Изобретение относится к вирусологии, биотехнологии и иммунологии. Предложенное иммунобиологическое средство может применяться в качестве одного из компонентов для профилактики и лечения заболевания, вызванного вирусом гепатита С.
Далее приведены использованные в настоящей разработке сокращения и их расшифровка.
ДНК
дезоксирибонуклеиновая кислота
мМСК
генетически модифицированные МСК
ХГС
ЭТС
хронический гепатит С
эмбриональная телячья сыворотка
CTL
DC
Е1, Е2
ELISpot
IFN
IL
GFP
MDSC
цитотоксические лимфоциты CD8
дендритные клетки
поверхностные белки ВГС
метод иммуноферментных пятен
интерферон
интерлейкин
зеленый флуоресцентный белок
супрессорные клетки миелоидного происхождения
PBS
SD
0,1 М фосфатно-солевой буфер, рН 7,4
стандартное отклонение
TNF-α
VEGF
фактор некроза опухолей альфа
фактор роста эндотелия сосудов
Уровень техники
Гепатит С, вызываемый вирусом гепатита С (ВГС), признан одной из главных причин хронических заболеваний печени. В мире насчитывается более 71 млн инфицированных, ежегодно 3-4 млн заражаются, около 400 000 больных каждый год умирают от терминальных стадий хронического гепатита С (ХГС) - цирроза печени и гепатокарциномы. До 80% случаев острого гепатита С переходят в хроническую форму, что объясняется повышенной частотой мутаций ВГС, интерференцией вирусных белков с факторами врожденного и адаптивного иммунитета хозяина, образованием «эскейп»-вариантов ВГС, ускользающих от иммунного ответа [1]. Терапия гепатита С, основанная на сочетании пегилированного рекомбинантного IFN-α и рибавирина, позволяла элиминировать вирус не более чем у 50% пациентов [2]. Современная терапия с применением новых препаратов прямого противовирусного действия на основе ингибиторов ферментов ВГС - неструктурных белков NS3 и NS5B, а также блокаторов белка NS5A, позволяет добиться успеха в лечении большинства пациентов, но ее крайне высокая стоимость делает лечение малодоступным. Кроме того, недостаточно изучены отдаленные последствия блокирования вирусных ферментов, так как ВГС может оставаться в клетках печени после исчезновения вирусной РНК в периферической крови. Уже известно о некоторых побочных эффектах лечения этими препаратами, таких, как мутации ВГС, рецидивы и реинфекция [3, 4].
Отсутствие вакцины - главное препятствие в контроле за гепатитом С. Доказано, что эффективная вакцина должна стимулировать мощный мультиэпитопный и функциональный клеточный иммунный ответ к относительно консервативным неструктурным белкам NS3, NS4A, NS4B, NS5A, NS5B, а также индуцировать вируснейтрализующие антитела к вариабельным поверхностным белкам Е1 и Е2 [5].
В связи с отсутствием клеточных линий для эффективного размножения вируса in vitro основой для разработки вакцин служат генно-инженерные продукты, имитирующие последовательности ВГС: рекомбинантные белки, пептиды, представляющие В- и Т-клеточные эпитопы, ДНК в составе плазмид и вирусных векторов [6].
В настоящее время мезенхимальные стволовые клетки (МСК) успешно применяются в разных областях регенеративной медицины [7]. МСК - иммунопривилегированные клетки, т.е. их введение не приводит к воспалительному иммунному ответу. МСК могут быть получены почти из всех тканей, в том числе из костного мозга, жировой ткани, пуповины, дентальной пульпы. Клеточная терапия основана на способности МСК к направленной миграции к очагам патологии. Получение генетически модифицированных МСК (мМСК), экспрессирующих введенные гены, существенно расширяет возможности как клеточной, так и генной терапии, обеспечивая доставку терапевтических молекул к местам повреждения и воспаления [8, 9]. мМСК, содержащие вирусные гены, можно использовать для создания противовирусных вакцин. Такой подход имеет ряд преимуществ перед традиционными технологиями создания вакцин. мМСК потенциально способны экспрессировать одновременно десятки белков, тем самым формируя широкий спектр эпитопов с правильной посттрансляционной конформацией, имитируя естественную инфекцию, а также способны доставлять, экспрессировать и депонировать антиген в течение длительного времени. Таким образом, создание МСК-вакцин на основе генных и клеточных технологий может быть одним из наиболее перспективных достижений биотехнологии для иммунопрофилактики инфекционных заболеваний.
Проведенный анализ существующего уровня техники по научной и патентной информации в области разработки вакцины против ВГС показал, что на дату представления заявочных материалов наиболее широко апробированы рекомбинантные белки, синтетические пептиды, вирусоподобные частицы, ДНК вакцины и живые вирусные векторы. Однако оптимальный состав генов ВГС (или их участков) для включения в вакцину не определен. Неструктурные белки репликативного комплекса ВГС (NS2-NS5B) играют критическую роль в клеточном противовирусном иммунном ответе. Так, у спонтанно выздоровевших больных острым гепатитом С обнаружены Т-клетки фенотипов CD4+ (T-хелперные лимфоциты, Th) и CD8+ (цитотоксические лимфоциты, CTL), реагирующие на эпитопы белков NS3, NS4 и NS5, которые не выявляются у больных ХГС. К тому же, неструктурные белки, в отличие от вариабельных поверхностных белков, содержат высоко консервативные области, в которых локализован ряд Т-клеточных эпитопов [10 - 12]. Таким образом, высокий ВГС-специфический клеточный ответ к CD4+ и CD8+ Т-клеточным эпитопам неструктурных белков может ограничить инфекцию, элиминировать ВГС-инфицированные клетки и предотвратить хронизацию острого гепатита. В связи с этим гены неструктурных белков, по нашему мнению, являются наиболее перспективными кандидатами для включения в вакцину. Известно, что неструктурный белок NS5A - мультифункциональный фосфопротеин. Он играет важную роль в вирусном патогенезе и в индукции иммунных реакций, а также выполняет ряд важных функций в жизненном цикле вируса. Этот белок может быть активатором транскрипции и участвует во многих клеточных регуляторных процессах. NS5A, кроме непосредственного участия в репликации вируса, играет ключевую роль на ранних стадиях сборки и созревания вирусных частиц [13, 14]. NS5A участвует в модуляции ответа на лечение интерфероном (IFN); в частности, устойчивость ВГС к IFN-α коррелирует с числом мутаций в NS5A [15]. Ряд современных пангенотипных препаратов прямого противовирусного действия (Ledipasvir, Pibrentasvir, Velpatasvir и др.) нацелены на блокирование функций NS5A, и ведется активный поиск новых ингибиторов этого белка [16].
Несмотря на положительные результаты, полученные с вакцинами, основанными на белковых субъединицах ВГС, индуцированный ими иммунный ответ является, главным образом, гуморальным, кратковременным и изолят-специфичным, который не защищает от инфицирования вирусами другого генотипа.
Вакцины на основе ДНК-конструкций могут стимулировать клеточный иммунный ответ, нетоксичны, однако для увеличения клеточного ответа необходимо применение высокоэффективных адъювантов. ДНК-иммунизация практически не индуцирует гуморальный ответ. Проблема состоит также в способе введения.
Известно решение [17], в котором в качестве вакцины против ВГС предлагается полинуклеотид, кодирующий полипептидные последовательности либо индивидуальных белков ВГС core, NS3, NS4B и NS5B, либо их сочетание в одной открытой рамке считывания. Предлагается способ вакцинации, при котором полинуклеотид наносят на золотые гранулы и вводят подкожно.
Известно изобретение [18], в котором для генерирования ВГС-специфического клеточного иммунного ответа используется ДНК, кодирующая полипротеин NS3-NS4A-NS4B-NS5A-NS5B ВГС, встроенная в аденовирусный вектор Ad6. Вакцины на основе вирусных векторов повышают иммуногенность кодируемых последовательностей ВГС, однако могут быть потенциально канцерогенными, а при использовании аденовирусов предсуществующий иммунный ответ может снижать иммунитет к ВГС.
Ни одна из кандидатных вакцин пока не способна обеспечить полноценный профилактический и терапевтический эффекты против ВГС [19].
Проведенный анализ выявленного уровня техники по научной и патентной информации в области применения МСК показал, что обнаруженная патентная документация описывает терапевтические свойства МСК человека.
Из исследованного уровня техники выявлено изобретение, в котором предлагается препарат для лечения метахроматической лейкодистрофии, состоящий из МСК человека, генетически модифицированных рекомбинантным аденоассоциированным вирусом 9 серотипа, содержащим кодон-оптимизированную последовательность гена ARSA, представленную в SEQ ID NO: 1 (патент RU 2769577 [20]).
Проведенный анализ выявленного уровня техники по научной и патентной информации в области разработки иммунобиологического средства на основе мМСК человека, экспрессирующих неструктурный белок ВГС, для повышения клеточного ответа против ВГС показал, что сведения о получении и использовании мМСК в качестве профилактических и терапевтических вакцин против инфекционных заболеваний единичны. В настоящее время известна одна работа, в которой получены мМСК человека, экспрессирующие гены вируса SARS-CoV-2, и показана индукция вирусспецифических антител у мышей, иммунизированных мМСК. Авторы сообщили, что подали заявку на патент PCT, относящийся к МСК-вакцине против COVID-19 [21].
Известна возможность использования мМСК мыши в качестве инновационных вакцин, усиливающих иммунные реакции против ВИЧ [22], против гепатита С [23, 24] и против вируса простого герпеса 1 типа [25], которые можно принять за аналоги. Результаты, опубликованные в 2020 г., выбраны авторами настоящего изобретения в качестве наиболее близкого аналога заявляемому составу иммунобиологического средства и способу его использования [24]. В данном исследовании МСК для иммунизации мышей получали из костного мозга сингенных мышей, МСК предварительно трансфицировали плазмидой, кодирующей комплекс неструктурных белков NS3-NS4A-NS4B-NS5A-NS5B. Показано существенное повышение иммунного ответа на мМСК по сравнению с плазмидой, несущей те же гены. Недостатком прототипа является то, что мМСК были получены на основе мышиных МСК, трансфицированных вирусными генами. Однако положительные результаты, полученные с мышиными МСК, при переходе к экспериментам с МСК человека не всегда подтверждаются, что может быть вызвано различиями в иммунных реакциях [26].
Раскрытие настоящего изобретения
Технической задачей данного изобретения является разработка иммунобиологического средства для повышения клеточного ответа против ВГС на основе мМСК человека, экспрессирующих неструктурный белок ВГС.
Технический результат заключается в повышении клеточного иммунного ответа, характеризующегося усилением антиген-специфической пролиферации лимфоцитов, продукции интерферона-гамма и формированием Т-клеток памяти.
Указанный технический результат достигается за счет того, что создано иммунобиологическое средство для повышения клеточного ответа против вируса гепатита С, состоящее из генетически модифицированных мезенхимальных стволовых клеток человека, экспрессирующих неструктурный белок NS5A вируса гепатита С.
Заявленное изобретение можно применять как в качестве профилактического средства, нацеленного на предотвращение перехода острого гепатита С в хронический за счет быстрой активации клеток памяти, индуцированных вакциной, так и в качестве терапевтического средства, используемого совместно с противовирусными лечебными препаратами, для усиления адаптивного иммунитета к ВГС. В заявленном изобретении могут быть использованы МСК, полученные самостоятельно по любому известному из уровня техники способу (например, из липоаспирата, хирургических биопсий и др.), так и в виде коммерческих препаратов МСК, имеющих сертификат качества.
Краткое описание чертежей
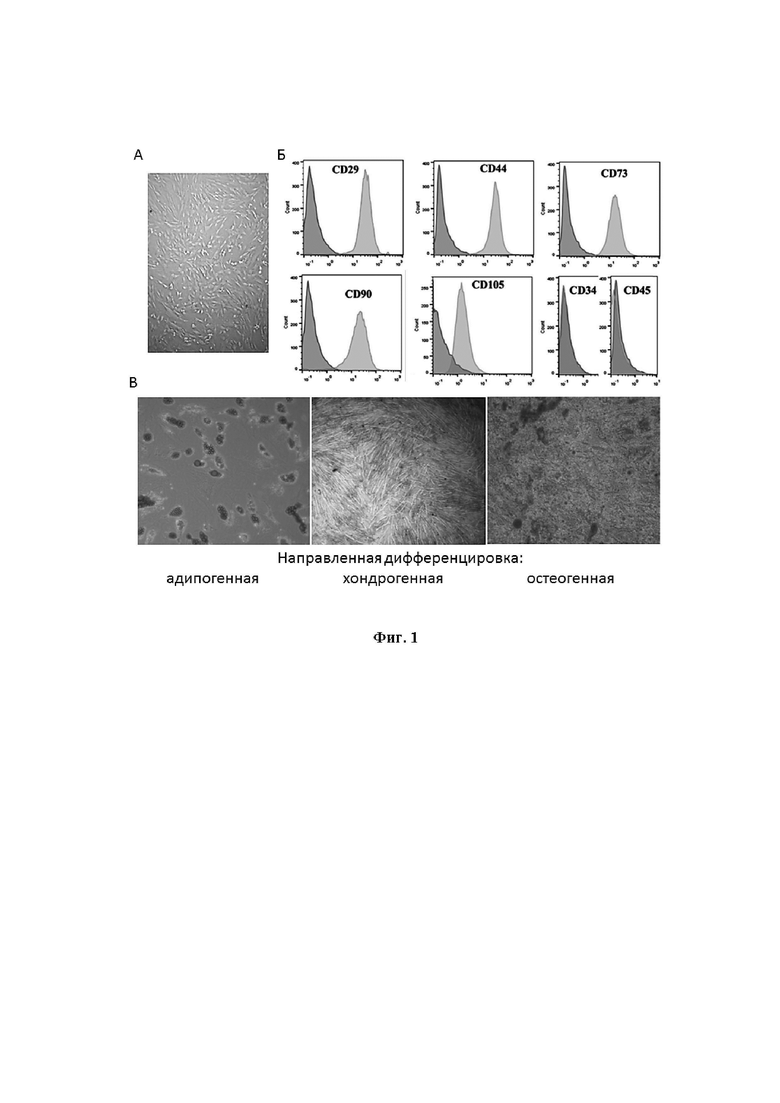
На Фиг. 1 приведена характеристика МСК, полученных из дентальной пульпы. На фиг. 1А - Морфология первичной культуры МСК, световая микроскопия, увеличение 200 X; фиг 1Б - Фенотипирование поверхностных рецепторов МСК методом проточной цитометрии. По вертикали - количество клеток, по горизонтали - относительная интенсивность флуоресценции. Светло-серые гистограммы - распределение МСК, окрашенных указанными антителами, темно-серые - контрольными изотипическими антителами; фиг. 1В - Морфология МСК и характерная окраска клеток после направленной дифференцировки, трансмиссионная микроскопия, увеличение 200 X.
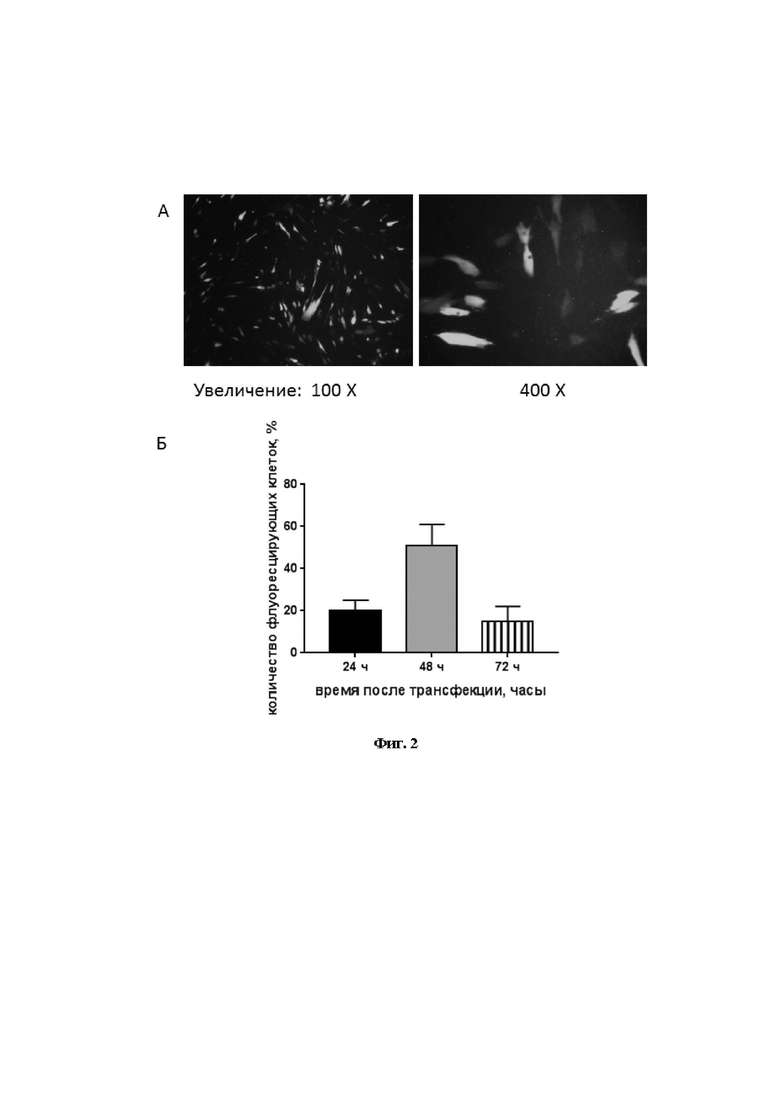
На Фиг. 2 продемонстрирована визуализация флуоресцирующих клеток при трансфекции МСК химерной плазмидой pcNS5A-GFP. На фиг. 2А - флуоресценция через 48 ч после трансфекции; на фиг 2Б - динамика экспрессии гена NS5A-GFP.
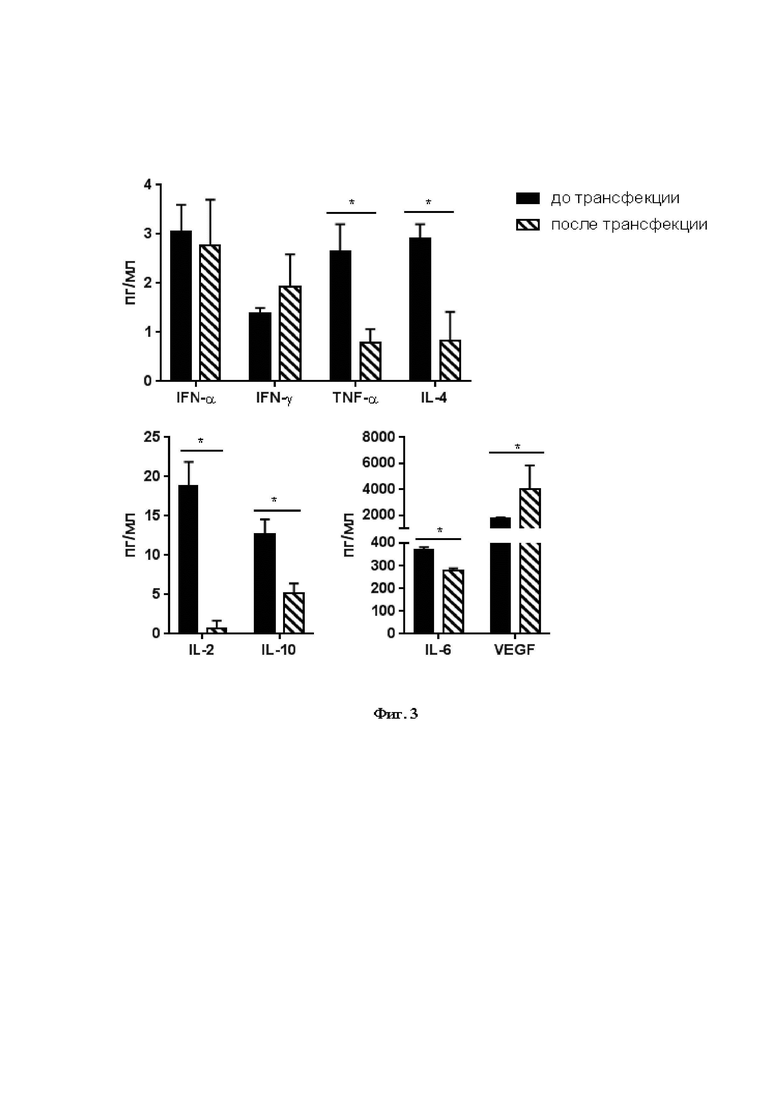
На Фиг. 3 показаны изменения концентрации цитокинов, секретируемых трансфицированными мезенхимальными клетками, выделенными из дентальной пульпы. Данные представлены как средние + стандартное отклонение (SD); * p<0,05.
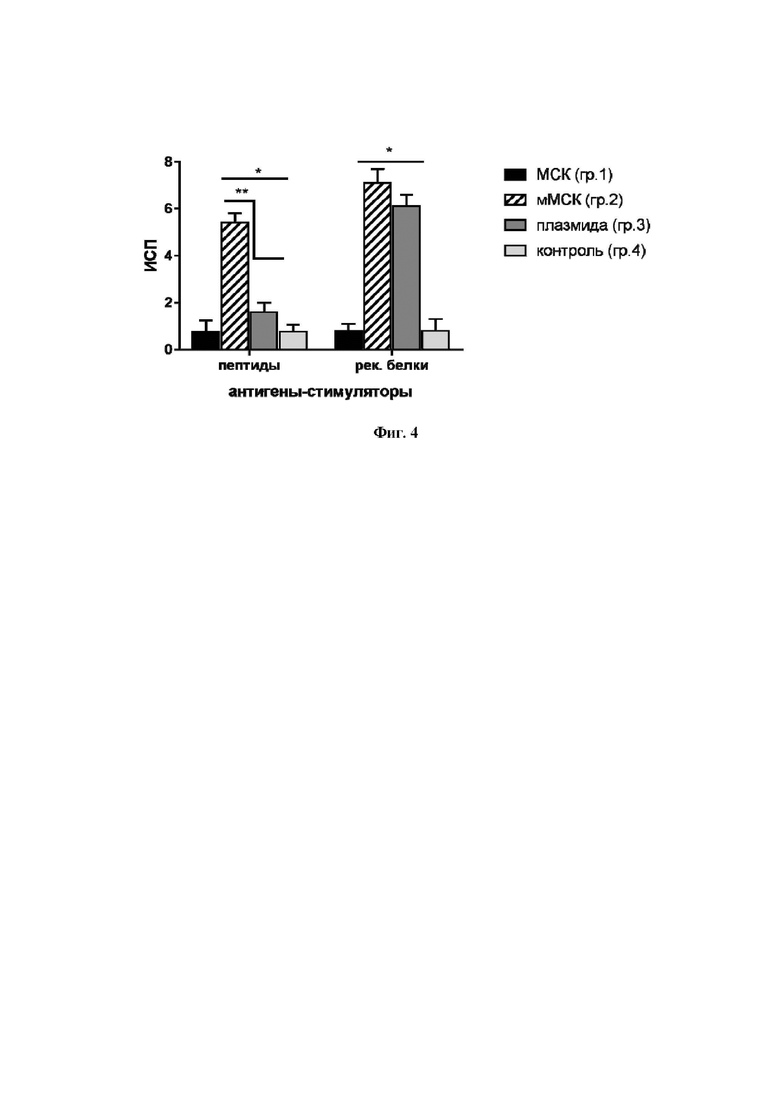
На Фиг. 4 приведена пролиферативная активность лимфоцитов иммунизированных мышей в ответ на стимуляцию антигенами in vitro. Данные представлены как средние + стандартное отклонение (SD); * p < 0,05 по сравнению с контрольной группой; ** p < 0,05 по сравнению с указанными группами.
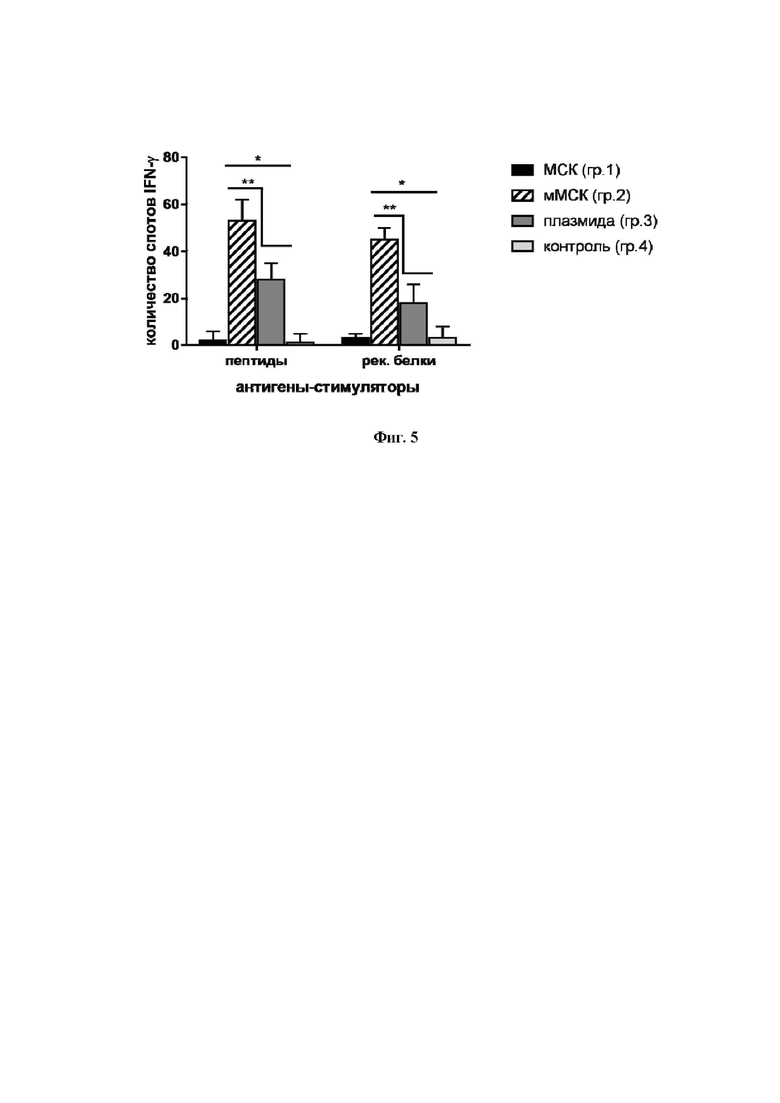
На Фиг. 5 показана сравнительная частота индукции лимфоцитами IFN-γ-синтезирующих клеток в ответ на антигены из состава белка NS5A, определенная методом ELISpot. Данные представлены как средние + стандартное отклонение (SD); * p < 0,05 по сравнению с контрольной группой; ** p < 0,05 по сравнению с указанными группами.
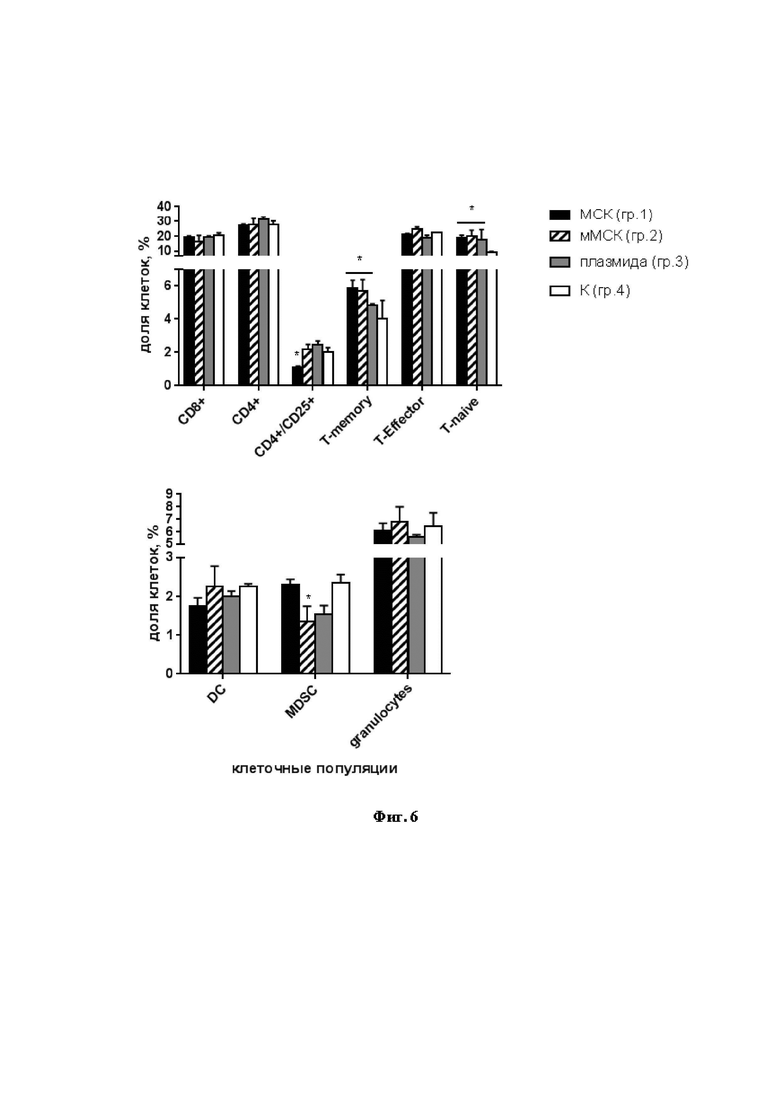
На Фиг. 6 продемонстрированы изменения относительного содержания клеточных популяций в селезенках мышей, иммунизированных МСК, мМСК и плазмидой. Данные представлены как средние + стандартное отклонение (SD); * p < 0,05 по сравнению с контрольной группой.
Осуществление изобретения
Поставленная цель и заявленный технический результат достигаются путем трехкратного внутривенного введения иммунобиологического средства, представляющего собой мМСК человека, экспрессирующие неструктурный белок NS5A ВГС.
Практическая реализация заявленного изобретения осуществляется в 3 этапа в нижеприведённой последовательности, а именно:
1 этап: получение МСК человека;
2 этап: получение плазмидной конструкции; трансфекция МСК человека плазмидой и получение мМСК, экспрессирующих неструктурный белок ВГС;
3 этап: анализ эффективности заявленного способа увеличения ВГС-специфического клеточного иммунного ответа на лабораторных мышах.
Далее приведены примеры каждого из этапов осуществления заявленного технического решения.
Пример 1. Получение и характеристика МСК из дентальной пульпы
Первичные клетки дентальной пульпы получены из содержимого пульповой камеры удаленных по ортодонтическим показаниям здоровых зубов. Вскрытую в стерильных условиях пульповую камеру промывали в клеточной культуральной среде DMEM/F-12 (Gibco, США) с двухкратным раствором антибиотика-антимикотика (Gibco, США). Дентальную пульпу извлекали тонким шпателем, измельчали, инкубировали в растворе коллагеназы I (Gibco, США) в концентрации 1,0 мг/мл в течение 3 ч при 37°С, затем центрифугировали (400g). Выделенные клетки культивировали в среде DMEM/F-12 с 10% ЭТС (Gibco, США), 100,0 Ед/мл пенициллина и 100,0 мкг/мл стрептомицина (ПанЭко, Россия), при достижении 90% конфлюэнтности клетки пассировали в соотношении 1:3.
Полученные МСК характеризовали по морфологическим свойствам, по экспрессии поверхностных рецепторов и способности к направленной дифференцировке. Как показано на Фиг. 1А, клетки были адгезионные и имели характерную для МСК веретенообразную форму.
С помощью проточной цитофлюориметрии в культуре МСК исследовали экспрессию маркеров МСК: CD29, CD44, CD73, CD90 и CD105 и отсутствие экспрессии маркеров CD34 и CD45. Первичные антитела, меченные флуорохромами FITC или PE, получены из Miltenyi Biotec (Германия). В этом эксперименте показано, что полученные МСК экспрессировали базовые маркеры, характерные для МСК, и не экспрессировали маркеры гемопоэтических клеток (Фиг. 1Б).
Культуру МСК (5 пассаж) дифференцировали в остеогенном, адипогенном и хондрогенном направлении с помощью остеогенной среды (среда Dulbecco's modified Eagle's medium (DMEM) с добавлением 10−8 M дексаметазона (Sigma, США), 10,0 мМ b-глицерофосфата (Sigma), и 50,0 мкг/мл аскорбиновой кислоты), адипогенной среды (DMEM с 10,0 мМ 3-изобутил-1-метилксантин (Sigma), 0,1 мМ индометацин (Sigma), 10,0 мкг/мл инсулин (Sigma), 10−6 М дексаметазон) и хондрогенной среды (Stempro, Invitrogen, Life Technologies Ltd Paisley, Великобритания), соответственно. Для проявления остеогенной, адипогенной и хондрогенной дифференцировки культуры МСК, выдержанные в течение недели в дифференцировочных средах, окрашивали с помощью Alizarin red (Sigma), Oil Red O (Sigma), и Alcian blue (Sigma), соответственно, с последующей микроскопией в просвечивающем режиме на микроскопе Nikon Eclipse Ci microscope (Nikon Instruments Inc.). Как показано на Фиг. 1В, после воздействия вышеописанных реагентов и процедур первичные МСК дифференцировались в трех основных направлениях: остеогенном, адипогенном и хондрогенном.
Таким образом, в Примере 1 показано, что по адгезивной способности, поверхностным маркерам и дифференцировочному потенциалу полученные клетки соответствуют критериям определения МСК Международного общества клеточной терапии (The International Society for Cellular Therapy) [27].
Пример 2. Характеристика плазмиды и получение мМСК
В качестве кандидатной ДНК-вакцины использовали химерную плазмиду pcNS5A-GFP, кодирующую полноразмерный неструктурный белок NS5A ВГС (1973-2419 а.о.) в комплексе с зеленым флуоресцирующим белком GFP, как описано ранее [28]. Образование химерного комплекса GFP-NS5A позволяет визуально оценивать число трансфицированных клеток с помощью флуоресцентного микроскопа, не используя иммуноцитохимическое окрашивание продуктов экспрессии генов ВГС. Плазмиду конструировали на основе вектора pcDNA-3.1(+) (Invitrogen, США). Плазмидную ДНК получали, трансформируя компетентные клетки Escherichia coli (штамм XL-1 blue). Для препаративного выделения и очистки плазмид из культуры бактерий использовали щелочной метод, для аналитического - набор Qiagen Inc. (США) в соответствии с инструкцией. Концентрацию очищенной ДНК измеряли на спектрофотометре при длине волны 260/280 нм.
Генетически модифицированные клетки человека получали путем трансфекции. За 24 ч до трансфекции вносили 1,5×105 МСК в 24-луночные культуральные панели или 2,5×106 клеток в культуральные чашки Петри с ростовой площадью 60,1 см2, после достижения субконфлюэнтного монослоя (около 80%) на МСК наносили комплексы плазмиды pcNS5A-GFP со сливающим агентом GenJect™-39 (Molecta, Россия) в соотношении 1:2 (5,0 мкг плазмиды плюс 10,0 мкл сливающего агента). Через 18 ч для снижения токсичности данный комплекс удаляли путем смены среды. Все манипуляции с трансфекцией МСК выполняли стерильно в полной ростовой среде в отсутствии антибиотиков. Анализ трансфицированных клеток проводили с помощью инвертированного флуоресцентного микроскопа Primovert A1. Zeiss с камерой AxioCam MRc5 (Германия). Эффективность трансфекции оценивали по доли флуоресцирующих клеток в общем количестве клеток (в %), учитывая не менее семи полей зрения.
На Фиг. 2А показан пример визуализации трансфицированных клеток с помощью флуоресценции GFP. Изучение динамики экспрессии гена NS5A-GFP показало, что наибольшая эффективность трансфекции МСК наблюдалась через 48 ч (Фиг. 2Б).
Таким образом, в этом эксперименте показано, что белок NS5A ВГС экспрессируется в МСК, трансфицированных плазмидой pcNS5A-GFP, при этом эффективность трансфекции (около 50%) максимальна через 48 ч после трансфекции.
Пример 3. Секреция цитокинов нативными МСК и мМСК
Известно, что МСК - источник целого спектра цитокинов и ростовых факторов, играющих регуляторную роль в гемопоэзе, пролиферации, апоптозе, ангиогенезе, иммунном ответе и других важных физиологических процессах. Концентрации восьми цитокинов, секретируемых МСК, определяли в культуральных жидкостях, отобранных:
а) при культивировании МСК,
б) после трансфекции клеток.
Для этого эксперимента использовали ИФА-наборы производства Вектор-Бест (г. Новосибирск, Россия) для следующих цитокинов: IFN-α, IFN-γ, TNF-α, IL-2, IL-4, IL-6, IL-10 и VEGF (фактор роста эндотелия сосудов). Концентрацию цитокинов определяли по калибровочным кривым стандартных образцов. Обнаружено, что экспрессия NS5A ВГС в результате трансфекции МСК оказала существенное влияние на профиль секретируемых цитокинов. Через 24 - 48 ч после трансфекции плазмидой pcNS5A-GFP МСК, выделенных из дентальной пульпы, концентрации шести цитокинов изменялись. Статистически значимо снижалась секреция IL-2, IL-6, TNF-α, IL-4 и IL-10, повышалась - VEGF (Фиг. 3). Данные изменения принципиальны: показано, что МСК могут поляризоваться в фенотип МСК 2, который супрессирует иммунный ответ, или в фенотип МСК 1, который индуцирует иммунный ответ. Направленность иммуномодуляции зависит от концентрации про- и противовоспалительных цитокинов и других растворимых факторов в среде. Показано, что высокая концентрация ряда цитокинов, в первую очередь, IFN-γ и TNF-α, связана с супрессией иммунного ответа [29]. Поэтому снижение уровня TNF-α, IL-2, IL-4, IL-6 и IL-10, секретируемых мМСК, способствует активации различных звеньев врожденного и адаптивного иммунного ответа.
Таким образом, данный эксперимент показывает, что снижение секреции мМСК провоспалительных цитокинов TNF-α, IL-2, IL-6 и противовоспалительных IL-4 и IL-10 - один из факторов, направляющих иммунный ответ в сторону активации.
Пример 4. Оценка эффективности иммунного ответа мышей на введение мМСК
Для экспериментов in vivo использовали самок мышей инбредной линии C57BL/6 (Н-2b) в возрасте 6-8 недель (средний вес 20±3 г), полученных из питомника лабораторных животных «Столбовая». Было сформировано 4 группы мышей по 6 - 8 животных в группе:
группу 1 иммунизировали нативными (немодифицированными) МСК (МСК) (контроль 1);
группу 2 иммунизировали мМСК, трансфицированными плазмидой pcNS5A-GFP;
группу 3 иммунизировали плазмидой pcNS5A-GFP;
группу 4 иммунизировали ф/р (контроль 2).
Клетки вводили в/в в дозе 5,0×105/мышь, плазмиду - 100,0 мкг/мышь в/м в четырехглавую мышцу бедра задних лап. Иммунизации проводили трехкратно с интервалом 2 недели, через 8 сут после последней инъекции учитывали иммунный ответ.
При проведении экспериментов in vivo не зарегистрировано ни падежа иммунизированных мышей, ни потери ими массы тела. Поведенческие реакции у животных контрольных и опытных групп не различались, что, по нашему мнению, свидетельствует о безопасности введенных препаратов.
Для оценки гуморального и клеточного ответа использовали рекомбинантные белки и пептиды, имитирующие последовательности белка NS5A ВГС (Таблица 1).
Синтезированы в Институте биоорганической химии РАН.
Гуморальный иммунный ответ на белки NS5A определяли в твердофазном иммуноферментном анализе (ИФА), в лунки сорбировали рекомбинантные белки NS5A ВГС в концентрации 1,0 мкг/мл. В сыворотках крови мышей контрольных групп 1 и 4 антитела к NS5A отсутствовали (титр < 1:10). В группах 2 и 3 титр антител к NS5A составлял 1:10 - 1:20.
Важной характеристикой клеточного иммунного ответа служит пролиферация лимфоцитов в ответ на рестимуляцию in vitro антигенами из состава белка NS5A. Анализ пролиферации лимфоцитов иммунизированных мышей проводили in vitro по включению в лимфоциты [3H]-тимидина. Для получения спленоцитов селезенки диспергировали, клеточные суспензии процеживали через ситечки с размером пор 100 мкм (BD Falcon Cell srtainers), дважды отмывали в среде RPMI-1640 (ПАНЭКО, Россия). Спленоциты помещали в 96-луночные культуральные панели по 5×106 клеток в лунку в среде RPMI-1640, содержащую 20% ЭТС (Invitrogen, США), 2,0 мМ глутамина, 4,5 г/л глюкозы, 50,0 мкг/мл гентамицина, 0,2 ед/мл инсулина. К клеткам добавляли стимуляторы - смесь трех рекомбинантных белков NS5A и смесь пептидов в конечной концентрации 1,0 мкг/мл каждого компонента и культивировали при 37°С в атмосфере 5% СО2 в течение 4 сут, затем добавляли свежую среду с [3H]-тимидином (1,0 мкКи/лунка) (получен из ФГБУ Институт молекулярной генетики НИЦ «Курчатовский институт» РАН). Через 18 ч радиоактивность в клетках измеряли с помощью β-счетчика MicroBeta2 (PerkinElmer, США). Рассчитывали индекс стимуляции пролиферации (ИСП) как отношение радиоактивности (в имп/мин) в лунках со стимуляторами к радиоактивности в лунках со средой.
Выявлено, что ИСП в ответ на антигены ВГС в группах 2 и 3 статистически значимо превышали значения ИСП в контрольных группах 1 и 4, при этом ИСП в ответ на пептиды был более чем в 3 раза больше в группе мышей, иммунизированных мМСК, по сравнению с группой, которой вводили плазмиду (Фиг. 4).
Таким образом, в данном эксперименте показано, что иммунизация мышей мМСК, экспрессирующими ген NS5A, усиливает антиген-специфическую пролиферацию спленоцитов in vitro.
Пример 5. Иммунизация мМСК повышает количество IFN-γ-синтезирующих клеток
Наиболее информативный метод, рекомендуемый для валидации Т-клеточного ответа к новым вакцинам против ВГС - определение количества IFN-γ-синтезирующих клеток методом ELISpot [30]. Данный тест проводили c помощью набора Mouse IFN-γ ELISpotPLUS kit (HRP) (Mabtech, Швеция). Изолированные спленоциты (4х105 клеток в лунке) инкубировали с иммобилизованными на 96-луночных планшетах антителами к IFN-γ мыши течение 2,5 сут при 37°С в атмосфере 5% СО2 в присутствии стимуляторов - смесей рекомбинантных белков NS5A и пептидов. Окрашивание клеток проводили в соответствии с инструкцией производителя. Окрашенные пятна (spots, споты) детектировали визуально с помощью стереоскопического микроскопа МБС-10 (ОАО «ЛЗОС», Россия). Результаты выражали как разницу в количестве спотов на 106 клеток в лунках в присутствии стимуляторов и в контрольных лунках со средой.
На Фиг. 5 показано, что число спотов в группах 2 и 3 превышало таковое в контрольных группах 1 и 4, соответственно. При сравнении групп 2 и 3 между собой установлено, что спотов образовалось в 2 - 2,5 раза больше в группе 2.
Таким образом, в данном опыте показано, что иммунизация мышей мМСК, экспрессирующими ген NS5A, усиливает образование клеток, синтезирующих IFN-γ, в ответ на стимуляцию пептидами и рекомбинантными белками из состава последовательностей белка NS5A. Это свидетельствует об активации антивирусного ответа. Важно, что клеточный ответ на мМСК (группа 2) превосходил ответ на плазмиду (группа 3) несмотря на то, что доза введенной плазмиды в 100 раз превышала дозу той же плазмиды, трансфицированной в МСК (100,0 мкг и 1,0 мкг, соответственно).
Пример 6. Анализ клеточных популяций в селезенках иммунизированных животных
Анализ клеточных популяций в селезенках мышей, иммунизированных МСК, мМСК и плазмидой, проводили методом проточной цитометрии. Использовали меченные флуорохромами антитела к кластерам дифференцировки: CD4, CD8, CD25, CD11b, CD11c и Gr1 (BD Biosciences, США). Содержание различных популяций Т-клеток - наивных, эффекторных и памяти, определяли с помощью набора Mouse Naïve/Memory T Cell Panel (BD Pharmingen, BD Biosciences, США). Характеристика антител и определяемых клеточных популяций представлена в таблице 2. Окрашивание клеток (106 клеток/пробу) проводили по стандартной методике, рекомендованной производителем. Измерения выполняли на проточном цитофлуориметре BD FACSCanto™ II (США), результаты анализировали с помощью программы BD FACSDiva v.6.1.3 (BD Biosciences, США).
CD11b - APC
CD4 - PerCP-Cy5.5
CD62L - APC
CD44 - PE
Как показано на Фиг. 6, не было обнаружено различий в относительном количестве лимфоцитов CD8+ (CTL) и CD4+ (Th), однако содержание некоторых субпопуляций CD4+ отличалось. Так, в группе 1 (МСК) статистически значимо снижалась доля активированных лимфоцитов CD4+/CD25+ по сравнению с контролем и другими группами. В селезенках мышей, получивших МСК и мМСК, статистически значимо увеличивалось относительное количество Т-клеток памяти (T-memory). Содержание наивных Т-клеток (T-naïve) самым низким было в контрольной группе, тогда как эффекторных Т-клеток (T-effector) - сходным во всех группах.
Что касается клеток миелоидного ряда, то содержание дендритных клеток (DC), гранулоцитов и макрофагов было сходным (Фиг. 6). Важно, что в группе 2 (мМСК) статистически значимо снижалось количество супрессорных клеток миелоидного происхождения (MDSC). MDSC - гетерогенная популяция незрелых миелоидных клеток с мощным супрессорным потенциалом. Роль MDSC в вирусных инфекциях изучена недостаточно. У пациентов, инфицированных ВГС, наблюдается повышение популяции MDSC; эти клетки подавляют пролиферацию CD4+ и CD8+ лимфоцитов, NK-клеток и продукцию ИФН-γ [31, 32]. Стимуляции адаптивного иммунного ответа на модифицированные МСК, показанная в примерах 4 и 5, может быть результатом подавления MDSC.
Таким образом, пример 6 демонстрирует, что мМСК, экспрессирующие белок NS5A ВГС, стимулируют формирование CD4+ Т-клеток памяти и снижение содержания клеток-супрессоров MDSC, что способствует эффективному клеточному ответу против ВГС.
Библиография:
1. Dustin LB. Innate and Adaptive Immune Responses in Chronic HCV Infection. Curr Drug Targets. 2017;18(7):826-843. doi: 10.2174/1389450116666150825110532.
2. Pawlotsky JM. Treatment of hepatitis C: how will we use viral kinetics, response-guided therapy? Curr Gastroenterol Rep. 2013 Feb;15(2):309. doi: 10.1007/s11894-012-0309-x.
3. Wang Y., Rao H., Chi X. et al. Detection of residual HCV-RNA in patients who have achieved sustained virological response is associated with persistent histological abnormality. EBioMedicine 2019, 46, 227-235.
4. Huang C.F., Yu M.L. Unmet needs of chronic Hepatitis C in the era of direct-acting antiviral therapy. Clin. Mol. Hepatol. 2020, 26, 251-260.
5. Todryk S.M., Bassendine M.F., Bridge S.H. Revisiting the elusive Hepatitis C vaccine. Vaccines 2021, 9, 114.
6. Andrianov A.K., Fuerst T.R. Immunopotentiating and Delivery Systems for HCV Vaccines. Viruses 2021, 13, 981.
7. Yuan X., Logan T.M., Ma T. Metabolism in human mesenchymal stromal cells: A missing link between hMSC biomanufacturing and therapy? Front. Immunol. 2019, 10, 977.
8. Cobo M.; Anderson, P.; Benabdellah, K. et al. Mesenchymal stem cells expressing vasoactive intestinal peptide ameliorate symptoms in a model of chronic multiple sclerosis. Cell Transp. 2013, 22, 839-854.
9. Wyse R.D.; Dunbar G.L.; Rossignol J. Use of genetically modified mesenchymal stem cells to treat neurodegenerative diseases. Int. J. Mol. Sci. 2014, 15, 1719-1745.
10. Neumann-Haefelin C, Thimme R. Success and failure of virus-specific T cell responses in hepatitis C virus infection. Dig Dis. 2011;29(4):416-22. doi: 10.1159/000329807.
11. Folgori A, Spada E, Pezzanera M et al. Acute Hepatitis C Italian Study Group. Early impairment of hepatitis C virus specific T cell proliferation during acute infection leads to failure of viral clearance. Gut. 2006 Jul;55(7):1012-9. doi: 10.1136/gut.2005.080077.
12. Chigbu DI, Loonawat R, Sehgal M et al. Hepatitis C Virus Infection: Host-Virus Interaction and Mechanisms of Viral Persistence. Cells. 2019 Apr 25;8(4):376. doi: 10.3390/cells8040376.
13. Kandangwa M, Liu Q. HCV NS5A hyperphosphorylation is involved in viral translation modulation. Biochem Biophys Res Commun. 2019 Nov 26;520(1):192-197. doi: 10.1016/j.bbrc.2019.09.105.
14. Shanmugam S, Nichols AK, Saravanabalaji D et al. HCV NS5A dimer interface residues regulate HCV replication by controlling its self-interaction, hyperphosphorylation, subcellular localization and interaction with cyclophilin A. PLoS Pathog. 2018 Jul 23;14(7):e1007177. doi: 10.1371/journal.ppat.1007177.
15. Muñoz de Rueda P, Fuentes Rodríguez JM, Quiles Pérez R et al. Hepatitis C virus NS5A region mutation in chronic hepatitis C genotype 1 patients who are non-responders to two or more treatments and its relationship with response to a new treatment. World J Gastroenterol. 2017 Jul 7;23(25):4538-4547. doi: 10.3748/wjg.v23.i25.4538.
16. Hamdy J, Emadeldin N, Hamed MM et al. Design and Synthesis of Novel Bis-Imidazolyl Phenyl Butadiyne Derivatives as HCV NS5A Inhibitors. Pharmaceuticals (Basel). 2022 May 20;15(5):632. doi: 10.3390/ph15050632.
17. Brett S., Hamblin P. A., Ogilvie L. Vaccine against HCV. Патент WO 2004046176, 13.11.2003.
18. Armin L., Stefano C., Shiver J.W. et al. Hepatitis C virus vaccine, CN 101988071 (A), 23.03.2011.
19. Bailey, J.R.; Barnes, E.; Cox, A.L. Approaches, Progress, and Challenges to Hepatitis C Vaccine Development. Gastroenterology 2019, 156, 418-430.
20. Муллагулова А.И., Шаймарданова А.А., Чулпанова Д.С. и др. Препарат для лечения метахроматической лейкодистрофии и способ ее лечения. Патент RU 2769577 (заявка № 2021115696, 01.06.2001).
21. Junhua Liu, Huping Jiao, Xiushan Yin. Engineered human mesenchymal stem cells as new vaccine platform for COVID-19. bioRxiv preprint doi: https://doi.org/10.1101/2020.06.20.163030.
22. Tomchuck S.L., Norton E.B., Garry R.F. et al. (2012) Mesenchymal stem cells as a novel vaccine platform. Front. Cell. Infect. Microbiol. 2, 140.
23. Masoudi MR, Rafati A. Immunogenicity against hepatitis C virus with mesenchymal stem cells of inbreed BALB/c mice sub cloned with HCVcp protein gene. Transpl Immunol. 2022 Oct; 74:101651. doi: 10.1016/j.trim.2022.101651.
24. Masalova OV, Lesnova EI, Klimova RR et al. Genetically Modified Mouse Mesenchymal Stem Cells Expressing Non-Structural Proteins of Hepatitis C Virus Induce Effective Immune Response. Vaccines (Basel). 2020 Feb 2;8(1):62. doi: 10.3390/vaccines8010062.
25. Климова Р.Р., Демидова Н.А., Масалова О.В., Кущ А.А. Профилактическая вакцинация мезенхимальными стволовыми клетками защищает мышей от летальной инфекции, вызванной вирусом простого герпеса 1 типа. Молекулярная биология, 2021, том 55, № 3, с. 478-490.
26. Galipeau J, Sensébé L. Mesenchymal stromal cells: clinical challenges and therapeutic opportunities. Cell Stem Cell. 2018; 22(6): 824-833. doi: 10.1016/j.stem.2018.05.004.
27. Dominici M., Le Blanc K., Mueller I. et al. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy. 2006. 8(4), 315-317.
28. Масалова О.В., Леснова Е.И., Грабовецкий В.В. и др. ДНК-иммунизация плазмидой, содержащей ген белка NS5A вируса гепатита C, индуцирует эффективный клеточный иммунный ответ. Молекулярная биология, 2010, т. 44, № 2, с. 275-283.
23. Cagliani J, Grande D, Molmenti EP et al. Immunomodulation by Mesenchymal Stromal Cells and Their Clinical Applications. J Stem Cell Regen Biol. 2017;3(2):10.15436/2471-0598.17.022. doi: 10.15436/2471-0598.17.022.
30. Ahlen G., Frelin L. Methods to Evaluate Novel Hepatitis C Virus Vaccines. Methods Mol. Biol. 2016, 1403, 221-244.
31. Cai W., Qin A., Guo P. et al. Clinical significance and functional studies of myeloid-derived suppressor cells in chronic hepatitis C patients. J. Clin. Immunol. 2013, 33, 798-808.
32. Goh C.C., Roggerson K.M., Lee H.C. et al. Hepatitis C virus-induced myeloid-derived suppressor cells suppress NK cell IFN-gamma production by altering cellular metabolism via arginase-1. J. Immunol. 2016, 196, 2283-2292.
| название | год | авторы | номер документа |
|---|---|---|---|
| ИММУНОСТИМУЛИРУЮЩАЯ КОМБИНАЦИЯ ДЛЯ ПРОФИЛАКТИКИ И ЛЕЧЕНИЯ ГЕПАТИТА С | 2006 |
|
RU2431499C2 |
| ВАКЦИНА ПРОТИВ HCV | 2003 |
|
RU2323744C2 |
| СПОСОБ ИММУНОТЕРАПИИ ХРОНИЧЕСКОГО ВИРУСНОГО ГЕПАТИТА С | 2016 |
|
RU2637631C2 |
| АГЕНТЫ И СПОСОБЫ, ОСНОВАННЫЕ НА ПРИМЕНЕНИИ ДОМЕНА EDA ФИБРОНЕКТИНА | 2006 |
|
RU2430738C2 |
| Рекомбинантный штамм вируса гриппа A/PR8/HK-NS80E85A, экспрессирующий фрагменты антигенов ESAT-6 и Ag85A M.tuberculosis, для получения векторной вакцины против туберкулеза | 2018 |
|
RU2678175C1 |
| Искусственный ген, кодирующий белок-иммуноген BSI-COV-Ub, рекомбинантная плазмидная ДНК pBSI-COV-Ub, обеспечивающая экспрессию целевого гена, и искусственный полиэпитопный белок-иммуноген BSI-COV-Ub, содержащий убиквитин и эпитопы антигенов вируса SARS-CoV-2 и индуцирующий SARS-CoV-2-специфический Т-клеточный иммунитет | 2023 |
|
RU2806590C1 |
| ВАКЦИННАЯ КОМПОЗИЦИЯ ПРОТИВ ВИРУСА ГЕПАТИТА С | 2005 |
|
RU2351363C2 |
| Плазмидная ДНК-матрица и молекула мРНК, используемая для создания вакцины против коронавируса, полученная с использованием ДНК-матрицы и обеспечивающая экспрессию гена искусственного полиэпитопного белка-иммуногена, содержащего консервативные эпитопы антигенов вируса SARS-Cov-2 и индуцирующего SARS-Cov-2-специфический иммунитет | 2024 |
|
RU2839372C1 |
| РЕКОМБИНАНТНЫЕ ВИРУСЫ ГРИППА А | 2001 |
|
RU2280690C2 |
| ХИМЕРНЫЕ АНТИГЕНЫ ДЛЯ ВАКЦИНЫ ПРОТИВ ВИРУСА ГЕПАТИТА С | 2013 |
|
RU2639504C2 |
Изобретение относится к области вирусологии, биотехнологии и иммунологии и касается иммунобиологического средства и способа его использования. Иммунобиологическое средство получено на основе генетически модифицированных мезенхимальных стволовых клеток человека, экспрессирующих неструктурный белок NS5A вируса гепатита С. Представленное изобретение позволяет повысить клеточный иммунный ответ, характеризующийся усилением антиген-специфической пролиферации лимфоцитов, продукции интерферона-гамма и формированием Т-клеток памяти. Настоящее изобретение может быть использовано в качестве одного из компонентов специфического профилактического и лечебного средства против заболевания, вызванного вирусом гепатита С. 6 ил, 2 табл., 6 пр.
Иммунобиологическое средство для повышения клеточного ответа против вируса гепатита С, состоящее из генетически модифицированных мезенхимальных стволовых клеток человека, экспрессирующих неструктурный белок NS5A вируса гепатита С.
| Masalova OV, Lesnova EI, Klimova RR et al | |||
| Genetically Modified Mouse Mesenchymal Stem Cells Expressing Non-Structural Proteins of Hepatitis C Virus Induce Effective Immune Response | |||
| Vaccines (Basel) | |||
| Способ восстановления спиралей из вольфрамовой проволоки для электрических ламп накаливания, наполненных газом | 1924 |
|
SU2020A1 |
| Масалова О.В., Леснова Е.И., Грабовецкий В.В | |||
| и др | |||
| ДНК-иммунизация плазмидой, содержащей ген белка NS5A вируса гепатита C, | |||
