Изобретение относится к биохимии и молекулярной биологии эукариотической клетки и может быть применено к анализу молекулярно-генетических механизмов формирования структур клеточного ядра и роли белковых компонентов в их организации, что необходимо для получения дополнительной информации в разработках и построении компьютерных моделей организации генных и эпигенных сетей управления.
Известен способ препаративного выделения основных белков клеточных ядер из тимуса теленка [1], в котором был описан способ фракционирования гистонов с помощью ионообменной хроматографии на колонке с амберлитом ИРЦ-50. Недостаток этого метода заключается в том, что были использованы колонки размером 7,5×55 см и 4,5×49 см, для загрузки которых необходимо достаточно большое количество белка (2-3 г и 0,5-1 г соответственно), в то время как при работе с растительным объектом выделение белка из клеточных ядер в таком объеме чрезвычайно трудоемко.
Известен способ фракционирования растительных гистонов на колонках с амберлитом ИРЦ-50 [2], в котором был описан способ разделения основных белков клеточных ядер проростков кукурузы. Недостаток этого метода заключается в том, что с его помощью невозможно расфракционировать микроколичества белков из супраструктур клеточных ядер (20-50 мкг).
Вышеуказанный способ выделения растительных гистонов на колонках с амберлитом ИРЦ-50 [2] был принят за основу, в нем первоначально из проростков кукурузы изолируют клеточные ядра по методу Ро и Чипчейза [см. 2], получают ядерный экстракт с помощью 2 М хлористого натрия и добавления 1 н. соляной кислоты, с последующим нанесением полученного белка на колонку с амберлитом ИРЦ-50 и элюированием гистонов в линейном градиенте гуанидин гидрохлорида, определением белка в элюатах турбидиметрически. Недостаток этого метода заключается в том, что ядра были изолированы методами Ро и Чипчейза [см. 2], что ведет к низкому выходу нативных ядер, из которых с помощью 2 М хлористого натрия была выделена только одна фракция белков, содержание белка в которой определялось турбидиметрически, методом с низким порогом чувствительности (до 3-5 мкг белка в 0,1 мл раствора).
Цель изобретения - предлагается способ для препаративного выделения основных белков из супраструктур клеточных ядер растений с помощью ионообменной хроматографии на колонке с амберлитом ИРЦ-50.
Указанная цель достигается тем, что в способе выделения основных белков из супраструктур клеточных ядер растений первоначально в определенные интервалы времени от начала замачивания 0 ч, 3 ч, 6 ч, 9 ч, 12 ч, 15 ч, 18 ч, 21 ч проводят отделение зародышей от эндосперма с последующей консервацией зародышей в забуференном 80-90% глицерине при минус 25°С, выделяют клеточные ядра, проводят экстракцию ядерных фракций возрастающими концентрациями 0,14 М, 0,35 М, 2 М хлористого натрия и 6 М гуанидин гидрохлорида с 0,1% β-меркаптоэтанолом и выделяют из вышеперечисленных фракций основные белки с помощью ионообменной хроматографии с амберлитом ИРЦ-50 в прерывистом градиенте гуанидин гидрохлорида: 6%, 8,9%, 10,6%, 13%, 40% на 0,1 М калий-фосфатном буфере рН 6,8. Изобретение иллюстрируется следующим примером.
Пример. Опыты проводили на элитных семенах пшеницы (Triticum aestivum L.) сорта Мироновская яровая (любезно присланных нам из коллекции Мироновского научно-исследовательского института селекции и семеноводства пшеницы им. В.Н.Ремесло). Проращивание зародышей осуществляли в темноте при 22±1°С. В определенные интервалы времени - 0 ч (воздушно-сухое семя) и от начала замачивания семян: 3 ч, 6 ч, 9 ч, 12 ч, 15 ч, 18 ч, 21 ч проводили отделение зародышей от эндосперма. Клеточные ядра выделяли по методу [3]. Надмолекулярные структуры: нуклеоплазму (Нп), хроматин, непрочно (Хр-I) и прочно (Хр-II) связанный с ядерным матриксом (ЯМ), а также ЯМ выделяли из очищенных клеточных ядер соответственно при повышении ионной силы раствора: 0,14 М NaCl, 0,35 M NaCl, 2 M NaCl в 0,01 М трис-HCl буфере, рН 6,8. ЯМ извлекали 6 М гуанидин гидрохлоридом (Gu·HCl) с 0,004%-ным β-меркаптоэтанолом в том же буфере. Количество белка в ядрах и ядерных фракциях определяли методом Бредфорда в нашей модификации [4]. Полученные супраструктуры клеточных ядер пропускали через колонку с амберлитом ИРЦ-50 (полиментакриловая синтетическая смола со свободными карбоксильными группами). Смола использовалась в виде порошка, получаемого при размалывании в шаровой мельнице. Размельченная смола просеивалась через сито 200 меш, после чего многократно промывалась водой для удаления мельчайших частиц, высушивалась и промывалась ацетоном. Для достижения большей хроматографической эффективности проводили циклизацию. К 100 г смолы добавляли 500 мл 4 н. NaOH, перемешивали в течение 3 ч с последующим отмыванием на фильтре водой до нейтральной реакции и переводили в кислую форму, пропуская через нее 500 мл 4 М НС1, избыток HCl удаляли промыванием водой. Этой смолой заполняли колонку размером 0,4×4,5 см. На колонку наносили белок в количестве 20-50 мкг, растворенный в 6% гуанидин гидрохлориде на 0,1 М калий-фосфатном буфере рН 6,8. Скорость элюции составляла 6 мл/ч. С помощью элюирования основных белков хроматина в линейном градиенте гуанидин гидрохлорида (6%-40%) были установлены точные концентрации выхода гистоновых фракций: 6%, 8,9%, 10,6%, 13%, 40% раствора гуанидин гидрохлорида на 0,1 М калий-фосфатном буфере рН 6,8. Последующее препаративное отделение основных белков протеома клеточных ядер проводили в прерывистом градиенте гуанидин гидрохлорида: 6%, 8,9%, 10,6%, 13%, 40% на 0,1 М калий-фосфатном буфере рН 6,8. Описанным выше методом удалось разделить суммарные основные белки на 5 фракций, которые по концентрации гуанидин гидрохлорида в элюате соответствуют фракциям: 0-фракция (негистоновые белки и белок HMG), гистон H1, гистоны Н2А и Н2В, гистоны Н3 и Н4, остаток гистонов Н3 и Н4 с примесью белков ядерного матрикса. Содержание белка в элюатах определяли методом Бредфорда в нашей модификации [4]. В работе использовался отечественный препарат гуанидин гидрохлорида («Реахим»), который предварительно был очищен. Неперекристаллизованный препарат имеет высокое поглощение в ультрафиолете. Перекристаллизация проводилась по методу, описанному Луком [1]. Концентрацию гуанидин гидрохлорида определяли рефрактометрически при комнатной температуре. Расчет концентрации велся исходя из рефрактометрического индекса [5], используя следующее соотношение:
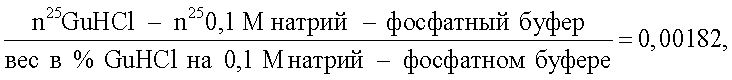
где n25GuHCl - показатель преломления гуанидин гидрохлорида (величина, зависящая от концентрации препарата); n250,1 М натрий-фосфатный буфер - показатель преломления этого буфера (величина постоянная для данной концентрации); показатель 25 указывает на температуру, при которой проводились рефрактометрические исследования.
На табл. показан аминокислотный состав фракций, полученных после элюции основных белков хроматина возрастающими концентрациями гуанидин гидрохлорида, доказывающий соответствие полученных белков гистонам Н1, Н2А и Н2В, Н3 и Н4. Что касается 0-фракции, не задержавшейся на колонке, то это ядерные белки, обогащенные лизином и аланином.
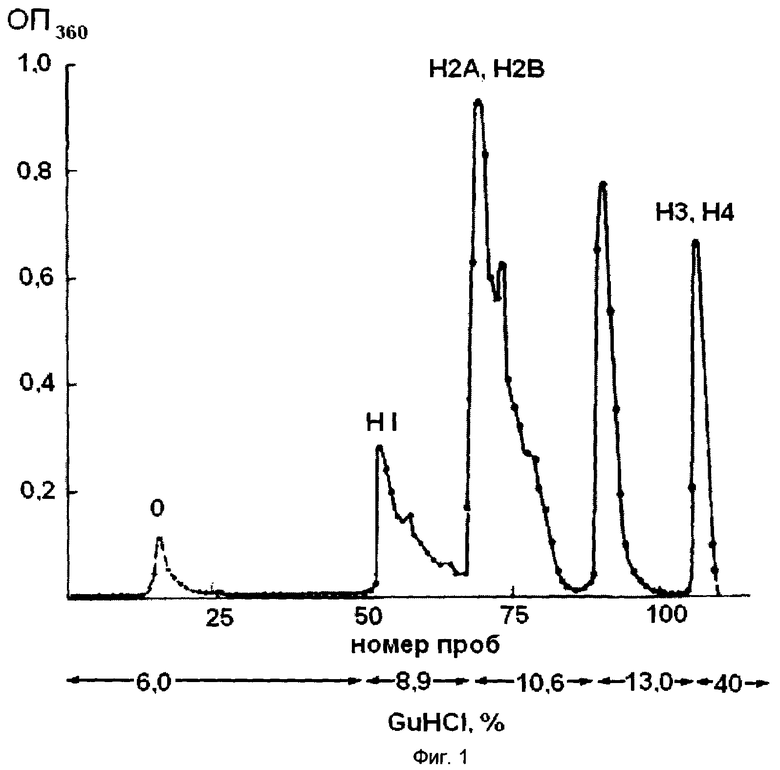
На фиг.1 показан хроматографический профиль элюции основных белков хроматина на колонке с ИРЦ-50 с помощью линейного градиента гуанидин гидрохлорида. На оси ординат указана оптическая плотность элюата, на оси абсцисс - номера проб. Из фигуры видно, что фракции гистонов выделяются при следующих концентрациях гуанидин гидрохлорида: 6%, 8,9%, 10,6%, 13%, 40% на 0,1 М калий-фосфатном буфере рН 6,8.
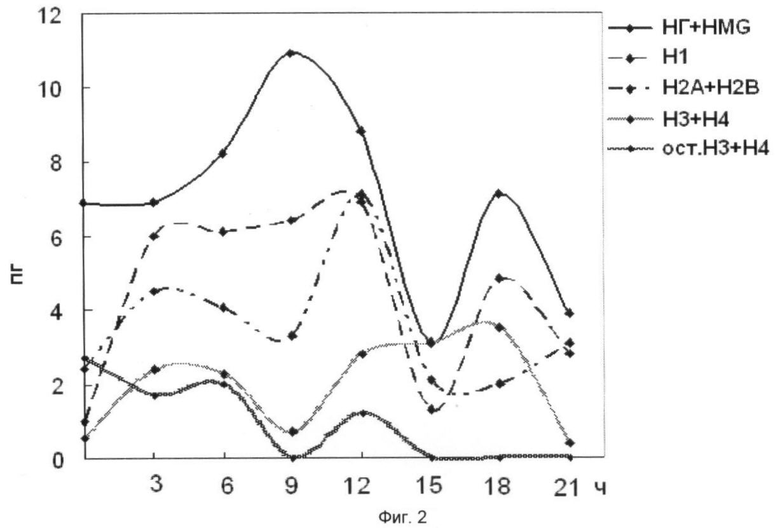
На фиг.2 представлена динамика содержания основных белков хроматина зрелых зародышей Мироновской яровой пшеницы, полученная после прерывистого градиента глицерина. Использованы следующие обозначения: НГ+HMG - негистоновые белки и белки HMG; H1, Н2А, Н2В, Н3, Н4 - гистоны. На оси абсцисс показано время прорастания зародышей пшеницы, на оси ординат - содержание белка на 1 ядро, пг.
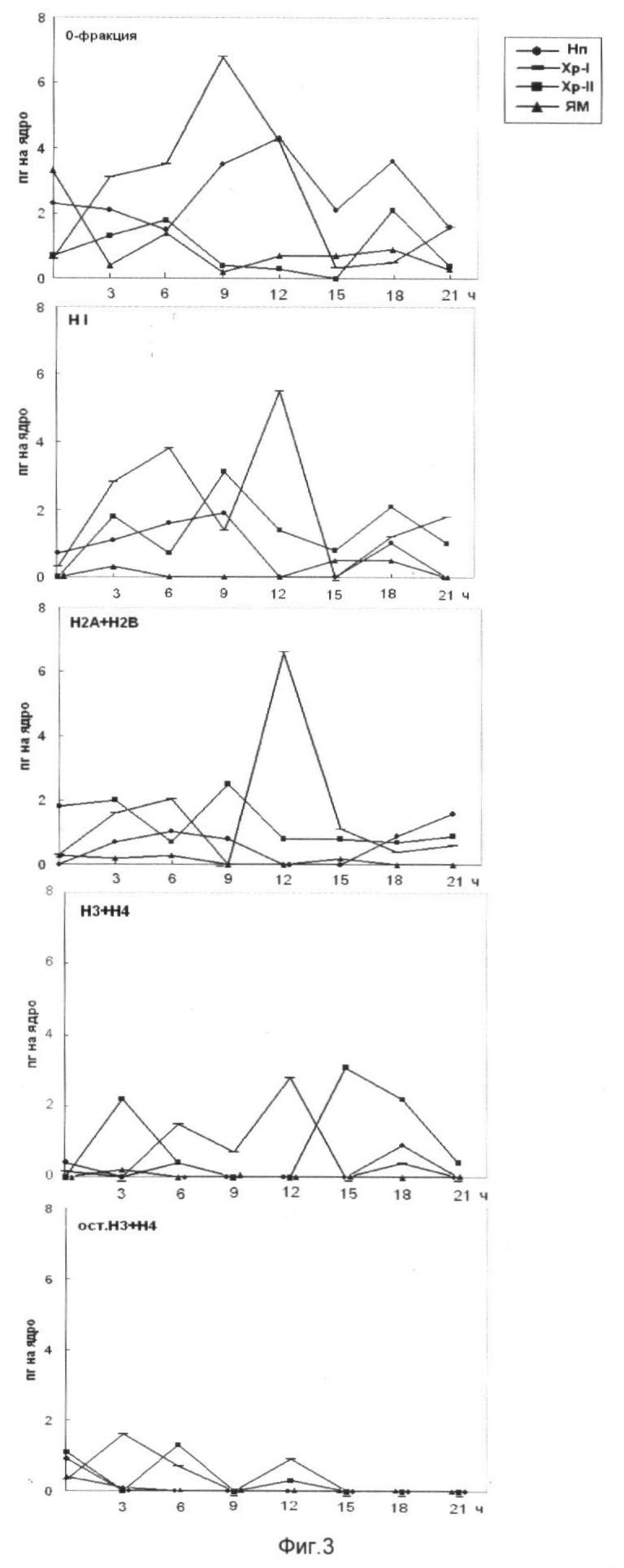
На фиг.3 представлен выход основных белков из фракций клеточных ядер зрелых зародышей Мироновской яровой пшеницы. Использованы следующие обозначения: Нп - нуклеоплазма, Xp-I - хроматин, непрочно связанный с ядерным матриксом, Хр-II - прочно связанный с ядерным матриксом, ЯМ - ядерный матрикс. HГ+HMG - негистоновые белки и белки HMG; H1, H2A, Н2В, Н3, Н4 - гистоны. На оси абсцисс показан возраст зародышей пшеницы, по оси ординат - содержание белка на 1 ядро, пг.
Анализ фиг.2 показал увеличение содержания основных белков хроматина при инициации ростовых процессов (3 ч) с последующим понижением во фракциях НГ и HMG, H1, H2A и Н2В (15 ч). Фаза 9-15 ч клеточного цикла у пшеницы интересна тем, в этот временной период наблюдается возрастание матричной доступности хроматина для транскрипции. Возрастание содержания основных белков хроматина, приходящееся на 18 ч роста, по-видимому, можно объяснить тем, что в эту фазу клеточного цикла активно осуществляются процессы репликации и переход к синтезу ДНК. На фиг.3 указана динамика содержания основных белков хроматина в супраструктурах клеточных ядер и фазы клеточного цикла, в которых содержание основных белков хроматина максимально.
Данное изобретение рекомендуется для молекулярно-генетического анализа механизмов «молекулярного» морфогенеза растений, а именно при исследовании процессов, происходящих на уровне супраструктур интерфазного клеточного ядра.
Источники информации
1. Luck J.M., Rasmussen P.S., Satake K., Tsvetikov A.N. Further studies on the fractionation of calf thymus histone //The J. of Biological Chemistry. 1958, V.233, N6, P.1407-1414.
2. Иванова Э.А. Фракционирование растительных гистонов на колонках с амберлитом ИРЦ-50// Материалы третьей научной конференции молодых ученых. Уфа: Башкирский филиал АН СССР, 1972, С.54-55.
3. Иванова Э.А., Вафина Г.Х. Способ выделения растительных клеточных ядер. Авторское свидетельство 1701747// БИ 1991. №48. С.98.
4. Иванова Э.А., Вафина Г.Х. Способ получения ядерных фракций, обладающих протеиназной и ингибирующей активностью. Авторское свидетельство 1733471 БИ 1992. №18, С.96.
5. Bonner J., Chalkley G.R., Dahmus М., Fambrough D., Fujimura F., Huang R.C., Huberman J., Jensen R., Marushige K., Ohienbusch H., Olivera В., Widholm J. Isolation and characterization of chromosomal nucleoproteins// Methods Enzymology. 1968, Acad. Press. New York. V. XII, part B, sec. V,ch.VII,P.25-31.
Из клеток растений экстрагируют ядерные фракции, из которых с помощью ионообменной хроматографии с амберлитом ИРЦ-50 в прерывистом градиенте гуанидин гидрохлорида выделяют гистоновые белки хроматина. Способ позволяет глубже изучить молекулярно-генетические механизмы структур клеточного ядра и роль белковых компонентов в их организации. 3 ил., 1 табл.
Способ препаративного выделения основных белков из супраструктур клеточных ядер растений, включающий консервацию зародышей в забуференном 80-90% глицерине при минус 25°С, изолирование клеточных ядер, проведение экстракции ядерных фракций возрастающими концентрациями 0,14 М, 0,35 М, 2 М хлористого натрия и 6 М гуанидин гидрохлорида с 0,1% β-меркаптоэтанолом и выделение из вышеперечисленных фракций основных белков хроматина с помощью ионообменной хроматографии с амберлитом ИРЦ-50 в прерывистом градиенте гуанидин гидрохлорида: 6%, 8,9%, 10,6%, 13%, 40% на 0,1 М калий-фосфатном буфере рН 6,8.
| СПОСОБ ОПРЕДЕЛЕНИЯ ОКИСЛИТЕЛЬНО-ВОССТАНОВИТЕЛЬНОЙ АКТИВНОСТИ ПЕРОКСИДАЗНОЙ СИСТЕМЫ В КЛЕТОЧНЫХ ЯДРАХ ПРОРОСТКОВ ПШЕНИЦЫ | 1993 |
|
RU2127761C1 |
| Способ получения ядерных фракций, обладающих протеиназной и ингибирующей активностью | 1990 |
|
SU1733471A1 |
| Способ выделения растительных клеточных ядер | 1990 |
|
SU1701747A1 |
| СПОСОБ ОЧИСТКИ ПРЕПАРАТА ГИСТОНА Н4 ИЗ ТКАНИ ТИМУСА | 1985 |
|
SU1319352A1 |
| Молекулярно-электронное интегрирующее устройство | 1978 |
|
SU682768A1 |

