Изобретение относится к иммунологии, вирусологии, биотехнологии и направлено на повышение специфической активности иммунобиологических лекарственных препаратов (ИЛП), может быть использовано для специфической иммунопрофилактики управляемых инфекций различной этиологии.
Изобретение реализуется путем использования деривата фталгидразида, представляющего собой средство с иммунотропными и противовоспалительными свойствами в качестве адъюванта. Противовоспалительные средства относятся к лекарственным средствам, регулирующим функционально-метаболическую активность макрофагов и нейтрофильных гранулоцитов, обладает выраженным противовоспалительным эффектом.
Формула адьюванта Абидова - дериват синтетических производных фталгидразида; другие дериваты фталгидразида, имеющие схожую формулу, полученные синтетическим или другими способами в виде дигидрата, моногидрата, ангидрата и с другими любыми солями (фиг. 1).
Адьювант Абидова - химическая формула

Основные механизмы иммунотропного действия:
система неспецифической иммунологической резистентности - регулирует активность макрофагов, регулирует продукцию активных форм кислорода и других острофазовых белков, ответственных за развитие токсического синдрома; нормализует функциональное состояние макрофагов, приводит к восстановлению их антиген-представляющей и регулирующей функции, способствуя тем самым усилению не менее чем в 3-4 раза процессов специфического антителообразования; усиливает функциональную антибактериальную активность нейтрофильных гранулоцитов, фагоцитоз завершенный и неспецифическую иммунологическую резистентность организма;
система цитокинов - снижает избыточный синтез ФНО, интерлейкина (ИЛ)-1; усиливает взаимодействие между Т- и В-клетками и, тем самым специфический иммунный ответ, повышает антигенпрезентирующую активность клеток.
Инфекционные заболевания различной природы продолжают оставаться заметной проблемой современной инфектологии и медицины. В этом аспекте немаловажное значение придается опасным и особо опасным инфекциям (ОООИ), к числу которых относят чуму, сибирскую язву, туляремию, вирусные геморрагические лихорадки, энцефалиты и др. [1, 2]. Согласно данным доступных публикаций, вспышки этих инфекций регистрируют, прежде всего, в развивающихся странах с нестабильными социально-экономическими условиями [3-6]. Для развитых стран, включая Россию, данные инфекции также не потеряли на сегодняшний день своей актуальности, о чем свидетельствуют:
- периодическая активизация на территории страны природных очагов сибирской язвы, геморрагической лихорадки с почечным синдромом, Конго-крымской геморрагической лихорадки, клещевой энцефалит и др.;
- периодические вспышки лихорадки Западного Нила, птичьего гриппа, Калифорнийского энцефалита, Японского энцефалита, лихорадки Сахалин, лихорадки Иссык-Куль и других опасных инфекций. [7].
Анализ современного состояния санитарно-эпидемиологической обстановки показывает, что в некоторых сопредельных РФ странах сохраняется эпидемиологически неблагополучная ситуация по целому ряду бактериальных и вирусных инфекций. Помимо этого, в настоящее время не исключается несанкционированное применение возбудителей инфекций, в том числе ОООИ, для осуществления биотеррористических атак. По мнению отечественных и зарубежных специалистов приоритет в подобных условиях и для осуществления подобных целей будет отдаваться нетрадиционным в генетическом отношении возбудителям инфекций, против которых существующие средства специфической иммунопрофилактики могут оказаться недостаточно эффективными [7].
В связи с изложенным, актуальными представляются исследования по поиску эффективных средств защиты от инфекций, особенно тех, которые внезапно появляются в человеческой популяции и сразу же вызывают панику, хаос, социально-экономические катаклизмы и т.д. К числу последних можно отнести зоонозные трансмиссивные вирусы птичьего гриппа, вирусы свиного гриппа, коронавирусы, вызывающие SARS, MERS, SARS-CoV-2, COVID-19 и др., способные в кратчайшие сроки инфицировать большие группы населения и вызвать эпидемии. Основная опасность этих биоагентов не в массовости вызываемых ими поражений, а в отсутствии адекватных мер противодействия вследствие их внезапности появления. Более того, имеющиеся средства, в частности, антибактериальные и противовирусные препараты не всегда оказываются эффективными.
Общеизвестно, что устойчивость организма к патогенам - это способность организма переносить воздействие патогенов и формировать иммунный ответ к ним. Жизнестойкий организм способен вернуться к состоянию здоровья после ответа на инфекцию [8-10]. Большинство доступных в настоящее время средств лечения инфекционных заболеваний являются противомикробными препаратами и, таким образом, нацелены на сам возбудитель. Учитывая ущерб, который могут нанести патогены организму, повышенное внимание к быстрому их клиренсу представляется наиболее приоритетной задачей. Вместе с тем, при заражении различными патогенами, не менее важным медицинским вмешательством является повышение способности макроорганизма адекватно переносить прямое и/или косвенное воздействия патогена [11-14]. В определенном аспекте это возможно в случае предварительного заблаговременного проведения цикла специфической иммунопрофилактики с использованием ИЛП.
ИЛИ относят к наиболее действенным и широко применяемым средствам противоинфекционной защиты. Они представляют собой специфический в отношении конкретной инфекции биоагента антигенный раздражитель для макроорганизма, направленный исключительно на формирование его иммунной защиты против аналогичного, но вирулентного возбудителя биоагента. Базовым критерием, разрешающим применение ИЛП в клинической практике является безвредность: ИЛП не должны обладать побочными эффектами и вызывать у привитых развитие и прогрессирование соответствующего инфекционного заболевания. Вместе с тем, нельзя не отметить, что, основным критерием, характеризующим ИЛП в плане его практического применения, является эффективность. Упомянутый критерий свидетельствует, что ИЛП должен обеспечивать выработку напряженного иммунитета к конкретной инфекции и его сохранение в течение длительного (не менее 12 мес.) периода времени после вакцинации [15-19].
Состав современных ИЛП представляет собой собственно специфический антиген (иммуноген) и вспомогательные вещества, не оказывающие отрицательного действия как на сам иммуноген, так и на прививаемый организм. В состав отдельных ИЛП могут входить антиген - активное начало, вещества, активирующие действие антигена, - адъюванты, а также стабилизаторы и консерванты. Очевидно, что эффективность ИЛП определяется, прежде всего, свойствами входящего в него иммуногена. Имеющиеся ИЛП существенным образом различаются по природе используемого иммуногена [16, 20]. Наиболее широко используемыми в инфектологии ИЛП являются препараты на основе живых аттенуированных штаммов соответствующих микроорганизмов (например, вакцины против сибирской язвы, чумы, туляремии, желтой лихорадки и др.) и препараты на основе инактивированных убитых штаммов микроорганизмов, а также химических структур отдельных микроорганизмов, рекомбинантные структуры, ДНК-вакцины и др.
ИЛП на основе аттенуированных штаммов микроорганизмов характеризуются наличием в их составе цельных штаммов микроорганизмов, не утративших антигенные свойства, но лишенных вирулентности. Как правило, эти ИЛП высокоэффективны, но требуют постоянного контроля вследствие реверсии вакцинного штамма и приобретения им вирулентных свойств. Аттенуированные вакцины могут быть использованы не только для иммунизации против конкретного возбудителя, но и против гетерологичной инфекции вследствие определенной общности между возбудителями (например, вакцины против чумы могут быть применены для специфической иммунопрофилактики туляремии в виду наличия у возбудителей общности антигенной структуры).
ИЛП, сконструированные на основе убитых штаммов возбудителей, их антигенных детерминант, генетических последовательностей являются в плане безопасности более предпочтительными в сравнении с живыми аттенуированными вакцинами, то есть менее реактогенны. Однако по эффективности упомянутые препараты им уступают. Более того, длительность формируемого при их применении специфического иммунитета примерно в 1,5-2 раза короче, чем при иммунизации живыми вакцинами. В настоящее время практически все разрешенные к применению на территории РФ ИЛП включены в Национальный календарь профилактических прививок.
Благодаря расшифровке генетической последовательности многих возбудителей в настоящее время можно получать ИЛП известной специфичности и структуры [21, 22]. Изменяя генотип возбудителей путем делеций или точечных мутаций генов вирулентности, направленно конструируют аттенуированные (ослабленные) штаммы высокой эффективности и безопасные для человека. В частности, таким образом разработаны и продолжают разрабатываться вакцины против гриппа [20]. Клонирование генов протективных антигенов различных возбудителей открыло возможности получения рекомбинантных продуцентов ИЛП. К числу таковых отнесены рекомбинантные штаммы вирусов, бактерий, дрожжей, обеспечивающих экспрессию указанных генов и соответственно синтез антигенов известной структуры. Наиболее удобными для использования в качестве вектора оказались вирус осповакцины, аденовирусы, вирус везискулярного стоматита и др. С их помощью, вернее на их платформе разработаны вакцины против гепатита В (Ревакс-В), геморрагической лихорадки Эбола и др. инфекций. Кроме того, при культивировании рекомбинантных штаммов-продуцентов можно получать достаточные количества определенных протективных антигенов, на основе которых в дальнейшем конструировать молекулярные вакцины.
В некоторых случаях из-за высокой токсичности антигена для иммунизации используют идиотипические вакцины, содержащие в качестве активного начала антиидиотипические антитела, воспроизводящие конфигурацию антигена. На этом принципе сконструирована эффективная вакцина против вируса гепатита В.
Другой путь создания специфических вакцин известной структуры основан на использовании химически синтезированных пептидов, соответствующих определенным эпитопам антигена. Обычно эти пептиды неиммуногенны, поэтому их коньюгируют с носителями в виде белка или высокомолекулярного полимера. Синтетические вакцины такого типа созданы против сальмонеллеза и гриппа. Возникновение и развитие генотерапии открыло качественно новый путь иммунопрофилактики, основанный на создании ДНК-вакцин. Методами генной инженерии можно получать любые генетические конструкции, включающие ген протективного антигена. Для переноса и экспрессии цитокиновых генов в качестве вектора используется плазмиды, а также ряд вирусов. При внутримышечном введении вектора в миоциты происходит экспрессия указанного гена, что обеспечивает развитие специфического иммунного ответа. Безопасность метода и возможность целенаправленной иммунопрофилактики открывает большие перспективы применения ДНК-вакцин [23]. Необходимо также упомянуть о так называемых «терапевтических вакцинах». Это понятие используют для обозначения бактериальных вакцин и анатоксинов, которые индуцируют не только протективный иммунитет, но и вызывают выраженную иммуномодуляцию [24, 25]. Вакцинотерапия нашла широкое применение при лечении онкологических больных, устранения рецидивов хронических заболеваний, а также при купировании проявлений аллергических болезней.
Сила иммунного ответа зависит от двух основных факторов: свойств макроорганизма и особенностей антигенов, используемых для иммунизации. Иммуногенность антигенов, получаемых из возбудителей инфекционных болезней, неодинакова. Наиболее иммуногенны экзотоксины и поверхностные антигены микроорганизмов. Иммуногенность вакцины во многом зависит от того, насколько удачно выбраны антигены для конструирования препарата. При недостаточной его иммуногенности используют неспецифические иммуностимуляторы (адъюванты). В практике вакцинации в качестве иммуностимуляторов используют гидроокись алюминия, фосфат алюминия, фосфат кальция, полиоксидоний и белковые носители. В качестве примера можно привести ИЛП «Гриппол», в составе которого находится в качестве адъюванта полиоксидоний. Трудности в создании высокоэффективных вакцин связаны с особенностями макроорганизма, его генотипа, фенотипа, с существованием двух видов иммунитета (гуморального и клеточного), которые регулируются разными субпопуляциями клеток-хелперов (Tx1 и Тх2). Поствакцинальный иммунитет складывается из двух видов иммунных реакций: гуморального и клеточного. Отсутствие циркулирующих антител еще не является доказательством слабости иммунитета, при новой встрече с антигеном иммунный ответ развивается за счет иммунологической памяти. Кроме того, в основе резистентности к некоторым видам инфекций лежат клеточные механизмы, поэтому вакцины, используемые для профилактики этих инфекций, должны формировать клеточный иммунитет.
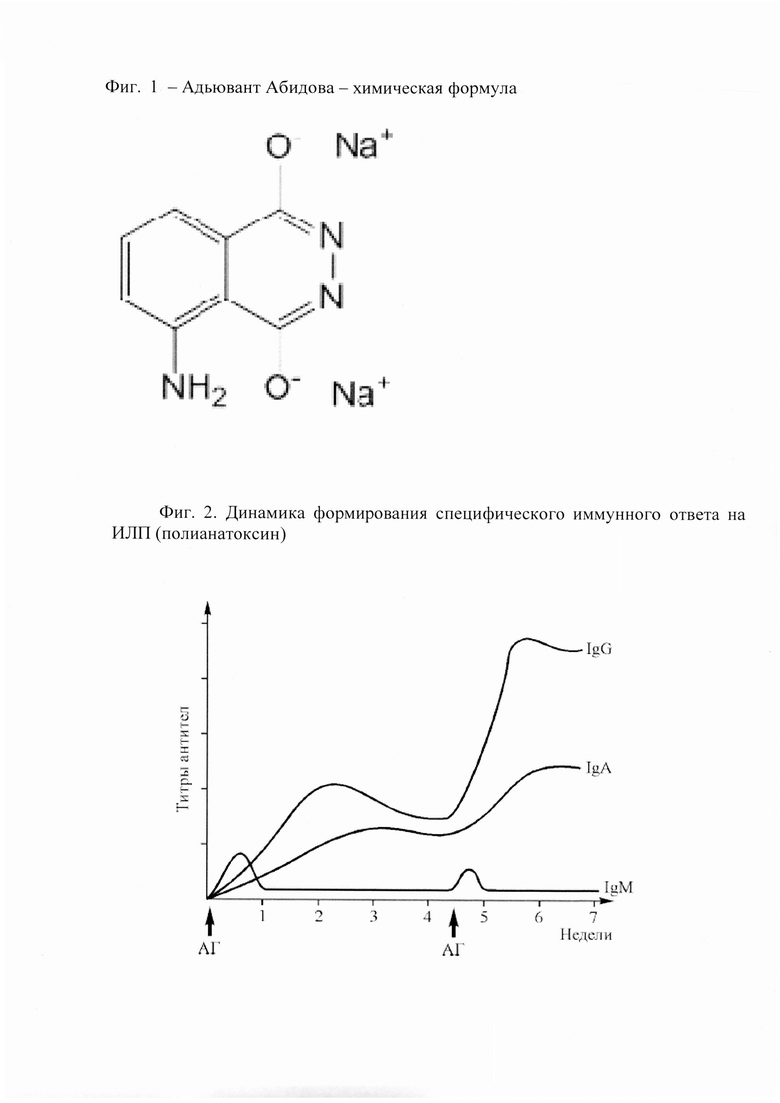
Иммуногенность ИЛП составляет основу ее эффективности [17, 18]. Как правило, корпускулярность ИЛП (живых, убитых) обеспечивает необходимую иммуногенность, в остальных случаях часто приходится использовать дополнительные методы повышения иммуногенности ИЛП. К основным способам повышения иммуногенности ИЛП относят: использование оптимальной концентрации антигена; очистка вакцин от низкомолекулярных веществ, способных вызывать специфическую или неспецифическую супрессию иммунного ответа; агрегация антигена с помощью ковалентного связывания и других методов комплексообразования; включение в вакцину максимального количества эпитопов антигена; сорбция на веществах, создающих депо антигена (гидроокись алюминия, фосфат кальция и др.); использование липосом (водно-масляной эмульсии); добавление микробных, растительных, синтетических и других видов адъювантов; связывание слабого антигена с белковым носителем (столбнячным, дифтерийным анатоксином и др.); включение антигена в микрокапсулы, обеспечивающие выброс антигена через заданный промежуток времени; улучшение условий процессинга и представления антигена. Использование антигенов гистосовместимости I и II классов или антител к этим антигенам. Подходы к созданию ИЛП, обеспечивающих формирование клеточного и гуморального иммунитета, различны. Это обусловлено участием в иммунном ответе двух регуляторных клеток: Tx1 и Тх2. Между ними существует определенная степень антагонизма, хотя они и образуются из одного и того же вида клеток-предшественников. Получить ИЛП, который бы вызывала клеточный иммунитет, достаточно трудно. Во многих случаях не удается переключить иммунный ответ Тх2 на вакцину, которая стимулирует образование антител, на клеточный ответ Tx1. Крайне важно, чтобы ИЛП вызывал Т-зависимый иммунный ответ. В противном случае ответ будет кратковременным, а повторное введение препарата не будет вызывать вторичный ответ. Первичный и вторичный иммунный ответ отличаются друг от друга по динамике формирования иммунитета (фиг. 2). Вторичный иммунный ответ недостаточно выражен, если для иммунизации используется слабоиммуногенный антиген, если в организме присутствуют пассивно введенные или активно приобретенные антитела, если антиген вводят пациенту с иммунодефицитом. Вторичный иммунный ответ характеризуется следующими признаками: более раннее (по сравнению с первичным ответом) развитие иммунных реакций; уменьшение дозы антигена, необходимой для достижения оптимального ответа; увеличение силы и продолжительности иммунного ответа; усиление гуморального иммунитета посредством: увеличения количества антителообразующих клеток и циркулирующих антител; активации Тх2 и усиления выработки цитокинов (ИЛ-3, -4, -5, -6, -9, -10, -13, ГМ-КСФ и др.); сокращения периода образования IgM-ангител, преобладание IgG- и IgA-антител; повышение аффинности антител; усиление клеточного иммунитета посредством: увеличения числа антигенспецифических Т-киллеров и Т-эффекторов гиперчувствительности замедленного типа; активации Tx1 и усиления выработки цитокинов (ИФ-γ, ФИО, ИЛ-2, ГМ-КСФ и др.); повышения аффинности антигенспецифических рецепторов Т-клеток; повышение устойчивости к заражению.
Способность быстро реагировать на повторный контакт с антигеном организм приобретает благодаря иммунологической памяти. Она характерна для клеточного и гуморального иммунитета, зависит от формирования Т- и В-клеток памяти. Иммунологическая память развивается после перенесенной инфекции или вакцинации и сохраняется длительное время. При некоторых инфекциях антитела в сыворотке крови присутствуют на протяжении десятилетий. Вместе с тем полупериод жизни самого устойчивого иммуноглобулина составляет в среднем 25 дней. Таким образом, в организме постоянно происходит ресинтез специфического иммуноглобулина. Длительность постинфекционного иммунитета зависит от свойств возбудителя, инфицирующей дозы, состояния иммунной системы, генотипа, возраста и других факторов. Иммунитет может быть кратковременным, например, при гриппе, дизентерии, возвратном тифе, достаточно продолжительным, например при сибирской язве, риккетсиозе, лептоспирозе, и даже пожизненным, например при полиомиелите, кори, коклюше. Приобретенный иммунитет является хорошей защитой против заражения соответствующим возбудителем. Если основным механизмом иммунитета при данной инфекции является эффект нейтрализации, то наличие определенного уровня циркулирующих антител достаточно для предупреждения реинфекции. Для достижения стойкого иммунитета вакцины приходится вводить 2 раза и более. Первичная вакцинация может состоять из нескольких доз вакцины, интервалы между дозами строго регламентированы. График проведения ревакцинации более свободный, ревакцинацию можно проводить через год и даже через несколько лет. Интервал между введениями вакцины должен быть не менее 4 недели. В противном случае развивается менее стойкий иммунитет. Наоборот, некоторое увеличение 4-недельного интервала может усилить вторичный иммунный ответ. Максимальное повышение концентрации антител при вторичном ответе на вакцины возникает при невысоких исходных титрах антител. Высокий предшествующий уровень антител препятствует дополнительной выработке антител и длительному их сохранению, а в некоторых случаях наблюдается снижение титров антител [26-40].
Вместе с тем, традиционно используемые адъюванты в плане усиления иммуногенности ИЛП не лишены недостатков. Так, адъюванты бактериального происхождения (например, адъювант Фрейнда) при попадании в организм способствует развитию большого количества побочных реакций. Соединения алюминия, несмотря на широкое применение, резко повышают аллергезирующие свойства ИЛП, кроме того, они эффективны только в отношении тех антигенов, которые требуют индукции гуморального иммунитета.
В настоящее время при поиске эффективных адъювантов отдается предпочтение препаратам избирательного целенаправленного действия, и этот поиск значительно расширился в связи с успехами, достигнутыми на пути расшифровки механизмов иммунореактивности. Исследования показали, что все ее звенья находятся под контролем цитокинов. Последние синтезируются различными типами клеток, участвующими в развитии иммунного ответа, и характеризуются определенной специфичностью действия [40-42]. Достижения иммунофармакологии последних лет позволяют по-новому подойти к этой проблеме. В частности, известно, что при совместном применении с вакцинными препаратами адъюванты по-разному влияют на компоненты и этапы иммунного ответа (макрофаги, В-, Т-клетки и их субпопуляции, естественные киллеры, процессы миграции и кооперации и др.). Это создает принципиальную возможность дифференцированного применения адъювантов, ориентированного на конкретные клетки-мишени. Селективная модуляция тех или иных звеньев иммунитета адъювантами при их введении с вакцинами для повышения иммуногенности последних может оказаться весьма перспективным направлением вакцинологии. С этой целью можно использовать препараты, оказывающие регулирующее влияние на функционально состояние клеток моноцитарно-макрофагального звена иммунной системы, обеспечивая в этом плане сбалансированность, специфичность и эффективность иммунных реакций организма на антигенное воздействие.
Учитывая вышеизложенное, можно заключить, что в настоящее время некоторые иммуномодулирующие средства могут быть использованы в качестве адъювантов ИЛП. Вместе с тем, эти препараты, в основном, оказывают иммуномодулирующее действие, активируя процессы специфического антителогенеза применительно к используемому в сочетании ИЛП, но при этом запуская как иммуногенез, так и выраженную воспалительную реакцию, без которой, по мнению исследователей не возможен полноценный иммунный ответ. Однако повышенная воспалительная реакция может привести и к нежелательным последствиям, а именно реактогенности прививок, формированию в поствакцинальном периоде иммунодефицита и др. В этой связи, остается актуальной задача поиска вакцинных адъювантов, которые с одной стороны способствовали повышению иммуногенности ИЛП, а с другой нивелируют его побочные эффекты, в частности, сопряженные с поствакцинальным воспалением. Таким образом, использование в качестве адъювантов синтетических дериватов производных фталгидразида, в частности, адьюванта Абидова, является неочевидным для специалиста. Для таких веществ известно, что на уровне неспецифической иммунологической резистентности они способны регулировать активность макрофагов, соответственно, продукцию активных форм кислорода и других острофазовых белков, ответственных за развитие токсического синдрома. Увеличивают функциональную антибактериальную активность нейтрофильных гранулоцитов, усиливает фагоцитоз и повышает неспецифическую защиту организма. На уровне системы цитокинов - снижает избыточный синтез ФНО и ИЛ-1. На уровне Т- и В-систем иммунологической защиты - активируются как Т-, так и В-лимфоциты. При различных инфекциях усиливает иммунный ответ (Абидов М.Т. Токсический синдром, патогенез, методы коррекции, 1994, Бюллетень экспериментальной биологии и медицины, 2000) Учитывая вышеизложенное, можно предположить возможность использования производных фталгидразида в качестве адъювантов ИЛП (далее - адьювант Абидова) с целью не только повышения их иммуногенности, но и, что особенно важно, для снижения побочного действия ИЛП, о чем упоминание в доступной литературе отсутствует.
Среди данных веществ известны соединения, обладающие определенной противоинфекционной активностью [43], в частности, фурункулеза стафилококковой и стрептококковой этиологии, герпетической инфекции, а также Mycobacterium tuberculosis [44, 45]. Некоторые производные фталгидразида применяются в схемах терапии панкреонекроза [46], бронхиальной астмы [47], пародонтита [48, 49], вирусной инфекции [50].
Техническими результатами заявленного изобретения являются:
- проявление адъювантного эффекта, т.е. повышения иммунного ответа, при совместном, раздельном (в разные анатомические области тела организма) введении адьюванта Абидова с ИЛП, что является неожиданным в свете существующего уровня техники. Специалист не мог бы ожидать, как проявления самого адъювантного эффекта, так и, тем более, его проявления именно при одновременном (под одновременным введением понимается минимально возможный интервал между введением адьюванта Абидова и иммуногена, антигена) раздельном введении с ИЛП;
- повышение иммуногенности ИЛП, а с другой нивелирование побочных эффектов ИЛП, в частности, сопряженных с поствакцинальным воспалением;
- снижение реактогенности прививок, снижение случаев формирования в поствакцинальном периоде иммунодефицита;
- повышение иммуногенности не менее чем в 3 раза инактивированных и химических вакцин;
- повышение безопасности живых вакцин, не влияя на иммуногенность, по средствам снижения реактогенности;
- повышение иммуногенности ассоциаций полианатоксинов не менее чем на 30% каждого из входящих в ассоциацию антигена (анатоксина);
- активация регенеративных процессов на уровне тканей при повреждении, посредством вируса, например, легких. Из уровня техники невозможно было предположить влияние адьюванта Абидова на регенерацию, например легких.
Целью изобретения является повышение эффективности борьбы с инфекционными заболеваниями, вызываемыми возбудителями опасных и особо опасных инфекций различной этиологии, а также иммунологической эффективности используемых для этого иммунобиологических лекарственных препаратов.
Цель достигается тем, что для повышения эффективности иммунопрофилактики опасных и особо опасных инфекций различной этиологии используют производное фталгидразида - адьювант Абидова в разовой дозе 150 мкг/особь и, или в иной дозе, от 0,01 до 1000 мкг/особь в зависимости от нуждающегося субъекта, вводимый однократно или многократно, внутримышечно, совместно и/или раздельно с соответствующим конкретной инфекции иммунобиологическим лекарственным препаратов.
В таблице 1 представлена оценка адъювантной активности адьюванта Абидова в отношении ИЛП против герпетической инфекции.
В таблице 2 представлена оценка адъювантной активности адьюванта Абидова в отношении ИЛП против венесуэльского энцефаломиелита лошадей.
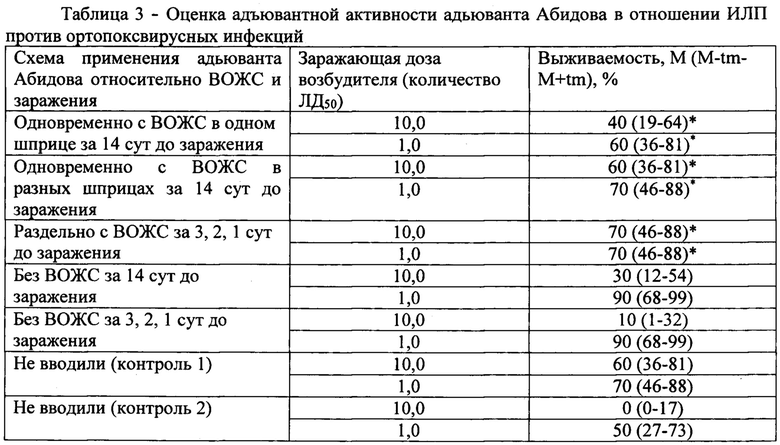
В таблице 3 представлена оценка адъювантной активности адьюванта Абидова в отношении ИЛП против ортопоксвирусных инфекций.
В таблице 4 представлена оценка адъювантной активности адьюванта Абидова в отношении ИЛП против ботулизма и столбняка.
В таблице 5 представлена оценка адъювантной активности адьюванта Абидова в отношении ИЛП против брюшного тифа и дизентерии.
В таблице 6 представлена оценка адъювантной активности адьюванта Абидова в сравнительном аспекте в отношении ИЛП против оспы коров и венесуэльского энцефаломиелита лошадей.
Способ реализуется использованием для повышения иммуногенности ИЛП адьюванта Абидова в разовой дозе 150 мкг/особь внутримышечно одновременно или раздельно с ИЛП.
Адьювант Абидова представляет собой натриевую соль 5-амино-1,2,3,4-тетрагидрофталазин-1,4-диона, натриевая соль, иммуностимулирующее средство, оказывает адьювантное действие. Влияет на уровни провоспалительных цитокинов, ответственных за развитие токсического синдрома. Снижает так же уровни реакционно способных кислородных анион-радикалов ответственных за образования фиброза в легких. Нормализует функциональное состояние макрофагов, приводит к восстановлению их антиген-представляющей и регулирующей функции. Увеличивая функциональную антибактериальную активность нейтрофильных гранулоцитов, усиливает фагоцитоз и повышает неспецифическую защиту организма. Препарат может быть использован при острых и хронических заболеваниях, различного генеза сопровождающихся интоксикацией, а также при гнойно-септических осложнениях - в составе комплексной терапии.
Приводим конкретные примеры реализации способа.
Пример 1. Адъювантные свойства адьюванта Абидова в отношении ИЛП против герпетической инфекции.
Для моделирования герпетической инфекции использовали вирус герпеса простого (ВГП) 1 типа, штамм УС, исходный титр вируса 102-103 ЛД50/МЛ или ВГП 2 типа, штамм ВН, исходный титр вируса 102-103 ЛД50/МЛ. Исследования выполнены на белых беспородных мышах, полученных из питомника «Рапполово» РАН. Всего в исследованиях было использовано 300 белых беспородных мышей-самцов массой 16-18 г. Опыты проводили на животных, в обязательном порядке выдержавших карантин в течение 1 недели. Для внутримозгового накопления модельного вируса, а также определения инфекционной активности вируссодержащего материала использовали мышей-сосунков. Накопление ВГП проводили посредством внутримозгового церебрального заражения мышей-сосунков. Инфекционную активность вируссодержащих материалов определяли по методу Рида-Менча. Величину ЛД50 применительно к каждому возбудителю определяли на белых беспородных мышах по методу Кербера в модификации И.П. Ашмарина и А.А. Воробьева (1962). Заражение осуществляли путем подкожного заражения животных суспензией вируса простого герпеса 1 типа в объеме 0,5 мл/особь в дозе 2,5 ЛД50. Для повышения восприимчивости животных к инфекции их подвергали однократному облучению за 4 суток до заражения на установке ИГУР-1 при мощности дозы 1,5 Гр/мин. Совокупная доза облучения составила 5,5 Гр. Численность животных в опытных и контрольных группах составляла по 10 особей в каждой. Заражение осуществляли спустя 28 суток после иммунизации животных. Для иммунизации использовали вакцину герпетическую культуральную инактивированную сухую (ГВ) (сер. 126, контр. №718). Иммунизирующая доза ИЛП составляла 0,1 человеческой дозы. Вакцину вводили однократно подкожно в объеме 0,5 мл в холку животным. Адьювант Абидова вводили животным внутримышечно в правое бедро задней лапы в дозе 150 мкг/особь. Было апробировано две схемы иммунизации:
- адъювант Абидова вводили одновременно раздельно с ИЛП;
- адъювант Абидова вводили на фоне текущего иммунизационного процесса - за 3,2, 1 суток до заражения.
Животным контрольных групп вводили: только ИЛП, только адьювант Абидова, только физиологический раствор хлорида натрия.
Полученные результаты представлены в таблице 1.


Полученные результаты свидетельствуют о том, что адьювант Абидова обладает адъювантным действием в отношении герпесвирусной вакцины при применении по всем изученным схемам. Выживаемость в этих группах составила 80% и достоверно превышала аналогичный показатель в контроле заражения (р≤0,05). По сравнению с иммунизацией только ИЛП или применением только адьюванта, показатель выживаемости в группах животных, которым вводили и ИЛП, и адъювант Абидова оказался выше на 30-60%. В случае применения адъюванта в схемах иммунизации эффективность ИЛП повышалась на 30%, что свидетельствует о наличии у адьюванта Абидова адъювантной активности в отношении вакцины герпетической культуральной инактивированной сухой. В доступных источниках информационных материалах подобные сведения отсутствуют. Новая функция адьюванта Абидова не вытекает с очевидностью из его известных свойств и состава. Активность препарата в опытах in vivo против возбудителей, вызывающих фурункулез, острые и хронические инфекции ЖКТ, генерализованную герпетическую инфекцию без использования ИЛП, не означает, что препарат непременно будет обладать адъювантными свойствами в отношении герпетической вакцины, равно как и в отношении других ИЛП.
Для доказательства соответствия заявляемого способа критерию изобретения «промышленная применимость» приводим следующие примеры. Для моделирования инфекций использовали вирулентные штаммы возбудителей: венесуэльского энцефаломиелита лошадей (ВЭЛ), оспы коров (ОК), ботулизма, столбняка, брюшного тифа (БТ), дизентерии. Исследования выполнены на белых беспородных мышах-самцах массой 16-18 г. В исследование брали животных, в обязательном порядке выдержавших карантин течение 1 недели в условиях вивария. Величину инфекционной активности возбудителей (ЛД50) возбудителей оценивали по методу Кербера в модификации И.П. Ашмарина и А.А. Воробьева [51].
Эффективность соответствующих ИЛП при применении одновременно с адъювантом Абидова и без него определяли по сопоставлению величин показателей выживаемости животных в подопытных (иммунизированные животные ИЛП с адъювантом Абидова) и контрольных группах. Процент выживших животных в подопытных и контрольных группах определяли по таблицам Генеса B.C. [52]. Наблюдение за инфицированными животными проводили в течение 21 суток, ежедневно регистрируя число живых и павших в подопытных и контрольных группах. Помимо этого, определяли защитную эффективность (%) адьюванта Абидова, представляющую собой разность между количеством выживших инфицированных животных в подопытной группе (вводили адъювант Абидова, %) и количеством выживших инфицированных животных в контрольной группе (без адъюванта Абидова, %), а также среднюю продолжительность жизни Т, (СПЖ, суток) подопытных и контрольных животных. Последнюю вычисляли по формуле

где N - количество выживших животных, гол.;
S - срок наблюдения, сутки.;
N1 - количество зараженных мышей в группе, гол.
Статистический анализ результатов исследования проведен с помощью компьютерной программы статистической обработки данных Statistica 6.0 for Windows. Для оценки количественных показателей определялись стандартные количественные характеристики: среднее значение показателя (М), среднеквадратичное отклонение, стандартная ошибка средней величины (m). Сравнение количественных данных проводили при помощи парного и непарного теста Стьюдента с использованием t-критерия Стьюдента. Результаты представлены как М±m. Различия считали достоверными при р≤0,05.
Пример 2. Адъювантный эффект адьюванта Абидова в отношении ИЛП против венесуэльского энцефаломиелита лошадей
Оценку адъювантных свойств адьюванта Абидова в отношении вакцины венесуэльского энцефаломиелита лошадей (ВВЭЛ) проводили в «точечных» экспериментах, в каждой точке использовали по 10 мышей массой от 16 до 18 г. Вируссодержащий материал вводили животным подкожно в объеме 0,3 мл/мышь. Заражающие дозы вируса составляли 2 и 10 ЛД50. Для заражения использовали вирус венесуэльского энцефаломиелита лошадей (ВЭЛ), штамм Тринидад. Исходный титр вируса составил 107-108 ЛД50/МЛ. Для иммунизации использовали ВВЭЛ культуральную инактивированную жидкую (ВВЭЛ) (сер. 145; контр. N 1244). Адьювант применяли в дозе 150 мкг/особь, вводили внутримышечно по различным схемам. ИЛП применяли однократно, вводили внутримышечно в объеме 0,5 мл, иммунизирующая доза ИЛП составляла 0,1 человеческой дозы.
Как следует из представленных данных, ВВЭЛ, введенная однократно в дозе равной 0,1 человеческой дозы за 21 сутки до заражения в зависимости от заражающей дозы вируса обеспечивала выживаемость 15% (10 ЛД50) и 70% (2 ЛД50) инфицированных животных.
Результаты проведенных исследований представлены в таблице 2.
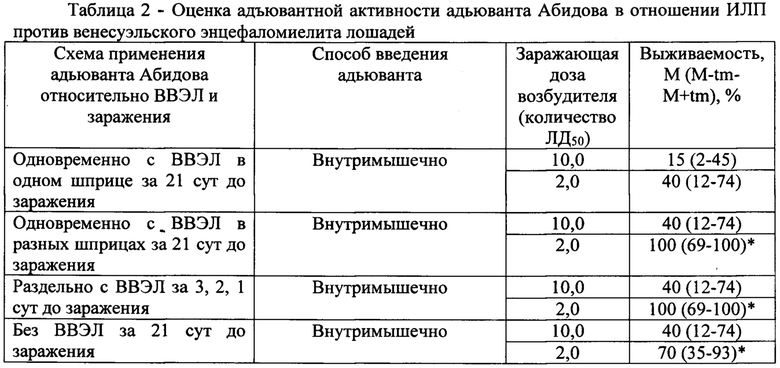

Введение в эти же сроки только адьюванта Абидова обеспечивало выживаемость 40% (10 ЛД50) и 70% (2 ЛД50) инфицированных вирусом ВЭЛ мышей. Более эффективным оказалось использование адьюванта Абидова по многократной схеме. При этом в зависимости от заражающей дозы вируса выживаемость инфицированных мышей составила 60-100%.
При одновременном введении ВВЭЛ и адьюванта Абидова в одном шприце показатели выживаемости инфицированных животных колебались в диапазонео от 15 до 40% и были либо на уровне, либо несколько меньшими, чем в группах животных, которым вводили только ВВЭЛ или только адьювант. Более эффективным оказалось одновременное применение ВВЭЛ и адьюванта, но в разных шприцах. В этом случаев в зависимости от величины заражающей дозы вируса ВЭЛ показатель выживаемости инфицированных животных составил 40% и 100%, соответственно при заражении вирусом в дозе 10 ЛД50 и 2 ЛД50, что оказалось на 25-30% выше аналогичного показателя, зарегистрированного в группах животных, иммунизированных только ВВЭЛ.
Пример 3. Адъювантный эффект адьюванта Абидова в отношении ИЛП против ортопоксвирусной инфекции
Исследования выполнены на белых беспородных мышах-самцах массой от 10 до 12 г. Численность групп - по 20 животных в каждой. Для иммунизации использовали вакцину оспенную живую сухую (ВОЖС) (сер. 80, контр. N 1841). ИЛП вводили животным в дозе 0,2 человеческой дозы, иммунизацию осуществляли однократно подкожно, объем вводимой вакцины составил 0,5 мл. Адьювант Абидова применяли в дозе 150 мкг/особь, вводили подкожно по различным схемам. Заражение мышей проводили интраназально через 14 суток после иммунизации. Для этого использовали вирус оспы коров (ВОК), штамм Пуменок. Исходный титр вируса составлял 103-104 ЛД50/мл. Вируссодержащую суспензии вводили в объеме 0,1 мл. Заражающие дозы вируса составляли 1 и 10 ЛД50. Результаты исследований представлены в таблице 3.
Установлено, что независимо от схемы сочетанного применения? не удалось выявить выраженного адъювантного действия адъюванта Абидова в отношении ВОЖС. Величины показателей выживаемости инфицированных животных, которым вводили адьювант Абидова и ВОЖС, практически не отличались от таковых, зарегистрированных в группах мышей, иммунизированных только ВОЖС. Возможно это обусловлено тем, что адъювантный эффект в отношении такого сильного антигена, каким является ВОЖС, получить достаточно сложно. В то же время, какого-либо отрицательного воздействия адьюванта на формирование специфического ортопоксвирусного иммунитета при совместном применении с вакциной не установлено. Поэтому такой вариант при проведении специфической иммунопрофилактики упомянутого заболевания с использованием ВОЖС исключить нельзя.

Пример 4. Адъювантный эффект адьюванта Абидова в отношении ИЛП против ботулизма и столбняка.
Как известно, тетраанатоксин (ТА) включает в себя анатоксины против ботулизма типов А, В, Е и столбняка. В данном препарате упомянутые анатоксины сорбированы на гидрате окиси алюминия. Для формирования специфической защиты ТА применяется трехкратно, при этом между первой и второй инъекциями интервал составляет 28-30 суток, а между второй и третьей - 4-6 мес. Исследования по оценке адъювантных свойств адьюванта Абидова в отношении ТА проводили на белых мышах и морских свинках. Доза адьюванта составляла 150 мкг/особь.
Иммунизацию осуществляли двукратно с интервалом 30 суток. Адьювант Абидова и ТА вводили в одном шприце подкожно. Объем вводимой смеси препаратов составлял 0,5 мл. В качестве инфекционной модели использовали отравление животных ботулиническим токсином типа А в дозе 100 ЛД50. Моделирование интоксикации осуществляли спустя 14 суток после второй иммунизации путем внутрибрюшинного введения токсина в объеме 0,5 мл. Результаты проведенных исследований представлены в таблице 4.


Применение одного адьюванта Абидова не оказывало существенного влияния на устойчивость животных к отравлению ботулотоксином типа А. Выживаемость мышей в данной группе составила 0%. Иммунизация одним ТА обеспечивала защиту 60-70% отравленных животных в зависимости от их вида (белые мыши или морские свинки) на фоне 100% летальности в контроле (р<0,05). Наиболее эффективным оказалось сочетанное применение ТА и адьюванта Абидова. В этом случае удалось достигнуть 100% выживаемости животных на фоне полной летальности в контроле (р<0,05). Более того, этот показатель был достоверно выше, чем в группе животных, привитых только ТА (р<0,05). Практически полностью идентичные результаты были получены на модели интоксикации столбнячным токсином.
Пример 5. Адъювантный эффект адьюванта Абидова в отношении ИЛП против острых кишечных инфекций
Для иммунопрофилактики брюшнотифозной инфекции использовали вакцину брюшнотифозную Ви-полисахаридную жидкую («ВИАНВАК»). Препарат вводили однократно подкожно в объеме 0,5 мл, в дозе, эквивалентной 0,02 и 0,2 человеко-дозы. Для иммунопрофилактики дизентерии использовали вакцину дизентерийную «Шигеллвак» липополисахаридную из штамма Зонне. Препарат вводили однократно подкожно в объеме 0,5 мл в дозе, эквивалентной 0,2 человеко-дозы.
Характеристика модельных биопатогенов.
Возбудитель брюшного тифа - штамм Breslau. Суспензию возбудителя готовили следующим образом: на агаре Мюллер-Хинтона при температуре 37°С в течение 24 ч выращивали микробную культуру. По окончании культивирования готовили серийные разведения выращенной микробной суспензии по стандарту мутности с шагом 10 (начиная с разведения 109 до 102 КОЕ/мл). Полученную суспензию использовали для дальнейшего подкожного заражения экспериментальных животных.
Возбудитель дизентерии - штамм Sh. Sonnei II. Суспензию возбудителя готовили следующим образом: на агаре Мюллер-Хинтона при температуре 37°С в течение 24 ч выращивали микробную культуру. Для подтверждения чистоты высев осуществляли и на среду Эндо. По окончании культивирования готовили серийные разведения выращенной микробной суспензии по стандарту мутности с шагом 10 (начиная с разведения 109 до 102 КОЕ/мл). Полученную суспензию использовали для дальнейшего подкожного заражения экспериментальных животных. Исследования по оценке иммунотропных эффектов адьюванта Абидова при его совместном применении с ИЛП выполнены на белых не инбредных мышах-самцах массой 16-18 г (600 голов). В исследования брали животных, в обязательном порядке выдержавших карантин в течение 1 недель.
Моделирование брюшнотифозной инфекции in vivo осуществляли на белых беспородных мышах массой 16,0-18,0 г путем внутрибрюшинного заражения взвесью спор штамма Breslau в объеме 0,5 мл. Наблюдение за инфицированными животными осуществляли в течение 14 суток, ежедневно регистрируя число живых и павших в опытных и контрольных группах. Специфичность гибели животных подтверждали бактериологически - посевом селезенки павших животных на специальную питательную среду. Моделирование дизентерийной инфекции in vivo осуществляли на белых беспородных мышах массой 16,0-18,0 г путем внутрибрюшинного заражения взвесью спор штамма Sh. Sonnei II в объеме 0,5 мл. Наблюдение за инфицированными животными осуществляли в течение 14 суток, ежедневно регистрируя число живых и павших в опытных и контрольных группах. Специфичность гибели животных подтверждали бактериологически - посевом селезенки павших животных на специальную питательную среду.
Антитела к брюшнотифозным антигенам у животных, иммунизированных вакциной брюшнотифозной Ви-полисахаридной жидкой «ВИАНВАК», определяли в сыворотке крови на 21 сутки после иммунизации с помощью набора реагентов для определения антител к антигенам бактерий тифо-паратифозной группы, бруцеллам и протею в реакции агглютинации («Анти-Бактантиген-Тест») (сер. 04/15.). Постановку реакции осуществляли в соответствии с «Инструкцией…» по применению набора. Антитела к шигеллезным антигенам Зонне определяли в сыворотке крови иммунизированных соответствующим ИЛП животных на 21 суток после иммунизации с помощью препарата «Реплан» - диагностикум эритроцитарный шигеллезный Зонне антигенный (лиофилизат для диагностических целей) (серия 950914). Диагностический препарат применяли согласно «Инструкции…» по применению. Определение титра противошигеллезных антител осуществляли в РПГА.
В ходе проведенных исследований иммунизацию животных осуществляли по схеме: первоначально животных иммунизировали соответствующим ИЛП, затем одновременно с ИЛП и далее через 3 суток и 6 суток после иммунизации ИЛП животным вводили адьювант Абидова внутрибрюшинно в объеме 0,5 мл в разовой дозе 150 мкг/особь.
До иммунизации и спустя 21 сутки после окончания последнего введения адьюванта у животных методом декапитации отбирали кровь, в сыворотке крови производили определение титров соответствующих специфических антител.
Результаты проведенных исследований сведены в таблицу 5.

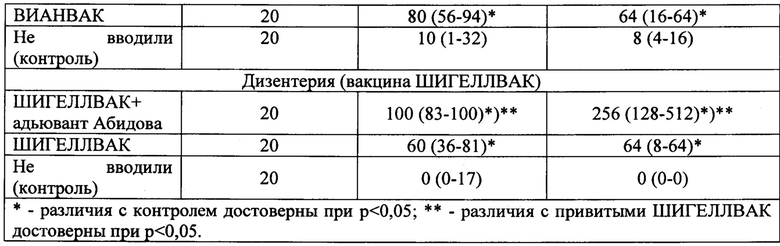
Иммунизация брюшнотифозной вакциной. Результаты, приведенные в таблице 5, показали, что иммунизация брюшнотифозной вакциной «ВИАНВАК» обеспечивала формирование устойчивого антителогенеза к сальмонеллезным антигенам (в частности О-антигену S. Typhi) у 80% иммунизированных животных. При этом титр специфических антител в сыворотке крови (Me) составил 1:64. В обоих случаях различия с контролем были статистически достоверными (р<0,05). При комбинированном применении брюшнотифозной вакцины «ВИАНВАК» с адьювантом Абидова процент положительных сероконверсий достигал 100%, что на 20% выше, чем среди привитых только ИЛП, а уровень специфических сывороточных антител (Me) достигал 1:256, что достоверно превышало как контрольные значения, так и аналогичный показатель у привитых только ИЛП (р<0,05).
Иммунизация шигеллезной вакциной. Результаты, приведенные в таблице 5, показали, что иммунизация вакциной «Шигеллвак» обеспечивала формирование иммунного ответа в отношении шигелл Зонне у 60% иммунизированных животных, то есть уровень положительных сероконверсий составил 60%, что достоверно превышало контрольные значения (р<0,05). При этом средняя величина титра специфических антител в сыворотке крови привитых мышей (Me) составила 1:64, что также достоверно превышало контрольные значения (р<0,05). В случае комбинированной иммунизации препаратом «Шигеллвак» и адьювантом Абидова показатель положительных сероконверсий достигал 100%, что достоверно превышало как контрольные значения, так и аналогичный показатель у привитых только Шигеллвак. В этих условиях происходило и более интенсивное образование специфических сывороточных антител, о чем свидетельствовала величина среднего титра (Me), составившая 1:256, что было достоверно выше в 4 раза, чем в контроле и у привитых только Шигеллвак (р<0,05).
Пример 6. Адъювантная активность адьюванта Абидова в отношении ИЛП против венесуэльского энцефаломиелита лошадей и ортопоксвирусной инфекции, примененных в составе ассоциации (ВЭЛ + ВОЖС).
Опасные инфекционные заболевания (ОИЗ) представляют достаточно серьезную в эпидемиологическом отношении группу инфекций, возбудители которых характеризуются выраженной патогенностью, вирулентностью и контагиозностью применительно к человеческой популяции, что способствует их достаточно быстрому распространению, охвату больших контингентов и нанесению серьезного урона здравоохранению и национальной безопасности государств, на территории которых они регистрируются. В качестве примера можно привести периодически возникающие эпидемии лихорадок Эбола, Зика, денге, Западного Нила, коронавирусных инфекций и др. Поэтому вопросам профилактики и лечения ОИЗ уделялось и продолжает уделяться пристальное внимание эпидемиологов и инфекционистов, причем приоритет отдается иммунопрофилактике как наиболее действенному мероприятию в отношении данной группы инфекций. Причем первостепенное значение имеет комплексность в применении ИЛП, поскольку в результате резкого ухудшения санитарно-эпидемической обстановки в том или ином регионе высока вероятность возникновения и быстрого прогрессирования не одной, а нескольких инфекций одновременно. В последнее время внимание исследователей приковано к так называемым «комплексным вакцинальным системам» (КВС), которые призваны явиться альтернативой существующих средств иммунопрофилактики (ИП). Подобные системы включают в себя ассоциации и комплексы ИЛП, способствующие одновременному введению в организм нескольких антигенных раздражителей, а также ИЛП, основанные на наночастицах, покрытых несколькими антигенными детерминантами в отношении нескольких инфектов.
В структурном отношении КВС представляют собой наночастицы различной природы, нагруженные наиболее активными в иммунологическом отношении детерминантами возбудителей инфекций [53-55]. Основным принципом конструирования КВС является повышение защищенности иммуногенных субстанций от воздействия ферментных систем организма и, как следствие, их стабильности и биодоступности в отношении иммунокомпетентных клеток [56].
Учитывая вышеизложенное и для полноты доказательности адъювантных свойств адьюванта Абидова мы провели исследования по оценке его адъювантных свойств в условиях одномоментного раздельного введения с ассоциацией ИЛП против ВЭЛ и ортопоксвирусной инфекции.
Для иммунопрофилактики ВЭЛ использовали ВВЭЛ культуральную инактивированную жидкую (сер. 145; контр. №1244). Животных иммунизировали однократно подкожно, иммунизирующая доза препарата составила 0,1 человеко-дозы. Для иммунопрофилактики ортопоксвирусной инфекции использовали вакцину оспенную живую сухую (сер. Т. 15, контр. №1542). Препарат применяли однократно подкожно в объеме 0,5 мл в прививочной дозе, эквивалентной 0,2 человеко-дозы.
В качестве инфицирующих возбудителей:
- вирус ВЭЛ - патогенный штамм Тринидад. Накопление вируссодержащего материала для последующего заражения лабораторных животных осуществляли с использованием 9-11 дневных развивающихся куриных эмбрионов - 30-50 шт. Первоначально готовили пять последовательных десятикратных разведений вируссодержащей суспензии. По 0,2 мл каждого разведения вируссодержащей суспензии вносили в аллантоисную полость развивающихся куриных эмбрионов. Место инъекции вируссодержащей суспензии покрывали расплавленным парафином. Затем развивающиеся куриные эмбрионы помещают в термостат при температуре (37±0,5)°С на 18 ч, периодически оценивая их жизнеспособность с помощью овоскопа. По истечении времени инкубации в термостате оценивали жизнеспособность развивающихся куриных эмбрионов и из «тушек» живых эмбрионов готовили 10% суспензию вируссодержащего материала с использованием физиологического раствора с добавлением антибиотиков (пенициллин из расчета 100 ЕД на 1 мл, стрептомицин - 200 ЕД на 1 мл). Полученную суспензию центрифугировали в течение 10 мин при 1,5-2,0 тыс.об./мин и температуре плюс (3±0,5)°С. Надосадочную жидкость разливали во флаконы объемом 1,0 мл и использовали для дальнейшего заражения экспериментальных животных мышей. Исходный титр вируса 107-108 ЛД50/МЛ.
- вирус ОК - патогенный штамм Пуменок. Накопление вируссодержащего материала для заражения лабораторных животных осуществляли с использование 11-12-дневных развивающихся куриных эмбрионов - 30-50 шт. Первоначально готовили пять последовательных десятикратных разведений вируссодержащего материала. По 0,2 мл каждого разведения вносили на хорион-аллантоисную оболочку развивающихся куриных эмбрионов. Место инъекции покрывали расплавленным парафином. Инфицированные развивающиеся куриные эмбрионы помещали в термостат при температуре (35±0,5)°С на 72 ч, ежедневно оценивая их жизнеспособность с помощью овоскопа. По окончании времени инкубации оценивали жизнеспособность развивающихся куриных эмбрионов и из хорион-аллантоисной оболочки живых готовили 60 мл 10% суспензии вируссодержащего материала с использованием физиологического раствора с добавлением антибиотиков (пенициллин из расчета 100 ЕД на 1 мл, стрептомицин - 200 ЕД на 1 мл). Полученную суспензию вируссодержащего материала центрифугировали в течение 10 мин при 1,5-2,0 тыс.об./мин и температуре (3±0,5)°С. Надосадочную жидкость разливали во флаконы объемом 10 мл (пенициллиновые флаконы) и использовали для заражения лабораторных животных. Исходный титр вируса 103-104 ЛД50/мл.
Исследования выполнены на белых не инбредных мышах-самцах массой 16-18 г (350 голов) и 10-12 г (180 голов). В исследования брали животных, в обязательном порядке выдержавших карантин в течение 1 недели.
Моделирование ВЭЛ in vivo проводили на белых беспородных мышах массой 16,0-18,0 г путем интраназального заражения вируссодержащим материалом в объеме 0,3 мл. Использовали две заражающие дозы возбудителя, вызывающие в контроле гибель 57-100% инфицированных животных. После заражения за мышами устанавливали ежедневное наблюдение в течение 21 суток с регистрацией количества живых и павших особей. Инкубационный период моделируемой инфекции составлял в среднем 5 суток. Заболевание характеризовалось следующими признаками: экспериментальные животные становились малоподвижными, отказывались от еды и питья, шерсть у них была взъерошена. В последующем развивались и нарастали явления энцефалита (нарушение координации, появление парезов и параличей) и наступала гибель инфицированных животных. Максимальная гибель мышей отмечалась, как правило, на 7-9 день после заражения.
Моделирование ОК проводили на белых беспородных мышах массой 10,0-12,0 г, предварительно проводя им эфирный рауш-наркоз и в дальнейшем проводили интраназальное заражение вирусом ОК в объеме 0,03 мл в каждую ноздрю. В эксперименте использовали две заражающие дозы возбудителя, которые различались на один порядок. После заражения за инфицированными животными устанавливали ежедневное наблюдение в течение 14 суток с регистрацией количества живых и павших особей.
Антитела к оспенным антигенам определяли в сыворотке крови иммунизированных соответствующим ИЛП животных на 21 суток после иммунизации. Наличие специфических противооспенных антител в сыворотках крови определяли с помощью общепринятых серологических реакций - реакции нейтрализации (РН), реакции пассивной гемагглютинации (РПГА), реакции торможения гемагглютинации (РТГА), реакций непрямой гемагглютинации (РИГА). Антитела к антигенам вируса ВЭЛ определяли в сыворотке крови иммунизированных соответствующим ИЛП животных на 21 суток после иммунизации. Наличие специфических антител к вирусу ВЭЛ определяли с помощью общепринятых серологических реакций - РН, РПГА, РТГА, РИГА.
В ходе проведенных исследований иммунизацию животных осуществляли по схеме: первоначально животных иммунизировали комбинацией ИЛП (ВЭЛ + ВОЖС), а адъювант вводили одновременно с комбинацией ИЛП и далее на 3 сутки и 6 сутки после иммунизации комбинацией ИЛП внутрибрюшинно в объеме 0,5 мл в дозе 150 мкг/особь. До иммунизации и спустя 21 сутки после окончания последнего введения адьюванта Абидова у животных методом декапитации отбирали кровь, в сыворотке крови производили определение титров соответствующих специфических антител. Совокупность полученных результатов приведена в таблице 6. Как следует из представленных в таблице 6 данных, ВВЭЛ обеспечивала 70% сероконверсий, что достоверно превышало контрольные параметры, поскольку у животных контрольной группы специфические антитела к вирусу ВЭЛ в сыворотке крови не определялись.
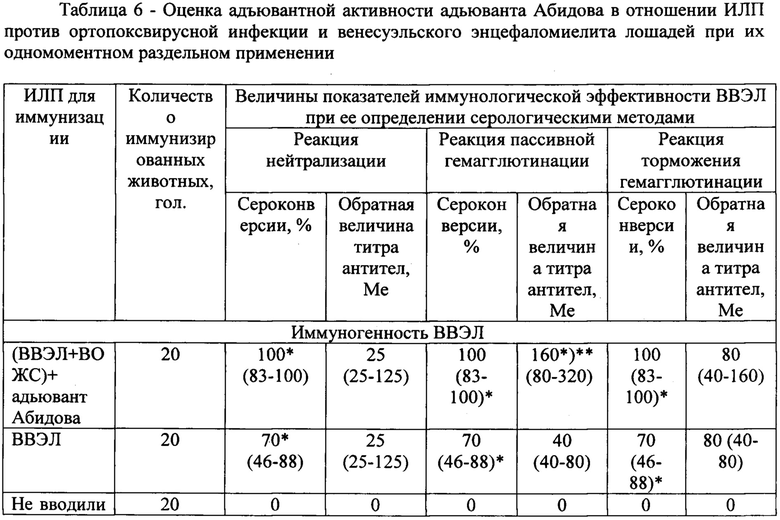

В случае, когда на фоне применения ИЛП животным вводили адьювант Абидова, то иммуногенные свойства вакцины возрастали. Это проявлялось как увеличением уровня положительных сероконверсий (на 30% по сравнению с показателями у привитых только ВВЭЛ), так и титров специфических сывороточных антител, что наиболее отчетливо проявлялось при титровании иммунных сывороток в РПГА (р<0,05). Сравнение полученных результатов с аналогичными по защитной эффективности (таблица 2) позволяет заключить, что выявленные более высокие показатели выживаемости животных, инфицированных вирусом ВЭЛ, предварительно иммунизированных и получавших совместно с вакциной адьювант Абидова, могут быть обусловлены более выраженным антителогенезом под влиянием препарата.
Применительно к иммунизации ВОЖС в составе ассоциации ВЭЛ + ВОЖС оказалось, что оспенная вакцина обеспечивала формирование противооспенного иммунитета у всех 20 иммунизированных животных к 21 сутки поствакцинального периода как при самостоятельном применении, так и в ассоциации с ВВЭЛ. Уровень сероконверсий составил 100%, что достоверно отличалось от контрольного уровня. При этом в зависимости от метода выявления специфических противооспенных антител в сыворотке крови титры колебались от 1:25 (в реакции нейтрализации) до 1:40 (в реакции пассивной гемагглютинации). Поскольку среди контрольных животных ни у одной особи специфических противооспенных антител в сыворотке крови не выявлено, то различия в титрах между иммунизированными ВОЖС и контрольными мышами достоверны при р<0,05. Эти данные согласуются с исследованиями по оценке напряженности поствакцинального иммунитета, проведенными на модели ОК, вызванной одноименным вирусом, штамм Пуменок. При этом также не было зарегистрировано ощутимого вклада препарата в повышение иммуногенности ИЛП, однако и не было выявлено отрицательного влияния адьюванта Абидова на иммунологическую эффективность ВОЖС. Нельзя, однако, не признать, что под его влиянием происходила интенсификация процессов противооспенного антителогенеза, что проявлялось в увеличении титра специфических противооспенных антител в сыворотке крови иммунизированных комбинацией препаратов животных в 4 раза по сравнению с аналогичными показателями у привитых только ВОЖС, причем различия были статистически достоверными (р<0,05).
Суммируя результаты проведенных исследований, можно заключить, что:
- адьювант Абидова - перспективный иммунотропный препарат с адъювантной активностью в отношении ИЛП различной природы против инфекционных агентов, вызывающих опасные и особо опасные инфекции бактериальной, вирусной и токсинной природы;
- адъювант Абидова проявляет адъювантные свойства при использовании в дозе 150 мкг/особь, введении однократно, внутримышечно как одновременно раздельно с ИЛП, так и на фоне уже текущего иммунизационного процесса либо за 3-1 суток до предполагаемого заражения, либо одновременно с ИЛП и далее спустя 3 суток, 6 суток после введения ИЛП;
- адъювантые свойства адъюванта Абидова, заключаются в стимуляции специфического антителогенеза как минимум на 30% по сравнению с таковым под влиянием только ИЛП, причем подобный эффект наиболее отчетливо проявляется при использовании адъюванта в сочетании с инактивированными и химическими вакцинами, а также полианатоксинами;
- адъювант Абидова, при сочетанном применении с живыми вакцинами, на примере ВОЖС, не приводит к повышению иммуногенных свойств подобных препаратов, однако и не оказывает и отрицательного иммунотропного действия. Учитывая наличие у адъюванта Абидова противовоспалительного действия, его применение с живыми вакцинами оправдано в плане снижения их реактогенности, обусловленной провоспалительными эффектами подобных ИЛП в начале поствакцинального периода, особенно на этапе первичного IgM ответа;
- адъювант Абидова обладает положительным иммунотропным действие не только в отношении моновакцин, но и вакцинных препаратов или иммуноантигенов в составе ассоциированных препаратов (ТА) и комплексных вакцинальных систем (ассоциация ВЭЛ + ВОЖС). Причем, повышение иммуногенности практически в одинаковой степени имеет место в отношении каждого из антигенов, входящих в ассоциацию или комплекс, за исключением живых вакцин. Применительно к последним использование адъюванта не столько направлено на повышение их иммуногенности, сколько на снижение реактогенности прививки и, как следствие, повышение Безопасности подобной иммунизации;
- механизм адъювантного действия адъюванта Абидова обусловлен одновременным запуском под его влиянием систем неспецифической иммунологической резистентности, цитокинов и хемокинов, повышению интенсивности перерабатывания антигенное до иммуногенных форм и, как следствие, более выраженной стимуляции специфического антителогенеза. Более того, попадая в организм вместе с ИЛП, адьювант Абидова обладает способностью не только выступать в роли иммунологического адъюванта, но и, возможно, препятствует развитию патологического процесса инфекций на уровне иммунной системы. В частности, последний, как известно, сопряжен с иммунными дисфункциями, которые развиваются под влиянием либо самих возбудителей инфекций, либо продуктов их жизнедеятельности. При этом имеет место дисбаланс в функционировании как иммунокомпетентных клеток и клеток системы неспецифической иммунологической резистентости, так и нарушение их секреторной активности, приводящее в конечном итоге к дисбалансу медиаторной составляющей иммунной системы;
- учитывая полученные результаты, доказывающие принадлежность адъюванта Абидова к иммунологическим адъювантам, проявляющим подобное действие в отношении ИЛП различной природы и направленности, предполагаются следующие приоритетные области его применения в иммунологии и вакцинологии:
• в составе схем иммунопрофилактики инфекционных заболеваний бактериальной, вирусной и другой природы, включающих использование существующих ИЛП или их ассоциаций и комплексов на основе инактивированных, химических рекомбинантных препаратов вследствие недостаточной иммуногенности подобных препаратов;
• в составе иммунобиологических лекарственных препаратов на основе слабоиммуногенных антигенов (антигенных детерминант, протективных антигенов, компонентов наружных мембран бактерий и вирусов, отвечающих за их патогенные и вирулентные свойства и др.) в качестве одного из компонентов структуры ИЛП;
• в качестве препарата, повышающего безопасность без снижения иммуногенности препарата в схемах иммунопрофилактики инфекций различной природы с использование живых вакцин (при иммунизации против чумы, туляремии, сибирской язвы, бруцеллеза, желтой лихорадки и др.);
• в качестве препарата замены традиционным сорбирующим субстанциями (гидрат окиси алюминия и др.) при производстве сорбированных моно или полианатоксинов, инактивированных вакцин и др.
Полученные результаты следует рассматривать в качестве обоснования использования адьюванта Абидова при проведении, как взрослым, так и детям, иммунопрофилактики инфекционных заболеваний различной природы, в том числе герпеса простого, венесуэльского энцефаломиелита лошадей, ортопоксвирусных инфекций, ботулизм, столбняк, брюшной тиф, дизентерия Зонне и др.
Список литературы
1. Руководство по военной микробиологии / Под общ. ред. П.И.Мельниченко [и др.]. - М.: Военное издательство, 2005. - 511 с.
2. Den biologiske trussel og det biologiske beredskab i Danmark / E.D. Heegaard [et al.] // Ugeskr. Laeger. - 2005. - Vol. 167, №36. - P. 3381-3384.
3. Спирин A.C. Современная биология и биологическая безопасность / А.С. Спирин // Человек. - 1998. - №5. - С. 5-11.
4. CBR Threats: A case study / K. Cuneo [et al.] // Crisis Res. J. - 2005. - Vol. 1, №3. - P. 50-52.
5. Public Health Assessment of Potential Biological Terrorism Agents / L.D. Ron [et al.] // Emerging Infectious Diseases. - 2002. - Vol. 8, №2. - P. 18-24.
6. VienPoint: Terrorism and dispelling the myth of a panic prone public / B. Sheppard [et al.] // J. Public Health Policy. - 2006. - Vol. 27, №3. - P. 219-245.
7. Биотерроризм: национальная и глобальная угроза / Г.Г. Онищенко и др. // Вестн. Рос. Акад. Наук. - 2003. - Т. 73, №3. - С. 194-204.
8. Medzhitov R, Schneider DS, Soares MP. Disease tolerance as a defense strategy. Science. 2012; 335(6071):936-941.
9. Ashida H, Mimuro H, Ogawa M, et al. Host-pathogen interactions: cell death and infection: a double-edged sword for host and pathogen survival. J. Cell. Biol. 2011;195(6):931-942.
10. Torres BY, Oliveira JHM, Thomas Tate A, Rath P, Cumnock K, Schneider DS. Tracking resilience to infections by mapping disease space. PLoS Biol. 2016;14(4):e1002436-19.
11. Maeurer M, Rao M, Zumla A. Host-directed therapies for antimicrobial resistant respiratory tract infections. Curr. Opin. Pulm. Med. 2016; 22(3):203-211.
12. Monticelli LA, Sonnenberg GF, Abt MC, et al. Innate lymphoid cells promote lung-tissue homeostasis after infection with influenza virus. Nature. 2011; 12(11): 1045-1054.
13. Jamieson AM, Pasman L, Yu S, et al. Role of tissue protection in lethal respiratory viral-bacterial coinfection. Science. 2013; 340(6137):1230-1234.
14. Rock JR, Hogan BLM. Epithelial progenitor cells in lung development, maintenance, repair, and disease. Annu. Rev. Cell. Dev. Biol. 2011;27(1):493-512.
15. Абидов M.T. Бюллетень экспериментальной биологии и медицины 2000.
16. Ярилин А.А. Основы микробиологии. - М.: Медицина, 1999. - 607 с.
17. Hackett C.J. Focus on the immunological of effective vaccines // Immunologist - 1997. - Vol.5, №5. - P.171-174.
18. Lee J.J., Sinha K.A., Harrison J.A., et al. Tetanus toxin fragment С expressed in live Salmonella vaccines enhances antibody responses to its fusion partner Schistosoma haematobium glutathione S-transferase // Infect. Immun. - 2000. - Vol. 68, №5. - P.2503-2512.
19. Петров P.B., Хаитов P.M. Искусственные антигены и вакцины. - М.: Медицина, 1988. - 288 с.
20. Медуницин Н.В. Вакцинология. - М: Триада, 1999. - 279 с.
21. Ertl H.C.J., Xiang Z. Novel vaccine approaches // J. Immunol. - 1996. - Vol. 156. - P. 3579-3582.
22. Pardoll D.M. Paracrine cytokine adjuvants in cancer immunotherapy // Ann. Rev. Immunol. - 1995. - Vol. 13. - P. 399-415.
23. Kim J.J., Yang J.S., Dentchev Т., Dang K., Weiner D.B. Chemokine gene adjuvants can modulate immune responses induced by DNA vaccines // J. Interferon Cytokine Res. - 2000. - Vol. 20, №5. - P. 487-498.
24. Семенова И.Б., Семенов Б.Ф. Закономерности коррекции вторичных иммунодефицитов разными по своей природе иммуномодуляторами // Intern J. Immunorehabilitation. - 1997. - №6. - С. 35-40.
25. Pasquini S., Xiang Z., Wang Y., et al. Cytokines and cjstimulatory molecules as genetic adjuvants // Immunol. Cell Biol. - 1997. - Vol. 75. №4. - P. 397-401.
26. Fleo J., Tisminetzky S., Baralle F. Modulation of the immune response to DNA vaccine by costimulatory molecules // Immunology. - 2000. - Vol. 100, №2. - P. 259-267.
27. Kim J.J., Yang J.S., Montaner L., Lee D.J., Chalian A.A., Weiner D.B. Coimmunization with IFN-gamma or IL-2, but not IL-13, or IL-4 cDNA can enhance Th-1 type DNA vaccine-induced immune responses in vivo // J. Interferon Cytokine Res. - 2000. - Vol. 20, №3, - P. 311-319.
28. Iwasaki A., Stiernholm В.J.N., Chan A.K., Berinstrein N.L., and Barber B.H. Enhanced CTL responses mediated by plasmid DNA immunogens encoing costimulatory molecules and cytokines // J. Immunol. - 1997. - Vol. 158. - P.4591.
29. Tough D.F., Sun S., Zhang X., Sprent J. Stimulation of  and memory T cells by cytokines // Immunol. Rev. - 1999. - Vol. 170. - P. 39-47.
and memory T cells by cytokines // Immunol. Rev. - 1999. - Vol. 170. - P. 39-47.
30. Петров P.B., Хаитов P.M. Вакцины нового поколения на основе синтетических полионов: история создания, феноменология и механизмы действия, внедрение в практику // Intern. J. Immunorehabilitation. - 1999. -№11. - С 13-25.
31. Mebra N.K. Major histocompatibility compex and future vaccination strategies // Proc. Indian. Nat. Sci. Acad. B. - 1998. - Vol. 64. - №2. - P. 81-100.
32. Lindblad E.B., Elhay M.J., Silva R., Appelberg R., Andersen P. Adjuvant modulation of immune responses to tuberculosis subunit vaccines // Infect. Immun. - 1997. - Vol. 65, №2. - P. 623-629.
33. Sjolander A., Bengtsson K.L., Morein B. Kinetics, localisation and cytokine profile of T-cell responses to immune stimulating complexes (iscoms) containing human influenza virus envelope glycoproteins // Vaccine. - 1997. - Vol. 15, №9. - P. 1030-1038.
34. Gursel M., Gregoriadis G. The immunological co-adjuvant action of liposomal interleukin-2: the role of mode of localisation of the cytokine and antigen in the vesicles // J. Drug Target. - 1998. - Vol. 5, №2. - P. 93-98.
35. Krup O.C., Kroll I., Bose G., Falkenberg F.W. Cytokine depot formulations as adjuvants for tumor vaccines. I. Liposome-encapsulated IL-2 as a depot formulation // J. Immunother. - 1999. - Vol. 22, №6. - P.525.
36. Katz J.M., Lu X., Young S.A., Galphin J.C. Adjuvant activity of the heat-labile enterotoxin from enterotoxigenic Escherichia coli for oral administration of inactivated influenza virus vaccine // J. Infect. Dis. - 1997. - Vol. 175, №2. - P. 352-363.
37. Rajananthanan P., Attard G.S., Sheikh N.A., Morrow W.J. Novel aggregate structure adjuvants modulate lymphocyte proliferation and Th1 and Th2 cytokine profiles in ovalbumin immunized mice // Vaccine. - 1999. - Vol. 18, №1-2. - P. 140-152.
38. Кетлинский C.A., Симбирцев A.C., Воробьев A.A. Эндогенные иммуномодуляторы. - СПб.: Гиппократ, 1992. - С. 256.
39. Кетлинский С.А., Калинина Н.М. Иммунология для врача. - СПб.: Гиппократ, 1998. - 156 с.
40. Фрейдлин И.С. Иммунная система и ее дефекты. - СПб.: НТФФ «Полисан», 1998. - 112 с.
41. Pighetti G.M., Sordillo L.M. Enhanced antigen-specific responses in bovine mammary glands following administration of interleukin-2 // J. Dairy Sci. -1995. - Vol. 78, №3. - P. 528-537.
42. Souberbielle B.E., Knight B.C., Morrow W.J., et al. Comparison of IL-2- and IL-4-transfected B16-F10 cells with a novel oil-microemulsion adjuvant for B16-F10 whole cell tumour vaccine // Gene Ther. - 1996. - Vol., №10. - P. 853-858.
43. Санин A.B. и др., Особенности применения иммумодуляторов при паразитарных инвазиях // Ветеринария Кубани. - 2010. - №2. – С. 15-18.
44. Гетероциклические соединения, под ред. Р. Эладерфилда, пер. с англ., Т. 6, М., 1960, с. 160.
45. Келарев В.И., Караханов Р.А., Общая органическая химия, пер. с англ., т. 8, М., 1985, с. 160-84.
46. RU 2492527 от 10.09.2013.
47. RU 2345774 от 10.02.2009.
48. RU 2332998 от 10.09.2008.
49. RU 56185 от 10.09.2006.
50. RU 2011107380 от 27.08.2012.
51. Ашмарин И.П., Воробьев А.А. Статистические методы в микробиологических исследованиях. - Л.: Гос. Изд-во мед. Лит., 1962. -178 с.
52. Генес B.C. Некоторые простые методы кибернетической обработки данных диагностических и физиологических исследований. - М.: Наука, 1967. - 208 с.
53. Fan, Y. Cationic liposome-hyaluronic acid hybrid nanoparticles for intranasal vaccination with subunit antigens / Y. Fan [et al.] // J. Control Rel. - 2015. - Vol. 208. - P. 121-129.
54. Haughney, S.L. Effect of nanovaccine chemistry on humoral immune response kinetics and maturation / S.L. Haughney [et al.] // Nanoscale. - 2014. - Vol. 6. - P. 13770-13778.
55. Irvine D.J. Synthetic Nanoparticles for Vaccines and Immunotherapy / D.J. Irvine [et al.] // Chem. Rev. - 2015. - Vol. 115. - P. 11109-11146.
56. Sahdev, P. Biomaterials for nanoparticle vaccine delivery systems / P. Sahdey [et al.] // Pharm. Res. - 2014. - Vol. 31. - P. 2563-2582.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ЗАЩИТЫ ОТ КОРОНАВИРУСНОЙ ИНФЕКЦИИ | 2020 |
|
RU2747467C1 |
| СПОСОБ ИММУНОПРОФИЛАКТИКИ ЭКСПЕРИМЕНТАЛЬНОГО МЕЛИОИДОЗА ИНКАПСУЛИРОВАННЫМИ АНТИГЕНАМИ Burkholderia pseudomallei | 2008 |
|
RU2373955C1 |
| НОСИТЕЛЬ И АДЪЮВАНТ ДЛЯ АНТИГЕНОВ | 2006 |
|
RU2311926C2 |
| АДЪЮВАНТ НА ОСНОВЕ НИЗКОМОЛЕКУЛЯРНОГО ПЕПТИДОГЛИКАНА КЛЕТОЧНОЙ СТЕНКИ БАКТЕРИЙ | 2013 |
|
RU2563354C2 |
| СПОСОБ ПОЛУЧЕНИЯ ИНАКТИВИРОВАННОЙ ВАКЦИНЫ ДЛЯ ПРОФИЛАКТИКИ ВЕНЕСУЭЛЬСКОГО ЭНЦЕФАЛОМИЭЛИТА | 1987 |
|
RU2078816C1 |
| ВАКЦИНА ПРОТИВ БОЛЕЗНИ АУЕСКИ И РОЖИ СВИНЕЙ | 2009 |
|
RU2403062C1 |
| ИММУНОГЕННАЯ КОМПОЗИЦИЯ | 2005 |
|
RU2308289C2 |
| СПОСОБ ПОЛУЧЕНИЯ ТЕТРАВАЛЕНТНОЙ СУБЪЕДИНИЧНОЙ ПРОТИВОГРИППОЗНОЙ ВАКЦИНЫ | 2019 |
|
RU2740751C1 |
| ИММУНОГЕННЫЕ ВЕЩЕСТВА, СОДЕРЖАЩИЕ АДЪЮВАНТ НА ОСНОВЕ ПОЛИИНОЗИНОВОЙ КИСЛОТЫ-ПОЛИЦИТИДИЛОВОЙ КИСЛОТЫ | 2006 |
|
RU2462264C2 |
| Адъювант для противовирусных вакцин | 2017 |
|
RU2676266C2 |
Изобретение относится к области микробиологии, вирусологии и иммунологии. Предназначено для формирования невосприимчивости организма к инфекциям различной природы и повышения специфической активности иммунобиологических лекарственных препаратов. Повышение невосприимчивости организма к инфекциям различной природы и специфической активности иммунобиологических лекарственных препаратов (ИЛП) обеспечивается введением животным деривата фталгидразида (далее - адьювант Абидова) в разовой дозе 150 мкг/особь в качестве адъюванта (ИЛП). Использование изобретения позволяет повысить эффективность специфической профилактики инфекций независимо от природы ИЛП и тем самым усилия по борьбе с актуальными управляемыми инфекционными заболевания различной этиологии. 3 н. и 1 з.п. ф-лы, 6 табл., 6 пр., 2 ил.
1. Вакцина для иммунопрофилактики инфекционных заболеваний, содержащая в качестве адьюванта дериват синтетических производных фталгидразида (адьюванта Абидова), а также фармацевтически приемлемый носитель и/или разбавитель.
2. Применение деривата синтетических производных фталгидразида (адьюванта Абидова) в качестве адьюванта для повышения иммуногенности средств специфической иммунопрофилактики инфекционных заболеваний вне зависимости от их природы.
3. Способ повышения иммуногенности средств специфической иммунопрофилактики инфекционных заболеваний вне зависимости от их природы, включающий использование деривата синтетических производных фталгидразида (адьюванта Абидова):
путем введения его внутримышечно одновременно с антигеном или иммуногеном, но в раздельных шприцах; либо
путем введения его внутримышечно трехкратно: за трое суток, за двое суток, за сутки до введения антигена или иммуногена;
либо введение его вместе с антигеном или иммуногеном.
4. Способ по п. 3, где адьювант Абидова используют в разовой дозе 300 мкг/мл (150 мкг/особь) внутримышечно в шприце при раздельном с антигеном или иммуногеном введении; используют в дозе от 0,01 до 1000 мг/мл одновременно с введением антигена или иммуногена, при этом иммуногенность инактивированных и химических вакцин повышается не менее чем в 3 раза, иммуногенность живых вакцин повышается, не влияя на их безопасность по средствам снижения реактогенности, иммуногенность ассоциаций полианатоксинов повышается не менее чем на 30% для каждого из входящих в ассоциацию антигена (анатоксина).
| АДЪЮВАНТ НА ОСНОВЕ НИЗКОМОЛЕКУЛЯРНОГО ПЕПТИДОГЛИКАНА КЛЕТОЧНОЙ СТЕНКИ БАКТЕРИЙ | 2013 |
|
RU2563354C2 |
| АДЬЮВАНТ ДЛЯ БИОПРЕПАРАТОВ | 2002 |
|
RU2214278C1 |
| СПОСОБ ПОЛУЧЕНИЯ ВАКЦИННОЙ КОМПОЗИЦИИ, СОДЕРЖАЩЕЙ ПО МЕНЬШЕЙ МЕРЕ ОДИН АНТИГЕН И ПО МЕНЬШЕЙ МЕРЕ ОДИН АДЬЮВАНТ | 2008 |
|
RU2500421C2 |
| KR 101586468 B1, 18.01.2016 | |||
| EP 2929894 B1, 21.02.2018. | |||

