Область и предпосылки к созданию изобретения
Настоящее изобретение относится к вакцине и способам лечения болезней двигательных нейронов (MND), особенно бокового амиотрофического склероза (ALS).
Болезнь двигательных нейронов (MND) - это название, присвоенное группе родственных заболеваний, поражающих двигательные нейроны головного мозга (верхние двигательные нейроны) и спинного мозга (нижние двигательные нейроны). Двигательные нейроны (или мотонейроны) представляют собой нервные клетки, по которым головной мозг посылает команды мышцам в форме электрических импульсов. Дегенерация двигательных нейронов приводит к слабости и гипотрофии мышц. Это обычно наблюдается сначала на руках и ногах, причем некоторые группы мышц поражаются более других.
Существует несколько классификаций MND. В большинстве случаев MND наблюдается дегенерация как верхних, так и нижних двигательных нейронов. Указанное состояние называется боковым амиотрофическим склерозом (ALS), известным также как болезнь Шарко, которая характеризуется мышечной слабостью, ригидностью и фасцикуляциями (мышечными подергиваниями). Существуют также менее распространенные формы, при которых наблюдается более выборочная дегенерация верхних двигательных нейронов (такие как первичный боковой склероз, PLS) или нижних двигательных нейронов (такие как прогрессивная мышечная атрофия, РМА). Прогрессивный бульбарный паралич (РВР или бульбарное начало) представляет собой разновидность ALS, который начинается с затруднения глотания, жевания и речи и поражает приблизительно 25% пациентов с ALS.
Указанные формы MND во многом не имеют четких разграничений. У людей, страдающих РМА, со временем в болезнь вовлекаются верхние двигательные нейроны, а при РМА и ALS у некоторых людей в конечном итоге могут появиться затруднения речи и глотания различной степени тяжести (бульбарное начало ALS или РМА).
ALS представляет собой хроническое прогрессирующее нейродегенеративное заболевание, которое характеризуется постепенной дегенерацией нервных клеток центральной нервной системы (ЦНС), которая контролирует волевые движения мускулатуры. Прогрессирующая утрата двигательных нейронов приводит к постепенной атрофии скелетной мускулатуры и к неизбежной смерти, обычно через 2-3 и до десяти лет после начала болезни. Слабость и атрофия мышц, а также признаки дисфункции клеток переднего рога сначала наиболее часто отмечаются в кистях рук и менее часто - в стопах. Область начала заболевания является случайной, а прогрессирование - асимметричным. Только в США в настоящее время больны ALS 30000 человек, и каждый год регистрируется около 8000 новых случаев.
ALS наблюдается в спорадической (SALS) и семейной (FALS) формах (Mulder et al., 1986; Munsat, 1989). Главные факторы риска по большей части неизвестны, однако от 5 до 10% всех пациентов с ALS имеют наследственную форму (FALS). Около 20% всех семейных форм, как было установлено, имеют мутации в гене, кодирующем Cu/Zn супероксиддисмутазу 1 типа, в 21 хромосоме (Rosen et al., 1993; Brown, 1995). SOD представляет собой фермент, который катализирует превращение супероксид-анионов в пероксид водорода, и, таким образом, SOD может защищать клетки от повреждающего действия указанных токсичных радикалов. Представляется, что токсичность различных мутантов SOD не обусловлена уменьшением активности устранения свободных радикалов, поскольку не было обнаружено корреляции между ферментативной активностью, полупериодом существования полипептида и резистентности к протеолизу и возрастом начала или скоростью прогрессирования заболевания у человека (см. обзор Julien, 2001). У трансгенных мышей, экспрессирующих различные мутанты SOD1, развивается болезнь двигательных нейронов, и, таким образом, они представляют собой принятую модель для испытаний лекарственных средств для лечения ALS и других болезней двигательных нейронов.
Недавно двумя независимыми группами ученых был обнаружен новый ген ALS (Hadano et al., 2001; Yang et al., 2001). Указанный новый ген, названный ALS2, располагается во 2 хромосоме и кодирует белок, названный алсином. Новый ген ALS2 мутирован у людей с ювенильным боковым амиотрофическим склерозом (JALS), известным также как ALS2, и у людей с ювенильным первичным боковым склерозом (JPLS). Мутации в различных областях указанной хромосомы связаны с различными болезнями двигательных нейронов. Конкретно, у людей с ALS обнаружена мутация в одной области, в то время как у людей с JPLS обнаружены мутации в двух других областях. В будущем трансгенные мыши, несущие указанные мутации, будут несомненно служить еще одной моделью для испытаний лекарственных средств для лечения ALS.
В течение последнего десятилетия многочисленные исследования посвящались изучению этиологии, прогноза и прогрессирования болезни. Консенсус достигнут не был, за исключением принятой идеи о том, что данное заболевание является многофакторным с точки зрения обстоятельств, которые приводят к его прогрессированию, в то время как этиология остается неясной.
В настоящее время очевидно, что многие факторы, которые вносят свой вклад в прогрессирование ALS, обнаруживаются и при многих других хронических и острых нейродегенеративных расстройствах. Указанные факторы включают в себя окислительный стресс, токсические факторы, лишение трофической поддержки и ионный дисбаланс. Многие годы предпринимались попытки остановить прогрессирование ALS, как при других хронических и острых нейродегенеративных расстройствах, путем блокирования различных медиаторов цитотоксичности. Большинство указанных клинических испытаний дали отрицательные результаты (Turner et al., 2001).
Окислительный стресс характеризуется накоплением свободных радикалов, которое может приводить к гибели двигательных нейронов. Свободные радикалы повреждают компоненты клеточных мембран, белков или генетического материала путем их «окисления». Указанные свободные радикалы могут вырабатываться, когда нарушается функция фермента SOD, в силу генетической мутации, как это происходит у некоторых пациентов с семейным ALS, или из-за химической среды, окружающей нервные клетки, или они могут вырабатываться при повышении токсичности, опосредованной глутаматом, или по какой-либо другой причине. Многие пациенты с ALS принимают кофермент Z Q10 и витамин Е в попытке нейтрализовать свободные радикалы.
Глутамат представляет собой один из наиболее распространенных медиаторов токсичности при острых и хронических дегенеративных расстройствах (Pitt et al., 2000), таких как эпилептический статус, ишемия головного мозга, травматическое повреждение головного мозга, ALS, хорея Гентингтона, латиризмы и болезнь Альцгеймера. Глутамат представляет собой главный нейромедиатор возбуждения в ЦНС человека. L-глутамат присутствует в большинстве синапсов и способен проявлять двойственную активность: он играет основную роль в нормальном функционировании как главный нейромедиатор, но становится токсичным, когда его физиологические уровни превышены.
Что касается спинальных двигательных нейронов, быстрое удаление глутамата после срабатывания синапса осуществляется переносчиком глутамата ЕААТ2, присутствующим в астроцитах. Уменьшение активности ЕААТ2 и уровня белка было обнаружено в ткани головного мозга пациентов с ALS (Rothstein et al., 1992). Это может приводить к повышению внеклеточной концентрации глутамата и гибели двигательных нейронов. В клинике благоприятное действие препарата Riluzole, ингибитора высвобождения глутамата, на течение заболевания у людей и трансгенных мышей приводит к улучшению медикаментозного лечения ALS. Однако при нейтрализации токсического эффекта он, возможно, препятствует физиологическому функционированию глутамата как повсеместно действующего нейромедиатора ЦНС.
Роль иммунных факторов, клеточных и молекулярных, в ALS обсуждается годами. Было доказано, как и при многих других нейродегенеративных заболеваниях, что воспаление связано с развитием болезни, и было предложено использовать иммунодепрессантные лекарственные средства для лечения ALS. Также у многих пациентов с ALS наблюдается корреляция с наличием антиганглиозидных антител, что навело некоторых исследователей на мысль о том, что ALS представляет собой аутоиммунное заболевание. Однако в поддержку данной гипотезы исчерпывающих доказательств нет.
В лаборатории авторов настоящего изобретения недавно было установлено, что при нейродегенеративных состояниях, вызванных механическими (аксотомия) или биохимическими (глутамат, окислительный стресс) повреждениями, иммунная система играет главную роль. Так, было установлено, что активированные Т-клетки, которые распознают антиген нервной системы (НС), способствуют регенерации нервов или обеспечивают нейропротекцию. Приводится ссылка на публикацию РСТ № WO 99/60021, содержание которой целиком включено в настоящий документ в качестве ссылки. Более конкретно, Т-клетки, реагирующие на МВР, как было показано, обладают нейропротективным действием у крыс с частично разрушенным зрительным нервом (Moalem et al., 1999) и повреждением спинного мозга (Hauben et al., 2000). До недавнего времени полагали, что иммунная система исключает иммунные клетки из участия в восстановлении нервной системы. Было достаточно неожиданным открытие, что специфичные в отношении НС активированные Т-клетки можно использовать для ускорения регенерации нервов или для защиты ткани нервной системы от вторичной дегенерации, которая может следовать за повреждением, вызванным травмой или болезнью ЦНС или периферической нервной системы (ПНС).
Авторы также установили, что в стрессовых условиях в ЦНС запускается адаптивный иммунный ответ для преодоления стресса, и что данный ответ контролируется на генетическом уровне. Так, коэффициент выживания ганглионарных клеток сетчатки у взрослых мышей или крыс после повреждения зрительного нерва в результате сдавливания или инъекции в стекловидное тело токсической дозы глутамата, как было показано, был до двух раз выше у животных тех линий, которые являются резистентными к аутоиммунным заболеваниям ЦНС, чем в чувствительных к ним линиях. Как было установлено, данное различие было обязано своим происхождением благоприятному аутоиммунному Т-клеточному ответу, который спонтанно возникал после повреждения ЦНС, у резистентных, но не у чувствительных линий. Так, коэффициент выживания нейронов после подобного повреждения выше, когда возникает аутоиммунный Т-клеточный ответ, при условии, что он должным образом регулируется. Иными словами, было показано, что защитный аутоиммунный ответ возникает как противодействие стрессовым условиям, чтобы защитить животное от последствий повреждения. Было установлено также, что у животных с нарушенной способностью регулировать указанный ответ или у животных, лишенных зрелых Т-клеток (в результате удаления тимуса после рождения), способность справляться со стрессовыми условиями снижена. Как следствие, коэффициент выживания нейронов после повреждения ЦНС у данных животных значительно ниже, чем у животных, обладающих эффективным механизмом для стимуляции защитного аутоиммунного, опосредованного Т-клетками, ответа (Kipnis et al., 2001).
Далее заявителями было также установлено, что вакцинация непатогенными синтетическими сополимерами, которые напоминают собственные белки, такими как сополимер 1 (Сор-1 или Glatiramer), сополимер, составленный случайным образом из четырех аминокислот: тирозин-глутамат-аланин-лизин (далее в настоящем документе упоминается как «Сор-1»), и poly-Glu,Tyr (далее в настоящем документе упоминается как «Poly-YE»), и активированные им Т-клетки, после травматического повреждения ЦНС можно использовать для усиления защитного аутоиммунитета и, следовательно, для уменьшения дальнейшего индуцированного повреждением поражения, и можно в дальнейшем защищать клетки ЦНС от токсичности глутамата. Приводится ссылка на предыдущие патентные заявки авторов США №№ 09/756301 и 09/765644, датированные 22 января 2001 г., включенные целиком в настоящий документ в качестве ссылок, как если бы они были описаны здесь полностью, соответствующие WO 01/93893, в которой описано, что Сор-1, родственные Сор-1 пептиды и полипептиды, а также активированные ими Т-клетки защищают клетки ЦНС от токсичности глутамата (USSN 09/756301) и предотвращают или ингибируют дегенерацию нейронов или способствуют регенерации нервов в ЦНС или ПНС (USSN 09/765644). Приводится также ссылка на предыдущую патентную заявку Соединенных Штатов, принадлежащую авторам, № 09/893344, датированную 28 июня 2001 г., включенную целиком в настоящий документ в качестве ссылки, как если бы она была описана здесь полностью, в которой описано, что сополимер poly-Glu50Tyr50, прежде называвшийся polyGT и обозначаемый также Poly-YE, а также активированные им Т-клетки защищают клетки ЦНС от токсичности глутамата и предотвращают или ингибируют дегенерацию нейронов или способствуют регенерации нервов в ЦНС или ПНС. Конкретно, в указанных заявках было показано, что в волокнах зрительного нерва количество выживших ганглионарных клеток сетчатки было достоверно больше у мышей, иммунизированных Сор-1 или poly-Glu,Tyr, чем у мышей, которым инъецировали PBS.
Единственным лекарственным средством, разрешенным и имеющимся в настоящее время для лечения ALS, является Riluzole (2-амино-6-(трифторметокси)бензотиазол), предполагаемый блокатор высвобождения глутамата, который, как представляется, обладает некоторым уменьшающим спазмы действием при данном состоянии, возможно, посредством ингибирования глутаматергической передачи в ЦНС. Его вводят перорально в форме таблеток. Riluzole не излечивает болезнь и не облегчает симптомы. Он оказывает действие от умеренного до значительного на пациентов с ALS, удлиняя срок их жизни приблизительно на 3 месяца, но не улучшает мышечную силу или неврологическую функцию.
Весьма желательным является создание других лекарственных средств для лечения болезней двигательных нейронов, включая ALS.
Цитирование или идентификация любой ссылки в данном разделе или любой другой части данной заявки не должны истолковываться как допущение, что данная ссылка является доступной в качестве прототипа настоящего изобретения.
Краткое описание существа изобретения
В настоящее время установлено в соответствии с настоящим изобретением, что иммунизация Сор-1 может защищать трансгенных мышей с избыточной экспрессией SOD1 и мышей после аксотомии лицевого нерва, обе модели ALS, от дегенерации двигательных нейронов. Это, а также и тот факт, что Сор-1 и Poly-YE являются эффективными для защиты ганглионарных клеток сетчатки от токсичности глутамата, указывает на то, что данные полимеры являются подходящими для лечения болезней двигательных нейронов, особенно ALS.
Настоящее изобретение, таким образом, относится, в одном аспекте, к способу уменьшения прогрессирования болезни, защиты от дегенерации двигательных нейронов и/или защиты от токсичности глутамата у пациента, страдающего болезнью двигательных нейронов (MND), который включает в себя иммунизацию указанного пациента вакциной, содержащей активный агент, выбранный из группы, состоящей из Сор-1, родственного Сор-1 пептида, родственного Сор-1 полипептида и Poly-YE.
Болезнь двигательных нейронов (MND) представляет собой любую болезнь, поражающую двигательные нейроны головного мозга и спинного мозга, и включает в себя боковой амиотрофический склероз (ALS), как семейный (FALS), так и спорадический (SALS) ALS, первичный боковой склероз (PLS), прогрессивную мышечную атрофию (РМА), прогрессивный бульбарный паралич (РВР или бульбарное начало) и их комбинированные формы, такие как бульбарное начало ALS и бульбарное начало РМА.
В одном варианте осуществления способ согласно изобретению включает в себя лечение также препаратом Riluzole или любым другим лекарственным средством, подходящим для лечения MND, особенно ALS.
В другом аспекте настоящее изобретение относится к вакцине для уменьшения прогрессирования болезни, для защиты от дегенерации двигательных нейронов и/или для защиты от токсичности глутамата при болезни двигательных нейронов (MND), особенно ALS, содержащей активный агент, выбранный из группы, состоящей из Сор-1, родственного Сор-1 пептида, родственного Сор-1 полипептида и poly-Glu, Tyr.
В еще одном аспекте настоящее изобретение относится к применению активного агента, выбранного из группы, состоящей из Сор-1, родственного Сор-1 пептида, родственного Сор-1 полипептида и poly-Glu,Tyr, для производства вакцины для уменьшения прогрессирования болезни, для защиты от дегенерации двигательных нейронов и/или для защиты от токсичности глутамата при болезни двигательных нейронов (MND), особенно ALS.
Активный агент можно вводить без адъюванта или его можно эмульгировать в адъюванте, подходящем для клинического применения у человека. Адъювант, подходящий для клинического применения у человека, выбирают из группы, состоящей из гидроксида алюминия, геля гидроксида алюминия и гидроксифосфата алюминия. В предпочтительном варианте осуществления настоящего изобретения вакцинный адъювант представляет собой аморфный гидроксифосфат алюминия, имеющий кислую изоэлектрическую точку и соотношение Al:P 1:1 (в настоящем документе упоминается как Alum-phos).
В одном предпочтительном варианте осуществления настоящего изобретения активный агент вакцины согласно изобретению представляет собой Сор-1. В другом предпочтительном варианте осуществления настоящего изобретения активный агент представляет собой poly-Glu, Tyr.
Помимо этого, вакцину можно вводить по схеме, которая включает в себя введение Riluzole или другого лекарственного средства, подходящего для лечения ALS.
Краткое описание фигур
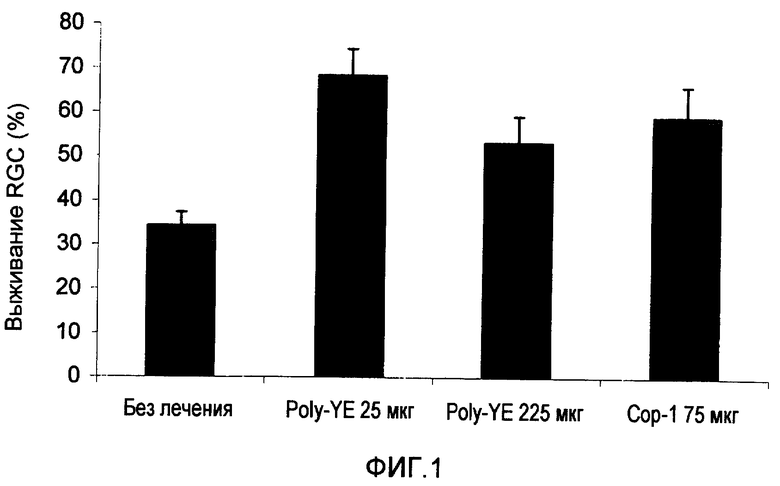
На фиг.1 показано, что иммунизация Сор-1 или Poly-YE без адъюванта защищает ганглионарные клетки сетчатки (RGC) мышей от токсичности глутамата.
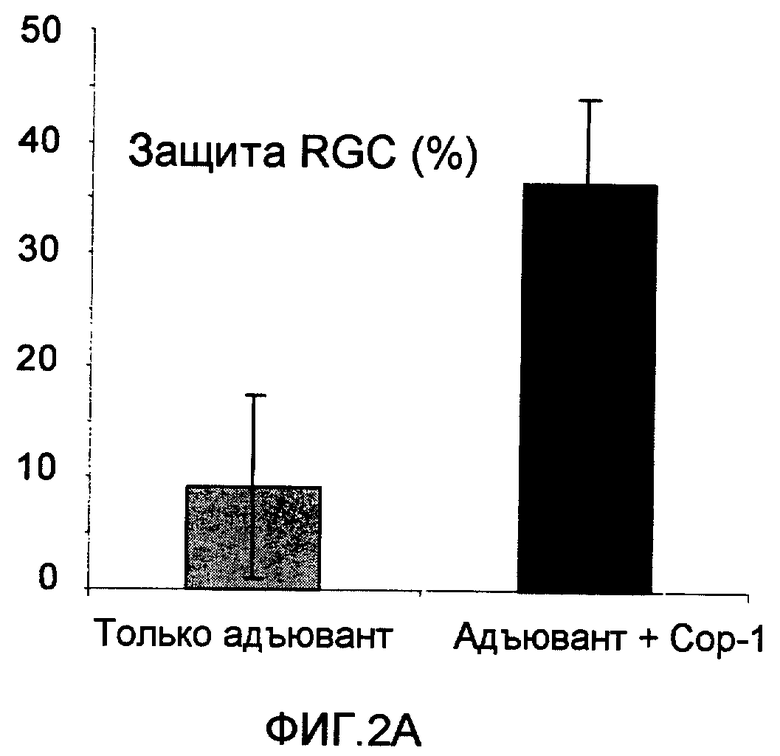
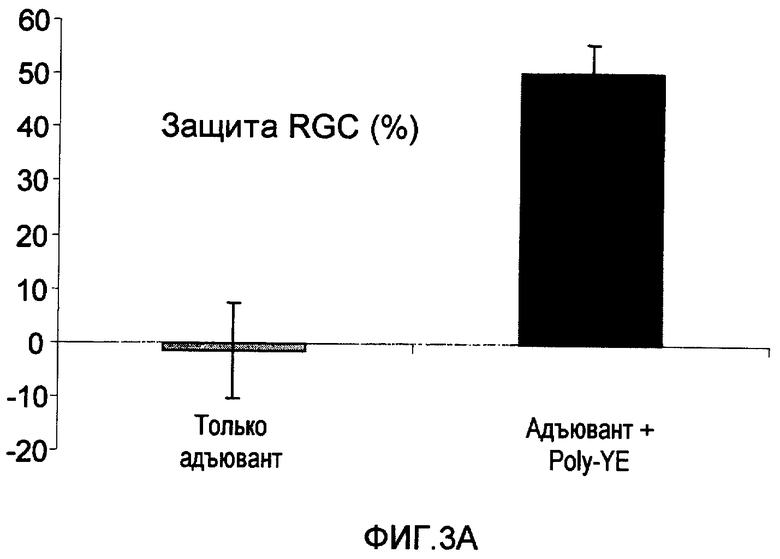
На фиг.2А-В показано, что иммунизация Сор-1 (фиг.2А) или Poly-YE (фиг.2В) в адъюванте (CFA) защищает RGC мышей от токсичности глутамата.
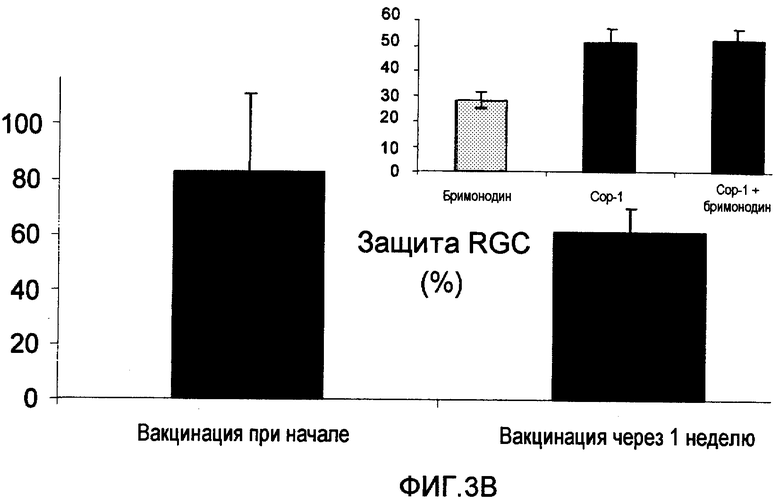
На фиг.3А-В показано влияние иммунизации Poly-YE (фиг.3А) или Сор-1 (фиг.3В) на выживание RGC на модели внутриглазного давления (IOP) при глаукоме.
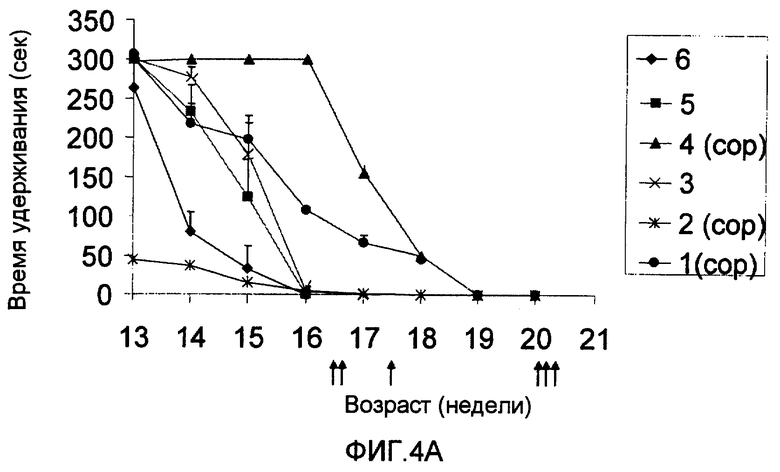
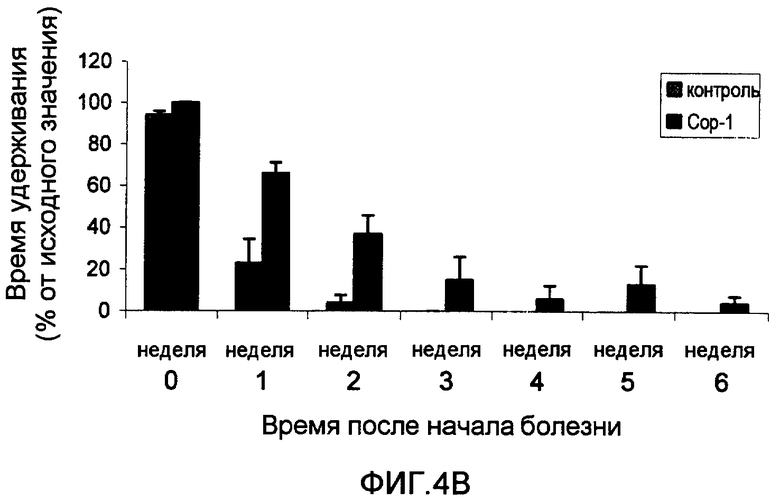
На фиг.4А-В приведены результаты теста на мышечную силу, выполненного на трансгенных мышах, экспрессирующих избыточное количество человеческой мутантной SOD1 (далее в настоящем документе упоминаются как «мыши с ALS»). На фиг.4А показано среднее время (в секундах) за неделю, в течение которого мыши с ALS, иммунизированные Сор-1, эмульгированным в Alum-phos (мыши 1, 2 и 4), и неиммунизированные трансгенные мыши (мыши 3, 5 и 6) удерживаются на вращающемся вертикальном стержне. На фиг.4В изображено среднее время (в % от исходного значения), в течение которого удерживаются 3 мыши с ALS, иммунизированные Сор-1 в Alum-phos (черные столбцы), и 3 трансгенные неиммунизированные мыши (контроль, серые столбцы). Для сравнения скорости прогрессирования болезни всех животных синхронизировали по времени начала мышечной слабости (время 0), стандартизируя время удерживания каждого животного по его собственному исходному времени перед началом заболевания (исходное время - 100%). На фигуре изображено среднее ±SEM время удерживания для каждой группы в течение следующих недель прогрессирования болезни.
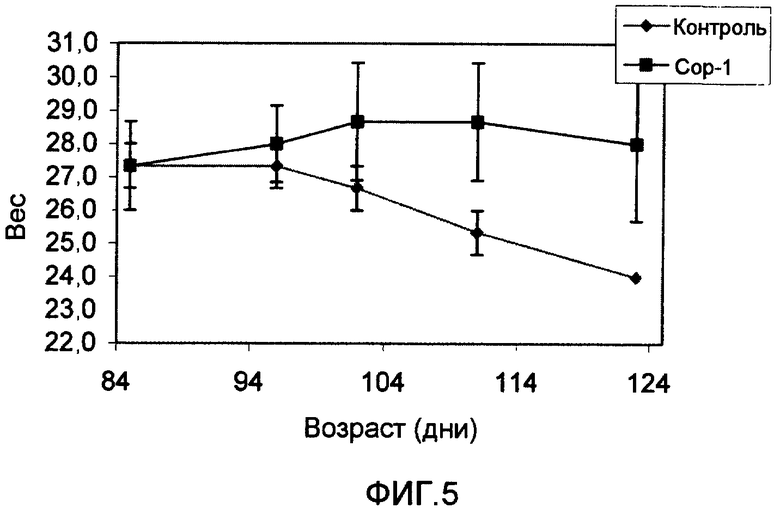
На фиг.5 показано сохранение массы тела мышей с ALS, иммунизированных Сор-1 в Alum-phos (черные квадраты), по сравнению с неиммунизированными мышами (серые звездочки).
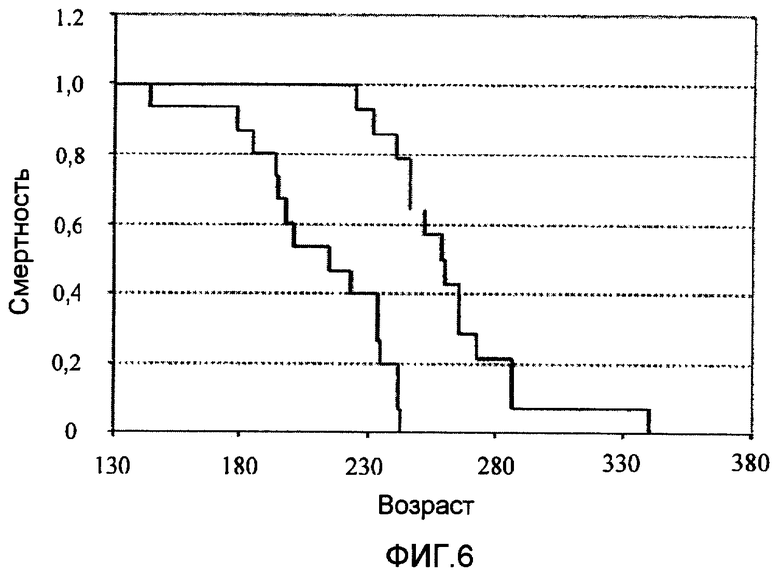
Фиг.6 представляет собой график, показывающий предполагаемую продолжительность жизни мышей с ALS, иммунизированных Сор-1 в CFA. Паралич вызван прогрессирующей утратой спинным мозгом двигательных нейронов. У невакцинированных контролей (n=15) развился паралич одной или более конечностей, и они погибли в возрасте 211±7 дней (среднее ±SD). Мыши, получившие Сор-1, выживали в течение 263±8 дней.
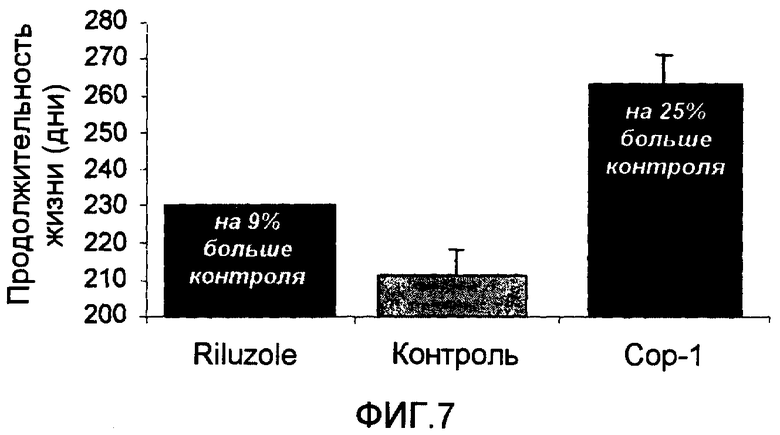
На фиг.7 показана предполагаемая продолжительность жизни мышей с ALS, иммунизированных Сор-1 в CFA, и мышей с ALS, получавших Riluzole. Мыши с ALS, получавшие Riluzole, и мыши с ALS, иммунизированные Сор-1, показали, соответственно, 9% и 25% увеличение, по сравнению с невакцинированными контрольными мышами.
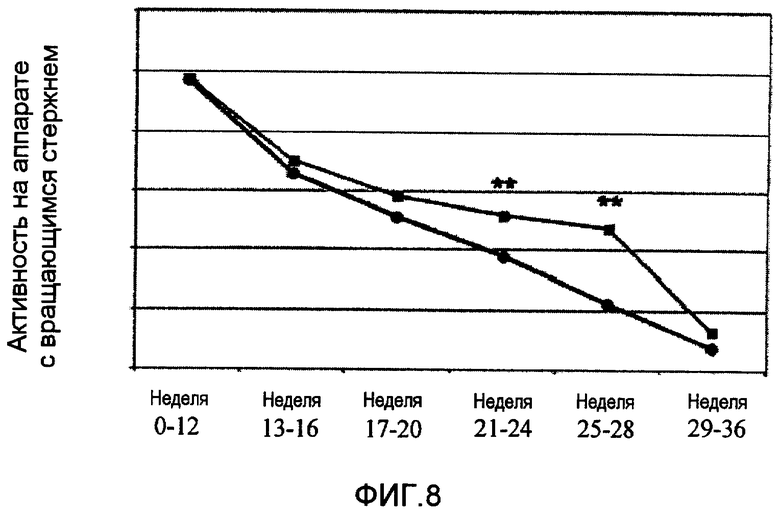
На фиг.8 показана средняя активность вращения, измеренная в указанные временные точки, у получавших Сор-1 и ничего не получавших мышей с ALS. Мышам давали схватиться и держаться на вертикальной проволоке (диаметром 2 мм) с маленькой петлей на нижнем конце. Их активность отмечали индивидуально с помощью компьютеризованной системы и оценивали ежедневно. Для статистической оценки активность на вращающемся стержне стандартизировали по средней активности каждой мыши от 40 дня до 60 дня. Данные выражали как среднее ± стандартная ошибка среднего (SEM). Наблюдалась достоверная разница между леченными и нелеченными мышами в следующие периоды времени: между 12 и 20 днями (P<0,058), между 21 и 24 днями (P<0,0079) и между 25 и 28 днями (P<0,0017).
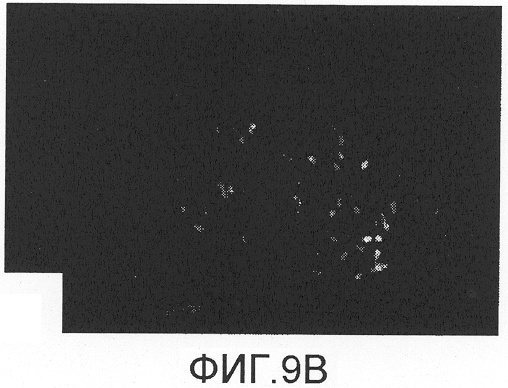
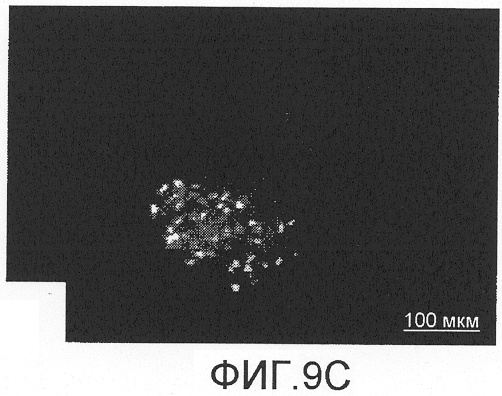
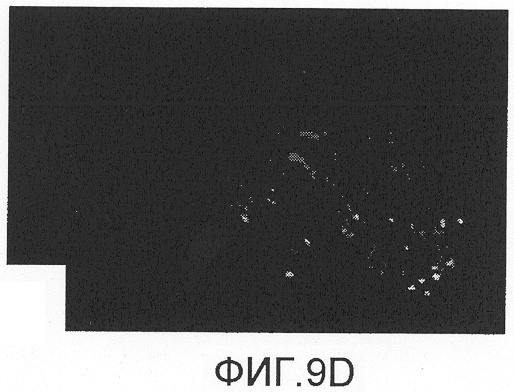
На фиг.9А-D показано спасение двигательных нейронов с помощью Сор-1, введенного мышам после аксотомии лицевого нерва. Через восемь недель после аксотомии количество двигательных нейронов, меченных FluoroGold, в стволе головного мозга мышей, вакцинированных Сор-1 (фиг.9D), было достоверно больше, чем в группе, которой инъецировали PBS в CFA (фиг.9В). Лечение Сор-1 не оказывало влияния на количество двигательных нейронов в неповрежденном ядре лицевого нерва (фиг.9А, 9С). Контрольная иммунизация PBS в CFA не показывала защитного действия.
Подробное описание изобретения
Настоящее изобретение относится к вакцине и способу уменьшения прогрессирования болезни, защиты от дегенерации двигательных нейронов, увеличения продолжительности жизни и улучшения качества жизни и/или защиты от токсичности глутамата у пациента, страдающего MND, особенно ALS, который включает в себя иммунизацию указанного пациента вакциной, содержащей активный агент, выбранный из группы, состоящей из Сор-1, родственного Сор-1 пептида, родственного Сор-1 полипептида или Poly-YE, без адъюванта или эмульгированного в адъюванте, подходящем для клинического применения у человека.
Используемые в настоящем документе термины "двигательные нейроны" и "мотонейроны", термины "Poly-YE" и "poly-Glu,Tyr" и термины «Сор-1» и «сополимер 1» являются взаимозаменяемыми.
Для целей настоящего изобретения «Сор-1 или родственный Сор-1 пептид или полипептид» включает в себя любой пептид или полипептид, включая случайный сополимер, который функционально перекрестно реагирует с основным белком миелина (МВР) и способен конкурировать с МВР или МНС класса II в антигенной презентации.
Вакцина согласно изобретению может включать в себя в качестве активного агента сополимер, составленный случайным образом, содержащий подходящее количество положительно заряженных аминокислот, таких как лизин или аргинин, в комбинации с отрицательно заряженной аминокислотой (предпочтительно в меньшем количестве), такой как глутаминовая кислота или аспарагиновая кислота, необязательно в комбинации с незаряженной нейтральной аминокислотой, такой как аланин или глицин, служащей наполнителем, и, необязательно, с аминокислотой, адаптированной для сообщения сополимеру иммуногенных свойств, такой как ароматическая аминокислота, подобная тирозину или триптофану. Указанные вакцины могут включать в себя любой из сополимеров, описанных в WO 00/05250, который целиком включен в настоящий документ в качестве ссылки.
Более конкретно, вакцина для применения согласно изобретению включает в себя по меньшей мере один сополимер, выбранный из группы, состоящей из составленных случайным образом сополимеров, содержащих одну аминокислоту, выбранную по меньшей мере из трех из следующих групп: (а) лизин и аргинин; (b) глутаминовая кислота и аспарагиновая кислота; (с) аланин и глицин и (d) тирозин и триптофан.
Сополимеры для применения согласно изобретению могут состоять из L- или D-аминокислот или их смесей. Как известно специалистам, L-аминокислоты встречаются в большинстве нативных белков. Однако D-аминокислоты являются коммерчески доступными и могут заменять некоторые или все аминокислоты, используемые для изготовления тройных сополимеров и других сополимеров, используемых в настоящем изобретении. Настоящее изобретение предполагает применение сополимеров, содержащих как D-, так и L-аминокислоты, а также сополимеров, содержащих преимущественно L- или D-аминокислоты.
В одном варианте осуществления настоящего изобретения сополимер содержит четыре различные аминокислоты, каждая из разных групп от (а) до (d). Предпочтительный сополимер, согласно указанному варианту осуществления настоящего изобретения, включает в себя в комбинации аланин, глутаминовую кислоту, лизин и тирозин, с суммарным положительным электрическим зарядом и молекулярной массой приблизительно 2000-40000 Да, предпочтительно приблизительно 2000-13000 Да, и наиболее предпочтительно сополимер 1 со средней молекулярной массой приблизительно 4700-13000 Да. Предпочтительные пределы молекулярных масс и способы изготовления предпочтительной формы Сор-1 описаны в патенте США № 5800808, который целиком включен в настоящий документ в качестве ссылки. Очевидно, что это является только примером, и что вакцину можно изменять как с точки зрения ее составляющих, так и относительных пропорций составляющих, если удовлетворяются указанные выше общие критерии. Так, сополимер может представлять собой полипептид длиной приблизительно от 15 до 100, предпочтительно приблизительно от 40 до 80 аминокислот, и предпочтительно представляет собой сополимер, имеющий родовое название глатирамер (Glatiramer) ацетат.
В другом варианте осуществления настоящего изобретения сополимер содержит три различные аминокислоты, каждая из разных трех групп от (а) до (d). Указанные сополимеры в настоящем документе упоминаются как тройные сополимеры.
В одном варианте осуществления тройные сополимеры для применения согласно изобретению содержат тирозин, аланин и лизин, и далее обозначаются как YAK. Средняя молярная фракция аминокислот в указанных тройных сополимерах может варьировать. Например, тирозин может присутствовать в молярной фракции приблизительно 0,005-0,250; аланин может присутствовать в молярной фракции приблизительно 0,3-0,6, и лизин может присутствовать в молярной фракции приблизительно 0,1-0,5. Средняя молекулярная масса составляет от 2000 до 40000 Да и предпочтительно приблизительно от 3000 до 35000 Да. В более предпочтительном варианте осуществления настоящего изобретения средняя молекулярная масса составляет приблизительно от 5000 до 25000 Да. Возможна замена лизина на аргинин, аланина на глицин и/или тирозина на триптофан.
В другом варианте осуществления тройные сополимеры для применения согласно изобретению содержат тирозин, глутаминовую кислоту и лизин, и далее обозначаются как YEK. Средняя молярная фракция аминокислот в указанных тройных сополимерах может варьировать: глутаминовая кислота может присутствовать в молярной фракции приблизительно 0,005-0,300, тирозин может присутствовать в молярной фракции приблизительно 0,005-0,250, и лизин может присутствовать в молярной фракции приблизительно 0,3-0,7. Средняя молекулярная масса составляет от 2000 до 40000 Да и предпочтительно приблизительно от 3000 до 35000 Да. В более предпочтительном варианте осуществления настоящего изобретения средняя молекулярная масса составляет приблизительно от 5000 до 25000 Да. Возможна замена глутаминовой кислоты на аспарагиновую кислоту, лизина на аргинин и/или тирозина на триптофан.
В другом варианте осуществления тройные сополимеры для применения согласно изобретению содержат лизин, глутаминовую кислоту и аланин, и далее обозначаются как КЕА. Средняя молярная фракция аминокислот в указанных полипептидах также может варьировать. Например, глутаминовая кислота может присутствовать в молярной фракции приблизительно 0,005-0,300, аланин может присутствовать в молярной фракции приблизительно 0,005-0,600, лизин может присутствовать в молярной фракции приблизительно 0,2-0,7. Средняя молекулярная масса составляет от 2000 до 40000 Да и предпочтительно приблизительно от 3000 до 35000 Да. В более предпочтительном варианте осуществления настоящего изобретения средняя молекулярная масса составляет приблизительно от 5000 до 25000 Да. Возможна замена глутаминовой кислоты на аспарагиновую кислоту, аланина на глицин и/или лизина на аргинин.
В другом варианте осуществления тройные сополимеры для применения согласно изобретению содержат тирозин, глутаминовую кислоту и аланин, и далее обозначаются как YEA. Средняя молярная фракция аминокислот в указанных полипептидах может варьировать. Например, тирозин может присутствовать в молярной фракции приблизительно 0,005-0,250, глутаминовая кислота может присутствовать в молярной фракции приблизительно 0,005-0,300, и аланин может присутствовать в молярной фракции приблизительно 0,005-0,800. Средняя молекулярная масса составляет от 2000 до 40000 Да и предпочтительно приблизительно от 3000 до 35000 Да. В более предпочтительном варианте осуществления настоящего изобретения средняя молекулярная масса составляет приблизительно от 5000 до 25000 Да. Возможна замена тирозина на триптофан, глутаминовой кислоты на аспарагиновую кислоту и/или аланина на глицин.
В более предпочтительном варианте осуществления настоящего изобретения молярная фракция аминокислот в тройных сополимерах является приблизительно такой же, которая предпочтительна для сополимера 1. Молярная фракция аминокислот в сополимере 1 составляет для глутаминовой кислоты приблизительно 0,14, для аланина - приблизительно 0,43, для тирозина - приблизительно 0,10 и для лизина - приблизительно 0,34. Наиболее предпочтительная средняя молекулярная масса для сополимера 1 составляет приблизительно от 5000 до 9000 Да. Активность сополимера 1 в вакцине, описанной в настоящем изобретении, как ожидается, сохранится, если будет произведена одна или более из следующих замен: глутаминовой кислоты на аспарагиновую кислоту, аланина на глицин, лизина на аргинин и тирозина на триптофан.
Молярное соотношение мономеров в более предпочтительном тройном сополимере глутаминовой кислоты, аланина и тирозина, или YEA, составляет приблизительно 0,21:0,65:0,14.
Молярное соотношение мономеров в более предпочтительном тройном сополимере глутаминовой кислоты, аланина и лизина, или KEA, составляет приблизительно 0,15:0,48:0,36.
Молярное соотношение мономеров в более предпочтительном тройном сополимере глутаминовой кислоты, тирозина и лизина, или YEK, составляет приблизительно 0,26:0,16:0,58.
Молярное соотношение мономеров в более предпочтительном тройном сополимере тирозина, аланина и лизина, или YAK, составляет приблизительно 0,10:0,54:0,35.
Тройные сополимеры можно изготавливать любым способом, доступным специалисту. Например, тройные сополимеры можно изготавливать в условиях конденсации, с использованием желательного молярного соотношения аминокислот в растворе, или с помощью твердофазных способов синтеза. Условия конденсации включают должную температуру, рН и растворитель для конденсации карбоксильной группы одной аминокислоты с аминогруппой другой аминокислоты с образованием пептидной связи. Конденсирующие агенты, например, дициклогексилкарбодиимид, можно использовать для облегчения образования пептидной связи. Блокирующие группы можно использовать для защиты функциональных групп, таких как части боковых цепей и некоторые амино- или карбоксильные группы, от нежелательных побочных реакций.
Например, можно использовать способ, описанный в патенте США № 3849650, в котором N-карбоксиангидриды тирозина, аланина, γ-бензилглутамата и N-ε-трифторацетиллизина полимеризуются при температуре окружающей среды в безводном диоксане с диэтиламином в качестве инициирующего агента. γ-карбоксильную группу глутаминовой кислоты можно деблокировать бромидом водорода в ледяной уксусной кислоте. Трифторацетильные группы удаляют из лизина 1 молярным пиперидином. Специалист легко поймет, что процесс можно модифицировать, чтобы получать пептиды и полипептиды, содержащие желательные аминокислоты, т.е. три из четырех аминокислот в сополимере 1 путем избирательного устранения реакций, которые относятся к любой из глутаминовой кислоты, аланина, тирозина или лизина. Для целей настоящей заявки термин «температура окружающей среды» или «комнатная температура» означает температуру приблизительно от 20 до 26°С.
Молекулярную массу тройных сополимеров можно подбирать во время полипептидного синтеза или после изготовления тройных сополимеров. Для подбора молекулярной массы во время полипептидного синтеза условия синтеза или количества аминокислот подбирают таким образом, что синтез останавливается, когда полипептид достигает приблизительной желаемой длины. После синтеза полипептиды с желательной молекулярной массой можно получать с помощью любой доступной процедуры селекции по размеру, такой как хроматография полипептидов на колонке, пропускающей вещества с определенной молекулярной массой, или колонке с гелем, и сбор полипептидов, имеющих молекулярную массу в желательных пределах. Настоящие полипептиды можно также частично гидролизовать для удаления высокомолекулярных видов, например, путем кислотного или ферментативного гидролиза, а затем очищать для удаления кислоты или ферментов.
В одном варианте осуществления настоящего изобретения тройные сополимеры с желаемой молекулярной массой можно получать способом, который включает в себя взаимодействие защищенного полипептида с бромистоводородной кислотой, с получением трифторацетил-полипептида, имеющего желаемый профиль молекулярных масс. Реакцию проводят в течение времени и при температуре, которые определяют заранее путем одной или более пробных реакций. Во время пробной реакции время и температуру изменяют, и определяют пределы молекулярных масс данной партии пробных полипептидов. Условия пробных реакций, которые обеспечивают оптимальные пределы молекулярных масс для данной партии полипептидов, используют для партии. Так, трифторацетил-полипептид, имеющий желаемый профиль молекулярных масс, можно получать способом, который включает в себя взаимодействие защищенного полипептида с бромистоводородной кислотой в течение времени и при температуре, которые определяют заранее путем пробной реакции. Трифторацетил-полипептид с желаемым профилем молекулярных масс затем подвергают воздействию водного раствора пиперидина для получения малотоксичного полипептида, имеющего желаемую молекулярную массу.
В предпочтительном варианте осуществления настоящего изобретения пробный образец защищенного полипептида из данной партии взаимодействует с бромистоводородной кислотой в течение приблизительно 10-50 часов при температуре приблизительно 20-28°С. Оптимальные условия для данной партии определяют путем выполнения нескольких пробных реакций. Например, в одном варианте осуществления настоящего изобретения защищенный полипептид взаимодействует с бромистоводородной кислотой в течение приблизительно 17 часов при температуре приблизительно 26°С.
Поскольку мотивы связывания Сор-1 с MS-связанными молекулами HLA-DR известны (Fridkis-Hareli et al., 1999), полипептиды фиксированной последовательности можно легко изготовить и испытать на предмет связывания со связывающей пептиды бороздкой молекул HLA-DR, как описано в публикации Fridkis-Hareli et al. (1999). Примеры указанных пептидов описаны в WO 005249, содержание которой целиком включено в настоящий документ в качестве ссылки. Тридцать два из пептидов, конкретно описанных в указанной заявке, воспроизведены в таблице 1. Указанные пептиды и другие сходные пептиды, как можно ожидать, будут обладать такой же активностью, что и Сор-1. Указанные пептиды и другие сходные пептиды, как полагают, также относятся к родственным Сор-1 пептидам или полипептидам, а их применение рассматривается как часть настоящего изобретения.
Определение «родственный Сор-1 полипептид», согласно настоящему изобретению подразумевает другие синтетические аминокислотные сополимеры, такие как составленные случайным образом из четырех аминокислот сополимеры, описанные Fridkis-Hareli et al., 2002, как кандидаты для лечения рассеянного склероза, такие как сополимеры (14-, 35- и 50-меры), содержащие аминокислоты фенилаланин, глутаминовую кислоту, аланин и лизин (Poly-FEAK) или тирозин, фенилаланин, аланин и лизин (Poly-YFAK) и любой другой сходный сополимер, который еще предстоит открыть, который можно рассматривать как универсальный антиген, аналогичный Сор-1 и Poly-YE.
Согласно настоящему изобретению предпочтительным сополимером для применения в вакцине согласно изобретению является сополимер 1, обозначаемый в настоящем изобретении так же, как Сор-1, наиболее предпочтительно, в форме своей ацетатной соли, известной под родовым названием глатирамер ацетат. Глатирамер ацетат разрешен в некоторых странах для лечения рассеянного склероза (MS) под торговым наименованием COPAXONE® (товарный знак Teva Pharmaceuticals Ltd., Petah Tikva, Израиль). Несколько клинических испытаний показали, что Сор-1 хорошо переносится и имеет минимальные побочные эффекты, которые в большинстве случаев являются легкими реакциями в месте инъекции (Johnson et al., 1995).
Как было упомянуто выше, мутации в гене SOD1 являются одной генетической причиной семейного ALS (Rosen et al., 1993; Brown, 1995). У нескольких мышиных моделей, которые экспрессировали мутированные гены SOD1, развивалась дегенерация двигательных нейронов, подобная той, которая наблюдается у человека (Gurney et al., 1994; Ripps et al., 1995; Kong and Xu, 1998). Исходное изучение свойств данных мышиных линий показало, что доминантное приобретение неблагоприятного свойства мутированными ферментами вызывает дегенерацию двигательных нейронов (см. обзор Bruijn and Cleveland, 1996). Помимо этого, данные анализы подтвердили многочисленные патологические признаки, которые наблюдаются у человека (Hirano, 1991; Chou, 1992). Понимание данной мутации, названной альтерацией SOD1, привело к созданию экспериментальной модели на животных (мышей с ALS) для испытания лекарственных средств для лечения семейного ALS. Поскольку связанный с SOD1 семейный ALS и спорадический ALS (который составляет 90% всех случаев ALS) имеют сходные симптомы и патологические признаки, трансгенные мыши, несущие мутантный ген SOD1, являются принятой моделью для испытания лекарственных средств для лечения как семейной, так и спорадической формы ALS, и являются моделью, используемой фондом разработки лечения ALS (ALS-TDF). У мышей с ALS развивается моторное заболевание, которое очень напоминает ALS. Двигательная дисфункция в конечном итоге приводит к их гибели.
Согласно настоящему изобретению мыши с ALS, которых иммунизировали вакциной Сор-1, эмульгированного в CFA или в адъюванте, подходящем для применения у человека, как было показано, были защищены от дегенерации двигательных нервов, несмотря на условия окислительного стресса, созданные избыточной экспрессией SOD. Так, вакцинация «универсальным» слабым самореакционным антигеном Сор-1 в CFA продлевала на 52 дня жизнь мышей с ALS (среднее ±SD, 263±8 дней, n=14) по сравнению с нелеченными парными контролями (211±7 дней, n=15, P<0,0001). Вакцинация достоверно улучшала двигательную активность на клинической и доклинической стадии. Кроме того, вакцинация Сор-1 также предупреждала острую дегенерацию двигательных нейронов после аксотомии лицевого нерва: почти на 200% больше двигательных нейронов выживало у вакцинированных мышей, чем у контролей с аксотомией (P<0,05). Указанные результаты предполагают, что концепцию аутоиммунитета как защитного механизма можно распространить на болезни двигательных нейронов. Они также имеют потенциально драматичное клиническое применение.
Адъюванты, использованные для иммунизации согласно изобретению, представляют собой адъюванты на основе алюминия. Более широко используемые в вакцинах, содержащих антигены вирусного происхождения, такие как поверхностный антиген вируса гепатита В или капсулярный полисахарид Haemophilus influenza типа b, указанные адъюванты впервые используются вместе с синтетическими сополимерами, особенно с Сор-1.
Дозу Сор-1 или Poly-YE, которую предстоит ввести, будет определять врач, в соответствии с возрастом пациента и стадией заболевания, и ее можно выбирать из пределов 10-80 мг, хотя любая другая подходящая доза входит в объем настоящего изобретения. Введение можно осуществлять по меньшей мере один раз в месяц или по меньшей мере один раз каждые 2 или 3 месяца, или реже, но любой другой подходящий интервал между иммунизациями рассматривается настоящим изобретением в соответствии с состоянием пациента.
Вакцину согласно изобретению можно вводить любым подходящим способом, включая пероральный, внутримышечный, подкожный и внутрикожный, с адъювантом или без него.
При введении вместе с препаратом Riluzole или любым другим лекарственным средством, подходящим для лечения MND, особенно ALS, дополнительное лекарственное средство вводят в тот же день, что и вакцину, и далее ежедневно, в соответствии в инструкциями производителя, вне связи со схемой вакцинации. Например, суточная доза Riluzole составляет 100 мг.
Следующие примеры иллюстрируют некоторые признаки настоящего изобретения, но не предназначены для ограничения объема настоящего изобретения.
ПРИМЕРЫ
Материалы и методы
Животные. Мыши линии C57BL/6J в возрасте 8-13 недель поступали из центра по разведению животных научно-исследовательского института Weizmann (Rehovot, Израиль). До использования в экспериментах мышам давали наркоз путем интраперитонеального введения 80 мг/кг кетамина и 16 мг/кг ксилазина. Трансгенных мышей, экспрессирующих избыточное количество дефектного человеческого мутантного аллеля SOD1, содержащего ген Gly93→Ala (G93A) (B6SJL-TgN (SOD1-G93A)1Gur (здесь «мыши с ALS»), приобретали в лаборатории The Lackson Laboratory (Bar Harbor, ME, США). Всех животных содержали согласно нормам, сформулированным Institutional Animal Care and Use Committee (IACUC).
Материалы. Сор-1 (медиана ММ: 7200 дальтон) приобретали у Teva Pharmaceuticals Ltd. (Petah Tikva, Израиль). Гель гидроксифосфата алюминия (вакцинный адъювант REHYDRAPHOS™, здесь Alum-phos) приобретали у Reheis (NL, США). Полный адъювант Фрейнда, содержащий 0,5 мг/мл Mycobacterium tuberculosis (CFA) приобретали у Difco (Детройт, Мичиган, США), если не указано иное.
Иммунизация. Мышей иммунизировали Сор-1, эмульгированным в CFA, или Сор-1-Alum-phos (100 мкг в общем объеме 100 мкл). Alum-phos интенсивно смешивали с Сор-1 в соотношении 1:4. Каждую вакцину инъецировали подкожно (п/к) в одну область на боку мышей. Контрольным мышам инъецировали маннит в CFA или в Alum-phos.
Инъекция глутамата. Правый глаз находящихся под наркозом мышей C57B BL/6J, пунктировали иглой калибра 27 в верхней части склеры, и вводили 10-мкл шприц Гамильтона с иглой калибра 30 до стекловидного тела. Мышам инъецировали общий объем 1 мкл (200 нмоль) L-глутамата, растворенного в физиологическом растворе.
Нанесение метки на ганглионарные клетки сетчатки (RGC) мышей. RGC метили за 72 часа до окончания эксперимента. Мышам давали наркоз и помещали их в стереотактическое устройство. Обнажали череп и держали его сухим и чистым. Идентифицировали и отмечали брегму. Обозначенная точка инъекции находилась на глубине 2 мм от поверхности головного мозга, на расстоянии 2,92 мм позади брегмы на передне-задней оси и на расстоянии 0,5 мм латеральнее срединной линии. Просверливали окно в волосистой части головы над обозначенными координатами в правом и левом полушариях. Затем наносили индикатор для нервной ткани FluoroGold (5% раствор в физиологическом растворе; Fluorochrome, Denver, CO) (1 мкл со скоростью 0,5 мкл/мин в каждое полушарие) с помощью шприца Гамильтона, и кожу вокруг раны зашивали. Ретроградный захват индикатора обеспечивал маркер живых клеток.
Оценка выживания RGC у мышей. Мышам вводили смертельную дозу пентобарбитона (170 мг/кг). Производили энуклеацию глаз, отпрепаровывали сетчатку и изготавливали в виде плоских цельных препаратов в параформальдегиде (4% в PBS). Подсчитывали меченые клетки из 4-6 выбранных полей идентичного размера (0,7 мм2). Выбранные поля находились приблизительно на одном и том же расстоянии от диска оптического нерва (0,3 мм), чтобы преодолеть изменчивость плотности RGC в зависимости от расстояния от диска оптического нерва. Поля обсчитывали под флюоресцентным микроскопом (увеличение х800), причем лица, производившие исследование, не были осведомлены о лечении, которое получала мышь. Подсчитывали среднее количество RGC в каждом поле каждой сетчатки.
Модель бокового амиотрофического склероза. Трех мышей с ALS в возрасте 75 дней вакцинировали Сор-1, эмульгированным в Alum-phos (100 мкг Сор-1 в общем объеме 100 мкл, одна подкожная инъекция в область бока). Мышам производили бустерную инъекцию неделей позже, а после этого - ежемесячные инъекции. Еще трех трансгенных мышей не иммунизировали, и они служили контролем спонтанного прогрессирования болезни. Мышечную силу оценивали испытанием вслепую времени удерживания каждой мыши на вращающемся вертикальном стержне. Поскольку максимальное время, в течение которого большинство животных могло висеть на вращающемся стержне, составляло 5 минут, каждый эксперимент продолжали до 5 минут.
Тест на мышечную силу. Тест выполняли как описано ранее (Kong and Xu, 1998). Мышам давали схватиться и держаться на вертикальной проволоке (диаметром 2 мм) с маленькой петлей на нижнем конце. Вертикальная проволока позволяла мышам использовать как передние, так и задние конечности, чтобы хвататься за проволоку. Проволоку поддерживали в вертикально ориентированном круговом движении (радиус круга составлял 10 см) со скоростью 24 об/мин. Время, в течение которого мышь была способна висеть на проволоке, отмечали с помощью таймера. Поскольку большинство мышей падало не позднее 5 мин, испытание прекращали через 5 мин. Мышей обычно подвергали испытанию один раз в неделю, и испытания продолжали до тех пор, пока они больше не могли висеть на проволоке.
Анализ данных. Данные по выживанию анализировали с помощью критерия Mantel-Cox или с помощью регрессионного анализа пропорциональных рисков Cox. Статистическую достоверность определяли с помощью однофакторного ANOVA, с последующей post-hoc процедурой Student-Neuman-Keuls c использованием компьютерной программы SPSS-PC (SPSS, Chicago, IL).
Пример 1. Защита нейронов от токсичности глутамата активной вакцинацией Сор-1, эмульгированным в Alum-phos
Впервые изучали, можно ли токсичность, индуцированную глутаматом, блокировать активной вакцинацией Сор-1, эмульгированным в CFA или в Alum-phos. CFA представляет собой адъювант, не разрешенный для использования у человека, и он часто используется только в экспериментах на лабораторных животных. Alum-phos и другие адъюванты на основе гидроксида алюминия разрешены FDA и другими уполномоченными учреждениями и широко используются в ветеринарных и медицинских вакцинах.
Сор-1, эмульгированный в CFA или в Alum-phos (100 мкг Сор-1 в общем объеме 100 мкл), инъецировали подкожно в одну область бока мышей C57BL/6J, и семью днями позже инъецировали глутамат (200 нмоль) в стекловидное тело мышей. Через семь дней подсчитывали количество выживших RGC. Выживание RGC после токсичности глутамата без предварительной иммунизации принимали за 100%.
Как показано в таблице 2, предварительная иммунизация Сор-1 в CFA или в Alum-phos за семь дней до инъекции глутамата обеспечивала достоверную защиту ганглионарных клеток сетчатки от токсичности глутамата, но защита Сор-1, эмульгированным в Alum-phos, была достоверно выше, чем в случае CFA.
Защита нейронов от токсичности глутамата активной вакцинацией Сор-1, эмульгированным в CFA или в Alum-phos
Пример 2. Защита нейронов от токсичности глутамата вакцинацией Сор-1 или Poly-YE без адъюванта или с адъювантом
Токсичность глутамата представляет собой один из факторов риска нейродегенерации при ALS. Для изучения эффективности иммунизации Сор-1 и Poly-YE без адъюванта для защиты нейронов от токсичности глутамата сетчатку мышей C57BL подвергали воздействию избыточного количества глутамата. Мышей C57BL разделяли на 4 экспериментальные группы:
1. Животные, которых не иммунизировали - отрицательный контроль, n=9.
2. Животные, которых иммунизировали 25 мкг Poly-YE на каждую мышь, n=10.
3. Животные, которых иммунизировали 225 мкг Poly-YE на каждую мышь, n=10.
4. Животные, которых иммунизировали 75 мкг Сор-1 на каждую мышь, n=7.
Экспериментальные группы иммунизировали Poly-YE или Сор-1, растворенными в 100 мкл PBS за 7 дней до интраокулярной инъекции глутамата. Подсчитывали количество RGC, которые выживали через 7 дней после воздействия повышенным уровнем глутамата, и рассчитывали как процентную долю по отношению к нормальным глазам. Результаты представлены на фиг.1. Выживание RGC во всех экспериментальных группах (группы 2-4) было достоверно (p<0,001, t-критерий) выше, чем в группе отрицательного контроля.
В дополнительных экспериментах мышам C57B1 вводили Сор-1 (100 мкг), эмульгированный в Alum-phos (n=8), или только Alum-phos (n=8) или Poly-YE (100 мкг), эмульгированный в CFA (n=24), или только адъювант (отрицательный контроль) (n=27) (100 мкл), за 7 дней до интраокулярной инъекции глутамата. Подсчитывали количество RGC, которые выживали через 7 дней после воздействия повышенным уровнем глутамата. Защиту рассчитывали как процентную долю RGC, которые выживали из всей потери RGC в нелеченной группе. Результаты представлены на фиг.2А-В. Выживание RGC в группе, леченной Сор-1 (фиг.2А), и в группе, леченной Poly-YE (фиг.2В), было достоверно выше, чем в группах отрицательного контроля, которые получали только адъювант.
Сор-1 с высокими молекулярными массами (медиана ММ: 12600, 15500 и 22000 дальтон) испытывали на модели токсичности глутамата. Эффективность в обеспечении специфичного нейропротективного ответа определяли на модели острой токсичности глутамата на RGC, как описано выше. Мышей C57BL/6 (всего 5 групп на эксперимент, по 10 животных в группе) иммунизировали за 14 дней до интраокулярной инъекции глутамата (200 нмоль), и выживание RGC оценивали через 7 дней после инъекции глутамата. Испытывали три дозы Сор-1 каждой ММ и сравнивали с отрицательным контролем (только глутамат) и положительным контролем (75 мкг Сор-1 с ММ 7200 Да, за 7 дней до воздействия токсичности глутамата).
Пример 3. Нейропротективный эффект вакцинации Сор-1 и Poly-YE на модели глаукомы
Глаукома представляет собой хроническое нейродегенеративное заболевание с прогрессирующей утратой нейронов зрительного нерва, которая в конечном итоге приводит к слепоте. Повышенное внутриглазное давление (IOP) считается главным фактором риска и, как полагают, является главной причиной гибели нейронов. Соответственно, биохимические агенты или оперативное вмешательство, призванные снижать IOP, в настоящее время представляют собой обычное лечение. Тем не менее, снижения IOP не всегда достаточно для того, чтобы остановить утрату нейронов. Более того, дегенерация зрительного нерва иногда наблюдается при отсутствии повышенного IOP, и данное состояние называют глаукомой нормального давления (наблюдается приблизительно у одной трети пациентов с глаукомой). Таким образом, нейропротективная терапия считается уместной. Авторы изобретения использовали модель хронического повышения IOP у крыс для изучения способности вакцинации Сор-1 или Poly-YE уменьшать гибель нейронов, которые находятся в условиях стресса, как это может наблюдаться у пациентов с ALS. Поскольку глаукома представляет собой хроническое нейродегенеративное заболевание, как и ALS, нейропротекция, достигнутая на модели глаукомы, может быть показателем сходной нейропротекции при ALS.
Индукцию высокого IOP осуществляли следующим образом: используя щелевую лампу Haag-Streit, испускающую лазерное излучение сине-зеленого спектра, правый глаз находящихся под наркозом взрослых самцов крыс Lewis обрабатывали 80-120 нанесениями, направленными на три из четырех эписклеральные вены и в направлении 270 градусов от сегментарного сплетения. Применяли лазерный луч с мощностью 1 ватт в течение 0,2 секунды, получая пятно размером 100 мм на эписклеральных венах и 50 мм на сегментарном сплетении. Во время второй обработки лазером неделей позже использовали те же параметры, за исключением того, что размеры пятна составляли 100 мм для всех нанесений. Облучение было направлено на все четыре эписклеральные вены и в направлении 360 градусов от сегментарного сплетения 24.
Для измерения повышения IOP крысам интраперитонеально инъецировали 10 мг/мл ацепромазина, седативного лекарственного средства, которое не понижает IOP, и через 5 минут измеряли давление в обоих глазах с помощью тонометра Tono-Pen XL (Automated Ophthalmics, Ellicott City, MD, США) после нанесения локалина на роговицу. Подсчитывали среднее значение от 10 измерений по каждому глазу. Через одну неделю после первой обработки лазером IOP достигало уровней приблизительно 30 мм рт.ст. без каких-либо значимых изменений до конца эксперимента (через 3 недели после первой обработки лазером), как показано в таблице 3.
Для определения выживания RGC через 3 недели после первой обработки лазером наносили гидрофильную краску для нервной ткани декстран тетраметилродамин (Rhodamine Dextran) (Molecular Probes, Oregon, США) непосредственно на интраорбитальную часть зрительного нерва. Только те аксоны, которые пережили высокое IOP и остались функциональными, и у которых клеточные тела все еще живы, могут воспринять краску и продемонстрировать меченые RGC. Крыс умерщвляли через 24 часа и вырезали сетчатки, из которых готовили цельные препараты, и подсчитывали меченые RGC под увеличением х800 цейссовского флюоресцентного микроскопа. На каждой сетчатке обсчитывали по четыре поля, все одного диаметра (0,076 мм2) и находящиеся на одном и том же расстоянии от диска оптического нерва. RGC подсчитывал исследователь, не знавший о принадлежности сетчаток.
В таблице 3 суммировано выживание RGC у крыс с нормальным IOP и у крыс с индуцированным лазером повышением IOP спустя 3 недели.
3а. Влияние вакцинации Poly-YE на выживание RGC на модели глаукомы IOP
Крыс SPD иммунизировали Poly-YE (500 мкг), эмульгированным в CFA, спустя один час после первой обработки лазером (n=9). Одну контрольную группу иммунизировали CFA без антигена (n=7), а второй контрольной группе инъецировали только PBS (n=5). Как показано на фиг.3А, несмотря на то, что IOP оставалось повышенным в течение всего периода эксперимента, у крыс, иммунизированных Poly-YE, но не PBS, наблюдалось достоверное увеличение выживаемости из RGC по сравнению с неиммунизированными крысами. Защиту RGC рассчитывали как процентную долю клеток, выживших в леченных группах, от общего количества потери клеток в неиммунизированной группе.
3b. Влияние вакцинации Сор-1 на выживание RGC на модели глаукомы IOP
При использовании крысиной модели повышения IOP было показано, что Сор-1 уменьшал утрату нейронов, когда его вводили (500 мкг в CFA) в начале повышения IOP или неделю спустя (см. фиг.3В), несмотря на тот факт, что IOP оставалось высоким, и дегенерация нерва уже началась. Помимо этого, вакцинация Сор-1 вместе с введением понижающего IOP лекарственного средства бримонодина обеспечивала более выраженную защиту RGC, чем при использовании только бримонодина (см. фиг.3В, вставка).
Пример 4. Иммунизация Сор-1 защищает от дегенерации двигательных нервов трансгенных мутантных SOD1 мышей (мышей с ALS)
Для того, чтобы установить, может ли иммунизация Сор-1 защищать от прогрессирования дегенерации двигательных нейронов, мышей SOD1 с ALS (n=3) иммунизировали Сор-1 в Alum-phos в возрасте 75 дней, а бустер-дозу вводили неделей позже. Затем их иммунизировали каждые 30 дней. Контрольную группу (n=3) мышей с ALS не иммунизировали Сор-1. Мышей затем несколько раз в неделю испытывали на мышечную силу, слепым тестом на время удерживания на вращающемся вертикальном стержне. Каждый эксперимент продолжался 5 мин.
Развитие мышечной слабости у мышей изображено на фиг.4А-В. На фиг.4А изображено среднее время удерживания каждого животного в неделю (результаты представляют собой среднее ±SEM). Как показано, два иммунизированных Сор-1 животных (мыши 1 и 4) показали более продолжительное время удерживания, чем неиммунизированные мыши.
Начало уменьшения мышечной силы у мышей варьировало индивидуально. Для оценки влияния вакцинации на скорость уменьшения у каждой мыши мышечную силу в любое данное время сравнивали с мышечной силой, установленной за одну неделю до начала уменьшения.
На фиг.4В показан синхронизированный график уменьшения мышечной силы у трансгенных мышей. Очевидно, что мыши, иммунизированные Сор-1 (черные столбцы), показали достоверно более низкую скорость уменьшения мышечной силы, вне зависимости от своей силы на день иммунизации. Таким образом, они сохраняли двигательную силу в течение более длительного периода времени по сравнению с неиммунизированными животными.
Благоприятный эффект иммунизации Сор-1 отражается также на массе тела мышей. Как показано на фиг.5, по мере прогрессирования болезни у трансгенных мышей, иммунизированных Сор-1, также наблюдалась более медленная потеря массы тела. В промежутке между 86 и 111 днями жизни все неиммунизированные трансгенные мыши потеряли 2 грамма массы своего тела. Напротив, в группе, иммунизированной Сор-1, у одной мыши изменений не наблюдалось, а две прибавили 2 грамма массы тела.
Иммунизация Сор-1 также влияла на коэффициент смертности трансгенных мышей. По мере прогрессирования болезни у мышей развивался паралич, и они погибали. Иммунизация Сор-1 достоверно продлевала жизнь трансгенных мышей: в то время как нелеченные мыши умирали через 2, 3 и 4 недели после начала болезни, одна мышь, иммунизированная Сор-1, оставалась живой через 4 недели, а другие две - через 7 недель после начала болезни (таблица 4). К моменту гибели трансгенные мыши, иммунизированные Сор-1, были, в среднем, на 3 недели старше, чем неиммунизированные мыши.
Иммунизация Сор-1 увеличивает продолжительность жизни трансгенных мышей, экспрессирующих избыточное количество мутантной человеческой SOD1
Пример 5. Лечение Сор-1 увеличивает предполагаемую продолжительность жизни мышей с ALS
Четырнадцать мышей с ALS в возрасте 60 дней вакцинировали Сор-1 (75 мкг), эмульгированным в CFA (Difco Laboratories, Heidelberg, Германия), содержащем 5 мг/мл Mycobacterium tuberculosis. Эмульсию (общий объем 200 мкл) инъецировали в подушечку задней лапы, и мышей впоследствии лечили ежедневно пероральным введением Сор-1 (12,5 мг/кг/день) с питьевой водой. Мышей иммунизировали Сор-1 в возрасте 60 дней, а нелеченных контрольных мышей наблюдали ежедневно и взвешивали раз в неделю. Мониторировали их двигательную активность и смертность. Возраст начала симптомов определяли как возраст (в днях) времени первого появления тремора и/или тряски конечностей или свисания (скорее чем выворачивания носков наружу при ходьбе) задних конечностей, когда мышей держали в воздухе за хвост. Утрата установочного рефлекса была принята за показатель конечной стадии болезни. Паралич вызывается прогрессирующей утратой двигательных нейронов в спинном мозге. Как показано на фиг.6, невакцинированные контроли (n=14) были парализованы в одной или более конечностей и погибали в возрасте 211±7 дней (среднее ±SD). Мыши, леченные Сор-1, выживали в течение 263±8 дней. Таким образом, вакцинация Сор-1 драматически увеличивала предполагаемую продолжительность жизни мышей с ALS (фиг.6).
В качестве положительного контроля 15 мышам с ALS давали ежедневную дозу (30 мг/кг) препарата Riluzole, единственного лекарственного средства, которое назначают в настоящее время пациентам с ALS. Как показано на фиг.7, выживаемость мышей, леченных Riluzole, была выше на 9% по сравнению с контролем, в то время как выживаемость мышей, леченных Сор-1, была выше на 25% по сравнению с контролем.
Помимо увеличения почти на 25% продолжительности жизни, было отсрочено начало болезни (проявляющееся двигательными нарушениями), что указывает на то, что преимущество выражалось также в качестве жизни как на доклинической, так и на клинической стадиях (фиг.8). Мышам давали схватиться и держаться на вертикальной проволоке (диаметром 2 мм) с маленькой петлей на нижнем конце. Нормальные значения для каждой мыши получали путем оценки ночной двигательной активности (от 8 часов вечера до 8 часов утра) в возрасте 40 и 60 дней, с использованием аппарата с вращающимся стержнем (LMTB, Берлин). Их активность отмечали индивидуально с помощью компьютеризованной системы и оценивали ежедневно. Для статистической оценки активность на вращающемся стержне стандартизировали по средней активности каждой мыши от 40 дня до 60 дня. Данные выражали как среднее ± стандартная ошибка среднего (SEM). Результаты испытаний на вращающемся стержне и массу тела сравнивали с помощью дисперсионного анализа (ANOVA). Статистическую достоверность оценивали с помощью однофакторного ANOVA, с последующей post-hoc процедурой Student-Neuman-Keuls c использованием компьютерной программы SPSS-PC (SPSS, Chicago, IL). Наблюдалась достоверная разница между леченными Сор-1 и нелеченными мышами в следующие периоды времени: между 12 и 20 днями (p<0,058), между 21 и 24 днями (p<0,0079) и между 25 и 28 днями (p<0,0017).
Пример 6. Лечение мышей с ALS Сор-1 без адъюванта
Мышей с ALS (15 животных на группу) разделяли на 11 экспериментальных групп:
1. Нелеченные мыши - группа отрицательного контроля.
2. Мыши, леченные препаратом Riluzole, - 30 мг/кг/день.
3. Мыши, иммунизированные Сор-1/CFA, - первичная вакцинация 75 мкг с последующим ежедневным пероральным введением Сор-1 (12,5 мг/кг) - группа положительного контроля.
4. Мыши, иммунизированные двумя инъекциями 75 мкг Сор-1: первый раз на 45 день и второй раз на 59 день.
5. Мыши, иммунизированные как в группе № 4, с последующей однократной инъекцией 100 мкг Сор-1 на 87 день.
6. Мыши, иммунизированные двумя инъекциями 150 мкг Сор-1: первый раз на 45 день и второй раз на 59 день.
7. Мыши, иммунизированные двумя инъекциями 75 мкг Сор-1: первый раз на 83 день и второй раз на 97 день.
8. То же, что и в группе № 4, с 30 мг/кг/день Riluzole.
9. То же, что и в группе № 5, с 30 мг/кг/день Riluzole.
10. То же, что и в группе № 6, с 30 мг/кг/день Riluzole.
11. То же, что и в группе № 7, с 30 мг/кг/день Riluzole.
Мониторировали двигательную активность и массу тела мышей один раз в неделю, начиная за две недели до начала лечения. Критерий конечной стадии для умерщвления животных определялся как неспособность подняться в течение 30 секунд после того, как их положили на бок на плоскую поверхность. Решение принималось независимым ветеринарным врачом, как это требуется протоколом по обращению с экспериментальными животными.
Пример 7. Введение Сор-1 защищает от дегенерации двигательных нейронов после аксотомии лицевого нерва
Пересечение лицевого нерва у взрослой мыши, как известно, вызывает легко обнаруживаемую позднюю дегенерацию от 20% до 35% аксотомизированных двигательных нейронов. Следовательно, аксотомия лицевого нерва обеспечивает модель ALS, который представляет собой заболевание, характеризующееся прогрессирующей утратой мотонейронов. Влияние иммунизации на выживание и функцию нейронов на модели с аксотомией лицевого нерва является показательным в смысле возможности лечения для уменьшения утраты нейронов у пациентов с ALS.
В данном эксперименте использовали тридцать четыре взрослых самок мышей (возраст 12 недель, 20-25 г) линии C57BL/6JOlaHsd (Harlan Winkelmann, Borchen, Германия). Контрольным животным производили одностороннюю аксотомию лицевого нерва и не проводили лечение, или инъецировали им PBS, эмульгированный в CFA. Мышей в экспериментальной группе (n=10) иммунизировали Сор-1 (всего 100 мкг) или инъецировали им PBS (n=9), оба эмульгированные в CFA, и 7 днями позже производили аксотомию лицевого нерва. Мышей в третьей группе (n=8) подвергали аксотомии без предварительной иммунизации, а мышей в четвертой группе (n=7) оставляли интактными.
Через семь дней мышам под наркозом (100 мг Ketanest® плюс 5 мг Rompun® на кг массы тела) накладывали фациально-фациальный анастомоз (FFA) путем микрохирургического повторного соединения проксимальной культи и дистальной культи двумя 11-0 эпиневральными швами (Ethicon EH 7438G, Norderstedt, Германия). Рану закрывали тремя 4-0 кожными швами. Для оценки восстановления двигательные нейроны лицевого нерва, иннервирующие мышцы подушечек вибрисс, ретроградно метили путем инъецирования 30 мкл 1% водного раствора флюоресцентного ретроградного индикатора FluoroGold плюс 2% диметилсульфоксид (ДМСО) инъецировали в мышцы каждой подушечки вибрисс. Семью днями позже мышам повторно давали наркоз и производили транскардиальную перфузию 0,9% NaCl, с последующей фиксацией 4% параформальдегидом в 0,1 М фосфатном буфере рН 7,4 в течение 20 мин. Головной мозг удаляли, и изготавливали срезы относительно венечного шва толщиной 50 мкм через ствол головного мозга с помощью вибратома. Срезы изучали под эпифлюоресцентным микроскопом Zeiss Axioskop 50 через изготовленное на заказ устройство HQ-Schmalband-filter для FluoroGold (AHF Analysentechnik, Tubingen, Германия).
Через восемь недель после аксотомии, как показано на фиг.9A-D и в таблице 5, среднее количество меченных FluoroGold двигательных нейронов у мышей, вакцинированных Сор-1, было достоверно больше количества, полученного в группе, которой инъецировали PBS в CFA, или в нелеченной контрольной группе (p<0,05). Лечение Сор-1 не оказывало влияния на количество двигательных нейронов в ядре неповрежденного лицевого нерва. Контрольная иммунизация PBS в CFA не оказывала защитного действия.
Ретроградное нанесение метки на нейроны после инъекции FluoroGold в подушечки вибрисс не выявило разницы в локализации или количестве двигательных нейронов в ядре интактного лицевого нерва у мышей, иммунизированных Сор-1 в CFA (фиг.9А), и у мышей, которым инъецировали PBS в CFA (фиг.9С). Напротив, ядро поврежденного лицевого нерва после предварительного лечения мышей Сор-1 в CFA (фиг.9В) содержало достоверно больше меченых двигательных нейронов, чем ядро поврежденного лицевого нерва у контрольных животных, предварительно получавших PBS в CFA (фиг.9D). Данные представлены как средние ± стандартное отклонение (SD). Разницу между различными экспериментальными группами выявляли с помощью однофакторного дисперсионного анализа (ANOVA) и post-hoc t-критерия для непарных данных с коррекцией Bonferroni-Holm. Величины р менее 0,05 считались статистически достоверными.
Пример 8. Введение Сор-1 сохраняет активность двигательных нейронов после острой аксотомии
Для того чтобы определить было ли большее количество двигательных нейронов, обнаруженных у мышей с аксотомией, леченных Сор-1, чем у контрольных животных, связано с функциональным улучшением, был произведен биометрический анализ поведения, включающего активность вибрисс. Основные параметры поведения, включающего активность вибрисс, были документированы у интактных контрольных мышей. В нормальных физиологических условиях вибриссы («усы») распрямляются с передней ориентацией. Их одновременные взмахи, известные как «ощупывание» или «принюхивание», наблюдаются 5-11 раз в секунду. Главными движениями указанной двигательной активности являются вытягивание и втяжение волосков вибрисс, производимое пилоэректорными мышцами, которые иннервируются щечной ветвью лицевого нерва. Когда лицевой нерв пересечен, вибриссы приобретают каудальную ориентацию и остаются неподвижными.
Используя данную модель, оценивали следующие параметры: (i) вытягивание (движение вибрисс вперед), измеряемое рострально открытым углом между срединно-сагиттальной плоскостью и стержнем волоса (большие вытягивания представлены малыми углами); (ii) частоту движения вибрисс, представленную циклами вытягивания и втяжения (пассивного движения в заднем направлении) за секунду; (iii) амплитуду - разницу, в градусах, между максимальным втягиванием и максимальным вытягиванием; (iv) угловую скорость во время вытягивания, в градусах за секунду; и (v) угловое ускорение во время вытягивания, в градусах за секунду.
У мышей с аксотомией лицевого нерва, вакцинированных Сор-1, наблюдалась более высокая активность вибрисс, чем в других группах мышей. Лучше всего на это указывала амплитуда, угловая скорость во время вытягивания и угловое ускорение во время вытягивания (таблица 6).
Влияние вакцинации Сор-1 на выживание двигательных нейронов
Численные значения результатов показаны на фиг.9. Представлены количества (средние ±SD) перикарионов, ретроградно меченных инъекцией 1% FluoroGold (30 мкл), в группе интактных мышей (группа А) и у мышей, подвергавшихся только FFA (группа В), FFA после инъекции PBS в CFA (группа С) и FFA после вакцинации Сор-1 в CFA (группа D). Надписанные буквы указывают группы с достоверно различающимися величинами (*p<0,05). Для визуального анализа использовали систему с видеокамерой CCD (Optronics Engineering Model DEI-470, Goleta, CA), комбинированную с компьютерной программой для анализа изображения Optimas 6.5 (Optimas, Bothell, WA), для мануального подсчета ретроградно меченных двигательных нейронов лицевого нерва на экране компьютера (42). Используя принцип ректификационной колонны (43), все ретроградно меченные двигательные нейроны с видимым ядром клетки подсчитывали в каждом втором срезе 50-мкм срезов ядра лицевого нерва как на оперированной, так и на неоперированной стороне. Подсчет производили двое исследователей, которые не были осведомлены о лечении, полученном крысами.
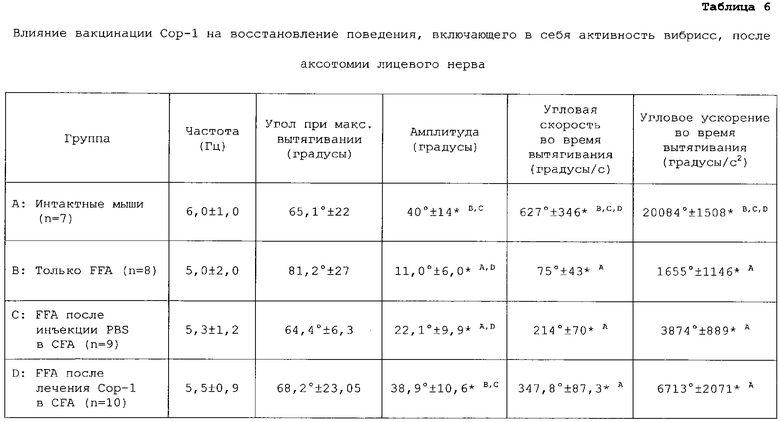
Биометрические данные нормального и восстанавливающегося поведения, включающего в себя активность вибрисс, у интактных мышей (группа А) и у мышей, подвергавшихся только FFA (группа В), мышей, подвергавшихся FFA после инъекции PBS в CFA (группа С) и мышей, подвергавшихся FFA после инъекции Сор-1 в CFA (группа D). Величины представляют собой средние ±SD. Надписанные буквы указывают на группы с достоверно различающимися величинами (*p<0,05). Для биометрического анализа использовали два больших волоска ряда С на каждой стороне морды животного. У мышей под легким эфирным наркозом все остальные вибриссы подстригали маленькими тонкими ножницами. Для видеозаписи активного исследовательского поведения мышей в течение 3-5 мин использовали цифровую видеокамеру (Panasonic NV DX-110 EG). После калибровки отбирали образцы видеоизображения поведения, включающего в себя активность вибрисс, при 50 Гц (50 полей в сек) при открытом состоянии затвора видеокамеры 4 мсек. Изображения записывали на миникассеты AY-DVM 60 EK. Видеопоследовательности медленно просматривали и отбирали фрагменты 1,5-сек продолжительности для каждой мыши, чтобы произвести сбор биометрических данных, касающихся поведения, включающего в себя активность вибрисс. Использовавшиеся критерии отбора включали стабильное положение головы, частоту движений вибрисс и угол вытягивания вибрисс. Выбранные последовательности улавливались 2D/ручной усовершенствованной видеосистемой PEAK Motus 2000 (PEAK Performance Technologies, Englewood, CO). Пространственная модель состояла из трех стандартных точек (кончик носа и внутренние углы обоих глаз). Каждая вибрисса была представлена в пространственной модели двумя точками: своим основанием и точкой на стрежне в 0,5 см от основания.
Ссылки
Brown R.H. (1995) "Amyotrophic lateral sclerosis: recent insights from genetics and transgenic mice" Cell 80: 687-692.
Bruijn L.I., Cleveland D. W. (1996) "Mechanisms of selective motor neuron death in ALS: insights from transgenic mouse models of motor neuron disease" Neuropathol Appl Neurobiol 22: 373-387.
Chou S.M. (1992) "Pathology-light microscopy of amyotropic lateral sclerosis". In: Handbook of Amyotrophic Lateral Sclerosis (Smith R.A., ed.), pp. 133-181. New York: Marcel Dekker.
Fridkis-Hareli et al. (1999) "Binding motifs of copolymer 1 to multiple sclerosis- and rheumatoid arthritis-associated HLA-DR molecules" J Immunol. 162 (8): 4697-4704.
Fridkis-Hareli et al. (2002) "Novel synthetic amino acid copolymers that inhibit autoantigen-specific T cell responses and suppress experimental autoimmune encephalomyelitis" J Clin Invest 109 (12):1635-1643.
Gurney M.E. et al. (1994) "Motor neuron degeneration in mice that express a human Cu1Zn superoxide dismutase" Science 264: 1772-1775.
Hadano et al. (2001) "A gene encoding a putative GTPase regulator is mutated in familial amyotrophic lateral sclerosis 2" Nature Genetics 29: 166-73.
Hauben et al. (2000) "Autoimmune T cells as potential neuroprotective therapy for spinal cord injury" Lancet 355: 286-287.
Hirano A. (1991) "Cytopathology of amyotrophic lateral sclerosis" Adv Neurol 56: 91-101.
Johnson et al. (1995) "Copolymer 1 reduces relapse rate and improves disability in relapsing-remitting multiple sclerosis: results of a phase III multicenter, double-blind placebo-controlled trial. The Copolymer 1 Multiple Sclerosis Study Group," Neurology 1: 65.
Julien J.P. (2001) "Amyotrophic lateral sclerosis: unfolding the toxicity of the misfolded". Cell 104: 581-591.
Kipnis et al. (2001) "Neuronal survival after CNS insult is determined by a genetically encoded autoimmune response". J Neurosci 21(13): 4564-71.
Kong, J. and Xu, Z. (1998) "Massive mitochondrial degeneration in motor neurons triggers the onset of amyotrophic lateral sclerosis in mice expressing a mutant SOD1" J. Neuroscience 18: 3241-3250.
Moalem G. et al. (1999) "Autoimmune T cells protect neurons from secondary degeneration after central nervous system axotomy", Nature Medicine 5: 49-55.
Mulder D.W. et al. (1986) "Familial adult motor neuron disease: amyotrophic lateral sclerosis" Neurology 36: 511-517.
Munsat T.L. et al. (1989) "Adult motor neuron disease". In: Merritt's Textbook of Neurology (Rowland L.P., ed.), pp. 682-687. Philadelphia: Lea & Febiger.
Pitt et al. (2000) "Glutamate excitotoxicity in a model of multiple sclerosis" Nature Medicine 6: 67-70.
Ripps M.E. et al. (1995) "Transgenic mice expressing an altered murine superoxide dismutase gene provide an animal model of amyotrophic lateral sclerosis" Proc Natl Acad Sci, USA 92: 689-693.
Rosen D.R. et al. (1993) "Mutations in Cu1Zn superoxide dismutaso gene are associated with familial amyotrophic lateral sclerosis" Nature 362: 59-62.
Rothstein J.D., et al. (1992) "Decreased glutamate transporter by the brain and spinal cord in amyotrophic lateral sclerosis". N. Eng. J. Med; 326: 1464-68.
Turner et al. (2001) "Clinical trials in ALS: An overview. Seminars in Neurology" 21: 167-175.
Yang et al. (2001) "The gene encoding alsin, a protein with three guanine-nucleotide exchange factor domains, is mutated in a form of recessive amyotrophic lateral sclerosis", Nature Genetics 29: 160-165.










Изобретение относится к медицине, неврологии и касается способа уменьшения прогрессирования болезни и/или защиты двигательных нейронов от дегенерации и/или защиты от токсичности глутамата у пациента, страдающего болезнью двигательных нейронов (MND). Способ включает иммунизацию указанного пациента вакциной, содержащей сополимер на основе четырех аминокислот тирозин-глутамат-аланин-лизин Сор-1 в качестве активного агента в эффективной для иммунизации дозе. Использование данного сополимера обеспечивает наиболее высокую активность в отношении восстановления двигательных нейронов у пациента. 9 з.п. ф-лы, 9 ил., 6 табл.
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| RU 99106243 A, 10.04.2001, формула: пп.1-3, 36-44, 72-81 | |||
| ТЕТРАПЕПТИД, СТИМУЛИРУЮЩИЙ ФУНКЦИОНАЛЬНУЮ АКТИВНОСТЬ НЕЙРОНОВ, ФАРМАКОЛОГИЧЕСКОЕ СРЕДСТВО НА ЕГО ОСНОВЕ И СПОСОБ ЕГО ПРИМЕНЕНИЯ | 1999 |
|
RU2155063C1 |
| Прибор, замыкающий сигнальную цепь при повышении температуры | 1918 |
|
SU99A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| / Под ред | |||
| Н.Н.Яхно и др | |||
| - М.: Медицина, 1995, т.2, с.218-227 | |||
| Schori H | |||
| et al | |||
| Vaccination for protection of | |||

