Изобретение относится к физиологии и биохимии растений, но может быть применено в молекулярной биологии развития при оценке электрон-транспортного состояния интерфазных ядер.
Известен способ определения пероксидазной активности крови [1], в котором окислительно-восстановительная пероксидазная система была представлена из разбавленного водного раствора перекиси водорода и аликвота крови. Недостаток этого метода заключается в том, что гемин крови сам обладает пероксидазной активностью в отсутствии фермента пероксидазы.
Известен способ определения пероксидазной активности почв [2], в котором окислительно-восстановительная система in vitro была представлена из разбавленного водного раствора перекиси водорода и аликвота почвенной вытяжки. Недостаток этого метода заключается в том, что трудно установить природу проявления пероксидазной активности, то ли это окислительно-восстановительная активность - пероксидазы микроорганизмов почвы, отживших растений, грибов или животных; то ли это какие-то неорганические элементы почвы, проявляющие пероксидазную активность.
Известен способ определения активности пероксидаз в суспензионных культурах тканей табака и сои [3], в котором окислительно-восстановительная пероксидазная система in vitro была представлена из разбавленного водного раствора перекиси водорода и аликвота суспензионной культуральной жидкости. Недостаток этого метода заключается в том, что окислительно-восстановительная активность пероксидазной системы определяли в тканевых экстрактах растений, без учета эффекта и полноты извлечения пероксидазной активности отдельных органелл клеток.
Из всех перечисленных способов определения окислительно-восстановительной активности пероксидазной системы за основу был принят способ определения К.З. Гамбурга, Л.А. Подолякина, В.М. Ситнева [3], в котором для определения окислительно-восстановительной активности используют растительные тканевые экстракты табака и сои с последующей экстракцией 0,05 М ацетатным буфером pH 5,0 и 0,2 М ацетатным буфером с добавлением 1 М KCl. Недостатком способа является то, что не представлены физиологические фазы эффективного выхода окислительно-восстановительной активности пероксидазной системы растительных тканей и клеточных органелл путем их последовательной расширенной ступенчатой экстракции повышением ионной силы растворов, а также приведенные расчеты пероксидазной активности не учитывают прямолинейную зависимость образования в пероксидазной системе первого окрашенного продукта конденсации - бензидинового голубого. В приведенной формуле [3]:
A = 0,125 • V/t,
A - активность пероксидазы в условных единицах;
0,125 - заданная экстинкция;
V - объем экстракта в мл с учетом разведения;
t - время, с.
Заданность экстинкции - 0,125 может отражать начало образования бензидинового голубого--->бензидинового синего-->-бензидиново коричневого---> и т. д.
Цель изобретения - определение эффективного и полного выхода окислительно-восстановительной активности пероксидазной системы клеточных ядер проростков пшеницы, с учетом оценки прямолинейного характера ферментативной реакции.
Указанная цель достигается тем, что в способе определения ядерной окислительно-восстановительной активности пероксидазной системы из консервированных тканей проростков пшеницы выделяют в глицериновой среде клеточные ядра с последующей их промывкой 0,4 - 0,5% тритоном X-100 и последовательной ступенчатой экстракции ядерных фракций: 0,14 М; 0,35 М; 2 М хлористым натрием; 6 М гуанидингидрохлоридом + 0.1% β- меркаптоэтанолом и 0,5 н. NaOH, в которых при применении пероксидазной системы: 0,008 М H2O2 (акцептора водорода) и 0,038% бензидина (донора водорода) регистрируют образование первого продукта прямолинейной окислительно-восстановительной активности пероксидазной системы - бензидинового голубого.
Изобретение иллюстрируется следующим примером.
Пример. Опыты проводили на покоящихся, 1-2 дневных проростках, 3-4 дневных тканях колеоптилей и листьев пшениц Московской-35, Мироновской озимой и яровой. Проростки выращивали при температуре 25oС в термостатированной камере между слоями фильтровальной бумаги, смоченной дистиллированной водой. Отделенные от эндосперма зародыши, прорости, колеоптили и листья в течение 3с промывали холодным эфиром и многократно дистиллированной водой. Проростки и ткани консервировали при -25oC в 80-90% глицерине. Затем зародыши, проростки и ткани заливали гомогенизационной средой: 20% глицерин; 0,005 М MgCl2; 0,025 M KCl; 0,003 M CaCl2; 0,005 M NaCl; 0,004 M β-меркаптоэтанол; 0,02% триэтанолами (ТЭА) HCl pH 6,8; 0,004 M н-октиловый спирт. Гомогенизацию проростков проводили 7 с при 15000 об/мин (гомогенизатор МПВ-302, Польша). Гомогенат фильтровали через 1 слой фланели, флюзелина и 3 слоя капрона (размер пор 70 мкм). С грубым остатком растительного материала гомогенизацию и фильтрование проводили еще 2 раза 15 с и 30 с при 15000 об/мин. Объединенные гомогенаты центрифугировали при 500 об/мин (ЦПР-1, СССР) в течение 5 мин, с целью освобождения от грубых неразрушенных тканей и целых клеток, затем надосадок центрифугировали при 2700 об/мин (К-23, ГДР) в течение 20 мин. Осадок ядер собирали суспендированием средой гомогенизации (50 мл) и наслаивали на прерывистый глицериновый градиент, состоящий из 5 слоев (по 30 мл) возрастающей концентрации глицерина (50%, 60%, 70%, 80%, 90% вес/объем) приготовленном на ТЭА•HCl pH 6,8 буфера со всеми вышеперечисленными компонентами гомогенизационной среды, исключая 20% глицерин. Градиентное центрифугирование проводили при 2000 об/мин в течение ч (ЦПР-1, СССР). Осадок ядер промывали 0,5% тритоном Х-100 на среде гомогенизации, но без глицерина, с последующим центрифугированием при 2700 об/мин (К-23, ГДР) в течение 15 мин, после чего ядра промывали трижды в среде следующего состава: 0,005 M MgCl2; 0,025 M KCl; 0,003 M CaCl2; 0,005 M NaCl; 0,01 M трис-HCl pH 6,8 с последующим центрифугированием при вышеуказанных условиях. 1% тритоном Х-100 ядра промывать нельзя, т.к. они набухают и разваливаются.
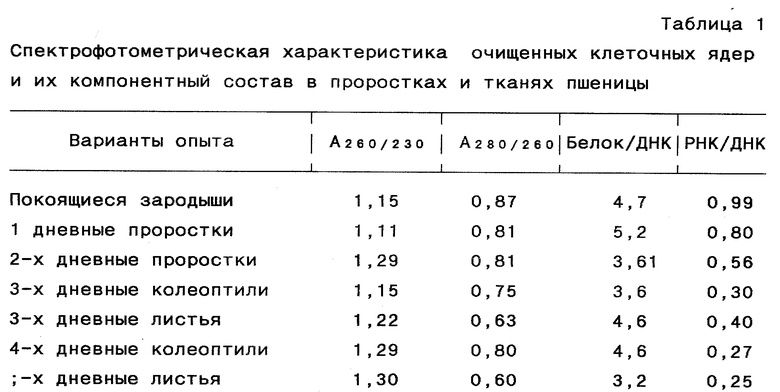
Степень чистоты выделенных препаратов клеточных ядер определяли микроскопически, спектрофотометрически и по компонентному составу клеточных ядер (белок/ДНК, РНК/ДНК) (табл. 1).
Из очищенных препаратов ядер нуклеоплазматические белки экстрагировали при низкой ионной силе 0,14 M NaCl, 0,01 M трис-HCl pH 6,8 буфером. Непрочносвязанный хроматин с ядерным матриксом выделяли путем трех последовательных экстракций 0,35 M NaCl на том же буфере.
Далее осадок фракционировали суспендированием в трис-HCl буфере с 2 M NaCl. В осадке оставалась фракция, содержащая ядерный матрикс. Последующую экстракцию проводили 6 M гуанидин гидрохлоридом с 0,1% β- меркаптоэтанолом на трис-HCl буфере pH 6,8 (ядерный матрикс), остаточные компоненты экстрагировали 0,5 н. NaOH (ядерный остаток). Все фракции ядерных компонентов получали путем трех последовательных экстракций. Достаточность экстракций контролировали по выходу белка. Ядерные фракции хранили при - 196o в азоте.
Количество белка определяли по связыванию белка с кумасси ярко-синим G (Loba, Австрия) [4]. Метод использовался в случае микро-наноколичественного определения белка: 0,3 мл кумасси (1 мг кумасси растворяли в 5 мл 85% H3PO4, доводили до 15 мл бидистиллированной водой и фильтровали для удаления нерастворившегося красителя); оптическую плотность раствора измеряли при 596 нм. Количественную оценку белка давали в соответствии с калибровочной кривой по бычьему сывороточному альбумину (марка Б, "Реахим").
Для определения ДНК и РНК использовали метод А.С. Спирина [5].
Анализ полноты выделения ядерных фракций (табл. 2) показал, что все фракции представляют собой не чистый белок, а комплексы белка и нуклеиновых кислот.
Определение окислительно-восстановительной активности пероксидазной системы в ядерных фракциях оценивали, используя в качестве субстрата o-фенилендиамин (10 мг o-фенилендиамина с добавлением 10 мкл концентрированной перекиси водорода в 25 мл 0,06 M фосфатного буфера pH 5) в лунках полистеролового планшета (Linbro, США). Через 5-10 мин по мере развития окраски реакцию останавливали добавлением 4 н. серной кислоты. Интенсивность окраски измеряли на Uniskan 11 (Labsystems) при 492 нм. Пероксидазную активность рассчитывали в условных единицах оптической плотности на 10 мкг белка (фиг. 1). Учет сорбции ядерных фракций проводили путем их инкубации в течение часа в плашках Linbro с последующей их промывкой и внесением в лунки полистеролового планшета вышеуказанных реактивов для определения пероксидазной реакции (фиг. 2). Параллельно проводили биохимическую оценку проявления окислительно-восстановительной активности пероксидазной системы в ядерных фракциях с применением субстрата бензидина (Reanal): к 0,1 мл исследуемого материала добавляли 2 мл 0,2 М ацетатного буфера с 0,038% бензидином pH 4,7; затем добавляли 1,9 мл дистиллированной воды и 1 мл 0,008 M H2O. Окраску бензидинового голубого оценивали на ФЭК-56 М (СССР) при 610 нм [1]. Расчет пероксидазной активности проводили по формуле:
где A - окислительно-восстановительная активность пероксидазной системы в условных единицах коэффициента светопропускания на 10 мкг белка•с-1;
10 - мкг белка;
τ - коэффициент светопропускания, берется по левому барабану (ФЭК-56 М), в зависимости от прямолинейности образования первичного продукта ферментативной реакции, стандартизированной по активности пероксидазы хрена (Reanal, 1979);
t -время в секундах, отмечают по секундомеру;
C - концентрация белка в мкг, взятая для анализа.
В пероксидазной системе с бензидином использовали ацетатный буфер вместо фосфатного, так как последний в присутствии бензидина дает осадок (фиг. 3).
Пероксидазная система в ацетатном буфере в присутствии o-фенилендиамина проявляет низкую ферментативную активность, поэтому применяли фосфатный буфер.
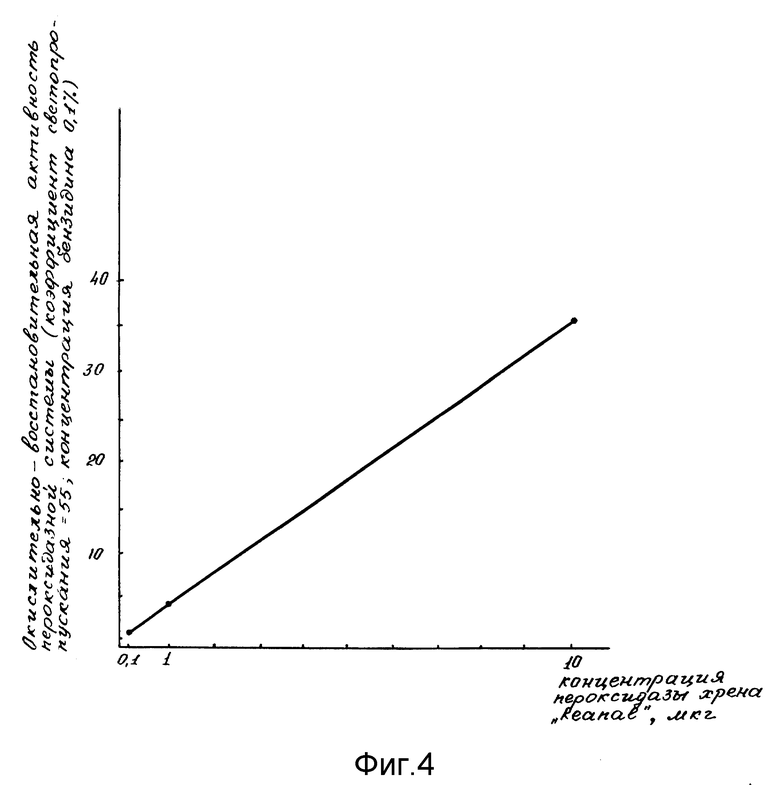
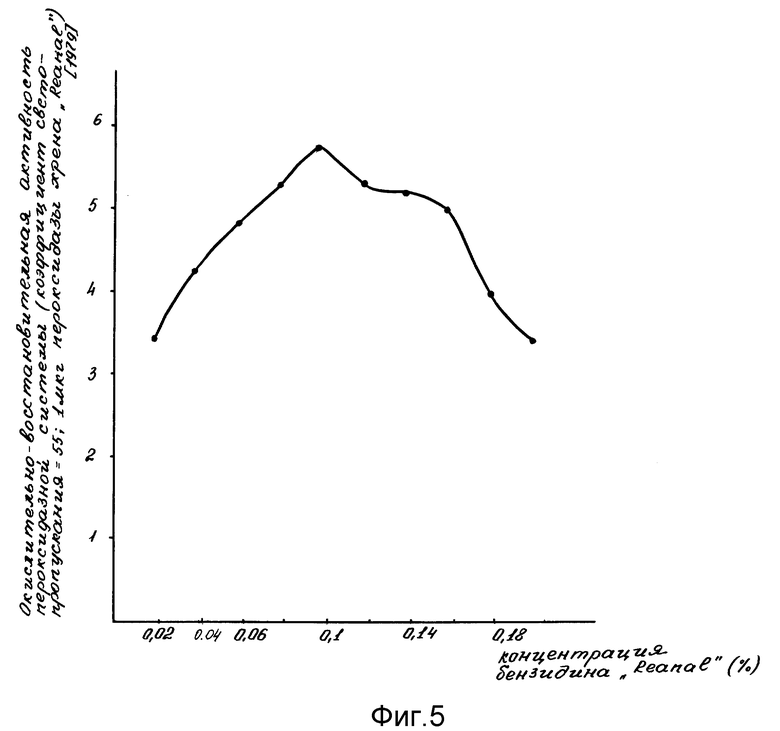
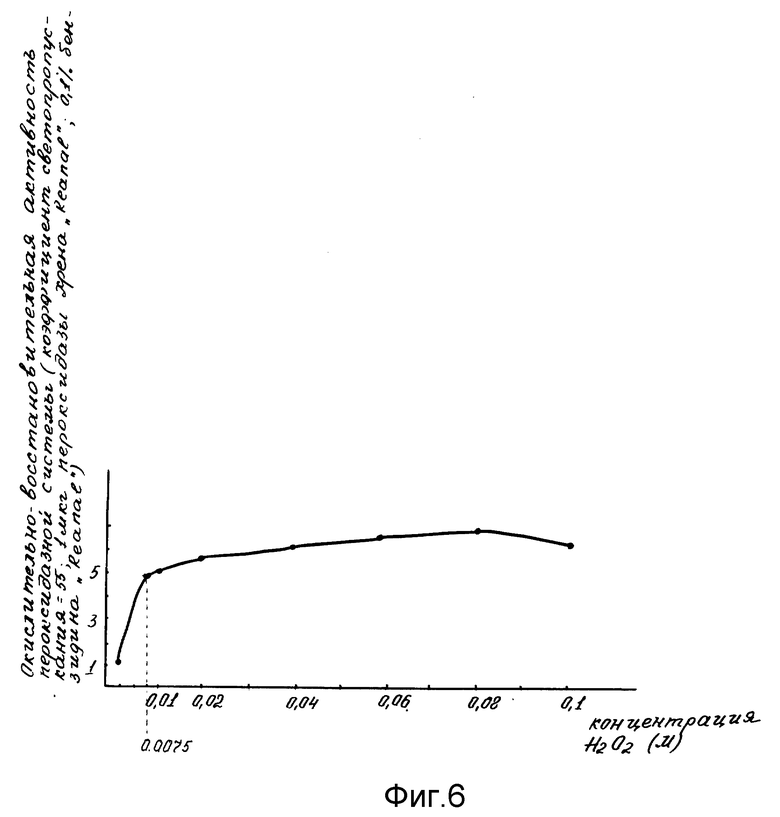
Проявление окислительно-восстановительной активности пероксидазной системы проводили в сравнении со стандартной пероксидазной системой, которую готовили 0,1 мл стандартной пероксидазы в количестве от 0,1; 1; 10 мкг (фиг. 4); 2 мл и 0,2 М ацетатного буфера с концентрацией бензидина от 0,02% до 0,2% (фиг. 5) pH 4,7; затем добавляли 1,9 мл дистиллированной воды и 1 мл от 0,01 до 0,1 М H2O2 (фиг. 6). Прямолинейную реакцию образования бензидинового голубого оценивали на ФЭК-56 М (СССР) при 610 нм.
Расчет пероксидазной активности проводили по формуле: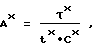
где Ax - активность стандартной пероксидазы (Reanal) в условных единицах коэффициента светопропускания в с-1;
τx - коэффициент светопропускания берется на прямолинейному участке ферментативной реакции образования бензидинового голубого в пределах 5 секундной реакции. Этот показатель (коэффициент светопропускания левого барабана ФЭК-56 М) реакции стандартной пероксидазы (Reanal), мы использовали для опытных реакций окислительно-восстановительной активности пероксидазной системы клеточных ядер;
tx - время в секундах: (обычно концентрацию стандартной пероксидазы выбираем таким образом, чтобы время реакции образования бензидинового голубого было в пределах 5-10 сек; с учетом этого времени и подбирается коэффициент светопропускания τx = τ;
Cx - концентрация стандартной пероксидазы (Reanal), взятой для анализа.
Оптимальные процентные концентрации бензидина, H2O2 и минимальная чувствительность к концентрации стандартной пероксидазы (Reanal) были подобраны в специальных опытах (фиг. 4-6). На оси ординат фиг.4-6 указана пероксидазная активность в условных единицах коэффициента светопропускания; на оси абсцисс фиг. 4 указана концентрация стандартной пероксидазы из хрена; на фиг. 5 - концентрации бензидина; на фиг. 6 - концентрации H2O2.
На фиг. 7 по оси ординат указана оптическая плотность чувствительности определения белка по методу Бредфорд; по оси абсцисс указано количество стандартной пероксидазы, взятой в эксперимент.
Числа в таблицах и точки на графиках представляют собой среднеарифметическое не менее 3-5-ти аналитических повторов из 2-х биологических.
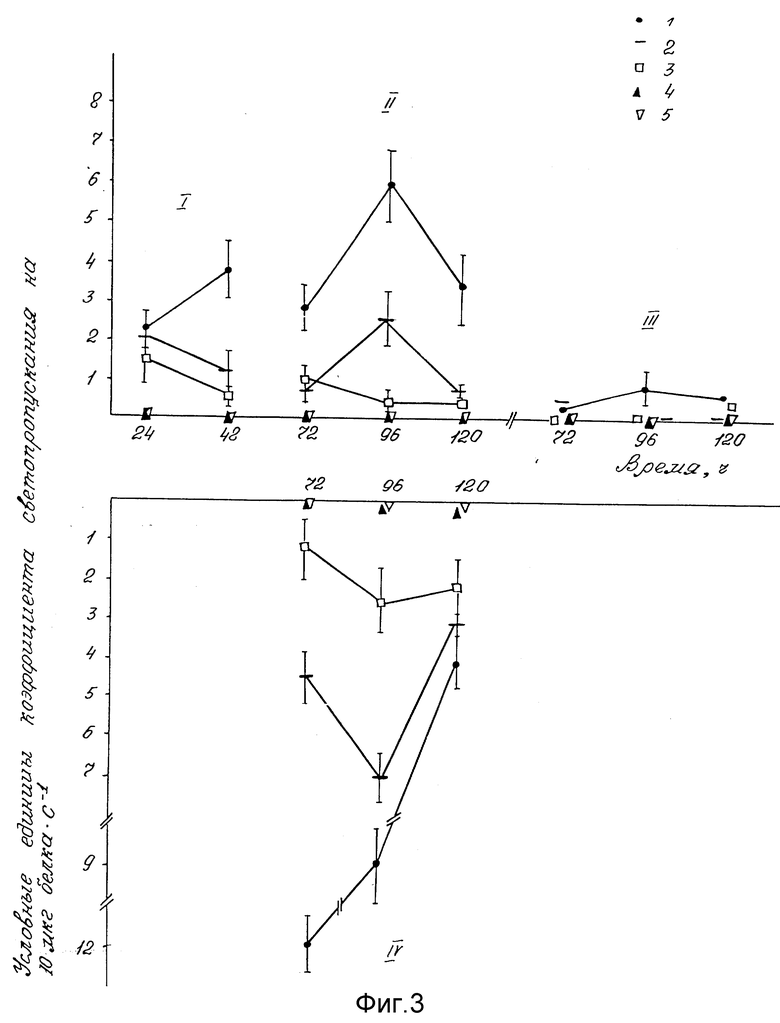
Эффективность последовательной ступенчатой экстракции ядерной окислительно-восстановительной активности пероксидазной системы продемонстрирована на фиг. 1, 3.
На оси ординат фиг. 1, 2 указаны условные единицы оптической плотности на 10 мкг белка.
На оси ординат фиг. 3 указаны условные единицы коэффициента светопропускания на 10 мкг белка•с-1.
На оси абсцисс фиг. 1, 2, 3 указан физиологический возраст проростков от (0) покоящихся зародышей до 2-х суточных проростков; 3-5-ти суточные колеоптили; 3-5-ти суточные листья; 3-5-ти суточные корни. В фигурах даны следующие обозначения: 1 - нуклеоплазма (0,14 M NaCl); 2 - хроматин, непрочно связанный с ядерным матриксом (0,35 M NaCl); 3 - хроматин, прочно связанный с ядерным матриксом (2 M NaCl); 4 - ядерный матрикс (6 M гуанидин гидрохлорид + 0,1% β- меркаптоэтанол); 5 - ядерный остаток (0,5 н. NaOH. I - (0 ч, 24 ч, 48 ч) целые зародыши и проростки; II - (72 ч, 96 ч, 120 ч) - колеоптили; III - (72 ч, 96 ч, 120 ч) - первый настоящий лист; IV - (72 ч, 96 ч, 120 ч) - корневая система.
Сравнение окислительно-восстановительной активности пероксидазной системы в пробирках (фиг. 3) и иммунологических плашках (фиг. 1) показало, что пробирочный метод более точный, так как дает возможность точно (по секундомеру) регистрировать прямолинейный участок образования первого продукта реакции пероксидазной системы - бензидинового голубого [6]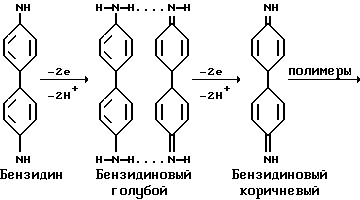
В случае определения пероксидазной активности на одной плашке (фиг. 1) (~ 96 проб) трудно для каждой пробы точно оценить время образования первого окрашенного продукта конденсации соответствующего прямолинейному участку зависимости активности фермента от времени его инкубации в плашке.
С целью доказательства сорбции ядерных фракций в плашках Linbro (фиг. 2) в связи с этим, возможной неэкранируемости активного центра фермента, был проведен эксперимент на возможность сорбции ядерных фракций. Эксперимент показал, за исключением слабой сорбции фракции непрочносвязанной с хроматином 96 ч колеоптилей и корней; что ядерные фракции не сорбируются в плашках Linbro.
Исследование ядерных фракций, обладающих окислительно-восстановительной активностью в пероксидазной системе, включает предварительную консервацию растительных тканей и выделение из них клеточных ядер в глицериновой среде с последующей ступенчатой экстракцией ядерных фракций при повышении ионной силы растворов, в которых при применении пероксидазной системы: 0,008 M H2O2 и 0,038% бензидина учитывали эффективность и полноту определения окислительно-восстановительной активности ядер. Таблица 1 иллюстрирует, что структура клеточных ядер по содержанию генетического материала не нарушена; таблица 2 иллюстрирует полноту выделения ядерных экстракций.
Фиг. 1 иллюстрирует влияние ступенчатой последовательности экстракций ядерных фракций на выход окислительно-восстановительной активности пероксидазной системы при определенном времени (5 мин) инкубации фермента с субстратом для всех 96 проб. Эти данные свидетельствуют о том, что минимальное время (5 мин) для окислительно-восстановительной активности в 96 пробах плашки очень велико, так как прямолинейная ферментативная реакция для многих проб практически заканчивается в пределах от 5 до 15 сек, а иногда и меньше. Мы считаем, что этот способ масштабного определения окислительно-восстановительной активности пероксидазной системы не всегда корректен.
Фиг. 2 иллюстрирует наличие сорбции ядерных фракций на иммунологических плашках Linbro.
Фиг. 3 иллюстрирует влияние физиологического возраста растений и ступенчатой последовательности экстракций ядерных фракций повышающими ионными силами растворов на выход окислительно-восстановительной активности пероксидазной системы при точном времени (по секундомеру) определении образования первого продукта прямолинейной реакции для каждой пробы.
Фиг. 4 иллюстрирует минимальную концентрацию пероксидазы хрена, для проявления пероксидазной активности.
Фиг. 5 иллюстрирует оптимальное количество бензидина, необходимого для проявления окислительно-восстановительной активности пероксидазной системы.
Фиг. 6 иллюстрирует концентрацию H2O2, необходимую для проведения оптимальной окислительно-восстановительной активности пероксидазной системы.
Фиг. 7 иллюстрирует чувствительность метода Бредфорд [см. 7] определяющего количество белка в стандартной пероксидазе (Reanal).
Предложенный способ рекомендуется в исследовании молекулярно-генетических механизмов эндогенной регуляции организации генетических, морфологических программ развития организма на разных онтогенетических стадиях дифференцированного роста растений.
Источники информации
1. Иванова Э.А. Метод определения пероксидазной активности в крови для внедрения в практику клинико-биохимических лабораторий // Удостоверение на рационализаторское предложение N 59 (331), выданное Башкирским мединститутом, г. Уфа, от 16 декабря 1980 г.
2. Иванова Э.А., Хазиев Ф.Х. Пероксидазная активность почв // Депонирована в ВИНИТИ. М.: N 6941, 1983.
3. Гамбург К.З., Подолякина Л.А., Ситнева В.М. Изучение активности пероксидазы и ИУК-оксидазы в суспензионных культурах тканей табака и сои // Физиология растений. 1977, Т. 24, N 3, с. 542.
4. Иванова Э.А., Вафина Г.Х. Методика выделения протеиназ и ингибиторов из клеточных ядер проростков пшеницы // Физиология растений, 1990, Т. 37, С. 609.
5. Спирин А.С. Спектрофотометрическое определение суммарного количества нуклеиновых кислот // Биохимия, 1968, Т. 23, N 5, с. 656.
6. Саундерс Б.К. Пероксидазы и каталазы // Неорганическая биохимия. М.: Мир, 1978. Т. 2, С. 434.
7. Скоупс Р. Методы очистки белков. - М.: Мир, 1985, С. 342.



Изобретение относится к области физиологии и биохимии растений. Из предварительно консервированных при -25oC в глицериновой среде тканей проростков пшеницы выделяют клеточные ядра, подвергают их ступенчатой экстракции 0,14 М; 0,35 М; 2 М NaCl, 6 M гуанидинхлоридом с 0,1%-ным β-MЭ и 0,5 н. NаOH и получают фракции, в которых затем определяют пероксидазную активность с бензидином в присутствии H2O2 в интервале времени 5 - 10 с от начала ферментативной реакции. Предложенный способ обеспечивает повышение точности и чувствительности анализа за счет полноты извлечения ферментсодержащего ядерного материала и выбора оптимальных условий проведения цветной количественной реакции. 2 табл., 7 ил.
Способ определения окислительно-восстановительной активности пероксидазной системы в клеточных ядрах проростков пшеницы, предусматривающий консервирование тканей проростков на гетеротрофной фазе роста в 80 - 90% глицерине при -25oС, их гомогенизацию, выделение клеточных ядер, последовательную экстракцию ядерного материала 0,14 М, 0,35 М и 2 М NaCl, 6 М гуанидин гидрохлоридом с 0,1% β-меркаптоэтанолом и 0,5 н NaOH с получением соответствующих фракций, которые используют для определения пероксидазной активности путем колориметрирования при 610 нм проб, содержащих, кроме исследуемого образца фракции, 0,038% бензидин и 0,008 М H2O2 в ацетатном буфере при pH 4,7, а расчет проводят по формуле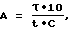
где А - пероксидазная активность, рассчитанная на 10 мкг белка, в условных единицах коэффициента светопропускания, с-1;
τ - устанавливаемая на барабане колориметра величина коэффициента светопропускания, предварительно определяемая в реакции с коммерческим препаратом пероксидазы хрена, проводимой в течение 5 - 10 с в аналогичных условиях;
t - измеряемое по секундомеру время реакции, сек;
С - концентрация белка в анализируемой пробе, мкг.
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Гамбург К.З | |||
| и др | |||
| Изучение активности пероксидазы и ИУК - оксидазы в суспензионных культурах тканей табака и сои | |||
| Физиология растений | |||
| Шеститрубный элемент пароперегревателя в жаровых трубках | 1918 |
|
SU1977A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Методы биохимического исследования растений./Под ред | |||
| А.И.Ермакова | |||
| - Л.: Здоровье, 1987, с.42-43 | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| SU, авторское свидетельство 1733471, A1, C 12 N 9/50, 1992. | |||
