Изобретение относится к медицине, к способу количественного анализа биологического материала и может быть использовано для оценки генеративной потенции яичников, для диагностики нарушений оогенеза, фолликулогенеза и бесплодия в гинекологии, ветеринарии и животноводстве.
Для яичника стареющего женского организма примерно после 35 лет характерно снижение числа примордиальных фолликулов в результате истощения яичника ооцитами. В каждом цикле до 10-15 фолликулов вступают в развитие и рост, что обеспечивает созревание в них ооцита 1 до зрелой, готовой к выходу из фолликула и к оплодотворению яйцеклетке. Из 2-3 млн ооцитов 1, в норме формирующих пул половых клеток у 4-месячного плода человека женского пола, ко времени наступления менархе (менструального периода, к 12-14 годам) остается двести-триста тысяч, а к концу репродуктивной жизни женщины (40-45 лет) остается несколько тысяч или даже сотен ооцитов 1, заключенных в фолликулы, что исключает оплодотворение яйцеклетки и развитие эмбриона. Формирующийся еще во внутриутробный период развития женского организма, этот пул примордиальных фолликулов может быть истощен преждевременно при различных патологических состояниях организма или внешних на него воздействиях. При этом пул фолликулов и ооцитов в них не восполняется, что приводит к развитию синдрома преждевременного истощения яичников, т.е. к недостаточности овариального резерва яичников и бесплодию уже в детородном возрасте.
У 6-недельного эмбриона половые железы (яичники и яички) различаются по морфологии. Они продуцируют половые клетки, т.е. выполняют генеративную функцию. Формирование в яичнике ооцитов, примордиальных и прогрессирующих в развитии фолликулов, обеспечивает созревание яйцеклеток - источников здорового потомства - generation. Для выявления причин женского бесплодия необходимо диагностировать состояние фолликулярного аппарата яичника с начальных стадий развития ооцитов.
Традиционно, для диагностики бесплодия, состояние оогенеза, дофолликулярных стадий развития женских гамет, не определяют. Для оценки состояния фолликулогенеза - генеративной потенции яичника женщины, судят: а) по концентрации ряда половых гормонов, например фолликулостимулирующего гормона (ФСГ), эстрадиола (Е2), прогестерона (Прг) и др.; б) по изменению объема яичника, определяемого с помощью ультразвукового исследования (УЗИ) и ультразвукового мониторинга (УЗМ) [1].
Кроме того, с помощью УЗИ и УЗМ проводят исследование фолликулогенеза в яичниках и анализ эндометрия, что является основным методом динамического контроля за развитием фолликулов и эндометрия матки, в том числе и во время индукции овуляции гормонами. В процессе УЗИ в яичнике определяют число полостных фолликулов, проводят измерение их среднего диаметра (от 1 мм) по сумме двух измерений, определяют толщину эндометрия.
При гормональном мониторинге выполняют динамическое определение концентрации эстрадиола (Е2), прогестерона (Прг), фолликулостимулирующего гормона (ФСГ), ингибина В, антимюллерова гормона в сыворотке крови, и эта информация дополняет данные УЗИ (УЗМ) в оценке функциональной зрелости фолликулов. Критериями завершенности гормональной стимуляции суперовуляции, например при экстракорпоральном оплодотворении - ЭКО, являются: диаметр лидирующих преовуляторных фолликулов более 17 мм и толщина эндометрия 8 мм и более. Дополнительным маркером степени зрелости фолликулов яичников служит информация о концентрации Е2 в сыворотке крови, свидетельствующая об активности стероидогенеза в оболочке созревающих фолликулов [1]. Т.е. к наиболее информативным сведениям относят данные о концентрации ФСГ, объеме яичника и количестве полостных фолликулов. Однако разрешающая способность УЗИ не позволяет выявлять примордиальные и первичные фолликулы диаметром до 1 мм, из которых примордиальные с заключенными в них ооцитами характеризуют имеющийся в яичнике резерв (пул) ооцитов.
Известен способ дифференциальной диагностики задержки полового развития у девочек препубертатного и подросткового возраста, включающий определение УЗИ размеров матки, просветов маточных сосудов, яичников, количество и диаметр в них полостных фолликулов минимального и максимального размеров. Определены критерии соотношений размеров органов половой системы, которые позволяют выявлять степень задержки полового развития девочек, осложненного сахарным диабетом 1 типа (2). Т.е. при общепринятом диагностическом обследовании с помощью УЗИ могут определять число растущих и полостных преовуляторных фолликулов, но разрешающая способность аппаратов УЗИ не обеспечивает определение пула исходного состава фолликулярного аппарата яичников - пула примордиальных фолликулов, из которых постепенно развиваются все остальные типы фолликулов, что является недостатком метода.
Известны способы обработки фрагментов яичников животных или человека на постсмертном материале для получения обзорных картин структур яичника, включающие получение ткани яичника, проведение ее через спирты, ксилолы, парафины, приготовление парафиновых срезов, удаление парафина и окрашивание срезов, микроскопирование [3, 4].
Возможность охарактеризовать фолликулярную систему яичников при анализе гистологических препаратов срезов биоптата яичника для оценки стадий развития фолликулов яичников используют редко, по медицинским показаниям, либо на операционном материале, при вылущивании кисты яичника, резекции части яичника или удалении яичника [3]. Но такую работу проводят не столько для оценки фолликулярной системы и прогноза деторождения, возможности зачатия для данной женщины, сколько для оценки данного клинического случая, с определенным диагнозом заболевания. В медицинской практике соответствующих служб не используется: взятие биоптата яичника, гистологическая проводка, приготовление гистопрепарата биоптата яичника и его оценка для прогноза деторождения.
В ветеринарии и животноводстве, при разведении ценных пород, исчезающих видов и экспериментальных животных, также не используют предложенный способ диагностики генеративной функции яичников и тестирования повреждающих факторов.
До разработки и апробации указанного способа отсутствовало представление об эталонных значениях хронологии и соотношения ооцитов и фолликулов разных этапов развития, у практически здоровых женщин различных возрастных групп, т.е. контрольной возрастной нормы, с которой необходимо сравнивать в каждом клиническом случае полученные данные о количестве ооцитов и фолликулов разных стадий развития.
Известен способ приготовления гистопрепаратов органов половой системы животных для их сравнительного морфофункционального изучения яичника и семенника различных видов. При этом способе берут часть коры яичника, фиксируют смесью Ценкера, окрашивают гематоксилином и эозином и под большим увеличением изучают фолликулярные клетки на разных стадиях их развития [5]. Данный способ принят за прототип.
При известных способах проводили анализ и описание только единичных фолликулов яичника, без анализа ооцитов в них, без количественного учета дофолликулярных стадий оогенеза и без описания состава разных типов фолликулов, что не позволяет полно определить истинный пул ооцитов и фолликулов у обследуемого.
Вместе с тем, такие проблемы оценки состояния гамет и фолликулов для диагностики нарушений в яичнике являются актуальными не только в медицине, для человека, но и для животноводства и ветеринарии.
Для диагностики генеративной потенции и возможной патологии яичников требуется выбор оптимальных размеров биоптата, параметров и веществ для его обработки, цитологических критериев идентификации ооцитов и фолликулов разных стадий развития, разработка количественных критериев и определение нормы или патологии стадий развития ооцитов и фолликулов, их резерва, в виде эталонных значений, индексов, в зависимости от возраста обследуемого.
Технической задачей предложенного способа, на которую направлено изобретение, является повышение эффективности и информативности диагностики генеративной потенции и возможной патологии обследуемого женского организма.
Техническая задача решена тем, что в способе диагностики генеративной потенции яичников, включающем биопсию коркового вещества яичника, приготовление гистопрепаратов и микроскопический анализ состава ооцитов 1, числа примордиальных фолликулов и фолликулов последующих стадий развития, СОГЛАСНО ИЗОБРЕТЕНИЮ, при биопсии выделяют фрагмент коры яичника площадью 1-2 мм2, толщиной 1,5-2 мм, после фиксации изъятого биоптата в растворе Буэна, приготовленном ex tempore, содержащем 15 мл насыщенного раствора пикриновой кислоты +5 мл 40% нейтрального формалина +1 мл ледяной уксусной кислоты, биоптат промывают в 2-5 сменах 70° этилового спирта при t 18-25°С, проводят через спирты восходящей концентрации: 80°, 96°, 100°1, 100°2 по 30-60 мин в каждом и через смеси: 100° спирт-ксилол в соотношении 1:1, ксилол 1 и 2, ксилол-парафин в соотношении 1:1, парафин 1 и 2 при 56°-58°С, по 30-60 мин в каждом, заливают в парафин и приготавливают парафиновые гистологические срезы толщиной 7-8 мкм, после депарафинизации их окрашивают гематоксилином Майера, Караччи или Эрлиха, с подкрашиванием в 1% спиртовом или водном растворе эозина, затем обезвоживают срезы спиртами восходящей концентрации и двумя сменами ксилола и заключают в канадский бальзам, после чего микроскопируют при увеличении ×400 и подсчитывают на каждом 3-м срезе число ооцитов 1 на дофолликулярных - предыдущих диплотене стадиях профазы 1 мейоза: в прелептотене, лептотене, зиготене, пахитене и на стадии кариосферы в ооците 1, идентифицируя каждую стадию по состоянию хроматина в ядре ооцитов, а также при увеличении ×240 - подсчитывают число фолликулов на каждом 6-м срезе, через 42-48 мкм, соответствующих диаметру примордиального фолликула, и число ооцитов 1 в диплотене или диктиотене, заключенных в примордиальные фолликулы или вне их, а также подсчитывают число фолликулов следующих после примордиального фолликула стадий развития: первичных однослойных и многослойных, вторичных полостных, предовуляторных, зрелого овуляторного фолликула, атретические, желтые и белые тела, а затем определяют долю половых клеток каждой стадии развития от числа всех подсчитанных половых клеток: для ооцитов - в %% от общего числа подсчитанных ооцитов; а для фолликулов каждой из названных стадий их развития и их производных - их количество на 1 поле зрения микроскопа; затем, сравнивая с контрольными индексами, определяют генеративную потенцию яичников и возможную их патологию.
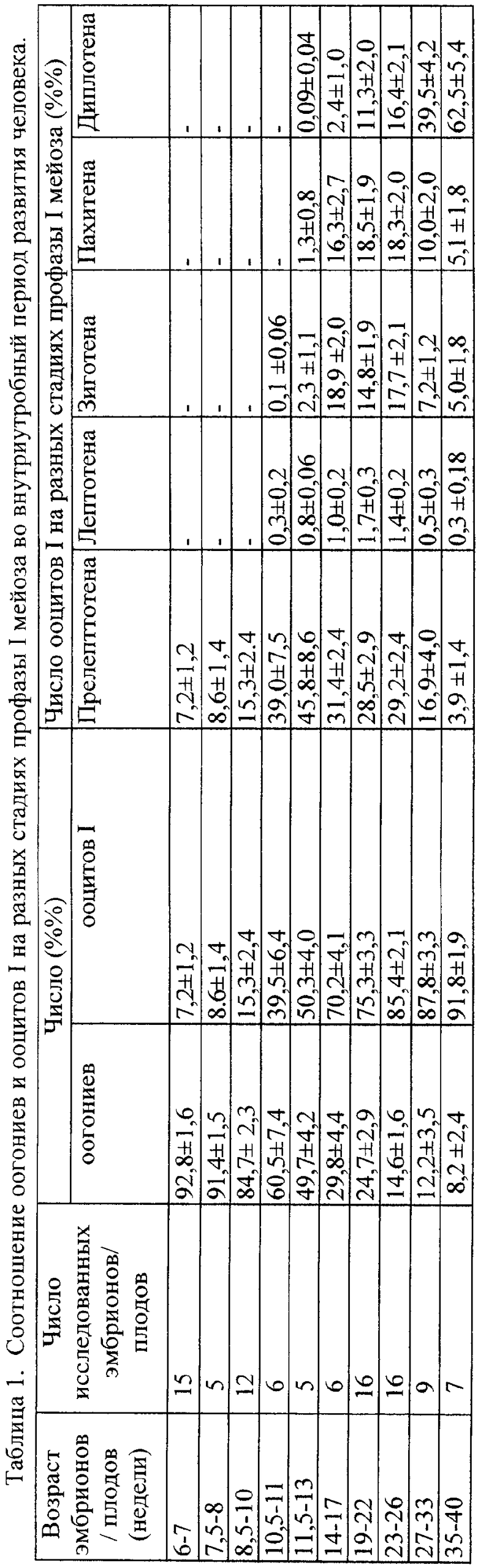
Техническая задача решена также тем, что для определения нормы формирования ооцитов дофолликулярных стадий у человека подсчитывают только число ооцитов 1 на предыдущих диплотене стадиях профазы 1 мейоза: от прелептотены до пахитены, число ооцитов 1 в диплотене, вне примордиальных фолликулов, а также - на стадии кариосферы из полостных фолликулов, идентифицируя каждую стадию по состоянию хроматина в ядре ооцитов, при этом контрольные индексы ооцитов на последовательных стадиях профазы 1 мейоза и сроки перехода одной стадии на следующую, относительно времени внутриутробного развития плода человека, соответствуют следующим значениям: у 6-10-недельных плодов ооциты в прелептотене составляют 7,2-15,3%, остальные половые клетки - оогонии; у плодов 10,5-11 недель число ооцитов достигает 39,0%, появляются ооциты в лептотене - 0,3% и зиготене - 0,1%; у плодов 11,5-13 недель - ооциты в прелептотене достигают 45,8%, на стадии лептотены - 0,8%, в зиготене - 2,3%, появляются ооциты в пахитене - 1,3% и в диплотене, не заключенных в фолликулы - 0,09%; у плодов 14-17 недель - ооциты в прелептотене - 31,4%, в лептотене - 1,0%, в зиготене - 18,9%, в пахитене - 16,3%, в диплотене - 2,4%; у плодов 19-22 недель - в прелептотене - 28,5%, в лептотене - 1,7%, в зиготене - 14,8%, в пахитене - 18,5%, в диплотене - 11,3%; к 23-26 неделям ооциты в прелепотене - 29,2%, в лептотене - 1,4%, в зиготене - 17,7%, в пахитене - 18,3%, в диплотене - 16,4%; у плодов 27-33 недель ооциты в прелептотене - 16,9%, в лептотене - 0,5%, в зиготене - 7,2%, в пахитене - 10,0%, в диплотене - 39,5%; к 35-40 неделям беременности число ооцитов на стадиях в прелептотене снижается до 3,9%, в лептотене - 0,3%, в зиготене - 5,0%, в пахитене - 5,1%, в диплотене - 62,5%.
Техническая задача решена также тем, что для оценки прогрессивного перехода фолликулов человека на последующие стадии, подсчитанных в 1 поле зрения; контрольные индексы фолликулов разных стадий развития в яичниках, в зависимости от возраста, соответствуют следующим значениям: у девочек 6 недель - 6 лет - 20-60 примордиальных фолликулов и единичные бесполостные фолликулы; у девочек 9-13 лет - до 40 примордиальных фолликулов при наличии единичных бесполостных и полостных фолликулов; у женщин детородного возраста 18-35 лет - среднее число примордиальных фолликулов - 5, число первичных и вторичных полостных - 10-11; у женщин 36-55 лет среднее число примордиальных не превышает 2-х фолликулов, и единичные полостные фолликулы.
Техническая задача решена также тем, что для определения генеративной возможности яичников коров и нормы формирования ооцитов дофолликулярных стадий в разные сроки внутриутробного развития контрольные индексы ооцитов соответствуют следующим значениям: у 1,5 месячных эмбрионов ооциты в прелептотене составляют 3,8%, остальные половые клетки - оогонии; у плодов 2-2,5 месяцев - ооциты в прелептотене достигают 44,0%, на стадии лептотены - 2,1%; в зиготене - 3,0%; в пахитене - 0,6%; у плодов 3-4 месяцев развития - ооциты в прелептотене - 35,0-33,1%; в лептотене - 2,8-3,1%; в зиготене - 6,4-18%; в пахитене - 5,7-22,0%; в диплотене - 0,1-4,5%; у плодов 5-6 месяцев беременности число ооцитов на стадии прелептотены - 40,3-15,5%, в лептотене - 8,9-0,9%; в зиготене - 16,9-16,1%, в пахитене 18,9-21,1%; в диплотене - 11,9-46,0%; у 7-месячных плодов коров ооциты в прелептотене составляют 13,9%, в лептотене - 1,4%; в зиготене - 10,8%; в пахитене - 11,4%; в диплотене - 62,3%; к рождению, у 9-месячных плодов отсутствуют ооциты в прелептотене, лептотене, зиготене и пахитене, число ооцитов в диплотене составляет 100%.
Техническая задача решена также тем, что для определения генеративной возможности яичников экспериментальных мышей и нормы формирования ооцитов дофолликулярных стадий в разные сроки внутриутробного развития контрольные индексы ооцитов соответствуют следующим значениям: у 12-дневных плодов ооциты в прелептотене составляют 3,3%, у 13-дневных -31,2%, остальные половые клетки - оогонии; у плодов 14 дней - ооциты в прелептотене составляют 17,2%, на стадии лептотены - 17,7%; у плодов 15 дней-ооциты в прелептотене не превышают 6,0%; в лептотене - 31,8%; в зиготене - 30,8%; в пахитене - 5,8%; у плодов 16 дней - в прелептотене - 0,1%; в лептотене - 7,0%; в зиготене - 61,0%; в пахитене - 26,6%; в диплотене - 0,9%; у плодов 17 дней - ооциты на стадиях прелептотены и лептотены уже отсутствуют; число ооцитов на стадии зиготены составляет 41,0%; на стадии пахитены 50,3%; в диплотене - 6,2%; у плодов 18 дней внутриутробного развития, перед рождением, ооциты в зиготене составляют 0,9%; в пахитене - 49,7%; в диплотене - 42,3%; появляются ооциты в диктиотене - 3,8%.
Техническая задача решена также тем, что при воздействии внешних факторов: физических, химических и биологических на женский беременный организм у плодов снижается число ооцитов в 1,5-2 раза, число примордиальных фолликулов в 1,5-2 раза.
Положительный результат, получаемый при реализации предложенного способа, заключается в следующем:
- повышение точности и информативности поставленного диагноза за счет увеличения числа критериев оценки нарушения резерва примордиальных фолликулов и/или ооцитов для расширения возможности терапии женского бесплодия;
- доступность отработанной методики с использованием стандартных препаратов и оборудования;
- определены минимальные размеры биоптата и оптимальные параметры его обработки;
- определены эталонные значения - соотношения ооцитов 1 на разных дофолликулярных стадиях профазы 1 мейоза в яичниках на всех сроках внутриутробного развития женского организма (у человека, коровы и мыши);
- определены эталонные значения числа примордиальных фолликулов и фолликулов более прогрессивных стадий развития в яичниках в разных возрастных группах здоровых девочек и женщин;
- условно-контрольные цифры прохождения оогенеза и фолликулогенеза в яичниках женского организма по каждой возрастной группе от эмбриона и плода до наступления менопаузы (климакса) расширяют спектр диагностируемых заболеваний и состояний;
- универсальность метода позволяет выявлять норму и нарушение развития дофолликулярных ооцитов и фолликулов яичника человека и разных видов млекопитающих животных.
По сравнению с прототипом [5] предложенный способ отличается операциями забора биопробы, приготовления препарата дофолликулярных ооцитов и фолликулов разных типов для анализа и операциями количественного исследования фолликулярного аппарата яичников с идентификацией и учетом разных последовательных стадий развития дофолликулярных ооцитов и фолликулов, что подтверждает критерий изобретения: «новизна».
Предлагаемый способ диагностики генеративной потенции яичников апробирован на биопсии яичников 40 женщин с заболеваниями яичников и на постсмертном (аутопсии) материале яичников после внезапной гибели 68 здоровых девочек и женщин разных возрастов; яичники при аутопсии от 97 эмбрионов и плодов человека всех сроков внутриутробного развития, 34 эмбрионов и плодов коровы (6-40 недель развития), 151 эмбриона и плода нелинейных мышей (12-18 дней развития), 135 самок мышей половозрелого возраста; 20 зеленых мартышек-самок половозрелого возраста, 4 коровы и 34 зрелых ооцита от 24 коров - для определения состояния оогенеза и фолликулярной системы.
Проведенные вышеуказанные клинические испытания позволили выявить причины нарушения фолликулогенеза, снижение числа примордиальных фолликулов и числа прогрессирующих в развитии фолликулов и бесплодия. Исследование состава ооцитов и фолликулов на аутопсийном (из моргов) и абортивном материале, в экспериментах на животных позволяют: определить закономерности оогенеза и фолликулогенеза; определить индексы соотношения чисел ооцитов и фолликулов в яичниках человека и исследованных видов животных в разных возрастных группах и на разных стадиях менструального (эстрального для животных) цикла, которые служат как исходные контрольные, соответственно, для каждого вида, для сравнения их со значениями, получаемыми при диагностике. Это позволяет считать предложение заявителя соответствующим критерию «промышленная применимость».
Суть изобретения заключается в следующем: Для диагностики генеративной потенции яичников автором, используя известные методы обработки ткани яичника разных видов млекопитающих, экспериментально отработаны оптимальные параметры протокола приготовления гистопрепаратов биоптата яичника. Указанный протокол отрабатывали прежде всего для биопсии яичника женщин с целью наименьшего травматизма. Выполнение данного протокола показано при проведении диагностики яичников разных видов млекопитающих.
Площадь фрагмента яичника 1,5-2 мм2 является необходимой и достаточной для полного и глубокого анализа состава половых клеток и фолликулов яичника. Отработан состав фиксатора - использование раствора Буэна. Раствор Буэна, приготовленный ex tempore, содержит: 15 мл насыщенного раствора пикриновой кислоты +5 мл 40% нейтрального формалина +1 мл ледяной уксусной кислоты, которые обеспечивают быстрое проникновение и консервирование ткани яичника, без изменений его морфологии. Экспериментально отработаны время и условия хранения в растворе Буэна фрагмента яичника, сроки проведения его через спирты восходящих концентраций, соотношения ксилолов и парафинов и время обработки ими фрагмента яичника: фрагмент промывают в 2-5 сменах 79° этилового спирта при t 18-20°С, проводят для 100% обезвоживания ткани через спирты восходящей концентрации: 80°, 96°, 100°1, 100°2 по 30-60 мин в каждом и через смеси: 100° спирт-ксилол, в соотношении 1:1, ксилол 1 и 2, ксилол - парафин, в соотношении 1:1. Обезвоживание обеспечивает проникновение парафина в ткань и пропитывание ее парафином. Затем фрагмент яичника проводят через парафин 1 и 2 при 56-58°С, по 30-60 мин в каждом, заливают в парафин. Прочность и пластичность парафина позволяет готовить из парафиновых блоков, с заключенными в них фрагментами яичника, тонкие гистологические срезы 7-8 мкм с помощью бритвы фирмы Рейхерт. После депарафинизации их окрашивают гематоксилином Майера, Караччи или Эрлиха, с подкрашиванием в 1% спиртовом или водном растворе эозина. Именно срезы такой толщины, после подобранного режима окрашивания красителями: для хроматина ядра (гематоксилином) и цитоплазмы клетки (эозином), позволяют исследовать половые клетки-ооциты и фолликулы под микроскопом. При увеличении ×400 оценивают стадию по морфологии хроматина и ядер ооцитов и ее нарушение. Увеличение ×240 обеспечивает оптимальную обзорность оценки и стадии фолликула. Ооциты 1 идентифицируют по стадиям и подсчитывают на каждом 3-м срезе, т.е. через 21-24 мкм потому, что диаметр дофолликулярных ооцитов не превышает 20 мкм. При этом подсчитывают лишь те ооциты, ядра которых, диаметром до 10 мкм, входят в поле зрения микроскопа. Диаметр примордиального и первичного фолликула не превышает 40 мкм, поэтому их подсчитывают на каждом 6-м срезе, через 42-48 мкм и проходящем через ооцит.
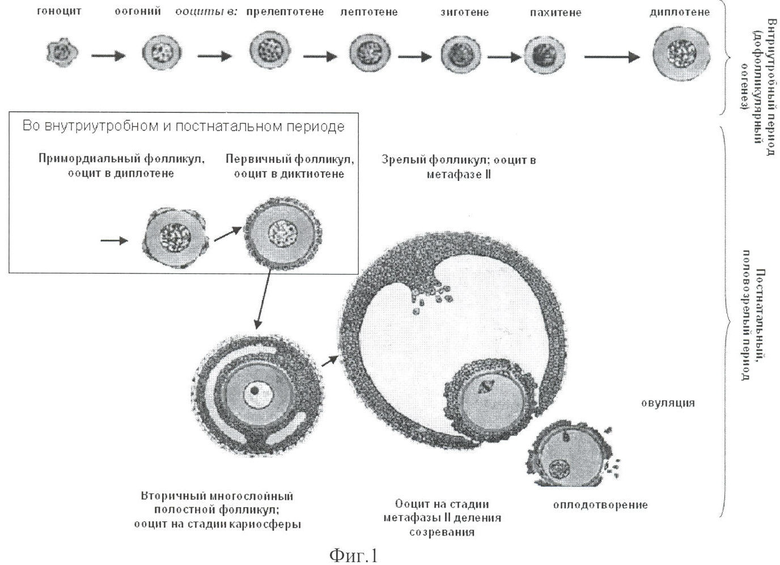
Схема оогенеза - длительного и многостадийного развития женских половых клеток, гамет, указана на фиг.1. Фотографии ооцитов и фолликулов на различных стадиях развития приведены на фиг.2, 3 и 4.
На фиг.1 показаны последовательные стадии развития женских половых клеток, начиная с эмбрионального периода развития организма и самих гамет. Первичные половые клетки - гоноциты - мигрируют из эпибласта - зоны их формирования в период от 15-ти дней до 4-х недель эмбрионального развития человека и заселяют зачаток яичников. В яичнике гаметы определяют как оогонии, которые делятся митозом и переходят в ооциты 1. Вначале ооциты проходят дофолликулярные стадии профазы 1 мейоза: прелептотену (последний синтез ДНК), лептотену (начинают синтезироваться белки синаптонемного комплекса для конъюгации), зиготену (развивается конъюгация между гомологичными родительскими хромосомами), пахитену (происходит обмен генами между гомологичными родительскими хромосомами), диплотену. Ооциты проходят вышеуказанные стадии во внутриутробный период жизни формирующегося женского организма, человека или млекопитающих животных. Данная схема оогенеза универсальна для разных видов млекопитающих.
Ооцит 1 на стадии диплотены, как впервые количественным методом показано автором на плодах человека, коровы и мыши, заключается в примордиальный фолликул. И далее ооциты развиваются и созревают в фолликулах - фолликулярное развитие. При нормально протекающей беременности у рожденной девочки подавляющее большинство ооцитов находятся на стадии диплотены и заключены в примордиальные фолликулы, а единичные - в первичные. При наступлении полового созревания в начале каждого менструального (для животных - эстрального) цикла в правом или левом яичнике попеременно, несколько примордиальных фолликулов вступают в рост и трансформируются в первичные, в которых ооциты переходят на стадию диктиотены. По мере нарастания числа слоев фолликулярных клеток ооцит и фолликул увеличиваются в размере. В многослойном вторичном фолликуле формируется полость, а в его ооците 1 хромосомы конденсируются и локализуются в виде плотной массы вокруг ядрышка - кариосферы. Эту стадию кариосферы в ооцитах полостных фолликулов человека, коровы и зеленой мартышки впервые исследовала и описала для этих трех видов автор данного изобретения. При достижении созревания фолликула с большой полостью в ооците развивается короткий диакинез, переходящий в метафазу 1, далее протекают два деления мейоза, которые отмечают по наличию метафазы 1 и 2 мейоза. После метафазы 1, завершающейся ана- и телофазой 1 из одного ооцита 1, формируются: один ооцит 2 (вторичный) и первое полярное тельце (абортивная клетка, уходящая в гибель). Затем из ооцита 2 в зрелом полостном фолликуле формируются одна зрелая яйцеклетка и второе полярное тельце. Яйцеклетка овулирует из фолликула и готова к оплодотворению. Таким образом, незрелые половые клетки от гоноцитов и оогониев до ооцитов 1, и ооцитов 2 и зрелые женские гаметы - яйцеклетки являются предметом исследования и диагностики состояния генеративных потенций яичников и патологии половых клеток и фолликулов.
На фиг.2 и фиг.3 приведены фото половых клеток с гистопрепаратов половых клеток - ооцитов яичников 3-месячных плодов человека, полученных из аутопсийного материала, приготовленных по разработанному способу.
На фиг.2 показаны: 2а - ооцит в прелептотене (ГШ), зиготене (ЗТ); 2б - ооцит в зиготене (ЗТ), пахитене (ПТ), диплотене (Дт); 2в - ооцит в прелептотене (ГШ), оогонии (ОГ).
На фиг.3 показаны: 3а - ооциты в диплотене (Дт), заключенные в примордиальные фолликулы (ПФ); 3б - фрагмент ооцита на стадии кариосферы (КФ), заключенный в многослойный полостной фолликул (стрелки - три слоя фолликулярных клеток фолликула).
В медицинских и ветеринарных клиниках анализ и оценка фолликулогенеза по биоптату яичника не выполняются; по УЗИ не выявляются примордиальные и первичные без полости фолликулы и дофолликулярные ооциты 1, т.е. не выявляется пул - резерв примордиальных фолликулов с ооцитами. Автором предлагается разработанный способ оценки: состояния оогенеза по количественному цитологическому анализу клеточного состава гамет по стадиям их дофолликулярного развития, т.е. по анализу половых клеток на всех последовательных дофолликулярных и на стадии кариосферы полостного фолликула, идентифицируя по цитологической картине хроматина, ядер ооциты 1 на разных стадиях оогенеза. В предлагаемом способе проводят также детальный и количественный анализ фолликулогенеза, начиная с определения пула примордиальных фолликулов и учитывая количество фолликулов на всех последующих стадиях их развития, а также - их производные - атретические, желтые и белые тела в яичнике. Для критериев оценки резерва определены условно-контрольные индексы для человека, коровы, мыши.
Автор, впервые в научном мире, разработала и практически использовала схему такой оценки идентификации и исследования количественного соотношения ооцитов именно всех перечисленных стадий оогенеза, у человека и нескольких видов животных, в многочисленных экспериментах и на клиническом материале.
Были проведены исследования для определения хронологии и динамики формирования ооцитов дофолликулярных стадий. Предлагаемый способ диагностики генеративной потенции яичников апробирован: на гистологических препаратах яичников при аутопсии от 97 эмбрионов и плодов человека всех сроков внутриутробного развития, после спонтанных абортов или медицинского прерывания беременности (табл.1, пункт формулы 1, 2); от 34 эмбрионов и плодов коровы, 6-40 недель развития (табл.2, пункт формулы 1, 2, 5); от 151 эмбриона и плода нелинейных мышей, 12-18 дней развития (табл.3, пункт формулы 1,2,5). Эти исследования позволили получить новую информацию о динамике формирования ооцитов каждой стадии развития, хронологии их перехода на последующие стадии на разных сроках внутриутробного развития в яичниках человека, коровы, мыши, а также информацию о составе ооцитов разных стадий развития на каждом сроке внутриутробного периода развития яичника. Эти количественные данные используются как эталон для сравнения при проведении исследований по тестированию повреждающих факторов, при диагностике нарушений развития ооцитов и фолликулов в яичниках у человека и животных.
Так, например, в таблице 1 показаны периоды начала появления ооцитов человека на стадии прелептотены, их перехода на следующую стадию - лептотены, сроки - недели развития и конкретную долю (от общего числа ооцитов) максимального количества ооцитов на каждой из стадий. Конкретно - период максимального количества ооцитов в прелептотене - от 10 до 26 недель, появление ооцитов в лептотене и зиготене - у 10-недельных плодов, появление ооцитов на стадии пахитены (т.е. их развитие после зиготены) начинается у плодов не ранее 10-11 недель. Но если эти стадии у плода человека появляются у плодов более поздних сроков развития или если их доля будет большей на любых сроках развития, это будет свидетельствовать о блоке - задержке развития ооцита на этой стадии. Такая же закономерность - запаздывание или блок перехода ооцитов от более ранней на более позднюю стадию всегда свидетельствует о нарушении времени и стадии развития женских половых клеток. Например, воздействие повреждающими факторами: алкоголем, никотином, женским гормоном 17-в-эстрадиолом, антибиотиком вызывает задержку перехода ооцитов с одной стадии на другую и снижение числа половых клеток, резерв которых в женском организме не пополняется (пример 1).
Эти нарушения являются универсальными в биологии и происходят у человека и разных видов животных (таблицы 1, 2, 3). Такая же закономерность сроков перехода ооцитов на следующие стадии развития, проявление максимума такого перехода для каждой стадии в определенные сроки внутриутробного развития плода и к моменту его рождения накопление ооцитов на поздних стадиях дофолликулярного развития - в пахитене и затем - в диплотене, характерна для всех видов млекопитающих, в том числе и у изученных нами коровы и мыши.
Для оценки образования резерва примордиальных фолликулов в яичниках человека и животных, закономерности перехода примордиальных на последующие стадии развития фолликулов, динамики формирования стадии кариосферы в ооцитах полостных фолликулов исследовано: яичники при аутопсии от 97 эмбрионов и плодов человека всех сроков внутриутробного развития (после спонтанных абортов или медицинского прерывания беременности) (табл.4, пункт формулы 3); на постсмертном материале аутопсии яичников после внезапной гибели 68 здоровых девочек и женщин разных возрастов (табл.5, 6, пункт формулы 3); 20 зеленых мартышек-самок половозрелого возраста (табл. 8, пункт формулы 3), 4 коровы и 34 зрелых ооцита от 24 коров (табл.7, пункт формулы 3); 135 самок мышей половозрелого возраста (пункт формулы 5). Это позволяет определить резерв примордиальных фолликулов и генеративную потенцию яичников. Это позволяет определить норму развития ооцита в фолликуле до стадии кариосферы, означающей готовность ооцита вступить в созревание и в дальнейшую овуляцию.
Полученные количественные данные используются как эталон для определения состояния фолликулярной системы и резерва - количества примордиальных фолликулов, перехода фолликулов на последующие стадии фолликулогенеза, динамики формирования стадии кариосферы у ооцитов из полостных фолликулов человека и животных.
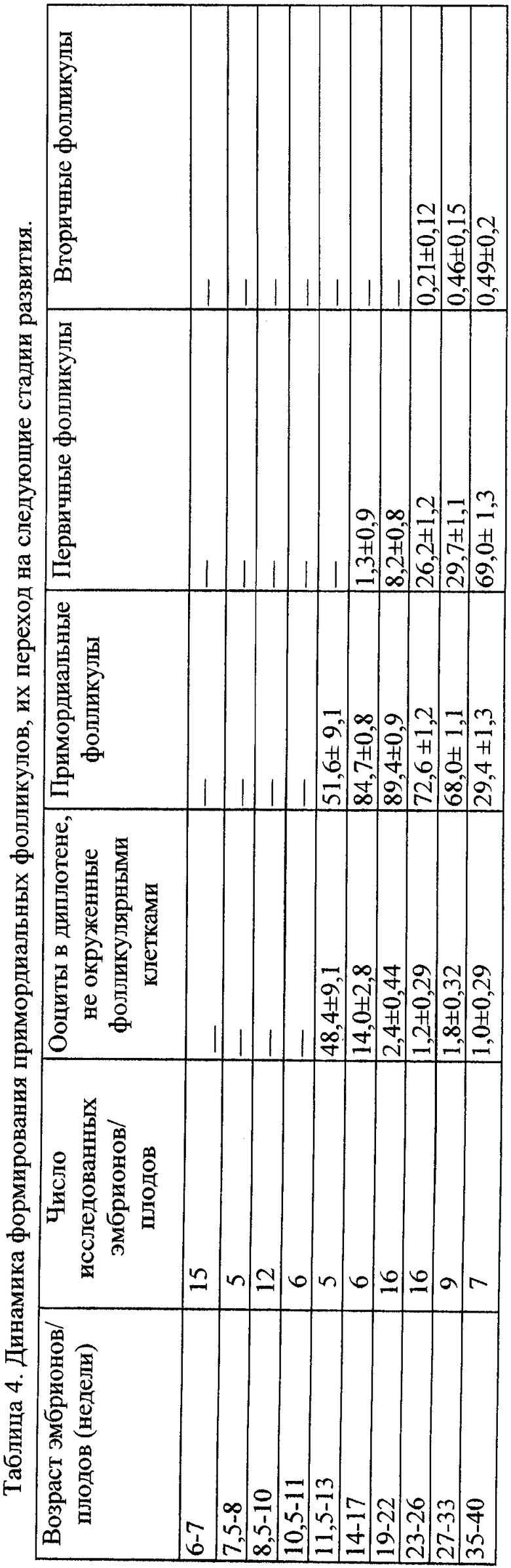
В таблице 4 дана динамика формирования примордиальных фолликулов у плодов человека и их переход на следующие стадии развития. Показано, что формирование примордиального (исходного) фолликула начинается у 11-недельного плода вокруг ооцита в диплотене. Максимальное фолликулообразование происходит к сроку 19-22 неделям и завершается ко времени рождения - 40 неделям развития. Поскольку яичник плодов человека ко времени рождения имеет малый размер (0,5 см×1,5 см), а у растущего далее половозрелого организма яичники значительно увеличиваются в размерах (до 2,5 см×4 см и более), частота распределения примордиальных фолликулов после рождения у увеличивающихся в размерах яичников становится реже - и подсчет примордиальных фолликулов проводится в полях зрения.
В таблице 5 приведены результаты подсчета разных стадий развития фолликулов в яичниках 68 здоровых девочек и женщин, погибших в острых ситуациях. При этом контрольные индексы стадий развития фолликулов здоровых девочек и женщин соответствуют следующим значениям: у девочек 6 недель - 6 лет - 20-60 примордиальных фолликулов (на 1 поле зрения) и единичные бесполостные фолликулы; у девочек 9-13 лет - до 40 примордиальных фолликулов при наличии единичных бесполостных и полостных фолликулов; у женщин детородного возраста 18-35 лет - среднее число примордиальных фолликулов - 5, число первичных и вторичных полостных - 10-11; у женщин 36-55 лет среднее число примордиальных не превышает 2-х фолликулов.
Указанные контрольные индексы динамики и хронологии оогенеза, фолликулогенеза, преобразования хромосомно-ядрышкового комплекса в кариосферу ооцита 1 полостного фолликула приведены в таблицах 1-8 и могут быть использованы для сравнения в качестве эталона - условно-контрольной информации по изученным процессам у эмбрионов, плодов, девочек и женщин разных возрастных групп, а также - у коровы, мыши и зеленой мартышки.
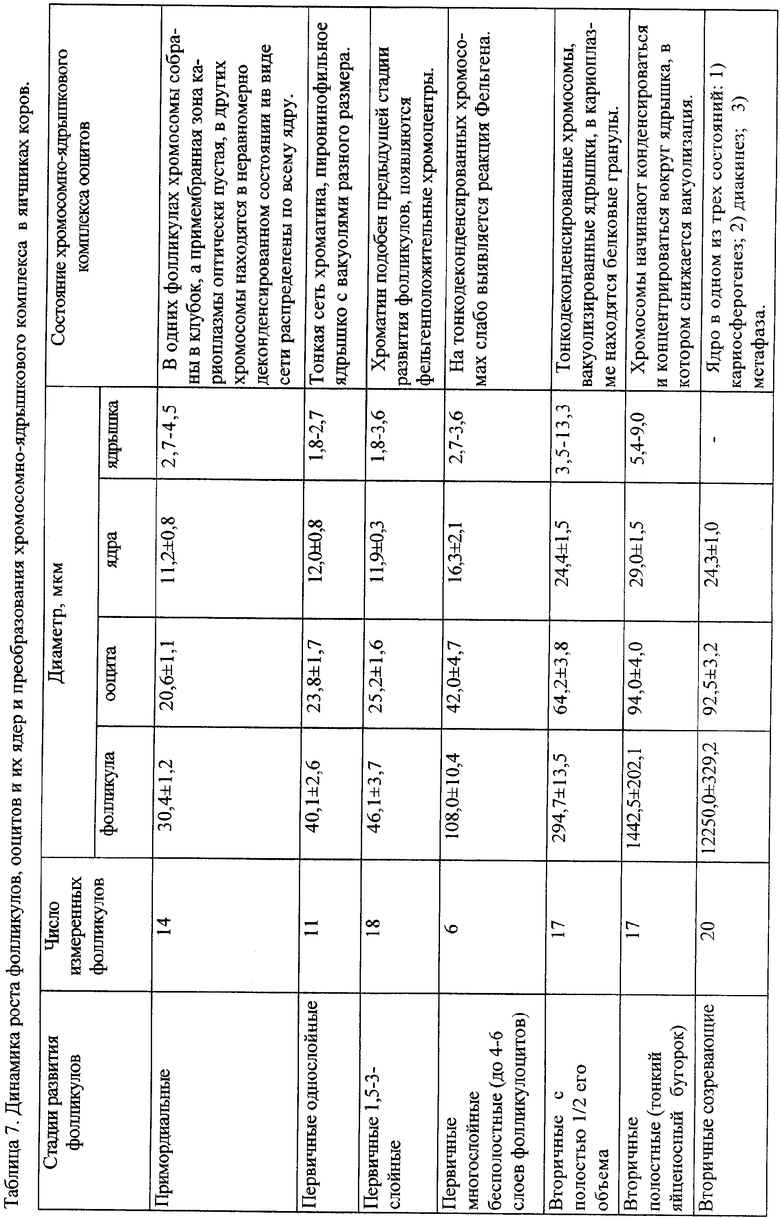
Важным критерием заключительного созревания ооцитов в развивающихся фолликулах являются цитологические преобразования хроматина и ядра в ооците, завершающиеся наступлением стадии кариосферы. После этого развивается диакинез и два деления мейоза - обязательные события для овуляции яйцеклетки. В таблице 6, 7 и 8 приведены результаты морфометрического исследования на гистопрепаратах яичников корреляции между стадией роста и развития фолликулов и изменением структуры ядра и хромосом, их постепенным перемещением вокруг ядрышка и формированием особой структуры - кариосферы. Такая локализация хромосом в крайне компактную структуру - кариосферу предотвращает потерю хромосом после обязательного разрушения оболочки ядра при вступлении ооцита в деления созревания. Согласно таблицам 6, 7 и 8 установлена универсальность преобразований хромосом и ядра ооцитов в фолликулах разных стадий развития у разных видов млекопитающих, т.е. показано полное сходство схемы созревания ооцитов и фолликулов у человека, коровы и мартышки. Идентификация стадии кариосферы необходима для диагностики физиологичности созревания ооцитов - яйцеклеток в полостных фолликулах и выявления их патологии.
Следует отметить, что при разработке предложенного способа диагностики генеративной потенции яичников и нарушения фолликулогенеза на гистопрепаратах биоптата яичников удалось выявить, что у половозрелых женщин с трансексуализмом (3 человека) и с эндометриозом яичников (37 человек) в их гонадах определены лишь единичные примордиальные фолликулы, что свидетельствует о значительном снижении числа примордиальных и первичных фолликулов, т.е. практически - отсутствии резерва фолликулов в каждом клиническом случае. Предложенный способ позволяет более точно выявлять нарушение конкретных стадий формирования фолликулов. Значительно сниженное количество примордиальных фолликулов и развивающихся, в том числе - полостных фолликулов свидетельствует о том, что лишь малая доля половых клеток, заключенных в фолликулы, в случае нарушения репродуктивной функции у женщин проходит через эти стадии (фолликулообразования в примордиальный фолликул), у некоторых пациенток фолликулы отсутствовали вообще. Таким образом, подтвержденные на обширном экспериментальном и клиническом материале, обеспечившем достоверность результатов, полученные с помощью отработанного и апробированного метода (способа) данные количественного анализа состояния оогенеза и фолликулогенеза по предложенному способу позволяют в каждом клиническом случае найти причины патологии развития гамет и фолликулогенеза, т.е. нарушения резерва гамет и/или фолликулов и бесплодия.
Предлагаемый способ иллюстрируется следующими примерами.
Пример 1.
При культивировании in vitro выделенных гонад абортированных 7-10-недельных плодов человека и воздействии на них терапевтических доз антибиотика окситетрациклина (ОТЦ) или женского полового гормона 17-в-эстрадиола на гистопрепаратах яичников выявлено их повреждающее воздействие: эти соединения блокировали деление оогониев митозом и переход оогониев в ооцит на стадии прелептотены. Введение на 12-е и 13-е сутки беременности ОТЦ в терапевтической дозе приводило к уменьшению числа оогониев и ооцитов у их 14-суточных плодов (в 2 раза); снижению числа примордиальных фолликулов и овулирующих яйцеклеток у половозрелого потомства тех мышей, которым вводили ОТЦ во время их беременности этим потомством (в 1,5-2 раза).
Пример 2.
У 68 девочек и женщин (яичники получены при их вскрытии в моргах) возрастных 4-х групп условной нормы были определены на гистопрепаратах яичников контрольные индексы фолликулов разных стадий развития (таблица 5). По сравнению с этими показателями у обследуемых у женщин с эндометриозом яичников (37 человек), кистозный или железисто-кистозный варианты: 1-я группа - возраст 21-35 лет (17 человек); 2-я группа - 36-48 лет (20 человек) - показатели пула примордиальных фолликулов и развивающихся полостных соответственно значительно снижены. У женщин 1-й группы частота встречаемости примордиальных фолликулов в среднем составила 0,01 на 1 поле зрения, а у женщин 2-й группы - 0,002 фолликула на 1 пае зрения. Половых клеток дофолликулярных стадий развития не выявлено. Сохранена тенденция снижения числа примордиальных фолликулов и полостных фолликулов в зависимости от возраста женщины (таблица 5). Во всех проведенных испытаниях в каждом случае выявлены причины женского бесплодия - нарушение резерва ооцитов и фолликулогенеза.
Пример 3.
У трех пациенток с женским транссексуализмом, с нормальным женским кариотипом 46,ХХ и женским фенотипом - на биопсии яичника по сравнению с контрольными показателями (у погибших женщин того же возраста) выявлены: у 1-й женщины - единичные примордиальные фолликулы с признаками дегенерации (фиг.4); у двух других - отсутствие примордиальных фолликулов. У всех трех были выявлены кистозно-перерожденные полостные фолликулы (фиг.4) до 5,5 мм в диаметре. Половых клеток дофолликулярных стадий развития не выявлено. Эти факты свидетельствуют о том, что постоянные психологические стрессы и нарушение уровня гормонов у данных пациенток отражается на развитии числа половых клеток и пуле примордиальных фолликулов и это объясняет причину бесплодия.
Без проведения цитологической диагностики генеративной потенции женщин без подсчета пула примордиальных фолликулов по предложенному способу невозможно было объяснить нарушение фолликулярной системы яичника и бесплодие.
На фиг.4 - гистологический препарат биоптата яичника женщины с мужским транссексуализмом, показаны: 4а - в корковом слое яичника локализованы 2 примордиальных фолликула с признаками дегенерации (ПФ); 4б - в корковом слое яичника локализованы 2 кистозно-перерождающихся полостных фолликула.
Отработан способ диагностики генеративной потенции яичников, включающий забор материала яичников, приготовление гистологических препаратов, их анализ и сравнение результатов с установленными авторам эталонными значениями. Проведенные вышеуказанные клинические испытания позволили выявить причины нарушения фолликулогенеза, снижение числа примордиальных фолликулов и числа развивающихся фолликулов и бесплодия. Исследование состава ооцитов и фолликулов на аутопсийном и абортивном материале, в экспериментах на мышах позволяет: определить закономерности оогенеза и фолликулогенеза; определить индексы соотношения чисел ооцитов на разных стадиях и фолликулов в яичниках человека и исследованных видов животных, в разных возрастных группах и на разных стадиях менструального (эстрального для животных) цикла.
Эти количественные данные являются контрольными соответственно для каждого вида для сравнения их со значениями, получаемыми при диагностике.
Источники информации
1. Приказ МЗ РФ от 26/2-2003 №67. О применении вспомогательных репродуктивных технологий (ВРТ) в терапии женского и мужского бесплодия, с.1, 3, 4.
2. Патент РФ №02264789 С2 Опублик. 27.11.2005 г. Кл. А61В - 8/00, 5/107.
3. Лапароскопия и гистероскопия в гинекологии и акушерстве. Ред. В.И.Кулаков, Л.В.Адамян. М: Пантори, 2002, с.148-149.
4. Ромейс Б. Микроскопическая техника. М.: ИЛ, 1954, с.518-519.
5. Кирпичникова Е.С., Левинсон Л.Б. Практикум по частной гистологии. М.: Высшая школа, 1963, с.118-121 - прототип.





| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ЦИТОЛОГИЧЕСКОЙ ДИАГНОСТИКИ НАРУШЕНИЯ СПЕРМАТОГЕНЕЗА | 2007 |
|
RU2328736C1 |
| Средство для лечения женского бесплодия и бесплодия самок животных | 2017 |
|
RU2740922C2 |
| СПОСОБ КУЛЬТИВИРОВАНИЯ IN VITRO ОВАРИАЛЬНЫХ ФОЛЛИКУЛОВ | 2001 |
|
RU2286384C2 |
| Белково-пептидный комплекс, повышающий жизнеспособность фолликулов в яичниках млекопитающих | 2017 |
|
RU2660587C1 |
| Способ хирургической активации функции яичников при низком овариальном резерве | 2021 |
|
RU2754060C1 |
| ТЕХНОЛОГИЯ ДОЗРЕВАНИЯ ООЦИТОВ ЧЕЛОВЕКА НА СТАДИИ GV С ПОМОЩЬЮ ВНЕКЛЕТОЧНЫХ ВЕЗИКУЛ ФОЛЛИКУЛЯРНОЙ ЖИДКОСТИ В ПРОГРАММАХ ЭКСТРАКОРПОРАЛЬНОГО ОПЛОДОТВОРЕНИЯ: EV-IVM (EXTRACELLULAR VESICLES IN VITRO MATURATION) | 2023 |
|
RU2807492C1 |
| Способ определения атретических изменений в ооцитах из полостных фолликулов яичников крупного рогатого скота | 1986 |
|
SU1386189A1 |
| СПОСОБЫ СОЗРЕВАНИЯ ФОЛЛИКУЛОВ ЯИЧНИКА IN VITRO | 2009 |
|
RU2492866C2 |
| СПОСОБ ОТБОРА ПАЦИЕНТОК С СИНДРОМОМ "ПУСТЫХ" ФОЛЛИКУЛОВ ДЛЯ ПРОВЕДЕНИЯ ПРОГРАММЫ ЭКО И ПЭ ДОНОРСКИМИ ООЦИТАМИ | 2005 |
|
RU2309411C2 |
| Препарат для стимуляции фолликулогенеза и способ его применения | 2016 |
|
RU2629871C1 |



Изобретение относится к медицине и биологии, а именно к диагностике бесплодия. Предложен способ оценки генеративной потенции яичников. Ткань яичника фиксируют в растворе Буэна, приготавливают гистологические срезы толщиной 7-8 мкм, окрашивают гематоксилином и эозином. Определяют долю половых клеток каждой стадии их развития от числа всех подсчитанных гамет: для ооцитов в %% от общего числа подсчитанных ооцитов; для фолликулов и их производных - их количество на каждой стадии на 1 поле зрения микроскопа. Для здоровых представителей разных видов млекопитающих определяют контрольные индексы по различным стадиям оогенеза и/или фолликулогенеза. По результатам сравнения с контрольными индексами определяют генеративную потенцию яичников и возможную их патологию. Способ позволяет диагносцировать нарушения оогенеза и фолликулогенеза в гинекологии, ветеринарии и животноводстве. 4 ил., 8 табл.
Способ оценки генеративной потенции яичников, характеризующийся тем, что ткань яичника фиксируют в растворе Буэна, приготовленном ex tempore, содержащем 15 мл насыщенного раствора пикриновой кислоты +5 мл 40%-ного нейтрального формалина +1 мл ледяной уксусной кислоты, промывают в 2-5 сменах 70° этилового спирта при 118-25°С, проводят через спирты восходящей концентрации: 80°, 96°, 100°1, 100°2 по 30-60 мин в каждом и через смеси: 100° спирт-ксилол, в соотношении 1:1, ксилол 1 и 2, ксилол-парафин, в соотношении 1:1, парафин 1 и 2 при 56-58°С, по 30-60 мин в каждом, заливают в парафин и приготавливают парафиновые гистологические срезы толщиной 7-8 мкм, после депарафинизации их окрашивают гематоксилином Майера, Караччи или Эрлиха, с подкрашиванием в 1%-ном спиртовом или водном растворе эозина, затем обезвоживают срезы спиртами восходящей концентрации и двумя сменами ксилола и заключают в канадский бальзам, после чего микроскопируют при увеличении ×400 и подсчитывают на каждом 3-м срезе число ооцитов 1 на дофолликулярных - предыдущих диплотене стадиях профазы 1 мейоза: в прелептотене, лептотене, зиготене, пахитене и на стадии кариосферы в ооците 1 полостного фолликула, идентифицируя каждую стадию по состоянию хроматина в ядре ооцитов, а также при увеличении ×240 - подсчитывают число фолликулов на каждом 6-м срезе, через 42-48 мкм, соответствующих диаметру примордиального фолликула, и число ооцитов 1 в диплотене или диктиотене, заключенных в примордиальные фолликулы или вне их, а также подсчитывают число фолликулов следующих после примордиального фолликула стадий развития: первичных однослойных и многослойных, вторичных полостных, предовуляторных, зрелого овуляторного фолликула, атретические, желтые и белые тела, а затем определяют долю половых клеток каждой стадии развития от числа всех подсчитанных половых клеток: для ооцитов - в %% от общего числа подсчитанных ооцитов; а для фолликулов каждой из названных стадий их развития и их производных - их количество на 1 поле зрения микроскопа, и сравнивают полученные данные с контрольными индексами, определенными для здоровых представителей разных видов млекопитающих по вышеуказанным стадиям оогенеза и/или фолликулогенеза.
| Способ определения функционального состояния репродуктивной системы у женщин при болезни Иценко-Кушинга | 1990 |
|
SU1835078A3 |
| КИРПИЧНИКОВА Е.С | |||
| и др | |||
| Практикум по частной гистологии | |||
| - М.: Высшая школа, 1963, с.118-121 | |||
| Способ диагностики заболеваний яичников | 1991 |
|
SU1803155A1 |
| СПОСОБ ПРОГНОЗИРОВАНИЯ ФЕРТИЛЬНОСТИ ЖЕНЩИН С ЕДИНСТВЕННОЙ МАТОЧНОЙ ТРУБОЙ ПОСЛЕ ВНЕМАТОЧНОЙ БЕРЕМЕННОСТИ | 2002 |
|
RU2219552C2 |

