Перекрестная ссылка на родственные заявки
В настоящей заявке испрашивается приоритет предварительной заявки на патент США 60/489840, поданной 24 июля 2003, и предварительной заявки на патент США 60/520115, поданной 14 ноября 2003, каждая из которых приведена здесь в качестве ссылки.
Предшествующий уровень техники
Цитируемые в настоящей заявке работы не должны рассматриваться как прототипы заявленного изобретения.
Staphylococcus aureus представляет собой патоген, ответственный за развитие большого числа заболеваний и состояний. Примерами заболеваний и состояний, вызываемых S. aureus, являются бактериемия, инфекционный эндокардит, фолликулит, фурункулы, карбункулы, импетиго, буллезное импетиго, целлюлит, ботриомиозит, синдром токсического шока, синдром обожженной кожи, инфекции центральной нервной системы, инфекционные и воспалительные заболевания глаз, остеомиелит и другие инфекционные заболевания суставов и костей, и инфекции дыхательных путей (The Staphylococci in Human Disease, Crossley & Archer (eds.), Churchill Livingstone Inc. 1997).
Для борьбы против инфекций, вызываемых S. aureus, и их распространения могут быть применены иммунологические стратегии. Иммунологические стратегии включают пассивную и активную иммунизацию. Пассивная иммунизация предусматривает использование иммуноглобулинов, конъюгированных с S. aureus. Активная иммунизация предусматривает индукцию иммунного ответа против S. aureus.
Предполагаемые вакцины нацелены на полисахариды и полипептиды S. aureus. Нацеливание может быть достигнуто с использованием подходящих полисахаридов или полипептидов S. aureus в качестве компонентов вакцины. Примеры возможных полисахаридных компонентов вакцины включают капсульные полисахариды S. aureus типа 5 и типа 8 (Shinefield et al., N. Eng. J. Med. 346:491-496, 2002). Примеры полипептидов, которые могут быть использованы в качестве возможных компонентов вакцины, включают коллаген, адгезин, фибриноген-связывающие белки и фактор свертывания крови (Mamo et al., FEMS Immunology and Medical Microbiology 10:47-54, 1994, Nilsson et al., J. Clin. Invest. 101:2640-2649, 1998, Josefsson et al., The Journal of Infectious Diseases 184:1572-1580, 2001).
Сведения о полипептидных последовательностях S. aureus были получены из данных секвенирования генома S. aureus (Kuroda et al., Lancet 357:1225-1240, 2001, Baba et al., Lancet 359:1819-1827, 2000, Kunsch et al., европейская патентная заявка ЕР 0786519, опубликованная 30 июля 1997). Для характеристики полипептидных последовательностей, полученных на основе данных секвенирования генома, были предприняты попытки применения некоторых методов биоинформатики (Kunsch et al., Европейская патентная заявка ЕР 0786519, опубликованная 30 июля 1997).
Для идентификации генов, кодирующих возможные антигены, могут быть частично применены такие методы, как технологии молекулярного дисплея и технологии с использованием сыворотки от инфицированных пациентов (Foster et al., Международная заявка № WO 01/98499, опубликованная 27 декабря 2001, Meinke et al., Международная заявка № WO 02/059148, опубликованная 1 августа 2002).
Описание сущности изобретения
Настоящее изобретение относится к полипептидам, содержащим аминокислотную последовательность, структурно относящуюся к SEQ ID NO:1, к использованию таких полипептидов и к экспрессионным системам для получения таких полипептидов. SEQ ID NO:1 представляет собой усеченное производное полноразмерного полипептида S. aureus. Этот полноразмерный полипептид обозначен здесь как полноразмерный "ORF0657n". Было обнаружено, что полипептиды, содержащие аминокислотную последовательность SEQ ID NO:1, вызывают защитный иммунный ответ против S. aureus.
Термин "защитный" иммунитет или иммунный ответ означает определяемый уровень защиты против инфекции S. aureus. Уровень защиты может быть оценен на моделях животных, например на моделях, описанных в настоящей заявке.
Таким образом, в первом своем аспекте настоящее изобретение относится к полипептидному иммуногену, содержащему аминокислотную последовательность, которая по крайней мере на 90% идентична SEQ ID NO:1, где полипептид не содержит карбокси-концевой конец, состоящий из аминокислот 609-645 SEQ ID NO:2, и где полипептид обеспечивает защитный иммунитет против S. aureus. SEQ ID NO:2 представляет собой полноразмерный полипептид ORF0657n, где аминокислоты 609-645 составляют карбокси-концевой домен, начинающийся с мотива LPXTG (называемый здесь "сигналом сортировки белков клеточной стенки").
Термин "иммуноген" означает вещество, обладающее способностью вызывать защитный иммунитет.
Выражение "содержащий аминокислотную последовательность, которая по крайней мере на 90% идентична SEQ ID NO:1" подразумевает присутствие области, родственной SEQ ID NO:1, и возможное присутствие других полипептидных областей. Если присутствуют другие полипептидные области, то указанный полипептид не содержит карбокси-концевой мотив LPXTG, состоящий из аминокислот 609-645 SEQ ID NO:2.
В другом своем аспекте настоящее изобретение относится к иммуногену, содержащему аминокислотную последовательность, которая вызывает защитный иммунитет против S. aureus. Иммуноген содержит аминокислотную последовательность, которая по крайней мере на 90% идентична SEQ ID NO:1, и одну или несколько дополнительных областей или частей, ковалентно связанных на карбоксильном конце или амино-конце, где каждая указанная область или часть независимо выбрана из области или части, обладающих по крайней мере одним из следующих свойств: способностью усиливать иммунный ответ, способностью облегчать очистку полипептида или способностью обеспечивать его стабильность.
Термин "дополнительная область или часть" обозначает область или часть, отличную от ORF0657n-родственного полипептида, который может продуцироваться в биологическом хозяине, таком как прокариотический или эукариотический хозяин. Такой дополнительной областью или частью могут быть, например, дополнительная полипептидная область или не-пептидная область.
В другом своем аспекте, настоящее изобретение относится к композиции, способной индуцировать у пациента защитный иммунитет против S. aureus. Указанная композиция содержит фармацевтически приемлемый носитель и иммунологически эффективное количество иммуногена, обеспечивающего выработку защитного иммунитета против S. aureus.
Иммунологически эффективным количеством является количество, достаточное для выработки защитного иммунитета против инфекции, вызываемой S. aureus. Такое количество должно быть достаточным для надежного предупреждения вероятности возникновения или обострения инфекции, вызываемой S. aureus.
В другом своем аспекте, настоящее изобретение относится к нуклеиновой кислоте, содержащей рекомбинантный ген, кодирующий полипептид, который вызывает защитный иммунитет против S. aureus. Рекомбинантный ген содержит рекомбинантную нуклеиновую кислоту, кодирующую полипептид, вместе с регуляторными элементами для "правильной" транскрипции и процессинга (которыми могут быть трансляционные и посттрансляционные элементы). Указанный рекомбинантный ген может присутствовать независимо от генома хозяина, либо он может быть частью генома хозяина.
Рекомбинантная нуклеиновая кислота представляет собой нуклеиновую кислоту, которая имеет последовательность и/или форму, не встречающуюся в природе. Примеры рекомбинантной нуклеиновой кислоты включают очищенную нуклеиновую кислоту; нуклеиновую кислоту, в которой две или несколько областей нуклеиновых кислот, объединены вместе и образуют нуклеиновую кислоту, не встречающуюся в природе; и нуклеиновую кислоту, в которой отсутствует одна или несколько областей нуклеиновых кислот (например, в 3′-5′ направлении или в 5′-3′ направлении), связанных друг с другом в природе.
В другом своем аспекте настоящее изобретение относится к рекомбинантной клетке. Эта клетка содержит рекомбинантный ген, кодирующий полипептид, который вызывает защитный иммунитет против S. aureus.
В другом своем аспекте настоящее изобретение относится к способу получения полипептида, вызывающего защитный иммунитет против S. aureus. Указанный способ предусматривает культивирование рекомбинантной клетки, содержащей рекомбинантную нуклеиновую кислоту, кодирующую указанный полипептид, и очистку полипептида.
В другом своем аспекте настоящее изобретение относится к полипептиду, который вызывает защитный иммунитет у хозяина против S. aureus и который получают способом, предусматривающим стадию культивирования рекомбинантной клетки, содержащей рекомбинантную нуклеиновую кислоту, кодирующую указанный полипептид, и очистку этого полипептида. Для этих целей могут быть использованы различные клетки-хозяева. В одном из вариантов настоящего изобретения указанной клеткой-хозяином является дрожжевая клетка.
В другом своем аспекте настоящее изобретение относится к способу индукции у пациента защитного иммунного ответа против S. aureus. Указанный способ предусматривает стадию введения пациенту иммунологически эффективного количества иммуногена, вызывающего защитный иммунитет против S. aureus.
В другом своем аспекте настоящее изобретение относится к способу индукции анамнетической реакции у пациента. Указанный способ предусматривает стадию введения пациенту эффективного количества иммуногена для продукции анамнетической реакции.
В другом своем аспекте настоящее изобретение относится к нуклеиновой кислоте, кодирующей ORF0657n-родственный полипептид, который оптимизирован для экспрессии в дрожжах. Для экспрессии в дрожжах оптимизируют один или несколько кодонов.
В другом своем аспекте настоящее изобретение относится к способу получения полипептида, вызывающего защитный иммунитет против S. aureus в рекомбинантной дрожжевой клетке. Указанный способ предусматривает:
(а) стадию культивирования рекомбинантной дрожжевой клетки в условиях, благоприятных для экспрессии полипептида, где указанная рекомбинантная дрожжевая клетка содержит рекомбинантный ген, кодирующий указанный полипептид, и где указанным полипептидом является полноразмерный полипептид ORF0657n, вызывающий защитный иммунитет против инфекции, вызываемой S. aureus, или его фрагмент, содержащий аминокислотную последовательность, которая по крайней мере на 90% идентична последовательности SEQ ID NO:1; и
(b) стадию очистки указанного полипептида.
Если конкретные термины являются взаимоисключающими, то слово "или" означает одну из двух возможностей. Используемое иногда сочетание слов "и/или" означает любую или обе возможности.
Неограничивающие термины, такие как "включает" подразумевают присутствие дополнительных элементов или стадий. Используемое иногда сочетание слов "один или несколько" вместе с ограничивающим или неограничивающим условием подразумевает возможность наличия дополнительных элементов или стадий.
Если это не оговорено особо, то употребление артиклей "а" или "an" не подразумевает ограничение лишь единственным числом. Так, например, употребление слова "a cell" (клетка) не исключает множественного числа, то есть "cells" (клетки). Используемое иногда сочетание слов, такое как "один или несколько", может означать присутствие множества элементов.
Другие отличительные признаки и преимущества настоящего изобретения будут очевидны из приведенного ниже дополнительного описания, включая различные примеры. Приведенные примеры иллюстрируют различные компоненты и методику, применяемую для осуществления настоящего изобретения. Эти примеры не ограничивают объема притязаний настоящего изобретения. Исходя из описания настоящего изобретения, любой средний специалист может идентифицировать и использовать другие компоненты и методы, которые могут быть применены для практического осуществления настоящего изобретения.
Краткое описание графического материала
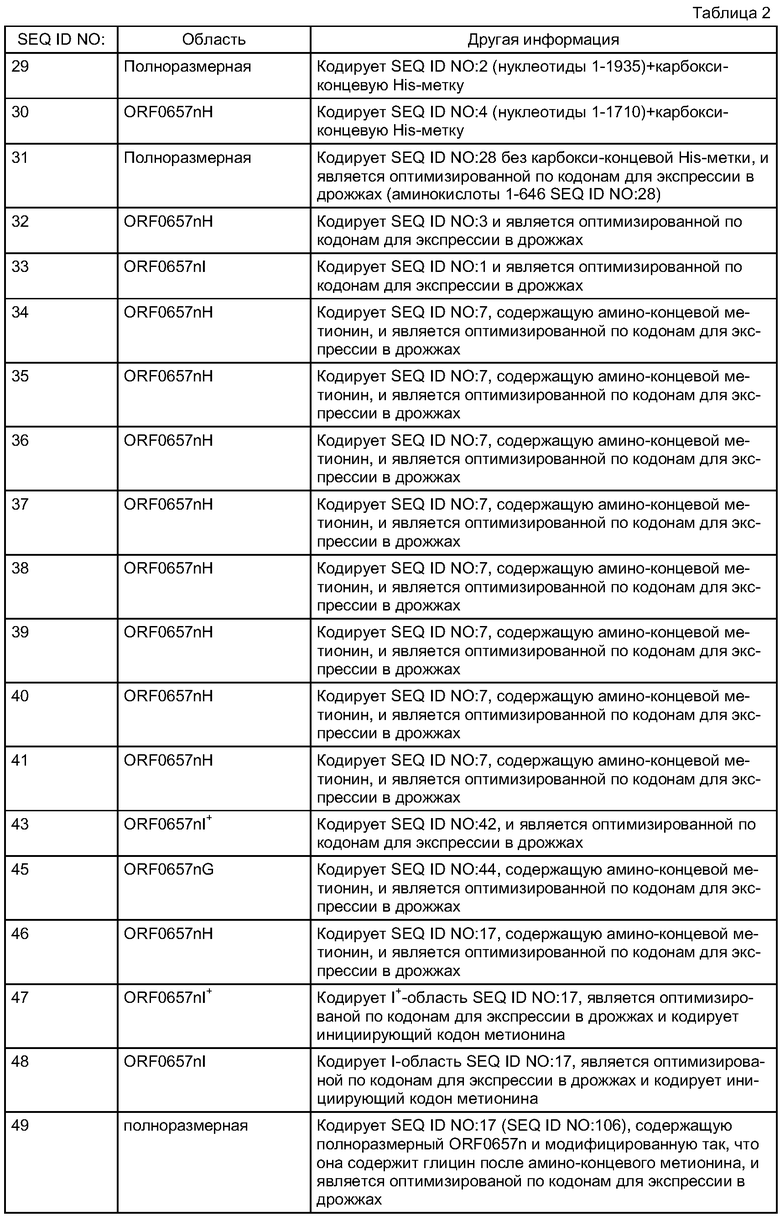
На фиг.1А, 1В, 1С и 1D схематически проиллюстрированы полипептидные области, родственные полипептиду ORF0657n и скринированные на наличие защитного ответа у животных, а также некоторые другие последовательности ORF0657n. На фиг.1А схематически проиллюстрированы протестированные полипептиды, которые, как было обнаружено, являются защитными (показаны черными прямоугольниками); протестированные полипептиды, которые, как было обнаружено, не являются защитными (показаны незаштрихованными прямоугольниками); и не протестированные полипептиды (показаны штрихом). На фиг.1В представлена полноразмерная последовательность, используемая на фиг.1А как эталонная (SEQ ID NO:2). На фиг.1С представлена последовательность SEQ ID NO:28. SEQ ID NO:28 содержит карбокси-концевую "His-метку" (LEHHHHHH; SEQ ID NO:64). Последовательность SEQ ID NO:28, содержащая карбокси-концевую His-метку, также обозначена здесь "His-tag-ORF0657n". На фиг.1D представлена последовательность ORF0657nI+.
На фиг.2А-2Е проиллюстрировано сравнение последовательностей различных ORF0657n-родственных последовательностей в области ORF0657nН. На фигурах SEQ ID NO: обозначают "ID".
На фиг.3А, 3В и 3С проиллюстрирована способность ORF0657n-родственных полипептидов, содержащих полноразмерную последовательность, а именно область ORF0657nН и область ORF0657nI, вызывать защитный иммунитет против S. aureus Becker. Эти полипептиды использовали вместе с адъювантом, а именно с гидроксифосфатом алюминия (АНР). На фиг.3А представлены результаты, полученные с использованием SEQ ID NO:28. На фиг.3В представлены результаты, полученные с использованием SEQ ID NO:4, содержащей карбокси-концевую His-метку. На фиг.3С представлены результаты, полученные с использованием SEQ ID NO:5, содержащей карбокси-концевую His-метку. Термин "карбоксильная His-метка" означает His-метку группы LEHHHHHH (SEQ ID NO:64), присутствующую на карбоксильном конце.
На фиг.4А-4Н проиллюстрирована способность ORF0657n-родственных полипептидов вызывать защитный иммунитет против заражения другими S. aureus. Полипептид SEQ ID NO:28 использовали в качестве иммуногена. На фиг.4А представлены результаты, полученные при провокационном заражении штаммом CL-10 (2,2х108 к.о.е./мл). На фиг.4В представлены результаты, полученные при провокационном заражении штаммом CL-10 (2,1х108 к.о.е./мл). На фиг.4С представлены результаты, полученные при провокационном заражении штаммом CL-13 (2,9х108 к.о.е./мл). На фиг.4D представлены результаты, полученные при провокационном заражении штаммом CL-13 (2,8х108 к.о.е./мл). На фиг.4Е представлены результаты, полученные при провокационном заражении штаммом CL-30 (3,1х108 к.о.е./мл). На фиг.4F представлены результаты, полученные при провокационном заражении штаммом CL-30 (3,0х108 к.о.е./мл). На фиг.4G представлены результаты, полученные при провокационном заражении штаммом CL-18 (1,0х108 к.о.е./мл). На фиг.4Н представлены результаты, полученные при провокационном заражении штаммом CL-21 (1,6х108 к.о.е./мл).
На фиг.5А и 5В проиллюстрированы карты экспрессионных плазмид S. cerevisiae. На фиг.5А представлена плазмидная карта вектора рGAL110. На фиг.5В представлена плазмидная карта для piUC-1, где показана последовательность, оптимизированная по кодонам для экспрессии дрожжей и клонированная под контролем промотора гена GAL1 вектора рGAL110.
На фиг.6А и 6В представлены Вестерн-блоты, иллюстрирующие внутриклеточную экспрессию полноразмерного ORF0657n, имеющего аминокислоты 1-646 SEQ ID NO:28 (SEQ ID NO:28 без карбокси-концевой His-метки). Дорожка 1: стандарты молекулярных масс; дорожка 2: рекомбинантная полноразмерная область ORF0657n (SEQ ID NO:28), продуцированная в очищенной E.coli, 100 нг; дорожки 3-6 содержат 20 мкг лизата дрожжевых клеток; дорожки 3-4: клеточные лизаты, полученные после двух дубликатных ферментаций трансформантов 1260 (фиг.6А) и 1309 (фиг.6В), содержащих только вектор рGAL110; дорожки 5-6: клеточные лизаты, полученные после двух дубликатных ферментаций трансформантов 1260 (фиг.6А) и 1309 (фиг.6В), содержащих pRUnkC-рGAL110, которые экспрессируют полноразмерную область ORF0657n (SEQ ID NO:28 без карбокси-концевой His-метки).
На фиг.7А и 7В проиллюстрированы кумасси-окрашивание ДСН-ПААГ-геля и Вестерн-блот-анализ, с помощью которых была продемонстрирована внутриклеточная экспрессии S. cerevisiae из нуклеиновой кислоты, кодирующей SEQ ID NO:3. Дорожка 1, панель А, BSA, 1,25 мкг; панель В, рекомбинантный полноразмерный ORF0657n в очищенной E.coli (SEQ ID NO:28), 100 нг; дорожка 2, клеточный лизат продуцента ORF0657nН (SEQ ID NO:4 с карбокси-концевой His-меткой) в E.coli; панель А, 1,25 мкг, панель В, 0,5 мкг. Дорожки 3-7, панели А и В содержат 1,25 и 0,5 мкг лизата дрожжевых клеток соответственно; дорожка 3, трансформант, содержащий только вектор рGAL110; дорожка 4, трансформант, содержащий полноразмерный ORF0657n (SEQ ID NO:28 без карбокси-концевой His-метки); дорожки 5, 6 и 7, трансформант 1-1, содержащий piUC-S(-), экспрессирующий область зрелого ORF0657nН (SEQ ID NO:3); дорожка 7 содержит клеточный лизат (трансформанта 1-1), который замораживали, а затем оттаивали. Дорожка 8 содержит стандарты молекулярной массы.
На фиг.8А-8U проиллюстрированы примеры различных последовательностей нуклеиновой кислоты, кодирующих ORF0657n-родственные полипептиды. На фиг.8А представлена последовательность нуклеиновой кислоты (SEQ ID NO:29), кодирующая SEQ ID NO:2 плюс карбокси-концевая His-метка. На фиг.8В представлена последовательность нуклеиновой кислоты (SEQ ID NO:30), кодирующая SEQ ID NO:4 плюс карбокси-концевая His-метка. На фиг.8С представлена последовательность нуклеиновой кислоты (SEQ ID NO:31), оптимизированная по кодонам для экспрессии в дрожжах и кодирующая SEQ ID NO:28 без карбокси-концевой His-метки. На фиг.8D представлена последовательность нуклеиновой кислоты (SEQ ID NO:32), оптимизированная по кодонам для экспрессии в дрожжах и кодирующая SEQ ID NO:3. На фиг.8Е представлена последовательность нуклеиновой кислоты (SEQ ID NO:33), оптимизированная по кодонам для экспрессии в дрожжах и кодирующая SEQ ID NO:1. На фиг.8F-8M представлены последовательности нуклеиновой кислоты (SEQ ID NO:34, 35, 36, 37, 38, 39, 40 и 41), оптимизированные по кодонам для экспрессии в дрожжах и кодирующие последовательность SEQ ID NO:7, содержащую метионин у амино-конца. На фиг.8N-8U представлены последовательности нуклеиновой кислоты (SEQ ID NO:46-53), оптимизированные по кодонам для экспрессии в дрожжах и кодирующие различные ORF0657n-родственные полипептиды на основе последовательностей SEQ ID NO:17 или SEQ ID NO:20.
На фиг.9 проиллюстрирован Вестерн-блот-анализ для сравнения внутриклеточной экспрессии ORF0657n-родственных полипептидов в S. cerevisiae. Дорожки 1 и 18: стандарты молекулярной массы. Дорожки 2 и 5: 50 и 100 нг соответственно очищенного ORF0657n, продуцированного в E.coli (SEQ ID NO:4 плюс карбокси-концевая His-метка). Дорожка 5 содержит 500 нг белка клеточных лизатов контроля, трансформированного вектором S. cerevisiae. Дорожки 7, 8 и 9 содержат 1,0, 2,0 и 4,0 мкг белка клеточных лизатов от S. cerevisiae-трансформанта, продуцирующего SEQ ID NO:28 без карбокси-концевой His-метки. Дорожки 11 и 12 содержат 50 и 100 нг соответственно белка клеточного лизата от S. cerevisiae-трансформанта, продуцирующего ORF0657nН (SEQ ID NO:3). Дорожки 14 и 15 содержат 250 и 500 нг соответственно белка клеточного лизата от S. cerevisiae-трансформанта, продуцирующего ORF0657nG (SEQ ID NO:44). Дорожка 17 содержит 250 нг белка клеточного лизата от E.coli, продуцирующей ORF0657nG (SEQ ID NO:44 плюс карбокси-концевую His-метку). Дорожки 4, 6, 10, 13 и 16 являются пустыми.
На фиг.10 представлены данные, демонстрирующие, что ORF0657n-родственные полипептиды, продуцированные в E.coli и в дрожжах, вызывают защитный иммунный ответ. Обозначение "ORF0657nН (E.coli)" соответствует последовательности SEQ ID NO:4 с карбокси-концевой His-меткой. Обозначение "ORF0657nI (E.coli)" соответствует последовательности SEQ ID NO:5 с карбокси-концевой His-меткой. Обозначение "ORF0657nС (E.coli)" соответствует последовательности SEQ ID NO:28. Обозначение "ORF0657nН (дрожжи)" соответствует последовательности SEQ ID NO:3.
На фиг.11 проиллюстрирован Вестерн-блот-анализ для сравнения внутриклеточной экспрессии ORF0657nI в S. cerevisiae. Дорожки 1 и 25, стандарты молекулярной массы. Дорожки 2, 3 и 24: 25, 50 и 100 нг соответственно очищенной области полипептида ORF0657nН (SEQ ID NO:4 с карбокси-концевой His-меткой), продуцированного в E.coli. Дорожки 4-23 содержат белок клеточных лизатов от дрожжевых трансформантов. Дорожки 13-21 представляют дубликаты клеточных лизатов, полученных из того же самого ферментированного образца, как и лизаты на дорожках 4-12. Дорожки 4 и 13 содержат 200 нг белка контрольного векторного трансформанта, содержащего рGAL110. Дорожки 5 и 14 содержат 100 нг белка от трансформанта 1-1, продуцирующего область ORF0657nН (SEQ ID NO:3). Дорожки 6 и 15 содержат 200 нг белка от трансформанта 1-1. Дорожки 7 и 16 содержат 100 нг белка, а дорожки 8 и 17 содержат 200 нг белка от трансформанта I1. Дорожки 9 и 18 содержат 100 нг белка, а дорожки 10 и 19 содержат 200 нг белка трансформанта I2. Дорожки 11 и 20 содержат 100 нг белка, а дорожки 12 и 21 содержат 200 нг белка от трансформанта I3. Дорожки 22 и 23 содержат 100 и 200 нг белка клеточного лизата, полученного ранее путем предварительной ферментации трансформанта 1-1.
На фиг.12 приводятся данные иммунизации макак-резусов. Макак-резусов иммунизировали либо продуцируемым в дрожжах ORF0657n-родственным полипептидом (ORF0657nН, SEQ ID NO:3), либо экспрессируемым в E.coli ORF0657n-родственным полипептидом (полноразмерным полипептидом ORF0657nС, SEQ ID NO:28), приготовленными в АНР или без него. Обезьянам, входящим в группу вакцинации, внутримышечно вводили 50 мкг ORF0657n-родственных полипептидов.
Подробное описание изобретения
Было обнаружено, что ORF0657n-родственные полипептиды, включая полноразмерные и более короткие производные, содержащие область ORF0657nI, вызывают у животных-моделей защитный иммунитет против S. aureus. На фиг.1А показана локализация различных ORF0657n-родственных полипептидных областей, вызывающих защитный иммунитет против инфекции, вызываемой S. aureus, а также локализация областей, которые не вызывают защитного иммунитета. На фиг.1А ORF0657n означает полноразмерную последовательность, соответствующую SEQ ID NO:2; ORF0657nI означает область, соответствующую SEQ ID NO:1 (без амино-концевого метионина), а ORF0657nН означает область, соответствующую SEQ ID NO:3 (без амино-концевого метионина).
"ORF0657n-родственный" полипептид содержит область, которая является структурно родственной полноразмерному ORF0657n или его фрагменту. ORF0657n-родственные полипептиды представляют собой полипептиды, последовательность которых по крайней мере примерно на 90% идентична последовательности соответствующей области природного ORF0657n. Сравниваемый полипептид ORF0657n, показанный на фиг.1, соответствует полипептиду ORF0657n, происходящему от COL S. aureus (SEQ ID NO:2).
Процент идентичности сравниваемой последовательности определяют путем выравнивания данной полипептидной последовательности со сравниваемой последовательностью и определения числа идентичных аминокислот. Это число делят на общее число аминокислот в сравниваемых последовательностях, а затем результат умножают на 100 и округляют до ближайшего целого числа.
Фиг.1А позволяет проиллюстрировать важное значение коровой области, содержащей аминокислотную последовательность, структурно родственную последовательности SEQ ID NO:1. SEQ ID NO:1 содержит аминокислоты 42-486 ORF0657n COL. SEQ ID NO:1 также содержит амино-концевой метионин для облегчения экспрессии. Полипептидные фрагменты, состоящие из аминокислот 461-609, аминокислот 82-486 или аминокислот 42-196 SEQ ID NO:2, не являются защитными.
В описании настоящей заявки приводятся и другие аминокислотные последовательности и последовательности нуклеиновых кислот. В таблице 1 систематизированы некоторые полипептидные последовательности с указанием области, показанной на фиг.1, и дополнительных модификаций. В таблице 2 в систематизированном виде представлены некоторые последовательности нуклеиновых кислот.

SEQ ID NO:1 Родственные полипептиды
Полипептидная область, структурно родственная SEQ ID NO:1, содержит аминокислоту по крайней мере на 90% идентичную последовательности SEQ ID NO:1. Полипептиды, содержащие область, структурно родственную SEQ ID NO:1, могут быть сконструированы в соответствии с настоящим изобретением, в результате чего могут быть получены полипептиды, вызывающие защитный ответ против S. aureus.
С использованием SEQ ID NO:1 в качестве основы и принимая во внимание аминокислотную последовательность различных природных полипептидов ORF0657n и известные свойства аминокислот, могут быть созданы различные модификации. Такими модификациями являются одно или несколько из добавлений, делеций и/или замен. Общий эффект различных модификаций может быть оценен описанными здесь методами для подтверждения способности конкретного полипептида вызывать защитный иммунитет.
Было обнаружено, что ORF0657n является достаточно консервативным по всей совокупности патологически и таксономически отличающихся клинических изолятов S. aureus. (см. ниже, пример 5). На фиг.2 проиллюстрировано сравнение различных аминокислотных последовательностей, содержащих SEQ ID NO:1. Сравнение этих последовательностей представлено для области ORF0657nН. Область ORF0657nН включает меньшую по размеру защитную область ORF0657nI.
На фиг.2 приводится сравнение последовательностей SEQ ID NO:1 и 3-27. Это сравнение иллюстрирует различие между аминокислотными последовательностями клинических изолятов S. aureus, которые могут быть использованы для внесения потенциальных модификаций в полипептиды, родственные полипептидам S. aureus, таким как SEQ ID NO:1 и 3. Кроме того, такие модификации могут быть внесены с учетом известных свойств аминокислот. Последовательности SEQ ID NO:1, 3-6 и 8-26 являются природными последовательностями, в которые, начиная с положения 3, а в некоторых последовательностях, с положений 1 и 2 со стороны амино-конца были введены метионин или метионин-глицин. Последовательности SEQ ID NO:11-26 были получены из различных клинических изолятов. SEQ ID NO:7 и 27 представляют собой варианты области ORF0657nН SEQ ID NO:4, которая содержит пять аминокислотных замен в области, расположенной за пределами коровой области SEQ ID NO:1.
При сравнении последовательностей, проводимом для поиска нужных модификаций, могут быть использованы дополнительные последовательности ORF0657n. Примеры дополнительных последовательностей областей ORF0657nН S. aureus представлены в SEQ ID NO:54-63, а полноразмерные последовательности SEQ ID NO:17 и 20 представлены в SEQ ID NO:106 и 107.
В общих чертах, для сохранения активности при заменах различных аминокислот предпочтительно, чтобы заменяемые аминокислоты имели аналогичные свойства. При замене аминокислот следует учитывать такие факторы, как размер, заряд, полярность и гидрофобность аминокислот. Влияние различных аминокислотных R-групп на свойства аминокислот хорошо известно специалистам (см., например, Ausubel, Current Protocols in Molecular Biology, John Wiley, 1987-2002, Appendix IC).
Для сохранения активности при замене аминокислот заменяемая аминокислота должна иметь одно или несколько свойств, аналогичных свойствам замененной аминокислоты, она должна иметь примерно такой же заряд, и/или размер, и/или такую же полярность, и/или гидрофобность. Так, например, хорошими кандидатами на замену, которая не приводит к изменению функции полипептида, являются замены лейцина на валин, лизина на аргинин и глутамина на аспарагин.
Модификациями, вносимыми для достижения конкретной цели, являются модификации, создаваемые для стимуляции продуцирования или повышения эффективности данного полипептида или для клонирования кодирующей нуклеиновой кислоты. Продуцирование полипептида может быть стимулировано с использованием инициирующего кодона (например, кодирующего метионин), подходящего для рекомбинантной экспрессии. Такой метионин может быть затем удален во время клеточного процессинга. Процесс клонирования может быть облегчен, например, путем введения рестрикционных сайтов, которое может быть осуществлено вместе с добавлением или заменой аминокислот.
Эффективность индуцирования иммунного ответа полипептидом может быть увеличена путем "усиления" эпитопа. Усиление эпитопа может быть осуществлено различными методами, такими как введение модификаций якорных остатков для повышения аффинности пептида по отношению к молекулам МНВ и увеличения аффинности комплекса "пептид-МНС" по отношению к Т-клеточному рецептору (Berzofsky et al., Nature Review 1:209-219, 2001).
Что касается области полипептида, родственной последовательности SEQ ID NO:1, то в различных вариантах осуществления изобретения эта область по крайней мере на 90%, по крайней мере на 94% или по крайней мере на 99% идентичная последовательности SEQ ID NO:1 и отличается от последовательности SEQ ID NO:1 модификациями, число которых может составлять 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 или до 50, либо указанная область состоит или, в основном, состоит из ORF0657nI-родственной области, выбранной из группы, состоящей из:
аминокислот 1-442 SEQ ID NO:11, 15, 16, 18 или 54;
аминокислот 1-443 SEQ ID NO:63,
аминокислот 1-444 SEQ ID NO:57 или 59,
аминокислот 1-445 SEQ ID NO:7, 8, 9, 10, 12, 13, 14, 17, 19, 20, 55, 56 или 58,
аминокислот 1-446 SEQ ID NO:23 или 24,
аминокислот 1-446 или 2-446 SEQ ID NO:1 или 3,
аминокислот 1-447 SEQ ID NO:25 или 26,
аминокислот 1-447, 2-447 или 3-447 SEQ ID NO:4, 5 или 27,
аминокислот 1-449 SEQ ID NO:61 или 62,
аминокислот 1-453 SEQ ID NO:60 и
аминокислот 1-454 SEQ ID NO:6, 21 или 22.
Термин "в основном, состоит" из указанных аминокислот означает, что помимо указанных аминокислот могут также присутствовать дополнительные аминокислоты. Такими дополнительными аминокислотами могут быть аминокислоты, находящиеся у карбокси- или у амино-конца. В различных вариантах настоящего изобретения могут присутствовать 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 дополнительных аминокислот. В предпочтительных вариантах изобретения у амино-конца присутствует метионин либо метионин-глицин.
В одном из вариантов настоящего изобретения указанный полипептид состоит из аминокислотной последовательности, которая по крайней мере на 90% идентична последовательности SEQ ID NO:42 или ее фрагменту, содержащему аминокислотную последовательность, структурно родственную последовательности SEQ ID NO:1. Что касается SEQ ID NO:42, то в других вариантах настоящего изобретения указанный полипептид по крайней мере на 94% или по крайней мере на 99% идентичен SEQ ID NO:42 и отличается от SEQ ID NO:42 модификациями, число которых может составлять 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 или до 50 или до 65, либо указанный полипептид состоит или, в основном, состоит из SEQ ID NO:42 или ORF0657nI+-родственной области, выбранной из группы, состоящей из:
аминокислот 1-477 SEQ ID NO:11, 15, 16, 18 или 54;
аминокислот 1-478 SEQ ID NO:63,
аминокислот 1-479 SEQ ID NO:57 или 59,
аминокислот 1-480 SEQ ID NO:7, 8, 9, 10, 12, 13, 14, 17, 19, 20, 55, 56 или 58,
аминокислот 1-481 SEQ ID NO:23 или 24,
аминокислот 1-481 или 2-481 SEQ ID NO:1 или 3,
аминокислот 1-482 SEQ ID NO:25 или 26,
аминокислот 1-482, 2-482 или 3-482 SEQ ID NO:4, 5 или 27,
аминокислот 1-484 SEQ ID NO:61 или 62,
аминокислот 1-488 SEQ ID NO:60 и
аминокислот 1-489 SEQ ID NO:6, 21 или 22.
В другом варианте осуществления настоящего изобретения указанный полипептид состоит из аминокислотной последовательности, которая по крайней мере на 90% идентична последовательности SEQ ID NO:3 или ее фрагменту, содержащему аминокислотную последовательность, структурно родственную последовательности SEQ ID NO:1. Что касается SEQ ID NO:3, то в других вариантах настоящего изобретения указанный полипептид по крайней мере на 94% или по крайней мере на 99% идентичен SEQ ID NO:3; отличается от SEQ ID NO:3 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, или 25 модификациями, или 50 модификациями, или 65 модификациями либо состоит или, в основном, состоит из аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 3, 4, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 54, 55, 56, 57, 58, 59, 60, 61, 62 и 63.
В другом варианте изобретения указанный полипептид состоит или, в основном, состоит из полипептида SEQ ID NO:2, модифицированного путем включения глицина после инициирующего метионина, или из указанного полипептида без инициирующего метионина.
В другом варианте изобретения таким полипептидом является очищенный полипептид. Термин "очищенный полипептид" означает, что этот полипептид присутствует в окружении, не содержащем одного или нескольких других полипептидов, с которыми он обычно ассоциирован в природе, и/или содержащем другие полипептиды в количестве, составляющем по крайней мере примерно 10% от общего присутствующего белка. В других вариантах изобретения очищенный полипептид составляет по крайней мере примерно 50%, по крайней мере примерно 75% или по крайней мере примерно 95% от общего белка, присутствующего в образце или в препарате.
В другом варианте изобретения указанный полипептид является "в основном, очищенным". "В основном, очищенным" является полипептид, который присутствует в окружении, не содержащем всех или большинства других полипептидов, с которыми он обычно ассоциирован в природе. Так, например, в основном, очищенный полипептид S. aureus присутствует в окружении, не содержащем всех или большинства других полипептидов S. aureus. Таким окружением может быть, например, образец или препарат.
Термин "очищенный" или "в основном, очищенный" не означает, что данный полипептид должен быть подвергнут любой очистке; так, например, он может представлять собой химически синтезированный полипептид, который не был очищен.
Стабильность полипептида может быть увеличена путем модификации карбоксильного или амино-конца полипептида. Примерами возможных модификаций является введение амино-концевых защитных групп, таких как ацетил, пропил, сукцинил, бензил, бензилоксикарбонил или трет-бутилоксикарбонил, и карбокси-концевых защитных групп, таких как амид, метиламид и этиламид.
В одном из вариантов настоящего изобретения защитный полипептид является частью иммуногена, состоящего из полипептида и одной или нескольких дополнительных областей или частей, ковалентно присоединенных к полипептиду у карбоксильного конца или у амино-конца. Каждая область или часть должна быть независимо выбрана из области или части, обладающей по крайней мере одним из нижеследующих свойств: способности усиливать иммунный ответ, способности облегчать очистку или способности повышать стабильность полипептидов. Стабильность полипептида может быть увеличена, например, с использованием таких групп, как полиэтиленгликоль, который может присутствовать у амино-конца или у карбоксильного конца.
Очистка полипептида может быть облегчена путем присоединения к карбоксильному концу или к амино-концу группы, способствующей облегчению очистки. Примерами групп, которые могут быть использованы для облегчения очистки, являются полипептиды, представляющие собой аффинные метки. Примерами таких аффинных меток являются метка из шести гистидинов, trpE, глутатион и белок, связывающийся с мальтозой.
Способность полипептида продуцировать иммунный ответ может быть усилена с использованием групп, которые обычно стимулируют иммунный ответ. Примерами групп, которые могут быть присоединены к полипептиду для усиления иммунного ответа против указанного полипептида, являются такие цитокины, как IL-2 (Buchan et al., 2000, Molecular Immunology 37:545-552).
Продуцирование полипептидов
Полипептиды могут быть получены стандартными методами, включая методы химического синтеза полипептидов и методы их очистки из клетки, продуцирующей эти полипептиды. Методы химического синтеза полипептидов хорошо известны специалистам (см., например, Vincent, Peptide and Protein Drug Delivery, New York, N.Y., Decker, 1990).
Способы очистки полипептида из клетки проиллюстрированы в примерах, описанных ниже. Другие примеры методов очистки хорошо известны специалистам (см., например, Ausubel, Current Protocols in Molecular Biology, John Wiley, 1987-2002).
Выделение полипептидов из клетки может быть осуществлено с использованием техники рекомбинантных нуклеиновых кислот для продуцирования полипептидов. Техника рекомбинантных нуклеиновых кислот, применяемая для продуцирования полипептида, предусматривает введение в клетку или продуцирование в этой клетке рекомбинантного гена, кодирующего полипептид, и экспрессию данного полипептида.
Рекомбинантный ген содержит нуклеиновую кислоту, кодирующую полипептид вместе с регуляторными элементами для экспрессии полипептида. Рекомбинантный ген может присутствовать в клеточном геноме, либо он может быть частью экспрессирующего вектора.
Регуляторными элементами, которые могут присутствовать как часть рекомбинантного гена, являются элементы, обычно ассоциированные с последовательностью, кодирующей данный полипептид, и экзогенные регуляторные элементы, которые обычно не ассоциированы с последовательностью, кодирующей этот полипептид. Экзогенные регуляторные элементы, такие как экзогенный промотор, могут быть использованы для экспрессии рекомбинантного гена в конкретном хозяине или для повышения уровня его экспрессии. В общих чертах, регуляторными элементами, которые присутствуют в рекомбинантном гене, являются транскрипционный промотор, сайт связывания с рибосомой, терминатор и необязательно, присутствующий оператор. Предпочтительным элементом процессинга в эукариотических клетках является сигнал полиаденилирования.
Экспрессия рекомбинантного гена в клетке может быть стимулирована с использованием экспрессирующего вектора. Предпочтительно, чтобы экспрессирующий вектор, помимо рекомбинантного гена, также содержал сайт инициации автономной репликации в клетке-хозяине, селективный маркер, ограниченное число подходящих рестрикционных сайтов и, по возможности, большое число копий. Примерами экспрессирующих векторов являются клонирующие векторы, модифицированные клонирующие векторы, конкретно сконструированные плазмиды и вирусы.
Из-за вырожденности генетического кода для кодирования конкретного полипептида может быть использовано большое число различных кодирующих последовательностей нуклеиновой кислоты. Вырожденность генетического кода обусловлена тем, что почти все аминокислоты кодируются различными комбинациями нуклеотидных триплетов или "кодонов". Аминокислоты кодируются следующими кодонами:
А=Ala=аланин: кодоны GCA, GCC, GCG, GCU
С=Cys=цистеин: кодоны UGC, UGU
D=Asp=аспарагиновая кислота: кодоны GAC, GAU
Е=Glu=глутаминовая кислота: кодоны GAA, GAG
F=Phe=фенилаланин: кодоны UUC, UUU
G=Gly=глицин: кодоны GGA, GGC, GGG, GGU
H=His=гистидин: кодоны CAC, CAU
I=Ile=изолейцин: кодоны AUA, AUC, AUU
К=Lys=лизин: кодоны ААА, AAG
L=Leu=лейцин: кодоны UUA, UUG, CUA, CUC, CUG, CUU
М=Met=метионин: кодон AUG,
N=Asn=аспарагин: кодоны AAC, AAU
Р=Pro=пролин: кодоны CCA, CCC, CCG, CCU
Q=Gln=глутамин: кодоны CAA, CAG
R=Arg=аргинин: кодоны AGC, AGG, CGA, CGC, CGG, CGU
S=Ser=серин: кодоны AGC, AGU, UCA, UCC, UCG, UCU
Т=Thr=треонин: кодоны ACA, ACC, ACG, ACU
V=Val=валин: кодоны GUA, GUC, GUG, GUU
W=Trp=триптофан: кодон UGG
Y=Tyr=тирозин: кодоны UAC, UAU
Подходящими клетками для рекомбинантной экспрессии нуклеиновой кислоты, кодирующей ORF0657n-родственные полипептиды, являются прокариоты и эукариоты. Примерами прокариотических клеток являются E.coli, члены рода Staphylococcus, такие как S. aureus; члены рода Lactobacillus, такие как L.plantarum; члены рода Lactococcus, такие как L. lactis; и члены роды Bacillus, такие как B. subtilis. Примерами эукариотических клеток являются клетки млекопитающих, клетки насекомых, дрожжевые клетки, такие как члены рода Saccharomyces (например, S. cerevisiae); члены рода Pichia (например, P. pastoris), члены рода Hansenula (например, H. polymorpha), члены рода Kluyveromyces (например, K. lactis или K.fragilis) и члены Schizosaccharomyces (например, S. pombe).
Методы продуцирования рекомбинантного гена, введения его в клетку и его экспрессии в клетке хорошо известны специалистам. Примеры таких методов описаны в руководствах, например в руководстве Ausubel, Current Protocols in Molecular Biology, John Wiley, 1987-2002 и Sambrook et al., Molecular Cloning, A Laboratory Manual, 2nd Edition, Cold Spring Harbor Laboratory Press, 1989.
Если это необходимо, то экспрессия в конкретном хозяине может быть усилена путем оптимизации кодонов. Оптимизация кодонов предусматривает использование более предпочтительных кодонов. Методы оптимизации кодонов для экспрессии в различных хозяевах хорошо известны специалистам.
ORF0657n-родственные полипептиды могут содержать посттрансляционные модификации, введенные, например, путем N-гликозилирования, О-гликозилирования или ацетилирования. Термины "полипептид" или "аминокислотная последовательность" полипептида означают полипептиды, содержащие одну или несколько аминокислот и имеющие структуру, образованную посттрансляционной модификацией в клетке-хозяине, такой как хозяйская клетка млекопитающего, насекомого или дрожжей.
Посттрансляционные модификации могут быть введены химическим методом или с использованием подходящих хозяев. Так, например, в S. cerevisiae природа предпоследней аминокислоты, очевидно, определяется независимо от того, отсутствует ли N-концевой метионин или нет. Кроме того, природа предпоследней аминокислоты также определяется независимо от того, является ли N-концевая аминокислота Nα-ацетилированной или нет (Huang et al., Biochemistry 26:8242-8246, 1987). Другим примером является полипептид, который может секретироваться благодаря присутствию секреторной лидерной последовательности (например, сигнального пептида), где указанный полипептид является модифицированным путем N-или О-гликозилирования (Kukuruzinska et al., Ann. Rev. Biochem. 56:915-944, 1987).
Экспрессия в дрожжах
Экспрессию ORF0657n-родственных полипептидов предпочтительно осуществляют в дрожжах с использованием кодирующей нуклеиновой кислоты, содержащей кодоны, оптимизированные для экспрессии в дрожжах. Экспрессия в дрожжах может быть осуществлена с использованием рекомбинантного гена, кодирующего ORF0657n-родственный полипептид и регуляторные области для экспрессии в дрожжах. В зависимости от используемой экспрессионной системы продуцируемый белок может оставаться либо внутри клетки или он может секретироваться из клетки.
При этом предпочтительно, чтобы промотором для экспрессии рекомбинантного гена был индуцибельный промотор, такой как промотор, происходящий от генного кластера дрожжевой галактозы (промотор GAL, такой как GAL1, GAL7, GAL10, МЕL1), или промотор кислой фосфатазы РН05, или промотор алкоголь-дегидрогеназы II ADH2, или регулируемый медью промотор металлотионеина CUP1. Примерами "конститутивных" промоторов, которые могут быть использованы, являются промоторы GAP (TDH), PGK или TP1 (Romanos et al., YEAST 8:423-488, 1992).
Дрожжевые клетки-хозяева, используемые для рекомбинантной экспрессии, могут быть выбраны или сконструированы так, чтобы они стимулировали экспрессию рекомбинантного гена. Обычно желательными являются такие мутации, как mnn9, prb1 и/или pep4. Для усиления экспрессии от промоторов GAL может быть достигнута сверхэкспрессия фактора транскрипции GAL4 (Hopper et al., патент США № 5068185).
Оптимизацию кодонов для конкретного хозяина осуществляют путем замены кодонов, имеющих низкую или умеренную частоту встречаемости в данном хозяине, кодонами, имеющими высокий уровень экспрессии в данном хозяине. Процент оптимальных кодонов, присутствующих в кодирующей последовательности, может варьироваться. В других вариантах осуществления изобретения число оптимальных кодонов (включая первоначально присутствующие кодоны и введенные кодоны) составляет по крайней мере 50%, по крайней мере 75%, по крайней мере 95% или 100% от общего числа кодонов.
Оптимизация кодонов может быть осуществлена следующим образом.
1. Для данного кодона сравнивают частоту встречаемости кодона дикого типа с частотой встречаемости всех кодонов в генах дрожжей.
2. Если данный кодон не является одним из кодонов, которые наиболее часто встречаются в дрожжах, то его заменяют оптимальным кодоном, имеющим высокий уровень экспрессии в дрожжевых клетках.
3. Повторяют стадии (2) и (2) для других кодонов до тех пор, пока не будет достигнут желаемый уровень оптимизации кодонов.
4. Новую кодирующую последовательность исследуют на наличие нежелательных генерированных последовательностей, таких как нежелательные рестрикционные сайты, сайты сплайсинга, промоторы, нежелательные палиндромные или повторяющиеся последовательности, последовательности терминации транскрипции, и на высокую частоту встречаемости GC. Удаление нежелательных последовательностей осуществляют с использованием альтернативного кодона.
Встречаемость альтернативных кодонов описана Lathe J. Molec. Biol. 183:1-12, 1985. Частота встречаемости кодонов в различных дрожжевых хозяевах хорошо известна специалистам. Так, например, в работе Sharp et al., Yeast 7:657-678, 1991, описана частота встречаемости синонимичных кодонов в Saccharomyces cerevisiae.
На фиг.8С-8М представлены последовательности нуклеиновых кислот, оптимизированные для экспрессии в дрожжах. На фиг.8С представлена оптимизированная для дрожжей последовательность нуклеиновой кислоты (SEQ ID NO:31), кодирующая SEQ ID NO:28 без His-метки на карбоксильном конце. На фиг.8D представлена оптимизированная для дрожжей последовательность нуклеиновой кислоты (SEQ ID NO:32), кодирующая SEQ ID NO:3. На фиг.8E представлена оптимизированная для дрожжей последовательность нуклеиновой кислоты (SEQ ID NO:33), кодирующая SEQ ID NO:1. На фиг.8F-8М представлены оптимизированные для дрожжей последовательности нуклеиновых кислот (SEQ ID NO:34-41), кодирующие SEQ ID NO:7, содержащую амино-концевой метионин. На фиг.8N-8U представлены оптимизированные для дрожжей последовательности нуклеиновых кислот (SEQ ID NO:46-53), кодирующие различные ORF0657n-родственные полипептиды, полученные на основе SEQ ID NO:17 или SEQ ID NO:20.
Экспрессия в дрожжах может быть достигнута с использованием последовательностей, оптимизированных или не оптимизированных для экспрессии в дрожжах (например, нуклеотид 1-1935, или 124-1458 SEQ ID NO:29, или нуклеотиды 1-1710 SEQ ID NO:30). Методы экспрессии в дрожжах с использованием оптимизированных или неоптимизированных последовательностей описаны в нижепривиденных примерах.
ORF0657n представляет собой поверхностный белок, содержащий С-концевой сигнал сортировки белков клеточной стенки, состоящий из 36 аминокислот и имеющий консервативный мотив "LPXTG" (Schneedwind et al., 1993, EMBO, 12:4803-4811, 1993). Белки, содержащие сигнал сортировки белков клеточной стенки, связаны с оболочкой клеточной стенки посредством механизма транспептидизации, катализируемого мембрано-связанным белком, сортазой (Mazmanian et al., Science 299:906-909, 2001). Для связывания поверхностный белок должен также содержать N-концевой сигнальный пептид для направления на секреторный путь. В этом секреторном пути сигнальный пептид удаляется, и сигнал сортировки белков клеточной стенки способствует прохождению белком этого секреторного пути. Затем сортаза расщепляет последовательность между треонином и глицином мотива LPXTG и катализирует образование амидной связи между карбоксильной группой треонина и аминогруппой пептидогликановых поперечных мостиков.
Было обнаружено, что при удалении последовательности сортировки белков клеточной стенки экспрессия в дрожжах значительно усиливается. В других вариантах осуществления изобретения полипептид-кодирующие конструкции не содержат последовательности сортировки белков клеточной стенки, а более предпочтительно они не содержат функциональной последовательности сортировки белков клеточной стенки и последовательности, кодирующей сигнальный пептид. Соответствующие ORF0657n-родственные полипептиды не содержат функциональной последовательности сортировки белков клеточной стенки или обеих последовательностей сортировки белков клеточной стенки и последовательности сигнального пептида.
В других вариантах осуществления изобретения в полипептиде или в нуклеиновой кислоте, кодирующей этот полипептид, отсутствует по крайней мере почти вся последовательность сортировки белков клеточной стенки или обе последовательности сортировки белков клеточной стенки и сигнального пептида. В других вариантах изобретения уровень экспрессии белка повышается по крайней мере в 10 раз, по крайней мере примерно в 15 раз или по крайней мере в 20 раз после удаления по крайней мере почти всей последовательности сортировки белков клеточной стенки или обеих последовательностей сортировки белков клеточной стенки и сигнального пептида. Более предпочтительно, чтобы кодирующая конструкция также содержала один или несколько кодонов, оптимизированных для экспрессии в дрожжах (например, в S. cerevisiae).
Примеры приблизительных областей для последовательностей сортировки белков клеточной стенки ORF0657n и последовательностей сигнального пептида могут быть проиллюстрированы для SEQ ID NO:2. Аминокислотная область 1-42 содержит последовательность сигнального пептида. Аминокислотная область 609-645 содержит последовательность сортировки белков клеточной стенки.
Адъюванты
Адъюванты представляют собой вещества, которые усиливают продуцирование иммуногеном иммунного ответа. Адъюванты могут функционировать в соответствии с одним или несколькими различными механизмами, такими как увеличение биологического или иммунологического времени полужизни; стимуляция доставки антигена к антиген-презентирующим клеткам; стимуляция процессинга антигена и его презентации антиген-презентирующими клетками и стимуляция продуцирования иммуномодуляторных цитокинов (Vogel, Clinical Infectious Diseases 30(suppl.3):S266-270, 2000).
Для стимуляции продуцирования иммунного ответа могут быть использованы адъюванты различных типов. Примерами конкретных адъювантов являются гидроксид алюминия, фосфат алюминия или другие соли алюминия, фосфат кальция, ДНК-мотив CpG, монофосфорил-липид А, холерный токсин, термолабильный токсин E.coli, коклюшный токсин, мурамиловый дипептид, неполный адъювант Фрейнда, MF59, SAF, иммуностимулирующие комплексы, липосомы, биоразлагаемые микросферы, сапонины, неионогенные блок-сополимеры, аналоги мирамилового пептида, полифосфацен, синтетические полинуклеотиды, IFNγ, IL-2, IL-12 и ISCOMS (Vogel Clinical Infectious Diseases 30(suppl.3):S266-270, 2000, Klein et al., Journal of Pharmaceutical Sciences 89:311-321, 2000, Rimmelzwaan et al., Vaccine 19:1180-1187, 2001, Kersten Vaccine 21:915-920, 2003, O'Hagen Curr.Drug.Target.Infect.Disord., 1:273-286, 2001).
Пациенты, нуждающиеся в индуцировании защитного иммунитета
Термин "пациент" означает млекопитающее, которое может быть инфицировано S. aureus. Этот пациент может быть подвергнут профилактическому или терапевтическому лечению. При профилактическом лечении происходит индукция защитного иммунитета у данного пациента для снижения вероятности возникновения или обострения инфекции S. aureus. Терапевтическое лечение может быть осуществлено для ослабления тяжести инфекции S. aureus.
Профилактическое лечение может быть проведено с использованием вакцины, содержащей описанный здесь иммуноген. Предпочтительно, чтобы такое лечение было проведено человеку. Вакцины могут быть введены всему населению или конкретным индивидуумам с повышенным риском инфицирования S. aureus.
Индивидуумами с повышенным риском инфицирования S. aureus являются медицинские работники; госпитализированные больные; пациенты с ослабленной иммунной системой; пациенты, перенесшие хирургическую операцию; пациенты, которым был введен экзогенный имплантат, такой как катетер или сосудистое устройство; пациенты, подвергаемые терапии, приводящей к ослаблению иммунитета; пациенты с ожогами или ранами и пациенты, профессии которых связаны с повышенным риском получения ожогов или ран (The Staphylococci in Human Disease, Crossley & Archer (ed.), Churchill Livingstone Inc. 1997).
Пациентами-животными, которые могут быть инфицированы S. aureus, являются коровы, свиньи, овцы, козы, кролики, лошади, собаки, кошки и мыши. Такое лечение пациентов-животных и последующая оценка эффективности данного лечения могут быть проведены в целях защиты от инфекций домашних питомцев и домашнего скота.
Анамнестическая реакция
Было обнаружено, ORF0657n-родственные полипептиды продуцируют быстрый и эффективный иммунный ответ у макак резус после введения одной дозы (см. ниже пример 17). Наблюдаемый ответ соответствовал анамнестической реакции.
Продуцирование анамнестической реакции имеет значительные преимущества, такие как способность индуцировать эффективный иммунитет при введении одной дозы и способность индуцировать эффективный иммунитет за короткий промежуток времени. В других вариантах изобретения анамнестическая реакция приводит по крайней мере к 3-кратному, по крайней мере к 5-кратному или по крайней мере к 6-кратному увеличению геометрически средних титров по сравнению с исходными титрами, и такое многократное увеличение титров продуцируется в течение 3, 5, 7, 9, 14 или 21 дней.
Возможность быстрого генерирования эффективного иммунного ответа позволяет снизить расходы на вакцинацию по сравнению с вакцинацией путем введения нескольких доз и позволяет проводить вакцинацию пациентов с повышенным риском инфицирования S. aureus. Индивидуумами с повышенным риском инфицирования S. aureus являются медицинские работники; госпитализированные больные; пациенты с ослабленной иммунной системой; пациенты, перенесшие хирургическую операцию; пациенты, которым был введен экзогенный имплантат, такой как катетер или сосудистое устройство; пациенты, подвергнутые терапии, приводящей к ослаблению иммунитета; пациенты с ожогами или ранами и пациенты, профессии которых связаны с повышенным риском получения ожогов или ран (The Staphylococci in Human Disease, Crossley & Archer (ed.), Churchill Livingstone Inc. 1997). В других вариантах осуществления изобретения, пациентов подвергают немедленной вакцинации или в дни 3, 5, 7, 9, 14, или 21 после проведения медицинской процедуры.
Комбинированная вакцина
ORF0657n-родственные полипептиды, обеспечивающие защитный иммунитет, могут быть использованы отдельно или в комбинации с другими иммуногенами в целях продуцирования иммунного ответа. Такими дополнительными иммуногенами, которые могут быть использованы в этих целях, являются один или несколько дополнительных иммуногенов S. aureus, которые упоминались выше в разделе "Предшествующий уровень техники" настоящего описания; один или несколько иммуногенов, вызывающих иммунный ответ против одного или нескольких микроорганизмов Staphylococcus, таких как S. epidermidis, S. haemolyticus, S. warneri или S.lugunensis; и один или несколько иммуногенов, вызывающих иммунный ответ против других инфекционных микроорганизмов.
Системы животных-моделей
Для оценки эффективности полипептида в продуцировании защитного иммунного ответа против S. aureus используют систему животных-моделей. Индукция защитного ответа у животного-модели связана с двумя проблемами: (1) для преодоления врожденного иммунитета необходимо введение очень высокой провокационной дозы и (2) время наступления гибели животных слишком мало для детекции защитного ответа. Более конкретно, мыши погибали от инфекции в течение 24 часов после бактериального заражения, и этого периода времени было недостаточно для индукции специфических иммунных ответов для излечения от инфекции. При снижении дозы контрольные и иммунизованные мыши выживали после инфицирования.
Для решения этих проблем были использованы модели, которые имели медленную кинетику летальности и которые были заражены микроорганизмом S. aureus, полученным из клеток в стационарной фазе роста, соответствующим образом титрованным и введенным внутривенно. Такая медленная кинетика летальности дает достаточное время для обеспечения специфической иммунной защиты от бактериальной инфекции (например, 10 дней вместо 24 часов).
Клетки S. aureus в стационарной фазе роста могут быть получены из клеток, культивированных на твердой среде. Они могут быть также получены из жидкости, однако результаты, полученные при использовании клеток, культивированных на твердой среде, являются более воспроизводимыми. Клетки удобнее всего культивировать в течение ночи на твердой среде. Так, например, клетки S. aureus могут быть выращены примерно в течение 18-24 часов в условиях, при которых время их удвоения составляет примерно 20-30 минут.
Стафилококки (Staphylococcus) могут быть выделены из твердой или жидкой среды стандартными методами, обеспечивающими их иммуногенность. Выделенные стафилококки могут храниться, например, при -70°С в виде промытой суспензии высокой плотности (>109 колониеобразующих единиц (к.о.е.)/мл) в забуференном фосфатом физиологическом растворе, содержащем глицерин.
Титр заражения стафилококками должен быть таким, чтобы он приводил к гибели примерно 80-90% животных-моделей за период времени примерно 7-10 дней, начиная с первого или второго дня после заражения. Для мониторинга иммуногенности инокулята Staphylococcus, находящегося на хранении, для животных-моделей могут быть проведены эксперименты по титрованию. Такие эксперименты по титрованию могут быть проведены примерно за одну или две недели до проведения эксперимента по инокуляции.
Начальные титры для экспериментов по титрованию могут быть определены исходя из предыдущих экспериментов. Для заражения животных моделей штаммом Becker S. aureus подходящий титр заражения должен составлять в пределах 5×108-8×108 к.о.е./мл.
Введение
Иммуногены могут быть приготовлены и введены пациенту в соответствии с описанной здесь методикой и в соответствии с методикой, хорошо известной специалистам. Методика введения фармацевтических препаратов в общих чертах описана, например, в руководствах: Vaccines Eds. Plotkin and Orenstein, W.B. Sanders Company, 1999; Remington's Pharmaceutical Sciences, 20th Edition, Ed.Gennaro, Mack Publishing. 2000; и Modern Pharmaceutics 2nd Edition, Eds. Banker and Rhodes, Marcel Dekker, Inc., 1990, каждое из которых вводится в настоящее описание посредством ссылки.
Для лучшего хранения и введения иммуногена пациенту обычно используют фармацевтически приемлемые носители. Фармацевтически приемлемые носители могут содержать различные компоненты, такие как буфер, стерильная вода для инъекций, нормальный физиологический раствор или забуференный фосфатом физиологический раствор, сахароза, гистидин, соли и полисорбат.
Иммуноген может быть введен различными путями, например подкожно, внутримышечно или через слизистую. Подкожное и внутримышечное введение может быть осуществлено, например, с помощью иглы или с помощью безыгольного шприца.
Подходящую схему введения доз определяют предпочтительно с учетом хорошо известных факторов, включая возраст, вес, пол и клиническое состояние пациента, способ введения, желаемый эффект и конкретно вводимое соединение. Иммуноген может быть введен в формате вакцинации многократными дозами. Доза общего полипептида должна составлять предположительно в пределах от 1,0 мкг до 1,0 мг. В некоторых вариантах изобретения эта доза составляет от 0,01 мг до 1,0 мг и 0,1 мг до 1,0 мг.
Время введения доз зависит от факторов, хорошо известных специалистам. Если необходимо, то после начального введения могут быть последовательно введены одна или несколько бустер-доз для поддержания или увеличения титров антител. Примером схемы введения доз может служить следующий режим: введение в 1-й день; затем введение через 1 месяц и введение третьей дозы - через 4, 6 или 12 месяцев и, если необходимо, введение дополнительных бустер-доз через длительные промежутки времени.
Генерирование антител
ORF0657n-родственный полипептид, способный индуцировать защитный иммунитет, может быть использован для продуцирования антител и фрагментов антител, которые связываются с данным полипептидом или с S. aureus. Такие антитела или их фрагменты используются в различных целях, включая очистку полипептида, идентификацию S. aureus либо лечение или профилактика инфекций, вызываемых S. aureus.
Антитела могут быть поликлональными или моноклональными. Способы продуцирования и применения антител хорошо известны специалистам. Примеры таких способов описаны у Ausubel, Current Protocols in Molecular Biology, John Wiley, 1987-1998, Harlow et al., Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory Press, 1988, и Kohler et al., Nature 256: 495-497, 1975.
Примеры
Ниже приводятся примеры, которые дополнительно иллюстрируют отличительные признаки настоящего изобретения. В этих примерах также проиллюстрирована методика, подходящая для осуществления настоящего изобретения. Эти примеры не ограничивают заявленное изобретение.
Пример 1: Использование области ORF0657n для индукции защитного иммунитета
В этом примере проиллюстрирована способность полноразмерной области ORF0657n вызывать защитный иммунитет.
Клонирование и экспрессия ORF0657n
ПЦР-праймеры конструировали для амплификации гена, кодирующего ORF0657n и простирающегося от первого аспарагинового остатка и до концевого аспарагинового остатка, расположенного перед стоп-кодоном. Эти ПЦР-праймеры также содержали дополнительные NcoI-сайт (прямой праймер) и XhoI-сайт (обратный праймер) для облегчения клонирования в экспрессирующий вектор.
Кодируемый белок конструировали так, чтобы он экспрессировался из вектора рЕТ28 с концевыми остатками His и стоп-кодоном, кодируемым этим вектором. Кроме того, в этот белок после инициирующего метионина был добавлен глициновый остаток. Полученная амплифицированная ДНК-последовательность кодирует ORF0657n с His-меткой на карбокси-конце (SEQ ID NO:28).
ПЦР-амплифицированные последовательности лигировали в вектор рЕТ28 (Novagen) с использованием NcoI/XhoI-сайтов, которые были сконструированы в ПЦР-праймерах и введены в DH5α E.coli (Invitrogen) посредством теплового шока. Смесь для трансформации культивировали при 37оС в течение ночи на планшетах с агаром Лурия-Бертани (LB), содержащим с 100 мкг/мл канамицина. Колонии отбирали, культивировали в LB с 30 мкг/мл канамицина и приготавливали мини-препараты ДНК (Promega), после чего определяли целостность вставки с помощью гидролиза рестриктирующими ферментами и ПЦР. Четыре мини-препарата, содержащие вставку нужного размера, секвенировали с использованием праймеров М13F (SEQ ID NO:65), М13R (SEQ ID NO:66), ORF0657nF (SEQ ID NO:67) и ORF0657nR (SEQ ID NO:68). Затем из нужной последовательности отбирали клон, не содержащий ДНК-модификаций.
Клетки HMS174 (DЕ3) E.coli (Novagen) трансформировали и культивировали на планшетах со средой LB, содержащей канамицин (30 мкг/мл), и 3 колонии (UnkC-1, UnkC-2 и UnkC-3) отбирали для тестирования на экспрессию. Жидкие LB-культуры (канамицин) инкубировали при 37оС, при 250 об/мин, до тех пор, пока оптическая плотность А600 не достигала 0,6-1,0, после чего эти культуры индуцировали добавлением IPTG до конечной концентрации 1 мМ, а затем снова инкубировали в течение трех часов. Культуры собирали центрифугированием при 5000×g в течение 5 минут при 4°С. Клетки ресуспендировали в 500 мкл буфера для лизиса (Bug Buster, с ингибиторами протеазы, Novagen). Затем добавляли равный объем буфера для загрузки (в который был добавлен β-меркаптоэтанол до конечного объема 5%) и образцы нагревали при 70°С в течение 5 минут. Экстракты подвергали электрофорезу в 4-20% трис-глициновом геле Novex и белки визуализировали (путем окрашивания кумасси синим), а затем подвергали блоттингу на нитроцеллюлозе и зондировали антителами против His-6 (Zymed).
Очистка ORF0657n
Описанную выше лабораторную процедуру пропорционально масштабировали и эту процедуру проводили в перемешиваемых ферментерах (в объеме 75 литров) с рабочим объемом 50 литров. Инокулят культивировали в 250-миллилитровой колбе, содержащей 50 мл среды Лурия-Бертани (LB)(плюс канамицин), инокулировали 1 мл замороженной посевной культуры и культивировали в течение 3 часов. Один миллилитр этой посевной культуры использовали для инокуляции 2-литровой колбы, содержащей 500 мл среды LB (плюс канамицин) и инкубировали в течение 16 часов. В промышленном ферментере (объемом 75 литров) культивировали 50 литров среды LB (плюс канамицин). При этом были использованы следующие параметры ферментации: давление=5 фунт/кв.дюйм, скорость перемешивания=300 об/мин, поток воздуха=15 л/мин и температура=37оС. Клетки инкубировали до оптической плотности (OD), равной 0,8 единиц, при длине волны 600 нм, и индуцировали изопропил-β-K-тиогалактозидом (IPTG) при концентрации 1 мМ. Время IPTG-индуцирования составляло три часа. Клетки собирали при понижении температуры до 15°С, затем концентрировали путем пропускания через картридж из полого волокна 500КMWCO и центрифугировали при 9000×g и при 4°С в течение 20 минут. Супернатанты декантировали и мокрый клеточный осадок рекомбинантных E.coli замораживали при -70°С.
Рекомбинантные клетки E.coli (19,2 г сырой клеточной массы) суспендировали в буфере для лизиса (50 мМ Трис-HCl, рН 8,0, 0,1М NaCl, 2 мМ MgCl2, 10 мМ имидазола, 0,1% твинаТМ 80 и 6М гуанидина-HCl) при концентрации 8 мл/г сырой клеточной массы. Затем в суспензию добавляли смесь ингибиторов протеазы, используемую с белками, меченными полигистидином (Sigma, Р8849), при концентрации 0,05 мл/г клеточной пасты. Кроме того, добавляли лизозим до 1 мг/мл и бензоназуТМ (ЕМ Ind.) до 1 мкл/мл. Лизис клеток осуществляли путем 4-кратного пропускания суспензии через микрофлюидизатор при 14000 фунт/кв.дюйм (Microfluidics Model 110S) при 4°С. Клеточный дебрис осаждали при 11000×g в течение 30 минут при 4°С и супернатант сохраняли.
Белки, несущие His-метку, выделяли из супернатанта. Супернатант смешивали с 20 мл Ni+-NTA-агарозы (Qiagen) при 4°С, осторожно переворачивая, в течение 2 часов. Смесь выливали в открытую колонку (1,5 см × 20 см) и несвязанную фракцию собирали в пул. Колонку промывали буфером для промывки (20 мМ Трис-HCl, рН 8,0, 0,15М NaCl, 0,1% твинаТМ 80). His-меченный ORF0657n элюировали ступенчатым градиентом 300 мМ имидазола, 20 мМ Трис-HCl, рН 7,5, 0,15М NaCl, 0,1% твинаТМ 80.
Фракции, содержащие His-меченный ORF0657n (SEQ ID NO:28), детектировали с помощью электрофореза в окрашенном кумасси ДСН-ПААГ и объединяли. Объединенные фракции фильтровали через 0,2-микронный фильтр для удаления крупных частиц и наносили на эксклюзионную колонку (колонку с сефакрилом S-300 26/60, Amersham Bioscience), а затем элюировали при 1 мл/мин 10 мМ MOPS, рН 7,1, 150 мМ NaCl. Фракции, содержащие His-меченный ORF0657n, детектировали с помощью электрофореза в окрашенном кумасси ДСН-ПААГ и с помощью Вестерн-блоттинга (с использованием Mab против тетра-His, Qiagen). Эндотоксин удаляли путем фильтрации через биофильтр Zeta-PlusТМ (CUNO). Концентрацию белка определяли с помощью ВСА (Pierce). Чистоту определяли путем денситометрии на гелях, окрашенных кумасси.
Пример 2: Получение препарата для контрольного заражения S. aureus
S. aureus культивировали на планшетах с триптиказо-соевым агаром (TSA)(Becton Dickinson, Sparks, MD) при 37оС в течение ночи. Бактерии вымывали из TSA-планшетов путем добавления в планшет 5 мл PBS, после чего бактерии слегка ресуспендировали стерильным шпателем. Бактериальную суспензию центрифугировали при 6000 об/мин в течение 20 минут на центрифуге Sorvall RC-5B (DuPont Instrument). Осадок ресуспендировали в 16% глицерине и аликвоты хранили в замороженном виде при -70°С.
Перед использованием инокуляты оттаивали, соответствующим образом разводили и использовали для инфицирования. Для определения дозы, подходящей для индуцирования замедленной кинетики гибели неиммунизованных мышей, каждый маточный раствор титровали по крайней мере 3 раза. Затем проводили непрерывный мониторинг иммуногенности бактериального инокулята (способного вызывать гибель 80-90% мышей) для подтверждения воспроизводимости данной модели. За десять дней до проведения каждого эксперимента по инокуляции осуществляли контрольное заражение и проводили мониторинг группы из 10 контрольных животных (иммунизованных только адъювантом).
Пример 3: Исследования по индуцированию иммунитета с использованием His-меченных ORF0657n-родственных полипептидов
Двадцать пять мышей BALB/с иммунизировали тремя дозами His-меченного ORF0657n (SEQ ID NO:28), 20 мкг на дозу, с использованием адъюванта на основе гидроксифосфата алюминия (450 мкг на дозу). Адъювант на основе гидроксифосфата алюминия (АНР) описан Klein et al., Journal of Pharmaceutical Sciences 89, 311-321, 2000. Эти дозы вводили в виде двух внутримышечных 50 мкл-инъекций в дни 0, 7 и 21. На 28-й день у мышей брали кровь и их сыворотку скринировали с помощью ELISA на реакцию с His-меченным ORF0657n.
На 35-й день эксперимента мышей заражали путем внутривенной инъекции культивированного S. aureus в дозе (7,3×108 к.о.е./мл), определенной в экспериментах по титрованию дозы, приводящей к гибели примерно через 2-7 дней. Выживаемость этой летальной модели с низкой кинетикой гибели оценивали путем сравнения с выживаемостью контрольной группы мышей, которые были подвергнуты лишь ложной иммунизации АНР. Затем проводили мониторинг на выживание мышей в течение 14 дней (фиг.3А). По окончании эксперимента в ORF0657n-иммунизованной группе выживало 11 мышей, а в контрольной АНР-группе выжило только 3 мыши.
На фиг.3В и 3С проиллюстрировано индуцирование иммунитета с использованием полипептидов, соответствующих областям ORF0657nН и ORF0657nI. На фиг.3В проиллюстрировано индуцирование иммунитета с использованием SEQ ID NO:4, содержащей карбокси-концевую His-метку. На фиг.3С проиллюстрированы результаты, полученные с использованием SEQ ID NO:5, содержащей карбокси-концевую His-метку.
Пример 4: Получение последовательностей ORF0657n
Считается, что ORF0657n играет определенную роль в утилизации железа бактерией S. aureus (Andrade et al., Genome Biology 3(9):47.1-47.5, 2003). Последовательности ORF0657n, некоторые из которых были получены из различных источников, в различных работах имеют различные обозначения (см. например, Etz et al., PNAS USA, 99:6573-6578, 2002 (LPXTGVI); Baba et al., The Lancet 359:1819-1827, 2002 (MW 1011); Kuroda et al., The Lancet 357, 1225-1240, 2001 (SA0976); Andrade et al., Genome Biology 3(9):47.1-47.5, 2003 (S_aur2); Mazmanian et al., Science 299:906-909, 2003 (isdB); Mazmanian et al., Molecular Microbiology 40:1049-1057, 2001 (sasJ) & Taylor et al., Mol. Microbiol. 43:1603-1614, 2002 (sirH).
Полипептидная последовательность, соответствующая последовательности белка ORF0657n, описана в различных публикациях патентных заявок (Meinke et al., Международная заявка № WO 02/059148, опубликованная 1 августа 2002, Wang et al., Международная заявка № WO 02/077183, опубликованная 3 октября 2002, Masignani et al., Международная заявка № WO 02/094868, опубликованная 28 ноября 2002, Foster et al., Международная заявка № WO 02/102829, опубликованная 27 декабря 2002, и Foster et al., Международная заявка № WO 03/011899, опубликованная 13 февраля 2003).
Геномную ДНК получали из различных клинических изолятов S. aureus. Клинические изоляты добавляли в 3 мл триптиказо-соевого бульона Difco (Becton Dickinson, Sparks, MD) и инкубировали в течение ночи при 37оС и при 150 об/мин. Ночные культуры центрифугировали в 1,5 мл-пробирках Эппендорфа при 14000 об/мин в течение 5 минут. Затем бульон декантировали и осадок ресуспендировали в 500 мкл буфера для ресуспендирования (25% сахароза, 10 мМ Трис, рН 7,5). Затем в каждый ресуспендированный осадок добавляли 5 мкл-аликвоту 2 мг/мл раствора лизостафина (Sigma-Aldrich, St.Louis, MO). После этого суспензии инкубировали при 37оС в течение 1 часа.
По окончании инкубирования в каждую пробирку добавляли 250 мкл 2% ДСН и содержимое перемешивали до тех пор, пока не наблюдалось заметное снижение вязкости раствора. После этого добавляли 250 мкл раствора фенола-хлороформа-изоамилового спирта (25:24:1, об/об)(Gibco/Invitrogen Corporation, Grand Island, NY). Смесь перемешивали в течение 30 секунд и центрифугировали в течение 5 минут при 14000 об/мин. Верхнюю водную фазу удаляли и стадии осаждения повторяли до тех пор, пока не оставалась едва заметная граница раздела фаз. В каждую пробирку добавляли 0,1 объема 3М NaOAC, рН 4,8, и содержимое перемешивали. Затем добавляли один объем изопропанола и содержимое снова перемешивали. После этого пробирки оставляли на 5 минут при комнатной температуре для инкубирования, а затем центрифугировали при 14000 об/мин в течение 15 минут. Супернатант декантировали и пробирки ставили на ткань вверх дном для осушки. Осадок ресуспендировали в 50 мкл стерильной H2O.
Выделенную ДНК использовали в качестве матрицы для ПЦР. Ген амплифицировали с использованием прямого праймера (ORF0657nF, SEQ ID NO:67) и обратного праймера (ORF0657nR, SEQ ID NO:68). ПЦР-продукты секвенировали в соответствии со стандартными протоколами Big Dye.
Пример 5: Сравнение ORF0657n от различных изолятов S. aureus
Было обнаружено, что ORF0657n является достаточно консервативным во всей совокупности патологически и таксономически отличающихся клинических изолятов S. aureus. В таблице 3 систематизированы величины процента идентичности различных изолятов, включая клинические изоляты.
Процент идентичности (% ID) последовательности определяли путем выравнивания данной полипептидной последовательности с последовательностью SEQ ID NO:2 и определения числа идентичных аминокислот. Это число делят на общее число аминокислот (в SEQ ID NO:2), а затем результат умножают на 100 и округляют до ближайшего целого числа.
Пример 6: Иммунитет против различных клинических изолятов S. aureus
Иммуногенную эффективность ORF0657n как иммуногена против различных клинических изолятов S. aureus оценивали с использованием His-меченного ORF0657n в качестве иммуногена (SEQ ID NO:28). В качестве инокулята для заражения использовали субсерию таксономически отличающихся изолятов, описанных в таблице 3. Последовательности ORF0657n в этих различных изолятах отличались от последовательности ORF0657n, используемой в вакцине.
Штаммы, используемые для заражения, получали методами, описанными в примере 2. Мышей иммунизовали и подвергали контрольному заражению методами, описанными в примере 3, а затем проводили мониторинг в течение 10 дней. Количество инокулята для заражения, вводимое животным-моделям, значительно превышало уровень инфицирования, обычно наблюдаемый у человека.
Было продемонстрировано индуцирование иммунитета против восприимчивых и резистентных к метициллину штаммов. Эти результаты представлены на фиг.4А-4Н.
Пример 7: Оптимизация кодонов SEQ ID NO:28 для экспрессии в дрожжах
Последовательность нуклеиновой кислоты, кодирующей аминокислоты 1-646 SEQ ID NO:28, оптимизировали по кодонам для экспрессии в дрожжах. Оптимизированная по кодонам последовательность для аминокислот 1-646 SEQ ID NO:28 представлена на фиг.8С (SEQ ID NO:31).
В качестве исходной конструкции для оптимизации была использована последовательность SEQ ID NO:29, не содержащая области, кодирующей карбокси-концевую His-метку. Встречаемость всех кодонов, кодирующих данную последовательность до ее оптимизации, была следующей: очень редкие кодоны или кодоны, которые никогда не встречаются в высоко экспрессируемых генах дрожжей, составляли 28% (179), а относительно редкие кодоны составляли 20% (126).
Оптимизацию кодонов нуклеиновой кислоты, кодирующей аминокислоты 1-646 SEQ ID NO:28, осуществляли в целях замены кодонов, имеющих низкий или умеренный уровень экспрессии в дрожжах, на кодоны, имеющие высокий уровень экспрессии в дрожжах. Кроме того, был добавлен глициновый кодон во втором положении. Оптимизацию кодонов осуществляли с использованием компьютерной программы MacDNAsis Pro V3.0. В таблице применяемых параметров, используемых для обратной трансляции белков, указаны кодоны с высоким уровнем экспрессии в S. cerevisiae. В программе MacDNAsis Pro V3.0. была использована функция "Translate>[Protein-->DNA]". Выходные данные были озаглавлены "Amino Acid Conversion" ("Преобразование аминокислот").
В некоторых случаях для данной аминокислоты имелось более одного высоко экспрессирующегося кодона. Так, например, серин кодируется либо "ТСТ", либо "ТСС". В этих случаях можно использовать приблизительно равное число этих двух различных кодонов. В таблице 4 приводятся кодоны с высоким уровнем экспрессии в S. cerevisiae.
Все кодоны, которые не являются оптимальными, были заменены кодонами, обнаруженными в высокой степени экспрессирующихся генах дрожжей, за исключением двух аланиновых кодонов, которые были заменены кодонами, обнаруженными в умеренно экспрессирующихся генах (кодоны GCG, начинающиеся с нуклеотидов 505 и 1546) SEQ ID NO:31.
Полную последовательность, кодирующую ORF0657n, получали путем отжига и удлинения на 25 олигомеров (SEQ ID NO:69-93), сконструированных так, чтобы они кодировали нужную конечную последовательность. Эти олигомеры имеют длину 8-110 п.н. Такие олигомеры представляют собой чередующуюся ORF0657n-кодирующую последовательность. Каждый олигомер имеет комплементарную область в 25-29 п.н., перекрывающуюся с примыкающим олигомером, и этот дуплекс имеет Tm 80-84°С. Эта температура была вычислена вручную путем присваивания паре оснований GC значения 4°С, а паре оснований АТ - значения 2°С.
Проводили семь отдельных реакций удлинения с использованием трех или четырех смежных и перекрывающихся олигомеров и смысловых и антисмысловых ПЦР-праймеров длиной 23-26 нуклеотидов с Tm дуплекса 70-72°С. Нативную ДНК-полимеразу Pfu (STRATAGENE, La Jolla, CA) использовали для осуществления ПЦР-реакций с применением стратегии "фиксации", при следующих реакционных условиях: 1 цикл: 95°С, 90 секунд; 5 циклов: 95°С, 30 секунд, 55°С, 30 секунд, 68°С, 3 минуты; и непосредственно за ними проводили вторую серию реакции; 20 циклов: 95°С, 30 секунд, 52°С, 30 секунд, 68°С, 3 минуты. Эту реакцию завершали путем инкубирования при 68°С в течение 7 минут. В результате проведения этих ПЦР-реакций получали семь коллинеарных фрагментов гена (для удобства, обозначенных здесь 1, 2, 3, 4, 5, 6 и 7).
Эти фрагменты выделяли путем электрофореза в агарозном геле и продукты соответствующего размера вырезали и очищали методом GENECLEAN® II (QBIOgene, Carlsbad, CA) в соответствии с рекомендациями производителей. Коллинеарные фрагменты 1, 2 и 3, коллинеарные фрагменты 4 и 5 и коллинеарные фрагменты 6 и 7 объединяли в последующих ПЦР-реакциях с использованием соответствующих праймеров и получали фрагменты А, В и С соответственно. Затем осуществляли сборку полного гена ORF0657n путем проведения дополнительной ПЦР-реакции, в которой фрагменты А, В и С объединяли с использованием дистальных смысловых и антисмысловых праймеров. Конечный ПЦР-продукт выделяли из геля и клонировали в pCR®-Blunt II-TOPO® (INVITROGEN, Carlsbad, CA) в соответствии с рекомендациями производителя. Таким образом, получали ДНК-последовательность нескольких независимых клонов и идентифицировали ошибки.
Ошибки корректировали на различных сегментах трех независимых клонов, pUC3, pUC4 и pUC6, либо последовательно с использованием набора для сайт-направленного мутагенеза QUIK-CHANGE, либо одновременно с использованием набора для сайт-направленного мультимутагенеза QUIK-CHANGE (STRATAGENE, La Jolla, CA) в соответствии с рекомендациями производителя. Конечную скорректированную последовательность получали путем замены репарированных рестрикционных фрагментов из трех клонов. Репарированный 1,1 т.п.н.-XmnI-фрагмент клона pUC4 заменяли на соответствующий XmnI-фрагмент pUC3 для конструирования pUnkC13. Репарированный 456 т.п.н.-AccI-фрагмент клона pUC6 заменяли на соответствующий фрагмент pUnkC13 для конструирования pUnkCR1 и последовательность ДНК подтверждали.
Пример 8: Конструирование штаммов Saccharomyces cerevisiae для экспрессии рекомбинантного гена
В этом примере проиллюстрированы методы, которые могут быть применены в целях получения штаммов Saccharomyces cerevisiae для экспрессии рекомбинантного гена. Создание штаммов, обозначенных 1260 и 1309, описано ниже. Поскольку генетический фон штамма может сильно влиять на способность штамма экспрессировать гетерологичный белок, то желательно сконструировать дрожжевые штаммы с различными генетическими фонами, которые также имеют несколько нужных генетических маркеров: (1) мутацию mnn9 для предупреждения гипергликозилирования секретируемых белков; (2) мутации протеазы prb1 и/или pep4 для решения проблемы, связанной с протеолизом, и (3) сверхэкспрессию фактора транскрипции GAL4 для повышения уровня экспрессии с промоторов GAL.
Конструирование штамма S. cerevisiae, обозначенного 1260
Исходный штамм S. cerevisiae конструировали следующим образом. Штамм Y379-5D S. cerevisiae (MATα, cyh2, nib1, rho-, ciro)(Livingston, Genetics 86:73-84, 1977) скрещивали со штаммом DC04 (МАТа, ade1, adeX, leu2-04, ciro)(Broach et al., Cell 21:501-508, 1980). Полученный диплоидный штамм подвергали споруляции и тетрады иссекали стандартными способами. Одна из гаплоидных спор продуцировала штамм 2150-2-3 (МАТа, ade1, leu2-04, ciro). Штамм S. cerevisiae типа LB-347-1С (МАТα, mnn9), предназначенный для α-скрещивания, представляет собой штамм X2180-1B (МАТα, SUC2, mal, mel, gal2, CUP1; АТСС рег.№ 204504), содержащий мутацию mnn9. LB-347-1С скрещивали со штаммом типа 2150-2-3 (MATa, leu2-04, ade1) путем смешивания указанного штамма на планшете с агаром, содержащим полную среду YEHD (Carty et al., J. Ind. Micro. 2:117-121, 1987). Для отбора диплоидов скрещенные штаммы высевали в виде реплик на минимальную среду, не содержащую лейцина и содержащую 2% сахарозу в качестве единственного источника углерода. После выделения одиночных колоний диплоиды подвергали споруляции и сумки иссекали стандартными методами. Штамм KHY-107 выделяли в виде одной гаплоидной споры и характеризовали как ADE1, leu2 и mnn9 (путем окрашивания по Шиффу). Затем приготавливали замороженный маточный раствор в глицерине и хранили при -70°С.
KHY-107 (cir+) культивировали из маточного раствора (-70°С) и трансформировали с использованием дрожжевого вектора рС1/1, который содержит дрожжевой ген LEU2-d и является родственным вектору рJDB219 (Beggs, Nature 275:104-109, 1978), за исключением того, что ДНК рМВ9 заменяли ДНК рВR322. Выделенный трансформант культивировали на селективной жидкой среде (не содержащей лейцин и содержащей 1М сорбит), а затем культивировали путем многократных пассажей в обогащенной среде (содержащей 1М сорбит) для удаления рС1/1 и 2 мкм ДНК. Колонии, которые не содержали рС1/1 (то есть, были неспособны расти на среде, не содержащей лейцин), оценивали с помощью ДНК-ДНК-гибридизации на присутствие 2 мкм ДНК. Выделенный клон, KHY-107 (cirо)-1, не обнаруживающий 2 мкм ДНК, отбирали и приготавливали как замороженный маточный раствор (-70°С) в глицерине.
KHY-107 (cirо)-1 создавали путем дизрупции гена ura3. Для этого конструировали плазмиду, содержащую ген URA3 S. cerevisiae, подвергнутый дизрупции 2 копиями гена Aph3'I из Tn903. 5'-URA3-Aph3'I-URA3-3'-кластер вырезали из вектора и использовали для трансформации KHY-107 (cirо)-1. Трансформанты, которые содержали интегрированный в них кластер с подвергнутым дизрупции геном ura3, отбирали на чашках с 5-фтороротовой кислотой (Kaiser C. et al., Methods in Yeast Genetics - 1994 Edition, Cold Spring Harbor Laboratory Press (Cold Spring Harbor, NY, 1994)pages 214-215), и было обнаружено, что они не способны расти на среде, не содержащей урацил. Один выделенный клон, KHY-107ura3(PN2), отбирали и приготавливали как замороженный маточный раствор (-70°С) в глицерине.
KHY-107ura3(PN2) культивировали на комплексной среде (содержащей 1М сорбит), а затем высевали на синтетическую среду, содержащую канаванин вместо аргинина, и получали мутанты, резистентные к канаванину (canR). Для получения отдельных колоний спонтанные canR-мутанты высевали штрихом на твердую синтетическую среду, содержащую канаванин вместо аргинина. Выделенные колонии были идентифицированы с помощью стандартных тестов на генетическую комплементацию как can1-колонии. Одну выделенную can1-колонию, DМY10, отбирали и хранили в виде замороженного (-70°С) маточного раствора в глицерине.
Для сверхэкспрессии фактора транскрипции GAL4 в дрожжах гибридный ген GAL10р-GAL4 интегрировали в ген HIS3 DМY10. Кластер 5'-HIS3-GAL10р-GAL4-URA3-HIS3-3' вырезали из pKHint-C (Schultz et al., Gene 61:123-133, 1987) и использовали для трансформацми DМY10. Трансформанты, которые содержали указанный интегрированый кластер, отбирали на твердой синтетической среде, не содержащей урацил, и было показано, что они не способны расти на среде, не содержащей гистидин. Интеграцию этого кластера только в локусе HIS3 подтверждали с помощью Саузерн-гибридизации. Один выделенный интегрант, DМY10int-3, отбирали и приготавливали как замороженный маточный раствор (-70°С) в глицерине. Этот штамм обозначали CF52.
Плазмиду FP8ΔН, несущую ген PRB1 S. cerevisiae (Moehle et al., Genetics 115:255-263, 1987), гидролизовали ферментами HindIII и XhoI и 3,2 т.п.н-ДНК-фрагмент, несущий ген PRB1, подвергали гель-очистке и концы затупляли путем обработки ДНК-полимеразой Т4. Плазмиду pUC18 гидролизовали ферментом BamHI, подвергали гель-очистке и затупляли по концам путем обработки ДНК-полимеразой Т4. Полученный фрагмент вектора лигировали с вышеуказанным фрагментом гена PRB1 и получали плазмиду pUC18-PRB1. Плазмиду YEp6 (Struhl et al., Proc.Natl. Acad. Sci., USA, 76:1035, 1979), содержащую ген HIS3, гидролизовали ферментом BamHI. Полученный 1,7-т.п.н.-BamHI-фрагмент, несущий функциональный ген HIS3, подвергали гель-очистке, а затем затупляли по концам путем обработки ДНК-полимеразой Т4. Вектор pUC18-PRB1 гидролизовали ферментами EcoRV и NcoI, которые разрезали сайт в PRB1-кодирующей последовательности и удаляли активный центр протеазы В и фланкирующую последовательность. 5,7 т.п.н-EcoRV-NcoI-фрагмент, несущий остаточные 5'- и 3'-части PRB1-кодирующей последовательности в pUC18, подвергали гель-очистке, затупляли по концам путем обработки ДНК-полимеразой Т4, дефосфорилировали щелочной фосфатазой и лигировали с затупленным по концам HIS3-фрагментом, описанным выше. Полученная плазмида (обозначенная pUC18-prb1::HIS3, штамм #1245) содержала функциональный ген HIS3 вместо части гена PRB1, который был делетирован, как описано выше.
Вектор с дизрупцией гена PRB1 (pUC18-prb1::HIS3) гидролизовали ферментами SacI и XbaI, в результате чего получали линейный кластер с дизрупцией гена prb1::HIS3 и этот кластер использовали для трансформации штамма CF52 методом с применением ацетата лития (Ito et al., J. Bacteriol. 153:163, 1983). His+-трансформанты отбирали на синтетической агаровой среде, не содержащей гистидина, и снова высевали штрихом на ту же самую среду для получения клональных изолятов. Геномную ДНК выделяли из числа полученных His+-изолятов, гидролизовали ферментом EcoRI, а затем подвергали электрофорезу в 0,8% агарозных гелях. Саузерн-блот-анализы подтвердили присутствие нужной конструкции с дизрупцией гена prb1Δ::HIS3.
Один из изолятов, содержащих нужную дизрупцию гена prb1Δ::HIS3, отбирали для последующего использования и обозначали как штамм #1260. Замороженные маточные растворы в глицерине приготавливали для хранения штамма #1260 при -70°С. Полученный генотип штамма 1260 представлял собой MATa, leu2-2, 112, mnn9, ura3Δ, can1, his3Δ::GAL10p-GAL4-URA3, prb1Δ::HIS3, ciro.
Конструирование штамма S. cerevisiae, обозначенного 1309
Штамм BJ1995 S. cerevisiae (MATα, leu2, trp1, ura3-52, prb1-1122, pep4-3, gal2) был описан ранее (Jones, E.W. Tackling the Protease Problem in Saccharomyces cerevisiae, Methods in Enzymology 194(1991), pages 428-453). ciro-Производное BJ1995, не содержащее эндогенную 2 мк-ДНК-плазмиду, выделяли в соответствии с процедурой, описанной для конструирования штамма 1260. Полученный ciro-изолят обозначали штаммом 91 и хранили в виде замороженного маточного раствора (-70°С) в глицерине.
Затем конструировали производное штамма 91, которое обладало способностью к сверхпродуцированию фактора транскрипции GAL4, для повышения уровня транскрипции с промоторов GAL. Плазмиду pKHint-C (Schultz et al., Gene 61:123-133, 1987) гидролизовали ферментом BamHI и полученный кластер 5'-HIS3-GAL10p-GAL4-URA3-HIS3-3' использовали для трансформации штамма 91. Трансформанты, которые содержали и указанный интегрированный кластер, отбирали на твердой синтетической среде, не содержащей урацил, и было обнаружено, что они не способны расти на среде, не содержащей гистидин. Желательная интеграция этого кластера в локусе HIS3 была подтверждена с помощью Саузерн-гибридизации с использованием зонда для гена HIS3 дрожжей. Один выделенный интегрант, BJ1995ciroint#22, отбирали и приготавливали как замороженный маточный раствор (-70°С) в глицерине. Этот изолят обозначали штаммом 1282.
Затем производное штамма 1282, содержащее дизрупцию гена MNN9, выделяли в последующих стадиях. Для дизрупции гена MNN9 необходимо сначала клонировать ген MNN9 из геномной ДНК S. cerevisiae в целях получения вектора с дизрупцией гена. Это было осуществлено с помощью стандартной ПЦР-техники. 5'-Смысловой праймер и 3'-антисмысловой праймер для ПЦР полноразмерной MNN9-кодирующей последовательности конструировали исходя из опубликованной последовательности гена MNN9 дрожжей (MacKay et al., Европейская международная патентная заявка № ЕР0314096, опубликованная 3 мая 1989). Были использованы нижеследующие олигодезоксинуклеотидные праймеры, содержащие фланкирующие HindIII-сайты (подчеркнуты):
смысловой праймер (SEQ ID NO:94): 5'-CTT AAA GCT TAT GTC ACT TTC TCT TGT ATC G-3'
антисмысловой праймер (SEQ ID NO:95): 5'-TGA TAA GCT TGC TCA ATG GTT CTC TTC CTC-3'.
Инициирующий метиониновый кодон для гена MNN9 выделен жирным шрифтом.
ПЦР проводили с использованием геномной ДНК от S. cerevisiae в качестве матрицы. Полученный 1,2-т.п.н.-ПЦР-фрагмент, несущий ген MNN9, гидролизовали ферментом HindIII, подвергали гель-очистке и лигировали с HindIII-гидролизованным и обработанным щелочной фосфатазой вектором pUC13. Полученную плазмиду обозначали р1183. Ген TRP1 дрожжей выделяли из YRр7 (Struhl et al., Proc.Natl. Acad. Sci., USA, 76:1035-1039, 1979) в виде 0,85 т.п.н. EcoRI-BglII-фрагмента с достроенными концами, а затем встраивали в PmlI-сайт р1183. PmlI-сайт находится в середине кодирующей последовательности MNN9, в результате чего встраивание этого гена TRP1 в указанный сайт приводит к дизрупции гена. Затем полученную плазмиду pUC13-mnn9::TRP1 (p11885-239-1) гидролизовали ферментами HpaI и EcoRI и кластер 5'-mnn9-TRP1-mnn9-3' использовали для трансформации штамма 1282 методом с применением ацетата лития (Ito et al., J. Bacteriol. 153:163, 1983).
Trp+-трансформанты отбирали на чашках с синтетической агаровой средой, не содержащей триптофана, и пересевали штрихом на те же самые чашки для получения изолятов одиночных колоний. Геномную ДНК выделяли из ряда полученных Trp+-изолятов, гидролизовали ферментом HindIII, а затем подвергали электрофорезу в 0,8% агарозных гелях. Саузерн-блот анализ подтвердил присутствие нужной дизрупции гена mnn9::TRP1. Один из этих изолятов, содержащих нужную дизрупцию гена mnn9::TRP1, отбирали для последующего использования и обозначали штаммом 1309. Замороженные маточные растворы в глицерине приготавливали для хранения штамма 1309 при -70°С. Генотип штамма 1309 представлял собой MATα, leu2, mnn9::TRP1, trp1, ura3-52, his3Δ::GAL10p-GAL4-URA3, prb1-1122, pep4-3, gal2, ciro.
Пример 9: Экспрессия полноразмерной области ORF0657n
Полноразмерную область ORF0657n экспрессировали в E.coli (SEQ ID NO:28; His-меченный ORF0657n) и в S. cerevisiae (аминокислоты 1-646 SEQ ID NO:28) и продукты экспрессии сравнивали.
Экспрессия в S. cerevisiae
ДНК, кодирующую белок ORF0657n с кодонами, оптимизированными для экспрессии в дрожжах, амплифицировали из конечного скорректированного клона, pUnkCR1 (пример 7), с использованием следующих смыслового и антисмыслового праймеров соответственно: UnkCY-F (SEQ ID NO:96), 5'-AAC CGG TTT GGA TCC CAC AAA ACA AAA TGG GTA ACA AGC AAC AAA AGG AAT TC-3' и UnkCY-R (SEQ ID NO:97), 5'-AAC CGG TTT GGA TCC TTA GTT CTT TCT CTT TCT TGG CAA GAC-3', содержащих фланкирующие рестрикционные BamHI-сайты. Смысловой праймер содержит 5'-нетранслируемую последовательность и кодон ATG, а антисмысловой праймер содержит ТАА в качестве стоп-кодона. Полученный 1,9 т.п.н-продукт выделяли из геля и клонировали в вектор рСR2.1 ТОРО ТА (INVITROGEN CORPORATION, Carlsbad, CA) в соответствии с инструкциями производителей и получали плазмиду UnkC-B1. Затем подтверждали ДНК-последовательность вставки. BamHI-фрагмент выделяли из геля и субклонировали в "правильной" ориентации в дрожжевой вектор (pGAL110, фиг.5А), в результате чего получали конструкцию pRUnkC-pGAL110. Последовательность вставки подтверждали путем секвенирования ДНК.
Плазмидную ДНК от pRUnkC-pGAL110 использовали для трансформации штаммов S. cerevisiae, содержащих leu2-мутацию, приводящую к прототрофии по лейцину (Leu+), в соответствии с протоколом трансформации сферопластов (Hinnen et al., Proc.Natl. Acad. Sci., USA, 75:1929-33, 1978). Конструирование штаммов 1260 и 1309 описано выше в примере 8.
Трансформанты отбирали на синтетической агаровой среде, не содержащей лейцина и содержащей 1М сорбит. Полную и минимальную синтетическую агаровую среду, не содержащую лейцина и содержащую 1М сорбит, получали от REMEL, Lenexa, KS (cat # 09459 и 92155 соответственно). Клональные Leu+-изоляты получали путем серийного размножения на SD-чашках без лейцина. (KD MEDICAL, Columbia MD).
Для скрининга множественных трансформантов 5,0 мл культур-продуцентов культивировали в культуральных пробирках с медленным вращением при 30°С. Затем продуцирование отобранных трансформантов подтверждали в 25 мл-культурах в 125 мл-колбах. В обоих случаях 5 мл посевных культур культивировали при 30°С в 5х среде, не содержащей лейцина и содержащей 4,0% глюкозы и 0,1М сорбита, в течение 18-24 часов до достижения OD600=1,5-3,0/мл. 5Х среда без лейцина содержала следующие компоненты на литр: дрожжевое азотистое основание без добавления аминокислот или сульфата аммония 8,5 г; аденин 0,40 г; L-тирозин 0,25 г; урацил 0,20 г; янтарная кислота 10,0 г; сульфат аммония 5,0 г и 50 мл раствора #3, не содержащего лейцина. Раствор #3 без лейцина содержал на один литр дистиллированной воды: L-аргинин 2 г; L-гистидин 1,0 г; L-изолейцин 6 г; L-лизин 4,0 г; L-метионин 1,0 г; L-фенилаланин 6,0 г; L-триптофан 4,0 г. рН среды доводили до 5,3 добавлением 50%-ного гидроксида натрия.
Для продуцирования трансформантов в пробирках 0,3 мл аликвот посевной культуры переносили либо в 5,0 среды 5Х, не содержащей лейцина и содержащей 2% глюкозу, 4% галактозу, либо в среду YEHDG, на 72 часа до достижения OD600 5-16,0/мл. Среда YEHDG содержала на 1 литр: L-Hy-соевый пептонный бульон Шеффилда 10 г; дрожжевой экстракт 20 г; L-декстроза 16 г; D(+)-галактоза 40 г. Для продуцирования трансформанта в колбах 1,5 мл аликвоту посевной культуры переносили в 25 мл среды и культивировали, как описано выше, со встряхиванием при 220 об/мин.
После сбора 10 единиц OD на образец клеточный осадок разрушали с помощью стеклянных сфер в 0,3 мл буфере для лизиса (0,1М натрийфосфатного буфера, рН 7,2, 0,5 М NaCl, 2 мМ PMSF). Лизат выделяли путем центрифугирования, неразрушенные клетки/сферы промывали 0,3 мл буфера для лизиса и осветленные супернатанты объединяли. Концентрацию белка определяли с использованием системы реагентов-красителей для анализа на белок BIO-RAD (BIO-RAD, Hercules, CA) в соответствии с инструкциями производителей. Клеточные лизаты анализировали на экспрессию ORF0657n путем проведения иммуноблот-анализа после электрофореза в 4-20% градиенте трис-глициновых гелей (INVITROGEN, Carlsbad, CA) в 1х трис-глициновом буфере с ДСН (BIO-RAD) в условиях восстановления и денатурации. Образцы содержали 20 мкг общего клеточного белка. Гели подвергали электроблоттингу на 0,45-микронных нитроцеллюлозных мембранных фильтрах (Optitran from Schleicher and Schuell, Keene, NH). Для оценки размера белка параллельно проводили электрофорез предварительно окрашенных стандартов с массой в пределах 6,4-203 кДа (Broad Range Prestained SDS-PAGE Standard, BIO-RAD) и лизатов.
E.coli-стандарт
His-меченный ORF0657n (SEQ ID NO:28) выделяли из культуры-продуцента E.coli (штамма-хозяина E.coli НМS174(DЕ3), Madison, NOVAGEN, WI), стимулированной для продуцирования ORF0657n, и клеточный лизат использовали в качестве стандартов. E.coli трансформировали экспрессионным вектором, экспрессирующим His-меченный ORF0657n. Культуру E.coli выращивали в течение ночи на LB-бульоне, содержащем 30 мкг/мл канамицина при 37оС. На следующий день 60 мкл ночной культуры использовали для инокуляции 6,0 мл LB плюс 30 мкг на мл канамицина. Эту культуру выращивали при 37оС в течение приблизительно 3 часов до тех пор, пока OD600 не достигала 0,4-1,0. Экспрессию индуцировали 1 мМ IPTG в течение 2 часов при 37оС. Затем клетки собирали и клеточный осадок хранили при -80°С.
Лизат E.coli получали с использованием реагента для экстракции белка (Bugbuster) (NOVAGEN, Madison, WI) в соответствии с протоколом производителя. Белки подвергали иммунодетекции с помощью Вестерн-блоттинга с использованием мышиного моноклонального антитела (обозначенного "2Н2В8") против ORF0657n в качестве "первого" антитела и конъюгированного с пероксидазой хрена целого козьего антитела против мышиного IgG (Н+L)(ZYMED LABORATORIES, South San Francisco, CA) в качестве "второго" антитела. Mab 2Н2В8 генерировали путем иммунизации мышей очищенным и продуцированным в E.coli полноразмерным ORF0657n. Mab 2Н2В8 отбирали с помощью ELISA, и было показано, что это антитело обладает специфичностью к ORF0657n. Фильтры обрабатывали с использованием набора, содержащего ПХ-конъюгированный субстрат BIO-RAD.
Продукты экспрессии из E.coli и S. cerevisiae
Исходя из начального скрининга дрожжевых трансформантов, один трансформант от каждого из дрожжевых штаммов 1260 и 1309 отбирали как наилучший из продуцентов полноразмерной области ORF0657n (SEQ ID NO:28 без His-метки на карбокси-конце). Пример продуцирования этим штаммом полноразмерной области ORF0657n после 72-часовой ферментации в среде YEHDG по сравнению с продуцированием области ORF0657n (SEQ ID NO:28) в E.coli приводится на фиг.6А-6В.
Мажорный белок, детектированный с помощью Вестерн-блот-анализа с использованием mab 2H2B8, имел молекулярную массу ˜105-110 кДа, как показано на фиг.6А (штамм 1260) и 6В (штамм 1309): дорожки 5 и 6. ˜105-110 кДа-белок по своему размеру соответствовал самому крупному белку, детектированному в образце очищенного рекомбинантного ORF0657n, продуцированного в E.coli (SEQ ID NO:28; дорожка 2) или в экстракте индуцированной ORF0657n-продуцирующей культуры E.coli (последняя не включена в гели, показанные на фиг.6). 105-110 кДа-белки, продуцируемые в E.coli и S. cerevisiae, имели более высокую молекулярную массу, чем это было предсказано в анализе, проведенном авторами настоящего изобретения с использованием системы гель-электрофореза. При этом следует отметить, что преобладающие молекулы, детектируемые в E.coli-контролях, имели меньший размер, ˜95 кДа, и мигрировали вместе с небольшим количеством белка, продуцированного в дрожжах. Очевидно, что ˜105-110 кДа-белок соответствует полноразмерному ORF0657n, который экспрессировался с лидерной последовательностью E.coli как в E.coli, так и в S. cerevisiae, а 95 кДа-белок, очевидно, является продуктом деградации. В экстракте контрольного трансформанта, полученного с использованием только вектора рGAL110 (дорожки 3 и 4) белок не детектировался.
Оценка количества области ORF0657n (SEQ ID NO:28 без метки His на карбокси-конце), продуцированной трансформантами 1260 и 1309, показала, что это количество составляло ˜10 микрограммов ORF0657n/мл среды YEHDG, где область ORF0657n составляла ˜2% от общего белка, как было определено с помощью полуколичественного Вестерн-блот-анализа с использованием 100 нг очищенного ORF0657n (SEQ ID NO:28) в качестве стандарта. Количества обоих экспрессируемых трансформантов и % области ORF0657n (SEQ ID NO:28 без карбокси-концевой His-метки) от общего белка через 72 часа были выше, чем указанные количества в обоих штаммах через 48 часов (данные не приводятся). Титр области ORF0657n (SEQ ID NO:28 без карбокси-концевой His-метки), продуцированной трансформантами 1260 в 5х среде с лейцином, содержащей 2% глюкозу и 4% галактозу, составлял ˜8,0 мкг/мл, а % общего белка составлял 2,0.
Пример 10: Экспрессия секретированной и оптимизированной по кодонам последовательности, кодирующей область ORF0657nН
Вектор рКН4-3В (Carty et al., Biotechnol. Lett. 12:879-884, 1990) содержит секреторную лидерную пре-про-последовательность дрожжевого альфа-фактора (MFα1). Образование гибрида нужного белка с этой лидерной последовательностью приводит к трансляции продукта, который сначала отщепляется сигнальной пептидазой S. cerevisiae. Затем протеаза Кех2 отщепляет последовательность, расположенную за остатками "Lys Arg" в секреторной лидерной последовательности, в результате чего высвобождается зрелый белок. Для запуска экспрессии белка использовали индуцибельный промотор GAL10 S. cerevisiae.
Для экспрессии оптимизированного по кодонам гена, кодирующего модифицированную SEQ ID NO:3, использовали вектор рКН4-3В. SEQ ID NO:3 модифицировали путем удаления амино-концевого метионина и добавления амино-концевой лидерной последовательности. Оптимизированная по кодонам SEQ ID NO:3-кодирующая область представлена нуклеотидами 4-1710 SEQ ID NO:32, а последовательность, кодирующая лидерную последовательность, обеспечивалась вектором рКН4-3В, полученным, как описано ниже.
Был получен гибрид секреторной лидерной пре-про-последовательности альфа-фактора рКН4-3В с областью ORF0657nН SEQ ID NO:3 с сохранением рамки считывания. Это было осуществлено путем гидролиза вектора рКН4-3В ферментом SphI, а затем путем обработки ДНК-полимеразой Т4 для удаления SphI-выступающих концов и создания тупого 5'-конца с соответствующим кодоном лидерной последовательности. Затем вектор гидролизовали ферментом BamHI для создания 3'-сайта клонирования. Для этой конструкции требовался ПЦР-продукт с затупленным 5'-концом, соответствующим второму кодону реконструированной области ORF0657nН (SEQ ID NO:32), и с 3'-концом, имеющим стоп-кодон и рестрикционный BamHI-сайт.
ДНК, кодирующую область ORF0657nН с кодонами, оптимизированными для экспрессии в дрожжах, амплифицировали из pUnkCR1 (пример 7) с использованием нижеследующих смыслового и антисмыслового праймеров соответственно: UCKHS2 (SEQ ID NO:98), 5'GCTGAAGAAACTGGTGGTACCAAC-3' и UCKHA2 (SEQ ID NO:99), 5'-GTCACGGATCCTTAAGACTTAGCCTTGTTTTCTTGAGTGTTC-3'. 5'-конец смыслового праймера, GCT, кодирует аланин, предсказанный N-конец области ORF0657nН. Антисмысловой праймер содержит стоп-кодон ТАА и BamHI-сайт (подчеркнут). Полученный 1,7 т.п.н.-ПЦР-продукт выделяли из геля и клонировали в рКН4-3В (полученный, как описано выше) с образованием конструкции pUS38. Затем подтверждали всю ДНК-последовательность вставки и неполную последовательность вектора, и было показано, что в результате был создан нужный гибрид с секреторной лидерной последовательностью. Соответствующее расщепление протеазой Кех2р исходного белка, экспрессированного из этой конструкции, должно приводить к образованию белка, соответствующего последовательности SEQ ID NO:3 и не содержащей N-концевого метионина.
Плазмиду pUS38 использовали для трансформации штаммов 1260 и 1309 S. cerevisiae, так, чтобы они обладали прототрофией по лейцину (Leu+), в соответствии с протоколом трансформации сферопластами (Hinnen et al., Proc.Natl. Acad. Sci., USA, 75:1929-33, 1978). Трансформанты отбирали на синтетической агаровой среде, не содержащей лейцина и содержащей 1М сорбит. Полную и минимальную синтетическую агаровую среду, не содержащую лейцина и содержащую 1М сорбит, получали от REMEL, Lenexa, KS (cat # 09459 и 92155 соответственно). Клональные Leu+-изоляты получали путем серийного размножения на SD-чашках без лейцина (KD MEDICAL, Columbia MD).
Для скрининга множественных трансформантов 5 мл посевных культур культивировали при 30°С в 5х среде, не содержащей лейцина и содержащей 4,0% глюкозы и 0,1М сорбита, в течение 18-24 часов до достижения OD600 1,5-3,0/мл. 5Х Среда без лейцина содержала следующие компоненты на литр: дрожжевое азотистое основание без добавления аминокислот или сульфата аммония 8,5 г; аденин 0,40 г; L-тирозин 0,25 г; урацил 0,20 г; янтарная кислота 10,0 г; сульфат аммония 5,0 г и 50 мл раствора #3, не содержащего лейцина. Раствор #3 без лейцина содержит на один литр дистиллированной воды: L-аргинин 2 г; L-гистидин 1,0 г; L-изолейцин 6 г; L-лизин 4,0 г; L-метионин 1,0 г; L-фенилаланин 6,0 г; L-триптофан 4,0 г. рН среды доводили до 5,3 добавлением 50%-ного гидроксида натрия. 0,3 мл-аликвоту переносили либо в 5,0 мл 5х среды, не содержащей лейцин и содержащей 2% глюкозу + 4% галактозу, либо в среду YEHDG, на 72 часа до достижения конечной оптической плотности OD600 10-20,0/мл. Среда YEHDG содержала на 1 литр: L-Hy-соевый пептонный бульон Шеффилда 10 г; дрожжевой экстракт 20 г; L-декстрозу 16 г; D(+)-галактозу 40 г.
Продукты ферментации собирали путем удаления дрожжевых клеток и анализа супернатантов непосредственно на экспрессию области ORF0657nН (SEQ ID NO:3) с помощью Вестерн-блот-анализа и кумасси-окрашивания ДСН-ПААГ-гелей. Для иммуноблот-анализа 10-25 микролитров образцов подвергали электрофорезу в 4-15% градиенте Трис-HCl-гелей (BIO-RAD, Hercules, CA) в 1х трис-глициновом буфере с ДСН (BIO-RAD, Hercules, CA) в условиях восстановления и денатурации. Гели подвергали электроблоттингу на 0,2-микронных мембранных ПВДФ-фильтрах. Белки подвергали иммунодетекции с использованием моноклонального антитела 2Н2В8 в качестве "первого" антитела и конъюгированного с пероксидазой хрена целого козьего антитела против мышиных IgG (Н+L)(ZYMED LABORATORIES, South San Francisco, CA) в качестве "второго" антитела. Mab 2Н2В8 описано в примере 9. Фильтры обрабатывали с использованием набора для хемилюминесценции WESTERN LIGHTNINGTm Chemiluminescence Reagent Plus Kit (PERKIN ELMER, Wellesley, MA).
Для анализа на экспрессию рекомбинантной дрожжевой области ORF0657nН (SEQ ID NO:3) путем кумасси-окрашивания ДСН-ПААГ-гелей образцы подвергали электрофорезу в 4-15% градиенте трис-HCl-гелей (BIO-RAD) в 1х трис-глициновом буфере с ДСН (BIO-RAD) в условиях восстановления и денатурации. Гели окрашивали биологически безопасным красителем кумасси (Bio-Safe) G250 в соответствии с протоколом производителя (BIO-RAD).
Клеточный лизат культуры E.coli, продуцирующей ORF0657n (SEQ ID NO:4 с карбокси-концевой His-меткой), использовали как качественный стандарт. Предсказанная аминокислотная последовательность области ORF0657nН, экспрессируемой в E.coli, была идентична предсказанной аминокислотной последовательности области ORF0657nН, экспрессируемой во внутренней части S. cerevisiae (SEQ ID NO:3), за исключением того, что в E.coli-конструкции глицин находился за N-концевым метионином.
Для продуцирования области ORF0657nН в E.coli культуру-продуцент культивировали в течение ночи на бульоне LB, содержащем 50 мкг/мл канамицина при 37оС. Для экспрессии белка использовали рЕТ28, кодирующий SEQ ID NO:4 с карбокси-концевой His-меткой. На следующий день 500 мкл ночной культуры использовали для инокуляции 5,0 мл бульона LB плюс 50 мкг на мл канамицина. Эту культуру выращивали при 37оС в течение приблизительно 3 часов до тех пор, пока OD600 не достигала 0,6. Экспрессию индуцировали 1 мМ IPTG в течение 3,5 часов при 37оС. Эти клетки собирали и клеточный осадок хранили при -80°С. Лизат E.coli получали с использованием реагента для экстракции белка (Bugbuster)(NOVAGEN, Madison, WI) в соответствии с протоколом производителя.
Белок трансформанта 1-1 клеточного лизата S. cerevisiae, экспрессирующего внутреннюю область ORF0657nН (SEQ ID NO:3), описанную и полученную, как показано ниже в примере 11, был также включен в качестве стандарта. Для оценки размера белка параллельно проводили электрофорез предварительно окрашенных стандартов с массой в пределах 10-250 кДа (BIO-RAD) и образцов на ДСН-ПААГ-гелях для их анализа путем окрашивания кумасси и Вестерн-блоттинга.
Продуцирование нужных молекул осуществляли в 5Х среде, не содержащей лейцин и содержащей 2% глюкозу и 4% галактозу. Трансформанты, содержащие рUS38 обоих штаммов 1260 и 1309, секретировали ˜80 кДа-белок, который очень тесно мигрировал с областью ORF0657nН, экспрессируемой внутри дрожжевой клетки (из нуклеиновой кислоты, кодирующей SEQ ID NO:3), и с областью ORF0657nН, экспрессируемой в E.coli (из нуклеиновой кислоты, кодирующей SEQ ID NO:4 с карбокси-концевой His-меткой), которые детектировали с помощью Вестерн-блоттинга и окрашивания кумасси. В обычном эксперименте 500 нг этих контрольных лизатов и 25 микролитров супернатанта среды подвергали электрофорезу. Детекция, проводимая с помощью Вестерн-блоттинга и окрашивания кумасси, была специфической, то есть ˜80 кДа-белок не детектировался в супернатанте трансформанта, содержащего только вектор. Секретированный ˜80 кДа-белок должен соответствовать зрелой не гликозилированной области ORF0657nН (SEQ ID NO:3), или, альтернативно, он должен содержать несколько гликозильных остатков. Высокомолекулярные белки в супернатантах, а также два низкомолекулярных белка, каждый из которых был окрашен кумасси синим, детектировали с помощью антитела. Высокомолекулярные белки должны содержать непроцессированную лидерную последовательность, и/или они должны быть гликозилированы. Низкомолекулярные продукты, вероятно, являются продуктами деградации.
Оценка количества 80 кДа-продуктов, секретированных трансформантами обоих штаммов 1260 и 1309, показала, что это количество составляет ˜2 мкг/мл культуральной среды. Это было определено путем сравнения Вестерн-сигнала при 80 кДа с сигналом образца дрожжевого клеточного лизата, содержащего область ORF0657nН (SEQ ID NO:3), что позволяет предположить, что область ORF0657nН составляет по крайней мере 50% от общего белка. Было определено, что объединенный титр остальных молекул области ORF0657n составлял приблизительно 50 мкг на мл культуральной среды.
Пример 11. Внутриклеточная экспрессия области ORF0657nН (SEQ ID NO:3) в S. cerevisiae
Продуцируемый на очень высоком уровне белок в виде отдельных фракций был получен путем внутриклеточной экспрессии SEQ ID NO:3 в S. cerevisiae. Экспрессию осуществляли с использованием кодирующей последовательности нуклеиновой кислоты с кодонами, оптимизированными для экспрессии в дрожжах.
ДНК, кодирующую область ORF0657nН с кодонами, оптимизированными для экспрессии в дрожжах (SEQ ID NO:32), амплифицировали из pUnkCR1 (пример 7) с использованием следующих смыслового и антисмыслового праймеров соответственно: (SEQ ID NO:100) 5'-GGGG GGATCC CACAAAACAAA ATG GCT GAA GAA ACT GGT GG -3' и (SEQ ID NO:101) 5'-GGG GGG GGATCC TTA AGA CTT AGC CTT GTT TTC TTG AGT-3', содержащих фланкирующие рестрикционные BamHI-сайты (подчеркнуты). Смысловой праймер содержал 5'-нетранслируемую лидерную последовательность и кодон ATG, а антисмысловой праймер содержал ТАА в качестве стоп-кодона.
Полученный 1,7 т.п.н-продукт выделяли из геля и клонировали в вектор рСR2.1 ТОРО ТА (INVITROGEN CORPORATION, Carlsbad, CA) в соответствии с инструкциями производителей и получали плазмиду pCR_iUC-S. Затем ДНК-последовательность вставки подтверждали исходя из двух независимых клонов, pCR_iUCS2.2 и pCR_iUCS-2.4. В каждой из этих вставок, локализованных на различных HindIII-фрагментах каждой плазмиды, была обнаружена одна ПЦР-ошибка. Плазмиду со скорректированной последовательностью получали путем замены 1,3 т.п.н.-HindIII-фрагмента плазмиды pCR_iUC-S2.2 со скорректированной последовательностью на соответствующий фрагмент pCR_iUCS-2.4.
1,3 т.п.н.-фрагмент pCR_iUC-S2.2 и 4,3 т.п.н.-фрагмент pCR_iUCS-2.4 выделяли из геля и лигировали. Клон с нужными фрагментами, pCR_iUC-S, отбирали, и соответствующую ориентацию и нуклеотидную последовательность подтверждали путем секвенирования ДНК. Затем 1,7 т.п.н.-BamHI-фрагмент субклонировали в рGAL110 (фиг.5А) для конструирования piUCS-S(-).
Плазмидную ДНК от piUC-S(-)использовали для трансформации штаммов 1260 и 1309 S. cerevisiae, приводящей к прототрофии по лейцину (Leu+), в соответствии с протоколом трансформации сферопластов (Hinnen et al., Proc.Natl. Acad. Sci., USA, 75:1929-33, 1978). Трансформанты отбирали на синтетической агаровой среде, не содержащей лейцина и содержащей 1М сорбит. Полную и минимальную синтетическую агаровую среду, не содержащую лейцина и содержащую 1М сорбит, получали от REMEL, Lenexa, KS (cat # 09459 и 92155 соответственно). Клональные Leu+-изоляты получали путем серийного размножения на SD-чашках без лейцина (KD MEDICAL, Columbia MD).
Множество трансформантов скринировали на продуцирование области ORF0657nН в условиях ферментации, описанных в примере 9. Клеточные лизаты анализировали на продуцирование области ORF0657nН с помощью Вестерн-блот-анализа или с помощью анализа в ДСН-ПААГ-гелях, окрашенных кумасси синим.
Для Вестерн-блот-анализа образцы подвергали электрофорезу в 4-15% градиенте трис-HCl-гелей Criterion (BIO-RAD, Hercules, CA) в 1х трис-глициновом буфере с ДСН (BIO-RAD) в условиях восстановления и денатурации. Гели подвергали электроблоттингу на 0,2-микронных Мембранных ПВДФ-фильтрах. Белки подвергали иммунодетекции с помощью Вестерн-блоттинга с использованием моноклонального антитела 2Н2В8 против полноразмерного ORF0657n (SEQ ID NO:28) в качестве "первого" антитела и конъюгированного с пероксидазой хрена целого козьего антитела против мышиных IgG (Н+L)(ZYMED LABORATORIES, South San Francisco, CA) в качестве "второго" антитела. Mab 2Н2В8 описано в примере 9. Фильтры обрабатывали с использованием набора для хемилюминесценции WESTERN LIGHTNINGTm Chemiluminescence Reagent Plus Kit (PERKIN ELMER, Wellesley, MA).
Для анализа на экспрессию рекомбинантной дрожжевой области ORF0657nН (кодируемой SEQ ID NO:3) путем кумасси-окрашивания ДСН-ПААГ-гелей образцы подвергали электрофорезу в 4-15% градиенте Трис-HCl-гелей Criterion (BIO-RAD) в 1х трис-глициновом буфере с ДСН (BIO-RAD) в условиях восстановления и денатурации. Гели окрашивали биологически безопасным красителем кумасси (Bio-Safe) G250 в соответствии с протоколом производителя (BIO-RAD).
Образцы дрожжевых клеточных лизатов, подвергаемых электрофорезу, содержали 0,5 и 1,25 мкг общего клеточного дрожжевого белка соответственно для Вестерн-блот-анализа и для окрашивания кумасси. В качестве количественного стандарта для окрашивания кумасси использовали очищенный BSA (100X, NEW ENGLAND BIOLABS, Beverly, MA). В качестве количественного стандарта для Вестерн-блот-анализа использовали очищенную из E.coli полноразмерную рекомбинантную область ORF0657n (SEQ ID NO:28). Качественным стандартом для обоих анализов был клеточный лизат, выделенный из культуры E.coli, продуцирующей область ORF0657nН (SEQ ID NO:4 с His-меткой на карбокси-конце), индуцированную для получения ORF0657nН.
Для продуцирования области ORF0657nН (SEQ ID NO:4 с His-меткой на карбокси-конце) в E.coli, культуру-продуцент культивировали в течение ночи на бульоне LB, содержащем 50 мкг/мл канамицина при 37оС. На следующий день 500 мкл ночной культуры использовали для инокуляции 5,0 мл бульона LB плюс 50 мкг на мл канамицина. Эту культуру выращивали при 37оС в течение приблизительно 3 часов до тех пор, пока OD600 не достигала 0,6. Экспрессию индуцировали 1 мМ IPTG в течение 3,5 часов при 37оС. Эти клетки собирали и клеточный осадок хранили при -80°С. Лизат E.coli получали с использованием реагента для экстракции белка (Bugbuster)(NOVAGEN, Madison, WI) в соответствии с протоколом производителя. Для оценки размера белка параллельно проводили электрофорез предварительно окрашенных стандартов с массой в пределах 10-250 кДа и лизатов (BIO-RAD).
Исходя из начального скрининга дрожжевых трансформантов, один трансформант дрожжевого штамма 1260, обозначенный 1-1, отбирали как наилучший продуцент области ORF0657nН (SEQ ID NO:3). Пример продуцирования области ORF0657nН после 72-часовой ферментации в комплексной среде YEHDG в шейкерных колбах проиллюстрирован на фиг.7А и 7В. Мажорный белок, детектированный с помощью окрашивания кумасси (А) или антителом (В), имел молекулярную массу ˜85 кДа (см. дорожки 5, 6 и 7) и мигрировал вместе с ORF0657nН E.coli (SEQ ID NO:4, с карбокси-концевой His-меткой; дорожка 2). Молекулярная масса ORF0657n, экспрессируемого в E.coli, и ORF0657n, экспрессируемого в дрожжах, была выше, чем масса, предсказанная в анализе, проведенном авторами с использованием системы гель-электрофореза. Детекция ˜85 кДа-белка была специфической, то есть этот белок не детектировался в клеточном лизате трансформанта, содержащего только вектор рGAL110 (дорожка 3). Область ORF0657nН (SEQ ID NO:3) продуцировалась в значительно большем количестве, чем полноразмерная область ORF0657n (SEQ ID NO:28, без карбокси-концевой His-метки), ср. дорожки 5, 6 и 7 с дорожкой 4. Трансформант, содержащий полноразмерный ORF0657n, ферментировали одновременно с трансформантом, содержащим область ORF0657nН (SEQ ID NO:3), в идентичных условиях. При этом наблюдалась менее заметная деградация зрелой экспрессируемой в дрожжах области ORF0657nН.
Для оценки стабильности белка образец клеточного лизата трансформанта 1-1, который замораживали на несколько дней, но затем не оттаивали, был включен в гель (фиг.7, дорожка 7). Можно видеть, что целостность и количество этого образца аналогичны целостности и количеству свежих образцов (дорожки 5 и 6). Отсутствие деградации и стабильности области ORF0657nН (SEQ ID NO:3) подтверждали с помощью Вестерн-блот-анализа с использованием козьего поликлонального антитела, вырабатываемого против полноразмерного ORF0657n E.coli (SEQ ID NO:28)(данные не приводятся).
Оценка количества области ORF0657nН (SEQ ID NO:3) с помощью Вестерн-блот-анализа и окрашивания геля кумасси показала, что это количество составляло ˜360 мкг/мл среды YEHD, а % общего белка составлял ˜50%. Количество этой области, продуцируемое в определенной среде, составляло 225 мкг/мл культуральной среды, а % общего белка составлял ˜50%. Количество области ORF0657nН (SEQ ID NO:3) (350 мкг/мл) и % общего белка (50%) были выше, чем количество полноразмерной области ORF0657n (SEQ ID NO:28, без карбокси-концевой His-метки)(10 мкг/мл) и % общего белка (2,0%).
Пример 12. Внутриклеточная экспрессия области ORF0657nG (SEQ ID NO:44) в S. cerevisiae
В нижеследующем примере оценивали внутриклеточную экспрессию ORF0657n-родственных полипептидов с использованием ДНК-конструкции, кодирующей область ORF0657nG (SEQ ID NO:44). ORF0657nG (SEQ ID NO:44) представляет собой усеченный вариант SEQ ID NO:2, в котором отсутствует амино-концевая сигнальная последовательность. В SEQ ID NO:44 сохраняются N-концевой метионин и аминокислоты 42-645 SEQ ID NO:2.
ДНК, кодирующую область ORF0657nG (SEQ ID NO:44), оптимизированную для экспрессии в дрожжах, амплифицировали из pUnkCR1 (пример 7) с использованием следующих смыслового и антисмыслового праймеров соответственно 5'-GGGGGGATCCCACAAAACAAAATGGCTGAAGAAACTGGTGG-3' (SEQ ID NO:102) и 5'-GGGGGGGATCCTTAGTTCTTTCTCTTTCTTGG-3' (SEQ ID NO:103), содержащих фланкирующие рестрикционные BamHI-сайты. Смысловой праймер содержал 5'-нетранслируемую лидерную последовательность и кодон ATG, а антисмысловой праймер содержал ТАА в качестве стоп-кодона. Полученный 1,8 т.п.н-продукт выделяли из геля и клонировали в вектор рСR2.1 ТОРО ТА (INVITROGEN CORPORATION, Carlsbad, CA) в соответствии с инструкциями производителей и получали плазмиду pCR_iUC-L4. Анализ ДНК-последовательности подтвердил присутствие одной делеции у 5'-конца.
Клон со скорректированной последовательностью получали путем замены 1,3 т.п.н.-HindIII-HindIII-фрагмента pCR_iUC-S2.2, содержащего скорректированную последовательность (см. пример 11), на соответствующий фрагмент pCR_iUC-L4. 1,3 т.п.н-фрагмент pCR_iUC-S2.2 и 4,4 т.п.н.-фрагмент pCR_iUC-L4 выделяли из геля и лигировали. Клон с нужными фрагментами pCR_iUC-L отбирали и соответствующую ориентацию и нуклеотидную последовательность подтверждали с помощью секвенирования ДНК. Затем 1,8 т.п.н.-BamHI-фрагмент субклонировали в рGAL110 с получением конструкции piUC-L(-) и ДНК-последовательность подтверждали.
Плазмидную ДНК от piUC-L(-) использовали для трансформации штамма 1260 S. cerevisiae, приводящей к прототрофии по лейцину (Leu+), как описано в примере 9. Несколько дрожжевых трансформантов скринировали на внутриклеточное продуцирование области ORF0657nG (SEQ ID NO:44) в условиях ферментации и разрушения клеток, описанных в примере 11. 0,25-0,5 мкг белка клеточного лизата анализировали на продуцирование области ORF0657nG с помощью Вестерн-блот-анализа, описанного в примере 11. Выделенную из E.coli область ORF0657nН (SEQ ID NO:4 с карбокси-концевой His-меткой) использовали в качестве количественного стандарта, а клеточный лизат, выделенный из культуры E.coli, продуцирующей область ORF0657nG (SEQ ID NO:44 с карбокси-концевой His-меткой) и стимулированной для продуцирования области ORF0657nG, использовали как качественный стандарт. Образцы подвергали электрофорезу в 4-15% градиенте трис-HCl-гелей Criterion (BIO-RAD, Hercules, CA) в 1х трис-глициновом буфере с ДСН (BIO-RAD) в условиях восстановления и денатурации. Гели подвергали электроблоттингу на 0,2-микронных мембранных ПВДФ-фильтрах (BIO-RAD).
Белки подвергали иммунодетекции с помощью Вестерн-блоттинга с использованием поликлонального антитела против выделенного из E.coli полноразмерного ORF0657n (SEQ ID NO:28, без карбокси-концевой His-метки) в качестве "первого" антитела и конъюгированного с пероксидазой хрена целого козьего антитела против мышиных IgG (Н+L)(ZYMED LABORATORIES, South San Francisco, CA) в качестве "второго" антитела. Это поликлональное антитело генерировали путем иммунизации выделенной из E.coli полноразмерной областью ORF0657n (SEQ ID NO:28). Фильтры обрабатывали с использованием набора для хемилюминесценции WESTERN LIGHTNINGTm Chemiluminescence Reagent Plus Kit (PERKIN ELMER, Wellesley, MA).
Для сравнения, область ORF0657nG (SEQ ID NO:44 с карбокси-концевой His-меткой) клонировали в экспрессирующий вектор E.coli, как описано в примере 1, и экспрессировали. Для продуцирования области ORF0657nG в E.coli культуру-продуцент культивировали в течение ночи на бульоне LB, содержащем 50 мкг/мл канамицина, при 37оС. На следующий день 500 мкл ночной культуры использовали для инокуляции 5,0 мл бульона LB плюс 50 мкг на мл канамицина. Эту культуру выращивали при 37оС в течение приблизительно 3 часов до тех пор, пока OD600 не достигала 0,6. Экспрессию индуцировали 1 мМ IPTG в течение 3,5 часов при 37оС. Эти клетки собирали и клеточный осадок хранили при -80°С. Лизат E.coli получали с использованием реагента для экстракции белка (Bugbuster)(NOVAGEN, Madison, WI) в соответствии с протоколом производителя. Для оценки размера белка параллельно проводили электрофорез предварительно окрашенных стандартов с массой в пределах 10-250 кДа (BIO-RAD) и лизатов.
Исходя из начального скрининга дрожжевых трансформантов, один трансформант дрожжевого штамма 1260, обозначенный iUC-L3, был отобран как хороший продуцент области ORF0657nG после 72-часовой ферментации в комплексной среде YEHDG в колбах для культивирования, как описано в примере 11. Мажорный белок, детектированный либо с помощью окрашивания кумасси, либо антителом, имел молекулярную массу ˜ 85 кДа и мигрировал вместе с экспрессируемой в E.coli областью ORF0657nG (SEQ ID NO:44, с карбокси-концевой His-меткой). Молекулярная масса области ORF0657nG, экспрессируемой в E.coli, и области ORF0657nG, экспрессируемой в дрожжах, была выше, чем масса, предсказанная в анализе, проведенном авторами с использованием системы гель-электрофореза. Детекция ˜85 кДа-белка была специфической, то есть этот белок не детектировался в клеточном лизате трансформанта, содержащего только вектор рGAL110. Средний титр ORF0657nG, определенный после проведения трех Вестерн-блот-анализов, составлял 30 мкг/мл среды YEHDG, а оцененный % общего белка составлял 10,0 (см. ниже пример 13).
Пример 13: Удаление сигнала сортировки белков клеточной стенки приводит к повышению уровня внутриклеточной экспрессии
Эффект удаления одной С-концевой области ORF0657n, кодирующей клеточную сортазу, и в комбинации с удалением N-концевой сигнальной области оценивали с использованием оптимизированных по кодонам последовательностей генов ORF0657n, SEQ ID NOs: 31, 32 и 45. SEQ ID NO:31 кодирует полноразмерную область ORF0657n. SEQ ID NO:32 кодирует область ORF0657nН, в которой отсутствуют N-концевая сигнальная последовательность и С-концевая область сортировки белков клеточной стенки. SEQ ID NO:45 кодирует область ORF0657nG, в которой отсутствует только сигнальная пептидная последовательность.
Один трансформант, экспрессирующий полноразмерную область ORF0657n (SEQ ID NO:28 без карбокси-концевой His-метки; трансформант rUnkC1), один трансформант, экспрессирующий область ORF0657nН (SEQ ID NO:3, трансформант 1-1), и один трансформант, экспрессирующий область ORF0657nG (SEQ ID NO:44, трансформант iUC-L3), подвергали ферментации и получали клеточные лизаты, которые анализировали, как описано в примере 11. Репрезентативный Вестерн-блот-анализ проиллюстрирован на фиг.9. Количество белка клеточного лизата, загруженного на гель от различных трансформантов, не было одинаковым, и это было сделано для того, чтобы компенсировать различные уровни экспрессии трех конструкций, кодирующих области ORF0657n. В каждом случае были загружены различные количества по крайней мере двух белков клеточного лизата от каждого из ORF0657n-содержащих трансформантов.
Типичные результаты, указывающие на то, что область ORF0657nН (SEQ ID NO:3) экспрессировалась на гораздо более высоком уровне, чем полноразмерная область ORF0657n (SEQ ID NO:28 без карбокси-концевой His-метки) или область ORF0657nG (SEQ ID NO:44), представлены на фиг.9. Заметный сигнал детектировался лишь от 50 и 100 нг белка клеточного лизата продуцентов области ORF0657nН, как показано на дорожках 11 и 12. Для сравнения использовали 1,0 и 2,0 мкг белка клеточного лизата продуцента полноразмерной области ORF0657nС, дорожки 7 и 8, соответственно. Хороший сигнал от области ORF0657nG детектировался с меньшим количеством белка, при загрузке 250 и 500 нг белка, как видно на дорожках 14 и 15.
В таблице 5 проиллюстрировано сравнение среднего титра каждой из трех ОРС и % общего белка, определенных от трех независимых Вестерн-блотов. Для каждого Вестерн-блота получали свежие клеточные лизаты. Средние величины представляют собой результаты от двух независимых ферментаций. Количество специфического ORF0657n определяли по отношению к очищенному стандарту ORF0657nН E.coli (SEQ ID NO:4 с карбокси-концевой His-меткой).
Удаление сигнальной пептидной последовательности приводило к увеличению % общего белка и к 2,5-3-кратному увеличению титра, как было определено путем сравнения экспрессии полноразмерной области ORF0657n (SEQ ID NO:28 без карбокси-концевой His-метки) и области ORF0657nG (SEQ ID NO:44). Удаление сигнального пептида и сигнала сортировки белков клеточной стенки приводило к 8-14-кратному увеличению уровня экспрессии по сравнению с удалением только сигнальной пептидной последовательности (при сравнении областей G и Н) и к 20-42,5-кратному увеличению уровня экспрессии по сравнению с удалением полноразмерного белка (при сравнении областей С и Н). Величина повышения уровня экспрессии после удаления последовательности сортировки белков клеточной стенки не была предсказана.
Пример 14: Внутриклеточная экспрессия области ORF0657nI в S. cerevisiae
ДНК, кодирующую область ORF0657nI с кодонами, оптимизированными для экспрессии в дрожжах (SEQ ID NO:33), амплифицировали из pUnkCR1 (пример 7) с использованием следующих смыслового и антисмыслового праймеров соответственно: 5'-CTCCGGATCCCACAAAACAAAATGGCTGAAGAAACTGGT-3' (SEQ ID NO:104) и 5'-GCTGCCGGGATCCTTATGGGGTTGGCTTAGATGGGGTAG-3' (SEQ ID NO:105), содержащих фланкирующие рестрикционные BamHI-сайты (подчеркнуты). Смысловой праймер содержит 5'-нетранслируемую лидерную последовательность и кодон ATG, а антисмысловой праймер содержит ТАА в качестве стоп-кодона. Полученный 1,3 т.п.н-продукт выделяли из геля и клонировали в вектор рGAL110 (фиг.5А) с получением конструкции piUC-I (фиг.5В) и ДНК-последовательность подтверждали.
Плазмидную ДНК от piUC-I использовали для трансформации штамма 1260 S. cerevisiae, приводящей к прототрофии по лейцину (Leu+), как описано в примере 9. Три Leu+-трансформанта скринировали на внутриклеточное продуцирование области ORF0657nI (SEQ ID NO:1) в лабораторных условиях ферментации (5,0 мл культуры), как описано в примере 9.
Продуцирование указанной области сравнивали с продуцированием области ORF0657nН (SEQ ID NO:3) трансформантом 1-1 S. cerevisiae (см. пример 11). Клеточные лизаты получали, как описано в примере 9, и 100 и 200 нг белка дрожжевого клеточного лизата анализировали на продуцирование ORF0657nI и ORF0657nН с помощью полуколичественного Вестерн-блот-анализа, описанного в примере 11. Выделенный из E.coli ORF0657nН (SEQ ID NO:4 с карбокси-концевой His-меткой) использовали в качестве количественного стандарта. Образцы подвергали электрофорезу в 10-20% градиенте трис-HCl-гелей Criterion (BIO-RAD, Hercules, CA) в 1х трис-глициновом буфере с ДСН (BIO-RAD) в условиях восстановления и денатурации. Гели подвергали электроблоттингу на 0,2-микронных мембранных ПВДФ-фильтрах (BIO-RAD). Белки подвергали иммунодетекции с помощью Вестерн-блоттинга с использованием поликлонального антитела против выделенного из E.coli полноразмерного ORF0657n (SEQ ID NO:28) в качестве "первого" антитела и конъюгированного с пероксидазой хрена целого козьего антитела против мышиных IgG (Н+L)(ZYMED LABORATORIES, South San Francisco, CA) в качестве "второго" антитела. Поликлональное антитело генерировали путем иммунизации выделенной из E.coli полноразмерной областью ORF0657n (SEQ ID NO:28). Фильтры обрабатывали с использованием набора для хемилюминесценции WESTERN LIGHTNINGTm Chemiluminescence Reagent Plus Kit (PERKIN ELMER, Wellesley, MA). Для оценки размера белка параллельно проводили электрофорез предварительно окрашенных стандартов с массой в пределах 10-250 кДа и (BIO-RAD) лизатов.
На фиг.11 представлен результат 72-часовой ферментации в комплексной среде YEHDG в пробирках для культивирования. Был детектирован сильный сигнал экспрессируемой в дрожжах области ORF0657nН (SEQ ID NO:3), как показано на дорожках 5, 6, 14, 15, 21 и 22. ˜60 кДа-белок в трансформантах, сконструированных для экспрессии ORF0657nI (SEQ ID NO:1), также легко детектировался, как показано на дорожках 7-12 и 16-21. Экспрессируемый в дрожжах ORF0657nI (SEQ ID NO:1) мигрировал вместе с областью ORF0657nI, экспрессируемой и выделенной из E.coli (SEQ ID NO:5 с карбокси-концевой His-меткой) (данные не приводятся). Молекулярная масса области ORF0657nI, экспрессируемой в E.coli, и области ORF0657nI, экспрессируемой в дрожжах, была выше, чем масса, предсказанная в анализе, проведенном авторами с использованием системы гель-электрофореза. 60 кДа-белок, детектированный в трансформантах, сконструированных для экспрессии области ORF0657nI (SEQ ID NO:1), был специфическим, то есть этот белок не детектировался в клеточном лизате от контрольного трансформанта, содержащего только вектор (дорожки 4 и 13).
Количество областей ORF0657nН (SEQ ID NO:3) и ORF0657nI (SEQ ID NO:1) на геле оценивали с помощью полуколичественного Вестерн-блот-анализа путем сравнения с известным количеством очищенного ORF0657nН (SEQ ID NO:4 с карбокси-концевой His-меткой). Титры и процент общего белка вычисляли как среднюю величину, определенную исходя из лизатов, взятых в дубликате, содержащих два различных количества белка в геле. Волюметрические титры области ORF0657nН (SEQ ID NO:3), определенные для свежеферментированного образца и замороженного клеточного лизата, были сравнимыми, т.е. они составляли ˜500 и ˜550 мкг/мл культуральной среды, а титр области ORF0657nI (SEQ ID NO:1) составлял 320 мкг на 1 мл культуральной среды (титр области ORF0657nI для трансформанта II). Вычисленный процент области ORF0657nН (SEQ ID NO:3) составлял 78% и 80% от общего белка соответственно, а процент области ORF0657nI (SEQ ID NO:1) составлял 45% от общего белка.
Таким образом, область ORF0657nI хорошо экспрессировалась в S. cerevisiae, при этом ее титр был примерно в 1,5 раз ниже титра области ORF0657nН (SEQ ID NO:3), а % общего белка был ниже примерно в 1,3 раза. Хорошая экспрессия области ORF0657nI в дрожжах была подтверждена с помощью окрашивания кумасси ДСН-ПААГ-геля, содержащего клеточные лизаты. Экспрессия области ORF0657nI в среде YEDHG была масштабируемой: продуцирование в шейкерных колбах емкостью либо 125 мл, либо 2 л было сравнимо с продуцированием при лабораторной экспрессии в культуральных пробирках.
Область ORF0657nI (SEQ ID NO:1) также хорошо экспрессировалась в определенной 5Х среде, не содержащей лейцина (в среде, описанной в примере 9). Титр этой области был всего в ˜1,5 раз ниже, чем титр указанной области в комплексной среде YEHDG, а % общего белка был сравним в обеих средах. Целостность ORF0657nI в обеих средах была хорошей, и какой-либо заметной деградации не наблюдалось. Уровень продуцирования области ORF0657nI через 72 часа был выше, чем уровень продуцирования в комплексной среде и в 5х среде без лейцина через 96 часов, как было обнаружено при тестировании в шейкерных колбах-ферментерах.
Пример 15: Крупномасштабная ферментация дрожжевого штамма, продуцирующего ORF0657nН (SEQ ID NO:3)
Замороженный посевной материал дрожжевого штамма 1-1 (штамма 1260, трансформированного плазмидой piUC-S(-), описанного в примере 11) использовали для крупномасштабной ферментации и очистки. Сосуд с посевным материалом оттаивали и 1,0 мл использовали для инокуляции 250 мл-колбы Эрленмейера, содержащей 50 мл селективной среды без лейцина (5Х Leu--среды, Bayne et al., Gene 66(2):235-44, 1988), содержащей 4% глюкозу. Содержимое колбы инкубировали при 28°С, 250 об/мин, на роторном шейкере. Через 24 часа (23,5 г/л остаточной глюкозы) в 2-литровую колбу, содержащую 877 мл той же среды, добавляли 13 мл-объем культуры. И снова колбу инкубировали при 28°С и перемешивали при 250 об/мин. Через 24 часа (4,04 г/л остаточной глюкозы) содержание 2-литровой колбы использовали для инокуляции 20-литрового реактора, содержащего среду с определенным химическим составом (Oura, Biotechnol. Bioengineer. 16:1197, 1974), которая была оптимизирована для используемых штаммов. Эта среда содержала 20 г/л глюкозы, а затем в нее добавляли 25 г/л галактозы для индуцирования. Реактор работал при 28°С, 4,7 л/мин, 15 фунт/кв.дюйм и 300 об/мин. В этих условиях растворенный кислород поддерживали на уровне насыщения более чем 30%. Затем проводили мониторинг клеточного роста на утилизацию глюкозы, оптическую плотность (А600 нм, 1 см-кюветы), сухую массу клеток, утилизацию галактозы и на продуцирование этанола. Культивирование продолжали в течение 90 часов до тех пор, пока А600 не достигала 33,9, а сухая масса клеток не составляла 17,5 г/л.
Культуру собирали путем фильтрации в тангенциальном потоке на фильтре из полого волокна (картридж AMICON H5MP01-43) и на подставке для сбора AMICON DC-10 (MILLIPORE, Billerica, MA). Затем пермеат отбрасывали и клетки концентрировали, подвергали диафильтрации PBS и собирали путем центрифугирования при 8000 об/мин, 4°С в течение 20 минут с использованием центрифуги RC Sorvall Evolution (ротор SLA-3000). Клетки хранили при -70°С.
Для оценки продуцирования ORF0657nН штаммом 1-1 при крупномасштабной ферментации клеточные лизаты получали из собранной культуры с оптической плотностью OD600 = 10 единиц, как было подробно описано в примере 11 (результаты не приводятся). Также оценивали продуцирование ORF0657nН в шейкерной колбе-ферментере (72 часа, комплексная среда YEHDG). Результаты Вестерн-блот-анализа (осуществляемого, как описано в примере 11) показали, что белок, продуцированный при крупномасштабной ферментации, мигрировал вместе с ORF0657nН, продуцированным в шейкерной колбе-ферментере (результаты не приводятся). Было определено, что титр ORF0657nН, продуцированного при крупномасштабной ферментации (20 л), составлял 739 мкг/мл, а % общего белка составлял 55% по сравнению с ферментацией в шейкерных колбах, где титр ORF0657nН составлял 732 мкг/мл, а % общего белка составлял 58% (оценка проводилась методом полуколичественного Вестерн-блот-анализа, описанного в примере 11). Эти результаты подтвердили, что продуцирование ORF0657n в дрожжах может быть увеличено.
Пример 16: Защитный иммунитет, вызываемый ORF0657n-родственными полипептидами, продуцируемыми в дрожжах
Способность ORF0657n-родственных полипептидов, продуцируемых в дрожжах, вызывать защитный иммунитет оценивали по экспрессии полипептида SEQ ID NO:3 в дрожжах. В качестве контроля использовали E.coli, которая экспрессирует полноразмерный ORF0657nС (SEQ ID NO:28), ORF0657nН (SEQ ID NO:4 с карбокси-концевой His-меткой) и ORF0657nI (SEQ ID NO:5 с карбокси-концевой His-меткой), и один лишь адъювант.
ORF0657n-родственный полипептид SEQ ID NO:3 получали из дрожжей и использовали у животного-модели для индукции защитного иммунитета. Полипептид SEQ ID NO:3 экспрессировали, как описано в примере 11.
Замороженные клетки рекомбинантного S. cerevisiae, экспрессирующего ORF0657nН (SEQ ID NO:3), ресуспендировали в 5 мл на грамм сырой клеточной массы в 0,2М MOPS, рН 7,0, с ингибиторами протеазы (без EDTA). Лизат получали путем четырех пассажей через микрофлюидизатор при 14000 фунт/кв.дюйм (Microfluidics Model 110S). Затем лизат осветляли центрифугированием (10000×g, 20 минут, 2-8°С), после чего проводили грубую фильтрацию (через стекловолоконный предварительный фильтр, Millipore) и тонкую фильтрацию (через 0,2 мкм ацетата целлюлозы, Whatman).
Осветленный лизат фракционировали на эксклюзионной хроматографической колонке (SEC)(Pharmacia HiPrep 26/60 Sephacryl S-300 HR, подвижная фаза: 0,2М MOPS, pH 7,0). Фракции анализировали с помощью электрофореза в ДСН-ПААГ с окрашиванием кумасси и Вестерн-блоттингом с использованием антисыворотки против белка ORF0657n (против полноразмерного ORF0657n, SEQ ID NO:28). Фракции, содержащие продукт, объединяли.
SEC-продукт подвергали стерильной фильтрации через 0,2 мкм ацетата целлюлозы в асептических условиях. Чистота стерильно отфильтрованного продукта составляла ≥94% или выше при оценке с помощью электрофореза в ДСН-ПААГ и Вестерн-блот-анализа. Стерильно отфильтрованный продукт разводили до 0,2 мг/мл добавлением АНР и тимерозала.
Способность ORF0657n-родственных полипептидов, продуцируемых в дрожжах, вызывать защитный иммунитет проиллюстрирована на фиг.10. Экспрессируемый в дрожжах ORF0657nН (SEQ ID NO:3) продуцировал защитный иммунитет, эквивалентный иммунитету, продуцируемому полипептидами, экспрессируемыми в E.coli.
Также были получены экспериментальные данные в эксперименте с дрожжами, экспрессирующими ORF0657nI. Дрожжи, экспрессирующие ORF0657nI, обеспечивали защиту мышей ICR, мышей BALB/C, в сочетании с эндотоксином, но не обеспечивали защиту мышам BALB/C без эндотоксина (см. таблицу 6).
Сам по себе эндотоксин не давал защитной реакции. В каждом случае в качестве адъюванта использовали гидроксифосфат алюминия.
Процедуры очистки для получения ORF0657nI E.coli давали продукт, содержащий как ORF0657nI, так и эндотоксин. Дрожжи, продуцирующие ORF0657nI, не содержали эндотоксин. Таким образом, на модели BALB/C мышей было показано, что эндотоксин дает дополнительный эффект для ORF0657nI.
Пример 17: Анамнестический ответ у приматов за исключением человека
Три группы макак резусов иммунизировали либо продуцируемым в дрожжах ORF0657n-родственным полипептидом (ORF0657nH, SEQ ID NO:3), либо продуцируемым в E.coli ОРГ0657п-родственным полипептидом (полноразмерным ORF0657n, SEQ ID NO:28), полученными с АНР или без него. Обезьянам в вакцинируемой группе вводили внутримышечно 50 мкг ORF0657n-родственных полипептидов.
Как показано на фиг.12, у животных, подвергнутых вакцинации, наблюдался ответ после введения одной дозы, при этом у этих животных наблюдалось 3-6-кратное увеличение геометрически средних титров по сравнению с низкими исходными титрами, что позволяет предположить об индуцировании анамнестического ответа. В отличие от этого геометрически средние титры у животных контрольной группы оставались на том же уровне (с некоторой вариабельностью при анализе с использованием антитела). После введения второй дозы вакцины вакцинируемой группе и контрольной группе геометрически средние титры очень мало изменялись по сравнению с титрами, которые наблюдались после введения первой дозы.
Для мониторинга индуцирования гуморального ответа после введения только одной дозы группу, которой вводили экспрессируемую в дрожжах область ORF0657nH (SEQ ID NO:3) наблюдали в течение 3 месяцев (до окончания испытания). Титры антител продолжали увеличиваться через 9 дней, достигая уровней, которые не повышались даже после введения 2 или 3 доз вакцины.
Эти наблюдения дают основание предположить, что одна доза вакцины может индуцировать значительные и устойчивые гуморальные ответы после естественной стимуляции иммунной системы, вероятно, обусловленной внешним воздействием микроорганизма S. aureus. Наблюдение исходных титров антитела у человека показало присутствие низких титров антител против ORF0657n во всех протестированных образцах (данные не приводятся).
Объем нижеследующей формулы изобретения включает и другие варианты его осуществления. Хотя настоящее изобретение проиллюстрировано и описано на нескольких его вариантах, в него могут быть внесены различные модификации, не выходящие за рамки объема и сущности изобретения.






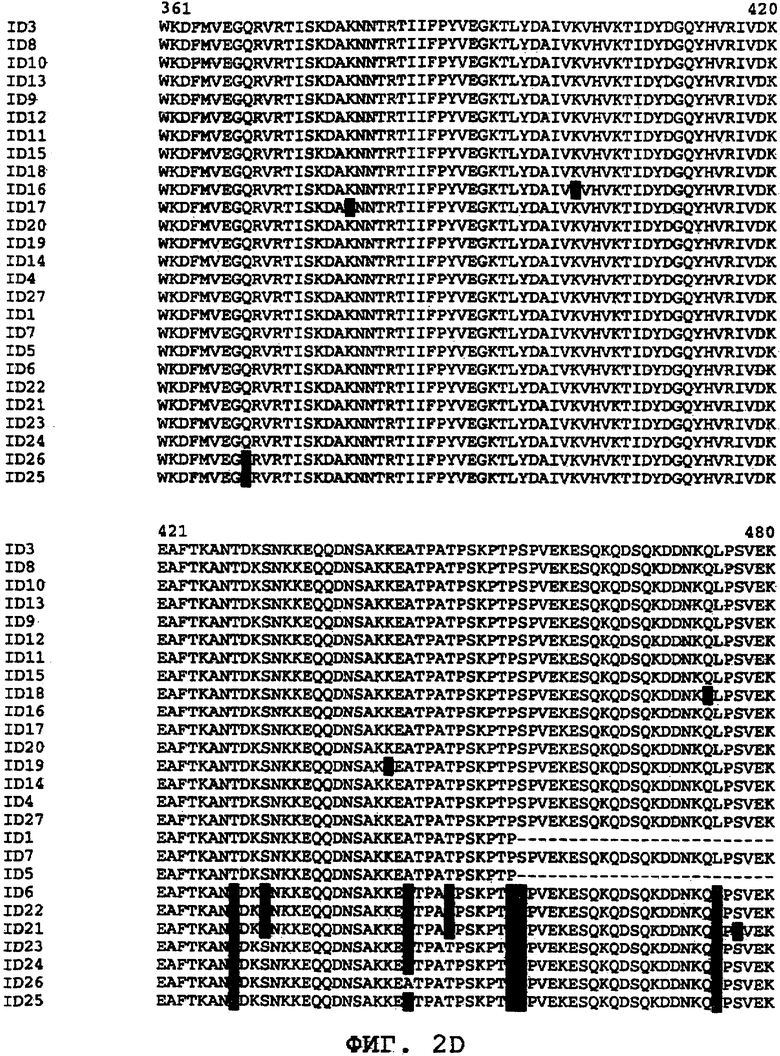
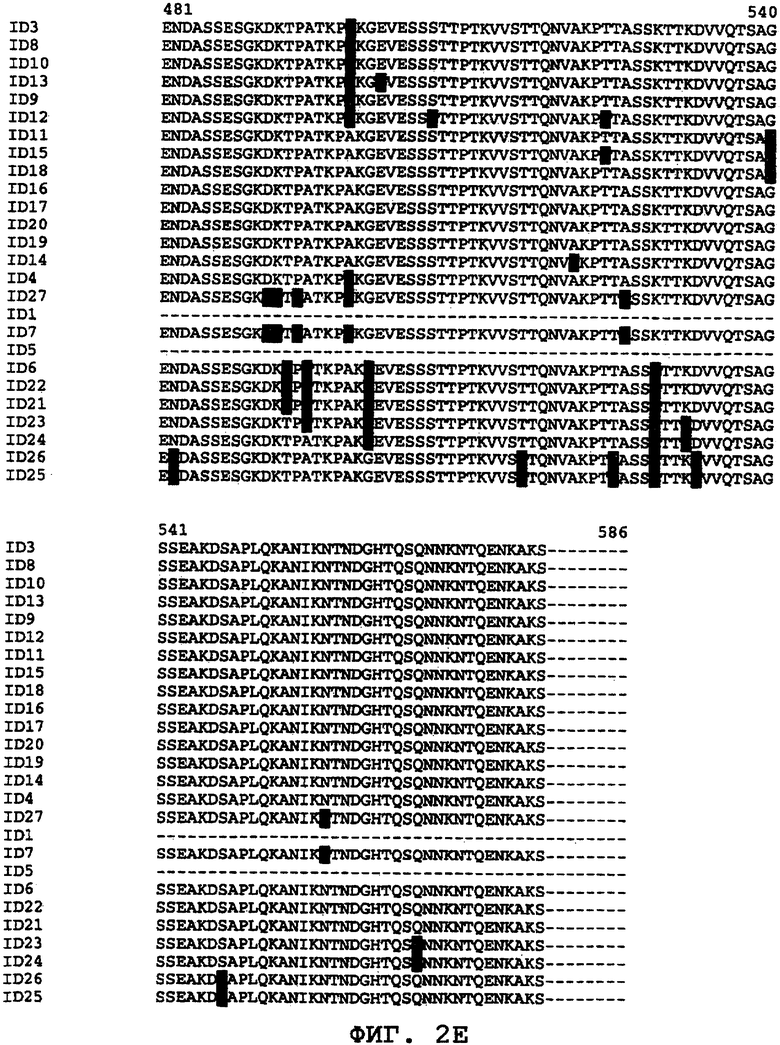
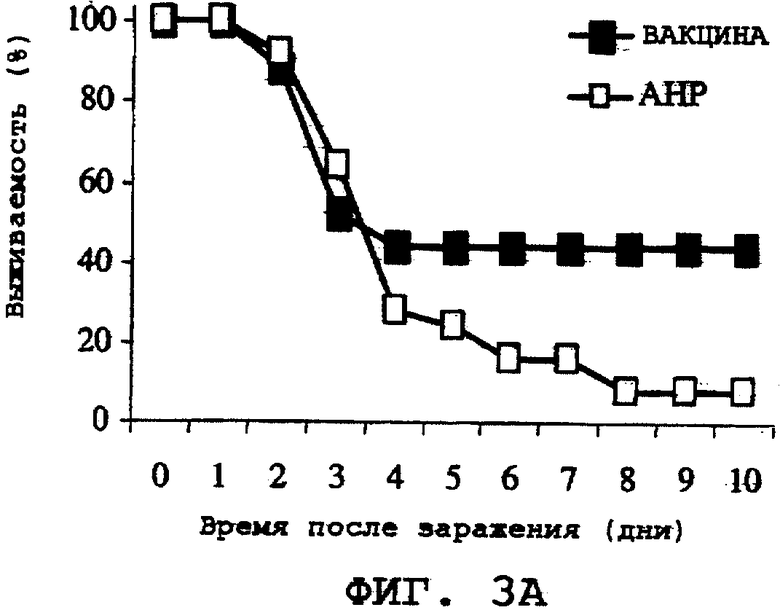
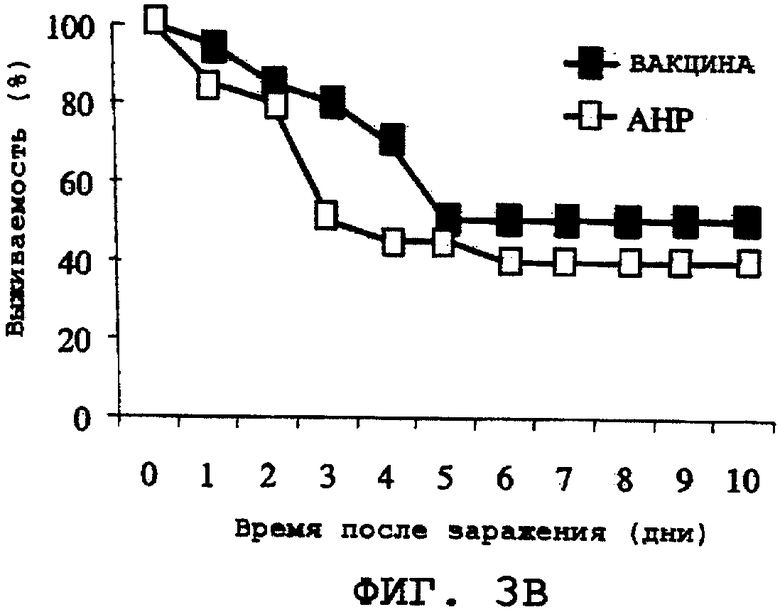


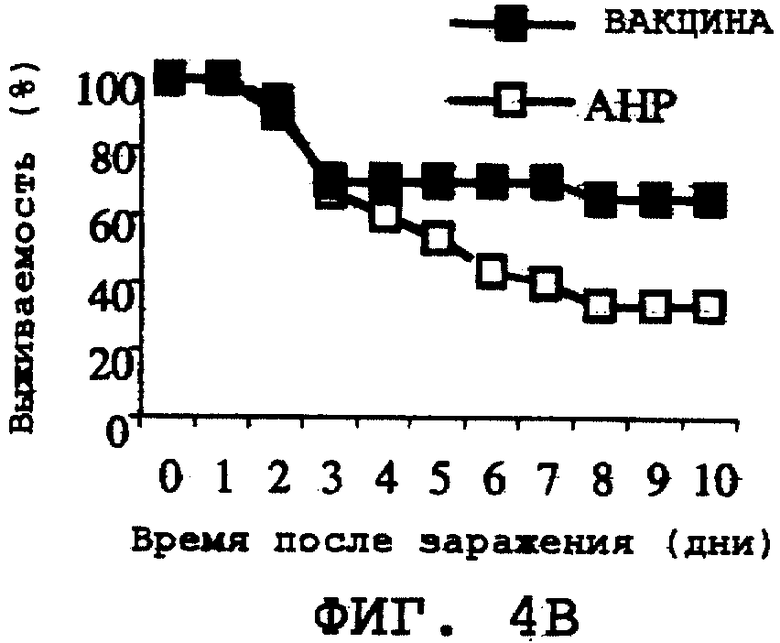
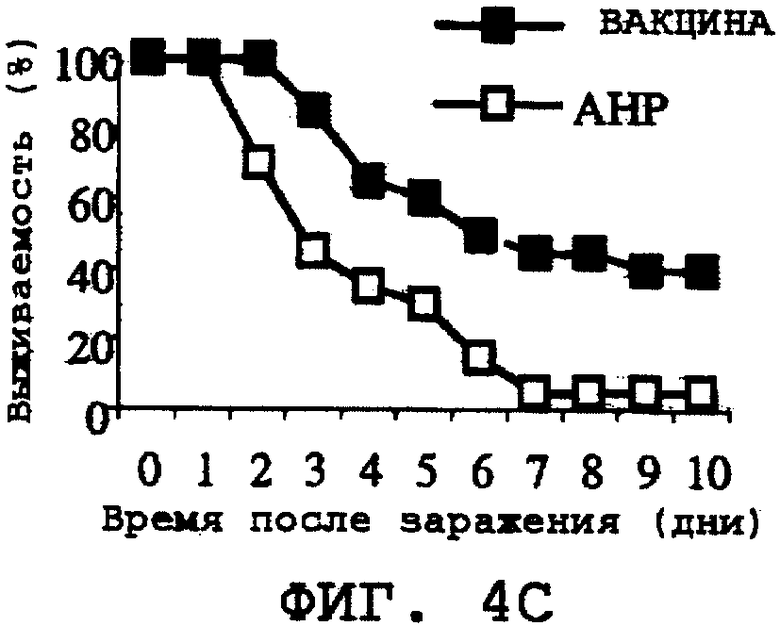

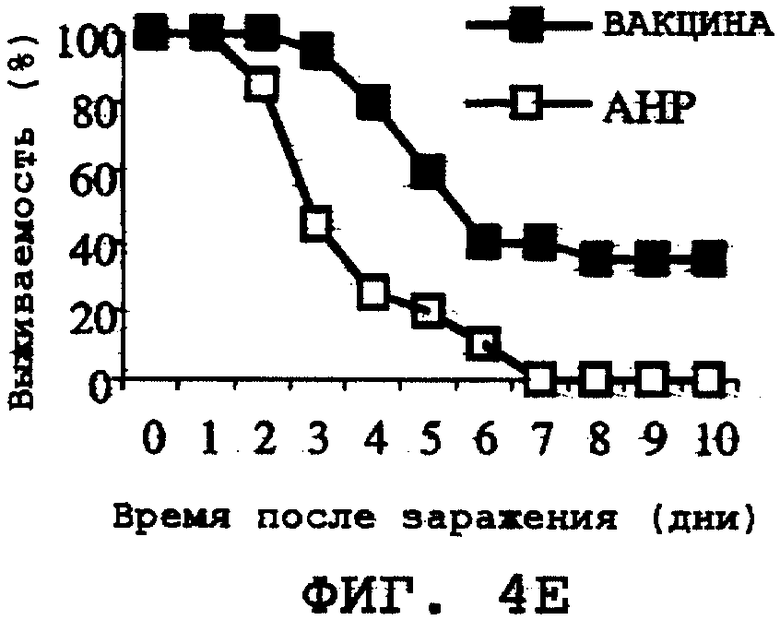
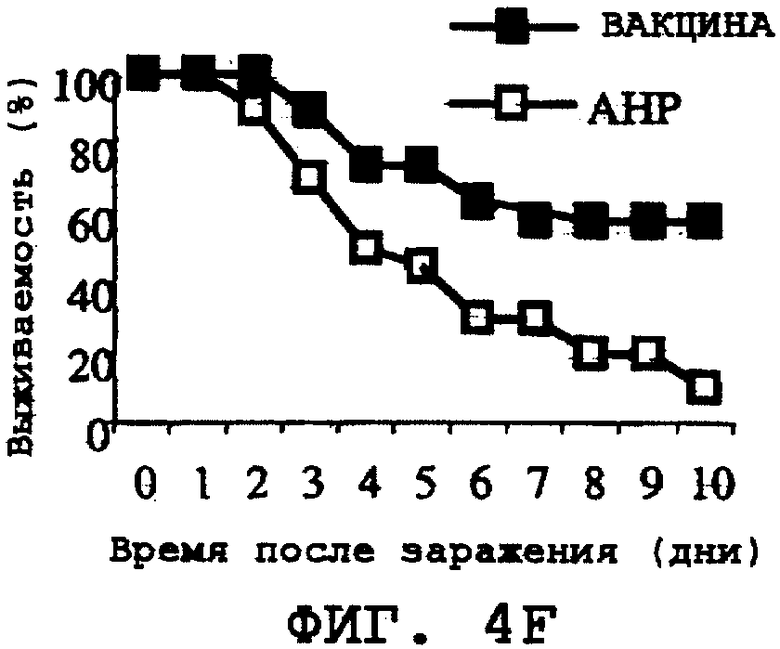
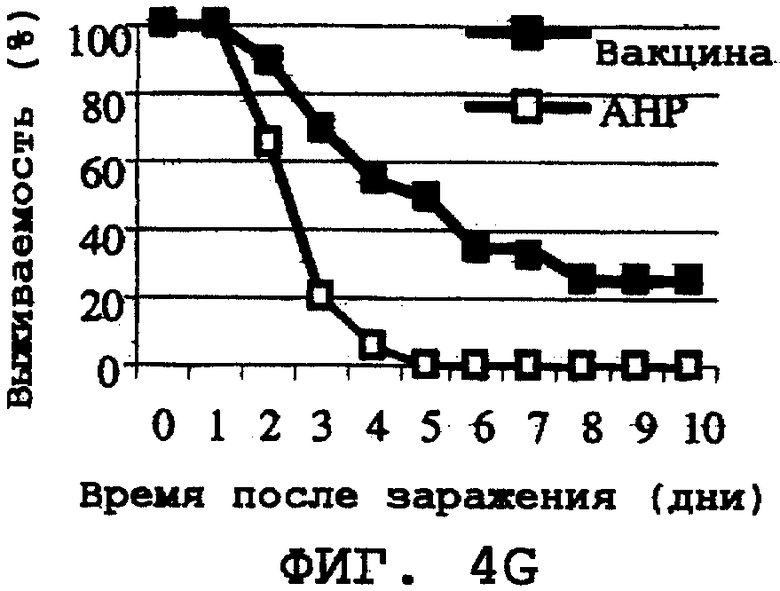
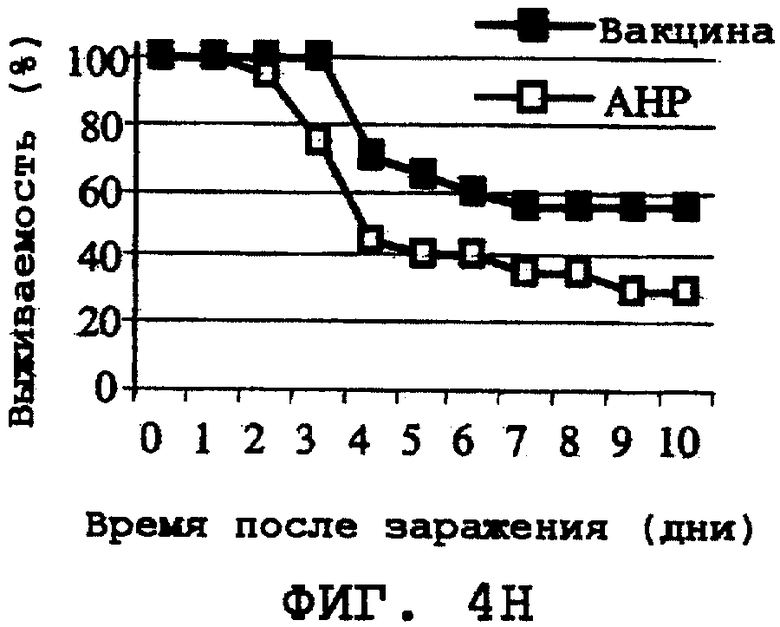

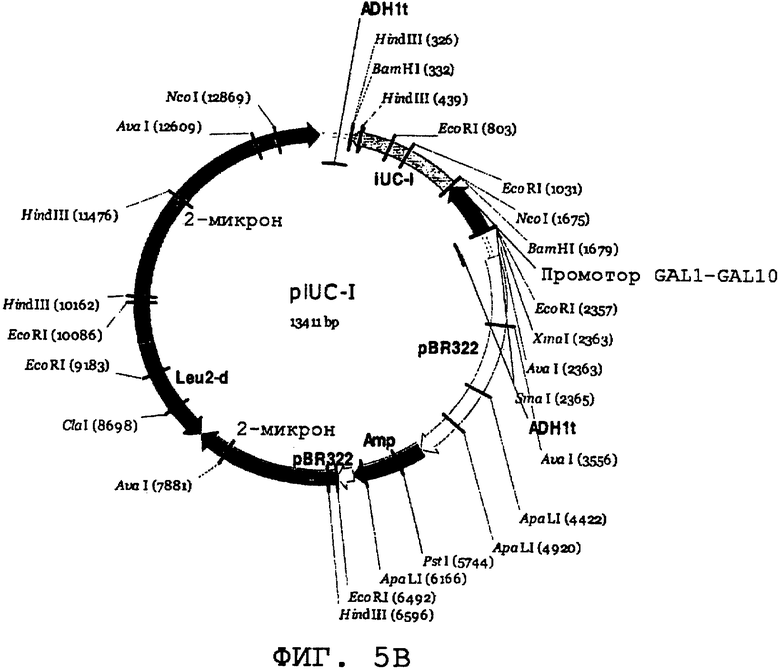
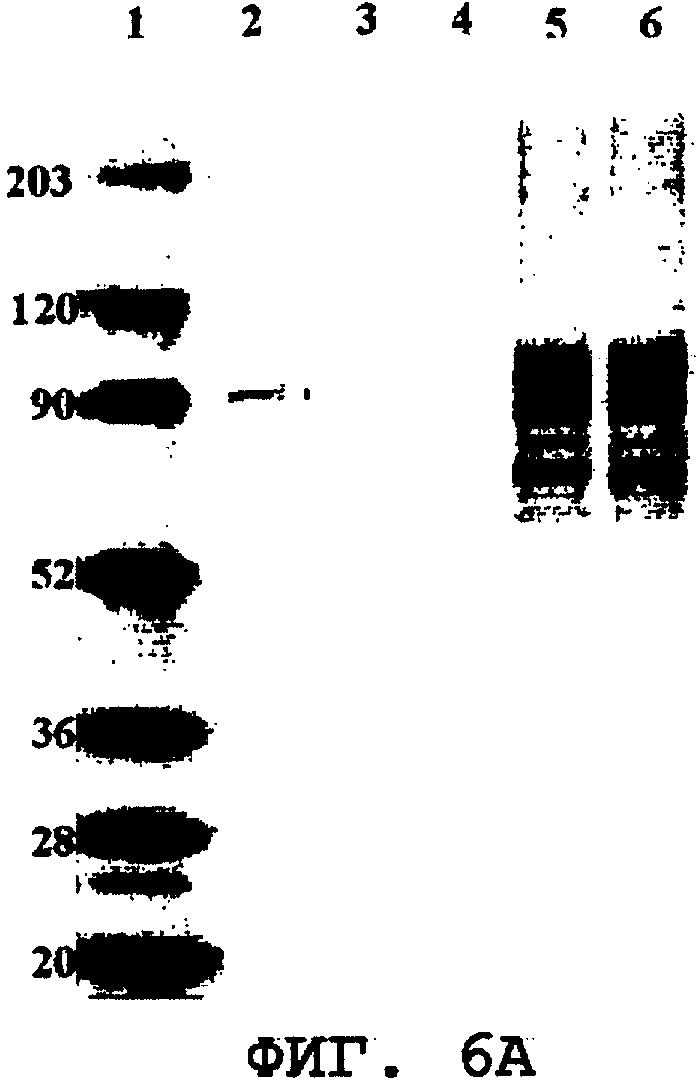

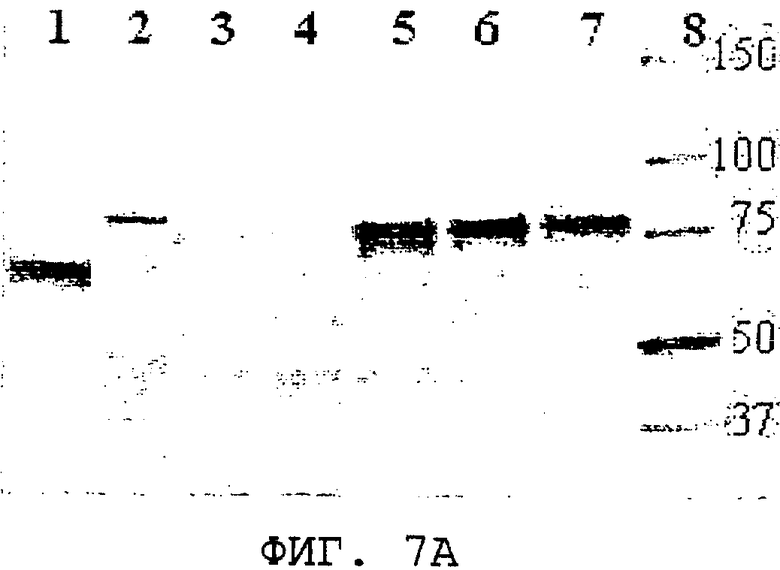
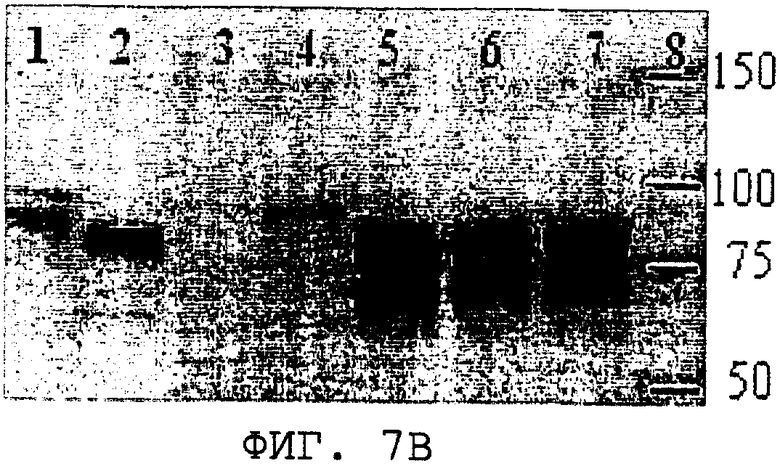
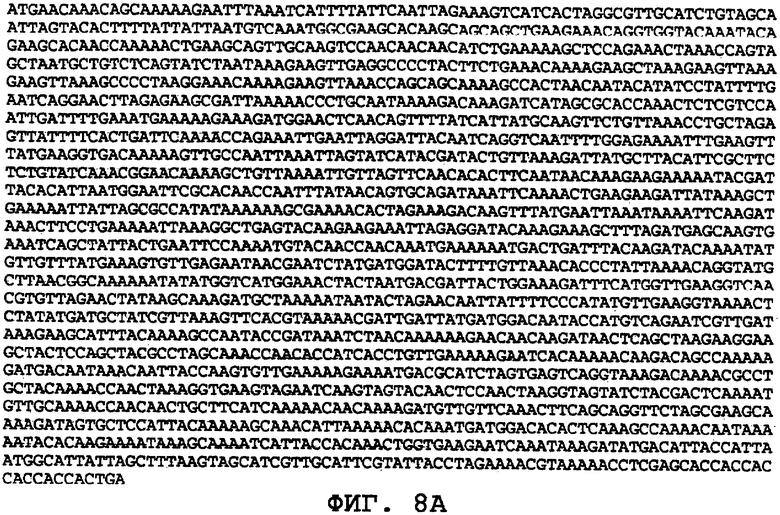
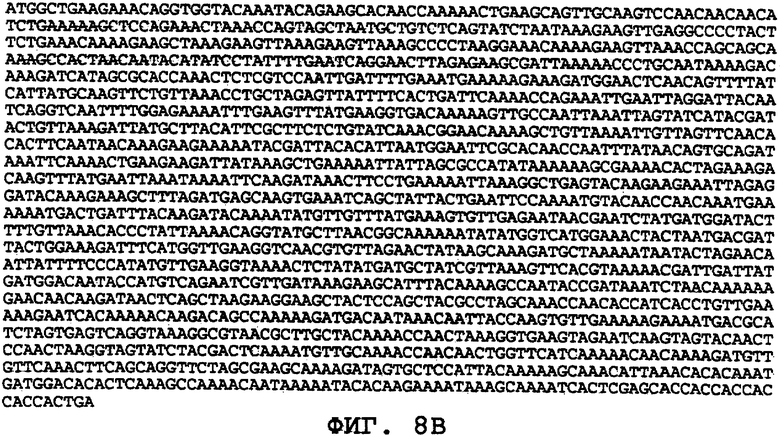






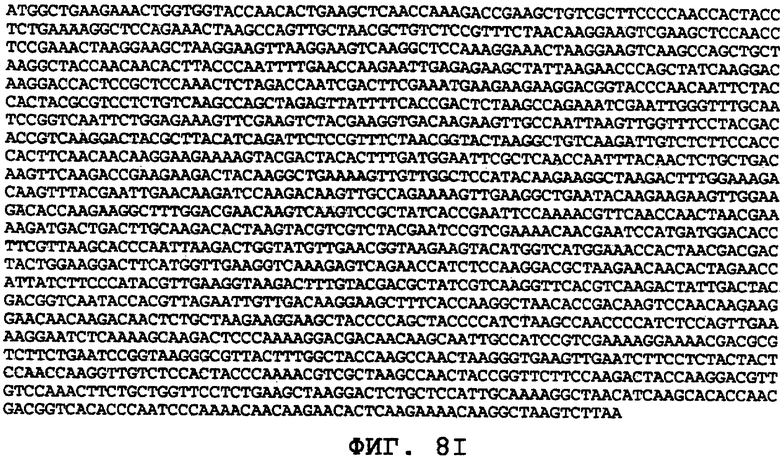




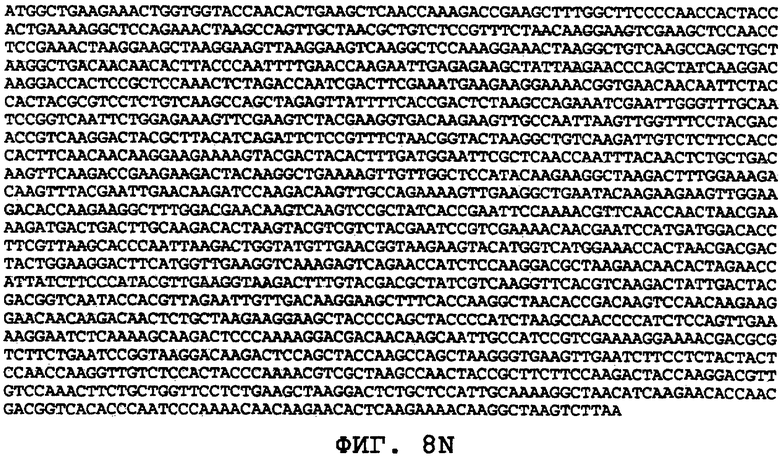
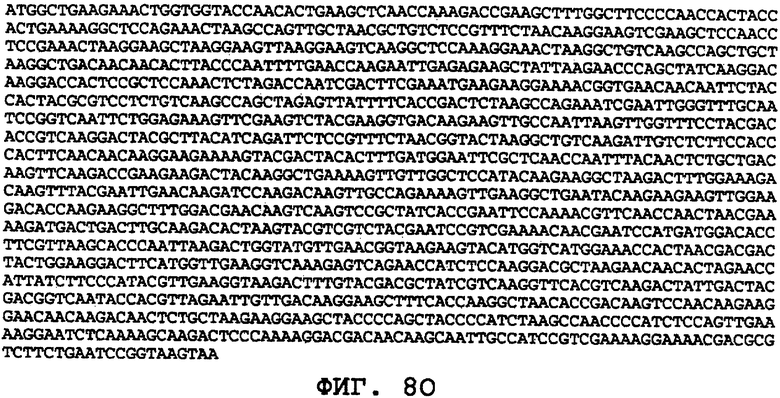
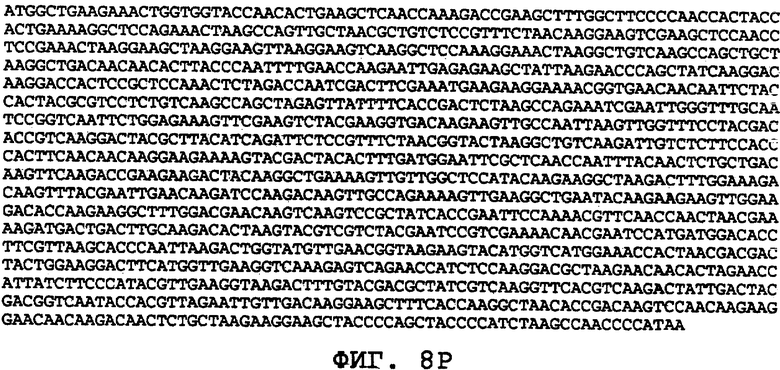
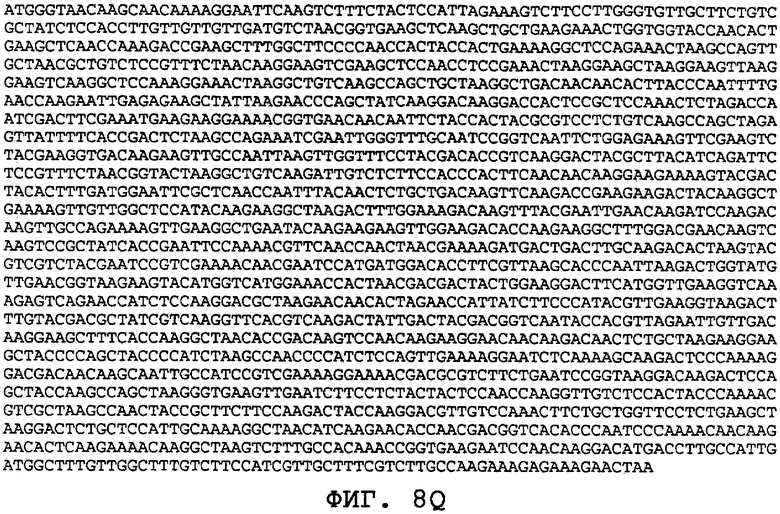

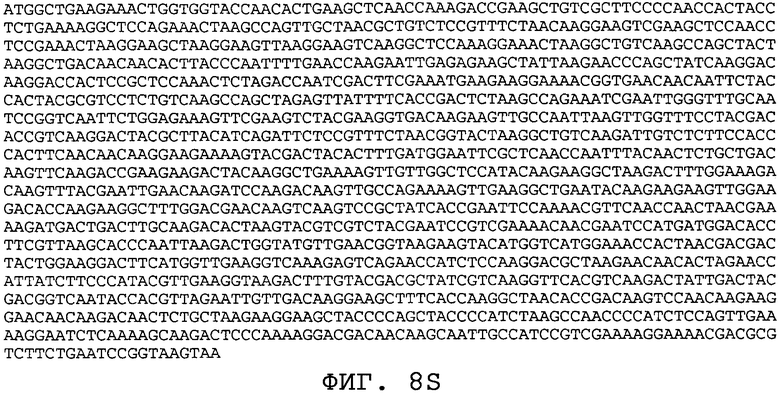
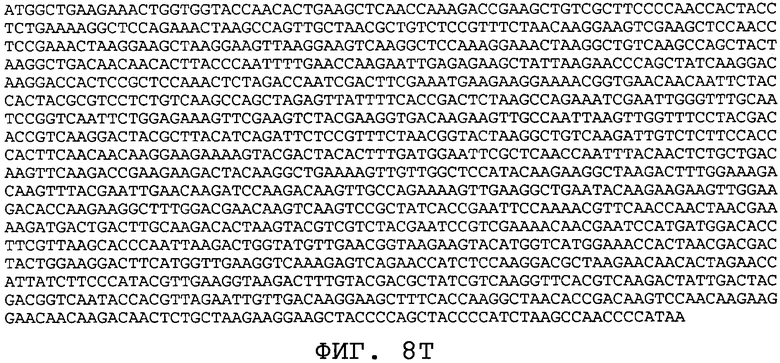

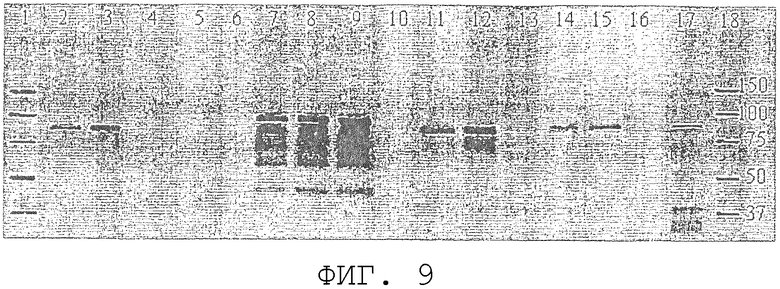
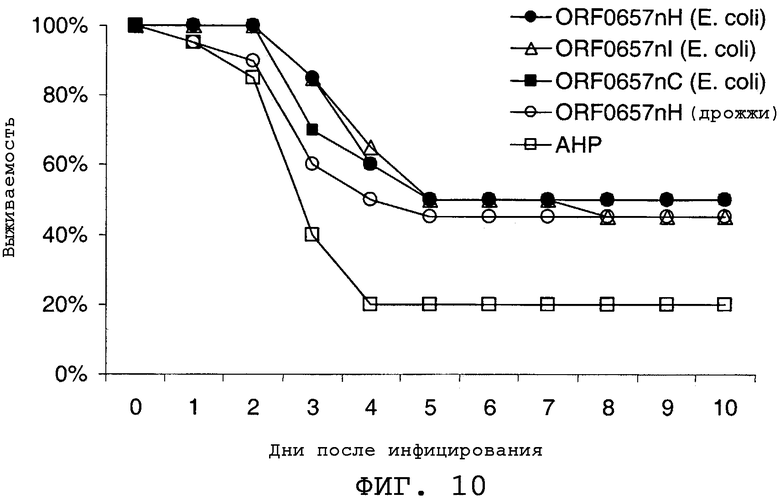
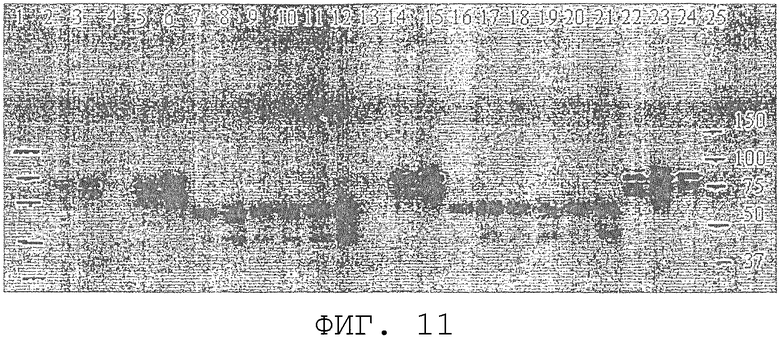
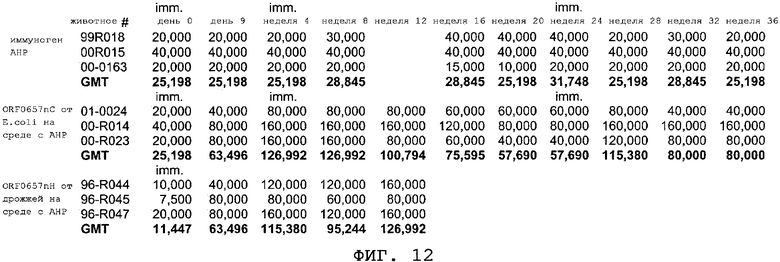


























































































Изобретение относится к полипептидным иммуногенам, способным вызывать защитный иммунный ответ против Staphylococcus aureus. Полипептиды по изобретению включают аминокислотную последовательность, структурно родственную последовательности SEQ ID NO:1, приведенной в тексте описания. Изобретение относится к использованию таких полипептидов в качестве иммуногенов, в том числе в составе композиций. Изобретение раскрывает также экспрессионные системы для продуцирования таких полипептидов. Последовательность SEQ ID NO:1 представляет собой усеченное производное полноразмерного полипептида S. aureus. Этот полноразмерный полипептид обозначается в изобретении как полноразмерный "ORF0657n". Установлено, что полипептиды, описанные в изобретении и содержащие аминокислотную последовательность SEQ ID NO:1, вызывают защитный иммунный ответ против S. aureus. 11 н. и 32 з.п. ф-лы, 12 ил., 6 табл.
(a) культивирования рекомбинантной клетки по п.29 в условиях, благоприятных для экспрессии полипептида; и
(b) очистки указанного полипептида.
(a) культивирования рекомбинантной дрожжевой клетки в условиях, благоприятных для экспрессии указанного полипептида, где указанная рекомбинантная дрожжевая клетка содержит рекомбинантный ген, кодирующий указанный полипептид, а указанным полипептидом является полноразмерный ORF0657n-родственный полипептид, вызывающий защитный иммунитет против инфекции, вызываемой Staphylococcus aureus, или его фрагмент, содержащий аминокислотную последовательность, которая по крайней мере на 90% идентична последовательности SEQ ID NO:1; и
(b) очистки указанного полипептида.
Приоритет по пунктам и признакам:
| DE 19917098 А, 19.10.2000 | |||
| Комбинированная печь | 1946 |
|
SU71585A1 |
| US 2002031528 А, 14.03.2002 | |||
| WO 03053462 A3, 03.07.2003 | |||
| РОЙТ А | |||
| и др | |||
| Иммунология, М., Изд | |||
| Мир, 2000 г., стр.237-238. | |||

