Настоящее изобретение относится к новому способу получения эволюционирующего микроорганизма для получения 1,2-пропандиола, эволюционирующему микроорганизму, полученному таким способом и его применению для получения 1,2-пропандиола.
1,2-Пропандиол или пропиленгликоль, С3-гликоль, является химическим продуктом, применяемым во многих областях. Он является компонентом ненасыщенных полиэфирных смол, жидких детергентов, хладагентов, антифризов и антиобледенительной жидкости для самолетов. С 1993-1994 года пропиленгликоль стали применять вместо этиленовых производных, которые были признаны более токсичными, чем пропиленовые производные.
В настоящее время 1,2-пропандиол получают химическим путем способом гидратации пропиленоксида с использованием большого количества воды. Пропиленоксид можно получить двумя способами: в одном используют эпихлоридин, в другом - гидропероксид. В обоих способах используют высокотоксичные продукты. Кроме того, в способе с использованием гидропероксида образуются побочные продукты, такие как трет-бутанол или 1-фенилэтанол, которые необходимо использовать, чтобы производство пропиленоксида было рентабельным. Химическим способом обычно получают рацемический 1,2-пропандиол, поскольку существуют две формы стереоизомеров: (R) 1,2-пропандиол и (S) 1,2-пропандиол, применение которых в других областях представляет интерес.
Биологический путь получения 1,2-пропандиола представляет собой перспективную альтернативу химическому пути получения в связи с указанными выше недостатками последнего. Многие микроорганизмы способны к естественному продуцированию (R) или (S) 1,2-пропандиола из сахара, такого как глюкоза или ксилоза, метаболизируя их путем гликолиза, или из деоксигексоз с образованием (S) 1,2-пропандиола (Cameron D.C. et coll. (1998) Biotechnol.Prog.). Из наиболее эффективных микроорганизмов следует назвать Clostridium sphenoides (Tran Din K. еt coll. 1986) Thermoanaerobium thermosaccharolyticum (Altaras N.E. and Cameron D.C 2001). Последний способен превращать путем ферментации несколько видов сахаров в (R) 1,2-пропандиол с выходом от 0,13 до 0,28 г полученного 1,2-пропандиола/г использованной глюкозы. Ферменты, ответственные за синтез 1,2-пропандиола в этих двух микроорганизмах, не были идентифицированы и улучшение их свойств ограничено недостатком имеющихся генетических инструментов. К тому же E. coli имеет все гены, необходимые для производства 1,2-пропандиола, даже если E. coli не продуцирует 1,2-пропандиол естественным путем. Действительно 1,2-пропандиол должен продуцироваться из метилглиоксаля, соединения, даже в малой концентрации высокотоксичного для клетки. Способы, в которых используют штаммы E. coli, генетически модифицированные для того, чтобы продуцировать 1,2-пропандиол, были также описаны, в частности, в US 6303352, US 6087140 и WO 98/37204. В этих способах, в частности, используют сверхэкспрессию одного или нескольких ферментов, участвующих в метаболическом пути продуцирования 1,2-пропандиола, путем клонирования их генов в плазмидах и, следовательно, оказывают селекционное давление с помощью антибиотиков. Для улучшения свойств штаммов некоторые эндогенные гены также подвергают делеции (см. например, Altaras N.E. and Cameron D.C. (2000) Biotechnol. Prog. 16, 940-946: Altaras N.E. and Cameron D.C. (1999) Appl. Env. Microb., 65, 1180-1185).
Способ с использованием эволюционирующего микроорганизма, продуцирующего совместно 1,2-пропандиол и ацетон, два ценных продукта, до настоящего времени не был описан.
Таким образом настоящее изобретение относится к способу получения штамма эволюционирующих микроорганизмов для продуцирования 1,2-пропандиола путем метоболизма источника простого углерода, причем указанный способ включает культивирование в условиях селекционного воздействия в соответствующей культуральной среде, содержащей источник простого углерода, исходного бактериального штамма, имеющего делецию гена tpiA и делецию по меньшей мере одного гена, участвующего в превращении метилглиоксаля (пропаналя) в лактат для эволюционирования в указанном исходном штамме одного или нескольких генов, участвующих в пути биосинтеза DHAP в метилглиоксаль, затем в 1,2-пропандиол, до получения эволюционирующих генов, обладающих более высокой активностью «1,2-пропандиолсинтазы», затем селекционируют и выделяют один или несколько штаммов эволюционирующих микроорганизмов, обладающих более высокой активностью «1,2-пропандиолсинтазы».
Ген tpiA кодирует триозафосфат изомеразу, которая катализирует превращение DHPA в глицериновый альдегид-3-фосфат. Делеция этого гена обеспечивает синтез достаточного количества метилглиоксаля. Теоретически делеция гена tpiA должна обеспечивать, чтобы 50% углерода глюкозы, метаболизированного клетками, использовались для получения метилглиоксаля из дигидроксиацетонфосфата.
Делеция по меньшей мере одного гена, участвующего в превращении метилглиоксаля (пропаналя) в лактат, ингибирует превращение метилглиоксаля в лактат так, что основная часть метилглиоксаля, присутствующего и продуцированного исходным штаммом, таким как полученный эволюционирующий штамм, использовалась клеточным механизмом указанных штаммов для получения 1,2-пропандиола.
Гены, участвующие в превращении метилглиоксаля в лактат, выбирают из гена gloA, кодирующего глиоксилазу I (катализирующего синтез лактоилглютатиона из метилглиоксаля), и генов aldA и aldB, кодирующих лактальдегиддегидрогеназу (катализирующую синтез (S) лактата из (S) лактальдегида). Предпочтительно исходный штамм имеет делецию трех генов gloA, aldA и aldB.
Преимущественно проводят дополнительное модифицирование исходного штамма, заключающееся в том, что устраняют естественные пути ферментации глюкозы, потребляющие эквиваленты восстановителей в виде NADH с тем, чтобы устранить эти пути метаболизма, конкурирующие с продуцированием 1,2-пропандиола.
В частности, следует упомянуть делецию гена IdhA, кодирующего лактатдегидрогеназу, катализирующую синтез лактата из пирувата, и гена adhE, кодирующего спирт-альдегиддегидрогеназу, катализирующую синтез этанола из ацетил-СоА.
Также можно заставить микроорганизм использовать комплекс пируватдегидрогеназы для продуцирования путем анаэробиоза ацетил-СоА и NADH из пирувата. Этого можно достичь за счет делеции генов pflA и pflB, кодирующих пируватформиатлиазу.
В соответствии с частным способом осуществления изобретения исходный штамм таким образом имеет также делецию по меньшей мере в одном гене, выбранном из IdhA, pflA, pflB и adhE, предпочтительно делецию в четырех генах IdhA, pflA, pflB и adhE.
Еще более предпочтительно исходный штамм согласно изобретению содержит также по меньшей мере один ген, кодирующий фермент, способствующий метаболизму пирувата в ацетат посредством анаэробиоза.
Предпочтительно фермент способствует метаболизму пирувата до продуцирования ацетил-СоА и NADH в условиях анаэробиоза. Более предпочтительно этот фермент является комплексом пируватдегидрогеназы.
Преимущественно указанный ген, кодирующий фермент, способствующий метаболизму пирувата в ацетат посредством анаэробиоза, слабочувствителен к ингибированию посредством NADH.
Этот ген может быть эндогенным геном, кодирующим эндогенный белок, или экзогенным или гетерологичным геном, кодирующим эндогенный или экзогенный фермент.
В случае эндогенного гена, кодирующего эндогенный белок, чувствительный к ингибированию посредством NADH, способ эволюционирования согласно изобретению позволяет селекционировать штаммы, обладающие повышенной активностью “1,2-пропандиолсинтазы”, указанный ген которых, кодирующий фермент, способствующий метаболизму пирувата в ацетат в условиях анаэробиоза, кодирует эволюционирующий фермент, ставший слабочувствительным к ингибированию посредством NADH.
В соответствии с другим вариантом осуществления изобретения можно вводить в исходный штамм гетерологичный ген, кодирующий фермент, слабочувствительный к ингибированию посредством NADH, или кодирующий фермент, чувствительный, но ставший слабочувствительным в результате осуществления способа эволюционирования согласно изобретению.
Кроме того, предпочтительной также является делеция гена edd, кодирующего 6-фосфоглюконатдегидратазу, первый фермент, участвующий в реализации пути Энтера-Дурова, с тем, чтобы избежать прямой метаболизации глюкозы в глицериновый альдегид-3-фосфат и пируват и вызвать таким образом превращение глюкозы в 1,2-пропандиол и ацетат.
Преимущественно в предварительно выделенный эволюционирующий штамм, полученный способом эволюционирования согласно изобретению, вводят один или несколько гетерологичных генов, кодирующих один или несколько ферментов, участвующих в превращении ацетил-СоА и ацетата в ацетон для получения эволюционирующего модифицированного штамма.
Это новое модифицирование позволяет продуцировать совместно с 1,2-пропандиолом ацетон - совместный ценный продукт. Кроме того, преимуществом этого модифицирования является повышение эффективности продуцирования 1,2-пропандиола. Действительно, ацетат является соединением, ингибирующим бактериальный рост при слабой концентрации (15 г/л), и быстро блокирует эволюционирование свойств штамма, культивируемого в хемостате в анаэробных условиях.
Введение в эволюционирующий штамм генов, кодирующих ферменты, катализирующие трансформацию ацетата в ацетон, вызывает уменьшение остаточной концентрации ацетата при культивировании в хемостате. Получают ацетон - соединение, в значительно меньшей степени ингибирующее рост, чем ацетат, способствуя при этом росту штамма и продуцированию 1,2-пропандиола.
Преимущественно один или несколько гетерологичных генов, кодирующих один или несколько ферментов, участвующих в превращении ацетил СоА и ацетата, получают из C. аcetobutylicum. Гены, кодирующие один или несколько ферментов, участвующих в превращении ацетил-СоА и ацетата в ацетон, могут иметь хромосомную или внехромосомную экспрессию. При хромосомной экспрессии одна или несколько копий могут быть введены в геном методами рекомбинации, известными специалисту в данной области. При внехромосомной экспрессии гены могут нести разные типы плазмид, которые отличаются своей ориджин репликацией, своим числом копий и своей стабильностью в клетке. Они могут присутствовать в количестве от 1 до 5 копий, до 20 или более чем 500 копий, соответствуя плазмидам с малым числом копий с ориджин репликацией типа strict (pSC101, RK2), плазмидам с малым числом копий (pACYC, pRSF1010) или с большим числом копий (pSK bluescript II). Гены могут иметь экспрессию с использованием промоторов различных типов, индуцибельных или неиндуцибельных. Можно, например, назвать промоторы Ptrc, Ptac, Plac, или другие промоторы, известные специалисту. Экспрессию генов-мишеней можно увеличить или уменьшить с помощью элементов, стабилизирующих или дестабилизирующих ARN мессенджер (Carrier and Keasling (1998) Biotechnol. Prog., 15, 58-64) или белки (GSTtags, Amsterham Biosciences).
В соответствии с предпочтительным вариантом осуществления изобретения эволюционирующий модифицированный штамм, полученный ранее в условиях селекционного воздействия, культивируют в соответствующей культуральной среде, содержащей источник простого углерода, с тем, чтобы вызвать в указанном эволюционированном модифицированном штамме эволюционирование одного или нескольких генов, участвующих в превращении ацетил-СоА и ацетата в ацетон, до более высокой активности “ацетонсинтазы”, затем проводят отбор и выделяют штаммы эволюционирующих микроорганизмов второго поколения, обладающих более высокой активностью «1,2-пропандиолсинтазы» и более высокой активностью “ацетонсинтазы”.
Настоящее изобретение относится также к исходному штамму согласно изобретению, такому, как описан выше, ниже и в примерах.
Оно относится также к эволюционирующему штамму, обладающему более высокой активностью «1,2-пропандиолсинтазы», который можно получить способом согласно изобретению, такому, как описан выше и ниже в примерах, причем это определение охватывает эволюционирующие штаммы второго поколения, которые обладают, кроме того, более высокой активностью “ацетонсинтазы”.
Изобретение относится также к способу получения 1,2-пропандиола, в ходе которого культивируют эволюционирующий штамм согласно изобретению в соответствующей культуральной среде, содержащей источник простого углерода, затем выделяют полученный 1,2-пропандиол и в случае необходимости ацетон, которые возможно подвергают очистке.
Штаммы модифицированных, исходных и эволюционирующих микроорганизмов согласно изобретению могут быть прокариотическими или эукариотическими, которые можно трансформировать и культивировать в целях получения 1,2-пропандиола и в случае необходимости ацетона.
Специалист сможет выбирать указанные микроорганизмы в соответствии с уровнем своих общих знаний в области молекулярно-клеточной биологии и в случае необходимости идентифицировать гены этих микроорганизмов, соответствующие генам E. coli, упомянутым выше.
Под штаммом микроорганизма понимают согласно изобретению совокупность микроорганизмов одного вида, содержащую по меньшей мере один микроорганизм указанного вида. Таким образом признаки, описанные в отношении одного штамма, относятся к каждому из микроорганизмов указанного штамма. Также признаки, описанные в отношении одного из микроорганизмов штамма, относятся к совокупности указанных микроорганизмов, образующих штамм.
Модифицированные микроорганизмы согласно изобретению выбирают из бактерий, дрожжей и грибов, относящихся, в частности, к следующим видам: Aspergillus sp., Bacillus sp., Brevibacterium sp., Clostridium sp., Corynebacterium sp., Escherichia sp., Gluconobacter sp., Pseudomonas sp., Rhodococcus sp., Saccharomyces sp., Streptomyces sp., Xanthomonas sp., Candida sp.
В соответствии с предпочтительным вариантом осуществления изобретения бактериальный штамм является штаммом Escherichia, в частности, относящимся к E.coli. В соответствии с другим вариантом осуществления бактериальный штамм является штаммом Corynebacterium, в частности, C.glutamicum.
В соответствии с другим вариантом осуществления штамм дрожжей является штаммом Saccharomyces, в частности, S.cerevisiae.
Изобретение описано выше, ниже и в примерах в отношении E. coli. Таким образом, использование названия гена E. coli относится главным образом к генам, способным к делеции или сверхэкспрессии в эволюционирующих штаммах согласно изобретению. Однако такое использование имеет более широкое значение согласно изобретению и охватывает соответствующие гены других микроорганизмов. Действительно, используя ссылки GenBank на гены E. coli, специалист в данной области может определить гены, являющиеся эквивалентами в других бактериальных штаммах, кроме E. coli.
Средства идентификации гомологических последовательностей и их процента гомологии хорошо известны специалисту, к ним, в частности, относятся программы BLAST, которыми можно воспользоваться на сайте http://www.ncbi.nlm.gov/BLAST/ с параметрами, указанными по умолчанию на этом сайте. Полученные последовательности можно применять (выравнивать) с помощью программ СLUSTERW (http://www.ebi.ac.uk/clustalw/) или MULTALIN (http://www.prodes.toulouse.inra.fr/multalin/cgi-bin/multalin.pl), с параметрами, указанными по умолчанию на этом сайте.
Используя ссылки, указанные в GenBank в отношении известных генов, специалист может определять эквивалентные гены в других организмах, штаммах бактерий, дрожжах, грибах, млекопитающих, растениях и т.д. Эту рутинную работу преимущественно проводят с использованием последовательностей согласия, которые можно определять, выстраивая последовательности генов, полученных из других микроорганизмов, и изображая вырожденные, позволяющие клонировать соответствующий ген в другом организме. Эти рутинные технологии в области молекулярной биологии хорошо известны из уровня техники и описаны, например, Sambrook и др. (1989 Molecular cloning: a laboratory manual. 2 Ed.Cold Spring Harbor Lab., Cold Spring Harbor, New York).
Под “делецией” согласно изобретению понимают супрессию активности “делетированного” гена. Эта супрессия может являться инактивацией продукта экспрессии рассматриваемого гена соответствующим методом или ингибированием экспрессии рассматриваемого гена или делецией по меньшей мере части рассматриваемого гена таким образом, чтобы или его экспрессии не было (например, делеция части или всей промоторной области, необходимой для его экспресcии), или чтобы продукт экспрессии утратил свою функцию (например, делеция в кодирующей части определенного гена). Предпочтительно делеция гена содержит супрессию основной части указанного гена и в случае необходимости его замену геном-маркером селекции, что позволяет облегчить идентификацию, выделение и очистку эволюционирующих штаммов согласно изобретению.
Инактивация гена преимущественно происходит путем гомологической рекомбинации (Datsenko K.A., Wanner B.L. (2000) One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 97: 6640-6645). Следует кратко напомнить принцип протокола: фрагмент линейной ДНК вводят в клетку, указанный фрагмент был получен in vitro, содержит оба участка, находящиеся по обе стороны гена, и по меньшей мере один селекционный ген между этими двумя участками (обычно ген, резистентный к антибиотику), причем указанный линейный фрагмент, таким образом, является инактивированным геном. Клетки, подвергнувшиеся рекомбинации и интегрировавшие введенный фрагмент, отбирают путем распределения на селекционной среде. Затем отбирают клетки, подвергшиеся двойной рекомбинации, в которых исходный ген был заменен инактивированным геном. Такой протокол можно усовершенствовать, используя позитивную и негативную системы селекции для ускорения обнаружения двойных рекомбинаций.
Предпочтительно для введения этих генов в штамм используют метод электропорации, хорошо известный специалисту. Следует напомнить протокол: представляющие интерес гетерологичные гены клонируют в векторе экспресcии между промотором и терминатором. Этот вектор имеет, кроме того, ген, резистентный к антибиотику, позволяющий селекционировать клетки, его содержащие, и функциональный ориджин репликации в штамме клетки хозяина с тем, чтобы он мог ее сохранить. Протокол требует получения электрокомпетентных клеток хозяев, которые в дальнейшем трансформируются путем электропорации с помощью вектора.
Согласно изобретению гены, введенные путем электропорации, предпочтительно являются генами adc, ctfA и В, thl, кодирующими соответственно ацетоацетаткарбоксилазу, коэнзим А трансферазу и тиолазу, естественного пути получения ацетона из Clostridium acetobutylicum - микроорганизма, признанного чрезвычайно эффективным для получения ацетона биологическим путем.
Способ эволюционирования согласно изобретению представляет собой способ получения эволюционирующих микроорганизмов, позволяющий модифицировать пути метаболизма, содержащий преимущественно следующие стадии:
а) модифицирование микроорганизма для получения исходного микроорганизма таким образом, чтобы ингибировать продуцирование или потребление метаболита, т.е. потребляемого или продуцируемого, если клетки исходного микроорганизма культивируют на определенной среде,
b) культивирование исходных модифицированных микроорганизмов, полученных на предыдущей стадии на указанной определенной среде для ее эволюционирования, при этом определенная среда может также содержать косубстрат, необходимый для этого эволюционирования,
c) селекция клеток модифицированных микроорганизмов, способных развиваться на определенной среде, возможно с косубстратом.
В данном случае эволюционирующий путь метаболизма является путем биосинтеза 1,2-пропандиола и в случае необходимости путем биосинтеза ацетона.
Под «определенной средой» согласно изобретению понимают среду известного молекулярного состава, адаптированную к выращиванию микроорганизма. Определенная среда по существу не содержит метаболит, продуцирование или потребление которого устраняют путем модифицирования.
Под «косубстратом» согласно изобретению понимают углеродную или неуглеродную молекулу, отличную от субстрата, которая участвует в реакции и дает один или несколько атомов субстрату для формирования продукта. Косубстрат не обладает признанным мутагенным свойством.
Под «селекцией» согласно изобретению понимают способ культивирования, возможно непрерывный, осуществляемый с применением повышающихся степеней разведения таким образом, что в культуральной среде сохраняются только микроорганизмы, степень роста которых равна или больше используемой степени разведения. Таким образом сохраняют микроорганизмы, которые эволюционировали так, что проведенное модифицирование больше не влияет на рост.
Под «эволюционирующим геном» согласно изобретению понимают последовательность нуклеиновой кислоты, ограниченной соответственно старт и стоп кодонами, и содержащую по окончании селекции по меньшей мере одну нуклеиновую кислоту, отличающуюся от исходной последовательности.
Под «эволюционирующим белком» согласно изобретению понимают последовательность аминокислот (белковая последовательность), содержащую по окончании селекции по меньшей мере одну аминокислоту, отличающуюся от исходной белковой последовательности.
Гены и белки можно идентифицировать по их первичным последовательностям, а также по гомологиям последовательностей или выравниваний, которые определяют группы белков.
PFAM (Protein families database of alignements and Hidden Markov Models) содержит большую коллекцию выравнивания белковых последовательностей. Каждая PFAM позволяет визуализировать множественные выравнивания, видеть белковые домены, оценивать распределение между организмами, иметь доступ к другим базам данных, визуализировать известные структуры белков.
COG (Сluster of Orthologous Groups of proteins; http:/www.ncbi.nlm.nih.gov/COG/) получают, сравнивая белковые последовательности, полученные из 43 полностью секвенированных геномов, представляющих собой 30 основных филогенетических линий. Каждая COG определена по меньшей мере по трем линиям, что позволяет таким образом идентифицировать старые сохранившиеся домены.
Согласно изобретению термины “культивирование” и “ферментация” используют в одинаковом смысле для обозначения выращивания бактерий на соответствующей культуральной среде, содержащей простой источник углерода.
Под простым источником углерода понимают согласно настоящему изобретению источники углерода, используемые специалистом для нормального роста микроорганизма, в частности, бактерии, которыми могут являться арабиноза, фруктоза, галактоза, лактоза, мальтоза, сахароза и ксилоза. Наиболее предпочтительным простым источником углерода является глюкоза.
Специалист в данной области может определить условия культивирования (ферментации) микроорганизмов согласно изобретению. В частности, бактерии культивируют при температуре от 20 до 55°С, предпочтительно от 25 до 4°С, более предпочтительная температура для C. glutamicum и S. cerevisiae составляет примерно 30°С и для E. coli примерно 34°С.
Ферментацию обычно проводят в ферментерах, содержащих известную композицию среды для культивирования с минералами, определенную и адаптированную в зависимости от используемых бактерий, содержащую по меньшей мере один простой источник углерода и в случае необходимости кофактор, необходимый для продуцирования метаболита.
В частности, минеральной культуральной средой для E. coli может быть композиция, идентичная или подобная среде М9 (Anderson, 1946, Proc. Natl. Acad. Sci. USA 32:120-128), среда М63 (Miller, 1992, A Schort Course in Bacterial Genetics: A Laboratory Manuаl and Handbook for Escherichia coli and Related Bacteria, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York) или такая среда, определение которой дали Schaefer и др. (1999, Anal. Biochem.270: 88-96), более конкретно минимальная культуральная среда, описанная ниже:
∗раствор олигоэлемента: лимонная кислота - 4 г/л, MnSO4 - 3 г/л, NaCl - 1г/л, CoCl2 - 0,1 г/л, ZnSO4 - 0,10 г/л, CuSO4 - 10 мг/л, H3BO3 - 10 мг/л, NaMoO4 - 10 мг/л.
Подобным образом минеральной культуральной средой для C. glutamicum может быть композиция, идентичная или подобная среде BMCG (Leibl et al., 1989, Appl. Microbiol. Biotechnol. 32: 205-210), среде, описанной Reidel et al. (2001, J. Mol. Microbiol. Biotechnol, 3: 573-583).
Ферментацию предпочтительно проводят в анаэробиозе и в хемостате, т.е. с непрерывной подачей указанной минимальной культуральной среды с постоянной степенью разведения, содержащей постоянную концентрацию источника углерода и дегазированной азотом.
Концентрация источника углерода в среде ферментации повышается только в случае, если постоянный режим, ограниченный концентрацией остаточного углерода, достигается и остается стабильным в течение нескольких дней.
Метод культивирования в хемостате является предпочтительным методом культивирования, т.к. он способствует улучшению характеристик роста модифицированного штамма и продуцирования 1,2-пропандиола и позволяет выделять эволюционирующие микроорганизмы.
Под повышенной активностью «1,2-пропандиолсинтазы» понимают согласно изобретению совокупность повышенных ферментативных активностей, участвующих в путях превращения DHAР в 1,2-пропандиол. Повышенная ферментативная активность эволюционирующего микроорганизма заключается в увеличении количества 1,2-пропандиола, продуцируемого эволюционирующим микроорганизмом, по отношению к количествам, продуцируемым соответствующим исходным микроорганизмом в идентичных условиях культивирования.
Под повышенной активностью “ацетонсинтазы” согласно изобретению понимают совокупность повышенных ферментативных активностей, участвующих в пути превращения ацетата и ацетил-СоА в ацетон. Эволюционирующая ферментативная активность эволюционирующего микроорганизма второго поколения заключается в повышении количества ацетона, продуцируемого эволюционирующим микроорганизмом второго поколения по отношению к соответствующему трансформированному эволюционирующему микроорганизму в идентичных условиях культивирования.
Изобретение также касается выделения и характеристики эволюционирующих генов в эволюционирующих штаммах, полученных способом согласно изобретению, и эволюционирующих белков, кодируемых указанными эволюционирующими генами. Эти эволюционирующие гены затем могут быть введены в организм хозяина под контролем соответствующих элементов регуляции для его экспрессии в указанном организме с тем, чтобы обеспечить продуцирование соответствующего эволюционирующего белка.
Улучшение свойств модифицированных микроорганизмов, в частности, штамма E. coli MG16555 ΔtpiA, ΔpflAB, ΔadhE, ldhA::kana, ΔgloA, ΔaldA, ΔaldB в ходе культивирования в хемостате свидетельствует о том, что эти условия культивирования позволяют селекционировать эндогенный функциональный комплекс пируватдегидрогеназы в условиях анаэробиоза, условиях эффективного продуцирования NADH. Действительно известно, что комплекс пируватдегидрогеназы, катализирующий трансформацию пирувата в ацетил-СоА с высвобождением NADH, является функциональным только в аэробных условиях, тогда как в условиях анаэробиоза функциональной является пируват-формиатлиаза, которая катализирует превращение пирувата в ацетил-СоА и формиат (Snoep J.L., De Graef M.R., Westphal A.H., De Kok A.Teixeire de Mattos M.J. and Neijssel O.M. (1993). Таким образом одним из модифицирований, проведенных в отношении модифицированного штамма E. coli, созданного для продуцирования 1,2-пропандиола, в целях продуцирования NADH путем декарбоксилирования пирувата, является делеция генов pflA и pflB, кодирующих активность пируват-формиатлиазы. Единственной возможностью для модифицированной клетки является метаболизация пирувата в ацетил-СоА с помощью комплекса пируватдегидрогеназы, продуцируя NADH. Комплекс пируватдегидрогеназы эволюционирующего модифицированного штамма был охарактеризован и является менее чувствительным к NADH, чем комплекс пируватдегидрогеназы штамма дикого типа.
Настоящее изобретение приводит к селекции функционального комплекса пируватдегидрогеназы в условиях анаэробиоза, что позволяет продуцировать два NADH путем окисления глицеринового альдегида-3-фосфата в ацетат, NADH, которые могут повторно окисляться только путем восстановления дигидроксиацетонфосфата в 1,2-пропандиол. Селекция ферментативного комплекса, мало чувствительного к NADH, ускоряет продуцирование 1,2-пропандиола.
Настоящее изобретение преимущественно относится к селекции мутаций гена lpd (последовательность дикого типа которого известна hppt://genolist.pasteur.fr/Colibri), кодирующего липоамиддегидрогеназу комплекса пируватдегидрогеназы. В частности, было идентифицировано присутствие точечной мутации, вызывающей замещение аланина 55 валином. Этот фермент известен в качестве ответственного за ингибирование комплекса пируватдегидрогеназы с помощью NADH. Этот модифицированный фермент также относится к настоящему изобретению.
Настоящее изобретение позволяет улучшать свойства модифицированных микроорганизмов, в частности, штамма E. coli MG1655 ΔtpiA, ΔpflAB, ΔadhE, ldhA::kana, ΔgloA, ΔaldA, ΔaldB также за счет эволюционирования в процессе анаэробного культивирования в хемостате эндогенных ферментов, участвующих в пути превращения DHAP в 1,2-пропандиол. Результатом эволюционирования этих ферментов является повышение степени роста и конечного содержания 1,2-пропандиола.
Предпочтительно согласно изобретению эволюционирующий штамм не имеет эволюционирования в гене gldA. В соответствии с частным вариантом эволюционирующий штамм имеет делецию в гене gldA.
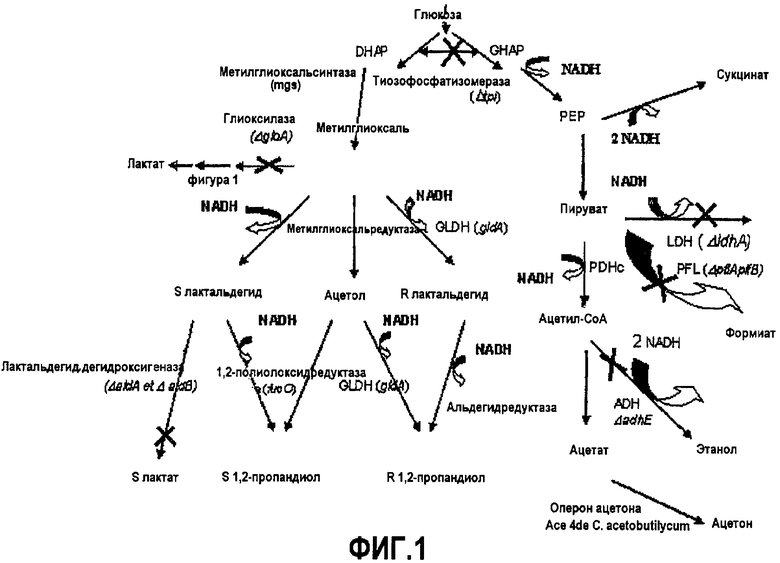
Описание фигур
Фиг.1 - схема метаболизма модифицированного штамма E. coli в целях продуцирования 1,2-пропандиола и ацетона согласно изобретению.
Список условных обозначений:
LDH: лактатдегидрогеназа
ADH: альдегид-алкоголь дегидрогеназа
PFL: пируват-форматлиаза
PDHc: комплекс пируватдегидрогеназа
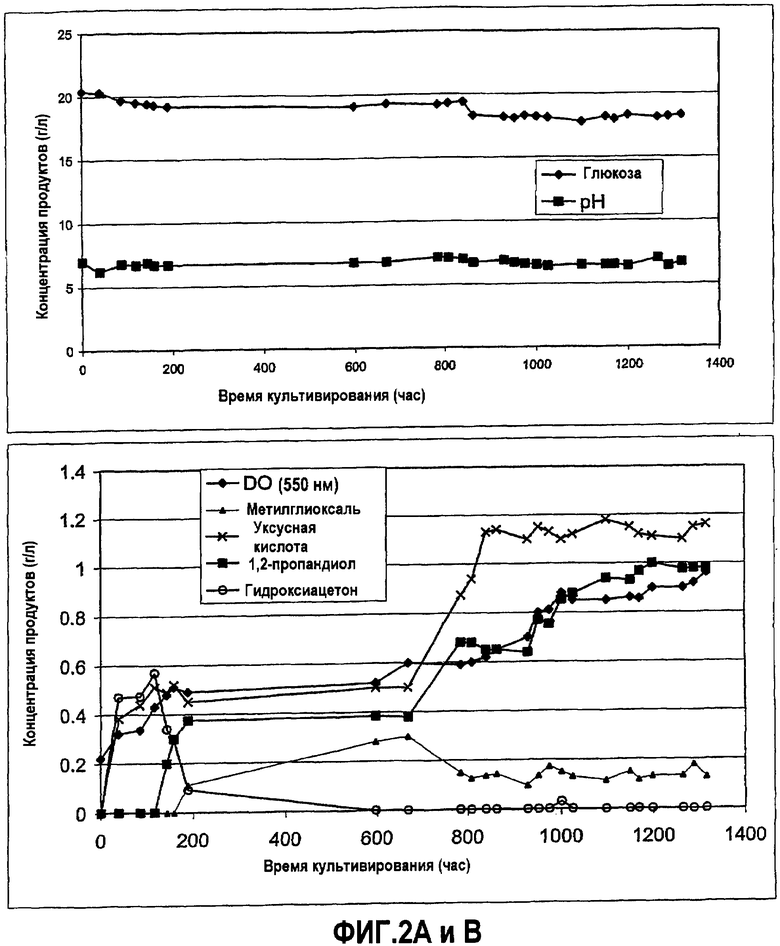
Фиг.2 - эволюционирование штамма E. coli MG1655 ΔtpiA, ΔpflAB, ΔadhE, ldhA::kana, ΔgloA, ΔaldA, ΔaldB при культивировании в хемостате на глюкозе: концентрация глюкозы (фиг.2А) и другие продукты (фиг.2В).
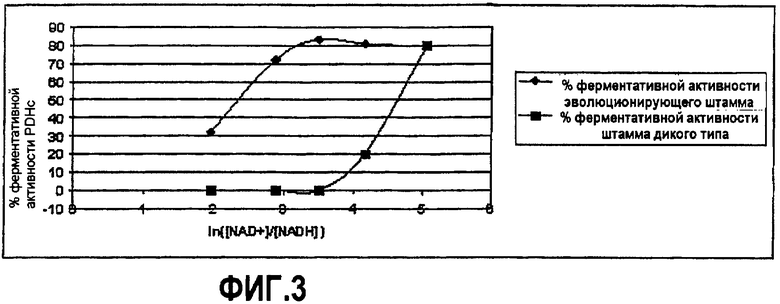
Фиг.3 - сравнение ферментативной активности комплекса пируватдегидрогеназы штамма дикого типа и эволюционирующего штамма согласно изобретению в зависимости от повышения концентраций NADH.
Примеры осуществления, указанные выше, иллюстрируют изобретение, не ограничивая его объем.
Пример 1. Конструирование модифицированного штамма E. coli MG1655 ΔtpiA, ΔpflAB, ΔadhE, ldhA::kana, ΔgloA, ΔaldA, ΔaldB, способного продуцировать только 1,2-пропандиол и ацетат путем ферментации глюкозы
а) Конструирование модифицированного штамма E. coli MG1655 ΔtpiA::cm
Инактивацию гена tpiA проводят путем введения кластера, резистентного к антибиотику хлорамфеноколу, проводя делецию большей части используемого гена. Используемая методика описана Datsenko K.A., Wanner B.L. (2000) One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 97: 6640-6645.
Для замещения гена tpiA используют два олигонуклеотида:
DtpiAr, содержащего 100 оснований (SEQ ID NO:1):
аtgcgacatcctttagtgatgggtaactggaaactgaacggcagccgccacatggttcacgagctggtttctaacctgcgtaCATATGAATATCCTCCTTAG
с:
участком (прописные буквы), гомологичным последовательности (4109007-4109087) гена tpiA (последовательность 4108320-4109087), последовательность для сравнения на сайте hppt://genolist.pasteur.fr/Colibri)
участком (заглавные буквы) для амплифицирования кластера, резистентного к хлорамфениколу, плазмиды pKD3 (Datsenko K.A., Wanner B.L. (2000) One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 97: 6640-6645).
DtpiAf, содержащего 100 оснований (SEQ ID NO:2):
сttaagcctgtttagccgcttctgcagctttaacgattactgcgaaggcgtcagctttcagagaagcaccaccaaccagcTGTAGGCTGGAGCTGCTTCG
с:
участком (прописные буквы), гомологичным последовательности (4108320-4108400) гена tpiA
участком (заглавные буквы) для амплифицирования кластера, резистентного к хлорамфениколу, находящегося в плазмиде pKD3.
Олигонуклеотиды DtpiA и DtpiAf использовали для амплифицирования кластера, резистентного к хлорамфениколу, из плазмиды pKD3. Полученный ПЦР-продукт затем вводили путем электропорации в штамм MG1655 (pKD46), в котором используемая система λ Red (γ,β, exо) в значительной степени способствовала гомологичной рекомбинации. Трансформанты, резистентные к антибиотику, затем селекционировали и инсерцию кластера резистентности проверяли с помощью анализа ПЦР с олигонуклеотидами cdh и YIIQ.
сdh (SEQ ID NO:3):
ggtgatgatagttatcgccg (гомологичная последовательности 4107536-4107555)
YIIQ (SEQ ID NO:4):
сgtgccatcgacagcagtcc (гомологичная последовательности 4109599-4109580)
Затем кластер, резистентный к антибиотику, элиминировали. Плазмиду рСР20, несущую рекомбиназу FLP, действующую на сайтах FRT кластера, резистентного к хлорамфениколу, затем вводили в рекомбинантные штаммы путем электропорации (Cheperanov P.P. and Wackernagel W. (1995) Gene disruption in Escherichia coli: TcR and KmR cassette wich option of Flp-catalysed excision of antibiotic-resistance detеrminant gene, 158, 9-14). После проведения серии культивирования при 42°С потерю кластера, резистентного к антибиотику, определяли анализом ПЦР с теми же олигонуклеидами, которые использовали ранее.
b) конструирование модифицированного штамма E. coli MG1655ΔpflAB::cm
Гены pflA и pflВ инактивировали путем инсерции кластера, резистентного к антибиотику хлорамфениколу, и делеции большей части используемых генов. Применяемая методика описана Datsenko K.A. and Wanner B.L. (2000).
Два олигонуклеотида использовали для осуществления замены генов pflA и pflВ:
DpflBr, содержащего 100 оснований (SEQ ID NO:5):
ccggacatcctgcgttgccgtaaatctggtgttctgaccggtctgccaga tgcatatggcСATATGAATATCCTCCTTAG
c:
участком (прописные буквы), гомологичным последовательности (952235-952315)) гена pflB (последовательность 950495-952777), последовательность для сравнения на сайте hppt://genolist.pasteur.fr/Colibri)
участком (заглавные буквы) для амплифицирования кластера, резистентного к хлорамфениколу, плазмиды pKD3 (Datsenko K.A., Wanner B.L. (2000) One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 97: 6640-6645).
DtpiAf, содержащего 100 оснований (SEQ ID NO:6):
gatgcactataagatgtgttaaaaacgctgtagcagaatgaagcgcggaataaaaaagcggcaactcaataaagttgccgCTGGAGCTGCTTCG
c:
участком (прописные буквы), гомологичным последовательности (949470-949550), находящимся перед геном pflА (последовательность 949563-950303),
участком (заглавные буквы) для амплифицирования кластера, резистентного к хлорамфениколу, находящегося в плазмиде pKD3.
Олигонуклеотиды pflАВ1 и pflАВ2 использовали для амплифицирования кластера, резистентного к хлорамфениколу, из плазмиды pKD3. Полученный ПЦР-продукт затем вводили путем электропорации в штамм MG1655 (pKD46), в котором используемый фермент Red рекомбиназа обеспечивала гомологичную рекомбинацию. Трансформанты, резистентные к антибиотику, затем селекционировали и инсерцию кластера резистентности проверяли с помощью анализа ПЦР с олигонуклеотидами pflАВ1 и pflАВ2.
pflАВ1 (SEQ ID NO:7)
agacattaaaaatatacgtgcagctacccg (гомологичная последовательности 948462-948491)
pflАВ2 (SEQ ID NO:8)
gtgaaagctgacaacccttttgatctttta (гомологичная последовательности 953660-983689)
с) конструирование модифицированного штамма E. coli MG1655 ΔtpiA, ΔpflAB:
Делеция генов pflA и pflВ путем замены генов кластером, резистентным к хлорамфениколу, в штамме MG1655 ΔtpiA была проведена методом трансдукции с помощью фага Р1. Протокол содержит две стадии: на первой получали лизат фага на штамме MG1655 ΔpflAB::cm, а на второй проводили трансдукцию штамма MG1655 ΔtpiA указанным лизатом фага.
Получение лизата фага:
- 100 мкм ночной культуры штамма MG1655 (ΔpflAB::cm) высевали в 10 мл LB + Cm 30 кмг/мл + глюкоза 0,2% + CaCl2 5 мМ;
- инкубировали в течение 30 минут при 37°С в условиях перемешивания;
- вводили 100 мкл лизата фага Р1, полученного на диком штамме MG1655 (примерно 1,109 фага/мл);
- перемешивали при 37°С в течение 3 часов до полного разложения клеток;
- добавляли 200 мкл хлороформа и перемешивали;
- центрифугировали 10 минут, 4500×g, для удаления клеточных остатков;
- перемещали отстоявшуюся жидкость в стерильную пробирку и добавляли 200 мкл хлороформа;
- лизат сохраняли при 4°С.
Трансдукция
- Центрифугировали в течение 10 минут, 1500×g, 5 мл ночной культуры штамма MG1655 (ΔtpiA) на среде LB;
- Cуспендировали клеточный осадок в 2,5 мл MgSO4 10 мM, CaCl2 5 мM;
- Контрольные пробирки: 100 мкл клеток;
100 мкл фагов Р1 штамма MG1655 (ΔpflAB::cm)
- Опытная пробирка: 100 мкл клеток + 100 мкл фагов Р1 штамма MG1655 (ΔpflAB::cm);
- Инкубировали в течение 30 мин при 30°С в условиях перемешивания;
- Вводили в каждую пробирку 100 мкл цитрата натрия 1М, затем перемешивали;
- Вводили 1 мл LB;
- Инкубировали в течение 1 часа при 30°С в условиях перемешивания;
- Помещали в кассеты LB + культуральная среда 30 мкг/мл после центрифугирования пробирок в течение 3 минут при 7000 об/мин;
- Инкубировали при 30°С до следующего дня.
Проверка штамма
Трансформанты, резистентные к антибиотику, селекционировали и инсерцию участка, содержащего (pflAB::cm), проверяли посредством анализа ПЦР с олигонуклеотидами pflAB1 и pflAB2, с одной стороны, и cdh и YIIQ, с другой стороны, с тем, чтобы проверить также делецию гена ΔtpiA в штамме ΔpflAB::cm. Полученный штамм назвали MG1655 Δ(pflAB::cm, ΔtpiA).
Как и ранее, кластер, резистентный к хлорамфениколу, затем элиминировали. Плазмиду рСР20, несущую рекомбиназу FLP, действующую на сайтах FRT кластера, резистентного к хлорамфениколу, затем вводили в рекомбинантные штаммы путем электропорации. После проведения серии культивирований при 42°С потерю кластера, резистентного к антибиотику, определяли анализом ПЦР с теми же олигонуклеотидами, которые использовали на предыдущей стадии. Полученный штамм назвали MG1655 ΔtpiA, ΔpflAB.
d) конструирование модифицированного штамма E. coli MG1655 ΔadhE::cm
Как и ранее ген adhE инактивировали путем инсерции кластера, резистентного к антибиотику хлорамфениколу, и делеции большей части используемого гена по методу, описанному Datsenko K.A. и Wanner B.L.(2000).
Два олигонуклеотида использовали для осуществления делеции:
DadhЕ r, содержащий 100 оснований (SEQ ID NO:9)
atggctgttactaatgtcgctgaacttaacgcactcgtagagcgtgtaaaaaaagcccagcgtgaatatgccagtttcactСATATGAATATCCTCCTTAG
c:
участком (прописные буквы), гомологичным последовательности (1297263-1297343) гена adhE (последовательность 1294669-1297344), последовательность для сравнения на сайте hppt://genolist.pasteur.fr/Colibri)
участком (заглавные буквы) для амплифицирования кластера, резистентного к хлорамфениколу, плазмиды pKD3 (Datsenko K.A., Wanner B.L. (2000))
DadhEf, содержащего 100 оснований (SEQ ID NO:10):
caataacgaatgatagcaattttaagtagttaggaggtgaaaaatgctgtcaaaaggcgtattgtcagcgcgtcttttcaTGTAGGCTGGAGCTGCTTCG
c:
участком (прописные буквы), гомологичным последовательности (1294694-1294774) гена adhE
участком (заглавные буквы) для амплифицирования кластера, резистентного к хлорамфениколу, находящегося в плазмиде pKD3.
Олигонуклеотиды DadhEr и DadhEf использовали для амплифицирования кластера, резистентного к хлорамфениколу, находящегося в плазмиде pKD3. Полученный ПЦР-продукт затем вводили путем электропорации в штамм MG1655 (pKD46), в котором используемый фермент Red рекомбиназа обеспечивал гомологичную рекомбинацию. Трансформанты, резистентные к антибиотику, затем селекционировали и инсерцию кластера резистентности проверяли с помощью анализа ПЦР с олигонуклеотидами ychGf и adhECr.
YchGf (SEQ ID NO:11)
ggctcattgcaccaccatccag (гомологичная последовательности 1294357-1294378)
adhECr (SEQ ID NO:12)
gaaaagacgcgctgacaatacgcc (гомологичная последовательности 1297772-1297749)
е) конструирование штамма MG1655 ΔtpiA, ΔpflAB, ΔadhE:
Делецию гена ΔadhE проводили в штамме MG1655 ΔtpiA, ΔpflAB, как указано выше, методом трансдукции с помощью фага Р1 (см. протокол стадии с). Лизат фага Р1 получали на штамме MG1655 ΔadhE::cm и трансдукцию штамма MG1655 ΔtpiA, ΔpflAB проводили с помощью указанного лизата. Трансдукты, резистентные к хлорамфениколу, контролировали с помощью олигонуклеотидов ychGf и adhECr для проверки мутации гена adhE, а также с помощью олигонуклеотидов pflAB1 и pflAB2 с одной стороны, и cdh и YIIQ с другой стороны, чтобы проверить также делецию генов pflA и B и tpiA в штамме ΔadhE::cm.
Как и ранее, кластер, резистентный к хлорамфениколу, затем элиминировали. Плазмиду рСР20, содержащую рекомбиназу FLP, действующую на сайтах FRT кластера, резистентного к хлорамфениколу, затем вводили в рекомбинантные штаммы путем электропорации. После проведения серии культивирований при 42°С потерю кластера, резистентного к антибиотику, определяли анализом ПЦР с теми же олигонуклеотидами, которые использовали на предыдущей стадии. Полученный штамм назвали MG1655 ΔtpiA, ΔpflAB, ΔadhE.
f) конструирование модифицированного штамма E. coli MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana
Инактивацию гена IdhA (координаты с 1439878 по 1440867) в штамме MG1655 ΔtpiA, ΔpflAB, ΔadhE осуществляли, как указано выше, методом с применением фага Р1 (см. протокол стадии с). Лизат фага получали с помощью штамма E. coli K12 NZN11 Δpfl::cam, IdhA::kana, описанного Clark D.P.(Bunch P.K., Mat-Jan F. and Clark D.P. (1997) The IdhA gene encoding the fermentative lactate dehydrogenase of Escherichia coli: microbiology, 143, 187-195). Трансдукты штамма MG1655 ΔtpiA, ΔpflAB, ΔadhE проводили с помощью лизата фага штамма E. coli K12 NZN11 Δpfl::cam, IdhA::kana. Трансдукты селекционировали на канамицине и инсерцию канамицинового кластера в гене IdhA проверяли с помощью олигонуклеотидов hsIJC и IdhAC2.
hsIJC (SEQ ID NO:13)
gccatcagcaggcttagccg (гомологичная последовательности 1439345-1439767)
IdhAC2 (SEQ ID NO:14)
gggtattgtggcatgtttaaccg (гомологичная последовательности
1441007-1441029)
Полученный штамм назвали MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana.
g) конструирование модифицированного штамма E. coli MG1655 gloA::cm
Инактивацию гена gloA осуществляли, как указано выше, путем инсерции кластера, резистентного к антибиотику хлорамфениколу, и делеции большей части используемых генов по методу, описанному Datsenko K.A. and Wanner B.L. (2000).
Для осуществления делеции использовали два олигонуклеотида:
GLOAD f, содержащий 100 оснований (SEQ ID NO:15)
atgcgtcttcttcataccatgctgcgcgttggcgatttgcaacgctccatcgatttttataccaaagtgctgggcatgaaGTGTAGGCTGGAGCTGCTTCG
с:
участком (прописные буквы), гомологичным последовательности (1725861-1725941) гена gloA (последовательность 1725861-176268), последовательность для сравнения на сайте hppt://genolist.pasteur.fr/Colibri)
участком (заглавные буквы) для амплифицирования кластера, резистентного к хлорамфениколу, плазмиды pKD3 (Datsenko K.A., Wanner B.L. (2000))
GLOAD R (SEQ ID NO:16)
ttagttgcccagaccgcgaccggcgtctttctcttcgattaactcaattttgtaaccgtccggatcttccacaaacgcgaCATATGAATATCCTCCTTAG
c:
участком (прописные буквы), гомологичным последовательности (1726188-1726268) гена gloA (последовательность 1725861-176268),
участком (заглавные буквы) для амплифицирования кластера, резистентного к хлорамфениколу, находящегося в плазмиде pKD3.
Олигонуклеотиды GLOADr и GLOADf использовали для амплифицирования кластера, резистентного к хлорамфениколу, из плазмиды pKD3. Полученный ПЦР-продукт затем вводили путем электропорации в штамм MG1655 (pKD46), в котором используемый фермент Red рекомбиназа обеспечивал гомологичную рекомбинацию. Трансформанты, резистентные к антибиотику, затем селекционировали и инсерцию кластера резистентности проверяли посредством анализа ПЦР с помощью олигонуклеотидов NemAQ d и Rnt Cr.
NemAQd (SEQ ID NO:17)
gaagtggtcgatgccgggattgaagaatggg (гомологичная последовательности 1725331-1725361)
Rnt Cr (SEQ ID NO:18)
gggttacgtttcagtgaggcgcgttctgcgg (гомологичная последовательности 1726765-1726795)
h) конструирование модифицированного штамма MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA:
Инактивацию гена gloA проводили в штамме MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, как указано выше, методом с использованием фага Р1 (см. протокол стадии с). Лизат фага Р1 получали на штамме MG1655 ΔgloA::cm и трансдукцию штамма MG1655 ΔtpiA, ΔpflAB, IdhA::kana проводили с помощью указанного лизата. Трансдукты, резистентные к хлорамфениколу, контролировали с помощью олигонуклеотидов Nem AQd и Rnt Cr для проверки мутации гена gloA, а также с помощью олигонуклеотидов pflAB1 и pflAB2, cdh и YIIQ, ychCf и adhECr, hsIJC и IdhAC2, чтобы проверить также соответственно делецию генов pflA и В, tpiA, adhE и IdhA в штамме ΔgloA::cm.
Как и ранее, кластер, резистентный к хлорамфениколу, затем элиминировали. Плазмиду рСР20, несущую рекомбиназу FLP, действующую на сайтах FRT кластера, резистентного к хлорамфениколу, затем вводили в рекомбинантные штаммы путем электропорации. После проведения серии культивирований при 42°С потерю кластера, резистентного к антибиотику, определяли анализом ПЦР с помощью тех же олигонуклеотидов, которые использовали ранее. Полученный штамм назвали MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA.
i) конструирование модифицированного штамма E. coli MG1655 IdhA::kana, ΔaldA::cm
Инактивацию гена aldA осуществляли, как указано выше, путем инсерции кластера, резистентного к антибиотику хлорамфениколу, и делеции большей части используемых генов по методу, описанному Datsenko K.A. and Wanner B.L.(2000).
Для осуществления делеции использовали два олигонуклеотида:
AldA D f, содержащий 100 оснований (SEQ ID NO:19)
atgtcagtacccgttcaacatcctatgtatatcgatggacagtttgttacctggcgtggagacgcatggattgatgtggtaGTGTAGGCTGGAGCTGCTTCG
с:
участком (прописные буквы), гомологичным последовательности (1486256-1486336) гена aldA (последовательность 1486256-1487695), последовательность для сравнения на сайте hppt://genolist.pasteur.fr/Colibri)
участком (заглавные буквы) для амплифицирования кластера, резистентного к хлорамфениколу, плазмиды pKD3 (Datsenko K.A., Wanner B.L. (2000))
aldA Dr, содержащий 100 оснований (SEQ ID NO:20)
ttaagactgtaaataaaccacctgggtctgcagatattcatgcaagccatgtttaccatctgcgccgccaataccggatttCATATGAATATCCTCCTTAG
c:
участком (прописные буквы), гомологичным последовательности (1487615-1487695) гена aldA (1486256-1487695),
участком (заглавные буквы) для амплифицирования кластера, резистентного к хлорамфениколу, находящегося в плазмиде pKD3.
Олигонуклеотиды AldA D r и AldA D f использовали для амплифицирования кластера, резистентного к хлорамфениколу, в плазмиде pKD3. Полученный ПЦР-продукт затем вводили путем электропорации в штамм MG1655 (pKD46), в котором используемый фермент Red рекомбиназа обеспечивал гомологичную рекомбинацию. Трансформанты, резистентные к антибиотику, затем селекционировали и инсерцию кластера резистентности проверяли посредством анализа ПЦР с помощью олигонуклеотидов Ydc F Cf и gapCCr.
Ydc F Cf (SEQ ID NO:21)
tgcagcggcgcacgatggcgacgttccgccg (гомологичная 1485722-1485752)
gapCCr (SEQ ID NO:22)
cacgatgacgaccattcatgcctatactggc (гомологичная последовательности 1488195-1488225)
h) конструирование модифицированного штамма MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA
Инактивацию гена aldA проводили в штамме MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, как указано выше, методом с использованием фага Р1 (см. протокол стадии с). Лизат фага Р1 получали на штамме MG1655 ΔgloA::cm и трансдукцию штамма MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA проводили с помощью указанного лизата. Трансдукты, резистентные к хлорамфениколу, контролировали с помощью олигонуклеотидов Ydc F Cf и gapCCr для проверки мутации гена aldA, а также с помощью олигонуклеотидов NemAQd и Rnt Cr, pflAB1 и pflAB2, cdh и YIIQ, ychCf и adhECr, hsIJC и IdhAC2, чтобы проверить также соответственно делецию генов gloA, pflA и В, tpiA, adhE в штамме ΔaldA::cm.
Как и ранее, кластер, резистентный к хлорамфениколу, затем элиминировали. Плазмиду рСР20, несущую рекомбиназу FLP, действующую на сайтах FRT кластера, резистентного к хлорамфениколу, затем вводили в рекомбинантные штаммы путем электропорации. После проведения серии культивирований при 42°С потерю кластера, резистентного к антибиотику, определяли анализом ПЦР с помощью тех же олигонуклеотидов, которые использовали ранее. Полученный штамм назвали MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA.
g) конструирование модифицированного штамма E. coli MG1655 ΔaldB::cm
Инактивацию гена aldВ осуществляли, как указано выше, путем инсерции кластера, резистентного к антибиотику хлорамфениколу, и делеции большей части используемых генов по методу, описанному Datsenko K.A. and Wanner B.L.(2000).
Для осуществления делеции использовали два олигонуклеотида:
AldB D f, содержащий 100 оснований (SEQ ID NO:23)
tcagaacagccccaacggtttatccgagtagctcaccagcaggcacttggtttgctggtaatgctccagcatcatcttgtGTGTAGGCTGGAGCTGCTTCG
с:
участком (прописные буквы), гомологичным последовательности (3752603-3752683) гена аldB (последовательность 3752603-3754141), последовательность для сравнения на сайте hppt://genolist.pasteur.fr/Colibri)
участком (заглавные буквы) для амплифицирования кластера, резистентного к хлорамфениколу, плазмиды pKD3 (Datsenko K.A., Wanner B.L. (2000))
аldB Dr, содержащий 100 оснований (SEQ ID NO:24)
atgaccaataatcccccttcagcacagattaagcccggcgagtatggtttccccctcaagttaaaagcccgctatgacaaCATATGAATATCCTCCTTAGG
c:
участком (прописные буквы), гомологичным последовательности (3754061-3754141) гена aldВ (3752603-3754141),
участком (заглавные буквы) для амплифицирования кластера, резистентного к хлорамфениколу, находящегося в плазмиде pKD3
Олигонуклеотиды AldВ D r и AldВ D f использовали для амплифицирования кластера, резистентного к хлорамфениколу, в плазмиде pKD3. Полученный ПЦР-продукт затем вводили путем электропорации в штамм MG1655 (pKD46), в котором используемый фермент Red рекомбиназа обеспечивал гомологичную рекомбинацию. Трансформанты, резистентные к антибиотику, затем селекционировали и инсерцию кластера резистентности проверяли посредством анализа ПЦР с помощью олигонуклеотидов aldB C f и YiaYCr.
aldB C f (SEQ ID NO:25)
catatttccctcaaagaatataaaaaagaacaattaacgc (гомологичная последовательности 3752057-3752095)
YiaYCr (SEQ ID NO:26)
tatgttcatgcgatggcgcaccagctgggcg (гомологичная последовательности 3754644-3754674)
h) конструирование модифицированного штамма MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ
Инактивацию гена aldВ проводили в штамме MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, aldA, как указано выше, методом с использованием фага Р1 (см. протокол стадии с). Лизат фага Р1 получали на штамме MG1655 ΔaldВ::cm и трансдукцию штамма MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA проводили с помощью указанного лизата. Трансдукты, резистентные к хлорамфениколу, контролировали с помощью олигонуклеотидов aldB C f и YiaYCr для проверки мутации гена aldВ, а также с помощью олигонуклеотидов NemAQd и Rnt Cr, pflAB1 и pflAB2, cdh и YIIQ, ychCf и adhECr, hsIJC и IdhAC2, YdcF C f и gapCCr, чтобы проверить также соответственно делецию генов gloA, pflA и В, tpiA, adhE, aldA в штамме ΔaldB::cm.
Как и ранее, кластер, резистентный к хлорамфениколу, затем элиминировали. Плазмиду рСР20, несущую рекомбиназу FLP, действующую на сайтах FRT кластера, резистентного к хлорамфениколу, затем вводили в рекомбинантные штаммы путем электропорации. После проведения серии культивирований при 42°С потерю кластера, резистентного к антибиотику, определяли анализом ПЦР с помощью тех же олигонуклеотидов, которые использовали на предыдущей стадии. Полученный штамм назвали MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ.
Пример 2. Культивирование и эволюционирование модифицированного штамма MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ в хемостате
Для оптимизации продуцирования 1,2-пропандиола из глюкозы штаммом MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ проводят культивирование штамма в хемостате в минимальной культуральной среде с добавлением нитрата натрия и экстракта дрожжей в течение нескольких недель в условиях анаэробиоза.
На начальной стадии культивирования первичное содержание глюкозы в емкости подачи культуры составляет 20 г/л, степень разведения составляет 0,04 час-1 и непрерывный приток азота поддерживают для обеспечения условий анаэробиоза. Клеточная концентрация и продуцирование 1,2-пропандиола и ацетата являются слабыми. Через несколько недель с начала культивирования рост и концентрации продуктов повышаются, достигается постоянный режим, характеризующийся содержанием остаточной глюкозы и постоянными концентрациями продуктов (фиг.2).
Пример 3. Характеристика эволюционирующего комплекса пируватдегидрогеназы, малочувствительного к NADH
Эволюционирование комплекса пируватдегидрогеназы (PDHc) в PDHc, мало чувствительный к NADH, определяют одновременно путем количественного анализа активности фермента, эволюционирующего in vitro, и путем сравнения последовательности одного из генов (lpd), кодирующих липоамиддегидрогеназу эволюционирующего PDHc, с последовательностью гена исходного PDHc.
а) количественный анализ ферментативной активности комплекса пируватдегидрогеназы
Количественный анализ ферментативной активности PDHc in vitro штамма MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ проводят в соответствии с протоколом, описанным Schwartz и Reed (Schwartz E.R. and Reed L.J. (1970) Regulation of the activity of piruvete dehydrogenase complex of Escherchia coli, Biochemistry, 6, 1434-1439).
100 мл клеточной культуры отбирают из хемостата в предварительно дегазированные флаконы, помещенные в анаэробный вытяжной шкаф. Клетки подвергают центрифугированию в течение 10 минут со скоростью 6000 об/мин. Осадок суспендируют в примерно 100 мл буферного раствора, содержащего фосфат калия в концентрации 50 мМ, рН 7,9, тиаминпирофосфат 0,5 мМ, затем повторно центрифугируют в течение 10 минут со скоростью 6000 об/мин. Клеточный осадок суспендируют в 800 мкл буферного раствора. Клеточную суспензию дезинтегрируют с помощью ультразвукового устройства, проводя 4 цикла обработки (30 секунд при 30%) с двухминутными перерывами выдерживания во льду. Клеточные остатки удаляют цетрифугированием в течение 5 минут, 13400 об/мин, отстоявшийся раствор представляет собой необработанный бесклеточный экстракт. Соли, присутствующие в бесклеточном экстракте, которые могут быть помехой при определении ферментативной активности, удаляют путем пропускания экстракта через колонку PD10, уравновешенную калиево-фосфатным буфером с рН 7,9, 0,5 мМ тиаминпирофосфата. Экстракт элюируют 3,5 мл того же буфера. Полученный элюат экстрагируют необработанным бесклеточным экстрактом.
Ферментативную активность бесклеточного экстракта измеряют сначала без NADH, затем в присутствии повышающихся от 0,14 мМ до 2,7 мМ концентраций NADH. Полученные результаты сравнивают с указанными в литературе в отношении дикого штамма E. coli, фиг.3 (Snoep J.L., De Graef M.R., Westphal A.H., De Kok A. Teixeira de Mattos M.J. and Neijssel O.M. (1993) Differences in sensitivity to NADH of purified pyruvate dehydrogenase complexes of Enterococcus faecalis, Lactococcus lactis, Azotobacter vinelandii and Escherichia coli: implications for their activity in vivo, FEMS microbiology letters, 114, 279-284).
Полученные результаты показывают, что PDHc эволюционирующего модифицированного штамма MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ менее чувствителен к NADH, чем дикий штамм E. coli. При отношении [NAD+]/[NADH]≅33 наблюдается тотальное ингибирование активности PDHc дикого штамма, тогда как измерения показывают 80% активности эволюционирующего PDHc.
b) Определение последовательности гена lpd, кодирующего липоамиддегидрогеназу комплекса пируватдегидрогеназы эволюционирующего штамма MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ
Хромосомную ДНК штамма MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ экстрагируют из 1 мл ночной культуры в LB. После центрифугирования клетки промывают стерильной водой, затем расщепляют тепловым ударом в течение 5 минут при 94°С. Хромосомную ДНК выделяют из отстоявшегося раствора после центрифугирования. Ген lpd (последовательность 127912-129336), кодирующий липоамиддегидрогеназу (Е3) комплекса пируватдегидрогеназы амплифицируют посредством ПЦР с помощью двух следующих олигонуклеотидов:
AceEf (SEQ ID NO:27)
cgcgtgatcgacggtgctgatggtgcccg (гомологичная последовательности 127504-127532)
YacH r (SEQ ID NO:28)
aagttcaggagagccgccc (гомологичная последовательности 127513-129531).
Получали и сeквенировали ПЦР-продукт в 2000 пар оснований, соответствующий гену lpd. Полученные результаты свидетельствуют о присутствии точечной мутации, вызывающей замену аланина 55 валином.
Пример 4. Путь превращения метилглиоксаля в 1,2-пропандиол эволюционирующего модифицированного штамма MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ не содержит глицеролдегидрогеназы
С тем, чтобы показать, что улучшение свойств эволюционирующего модифицированного штамма MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ не связано с эволюционированием глицеролдегидрогеназы, кодируемой геном glaA, провели конструирование эволюционирующего штамма MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ, имеющего делецию гена glaA.
а) конструирование модифицированного штамма MG1655Δgld::cm
Ген gldА инактивировали, как указано в примере 1, путем инсерции кластера, резистентного к антибиотику хлорамфениколу, и делеции большей части используемых генов по методу, описанному Datsenko K.A., Wanner B.L.(2000).
Для осуществления делеции использовали два олигонуклеотида:
GldА D f, содержащий 100 оснований (SEQ ID NO:29)
gttattcccactcttgcaggaaacgctgaccgtactggtcggctaccagcagagcggcgtaaacctgatctggcgtcgcgGTGTAGGCTGGAGCTGCTTCG
с:
участком (прописные буквы), гомологичным последовательности (4135512-4135592) гена gldА (последовательность 4135512-4136615), последовательность для сравнения на сайте hppt://genolist.pasteur.fr/Colibri)
участком (заглавные буквы) для амплифицирования кластера, резистентного к хлорамфениколу, плазмиды pKD3 (Datsenko K.A., Wanner B.L. (2000)
gldB Dr, содержащий 100 оснований (SEQ ID NO:30)
atggaccgcattattcaatcaccgggtaaatacatccagggcgctgatgtgattatcgtctgggcgaatacctgaagccCATATGAATATCCTCCTTAG
c:
участком (прописные буквы), гомологичным последовательности (4136535-4136615) гена gldA (4135512-4136615),
участком (заглавные буквы) для амплифицирования кластера, резистентного к хлорамфениколу, находящегося в плазмиде pKD3.
Олигонуклеотиды gldА D r и gldА D f использовали для амплифицирования кластера, резистентного к хлорамфениколу, в плазмиде pKD3. Полученный ПЦР-продукт затем вводили путем электропорации в штамм MG1655 (pKD46), в котором используемый фермент Red рекомбиназа обеспечивал гомологичную рекомбинацию. Трансформанты, резистентные к антибиотику, затем селекционировали и инсерцию кластера резистентности проверяли посредством анализа ПЦР с помощью олигонуклеотидов YijF D и TalCr.
YijF D (SEQ ID NO:31)
gcctggatttgtaccacggttggtggaacggcggg (гомолог последовательности 4135140-4135174)
TalCr (SEQ ID NO:32)
cacgcatattccccattgccgggg (гомолог последовательности 4137216-4137239).
ПЦР-продукты в 2100 п.н. получали на основании гена дикого типа, как в отношении гена, делетированного и замененного геном, резистентным к хлорамфениколу. Также полученные ПЦР-продукты затем обрабатывали ферментом рестрикции Sall. Два фрагмента примерно в 1000 п.н. получали из ПЦР-продукта дикого типа, тогда как ПЦР-продукт, содержащий ген, резистентный к хлорамфениколу, не обрабатывался.
b) конструирование эволюционирующего модифицированного штамма MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ, ΔgldA::cm
Ген gldA инактивировали в эволюционирующем штамме MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ, как указано в примере 1, методом с использованием фага Р1 (см. протокол стадии с)). Лизат фага Р1 получали на штамме MG1655 ΔgloA::cm и трансдукцию штамма MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA проводили с помощью указанного лизата. Трансдукты, резистентные к хлорамфениколу, контролировали с помощью олигонуклеотидов YijF D и TalCr для проверки мутации гена gldA.
с) культивирование модифицированных эволюционирующих штаммов E. coli *MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ, ΔgldA::cm и E. coli *MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ
Оба модифицированных эволюционирующих штамма E. coli *MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ, ΔgldA::cm и E. coli *MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ культивировали при нерегулируемом рН в минимальной культуральной среде с добавлением нитрата натрия и экстракта дрожжей с начальной концентрацией глюкозы 20 г/л в условиях анаэробиоза в течение 10 дней. Полученный профиль продуктов ферментации показывает, что делеция гена gldA не вызывает уменьшения продуцирования 1,2-пропандиола (таблица 1).
Сравнение концентраций субстрата и продуктов ферментации через 10 дней с начала культивирования модифицированных эволюционирующих штаммов E. coli *MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ, ΔgldA::cm и E. coli *MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ
(г/л)
(г/л)
Пример 5. Повышение выхода превращения глюкозы в 1,2-пропандиол путем делеции гена edd, кодирующего 6-фосфоглюконатдегидратазу в эволюционирующем штамме E. coli *MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ
а) конструирование модифицированного штамма MG1655Δedd::cm
Ген edd инактивировали, как указано в примере 1, путем инсерции кластера, резистентного к антибиотику хлорамфениколу, и делеции большей части используемых генов по методу, описанному Datsenko K.A. and Wanner B.L. (2000).
Для осуществления делеции использовали два олигонуклеотида:
edd D f, содержащий 100 оснований (SEQ ID NO:33)
ttaaaaagtgatacaggttgcgccctgttcggcaccggacagtttttcacgcaaggcgctgaataattcacgtcctgttcGTGTAGGCTGGAGCTGCTTCG
с:
участком (прописные буквы), гомологичным последовательности (1930817-4) гена edd (последовательность 1930817-1932628), последовательность для сравнения на сайте hppt://genolist.pasteur.fr/Colibri)
участком (заглавные буквы) для амплифицирования кластера, резистентного к хлорамфениколу, плазмиды pKD3 (Datsеnko K.A., Wanner B.L. (2000))
edd Dr, содержащий 100 оснований (SEQ ID NO:34)
atgaatccacaattgttacgcgtaacaaatcgaatcattgaacgttcgcgcgagatcgctctgcttatctcgcccggatCATATGAATATCCTCCTTAG
c:
участком (прописные буквы), гомологичным последовательности (1932548-1932628) гена edd (1930817-1932628),
участком (заглавные буквы) для амплифицирования кластера, резистентного к хлорамфениколу, находящегося в плазмиде pKD3.
Олигонуклеотиды edd D r и edd D f использовали для амплифицирования кластера, резистентного к хлорамфениколу, в плазмиде pKD3. Полученный ПЦР-продукт затем вводили путем электропорации в штамм MG1655 (pKD46), в котором используемый фермент Red рекомбиназа обеспечивал гомологичную рекомбинацию. Трансформанты, резистентные к антибиотику, затем селекционировали и инсерцию кластера резистентности проверяли посредством анализа ПЦР с помощью олигонуклеотидов eda d и zwf r:
eda d (SEQ ID NO:35)
ССССGGAATCAGAGGAATAGTCCC (гомолог последовательности 1930439-1930462)
zwf r (SEQ ID NO:36)
GGGTAGACTCCATTACTGAGGCGTGGGCG (гомолог последовательности 1932968-1932996)
b) конструирование эволюционирующего модифицированного штамма MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ, Δedd::cm
Ген edd инактивировали в эволюционирующем штамме MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ, как указано в примере 1, методом с использованием фага Р1 (см. протокол стадии с)). Лизат фага Р1 получали на штамме MG1655 Δedd::cm и трансдукцию штамма MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ проводили с помощью указанного лизата. Трансдукты, резистентные к хлорамфениколу, контролировали с помощью олигонуклеотидов eda d и zwf r для проверки мутации гена edd.
с) культивирование модифицированных эволюционирующих штаммов E. coli *MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ, Δedd::cm и E coli *MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ
Оба модифицированных эволюционирующих штамма E. coli *MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ, Δedd::cm и E. coli *MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ культивировали при нерегулируемом рН в минимальной культуральной среде с добавлением нитрата натрия и экстракта дрожжей с исходной концентрацией глюкозы 20 г/л в условиях анаэробиоза в течение 10 дней. Полученный профиль продуктов ферментации показывает, что делеция гена edd вызывает повышение выхода превращения глюкозы в 1,2-пропандиол от 0,13 г/г до 0,35 г/г (таблица 2). Делеция гена edd в эволюционирующем штамме E. coli *MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ обеспечивает таким образом улучшение рабочих характеристик штамма.
Сравнение концентраций субстрата и продуктов ферментации через 10 дней с начала культивирования модифицированных эволюционирующих штаммов E. coli *MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ, Δ edd::cm и E. coli *MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ
глюкозы
(г/г)
ΔtpiA, ΔpflAB, ΔadhE, dhA::kana, ΔgloA, ΔaldA, ΔaldВ,
Δedd::cm
ΔtpiA, ΔpflAB, ΔadhE, dhA::kana, ΔgloA, ΔaldA, ΔaldВ
Пример 6. Конструирование модифицированного штамма E. coli *MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ, Δedd::cm (pSOS95Т), способного продуцировать 1,2-пропандиол и ацетон
Плазмида, называемая pSOS95Т, является челночным вектором экспрессии E. coli С.acetobutylicum, несущим оперон ацетона Сlostridum acetobutylicum, состоящий из четырех генов adc, ctfAB, thl, кодирующих соответственно ацетоацетаткарбоксилазу, коэнзим А трансферазу и тиолазу в зависимости от промотора тиолазы. Эти три фермента катализируют трансформацию ацетил-СоА и ацетата в ацетон и диоксид углерода. Плазмида pSOS95Т была получена инсерцией в плазмиду pSOS95 (номер доступа в Gene Bank AY187686) гена thl C. Acetobutylicum, кодирующего тиолазу на сайте BamH1, находящемся между промотором тиолазы и геном ctfA. Плазмиду pSOS95Т вводили в эволюционирующий штамм E. coli *MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ, Δedd::cm путем электропорации.
Электрокомпетентные клетки штамма E. coli *MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ, Δedd::cm получали из ночной культуры штамма в LB. 10 мл культуры LB в колбе эрленмеера засевали 100-кратно разведенной ночной культурой и инкубировали при 37°С. Когда оптическая плотность при 500 нм культуры достигла значения от 0,4 до 0,6, 1 мл культуры подвергали центрифугированию. Клетки промывали водой, затем 10%-ным раствором глицерина, с повторным суспендированием в 0,05 мл 10%-ного раствора глицерина. Клетки сразу подвергали электропорации (25 мкФ, 200 Ом, 2,5 кВ)(Gene pulser, Biorad) с 5 мкл плазмидного препарата pSOS95Т (Qiagen, Hilden Germany). После 1 часа фенотипической экспрессии в среде SOC (Sambrook J., Fristch E.F. and Maniatis T. (1989) Molecular cloning: a laboratory manual, 2-nd ed Cold Spring Harbor Laboratory, cold Spring Harbor, N.Y.) при 37°С, трансформанты селекционировали на среде агар-агара с помощью карбенициллина 100 мкг/мл при 37°С.
Трансформанты повторно помещали на одну ночь в жидкую культуру в присутствии карбенициллина для проведения экстрагирования плазмидного ДНК (Qiagen, Hilden Germany) с тем, чтобы контролировать присутствие плазмиды pSOS95Т и проверять ее правильность путем ферментативной обработки.
Пример 7. Культивирование эволюционирующего модифицированного штамма E. coli *MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ, Δedd::cm (pSOS95Т), способного продуцировать 1,2-пропандиол и ацетон
Модифицированный штамм E. coli *MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ, Δedd::cm (pSOS95Т) культивировали при нерегулируемом рН в минимальной культуральной среде с добавлением нитрата натрия и экстракта дрожжей с исходной концентрацией глюкозы 20 г/л в условиях анаэробиоза (таблица 3). Анализ продуктов ферментации показывает, что эволюционирующий штамм E. coli *MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ, Δedd::cm (pSOS95Т) продуцирует смесь 1,2-пропандиола, ацетата и ацетона.
Сравнение концентраций субстрата и продуктов ферментации через 10 дней с начала культивирования модифицированных эволюционирующих штаммов E. coli *MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ, Δedd::cm и E. coli *MG1655 ΔtpiA, ΔpflAB, ΔadhE, IdhA::kana, ΔgloA, ΔaldA, ΔaldВ, Δedd::cm
(г/л)
(г/л)
ΔtpiA, ΔpflAB, ΔadhE, dhA::kana, ΔgloA, ΔaldA, ΔaldВ,
Δedd::cm
(pSOS95Т)
ΔtpiA, ΔpflAB, ΔadhE, dhA::kana, ΔgloA, ΔaldA, ΔaldВ
Δedd::cm
СПИСОК ЛИТЕРАТУРЫ
Altaras N. E. and Cameron D. (1999). Metabolic engineering of a 1,2-propanediol pathway in Escherichia coli: Appl. Env. Microb., 65, 1180-1185.
Altaras N. E. and Cameron D. C. (2000). Enhanced production of (R) 1,2-propanediol by metabolically engineered Escherichia coli: Biotechnol. Prog. 16, 940-946.
Altaras NE, Etzel MR and Cameron DC. (2001). Conversion of sugars to 1,2-propanediol by Thermoanaerobacterium thermosaccharolyticum HG-8: Biotechnol. Prog. 17, 52-56.
Bunch P. K., Mat-Jan F. and Clark D. P. (1997). The IdhA gene encoding the fermentative lactate dehydrogenase of Escherichia coli: microbiology, 143, 187-195.
Cameron D. C., Altaras N. E., Hoffman M. L. and Shaw A. J. (1998). Metabolic engineering of propanediol pathways: Biotechnol. Prog., 14, 116-125.
Cameron D. C., Shaw A. J. and Altaras N. E. (1998). Microbial production of 1,2-propanediol from sugar WO 98/37204.
Cameron D. C., Shaw A. J. and Altaras N. E. (2000). Microbial production of 1,2-propanediol from sugar US 6087140.
Cameron D. C., Shaw A. J. and Altaras N. E. (2001). Microbial production of 1,2-propanediol from sugar US 6303352.
Carrier Т. A. and Keasling J. D. (1999). Library of synthetic 5' secondary structures to manipulate mRNA stability in Escherichia coli, Biotechnol. prog., 15, 58-64.
Cheperanov P. P. and Wackernagel W. (1995). Gene disruption in Escherichia coli: Tc R and KmR cassettes with option of Flp-catalyzed excision of the antibiotic-resistance determinant, gene, 158, 9-14.
Datsenko, K.A. and Wanner, B.L. (2000). One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 97:6640-6645.
Sambrook J., Fristch E. F. and Maniatis T. (1989). Molecular cloning: a laboratory manual, 2nd ed Cold Spring Harbor Laboratory, cold Spring Harbor, N.Y.
Schwartz E. R. and Reed L. J. (1970). Regulation of the activity of the pyruvate dehydrogenase complex of Escherichia coli, Biochemistry, 6, 1434-1439.
Snoep J. L., De Graef M. R., Westphal A. H., De Kok A. Teixeira de Mattos M. J. and Neijssel O. M. (1993). Differences in sensitivity to NADH of purified pyruvate dehydrogenase complexes of Enterococcus faecalis, Lactococcus lactis, Azotobacter vinelandii and Escherichia coli: implications for their activity in vivo, FEMS microbiology letters, 114, 279-284)).
Tran Din К. and Gottschalk G. (1985). Formation of D(-)-1,2-propanediol and D(-)-lactate from glucose by Clostridium sphenoides under phosphate limitation: Arch. Microbiol. 142, 87-92.
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЙ МИКРООРГАНИЗМ И СПОСОБ ПОЛУЧЕНИЯ 1,2-ПРОПАНДИОЛА, ОСНОВАННЫЙ НА НАДФН-ЗАВИСИМОЙ АЦЕТОЛРЕДУКТАЗЕ И УЛУЧШЕННОЙ ДОСТАВКЕ НАДФН | 2015 |
|
RU2711870C2 |
| МИКРОБИОЛОГИЧЕСКИЙ СПОСОБ ПОЛУЧЕНИЯ 1,2-ПРОПАНДИОЛА | 2009 |
|
RU2521502C2 |
| ШТАММ БАКТЕРИЙ ESCHERICHIA COLI - ПРОДУЦЕНТ ФУМАРОВОЙ КИСЛОТЫ И СПОСОБ ПОЛУЧЕНИЯ ФУМАРОВОЙ КИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ ЭТОГО ШТАММА | 2014 |
|
RU2573936C1 |
| ШТАММ БАКТЕРИЙ ESCHERICHIA COLI - ПРОДУЦЕНТ ЯНТАРНОЙ КИСЛОТЫ (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ ЯНТАРНОЙ КИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ ЭТОГО ШТАММА | 2015 |
|
RU2603004C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ БАКТЕРИЙ Escherichia coli - ПРОДУЦЕНТ ЯНТАРНОЙ КИСЛОТЫ (ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ ЯНТАРНОЙ КИСЛОТЫ С ИСПОЛЬЗОВАНИЕМ ЭТОГО ШТАММА | 2012 |
|
RU2528056C2 |
| СПОСОБ ПОЛУЧЕНИЯ 2,4-ДИГИДРОКСИБУТИРАТА | 2013 |
|
RU2645260C2 |
| БАКТЕРИЯ Escherichia coli - ПРОДУЦЕНТ ЯНТАРНОЙ КИСЛОТЫ И СПОСОБ ПОЛУЧЕНИЯ ЯНТАРНОЙ КИСЛОТЫ | 2010 |
|
RU2466186C2 |
| МИКРООРГАНИЗМЫ ДЛЯ ПРОИЗВОДСТВА МЕТИОНИНА С УЛУЧШЕННЫМ ВЫХОДОМ МЕТИОНИНА | 2014 |
|
RU2678757C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА И L-АРГИНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ ESCHERICHIA, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН ybiV | 2005 |
|
RU2320719C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2008 |
|
RU2515044C2 |








Изобретение относится к биотехнологии. Способ получения бактерии Escherichia coli для продуцирования 1,2-пропандиола из исходной модифицированной бактерии, имеющей делецию в генах tpiA, gloA, aldA и aldB, включает культивирование указанной исходной модифицированной бактерии в соответствующей культуральной среде, содержащей источник углерода, с применением повышающихся степеней разведения таким образом, что в культуральной среде сохраняются только микроорганизмы, степень роста которых равна или больше используемой степени разведения, для эволюционирования в указанной исходной модифицированной бактерии одного или нескольких генов, участвующих в пути биосинтеза DHAP в метилглиоксаль, затем в 1,2-пропандиол, до получения эволюционирующих генов, кодирующих ферменты, обладающие более высокой активностью «1,2-пропандиолсинтазы», селекцию и выделение одной или нескольких бактерий Escherichia coli, обладающих более высокой активностью «1,2-пропандиолсинтазы». Изобретение также касается исходных микроорганизмов и таким образом полученных эволюционирующих микроорганизмов и способа получения 1,2-пропандиола и ацетона путем культивирования эволюционирующих микроорганизмов. Изобретение позволяет повысить эффективность продуцирования 1,2 пропандиола. 6 н. и 13 з.п. ф-лы, 3 ил., 3 табл.
1. Способ получения бактерии Escherichia coli для продуцирования 1,2-пропандиола путем метаболизма источника углерода из исходной модифицированной бактерии, имеющей делецию в генах tpiA, gloA, aldA и aldB,
где указанный способ включает следующие стадии:
культивирование указанной исходной модифицированной бактерии в соответствующей культуральной среде, содержащей источник углерода, с применением повышающихся степеней разведения таким образом, что в культуральной среде сохраняются только микроорганизмы, степень роста которых равна или больше используемой степени разведения, для эволюционирования в указанной исходной модифицированной бактерии одного или нескольких генов, участвующих в пути биосинтеза DHAP в метилглиоксаль, затем в 1,2-пропандиол, до получения эволюционирующих генов, кодирующих ферменты, обладающие более высокой активностью «1,2-пропандиолсинтазы»,
селекция и выделение одной или нескольких бактерий Escherichia coli, обладающих более высокой активностью «1,2-пропандиолсинтазы».
2. Способ по п.1, отличающийся тем, что исходная модифицированная бактерия имеет также делецию в генах IdhA, pflA, pflB, adhE и edd.
3. Способ по п.1, отличающийся тем, что исходная модифицированная бактерия содержит также по меньшей мере один ген, кодирующий фермент, способствующий метаболизму пирувата в ацетат.
4. Способ по п.3, отличающийся тем, что фермент, способствующий метаболизму пирувата в ацетат, является слабочувствительным к ингибированию посредством NADH.
5. Способ по п.3, отличающийся тем, что указанный фермент способствует метаболизму пирувата до продуцирования ацетил-СоА и NADH.
6. Способ по п.3, отличающийся тем, что фермент является комплексом пируватдегидроксигеназы.
7. Способ по п.3, отличающийся тем, что фермент, способствующий метаболизму пирувата в ацетат, является эндогенным ферментом.
8. Способ по п.1, отличающийся тем, что в бактерию Escherichia coli, обладающую более высокой активностью «1,2-пропандиолсинтазы», вводят один или несколько гетерологичных генов, кодирующих один или несколько ферментов, участвующих в превращении ацетил-СоА и ацетата в ацетон.
9. Способ по п.8, отличающийся тем, что один или несколько гетерологичных генов, кодирующих один или несколько ферментов, участвующих в превращении ацетил-СоА и ацетата, получают из С.acetobutylicum.
10. Способ по п.8, отличающийся тем, что культивируют в условиях селекционного давления в соответствующей культуральной среде, содержащей источник углерода, бактерию Escherichia coli, обладающую более высокой активностью «1,2-пропандиолсинтазы», полученную по п.8, для эволюционирования в указанной модифицированной бактерии одного или нескольких генов, участвующих в превращении ацетил-СоА и ацетата в ацетон, до получения более высокой активности «ацетонсинтазы», затем селекционируют и выделяют микроорганизмы второго поколения, обладающие более высокой активностью «1,2-пропандиолсинтазы» и более высокой активностью «ацетонсинтазы».
11. Исходная модифицированная бактерия Escherichia coli, имеющая делецию в генах tpiA, gloA, aldA и aldB, предназначенная для использования в способе по п.1 для получения штамма, предназначенного для продукции 1,2-пропандиола.
12. Бактерия Escherichia coli, обладающая более высокой активностью «1,2-пропандиолсинтазы», полученная способом по любому из пп.1-7, предназначенная для продукции 1,2-пропандиола.
13. Бактерия Escherichia coli, обладающая более высокой активностью «1,2-пропандиолсинтазы» и более высокой активностью «ацетонсинтазы», полученная способом по п.10, предназначенная для продукции 1,2-пропандиола и ацетона.
14. Бактерия Escherichia coli по любому из пп.12 или 13, в которой ген lpd несет точечную мутацию, вызывающую модификацию аланина 55 в валин.
15. Способ получения 1,2-пропандиола, в ходе которого культивируют бактерию Escherichia coli, обладающую более высокой активностью «1,2-пропандиолсинтазы», по п.12 в соответствующей культуральной среде, содержащей источник углерода, затем извлекают продуцированный 1,2-пропандиол.
16. Способ получения 1,2-пропандиола по п.15, отличающийся тем, что бактерию Escherichia coli, обладающую более высокой активностью «1,2-пропандиолсинтазы», культивируют в анаэробных условиях.
17. Способ получения 1,2-пропандиола и ацетона, в ходе которого культивируют бактерию Escherichia coli, обладающую более высокой активностью «1,2-пропандиолсинтазы» и более высокой активностью «ацетонсинтазы», по п.13 в соответствующей культуральной среде, содержащей источник углерода, затем извлекают продуцированный 1,2-пропандиол и ацетон.
18. Способ получения 1,2-пропандиола и ацетона по п.17, отличающийся тем, что бактерию Escherichia coli, обладающую более высокой активностью «1,2-пропандиолсинтазы» и более высокой активностью «ацетонсинтазы», культивируют в анаэробных условиях.
19. Способ по любому из пп.15-18, отличающийся тем, что 1,2-пропандиол и/или ацетон очищают.
| WO 9837204, 27.08.1998 | |||
| US 6303352 B1, 16.10.2001 | |||
| US 6514733, 04.02.2003 | |||
| ФРАГМЕНТ ДНК ИЗ ESCHERICHIA COLI, ОПРЕДЕЛЯЮЩИЙ ПОВЫШЕННУЮ ПРОДУКЦИЮ L-АМИНОКИСЛОТ (ВАРИАНТЫ), И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ | 1999 |
|
RU2175351C2 |

