ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к рекомбинантным антигенным рецепторам и их применению. Т-клетки, созданные методами генной инженерии для экспрессии таких антигенных рецепторов, применимы для лечения заболеваний, характеризующихся экспрессией одного или более антигенов, связываемых антигенными рецепторами.
УРОВЕНЬ ТЕХНИКИ
Т-клетки играют центральную роль в клеточно-опосредованном иммунитете у людей и животных. Распознавание и связывание конкретного антигена опосредуется Т-клеточными рецепторами (TCRs), экспрессируемыми на поверхности Т-4 клеток. TCR Т-клетки способен взаимодействовать с иммуногенными пептидами (эпитопами), связанными с молекулами главного комплекса гистосовместимости (МНС, ГКГС) и презентируемыми на поверхности клеток-мишеней. Специфическое связывание TCR запускает сигнальный каскад внутри Т-клетки, приводящий к пролиферации и дифференцировке в зрелую эффекторную Т-клетку.
TCR является частью комплексного механизма передачи сигнала, который включает модуль сигнальной трансдукции комплекс гетеродимера α- и β-цепей TCR, корецептор CD4 или CD8 и CD3. Гетеродимер TCR α/β отвечает за распознавание антигена и перенос сигнала активации через клеточную мембрану совместно с CD3, тогда как сами цепи CD3 передают входящий сигнал адаптерным белкам внутри клетки. Таким образом, перенос TCR α/β цепей дает возможность перенаправить Т клетки на любой нужный антиген.
В общих чертах иммунотерапию на основе адоптивного переноса клеток (ACT, АПК) можно определить как форму пассивной иммунизации предварительно сенсибилизированными Т-клетками, которые имплантируют не имеющим иммунитета реципиентам или аутологичному хозяину после ex vivo культивирования клеток от низкого исходного числа клеток до клинически релевантного числа клеток. Типы клеток, которые были использованы в ACT экспериментах, включают лимфокин-активированные киллерные (LAK) клетки (Mule, J.J. et al. (1984) Science 225, 1487-1489; Rosenberg, S.A. et al. (1985) N. Engl. J. Med. 313, 1485-1492), лимфоциты, инфильтрирующие опухоль (TILs) (Rosenberg, S.A. et al. (1994) J. Natl. Cancer Inst. 86, 1159-1166), донорские лимфоциты после трансплантации гемопоэтических стволовых клеток (HSCT), так же как линии или клоны опухолеспецифических Т-клеток (Dudley, М.Е. et al. (2001) J. Immunother. 24, 363-373; Yee, С. et al. (2002) Proc. Natl. Acad. Sci. U.S.A 99, 16168-16173). Было показано, что адоптивный перенос Т-клеток обладает терапевтической активностью против вирусных инфекций человека, таких как CMV. Для адоптивной иммунотерапии меланомы Rosenberg и сотрудники разработали метод ACT, основанный на инфузии культивированных in vitro аутологичных лимфоцитов, инфильтрирующих опухоль (TILs), выделенных из удаленных опухолей, в сочетании с немиелоаблативной противолимфоцитарной химиотерапией и высокодозной терапией IL2. Клиническое исследование дало частоту объективного ответа ~ 50% пролеченных пациентов, страдающих метастатической меланомой (Dudley, М.Е. et al. (2005) J. Clin. Oncol. 23: 2346-2357).
Альтернативным методом является адоптивный перенос аутологичных Т-клеток, перепрограммированных для экспрессии опухолереактивного иммунорецептора заданной специфичности в течение кратковременного ex vivo культивирования с последующей реинфузией пациенту (Kershaw М.Н. et al. (2013) Nature Reviews Cancer 13 (8):525-41). Эта стратегия делает ACT применимым к различным распространенным злокачественным опухолям, даже если опухолереактивные Т-клетки отсутствуют у пациента. Поскольку антигенная специфичность Т-клеток находится в состоянии полного покоя на гетеродимерном комплексе α- и β-цепи TCR, перенос клонированных TCR генов в Т-клетки обеспечивает возможность перенаправить их на любой нужный антиген. Следовательно, TCR генная терапия предоставляет заманчивую стратегию разработки антигенспецифической терапии с использованием аутологичных лимфоцитов в качестве метода лечения. Основными преимуществами переноса гена TCR являются получение терапевтических количеств антигенспецифических Т-клеток за несколько дней и возможность вводить специфичности, которые отсутствуют в репертуаре эндогенных TCR пациента. Несколько групп исследователей продемонстрировали, что перенос гена TCR является заманчивой стратегией для того, чтобы перенаправить антигенную специфичность первичных Т-клеток (Morgan, R.A. et al. (2003) J. Immunol. 171, 3287-3295; Cooper, L.J. et al. (2000) J. Virol. 74, 8207-8212; Fujio, K. et al. (2000) J. Immunol. 165, 528-532; Kessels, H.W. et al. (2001) Nat. Immunol. 2, 957-961; Dembic, Z. et al. (1986) Nature 320, 232-238). Возможность осуществления генной терапии с использованием TCR у людей была впервые продемонстрирована Rosenberg и его группой в клинических испытаниях по лечению злокачественной меланомы. Адоптивный перенос аутологичных лимфоцитов, трансдуцированных ретровирусной трансдукцией с использованием меланома/меланоцит антигенспецифических TCR, привел в результате к регрессии рака у до 30% пролеченных больных меланомой (Morgan, R.A. et al. (2006) Science 314, 126-129; Johnson, L.A. et al. (2009) Blood 114, 535-546). За прошедшее время клинические исследования генной терапии с использованием TCR были также распространены на многие различные антигены, нацеленные на раковые заболевания, отличные от меланомы (Park, T.S. et al., (2011) Trends Biotechnol. 29, 550-557).
Применение методов генной инженерии для включения в Т-клетки нацеленных на антиген рецепторов заданной специфичности значительно расширило потенциальные возможности ACT. Химерные антигенные рецепторы (CARs) представляют собой тип нацеленного на антиген рецептора, состоящего из внутриклеточных сигнальных доменов Т-клеточного рецептора, связанных с экстрацеллюлярными антиген-связывающими доменами, чаще всего одноцепочечными вариабельными фрагментами (scFv's) моноклональных антител. CARs непосредственно распознают антигены клеточной поверхности, независимо от МНС-опосредованной презентации, что позволяет применять единственную конструкцию рецептора, специфическую к любому данному антигену, у всех пациентов. Исходные CARs связывали домены антиген-распознающие домены с CD3ζ цепью активации комплекса Т-клеточного рецептора (TCR). Последующие итерации CAR включали вторичные костимулирующие сигналы вместе с CD3ζ, включающие внутриклеточные домены из CD28 или различные молекулы семейства TNF рецепторов, такие как 4-1ВВ (CD137) и ОХ40 (CD134). Далее, рецепторы третьего поколения включают два костимулирующих сигнала помимо CD3ζ, обычно из CD28 и 4-1ВВ. CARs второго и третьего поколения кардинально повышали противоопухолевую эффективность in vitro и in vivo (Zhao et al., (2009) J. Immunol., (183) 5563-5574), в некоторых случаях стимулируя полную ремиссию у пациентов с запущенным раковым заболеванием (Porter et al., (2011) N. Engl. J. Med., (365) 725-733).
Классический CAR состоит из фрагмента антигенспецифического одноцепочечного антитела (scFv), связанного с трансмембранным и сигнальным доменом, таким как CD3ζ. После введения в Т-клетки он экспрессируется в виде мембраносвязанного белка и индуцирует иммунные реакции при связывании со своим распознанным антигеном (Eshhar et al., (1993) PNAS, (90) 720-724). Индуцированный антигенспецифический иммунный ответ обусловливает активацию цитотоксических CD8+ Т-клеток, что в свою очередь приводит к эрадикации клеток, экспрессирующих специфический антиген, таких как опухолевые клетки или инфицированные вирусом клетки, экспрессирующие специфический антиген. Однако эти классические конструкции CAR не активируют / стимулируют Т-клетки посредством использования их эндогенного CD3 комплекса, который обычно является существенно важным для активации Т-клеток. Благодаря слиянию антигенсвязывающего домена с CD3ζ активация Т-клеток индуцируется с помощью биохимического "проскальзывания" (Aggen et al., (2012) Gene Therapy, (19) 365-374). Эта нефизиологическая активация Т-клеток представляет риск для пациента, проходящего лечение таким способом, так как сверхактивация Т-клеток может привести к нежелательным побочным эффектам. Например, вследствие экспрессии CAR наблюдалась in vitro продолжительная базальная активация 'рекомбинантных' Т-клеток ("тоническая передача сигнала"), которая привела к повышенной аккумуляции ингибирующих молекул, таких как LAG-3, TIM-3 и PD-1, на поверхности экспрессирующих рекомбинантные CAR Т-клеток, что в свою очередь имело следствием преждевременно истощенные Т-клетки, а это в дальнейшем оказывало сильное негативное воздействие на ответ на опухолевые клетки in vivo (Long et al., (2015) Nat. Med., (21) 581-590). Эта побочная реакция связана с нерегулярной кластеризацией scFv-фрагментов с помощью каркасных остатков этого антитела. Кроме того, хотя классические конструкции CAR данного типа были успешно испытаны против различных неоплазий, таких как лейкоз (Porter et al., (2011) N. Engl. J. Med., (365) 725-733), они также имели следствием смертельные аутоиммунные заболевания вследствие базальной экспрессии целевого антигена (целевого опухолевого антигена) в здоровых тканях (целенаправленная/внеопухолевая реакция, on-target/off-tumor-reaction; Morgan et al., (2010) Mol Ther., (18) 843-51).
Альтернативным методом, в котором активация Т-клетки происходит по более физиологическому механизму, заключается в предоставлении аналогичного одноцепочечного- TCR (scTv)- фрагмента, слитого с Cβ константным доменом, полученным из Т-клеточного рецептора (TCR), и его коэкспрессия с Сα константным доменом, происходящим из TCR (Voss et al., (2010) Blood, (115) 5154-5163), последним, который рекрутируется в незаменимый эндогенный CD3ζ гомодимер (Call et al., (2002) Cell, (111) 967-79.). Однако, для того, чтобы эти конструкции действовали как активаторы иммунной системы, было принципиально важно, чтобы их константные домены происходили из мышиных TCRs или необходимо ввести остатки мышиных последовательностей (Cohen et al., (2006) Cancer Res., (66) 8878-86; Bialer et al., (2010) J. Immunol. (184) 6232-41), чтобы осуществить спаривание цепей между scTCR и Сα. Тот факт, что эти конструкции для наличия функциональности должны иметь ксеногенные последовательности, повышает риск того, что иммунная система будет им противодействовать при введении и снижать или сводить к нулю их терапевтическую эффективность.
Следовательно, существует необходимость в альтернативных рекомбинантных антигенных рецепторах, при этом, например, рецептор, после связывания антигена, обладает достаточной способностью активировать Т-клетку, в которой он экспрессируется нормальным физиологическим способом с использованием комплекса эндогенного CD3 и, необязательно, без необходимости присутствия каких-либо аминокислотных последовательностей нечеловеческого происхождения, по меньшей мере в домене передачи сигнала антигенного рецептора, которые могли бы вызывать нежелательный иммунный ответ на сам рекомбинантный антигенный рецептор.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Сущность изобретения
Настоящее изобретение относится к рекомбинантным антигенным рецепторам, имеющим по меньшей мере два сайта связывания. Антигенные рецепторы содержат две пептидных цепи. Каждая пептидная цепь содержит по меньшей мере два домена, помимо вариабельной области цепи Т-клеточного рецептора, или ее участка, и домена передачи сигнала от иммунорецептора, при этом каждый из двух доменов на одной пептидной цепи образует антигенсвязывающий сайт с одним из доменов на другой пептидной цепи. Согласно одному варианту антигенный рецептор по изобретению имеет структуру Т-клеточного рецептора, в котором его цепи, каждая, содержат указанные по меньшей мере два домена, образующие антигенсвязывающие сайты, предпочтительно, на N-конце цепей Т-клеточного рецептора.
Согласно одному аспекту настоящее изобретение относится к антигенному рецептору, который содержит первую пептидную цепь и вторую пептидную цепь, причем первая пептидная цепь содержит первый домен, второй домен, вариабельную область цепи Т-клеточного рецептора или ее часть, и домен передачи сигнала от иммунорецептора; вторая пептидная цепь содержит первый домен, второй домен, вариабельную область цепи Т-клеточного рецептора или ее часть, и домен передачи сигнала от иммунорецептора; при этом первый домен из первой пептидной цепи совместно с одним из доменов из второй пептидной цепи образует первый антигенсвязывающий сайт, а второй домен из первой пептидной цепи совместно с другим доменом из второй пептидной цепи образует второй антигенсвязывающий сайт. В антигенном рецепторе согласно данному аспекту домены, образующие соответствующие антигенсвязывающие сайты, предпочтительно, расположены в разных пептидных цепях. Таким образом, антигенсвязывающие сайты образуются в результате межмолекулярного взаимодействия доменов.
Согласно одному варианту первый и/или второй домены, каждый, содержат вариабельную область цепи иммуноглобулина, или вариабельную область цепи Т-клеточного рецептора, или участок вариабельной области.
Согласно одному варианту один из доменов, образующих первый антигенсвязывающий сайт, содержит вариабельную область тяжелой цепи иммуноглобулина со специфичностью к антигену или ее участок, а другой домен, образующий первый антигенсвязывающий сайт, содержит вариабельную область легкой цепи иммуноглобулина со специфичностью к антигену или ее участок. Согласно одному варианту один из доменов, образующих второй антигенсвязывающий сайт, содержит вариабельную область тяжелой цепи иммуноглобулина со специфичностью к антигену или ее участок, а другой домен, образующий второй антигенсвязывающий сайт, содержит вариабельную область легкой цепи иммуноглобулина со специфичностью к антигену или ее участок.
Согласно одному варианту первый домен из первой пептидной цепи содержит вариабельную область тяжелой цепи иммуноглобулина со специфичностью к антигену или ее часть и домен из второй пептидной цепи, образующий антигенсвязывающий сайт с первым доменом первой пептидной цепи, содержит вариабельную область легкой цепи иммуноглобулина со специфичностью к антигену или ее часть. Согласно одному варианту второй домен из первой пептидной цепи содержит вариабельную область тяжелой цепи иммуноглобулина со специфичностью к антигену или ее часть, а домен из второй пептидной цепи, образующей антигенсвязывающий сайт со вторым доменом из первой пептидной цепи, содержит вариабельную область легкой цепи иммуноглобулина со специфичностью к антигену или ее часть.
Согласно одному варианту первый и второй домены из первой пептидной цепи, каждый, содержат вариабельную область тяжелой цепи иммуноглобулина или ее часть; и первый и второй домены из второй пептидной цепи, каждый, содержат вариабельную область легкой цепи иммуноглобулина или ее часть.
Согласно одному варианту N-концевой домен из первой пептидной цепи совместно с N-концевым доменом из второй пептидной цепи образует антигенсвязывающий сайт; и С-концевой домен из первой пептидной цепи совместно с С-концевым доменом из второй пептидной цепи образует антигенсвязывающий сайт.
Согласно одному варианту N-концевой домен из первой пептидной цепи совместно с С-концевым доменом из второй пептидной цепи образует антигенсвязывающий сайт; и С-концевой домен из первой пептидной цепи совместно с N-концевым доменом из второй пептидной цепи образует антигенсвязывающий сайт.
В одном варианте антигенных рецепторов по изобретению домен передачи сигнала от иммунорецептора содержит константную или консервативную область цепи Т-клеточного рецептора или константную или консервативную область Fc цепи иммунного рецептора или часть константной или консервативной области. В одном варианте антигенных рецепторов по изобретению (i) первая пептидная цепь содержит вариабельную область альфа цепи Т-клеточного рецептора или ее часть и константную область альфа цепи Т-клеточного рецептора или ее часть, а вторая пептидная цепь содержит вариабельную область бета цепи Т-клеточного рецептора или ее часть и константную область бета цепи Т-клеточного рецептора или ее часть, или (ii) первая пептидная цепь содержит вариабельную область бета цепи Т-клеточного рецептора или ее часть и константную область бета цепи Т-клеточного рецептора или ее часть, а вторая пептидная цепь содержит вариабельную область альфа цепи Т-клеточного рецептора или ее часть и константную область альфа цепи Т-клеточного рецептора или ее часть. Согласно этому варианту вариабельная область альфа цепи Т клеточного рецептора или ее часть и константная область альфа цепи Т клеточного рецептора или ее часть соответствует или по существу соответствует альфа цепи Т-клеточного рецептора, а вариабельная область бета цепи Т-клеточного рецептора или ее часть и константная область бета цепи Т-клеточного рецептора или ее часть соответствует или по существу соответствует бета цепи Т-клеточного рецептора, предпочтительно, того же Т-клеточного рецептора, из которого происходит альфа цепь Т-клеточного рецептора.
Домены, образующие антигенсвязывающие сайты, предпочтительно, сливаются по N-концу цепей, необязательно разделенных линкером.
Согласно одному варианту антигенных рецепторов по изобретению вариабельная область цепи Т-клеточного рецептора или ее часть, и/или домен передачи сигнала от иммунорецептора, такой как константная область Т-клеточного рецептора или ее часть, являются последовательностями человеческого происхождения. Так, цепь Т-клеточного рецептора, которой может соответствовать или по существу соответствовать вариабельная область цепи Т-клеточного рецептора или ее часть и константная область Т-клеточного рецептора или ее часть, может быть человеческого происхождения.
Согласно одному варианту антигенный рецептор по изобретению содержит (а) линкер(ы), связывающий(-ие) домены антигенного рецептора. Согласно одному варианту антигенный рецептор по изобретению содержит один или более линкеров между доменами, образующими антигенсвязывающие сайты, и/или между доменами, образующими антигенсвязывающие сайты, и вариабельными областями цепи Т-клеточного рецептора или ее части. Линкер может представлять собой произвольную аминокислотную последовательность любой протяженности при условии, что он не нарушает функции антигенного рецептора, такие, как способность антигенного рецептора связывать антиген или ассоциироваться с эндогенным CD3 комплексом, или нарушать способность антигенного рецептора стимулировать иммунный ответ на связывание антигена.
В соответствии с одним вариантом антигенных рецепторов по изобретению первый и второй антигенсвязывающие сайты связываются с одним и тем же антигеном или с разными антигенами. В одном варианте антигенных рецепторов по изобретению первый и второй антигенсвязывающие сайты связываются с различными эпитопами на одном и том же антигене. Соответственно, хотя домены, образующие первый антигенсвязывающий сайт, предпочтительно происходят из одного и того же иммуноглобулина, и домены, образующие второй антигенсвязывающий сайт, предпочтительно происходят из одного и того же иммуноглобулина, домены, образующие первый антигенсвязывающий сайт, и домены, образующие второй антигенсвязывающий сайт, получают из одного и того же иммуноглобулина или из разных иммуноглобулинов, причем указанные различные иммуноглобулины связываются с одним и тем же антигеном или с разными антигенами.
Согласно одному варианту антиген представляет собой антиген, специфический для конкретного заболевания, предпочтительно, опухолевый антиген. Согласно одному варианту антиген экспрессируется на поверхности клетки.
В одном аспекте настоящее изобретение относится к пептидной цепи антигенных рецепторов по изобретению. Согласно одному варианту настоящее изобретение относится к пептидной цепи, содержащей первый и второй домен, каждый из которых содержит вариабельную область тяжелой цепи иммуноглобулина или ее часть или каждый содержит вариабельную область легкой цепи иммуноглобулина или ее часть, и при этом пептидная цепь содержит далее вариабельную область цепи Т-клеточного рецептора или ее часть и домен передачи сигнала от иммунорецептора, такой как константная область цепи Т-клеточного рецептора или ее часть. Дополнительные варианты пептидных цепей по изобретению описаны в данной заявке для антигенных рецепторов по изобретению.
В одном аспекте настоящее изобретение относится к клетке, в частности, к иммунной эффекторной клетке, такой как Т-клетка, генетически модифицированная для экспрессии антигенного рецептора по изобретению. В одном аспекте настоящее изобретение относится к клетке, в частности, к иммунной эффекторной клетке, такой как Т-клетка, экспрессирующая первую пептидную цепь, вторую пептидную цепь или как первую, так и вторую пептидные цепи антигенного рецептора по изобретению, или экспрессирующая пептидную цепь по изобретению. Дополнительные варианты клетки или 'рекомбинантной' клетки по изобретению описаны в настоящей заявке для антигенных рецепторов по изобретению или пептидных цепей по изобретению.
В одном аспекте настоящее изобретение относится к способу получения клетки, экспрессирующей антигенный рецептор по изобретению, включающий: (а) предоставление клетки; (b) предоставление первой генетической конструкции, кодирующей первую пептидную цепь антигенного рецептора по изобретению; (с) предоставление второй генетической конструкции, кодирующей вторую пептидную цепь антигенного рецептора по изобретению; (d) введение первой и второй генетической конструкции в клетку; и (е) обеспечение возможности экспрессии конструкций в клетке. Согласно одному варианту настоящее изобретение относится к способу получения клетки, экспрессирующей антигенный рецептор, причем этот рецептор содержит первую пептидную цепь и вторую пептидную цепь, включающему: (а) предоставление клетки; (b) предоставление первой генетической конструкции, кодирующей первую пептидную цепь, содержащую первый домен, второй домен, вариабельную область цепи Т-клеточного рецептора или ее часть, и домен передачи сигнала иммуносупрессора; (с) предоставление второй генетической конструкции, кодирующей вторую пептидную цепь, содержащую по меньшей мере первый домен, второй домен, вариабельную область цепи Т-клеточного рецептора или ее часть, и домен передачи сигнала от иммунорецептора; (d) введение первой и второй генетической конструкции в клетку; и (е) обеспечение возможности экспрессии конструкций в клетке; причем первый домен из первой пептидной цепи способен образовывать, совместно с одним из доменов из второй пептидной цепи, первый антигенсвязывающий сайт, а второй домен из первой пептидной цепи способен образовывать, совместно с другим доменом из второй пептидной цепи, второй антигенсвязывающий сайт. Согласно одному варианту способов по изобретению экспрессия антигенного рецептора осуществляется на поверхности клетки. В одном варианте способов по изобретению первая пептидная цепь и вторая пептидная цепь предоставляются в единой генетической конструкции. В одном варианте способов по изобретению клетка является человеческой клеткой. В одном варианте способов по изобретению генетические конструкции содержат ДНК и/или РНК. Другие варианты способов по изобретению соответствуют способам, описанным в настоящей заявке для антигенных рецепторов по изобретению.
В одном аспекте настоящее изобретение относится к 'рекомбинантной' клетке, в частности, к иммунной эффекторной клетке, такой как Т клетка, полученной способами по изобретению, для получения клетки, экспрессирующей антигенный рецептор. Другие варианты 'рекомбинантной' клетки по изобретению соответствуют описанным в настоящей заявке для антигенных рецепторов по изобретению или в способах получения клетки, экспрессирующей антигенный рецептор по изобретению.
В одном аспекте настоящее изобретение относится к нуклеиновой кислоте, например, к ДНК или РНК, кодирующей первую пептидную цепь, вторую пептидную цепь, или как первую, так и вторую пептидную цепь антигенного рецептора по изобретению или кодирующие пептидную цепь по изобретению. Другие варианты нуклеиновой кислоты по изобретению соответствуют вариантам нуклеиновой кислоты по настоящему описанию для антигенных рецепторов по изобретению или пептидных цепей по изобретению.
Настоящее изобретение в общем охватывает лечение заболеваний путем таргетирования клеток, экспрессирующих один или более антигенов на поверхности клеток, например, больных клеток, экспрессирующих один или более специфических для конкретной болезни антигенов на клеточной поверхности, в частности, раковых клеток, экспрессирующих один или более опухолевых антигенов на поверхности клеток, с помощью антигенных рецепторов по изобретению. Способы предусматривают селективную эрадикацию клеток, которые экспрессируют на своей поверхности один или более антигенов, тем самым минимизируя побочное действие на нормальные клетки, не экспрессирующие антиген(-ы). Согласно одному варианту вводят Т-клетки, модифицированные для экспрессии антигенного рецептора по изобретению, таргетирующего клетки посредством связывания с антигеном(-ами). Т-клетки способны распознавать больные клетки, экспрессирующие антиген(-ы) на поверхности клеток, приводя в результате к эрадикации больных клеток. Согласно одному варианту популяция клеток-мишеней или ткань-мишень представляют собой опухолевые клетки или опухолевую ткань.
В одном аспекте настоящее изобретение относится к фармацевтической композиции, содержащей антигенный рецептор по изобретению, рекомбинантную клетку по изобретению или нуклеиновую кислоту по изобретению и фармацевтически приемлемый носитель. Фармацевтическая композиция по изобретению может применяться в качестве лекарственного средства, в частности для лечения заболевания, такого как рак, характеризующегося экспрессией одного или более антигенов, которые связываются с антигенным рецептором по изобретению, например, таким как один или более опухолевых антигенов.
В одном аспекте настоящее изобретение относится к способу лечения заболевания, такого как рак, включающему введение субъекту терапевтически эффективного количества фармацевтической композиции по изобретению, причем заболевание характеризуется экспрессией по меньшей мере одного антигена, такого как опухолевый антиген, который связывается антигенным рецептором.
В одном аспекте настоящее изобретение относится к способу лечения субъекта, у которого наблюдается заболевание, нарушение или патологическое состояние, ассоциированное с экспрессией или с повышенной экспрессией по меньшей мере одного антигена, причем способ включает введение субъекту Т-клеток, генетически модифицированных для экспрессии антигенного рецептора по изобретению, нацеленного по меньшей мере на один антиген. Согласно одному варианту заболевание, нарушение или патологическое состояние представляет собой рак. Согласно одному варианту Т-клетки могут быть аутологичными, гомологичными или сингенными для субъекта.
Согласно одному варианту настоящего изобретения антигенный рецептор связывается только с одним антигеном (например, будучи моноспецифическим и распознающим один и тот же эпитоп или будучи биспецифическим или мультиспецифическим и распознающим разные эпитопы на одном и том же антигене) или связывается с различными антигенами, в частности, с двумя различными антигенами.
Согласно одному варианту всех аспектов данного изобретения способ лечения также включает взятие образца клеток от субъекта, причем образец содержит Т-клетки или предшественники Т-клеток, и трансфицирование клеток с использованием нуклеиновой кислоты, кодирующей антигенный рецептор по изобретению, для предоставления Т-клеток, генетически модифицированных для экспрессии антигенного рецептора. Согласно одному варианту всех аспектов изобретения Т-клетки, генетически модифицированные для экспрессии антигенного рецептора, стабильно или транзиторно трансфицированы с использованием нуклеиновой кислоты, кодирующей антигенный рецептор. Таким образом, нуклеиновая кислота, кодирующая антигенный рецептор, является интегрированной или неинтегрированной в геном Т-клеток. Согласно одному варианту всех аспектов изобретения Т-клетки и/или образец клеток взяты от субъекта, которому вводятся Т-клетки, генетически модифицированные для экспрессии антигенного рецептора. Согласно одному варианту всех аспектов изобретения Т-клетки и/или образец клеток взяты от млекопитающего, отличного от млекопитающего, которому вводятся Т-клетки, генетически модифицированные для экспрессии антигенного рецептора.
Согласно одному варианту всех аспектов изобретения Т-клетки, генетически модифицированные для экспрессии антигенного рецептора, инактивируются для экспрессии эндогенного Т-клеточного рецептора и/или эндогенного HLA.
Согласно одному варианту всех аспектов изобретения антиген экспрессируется в больной клетке, например, такой как раковая клетка. Согласно одному варианту антиген экспрессируется на поверхности больной клетки, например, такой как раковая клетка. Согласно одному варианту антигенный рецептор связывается с внеклеточным доменом или с эпитопом во внеклеточном домене антигена. Согласно одному варианту антигенный рецептор связывается с нативными эпитопами антигена, присутствующими на поверхности жизнеспособных клеток. Согласно одному варианту всех аспектов изобретения антиген представляет собой опухолевый антиген. Согласно одному варианту всех аспектов изобретения антиген выбран из группы, состоящей из клаудинов, например, таких как клаудин 6 и клаудин 18.2, CD19, CD20, CD22, CD33, CD123, мезотелин, СЕА, с-Met, PSMA, GD-2 и NY-ESO-1. Согласно одному варианту всех аспектов изобретения антиген является антигеном из патогена. Патоген может представлять собой грибковый, вирусный или бактериальный патоген. Согласно одному варианту всех аспектов изобретения экспрессия антигена осуществляется на поверхности клетки. Согласно одному варианту антиген представляет собой клаудин (Claudin), в частности Claudin (клаудин) 6 или Claudin (клаудин) 18.2, и указанный антигенный рецептор связывается с первой внеклеточной петлей указанного клаудина. Согласно одному варианту связывание указанного антигенного рецептора, экспрессируемого Т-клетками и/или присутствующего на Т-клетках, с антигеном, присутствующим на клетках, обусловливает эффекторные функции указанных Т-клеток иммунной системы, такие как высвобождение цитокинов. Согласно одному варианту связывание указанного антигенного рецептора, экспрессируемого Т-клетками и/или присутствующего на Т-клетках, таких как антигенпрезентирующие клетки, с антигеном, присутствующим на клетках, таких как антигенпрезентирующие клетки, обусловливает стимуляцию, примирование и/или экспансию указанных Т-клеток. Согласно одному варианту связывание указанного антигенного рецептора, экспрессируемого Т-клетками и/или присутствующего на Т-клетках, с антигеном, присутствующим на больных клетках, обусловливает цитолиз и/или апоптоз больных клеток, при этом указанные Т-клетки предпочтительно высвобождают цитотоксические факторы, например, перфорины и гранзимы.
Согласно одному варианту всех аспектов изобретения домены антигенного рецептора, образующие антигенсвязывающие сайты, составлены эктодоменом антигенного рецептора. Согласно одному варианту всех аспектов изобретения антигенный рецептор по изобретению содержит трансмембранный домен. Согласно одному варианту трансмембранный домен представляет собой гидрофобную альфа-спираль, которая перекрывает мембрану.
Согласно одному варианту всех аспектов изобретения антигенный рецептор по изобретению содержит сигнальный пептид, который направляет растущий белок в эндоплазматический ретикулум. Согласно одному варианту сигнальный пептид предшествует домену, образующему антигенсвязывающие сайты.
Согласно одному варианту всех аспектов изобретения антигенный рецептор по изобретению, предпочтительно, является специфическим к антигену, на который он нацелен, в частности, находящийся на поверхности клетки, такой как больная клетка или антигенпрезентирующая клетка.
Согласно одному варианту всех аспектов изобретения антигенный рецептор по изобретению может экспрессироваться поверхностью и/или присутствовать на поверхности иммунореактивной клетки, например, такой как Т-клетка, предпочтительно, цитотоксическая Т-клетка. Согласно одному варианту Т-клетка способна реагировать с антигеном(-ами), на который(-е) нацелен антигенный рецептор по изобретению.
В соответствии с другим аспектом изобретения предусматриваются агенты и композиции по данному описанию для применения в способах по данному изобретению.
Другие признаки и преимущества настоящего изобретения будут очевидны на основании последующего подробного описания и формулы изобретения.
Подробное описание изобретения
Хотя настоящее изобретение подробно описано ниже, следует понимать, что данное изобретение не ограничивается конкретными методиками, протоколами и реагентами по данному описанию, поскольку таковые могут меняться. Также следует понимать, что терминология, применяемая в данной заявке, дается только с целью описания конкретных вариантов и не предназначена для ограничения объема настоящего изобретения, который ограничивается лишь прилагаемой формулой изобретения. Если особо не указано иное, все технические и научные термины, применяемые в данном описании, имеют те же значения, которые обычно подразумевает средний специалист в данной области техники.
Ниже описываются элементы настоящего изобретения. Эти элементы приводятся с конкретными вариантами, однако, следует понимать, что их можно комбинировать любым иным образом и в любом количестве, создавая дополнительные варианты. Разные описанные примеры и предпочтительные варианты не следует толковать как ограничивающие настоящее изобретение лишь в явном виде описанными вариантами. Следует понимать, что данное описание защищает и охватывает варианты, которые объединяют в явном виде описанные варианты с любым числом раскрываемых и/или предпочтительных элементов. Также следует рассматривать любые перестановки и комбинации всех описанных в настоящей заявке элементов как раскрываемые в описании настоящей заявки, если контекст не свидетельствует об ином.
Предпочтительно, термины, применяемые в настоящей заявке, определяются, как описано в глоссарии "A multilingual glossary of biotechnological terms: (IUPAC Recommendations)", H.G.W. Leuenberger, B. Nagel, and H.  , Eds., (1995) Helvetica Chimica Acta, CH-4010 Basel, Switzerland.
, Eds., (1995) Helvetica Chimica Acta, CH-4010 Basel, Switzerland.
При практическом осуществлении настоящего изобретения, если особо не указано иное, применяют обычные методы биохимии, биологии клетки, иммунологии и методы рекомбинантной ДНК, которые разъясняются в литературе, относящейся к данной области техники (ср., например, Molecular Cloning: A Laboratory Manual, 2nd Edition, J. Sambrook et al. eds., Cold Spring Harbor Laboratory Press, Cold Spring Harbor 1989).
По ходу данного описания и в формуле изобретения, которые приводятся далее, если по контексту не требуется иное, следует понимать, что слово "содержать" и его варианты, например, такие как "содержит" и "содержащий", предполагает включение указанного элемента, целого числа или стадии или группы элементов, целых чисел или стадий, хотя в некоторых вариантах такой другой элемент, целое число или стадия или группа элементов, целых чисел или стадий может быть исключена, т.е. предмет изобретения представляет собой включение указанного элемента, целого числа или стадии или группы элементов, целых чисел или стадий. Термины, определяющие единственное число, и аналогичное указание, применяемое в контексте описания изобретения (в особенности применительно к формуле изобретения), следует рассматривать как охватывающие как единственное, так и множественное число, если в данном описании не указано иное или если это явно не противоречит контексту. Предполагается, что перечисление интервалов величин в данном описании служит лишь для краткости при обращении индивидуально к каждой отдельной величине внутри интервала. Если в настоящем описании не указано иное, каждая индивидуальная величина включена в описание, как если бы она была перечислена индивидуально.
Все способы, описанные в настоящей заявке, можно осуществлять в любом подходящем порядке, если не указано иное, или если это иным образом явно не противоречит контексту. Применение любых примеров или вводных слов перед примером (например, "такие как") в настоящем описании предназначено только для того, чтобы более наглядно проиллюстрировать настоящее изобретение, и не ограничивает объем изобретения, заявленный в иных случаях. Никакие термины в описании не следует истолковывать как указывающие на какой-либо незаявленный элемент, крайне важный для практического осуществления настоящего изобретения.
Некоторые документы цитируются по тексту данного описания. Каждый из документов, цитируемых в данном описании (включая все патенты, заявки на патент, научные публикации, технические требования изготовителя, инструкции и т.д.), либо supra, либо infra, включены посредством отсылки во всей полноте. Данное изобретение не дает права рассматривать его как предшествующий более ранней дате раскрытия данного изобретения в предыдущем изобретении.
Термин "иммунный ответ" относится к общему системному ответу на антиген и предпочтительно относится к клеточному иммунному ответу или как к клеточному, так и к гуморальному иммунному ответу. Иммунный ответ может быть защитным/превентивным/профилактическим и/или терапевтическим.
"Обеспечение иммунного ответа" может означать, что не было никакого иммунного ответа на конкретный целевой антиген, клетку-мишень и/или ткань/мишень до обеспечения иммунного ответа, но это также может означать, что был определенный уровень иммунного ответа на конкретный целевой антиген, клетку-мишень и/или ткань/мишень до обеспечения иммунного ответа, а после формирования иммунного ответа указанный иммунный ответ усиливается. Таким образом "обеспечение иммунного ответа" включает "индукцию иммунного ответа" и "усиление иммунного ответа". Предпочтительно, когда после обеспечения иммунного ответа у субъекта указанный субъект защищен от развития заболевания, например, такого как рак, или интенсивность болезненного состояния уменьшается посредством обеспечения иммунного ответа. Например, иммунный ответ на опухолевый антиген можно сформировать у больного раком или у субъекта с риском развития ракового заболевания. Обеспечение иммунного ответа в данном случае может означать, что интенсивность болезненного состояния уменьшается, что у субъекта не развиваются метастазы, или что у субъекта с риском развития ракового заболевания раковое заболевание не развивается.
Предполагается, что термины "клеточно-опосредованный иммунитет", или "клеточный иммунитет", или аналогичные термины включают клеточный ответ, направленный на клетки, характеризующиеся экспрессией антигена, в частности характеризующиеся презентацией антигена молекулами МНС класса I или класса II. Клеточный ответ относится к клеткам, называемым Т-клетками или Т-лимфоцитами, которые действуют либо как "хелперы", либо как "киллерные клетки". Хелперные Т-клетки (также называемые CD4+ Т-клетками) играют главную роль за счет регулирования иммунного ответа, а киллерные клетки (также называемые цитотоксическими Т-клетками, цитолитическими Т-клетками, CD8+ Т-клетками или CTLs) убивают больные клетки, например, такие как раковые клетки, предупреждая продуцирование новых больных клеток.
Термин "антиген" относится к агенту, содержащему эпитоп, против которого должен формироваться и/или направляется иммунный ответ. Предпочтительно, антиген в контексте настоящего изобретения означает молекулу, которая, необязательно после процессирования, индуцирует иммунную реакцию, которая, предпочтительно, является специфической по отношению к антигену или клеткам, экспрессирующим антиген, предпочтительно, на поверхности клетки. Термин "антиген" включает в частности белки и пептиды. Антиген предпочтительно является продуктом, который соответствует антигену природного происхождения или получен с использованием антигена природного происхождения. Такие антигены природного происхождения могут включать или могут иметь источником происхождения аллергены, вирусы, бактерии, грибы, паразиты и другие инфекционные агенты и патогены, или антиген может также являться опухолевым антигеном. Согласно настоящему изобретению антиген может соответствовать продукту природного происхождения, например, вирусному белку, или его части.
Термин "патоген" относится к патогенным микроорганизмам и включает вирусы, бактерии, грибы, одноклеточные организмы и паразиты. Примерами патогенных вирусов являются вирус иммунодефицита человека (HIV, ВИЧ), цитомегаловирус (CMV), герпес-вирус (HSV), вирус гепатита A (HAV), HBV, HCV, папилломавирус и Т-лимфотропный вирус человека (HTLV). Одноклеточные организмы включают плазмодии, трипаносомы, амебы и т.д.
Согласно предпочтительному варианту антиген представляет собой антиген, специфический для конкретного заболевания, или ассоциированный с заболеванием (патогенный) антиген. Термин "антиген, специфический для конкретного заболевания" или "ассоциированный с заболеванием антиген" относится ко всем антигенам, которые имеют патологическую значимость. Согласно одному особенно предпочтительному варианту антиген присутствует в больных клетках, тканях и/или органах, в то же время он отсутствует или присутствует в уменьшенных количествах в здоровых клетках, тканях и/или органах, и, следовательно, его можно использовать для таргетирования больных клеток, тканей и/или органов, например, с использованием Т-клеток, несущих антигенный рецептор, нацеленный на антиген. Согласно одному варианту антиген, специфический для конкретного заболевания, или ассоциированный с заболеванием, присутствует на поверхности больной клетки.
Согласно предпочтительному варианту изобретения антиген представляет собой опухолевый антиген или опухолеассоциированный антиген, элемент раковой клетки, который может быть образован с использованием цитоплазмы, поверхности клетки и ядра клетки, в частности, такие антигены, которые продуцируются, предпочтительно, в большом количестве, в виде поверхностных антигенов на раковых клетках.
Применительно к настоящему изобретению термин "опухолевый антиген" или "опухолеассоциированный антиген" относится к белкам, которые в нормальных условиях специфически экспрессируются в ограниченном числе тканей и/или органов или на конкретных стадиях развития, например, опухолевый антиген в нормальных условиях может специфически экспрессироваться в ткани желудка, предпочтительно, в слизистой оболочке желудка, в репродуктивных органах, например, в яичках, в трофобластной ткани, например, в плаценте, или в клетках зародышевой линии, и экспрессируются или аберрантно экспрессируются в одной или более опухолевой или раковой ткани. В данном контексте термин "ограниченное число", предпочтительно, означает не более, чем 3, более предпочтительно, не более, чем 2. Опухолевые антигены, применительно к настоящему изобретению, включают, например, дифференцировочные антигены, предпочтительно, специфические по отношению к клеточному типу дифференцировочные антигены, т.е. белки, которые в нормальных условиях специфически экспрессируются в клетках определенного типа на определенной стадии дифференцировки, раково-тестикулярные антигены, т.е. белки, которые в нормальных условиях специфически экспрессируются в яичках и иногда в плаценте, и, специфические антигены зародышевой линии. Применительно к настоящему изобретению опухолевый антиген предпочтительно ассоциируется с клеточной поверхностью раковой клетки и предпочтительно не экспрессируется, или крайне редко экспрессируется в нормальных тканях. Предпочтительно, опухолевый антиген или аберрантная экспрессия опухолевого антигена позволяет идентифицировать опухолевые клетки. Применительно к настоящему изобретению опухолевый антиген, который экспрессируется раковой клеткой у субъекта, например, пациента, страдающего раковым заболеванием, обозначает, предпочтительно, собственный белок указанного субъекта. Согласно предпочтительным вариантам опухолевый антиген, применительно к настоящему изобретению, экспрессируется в нормальных условиях главным образом в тканях или в органах, которые не являются основными, т.е. в тканях или органах, повреждение которых, при повреждении иммунной системы, не приводит к смерти субъекта, или в органах или структурах организма, которые являются недоступными или разве что труднодоступными для иммунной системы. Предпочтительно, аминокислотная последовательность в опухолевом антигене, который экспрессируется в нормальных тканях, и в опухолевом антигене, который экспрессируется в раковых тканях, является идентичной.
Примерами опухолевых антигенов, которые могут применяться в настоящем изобретении, являются р53, ART-4, BAGE, бета-катенин/m, Bcr-abL, CAMEL, САР-1, CASP-8, CDC27/m, CDK4/m, СЕА, белки клеточной поверхности семейства клаудина, например, такие как CLAUDIN-6, CLAUDIN-18.2 и CLAUDIN-12, c-MYC, СТ, Сур-В, DAM, ELF2M, ETV6-AML1, G250, GAGE, GnT-V, Gap100, HAGE, HER-2/neu, HPV-E7, HPV-E6, HAST-2, hTERT (или hTRT), LAGE, LDLR/FUT, MAGE-А, предпочтительно, MAGE-A1, MAGE-A2, MAGE-A3, MAGE-A4, MAGE-A5, MAGE-A6, MAGE-A7, MAGE-A8, MAGE-A9, MAGE-A10, MAGE-A11, или MAGE-A12, MAGE-B, MAGE-C, MART-1/Melan-A, MC1R, Myosin/m, MUC1, MUM-1, -2, -3, NA88-A, NF1, NY-ESO-1, NY-BR-1, p190 минорный BCR-abL, Pm1/RARa, PRAME, протеиназа 3, PSA, PSM, RAGE, RU1 или RU2, SAGE, SART-1 или SART-3, SCGB3A2, SCP1, SCP2, SCP3, SSX, SURVIVIN, TEL/AML1, TPI/m, TRP-1, TRP-2, TRP-2/INT2, ТРТЕ и WT. Особенно предпочтительные опухолевые антигены включают CLAUDIN-18.2 (CLDN18.2) и CLAUDIN-6 (CLDN6).
Термин "CLDN" или просто "Cl" в контексте настоящей заявки означает клаудин и включает CLDN6 и CLDN18.2. Предпочтительно, клаудин означает человеческий клаудин. Клаудины представляют собой семейство белков, которые являются наиболее важными компонентами плотных контактов, поскольку они устанавливают парацеллюлярный барьер, который контролирует поток молекул во внутриклеточном пространстве между клетками эпителия. Клаудины представляют собой трансмембранные белки, перекрывающие мембрану 4 раза, причем как их N-концевая область, так и С-концевая область находятся в цитоплазме. Первая внеклеточная петля, обозначенная ЕС1 или ECL1, состоит в среднем из 53 аминокислот, а вторая внеклеточная петля, обозначенная ЕС2 или ECL2, включает около 24 аминокислот. Белки клеточной поверхности семейства клаудина экспрессируются в опухолях различного происхождения и особенно подходят в качестве целевых структур применительно к прицельной иммунотерапии рака благодаря их селективной экспрессии (отсутствие экспрессии в нормальной ткани релевантной токсичности) и локализации в плазматической мембране.
CLDN6 и CLDN18.2 были идентифицированы как дифференциально экспрессируемые в опухолевых тканях, причем единственной нормальной тканью, экспрессирующей CLDN18.2, является желудок (дифференцированные эпителиальные клетки слизистой оболочки желудка), а единственной нормальной тканью, экспрессирующей CLDN6, является плацента.
CLDN18.2 экспрессируется в злокачественных опухолях различного происхождения, таких как рак поджелудочной железы, рак пищевода, карцинома желудка, бронхиальная карцинома, карцинома молочной железы и ENT опухоли. CLDN18.2 представляет собой важную мишень для предупреждения и/или лечения первичных опухолей, таких как рак желудка, рак пищевода, рак поджелудочной железы, рак легкого, например, такой как немелкоклеточный рак легкого (NSCLC), рак яичников, рак толстой кишки, рак печени, рак головы и шеи и рак желчного пузыря, и их метастазов, в частности, метастазов рака желудка, например, таких как опухоли Крукенберга, метастазы брюшины и метастазы лимфатических узлов. Антигенные рецепторы, нацеленные по меньшей мере на CLDN18.2, применимы при лечении указанных раковых заболеваний.
Было обнаружено, что CLDN6 экспрессируется, например, при раке яичников, раке легкого, раке желудка, раке молочной железы, раке печени, раке поджелудочной железы, раке кожи, меланомах, раке головы и шеи, саркомах, раке желчного протока, почечно-клеточном раке и раке желчного пузыря. CLDN6 представляет собой особенно предпочтительную мишень для предупреждения и/или лечения рака яичников, в частности, аденокарциномы яичника и тератокарциномы яичника, рака легкого, включая мелкоклеточный рак легкого (SCLC) и немелкоклеточный рак легкого (NSCLC), в частности, плоскоклеточный рак легкого и аденокарциному, предупреждения и/или лечения рака желудка, рака молочной железы, рака печени, рака поджелудочной железы, рака кожи, в частности, базальноклеточного рака и плоскоклеточного рака, злокачественной меланомы, рака головы и шеи, в частности злокачественной плеоморфной аденомы, саркомы, в частности, синовиальной саркомы и карциносаркомы, рака желчного протока, рака мочевого пузыря, в частности, переходно-клеточного рака и папиллярного рака, рака почки, в частности почечно-клеточного рака, включая светлоклеточный почечно-клеточный рак и папиллярный рак почки, рака толстой кишки, рака тонкого кишечника, включая рак подвздошной кишки, в частности аденокарциному тонкого кишечника и аденокарциному подвздошной кишки, тестикулярного эмбрионального рака, хориокарциномы плацентарного ложа, рака шейки матки, рака яичников, в частности, семиномы яичка, тератомы яичка, тератомы яичка и эмбрионального тестикулярного рака, рака матки, эмбрионально-клеточных опухолей, например, таких как тератокарцинома или эмбриональная карцинома, в частности, эмбрионально-клеточных опухолей яичка и их метастатических форм. Антигенные рецепторы, нацеленные по меньшей мере на CLDN6, применимы при лечении таких раковых заболеваний.
В контексте вариантов настоящего изобретения антиген предпочтительно присутствует на поверхности клетки, предпочтительно, антигенпрезентирующей клетки или больной клетки. Согласно изобретению антиген, будучи связанным антигенным рецептором, предпочтительно способен индуцировать, необязательно в присутствии соответствующих костимулирующих сигналов, стимуляцию, примирование и/или экспансию Т-клетки, несущей антигенный рецептор, связывающий антиген. Распознавание антигена на поверхности больной клетки может привести к иммунной реакции против антигена (или клетки, экспрессирующей антиген).
Согласно различным аспектам изобретения целью предпочтительно является обеспечение иммунного ответа против больных клеток, экспрессирующих антиген, такой как опухолевый антиген, в частности CLDN6 или CLDN18.2, и лечение заболевания, например, такого как раковое заболевание, включающее клетки, экспрессирующие антиген, такой как опухолевый антиген. Предпочтительно, изобретение включает введение модифицированных с помощью антигенного рецептора эффекторных клеток иммунной системы, таких как Т-клетки, нацеленные на больные клетки, экспрессирующие антиген. Клетки, экспрессирующие антиген на поверхности, могут быть таргетированы эффекторными клетками иммунной системы, несущими антигенный рецептор, нацеленный на антиген.
Термин "клеточная поверхность" применяется в соответствии со значением, обычным в уровне техники, и, следовательно, включает наружную поверхность клетки, которая является доступной для связывания белками и другими молекулами. Антиген экспрессируется на поверхности клеток, если он локализован на поверхности указанных клеток и доступен для связывания антигенсвязывающими молекулами, например, такими как антигенные рецепторы или антитела, специфические по отношению к антигену, добавленные к клеткам. Согласно одному варианту настоящего изобретения антиген, экспрессируемый на поверхности клеток, представляет собой интегральный мембранный белок, имеющий внеклеточный участок, распознаваемый антигенным рецептором. Антигенный рецептор экспрессируется на поверхности клеток, если он локализован на поверхности указанных клеток и доступен для связывания, например, антигеном, к которому антигенный рецептор, добавленный к клеткам, является специфическим. Согласно одному варианту антигенный рецептор, экспрессируемый на поверхности клеток, представляет собой интегральный мембранный белок, имеющий внеклеточный участок, распознающий антиген.
Термин "внеклеточный участок" или "эктодомен" в контексте настоящего изобретения относится к части молекулы, такой как белок, которая расположена напротив внеклеточного пространства клетки и, предпочтительно, доступна снаружи указанной клетки, например, посредством связывания молекул, таких как антитела, локализованные снаружи клетки. Предпочтительно, термин относится к одной (или одному) или более внеклеточных петель или доменов или их фрагментов.
Термины "участок" или "часть" применяются в данной заявке в качестве синонимов и относятся к непрерывному или прерывистому элементу структуры, например, такому как аминокислотная последовательность. Термин "фрагмент" относится к непрерывному элементу структуры, например, такому как аминокислотная последовательность. Участок или часть белковой последовательности, предпочтительно, содержит по меньшей мере 6, в частности по меньшей мере 8, по меньшей мере 12, по меньшей мере 15, по меньшей мере 20, по меньшей мере 30, по меньшей мере 50 или по меньшей мере 100 последовательных и/или не являющихся последовательными аминокислот белковой последовательности. Фрагмент белковой последовательности, предпочтительно, содержит по меньшей мере 6, в частности по меньшей мере 8, по меньшей мере 12, по меньшей мере 15, по меньшей мере 20, по меньшей мере 30, по меньшей мере 50 или по меньшей мере 100 последовательных аминокислот белковой последовательности. Участок, часть или фрагмент структуры, предпочтительно, содержит одно или более функциональных свойств, например, антигенных, иммунологических и/или связывающих свойств указанной структуры. Например, участок вариабельной области цепи Т-клеточного рецептора, предпочтительно, способен образовывать сайт распознавания антигена и связывать антиген. Так, если вариабельная область цепи Т-клеточного рецептора представляет собой V альфа, ее участок, предпочтительно, все еще способен взаимодействовать с соответствующей областью V бета или ее участком с образованием функционального сайта распознавания антигена. Если вариабельная область цепи Т-клеточного рецептора представляет собой V бета, ее участок, предпочтительно, все еще способен взаимодействовать с соответствующей областью V альфа или ее участком с образованием функционального сайта распознавания антигена. Подобным образом участок константной области цепи Т-клеточного рецептора, предпочтительно, способен осуществлять свою функцию передачи сигнала.
Согласно изобретению антиген (по существу) не экспрессируется в клетке, если уровень экспрессии ниже предела обнаружения и/или если уровень экспрессии является слишком низким для того, чтобы допустить связывание антигенспецифическими антителами, добавленными к клетке. Согласно настоящему изобретению антиген экспрессируется в клетке, если уровень экспрессии выше предела обнаружения и/или если уровень экспрессии является достаточно высоким для того, чтобы допустить связывание антигенспецифическими антителами, добавленными к клетке. Предпочтительно, антиген, экспрессируемый в клетке, экспрессируется или экспонируется, т.е. присутствует, на поверхности указанной клетки и, следовательно, является доступным для связывания антигенспецифическими молекулами, например, такими как антитела или антигенные рецепторы, добавленные к клетке.
"Клетка-мишень" должна означать клетку, которая является мишенью для иммунного ответа, такого, как клеточный иммунный ответ. Клетки-мишени включают любую нежелательную клетку, такую как раковая клетка. Согласно предпочтительным вариантам клетка-мишень означает клетку, экспрессирующую целевой антиген, в частности, антиген, специфический в отношении конкретного заболевания, который, предпочтительно, находится на поверхности клетки.
Термин "эпитоп" относится к антигенной детерминанте в молекуле, такой как антиген, т.е. к части или фрагменту молекулы, которая (-ый) распознается, т.е. связывается, иммунной системой, например, распознается антителом или антигенным рецептором. Например, эпитопы представляют собой дискретные трехмерные сайты на антигене, которые распознаются иммунной системой. Эпитопы обычно состоят из химически активных поверхностных группировок молекул, таких как боковые цепи аминокислот или Сахаров, и обычно имеют специфические структурные характеристики трехмерных структур, а также специфические характеристики заряда. Конформационные и не-конформационные эпитопы различаются тем, что связывание с первым, но не с последним, утрачивается в присутствии денатурирующих растворителей. Предпочтительно, эпитоп способен вызывать иммунный ответ на антиген или клетку, экспрессирующую антиген. Предпочтительно, термин относится к иммуногенному участку антигена. Эпитоп белка, например, такого как опухолевый антиген, предпочтительно, содержит непрерывный или прерывистый участок указанного белка и имеет, предпочтительно, от 5 до 100, предпочтительно, от 5 до 50, более предпочтительно, от 8 до 30, наиболее предпочтительно, от 10 до 25 аминокислот в длину, например, эпитоп может иметь в длину, предпочтительно, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 аминокислот.
"Процессирование антигена" относится к расщеплению антигена до продуктов процессирования, которые являются фрагментами указанного антигена (например, расщепление белка до пептидов) и ассоциацию одного или более таких фрагментов (например, посредством связывания) с молекулами МНС для презентации клетками, предпочтительно, антигенпрезентирующим клетками, специфическим Т-клеткам.
Антигенпрезентирующая клетка (АРС) означает клетку, которая представляет антиген в контексте главного комплекса гистосовместимости (МНС) на своей поверхности. Т-клетки могут распознавать этот комплекс, используя свой Т-клеточный рецептор (TCR). Антигенпрезентирующие клетки процессируют антигены и представляют их Т-клеткам. Согласно изобретению термин "антигенпрезентирующая клетка" включает профессиональные антигенпрезентирующие клетки и непрофессиональные антигенпрезентирующие клетки.
Профессиональные антигенпрезентирующие клетки являются очень эффективными при интернализации антигена, либо посредством фагоцитоза, либо посредством опосредованного рецептором эндоцитоза, и последующей визуализации фрагмента антигена, связанного с молекулой класса II МНС на своей мембране. Т-клетка распознает комплекс антиген-молекула класса II МНС на мембране антигенпрезентирующей клетки и взаимодействует с ним. Затем антигенпрезентирующая клетка вырабатывает дополнительный костимулирующий сигнал, приводящий к активации Т-клетки. Экспрессия костимулирующих молекул является определяющим признаком профессиональных антигенпрезентирующих клеток. Основными типами профессиональных антигенпрезентирующих клеток являются дендритные клетки, которые имеют широчайший диапазон презентации антигена и, возможно, являются самыми важными антигенпрезентирующими клетками, макрофаги, В-клетки и некоторые активированные эпителиальные клетки.
Непрофессиональные антигенпрезентирующие клетки не экспрессируют постоянно белки МНС класса II, необходимые для взаимодействия с наивными Т-клетками; эти белки экспрессируются только в результате стимуляции непрофессиональных антигенпрезентирующих клеток некоторыми цитокинами, например, такими как IFNγ.
Дендритные клетки (DCs) представляют собой популяции лейкоцитов, которые презентируют антигены, захваченные в периферических тканях, Т-клеткам путем презентации антигена МНС как класса II, так и класса I. Хорошо известно, что дендритные клетки являются мощными индукторами иммунных реакций, а активация этих клеток является ключевой стадией для индукции противоопухолевого иммунитета. Дендритные клетки и предшественники можно получать из периферической крови, костного мозга, проникающих в опухоль клеток, окружающих опухоль проникающих в нее клеток, лимфатических узлов, селезенки, кожи, пуповинной крови или из любой другой подходящей ткани или жидкости. Например, дендритные клетки можно дифференцировать ex vivo путем добавления комбинации цитокинов, например, таких как GM-CSF, IL-4, IL-13 и/или TNFa, к культурам моноцитов, отобранных из периферической крови. В качестве альтернативы CD34 положительные клетки, отобранные из периферической крови, пуповинной крови или костного мозга, можно дифференцировать в дендритные клетки, добавляя в культуральную среду комбинации GM-CSF, IL-3, TNFα, CD40 лиганд, LPS, flt3 лиганд и/или другое(-ие) соединение(-я), которые индуцируют дифференцировку, созревание и пролиферацию дендритных клеток. Дендритные клетки условно относят к категории "незрелых" и "зрелых" клеток, которые можно применять как простой способ различать между собой два хорошо изученных фенотипа. Однако эти термины не следует толковать, как исключающие все возможные промежуточные стадии дифференцировки. Незрелые дендритные клетки характеризуются как антигенпрезентирующие клетки с высокой способностью захватывать и процессировать антиген, что коррелирует с высоким уровнем экспрессии Fcγ рецептора и маннозного рецептора. Зрелый фенотип обычно характеризуется пониженной экспрессией этих маркеров, но высоким уровнем экспрессии молекул клеточной поверхности, отвечающих за активацию Т-клеток, например, таких как молекулы класса I и класса II МНС, молекулы адгезии (например, CD54 и CD11) и костимулирующие молекулы (например, CD40, CD80, CD86 и 4-1ВВ). Созревание дендритных клеток называется статусом активации дендритных клеток, при котором такие антигенпрезентирующие дендритные клетки вызывают примирование Т-клеток, тогда как презентация незрелыми дендритными клетками вызывает толерантность. Созревание дендритных клеток обусловлено главным образом биомолекулами с микробными отличительными признаками, детектируемыми естественными рецепторами организма (бактериальная ДНК, вирусная РНК, эндотоксин и т.д.), провоспалительными цитокинами (TNF, IL-1, IFNs), лигированием CD40 на поверхности дендритных клеток посредством CD40L, и веществами, высвобождающимися из клеток, претерпевающих связанную со стрессом клеточную смерть. Дендритные клетки можно получать in vitro культивированием клеток костного мозга с цитокинами, такими как гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF) и фактор некроза опухолей альфа.
Термин "иммуногенность" относится к относительной эффективности стимулирования иммунной реакции антигеном.
Термин "эффекторные функции иммунитета" применительно к настоящему изобретению включает любые функции, опосредованные компонентами иммунной системы, которые обусловливают, например, киллинг больных клеток, таких как опухолевые клетки, или ингибирование роста опухоли и/или ингибирование развития опухоли, включающее ингибирование обсеменения и метастазирование опухоли. Предпочтительно, эффекторные функции иммунитета, применительно к настоящему изобретению, представляют собой опосредованные Т-клетками эффекторные функции. Такие функции включают, в случае хелперной Т-клетки, (CD4+ Т клетки), высвобождение цитокинов, таких как Интерлейкин-2, и/или активацию CD8+ лимфоцитов (CTLs) и/или В-клеток, и, в случае CTL, элиминацию клеток, т.е. клеток, характеризующихся экспрессией антигена, например, посредством апоптоза или опосредованного перфорином лизиса клеток, продуцирование цитокинов, таких как IFN-γ и TNF-α, и специфический цитолитический киллинг экспрессирующих антиген клеток-мишеней.
Термин "иммунореактивная клетка" или "иммунная эффекторная клетка", применительно к настоящему изобретению, относится к клетке, которая проявляет эффекторные функции в процессе иммунной реакции. "Иммунореактивная клетка", предпочтительно, способна связывать антиген, например, такой, как антиген, экспрессированный на поверхности клетки, и опосредовать иммунный ответ. Например, такие клетки секретируют цитокины и/или хемокины, убивают микробы, секретируют антитела, распознают инфицированные или раковые клетки и, необязательно, элиминируют такие клетки. Например, иммунореактивные клетки включают Т-клетки (цитотоксические Т-клетки, хелперные Т-клетки, Т-клетки, инфильтрирующие опухоль), В-клетки, природные киллерные клетки, нейтрофилы, макрофаги и дендритные клетки. Предпочтительно, применительно к настоящему изобретению, "иммунореактивные клетки" означают Т-клетки, предпочтительно, CD4+ и/или CD8+ Т-клетки. Согласно настоящему изобретению термин "иммунореактивная клетка" также включает клетку, которая может созревать до иммунной клетки (такой как Т-клетка, в частности хелперная Т-клетка или цитолитическая Т-клетка) с помощью соответствующей стимуляции. Иммунореактивные клетки включают CD34+ гемопоэтические стволовые клетки, незрелые и зрелые Т-клетки и незрелые и зрелые В-клетки. Дифференцировка предшественников Т-клеток в цитолитическую Т-клетку, при экспозиции с антигеном, аналогична клональной селекции с использованием механизмов работы иммунной системы.
Предпочтительно, "иммунореактивная клетка" или "иммунная эффекторная клетка" распознает антиген с определенной степенью специфичности, в частности, если он находится на поверхности антигенпрезентирующий клетки или больной клетки, такой как раковая клетка. Предпочтительно, указанное распознавание дает возможность клетке, которая распознает антиген, быть отвечающей или реактивной. Если клетка представляет собой хелперную Т-клетку (CD4+ Т-клетку), такая отвечаемость или реактивность может включать высвобождение цитокинов и/или активацию CD8+ лимфоцитов (CTLs) и/или В-клеток. Если клетка представляет собой CTL, такая отвечаемость или реактивность может включать элиминацию клеток, т.е. клеток, характеризующихся экспрессией антигена, например, посредством апоптоза или опосредованного перфорином лизиса клеток. Согласно изобретению CTL отвечаемость может включать устойчивый перенос кальция, деление клеток, продуцирование цитокинов, таких как IFN-γ и TNF-α, позитивную регуляцию маркеров активации, таких как CD44 и CD69, и специфический цитолитический киллинг экспрессирующих антиген клеток-мишеней. CTL отвечаемость можно также определять, используя искусственный репортер, который точно указывает на CTL отвечаемость. Такие CTL, которые распознают антиген и являются отвечающими или реактивными, в настоящем описании также называются "отвечающие на антиген CTL".
"Лимфоидная клетка" означает клетку, которая, необязательно после соответствующей модификации, например, после переноса Т-клеточного рецептора или антигенного рецептора, способна вызывать иммунный ответ, например, такой как клеточный иммунный ответ, или клетку-предшественник такой клетки, и включает лимфоциты, предпочтительно, Т-лимфоциты, лимфобласты и плазматические клетки. Лимфоидная клетка может представлять собой иммунореактивную клетку или иммунную эффекторную клетку по настоящему описанию. Предпочтительной лимфоидной клеткой является Т-клетка, которая может быть модифицирована для экспрессии Т-клеточного рецептора или антигенного рецептора на поверхности клетки. Согласно одному варианту настоящего изобретения в лимфоидной клетке отсутствует эндогенная экспрессия Т-клеточного рецептора.
Термины "Т-клетка" и "Т-лимфоцит" применяются в данной заявке в качестве синонимов и включают хелперные Т-клетки (CD4+ Т-клетки) и цитотоксические Т-клетки (CTLs, CD8+ Т-клетки), которые включают цитолитические Т-клетки.
Т-клетка относится к группе белых кровяных клеток, известных как лимфоциты, и играет центральную роль в клеточно-опосредованном иммунитете. Их можно отличить от лимфоцитов других типов, таких как В-клетки и натуральные киллерные клетки, по присутствию на их поверхности особых рецепторов, называемых Т-клеточными рецепторами (TCR). Тимус является основным органом, отвечающим за созревание Т-клеток. Было обнаружено несколько различных субпопуляций Т-клеток, каждая с индивидуальной функцией.
Хелперные Т-клетки содействуют другим белым кровяным клеткам в иммунологических процессах, включая, среди прочего, созревание В-клеток до плазматических клеток и активацию цитотоксических Т-клеток и макрофагов. Эти клетки известны также как CD4+ Т клетки, потому что они экспрессируют на своей поверхности белок CD4. Хелперные Т-клетки становятся активированными, когда они представляются с пептидными антигенами молекулами МНС класса II, которые экспрессируются на поверхности антигенпрезентирующих клеток (APCs). После активации они быстро делятся и секретируют малые белки, называемые цитокинами, которые регулируют активный иммунный ответ или содействуют ему.
Цитотоксические Т-клетки разрушают инфицированные вирусами клетки и опухолевые клетки, они также причастны к отторжению трансплантата. Эти клетки также известны как CD8+ Т-клетки, поскольку они на своей поверхности экспрессируют CD8 гликопротеин. Эти клетки распознают свои мишени посредством связывания с антигеном, ассоциированным с МНС класса I, которая присутствует на поверхности почти каждой клетки тела.
Большинство Т-клеток имеет Т-клеточный рецептор (TCR), существующий в виде комплекса нескольких белков. Реальный Т-клеточный рецептор состоит из двух отдельных пептидных цепей, которые получены с использованием независимых генов альфа и бета (TCRα и TCRβ) Т-клеточного рецептора и называются α- и β-TCR цепи, γδ Т-клетки (гамма дельта Т клетки) представляют небольшую субпопуляцию Т-клеток, на поверхности которых имеется отдельный Т-клеточный рецептор (TCR). При этом в γδ Т-клетках TCR состоит из одной γ-цепи и одной δ-цепи. Эта группа Т-клеток является значительно менее распространенной (2% от общего количества Т-клеток), чем αβ Т-клетки.
Каждая цепь Т-клеточного рецептора состоит из двух экстрацеллюлярных (внеклеточных) доменов: вариабельной (V) области и константной (С) области. Константная область является проксимальной к клеточной мембране, за ней следует трансмембранная область и короткий цитоплазматический хвост, тогда как вариабельная область связывается с комплексом пептид/МНС. В контексте настоящего изобретения термин "константная область цепи Т-клеточного рецептора или ее часть" включает также варианты, в которых за константной областью цепи Т-клеточного рецептора (от N-конца к С-концу) следует трансмембранная область и цитоплазматический хвост, например, трансмембранная область и цитоплазматический хвост, которые в естественных условиях связаны с константной областью цепи Т-клеточного рецептора.
Все Т-клетки ведут свое начало от гемопоэтических стволовых клеток в костном мозге. Гемопоэтические предшественники, происходящие из гемопоэтических стволовых клеток, "заселяют" тимус и размножаются путем деления клеток, создавая большую популяцию незрелых тимоцитов. 'Ранние' тимоциты не экспрессируют ни CD4, ни CD8 и по этой причине классифицируются как дважды негативные (CD4-CD8-) клетки. По мере их прогрессирования в ходе развития они становятся дважды позитивными тимоцитами (CD4+CD8+), и наконец созревают до монопозитивных (CD4+CD8- или CD4-CD8+) тимоцитов, которые затем высвобождаются из тимуса в периферические ткани.
Обычно Т-клетки можно получать in vitro или ex vivo, применяя стандартные методы. Например, Т-клетки можно выделять из костного мозга, периферической крови или фракции костного мозга или периферической крови млекопитающего, например, пациента, применяя коммерчески доступную систему сепарации клеток. Или же Т-клетки можно получать от связанных или не связанных родством людей, животных, отличных от человека животных, с использованием клеточных линий или культур. Образец, содержащий Т-клетки, может, например, представлять собой мононуклеарные клетки периферической крови (РВМС).
Т-клетки для применения по изобретению могут экспрессировать эндогенный Т-клеточный рецептор, или могут не экспрессировать эндогенный Т-клеточный рецептор.
Нуклеиновые кислоты, такие как РНК, кодирующие антигенный рецептор, можно ввести в Т-клетки с литическим потенциалом, в частности, в лимфоидные клетки.
Термин "антигенный рецептор, нацеленный на антиген" или аналогичные термины, относятся к антигенному рецептору, который, при его присутствии на иммунной эффекторной клетке, такой как Т-клетка, распознает антиген, который находится на поверхности антигенпрезентирующих клеток или больных клеток, таких как раковые клетки, вследствие чего иммунная эффекторная клетка стимулируется, примируется и/или размножается или воздействует на эффекторные функции иммунных эффекторных клеток, описанных выше.
Термин "антигенспецифическая Т-клетка" или аналогичные термины относятся к Т-клетке, которая, в частности, будучи предоставлена с антигенным рецептором, распознает антиген, на который нацелен антигенный рецептор, например, на поверхности антигенпрезентирующих клеток или больных клеток, таких как раковые клетки, и предпочтительно, воздействует на эффекторные функции Т-клеток, описанные выше.
Считают, что Т-клетки и другие лимфоидные клетки являются специфическими к антигену, если эти клетки убивают клетки-мишени, экспрессирующие антиген. Специфичность Т-клеток можно определять, применяя любые из ряда стандартных методов, например, тест с радиоактивным хромом или анализ пролиферации. В качестве альтернативы можно количественно определять синтез лимфокинов (таких как интерферон -γ).
Термин "главный комплекс гистосовместимости " и сокращение "МНС" включает молекулы МНС класса I и молекулы МНС класса II и относится к комплексу генов, который встречается у всех позвоночных. Белки или молекулы МНС являются существенными для передачи сигнала между лимфоцитами и антигенпрезентирующими клетками или больными клетками в иммунных реакциях, при этом МНС белки или молекулы связывают пептиды и представляют их для распознавания Т-клеточными рецепторами. Белки, кодированные МНС, экспрессируются на поверхности клеток и экспонируются Т клетке как аутоантигены (пептидные фрагменты из самой клетки), так и не аутоантигены (например, фрагменты инвазивных микроорганизмов).
Согласно настоящему изобретению термин "антигенный рецептор" включает генно-инженерные рецепторы, которые сообщают произвольную специфичность, например, такую как специфичность моноклонального антитела, иммунной эффекторной клетке, такой как Т-клетка. В результате этого можно получить большое количество антигенспецифических Т-клеток, которые могут быть созданы для адоптивного переноса клеток. Вследствие этого антигенный рецептор по изобретению может присутствовать на Т-клетках, например, вместо или помимо собственного Т-клеточного рецептора Т-клетки. Такие Т-клетки необязательно требуют процессирования и презентации антигена для распознавания клетки-мишени, но скорее могут распознавать, предпочтительно с использованием специфичности, любой антиген, присутствующий на клетке-мишени. Предпочтительно, указанный антигенный рецептор экспрессируется на поверхности клеток. Применительно к настоящему изобретению Т-клетки, содержащие антигенный рецептор, охватываются термином "Т-клетка" в контексте настоящей заявки. А именно, согласно настоящему изобретению термин "антигенный рецептор" включает искусственные рецепторы, содержащие одну молекулу или комплекс молекул, которые распознают, т.е. связываются с целевой структурой (например, антигеном) на клетке-мишени, такой как раковая клетка (например, посредством связывания антигенсвязывающего сайта или антигенсвязывающего домена с антигеном, экспрессированным на поверхности клетки-мишени), и могут сообщать специфичность иммунной эффекторной клетке, такой как Т-клетка, экспрессирующая указанный антигенный рецептор на поверхности клетки. Предпочтительно, распознавание целевой структуры антигенным рецептором приводит в результате к активации иммунной эффекторной клетки, экспрессирующей указанный антигенный рецептор. Антигенный рецептор может содержать один или более элементов белка, причем указанные элементы белка содержат один или более доменов по настоящему описанию. Термин "антигенный рецептор", предпочтительно, не включает Т-клеточные рецепторы. Согласно изобретению термин "антигенный рецептор", предпочтительно, является синонимом терминов "химерный антигенный рецептор (CAR)", "химерный Т-клеточный рецептор " и "искусственный Т-клеточный рецептор".
Согласно настоящему изобретению антиген может распознаваться антигенным рецептором через посредничество любого из доменов, отвечающих за распознавание антигена (также называемых в данной заявке просто "домены"), способных образовывать антигенсвязывающий сайт, например, с использованием антигенсвязывающих участков антител и Т-клеточных рецепторов, которые могут находиться на разных пептидных цепях. Согласно одному варианту два домена, образующих антигенсвязывающий сайт, получены с использованием иммуноглобулина. Согласно другому варианту два домена, образующие антигенсвязывающий сайт, получены из Т-клеточного рецептора. Особенно предпочтительными являются вариабельные домены антитела, например, такие как одноцепочечные вариабельные фрагменты (scFv), полученные из вариабельных доменов моноклональных антител и Т-клеточных рецепторов, в частности, одиночных цепей TCR альфа и бета. Фактически почти все, что связывает данную мишень с высокой аффинностью, можно применять в качестве антигенраспознающего домена.
Согласно одному варианту изобретения антигенный рецептор по изобретению содержит по меньшей мере четыре вариабельных домена иммуноглобулинов, образующих по меньшей мере два сайта связывания, где два сайта связывания могут связываться с одним и тем же эпитопом или с различными эпитопами, при этом каждый эпитоп может находиться на одном и том же антигене или на различных антигенах. Согласно одному варианту антигенный рецептор содержит вариабельный домен (или область) тяжелой цепи иммуноглобулина (VH) со специфичностью к первому эпитопу (VH(1)), вариабельный домен (или область) легкой цепи иммуноглобулина (VL) со специфичностью к первому эпитопу (VL(1)), вариабельный домен (или область) тяжелой цепи иммуноглобулина (VH) со специфичностью ко второму эпитопу (VH(2)), и вариабельный домен (или область) легкой цепи иммуноглобулина (VL) со специфичностью ко второму эпитопу (VL(2)), при этом первый и второй эпитопы могут быть одинаковыми и различными и могут находиться на одних и тех же или на различных антигенах. Согласно одному варианту VH(1) способен взаимодействовать и образовывать антигенсвязывающий сайт с VL(1), а VH(2) способен взаимодействовать и образовывать антигенсвязывающий сайт с VL(2), в то же время домен VH(1) не способен взаимодействовать и образовывать антигенсвязывающий сайт с VL(2), а домен VH(2) не способен взаимодействовать и образовывать антигенсвязывающий сайт с VL(1). Однако, согласно другому варианту домен VH(1) способен взаимодействовать и образовывать антигенсвязывающий сайт с VL(1), равно как и VL(2), и VH(2) способен взаимодействовать и образовывать антигенсвязывающий сайт с VL(2), равно как и VL(1). Согласно последнему варианту VH(1) и VH(2) могут быть идентичными или по меньшей мере полученными из одного и того же иммуноглобулина, и VL(1) и VL(2) могут быть идентичными или по меньшей мере полученными из одного и того же иммуноглобулина.
В одном аспекте данное изобретение относится к антигенному рецептору, также называемому в настоящем описании комбинированным антигенным рецептором, при этом такой рецептор содержит первую пептидную цепь и вторую пептидную цепь, причем первая пептидная цепь содержит первый домен, второй домен, вариабельную область цепи Т-клеточного рецептора или ее участок и домен передачи сигнала от иммунорецептора; вторая пептидная цепь содержит первый домен, второй домен, вариабельную область цепи Т-клеточного рецептора или ее участок и домен передачи сигнала от иммунорецептора; причем первый домен из первой пептидной цепи вместе с одним из доменов из второй пептидной цепи образует первый антигенсвязывающий сайт, а при этом второй домен из первой пептидной цепи вместе с другим доменом из второй пептидной цепи образует второй антигенсвязывающий сайт.
Согласно другому варианту комбинированный антигенный рецептор по изобретению содержит вариабельный домен тяжелой цепи, связанный с вариабельным доменом легкой цепи на каждой из обеих пептидных цепей, причем образование двух антигенсвязывающих сайтов происходит за счет взаимодействия между вариабельным доменом тяжелой цепи и вариабельным доменом легкой цепи на различных пептидных цепях. Согласно одному варианту комбинированный антигенный рецептор по изобретению содержит две пептидных цепи, причем одна пептидная цепь содержит VL(1) и VH(2), а вторая полипептидная цепь содержит VH(1) и VL(2). Согласно другому варианту комбинированный антигенный рецептор по изобретению содержит вариабельный домен тяжелой цепи, связанный с вариабельным доменом тяжелой цепи, на одной пептидной цепи, и вариабельный домен легкой цепи, связанный с вариабельным доменом легкой цепи, на другой пептидной цепи, при этом образование двух антигенсвязывающих сайтов происходит за счет взаимодействия между вариабельным доменом тяжелой цепи и вариабельным доменом легкой цепи на разных пептидных цепях. Согласно одному варианту комбинированный антигенный рецептор по изобретению содержит две пептидных цепи, причем одна пептидная цепь содержит VH(1) и VH(2), а другая пептидная цепь содержит VL(1) и VL(2).
Согласно одному варианту настоящего изобретения комбинированный антигенный рецептор по изобретению содержит первую пептидную цепь, где вариабельная область тяжелой цепи (VH) и вариабельная область легкой цепи (VL), предпочтительно, расположены, от N- конца к С-концу, в следующем порядке: VH(1)-VL(2), и вторую пептидную цепь, где вариабельная область тяжелой цепи (VH) и вариабельная область легкой цепи (VL), предпочтительно, расположены, от N-конца к С-концу, в следующем порядке: VL(1)-VH(2). Вариабельная область цепи Т-клеточного рецептора или ее участка и домен передачи сигнала от иммунорецептора, предпочтительно, расположены С-терминально к расположению вариабельных областей. Вариабельная область цепи Т-клеточного рецептора или ее участок и домен передачи сигнала от иммунорецептора, предпочтительно, содержит вариабельную область альфа цепи Т-клеточного рецептора или ее участок, и константную область альфа цепи Т-клеточного рецептора или ее участок, расположенные на одной из пептидных цепей, и вариабельную область бета цепи Т-клеточного рецептора или ее участок, и константную область бета цепи Т-клеточного рецептора или ее участок, расположенные на другой пептидной цепи. Согласно одному варианту вариабельная область альфа цепи Т-клеточного рецептора или ее участок и константная область альфа цепи Т-клеточного рецептора или ее участок содержат альфа цепь Т-клеточного рецептора. Согласно одному варианту вариабельная область бета цепи Т-клеточного рецептора или ее участок и константная область бета цепи Т-клеточного рецептора или ее участок содержат бета цепь Т-клеточного рецептора. Альфа цепь Т-клеточного рецептора и бета цепь Т-клеточного рецептора, предпочтительно, происходят из одного и того же Т-клеточного рецептора.
Согласно другому варианту комбинированный антигенный рецептор по изобретению содержит первую пептидную цепь, где вариабельная область тяжелой цепи (VH) и вариабельная область легкой цепи (VL), предпочтительно, расположены, от N-конца к С-концу, в таком порядке: VH(1)-VH(2), и вторую пептидную цепь, где вариабельная область тяжелой цепи (VH) и вариабельная область легкой цепи (VL), предпочтительно, расположены, от N-конца к С-концу, в таком порядке: VL(1)-VL(2). Вариабельная область цепи Т-клеточного рецептора или ее участок и домен передачи сигнала от иммунорецептора, расположены С-терминально к расположению вариабельных областей. Вариабельная область цепи Т-клеточного рецептора, или ее участок, и домен передачи сигнала от иммунорецептора, предпочтительно, содержит вариабельную область альфа цепи Т-клеточного рецептора или ее участок, и константную область альфа цепи Т-клеточного рецептора или ее участок, расположенные на одной из пептидных цепей, и вариабельную область бета цепи Т-клеточного рецептора или ее участок, и константную область бета цепи Т-клеточного рецептора или ее участок, расположенные на другой пептидной цепи. Согласно одному варианту вариабельная область альфа цепи Т-клеточного рецептора или ее участок и константная область альфа цепи Т-клеточного рецептора или ее участок содержат альфа цепь Т-клеточного рецептора. Согласно одному варианту вариабельная область бета цепи Т-клеточного рецептора или ее участок и константная область бета цепи Т-клеточного рецептора или ее участок содержит бета цепь Т-клеточного рецептора. Альфа цепь Т-клеточного рецептора и бета цепь Т-клеточного рецептора, предпочтительно, происходят из одного и того же Т-клеточного рецептора.
Антигенные рецепторы по изобретению имеют по меньшей мере два антигенсвязывающих сайта и, следовательно, являются по меньшей мере двухвалентными. Как отмечалось выше, связывающие сайты антигенных рецепторов по изобретению могут связываться с одним и тем же эпитопом или с разными эпитопами, каковые эпитопы могут находиться на одном и том же антигене или на различных антигенах. Если связывающие сайты связываются с одними и теми же эпитопами, в частности, на одном и том же антигене, два связывающих сайта могут быть идентичными или по существу идентичными и/или могут быть образованы идентичными или по существу идентичными доменами, при этом такие идентичные или по существу идентичные домены могут происходить из одного и того же иммуноглобулина. Если связывающие сайты связываются с различными эпитопами, либо на одном и том же, либо на различных антигенах, два связывающих сайта являются различными и образуются различными доменами, причем такие различные домены могут происходить из различных иммуноглобулинов. В случае таких различных доменов предпочтительно, чтобы домены, имеющие различные эпитопные специфичности, не взаимодействовали или по существу не взаимодействовали друг с другом, т.е. VH(1) не способен взаимодействовать и образовывать антигенсвязывающий сайт с VL(2), a VH(2) не способен взаимодействовать и образовывать антигенсвязывающий сайт с VL(1). Следовательно, VH(1) взаимодействует и образует антигенсвязывающий сайт с VL(1), a VH(2) взаимодействует и образует антигенсвязывающий сайт с VL(2). Если комбинированный антигенный рецептор по данному изобретению содержит две пептидных цепи, причем одна пептидная цепь содержит VH(1) и VL(2), а другая пептидная цепь содержит VH(2) и VL(1), то в результате получают пептидные цепи, не способные образовывать антигенсвязывающие сайты посредством внутримолекулярного взаимодействия доменов.
Два домена антигенного рецептора по изобретению, образующие антигенсвязывающий сайт, также могут иметь своим источником Т-клеточный рецептор и могут представлять собой его фрагменты или участки, которые сохраняют антигенспецифическое связывание, в частности, связывание с комплексом пептид-МНС, как например вариабельные области Т-клеточного рецептора.
Согласно изобретению термин "вариабельная область Т-клеточного рецептора" относится к вариабельным доменам цепей TCR. Вариабельная область как α-цепи, так и β-цепи TCR имеет три гипервариабельных области, или области, определяющей комплементарность (CDRs), тогда как вариабельная область β-цепи имеет дополнительную область гипервариабельности (HV4), которая обычно не контактирует с антигеном и, следовательно, не рассматривается как CDR. CDR3 является основной CDR, отвечающей за распознавание процессированного антигена, хотя также было показано, что CDR1 α-цепи взаимодействует с N-концевой частью антигенного пептида, тогда как CDR1 β-цепи взаимодействует с С-концевой частью пептида. Полагают, что CDR2 распознает МНС. Полагают, что CDR4 β-цепи не участвует в распознавании антигена, но показано, что эта область взаимодействует с суперантигенами.
Приведенная выше информация, относящаяся к вариабельным доменам иммуноглобулина, определенным образом применима к вариабельным доменам Т-клеточного рецептора. Антигенный рецептор по изобретению вместо вариабельного домена тяжелой цепи иммуноглобулина (VH) со специфичностью в отношении первого эпитопа (VH(1)) и вариабельного домена легкой цепи иммуноглобулина (VL) со специфичностью к первому эпитопу (VL(1)) может содержать вариабельный домен α-цепи TCR рецептора TCR со специфичностью к первому эпитопу и вариабельный домен β-цепи TCR рецептора TCR со специфичностью ко второму эпитопу. В качестве альтернативы или помимо этого антигенный рецептор по изобретению вместо вариабельного домена тяжелой цепи иммуноглобулина (VH) со специфичностью ко второму эпитопу (VH(2)), и вариабельного домена легкой цепи иммуноглобулина (VL) со специфичностью ко второму эпитопу (VL(2)) может содержать вариабельный домен α-цепи TCR рецептора TCR со специфичностью ко второму эпитопу и вариабельный домен β-цепи TCR рецептора TCR со специфичностью ко второму эпитопу.
Так как каждый антигенсвязывающий сайт образуется с использованием двух доменов, каждый домен может содержать участок или фрагмент иммуноглобулина или Т-клеточного рецептора, соответственно. Отдельный участок или фрагмент, самостоятельно, не сможет связывать антиген, но, когда два индивидуальных участка или фрагмента связываются, они совместно образуют или воссоздают антигенсвязывающую структуру оригинального иммуноглобулина или Т-клеточного рецептора и, вследствие этого, способны связывать тот же самый антиген, предпочтительно, с такой же аффинностью.
После распознавания антигена рецепторы, предпочтительно, кластеризуются и сигнал передается в клетку. В этом отношении "домен передачи сигнала от иммунорецептора" или "домен передачи сигнала Т-клетке" означает домен, который участвует в передаче активационного сигнала в Т-клетку после связывания антигена. Возможность такой передачи сигнала обеспечивается антигенными рецепторами по изобретению, содержащими константную или консервативную область цепи Т-клеточного рецептора или константную или консервативную область Fc цепи иммунного рецептора или участок константной или консервативной области, такой как константная область альфа цепи Т-клеточного рецептора или ее участок, на одной пептидной цепи, или содержащими константную или консервативную область цепи Т-клеточного рецептора или константную или консервативную область Fc цепи иммунного рецептора или участок константной или консервативной области, такой как константная область бета цепи Т-клеточного рецептора или ее участок, на другой пептидной цепи. При этом комплекс CD3 обозначает антиген, который экспрессируется на зрелых человеческих Т-клетках, тимоцитах и субпопуляции естественных клеток-киллеров как часть мультимолекулярного Т-клеточного рецепторного (TCR) комплекса. Корецептор Т-клетки представляет собой белковый комплекс и состоит из четырех отдельных цепей. У млекопитающих комплекс содержит CD3γ цепь, CD3δ цепь и две CD3ε цепи. Эти цепи ассоциируются с Т-клеточным рецептором (TCR) и с ζ-цепью, генерируя активационный сигнал в Т-лимфоцитах. Молекулы TCR, ζ-цепи и CD3, совместно, составляют TCR комплекс. CD3 отвечает за сигнальную трансдукцию TCR. Как описано у Lin and Weiss, Journal of Cell Science 114, 243-244 (2001), активация TCR комплекса посредством связывания МНС-презентированных специфических антигенных эпитопов приводит к фосфорилированию иммунорецепторных тирозиновых активационных мотивов (ITAMs) киназами семейства Src, запускающему рекрутинг других киназ, влечет за собой активацию Т-клеток, включая высвобождение Са2+. Кластеризация CD3 на Т клетках, например, с помощью иммобилизованных антител к CD3, ведет к активации Т-клеток, аналогичной вовлечению Т-клеточного рецептора, но независимо от обычной специфичности их клона.
Домен передачи сигнала антигенного рецептора, как минимум, предпочтительно, служит для взаимодействия с нативным комплексом сигнальной трансдукции клетки, например, CD3 комплексом, который отвечает за передачу сигнала связывания антигена с антигенным рецептором внутрь клетки, что приводит к активации иммунной клетки. Идентичность домена передачи сигнала ограничена только тем, что он обладает способностью взаимодействовать с нативным комплексом сигнальной трансдукции для индукции активации иммунной клетки при связывании антигена с антигенным рецептором.
Предпочтительно, домен передачи сигнала на одной пептидной цепи образует димер с доменом передачи сигнала на второй цепи, например, за счет дисульфидных мостиков. Предпочтительные домены передачи сигнала могут содержать константную или консервативную область цепи Т-клеточного рецептора или константную или консервативную область цепи Fc-рецептора иммунной клетки или участок константной или консервативной области. Предпочтительные домены передачи сигнала могут содержать альфа, бета, гамма или дельта цепи Т-клеточного рецептора или его участка, а также D2 или D3 консервативные области константного домена Fc рецептора иммунной клетки или их участок. Согласно предпочтительному варианту первая пептидная цепь содержит константную область альфа цепи Т-клеточного рецептора или ее участок, а вторая пептидная цепь содержит константную область бета цепи Т-клеточного рецептора или ее участок. Согласно данному варианту первая пептидная цепь предпочтительно содержит вариабельную область альфа цепи Т-клеточного рецептора или ее участок, а вторая пептидная цепь предпочтительно содержит вариабельную область бета цепи Т-клеточного рецептора или ее участок, причем вариабельные области расположены на N-конце константных областей. Или же первая пептидная цепь предпочтительно содержит константную область бета цепи Т-клеточного рецептора или ее участок, а вторая пептидная цепь предпочтительно содержит константную область альфа цепи Т-клеточного рецептора или ее участок. Согласно данному варианту первая пептидная цепь предпочтительно содержит вариабельную область бета цепи Т-клеточного рецептора или ее участок, а вторая пептидная цепь предпочтительно содержит вариабельную область альфа цепи Т-клеточного рецептора или ее участок, причем вариабельные области расположены на N-конце константных областей. Согласно другому варианту первая пептидная цепь содержит константную область гамма цепи Т-клеточного рецептора или ее участок, а вторая пептидная цепь содержит константную область дельта цепи Т-клеточного рецептора или ее участок. Согласно этому варианту первая пептидная цепь предпочтительно содержит вариабельную область гамма цепи Т-клеточного рецептора или ее участок, а вторая пептидная цепь содержит вариабельную область дельта цепи Т-клеточного рецептора или ее участок, причем вариабельные области расположены на N-конце константных областей. Или же первая пептидная цепь содержит константную область дельта цепи Т-клеточного рецептора или ее участок, а вторая пептидная цепь содержит константную область гамма цепи Т-клеточного рецептора или ее участок. Согласно этому варианту первая пептидная цепь предпочтительно содержит вариабельную область дельта цепи Т-клеточного рецептора или ее участок, а вторая пептидная цепь содержит вариабельную область гамма цепи Т-клеточного рецептора или ее участок, при этом вариабельные области расположены на N-конце константных областей. Необязательно, домены передачи сигнала или вариабельную область цепи Т-клеточного рецептора или ее участок можно модифицировать таким образом, чтобы между цепями создавались дополнительные дисульфидные связи, что приводит к более эффективному образованию и к повышенной стабильности димера.
Не желая ограничиваться конкретным механизмом действия, полагают, что две пептидных цепи антигенного рецептора по изобретению, будучи экспрессированы на поверхности иммунной клетки, образуют димер вследствие взаимодействия (например, связывания дисульфидной связью) по меньшей мере между отдельными доменами передачи сигнала от иммунорецептора на двух цепях, а также образуют комплекс с эндогенным CD3 комплексом, участвующим в физиологическом процессе сигнальной трансдукции от Т-клеточного рецептора. Однако изобретение может также включать непосредственное слияние с CD3ζ или любым другим доменом передачи сигнала в иммунную клетку (CD3, CD3 субъединица FcγR) вместо TCR Сα и Cβ-доменов. Полагают, что при связывании антигена сигнал передается внутрь клетки, приводя к активации иммунной клетки и к формированию антигенспецифического иммунного ответа. Также полагают, что межцепное связывание обусловливает более устойчивый модуль сигнальной трансдукции антиген-антигенный рецептор-эндогенный CD3, повышенная стабильность которого, в свою очередь, обеспечивает более эффективную стимуляцию антигенспецифического иммунного ответа, по сравнению с одновалентными рецепторами и двухвалентными рецепторами, способными лишь к внутрицепному связыванию с антигеном. Полагают, что эта повышенная стабильность дает возможность осуществить вариант применения доменов передачи сигнала от иммунорецептора только человеческого происхождения (например, с минимальной заменой или без замен в человеческой аминокислотной последовательности на аминокислотную последовательность из организма другого вида, например, мыши). Таким образом можно избежать любого возможного нежелательного иммунного ответа на сам антигенный рецептор.
Антигенные рецепторы согласно изобретению или их пептидные цепи, помимо доменов, образующих антигенсвязывающие сайты, вариабельных областей цепи Т-клеточного рецептора или их участка, доменов передачи сигнала от иммунорецептора, включающих CD3ζ, или любого другого домена передачи сигнала в иммунную клетку, могут также содержать один или более костимулирующих доменов. Костимулирующие домены служат для усиления пролиферации и повышения выживаемости Т-клеток, таких как цитотоксические Т-клетки, при связывании антигенного рецептора с компонентом-мишенью. Идентичность костимулирующих доменов ограничивается тем, что они обладают способностью усиливать клеточную пролиферацию и повышать выживаемость при связывании компонента-мишени антигенным рецептором. Соответствующие костимулирующие домены включают CD28, CD137 (4-1ВВ), члена семейства рецептора фактора некроза опухоли (TNF), CD134 (ОХ40), члена TNFR-суперсемейства рецепторов, и CD278 (ICOS), костимулирующую молекулу CD28-суперсемейства, экспрессируемую на активированных Т-клетках. Специалисту в данной области очевидно, что варианты последовательности этих вышеуказанных костимулирующих доменов можно применять, не оказывая отрицательное влияние на изобретение, поскольку варианты обладают такой же или сходной активностью, что и домен, на основании которого они смоделированы. Такие варианты обладают идентичностью последовательностей по меньшей мере около 80% с аминокислотной последовательностью домена, с использованием которого они образованы. Согласно некоторым вариантам изобретения конструкции антигенных рецепторов или их пептидные цепи содержат два костимулирующих домена. Хотя отдельные комбинации включают все возможные варианты четырех отмеченных доменов, конкретные примеры включают CD28+CD137 (4-1ВВ) и CD28+CD134 (ОХ40).
Антигенные рецепторы по настоящему изобретению или их пептидные цепи могут содержать один или более костимулирующих доменов и домены передачи сигнала от иммунорецептора, связанные в направлении от N-конца к С-концу. Однако антигенные рецепторы по настоящему изобретению или их пептидные цепи не ограничиваются расположением в таком порядке, и другие порядки расположения приемлемы и включают домены передачи сигнала от иммунорецептора и один или более костимулирующих доменов.
Понятно, что поскольку домены, образующие антигенсвязывающие сайты, должны быть свободными, чтобы связывать антиген, местоположение этих доменов в слитом белке обычно является таким, чтобы добиться экспонирования области на наружной поверхности клетки. Аналогично, поскольку костимулирующие домены и домены передачи сигнала от иммунорецептора способствуют стимуляции активности и пролиферации Т-клеток, слитый белок обычно экспонирует эти домены во внутренней части клетки. Антигенные рецепторы могут включать дополнительные элементы, такие как сигнальный пептид для обеспечения корректного экспорта слитого белка на поверхность клеток, трансмембранный домен для гарантии того, чтобы слитый белок сохранялся в виде интегрального мембранного белка, и шарнирный домен (или спейсерную область), который придает гибкость доменам, образующим антигенсвязывающие сайты и обеспечивает прочную связь с антигеном.
Необязательно, антигенные рецепторы по изобретению могут также содержать линкер, каковой линкер может представлять собой произвольную аминокислотную последовательность или другое химическое соединение, применимое в качестве спейсера между аминокислотными последовательностями. Линкер обычно предназначен для того, чтобы обеспечить гибкость и устойчивость к протеазам. Например, линкер может находиться между первым и вторым доменами на первой пептидной цепи и/или между первым и вторым доменами на второй пептидной цепи комбинированного антигенного рецептора по изобретению. Необязательно линкер может находиться между доменами, которые образуют антигенсвязывающие сайты, и вариабельной областью цепи Т-клеточного рецептора или ее участком. Любой тип линкера, известный в уровне техники, который позволяет доменам образовывать антигенсвязывающий сайт или не препятствует связыванию антигена, охватывается изобретением. Согласно конкретным вариантам линкер может представлять собой произвольную аминокислотную последовательность и его длина может составлять по меньшей мере 5, 10, 15, 20, 25 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95 или по меньшей мере 100 аминокислотных остатков. Аминокислотный линкер обычно обогащен глицином для гибкости, а также серином и треонином для растворимости. Согласно одному варианту линкер представляет собой один или более (1, 2, 3, 4, 5, 6, 7, 8 или 9) повторов четырех глициновых остатков с последующим сериновым остатком (Gly4Ser). Согласно некоторым вариантам линкер представляет собой шарнирную область антитела или ее фрагмент.
Антигенный рецептор по изобретению может также содержать другой домен, заякоривающий антигенный рецептор на мембране, например, такой как классический трансмембранный домен. Предпочтительно, трансмембранный домен включен в домен передачи сигнала или является его частью.
Согласно другим вариантам антигенные рецепторы или пептидные цепи антигенных рецепторов по изобретению могут также содержать другие домены, такие как дополнительные домены, участвующие в связывании антигена или стимулирующие связывание антигена, сигнальные последовательности для связанной с мембраной экспрессии или для секреции, домены, которые обеспечивают повышенную димеризацию, и трансмембранный домен, когда он уже не является частью домена передачи сигнала от иммунорецептора. Согласно некоторым вариантам трансмембранный домен может представлять собой гидрофобную альфа-спираль, которая пронизывает мембрану.
Предпочтительно, сигнальная последовательность или сигнальный пептид представляет собой последовательность или пептид, которая(-ый) обеспечивает возможность адекватного прохождения через секреторный путь и экспрессию на поверхности клетки таким образом, чтобы антигенный рецептор, например, мог связывать антиген, находящийся во внеклеточной среде. Предпочтительно, сигнальная последовательность или сигнальный пептид способна (или способен) отщепляться и удаляется из зрелых пептидных цепей. Сигнальная последовательность или сигнальный пептид, предпочтительно, выбрана (или выбран) с учетом клетки или организма, в которой (в котором) продуцируются пептидные цепи.
Согласно конкретному варианту настоящего изобретения пептидная цепь комбинированного антигенного рецептора по изобретению может представлять собой структуру: NH2-сигнальный пептид - первый домен, участвующий в связывании антигена - необязательный линкер - второй домен, участвующий в связывании антигена - необязательный линкер - вариабельная область цепи Т-клеточного рецептора или ее участок-домен передачи сигнала от иммунорецептора- СООН.
Примеры антигенных рецепторов согласно данному изобретению включают, но без ограничения, рецепторы, образованные первой и второй пептидными цепями, имеющими структуру, показанную в Таблице I ниже (VH обозначает вариабельную область тяжелой цепи иммуноглобулина или ее участок; VL обозначает вариабельную область легкой цепи иммуноглобулина или ее участок; V1 и V2 являются вариабельными областями цепи Т-клеточного рецептора или ее участками, которые образуют димер друг с другом, С1 и С2 являются доменами передачи сигнала иммунорецептора, которые образуют димер друг с другом, например, константную или консервативную область цепи Fc иммунного рецептора или константную или консервативную область цепи Т-клеточного рецептора или часть константной или консервативной области). Если С1 и С2 каждый являются константными областями цепи Т-клеточного рецептора, V1 и С1 происходят из одной и той же цепи Т-клеточного рецептора, и V2 и С2 предпочтительно происходят из той же самой цепи Т-клеточного рецептора, при этом цепи Т-клеточного рецептора предпочтительно происходят из одного и того же Т-клеточного рецептора. В частности, согласно одному из вариантов V1-С1 по существу соответствует последовательности цепи Т-клеточного рецептора (TCR альфа или TCR бета) и V2-C2 по существу соответствует последовательности комплементарной цепи Т-клеточного рецептора (TCR бета, если V1-С1 является TCR альфа, или TCR альфа, если V1-С1 является TCR бета).

Как указано выше, антигенный рецептор содержит вариабельный домен тяжелой цепи иммуноглобулина (VH) со специфичностью к первому эпитопу (VH(1)), вариабельный домен легкой цепи иммуноглобулина (VL) со специфичностью к первому эпитопу (VL(1)), вариабельный домен тяжелой цепи иммуноглобулина (VH) со специфичностью ко второму эпитопу (VH(2)) и вариабельный домен легкой цепи иммуноглобулина (VL) со специфичностью ко второму эпитопу (VL(2)), при этом первый и второй эпитопы могут быть одинаковыми или разными и могут быть расположены на одном и том же или разных антигенах. Согласно одному варианту VH(1) способен взаимодействовать и образовывать антигенсвязывающий сайт с VL(1) и VH(2) способен взаимодействовать и образовывать антигенсвязывающий сайт с VL(2), в то время как VH(1) не способен взаимодействовать и образовывать антигенсвязывающий сайт с VL(2) и VH(2) не способен взаимодействовать и образовывать антигенсвязывающий сайт с VL(1). Однако согласно другому варианту VH(1) способен взаимодействовать и образовывать антигенсвязывающий сайт с VL(1), а также VL(2), и VH(2) способен взаимодействовать и образовывать антигенсвязывающий сайт с VL(2), а также с VL(1). Согласно последнему варианту VH(1) и VH(2) могут быть идентичными или по меньшей мере происходить из одного и того же иммуноглобулина, и VL(1) и VL(2) могут быть идентичными или по меньшей мере происходить из одного и того же иммуноглобулина.
Согласно конкретным вариантам домены V1 и V2 первой и второй пептидных цепей, указанные в Таблице 1, являются вариабельными областями цепей альфа и бета Т-клеточного рецептора, соответственно, или их участками. Согласно конкретным вариантам домены С1 и С2 первой и второй пептидных цепей, перечисленные в Таблице 1, представляют собой константные области альфа цепей и бета цепей Т-клеточного рецептора, соответственно, или их участками.
Согласно одному из вариантов домены V1 и С1 первой пептидной цепи, перечисленные в Таблице 1, являются вариабельной и константной областями альфа цепи Т-клеточного рецептора или ее участками. В этом варианте домены V2 и С2 второй пептидной цепи, перечисленные в Таблице 1, предпочтительно являются вариабельной и константной областями бета цепи Т-клеточного рецептора или ее участками.
Согласно одному из вариантов домены V1 и С1 первой пептидной цепи, перечисленные в Таблице 1, являются вариабельной и константной областями бета цепи Т-клеточного рецептора или ее участками. В этом варианте домены V2 и С2 второй пептидной цепи, перечисленные в Таблице 1, предпочтительно являются вариабельной и константной областями альфа цепи Т-клеточного рецептора или ее участками.
Согласно предпочтительному варианту, когда два домена одной пептидной цепи оба представляют собой вариабельную область тяжелой цепи иммуноглобулина или ее участок и два домена другой цепи оба представляют собой вариабельную область легкой цепи иммуноглобулина или ее участок, между первым и вторым доменом обеих пептидных цепей находится линкер. Линкер может быть произвольной аминокислотной последовательностью длиной между 10 и 25 аминокислотами, более предпочтительно длиной 15 аминокислот. Согласно конкретному варианту линкер содержит 3 повтора 5-мерной аминокислотной последовательности (Gly4Ser).
Согласно некоторым вариантам данного изобретения аминокислотные последовательности первой и второй пептидных цепей, такие как содержащие один или более доменов, которые образуют антигенсвязывающие сайты, вариабельную область цепи Т-клеточного рецептора или ее часть или домен передачи сигнала иммунорецептора, являются последовательностями млекопитающих, предпочтительно мыши, и более предпочтительно имеют человеческое происхождение. Согласно одному из вариантов аминокислотные последовательности имеют человеческое происхождение, но в них были введены мышиные последовательности путем замены одной или более аминокислот в человеческой последовательности аминокислотой, находящейся в соответствующем положении в мышиной последовательности. Такая замена может способствовать в большей степени димеризации или стабильности, или способности передавать сигнал в клетку после связывания антигена. Согласно еще одному варианту аминокислотные последовательности имели мышиное происхождение и были гуманизированы.
Согласно настоящему изобретению антигенный рецептор может заменять функцию Т-клеточного рецептора, как описано выше, и в частности, может придавать реакционноспособность, например, цитолитическую активность клетке, такой как Т-клетка, как описано выше. Однако в противоположность связыванию Т-клеточного рецептора с комплексом антигенный пептид-МНС, как описано выше, антигенный рецептор согласно некоторым вариантам может связываться с антигеном, в особенности, когда экспрессирован на клеточной поверхности.
Аминокислотные последовательности пептидных цепей, включая любой из доменов или линкеров, могут быть модифицированы. Например, как очевидно для специалистов в данной области, последовательности вариабельных областей антител и Т-клеточных рецепторов могут быть модифицированы без потери способности связываться с мишенью, и вследствие этого аминокислотная последовательность антигенсвязывающих сайтов может быть подобным образом модифицирована без потери способности связываться с мишенью. Например, аминокислотная последовательность домена, образующего антигенсвязывающий сайт, может быть идентичной или высокогомологичной вариабельной области антитела, из которого она происходит. Под "высокогомологичной" подразумевается последовательность, в которой могут быть сделаны от 1 до 5, предпочтительно от 1 до 4, например, от 1 до 3 замен или 1 или 2 замены. Согласно одному варианту пептидная цепь может включать природные аминокислоты и неприродные аминокислоты. Согласно другому варианту пептидная цепь просто включает природные аминокислоты. Термин "неприродная аминокислота" относится к аминокислоте, имеющей структуру, отличающуюся от структур 20 природных аминокислот. Поскольку неприродные аминокислоты имеют структуру, похожую на структуру природных аминокислот, неприродные аминокислоты могут классифицироваться как производные или аналоги рассматриваемых природных аминокислот.
Согласно одному варианту аминокислотная последовательность одной или более вариабельных областей Т-клеточных рецепторов или их частей, в особенности не образующая антигенсвязывающие сайты, может быть модифицирована для удаления (остаточного) связывания с ее антигеном. В частности, такая "сайленсирующая" модификация может быть проведена путем интродукции одной или более мутаций в CDR3 вариабельной области TCR альфа и/или TCR бета.
Настоящее изобретение также охватывает производные антигенных рецепторов и пептидных цепей, описанных в данной заявке. Согласно изобретению "производные" представляют собой модифицированные формы белков и пептидов. Такие модификации включают любую химическую модификацию и включают единичную или многие замены, делеции и/или добавления любых молекул, ассоциированных с антигенным рецептором или пептидной цепью, таких как углеводы, липиды и/или белки или пептиды. Согласно одному варианту "производные" белков или пептидов включают модифицированные аналоги, полученные гликозилированием, ацетилированием, фосфорилированием, амидированием, пальмитоилированием, миристоилированием, изопренилированием, липидированием, алкилированием, дериватизацией, введением защитных/блокирующих групп, протеолитическим расщеплением или связыванием с антигеном. Термин "производное" также распространяется на все функциональные химические эквиваленты указанных антигенных рецепторов и пептидных цепей. Предпочтительно, когда модифицированные антигенный рецептор или его пептидная цепь обладают более прочным связыванием или способностью к димеризации и/или повышенной активации иммунной системы.
Клетки, используемые в связи с системой антигенных рецепторов по данному изобретению, предпочтительно являются Т-клетками, в частности цитотоксическими лимфоцитами, предпочтительно выбранными из цитотоксических Т-клеток, естественных киллерных клеток (NK-клеток) и лимфокин-активированных киллерных клеток (LAK). После активации каждый из этих цитотоксических лимфоцитов запускает деструкцию клеток-мишеней. Например, цитотоксические Т-клетки запускают деструкцию клеток-мишеней одним или обоими следующими способами. Во-первых, после активации Т-клетки высвобождают цитотоксины, такие как перфорин, гранзимы и гранулизин. Перфорин и гранулизин создают поры в клетке-мишени, и гранзимы попадают в клетку и запускают каспазный каскад в цитоплазме, который индуцирует апоптоз (программированную клеточную гибель). Во-вторых, апоптоз может быть индуцирован через взаимодействие системы лиганда Fas-Fas между Т-клетками и клетками-мишенями. Цитотоксические лимфоциты предпочтительно представляют собой аутологичные клетки, хотя могут быть использованы гетерологичные или аллогенные клетки.
Термин "иммуноглобулин" относится к белкам суперсемейства иммуноглобулинов, предпочтительно к антигенным рецепторам, таким как антитела или В-клеточный рецептор (BCR). Иммуноглобулины характеризуются структурным доменом, то есть, доменом иммуноглобулина, имеющим характерную укладку цепей иммуноглобулина (Ig). Этот термин охватывает иммуноглобулины, связанные с мембраной, а также растворимые иммуноглобулины. Связанные с мембранами иммуноглобулины называются также поверхностными иммуноглобулинами или мембранными иммуноглобулинами, которые обычно являются частью BCR. Растворимые иммуноглобулины обычно называются антителами. Иммуноглобулины обычно содержат несколько цепей, обычно две идентичные тяжелые цепи и две идентичные легкие цепи, которые соединены дисульфидными связями. Эти цепи в основном состоят из доменов иммуноглобулина, таких как the VL (вариабельный домен легкой цепи), CL (константный домен легкой цепи) и CH (константная область тяжелой цепи) доменов CH1, CH2, CH3 и CH4. Существуют пять типов тяжелых цепей иммуноглобулинов у млекопитающего, а именно, α, δ, ε, γ и μ, которые объясняют наличие разных классов антител, а именно, IgA, IgD, IgE, IgG и IgM. В противоположность тяжелым цепям растворимых иммуноглобулинов тяжелые цепи мембранных или поверхностных иммуноглобулинов содержат трансмембранный домен и короткий цитоплазматический домен на своем карбокси-конце. У млекопитающих имеются два типа легких цепей, а именно, лямбда и каппа. Цепи иммуноглобулинов содержат вариабельную область и константную область. Константная область является по существу консервативной у различных изотипов иммуноглобулинов, при этом вариабельная часть является в высшей степени разнообразной и отвечает за распознавание антигена.
Термин "антитело" относится к гликопротеину, содержащему по меньшей мере две тяжелых (Н) цепи и две легких (L) цепи, взаимосвязанные с помощью дисульфидных связей. Термин "антитело" включает моноклональные антитела, рекомбинантные антитела, человеческие антитела, гуманизированные антитела и химерные антитела. Каждая тяжелая цепь содержит вариабельную область тяжелой цепи (обозначаемую в данной заявке как VH) и константную область тяжелой цепи. Каждая легкая цепь содержит вариабельную область легкой цепи (обозначаемую в данной заявке как VL) и константную область легкой цепи. VH и VL области могут быть подразделены далее на области гипервариабельности, называемые областями, определяющими комплементарность (CDR), между которыми находятся области, которые являются более консервативными и называются каркасными областями (FR). Каждая VH и VL состоит из трех CDRs и четырех FRs, расположенных в направлении от амино-конца к карбокси-концу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области тяжелых и легких цепей содержат связывающий домен, который взаимодействует с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (Clq) классической системы комплемента.
Термин "моноклональное тело", используемый в данной заявке, относится к популяции молекул антитела одного молекулярного состава. Моноклональное антитело проявляет одну специфичность связывания и аффинность. Согласно одному варианту моноклональные антитела продуцируются гибридомой, которая включает В-клетку, полученную от животного, не относящегося к человеческому роду, например, от мыши, слитую с иммортализованной клеткой.
Термин "рекомбинантное антитело", используемый в данной заявке, включает все антитела, которые получены, экспрессированы, созданы или выделены рекомбинантным методом, например (а) антитела, выделенные из животного (например, мыши), которое является трансгенным или трансхромосомальным по отношению к генам иммуноглобулина или гибридоме, полученной из них, (b) антитела, выделенные из клетки-хозяина, трансформированной для экспрессии антитела, например, из трансфектомы, (с) антитела, выделенные из библиотеки рекомбинантных, комбинаторных антител, и (d) антитела, полученные, экспрессированные, созданные или выделенные любыми другими методами, которые включают сплайсинг последовательностей генов иммуноглобулина в другие последовательности ДНК,
Термин "человеческое антитело", используемый в данной заявке, включает антитела, имеющие вариабельные и константные области, происходящие из последовательностей зародышевых линий иммуноглобулинов человека. Человеческие антитела могут включать аминокислотные остатки, не кодируемые последовательностями зародышевой линии иммуноглобулина человека (например, мутации, интродуцированные случайным или сайт-специфическим мутагенезом in vitro или посредством соматической мутации in vivo).
Термин "гуманизированное антитело" относится к молекуле, имеющей антигенсвязывающий сайт, который по существу происходит из иммуноглобулина видов, не являющихся человеком, где остальная структура молекулы иммуноглобулина основана на структуре и/или последовательности человеческого иммуноглобулина. Антигенсвязывающий сайт может включать или полные вариабельные домены, слитые с константными доменами, или только области, определяющие комплементарность (CDR), привитые на соответствующие каркасные участки в вариабельных доменах. Антигенсвязывающие сайты могут быть дикого типа или модифицированными посредством замены одной или более аминокислот, например, модифицированными для достижения большего сходства с иммуноглобулином человека. Некоторые формы гуманизированных антител сохраняют все CDR последовательности (например, гуманизированное мышиное антитело, которое содержит все шесть CDRs из мышиного антитела). Другие формы содержат одну или более CDRs, которые изменены по сравнению с оригинальным антителом.
Термин "химерное антитело" относится к таким антителам, в которых один участок каждой аминокислотной последовательности тяжелой и легкой цепей является гомологичным соответствующим последовательностям в антителах, происходящих из конкретных видов или принадлежащих к конкретному классу, в то время как оставшийся сегмент цепи гомологичен соответствующим последовательностям из других видов. Обычно вариабельная область и легкой, и тяжелой цепей имитирует вариабельные области антител, происходящих из одного вида млекопитающих, в то время как константные области являются гомологичными последовательностям антител, происходящих из другого вида. Одним несомненным преимуществом химерных антител является то, что вариабельная область может быть легко получена из источников, известных к настоящему времени, с использованием легкодоступных В-клеток или гибридом из организмов хозяев, не относящихся к человеческому роду, в сочетании с константными областями, происходящими из, например, популяций человеческих клеток. В то время как вариабельная область имеет преимущество, заключающееся в легкости ее получения и отсутствии влияния источника на специфичность, константная область, имеющая человеческое происхождение, с меньшей долей вероятности будет вызывать иммунный ответ у человека при инъекции антител по сравнению с константной областью из нечеловеческого источника. Однако определение не ограничивается этим конкретным примером.
Антитела согласно настоящему изобретению могут быть получены из различных видов, включая, но без ограничения, мышь, крысу, кролика, хомячка и человека.
Антитела, описанные в данной заявке, включают IgA, такие как антитела IgA1 или IgA2, IgG1, IgG2, IgG3, IgG4, IgE, IgM и IgD. Согласно различным вариантам антитело представляет собой антитело IgG1, особенно изотипы IgG1, каппа или IgG1, лямбда (то есть, IgG1, κ, λ), антитело IgG2a (например, IgG2a, κ, λ), антитело IgG2b (например, IgG2b, κ, λ), антитело IgG3 (например, IgG3, κ, λ) или антитело IgG4 (например, IgG4, κ, λ).
Антигенные рецепторы, описанные в данной заявке, могут включать антигенсвязывающие участки одного или более антител. Термины "антигенсвязывающий участок" антитела (или просто "участок связывания") или " антигенсвязывающий фрагмент" антитела (или просто "фрагмент связывания") или подобные термины относятся к одному или более фрагментам антитела, которые сохраняют способность специфически связываться с антигеном. Было показано, что функция антитела связывать антиген может осуществляться фрагментами полноразмерного антитела. Примеры связывающих фрагментов, охваченных термином "антигенсвязывающий участок" антитела включают (i) Fab-фрагменты, одновалентные фрагменты, состоящие из доменов VL, VH, CL и СН; (ii) F(ab')2-фрагменты, бивалентные фрагменты, включающие два Fab-фрагмента, соединенные дисульфидным мостиком в шарнирном участке; (iii) Fd-фрагменты, состоящие из доменов VH и СН; (iv) Fv-фрагменты, состоящие из доменов VL и VH одного плеча антитела, (v) dAb-фрагменты (Ward et al., (1989) Nature 341: 544-546), которые состоят из домена VH; (vi) выделенные области, определяющие комплементарность(CDR), и (vii) комбинации двух или более выделенных CDRs, которые могут быть необязательно соединены синтетическим линкером. Кроме того, хотя два домена Fv-фрагмента, VL и VH, кодированы отдельными генами, они могут быть соединены с использованием рекомбинантных методов синтетическим линкером, который обеспечивает им возможность образовывать одну белковую цепь, в которой области VL и VH спариваются с образованием одновалентных молекул (известна как одноцепочечный Fv (scFv); см., например, Bird et al. (1988) Science 242: 423-426; и Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85: 5879-5883). Такие одноцепочечные антитела также охвачены термином "антигенсвязывающий фрагмент" антитела. Другим примером являются слитые белки "связывающий домен-иммуноглобулин", содержащие (i) полипептид связывающего домена, который слит с полипептидом шарнирной области иммуноглобулина (ii) константную область СН2 тяжелой цепи иммуноглобулина, слитую с шарнирным участком, и (iii) константную область СН3 тяжелой цепи иммуноглобулина, слитую с константной областью СН2. Полипептид связывающего домена может быть вариабельной областью тяжелой цепи или вариабельной областью легкой цепи. Слитые белки "связывающий домен-иммуноглобулин" описаны также в заявках США на патент №№2003/0118592 и 2003/0133939. Эти фрагменты антител получены с использованием обычных методов, известных специалистам в данной области, и такие фрагменты подвергали скринингу для определения применимости тем же методом, что и в случае интактных антител.
Одноцепочечный вариабельный фрагмент (scFv) представляет собой белок слияния вариабельных областей тяжелой (VH) и легкой (VL) цепей иммуноглобулинов, соединенных пептидным линкером. Этот линкер может соединять или N-конец VH с С-концом VL, или наоборот.Двухвалентные (или бивалентные) одноцепочечные вариабельные фрагменты (di-scFvs, bi-scFvs) могут быть получены путем связывания двух scFvs. Это может быть сделано путем получения одноцепочечной пептидной цепи с двумя областями VH и двумя областями VL с образованием тандемных scFvs.
Термин "связывающий домен" или просто "домен" в связи с настоящим изобретением характеризуется структурой, например, антитела, которая связывается/взаимодействует с данной (-ым) целевой (-ым) структурой/антигеном/эпитопом, необязательно при взаимодействии с другим доменом. Эти домены согласно данному изобретению называются "антигенсвязывающими сайтами".
Антитела и производные антител пригодны для создания связывающих доменов, таких как фрагменты антитела, в частности для получения областей VL и VH.
Связывающие домены для антигена, которые могут находиться в антигенном рецепторе, обладают способностью к связыванию с антигеном (таргетированию на антиген), то есть, способностью к связыванию с эпитопом (таргетированию на эпитоп), находящимся в антигене, предпочтительно с эпитопом, расположенным в экстрацеллюлярном домене антигена. Предпочтительно, когда связывающие домены для антигена являются специфичными в отношении антигена. Предпочтительно, когда связывающие домены для антигена связываются с антигеном, экспрессированным на клеточной поверхности. Согласно конкретным предпочтительным вариантам связывающие домены для антигена связываются с нативными эпитопами антигена, находящимися на поверхности живых клеток.
Все антитела и производные антител, такие как фрагменты антител, описанные в данной заявке, для целей данного изобретения охватываются термином "антитело".
Антитела можно получать различными методами, включая обычную процедуру с применением моноклональных антител, например, с использованием стандартного метода гибридизации соматических клеток, описанного в публикации Келера и Милстейна, см. Nature 256: 495 (1975). Хотя предпочтительными являются методы гибридизации соматических клеток, в принципе можно использовать другие методы получения моноклональных антител, например, методы вирусной или онкогенной трансформации В-лимфоцитов или метод фагового дисплея с использованием библиотек генов антител.
Предпочтительной системой для получения гибридом, которые секретируют моноклональные антитела, является мышиная система. Продуцирование гибридомы с использованием мыши является хорошо разработанной процедурой. Протоколы иммунизации и методика выделения иммунизированных спленоцитов для слияния известны из уровня техники. Партнеры по слиянию (например, клетки миеломы мыши) и методика слияния также являются известными.
Другими предпочтительными системами животных для продуцирования гибридом, которые секретируют моноклональные антитела, являются крысиная и кроличья системы (например, описанные в публикации Spieker-Polet et al., Proc. Natl. Acad. Sci. U.S.A. 92: 9348 (1995), см. также Rossi et al., Am. J. Clin. Pathol. 124: 295 (2005)).
Для получения антител мыши могут быть иммунизированы при помощи пептидов, конъюгированных с носителями, происходящими из последовательности антигена, то есть, последовательности, против которой должны быть направлены антитела, обогащенной популяцией рекомбинантно экспрессированных антигена или его фрагментов и/или клетками, экспрессирующими антиген, как описано в данной заявке. Альтернативно, мыши могут быть иммунизированы с помощью ДНК, кодирующей антиген или его фрагменты. В случае, когда иммунизация, использующая очищенную или обогащенную популяцию антигена, не приводит к продуцированию антител, мыши могут быть иммунизированы с помощью клеток, экспрессирующих антиген, например, клеточной линии, для промотирования иммунного ответа.
Иммунный ответ может быть подвергнут мониторингу в ходе протокола иммунизации, причем пробы плазмы и сыворотки получают с применением образцов плазмы и сыворотки, полученных из хвостовой вены или при заборе крови методом ретроорбитального кровопускания. Мышей с достаточными титрами иммуноглобулина можно использовать для слияния. Можно осуществить интраперитонеальную или внутривенную реиммунизацию мышей антигеном с экспрессией клеток за 3 дня перед умерщвлением и удалением селезенки для увеличения скорости секретирования гибридомами специфического антитела.
Для генерирования гибридом, продуцирующих моноклональные антитела, спленоциты и/или клетки лимфатических узлов из иммунизированных мышей могут быть выделены и слиты с подходящей иммортализованной клеточной линией, такой как линия мышиных миеломных клеток. Полученные гибридомы могут быть подвергнуты скринингу на продуцирование антигенспецифических антител. Затем отдельные лунки могут быть подвергнуты скринингу при помощи ELISA на гибридомы, секретирующие антитела. При помощи иммунофлуоресцентного метода и FACS-анализа с использованием клеток, экспрессирующих антигены, антитела со специфичностью к антигену могут быть идентифицированы. Гибридомы, секретирующие антитела, могут быть повторно высеяны, снова подвергнуты скринингу и, если они все еще являются положительными в отношении IgG человека, моноклональные антитела могут быть субклонированы с использованием лимитирующего разведения. Стабильные субклоны могут быть затем культивированы in vitro для генерирования антитела для характеристики в среде для культуры ткани.
Способность антител и других агентов связывания к связыванию с антигенами может быть определена с применением стандартных анализов связывания (например, ELISA, вестерн-блоттинга, иммунофлуоресцентного метода и метода проточного цитометрического анализа).
Согласно настоящему изобретению термин "связывание" предпочтительно относится к специфическому связыванию.
Согласно настоящему изобретению агент, такой как рецептор антигена, способен связываться с заданной мишенью (таргетировать на заданную мишень), если он обладает значительной аффинностью в отношении указанной заданной мишени и связывается с указанной заданной мишенью при проведении стандартного анализа. "Аффинность" или "аффинность связывания" часто измеряется при помощи константы равновесия диссоциации (KD). Предпочтительно, когда термин "высокая аффинность" относится к связыванию заданной мишени со значением константы диссоциации (KD) 10-5 М или менее, 10-6 М или менее, 10-7 М или менее, 10-8 М или менее, 10-9 М или менее, 10-10 М или менее, 10-11 М или менее, или 10-12 М или менее.
Агент (по существу) не способен связывать (таргетировать) мишень, если он не имеет высокой аффинности к указанной мишени и не связывается значительно, в особенности не связывается с возможностью определения, с указанной мишенью при проведении стандартного анализа. Предпочтительно, когда такой агент не связывается определимо с указанной мишенью, если присутствует в концентрации до 2, предпочтительно до 10, более предпочтительно до 20, в особенности до 50 или 100 мкг/мл или более. Предпочтительно, когда агент не обладает высокой аффинностью по отношению к мишени, если он связывается с указанной мишенью со значением KD, которое по меньшей мере в 10 раз, 100 раз, 103 раз, 104 раз, 105 раз или 106 раз больше, чем KD при связывании заданной мишени, с которой этот агент способен связываться. Например, если значение KD при связывании агента с мишенью, с которой этот агент способен связываться, составляет 10-7 М, KD при связывании с мишенью, в отношении которой этот агент не имеет значительной аффинности, будет составлять по меньшей мере 10-6 М, 10-5 М, 10-4 М, 10-3 М, 10-2 М или 10-1 M.
Агент является специфическим в отношении заданной мишени, если он способен связываться с указанной заданной мишенью, то есть, не имеет высокой аффинности к другим мишеням и не связывается в значительной степени с другими мишенями при проведении стандартных анализов. Предпочтительно, агент является специфическим по отношению к заданной мишени, если аффинность и связывание с такими другими мишенями не превышает в значительной степени аффинность к или связывание с белками, которые не являются родственными заданной мишени, такими как альбумин бычьей сыворотки (BSA), казеин или альбумин человеческой сыворотки (HSA). Предпочтительно, агент является специфическим по отношению к заданной мишени, если он связывается с указанной мишенью со значением KD, которое по меньшей мерее в 10 раз, 100 раз, 103 раз, 104 раз, 105 раз или 106 раз меньше, чем значение KD при связывании с мишенью, по отношению к которой он не является специфическим. Например, если значение KD при связывании агента с мишенью, по отношению к которой он является специфическим, составляет 10-7 М, величина KD при связывании с мишенью, по отношению к которой он не является специфическим, будет составлять по меньшей мере 10-6 М, 10-5 М, 10-4 М, 10-3 М, 10-2 М или 10-1 М.
Связывание агента с мишенью может быть определено с использованием подходящего метода; см., например, см. публикации Berzofsky et al., "Antibody-Antigen Interactions". In Fundamental Immunology, Paul, W.E., Ed., Raven Press New York, N Y (1984), Kuby, Janis Immunology, W.H. Freeman and Company New York, N Y (1992), и методов, описанных в данной заявке. Аффинность может быть легко определена с помощью стандартных методов, таких как равновесный диализ; с применением биосенсора BIAcore 2000, с использованием обычных методик, разработанных производителем; радиоиммунологическим методом анализа с использованием радиомеченого целевого антигена; или другого метода, известного специалисту в данной области. Результаты определения аффинности можно проанализировать, например, методом, описанным Scatchard et al. в Ann N.Y. Acad. Sci, 51: 660 (1949). Измеренная величина аффинности при взаимодействии конкретных антитела-антигена может варьироваться, если она измерена в разных условиях, например, это могут быть разная концентрация соли, разная величина рН. Таким образом, измерение аффинности и других параметров связывания антигена, например, KD, IC50, предпочтительно проводить с использованием стандартизованных растворов антитела и антигена и стандартизованного буфера.
Настоящее изобретение может включать введение, то есть, трансфекцию, нуклеиновых кислот, кодирующих рецепторы антигенов, в клетки, такие как Т-клетки, in vitro или in vivo.
Для целей данного изобретения термин "трансфекция" включает введение нуклеиновой кислоты в клетку или поглощение нуклеиновой кислоты клеткой, при этом такая клетка может находиться в организме субъекта, например, пациента. Таким образом, согласно настоящему изобретению клетка для трансфекции нуклеиновой кислоты, описанная в данной заявке, может находиться in vitro или in vivo, например, клетка может образовывать часть органа, ткани и/или организма пациента. В соответствии с данным изобретением трансфекция может быть транзиторной или стабильной. Для некоторых случаев использования трансфекции достаточно, если трансфицированный генетический материал экспрессировался только транзиторно. Поскольку нуклеиновая кислота, введенная в процессе трансфекции, обычно не интегрируется в нуклеарный геном, чужеродная нуклеиновая кислота будет разбавляться в ходе митоза или деградировать. Клетки, допускающие эписомальную амплификацию нуклеиновых кислот, значительно уменьшают скорость разбавления. Если желательно, чтобы трансфицированная нуклеиновая кислота действительно оставалась в геноме клетки и ее дочерних клетках, должна происходить стабильная трансфекция. РНК может быть трансфицирована в клетки для транзиторной экспрессии ее кодированного белка.
Согласно настоящему изобретению для введения, то есть, для переноса или трансфекции, нуклеиновых кислот в клетку может быть использован любой метод. Предпочтительно, нуклеиновая кислота, такая как РНК, трансфицируется в клетки стандартными методами. Такие методы включают электропорацию, липофекцию и микроинъекцию. В соответствии с одним особенно предпочтительным вариантом данного изобретения РНК вводят в клетки методом электропорации. Электропорация или электропермеабилизация относится к значительному увеличению электрической проводимости и проницаемости цитоплазматической оболочки, вызванной приложением внешнего электрического поля. Она обычно применяется в молекулярной биологии как метод введения некоторого вещества в клетку. Согласно настоящему изобретению предпочтительно, чтобы введение нуклеиновой кислоты, кодирующей белок или пептид, в клетки приводило к экспрессии указанных белка или пептида.
Для введения конструкций антигеновых рецепторов в Т-клетки могут быть использованы различные методы, включая трансфекцию с использованием невирусной ДНК, системы на основе транспозонов и системы на вирусной основе. Трансфекция с использованием невирусной ДНК характеризуется небольшим риском инсерционного мутагенеза. Системы на основе транспозонов могут интегрировать трансгены более эффективно, чем плазмиды, которые не содержат интегрирующего элемента. Системы на вирусной основе включают использование γ-ретровирусов и лентивирусных векторов. γ-Ретровирусы продуцируются сравнительно легко, эффективно и постоянно трансдуцируют Т-клетки и характеризуются предварительно доказанной безопасностью с точки зрения интеграции в первичные Т-клетки человека. Лентивирусные векторы также эффективно и постоянно трансдуцируют Т-клетки, но их получение является более дорогим. Они также потенциально являются более безопасными, чем системы на основе ретровирусов.
Для трансфекции клеток in vivo может быть использована фармацевтическая композиция, включающая нуклеиновую кислоту, кодирующую антигенный рецептор. Носитель для доставки, который таргетирует нуклеиновую кислоту в специфическую клетку, такую как Т-клетку, может быть введен пациенту, что приводит к трансфекции, которая происходит in vivo.
Согласно настоящему изобретению предпочтительно вводить нуклеиновую кислоту, кодирующую антигенный рецептор, в "оголенной форме" или на носителе. Носители, такие как липидные носители, предусмотренные для применения согласно настоящему изобретению, включают любые вещества или наполнители, с которыми нуклеиновая кислота, такая как РНК, может ассоциироваться, например, путем образования комплексов с нуклеиновой кислотой или путем образования везикул, в которые заключена или инкапсулирована нуклеиновая кислота. Это может привести к повышенной стабильности нуклеиновой кислоты по сравнению с оголенной нуклеиновой кислотой. В особенности, может быть повышена стабильность нуклеиновой кислоты в крови. Например, могут быть использованы составы РНК в форме наночастиц с определенным размером, такие как липоплексы из РНК и липосом, например, липоплексы, включающие DOTMA и DOPE или DOTMA и холестерин.
Используемый в данной заявке термин "наночастица" относится к любой частице, в частности к частице нуклеиновых кислот, имеющей диаметр, делающий эту частицу подходящей для системного, в частности парентерального, введения, обычно диаметр менее 1000 нанометров (нм). Согласно некоторым вариантам наночастица имеет диаметр менее 600 нм. Согласно некоторым вариантам наночастица имеет диаметр менее 400 нм.
Используемый в данной заявке термин "состав наночастиц" или похожие термины относятся к любому веществу, которое содержит по меньшей мере одну наночастицу. Согласно некоторым вариантам композиция наночастиц представляет собой однородную массу наночастиц. Согласно некоторым вариантам композиции наночастиц представляют собой дисперсию или эмульсию. В общем, дисперсия или эмульсия образуется, когда соединяются по меньшей мере два несмешивающихся материала.
Термин "липоплекс" или "липоплекс с нуклеиновой кислотой", в частности, "РНК-липоплекс", относится к комплексу липидов и нуклеиновых кислот, в частности, РНК. Липоплексы образуются спонтанно, когда катионные липосомы, которые часто включают также нейтральную жидкость "хелпер", смешиваются с нуклеиновыми кислотами.
Катионные липиды, катионные полимеры и другие вещества с положительными зарядами могут образовывать комплексы с отрицательно заряженными нуклеиновыми кислотами. Эти катионные молекулы могут быть использованы для комплексообразования с нуклеиновыми кислотами, образуя при этом, например, так называемые липоплексы или полиплексы, соответственно, и было показано, что эти комплексы осуществляют доставку нуклеиновых кислот в клетки.
Препараты нуклеиновых кислот в виде наночастиц для применения согласно настоящему изобретению могут быть получены по различным протоколам и из различных соединений, образующих комплексы с нуклеиновыми кислотами. Типичными комплексообразующими агентами являются липиды, полимеры, олигомеры или амфифилы. Согласно одному из вариантов комплексообразующее соединение включает по меньшей мере один агент, выбранный из группы, состоящей из протамина, полиэтиленимина, поли-L-лизина, поли-L-аргинина или гистона.
В соответствии с данным изобретением протамин пригоден в качестве катионного носителя. Термин "протамин" относится к любому из различных сильноосновных белков со сравнительно низкой молекулярной массой, которые богаты аргинином и особенно ассоциируются с ДНК вместо соматических гистонов в клетках спермы различных животных (таких как рыбы). В частности, термин "протамин" относится к белкам, обнаруженным в сперме рыб, которые являются сильноосновными, растворимыми в воде, не коагулируют при нагревании и при гидролизе образуют в основном аргинин. В чистом виде они используются в составе на основе инсулина с пролонгированным действием и для нейтрализации антикоагулирующего действия гепарина.
Согласно изобретению подразумевается, что термин "протамин", используемый в данной заявке, включает любую последовательность протамина аминокислоты, полученную или происходящую из нативных или биологических источников, включая ее фрагменты и мультимерные формы указанной аминокислотной последовательности или ее фрагмента. Кроме того, этот термин охватывает (синтезированные) полипептиды, которые являются искусственными и специально сконструированы для конкретных целей и не могут быть выделены из нативных или биологических источников.
Протамин, используемый согласно настоящему изобретению, может быть сульфированным протамином или протамина гидрохлоридом. Согласно предпочтительному варианту источник протамина, используемый для получения наночастиц, описанных в данной заявке, представляет собой протамин 5000, который содержит протамин в количестве более 10 мг/мл (5000 единиц, нейтрализующих гепарин, в мл) в изотоническом солевом растворе.
Липосомы являются микроскопическими липидными везикулами, часто имеющими один или более двойных слоев липида, образующего везикулы, такого как фосфолипид, и способны инкапсулировать лекарство. В контексте настоящего изобретения можно использовать различные типы липосом, включая, без ограничения, мультиламеллярные везикулы (MLV), малые одноламеллярные везикулы (SUV), большие одноламеллярные везикулы (LUV), стерически стабилизированные липосомы (SSL), мультивезикулярные везикулы (MV) и большие мультивезикулярные везикулы (LMV), а также другие двухслойные формы, известные в данной области. Размер и ламеллярность липосомы будет зависеть от метода получения, и выбор типа везикул, которые должны быть использованы, будет зависеть от предпочтительного способа введения. Существует несколько других форм надмолекулярной организации, в которых липиды могут находиться в водной среде, включающей ламеллярные фазы, гексагональные и инверсные гексагональные фазы, кубические фазы, мицеллы, обратные мицеллы, состоящие из монослоев. Эти фазы могут быть также получены в комбинации с ДНК или РНК, и взаимодействие с РНК и ДНК может существенно влиять на фазовое состояние. Описанные фазы могут присутствовать в составах на основе наночастиц нуклеиновых кислот по данному изобретению.
Для образования липоплексов нуклеиновой кислоты из нуклеиновой кислоты и липосом может быть использован любой подходящий способ, если он обеспечивает образование предусмотренных липоплексов нуклеиновой кислоты. Липосомы могут быть образованы с помощью стандартных методов, таких как метод обратного испарения (REV), метод с инъекцией этанола, метод дегидратации-регидратации (DRV), применение ультразвука или другие подходящие методы.
После образования липосом липосомы могут быть отсортированы по размеру для получения состава липосом, имеющих по существу однородный интервал размеров.
Липиды, образующие двойные слои, имеют обычно две углеводородных цепи, в особенности ацильные цепи, и головную группу, или полярную, или неполярную. Липиды, образующие двойные слои, состоят или из липидов природного происхождения, или из липидов синтетического происхождения, включая фосфолипиды, такие как фосфатидилхолин, фосфатидилэтаноламин, фосфатидовая кислота, фосфатидилинозитол и сфингомиелин, где две углеводородные цепи обычно содержат около 14-22 атомов углерода, и имеют различные степени ненасыщенности. Другие подходящие липиды для использования в композиции по данному изобретению включают гликолипиды и стеролы, такие как холестерин и его различные аналоги, которые могут быть также использованы в липосомах.
Катионные липиды обычно содержат липофильный фрагмент, такой как стерольную, ацильную и диацильную цепь, и несут суммарный положительный заряд. Головная группа липида обычно несет положительный заряд. Катионный липид предпочтительно содержит положительный заряд от 1 до 10, более предпочтительно положительный заряд от 1 до 3 и еще более предпочтительно положительный заряд 1. Примеры катионных липидов включают, но без ограничения, 1,2-ди-О-октадеценил-3-триметиламмонийпропан (DOTMA); диметилдиоктадециламмоний (DDAB); 1,2-диолеоил-3-триметиламмонийпропан (DOTAP); 1,2-диолеоил-3-диметиламмонийпропан (DODAP); 1,2-диацилокси-3-диметиламмонийпропаны; 1,2-диалкилокси-3-диметиламмонийпропаны; диоктадецилдиметиламмонийхлорид (DODAC), 1,2-димиристоилоксипропил-1,3-диметилгидроксиэтиламмоний (DMRIE) и 2,3-диолеоилокси-N-[2(сперминкарбоксамид)этил]-N,N-диметил-1-пропанаминтрифторацетат (DOSPA). Предпочтительными являются DOTMA, DOTAP, DODAC и DOSPA. Наиболее предпочтителен DOTMA.
Кроме того, наночастицы, описанные в данной заявке, предпочтительно также включают нейтральный липид ввиду структурной стабильности и т.п. Нейтральный липид может быть выбран соответствующим образом с учетом эффективности доставки комплекса нуклеиновая кислота-липид. Примеры нейтральных липидов включают, но без ограничения, 1,2-di-(9Z-октадеценоил)-sn-глицеро-3-фосфоэтаноламин (DOPE), 1,2-диолеоил-sn-глицеро-3-фосфохолин (DOPC), диацилфосфатидилхолин, диацилфосфатидилэтаноламин, церамид, сфингомиелин, цефалин, стерол и цереброзид. Предпочтительными являются DOPE и/или DOPC. Наиболее предпочтителен DOPE. В случае, когда катионная липосома включает как катионный липид, так и нейтральный липид, мольное отношение катионного липида к нейтральному липиду может быть соответствующим образом определено с учетом стабильности липосомы и т.п.
Согласно одному из вариантов наночастицы, описанные в данной заявке, могут включать фосфолипиды. Фосфолипидами могут быть глицерофосфолипиды. Примеры глицерофосфолипидов включают, но без ограничения, три типа липидов: (i) цвиттерионные фосфолипиды, которые включают, например, фосфатидилхолин (PC), фосфатидилхолин яичного желтка, PC из природных соевых бобов, в частично гидрогенизированной или полностью гидрогенизированной форме, димиристоилфосфатидилхолин (DMPC), сфингомиелин (SM); (ii) отрицательно заряженные фосфолипиды, которые включают, например, фосфатидилсерин (PS), фосфатидилинозитол (PI), фосфатидовую кислоту (РА), фосфатидилглицерин (PG) дипальмитоил-PG, димиристоилфосфатидилглицерин (DMPG); синтетические производные, в которых конъюгат представляет собой цвиттерионный фосфолипид, отрицательно заряженный, такой как в случае метокси-полиэтиленгликоль-дистеароил-фосфатидилэтаноламина (mPEG-DSPE); и (iii) катионные фосфолипиды, которые включают, например, фосфатидилхолин или сфингомиелин, фосфомоноэфиры которых были О-метилированы с образованием катионных липидов.
Ассоциация нуклеиновой кислоты с липидным носителем может происходить, например, путем заполнения нуклеиновой кислотой межклеточного пространства носителя, таким образом, что носитель физически захватывает нуклеиновую кислоту, или путем образования ковалентной, ионной или водородной связи, или посредством адсорбции при образовании неспецифических связей. Каков бы ни был механизм ассоциации, нуклеиновая кислота должна сохранять свои терапевтические, то есть кодирующие, свойства.
Согласно настоящему изобретению нуклеиновая кислота, кодирующая антигенный рецептор, в соответствии с одним вариантом представляет собой РНК, предпочтительно мРНК. РНК предпочтительно получают путем in-vitro транскрипции.
Термин "нуклеиновая кислота", используемый в данной заявке, включает ДНК и РНК, такие как геномные ДНК, кДНК, мРНК, продуцированные рекомбинантно и синтезированные химическим путем. Нуклеиновая кислота может быть однонитевой или двунитевой. РНК включает in vitro транскрибированную РНК (IVT RNA) или синтетическую РНК. Согласно данному изобретению нуклеиновая кислота предпочтительно является выделенной нуклеиновой кислотой.
Нуклеиновые кислоты могут быть заключены в вектор. Термин "вектор", используемый в данной заявке, включает любые векторы, известные специалисту в данной области, включая плазмидные векторы, космидные векторы, фаговые векторы, такие как фаг лямбда, вирусные векторы, такие как аденовирусные или бакуловирусные, или искусственные хромосомные векторы, такие как бактериальные искусственные хромосомы (ВАС), дрожжевые искусственные хромосомы (YAC) или искусственные хромосомы Р1 (РАС). Указанные векторы включают экспрессионные, а также клонирующие векторы. Экспрессионные векторы включают плазмиды, а также вирусные векторы и обычно содержат желательную кодирующую последовательность и соответствующие ДНК-последовательности, необходимые для экспрессии функционально связанной кодирующей последовательности в конкретном организме хозяина (например, бактерии, дрожжей, растения, насекомого или млекопитающего), или в in vitro системах экспрессии. Клонирующие векторы обычно используются для конструирования и амплификации некоторого желательного фрагмента ДНК и в них могут отсутствовать функциональные последовательности, необходимые для экспрессии желательных фрагментов ДНК.
В контексте настоящего изобретения термин "РНК" относится к молекуле, которая включает остатки рибонуклеотидов и предпочтительно полностью или существенно состоит из остатков рибонуклеотидов. Термин "рибонуклеотид" относится к нуклеотиду с гидроксильной группой в 2'-положении β-D-рибофуранозильной группы. Термин охватывает двунитевую РНК, однонитевую РНК, выделенную РНК, такую как частично очищенную РНК, практически чистую РНК, синтетическую РНК, полученную рекомбинантную РНК, а также модифицированную РНК, которая отличается от РНК природного происхождения добавлением, делецией, заменой и/или изменением одного или более нуклеотидов. Такие изменения могут включать добавление ненуклеотидного материала, например, по концу (-ам) РНК или внутри цепи, например, в состав одного или более нуклеотидов РНК. Нуклеотиды в молекулах РНК могут также включать нестандартные нуклеотиды, такие как нуклеотиды неприродного происхождения или нуклеотиды, синтезированные химическим путем, или дезоксинуклеотиды. Эти измененные РНК могут рассматриваться как аналоги или аналоги РНК природного происхождения.
Согласно настоящему изобретению термин "РНК" включает и предпочтительно относится к "мРНК", что означает "матричную РНК", и относится к "транскрипту", который может быть получен с использованием ДНК в качестве темплата и кодирует пептид или белок. мРНК обычно включает 5'-нетранслируемую область (5'-UTR), кодирующую пептид или белок и 3'-нетранслируемую область (3'-UTR). мРНК имеет ограниченный период полужизни в клетках и in vitro. Предпочтительно мРНК получается путем in vitro транскрипции с использованием матричной ДНК. Согласно одному варианту данного изобретения РНК получается путем in vitro транскрипции или путем химического синтеза. Методика in vitro транскрипции известна специалисту в данной области. Например, имеются разнообразные коммерчески доступные наборы для in vitro транскрипции.
Согласно одному из вариантов настоящего изобретения РНК является самореплицирующейся РНК, такой как однонитевая самореплицирующаяся РНК. Согласно одному варианту самореплицирующаяся РНК представляет собой однонитевую РНК с положительной полярностью. Согласно одному варианту самореплицирующаяся РНК представляет собой вирусную РНК или РНК, полученную из вирусной РНК. Согласно одному варианту самореплицирующаяся РНК представляет собой альфа-вирусную геномную РНК или получена из альфа-вирусной геномной РНК. Согласно одному варианту самореплицирующаяся РНК представляет собой вектор экспрессии вирусного гена. Согласно одному варианту вирус является вирусом леса Семлики. Согласно одному варианту самореплицирующаяся РНК содержит один или более трансгенов, по меньше мере один из указанных трансгенов, кодирующих агенты, описанные в данной заявке. Согласно одному варианту, если РНК является вирусной РНК или получена из вирусной РНК, трансгены могут частично или полностью заменять вирусные последовательности, такие как вирусные последовательности, кодирующие структурные белки. Согласно одному из вариантов самореплицирующаяся РНК представляет собой РНК, транскрибированную in vitro.
Для того, чтобы усилить экспрессию и/или увеличить стабильность РНК, используемой согласно настоящему изобретению, она может быть модифицирована, предпочтительно без изменения последовательности экспрессированного пептида или белка.
Термин "модификация" в контексте РНК, используемый согласно данному изобретению, включает любую модификацию РНК, которая само собой имеется в указанной РНК.
Согласно одному из вариантов данного изобретения РНК, используемая согласно изобретению, не содержит некэпированных 5'-трифосфатов. Удаление таких некэпированных 5'-трифосфатов может быть достигнуто путем обработки РНК фосфатазой.
Согласно настоящему изобретению РНК может содержать модифицированные природные или синтетические рибонуклеотиды с целью увеличения ее стабильности и/или уменьшения цитотоксичности. Например, согласно одному варианту в РНК, используемой согласно данному изобретению, 5-метилцитидин заменен частично или полностью, предпочтительно полностью, цитидином. Альтернативно или дополнительно, согласно одному варианту в РНК, используемой согласно данному изобретению, псевдоуридин может быть заменен частично или полностью, предпочтительно полностью, уридином.
Согласно одному варианту термин "модификация" относится к обеспечению РНК 5'-кэпом или 5'-кэп-аналогом. Термин "5'-кэп" относится к кэп-структуре, находящейся на 5'-конце молекулы мРНК и эта структура обычно состоит из гуанозиннуклеотида, соединенного с мРНК через нетипичную 5'-5'-трифосфатную связь. Согласно одному варианту этот гуанозин является метилированным в 7-положении. Термин "обычный 5'-кэп" относится к 5'-кэпу РНК природного происхождения, предпочтительно к кэпу 7-метилгуанозина (m7G). В контексте данного изобретения термин "5'-кэп" охватывает 5'-кэп-аналог, который напоминает структуру кэпа РНК и модифицирован для придания способности к стабилизации РНК, если присоединен к ней, предпочтительно in vivo и/или в клетке.
Обеспечение РНК 5'-кэпом или 5'-кэп-аналогом может быть достигнуто путем in vitro транскрипции матричной ДНК в присутствии указанного 5'-кэпа или 5'-кэп-аналога, причем указанный 5'-кэп вводится в генерированную нить РНК совместной транскрипцией или РНК может быть получена, например, путем in vitro транскрипции, и 5'-кэп может быть присоединен к РНК посттрансляционно с использованием кэпирующих ферментов, например, кэпирующих ферментов вируса осповакцины.
РНК может включать другие модификации. Например, модификацией РНК, используемой согласно данному изобретению, может быть вставка или усечение поли(А) хвоста природного происхождения или изменение 5'- или 3'-нетранслируемых областей (UTR), такое как введение UTR, которая не относится к кодирующему участку указанной РНК, например, инсерция одной или более, предпочтительно двух копий 3'-UTR, полученных из гена глобина, такого как альфа2-глобин, альфа1-глобин, бета-глобин, предпочтительно бета-глобин, более предпочтительно человеческий бета-глобин.
Следовательно, для того, чтобы увеличить стабильность и/или усилить экспрессию РНК, используемую согласно настоящему изобретению, она может быть модифицирована таким образом, чтобы находиться в совокупности с поли-А-последовательностью, предпочтительно, содержащей цепь длиной от 10 до 500, более предпочтительно от 30 до 300, еще более предпочтительно от 65 до 200 и особенно от 100 до 150 аденозиновых остатков. Согласно особенно предпочтительному варианту поли-А-последовательность имеет длину из приблизительно 120 аденозиновых остатков. Кроме того, введение двух или более 3'-нетранслируемых областей (UTR) в 3'-нетранслируемую область молекулы РНК может привести к повышению эффективности трансляции. Согласно одному конкретному варианту 3'-UTR получена из человеческого гена β-глобина.
Термин "стабильность" в отношении РНК относится к "периоду полужизни" РНК. Термин "период полужизни (полураспада)" относится к периоду времени, который необходим для исчезновения половины активности, количества или числа молекул. В контексте настоящего изобретения период полужизни РНК является показателем стабильности указанной РНК. Период полужизни РНК может влиять на "продолжительность экспрессии" РНК. Можно ожидать, что РНК с длительным периодом полужизни будет экспрессироваться в течение продолжительного периода времени.
В контексте настоящего изобретения термин "транскрипция" относится к процессу, когда генетический код в последовательности ДНК транскрибируется в РНК. Впоследствии РНК может транслироваться в белок. Согласно данному изобретению термин "транскрипция" включает "in vitro транскрипцию", причем термин "in vitro транскрипция" относится к процессу, в ходе которого РНК, в частности мРНК, синтезируется in vitro в системе, не содержащей клеток, предпочтительно при использовании соответствующих клеточных экстрактов. Для получения транскриптов предпочтительно применять клонирующие векторы. Эти клонирующие векторы обычно обозначаются как транскрипционные векторы и охвачены согласно данному изобретению термином "вектор".
Согласно настоящему изобретению термин "трансляция" относится к процессу в рибосомах клетки, при котором нить матричной РНК направляет сборку аминокислотных последовательностей для получения пептида или белка.
Нуклеиновые кислоты согласно данному изобретению могут находиться сами по себе или в комбинации с другими нуклеиновыми кислотами, которые могут быть гомологичными или гетерологичными. Согласно предпочтительным вариантам нуклеиновая кислота функционально связана с последовательностями, контролирующими экспрессию, которые могут быть гомологичными или гетерологичными по отношению к указанной нуклеиновой кислоте. Термин "гомологичный" означает, что нуклеиновые кислоты также функционально связаны естественным путем и термин "гетерологичный" означает, что нуклеиновые кислоты не связаны функционально естественным путем.
Нуклеиновая кислота и последовательность, контролирующая экспрессию, "функционально" связаны друг с другом, если они ковалентно связаны друг с другом таким образом, что экспрессия или транскрипция указанной кислоты находится под контролем или под влиянием указанной последовательности, контролирующей экспрессию. Если нуклеиновая кислота должна транслироваться в функциональный белок, тогда, с наличием последовательности, контролирующей экспрессию, функционально связанной с кодирующей последовательностью, индукция указанной последовательности, контролирующей экспрессию, приводит к транскрипции указанной нуклеиновой кислоты, не вызывая сдвига рамки считывания в кодирующей последовательности, или указанная кодирующая последовательность не способна транслироваться в желательный белок или пептид.
Термин "последовательность, контролирующая экспрессию" или "контролирующий экспрессию элемент" согласно изобретению включает промоторы, сайты связывания рибосом, энхансеры и другие контролирующие элементы, которые регулируют транскрипцию гена или трансляцию мРНК. Согласно конкретным вариантам изобретения можно регулировать контролирующие экспрессию последовательности. Точная структура последовательностей, контролирующих экспрессию, может меняться как функция видов или типов клеток, но обычно содержит 5'-нетранскрибированную и 5'- и 3'-нетранслируемые последовательности, которые участвуют в инициации транскрипции и трансляции, соответственно, такие как ТАТА-бокс, кэпирующая последовательность, САА-последовательность и т.п. Более конкретно, 5'-нетранскрибированные последовательности, контролирующие экспрессию, включают промоторную область, которая включает промоторную последовательность для контроля транскрипции функционально связанной нуклеиновой кислоты. Последовательности, контролирующие экспрессию, могут также включать последовательности энхансеров или активирующие последовательности.
Термин "экспрессия" согласно данному изобретению используется в его самом общем значении и включает продуцирование РНК и/или пептидов или белков, например, путем транскрипции и/или трансляции. В отношении к РНК термин "экспрессия" или "трансляция" относится в частности к получению пептидов или белков. Он также охватывает частичную экспрессию нуклеиновых кислот. Более того, экспрессия может быть временной или стабильной. Согласно изобретению термин "экспрессия" включает также "аберрантную экспрессию" или "аномальную экспрессию".
"Аберрантная экспрессия" или "аномальная экспрессия" согласно настоящему изобретению означает, что экспрессия изменена, предпочтительно усилена, по сравнению с контрольным вариантом, например, состоянием субъекта, не имеющего заболевания, ассоциируемого с аберрантной или аномальной экспрессией некоторого белка, например, опухолевого антигена. Усиление экспрессии относится к усилению по меньшей мере на 10%, в частности по меньшей мере на 20%, по меньшей мере на 50% или по меньшей мере на 100% или более. Согласно одному варианту экспрессия обнаруживается только в пораженной ткани, в то время как экспрессия в здоровой ткани репрессируется.
Термин "специфично экспрессируемый" означает, что белок по существу экспрессируется только в конкретной ткани или органе. Например, опухолевый антиген, специфично экспрессируемый в слизистой оболочке желудка, означает, что указанный белок преимущественно экспрессирован в слизистой оболочке желудка и не экспрессирован в других тканях или не экспрессирован в значительной степени в других типах ткани or органов. Таким образом, белок, который исключительно экспрессирован в клетках слизистой оболочки желудка и в значительно меньшей степени в любой другой ткани, например, в ткани яичка, специфично экспрессирован в клетках слизистой оболочки желудка. Согласно некоторым вариантам опухолевый антиген может также быть специфически экспрессирован при обычных условиях в более, чем одном типе ткани или органов, например, в 2 или 3 различных типах тканей или органов, но предпочтительно не более, чем в 3 различных типах тканей или органов. В этом случае опухолевый антиген специфично экспрессирован в этих органах. Например, если опухолевый антиген экспрессирован в обычных условиях, предпочтительно приблизительно в равной степени в легком и желудке, указанный опухолевый антиген специфично экспрессирован в легком и желудке.
Согласно данному изобретению термин "кодирующая нуклеиновая кислота" означает, что нуклеиновая кислота, если она находится в соответствующем окружении, предпочтительно в клетке, может экспрессироваться с получением белка или пептида, которые она кодирует.
Термин "пептид" согласно данному изобретению включает олиго- и полипептиды и относится к веществу, которое содержит две или более, предпочтительно 3 или более, предпочтительно 4 или более, предпочтительно 6 или более, предпочтительно 8 или более, предпочтительно 9 или более, предпочтительно 10 или более, предпочтительно 13 или более, предпочтительно 16 или более, предпочтительно 21 или более и предпочтительно до 8, 10, 20, 30, 40 или 50, в частности 100 аминокислот, соединенных ковалентно с помощью пептидных связей. Термин "белок" относится к большим пептидам, предпочтительно к пептидам с более чем 100 аминокислотными остатками, но в общем термины "пептид", "пептидная цепь" и "белок" являются синонимами и используются в данной заявке как взаимозаменяемые.
Как упоминалось выше, аминокислотные последовательности пептидных цепей и антигенных рецепторов, описанных в данной заявке, могут быть модифицированы таким образом, чтобы получить варианты указанных аминокислотных последовательностей. Соответственно, настоящее изобретение включает варианты пептидных и белковых последовательностей, описанных в данной заявке, и включает варианты аминокислотных последовательностей природного происхождения, приводящие к последовательностям, которые функционально эквивалентны указанным последовательностям, например, аминокислотным последовательностям, проявляющим свойства, идентичные или похожие на свойства указанных последовательностей. Важными являются свойства сохранять способность к связыванию антигенного рецептора с его мишенью или трансдукцию антигенсвязывающего фрагмента в клетку, такую как Т-клетка. Согласно одному из вариантов вариант молекулы или последовательности является иммунологически эквивалентным родительской молекуле или последовательности.
Термин "вариант" согласно изобретению относится, в частности, к мутантам, сплайс-вариантам, конформациям, изоформам, аллельным вариантам, вариантам видов и гомологам видов, в частности таким, которые находятся в природных условиях. Аллельный вариант относится к изменению в нормальной последовательности гена, значение которого часто является неясным. Полное секвенирование гена часто дает возможность идентифицировать многочисленные аллельные варианты данного гена. Гомолог вида представляет собой последовательность нуклеиновой кислоты или аминокислоты происхождения, отличающегося от происхождения последовательности данной нуклеиновой кислоты или аминокислоты. Термин "вариант" охватывает любые посттрансляционно модифицированные варианты и конформационные варианты.
Термин "иммунологически эквивалентный" означает, что иммунологически эквивалентная молекула, такая как иммунологически эквивалентная аминокислотная последовательность, проявляет одинаковые или по существу одинаковые иммунологические свойства и/или обеспечивает одинаковые или по существу одинаковые иммунологические эффекты, например, в отношении вида иммунологического эффекта. В контексте настоящего изобретения термин "иммунологически эквивалентный" предпочтительно используется в отношении иммунологических эффектов или свойств антигеновых рецепторов, применяемых для терапии.
Специалистам в данной области известно, что в особенности гипервариабельные и вариабельные области последовательностей CDR могут быть модифицированы без потери способности связываться с мишенью. Например, CDR области могут быть или идентичны, или в высокой степени гомологичны областям родительских антител. Под "высоко гомологичными" подразумевают, что в CDRs могут быть сделаны от 1 до 5, предпочтительно от 1 до 4, например, от 1 до 3 замен или 1, или 2 замены.
Для целей данного изобретения "варианты" аминокислотной последовательности включают варианты инсерций в аминокислоты, варианты аминокислотных добавлений, варианты аминокислотных делеций и/или варианты аминокислотных замен. Варианты аминокислотных делеций, которые включают делецию на N-конце и/или на С-конце белка, называются также N-концевыми и/или С-концевыми вариантами усечений.
Варианты аминокислотных инсерций включают инсерций одной или двух, или более аминокислот в конкретной аминокислотной последовательности. В случае вариантов аминокислотных последовательностей, имеющих инсерцию, один или более аминокислотных остатков инсерцированы (вставлены) в конкретный сайт в последовательности аминокислоты, хотя случайная инсерция с соответствующим скринингом полученного продукта также возможна.
Варианты аминокислотных добавлений включают аминоконцевые и/или карбоксиконцевые слияния одной или более аминокислот, например, 1, 2, 3, 5, 10, 20, 30, 50 или более аминокислот.
Варианты аминокислотных делеций характеризуются удалением одной или более аминокислот из последовательности, например, путем удаления 1, 2, 3, 5, 10, 20, 30, 50 или более аминокислот. Делеции могут быть в любом положении белка.
Варианты аминокислотных замен характеризуются тем, что по меньшей мере один остаток в последовательности удален и другой остаток инсерцирован на его место. Предпочтение отдается модификациям в положениях в аминокислотной последовательности, которые не являются консервативными, между гомологичными белками или пептидами, и/или замене аминокислот другими аминокислотами, имеющими сходные свойства. Предпочтительно аминокислотные изменения в белковых вариантах являются изменениями консервативных аминокислот, то есть, заменами заряженных сходным образом или незаряженных аминокислот. Изменение консервативной аминокислоты включает замену одной кислоты из семейства аминокислот, которые находятся в боковых цепях. Аминокислоты природного происхождения обычно подразделяются на четыре семейства: кислые (аспартат, глутамат), основные (лизин, аргинин, гистидин), неполярные (аланин, валин, лейцин, глицин, аспарагин, глутамин, цистеин, серин, треонин, тирозин) аминокислоты. Фенилаланин, триптофан и тирозин иногда совместно классифицируют как ароматические аминокислоты.
Предпочтительно, когда степень (процент) сходства, предпочтительно идентичности, между последовательностью данной аминокислоты и последовательностью аминокислоты, которая является вариантом указанной последовательности данной аминокислоты составляет по меньшей мере около 60%, 65%, 70%, 80%, 81%, 82%, 83%, 84%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%. Степень сходства или идентичности предпочтительно указывается для области аминокислоты, которая составляет по меньше мере приблизительно 10%, по меньшей мере приблизительно 20%, по меньшей мере приблизительно 30%, по меньшей мере приблизительно 40%, по меньшей мере приблизительно 50%, по меньшей мере приблизительно на 60%, по меньшей мере приблизительно 70%, по меньшей мере приблизительно на 80%, по меньше мере приблизительно 90% или приблизительно 100% от полной длины эталонной последовательности аминокислоты. Например, если эталонная последовательность аминокислоты состоит из 200 аминокислот, степень сходства или идентичности предпочтительно указана приблизительно для 20, по меньшей мере приблизительно для 40, по меньшей мере приблизительно 60, по меньшей мере приблизительно 80, по меньшей мере приблизительно 100, по меньшей мере приблизительно 120, по меньшей мере приблизительно 140, по меньшей мере приблизительно 160, по меньшей мере приблизительно 180, или приблизительно 200 аминокислот, предпочтительно аминокислот непрерывной цепи. Согласно предпочтительным вариантам степень сходства или идентичности приводится для полной длины последовательности эталонной аминокислоты. Выравнивание для определения сходства последовательностей, предпочтительно идентичности последовательностей, может быть сделано известными из уровня техники методами, предпочтительно с использованием оптимального метода выравнивания, например, с использованием программы Align, с пакетом стандартных параметров, предпочтительно EMBOSS::needle, Matrix: Blosum62, Gap Open 10.0, Gap Extend 0.5.
"Сходство последовательностей" показывает процент аминокислот, которые или идентичны, или представляют замены консервативных аминокислот. "Идентичность последовательностей" при рассмотрении последовательностей двух аминокислот показывает процент аминокислот, которые идентичны у этих последовательностей.
Термин "процент идентичности" обозначает количество аминокислотных остатков, которые идентичны у сравниваемых последовательностей, полученных после оптимального выравнивания, при этом эта величина в процентах является чисто статистической и различия между двумя последовательностями распределены хаотично и по всей их длине. Сравнение двух аминокислотных последовательностей обычно осуществляется путем сравнения этих последовательностей после их оптимального выравнивания, причем указанное сравнение проводится сегментарно или с помощью "окна сравнения" для того, чтобы идентифицировать и сравнить локальные области сходных последовательностей. Для сравнения можно провести выравнивание последовательностей для сравнения, кроме ручного выравнивания, с помощью алгоритма локальной аналогии Смита и Уотермана, 1981, Ads Арр. Math. 2, 482, посредством алгоритма локальной гомологии Нидлмана и Вунша, 1970, J. Mol. Biol. 48, 443, с помощью способа поиска сходства Пирсона и Липмана, 1988, Proc. Natl. Acad. Sci. USA 85, 2444, или с помощью компьютерных программ, которые используют эти алгоритмы (GAP, BESTFIT, FASTA, BLAST Р, BLAST N и TFASTA в Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Drive, Madison, Wis.).
Процент идентичности рассчитывается путем определения числа идентичных позиций в двух сравниваемых последовательностях, деления этого количества идентичных позиций на общее количество позиций в окне выравнивания и умножения полученного результата на 100, чтобы получить процент идентичности этих двух последовательностей.
Гомологичные последовательности аминокислот согласно данному изобретению характеризуются по меньшей мере 40%, в частности по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% и предпочтительно по меньшей мере 95%, по меньшей мере 98 или по меньшей мере 99%-ной идентичностью остатков аминокислот.
Согласно изобретению вариант, фрагмент, часть или производное последовательности аминокислоты, пептида или белка предпочтительно имеет функциональное свойство последовательности аминокислоты, пептида или белка, соответственно, из которых они получены, то есть, это функциональный эквивалент. Согласно одному варианту вариант, фрагмент, часть, область или производное аминокислотной последовательности, пептида или белка функционально эквивалентны, например, иммунологически эквивалентны аминокислотной последовательности, пептиду или белку, соответственно, из которых они получены. Согласно одному варианту функциональное свойство представляет собой способность связываться с антигеном или трансдуцировать сигнал связывания в клетку.
Термин "производное" согласно данному изобретению означает, что конкретный фрагмент, в частности конкретная последовательность, содержится в объекте, из которого он получен (происходит), в частности в организме или в молекуле. В случае аминокислотных последовательностей, особенно конкретных областей последовательностей, "производное" в частности означает, что релевантная последовательность аминокислоты получена (происходит) из аминокислотной последовательности, в которой она находится.
Термин "клетка" или "клетка-хозяин" предпочтительно относится к интактной клетке, то есть, к клетке с интактной мембраной, которая не высвободила свои нормальные внутриклеточные компоненты, такие как ферменты, органеллы или генетический материал. Интактная клетка предпочтительно является жизнеспособной клеткой, то есть, живой клеткой, способной выполнять свои нормальные метаболические функции. Предпочтительно согласно данному изобретению указанный термин относится к любой клетке, которая может быть трансфицирована экзогенной нуклеиновой кислотой. Предпочтительно, если клетка, будучи трансфицирована экзогенной нуклеиновой кислотой и перенесена в клетку, может экспрессировать нуклеиновую кислоту в реципиенте. Термин "клетка" включает бактериальные клетки; другие полезные клетки представляют собой дрожжевые клетки, клетки грибов или клетки млекопитающего. Подходящие бактериальные клетки включают клетки грамотрицательных бактериальных штаммов, таких как штаммы Escherichia coli, Proteus и Pseudomonas, и клетки грамположительных бактериальных штаммов, таких как штаммы Bacillus, Streptomyces, Staphylococcus и Lactococcus. Подходящие клетки грибов включают клетки видов Trichoderma, Neurospora и Aspergillus. Подходящие клетки дрожжей включают клетки видов Saccharomyces (например, Saccharomyces cerevisiae), Schizosaccharomyces (например, Schizosaccharomyces pombe), Pichia (например, Pichia pastoris и Pichia methanolicd), и Hansenula. Подходящие клетки млекопитающих включают, например, клетки СНО, клетки BHK, клетки HeLa, клетки COS, клетки HEK 293 и т.п. Однако могут быть также использованы клетки земноводных, клетки насекомых, клетки растений и любые другие клетки, используемые в уровне техники для экспрессии гетерологичных белков. Клетки млекопитающих предпочтительны для адоптивного переноса, это клетки людей, мышей, хомячков, свиней, коз и приматов. Эти клетки могут быть получены из большого числа типов тканей и включают первичные клетки и клеточные линии, такие как клетки иммунной системы, в частности антигенпрезентирующие клетки, такие как дендритные клетки и Т-клетки, стволовые клетки, например, гемопоэтические стволовые клетки и мезенхимальные стволовые клетки и другие виды клеток. Особенно предпочтительными клетками для применения согласно данному изобретению являются иммунореактивные или иммунные эффекторные клетки, в частности Т-клетки.
Клетка, которая содержит молекулу нуклеиновой кислоты, предпочтительно экспрессирует пептид или белок, кодированный нуклеиновой кислотой.
Термин "клональная экспансия" или "экспансия" относится к процессу, когда конкретное вещество размножается. В контексте данного изобретения термин предпочтительно используется в контексте иммунологического ответа, в котором лимфоциты стимулируются антигеном, пролифератом и конкретный лимфоцит, распознающий указанный антиген, амплифицируется. Предпочтительно клональная экспансия приводит к дифференцировке лимфоцитов. Термин "примирование" относится к процессу, когда Т-клетка имеет первый контакт с конкретным антигеном и вызывает дифференцировку в эффекторные Т-клетки.
Молекулы, такие как нуклеиновые кислоты, пептидные цепи и антигенные рецепторы или клетки, описанные в данной заявке, могут быть рекомбинантными и/или выделенными.
Термин "выделенное", использованный в данной заявке, относится к веществу, которое по существу не содержит других молекул, таких как другой клеточный материал. Термин "выделенное" предпочтительно означает, что выделенное вещество было отделено от его природного окружения. Выделенное вещество может быть по существу в очищенном состоянии. Термин "по существу очищенный" предпочтительно означает, что вещество по существу свободно от других веществ, с которыми оно ассоциировано в природе или in vivo.
Термин "рекомбинантный" в контексте настоящего изобретения означает "полученный с помощью генной инженерии". Предпочтительно "рекомбинантный объект", такой как рекомбинантная клетка, в контексте настоящего изобретения не имеет природного происхождения.
Термин "природное происхождение", используемый в данной заявке, относится к тому факту, что объект может быть обнаружен в природе. Например, пептид или нуклеиновая кислота, которые находятся в организме (включая вирусы) и могут быть выделены из источника в природе и которые не были намеренно модифицированы человеком в лаборатории, имеют природное происхождение.
Термин "аутологичный" используется для описания объекта, который происходит из того же субъекта. Например, "аутологичный трансплантат" относится к трансплантату ткани или органам, происходящим из того же самого субъекта. Такие методики являются преимущественными, потому что они преодолевают иммунологический барьер, который иначе приводит к отторжению.
Термин "аллогенный" используется для описания объекта, который происходит из разных индивидуумов одного и того же вида. Два или более индивидуумов считаются аллогенными по отношению один к другому, когда гены в одном и том же локусе не являются идентичными.
Термин "сингенный" используется для описания объекта, который происходит из разных индивидуумов или тканей, имеющих идентичные генотипы, то есть, идентичных близнецов или животных одного и того же инбредного штамма или их тканей.
Термин "гетерологичный" используется для описания объекта, состоящего из множества различных элементов. Примером является пересадка костного мозга одного индивидуума другому индивидууму, что представляет собой гетерологичный трансплантат.
Гетерологичный ген представляет собой ген, происходящий из источника, отличного от субъекта.
Термины "уменьшать" или "ингибировать", используемые в данной заявке, означают способность вызывать полное уменьшение, предпочтительно на 5% или более, на 10% или более, на 20% или более, более предпочтительно на 50% или более и наиболее предпочтительно на 75% или более от уровня. Термин "ингибирование" или похожие термины означают полное или по существу полное ингибирование, то есть, уменьшение до нуля или практически до нуля.
Термины, такие как "повышать" или "усиливать" предпочтительно относятся к повышению или усилению приблизительно по меньшей мере на 10%, предпочтительно по меньшей мере на 20%, предпочтительно по меньшей мере на 30%, более предпочтительно по меньшей мере на 40%, более предпочтительно по меньшей мере на 50%, еще более предпочтительно по меньшей мере на 80%, и наиболее предпочтительно по меньшей мере на 100%.
Поскольку антигенный рецептор по данному изобретению может быть получен в действительности для нацеливания на любой антиген, включая антигены, специфические для данной болезни, антигенный рецептор по данному изобретению характеризуется широким терапевтическим применением. Соответственно, настоящее изобретение направлено на применение антигенного рецептора по изобретению, его пептидных цепей, нуклеиновых кислот, кодирующих антигенный рецептор, и других родственных молекул при осуществлении терапевтических и профилактических способов. Одно такое применение представляет собой продуцирование антигенспецифических иммунных клеток, которые могут быть введены пациенту для предотвращения или лечения заболевания, причем это заболевание характеризуется экспрессией одного или более антигенов, которые могут быть связаны антигенным рецептором по изобретению, и экспрессированными в иммунных клетках. Предпочтительным заболеванием является раковое заболевание. Далее, антигенный рецептор по изобретению и родственные молекулы также могут быть использованы для селективной эрадикации клеток, экспрессирующих заданный антиген, а также для иммунизации или вакцинации против заболевания, при этом заданный антиген экспрессируется, этот антиген может быть связан с по меньшей мере одним сайтом связывания рецептора антигена по изобретению.
Согласно одному из вариантов изобретения способ лечения или профилактики заболевания включает введение пациенту эффективного количества нуклеиновой кислоты, кодирующей антигенный рецептор по изобретению, в котором по меньшей мере один сайт связывания антигенового рецептора способен связывать антиген, который ассоциирован с заболеванием (например, вирусный или опухолевый антиген), которое нужно подвергать лечению или профилактике. Согласно другому варианту способ лечения или профилактики заболевания включает введение пациенту эффективного количества рекомбинантной иммунной эффекторной клетки или размножившейся популяции указанных иммунных эффекторных клеток, при этом иммунная эффекторная клетка или популяция клеток рекомбинантно экспрессирует антигенный рецептор по изобретению, в котором по меньшей мере один сайт связывания антигенового рецептора способен связывать антиген, который ассоциирован с заболеванием, которое нужно подвергать лечению или профилактике. Согласно предпочтительным вариантам является раковое заболевание, и антиген представляет собой опухолеассоциированный антиген.
Согласно другому варианту настоящее изобретение предусматривает способ иммунизации или вакцинации против заболевания, ассоциированного со специфичным антигеном, или против организма, вызывающего заболевание и экспрессирующего специфичный антиген, причем этот способ включает введение пациенту эффективного количества нуклеиновой кислоты, кодирующей антигенный рецептор по изобретению, в котором по меньшей мере один сайт связывания антигенового рецептора способен связывать специфичный антиген. Согласно другому варианту настоящее изобретение предусматривает способ иммунизации или вакцинации против заболевания, ассоциированного со специфичным антигеном или против организма, вызывающего заболевание и экспрессирующего специфичный антиген, при этом способ включает введение пациенту эффективного количества рекомбинантной иммунной эффекторной клетки или популяции, при этом иммунная эффекторных клетка или популяция указанных размножившихся иммунных эффекторных клеток, при этом иммунная эффекторная клетка или популяция клеток рекомбинантно экспрессирует антигенный рецептор по изобретению, в котором по меньшей мере один сайт связывания антигенового рецептора способен связывать специфичный антиген.
Согласно некоторым вариантам популяция иммунных эффекторных клеток может быть клоногенно размножившейся популяцией. Рекомбинантные иммунный эффекторные клетки или их популяции обеспечивают терапевтическую или профилактическую иммунную эффекторную функцию антигенспецифическим способом. Предпочтительно, когда антигенный рецептор по изобретению экспрессирован на клеточной поверхности иммунной эффекторной клеток.
Клетки, используемые в связи с терапевтическим и профилактическим способами лечения согласно настоящему изобретению, предпочтительно являются иммунными эффекторными клетками и иммунные эффекторные клетки представляют собой предпочтительно Т-клетки. В частности, клетками, используемыми согласно изобретению, являются цитотоксические лимфоциты, предпочтительно выбранные из цитотоксических Т-клеток, естественные киллерные (NK) клетки и лимфокин-активированные киллерные (LAK) клетки. После активации/стимуляции каждый из этих цитотоксических лимфоцитов запускает деструкцию клеток-мишеней. Например, цитотоксические Т-клетки включают деструкцию клеток-мишеней с помощью одного или обеих следующих средств. Во-первых, после активации Т-клетки высвобождают цитотоксины, такие как перфорин, гранзимы и гранулизин. Перфорин и гранулизин приводят к образованию пор в клетке-мишени и гранзимы попадают в клетку и запускают каспазный каскад в цитоплазме, который инициирует апоптоз (программированную гибель клеток) клетки. Во-вторых, апоптоз может быть индуцирован через взаимодействие Fas-Fas-лиганда между Т-клетками и опухолевыми клетками-мишенями. Т-клетки и другие цитотоксические лимфоциты предпочтительно являются аутологичными клетками, хотя могут быть использованы гетерологичные клетки или аллогенные клетки.
Соответственно, агенты, композиции и способы, описанные в данной заявке, могут быть использованы для лечения субъекта, имеющего заболевание, например, заболевание, характеризуемое наличием пораженных клеток, экспрессирующих антиген. Особенно предпочтительными заболеваниями являются раковые заболевания.
Агенты, композиции и способы, описанные в данной заявке, могут быть использованы для иммунизации или вакцинации при профилактике заболевания, описанного в данной заявке.
Термин "заболевание" относится к аномальному состоянию, которое поражает организм индивидуума. Заболевание часто интерпретируется как медицинское состояние, ассоциированное со специфичными симптомами и признаками. Заболевание может быть первоначально вызвано факторами от внешнего источника, такими как инфекционное заболевание, или может быть вызвано дисфункциями внутренних органов, например, это могут быть аутоиммунные заболевания. У людей термин "заболевание" часто применяется более широко и относится к любому состоянию, которое вызывает боль, дисфункцию, тяжелое недомогание (дистресс), социальные проблемы или смерть больного индивидуума, или похожие проблемы для тех, кто находится в контакте с индивидуумом. В этом широком смысле термин иногда включает повреждения, инвалидность, расстройства, синдромы, инфекции, изолированные симптомы, отклонение от норм поведения и атипичные вариации структуры и функции, в то время как в другом контексте и для других целей эти понятия могут рассматриваться как различимые категории. Заболевания обычно поражают индивидуумов не только физически, но также эмоционально, так как заражение и жизнь со многими болезнями может изменить жизненную перспективу и личность субъекта. Согласно данному изобретению термин "заболевание" включает инфекционные болезни и раковые заболевания, в частности те виды рака, которые описаны в данной заявке. В данной заявке любая отсылка к раку или к конкретным видам рака включает также метастазирующий рак.
Заболевание, которое подвергается лечению согласно данному изобретению, предпочтительно представляет собой заболевание с участием антигена. Выражения "заболевание с участием антигена", "заболевание, ассоциированное с экспрессией или усиленной экспрессией антигена" или похожие выражения согласно настоящему изобретению означают, что этот антиген экспрессируется в клетках больной ткани или больного органа. Экспрессия в клетках больной ткани или больного органа может быть усилена по сравнению с состоянием здоровой ткани или здорового органа. Согласно одному варианту экспрессия обнаруживается только в пораженной ткани, в то время как экспрессия в здоровой ткани не выявляется, например, экспрессия может быть подавлена. Согласно данному изобретению, заболевания с участием антигена включают инфекционные болезни и раковые заболевания, при этом антиген, ассоциированный с заболеванием, предпочтительно представляет собой антиген инфекционного агента и опухолевый антиген, соответственно. Предпочтительно, когда заболевание с участием антигена является заболеванием с участием клеток, экспрессирующих антиген, предпочтительно на поверхности клеток.
Термин "здоровый" или "нормальный" относится к непатологическим состояниям и предпочтительно означает неинфекционный или нераковый.
Термины "раковое заболевание" или "рак" относятся к или описывают физиологическое состояние индивидуума, которое обычно характеризуется нерегулируемым ростом клеток. Примеры раковых заболеваний включают, но без ограничения, карциному, лимфому, бластому, саркому и лейкоз. Более конкретно, примеры таких раковых заболеваний включают рак кости, рак крови, рак легкого, рак печени, рак поджелудочной железы, рак кожи, рак головы или шеи, меланому кожи или интраокулярную меланому, рак матки, рак яичника, ректальный рак, рак заднего прохода, рак желудка, рак толстой кишки, рак молочной железы, рак простаты, рак матки, карциному половых и репродуктивных органов, болезнь Ходжкина, рак пищевода, рак тонкой кишки, рак эндокринной системы, рак щитовидной железы, рак паращитовидной железы, рак надпочечной железы, саркому мягких тканей, рак мочевого пузыря, рак почки, почечноклеточный рак, карциному почечной лоханки, новообразования центральной нервной системы (CNS), нейроэктодермальный рак, опухоли позвоночника, глиому, менингиому и аденому гипофиза. Термин "рак" согласно данному изобретению охватывает также метастазирующий рак. Предпочтительно "раковое заболевание" характеризуется наличием клеток, экспрессирующих опухолевый антиген, и раковая клетка экспрессирует опухолевый антиген.
Согласно одному варианту раковое заболевание является злокачественным заболеванием, которое характеризуется признаками анаплазии, инвазивности и наличием метастазов. Злокачественная опухоль может противопоставляться нераковой доброкачественной опухоли, так как само злокачественное образование не ограничивается ростом, способно внедряться в соседние ткани и может быть способно распространяться в удаленных тканях (метастазировать), в то время как доброкачественная опухоль не имеет ни одного из этих свойств.
Согласно данному изобретению термин "опухоль" или "опухолевая болезнь" относится к набуханию или опухолевому очагу, образованным за счет аномального роста клеток (называемых неопластическими клетками или опухолевыми клетками). Под "опухолевой клеткой" подразумевается аномальная клетка, которая растет за счет быстрой неконтролируемой клеточной пролиферации и продолжает расти после стимуляции, которая инициирует остановку развития новообразований. Опухоли демонстрируют частичное или полное отсутствие структурной организации и функциональной координации с нормальной тканью, и обычно образуют четко выраженную массу ткани, которая может быть или доброкачественной, или предзлокачественной, или злокачественной.
Согласно изобретению "карцинома" представляет собой злокачественную опухоль, развившуюся из клеток эпителиальной ткани. Эта группа опухолей представляет собой наиболее распространенные виды рака, включая обычные формы рака молочной железы, простаты, легкого и толстой кишки.
"Аденокарцинома" является опухолью, которая разрастается в железистой ткани. Эта ткань является также частью ткани, известной как эпителиальная ткань. Эпителиальная ткань включает кожу, железы и разнообразные другие ткани, которые обрамляют полости и органы тела. Эпителий образуется эмбриологически из эктодермы, эндодермы и мезодермы. При классификации как аденокарцинома клетки не должны обязательно быть частью железы, пока они имеют секреторные свойства. Эта форма карциномы может возникать у некоторых высших млекопитающих, включая людей. Хорошо дифференцированные аденокарциномы имеют тенденцию походить на железистую ткань, из которой они разрослись, в то время как плохо дифференцированные аденокарциномы могут не обладать этим свойством. Путем окрашивания клеток, полученных при биопсии, патолог определяет, является ли опухоль аденокарциномой или некоторым другим видом рака. Аденокарциномы могут развиться во многих тканях организма вследствие убиквитарной природы желез в организме. Хотя каждая железа может не секретировать одно и то же вещество, пока существует экзокринная функция в клетке, она считается железистой и поэтому ее злокачественная форма называется аденокарциномой. Злокачественные аденокарциномы проникают в другие ткани и часто метастазируют, если на это было достаточно времени. Аденокарцинома яичника является наиболее распространенным видом карциномы яичника. Она включает серозные и муцинозные аденокарциномы, аденокарциному прозрачных клеток и эндометриоидную аденокарциному.
Лимфома и лейкоз являются заболеваниями злокачественного характера, развивающимися из гемопоэтических (кроветворных) клеток.
Бластическая опухоль или бластома представляет собой опухоль (обычно злокачественную), которая напоминает незрелую или эмбриональную ткань. Многие из этих опухолей распространены у детей.
Под "метастазом" подразумевают распространение раковых клеток из их первоначального очага в другую часть организма. Образование метастаза является очень сложным процессом и зависит от отрыва злокачественных клеток от первичной опухоли, инвазии экстрацеллюлярной матрицы, проникновения эндотелиальной базальной мембраны в полость тела и в сосуды и затем после переноса кровью инфильтрации органов-мишеней. В результате рост новой опухоли в сайте-мишени зависит от ангиогенеза. Метастазы опухоли часто возникают даже после удаления первичной опухоли, потому что опухолевые клетки или компоненты могут оставаться и развивать метастатический потенциал. Согласно одному из вариантов данного изобретения термин "метастаз" относится к "отдаленному метастазу", который подразумевает метастаз, удаленный от первичной опухоли и системы регионарного лимфатического узла. Согласно одному из вариантов термин данного изобретения "метастаз" относится к метастазам в лимфоузлах.
Возврат болезни или рецидив возникает, когда субъект снова страдает от состояния, которое поражало его в прошлом. Например, если пациент страдал от опухолевой болезни, получил успешное лечение указанной болезни и у него снова развилась эта болезнь, указанная вновь развившаяся болезнь может рассматриваться как возврат или рецидив. Однако согласно данному изобретению возврат болезни или рецидив опухолевой болезни может, но не обязательно должен, возникать в сайте первоначальной опухолевой болезни. Таким образом, например, если пациент страдал от опухоли яичника и получил успешное лечение, возврат болезни или рецидив может быть возникновением опухоли яичника или возникновением опухоли в сайте не в области яичника. Возврат болезни или рецидив опухоли также включает ситуации, когда опухоль возникает в сайте, отличающемся от сайта первоначальной опухоли, а также в сайте первоначальной опухоли. Предпочтительно, когда исходная опухоль, от которой пациент получил лечение, представляет собой первоначальную опухоль, и опухоль в сайте, отличном от сайта первоначальной опухоли, является вторичной или метастатической опухолью.
Инфекционные болезни, лечение или профилактика которых возможны согласно настоящему изобретению, вызваны инфекционными агентами, включающими, но без ограничения, вирусы, бактерии, грибы, простейшие, гельминты и паразиты.
Инфекционные вирусы и людей, и позвоночных, не принадлежащих к человеческому роду, включают ретровирусы, РНК-вирусы и ДНК-вирусы. Примеры вирусов которые были обнаружены у людей, включают, но без ограничения: Retroviridae (например, вирусы иммунодефицита человека, такие как as HIV-1 (ВИЧ-1) (обозначаемые также как HTLV-III, LAV или HTLV-III/LAV, или HIV-III); и другие изоляты, такие как HIV-LP; Picornaviridae (например, полиовирусы, вирус гепатита А; энтеровирусы, вирусы Коксаки у человека, риновирусы, эховирусы); Calciviridae (например, штаммы, которые вызывают гастроэнтерит; Togaviridae (например, вирусы конского энцефалита, вирусы краснухи); Flaviridae (например, вирусы геморрагической лихорадки денге, вирусы энцефалита, вирусы желтой лихорадки); Coronaviridae (например, коронавирусы); Rhabdoviridae (например, вирусы везикулярного стоматита, вирусы бешенства); Filoviridae (например, вирусы Эбола); Paramyxoviridae (например, вирусы парагриппа, вирусы свинки, вирусы кори, респираторный синцитиальный вирус); Orthomyxoviridae (например, вирусы гриппа); Bungaviridae (например, хантавирусы, вирусы bunga, флебовирусы и вирусы Nairo); Arena viridae (вирусы геморрагической лихорадки); Reoviridae (например, реовирусы, орбивирусы и ротавирусы); Birnaviridae; Hepadnaviridae (вирус гепатита В); Parvovirida (парвовирусы); Papovaviridae (вирусы папилломы, полиомавирусы); Adenoviridae (в основном аденовирусы); Herpesviridae (вирус простого герпеса (HSV) (ВПГ) 1 и 2, вирус ветряной оспы, цитомегаловирус (CMV), вирус герпеса; Poxyiridae (вирусы оспы, вирусы осповакцины, поксвирусы); и Iridoviridae (например, вирус африканской лихорадки свиней); и неклассифицированные вирусы (например, этиологические агенты губчатой энцефалопатии животных, вирус гепатита дельта (считался дефектным сателлитом вируса гепатита В), агенты гепатита не А, гепатита не В (класс 1 = переданные внутренним органам; класс 2 = парентерально переданные (то есть, гепатита С); вирусы Norwalk и родственные вирусы, и астровирусы).
Ретровирусы, которые охвачены данным изобретением, включают и простые ретровирусы, и сложные ретровирусы. Сложные ретровирусы включают подгруппы лентивирусов, вирусов Т-клеточного лейкоза и спумавирусы (пенящие вирусы). Лентивирусы включают HIV-1, а также включают HIV-2, SIV, вирус Висна, вирус иммунодефицита кошек (FIV) и вирус инфекционной анемии у лошадей (EIAV). Вирусы Т-клеточного лейкоза включают HTLV-1, HTLV-II, вирус Т-клеточного лейкоза у обезьян (STLV) и вирус коровьего лейкоза (BLV). Пенящие вирусы включают пенящие вирусы человека (HFV), пенящие вирусы обезьян (SFV) и пенящие вирусы коров (BFV).
Бактериальные инфекции или болезни, лечение или профилактику которых можно проводить согласно настоящему изобретению, вызываются бактериями, включающими, но без ограничения, бактерии, которые обладают внутриклеточной стадией жизненного цикла, такие как микобактерии (например, Mycobacteria tuberculosis, М. bovis, М. avium, М leprae или М. africanum), риккетсии, микоплазма, хламидии и легионеллы. Другие примеры бактериальных инфекций по данному изобретению включают, но без ограничения, инфекции, вызванные грамположительными бациллами (например, Listeria, Bacillus, такие как Bacillus anthracis, Erysipelothrix species), грамотрицательными бациллами (например, Bartonella, Brucella, Campylobacter, Enterobacter, Escherichia, Francisella, Hemophilus, Klebsiella, Morganella, Proteus, Providencia, Pseudomonas, Salmonella, Serratia, Shigella, Vibrio и Yersinia species), спирохетами (например, род Borrelia, включая Borrelia burgdorferi, которые вызывают болезнь Лайма), анаэробными бактериями (например, Actinomyces и Clostridium species), грамположительными и грамотрицательными кокковыми бактериями, бактериями рода Enterococcus, рода Streptococcus, рода Pneumococcus, рода Staphylococcus, рода Neisseria. Конкретные примеры инфекционных бактерий включают, но без ограничений: Helicobacter pyloris, Borelia burgdorferi, Legionella pneumophilia, Mycobacteria tuberculosis, M. avium, M. intracellulare, M. kansaii, M. gordonae, Staphylococcus aureus, Neisseria gonorrhoeae, Neisseria meningitidis, Listeria monocytogenes, Streptococcus pyogenes (Streptococcus группы A), Streptococcus agalactiae (Streptococcus группы В), Streptococcus viridans, Streptococcus aecalis, Streptococcus bovis, Streptococcus pneumoniae, Haemophilus influenzae, Bacillus antracis, Corynebacterium diphtheriae, Erysipelothrix rhusiopathiae, Clostridium perfringers, Clostridium tetani, Enterobacter aerogenes, Klebsiella pneunmoniae, Pasturella multocida, Fusobacterium nucleatuin, Streptobacillus moniliformis, Treponema pallidium, Treponema pertenue, Leptospira, Rickettsia и Actinoyyces israelli.
Грибковые заболевания, лечение которых и профилактику можно проводить согласно настоящему изобретению, включают, но без ограничения, аспергилез, критококкоз, споротрихоз, кокцидиоидомикоз, паракокцидиоидомикоз, гистоплазмоз, бластомикоз, зигомикоз и кандидоз.
Паразитарные заболевания, лечение которых и профилактику можно проводить согласно настоящему изобретению, включают, но без ограничения, амёбиаз, малярию, лейшманиоз, кокцидиоз, лямблиоз, криптоспоридиоз, токсоплазмоз и трипаносомиаз. Данным изобретением охвачены также инфекции, вызываемые различными гельминтами, такие как, но без ограничения, аскаридоз, анкилостомидоз, трихуриаз, стронгилоидоз, токсокароз, трихиноз, онхоциркоз, филяриоз и дирофиляриоз. Охвачены также инфекции, вызванные различными двуустками, такие как, но без ограничения, шистосомиаз, парагонимоз и клонорхоз.
Термин "лечение" или "терапевтическое лечение" относится к любому виду лечения, который улучшает состояние здоровья и/или пролонгирует (увеличивает) продолжительность жизни субъекта. Такое лечение может устранять заболевание субъекта, останавливать или замедлять развитие заболевания у субъекта, ингибировать или замедлять развитие болезни у субъекта, снижать частоту или тяжесть симптомов у субъекта и/или уменьшать рецидивы у субъекта, который болен или ранее перенес болезнь.
Термины "профилактическое лечение" или "превентивное лечение" относятся к любому виду лечения, которое предназначено для предотвращения возникновения заболевания у субъекта. Термины "профилактическое лечение" или "превентивное лечение" используются в данной заявке как взаимозаменяемые.
Термины "индивидуум" и "субъект" используются в данной заявке как взаимозаменяемые. Они относятся к людям, приматам, не относящимся к человеческому роду, или другим млекопитающим (например, к мыши, крысе, кролику, собаке, кошке, крупному рогатому скоту, свинье, овце, лошади или примату), которые могут быть поражены болезнью или расстройством (например, раковым заболеванием) или восприимчивы к ним, но могут или не могут болеть этой болезнью или расстройством. Согласно многим вариантам индивидуум представляет собой человека. Если иное не указано, термины "индивидуум" и "субъект" не означают определенный возраст и поэтому охватывают взрослых людей, пожилых людей, детей и младенцев. Согласно предпочтительным вариантам настоящего изобретения "индивидуум" или "субъект" представляет собой "пациента". Термин "пациент" согласно данному изобретению означает субъекта для проведения лечения, в частности больного субъекта.
Под выражением "рискующий заболеть" подразумевают субъекта, то есть пациента, который идентифицирован как имеющий более высокую, чем обычная, вероятность развития заболевания, в частности рака, по сравнению с общей популяцией. Кроме того, субъект, который уже болел или который болеет в настоящее время заболеванием, в частности раком, является субъектом, который имеет повышенный риск к развитию заболевания, поскольку у этого субъекта может продолжать развиваться заболевание. Субъекты, которые в настоящее время больны раком или которые болели раком, также имеют повышенный риск развития раковых метастазов.
Термин "иммунотерапия" относится к лечению, включающему специфический иммунную реакцию или иммунный ответ.
В контексте настоящего изобретения такие термины как "защитить", "предотвратить", "профилактический", "превентивный" или "защитный" относятся к предотвращению или лечению, или и к возникновению, и/или распространению болезни у субъекта и, в частности, к минимизации вероятности того, что у субъекта разовьется заболевание, или к задержке развития заболевания. Например, субъект, рискующий заболеть опухолевым заболеванием, как описано выше, будет являться кандидатом для терапии для предотвращения развития опухоли.
Профилактическое применение иммунотерапии, например, профилактическое применение агента или композиции по изобретению, предпочтительно защищает реципиента от развития заболевания. Терапевтическое применение иммунотерапии, например, терапевтическое применение агента или композиции по изобретению, может привести к ингибированию развития/эскалации заболевания. Это включает замедление развития/эскалации заболевания, в частности нарушение прогрессирования заболевания, которое предпочтительно приводит к устранению заболевания.
Иммунотерапия может проводиться с использованием различных методов, в которых агенты, предусмотренные в данной заявке, предпочтительно функционируют для удаления антиген-экспрессирующих клеток у пациента. Такое удаление может иметь место в результате усиления или индуцирования иммунного ответа у пациента, специфического для антигена или клетки, экспрессирующей антиген.
Термин "иммунизация" или "вакцинация" описывает процесс лечения субъекта с целью индуцирования иммунного ответа по терапевтическим или профилактическим причинам.
Термин "in vivo" относится к действиям на живом организме.
Антигенные рецепторы, пептидные цепи, нуклеиновые кислоты, рекомбинантные клетки, иммунные эффекторные клетки, предпочтительно Т-клетки, по изобретению, а также другие соединения и агенты, описанные в данной заявке, могут быть введены в виде любой подходящей фармацевтической композиции.
Фармацевтические композиции по изобретению предпочтительно являются стерильными и содержат эффективное количество агентов, описанных в данной заявке, и необязательно других агентов, обсужденных в данной заявке и генерирующих желательную реакцию или желательный эффект.
Фармацевтические композиции обычно получаются в виде однородной лекарственной формы и могут быть приготовлены способом, известным per se. Фармацевтическая композиция может, например, быть в виде раствора или суспензии.
Фармацевтическая композиция может включать соли, буферные вещества, консерванты, носители, разбавители и/или эксципиенты, все из которых предпочтительно являются фармацевтически приемлемыми. Термин "фармацевтически приемлемый" относится к нетоксичному материалу, который не взаимодействует с активным компонентом фармацевтической композиции.
Соли, которые не являются фармацевтически приемлемыми, могут быть использованы для получения фармацевтически приемлемых солей и включены в объем изобретения. Фармацевтически приемлемые соли этого вида включают, без ограничения, соли, приготовленные из следующих кислот: соляной, бромистоводородной, серной, азотной, фосфорной, малеиновой, уксусной, салициловой, лимонной, муравьиной, малоновой, янтарной и т.п. Фармацевтически приемлемые соли могут также быть получены в виде солей щелочных металлов или солей щелочноземельных металлов, таких как соли натрия, соли калия или соли кальция.
Подходящие буферные вещества для применения в фармацевтической композиции включают уксусную кислоту и ее соль, лимонную кислоту и ее соль, борную кислоту и ее соль и фосфорную кислоту и ее соль.
Подходящие консерванты для применения в фармацевтической композиции включают бензалконийхлорид, хлорбутанол, парабен и тимеросал.
Состав для инъекций может включать фармацевтически приемлемый эксципиент, такой как лактат Рингера.
Термин "носитель" относится к органическому или неорганическому компоненту природного происхождения или синтетическому, в котором содержится активный компонент для того, чтобы облегчить, улучшить или обеспечить применение. Согласно данному изобретению термин "носитель" включает также один или более твердых или жидких наполнителей, разбавителей или инкапсулирующих веществ, которые подходят для введения пациенту.
Возможными носителями для парентерального введения являются, например, стерильная вода, раствор Рингера, лактат Рингера, стерильный раствор хлористого натрия, полиалкиленгликоли, гидрированные нафталины и, в особенности, биосовместимые полилактиды, сополимеры лактидов с гликолидами или сополимеры полиоксиэтилена/полиоксипропилена.
Термин "эксципиент" при использовании в данном изобретении предназначен для указания всех веществ, которые могут содержаться в фармацевтической композиции и которые не являются активными ингредиентами, таких как, например, носители, связующие, смазывающие вещества, загустители, поверхностно-активные агенты, консерванты, эмульгаторы, буферные вещества, вкусовые вещества или красители.
Агенты и композиции, описанные в данной заявке, могут быть введены любым обычным путем, таким как парентеральное введение, включая инъекции или инфузию. Введение осуществляют предпочтительно парентерально, например, внутривенно, интраартериально, подкожно, интрадермально или внутримышечно.
Композиции пригодные для парентерального введения, обычно содержат стерильные водный и неводный препараты активного соединения, которые предпочтительно являются изотоничными крови пациента. Примерами совместимых носителей и растворителей являются раствор Рингера и изотонический раствор хлористого натрия. Кроме того, в качестве растворимой или суспензионной среды используются обычно стерильные нелетучие масла.
Агенты и композиции, описанные в данной заявке, вводятся в эффективных количествах. Термин "эффективное количество" относится к количеству, которое обеспечивает желательный ответ или желательный эффект само по себе или вместе с дополнительными средствами. В случае лечения конкретной болезни или конкретного состояния желательный ответ предпочтительно относится к ингибированию развития болезни. Это включает замедление прогрессирования болезни и, в частности, перерыв или обращение прогрессирования болезни. Желательный ответ при лечении заболевания или состояния может также представлять собой задержку начала или предотвращение начала указанного заболевания или указанного состояния.
Эффективное количество агента или композиции, описанных в данной заявке, будет зависеть от состояния, подвергающегося лечению, степени серьезности заболевания, индивидуальных параметров пациента, включающих возраст, физиологическое состояние, рост и вес, продолжительность лечения, вид сопутствующей терапии (если она проводится), конкретного пути введения и подобных факторов. Соответственно, дозы вводимых агентов, описанных в данной заявке, могут зависеть от таких различных параметров. В случае, когда ответ пациента после введения первоначальной дозы является недостаточным, могут быть использованы более высокие дозы (или фактически более высокие дозы, полученные при другом, более локализованном способе введения).
Агенты и композиции, описанные в данной заявке, могут вводиться пациентам, например, in vivo, для лечения или предотвращения ряда заболеваний, таких как описанные в данной заявке. Предпочтительные пациенты включают людей, имеющих патологии, которые можно корригировать или ослабить их проявление путем введения агентов и композиций, описанных в данной заявке. Это включает патологические состояния с участием клеток, характеризуемых экспрессией антигена.
Например, согласно одному примеру в одном варианте агенты, описанные в данной заявке, могут быть использованы для лечения пациента с раковым заболеванием, например, раковым заболеванием, таким как описанные в данной заявке, характеризуемым наличием раковых клеток, экспрессирующих антиген.
Фармацевтические композиции и способы лечения согласно данному изобретению могут быть также использованы для иммунизации или вакцинации с целью предотвращения заболевания, описанного в данной заявке.
Фармацевтические композиции по изобретению могут вводиться вместе с веществами, повышающими и укрепляющими иммунитет, такими как один или более адъювантов, и могут содержать одно или более веществ, повышающих иммунитет для дальнейшего повышения эффективности, предпочтительно для достижения синергического эффекта иммуностимуляции. Термин "адъювант" относится к соединениям, которые пролонгируют или усиливают, или ускоряют иммунный ответ. В этом отношении возможны различные механизмы, зависящие от различных видов адъювантов. Например, соединения, которые обеспечивают созревание DC, например, липополисахариды или лиганд CD40, образуют первый класс подходящих адъювантов. Обычно любой агент, который влияет на иммунную систему, типа "молекулярного фрагмента, ассоциированного с опасностью" (LPS, GP96, dsRNA и т.д.) или цитокины, такие как GM-CSF, могут быть использованы в качестве адъюванта, который обеспечивает интенсификацию иммунного ответа и/или влияет регулируемым образом. CpG-олигодезоксинуклеотиды могут быть также использованы в этом контексте, хотя их побочные эффекты, которые проявляются при некоторых обстоятельствах, как объяснялось выше, должны приниматься во внимание. Особенно предпочтительными адъювантами являются цитокины, такие как монокины, лимфокины, интерлейкины или хемокины, например, IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-12, IFNα, IFNγ, GM-CSF, LT-α или факторы роста, например, hGH. Другими известными адъювантами являются гидроксид алюминия, адъювант Фрейнда или масло, такое как Montanide®, наиболее предпочтителен Montanide® ISA51. Липопептиды, такие как Pam3Cys, также пригодны для использования в качестве адъювантов в фармацевтических композициях по данному изобретению.
Фармацевтическая композиция может быть введена локально или системно, предпочтительно системно.
Термин "системное введение" относится к введению агента таким образом, что этот агент равномерно распределяется в организме индивидуума в значительных количествах и обеспечивает желательный эффект. Например, такой агент может обеспечить желательный эффект в крови и/или достигнуть желательного сайта действия через сосудистую систему. Типичные системные пути введения включают введение путем интродуцирования агента непосредственно в сосудистую систему или перорально, в легкое или внутримышечно, при этом этот агент адсорбируется, попадает в сосудистую систему и доставляется в один или более желательных сайтов действия через кровь.
Согласно настоящему изобретению предпочтительно, чтобы системное введение осуществлялось парентерально. Термин "парентеральное введение" относится к введению агента таким образом, что агент не проходит через кишечник. Термин "парентеральное введение" включает внутривенное введение, подкожное введение, интрадермальное введение или интраартериальное введение, но этим не ограничивается.
Введение может также осуществляться, например, перорально, интраперитонеально или внутримышечно.
Агенты и композиции, предусмотренные в данной заявке, могут быть использованы сами по себе или в комбинации с обычными терапевтическими схемами лечения, такими как хирургическое вмешательство, облучение, химиотерапия и/или трансплантация костного мозга (аутологичная, сингенная, аллогенная или несвязанная).
Настоящее изобретение описано подробно ниже с помощью фигур и примеров, которые использованы только для иллюстративных целей и не являются ограничивающими. Благодаря описанию и примерам специалисту в данной области доступны другие варианты, которые также включены в данное изобретение.
ФИГУРЫ
На Фигуре 1 схематически представлены все конструкции Т-клеточный рецептор (TCR)- и химерный антигенный рецептор (CAR), применяемые в экспериментах. А) и В) TCR состоит из гетеродимерного мембранного белка класса I, причем каждая цепь содержит консервативный С-домен и вариабельный V-домен, где последний специфически распознает процессированный пептид с ограничением по МНС. Гомология последовательностей и структурная гомология мышиного (Mu, А) и человеческого (Hu, В) TCR является повышенной. Мышиный TCR распознает опухолевый антиген человека, происходящий из белка плотных контактов Claudin 6, тогда как человеческий TCR распознает опухолевый антиген, полученный с использованием меланоцитарного дифференцировочного антигена gp100. С) Одновалентный одноцепочечный CAR содержит одноцепочечный (sc) Fv-фрагмент, прикрепленный к мышиному Сβ-домену, и автономный TCR Сα-домен с предшествующим сигнальным пептидом для экспорта в клеточную мембрану. Необязательно, он связан искусственной дисульфид ной связью между TCR С-доменами для повышения экспрессии на клеточной поверхности и функционирования этого CAR. scFv-Фрагмент в этом и всех последующих иллюстративных конструкциях направлен на Claudin 6. D) Классический scCAR Cl6 содержит гомодимер, причем каждый располагается на одной линии с scFv-фрагментом, шарнирной областью антитела, СН2СН3-доменами в качестве спейсерной области, клеточной мембраной и доменами передачи сигнала внутрь клетки костимулирующей молекулы CD28 и CD3ζ, соответственно. Гомодимеризация приводит к распознаванию двухвалентным рецептором антигена, причем каждая цепь связывает один антиген (т.е. образуется внутрицепная связь). Е) Прототипный комбинированный CAR несет связанные с аллелем V-домены, соединенные последовательно (т.е. VH-VH или VL-VL), и тот и другой прикреплены к TCR Сα или Cβ, соответственно. Распознавание неизбежно влечет за собой связывание антигена с участием обеих цепей комбинированным (т.е. межцепным) способом и в виде двухвалентного соединения. F) Гетеродимерный TCR-CAR Cl6, несущий scFv-фрагменты на каждом полноразмерном TCRα gp100 или TCRβ распознает родственный антиген в виде двухвалентного, но не комбинированного соединения. G) Чтобы исключить распознавание остатка родственного пептида gp 100(280-288) компонентом TCR gp100 некомбинированного TCR-CAR Cl6 (F), ‘молчащую’ (sil) S109Q (согласно номенклатуре IMGT) точечную мутацию вводили в петлю CDR3 цепи TCRα (silCDR3α). Н) Комбинированный TCR-CAR Cl6 получали последовательным соединением связанных с аллелем V-доменов, либо на цепи полноразмерного TCR α gp100, либо на цепи полноразмерного TCR β соответственно, как изложено в Е). Полноразмерный TCR, применяемый в данном случае в качестве партнера по связыванию, может обеспечить лучшую передачу сигнала в Т- клетку в физиологических условиях, чем усеченный TCR, содержащий только TCR С-домены вместо (Е). I) Чтобы исключить распознавание остатка родственного пептида gp100(280-288) компонентом TCR gp100 комбинированного TCR-CAR Cl6 (Н), ‘молчащую’ (sil) S109Q (согласно номенклатуре IMGT) точечную мутацию вводили в петлю CDR3 цепи TCR α (silCDR3 α), как в G).
На Фигуре 2А/В показана экспрессия Claudin 6 на APCs и различных CARs на человеческих Т-клетках, соответственно, перед постановкой сокультивирования. А) Незрелые дендритные клетки (iDCs) электропорировали с увеличивающимися количествами родственного полноразмерного антигена Claudin 6 или с единичной высокой дозой нерелевантного антигена gp100. В данном случае методом проточной цитометрии можно было наблюдать дозозависимый сдвиг массива данных по Cl6-экспрессии, указывающий, что iDCs этого донора являются в высшей степени пермиссивными в отношении захвата клеточной РНК, трансляции белка и экспорта на поверхность клетки. Высокий уровень экспрессии CD86 указывает на эффективную дифференцировку моноцитов в успешно антигенпрезентирующие iDCs. В) Предварительно активированные CD8+ Т-клетки электропорировали с различными CAR-кодирующими РНК и анализировали экспрессию CAR методом проточной цитометрии. Все CARs, за исключением одновалентного и классического scCAR, умеренно восстанавливались при антиидиотипическом окрашивании. Одновалентный CAR экспрессировался хуже всего, тогда как было найдено, что классический scCAR экспрессировался лучше всего. На Фигуре 2С показана эффективность CAR Cl6-перепрограммированных человеческих Т-клеток при распознавании экспрессирующих Claudin 6 клеток iDCs после постановки сокультивирования АРС/Т-клеток в IFNγ-ELISA. CAR-электропорированные Т-клетки совместно культивировали в течение ночи с Cl6-электропорированными APCs, как объясняется на Фигурах 2А/В при соотношении Е:Т (отношение эффекторных клеток к клеткам-мишеням), равном 10:1. По всему спектру Cl6 титрования все CARs показывали оптимальный результат при пониженных дозах Cl6 (0.02 мкг). При высоких уровнях экспрессии Claudin 6 классический scCAR Cl6 продемонстрировал наилучшую секрецию IFNγ по сравнению со всеми другими CARs, тогда как при самой низкой дозе Cl6 комбинированные CARs проявляли себя, как правило, лучше, чем классический scCAR. И в заключение, комбинированные CARs превосходили классический CAR по своим функциональным эффективностям от высокой до низкой экспрессии антигена: Они давали очень высокие количества IFNγ вплоть до 20-30.000 пг/мл IFNγ для комбинированных CARs и scCAR Cl6, соответственно, и заканчивая 12-15.000 пг/мл IFNγ для всех конструкций при самой низкой дозе Cl6. При этой дозе интеркомбинированный Сα/Сβ-CAR становился наиболее эффективным.
На Фигуре 3А/В показана экспрессия Claudin 6 на APCs и различных CARs на человеческих Т-клетках, соответственно, перед постановкой сокультивирования. А) Незрелые дендритные клетки (iDCs) электропорировали с увеличивающимися количествами родственного полноразмерного антигена Claudin 6. В данном случае методом проточной цитометрии можно было наблюдать только дозозависимый частичный сдвиг экспрессии Cl6, показывающий, что iDCs данного донора, бесспорно, являются менее пермиссивными в отношении захвата клеточной РНК, трансляции белка и экспорта на поверхность клетки. Высокий уровень экспрессии CD86 указывает на эффективную дифференцировку моноцитов в успешно антигенпрезентирующие iDCs. В) Предварительно активированные CD8+ Т-клетки электропорировали с различными РНК, кодирующими CAR, и анализировали экспрессию CAR методом проточной цитометрии. Все CARs, за исключением одновалентного и классического scCAR, лишь отчасти восстанавливались при антиидиотипическом окрашивании, опять же указывая на то, что клетки этого донора являются менее пермиссивными в отношении захвата РНК и процессирования. Однако плохо презентирующие антиген iDCs могут представлять состояние минимально антигенпозитивных опухолевых APCs, поскольку оно может имитировать ситуацию вариантов избавления клона от опухоли на ранней стадии или минимальной презентации опухолевого антигена. На Фигуре 3С показана эффективность CAR Cl6-перепрограммированных человеческих Т-клеток при распознавании экспрессирующих Claudin 6 клеток iDCs после постановки совместного культивирования АРС/Т-клеток в IFNγ-ELISA. CAR-электропорированные Т-клетки совместно культивировали в течение ночи с Cl6-электропорированными APCs, как объясняется на Фигурах 3А/В, при соотношении Е:Т, равном 10:1. По всему спектру Cl6 титрования все CARs показывали оптимальный результат при пониженных дозах Cl6 (0.02 мкг). Учитывая абсолютно минимальную экспрессию Claudin 6, как объясняется выше, комбинированные CARs Cl6 продемонстрировали наилучшую IFNγ-секрецию по сравнению с классическим scCAR Cl6 для всех доз, тенденцию, которая была даже более четко выраженной при самой низкой дозе электропорированных Cl6. И в заключение, для очень маленьких количеств антигена комбинированные CARs были еще лучше, чем классический scCAR Cl6 в отношении IFNγ-секреции. Общие количества секретированного IFNγ опускались ниже 1.000 пг/мл по всему интервалу титрования вследствие очень низкой экспрессии антигена, а также CARs.
На Фигуре 4 показана эффективность TCR gp100-CAR Cl6 перепрограммированных человеческих Т-клеток в распознавании экспрессирующих Claudin 6 клеток iDCs или нагруженных gp100 пептидом iDCs после постановки совместного культивирования HLA-A2.1+ АРС/Т-клеток в IFNγ-ELISA. CAR-электропорированные Т-клетки совместно культивировали в течение ночи с Cl6-электропорированными APCs или с нагруженными пептидом gp100(280-288) iDCs при соотношении Е:Т, равном 5:1. Главной целью этого эксперимента было проверить, действительно ли, во-первых (А) двухвалентные комбинированные TCR-CARs более эффективно секретируют IFNγ, чем двухвалентные некомбинированные TCR-CARs, и во-вторых (В), в какой степени они все еще распознают gp100 пептид применяемой в данном изобретении структуры TCR gp100. Остаточное связывание с антигеном пытались исключить посредством использования ‘молчащей’ мутации S109Q в CDR3 структуры TCRα gp100. TCR Cl6 и TCR gp100 служили в качестве положительного контроля для когнатного распознавания антигена. А) Комбинированные TCR-CAR, независимо от того, сделали их молчащими в CDR3α или нет, оказались более функциональными, чем некомбинированные CARs, особенно при самой низкой дозе антигена. В) Не-молчащий некомбинированный TCR-CAR Cl6 все еще распознавал gp100 при 10-6 М пептида. Введение мутации S109Q полностью аннулировало секрецию. Для комбинированного TCR-CARs Cl6 совсем не наблюдалось никакой секреции, вне зависимости от того, был ли осуществлен функциональный сайленсинг или нет. Весьма вероятно, что межцепное связывание последовательно соединенных VH-VH- и VL-VL-доменов на любой цепи создает пространственные затруднения для связывания gp100 пептида, презентируемого в виде ограниченного HLA-A2.1 на APCs.
На Фигуре 5 показана способность к пролиферации различных CARs Cl6 в результате столкновения с Cl6-экспрессирующими iDCs после постановки совместного культивирования АРС/Т-клеток. CAR-электропорированные Т-клетки совместно культивировали в течение 5 дней с Cl6-электропорированными и - титрованными APCs при соотношении Е:Т, равном 10:1. Априори Т-клетки окрашивали карбоксифлуоресцеина сукцинимидиловым эфиром (CFSE), чтобы количественно определить титр CFSE при делении клеток и, следовательно, число полученных в результате дочерних популяций, указанных справа от каждого графика плотности распределения, полученного в анализе методом проточной цитометрии. А) В случае одновалентно связанных с антигеном CARs наблюдалась наиболее слабая пролиферация, тогда как комбинированные TCR-CARs и классические CARs показывали схожие паттерны пролиферации. В) Гистограмма частоты встречаемости популяций Т- клеток показана в А). Т-клетки с высокой частотой встречаемости, ‘снабженные’ одновалентным CAR, не пролиферировали (40-60%), тогда как частота встречаемости не-пролиферирующих Т-клеток, несущих комбинированные TCR-CARs и классический CAR, была примерно в интервале 10-20%. Пролиферация Т-клеток, несущих классический CAR Cl6, была наивысшей в высокой дозе (0.2 мкг Cl6) антигена (90% по сравнению с 80% для комбинированных TCR-CARs), тогда как в самой низкой дозе (0.002 мкг) комбинированные TCR-CARs были по меньшей мере так же эффективны, как и классический CAR (почти 80%). И в заключение: комбинированные TCR-CARs опять же превзошли классический CAR по своей функциональной эффективности, в данном случае в том, что касается Т-клеточной пролиферации, в интервале от высоких до очень низких доз антигена. Наблюдается тенденция в сторону еще более высокой эффективности, чем в случае классического CAR.
На Фигуре 6 показана позитивная регуляция биомаркера костимуляции CD27 на комбинированный TCR-CAR Cl6-электропорированных дочерних Т-клеток после постановки совместного культивирования АРС/Т-клеток. CAR-электропорированные Т- клетки совместно культивировали в течение 6 дней с Cl6-электропорированными iDCs при соотношении Е:Т, равном 3:1. Априори Т-клетки окрашивали с помощью CPD-450, чтобы количественно определить титр этого флуорофора при делении клеток и, следовательно, число и частоту встречаемости полученных в результате популяций дочерних клеток, указанные слева от гистограмм, полученных в анализе методом проточной цитометрии. Т-клетки окрашивали CD8-специфическим антителом, чтобы проиллюстрировать на примере умеренную и равную позитивную регуляцию этого корецептора-маркера для комбинированных и классических CARs. Одновременно их окрашивали антителом, специфическим к костимулятору CD27 для оценки его регуляции в популяциях родительских /дочерних клеток G0-G6 для всех CARs. Хотя оба маркера позитивно регулировались, приблизительно в 2 раза, и помимо этого средняя экспрессия CD8 была одинаковой для всех CARs, средняя экспрессия CD27 была намного выше для комбинированных CARs, и в частности для интеркомбинированного TCR-CAR Cl6, чем для классического CAR. CD27, биомаркер длительной устойчивости Т- клеток in vivo, достигал значительно более высокого плато для Т- клеток с затухающей пролиферацией (G2-G4) до того, как уровни экспрессии падали до базового уровня в G6.
ПРИМЕРЫ
Технические приемы и методы, применяемые в настоящем изобретении, описаны в данной заявке или осуществляются хорошо известными способами, описанными, например, в лабораторном практикуме Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd Edition (1989) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. Все методы, включающие применение наборов и реагентов, осуществляются согласно информации, представленной изготовителями, если конкретно не указано иное.
Пример 1: Анализ экспрессии Claudin 6 в iDCs и антигенных рецепторов в Т-клетках методом проточной цитометрии.
РНК для различных конструкций были получены с использованием in vitro транскрипции (ivt-RNA) открытых рамок считывания (ORF), клонированных в РНК-вектор pST1, несущий Т7 промотор на своей 5'-лидирующей последовательности и оптимизированный полиА-‘хвост’ на своем 3'-конце. Экспрессию Claudin 6 в человеческих незрелых дендритных клетках анализировали по прошествии одного дня после электропорации РНК (2-0.002 мкг, 300 В, 12 мс, 1 импульс) в обработанные с помощью GM-CSF/IL-4 CD14+моноциты лейкоцитарной пленки, применяя антитело, специфическое к Claudin 6, меченное флуорофором Dylight-650. Экспрессию различных конструкций антигенных рецепторов в аутологичных человеческих Т-клетках анализировали через день после электропорации РНК (всего 10-30 мкг для обеих цепей, 495 В, 9 мс, 1 импульс) в CD8+ Т-клетки, преактивированные OKT3 (мышиным моноклональным антителом к CD3ε), с использованием Cl6 scFv специфического к идиотипу антитела, меченного флуорофором Dylight-649. Подробное описание получения человеческих iDCs и Т-клеток дано в Примере 2. CAR-конструкции, экспрессию которых проверяли, представляли собой (i) мышиный Т-клеточный рецептор TCR Cl6; (ii) человеческий TCR gp100; (iii) одновалентный некомбинированный CAR Cl6; (iv) классический scCAR) (двухвалентный); (v) интер- комбинированный CAR Cl6, слитый с человеческими TCR Сα/β-доменами (двухвалентный); (vi) некомбинированный CAR Cl6, слитый с полноразмерным TCR gp 100(280-288) (двухвалентный); (vii) соответствующий некомбинированный CAR Cl6, также ‘молчащий’ в CDR3 TCRα gp100 для исключения распознавания пептида (двухвалентный); (viii) интер- комбинированный CAR Cl6, слитый с полноразмерным TCR gp 100(280-288) (двухвалентный) и (ix) соответствующий комбинированный CAR Cl6, также 'молчащий' в CDR3 TCRα gp100 для исключения распознавания пептида (двухвалентный). Различные конструкции антигенных рецепторов схематически показаны на Фигурах 1A-I. Окрашивание клеток согласно обычной практике осуществляли для 0.2×106 клеток в буфере для проточной цитометрии в течение 20 мин при 4°С, отмывали и фиксировали с помощью 1% параформальдегид-содержащего буфера для проточной цитометрии. Результаты, приведенные на Фигуре 2А, показывают титрованную экспрессию Claudin 6. В данном случае iDCs этого донора были в высокой степени пермиссивными в отношении захвата Cl6 RNA и демонстрировали сдвиг массива данных по экспрессии Claudin 6 с увеличивающимися количествами электропорированной РНК. Экспрессия CD86 показала успешную дифференцировку моноцитов в активные антигенпрезентирующие iDCs. На Фиг. 2В показана экспрессия различных CARs, применяемых в данном эксперименте, в человеческих CD8-позитивных Т-клетках. Классический scCAR Cl6 продемонстрировал наивысшую экспрессию, возможно, благодаря его эндогенной CD3- независимой экспрессии на поверхности Т-клеток. Комбинированные CARs показали несколько более высокую экспрессию, чем одновалентный CAR, причем последний служил в качестве ‘слабого контроля’ вследствие того, что он связывает антиген только как одновалентное соединение.
Результаты на Фигуре 3А показывают титрованную экспрессию Claudin 6 в независимом эксперименте. В данном случае, iDCs от этого донора были слабо пермиссивными в отношении захвата Cl6 RNA и продемонстрировали только частичный сдвиг экспрессии Claudin 6 с увеличивающимися количествами электропорированной RNA. Но экспрессия CD86 показала успешную дифференцировку моноцитов в активные презентирующие антиген iDCs. На Фиг. 3В показана экспрессия различных CARs, используемых в данном эксперименте, в выбранных CD8-позитивных человеческих Т-клетках. Так как человеческие Т-клетки были взяты у того же донора, что и дифференцированные моноциты при аутологичных параметрах, Т-клетки также становились слабо пермиссивными в отношении электропорированной РНК, и в результате наблюдалась лишь более слабая экспрессия для CARs, чем в эксперименте, показанном на Фигуре 2В. Однако, классический scCAR Cl6 снова демонстрировал наивысший уровень экспрессии по той же самой причине, указанной ранее. В соответствии с предыдущими наблюдениями комбинированные CARs показали несколько лучшую экспрессию, чем одновалентный CAR. Так как порядок экспрессии CAR сохраняется по сравнению с экспериментом, представленным на Фигуре 2А/В, этот эксперимент пригоден для изучения активности различных CARs в случае экспрессии лишь весьма малых количеств Cl6 на антигенпрезентирующих клетках (APCs).
Пример 2: Анализ секреции IFN-γ титрованием антигеном
В день 1 эксперимента свежие мононуклеарные клетки периферической крови ("PBMCs") выделяли из лейкоцитарной пленки здорового донора. Из  PBMCs клетки CD14+ выделяли, используя метод магнитной сортировки клеток, MACS. Затем поток MACS и остаточные PBMCs сортировали методом MACS, выделяя CD8+ Т-клетки. Клетки CD14+ дифференцировали в незрелые дендритные клетки ("iDCs") посредством введения IL-4 & GM-CSF (1000 ед/мл) на 1, 3, 6 день. CD8+ Т-клетки переносили в 6-луночные планшеты, покрытые антителом OKT3. На 3 день Т-клетки переносили в новые 6- луночные планшеты. На 7 день iDCs подвергали электропорации нерелевантной и Cl6 ivt-РНК в зависимости от дозы в интервале 2-0.002 мкг РНК. Активированные антителом OKT3 Т-клетки электропорировали в один день с контрольными или с конструкциями антигенного рецептора, представленными на отдельных фигурах или описанными в Примере 1. Для обеспечения контроля качества экспрессию Cl6 на клетках iDCs и поверхностную экспрессию антигенного рецептора на Т-клетках анализировали с использованием специфических меченных флуоресцентной меткой антител, описанных ранее, на 8 день. Затем электропорированные Т-клетки и антиген-электропорированные iDCs совместно культивировали в 96-луночном планшете в течение 20 ч при соотношении Е:Т, равном от 3:1 до 10:1, в двойных повторах. Согласно обычной практике 2.5×104 iDCs засевали и совместно культивировали с 7.5×104 - 2.5×105 CAR-электропорированных Т- клеток в объеме 200 мкл среды для Т-клеток. На 9 день отбирали различные количества культуральных супернатантов (10-50 мкл) и анализировали количество секретированного IFNγ методом "сэндвич" - ELISA, используя набор теста IFN-γ Ready Set Go! от кампании eBioscience (#88-7316-88). Оптическую плотность детектировали на ридере Tecan Sunrise ELISA.
PBMCs клетки CD14+ выделяли, используя метод магнитной сортировки клеток, MACS. Затем поток MACS и остаточные PBMCs сортировали методом MACS, выделяя CD8+ Т-клетки. Клетки CD14+ дифференцировали в незрелые дендритные клетки ("iDCs") посредством введения IL-4 & GM-CSF (1000 ед/мл) на 1, 3, 6 день. CD8+ Т-клетки переносили в 6-луночные планшеты, покрытые антителом OKT3. На 3 день Т-клетки переносили в новые 6- луночные планшеты. На 7 день iDCs подвергали электропорации нерелевантной и Cl6 ivt-РНК в зависимости от дозы в интервале 2-0.002 мкг РНК. Активированные антителом OKT3 Т-клетки электропорировали в один день с контрольными или с конструкциями антигенного рецептора, представленными на отдельных фигурах или описанными в Примере 1. Для обеспечения контроля качества экспрессию Cl6 на клетках iDCs и поверхностную экспрессию антигенного рецептора на Т-клетках анализировали с использованием специфических меченных флуоресцентной меткой антител, описанных ранее, на 8 день. Затем электропорированные Т-клетки и антиген-электропорированные iDCs совместно культивировали в 96-луночном планшете в течение 20 ч при соотношении Е:Т, равном от 3:1 до 10:1, в двойных повторах. Согласно обычной практике 2.5×104 iDCs засевали и совместно культивировали с 7.5×104 - 2.5×105 CAR-электропорированных Т- клеток в объеме 200 мкл среды для Т-клеток. На 9 день отбирали различные количества культуральных супернатантов (10-50 мкл) и анализировали количество секретированного IFNγ методом "сэндвич" - ELISA, используя набор теста IFN-γ Ready Set Go! от кампании eBioscience (#88-7316-88). Оптическую плотность детектировали на ридере Tecan Sunrise ELISA.
На Фигуре 2С показаны количества секретированного IFNγ для iDCs и Т-клеток одного и того же донора, которые были в высокой степени пермиссивны в отношении РНК-электропорации и, следовательно, обусловливали массивную дозозависимую экспрессию Cl6 в iDCs и высокоуровневую экспрессию CARs в Т- клетках (Фигура 2А/В). Все CAR Т-клетки показывали максимальную IFNγ-секрецию при 0.02 мкг электропорированной Cl6 РНК, вплоть до 30.000 пг/мл IFNγ для классического scCAR и 20.000 пг/мл для комбинированных CARs, слитых либо с TCR Сα/β, либо с полноразмерным TCR gp100. По оценкам, модифицированные одновалентным CAR Т-клетки оказались наиболее "слабыми" эффекторными клетками. Весьма любопытен тот факт, что при высоком уровне Cl6-электропорации комбинированные CARs показали реактивность, составляющую приблизительно лишь 50% реактивности классического scCAR Cl6, которая повышалась до 70% при оптимальной дозе 0.02 мкг электропорированного Cl6. Важно то, что при самой низкой дозе Cl6, проверенной в данном примере, 0.002 мкг РНК, комбинированные CARs были так же эффективны, как и классический scCAR Cl6, или, в случае с комбинированным с CAR, слитым с TCR Сα/β, даже более эффективны. При этом очень низком уровне присутствующего в массе Cl6 комбинированные CARs все еще были способны секретировать высокие количества IFNγ в пределах 10.000 пг/мл.
На Фигуре 3С приводятся количества секретированного IFNγ для iDCs и Т-клеток одного и того же донора, которые были слабо пермиссивными в отношении РНК-электропорации и, следовательно, привели лишь к частичной дозозависимой экспрессии Cl6 в iDCs, а также к пониженной экспрессии CARs в Т-клетках (Фигура 3А/В). Это может создавать ситуацию, когда на iDCs находится даже меньше Cl6, чем при низкоуровневой экспрессии Cl6 в высокой степени пермиссивных титруемых антигеном iDCs (Фигура 3А/В в сравнении с 2А/В). В данном случае для всех титруемых очень малых количеств Cl6 комбинированные CARs оказались более эффективными в отношении секреции IFNγ, чем классический scCAR. Эта тенденция становится даже более явной для уменьшенных количеств Cl6 на поверхности клеток iDCs. Количество секретированного IFNγ было низким (<1000 пг/мл), но во-первых, его можно повысить за счет лучшей экспрессии CAR, и во-вторых, это может дать хорошие варианты выхода для пациентов клиник, имеющих опухоль (на ранней стадии) с очень низким уровнем экспрессии антигена, или для блокирования минимальной резидуальной болезни.
На Фигуре 4А сравнивается эффективность IFNγ-секреции для (двухвалентных) некомбинированных и комбинированных TCR-CARs в отношении CAR-специфического антигена Cl6. Все комбинированные CARs были более эффективны при распознавании антигена в большом диапазоне титруемого антигена. Для повышенных доз антигена эффекторные функции двухвалентных CARs были почти равными, за исключением ‘молчащего’ некомбинированного TCR-CAR. Известно, что точечная мутация S109Q в CDR3α немного нарушает связывание gp100(280-288)-антигена (Knies et al., Oncotarget 2016). Межцепное связывание V-доменов и связывание самого антигена ‘молчащим’ комбинированным TCR-CAR, по-видимому, возмещает потерю функции, вызванную этой мутацией. Важно отметить, что при низком уровне экспрессии Cl6 (0.02 мкг) комбинированные TCR-CARs, либо функционально молчащие, либо нет, превосходили некомбинированные TCR-CARs в отношении секреции цитокинов.
На Фигуре 4В иллюстрируется распознавание остаточных количеств gp100(280-288) антигена в титруемой пептидом секреции IFNγ. В данном случае TCRαβ gp100 служил в качестве положительного контроля, тогда как TCRαβ Cl6 служил в качестве контроля специфичности для оценки фоновой секреции цитокина. Некомбинированный TCR-CAR по-прежнему способен распознавать А2-1-эволюционно стабильный антиген при высокой пептидной нагрузке. Введение молчащей мутации S109Q отменяло распознавание совсем. Важно отметить, представлялось, что межцепное комбинированное расположение V-доменов, соединенных в виде тандемных повторов (VH-VH-, VL-VL-), полностью предупреждает распознавание родственного антигена молекулой TCR gp100. Следовательно, введение молчащей мутации может служить в качестве защитной меры для гарантии функциональной иммунологической толерантности молекулы TCR в этих CARs и для того, чтобы сосредоточиться на использовании ее скелета в качестве каркаса, стабилизирующего спаривание доменов и в качестве полноразмерной адаптерной молекулы для передачи сигнала Т-клетке в физиологических условиях.
Пример 3: Анализ титруемой антигеном пролиферации в сочетании с биомаркерным фенотипированием
В первый день эксперимента свежие PBMCs выделяли из лейкоцитарной пленки здорового донора. Из of PBMCs выделяли клетки CD14+, используя метод магнитной сортировки клеток MACS, и остаточные PBMCs вымораживали. Клетки CD14+ дифференцировали в iDCs посредством введения IL-4 & GM-CSF (1000 ед/мл) в 1, 3, 6 день. На 7 день iDCs электропорировали с нерелевантной и Cl6 IVT-РНК в зависимости от дозы в интервале 2-0.002 мкг РНК. Замороженные PBMCs подвергали оттаиванию в тот же день и проводили сортировку клеток методом MACS, отбирая CD8+ клетки. Затем без какой- либо предварительной активации (OKT3), наивные Т-клетки, приблизительно 7×106 клеток, электропорировали с использованием классических, одновалентных и интеркомбинированных TCR-CARs, указанных на Фигуре 1.
Для обеспечения контроля качества модифицированные с помощью CAR Т-клетки анализировали методом проточной цитометрии с окрашиванием на 8 день. Затем Т-клетки метили внутриклеточно флуоресцентным маркером пролиферации CFSE (0.8 мкМ) или CPD-450 (10 мкМ). Далее электропорированные Т-клетки и iDCs совместно культивировали в 96-луночном планшете в течение 5 дней при соотношении Е:Т 10:1 (или 3:1 для биомаркерного фенотипирования) в двойном повторе. Согласно обычной практике, 2.5×104 iDCs совместно культивировали в 2.5×105 CAR-электропорированных Т-клеток в объеме 200 мкл среды для Т-клеток в 96-луночном планшете. На 5 день культивированные клетки окрашивали в 96- луночных планшетах с помощью CD4 или CD8 антител, меченных посредством АРС-Су7. Пролиферацию Т- клеток детектировали методом проточной цитометрии по сдвигу влево сигнала флуорофора вследствие разведения в пролиферирующих дочерних клетках. Частоту встречаемости непролиферирующей родительской популяции G0 и дочерних популяций G1-G7 анализировали, используя программу для пролиферации в пакете прикладных программ для проточной цитометрии FlowJo v7.6.5. Частоту встречаемости для всех дочерних Т-клеток рассчитывали, исходя из суммы всех пролиферирующих популяций G1-G7. Фоновую пролиферацию Т-клеток анализировали для клеток, культивированных с iDCs, электропорированными с нерелевантными полноразмерными gp100 или Т-клетками, засеянными без APCs. Пролиферирующие клетки окрашивали на CD8, чтобы однозначно идентифицировать их как Т-клетки. Или же пролиферирующие Т- клетки окрашивали биомаркерами, такими как CD27, CD28, PD-1, CD95, CD45RA и CCR7, чтобы количественно определить статус дифференцировки изначально наивных непролиферирующих Т-клеток в G0 и появляющихся дочерних популяций G1-G7, соответственно, после 5 или 6 дней совместного культивирования с APCs. Антитела и соответствующие контроли изотипа титровали с целью оценить оптимальное отношение сигнал-шум. Количественный анализ проводят на цитофлуориметре системы FACS-Canto II-HTS (BD) в 96- луночном планшете.
На Фигуре 5А изображены графики плотности распределения пролиферирующих Т-клеток, модифицированных одновалентным CAR Cl6 в качестве ‘слабого контроля’, комбинированных TCR-CARs, содержащих и не содержащих молчащую мутацию S109Q в TCR CDR3α, и классического scCAR Cl6 в качестве эталонного CAR. Почти никакой неспецифической пролиферации нельзя наблюдать в отношении iDCs, нагруженных нерелевантным антигеном gp100. Пролиферация по отношению к APCs, нагруженным родственным антигеном, привела к шести различным дочерним популяциям, частоту встречаемости которых можно легко различить (перечислена справа от каждого графика). Все CARs, за исключением одновалентного CAR Cl6, показали высокие скорости пролиферации даже при низких плотностях антигенов. Максимальная частота встречаемости почти во всех случаях наблюдалась в G3.
На Фигуре 5В на гистограмме приведены количества родительских Т-клеток G0 и пролиферирующих Т-клеток G1-G7. При высокой антигенной нагрузке высокий столбик для G0 (40%) и более низкий столбик для G1-G7 (60%) по сравнению с другими CARs четко указывает на слабую предрасположенность модифицированных одновалентным CAR Т-клеток к пролиферации. Модифицированные классическим scCAR Т-клетки оказались лучше несколько лучше (90%), чем комбинированные CARs (80%), в соответствии с результатами анализов секреции IFNγ. Таким образом, при снижении количества антигена Т- клетки, модифицированные комбинированным CAR, становились по меньшей мере такими же эффективными, как и модифицированные классическим scCAR (почти 80%). На основании этой тенденции можно сделать предположение, что даже в случае пониженной антигенной плотности комбинированные CARs могут еще более превосходить классический scCAR в том, что касается активности пролиферации.
На Фигуре 6 слева показана частота встречаемости пролиферирующих Т-клеток, ‘запряженных’ либо классическим scCAR (вверху), либо комбинированным Cα/β-CAR Cl6 (центр) либо комбинированным TCR-CAR Cl6 silCDR3α (внизу). Справа показана средняя интенсивность минус неспецифическое связывание (т.е. связывание с изотипом) для корецептора CD8 и костимулирующего рецептора CD27 среди всех родительских и дочерних популяций. Т-клетки окрашивали CD8-специфическим антителом для визуализации умеренной и почти равной повышающей регуляции этого корецепторного маркера для комбинированных и классических CARs. Следовательно, CD8-окрашивание может служить в качестве маркера нормализации для того, чтобы подчеркнуть равную регуляцию этой 'инертной' молекулы среди всех CARs, рассмотренных в настоящем изобретении. Помимо этого, их окрашивали антителом, специфическим к костимулирующей молекуле CD27, в другом канале флуоресценции, для оценки их регуляции в родительских/дочерних популяциях G0-G6 для всех CARs. Хотя оба маркера активировались приблизительно в 2 раза и, помимо этого, средняя экспрессия CD8 была почти одинаковой для всех CARs, средняя экспрессия CD27 была значительно выше для комбинированных CARs, и в частности, для комбинированного TCR-CAR Cl6, чем для классического CAR. CD27, биомаркер длительной устойчивости Т- клеток in vivo, достигал значительно более высокого плато для Т-клеток с затухающей пролиферацией (G2-G4) до того, как уровни экспрессии падали до базового уровня в G6. Исходя из этого, можно выдвинуть гипотезу, что модифицированные комбинаторным CAR Т-клетки могут иметь меньше окончательно дифференцированного и истощенного фенотипа (т.е. даунрегуляция костимулирующих молекул) в популяциях с затухающей пролиферацией и, следовательно, могут дольше продолжать существовать in vivo. Размер (прямое светорассеяние) и грануляция (боковое светорассеяние) совместно культивированных комбинированный CAR-электропорированных Т-клеток были даже ниже, чем для классического CAR (данные не показаны). Следовательно, повышенная активация CD27 вызвана не увеличением клеточной поверхности, а повышенным количеством CD27 на большей поверхности.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИГЕННЫЕ РЕЦЕПТОРЫ И ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2792042C2 |
| ИММУННЫЕ КЛЕТКИ, ДЕФЕКТНЫЕ ПО Suv39h1 | 2018 |
|
RU2784531C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ К CD22 | 2012 |
|
RU2644243C2 |
| ИММУННЫЕ КЛЕТКИ, ЭКСПРЕССИРУЮЩИЕ ОБРАТНЫЙ УНИВЕРСАЛЬНЫЙ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР, ДЛЯ НАЦЕЛИВАНИЯ НА РАЗЛИЧНЫЕ МНОГОЧИСЛЕННЫЕ АНТИГЕНЫ И СПОСОБ ИХ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ ДЛЯ ЛЕЧЕНИЯ РАКА, ИНФЕКЦИЙ И АУТОИММУННЫХ ЗАБОЛЕВАНИЙ | 2019 |
|
RU2824391C2 |
| Модифицированные клетки с химерным антигеновым рецептором для лечения рака, экспрессирующего CLDN6 | 2020 |
|
RU2826058C2 |
| АГЕНТЫ ДЛЯ ЛЕЧЕНИЯ ЭКСПРЕССИРУЮЩИХ КЛАУДИН РАКОВЫХ ЗАБОЛЕВАНИЙ | 2013 |
|
RU2678127C2 |
| АНТИТЕЛА ПРОТИВ КЛАУДИНА 18.2 ДЛЯ ДИАГНОСТИКИ РАКА | 2013 |
|
RU2661772C2 |
| НЕ РЕСТРИКТИРОВАННЫЕ ПО HLA T-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2812917C2 |
| АНТИТЕЛА, ПРИГОДНЫЕ В ДИАГНОСТИКЕ РАКА | 2018 |
|
RU2815883C1 |
| СПОСОБ СОЗДАНИЯ Т-КЛЕТОК, ПРИГОДНЫХ ДЛЯ АЛЛОГЕННОЙ ТРАНСПЛАНТАЦИИ | 2015 |
|
RU2752933C2 |

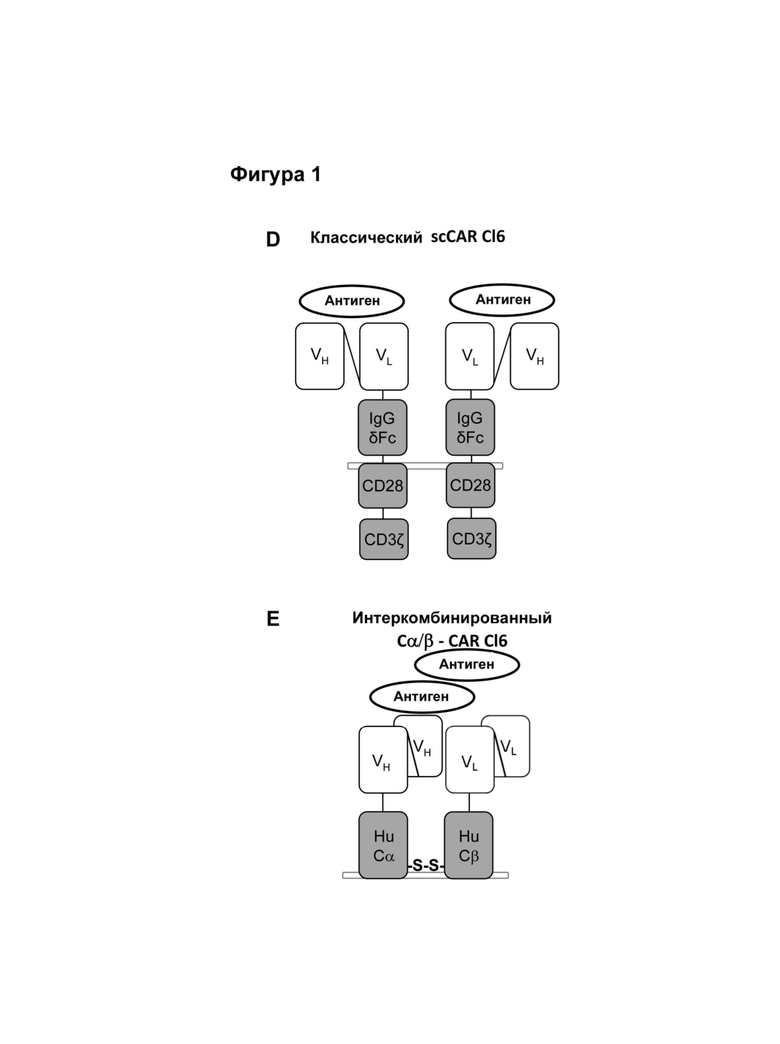

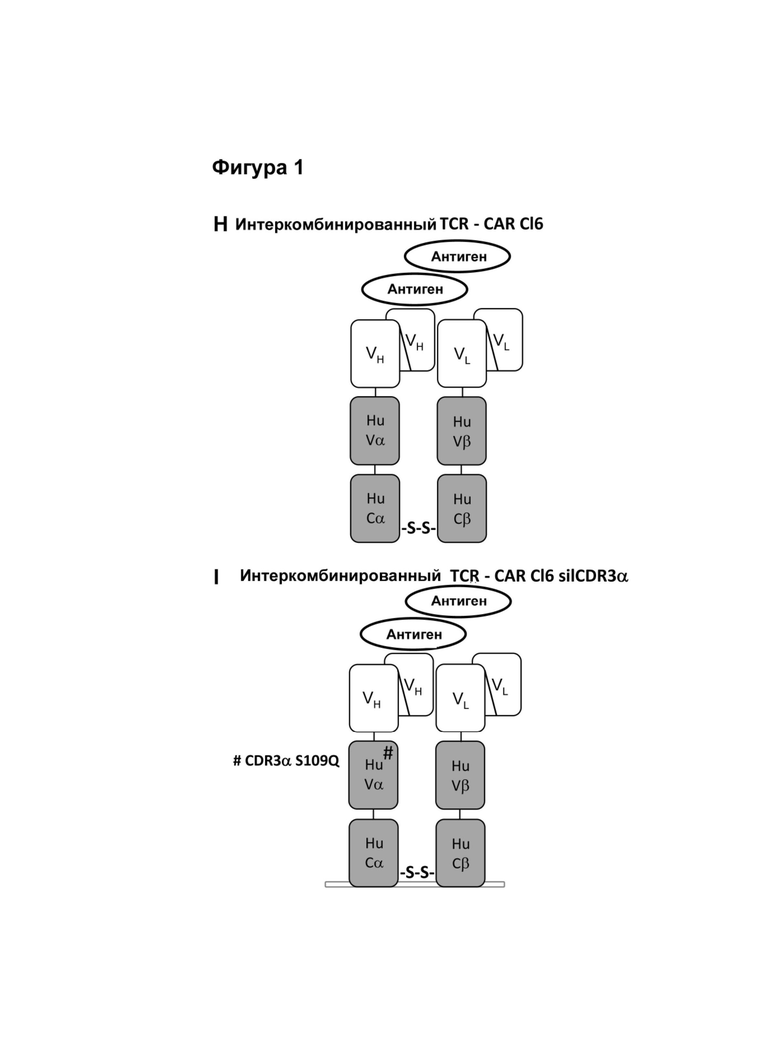
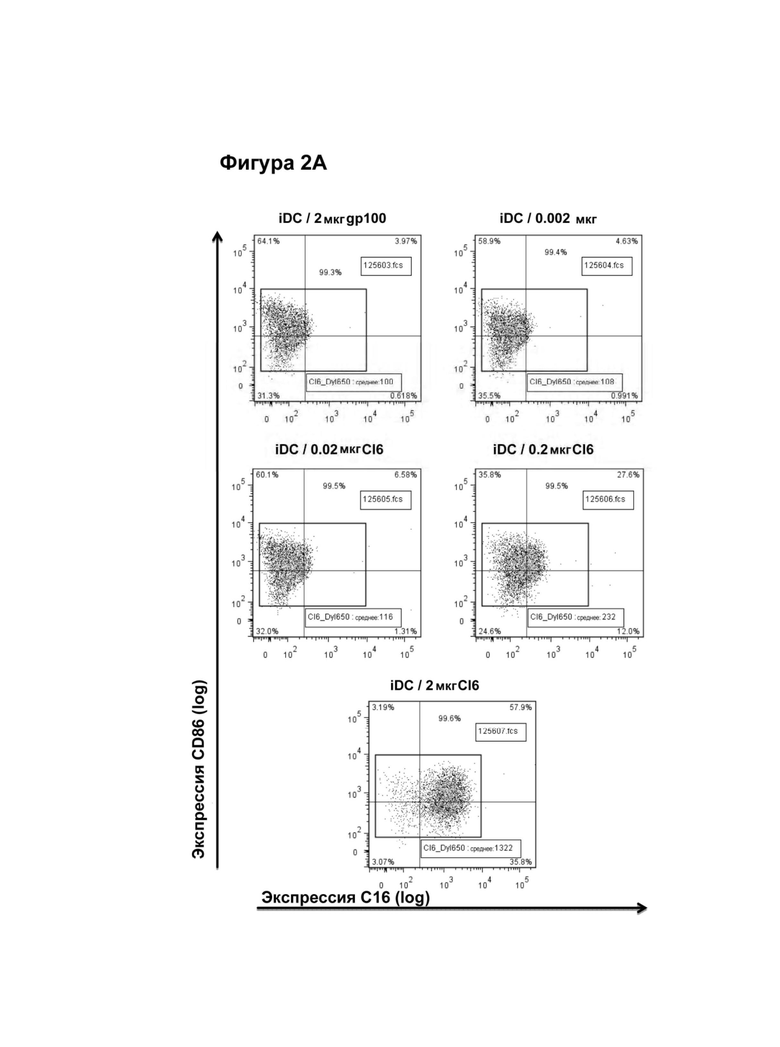

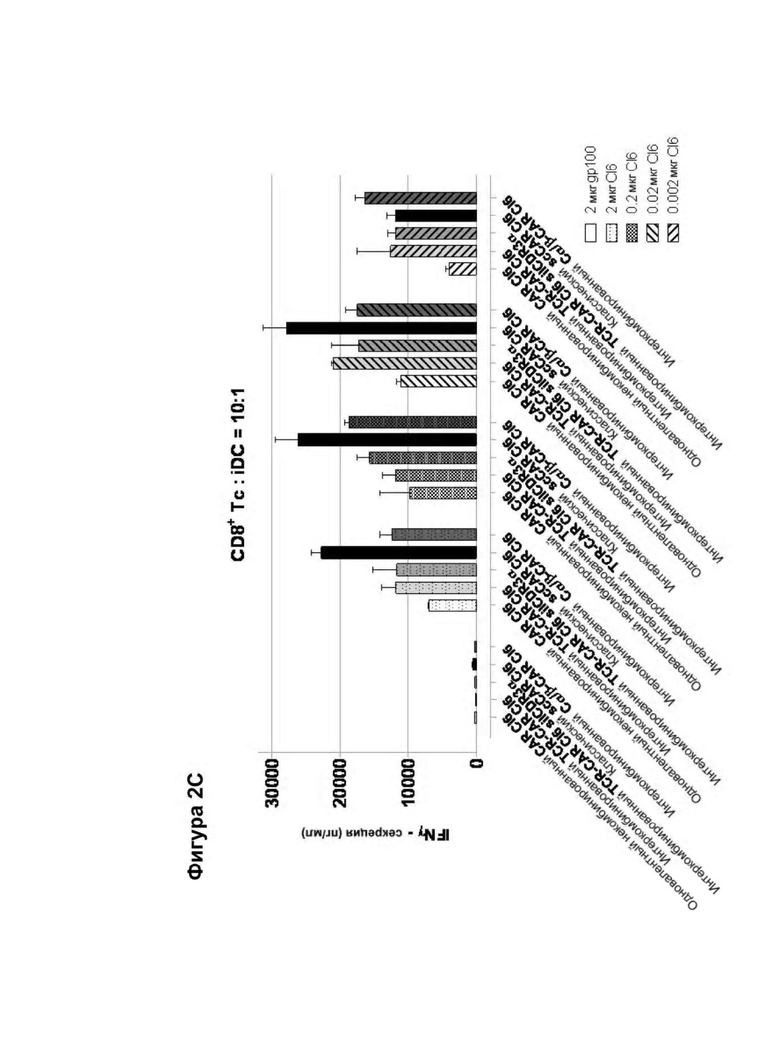
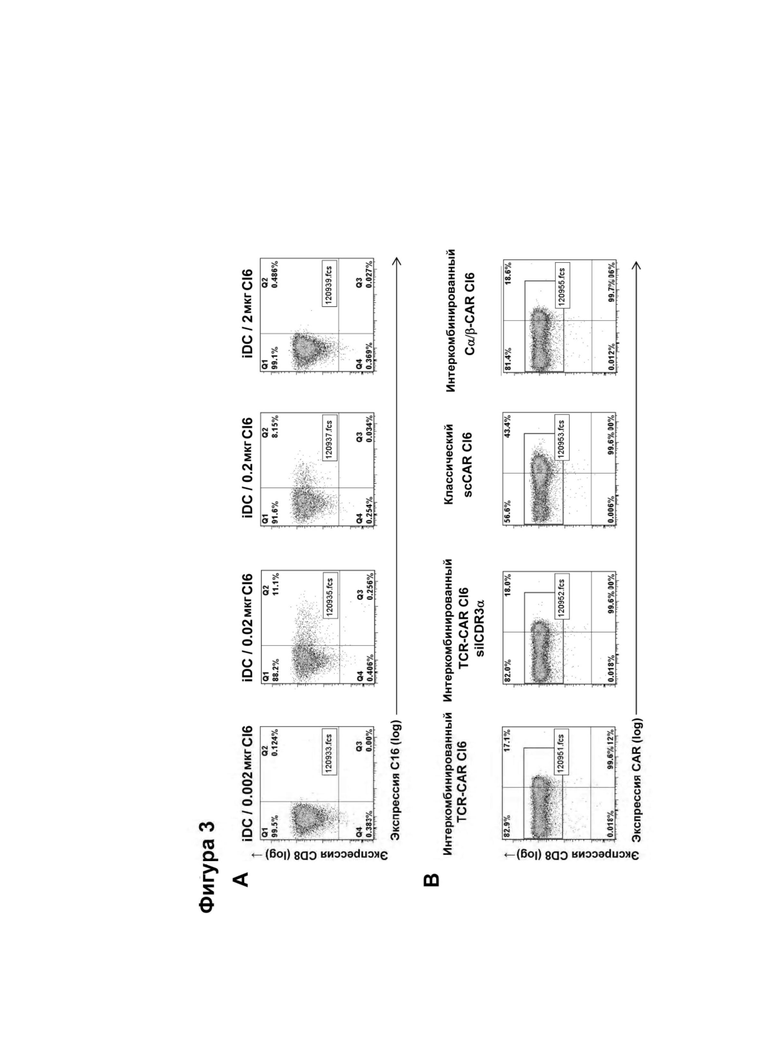
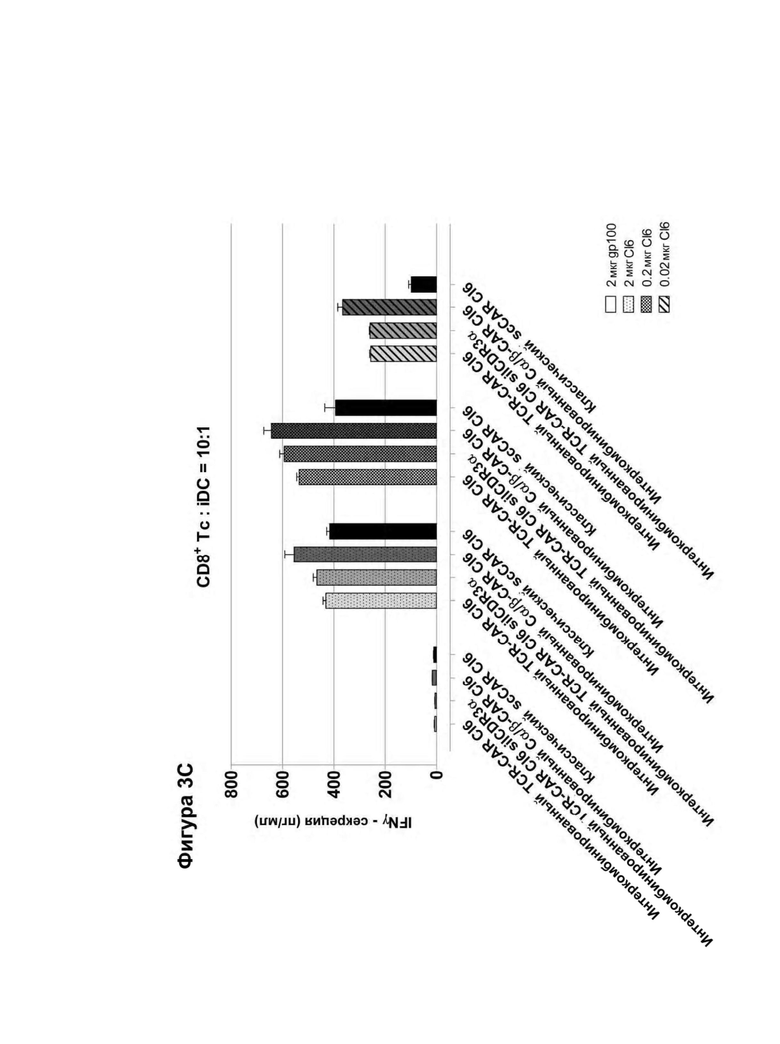
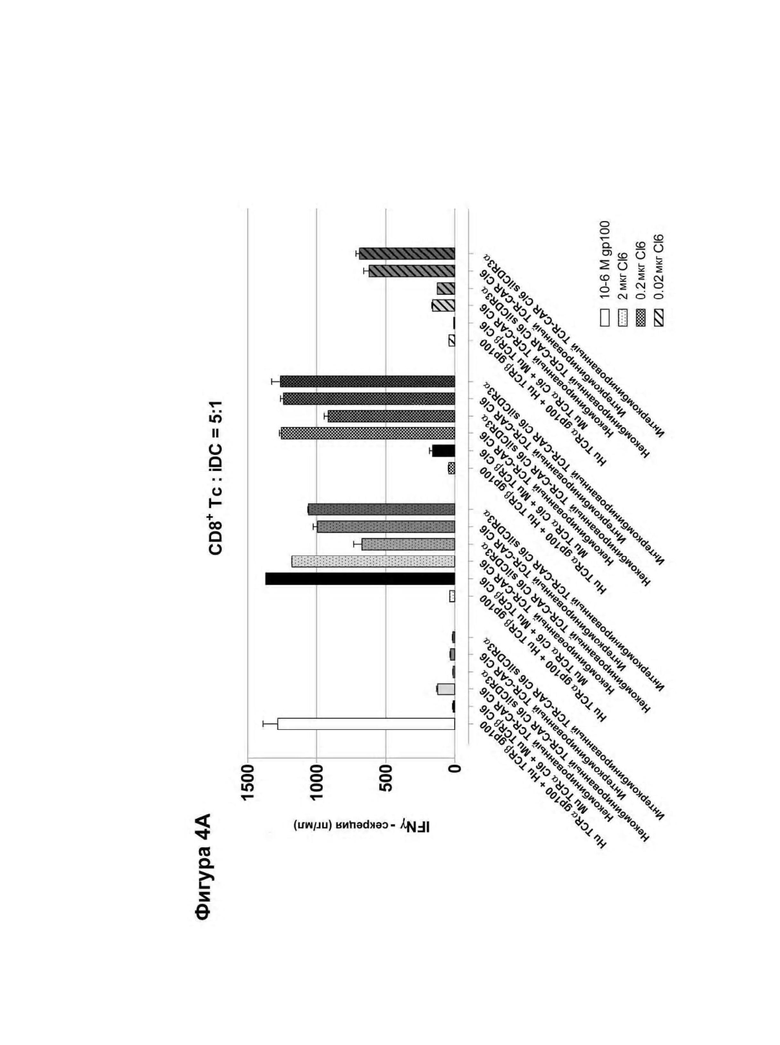



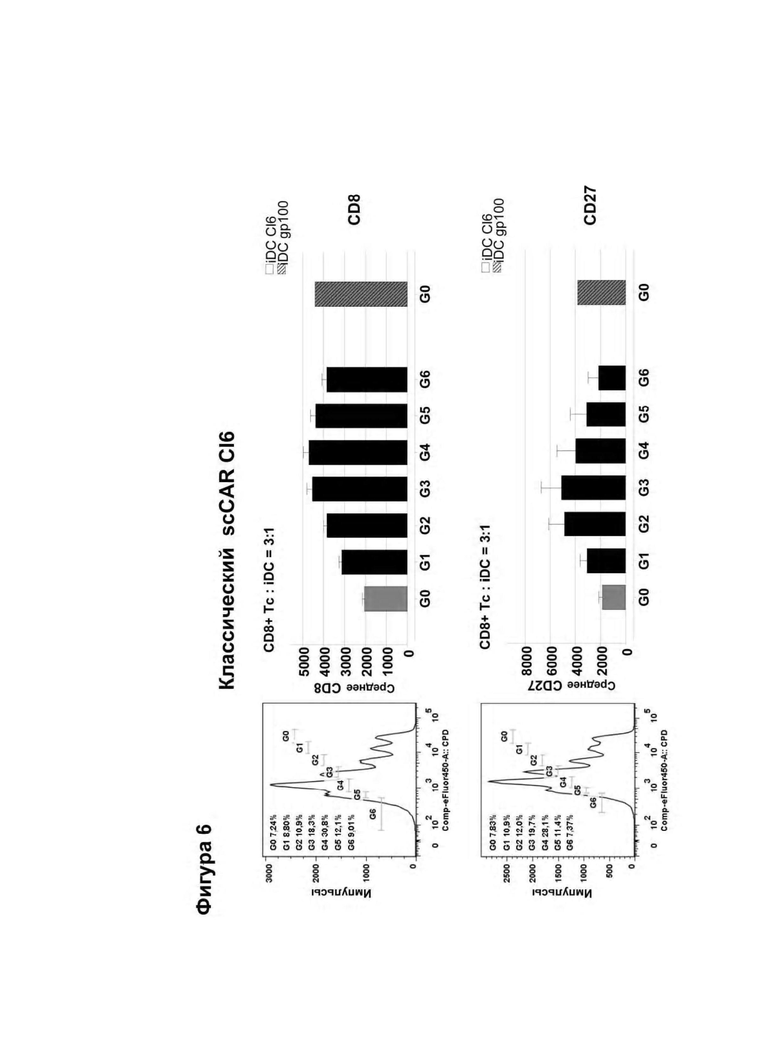

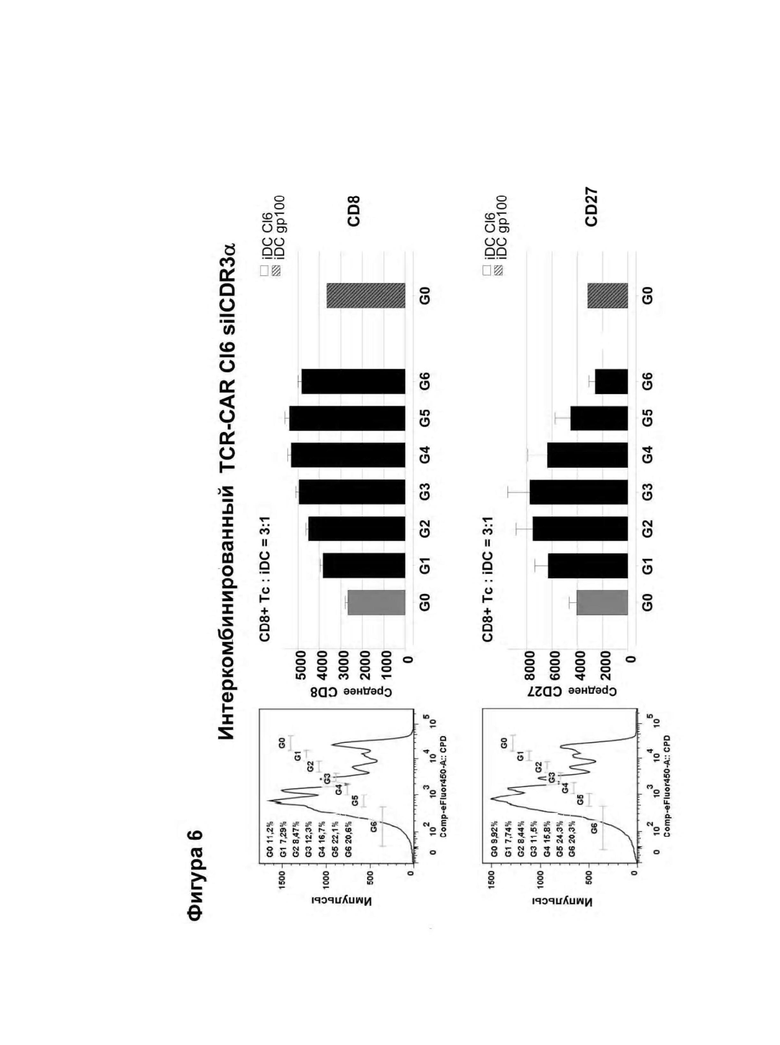
Настоящее изобретение относится к области биотехнологии, в частности в новым химерным рецепторам, и может быть применимо в медицине. Изобретение охватывает лечение заболеваний путем нацеливания Т-клеток на клетки организма, экспрессирующие опухолевый антиген на клеточной поверхности. Изобретение раскрывает различные комбинированные химерные антигенные рецепторы (см. фиг. 1). Т-клетки, модифицированные для экспрессии таких химерных антигенных рецепторов, могут быть использованы при лечении заболеваний, характеризующихся экспрессией одного или более опухолевых антигенов, в частности при терапии рака. 3 н. и 6 з.п. ф-лы, 6 ил., 1 табл., 3 пр.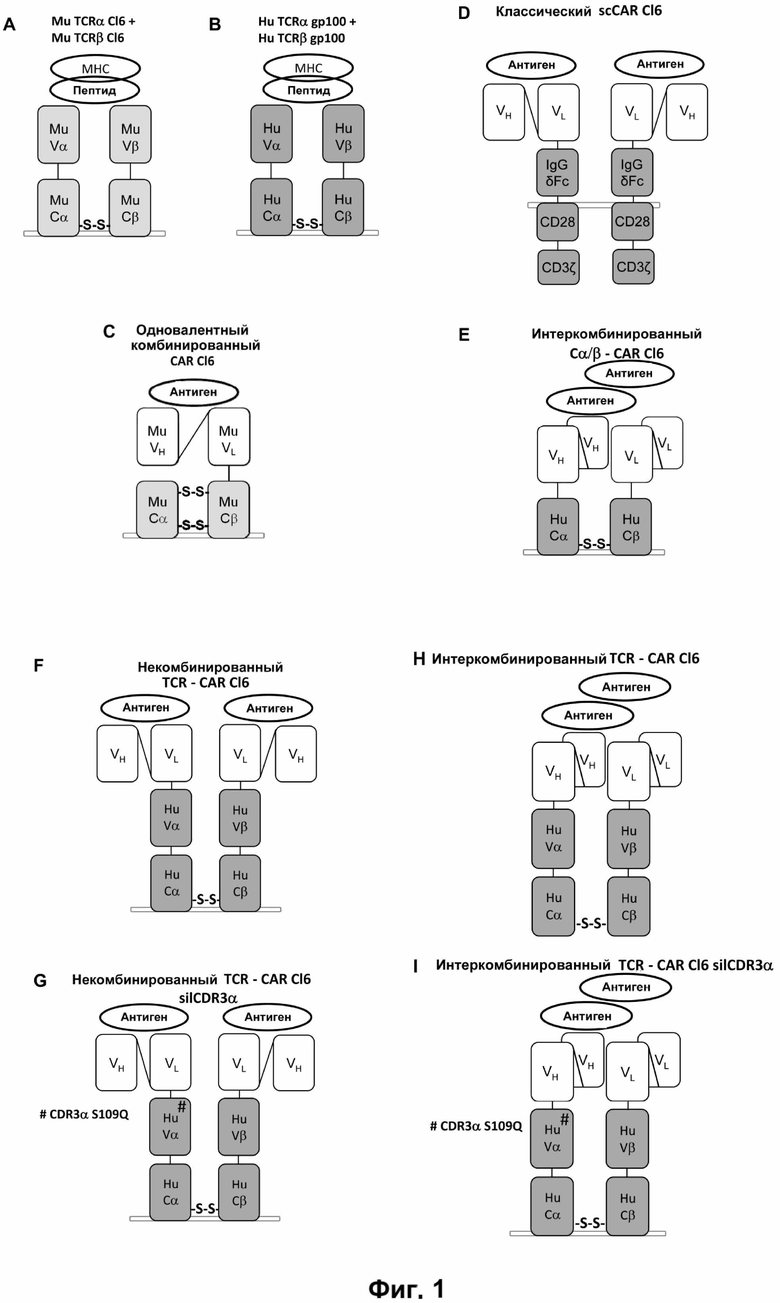
1. Опухолевый антигенный рецептор, обеспечивающий иммунный ответ на раковые клетки, экспрессирующие опухолевый антиген, который включает первую пептидную цепь и вторую пептидную цепь, при этом
первая пептидная цепь содержит первый домен со специфичностью к опухолевому антигену, второй домен со специфичностью к опухолевому антигену, вариабельную область цепи T-клеточного рецептора и домен передачи сигнала от иммунорецептора;
вторая пептидная цепь содержит первый домен со специфичностью к опухолевому антигену, второй домен со специфичностью к опухолевому антигену, вариабельную область цепи T-клеточного рецептора и домен передачи сигнала от иммунорецептора;
при этом первый домен первой пептидной цепи совместно с одним из доменов второй пептидной цепи образует первый опухолевый антигенсвязывающий сайт, и
при этом второй домен первой пептидной цепи совместно с другим доменом второй пептидной цепи образует второй опухолевый антигенсвязывающий сайт, где
(i) первая пептидная цепь содержит вариабельную область альфа-цепи Т-клеточного рецептора и константную область альфа-цепи Т-клеточного рецептора, и вторая пептидная цепь содержит вариабельную область бета-цепи Т-клеточного рецептора и константную область бета-цепи Т-клеточного рецептора, или
(ii) первая пептидная цепь содержит вариабельную область бета-цепи Т-клеточного рецептора и константную область бета-цепи Т-клеточного рецептора, и вторая пептидная цепь содержит вариабельную область альфа-цепи Т-клеточного рецептора и константную область альфа-цепи Т-клеточного рецептора,
где первая пептидная цепь содержит структуру VH(1)-VL(2)-V1-C1 и вторая пептидная цепь содержит структуру VL(1)-VH(2)-V2-C2,
где первая пептидная цепь содержит структуру VH(1)-VH(2)-V1-C1 и вторая пептидная цепь содержит структуру VL(1)-VL(2)-V2-C2,
где первая пептидная цепь содержит структуру VH(1)-VH(2)-V1-C1 и вторая пептидная цепь содержит структуру VL(2)-VL(1)-V2-C2, или
где первая пептидная цепь содержит структуру VH(1)-VL(2)-V1-C1 и вторая пептидная цепь содержит структуру VH(2)-VL(1)-V2-C2;
причем
VH(1) обозначает вариабельную область тяжёлой цепи иммуноглобулина (VH) со специфичностью к первому эпитопу;
VH(2) обозначает вариабельную область тяжёлой цепи иммуноглобулина (VH) со специфичностью ко второму эпитопу;
VL(1) обозначает вариабельную область лёгкой цепи иммуноглобулина (VL) со специфичностью к первому эпитопу;
VL(2) обозначает вариабельную область лёгкой цепи иммуноглобулина (VL) со специфичностью ко второму эпитопу;
V1 и V2 являются вариабельными областями альфа- и бета-цепей Т-клеточного рецептора, соответственно, или её участками,
C1 и C2 являются доменами передачи сигнала иммунорецептора, которые образуют димер друг с другом, причем
(i) домен передачи сигнала от иммунорецептора C1 первой пептидной цепи содержит константную область альфа-цепи T-клеточного рецептора и домен передачи сигнала от иммунорецептора C2 второй пептидной цепи содержит константную область бета-цепи T-клеточного рецептора, или
(ii) домен передачи сигнала от иммунорецептора C1 первой пептидной цепи содержит константную область бета-цепи T-клеточного рецептора и домен передачи сигнала от иммунорецептора C2 второй пептидной цепи содержит константную область альфа-цепи T-клеточного рецептора;
в котором первая и/или вторая пептидные цепи дополнительно содержат линкер между первым и вторым доменами и/или между первым и вторым доменами и вариабельной областью цепи T-клеточного рецептора;
и в котором первый и второй антигенсвязывающие сайты связываются с различными эпитопами на одном и том же опухолевом антигене.
2. Рецептор по п. 1, в котором домен передачи сигнала иммунорецептора содержит константную или консервативную область цепи Т-клеточного рецептора или константную или консервативную область Fc-цепи рецептора иммунной клетки или участок этой константной или консервативной области.
3. Рецептор по п. 1 или 2, в котором линкер представляет собой произвольную аминокислотную последовательность или линкер содержит 3 повтора 5-мерной аминокислотной последовательности (Gly4Ser).
4. Рецептор по любому из пп. 1-3, в котором домен передачи сигнала иммунорецептора C1 и C2 имеет человеческое происхождение.
5. Рецептор по любому из пп. 1-4, в котором аминокислотные последовательности первой и второй пептидных цепей имеют человеческое происхождение и в котором в домены, образующие антигенсвязывающие сайты, введены мышиные последовательности путём замены одной или более аминокислот в человеческой последовательности аминокислотой, находящейся в соответствующем положении в мышиной последовательности.
6. Способ получения клетки, экспрессирующей опухолевый антигенный рецептор по п. 1, который содержит первую пептидную цепь и вторую пептидную цепь, при этом способ включает:
(a) предоставление клетки;
(b) предоставление первой генетической конструкции, кодирующей первую пептидную цепь, содержащую по меньшей мере первый домен, второй домен, вариабельную область цепи Т-клеточного рецептора и домен передачи сигнала иммунорецептора;
(c) предоставление второй генетической конструкции, кодирующей вторую пептидную цепь, содержащую по меньшей мере первый домен, второй домен, вариабельную область цепи Т-клеточного рецептора и домен передачи сигнала иммунорецептора;
(d) введение первой и второй генетических конструкций в клетку; и
(e) обеспечение возможности экспрессии конструкций в клетке;
при этом первый домен из первой пептидной цепи способен образовать вместе с одним из доменов из второй пептидной цепи первый опухолевый антигенсвязывающий сайт, и
где второй домен из первой пептидной цепи способен образовать вместе с другим доменом из второй пептидной цепи второй опухолевый антигенсвязывающий сайт:
при этом
(i) первая пептидная цепь содержит вариабельную область альфа-цепи Т-клеточного рецептора и константную область альфа-цепи Т-клеточного рецептора, и вторая пептидная цепь содержит вариабельную область бета-цепи Т-клеточного рецептора и константную область бета-цепи Т-клеточного рецептора, или
(ii) первая пептидная цепь содержит вариабельную область бета-цепи Т-клеточного рецептора и константную область бета-цепи Т-клеточного рецептора, и вторая пептидная цепь содержит вариабельную область альфа-цепи Т-клеточного рецептора и константную область альфа-цепи Т-клеточного рецептора.
7. Способ по п. 6, где первая пептидная цепь и вторая пептидная цепь предоставляются в единой генетической конструкции.
8. Способ по п. 6 или 7, где клетка представляет собой человеческую клетку, предпочтительно Т-клетку.
9. Применение опухолевого антигенного рецептора по любому из пп. 1-5 для лечения заболевания, где заболевание характеризуется экспрессией по меньшей мере одного опухолевого антигена, который связан с опухолевым антигенным рецептором, и где заболевание предпочтительно представляет собой рак.
| СПОСОБ СТАБИЛИЗАЦИИ МОНОРЕЛЬСОВОЙ РАКЕТНОЙ ТЕЛЕЖКИ (ВАРИАНТЫ) И УСТРОЙСТВО ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ (ВАРИАНТЫ) | 2012 |
|
RU2502934C1 |
| GROSS G | |||
| et al.: "Endowing T-cells with antibody specificity using chimeric T-cell receptors", THE FASEB JOURNAL, 1992, v.6 (15): 3370-3378 | |||
| Устройство для демонстрирования кинолент при помощи солнечного света | 1930 |
|
SU23087A1 |
| BARRETT D.M | |||
| et al.: "Chimeric antigen receptor- and TCR-modified T-cells enter main street and wall street", THE JOURNAL OF IMMUNOLOGY, 2015, v.195 (3): | |||

