированных мышей через 2 недели после обработки, явно показала, что PBS-обработанные инфицированные животные обнаруживали индуцированную анти-CD3 антителом пролиферацию приблизительно 500 им/мин, а инфицированные мыши, которые получали мелоксикам в течение 14 дней, давали значительно более высокие Т-клеточные ответы на анти-CD3 антитело, чем неинфицированные мыши, обработанные лишь PBS (фиг.18a, более чем в 10 раз; р<0,05).
При дополнительном введении мелоксикама в клеточные культуры в течение 3-дневного in vitro-анализа на пролиферацию Т-клеток, который осуществляли для предупреждения высвобождения СОХ-2 в результате in vivo-ингибирования под действием мелоксама и, тем самым, реактивации этого фермента, иммунный ответ в обработанной мелоксамом группе в два раза превышал ответ, который был получен без добавления мелоксикама in vitro (р=0,005), и этот эффект был также значительно выше по сравнению с ответом, полученным от MAIDS-инфицированных мышей, которым вводили PBS in vivo (фиг.18b, р<0,05).
В противоположность этому, повышенные иммунные ответы у MAIDS-инфицированных мышей, получавших PBS in vivo, и у необработанных мелоксикамом мышей наблюдались лишь в том случае, когда в индуцированные анти-CD3 антителом in vitro-культуры смешанных клеточных культур лимфоузлов был добавлен РКА-(типа I)-селективный антагонист cAMP, Rp-8-Br-cAMPS (фиг.18c). Тот факт, что эффект от введения антагониста сАМР отсутствовал у обработанных мелоксикамом MAIDS-инфицированных мышей, указывал на то, что in vivo-обработка мелоксикамом приводит к снижению или отмене сАМР-индуцированного иммунодефицита при MAIDS и сохранению иммунной функции.
| название | год | авторы | номер документа |
|---|---|---|---|
| ЛИГАНД GITR И СВЯЗАННЫЕ С ЛИГАНДОМ GITR МОЛЕКУЛЫ И АНТИТЕЛА И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ | 2004 |
|
RU2369636C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ЛЕЧЕНИЯ ИНФЕКЦИЙ И ОПУХОЛЕЙ | 2007 |
|
RU2540490C2 |
| ПРОТИВОВИРУСНАЯ ИММУНОТЕРАПИЯ ПОСРЕДСТВОМ ЛИГИРОВАНИЯ МЕМБРАННЫХ РЕЦЕПТОРОВ | 2017 |
|
RU2773427C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ЛЕЧЕНИЯ ИНФЕКЦИЙ И ОПУХОЛЕЙ | 2007 |
|
RU2478400C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ ПЕРСИСТИРУЮЩИХ ИНФЕКЦИЙ | 2016 |
|
RU2746383C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ ПЕРСИСТИРУЮЩИХ ИНФЕКЦИЙ | 2006 |
|
RU2596491C2 |
| СПОСОБ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ ПЕРСИСТИРУЮЩИХ ИНФЕКЦИЙ | 2006 |
|
RU2434641C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ПРОФИЛАКТИКИ И(ИЛИ) ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ ВИЧ У ЛЮДЕЙ | 2012 |
|
RU2609769C2 |
| ДОСТАВКА БИОМОЛЕКУЛ В КЛЕТКИ ИММУННОЙ СИСТЕМЫ | 2015 |
|
RU2739794C2 |
| СПОСОБЫ ИНДУКЦИИ ДЛИТЕЛЬНОГО ИММУННОГО ОТВЕТА | 2002 |
|
RU2313364C2 |
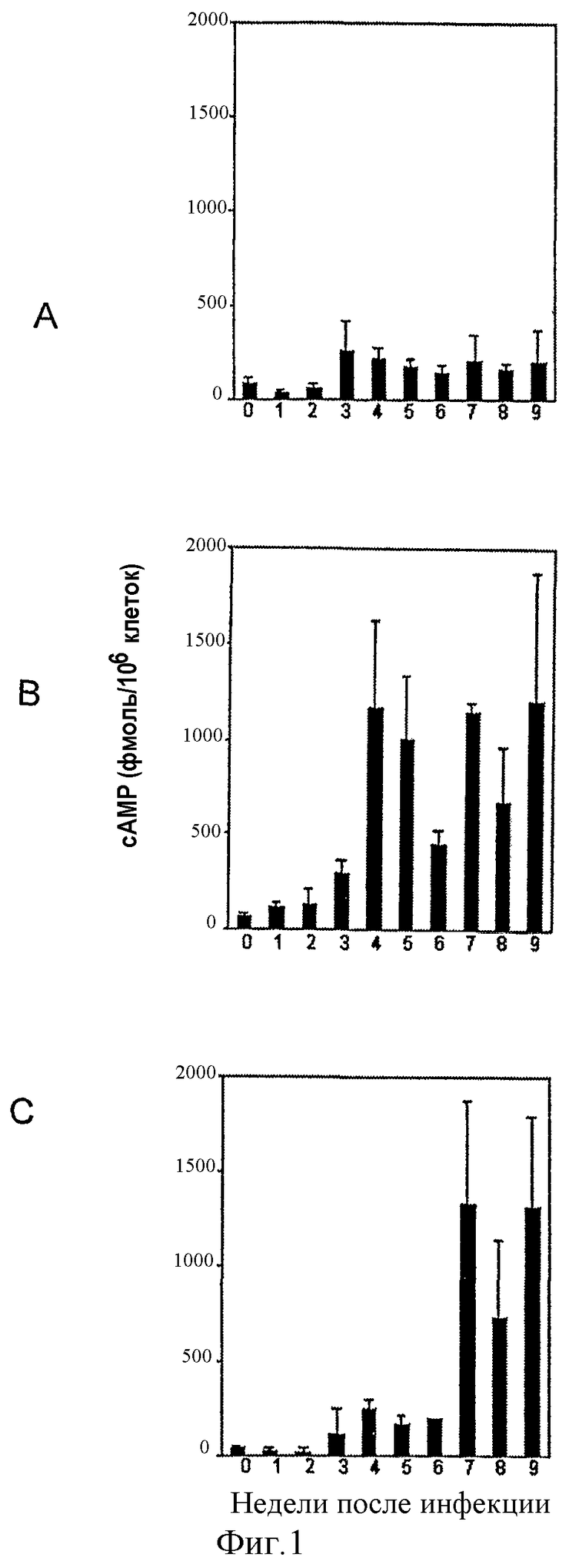
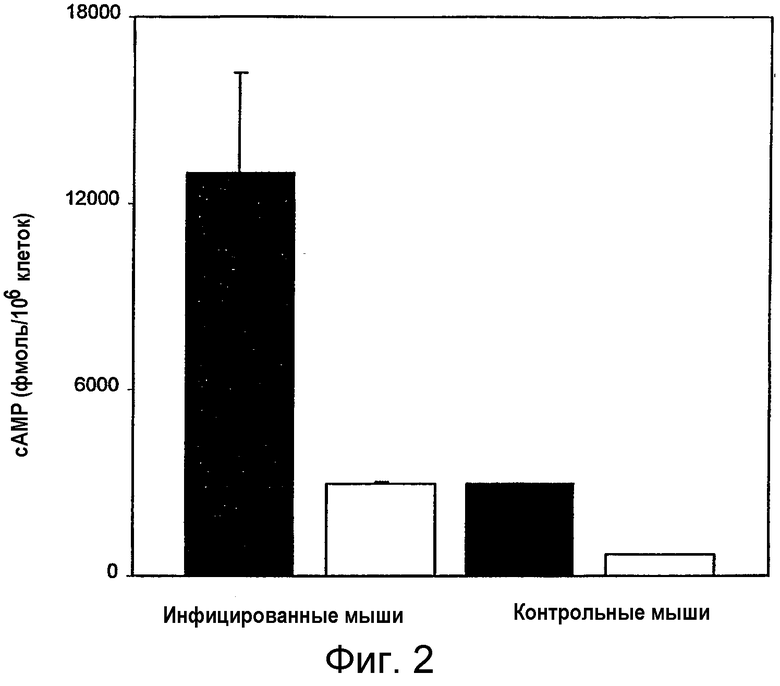

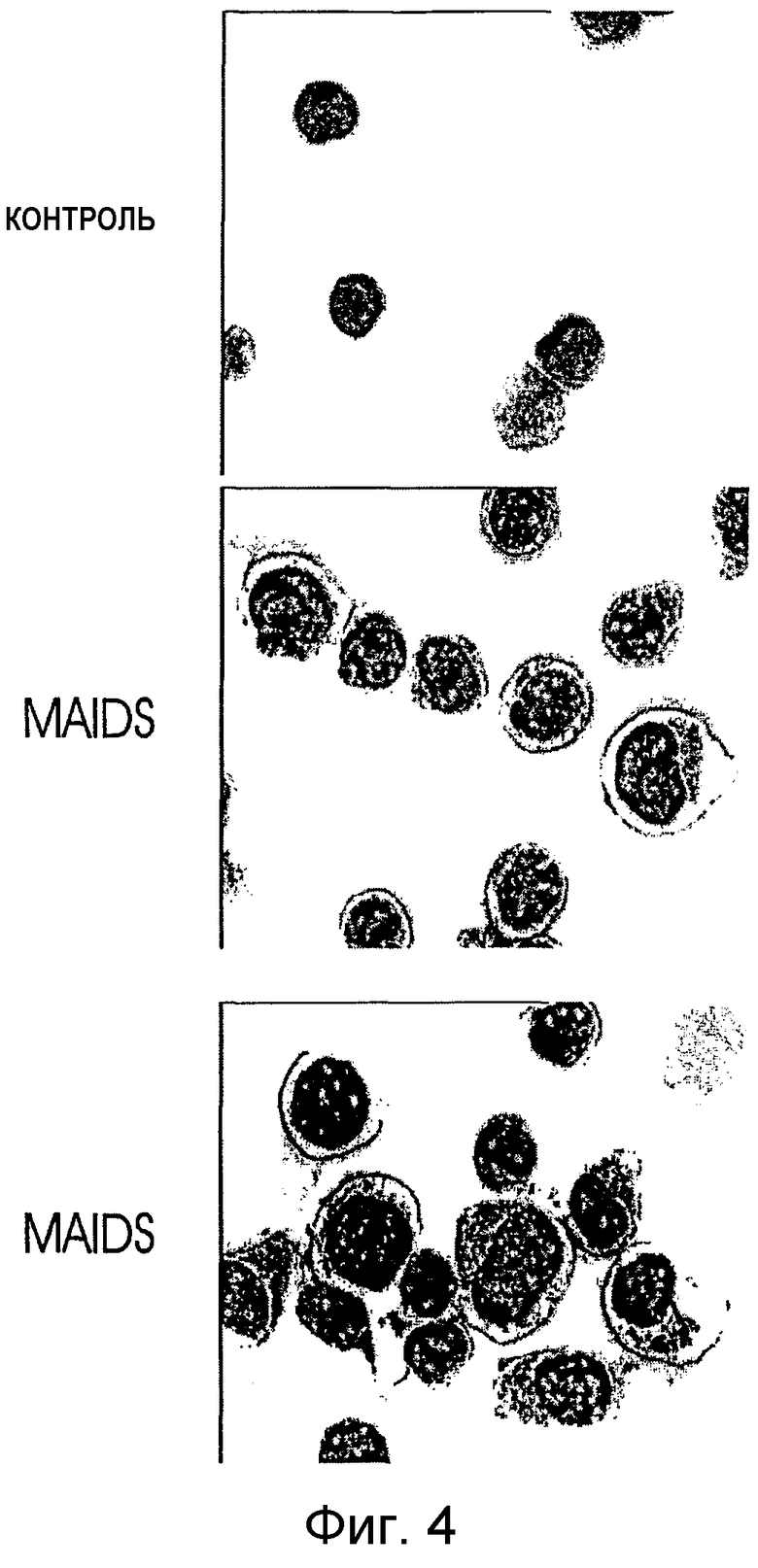
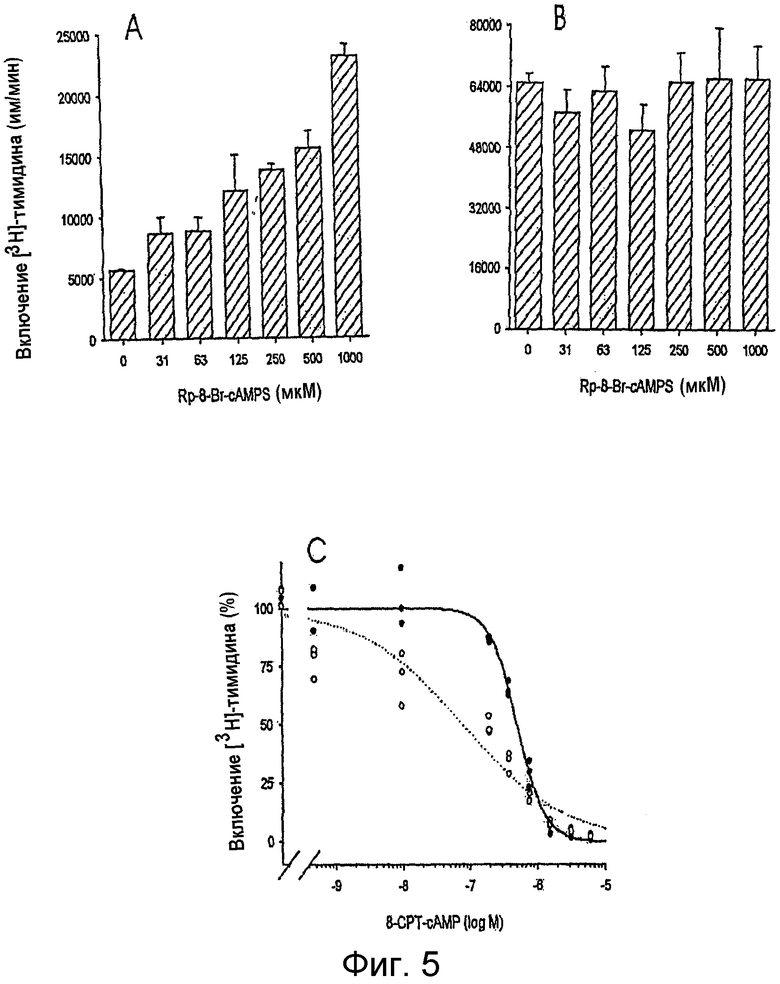

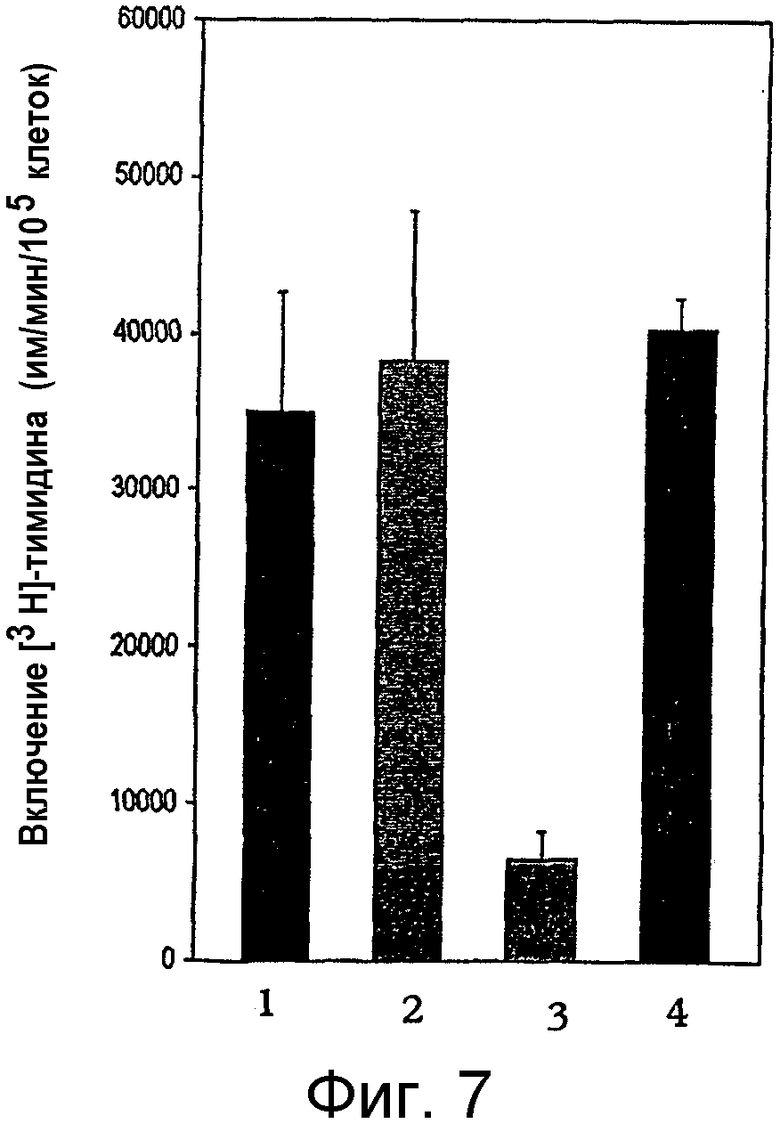
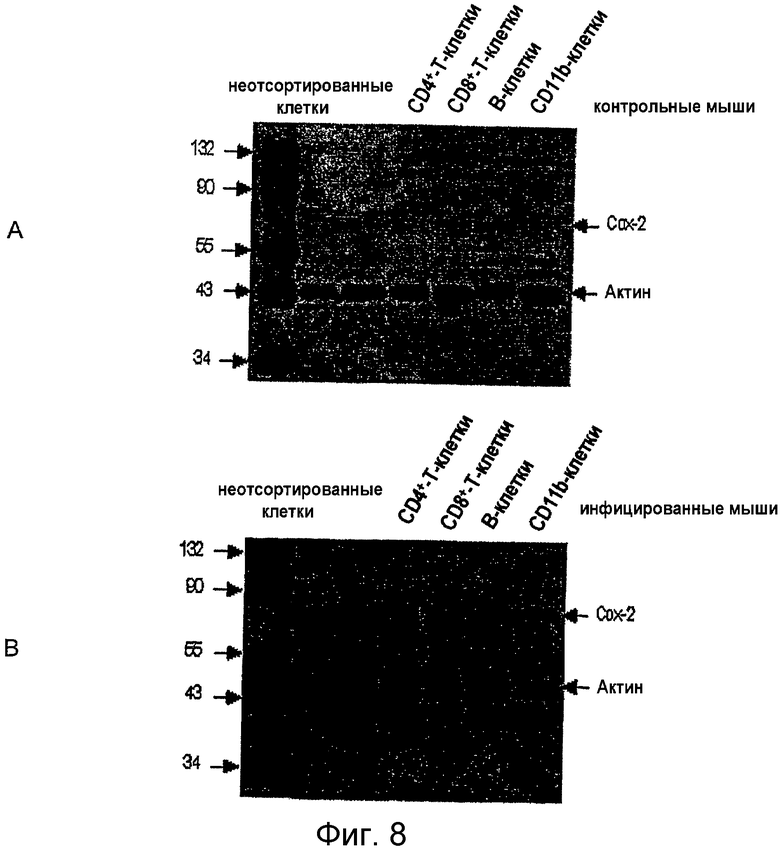
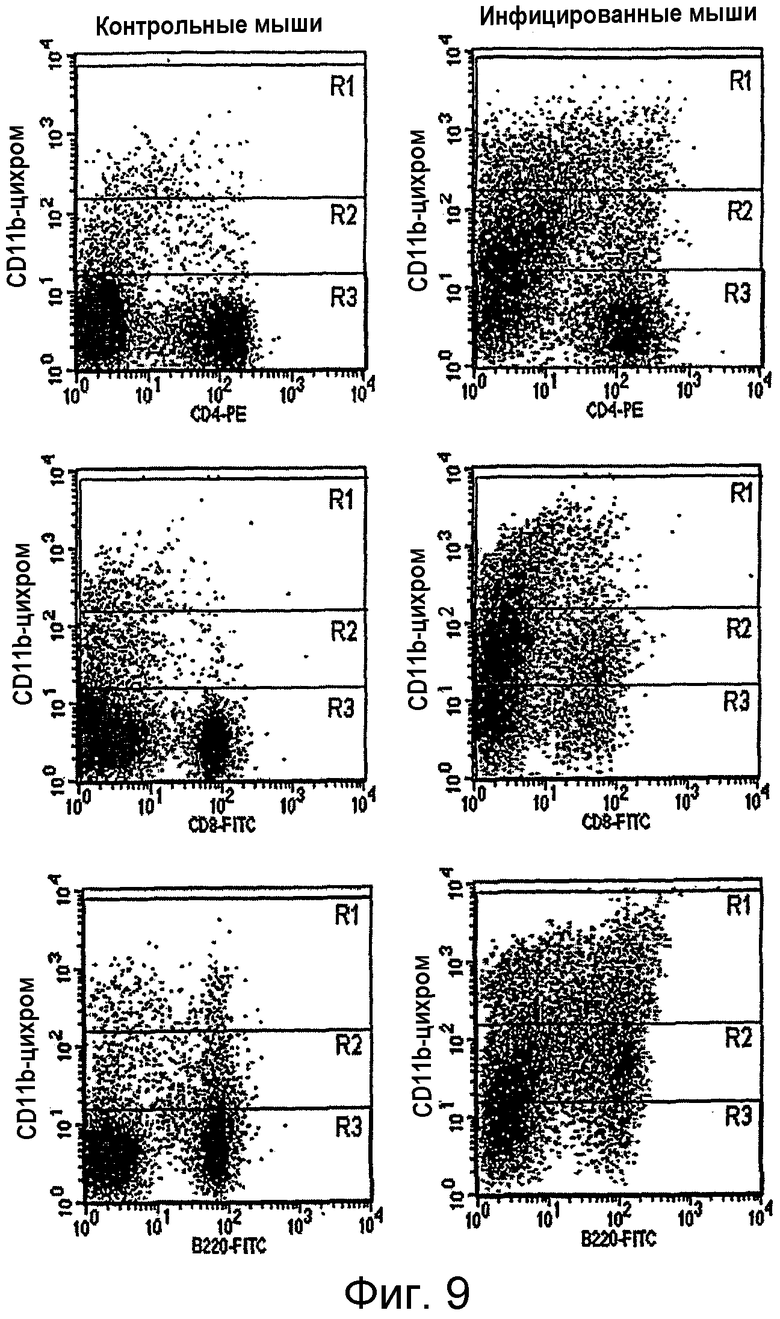

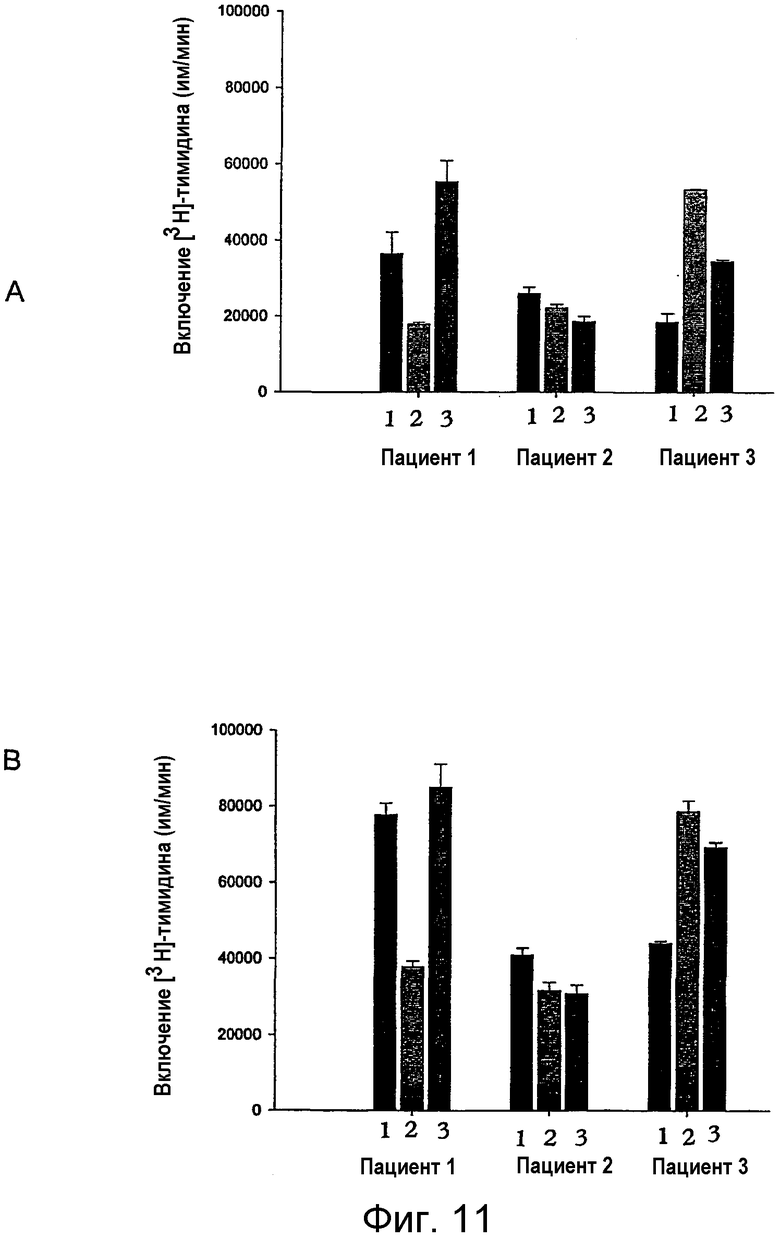

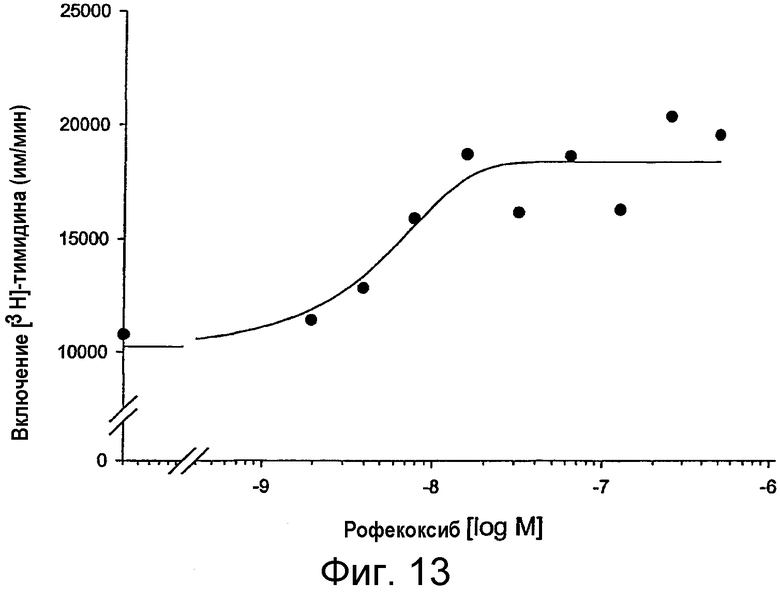
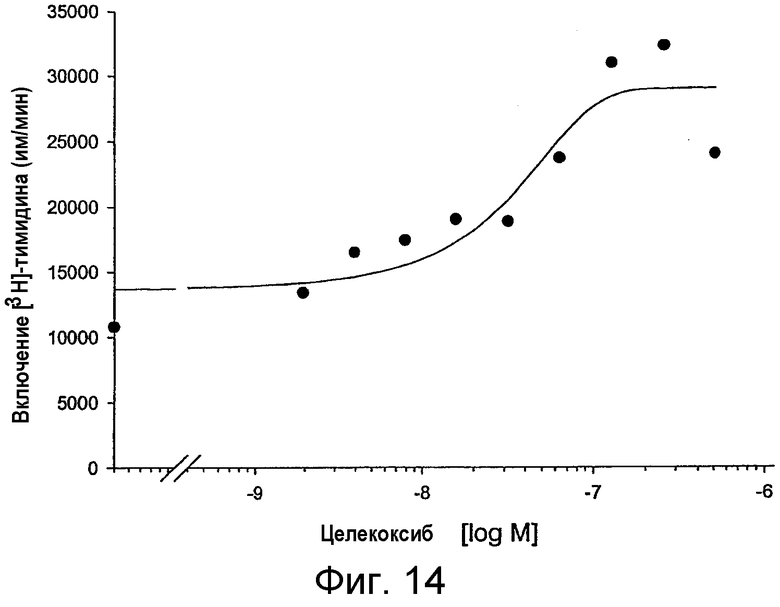
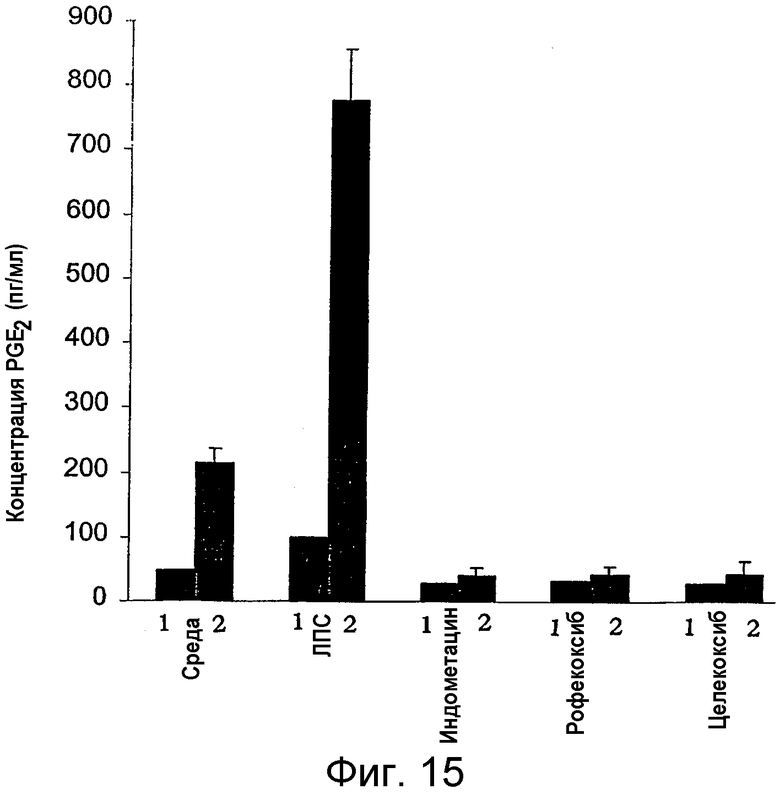
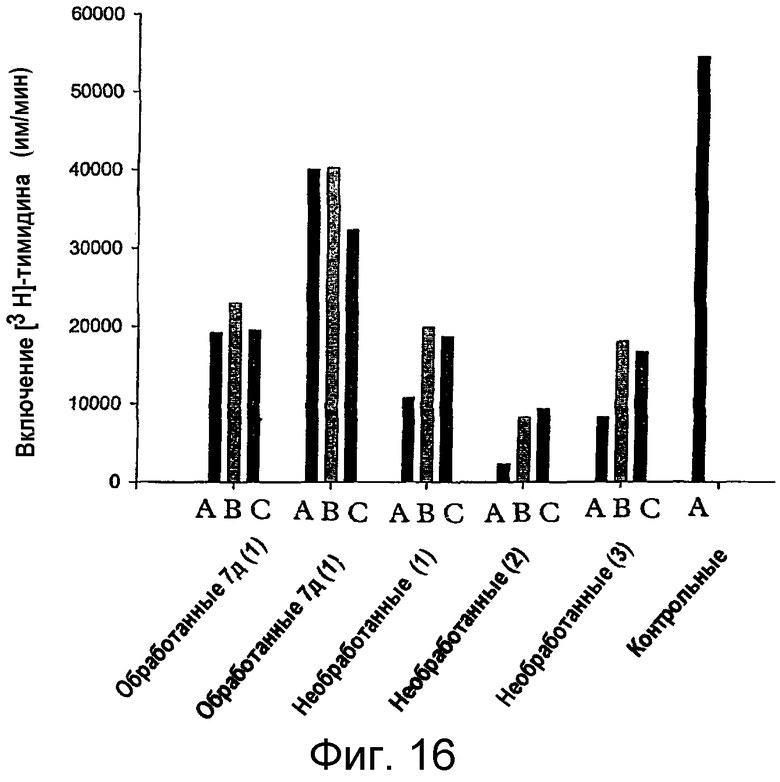
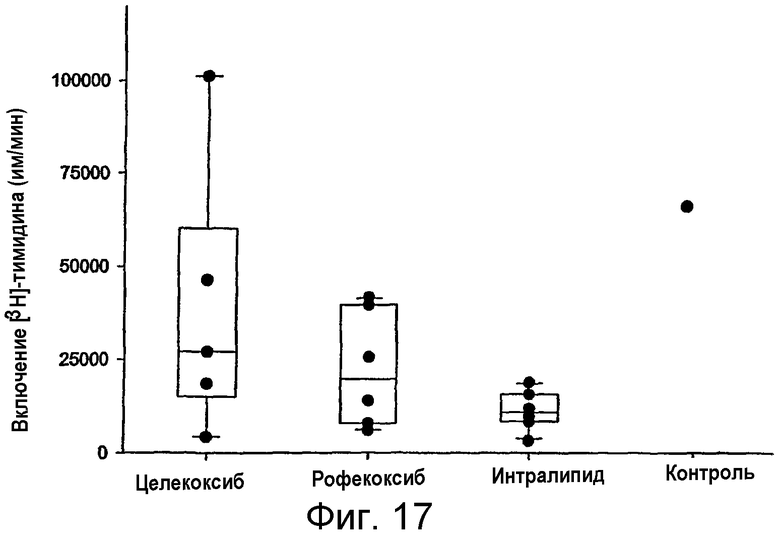
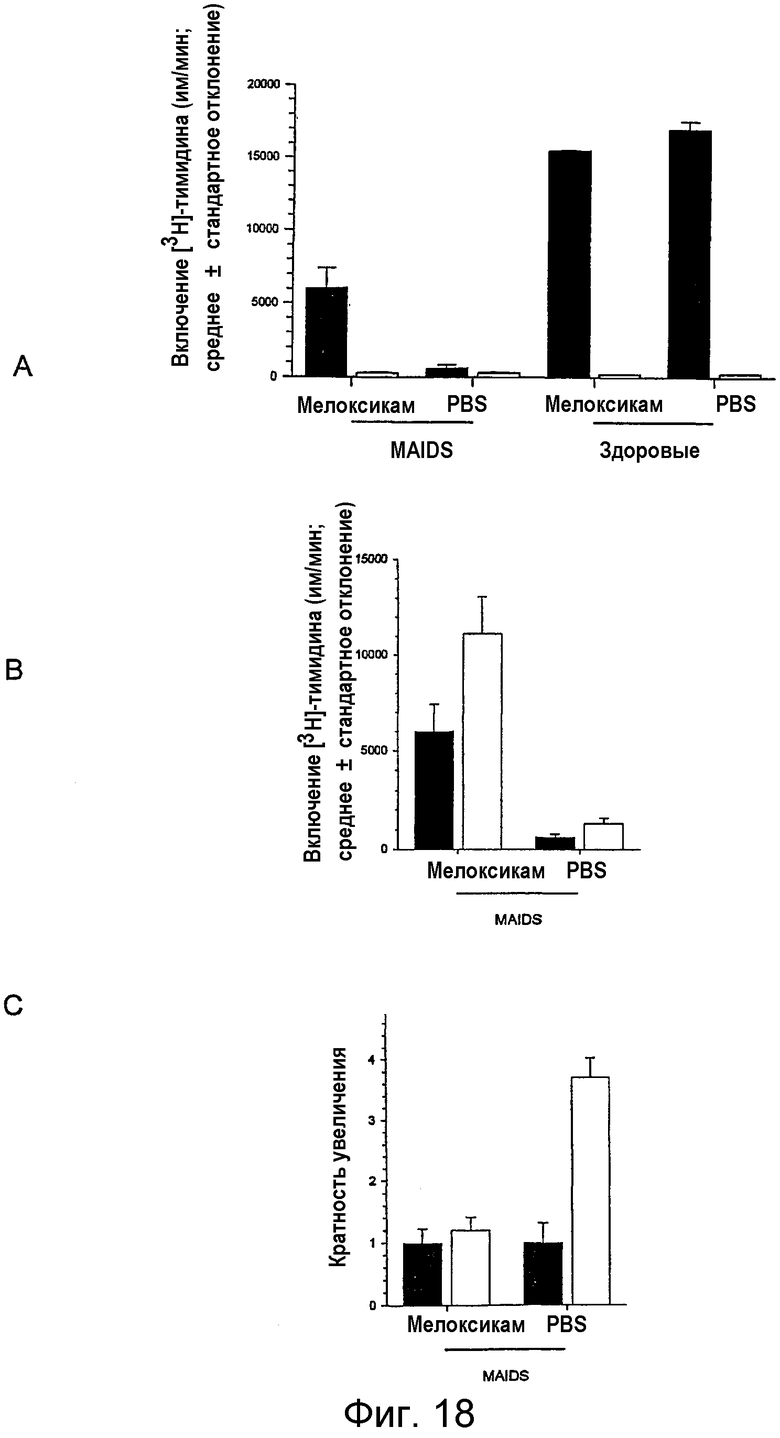
Настоящее изобретение относится к способу лечения или предупреждения инфекций, вызываемых родственным вирусом у животного, таких как SIV или FIV, или СПИД, а также к применению ряда ингибиторов циклооксигеназы изоформы 2 (СОХ-2) по указанному назначению. Предпочтительные ингибиторы COX-2: L-745337, рофекоксиб, NS398, SC58125, этодо-лак, мелоксикам, целекоксиб, DuP-697, DFU, трициклический MF, валдекоксиб, паракоксиб натрий, эторикоксиб или нимезулид - нестероидные противовоспалительные лекарственные средства. Они улучшают иммунные функции Т-клеток, снижают или отменяют иммунодефицит. 2 н. и 22 з.п. ф-лы, 18 ил., 6 табл.
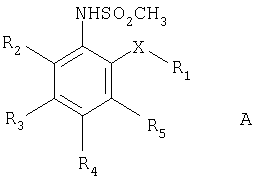
где Х представляет атом кислорода или серы или алкильную группу, предпочтительно группу -СН2-;
R1 представляет циклоалкильную или арильную группу, которые могут быть, но необязательно, замещены одной или более группами или атомами, предпочтительно одним или более атомами галогена; и
R2, R3, R4 и R5 независимо представляют атом водорода, нитро- или ацильную группу или алкильную группу, которая может быть, но необязательно, замещена одной или более группами или атомами, либо, альтернативно, R2 и R3, R3 и R4, или R4 и R5, взятые вместе с атомом углерода, с которым они связаны, образуют циклопентаноновую группу;
или его фармацевтически приемлемая соль, или указанным ингибитором СОХ-2 является соединение общей формулы В

где Y представляет циклическую группу, предпочтительно оксазолильную, изоксазолильную, тиенильную, дигидрофурильную, фурильную, пирролильную, пиразолильную, тиазолильную, имидазолильную, изотиазолильную, циклопентенильную, фенильную и пиридильную группу;
n представляет целое число от 0 до 3;
m представляет целое число от 0 до 4;
R6 представляет кетоциклильную, циклоалкильную или арильную группу, которая может быть, но необязательно, замещена одной или более группами или атомами, предпочтительно одним или более атомами галогена;
R7 каждый независимо представляет атом водорода или галогена или алкильную группу, которая может быть замещена одной или более группами или атомами; или ацильную группу; или оксогруппу;
R8 представляет алкильную группу или NHR10;
R9 представляет атом галогена и
R10 представляет атом водорода или алкильную группу, необязательно замещенную одной или более группами или атомами, предпочтительно ацильной группой;
или его фармацевтически приемлемая соль.
Приоритет от 20.07.2000 установлен по пп.15-17, 19-20, 22-24 в отношении инфекций, вызываемых вирусом СПИД, по пп.14, 21 - в отношении инфекций, вызываемых вирусом СПИД в части альтернатив L-74533 7, рофекоксиб, NS-398, SC-58125, этодолак, мелоксикам, целекоксиб, нимезулид, DuP-697, DFU, трициклический MF.
В отношении остальных пунктов и альтернатив пп.14,21 приоритет от 19.04.2001.
| Бесколесный шариковый ход для железнодорожных вагонов | 1917 |
|
SU97A1 |
| Приспособление в пере для письма с целью увеличения на нем запаса чернил и уменьшения скорости их высыхания | 1917 |
|
SU96A1 |
| СОЛЬ ЯНТАРНОЙ КИСЛОТЫ 4-[2-АМИНО-6-(ЦИКЛОПРОПИЛАМИНО)-9Н-ПУРИН-9-ИЛ]-2-ЦИКЛОПЕНТЕН-1-МЕТАНОЛА В КАЧЕСТВЕ ПРОТИВОВИРУСНОГО АГЕНТА | 1995 |
|
RU2145324C1 |
| Грэхам-Смит Д.Г | |||
| и др | |||
| Оксфордский справочник по клинической фармакологии и фармакотерапии | |||
| - М.: Медицина, 2000, с.25, 136-137 | |||
| WO 00/69255 A1, 23.11.2000 | |||
| US 6025353 A, 15.02.2000 | |||
| WO 00/00120 A1, 06.01.2000 | |||
| Бесколесный шариковый ход для железнодорожных вагонов | 1917 |
|
SU97A1 |
| US 5409944 А, 25.04.1995 | |||
| Прибор, замыкающий сигнальную цепь при повышении температуры | 1918 |
|
SU99A1 |
| US 5543297 А, 06.08.1996 | |||
| US 6025353 A, 15.02.2000. | |||

