Область техники, к которой относится изобретение
Настоящее изобретение относится к способу повышения роста и урожайности растений в условиях ограниченного уровня азота.
Уровень техники
Применение химических удобрений для снабжения азотом, как наблюдается в современном интенсивном сельском хозяйстве, приводит к существенным повышениям в урожайности, но высвобождает азот в окружающую среду в форме избыточных нитратов, что вызывает появление таких проблем для окружающей среды, как загрязнение рек и озер эвтрофикацией, опасность для здоровья из-за аккумуляции нитратов в сельскохозяйственных культурах и генерацию газов теплиц из-за остаточного азота в почве. Большое количество электричества, требуемого для производства азотистых удобрений, также вносит вклад в проблемы окружающей среды для сельского хозяйства. Другими словами, для поддержания урожайности требуются сельскохозяйственные способы, которые применяют меньше химического удобрения, особенно азотистого удобрения.
Однако за несколькими исключениями, такими как Leguminosae, растения не могут фиксировать азот из воздуха и полностью зависят в отношении азота от нитрата или аммиака, подаваемого извне. Следовательно, азот является фактором, наиболее ограничивающим рост и развитие растений, и современное сельское хозяйство встречается, очевидно, с противоречивой задачей поддержания и повышения современных урожаев при снижении вышеуказанной проблемы для окружающей среды при усилии поддержания урожайности. Усилия в этом направлении и особенно попытки вывести и культивировать растения с удовлетворительным ростом и урожайностью в условиях пониженной подачи азота до сих пор были недостаточными. Например, благоприятное для окружающей среды и способное поддерживать урожайность органическое сельское хозяйство было предложено в качестве альтернативы современному интенсивному сельскому хозяйству, но хотя внесение азотистого удобрения в таком благоприятном для окружающей среды сельском хозяйстве регулируется, здесь имеется много проблем в отношении урожайности.
С точки зрения улучшения сортов растений не было удачных примеров выведения растений для разрешения этих проблем обеспечением адекватного роста и/или урожайности в условиях ограниченного уровня азота, таких как условия, в которых внесение азотистого удобрения уменьшают. Следовательно, существует потребность в выведении сортов растений, имеющих сохраняемую или повышенную урожайность, посредством модификации или повышения эффективности утилизации азота растениями при меньшем внесении азотистого удобрения и в разработке технологий для выведения таких сортов растений.
Известно, что первой стадией ассимиляции неорганического азота в органическую форму в растениях в основном является включение аммиака в глутамат с образованием глутамина. Включение является реакцией, катализируемой глутаминсинтетазой (GS) и глутаматсинтазой (GOGAT), при этом две реакции объединяются с образованием одной молекулы глутамата ассимиляцией одной молекулы аммиака в одну молекулу 2-OG. Считается, что этот цикл CS/GOGAT является основным путем ассимиляции азота в растениях (Miflin and Lea: 1976, Phytochemistry 15: 873-885).
С другой стороны, в результате исследований с применением Cyanobacteria и Е. coli сообщается, что в микроорганизмах количественный баланс 2-OG и глутамина регулирует экспрессию генов ферментов, связанных с включением и ассимиляцией нитрата (Forchammer and Tandeau-Marsac, 1995, J. Bacteriol. 177, 2033-2040; Jiang et al, 1998, Biochemistry 37, 12795-12801). Однако из Arabidopsis thaliana (арабидопсис) выделен ген, имеющий строгую гомологию с белком PII, который, как предполагается, ассоциирован с такой регуляцией, и, когда такой ген вводили и сверхэкспрессировали в табак, не обнаружили повышения содержания нитрата, что позволило предположить, что аккумуляция азота в высших растениях происходит отличным образом по сравнению с микроорганизмами (Hsieh et al, 1998, Proc. Natl. Acad. Sci. USA 95: 13965-13970). С другой стороны, указывалось, что, когда исследовали экспрессию нитратредуктазы (далее обозначаемой аббревиатурой NR) с применением табачных растений, имеющих ген ферредоксинзависимой глутаматсинтазы, введенный в антисмысловом направлении так, чтобы снизить экспрессию глутаматсинтазы, уровень транскрипции NR повышался, когда поставляли нитрат, сахарозу и 2-OG, тогда как, когда поставляли глутамин, уровень транскрипции NR снижался (Ferrario-Mery et al, 2001 Planta 213: 265-271). В соответствии с данной ссылкой, количества 2-OG и глутамата также ассоциируется с регуляцией NR в высших растениях. Однако эти полученные данные относятся только к NR, и ничего не было указано о действии 2-OG на рост и развитие растений в условиях ограниченного содержания азота. Более того, 2-OG является относительно нестабильным соединением и рассматривается как непригодный, например, для целей разбрызгивания.
Разработаны также трансгенные растения, в которые был введен ген GDH, и сообщается, что, когда полученный из Е. coli NADP-зависимый ген GDH (gdhA) вводили в табак и кукурузу с той целью, чтобы все из факторов: придание устойчивости к гербициду фосфинотрицину, сухая масса, общее содержание аминокислот и содержание водорастворимых углеводов значительно повысились (Lightfoot et al. CA2180786, 1996). Аналогично этому, Thian et al (CN00109779.2), ввели полученный из Neurospora, NADP-зависимый ген GDH в табак и сообщили, что рост был лучше, чем рост табака, в который такой ген не был введен. Указано также, что аминокислотное содержание в плодах повышалось, когда полученный из Aspergillus nidulans NADP-зависимый ген GDH вводили в томатные растения (Kisaka et al, JP11-376710), и что масса клубней и число клубней повышались, когда тот же самый ген вводили в картофельные растения (Kisaka et al, JP2000-404322). Однако функцию введенного гена не объясняли ни в одном из этих сообщений, и не известно, как функционирует введенный ген или какие действия вызывают эти характеристики. Кроме того, ничего не упоминается о усилении роста и/или повышения урожайности растений в условиях ограниченного уровня азота.
Не имеется также сообщений, относящихся к введению гена аспартатаминотрансферазы в растения.
Сущность изобретения
Целью настоящего изобретения является предоставление способа выведения растений, имеющих повышенное включение в них азота и повышенную метаболическую активность растений и имеющих усиленный рост и/или повышенную урожайность в условиях культивации с ограниченным уровнем азота, или другими словами, с уровнем азота, ограниченным до уровня ниже нормальных условий культивации, а также способ культивации таких растений в условиях ограниченного уровня азота.
Авторы изобретения предположили наличие сигнальной молекулы, которая позволяет клеткам растения ощущать условия азотного потребления, и рассматривали 2-оксоглутарат (2-OG) в качестве сигнальной молекулы. То есть 2-OG является сигнальной молекулой, которая всестороннее регулирует включение и метаболизм азота в клетках растений, и было предположено, что посредством искусственного повышения или снижения содержания этого вещества может быть, возможно, поддержание в клетках растения включения и метаболизма азота в повышенном состоянии. Кроме того, поскольку считается, что растение в такой повышенном состоянии способно активно включать в себя азот из внешней стороны, полагают, что азот может быть пополнен в условиях ограничения уровня азота, в результате чего происходит усиление роста и повышение урожайности. Кроме того, авторы изобретения обнаружили, что содержание 2-OG клеток растений могло быть повышено индуцированием сверхэкспрессии активности глутаматдегидрогеназы (далее обозначаемой аббревиатурой GDH) или активности аспартатаминотрансферазы (далее обозначаемой аббревиатурой ASPC) и что растения с повышенным содержанием 2-OG имеют усиленный рост и/или повышенную урожайность в условиях ограниченного содержания азота, что привело к созданию настоящих изобретений.
Следовательно, настоящим изобретением является способ получения растения, проявляющего усиленный рост и/или повышенную урожайность в условиях культивации, когда содержание азота ограничивают до уровня ниже нормальных условий культивации повышением содержания 2-OG в растениях.
Кроме того, настоящим изобретением является растение или его семена, проявляющие усиленный рост и/или повышенную урожайность в условиях культивации, когда содержание азота ограничивают до уровней ниже нормальных условий культивации, введением гена GDH или гена ASPC и экспрессией гена в растительном организме, в результате чего происходит повышение содержание 2-OG. Кроме того, настоящим изобретением является способ получения растения, проявляющего усиленный рост и/или повышенную урожайность в условиях культивации, когда содержание азота ограничивают до уровней ниже нормальных условий культивации, ведением гена GDH или гена ASPC и экспрессией гена в растительном организме, в результате чего происходит повышение содержание 2-OG.
В частности, настоящим изобретением является способ получения растения, проявляющего усиленный рост и/или повышенную урожайность в условиях культивации, когда содержание азота ограничивают до уровня ниже уровня нормальных условий культивации, в котором рост и/или урожайность эквивалентные или выше, чем рост и/или урожайность, полученные в условиях культивации с обычно применяемым количеством (определенным для каждого растения) азотистого удобрения, полученные в условиях культивации с количеством азота, находящимся в интервале ниже, чем нижний предел, и 1/10-кратным нижним пределом стандартного количества внесения азотистого удобрения.
Настоящим изобретением является способ культивации растения, полученного указанным выше способом настоящего изобретения, в котором культивацию проводят в условиях ограниченного содержания азота. В частности, настоящим изобретением является способ культивации растения, выведенного указанным выше способом настоящего изобретения в условиях внесения азотистого удобрения в диапазоне между количеством ниже нижнего предела и 1/10-кратным нижним пределом стандартного количества внесения (определено для каждого растения).
Кроме того, настоящим изобретением является способ усиления роста и/или повышения урожайности растений в условиях культивации с содержанием азота, ограниченным до уровня ниже нормальных условий культивации, содержащий проведение некорневой подкормки растений пролином.
Краткое описание чертежей
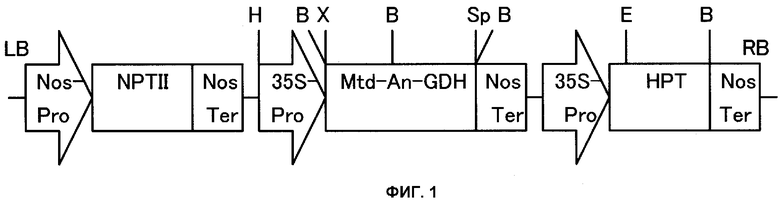
Фиг.1 представляет собой схематическую диаграмму сконструированного плазмидного вектора, в котором Nos-Pro представляет собой промотор нопалинсинтазы; NPTII представляет собой неомицинфосфотрансферазу; Nos-Ter представляет собой терминатор нопалинсинтазы; 35S-Pro представляет собой промотор 35S вируса мозаичной болезни цветной капусты; MTD-AN-GDH представляет собой полученный из Aspergillus nidulans ген глутаматдегидрогеназы (GDH) с присоединенным митохондриальным транзитным пептидом; НРТ представляет собой гигромицинфосфотрансферазу; Н представляет собой Hindill, B представляет собой BamHI, Х представляет собой Xbal, Sp представляет собой Spel, E представляет собой EcoRI, LB представляет собой левый пограничный край и RB представляет собой правый пограничный край.
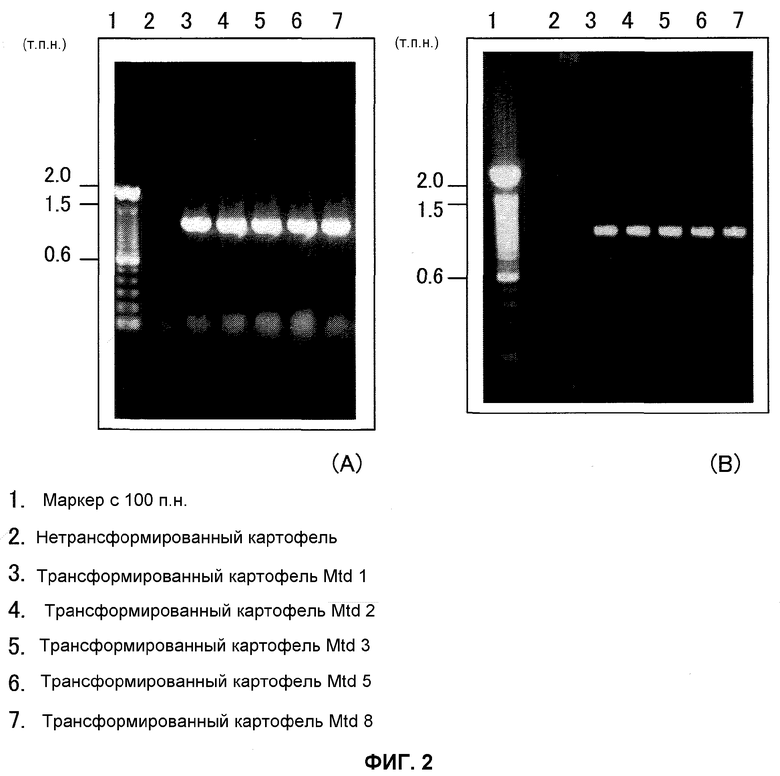
На фиг.2 показаны результаты анализа ПЦР трансформированного картофеля, причем А отражает применение An-GDH-ген-специфических праймеров и В отражает применение NPTII-ген-специфичных праймеров. Дорожка 1 представляет собой маркер со 100 п.н., дорожка 2 представляет собой нетрансформированный картофель, дорожка 3 представляет собой трансформированный картофель Mtd1, дорожка 4 представляет собой трансформированный картофель Mtd2, дорожка 5 представляет собой трансформированный картофель Mtd3, дорожка 6 представляет собой трансформированный картофель Mtd5, и дорожка 7 представляет собой трансформированный картофель Mtd8.
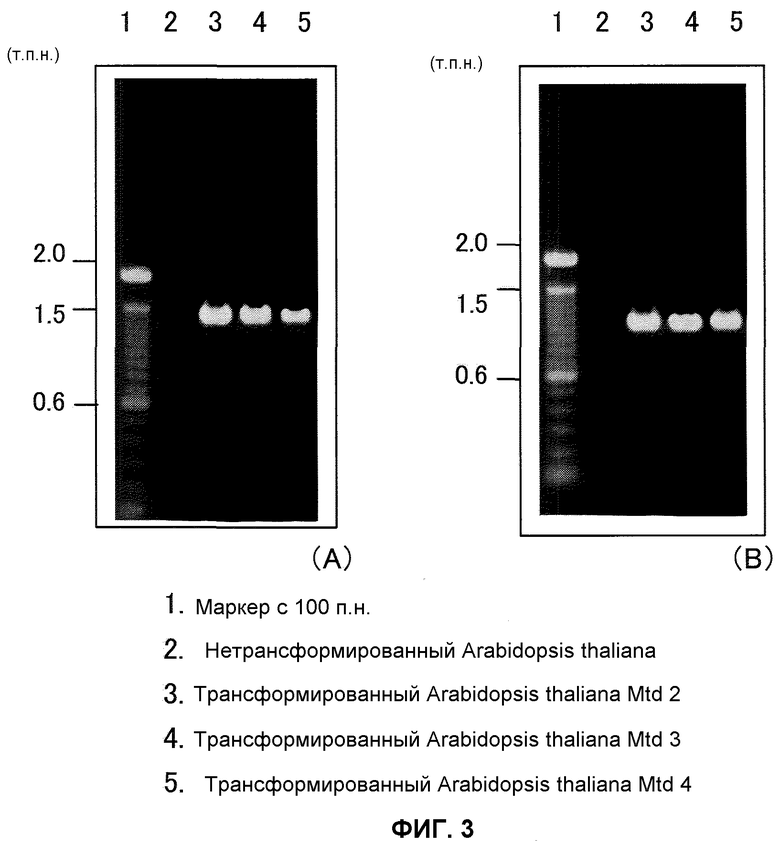
На фиг.3 показаны результаты анализа ПЦР трансформированного Arabidopsis thaliana, причем А отражает применение An-GDH-ген-специфичных праймеров и В отражает применение NPTII-ген-специфичных праймеров. Дорожка 1 представляет собой маркер со 100 п.н., дорожка 2 представляет собой нетрансформированный Arabidopsis thaliana, дорожка 3 представляет собой трансформированный Arabidopsis thaliana Mtd 2, дорожка 4 представляет собой трансформированный Arabidopsis thaliana Mtd 3, и дорожка 5 представляет собой трансформированный Arabidopsis thaliana Mtd 4.
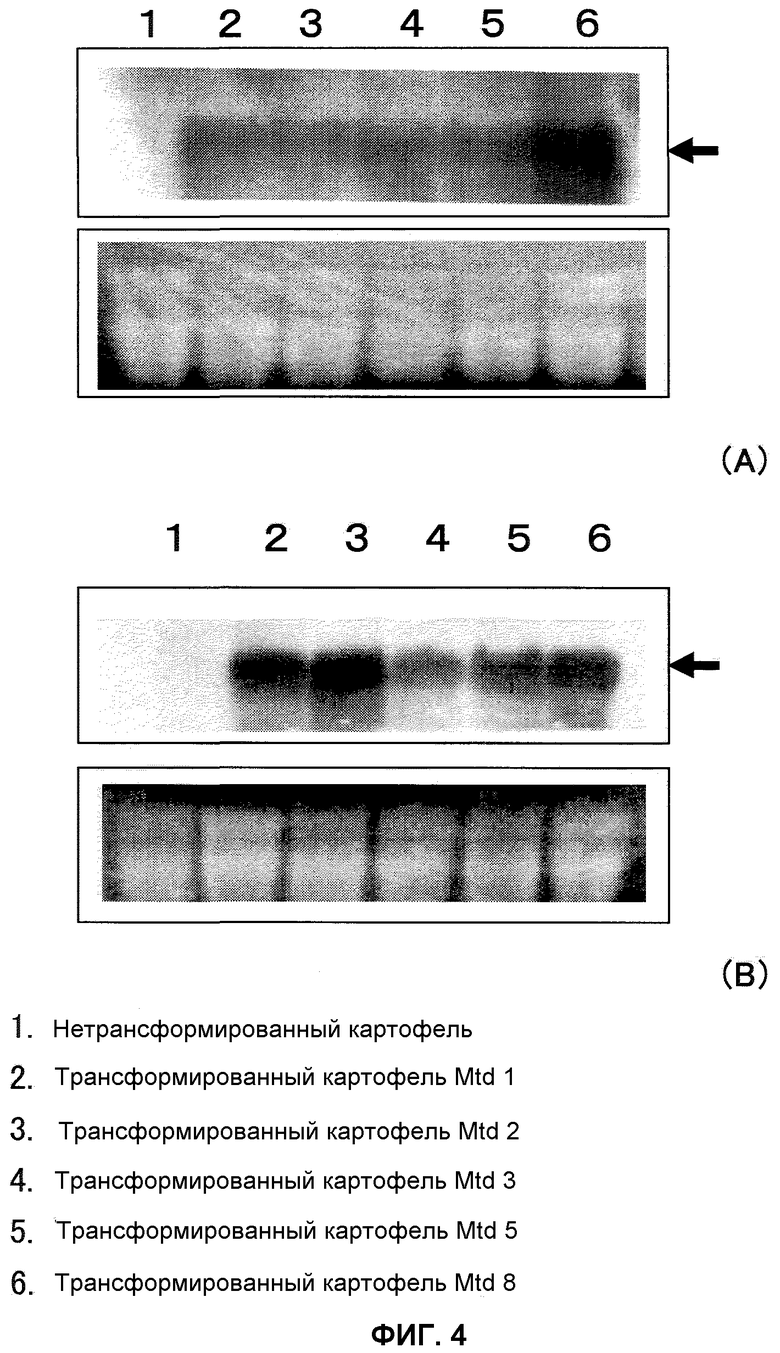
На фиг.4 показаны результаты нозерн-анализа трансформированных картофельных растений, причем А обозначает РНК, экстрагированную из ткани листьев, и В обозначает РНК, экстрагированную из клубней. Рисунки получали окрашиванием бромидом этидия 10 мкл тотальной РНК, которая была подвергнута электрофорезу (нижние рисунки как в А, так и в В). В качестве зонда применяли полноразмерный An-GDH-ген. Дорожка 1 представляет собой нетрансформированный картофель, дорожка 2 представляет собой трансформированный картофель Mtd1, дорожка 3 представляет собой трансформированный картофель Mtd 2, дорожка 4 представляет собой трансформированный картофель Mtd 3, дорожка 5 представляет собой трансформированный картофель Mtd 5 и дорожка 6 представляет собой трансформированный картофель Mtd 8.
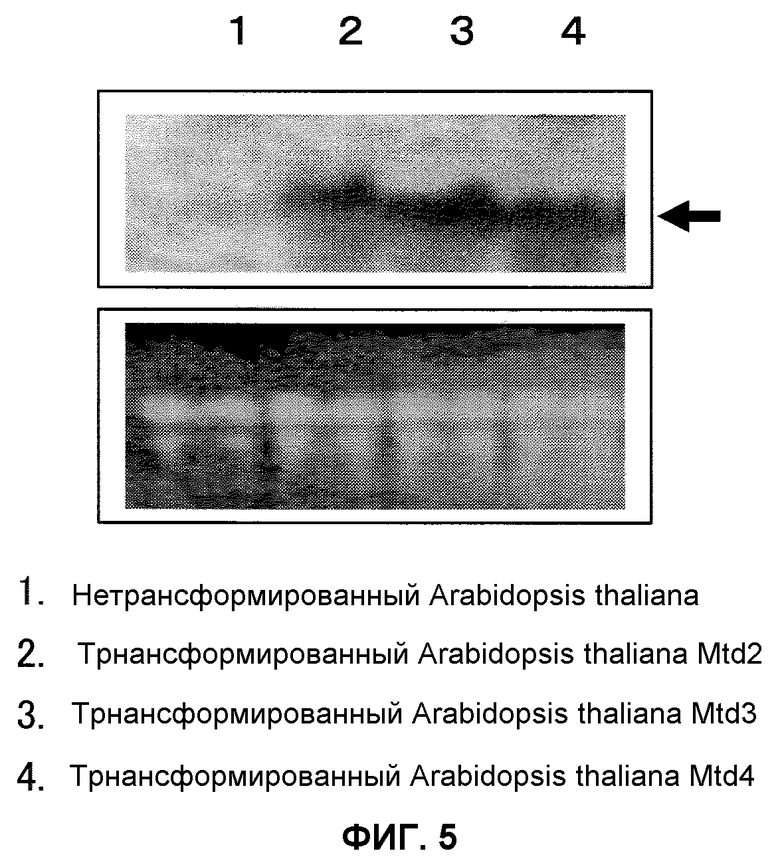
На фиг.5 показаны результаты нозерн-анализа трансформированного Arabidopsis thaliana. Применяли РНК, экстрагированную из тканей листьев. Рисунки (нижние рисунки) получали окрашиванием бромидом этидия 10 мкл тотальной РНК, которая была подвергнута электрофорезу. В качестве зонда применяли полноразмерный An-GDH-ген. Дорожка 1 представляет собой нетрансформированный Arabidopsis thaliana, дорожка 2 представляет собой трансформированный Arabidopsis thaliana Mtd 2, дорожка 3 представляет собой трансформированный Arabidopsis thaliana Mtd 3, и дорожка 4 представляет собой трансформированный Arabidopsis thaliana Mtd 4.
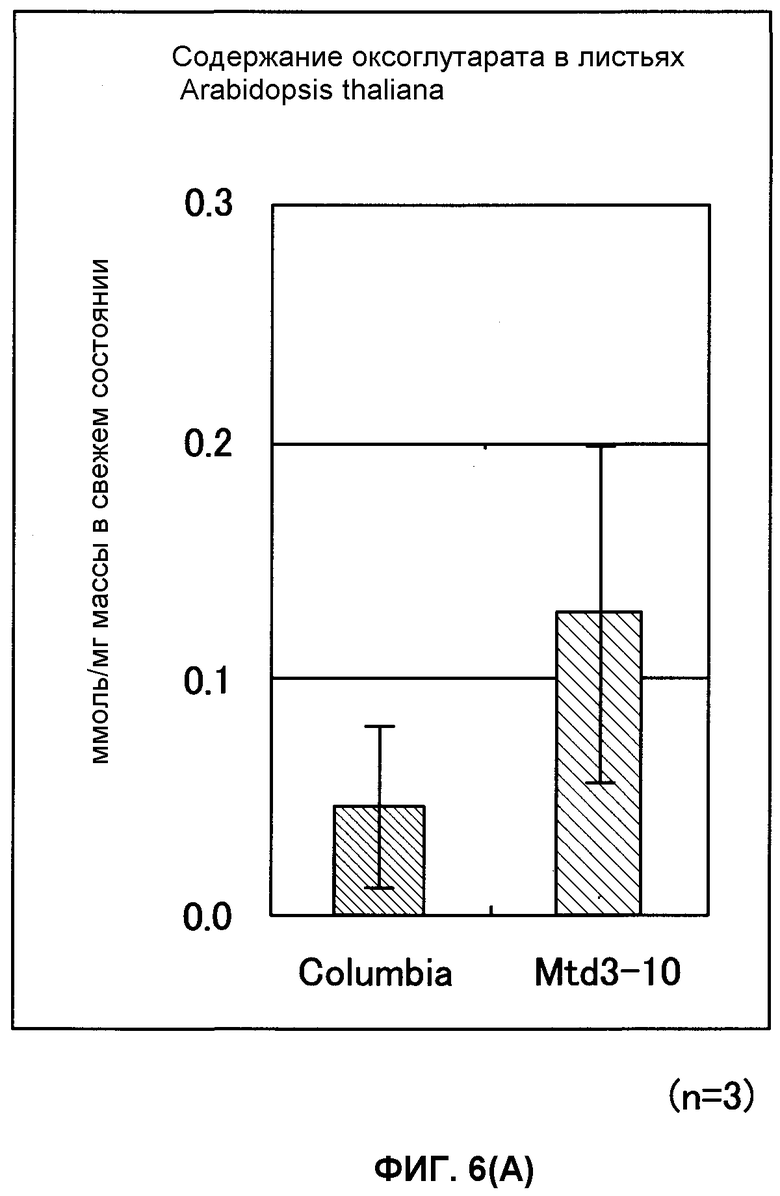
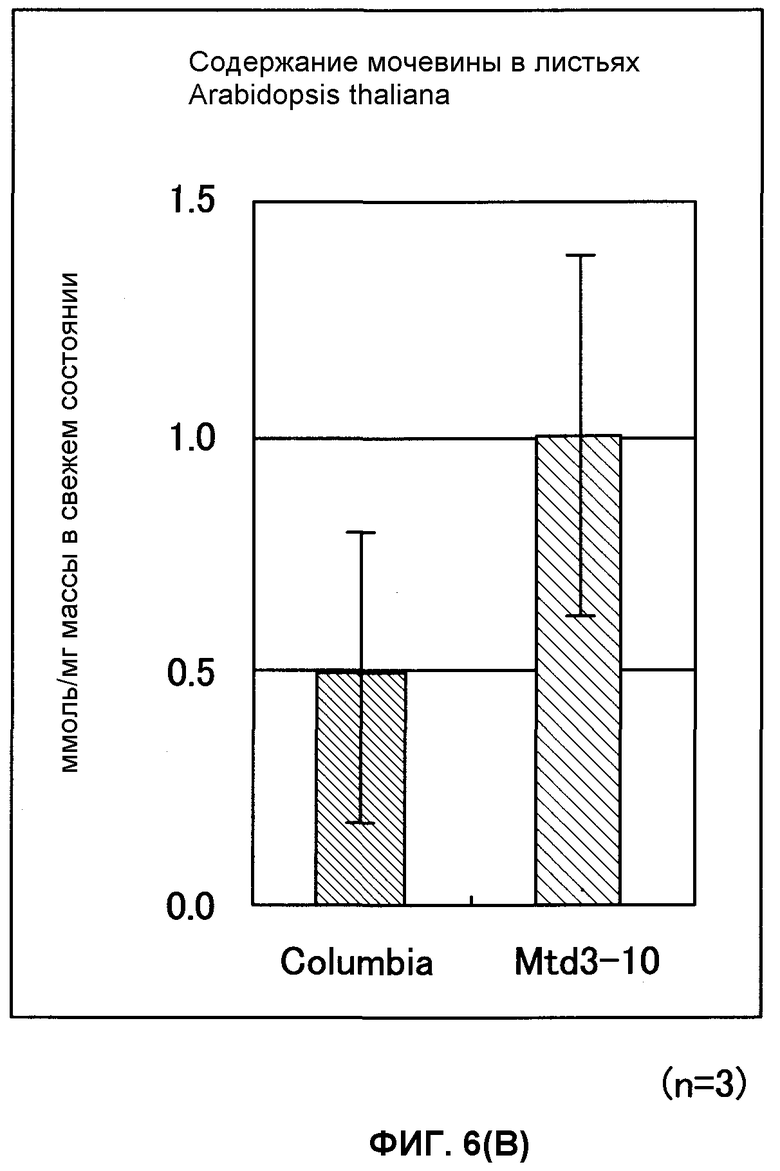
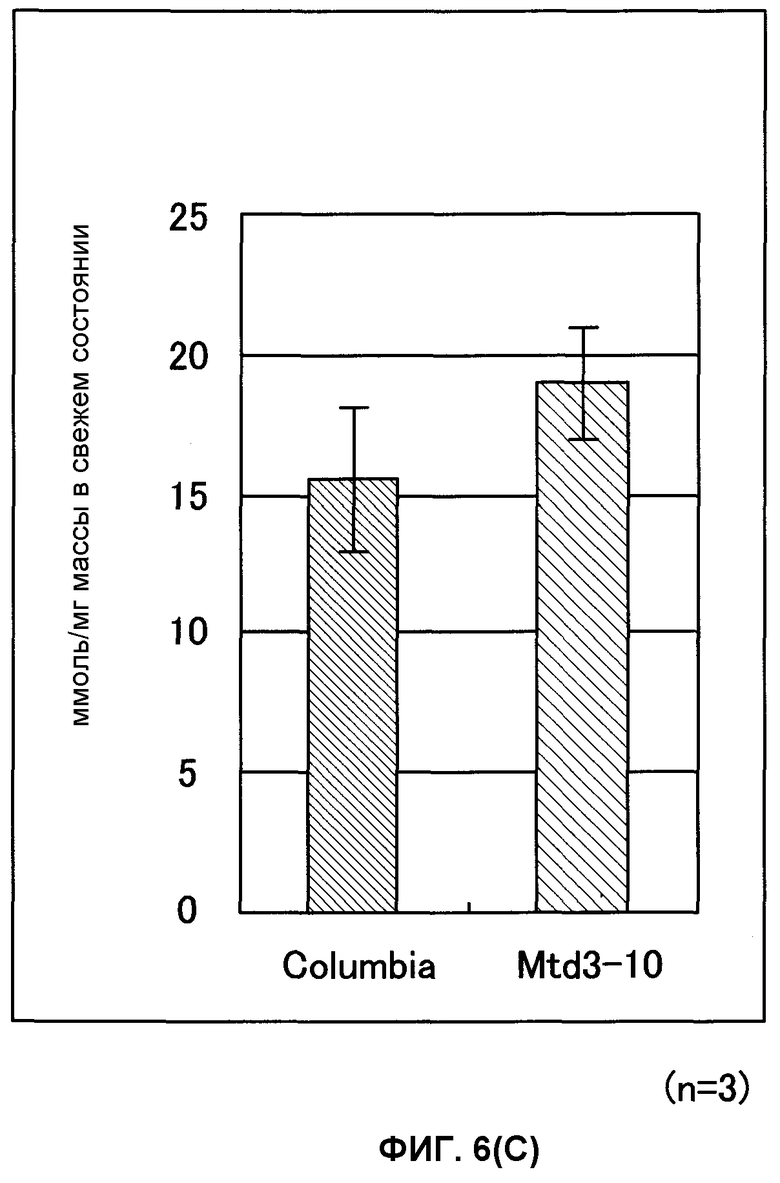
Фиг. от 6А до 6С представляют собой графики, показывающие содержание 2-оксоглутарата (2-OG), мочевины и нитрата GDH-трансформированных растений (Arabidopsis thaliana). Вертикальная ось указывает число молей (нмоль) на сырую массу соответственно, причем Mtd3-10 представляет собой трансформированный Arabidopsis thaliana (штамм № 3) и Columbia представляет собой нетрансформированный Arabidopsis thaliana (n = 3).
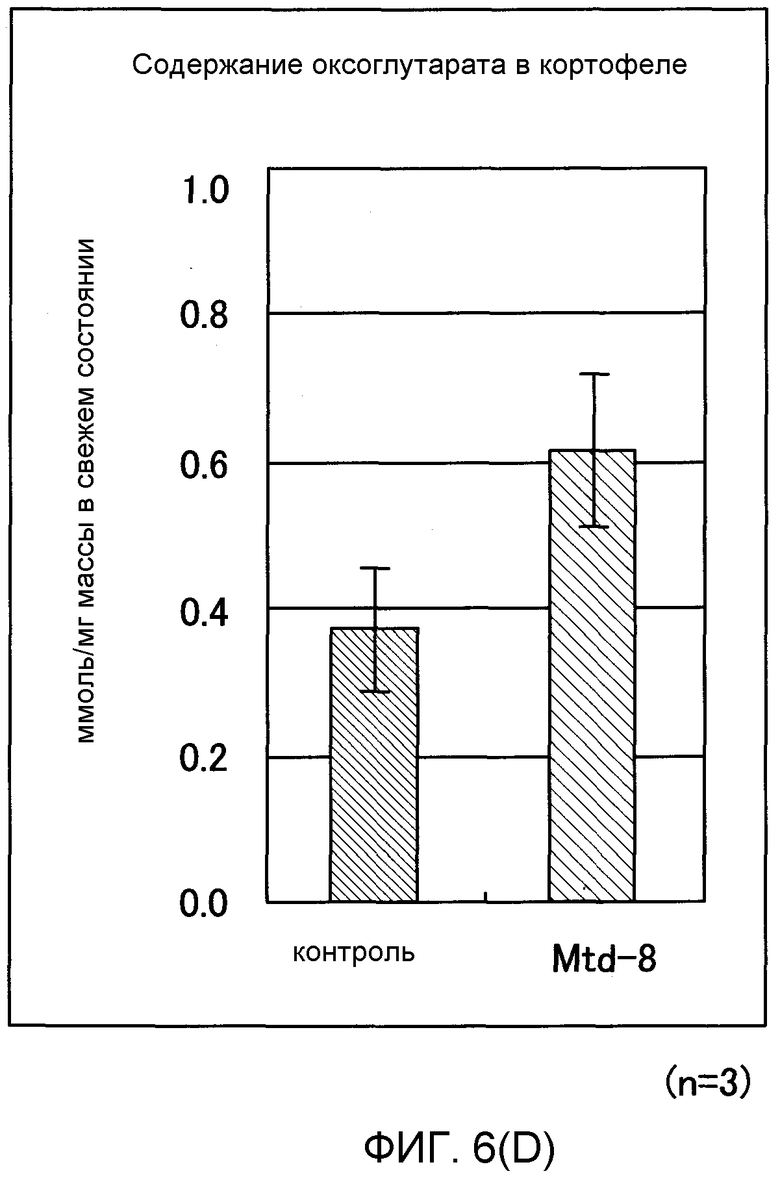
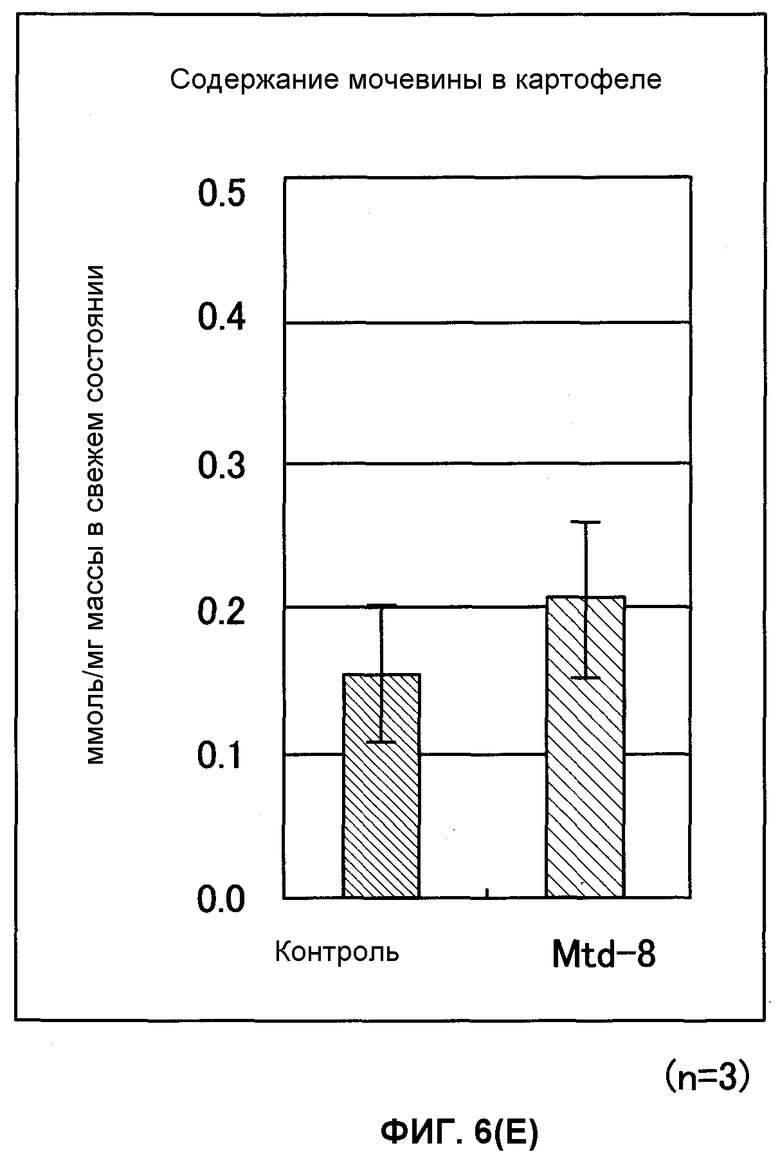
Фиг. от 6D до 6F представляют собой графики, показывающие содержание 2-оксоглутарата (2-OG), мочевины и нитрата GDH-трансформированных растений (картофеля). Вертикальная ось указывает число молей (нмоль) на сырую массу соответственно, причем Mtd-8 представляет собой трансформированный картофель и контроль представляет собой нетрансформированный картофель (n = 3).
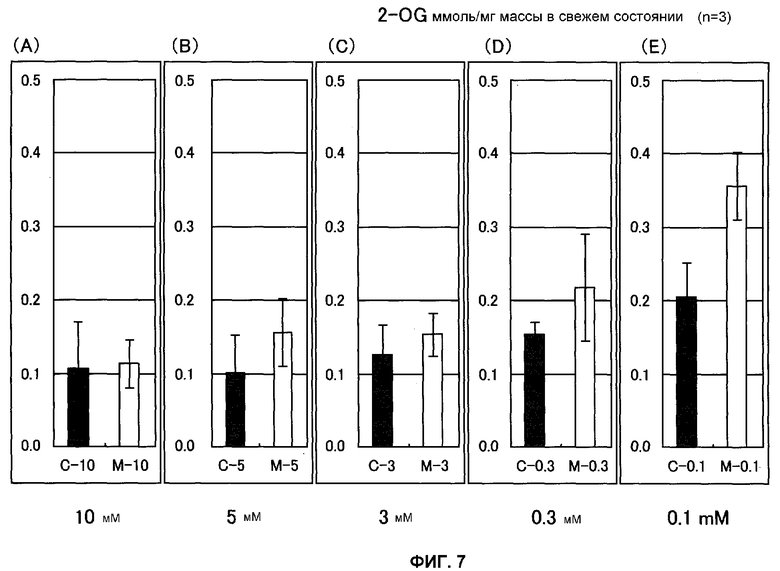
Фиг.7 представляет собой график, показывающий содержание 2-OG ткани листьев Arabidopsis thaliana спустя 3 недели культивации на среде PNS, содержащей различные количества KNO3. Число молей (нмоль) для каждой живой массы показано на вертикальной оси, причем С представляет собой нетрансформированный Arabidopsis thaliana и М представляет собой трансформированный Arabidopsis thaliana (штамм Mtd 3) (n = 3).
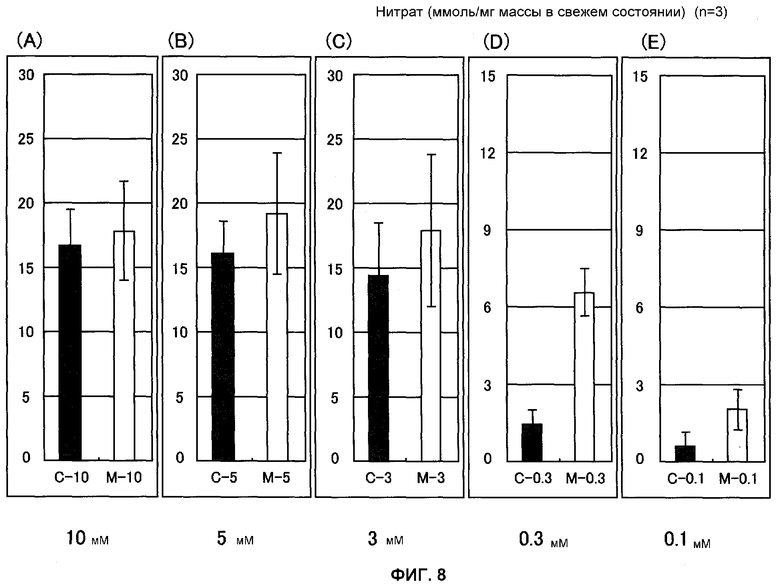
Фиг.8 представляет собой график, показывающий содержание нитрата в ткани листьев Arabidopsis thaliana после 3 недель культивации на среде PNS, содержащей различные количества KNO3. Число молей (нмоль) для каждой живой массы показано на вертикальной оси, причем С представляет собой нетрансформированный Arabidopsis thaliana и М представляет собой трансформированный Arabidopsis thaliana (штамм Mtd 3) (n = 3).
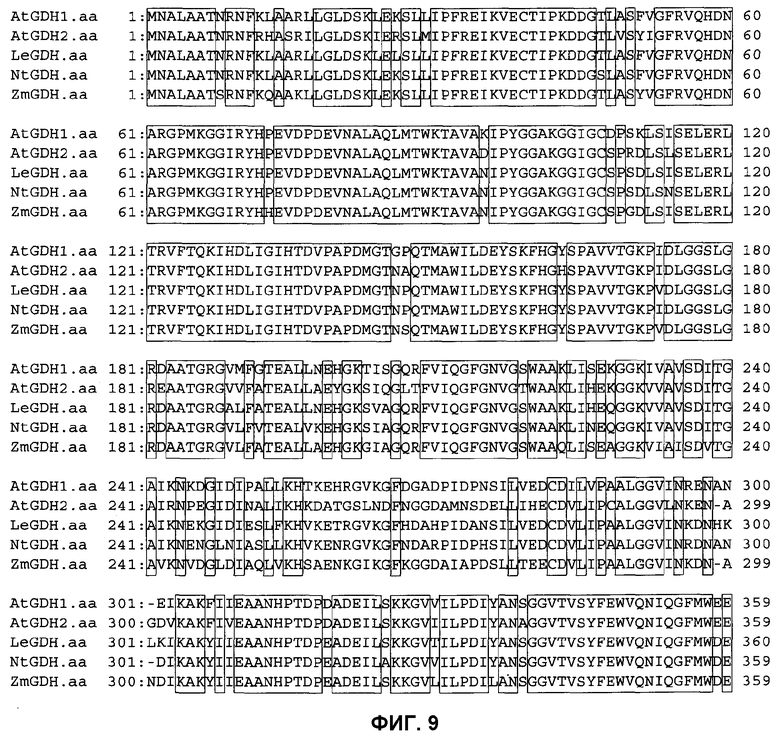
На фиг.9 показаны результаты сравнительного анализа первичной структуры аминокислотных последовательностей глутаматдегидрогеназы для высших растений, в которых AtGDH1 представляет собой GDH1 Arabidopsis thaliana, AtGDH2 представляет собой GDH2 Arabidopsis thaliana, LeGDH представляет собой GDH томата (Lycopersicon esculentum), NtGDH представляет собой GDH табака (Nicotiana tabacum) и ZmGDH представляет собой GDH кукурузы (Zea Mays).
На фиг.10 показаны результаты сравнительного анализа первичной структуры для аминокислотных последовательностей GDH плесени и GDH растения (томата), причем AaGDH представляет собой GDH Aspergillus awamori, AnGDH представляет собой GDH Aspergillus nidulans и LeGDH представляет собой GDH томата.
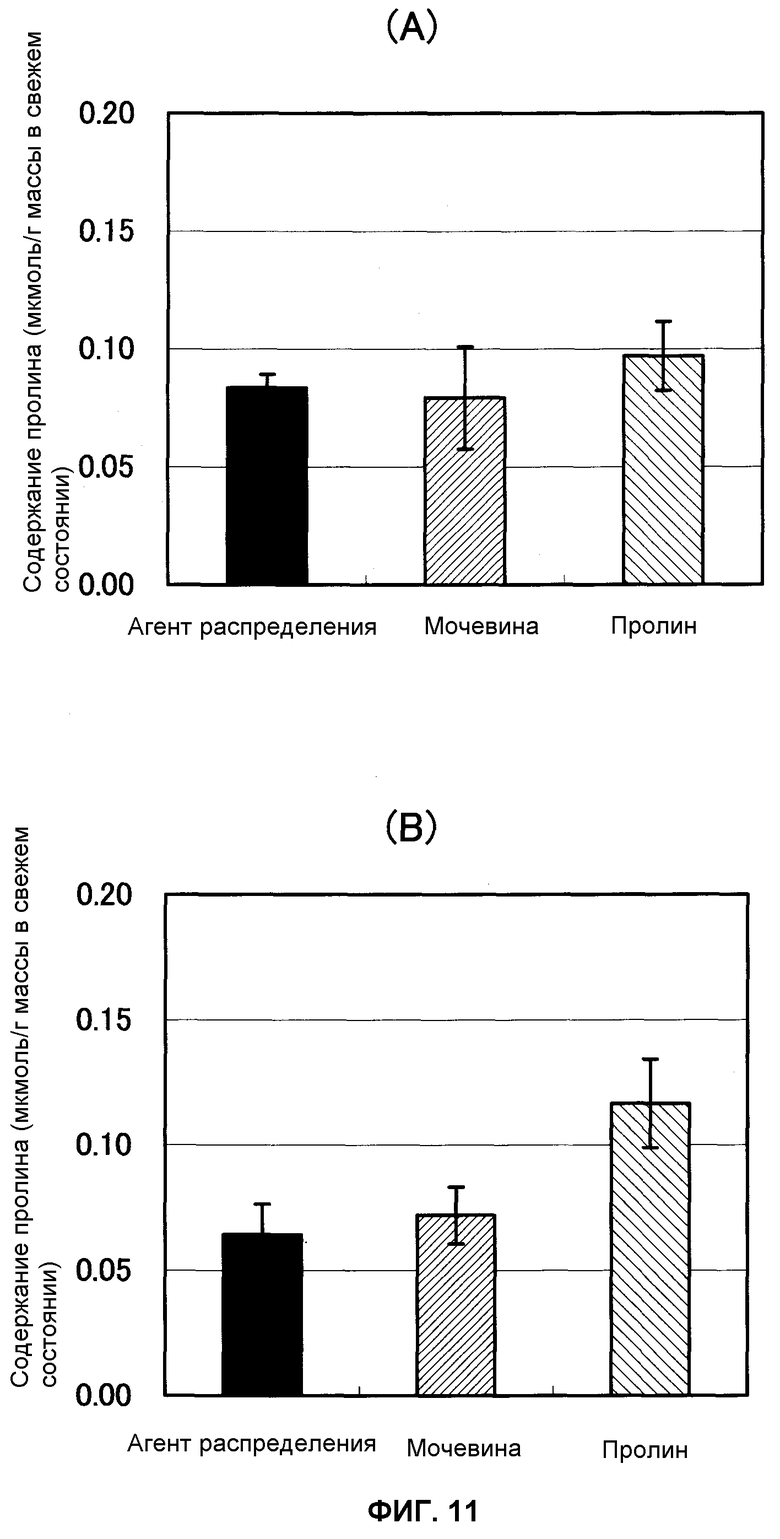
Фиг.11 представляет собой график, показывающий содержание пролина в ткани листьев спустя 1 и 5 часов после некорневой подкормки водным раствором диффундирующего агента, мочевины и пролина (n = 6) нанесением на ткани листьев картофеля соответственно, причем А указывает содержание через 1 час после нанесения и В указывает содержание через 5 часов после нанесения.
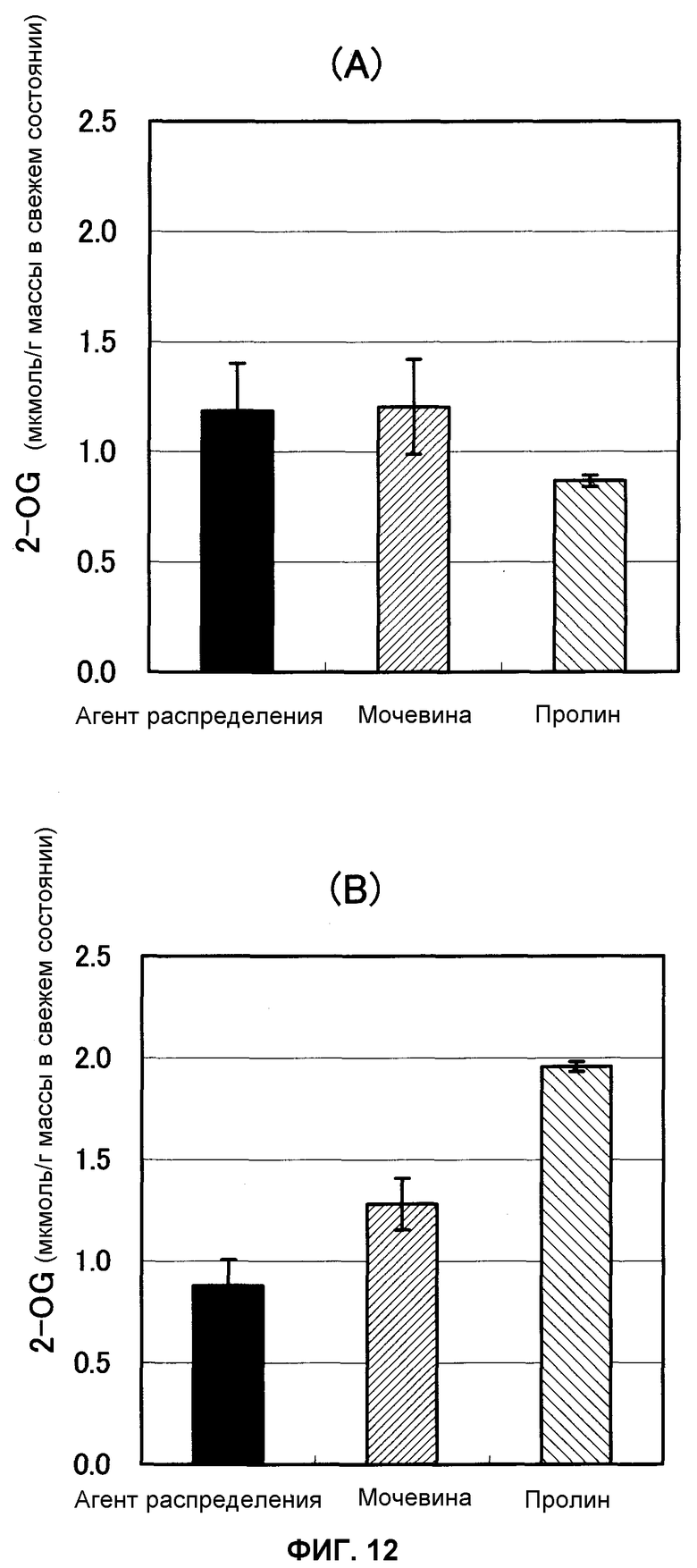
Фиг.12 представляет собой график, показывающий содержание 2-OG в ткани листьев спустя 1 и 24 часа после некорневой подкормки водным раствором диффундирующего агента, мочевины и пролина (n = 6) нанесением на ткани листьев картофеля соответственно, причем А указывает содержание через 1 час после нанесения и В указывает содержание через 24 часа после нанесения.
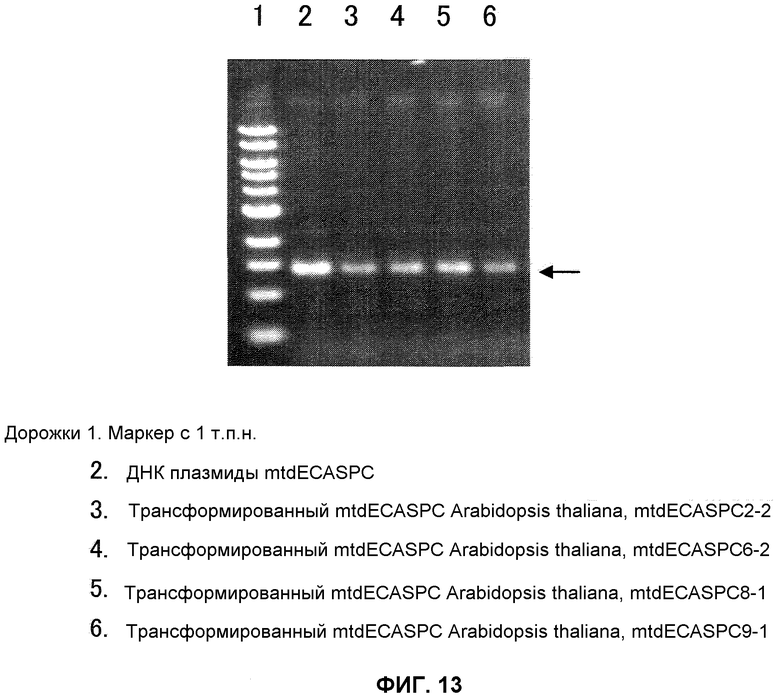
На фиг.13 показаны результаты анализа ПЦР генома Arabidopsis thaliana, трансформированного MtdECASPC. Анализ ПЦР проводили с применением ECASPC-специфичных праймеров в реакции из 30 циклов, состоящих из 1 минуты при 94°С, 1 минуты при 55°С и 2 минут при 72°С. После электрофореза в 1% геле агарозы проводили окрашивание бромидом этидия. Дорожка 1 представляет собой маркер с 1 т.п.н., дорожка 2 представляет собой ДНК плазмиды mtdECASPC, дорожка 3 представляет собой трансформированную mtdECASPC Arabidopsis thaliana mtdECASPC2-2, дорожка 4 представляет собой трансформированную mtdECASPC Arabidopsis thaliana mtdECASPC6-2, дорожка 5 представляет собой трансформированную mtdECASPC Arabidopsis thaliana mtdECASPC8-1 и дорожка 6 представляет собой трансформированную mtdECASPC Arabidopsis thaliana mtdECASPC9-1.
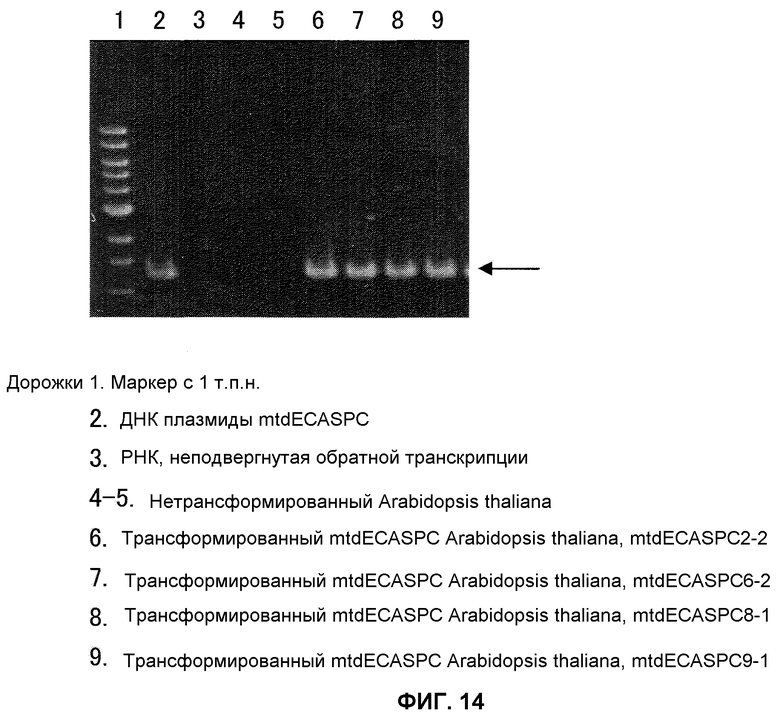
На фиг.14 показан ОТ-ПЦР-анализ трансформированного mtdECASPC Arabidopsis thaliana. РНК экстрагировали из ткани листьев Arabidopsis thaliana с применением набора Qiagen Plant Rneasy Mini-Kit. ОТ-ПЦР проводили с применением набора TaKaRa РНК-ПЦР с ECASPC-специфичными праймерами в реакции 30 циклов, состоящих из 1 минуты при 94°С, 1 минуты при 55°С и 2 минут при 72°С. После электрофореза в 1% геле агарозы проводили окрашивание бромидом этидия. Дорожка 1 представляет собой маркер с 1 т.п.н., дорожка 2 представляет собой ДНК плазмиды mtdECASPC, дорожка 3 представляет собой РНК без реакции обратной транскриптции, дорожки 4-5 представляют собой нетрансформированный Arabidopsis thaliana, дорожка 6 представляет собой трансформированную mtdECASPC Arabidopsis thaliana mtdECASPC2-2, дорожка 7 представляет собой трансформированную mtdECASPC Arabidopsis thaliana mtdECASPC6-2, дорожка 8 представляет собой трансформированную mtdECASPC Arabidopsis thaliana mtdECASPC8-1 и дорожка 9 представляет собой трансформированную mtdECASPC Arabidopsis thaliana mtdECASPC9-1.
Описание предпочтительных вариантов осуществления
Как описано выше, авторы изобретения предполагают существование сигнальной молекулы, которая дает возможность клеткам растений ощущать условия питания азотом. Она является сигнальной молекулой, которая всестороннее регулирует абсорбцию и метаболизм азота в клетках растений, и считается, что посредством искусственного повышения или снижения количества этого вещества возможно поддержание в клетках растения абсорбции и метаболизма азота в повышенном состоянии. Кроме того, поскольку считается, что растение в таком стимулированном состоянии способно активно включать в себя азот из внешней среды, полагают, что азот может быть пополнен в условиях ограничения уровня азота, в результате чего происходит усиление роста и повышение урожайности. Кроме того, авторы изобретения предположили, что 2-оксоглутарат (2-OG) является такой сигнальной молекулой.
Первой стадией ассимиляции неорганического азота в органическую форму в растениях является в основном включение аммиака в глутамат для образования глутамина, реакция, которая катализируется глутаминсинтетазой (GS) и глутаматсинтетазой (GOGAT), как уже указано выше, считается, что этот цикл GS/GOGAT является основным путем ассимиляции азота в растениях. Следовательно, считается, что степень ассимиляции азота или степень недостатка азота в клетках растения отражает количество 2-OG.
Авторы изобретения предложили индуцирование сверхэкспрессии активности глутаматдегидрогеназы (GDH) или активности аспартатаминотрансферазы (ASPC) в качестве одного способа повышения содержания 2-OG в клетках растений. GDH катализирует обратимые реакции включения аммиака в 2-OG с продуцированием глутамата и высвобождения аммиака из глутамата с образованием 2-OG. Однако считается, что вследствие того, что в общем GDH имеет высокую величину Km относительно аммиака, более вероятно, что она разлагает глутамат на 2-OG и аммоний, чем включает аммиак в 2-OG с образованием глутамата, так что ожидается повышение образования 2-OG как результат реакции. С другой стороны, аспартатаминотрансфераза является ферментом, который катализирует реакцию переноса аминогрупп оксалоацетата, что приводит к продуцированию аспарагиновой кислоты и 2-OG, поэтому предполагалось повышение содержания 2-OG.
Авторы изобретения получили трансформированные растения, в которые введен ген GDH или ген ASPC, и подтвердили действительное повышение содержания 2-OG, а также подтвердили, что рост и урожайность этих растений были повышенными в условиях культивации, когда содержание азота ограничивают до уровня ниже нормальных условий культивации.
В одном варианте осуществления настоящего изобретения конструкцию нуклеиновой кислоты, содержащую ген глутаматдегидрогеназы (GDH) или ген аспартатаминотрансферазы (ASPC), вводят в растение для получения растения, которое имеет повышенное содержание 2-OG вследствие экспрессии гена глутаматдегидрогеназы (GDH) или гена аспартатаминотрансферазы (ASPC) в образовавшемся трансформированном растении и которое имеет усиленный рост и/или повышенную урожайность в условиях культивации, когда содержание азота ограничивают до уровня ниже нормальных условий культивации.
В другом варианте осуществления настоящего изобретения число копий и/или транскрибированное количество эндогенного гена GDH или эндогенного гена ASPC увеличивают для повышения активности GDH или ASPC с повышенным содержанием 2-OG растительного организма, что приводит к получению растения с усиленным ростом и/или повышенной урожайностью в условиях культивации, когда содержание азота ограничивают до уровня ниже нормальных условий культивации. Для увеличения транскрибированного количества может быть введена конструкция гена, способная экспрессировать фактор активации транскрипции и/или энхансер или другой цис-функциональный элемент. Такие факторы активации транскрипции и цис-функциональные элементы являются хорошо известными специалистам в данной области.
Не существует определенных ограничений по происхождению глутаматдегидрогеназы, применяемой в настоящем изобретении, но предпочтительно применяют глутаматдегидрогеназу из микроорганизма, принадлежащего к роду Aspergillus. Более предпочтительно применяют глутаматдегидрогеназу, полученную из Aspergillus nidulans и Aspergillus awamori. Кроме того, в настоящем изобретении можно применять глутаматдегидрогеназу, имеющую одну или несколько аминокислотных делеций, замещений или добавок в ферментном белке, до тех пор, пока она имеет указанную выше активность глутаматдегидрогеназы и селективно катализирует реакцию продуцирования 2-OG и аммиака из глутамата в физиологических условиях в растительных организмах.
Не имеется также особых ограничений по происхождению аспартатаминотрансферазы, применяемой в настоящем изобретении, но предпочтительно применяют аспартатаминотрансферазу, полученную из Escherichia coli. Кроме того, можно применять аспартатаминотрансферазу, имеющую одну или несколько аминокислотных делеций, замещений или добавок в ферментном белке, до тех пор, пока она имеет указанную выше активность аспартатаминотрансферазы.
Что касается локализации глутаматдегидрогеназы или аспартатаминотрансферазы в клеточных органеллах, можно применять глутаматдегидрогеназу или аспартатаминотрансферазу, локализованную в митохондрии, хлоропласте, пероксисоме или тому подобное, а также глутаматдегидрогеназу или аспартатаминотрансферазу, локализованную в цитоплазме. Следовательно, в настоящем изобретении можно также применять ген глутаматдегидрогеназы или аспартатаминотрансферазы, имеющий добавленный к его N- или С-концу либо сигнальный, либо транзитный пептид для переноса ферментного белка к этим клеточным органеллам.
Такой ген глутаматдегидрогеназы или его кДНК могут быть получены относительно легко специалистом в данной области на основе опубликованных данных по последовательности. В случае гена глутаматдегидрогеназы Aspergillus nidulans, например, нуклеотидная последовательность геномного гена опубликована под номером доступа GenBank X16121. Нуклеотидная последовательность показана в SEQ ID №: 1 и аминокислотная последовательность показана в SEQ ID №: 2. кДНК глутаматдегидрогеназы может быть легко получена синтезом праймеров ПЦР для амплификации ДНК-фрагментов так, чтобы включить часть, кодирующую белок на основе его последовательности, и проведением ОТ-ПЦР с применением в качестве матрицы РНК, экстрагированную из культивированных клеток Aspergillus nidulans. Последовательность кДНК гена глутаматдегидрогеназы Aspergillus nidulans показана в SEQ ID №: 17. Aspergillus nidulans может быть получен, например, в качестве АТСС10074 или АТСС11267 из Американской коллекции типовых культур. Последовательность генома для гена глутаматдегидрогеназы Aspergillus awamori также опубликована под номером доступа GenBank Y15784. Эта нуклеотидная последовательность показана в SEQ ID №: 3 и аминокислотная последовательность показана в SEQ ID №: 4. Последовательность кДНК гена глутаматдегидрогеназы Aspergillus awamori показана в SEQ ID №: 18. Сам Aspergillus awamori может быть получен из Американской коллекции типовых культур в качестве АТСС10548 и АТСС11358. кДНК глутаматдегидрогеназы может быть получена способами, подобными способам, описанным выше, на основе их штаммов и информации о последовательности.
Гомология между GDH Aspergillus nidulans и GDH Aspergillus awamori составляет приблизительно 84% в аминокислотной последовательности (фигура 10).
Кроме того, сравнительные результаты для аминокислотных последовательностей GDH Aspergillus nidulans, Aspergillus awamori и томата (Lycopersicon esculentum) показаны на фиг.10, тогда как сравнительные результаты для аминокислотных последовательностей GDH1 Arabidopsis thaliana, GDH2 Arabidopsis thaliana, GDH томата, GDH табака (Nicotiana Tabacum) и GDH кукурузы (Zea mays) показаны на фиг.9.
В настоящем изобретении растения, которые имеют усиленный рост и/или повышенную урожайность в условиях культивации, когда содержание азота снижают ниже уровня обычных условий культивации, и которые имеют повышенное содержание 2-OG растения, могут быть получены с применением генов, которые кодируют различные ферменты, которые продуцируют 2-OG из глутамата и которые отличаются от гена глутаматдегидрогеназы. Примеры таких ферментов включают аспартатаминотрансферазу и аланинаминотрансферазу.
Например, нуклеотидная последовательность ДНК генома для гена аспартатаминотрансферазы штамма К-12 E. coli зарегистрирована под номером доступа GenBank Х03629. Ген аспартатаминотрансферазы может быть легко получен синтезом праймеров ПЦР на основании этой последовательности так, чтобы амплифицировать ДНК-фрагмент, содержащий район, кодирующий белок, и проведением ПЦР с применением этих праймеров с хромосомной ДНК, экстрагированной из штамма К-12 Е. coli в качестве матрицы.
В настоящем изобретении рост и/или урожайность растений в условиях ограничения уровня азота, где содержание азота ограничивают до уровня ниже нормальных условий культивации, могут быть повышены нанесением пролина на поверхность листьев в качестве средства повышения содержания 2-OG в растениях. Пролин, нанесенный на листья, будет включаться в ткань листьев и метаболизироваться в глутамат. Кроме того, поскольку этот глутамат метаболизируется в 2-OG эндогенной глутаматдегидрогеназой, он может повысить содержание 2-OG в ткани. Растение, содержание 2-OG которого повысилось этим путем, нанесением пролина на поверхности листьев, усиливало рост и/или повышало урожайность в условиях, в которых количество доступного азота во время периода культивации ограничивают ниже количества в обычных условиях культивации.
Конструкция нуклеиновой кислоты, которую можно применять в настоящем изобретении, может быть получена с применением обычных способов, известных специалисту в данной области. Например, Sambrook et al, Molecular Cloning-Laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory Press или тому подобное можно применять для консультации в отношении способов молекулярной биологии, включающих способы выделения конструкции нуклеиновой кислоты и определения ее последовательности. В противоположность этому, в некоторых случаях генетическая амплификация, такая как ПЦР, необходима для получения конструкции нуклеиновой кислоты, которую можно применять в настоящем изобретении, и публикацию F. M. Ausbel et al (eds.), Current Protocols in Molecular Biology, John Wiley & Sons, Inc. (1994) можно применять для консультации в случае таких способов.
Конструкции нуклеиновых кислот, которые можно применять в настоящем изобретении, обычно включают в себя подходящие промоторы, которые функционируют в клетках растений, такие как промотор гена нопалинсинтазы или промотор вируса мозаичности цветной капусты 35S (CaMV35S), подходящие терминаторы, такие как герминатор гена нопалинсинтазы, другие последовательности, которые являются необходимыми или полезными для экспрессии, и маркерные гены для выбора трансформантов, такие как гены устойчивости к лекарственным средствам, например устойчивости к канамицину, устойчивости к G418, устойчивости к гигромицину и тому подобное.
Промоторами, которые можно применять для таких конструкций, могут быть конститутивные промоторы или орган-специфические или специфические для стадии роста промоторы, они могут быть выбраны в зависимости от хозяина, необходимого количества экспрессии, органов, в которых экспрессия является особенно желательной, или стадии роста. В предпочтительном варианте осуществления настоящего изобретения применяют сильный промотор, который экспрессируется путем, не специфичным для органов и стадий роста, одним примером такого промотора является промотор CaMV35S. Орган-специфические промоторы, которые можно применять, включают в себя промотор гена фазеолина и промотор гена пататина. В наиболее желательном варианте осуществления настоящего изобретения применяют конструкцию, которая регулирует ген глутаматдегидрогеназы или ген аспартатаминотрансферазы при помощи сильного структурного промотора, такого как промотор CaMV35S.
Не существуют определенные ограничения по способу введения гена, применяемому в настоящем изобретении, и способ, известный специалисту в данной области для введения генов в клетки растений или организмы растений, может быть выбран в соответствии с хозяином. Например, в одном варианте осуществления настоящего изобретения применяют способ введения гена с применением Agrobacterium. В этой системе трансформации предпочтительно применяют бинарный вектор. При применении Agrobacterium конструкция нуклеиновой кислоты, применяемая в трансформации гена, может включать в себя также Т-ДНК-район, фланкирующий ДНК-последовательность, которую нужно ввести в клетки растения. В предпочтительном варианте осуществления введенную последовательность встраивают между левой и правой пограничными последовательностями T-ДНК. Подходящая разработка и конструкция вектора трансформации, основанная на такой Т-ДНК, хорошо известна специалисту в данной области. Условия инфицирования растения Agrobacterium, имеющей такую конструкцию нуклеиновой кислоты, также хорошо известны специалистам в данной области. Что касается таких способов и условий, см. Shujunsha Pub. Saibou Rongaku Supplement, "Experimental Protocols in model Plants: Rice and Arabidopsis thaliana" (1996).
В настоящем изобретении можно применять другой способ введения гена. Примеры способов введения гена, которые можно применять, включают в себя введение протопластной ДНК с применением полиэтиленгликоля или кальция, трансформацию протопласта с применением электропорации и введение с применением способа выстреливания частицами и тому подобное.
Не имеются определенные ограничения по растению, которое подвергают генетической манипуляции этим путем, желательно применять вид, который можно легко трансформировать и для которого установлена генеративная система, за исключением случаев, когда трансформацию проводят с применением цельного растения. Кроме указанных выше свойств, подходящим растением в настоящем изобретении является растение, для которого установлены способы массовой культивации, чтобы уменьшить нагрузку на окружающую среду сельского хозяйства. Примеры растений, которые являются подходящими для настоящего изобретения, включают в себя все растения семейства Brassica, а также томаты, картофель, кукуруза, пшеница, рис, сахарный тростник, соевые бобы, сорго и тому подобное. Примеры растений, которые трудно питать большими количествами азота, включают в себя деревья и фруктовые деревья, такие как тополь, эвкалипт и яблоневые деревья. Активности и свойства GDHs и ASPCs этих растений являются эквивалентными активностям и свойствам полученного из Aspergillus nidulans GDH (SEQ ID №№: 1 и 2) и полученного из штамма К-12 E. coli ASPC (SEQ ID №№: 19 и 20), применяемых в одном варианте осуществления настоящего изобретения. Следовательно, содержание 2-OG в этих растениях может быть повышено введением в них полученного из Aspergillus nidulans гена GDH (SEQ ID №: 1) или полученного из E. coli гена ASPC (SEQ ID №: 1). Ген GDH или ген ASPC, полученный из любого из этих растений, можно было также применять таким же образом в настоящем изобретении. Не имеется определенных ограничений по тому, какие органы или клетки подвергают генетической манипуляции, как описано выше, их можно выбрать в зависимости от хозяина, способа генетического введения и тому подобное. Примеры включают в себя орган-эксплантат, пыльцу, культивированные клетки, зародыш, растительные организмы и тому подобное, но не ограничиваются ими.
Для трансформации выбирают клетки растений или подобное, которые подвергали манипуляции, как описано выше. Выбор может быть основан на экспрессии маркерного гена, который присутствует на конструкции нуклеиновой кислоты, применяемой для трансформации. Например, когда маркерным геном является устойчивый к лекарственному средству ген, подвергнутые манипуляции клетки растений или подобные материалы могут быть культивированы или выращены на среде, содержащей подходящую концентрацию антибиотика, гербицида или тому подобное. Если маркерным геном является ген бета-глюкуронидазы, ген люциферазы или тому подобное, трансформант может быть выбран скринингом по его активности. Когда трансформант, идентифицированный этим путем, не является растительным организмом, но является протопластом, каллюсом, эксплантатом или тому подобное, его можно регенерировать в цельном растительном организме. Способ, известный специалисту в данной области, можно применять для регенерации в зависимости от растения-хозяина.
Образовавшееся цельное растение можно культивировать обычными способами или, другим словами, в тех же самых условиях, как и нетрансформат, и кроме указанной выше селекции, основанной на маркерных генах, для идентификации трансформированных растений, содержащих конструкцию нуклеиновой кислоты настоящего изобретения, можно применять различные способы молекулярной биологии. Для детекции присутствия или отсутствия структуры рекомбинантной ДНК-встроенных фрагментов можно применять, например, саузерн-гибридизацию или ПЦР. Для детекции и измерения продуктов транскрипции РНК, полученных из введенной конструкции нуклеиновой кислоты, можно применять назерн-гибридизацию, ОТ-ПЦР и тому подобное.
Затем экспрессию гена глутаматдегидрогеназы или гена аспартатаминотрансферазы образовавшимся трансформантом оценивают измерением количества белка, мРНК или ферментативной активности глутаматдегидрогеназы или аспартатаминотрансферазы. Например, количество белка глутаматдегидрогеназы или белка аспартатаминотрансферазы можно оценить вестерн-блоттингом или подобным образом и количество мРНК можно оценить назерн-блоттингом или количественной ОТ-ПЦР. Активность глутаматдегидрогеназы в экстракте растения можно измерить способом Ameziane et al (Ameziane, R., Bernhard, K. and Lightfood, D., Plant and Soil 221: 47-57, 2000). Активность аспартатаминотрансферазы в экстракте растения можно измерить способами Chao et al (Chao, Y. P., Lai, Z.I., Chen, P. and Chen. J.T., Biotechnology Progress 15:453-458, 1999).
Трансформированное растение, в котором экспрессию или повышенную экспрессию гена глутаматдегидрогеназы или гена аспартатаминотрансферазы подтвердили, дополнительно оценивают на содержание 2-OG. Содержание 2-OG можно анализировать, например, ферментным способом с применением экстракта растения, полученного дроблением всего количества или части трансформированного растительного организма. Экстракт растения может быть получен и анализ 2-OG может быть проведен, например, способами Usuda (Usuda, H. Plant Physiol. 78:859-864, 1985).
Если содержание 2-OG наземных частей (например, листьев, стеблей или обоих, цветковых органов или части плода) или подземных частей (например, корней или клубней) в нормальных условиях культивации и в условиях, когда содержание азота ограничивают до уровня ниже, чем уровень при нормальных условиях культивации, повышается в 1,2-раза или больше по сравнению с первоначальной линией, в которую был введен ген, считается, что содержание 2-OG повышается.
Трансформированное растение, в котором повышенное содержание 2-OG подтверждали таким путем, можно культивировать в условиях культивации, в которых содержание азота ограничивают до уровня ниже уровня нормальных условий культивации во время периода культивации, можно оценить также его рост или урожайность. Применяемое здесь выражение "условия, при которых содержание азота ограничивают до уровня ниже уровня нормальных условий культивации", относится к условию, в котором вносят количество азота, которое меньше (например, такое как 70% или ниже или, предпочтительно, 50% или ниже), чем стандартное количество азота, или, другими словами, стандартная норма внесения азота, которую обычно применяют при культивации каждого растения. В частности, по способу настоящего изобретения рост и/или урожайность, эквивалентную или выше, чем рост и/или урожайность, достигаемая в условиях культивации с внесением стандартного количества азота, достигают в условиях культивации, в которых обеспечивают количество азота, находящееся между количеством, которое ниже нижнего предела стандартного количества и 1/10-кратным нижним пределом. Стандартные количества внесения азота, например, устанавливают для индивидуальных сельскохозяйственных культур. В случае картофеля, например, стандартным количеством азота является 7 кг на 10 акров (JRT Japanese Society of Root and Tuber Crops home page: http://www.jrt.gr.jp/mini/pm_index.html; Minoru Yoshida. Enjov Learning All About Potatoes, Nobunkyo, 1988). тогда как для риса оно составляет 5-15 кг на 10 акров (A Manual of Experiments for Plant Bioloagy, Ed. Kokichi Hinata and Teruyoshi Hashiba, Soft Science Publications, 1995; Nara Agricultural Technology Center home page, http://www.naranougi.jp/sehi-kijyun/sehikijyun-index.html).Для Arabidopsis thaliana и других экспериментальных растений среда, содержащая 5 мМ или больше нитрата, считается стандартной средой, подходящей для роста и развития (Martinez-Zapater, J.M. and Salinas, J. Ed., Methods in Molecular Biology, Arabidopsis thaliana Protocols, Humana Press. New Jersey, 1998). Введение такой стандартной нормы азота или стандартное количество азота в среде хорошо известно специалисту в данной области.
Трансформированное растение культивируют в условиях с содержанием азота, ограниченным до уровня, которое ниже уровня нормальных условий культивации, или, другими словами, как описано выше, с количеством азота в диапазоне между уровнем ниже нижнего предела стандартной нормы внесения азота и 1/10 нижнего предела, и оценивают его рост и/или урожайность. Рост можно оценить измерением наземных частей или длины растения, числа листьев или массы всего растения в свежем состоянии или в сухом состоянии. Повышенный рост означает, что, по меньшей мере, один из параметров, относящихся ко всему растительному организму или части растительного организма (такой как длина растения, число листьев, масса в свежем состоянии или в сухом состоянии) выше в 1,2 раза или более по сравнению с параметром контрольного растения, культивированного приблизительно в то же самое время при стандартной норме внесения азота. Урожайность может быть оценена на основе собранной части растения, например, определением числа или массы отдельных клубней для картофеля или зерен для риса, или общей массы собранных частей. Следовательно, улучшенная (или повышенная) урожайность означает, что эта собранная часть выше в 1,2 раза или больше по сравнению с урожайностью контрольного растения, которое культивируют приблизительно в то же самое время при стандартной норме внесения азота.
После того, как трансформированное растение, которое имеет лучший рост и урожайность, чем нетрансформированное растение, применяемое в качестве контроля, в условиях, в которых применение азота ограничивают до уровня ниже уровней нормальных условий культивации, было идентифицировано, затем исследуют генетическую стабильность его характеристик. С этой целью растение выводят, культивируют, и семена собирают в нормальных условиях и характеристики и их расщепление анализируют в потомстве. Присутствие или отсутствие, расположение, экспрессию и тому подобное введенной конструкции нуклеиновой кислоты в потомстве можно анализировать таким же путем, как для первой генерации (генерация Т1) трансформанта.
В трансформированном растении, рост и урожайность которого повышаются в условиях содержания азота, ограниченного до уровня, который ниже уровня нормальных условий культивации, или, другими словами, в трансформированном растении, рост которого больше не ингибируется в условиях ограниченного содержания азота, последовательность, полученная из конструкции нуклеиновой кислоты, включенной в введенный геном, может быть либо гемизиготной, либо гомозиготной, и либо гемизигота, либо гомозигота может быть получена в потомстве, если необходимо, кроссбридингом или подобным образом. Последовательность, полученную из конструкции нуклеиновой кислоты, интегрированной в геном, выделяют в потомстве способами менделевской генетики. Следовательно, для получения потомственных растений со стабильными характеристиками предпочтительно применяют гомозиготное растение.
В большинстве случаев трансформант имеет экзогенный ген, встроенный только в один генетический локус, а поликопийные трансформанты со вставками в несколько локусов гена не являются необычными. Однокопийный трансформант является предпочтительным в настоящем изобретении для целей стабильности и тому подобное введенного гена. Например, однокопийный трансформант с введенным геном в один генный локус может быть выбран исследованием отношения расщепления резистентности к канамицину в Т2 (вторая генерация). Если Т1 является гемизиготной и введенный ген находится в одном генном локусе, резистентность и восприимчивость к канамицину могут быть расщеплены как отношение 3:1 в Т2 по закону Менделя. Когда имеется множество копий введенного гена, частота резистентного трансформанта увеличивается. Следовательно, однокопийный трансформант может быть получен высеванием в чашке образовавшихся семян Т2 в среде, содержащей канамицин, отбором линии, которая проявляет отношение расщепления 3:1, и отбором тех трансформантов, в которых, как считается, введенный ген присутствует в одном генном локусе.
Образовавшееся трансформированное растение может быть культивировано в тех же условиях, что и существующее в природе растение того же вида, и при культивации в условиях, когда внесение азота ограничивают до уровня ниже уровня нормальных условий культивации, можно обеспечить получение урожая, равного урожаю или больше, чем урожай, полученный из нетрансформированного растения. Кроме того, из трансформированного растения, полученного таким путем, могут быть получены семена. Семена могут быть легко получены такими же способами, какие применяют для нетрансформированного растения такого же вида. Если необходимо, образовавшиеся семена можно сохранять, стерилизовать и освобождать от вредных насекомых обычными способами, известными специалисту в данной области.
Содержание 2-OG растения можно, таким образом, повысить введением гена и содержание 2-OG растения можно также дополнительно повысить нанесением на листья аминокислоты пролина. В частности, водный раствор 20 мг/л - 500 мг/л пролина можно разбрызгать или нанести другим образом вместе с подходящим распределяющим средством на поверхности листьев растения. Содержание 2-OG растения, которое получило такую некорневую подкормку пролина, может быть оценено способами, сходными со способами, описанными выше. Кроме того, влияние на рост в условиях ограниченного уровня азота можно также оценить в условиях и способами, сходными с условиями и способами, описанными выше.
Настоящее изобретение объясняется более конкретно и более подробно ниже при помощи нижеследующих примеров, включающих в себя получение урожая и определение урожайности и проведение анализа компонентов растений, подвергнутых манипуляции для сверхэкспрессии гена NADP-GDH или гена аспартатаминотрансферазы, и посредством нижеследующих примеров, включающих в себя определение урожайности и анализ компонентов растений, которые получали листовую подкормку пролина.
Примеры
Пример 1 Выделение происходящего из Aspergillus nidulans, NADP-зависимого гена GDH и конструкция плазмидного вектора Ti
(1) Выделение происходящего из Aspergillus nidulans, NADP-зависимого гена GDH
А. nidulans высевали на среде из картофельного декстрозного агара и культивировали на протяжении ночи при 30°С и образовавшиеся колонии культивировали в течение 2 дней в среде жидкой декстрозы. Тотальную РНК получали из пролиферированных клеток.
мРНК очищали с применением набора выделения мРНК Poly(A) Quick (Stratagene) и кДНК первой цепи получали с применением набора синтеза кРНК первой цепи (Amesham Bioscience). Реакцию ПЦР проводили с применением полученной кДНК первой цепи в качестве матрицы. Условиями реакции ПЦР были 35 циклов из 3 минут при 94°С, 45 секунд при 94°С, 30 секунд при 59°С и 90 секунд при 72°С и последующих 10 минут при 72°С, проводимых с применением системы ПЦР Перкин-Элмера 2400. Применяемыми праймерами были 5'-TCT AGA ATG TCT AAC CCC CTT GTT GAG-3' (SEQ ID №: 5) и 5'-GAG CTC TCA CCA CCA GTC ACC CTG GTC-3' (SEQ ID №: 6). Полосу 1,4 т.п.н. грубо подтверждали совпадением размера с размером предполагаемого гена. Образовавшийся продукт ПЦР клонировали с применением набора клонирования ТА (Invitrogen). Нуклеотидную последовательность образовавшегося клона определяли секвенатором (ABI 377A). Она совпадала с последовательностью известного, происходящего из А. nidulans, NADP-зависимого гена GDH, и была представлена как An-GDH (SEQ ID №: 17).
(2) Конструкция плазмидного вектора Ti
Нуклеотидную последовательность, кодирующую митохондриальный транзитный пептид, присоединяли к образовавшемуся гену. Это была 5'-концевая последовательность происходящего из томата NAD-зависимого гена GDH, который получали ПЦР с применением праймеров 5'-CTG CAG ATG AAT GCT TTA GCA GCA AC-3' (SEQ ID №: 7) и 5'-TCT AGA TAA ACC AAG AAG CCT AGC TG-3' (SEQ ID №: 8). Лигирование образовавшегося гена An-GDH с нуклеотидной последовательностью, кодирующей митохондриальный транзитный пептид, выполняли с применением четырех праймеров 5'-TCT AGA ATG AAT GCT TTA GCA GCA AC-3' (SEQ ID №: 9), 5'-GGG AAG GTT TAG ACA TTA AAC CAA GAA GCC T-3' (SEQ ID №: 10), 5'-AGG CTT CTT GGT TTA ATG TCT AAC CTT CCC-3' (SEQ ID №: 11) и 5'-GAG CTC TTA CGC CTC CCA TCC TCG AA-3' (SEQ ID №: 12). Образовавшийся ген GDH с присоединенной последовательностью митохондриального транзитного пептида обозначали как Mtd-An-GDH. Район GUS плазмиды Ti pIG121-Hm, трансформирующий вектор Agrobacterium, заменяли на ген Mtd-An-GDH. Образовавшуюся плазмиду Ti вводили в Agrobacterium tumefaciens EHA101 и применяли при трансформации Arabidopsis thaliana и картофельных растений.
Пример 2 Создание трансформанта картофеля
Картофель (сорт May Qeen) трансформировали по способам Gordon et al (Ref: Plant Cell Reports, 1993, 12:324-327). Микроклубеньки с индуцированной стерильностью нарезали и получали кружочки клубеньков, помещали их на среду агара MS, дополненную 2 мг/л зеатина и 0,1 мг/л индола и культивировали в течение 24 часов при 25°С с продолжительностью дневного времени 16 часов. Agrobacterium, содержащий конструированный ген, инокулировали на среде YEP (10 г/л триптона Bacto, 10 г/л дрожжевого экстракта, 1 г/л глюкозы), содержащей 50 мг/л канамицина и 50 мг/л гигромицина, и культивировали при встряхивании на протяжении ночи при 28°С. Культуру Agrobacterium добавляли для инфицирования кружочков клубеньков, которые культивировали в течение 24 часов. Спустя 10 минут избыточный Agrobacterium удаляли с применением стерилизованной фильтровальной бумаги, переносили в вышеуказанную чашку и культивировали в течение 24 часов в таких же условиях. Кружочки клубеньков затем переносили на среду агара MS, содержащую 50 мг/л канамицина, 300 мг/л гидрохлорида цефотаксима, 2 мг/л зеатина и 0,1 мг/л индолуксусной кислоты и отбирали трансформанты. Генерированные ростки переносили еще раз в вышеуказанную среду для отбора и подтверждали резистентность. Ростки, которые явно проявляли резистентность к канамицину, переносили в агаровую среду укоренения MS, содержащую 50 мг/л канамицина и 300 мг/л гидрохлорида цефотаксима, для индуцирования генерации корней. Кончики корней отрезали, по меньшей мере, три раза от укорененного растения и переносили в селективную среду укоренения, подтверждали резистентность к канамицину и химеры исключали. Образовавшиеся 5 индивидов акклиматизировали к почве для получения клубней.
Пример 3 Получение трансформантов Arabidopsis thaliana
Введение гена в Arabidopsis thaliana выполняли по способам Bechtold et al (Ref: C. R. Acad. Sci. Paris, Life Science 316: 1194-1199, 1993). Семена Arabidopsis thaliana высевали на почве для культивации и через 10 дней культивации при 24°С с продолжительностью дня 16 часов проростки переносили один за другим в отдельные кусочки шлаковой ваты и культивировали в течение дополнительных двух недель в таких же условиях. Растения пасынковали, как только они начинали выбрасывать цветонос и затем культивировали в течение еще недели. Agrobacterium культивировали при встряхивании в течение 24 часов при 28°С в культуре YEP, содержащей 50 мг/л канамицина, и 50 мг/л гигромицина и клетки собирали центрифугированием (7000 об/мин, 10 минут). Клетки суспендировали в среде суспендирования для инфильтрации (1/2 соли MS, 1/2 витамина В5, 5% сахароза, 0,5 г/л MES, 0,044 мкМ бензиламинопурин, рН 5,7). После удаления органов цветения, которые уже цвели или образовывали завязь, растения погружали в жидкость суспензии Agrobacterium и обрабатывали в десикаторе при пониженном давлении (40 мм Hg) в течение 15 минут. Обработанные растения культивировали в течение 1 месяца и семена собирали. Семена высевали на среде агара MS, содержащей 50 мг/л канамицина и 100 мг/л цефотаксима, и отбирали. Три из образовавшихся трансформантов далее культивировали до 3-ей генерации (Т3) и линии, которые больше не проявляли расщепления резистентности к канамицину, применяли в анализе.
Пример 4 Подтверждение введенного гена
ДНК экстрагировали из выбранных особей, проявляющих резистентность к канамицину (5 картофельных растений и 3 растения Arabidopsis thaliana), и из неинфицированных растений. ДНК экстрагировали по способам Honda et al (Ref: Honda and Hirai, 1990 Jpn J Breed 40: 339-348). Анализ ПЦР проводили с применением экстрагированной ДНК, специфичных для гена An-GDH праймеров 5'-TCT AGA ATG TCT AAC CCC CTT GTT GAG-3' (SEQ ID №: 13) и 5'-GAG CTC TCA CCA CCA GTC ACC CTG GTC-3' (SEQ ID №: 14) и праймеров 5'-CCC CTC GGT ATC CAA TTA GAG-3' (SEQ ID №: 15) и 5'-CGG GGG GTG GGC GAA GAA CTC CAG-3' (SEQ ID №: 16) для амплификации гена NPTII в векторе. Реакция состояла из 3 минут при 94°С, последующих 35 циклов из 45 минут при 94°С, 30 секунд при 55°С и 90 секунд при 72°С и последующих 10 минут при 72°С, ее проводили применением системы ПЦР Перкин-Элмера 2400. Продукт ПЦР подвергали электрофорезу в 1% геле агарозы и окрашивали бромидом этидия.
В результате этого An-GDH-ген-специфичную полосу (приблизительно 1,5 т.п.н.) и NPTII-ген-специфичную полосу (приблизительно 1,1 т.п.н.) подтверждали в отобранных трансформированных картофельных растениях (фиг.2) и трансформированной Arabidopsis thaliana (фиг.2), тогда как никакие полосы не были подтверждены в неинфицированных растениях, что указывает на то, что Т-ДНК-район, содержащий ген An-GDH, был введен в трансформированные картофельные растения и трансформированный Arabidopsis thaliana.
Пример 5 Подтверждение экспрессии введенного гена (нозерн-анализ)
Экспрессию введенного гена подтверждали нозерн-анализом с применением трансформированных картофельных растений и трансформированного Arabidopsis thaliana, в которые введение гена An-GDH было подтверждено.
РНК экстрагировали способом SDS-фенола из ткани листьев трансформированного картофеля и ткани листьев трансформированного Arabidopsis thaliana. Экстрагированную РНК подвергали электрофорезу в 1,2% геле агарозы, содержащем 18% формальдегида, и окрашивали бромидом этидия. После блоттинга на найлоновой мембране (HybondN+) ее фиксировали УФ и гибридизировали. Полноразмерный ген An-GDH применяли в качестве зонда. Нозерн-блот и зонд получали с применением набора II Roche Diagnostics DIG-High Prime DNA Labeling and Detection Starter и набора PCR DIG Probe Synthesis.
В результате этого, специфичную для An-GDH-гена полосу подтвердили в ткани листьев и клубней трансформированных картофельных растений (фиг.4), причем было показало, что введенный ген был транскрибирован и экспрессирован в ткани листьев и в клубнях трансформированных картофельных растений. Специфичную для An-GDH-гена полосу также подтвердили в ткани листьев трансформированного Arabidopsis thaliana (фиг.5), причем было указано, что введенный ген был транскрибирован и экспрессирован в ткани листьев трансформированного Arabidopsis thaliana. Эти специфичные для An-GDH полосы не наблюдали в нетрансформированных картофельных растениях и растениях Arabidopsis thaliana.
Пример 6 Экспрессия введенного гена (активность NADP-GDH)
Активность NADP-GDH измеряли для подтверждения экспрессии введенного гена с применением трансформированных картофельных растений и растений Arabidopsis thaliana, в которых введение гена An-GDH было подтверждено.
Активность измеряли по способам Ameziane et al (Ref: Plant and Soil, 2000, 221:47-57). Ткань листьев трансформированного картофеля (приблизительно 0,2г) и трансформированного Arabidopsis thaliana (приблизительно 0,2г) сушили вымораживанием жидким азотом и измельчали в ступке и добавляли 5-кратное по массе количество буфера экстракции (200 мМ Трис (рН 8,0), 14 мМ бета-меркаптоэтанол, 10 мМ L-цистеин-HCl, 0,5 мМ PMSF). Смесь переносили в пробирку центрифуги и центрифугировали в течение 30 минут при 4°С, 12000 об/мин, и супернатант ультрафильтровали (Millipore, Ultrafree 0,5 filter unit, Biomax-10) и промывали 3 раза в буфере для экстракции.
Активность определяли измерением снижения в поглощении при длине волны 340 нм, когда реакционную смесь, содержащую 100 мМ Трис (рН 8,5), 20 мМ 2-оксоглутаровую кислоту, 10 мМ CaCl2, 0,2 мМ NADPH и 200 мМ NH4Cl, добавляли к предварительно экстрагированному неочищенному ферментному раствору. Концентрацию белка в экстрагированном неочищенном ферментном растворе измеряли способом Брэдфорда с применением бычьего сывороточного альбумина (BSA) в качестве стандартного белка.
В результате этого было показано, трансформированный картофель проявлял приблизительно 20-50-кратную активность нетрансформированного картофеля (таблица 1). Трансформированный Arabidopsis thaliana также проявлял приблизительно 5-12-кратную активность нетрансформированного Arabidopsis thaliana (таблица 2). Поскольку нетрансформанты почти не проявляли активность NADP-GDH, активность относили к экспрессии введенного гена An-GDH.
Активность NADP-GDH в трансформированных картофельных растениях и нетрансформированном картофеле
Активность NADP-GDH в трансформированных Arabidopsis thaliana и нетрансформированном Arabidopsis thaliana
Пример 7 Содержание 2-оксоглутарата (2-OG), мочевины и нитрата в Arabidopsis thaliana и картофеле с введенным геном GDH
Измеряли содержание 2-OG, мочевины и нитрата в тканях листьев трансформированного картофеля (Mtd 8) и трансформированного Arabidopsis thaliana (Mtd). Для картофеля ткани листьев энергично растущего растительного организма, который культивировали в течение 1 месяца в смешанной почве из 500 г почвы Power и 500 г вермикулита, применяемой в качестве материала. Ткань листьев растения, которое выращивали в течение 3 недель в среде PNS (содержащей 5 мМ KNO3), применяли в случае Arabidopsis thaliana. Экстракцию проводили по способам Agarie et al (Ref: Plant Science 2002, 162:257-265). Ткань листьев (приблизительно 0,1 г) сначала измельчали с применением жидкого азота и затем хорошо смешивали после добавления 200 мкл 3% НСО4. К оставшемуся осадку добавляли дополнительно 200 мкл 3% НСО4 и смесь хорошо перемешивали. После 10-минутного центрифугирования при 12000 об/мин супернатант переносили в предыдущую пробирку. К собранному супернатанту для нейтрализации жидкости добавляли 50-70 мкл 5 М К2СО3. После того, как с применением лакмусовой бумаги подтвердили, что рН жидкости является нейтральной, объем жидкости регулировали до 600 мкл стерильной водой для получения образца для измерения. Для измерения азотной кислоты и мочевины соответственно применяли F-наборы Roche Diagnostics nitric acid and urea/ammonia. Способ Ameziane et al (см. выше) модифицировали для измерения содержания 2-OG. 20 мкл экстрагированного образца добавляли к 475 мкл реакционной жидкости (0,1 М Трис-HCl (рН 8,5), 1,0 мМ CaCl2, 0,2 мМ NaDPH, 0,2 М NH4Cl), измеряли поглощение при 340 нм и добавляли 5 мкл (10 единиц) глутаматдегидрогеназы (GDH) и реакцию проводили в течение 10 минут при 37°С. Снова измеряли поглощение при 340 нм и содержание 2-OG вычисляли по разности между поглощением после добавления ферментной жидкости и поглощением до добавления фермента.
В результате этого было показано, что трансформированный Arabidopsis thaliana имел содержание 2-OG, в 2,6 раза превышающее содержание мочевины, превышающее в 2 раза и содержание нитрата, превышающее в 1,2 раза содержания их в нетрансформированном Arabidopsis thaliana. Таким же образом показано, что трансформированный картофель имел содержание 2-OG, в 1,7 раз превышающее, содержание мочевины, в 1,5 раз превышающее и содержание нитрата, в 1,3 раза превышающее содержания их в нетрансформированном картофеле (фиг.6А-6F).
Содержание 2-OG и содержание нитрата измеряли также с применением ткани растения Arabidopsis thaliana, которое культивировали в течение 3 недель в среде, концентрацию азота в которой регулировали до 10, 5, 3, 0,3 и 0,1 мМ. В результате этого наблюдали, что разность между содержанием 2-OG трансформированного Arabidopsis thaliana и содержанием 2-OG нетрансформированного Arabidopsis thaliana повышалась, когда содержание азота среды уменьшалось (фиг.7). Эти результаты позволяют предположить, что введенная GDH функционирует с разложением глутамата и синтезом 2-OG и аммиака. Аккумуляция нитрата в трансформированном Arabidopsis thaliana и нетрансформированном Arabidopsis thaliana оставалась грубо постоянной в среде, содержащей 3 мМ или больше азота, это подтверждает тот факт, что в таких условиях аккумуляция в трансформированном Arabidopsis thaliana была, самое большее, в 1,3 раза выше, чем в нетрансформированном Arabidopsis thaliana. Однако в условиях 0,3 и 0,1 мМ азота она была в 2,5-4,5 раза выше (фиг.8). Эти результаты позволяют предположить, что нитрат аккумулируется значительно больше в ткани листьев трансформированного Arabidopsis thaliana, чем нетрансформированного Arabidopsis thaliana, что отражает повышенную эффективность поглощения азота.
Пример 8 Относящиеся к аккумуляции азота активности ферментов в ткани листьев Arabidopsis thaliana и картофеля с введенным геном GDH
Ферментативную активность NADH-зависимой глутаматсинтазы (NADH-GOGAT), нитратредуктазы (NR) и глутаматсинтазы (GS) измеряли в ткани листьев (приблизительно 0,1 г) трансформированных картофельных растений и трансформированного Arabidopsis thaliana. Для картофеля материалами были ткани листьев энергично растущих растений, которые культивировали в течение 1 месяца в смешанной почве из 500 г почвы Power и 500 г вермикулита. Ткани листьев растений, которые выращивали в течение 3 недель в среде PNS (содержащей 5 мМ RNO3), применяли для Arabidopsis thaliana. Неочищенный ферментный раствор экстрагировали способами Groat et al (Ref: Plant Physiol, 1981, 67: 1198-1203). После измельчения ткани листьев жидким азотом добавляли 500 мкл охлажденного льдом буфера для экстракции (100 мМ Mes-NaOH (рН 7,5), 100 мМ сахароза, 2% (об./об.) меркаптоэтанола, 15% этиленгликоля, 0,1% PMSF) и смесь осторожно перемешивали. После центрифугирования в течение 20 минут при 0°С, 12000 об/мин супернатант очищали Ultrafree (Biomax-10K, Millipore Co.). Его промывали три раз тем же буфером и растворяли в 100 мкл буфера для получения раствора фермента.
Активность NADH-GOGAT также измеряли способами Groat et al (см. выше). Приготовляли 500 мкл реакционного раствора (100 мМ фосфат К (рН 8,2), 100 мкМ NaDH, 2,5 мМ 2-оксоглутарат, 10 мМ L-глутамин, 1 мМ аминооксиацетат), в него добавляли 10 мкл ферментной жидкости и смесь перемешивали. Поглощение измеряли в течение 3 минут при 340 нм и измеряли снижение сопутствующего расходования NADH.
Активность NR измеряли способами Ferrario-Mery et al (Ref: Planta, 197, 202: 510-521). Приготовляли 490 мкл реакционного раствора (50 мМ Mops-KOH (рН 7,8), 1 мМ NaF, 10 мМ KNO3, 0,17 мМ NaDH и 10 мМ MgCl2 или 5 мМ EDTA), к каждому раствору добавляли 10 мкл раствора фермента и смесь перемешивали. Спустя 16 минут добавляли 500 мкл 1% сульфониламида (в 3 н HCl) и затем добавляли 10 мкл 2% дихлоргидрата N-нафтил-1-этилендиамина. После центрифугирования при 12000 об/мин в течение 5 минут поглощение измеряли при 540 нм. Величину реакции с добавленной EDTA вычитали из величины реакции с добавленным MgCl2 для получения активности NR.
Активность GS также измеряли способами Ferrario-Mery et al (см. выше). 20 мкл раствора фермента добавляли к 480 мкл реакционной жидкости (80 мМ глутамат, 20 мМ MgSO4, 1 мМ EDTA, 100 мМ трицин, 6 мМ гидроксиламин, 8 мМ АТФ) и реакцию смеси проводили в течение 15 минут при 30°С. Для остановки реакции добавляли 500 мкл 0,37 М FeCl3, 0,2 М трихлоруксусную кислоту и 0,67 н раствор HCl и измеряли поглощение при 540 нм.
Концентрацию белка экстрагированного раствора фермента измеряли способом Брэдфорда с применением бычьего сывороточного альбумина (BSA) в качестве стандартного белка. В результате этого показали, что трансформированный Arabidopsis thaliana имел 1,38-кратную активность NADH-GOGAT, 1,40-кратную активность NR и 1,25-кратную активность GS относительно активностей нетрансформированного Arabidopsis thaliana (таблица 3). Трансформированный картофель также имел 1,33-кратную активность NADH-GOGAT, 1,36-кратную активность NR и 1,01-кратную активность GS относительно активностей нетрансформированного картофеля (таблица 4). Эти результаты показывают, что не только трансформированный Arabidopsis thaliana и трансформированный картофель имеют более высокую эффективность поглощения азота, чем нетрансформанты, но и ферменты, принимающие участие в метаболизме азота, также активируются, и эффективность метаболизма азота также повышалась в трансформированном Arabidopsis thaliana и трансформированных картофельных растениях.
Активности ферментов в трансформированном Arabidopsis thaliana (Mtd 3), единиц/мг белка
Активности ферментов в трансформированном картофеле (Mtd 8), единиц/мг белка
Пример 9 Определение урожайности трансформированного картофеля
Изучали урожайности трансформированных картофельных растений. В определении урожайности применяли посадочный картофель трансформированных картофельных растений и нетрансформированных картофельных растений. Рост, массу клубней, число клубней и тому подобное определяли в условиях культивации с различными концентрациями азота. Устанавливали две группы - одну применяли в стандартных условиях с 3 кг почвы Power (всего 1,2 кг азота) в горшке на 8,5 литра и другую применяли в условиях с низким содержанием азота с 300 г почвы Power (всего азота 0,12 кг) в горшке № 7, причем остальное количество почвы составлял вермикулит (без питательных веществ). Картофельные растения выращивали 3 месяца после посадки и затем измеряли массы надземной части растений, массы клубней и число клубней.
В группе с 1,2 г азота трансформированные картофельные растения имели 1,12-1,13-кратную массу клубней и 1,09-1,23-кратное число клубней относительно нетрансформированных картофельных растений (таблица 5). В группе с 0,12 г азота (условия с ограниченным содержанием азота) трансформированные картофельные растения имели 1,29-1,35-кратную массу клубней и 1,24-1,56-кратное число клубней относительно нетрансформированных картофельных растений (таблица 6).
Определение урожайности трансформированных картофельных растений в группе с 1,2 г азота (n ≥ 8)
Определение урожайности трансформированных картофельных растений в группе с 0,12 г азота (n≥8)
Пример 10 Определение урожайности трансформированного Arabidopsis thaliana
Рост изучали с применением трансформированного Arabidopsis thaliana (Mtd 3). Arabidopsis thaliana высевали на среде PNS (источник азота: только нитратный азот), дополненной варьируемыми концентрациями KNO3 0,1, 0,3, 3 и 5 мМ и 1% сахарозой с применением термочашек Петри 8 см, 8 растений на чашку. Спустя 3 недели после засевания измеряли массы в надземной части растений. Данные представляют собой общие массы надземных частей всех 8 растений, представленных как среднее значение и отклонение для 3 чашек Петри в каждой группе. В результате этого показано, что трансформированный Arabidopsis thaliana растет лучше в условиях низкого содержания азота, чем растет нетрансформированный Arabidopsis thaliana, демонстрируя рост, в 1,1-1,14 раз превышающий рост нетрансформированного Arabidopsis thaliana, в виде массы в свежем виде (таблица 7).
Масса (г) в свежем виде трансформированного Arabidopsis thaliana и нетрансформированного Arabidopsis thaliana, выращенных в среде PNS (с 1% сахарозы), содержащей различные концентрации KNO3
Пример 11 Некорневая подкормка водным раствором пролина картофельных растений
Влияния некорневой подкормки раствором пролина на рост и урожайность исследовали с применением картофельных растений (сорт May Queen). Клубни картофеля помещали в горшки № 7, содержащие 1,5 кг почвы Power (Kureha Chemical). Содержание азотистого удобрения почвы Power показано в таблице 8. В нормальных условиях культивации приблизительно 5 г азотистого удобрения применяли в качестве основного удобрения, хотя в этой культивации испытание на культивацию проводили в условиях с низким содержанием азота.
Получали три группы для некорневой подкормки - группу агента распределения (Approach B1 Kao), группу мочевины (агент распределения + 0,87 мМ мочевина) и группу пролина (агент распределения + 1,75 мМ пролина) и приблизительно 20 мл раствора на растение разбрызгивали один раз в начале недели приблизительно месяц после посадки (непосредственно перед цветением). Листья подвергали опрыскиванию 6 раз (6 недель) и урожай собирали через 3 месяца после посадки.
Условия удобрения азотом для испытания культивации картофеля
Пример 12 Изменение содержания свободного пролина и 2-оксоглутарата (2-OG) в тканях листьев картофеля после некорневой подкормки пролином
После разбрызгивания на поверхности листьев картофеля водных растворов распределяющего агента, мочевины и пролина содержание свободного пролина и содержание 2-OG изменяли на протяжении времени. 3-й лист от верхушек отбирали в качестве образцов через 1, 3, 5 и 8 часов после некорневой подкормки и тщательно промывали водой в качестве образцов. После измельчения листьев жидким азотом добавляли 80% этанол, который был нагрет до 80°С и был равен 5-кратной массе листьев в свежем состоянии, их инкубировали в течение 20 минут при 80°С. После центрифугирования в течение 10 минут при 12000 об/мин супернатант переносили в отдельную пробирку. Снова добавляли 80% этанол с последующей инкубацией в течение 20 минут при 80°С и центрифугированием и супернатант собирали и экстрагировали таким же образом всего 3 раза. Экстракт в этаноле сушили вымораживанием, добавляли 300 мкл стерильной воды и 200 мкл этилового эфира и смесь хорошо перемешивали и центрифугировали в течение 10 минут при 12 об./мин. После удаления верхнего слоя, слоя этилового эфира 200 мкл водного слоя переносили в отдельную пробирку и еще раз сушили вымораживанием. После растворения в 200 мкл 0,02 н хлористоводородной кислоты его фильтровали фильтром 0,22 мкм. Образовавшийся экстракт анализировали высокоскоростным аминокислотным анализатором Хитачи (L8800). Содержание 2-OG измеряли с применением аналогичных образцов растений. Экстракцию и определение содержания 2-OG проводили по способам, аналогичным способам, описанным в примере 7.
В результате этого не наблюдали значительного различия в содержании пролина в ткани листьев среди трех групп через час после некорневой подкормки, но через 5 часов после подкормки содержание свободного пролина в ткани листьев было значительно выше в группе подкормки пролином (фиг.11). Считали, что пролин, нанесенный разбрызгиванием на поверхность листьев, включался в ткани листьев. Аналогично этому, когда измеряли содержание 2-OG в ткани листьев, не наблюдали значительного различия среди трех групп в содержании 2-OG в ткани листьев спустя 1 час после подкормки, но через 24 часа после подкормки содержание 2-OG в ткани листьев было значительно выше в группе подкормки пролином (фиг.12). Считали, что пролин, распределенный по поверхностям листьев, включался в ткани листьев и затем метаболизировался в 2-OG и вступал в цикл ТСА.
Пример 13 Определение урожайности картофельных растений после некорневой подкормкой пролином
Водные растворы распределяющего агента, мочевины и пролина наносили 6 раз с интервалами одна неделя на поверхности листьев через 1 месяц после посадки. Указанные растения культивировали в течение 3 месяцев после посадки с применением 6 горшков в каждой группе и одного стебля на растение. Клубни собирали из каждого горшка и измеряли массу клубней и число клубней на горшок.
В результате этого обнаружили, что масса клубней на горшок была 145,8 г в группе агента распределения, 140,1 г - в группе мочевины и 158,4 г - в группе пролина, таким образом, группа пролина имела массу клубней, в 1,1 раз превышающую массу клубней группы распределяющего агента. Число клубней было 8,2 в группе агента распределения, 6,8 - в группе мочевины и 8,2 - в группе пролина, таким образом не наблюдали различия в числе клубней (таблица 9). Эти результаты указывают, что масса клубней повышается некорневой подкормкой пролином.
Урожайность картофельных растений, на которые разбрызгивали пролин (n = 6)
Пример 14 Получение трансформированного Arabidopsis thaliana, в который вводили ген аспартатаминотрансферазы (ECASPC) происхождения из Е. coli
Ген ECASPC изолировали на основе данных последовательности (под номером доступа GenBank: Х03629) для гена аспартатаминотрансферазы (ECASPC), уже указанной в E. coli (нуклеотидная последовательность: SEQ ID №: 19, аминокислотная последовательность: SEQ ID №: 20). Последовательность митохондриального транзитного пептида была присоединена к этому гену, как описано в примере 1 (mtdECASPC). Район GUS трансформирующего растения Ti-плазмидного вектора рВ|121 заменили на данный ген и продукт замены трансформировали в Agrobacterium tumefaciens EHA105. Ген mtdECASPC ввели в Arabidopsis thaliana способами, описанными в примере 3, с применением этого Agrobacterium. Его затем засевали в среде, содержащей канамицин, отбирали резистентные индивиды (генерация Т1) и линию, в которой резистентность и чувствительность к канамицину были расщеплены как 3:1, получили в следующей генерации (генерация Т2). В следующей генерации были отобраны четыре линии со всеми индивидами, проявляющими резистентность к канамицину (генерация Т3) (mtdECASPC2-2, mtdECASPC6-2, mtdECASPC8-1 и mtdECASPC9-1). Введенный ген подтверждали в образовавшихся трансформантах посредством PCR генома, как в примере 4. Транскрипцию и экспрессию введенного гена также анализировали ОТ-ПЦР. РНК экстрагировали из ткани листьев Arabidopsis thaliana с применением набора Qiagen Plant Rneasy Mini-Kit. ОТ-ПЦР проводили с применением набора TaKaRa RNA-PCR и ECASPC-специфичных праймеров. Реакция состояла из 30 циклов из 1 минуты при 94°С, 1 минуты при 55°С и 2 минут при 72°С, за которой следовал электрофорез в 1% геле агарозы и окрашивание бромидом этидия.
В результате этого показано, что трансформированный Arabidopsis thaliana, который проявляет резистентность к канамицину, образует полосу такого же размера, как полоса, полученная с применением в качестве матрицы плазмидной ДНК mtdECASPC, в качестве положительного контроля (фиг.13). Эти результаты позволяют предположить, что ген mtdECASPC введен в геном трансформированного Arabidopsis thaliana, который проявлял резистентность в канамицину. mtdECASPC-специфическую полосу подтверждали также при помощи ОТ-ПЦР в трансформированном Arabidopsis thaliana (фиг.14). Поскольку эту полосу не обнаружили с нетрансформированным Arabidopsis thaliana или когда РНК применяли без обратной транскрипции в качестве матрицы, представляется, что геном не был загрязнен. То есть эти результаты позволяют предположить, что введенный ген mtdECASPC был транскрибирован в трансформированном Arabidopsis thaliana.
Пример 15 Определение урожайности трансформированного геном ECASPC Arabidopsis thaliana в условиях ограниченного содержания азота
Рост определяли, как в примере 10, с применением образовавшихся трансформантов. Применяли PNS, имеющую только нитратный азот в качестве источника азота. Концентрации KNO3 в среде были 5 мМ и 10 мМ, каждую среду дополняли 1% сахарозой. Семена засевали в стерильных условиях и 10 растениям с одинаковым ростом предоставляли возможность оставаться после 1 недели. Спустя более 2 недель (3 недели после посева) измеряли массы надземных частей 10 растений. Средние значения и отклонения для 2 растений показаны в таблице 10. В результате этого показано, что Arabidopsis thaliana, содержащий введенный ген mtdECASPC, имел значительно более высокую массу надземной части, чем нетрансформированный Arabidopsis thaliana, в который ген не был введен, в 1,18-1,28 раза больше в группе с 5 мМ азота и 1,12-1,38 раза больше в группе с 10 мМ азота.
Определение урожайности Arabidopsis thaliana с введенным геном mtdECASPC
(мг, n = 2).
Пример 16 Измерение содержания 2-OG в Arabidopsis thaliana, трансфицированном геном mtdECASPC
Содержание 2-OG измеряли с применением четырех линий трансфицированного геном mtdECASPC Arabidopsis thaliana, выбранных в примере 14. Применяли надземные части растений, выращенных в течение 2 недель при 24°С при 16-часовой продолжительности дня из семян, которые засевали в стерильных условиях в среде PNS, содержащей 5 мМ KNO3 и 1% сахарозы. 2-OG экстрагировали и его содержание измеряли способами, аналогичными способам, описанным в примере 7.
Как результат, содержание 2-OG надземной ткани трансформированного Arabidopsis thaliana с введенным геном mtdECASPC было в 1,27-1,93 раза выше, чем содержание 2-OG нетрансформированного Arabidopsis thaliana, в который ген не был введен. Результаты являются очень близкими с результатами, полученными для трансформированных растений с введенным геном GDH.
Содержание 2-OG в Arabidopsis thaliana, трансфицированном геном mtdECASPC




















| название | год | авторы | номер документа |
|---|---|---|---|
| РАСТЕНИЕ, ХАРАКТЕРИЗУЮЩЕЕСЯ УМЕНЬШЕННЫМ ПЕРИОДОМ ПОКОЯ, И СПОСОБ ПРОДУКЦИИ УКАЗАННЫХ РАСТЕНИЙ | 2005 |
|
RU2298034C2 |
| КАРТОФЕЛЬ С ПОВЫШЕННЫМ ВЫХОДОМ КРАХМАЛА В ПЕРЕСЧЕТЕ НА ОТДЕЛЬНОЕ РАСТЕНИЕ И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2006 |
|
RU2326527C2 |
| КАРТОФЕЛЬ С ПОВЫШЕННЫМ ВЫХОДОМ КРАХМАЛА В ПЕРЕСЧЕТЕ НА ОТДЕЛЬНОЕ РАСТЕНИЕ И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2004 |
|
RU2289914C2 |
| ЭНХАНСЕР ПАЛОЧКОВИДНОГО ВИРУСА САХАРНОГО ТРОСТНИКА (SCBV) И ЕГО ПРИМЕНЕНИЕ В ФУНКЦИОНАЛЬНОЙ ГЕНОМИКЕ РАСТЕНИЙ | 2013 |
|
RU2639517C2 |
| СПОСОБ ИНДУЦИРОВАНИЯ ОБРАЗОВАНИЯ ЦВЕТКОВ У РАСТЕНИЙ | 1995 |
|
RU2145636C1 |
| РАСТЕНИЯ, ХАРАКТЕРИЗУЮЩИЕСЯ ПОВЫШЕННОЙ УРОЖАЙНОСТЬЮ, И СПОСОБ ИХ ПОЛУЧЕНИЯ | 2006 |
|
RU2463351C2 |
| БЕЛКИ, СВЯЗАННЫЕ С ФОРМОЙ ЗЕРЕН И ЛИСТЬЕВ РИСА, ГЕНЫ, КОДИРУЮЩИЕ УКАЗАННЫЕ БЕЛКИ, И ИХ ПРИМЕНЕНИЯ | 2010 |
|
RU2553206C2 |
| СПОСОБ РЕГУЛИРОВАНИЯ КЛУБНЕОБРАЗОВАНИЯ И ПРОДУКТИВНОСТИ РАСТЕНИЙ КАРТОФЕЛЯ В УСЛОВИЯХ ГИДРОПОНИКИ | 2017 |
|
RU2660918C1 |
| СПОСОБ БОРЬБЫ С НЕМАТОДАМИ | 1991 |
|
RU2129373C1 |
| ТРАНСГЕННЫЕ РАСТЕНИЯ С УЛУЧШЕННЫМИ ХАРАКТЕРИСТИКАМИ РОСТА | 2009 |
|
RU2582260C2 |

Изобретение относится к генной инженерии растений. Выращивают растение с повышенным содержанием 2-оксоглутарата (2-OG). Повышенное содержание 2-OG достигается экспрессией введенного в растение гена GDH или ECASPC либо опрыскиванием листьев растения пролином, что обеспечивает культивирование растения с увеличенным ростом и/или повышенной урожайностью в условиях пониженного содержания азота. 2 н. и 8 з.п. ф-лы, 19 ил., 11 табл.
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |

