Настоящее изобретение относится к способу получения 1,2-пропандиола из глицерина в клетках-хозяевах. Конкретнее, настоящее изобретение описывает рекомбинантные ферментативные активности, которые способны синтезировать исключительно 1,2-изомер пропандиола из чистых и неочищенных препаратов глицерина. Настоящее изобретение также относится к подходящим сочетаниям сверхэкспрессии и инактивации ключевых активностей для получения 1,2-пропандиола.
1,2-Пропандиол (пропиленгликоль; 1,2-PD) является основным объемным химическим продуктом, который широко используется в качестве компонента ненасыщенных полиэфирных смол, фармацевтических препаратов и косметических средств, жидких моющих средств, хладагентов и антифризов или антиобледенителей. Так как 1,2-PD является оптически активным, энантиомерно чистый препарат 1,2-PD может представлять особый интерес для медицинских, сельскохозяйственных или физиологических применений.
В настоящее время 1,2-пропандиол получают из нефтехимических продуктов химическим синтезом, который включает работу с большими количествами токсичных соединений, подобных эпихлоргидрину или гидропероксиду. В обычном химическом синтезе 1,2-PD получают гидратированием пропиленоксида, который получают из пропилена. Химический синтез дает рацемический 1,2-PD, и требуются большие количества воды для того, чтобы предотвратить образование полигликоля. Обычный химический синтез зависит от запасов ископаемых и ведет к получению больших количеств побочных продуктов; таким образом, он оказывается проблематичным в смысле воздействия на окружающую среду и экономических аспектов.
Известно, что 1,2-пропандиол может вырабатываться микроорганизмами из сахаров как субстратов (Kluyver and Schellen, 1937) (Heath B.C., 1962) (Altaras N.E., 2001) (Tran Din, K, 198) (Cameron D.C., 1986) (Cameron D.C., 1998) (Park Y.H., 2006; US 7049109 B2).
В US 6087140 и US 6303352 описывается получение 1,2-PD из сахаров, за исключением 6-дезоксигексоз, с помощью рекомбинантных организмов.
В WO 2005/073364, US 2007/072279, описывается способ, которым генерируют и отбирают микроорганизмы, которые показывают усиленные способности вырабатывать 1,2-PD из не указанных точно источников углерода. Конкретнее, описывается инактивация набора генов для создания штаммов, несущих одну или несколько мутаций, которые являются основой для последующей процедуры отбора ферментацией в хемостате. Центром заявки является инактивация генов, кодирующих активность aldA и gloA. Все раскрытые примеры даются для Е.coli MG1655, который имеет мутации в по меньшей мере следующих двух генах: тризофосфатизомеразы (triA) и обеих субъединицах пируватформиатлиазы (pflAB). Кроме того, примеры конкретно относятся к глюкозе как источнику углерода для ферментации.
Поэтому существует потребность в улучшении биотехнологических способов получения 1,2-пропандиола (1,2-PD). Также существует потребность в улучшенных микробиологических штаммах, которые можно использовать в таком способе.
Настоящее изобретение направлено на решение такой потребности, предлагая решения проблем, которые до сих пор препятствовали существенным улучшениям в данной области.
Настоящее изобретение относится к способу получения 1,2-пропандиола (1,2-PD) из неспособного к брожению дешевого углеродного субстрата, в соответствии с которым углеродный субстрат поддерживает выработку биомассы и в то же время служит в качестве субстрата для выработки 1,2-пропандиола (1,2-PD). Настоящее изобретение также относится к микробиологическим штаммам, которые специфически адаптированы к определенным требованиям способа по изобретению и поэтому специфически подходят для использования в способе по изобретению.
В частности, настоящее изобретение относится к клетке-хозяину, в особенности микроорганизму или штамму, которые созданы методами генной инженерии для выработки высоких уровней 1,2-пропандиола (1,2-PD), при выращивании на неспособном
к брожению углеродном субстрате, в соответствии с чем углеродный субстрат поддерживает выработку биомассы и в то же время служит в качестве субстрата для выработки 1,2-пропандиола (1,2-PD), особенно, когда клетки выращивают на глицерине как единственном источнике углерода.
В одном воплощении изобретение относится к клетке-хозяину, в особенности микроорганизму или штамму, которые созданы методами генной инженерии для выработки высоких уровней 1,2-пропандиола (1,2-PD), при выращивании на глицерине как единственном источнике углерода, где указанный глицерин имеет степень чистоты по меньшей мере 70%, в особенности по меньшей мере 75%, в особенности по меньшей мере 80%, в особенности по меньшей мере 85%, в особенности по меньшей мере 90%, в особенности по меньшей мере 95%, в особенности по меньшей мере 99% и до 100%, причем в связи с этим также рассматриваются все целые числа, попадающие в вышеуказанные интервалы.
В конкретном воплощении глицерин имеет степень чистоты от 80% до 90%, в особенности примерно 85%.
В одном воплощении изобретение относится к клетке-хозяину, в особенности микроорганизму или штамму, способным вырабатывать высокие уровни 1,2-пропандиола (1,2-PD), при выращивании на глицерине как единственном источнике углерода, где указанная клетка-хозяин, в особенности указанный микроорганизм или штамм, созданы методами генной инженерии для сверхэкспрессии пропандиолоксидоредуктазы (fucO), в особенности путем введения гена, кодирующего активность пропандиолоксидоредуктазы (fucO).
В одном воплощении изобретение относится к клетке-хозяину, в особенности микроорганизму или штамму, способным вырабатывать высокие уровни 1,2-пропандиола (1,2-PD), при выращивании на глицерине как единственном источнике углерода, где указанная клетка-хозяин, в особенности указанный микроорганизм или штамм, созданы методами генной инженерии для коэкспрессии по меньшей мере одного белка-фермента, выбранного из группы, состоящей из глицеролдегидрогеназы (gldA), дигидроксиацетонкиназы (dhaK) и метилглиоксальсинтазы (mgsA), вместе с активностью пропандиолоксидоредуктазы (fucO), в особенности путем совместного введения в указанную клетку-хозяина вместе с геном, кодирующим активность пропандиолоксидоредуктазы (fucO) по меньшей мере одного другого гена, кодирующего ферментативную активность, выбранную из группы, состоящей из глицеролдегидрогеназы (gldA), дигидроксиацетонкиназы (dhaK) и метилглиоксальсинтазы (mgsA), как, например, для экспрессии указанных активностей вместе с активностью пропандиолоксидоредуктазы (fucO).
В одном воплощении изобретение относится к клетке-хозяину, в особенности микроорганизму или штамму, способным вырабатывать высокие уровни 1,2-пропандиола (1,2-PD), при выращивании на глицерине как единственном источнике углерода, где указанная клетка-хозяин, в особенности указанный микроорганизм или штамм, созданы методами генной инженерии для коэкспрессии активности глицеролдегидрогеназы (gldA) вместе с активностью пропандиолоксидоредуктазы (fucO), в особенности путем совместного введения в указанную клетку-хозяина вместе с геном, кодирующим активность пропандиолоксидоредуктазы (fucO), гена, кодирующего активность глицеролдегидрогеназы (gldA), как, например, для экспрессии указанной активности глицеролдегидрогеназы (gldA) вместе с активностью пропандиолоксидоредуктазы (fucO).
В одном воплощении изобретение относится к клетке-хозяину, в особенности микроорганизму или штамму, способным вырабатывать высокие уровни 1,2-пропандиола (1,2-PD), при выращивании на глицерине как единственном источнике углерода, где указанная клетка-хозяин, в особенности указанный микроорганизм или штамм, созданы методами генной инженерии для коэкспрессии активности дигидроксиацетонкиназы (dhaK) вместе с активностью пропандиолоксидоредуктазы (fucO), в особенности путем совместного введения в указанную клетку-хозяина вместе с геном, кодирующим активность пропандиолоксидоредуктазы (fucO), гена, кодирующего активность дигидроксиацетонкиназы (dhaK), как, например, для экспрессии указанной активности дигидроксиацетонкиназы (dhaK) вместе с активностью пропандиолоксидоредуктазы (fucO).
В одном воплощении изобретение относится к клетке-хозяину, в особенности микроорганизму или штамму, способным вырабатывать высокие уровни 1,2-пропандиола (1,2-PD), при выращивании на глицерине как единственном источнике углерода, где указанная клетка-хозяин, в особенности указанный микроорганизм или штамм, созданы методами генной инженерии для коэкспрессии активности метилглиоксальсинтазы (mgsA) вместе с активностью пропандиолоксидоредуктазы (fucO), в особенности путем совместного введения в указанную клетку-хозяина вместе с геном, кодирующим активность пропандиолоксидоредуктазы (fucO), гена, кодирующего активность метилглиоксальсинтазы (mgsA), как, например, для экспрессии указанной активности метилглиоксальсинтазы (mgsA) вместе с активностью пропандиолоксидоредуктазы (fucO).
В одном воплощении изобретение относится к клетке-хозяину, в особенности микроорганизму или штамму, способным вырабатывать высокие уровни 1,2-пропандиола (1,2-PD), при выращивании на глицерине как единственном источнике углерода, где указанная клетка-хозяин, в особенности указанный микроорганизм или штамм, созданы методами генной инженерии для коэкспрессии активности глицеролдегидрогеназы (gldA) и дигидроксиацетонкиназы (dhaK) вместе с активностью пропандиолоксидоредуктазы (fucO), в особенности путем совместного введения в указанную клетку-хозяина вместе с геном, кодирующим активность пропандиолоксидоредуктазы (fucO), генов, кодирующих активность и дигидроксиацетонкиназы (dhaK), как, например, для экспрессии указанных активностей глицеролдегидрогеназы (gldA) и дигидроксиацетонкиназы (dhaK) вместе с активностью пропандиолоксидоредуктазы (fucO).
В одном воплощении изобретение относится к клетке-хозяину, в особенности микроорганизму или штамму, способным вырабатывать высокие уровни 1,2-пропандиола (1,2-PD), при выращивании на глицерине как единственном источнике углерода, где указанная клетка-хозяин, в особенности указанный микроорганизм или штамм, созданы методами генной инженерии для коэкспрессии активности глицеролдегидрогеназы (gldA) и метилглиоксальсинтазы (mgsA) вместе с активностью пропандиолоксидоредуктазы (fucO), в особенности путем совместного введения в указанную клетку-хозяина вместе с геном, кодирующим активность пропандиолоксидоредуктазы (fucO), генов, кодирующих активность глицеролдегидрогеназы (gldA) и метилглиоксальсинтазы (mgsA), как, например, для экспрессии указанных активностей глицеролдегидрогеназы (gldA) и метилглиоксальсинтазы (mgsA) вместе с активностью пропандиолоксидоредуктазы (fucO).
В одном воплощении изобретение относится к клетке-хозяину, в особенности микроорганизму или штамму, способным вырабатывать высокие уровни 1,2-пропандиола (1,2-PD), при выращивании на глицерине как единственном источнике углерода, где указанная клетка-хозяин, в особенности указанный микроорганизм или штамм, созданы методами генной инженерии для коэкспрессии активности дигидроксиацетонкиназы (dhaK) и метилглиоксальсинтазы (mgsA) вместе с активностью пропандиолоксидоредуктазы (fucO), в особенности путем совместного введения в указанную клетку-хозяина вместе с геном, кодирующим активность пропандиолоксидоредуктазы (fucO), генов, кодирующих активности дигидроксиацетонкиназы (dhaK) и метилглиоксальсинтазы (mgsA), как, например, для экспрессии указанных активностей дигидроксиацетонкиназы (dhaK) и метилглиоксальсинтазы (mgsA) вместе с активностью пропандиолоксидоредуктазы (fucO).
В одном воплощении изобретение относится к клетке-хозяину, в особенности микроорганизму или штамму, способным вырабатывать высокие уровни 1,2-пропандиола (1,2-PD), при выращивании на глицерине как единственном источнике углерода, где указанная клетка-хозяин, в особенности указанный микроорганизм или штамм, созданы методами генной инженерии для коэкспрессии активности глицеролдегидрогеназы (gldA), дигидроксиацетонкиназы (dhaK) и метилглиоксальсинтазы (mgsA) вместе с активностью пропандиолоксидоредуктазы (fucO), в особенности путем совместного введения в указанную клетку-хозяина вместе с геном, кодирующим активность пропандиолоксидоредуктазы (fucO), генов, кодирующих активности глицеролдегидрогеназы (gldA), дигидроксиацетонкиназы (dhaK) и метилглиоксальсинтазы (mgsA), как, например, для экспрессии указанных активностей глицеролдегидрогеназы (gldA), дигидроксиацетонкиназы (dhaK) и метилглиоксальсинтазы (mgsA) вместе с активностью пропандиолоксидоредуктазы (fucO).
В одном воплощении изобретение относится к клетке-хозяину, в особенности микроорганизму или штамму, способным вырабатывать высокие уровни 1,2-пропандиола (1,2-PD), при выращивании на глицерине как единственном источнике углерода, где указанная клетка-хозяин, в особенности указанный микроорганизм или штамм, созданы методами генной инженерии для коэкспрессии активности глицеролдегидратазы вместе с активностью пропандиолоксидоредуктазы (fucO), в особенности путем совместного введения в указанную клетку-хозяина вместе с геном, кодирующим активность пропандиолоксидоредуктазы (fucO), генов, кодирующих активность глицеролдегидратазы, как, например, для экспрессии указанной активности глицеролдегидратазы вместе с активностью пропандиолоксидоредуктазы (fucO).
В одном воплощении изобретение относится к клетке-хозяину, в особенности микроорганизму или штамму, способным вырабатывать высокие уровни 1,2-пропандиола (1,2-PD), при выращивании на глицерине как единственном источнике углерода, где указанная клетка-хозяин, в особенности указанный микроорганизм или штамм, созданы методами генной инженерии для коэкспрессии активности альдокеторедуктазы вместе с активностью пропандиолоксидоредуктазы (fucO), в особенности активности альдокеторедуктазы, которая вносится геном, в особенности геном микроорганизма, выбранным из группы, состоящей из dkgA, dkgB, yeaE и yghZ, в особенности путем совместного введения в указанную клетку-хозяина вместе с геном, кодирующим активность пропандиолоксидоредуктазы (fucO), генов, кодирующих активность альдокеторедуктазы, в особенности гена микроорганизма, выбранного из группы, состоящей из dkgA, dkgB, yeaE и yghZ, как, например, для экспрессии указанной активности альдокеторедуктазы вместе с активностью пропандиолоксидоредуктазы (fucO).
В одном воплощении изобретение относится к клетке-хозяину, в особенности микроорганизму или штамму, способным вырабатывать высокие уровни 1,2-пропандиола (1,2-PD), при выращивании на глицерине как единственном источнике углерода, где указанная клетка-хозяин, в особенности указанный микроорганизм или штамм, созданы методами генной инженерии рекомбинантных ДНК.
В одном воплощении изобретение относится к клетке-хозяину, в особенности микроорганизму или штамму, способным вырабатывать высокие уровни 1,2-пропандиола (1,2-PD), при выращивании на глицерине как единственном источнике углерода, где указанная клетка-хозяин, в особенности указанный микроорганизм или штамм, является дефективным по метаболизму арабинозы. В одном воплощении указанная клетка-хозяин, в особенности указанный микроорганизм или штамм, является дефектным по метаболизму арабинозы из-за уменьшенной или отсутствующей активности рибулозкиназы.
В одном воплощении изобретение относится к клетке-хозяину, в особенности микроорганизму или штамму по изобретению и описанным в данном описании выше, способным вырабатывать высокие уровни 1,2-пропандиола (1,2-PD), при выращивании на глицерине как единственном источнике углерода, где указанная клетка-хозяин, в особенности указанный микроорганизм или штамм, является дефектным по метаболизму метилглиоксаля. В одном воплощении указанная клетка-хозяин, в особенности указанный микроорганизм или штамм, является дефектным по метаболизму метилглиоксаля из-за уменьшенной или отсутствующей ферментативной активности, выбранной из группы, состоящей из активности системы глиоксилазы I, системы глиоксилазы II, лактатдегидрогеназы А, системы глиоксилазы III, альдегиддегидрогеназы А, но в особенности из-за уменьшенной или отсутствующей активности системы глиоксилазы I.
В одном воплощении изобретение относится к клетке-хозяину, в особенности микроорганизму или штамму по изобретению и описанным в данном описании выше, способным вырабатывать высокие уровни 1,2-пропандиола (1,2-PD), при выращивании на глицерине как единственном источнике углерода, где указанная клетка-хозяин, в особенности указанный микроорганизм или штамм, является дефектным по метаболизму дигидрооксиацетонфосфата. В одном воплощении указанная клетка-хозяин, в особенности указанный микроорганизм или штамм, является дефектным по метаболизму дигидрооксиацетонфосфата из-за уменьшенной активности триозофосфатизомеразы.
В одном воплощении изобретение относится к клетке-хозяину, в особенности микроорганизму или штамму по изобретению и описанным в данном описании выше, где указанный микроорганизм представляет собой Е.coli.
В одном воплощении изобретение относится к способу получения 1,2-пропандиола, в соответствии с которым клетку-хозяина, в особенности штамм микроорганизма по изобретению выращивают в соответствующей среде для выращивания, содержащей простой источник углерода, в особенности неочищенный препарат глицерина, после чего 1,2-пропандиол извлекают и при необходимости очищают.
В частности, изобретение относится к способу получения 1,2-пропандиола посредством выращивания клетки-хозяина, в особенности микроорганизма или штамма по изобретению на неспособном к брожению углеродном субстрате, включающему
i) культивирование указанной клетки, в особенности микроорганизма или штамма по изобретению и описанных в данном описании выше, которые сверхэкспрессируют пропандиолоксидоредуктазу (fucO), в среде, содержащей неспособный к брожению углеродный субстрат, в связи с чем углеродный субстрат поддерживает выработку биомассы и в то же время служит в качестве субстрата для продуцирования 1,2-пропандиола (1,2-PD), и неспособный к брожению источник углерода метаболизируется клеткой-хозяином, в особенности микроорганизмом или штаммом по изобретению в 1,2-пропандиол,
ii) извлечение 1,2-пропандиола, полученного на стадии i); и, необязательно,
iii) очистку извлеченного 1,2-пропандиола.
В одном воплощении изобретения указанный неспособный к брожению углеродный субстрат представляет собой неочищенный препарат глицерина, в особенности препарат, содержащий глицерин с чистотой по меньшей мере 70%, в особенности по меньшей мере 75%, в особенности по меньшей мере 80%, в особенности по меньшей мере 85%, в особенности по меньшей мере 90%, в особенности по меньшей мере 95%, в особенности по меньшей мере 99% и до 100%.
В конкретном воплощении глицерин имеет степень чистоты от 80% до 90%, в особенности примерно 85%.
В одном воплощении неспособный к брожению углеродный субстрат, в особенности неочищенный препарат глицерина, описанный в данном описании выше, селективно метаболизируется до 1,2-пропандиола.
В одном воплощении изобретение относится к способу получения 1,2-пропандиола, описанному в данном описании ранее, где в указанном способе используют клетку-хозяина, в особенности микроорганизм или штамм по изобретению и описанные в данном описании ранее, которые созданы методами генной инженерии для сверхэкспрессии пропандиолоксидоредуктазы (fucO).
В одном воплощении изобретение относится к способу получения 1,2-пропандиола, описанному в данном описании ранее, где в указанном способе используют клетку-хозяина, в особенности микроорганизм или штамм по изобретению и описанные в данном описании ранее, которые созданы методами генной инженерии для коэкспрессии с по меньшей мере одним другим белком-ферментом, выбранным из группы, состоящей из глицеролдегидрогеназы (gldA), дигидроксиацетонкиназы (dhaK) и метилглиоксальсинтазы (mgsA), вместе с активностью пропандиолоксидоредуктазы (fucO).
В одном воплощении изобретения используют клетку-хозяина, в особенности микроорганизм или штамм по изобретению и описанные в данном описании ранее, где по меньшей мере одну ферментативную активность, вовлеченную в непродуктивный путь, конкурирующий с продуцированием 1,2-PD, инактивируют.
В частности/используют мутантов микроорганизмов, в особенности мутантов Е.coli, в которых инактивированы один или несколько генов, кодирующих системы глиоксилазы I и II (gloA и gloB), лактатгидрогеназу A (IdhA) систему глиоксилазы III (косвенно, путем инактивации основного регулятора rpoS) и альдегиддегидрогеназу.
В другом воплощении используют мутанта микроорганизма или штамм, в особенности мутанта Е.coli, в котором ген, кодирующий активность gloA, частично или полностью инактивирован.
В другом воплощении в способе по изобретению используют мутанта микроорганизма или штамм, инактивированные по метаболизму арабинозы.
В одном воплощении изобретения в качестве организма-хозяина используют штамм Е.coli, в особенности штамм Е.coli MG1655 и DH5-альфа, соответственно.
В одном воплощении изобретения по меньшей мере один из генов, кодирующих ферментативную активность, выбранную из группы, состоящей из глицеролдегидрогеназы (gldA), дигидроксиацетонкиназы (dhaK) и метилглиоксальсинтазы (mgsA) и пропандиолоксидоредуктазы (fucO), находится под контролем индуцируемого промотора, в особенности арабинозаиндуцируемого промотора, в особенности промотора paraBAD.
В одном воплощении изобретение относится к синтетическому оперону, который используют в способе по изобретению для предоставления клетки-хозяина, в особенности микроорганизма или штамма, коэкспрессирующих по меньшей мере одну ферментативную активность, выбранную из группы, состоящей из активности глицеролдегидрогеназы (gldA), дигидроксиацетонкиназы (dhaK) и метилглиоксальсинтазы (mgsA) вместе с активностью пропандиолоксидоредуктазы (fucO). В одном воплощении изобретения гены, кодирующие вышеуказанные активности, находятся под контролем индуцируемого промотора, в особенности арабинозаиндуцируемого промотора, но в особенности промотора paraBAD.
В одном воплощении изобретение относится к синтетическому оперону, включающему ген, кодирующий пропандиолоксидоредуктазу (fucO) и по меньшей мере один другой ген, кодирующий белок-фермент, выбранный из группы, состоящей из глицеролдегидрогеназы, дигидроксиацетонкиназы и метилглиоксальсинтазы (mgsA), в особенности к синтетическому оперону, включающему гены, кодирующие пропандиолоксидоредуктазу (fucO), глицеролдегидрогеназу, дигидроксиацетонкиназу и метилглиоксальсинтазу (mgsA).
В одном воплощении синтетический оперон по изобретению находится под контролем индуцируемого промотора, в особенности арабинозаиндуцируемого промотора.
В одном воплощении изобретения гены, кодирующие ряд генов, транскрибированных после индукции от указанного оперона, следующий: mgsA, gldA, dhaK, fucO.
Изобретение также относится к полинуклеотидным молекулам или конструкциям, в особенности плазмидам и молекулам-векторам, включающим синтетический оперон по изобретению и описанный в данном описании выше, и к клеткам-хозяевам, в особенности клеткам-хозяевам микроорганизмов, включающим указанные полинуклеотидные молекулы.
Краткое описание чертежей
Фигура 1 иллюстрирует ингибирование роста штаммов Е.coli дикого типа (сплошные столбики) и мутантных штаммов (□gloA-мутант серые столбики; □gloB-мутант заштрихованные столбики) различными количествами метилглиоксаля, добавленного в культуральную жидкость.
Фигура 2 представляет собой схематическое изображение путей получения 1,2-PD, когда углеродными субстратами являются сахара или глицерин.
Фигуры 3 и 4 иллюстрируют карты плазмид, описанных в изобретении.
Определения
Подразумевается, что термин «полинуклеотид» в данном описании относится к полимерной молекуле высокой молекулярной массы, которая может быть одноцепочечной или двухцепочечной, состоящей из мономеров (нуклеотидов), содержащих сахар, фосфат и основание, которые находятся или в пурине или в пиримидине. Таким образом, термин «полинуклеотид» главным образом относится к полимеру ДНК или РНК, которые могут быть одно- или двухцепочечными, необязательно содержащими синтетические, неприродные или измененные нуклеотидные основания, способные включаться в полимеры ДНК или РНК. Если не указано иное, определенная нуклеотидная последовательность по данному изобретению также потенциально охватывает ее консервативно модифицированные варианты (например, замены вырожденных кодонов) и комплементарные последовательности, а также точно указанную последовательность. Конкретно, замен вырожденных кодонов можно достигнуть, генерируя последовательности, в которых третья позиция одного или нескольких (или всех) выбранных кодонов заменена смешанными основаниями и/или дезоксиинозиновыми остатками (Batzer et al. (1991); Ohtsuka et al. (1985); и Rossolini et al. (1994)). Термин «полинуклеотид» используется как взаимозаменяемый с нуклеиновой кислотой, нуклеотидной последовательностью и может включать гены, кДНК и мРНК, кодированные геном, и т.д.
Термин «конструкция» относится к плазмиде, вирусу, автономно реплицирующей последовательности, фагу или нуклеотидной последовательности, линейной или циклической одно- или двухцепочечной ДНК или РНК, полученным из любого источника, в которых ряд нуклеотидных последовательностей соединен или рекомбинирован в уникальную конструкцию, которая способна вводить в клетку фрагмент промотора и последовательность ДНК для выбранного генного продукта, кодирующего ферментативную активность по изобретению, вместе с соответствующей 3'-нетранслируемой последовательностью.
Термин «трансформация» или «трансфекция» относится к приобретению новых генов в клетке после включения нуклеиновой кислоты.
Термин «экспрессия» относится к транскрипции и трансляции генного продукта из гена, кодирующего последовательность генного продукта. При экспрессии цепь ДНК, кодирующей последовательность генного продукта, сначала транскрибируют до комплементарной РНК, которая часто представляет собой матричную РНК, и затем транскрибированную таким образом матричную ДНК транслируют в вышеуказанный генный продукт, если генный продукт представляет собой белок.
Термины «плазмида» или «вектор» или «космида», используемые в данном описании, относятся к дополнительному хромосомному элементу, обычно несущему гены, которые не являются частью основного метаболизма клетки, и обычно в форме циклических двухцепочечных молекул ДНК.
Термин «регулятор», используемый в настоящем описании, относится к последовательности оснований, имеющей функциональный промотор и любой родственный транскрипционный элемент (например, энхансер, бокс ССААТ, бокс ТАТА, группу SPI и т.п.).
Термин «функционально связанный», используемый в настоящем описании, означает, что различные регуляторные элементы, такие как промотер, энхансер и т.п., которые регулируют экспрессию генов, и ген, представляющий интерес, соединены в функциональном состоянии в клетке-хозяине, как, например, для возможности экспрессии указанного гена, представляющего интерес. Специалистам в данной области техники хорошо известно, что тип и вид регулятора может весьма изменяться в зависимости от хозяина.
Термин «делеция» обозначает супрессию активности гена, которая вообще заключается в супрессии активности, которая может представлять собой инактивацию, ингибирование или она может представлять собой делецию по меньшей мере части рассматриваемого гена (например, делецию всего или части промоторного участка, необходимого для его экспрессии), так что он не экспрессируется или нефункционален, или так, что продукт экспрессии теряет свою функцию (например, делеция в кодирующей части рассматриваемого гена). Преимущественно делеция гена, по существу, представляет собой супрессию такого гена, который может быть заменен маркерным геном селекции, что облегчает идентификацию, отделение и очистку штаммов по изобретению. Например, ген может быть инактивирован гомологичной рекомбинацией, опосредуемой recA-белком, например Е.coli (Cunningham et al. (1980)).
Коротко, протокол инактивации может быть следующим: линейный фрагмент ДНК вводят в клетку. Указанный фрагмент получают in vitro, и он включает два участка, фланкирующих ген, и ген, кодирующий селектируемый генный продукт (как правило, ген устойчивости к антибиотикам), расположенный между двумя фланкирующими участками. Таким образом, такой фрагмент представляет собой инактивированный ген. Клетки, которые подверглись рекомбинации и интегрировали синтетический фрагмент, отбирают культивированием в селективной среде для выращивания. Отбирают клетки, которые подверглись двойной рекомбинации, в которых нативный ген заменен инактивированным геном.
Термин «углеродный субстрат» обозначает любой источник углерода, способный метаболизироваться микроорганизмом, где субстрат содержит по меньшей мере один атом углерода.
Термин «неспособный к брожению углеродный субстрат», используемый в данном описании, относится к углеродным субстратам, которые не поддерживают окислительно-восстановительные процессы данного организма для образования биомассы в отсутствие экзогенных акцепторов электронов.
1,2-Пропандиол (пропиленгликоль; 1,2-PD) является основным объемным химическим продуктом, который широко используется в качестве компонента ненасыщенных полиэфирных смол, фармацевтических препаратов и косметических средств, жидких моющих средств, хладагентов и антифризов или антиобледенителей. Так как 1,2-пропандиол (1,2-PD) является оптически активным, энантиомерно чистые препараты 1,2-пропандиола (1,2-PD) могут представлять особый интерес для медицинских, сельскохозяйственных или физиологических применений.
1,2-Пропандиол (1,2-PD) может вырабатываться микроорганизмами из сахаров как субстратов и единственного источника углерода. В последние годы другие субстраты, такие как глицерин, привлекают значительное внимание для применения в качестве субстрата для ферментации вместо, например, источников углерода сахаров. Интерес к глицерину, по существу, является результатом возросшего производства биодизеля или биоэтанола. Оба способа генерируют глицерин как основной побочный продукт, который составляет до, например, 10% (мас./мас.) производимого дизельного биотоплива.
Настоящее изобретение теперь относится к улучшенному биотехнологическому способу получения 1,2-пропандиола (1,2-PD) из неспособного к брожению дешевого углеродного субстрата, в особенности неочищенного препарата глицерина, в связи с чем углеродный субстрат поддерживает выработку биомассы и в то же время служит в качестве субстрата для продуцирования 1,2-пропандиола (1,2-PD). Настоящее изобретение также относится к улучшенным штаммам микроорганизмов, которые специфически адаптированы к специфическим требованиям такой процедуры и поэтому специфически подходят для использования в способе по изобретению.
В предпочтительном воплощении настоящее изобретение относится к биоконверсии глицерина или неочищенных препаратов глицерина как неспособного к брожению источника углерода непосредственно в 1,2-пропандиол с использованием клетки-хозяина, в особенности микроорганизма или штамма, которые созданы методами генной инженерии как содержащие один или несколько генов, которые вовлечены в путь выработки 1,2-пропандиола (1,2-PD) из глицерина. В частности, клетка-хозяин, в особенности микроорганизм или штамм по изобретению создают методами генной инженерии рекомбинантных ДНК для получения рекомбинантной клетки-хозяина, в особенности рекомбинантного микроорганизма или штамма, включающих гены, вовлеченные в метаболизм дигидроксиацетонфосфата и метилглиоксаля - двух основных соединений-предшественников на пути выработки 1,2-пропандиола (1,2-PD). В частности, клетка-хозяин, в особенности микроорганизм или штамм, обеспечиваются укрывающими генами, выбранными из группы, состоящей из генов, кодирующих ферменты, проявляющие активность глицеролдегидрогеназы, активность дигидроксиацетонкиназы, активность метилглиоксальсинтазы и активность пропандиолоксидоредуктазы, которые способны превращать глицерин в 1,2-пропандиол (1,2-PD) с высокой селективностью.
В предпочтительном воплощении в объеме настоящего изобретения используют созданный методами генной инженерии штамм Е.coli, в особенности рекомбинантный штамм Е.coli.
В настоящем изобретении неожиданно обнаружено, что обычный неочищенный глицерин (чистота 85%) с производства биодизеля можно использовать в качестве субстрата для выращивания самых разных организмов различного происхождения в аэробных (oxic) и анаэробных (anoxic) условиях. Можно было бы показать что большинство испытанных организмов не ослабляются неочищенным глицерином в сравнении с очищенным глицерином в отношении выработки биомассы, что указывает, что можно использовать неочищенный глицерин, и он вообще нетоксичен для микроорганизмов. На выработку биомассы примеси, обнаруживаемые в неочищенных препаратах глицерина, не влияют. Следовательно, неочищенный глицерин с производства биодизеля или из других источников может в равной степени хорошо использоваться микроорганизмами в качестве источника углерода и, таким образом, может использоваться без дополнительной обработки как общий обновляемый источник углерода в процессах ферментации для выработки биомассы.
Поэтому неочищенные препараты глицерина, в особенности неочищенный глицерин с производства биодизеля или биоэтанола, можно использовать в способе по настоящему изобретению для получения 1,2-пропандиола (1,2-PD).
Первым ключевым соединением-предшественником при получении 1,2-пропандиола (1,2-PD) является дигидроксиацетонфосфат (DHAP). DHAP превращается в метилглиоксаль - 2-е существенное исходное соединение-предшественник через активность метилглиоксальсинтазы (mgsA). Метилглиоксаль окончательно превращается в S-лактальдегид. Идентифицированная на сегодняшний день активность глицеролдегидратазы может превратить глицерин в R- или S-лактальдегид, который метаболизируется далее до R- или S-1,2-PD соответственно. В то время как предполагается, что эндогенная восстановительная активность клетки-хозяина вырабатывает R-1,2-PD, оказывается, что S-лактальдегид является субстратом для пропандиолоксидоредуктазы (fucO), которая может превратить S-лактальдегид в S-1,2-PD (Altaras N.E., 1999; Applied and Environmental Microbiology (65), 1180-1185).
Поэтому предполагается, что посредством введения пропандиолоксидоредуктазы (fucO) в организм-хозяин маневренность схемы получения 1,2-пропандиола (1,2-PD) может быть расширена за счет акцептирования S-энантиомера лактальдегида для конверсии в 1,2-пропандиол (1,2-PD). Кроме того, делается вывод, что активность пропандиолоксидоредуктазы (fucO) может быть необходима для выработки 1,2-пропандиола (1,2-PD) из глицерина независимо от пути метаболизма метилглиоксаля.
Соответственно, штамм дикого типа, который не вырабатывает детектируемых количеств 1,2-пропандиола (1,2-PD) из глицерина, независимо от условий культивирования, снабжают полинуклеотидом, включающим нуклеотидную последовательность, кодирующую активность пропандиолоксидоредуктазы (fucO).
В одном воплощении изобретения ген, кодирующий активность пропандиолоксидоредуктазы (fucO), клонируют в организм-хозяина, который не вырабатывает детектируемые количества 1,2-пропандиола (1,2-PD) из глицерина, и сверхэкспрессируют в указанном хозяине в минимальной среде, содержащей глицерин, в анаэробных и микроанаэробных (semi anoxic) условиях. Результатом сверхэкспрессии активности пропандиолоксидоредуктазы (fucO) является выработка 1,2-пропандиола (1,2-PD).
Другим ключевым соединением-предшественником на пути выработки 1,2-пропандиола (1,2-PD) является дигидроксиацетонфосфат (DHAP). В одном воплощении изобретения альтернативным путем для получения дигидроксиацетонфосфата как предшественника для синтеза 1,2-пропандиола (1,2-PD) является встраивание конструкции в штамм микроорганизма, в особенности в штамм Е.coli, и на таком пути продуцируется необходимый предшественник DHAP независимо от эндогенной регуляторной сети, действующей на глицеролфосфаткиназу (glpK).
В частности, молекулу ДНК, включающую нуклеотидную последовательность, кодирующую активность глицеролдегидрогеназы (gldA) и дигидроксиацетонкиназы (dhaK), вводят в организм-хозяина, в особенности в хозяина Е.coli. Ген, кодирующий глицеролдегидрогеназу (gldA), можно выделить из штамма Е.coli, в особенности Е.coli K12, и клонировать в подходящую плазмиду.
В одном воплощении ген, кодирующий глицеролдегидрогеназу (gldA), можно клонировать в подходящую плазмиду вместе с геном, кодирующим активность дигидроксиацетонкиназы (dhaK). Ген, кодирующий активность дигидроксиацетонкиназы (dhaK), можно выделить из штамма Citrobacter, в особенности штамма Citrobacter freundii.
В другом воплощении изобретения ген глицеролдегидрогеназы (gldA) клонируют в подходящую плазмиду независимо от и отдельно от глицеролдегидрогеназы (gldA).
В одном воплощении изобретения введенные кодирующие последовательности, кодирующие активность глицеролдегидрогеназы (gldA) и/или дигидроксиацетонкиназы (dhaK), находятся под контролем индуцируемого промотора, в особенности арабинозаиндуцируемого промотора (paraBAD).
Гены такого альтернативного пути для получения дигидроксиацетонфосфата, кодирующие активность глицеролдегидрогеназы (gldA) и дигидроксиацетонкиназы (dhaK), соответственно, можно ввести в организм-хозяина дикого типа вместе с полинуклеотидом, включающим нуклеотидную последовательность, кодирующую активность пропандиолоксидоредуктазы (fucO), или отдельно в виде отдельных экспрессирующих кассет, где кодирующая последовательность находится под контролем ее собственного промотора и сигнала терминации, когда указанные экспрессирующие кассеты могут располагаться или на разных плазмидах, или на одной плазмиде, или в форме синтетического оперона, включающего два или большее число указанных генов под контролем общих промоторной и концевой последовательностей.
В одном воплощении изобретения гены, кодирующие активность глицеролдегидрогеназы (gldA) и дигидроксиацетонкиназы (dhaK), клонируют в одну плазмиду, которая уже включает ген, кодирующий активность пропандиолоксидоредуктазы (fucO), и создают плазмиду, включающую ген, кодирующий активность глицеролдегидрогеназы (gldA) вместе с активностью пропандиолоксидоредуктазы (fucO), или плазмиду, включающую ген, кодирующий активность дигидроксиацетонкиназы (dhaK) вместе с активностью пропандиолоксидоредуктазы (fucO).
В одном воплощении создают плазмиду, включающую ген, кодирующий активность глицеролдегидрогеназы (gldA) и дигидроксиацетонкиназы (dhaK) вместе с активностью пропандиолоксидоредуктазы (fucO).
Различные последовательности генов, кодирующие различные ферментативные активности, можно расположить на плазмиде так, чтобы создать синтетический оперон, где два или большее число генов располагаются под контролем общих регуляторных последовательностей, включая последовательности промотора и полиаденилирования. В одном воплощении синтетический оперон находится под контролем индуцируемого промотора, в особенности арабинозаиндуцируемого промотора.
DHAP является начальным промежуточным соединением на пути генерирования 1,2-пропандиола (1,2-PD). Триозофосфатизомераза (tpi) гликолитического пути конкурирует с метилглиоксальсинтазой за DHAP. Для того чтобы управлять выработкой 1,2-пропандиола (1,2-PD), в синтетический оперон можно ввести mgsA, кодирующую метилглиоксальсинтазу, для того, чтобы сдвинуть равновесие в сторону образования 1,2-пропандиола (1,2-PD).
Поэтому в одном воплощении изобретения предоставляется расширенный синтетический оперон, включающий, кроме генов, вовлеченных в путь с дигидроксиацетонфосфатом, дополнительный ген, вовлеченный в продуцирование метилглиоксаля, в особенности ген метилглиоксальсинтазы, в особенности ген метилглиоксальсинтазы Е.coli.
Затем полученную(ые) плазмиду(ы) вводят в клетку-хозяина, в особенности в клетку-хозяина микроорганизма или штамм, которые неспособны вырабатывать детектируемые количества 1,2-пропандиола (1,2-PD) из глицерина независимо от условий культивирования, в особенности в штамм Е.coli.
В одном воплощении изобретения организм-хозяин не имеет активности метаболизма арабинозы или предварительно инактивирован по метаболизму арабинозы путем делеции или инактивации по меньшей мере одного из необходимых генов, вовлеченных в метаболизм арабинозы, такого как, например, ген, кодирующий активность рибулозокиназы (araB). Соответствующие штаммы культивируют в минимальной среде, содержащей глицерин, в аэробных и микроаэробных условиях, и 1,2-пропандиол: (1,2-PD) отделяют от супернатантов и анализируют.
Сверхэкспрессия гена пропандиолоксидоредуктазы (fucO) приводит к получению 1,2-пропандиола (1,2-PD) из препаратов неочищенного глицерина. Количества 1,2-пропандиола (1,2-PD) можно увеличить коэкспрессией дигидроксиацетонкиназы (dhaK) и/или глицеролдегидрогеназы (gldA) вместе с геном пропандиолоксидоредуктазы (fucO). Других улучшений можно достигнуть коэкспрессией гена дигидроксиацетонкиназы (dhaK), и/или гена глицеролдегидрогеназы (gldA), и/или гена метилглиоксальсинтазы (mgsA) вместе с геном пропандиолоксидоредуктазы (fucO) и/или используя клетки-хозяева, в особенности клетки-хозяева микроорганизма или штаммы, которые дефектны в по меньшей мере одном из непродуктивных путей, конкурирующих в отношении ключевых соединений-предшественников на пути выработки 1,2-пропандиола.
Расположение генов, вовлеченных в катализ, в описанном виде, таком как 5'-mgsA, gldA, dahK, fucO-3', является предпочтительным, однако изобретение не ограничивается таким специфическим расположением. Любой порядок описанных генов может быть подходящим для получения 1,2-пропандиола (1,2-PD).
Показан путь, специфический для получения изомера 1,2-пропандиола (1,2-PD) пропандиола, так как 1,3-пропандиол не обнаружен.
Способы получения нужных генов из бактериального генома являются обычными и хорошо известными в области молекулярной биологии. Например, если последовательность гена известна, можно создать подходящие геномные библиотеки расщеплением эндонуклеазами рестрикции и скринировать зондами, комплементарными к нужной последовательности гена. Как только последовательность выделена, можно амплифицировать ДНК с использованием стандартных методов амплификации, управляемой праймерами, такими как полимеразная цепная реакция (ПНР, PCR) (пат. США №4683202), и получить количества ДНК, подходящие для трансформации с использованием соответствующих векторов. С другой стороны, можно создать библиотеки космид, где крупные сегменты геномной ДНК (35-45 кб) могут быть упакованы в векторы и использованы для трансформации соответствующих хозяев. Космиды уникальны по способности вмещать большие количества ДНК. Как правило, космиды имеют по меньшей мере одну копию последовательности ДНК cos, которая необходима для упаковки и последующего образования кольцевой молекулы чужеродной ДНК. Кроме последовательности cos такие векторы также будут содержать начало репликации, такое как ColEl, и маркеры лекарственной устойчивости, такие как ген, резистентный к ампицилину или неомицину. Способы применения космид для трансформации подходящих хозяев-бактерий хорошо описаны в Sambrook et al. (1989). Типично для космид чужеродную ДНК изолируют и лигируют с использованием соответствующих эндонуклеаз рестрикции, соседних с участком cos космиды. Затем космиды, содержащие линеаризованную чужеродную ДНК, вводят во взаимодействие с упаковывающим ДНК-носителем, таким как бактериофаг 1. В процессе упаковки сайты cos расщепляются и чужеродная ДНК упаковывается в головную часть бактериальной вирусной частицы. Затем такие частицы можно использовать для трансфекции подходящих клеток-хозяев, таких как Е.coli. Как только чужеродная ДНК инъецирована в клетку, она образует кольцевую молекулу под влиянием «липких» концов cos. Таким образом большие сегменты чужеродной ДНК могут быть введены в рекомбинантные клетки-хозяева и экспрессированы в них.
Как только ген выделен и его последовательности установлены в опубликованный домен, ссылки, сделанные, например, на GenBank для указанных известных генов, могут использоваться специалистами в данной области техники для определения эквивалентных генов в других организмах, бактериальных штаммах, дрожжах, грибах, млекопитающих и растениях, и т.д. Обычную работу выгодно выполнять с использованием консенсусных последовательностей, которые можно определить с использованием выравниваний последовательностей с генами из других микроорганизмов и создавая упрощенные зонды, с помощью которых соответствующий ген можно клонировать в другой организм. Такие обычные методы молекулярной биологии известны в технике и описаны, например, в Sambrook et al.(1989).
В другом воплощении настоящее изобретение относится к различным векторам и трансформационным и экспрессирующим кассетам, подходящим для клонирования, трансформации и экспрессии ферментативных активностей по изобретению.
Указанный вектор может представлять собой, например, фаг, плазмиду, вирусный или ретровирусный вектор. Ретровирусные векторы могут быть репликативно компетентными или репликативно недостаточными. В последнем случае размножение вируса, как правило, будет происходить только в дополняющих клетках-хозяевах.
Полинуклеотиды или гены по изобретению могут соединяться с вектором, содержащим селектируемые маркеры для размножения в хозяине. Как правило, плазмиду вводят в преципитат, такой как кальцийфосфатный преципитат или рубидийхлоридный преципитат, или в комплекс с заряженным липидом или в кластеры на основе углерода, такие как фуллерены. Если вектор представляет собой вирус, он может быть упакован in vitro с использованием соответствующей пакующей клеточной линии перед применением к клеткам-хозяевам.
В более предпочтительном воплощении вектора по изобретению полинуклеотид оперативно связан с регулирующими экспрессию последовательностями, допускающими экспрессию в прокариотных или эукариотных клетках, или их изолированными фракциями.
Экспрессия указанного полинуклеотида включает транскрипцию полинуклеотида, предпочтительно, в транслируемую мРНК. Регуляторные элементы, обеспечивающие экспрессию в эукариотных клетках, таких как клетки бактерий или грибов, клетки насекомых, клетки животных, клетки млекопитающих или клетки человека, но в особенности клетки бактерий или грибов, хорошо известны специалистам в данной области техники. Они обычно включают регуляторные последовательности, обеспечивающие инициацию транскрипции, и, необязательно, сигналы поли-А, обеспечивающие терминацию транскрипции и стабилизацию транскрипта. Другие регуляторные элементы могут включать энахансеры транскрипции, а также трансляции. Возможные регуляторные элементы, допускающие экспрессию в прокариотных клетках-хозяевах, включают, например, промотор lac, trp или tac в Е.coli, и примерами регуляторных элементов, допускающих экспрессию в эукариотных клетках-хозяевах, являются промотор AOXI или GALI в дрожжах. Кроме элементов, которые ответственны за инициацию транскрипции, такие регуляторные элементы также могут включать сигналы терминации транскрипции в прямом направлении полинуклеотида.
В данном контексте подходящие экспрессирующие векторы являются векторами, известными в технике, такими как экспрессирующий вектор Окаяма-Берга pvDV1 (Pharmacia), pCDM8, pRc/CMV, pcDNA1, pcDNA3 (Invitrogene), pSPORT1 (GIBCO BRL), pSE380 (Invitrogene) или любой pBR322 или плазмиды, образованные pUC18. Предпочтительно указанный вектор является экспрессирующим вектором и/или переносчиком гена или нацеливающим вектором. Экспрессирующие векторы, полученные из вирусов, таких как ретровирусы, вирус осповакцины, аденоассоциированный вирус, вирус герпеса или вирус папилломы коров, могут быть использованы для доставки полинуклеотидов или вектора по изобретению в популяцию клеток-мишеней. Способы, которые хорошо известны специалистам в данной области техники, могут быть использованы для конструирования рекомбинантных вирусных векторов; см., например, методы, описанные в Sambrook (1989), и Ausubel (1994). С другой стороны, полинуклеотиды и векторы по изобретению для доставки в клетки-мишени могут быть воспроизведены в липосомы.
Настоящее изобретение также относится к клетке-хозяину, созданной методами генной инженерии с помощью полинуклеотида по изобретению, гена по изобретению или вектора по изобретению. Подходящие клетки-хозяева для рекомбинантного получения 1,2-пропандиола могут являться или прокариотными или эукариотными и будут ограничиваться только способностью клеток-хозяев экспрессировать активные ферменты.
Указанная клетка-хозяин может представлять собой прокариотную или эукариотную клетку. Полинуклеотид или вектор по изобретению, который присутствует в клетке-хозяине, может быть или интегрирован в геном клетки-хозяина или он может сохраняться вне хромосомы. В таком аспекте также следует иметь в виду, что настоящее изобретение относится к молекулам рекомбинантных ДНК, которые можно использовать для «нацеливания гена» и/или «замены гена», для восстановления мутантного гена или для создания мутантного гена через гомологичную рекомбинацию; см., например, Mouellic (1990); Joyner, Gene Targetting, A Practical Approach, Oxford University Press.
Клетка-хозяин может представлять собой прокариотную или эукариотную клетку, такую как бактериальная клетка или клетка гриба, клетка насекомого, клетка животного, клетка млекопитающего или клетка человека, но в особенности бактериальная клетка или клетка гриба. Предпочтительными хозяевами будут хозяева, типично применимые для получения глицерина или 1,2-пропандиола. Предпочтительными клетками грибов являются, например, клетки родов Aspergillus, Saccharomyces, Schizosaccharomyces, Pichia, Kluyveromyces, Candida, Hansenula, Debaryomyces, Mucor или Torulopsis, в частности, клетки вида S. cerevisiae. Термин «прокариотный» предназначен для включения всех бактерий и архебактерий, которые могут быть трансформированы или трансфицированы полинуклеотидом для экспрессии ферментативной активности по настоящему изобретению. Прокариотные хозяева могут включать грамотрицательные, а также грамположительные бактерии, такие как, например, Citrobacter, Enterobacter, Clostridium, Klebsiella, Aerobacter, Lactobacillus, Methylobacter, Escherichia, Salmonella, Streptomyces и Pseudomonas. Наиболее предпочтительными в настоящем изобретении являются Е.coli, S. typhimurium, Serratia marcescens и Bacillus subilis, вид Klebsiella и вид Saccharomyces, но в особенности вид Е.coli.
Его конкретные примеры включают Escherichia coli MG1655 (АТСС 700926; Bachmann, В., pp.2460-2488 in Neidhardt et al. 1996), Escherichia coli XL1-Blue MRF' [manufactured by Stratagene, Strategies, 5, 81 (1992)], Escherichia coli C600 [Genetics, 39, 440 (1954)], Escherichia coli Y1088 [Science, 222, 778 (1983)], Escherichia coli Y1090 [Science, 222, 778 (1983)], Escherichia coli NM522 [J. Mol. Biol., 166, 1 (1983)], Escherichia coli K802 [J. Mol. Biol, 16, 118 (1966)], Escherichia coli JM109 [Gene, 38, 275 (1985)], Escherichia coli DH5α [J. Mol. Biol, 166, 557 (1983)], и т.п.
Других улучшений в получении 1,2-PD можно достигнуть за счет использования подходящих мутантов микроорганизмов, в которых некоторые или все ферментативные активности, вовлеченные в непродуктивные пути, ослаблены или устранены. Например, ферментативные активности, кодированные mgsA и tpi, конкурируют за DHAP. Показано, что активность метилглиоксальсинтазы (MgsA) инактивируется дифосфатом (Hopper D.J., 1972). Для того чтобы улучшить конверсию DHAP в метилглиоксаль, можно идентифицировать нечувствительный к фосфату мутант MgsA скринингом вариантных библиотек, полученных любым способом, генерируя вариацию в кодирующих последовательностях mgsA, например, подверженной ошибкам ПЦР. Штаммы микроорганизмов, в особенности клоны Е.coli, предварительно инактивированные в триозофосфатизомеразе (tpiA) и эндогенной mgsA, трансформируют библиотеками плазмид вариантов mgsA и выращивают на неселективных плотных средах. Путем получения реплик начальных трансформантов (пластинка А) на агаровых пластинках, содержащих высокие концентрации глицерина или DHAP (пластинка В) или высокие концентрации дифосфата и глицерина или DHAP (пластинка С), отбирают клоны, которые могут расти на пластинке А и В, но не на пластинке С. Указанные клоны кодируют варианты mgsA с существенной активностью, которые вырабатывают токсичные уровни метилглиоксаля в присутствии высоких концентраций фосфата.
Мутанты триозофосфатизомеразы можно получить так, как описано для мутантов метилглиоксальсинтазы. Tpi-мутанты, которые существенно ослаблены в отношении кинетики роста, идентифицируют, сравнивая кинетику роста на комплексной среде (например, среде Луриа, LB), глюкозе и глицерине. Мутант, представляющий интерес, показывает отсутствие роста или меньший рост по сравнению с немодифицированным штаммом с глицерином как единственным источником углерода и энергии, в то время как кинетика роста с LB или глюкозой как источником углерода не изменяется.
Существуют два других основных пути детоксикации MG, которые являются продуктивными в смысле биосинтеза 1,2-PD. Как начальная стадия, они катализируются так называемыми MG-редуктазами. Предполагается, что некоторые ферменты кодируют такую активность, которая должна усиливаться инактивацией конкурирующих непродуктивных путей.
В одном воплощении изобретения конструируют мутантов микроорганизмов, в особенности мутантов Е.coli, в которых инактивированы один или несколько генов, кодирующих системы глиоксилазы I и II (gloA и gloB), лактатдегидрогеназу A (ldhA), систему глиоксилазы III (косвенно, путем инактивации регулятора хозяина rpoS) и альдегиддегидрогеназу (aldA), например, до достаточного уменьшения или полного ингибирования экспрессии функциональных ферментативных активностей через, например, нокауты отдельных генов. Таким путем можно получить микробный мутант, который имеет один или несколько вышеуказанных инактивированных генов, в особенности мутанта, в котором инактивированы 2, в особенности 3, в особенности 4, в особенности 5 генов, выбранных из группы, состоящей из генов, кодирующих систему глиоксилазы I (gloA), систему глиоксилазы II (gloB), лактатдегидрогеназу А (ldhA), систему глиоксилазы III (косвенно, путем инактивации регулятора хозяина rpoS) и альдегиддегидрогеназу (aldA).
В одном воплощении изобретение относится к мутанту микроорганизма, в особенности мутанту Е.coli, в котором полностью или частично инактивирован ген, кодирующий активность gloA.
В одном воплощении настоящего изобретения клетка-хозяин, в особенности микроорганизм или штамм, в особенности прокариотный микроорганизм, например Е.coli, инактивирован в отношении своей способности метаболизировать метилглиоксаль (MG) в D- и/или L-лактат (метаболизм MG-лактат). Этого достигают, например, инактивацией глиоксилазы А, предпочтительно в сочетании с инактивацией одного или нескольких генов, кодирующих глиоксилазу В, альтернативный сигма-фактор rpoS и альдегидредуктазу А. В предпочтительном воплощении штамм является дефицитным по всем перечисленным выше активностям и/или генам.
В другом воплощении штамм, инактивированный по метаболизму арабинозы и метаболизму MG-лактат, например, инактивацией глиоксилазы А, трансформируют с помощью полинуклеотида, включающего гены, кодирующие ферментативную активность, выбранную из группы, состоящей из активности метилглиоксальсинтазы (mgsA), глицеролдегидрогеназы (gldA), дигидроксиацетонкиназы (dhaK) и пропандиолоксидоредуктазы (fucO), в особенности плазмидой pDP_mgdf, в то время как расположение генов в синтетическом опероне не ограничивается расположением, показанным в плазмиде pDP_mgdf, но может иметь другой порядок. Изобретение также относится к штамму, не являющемуся неспособным к метаболизму арабинозы, например, Е.coli дикого типа.
В другом воплощении штамм, предпочтительно, но не необходимо, инактивированный по метаболизму MG-лактат, трансформируют кодированными плазмидами генами, которые придают альдокеторедуктазную активность. Активность, кодированную генами, берут из группы генов Е.coli, включающих dkgA, dkgB, уеаЕ и yghZ.
В предпочтительном воплощении штамм, экспрессирующий глицеролдегидрогеназу (gldA), пропандиолоксидоредуктазу (fucO) и метилглиоксальсинтазу (например, с помощью плазмиды pDP_mgdf), трансформируют плазмидой, кодирующей активность альдокеторедуктазы (например, DkgA Е.coli) и культивируют в среде, содержащей неочищенный глицерин как источник углерода.
В особенно предпочтительном воплощении штамм, инактивированный по метаболизму MG-лактат и экспрессирующий глицеролдегидрогеназу (gldA), дигидроксиацетонкиназу, пропандиолоксидоредуктазу (fucO) и метилглиоксальсинтазу (например, кодированный на плазмиде pDP_mgdf), экспрессирует активность альдокеторедуктазы (например, dkgA Е.coli, кодированную на плазмиде pCR2.1) и культивируется в среде, содержащей глицерин как источник углерода.
Изобретение также относится к штамму, не являющемуся неспособным по метаболизму арабинозы, и/или к штаммам, экспрессирующим релевантные ферментативные активности, указанные в данном изобретении, из хромосомы микроорганизма.
Полинуклеотид, кодирующий ферментативную активность по настоящему изобретению, можно использовать для трансформации или трансфекции клетки-хозяина с использованием любых методов, обычно известных специалистам в данной области техники.
Методом, преимущественно используемым для введения указанных генов в штамм, является электропорация, которая хорошо известна специалистам в данной области техники. Вкратце протокол электропорации может быть следующим: гетерологичные гены, представляющие интерес, клонируют в экспрессирующий вектор между промотором и терминатором. Такой вектор также обладает геном устойчивости к антибиотикам для отбора клеток, которые его содержат, и точкой начала репликации (ori) в штамме-хозяине, так что он может быть сохранен. Протокол требует получения электрокомпетентных клеток-хозяев, которые затем конвертируют электропорацией вектором. Согласно изобретению гены, введенные электропорацией, преимущественно представляют собой гены по изобретению, кодирующие ферментативную активность, выбранную из группы, состоящей из активности глицеролдегидрогеназы (gldA), дигидрокеиацетонкиназы (dhaK), метилглиоксальсинтазы (mgsA) и пропандиолоксидоредуктазы (fucO).
Способы получения слитых функционально связанных генов и их экспрессии в клетках бактерий или животных, хорошо известны в технике (Sambrook, цитировано выше). Генетические конструкции и способы, описанные в данном описании, можно использовать для экспрессии полипептидов по изобретению, например, в прокариотных хозяевах. Вообще, экспрессирующие векторы, содержащие промоторные последовательности, которые облегчают эффективную транскрипцию встроенного полинуклеотида, используют в связи с хозяином. Экспрессирующий вектор типично содержит точку начала репликации, промотор и терминатор, а также специфические гены, которые способны обеспечивать фенотипический отбор трансформированных клеток. Трансформированных прокариотных хозяев можно выращивать в ферментерах и культивировать согласно методам, известным в технике для достижения оптимального роста клеток.
Типично клетки выращивают при 30°С в соответствующих средах. Предпочтительными средами для выращивания в настоящем изобретении являются определенные или синтетические среды, например минимальная среда М9, содержащая глицерин в качестве источника углерода. Обычные коммерчески полученные среды, такие как среда Луриа Бертани (LB), декстрозный бульон Sabouraud или дрожжевой солодовый экстракт (YM), также можно использовать, и соответствующая среда для выращивания определенных микроорганизмов будет известна специалистам в области микробиологии или ферментации. Использование известных агентов для прямой или косвенной модуляции катаболитной репрессии, например циклического аденозин-2':3'-монофосфата или циклического аденозин-2':5'-монофосфата, также может включаться в реакционные среды. Подобным образом, известные агенты для модуляции ферментативных активностей (например, сульфиты, бисульфиты и щелочи), что ведет к усилению продуцирования 1,2-PD, можно применять в сочетании с или в качестве альтернативы генетическим манипуляциям.
Подходящие интервалы рН для ферментации находятся между рН 5,0 и рН 9,0, где рН 6,0 - рН 8,0 являются предпочтительными как интервал для начальных условий. Взаимодействия можно осуществлять в аэробных, микроаэробных или анаэробных условиях, из которых микроаэробные условия являются предпочтительными.
Периодическая и непрерывная ферментации.
В способе по настоящему изобретению используют периодический способ ферментации. Классическая периодическая ферментация представляет собой закрытую систему, где состав сред устанавливают в начале ферментации и не подвергают искусственным изменениям во время ферментации. Таким образом, в начале ферментации среды инокулируют нужным организмом или организмами, и ферментации позволяют происходить без добавления чего-либо в систему. Однако типично периодическая ферментация является «периодической» в отношении добавления источника углерода, и часто берут пробы на регулирующие факторы, такие как рН и концентрация кислорода. Составы метаболитов и биомассы в периодической системе постоянно изменяются до тех пор, пока не закончится время ферментации. В периодических культурах клетки проходят через статическую лог-фазу до лог-фазы сильного роста и наконец до стационарной фазы роста, где скорость роста уменьшается или рост прекращается. Без обработки клетки в стационарной фазе будут в конечном счете погибать. Клетки в лог-фазе, как правило, ответственны за объем выработки конечного или промежуточного продукта. Вариантом стандартной периодической системы является периодическая система ферментации с подпиткой (Fed-Batch), которая также подходит для настоящего изобретения. В таком варианте типичной периодической системы субстрат добавляют поэтапно по мере продвижения ферментации. Периодические системы с подпиткой применимы, когда катаболитная репрессия склонна к ингибированию метаболизма клеток и когда желательно иметь ограниченные количества субстрата в средах. Измерение фактической концентрации субстрата в системах с подпиткой является трудным и поэтому оценку осуществляют на основании изменений факторов, поддающихся измерению, таких как рН, растворенный кислород и парциальное давление отходящих газов, таких как СО2. Периодическая ферментация и ферментация с подпиткой являются обычными и хорошо известны в технике, и их примеры можно найти в Brock, цитировано выше. Также предполагается, что способ должен адаптироваться к способам непрерывной ферментации. Непрерывная ферментация представляет собой открытую систему, в которой во время переработки определенные сбраживаемые среды добавляют непрерывно в биореактор и одновременно удаляют равное количество кондиционированных сред. Непрерывная ферментация, как правило, сохраняет культуры при постоянной высокой плотности, где клетки находятся, главным образом, в лог-фазе роста. Непрерывная ферментация допускает модуляцию одного фактора или любого числа факторов, которые влияют на рост клеток или концентрацию конечного продукта. Например, в одном способе будет сохраняться ограничение питания, например, как фиксированные скорость источника углерода или уровень азота, и все другие параметры снижаются. В других системах может непрерывно изменяться ряд факторов, влияющих на рост, в то время как концентрация клеток, измеряемая по мутности среды, поддерживается постоянной. В непрерывных системах стараются поддерживать неизменные условия выращивания, и, таким образом, клетки теряются из-за того, что при ферментации выпуск среды должен быть сбалансирован со скоростью роста клеток. Способы модуляции питательных веществ и факторов роста для процессов непрерывной ферментации, а также методы максимизации скорости образования продукта хорошо известны в области промышленной микробиологии, и ряд способов подробно описан, например, в Brock, цитировано выше. Настоящее изобретение может осуществляться на практике с использованием или периодических с подпиткой или непрерывных способов, и любой известный способ ферментации может быть подходящим. Кроме того, предполагается, что клетки могут быть иммобилизованы на субстрате вообще как клеточные катализаторы и подвергаться воздействию в условиях ферментации для получения 1,2-пропандиола.
Затем продукт 1,2-пропандиол можно отделить от среды для выращивания или клеточных лизатов. Отделение и очистку произведенных микробиологически или иначе пропандиолов можно осуществлять любыми обычными средствами. Способы очистки пропандиолов от сбраживаемых или культуральных сред известны в технике. Например, пропандиолы можно получить из клеточных сред, подвергая реакционную смесь экстракции органическим растворителем, перегонке и колоночной хроматографии (патент США №5356812). Особенно хорошим органическим растворителем для такого способа является циклогексан (патент США №5008473).
Для промышленных применений очистка 1,2-пропандиола от больших объемов культуральной жидкости требует способов нелабораторного масштаба. Трудности, с которым борются, включают удаление клеточного вещества, образующего среду (очистка), концентрирование 1,2-пропандиола или экстракцией или удалением воды и отделение остальных примесей от частично очищенного мономера. Очистка культуральной жидкости типично будет производиться фильтрацией, центрифугированием или проточной микрофильтрацией. Подходящие фильтры производят, например, Millipore (Millipore Corporation, 80 Ashby Road, Bedford, Mass.) или Filmtec (Dow Chemical Co.). Центрифугирование эффективно удаляет массу клеток, но, в зависимости от характера культуральной жидкости, полное удаление клеток достигается не всегда. Проточная микрофильтрация дает исключительно чистый фильтрат. Концентрат представляет собой взвесь, а не остаток на фильтре с высоким содержанием твердого вещества. Специалист сможет подобрать способ очистки, наиболее соответствующий используемым аппарату для ферментации и условиям ферментации. Снижение содержания воды в культуральной жидкости осложняется высокой растворимостью 1,2-пропандиола в воде. Экстракцию 1,2-пропандиола из очищенной культуральной жидкости можно осуществлять различными способами, включая выпаривание/перегонку, мембранную технологию, экстракцию органическим растворителем и адсорбцию. Сначала можно использовать роторные испарители для уменьшения объема воды в культуральной жидкости. Такой способ имеет особый успех у заявителей. Оказывается, что осаждение посторонних белков и солей не влияет на извлечение 1,2-пропандиола. Мембранную технологию можно использовать или отдельно или в сочетании с выпариванием. Подходящие мембраны будут или (i) пропускать 1,2-пропандиол, задерживая воду и другие молекулы загрузки, или (ii) пропускать воду и другие молекулы загрузки, задерживая 1,2-пропандиол, или (iii) пропускать воду и 1,2-пропандиол, в то время как другие молекулы задерживаются. В настоящем изобретении предпочтителен способ (iii). Особенно применимы осмотические мембраны, такие как SW-30 2540 (Filmtec, Dow Chemical Co.) и обратноосмотические мембраны серий DL и SH, изготовленные Millipore (Millipore Corporation, Bedford, Mass.). После выпаривания и мембранной концентрации частично очищенный 1,2-пропандиол можно экстрагировать органическими растворителями. Подходящие растворители будут включать спирты, такие как трет-амиловый спирт, циклопентанол, октанол, пропанол, метанол и этанол. Также можно использовать неспиртовые растворители, такие как октанон, циклогексан и валериановый альдегид. В контексте настоящего изобретения спирты являются предпочтительными, и наиболее предпочтителен этанол. С другой стороны, 1,2-пропандиол можно дополнительно сконцентрировать адсорбцией на различных промышленных адсорбентах. Особенно подходящими являются активированный углерод и полициклодекстрин, такие как производимые American Maize Products Company. После или экстракции или адсорбции частично очищенный 1,2-пропандиол необходимо рафинировать. Рафинирование можно осуществить электродиализом (в особенности применимым для обессоливания), в котором используют сочетание анионо- и катионообменных мембран или биполярных (анионных и катионных) мембран (см., например, Grandison, Alistair S. (1996)). Предпочтительным способом рафинирования в настоящем изобретении является перегонка. Перегонку можно осуществлять периодически, когда рабочее давление равно окружающему или ниже, например в вакууме примерно 85000 кПа (635 мм рт. ст., 25 дюймов). Контроль за перегонкой показывает, что материалы испаряются в порядке от первого до последнего материала, начиная с легких органических веществ, затем вода, диолы, включая 1,2-пропандиол, и наконец тяжелые вещества, такие как глицерин, и твердые вещества выпадают в осадок.
Сведения, подтверждающие возможность осуществления изобретения
Приведенные далее примеры представляют пояснительные воплощения. С учетом раскрытия настоящего изобретения и общего уровня специалистов в данной области техники специалисты будут иметь в виду, что приведенные далее примеры предназначены быть только примерами и что можно использовать многочисленные замены, модификации и изменения без отхода от объема заявляемого в данном случае предмета.
Все манипуляции и методы, необходимые для конструирования и размножения штаммов, описанных в данном описании, известны специалистам в данной области техники. Технические подробности описаны, например, в Ausubel et al., 1995; Sambrook J., 2001, и Miller J.H., 1992, и в соответствующих публикациях, цитированных в данном описании.
Пример 1. Общие методы
1.1. Культивирование штаммов
Е.coli культивируют в определенной минимальной среде, которую создают как содержащую низкие уровни фосфата, так как фосфат является известным ингибитором метилглиоксальсинтазы. Среда содержит, на литр,
(NH4)2SO4 - 3 г,
дрожжевой экстракт - 0,2 г,
CoCl2 - 1,9е-6 г,
бис(2-гидроксиэтил)иминотрис(гидроксиметил)метан - 10 г,
KH2PO4 - 0,002 г,
K2HPO4 - 0,0085 г,
MgSO4 - 0,225 г,
раствор микроэлементов [Pfennig, 1966] - 1 мл.
В соответствующем случае в среду добавляют антибиотики. Используют концентрации 5 мкг/л гентамицина, 10 мкг/л ампицилина.
Неочищенный глицерин получают с производства дизельного биотоплива, и его чистота 85%.
Штаммы Е.coli размножают обычным способом в культуральных пробирках (общий объем 30 мл; инокулят 5 мл) или стеклянных пузырьках, плотно закрывающихся резиновыми пробками (общий объем 12 мл, инокулят 10 мл) для создания микроанаэробных условий. Термин «культивирование в аэробных условиях» подразумевает культивирование в отсутствие герметичности при перемешивании. «Микроанаэробные условия» в данном контексте означает культивирование штаммов без перемешивания в среде, которую получают в аэробных условиях и в условиях герметичности, например пузырьках, закрытых резиновыми пробками после инокуляции для того, чтобы избежать диффузии кислорода извне. Время культивирования изменяется от 2 до 5 суток в аэробных и микроанаэробных условиях соответственно. Вообще, эксперименты прекращают, когда оптическая плотность прекращает повышаться далее.
1.2. Анализ образования 1,2-PD
Уровни 1,2-PD в супернатантах в культуральной среде определяют тремя различными способами, включающими колориметрический анализ, ВЭЖХ и ГХ-МС. В то время как ВЭЖХ с использованием катионообменной колонки не позволяет различить 1,2- и 1,3-изомеры пропандиола, колориметрический анализ специфичен для 1,2-PD. Анализ методом ГХ-МС допускает одновременное количественное определение обоих изомеров по отдельности. Уровни детекции составляют 0,5 г/л для анализа:методом ВЭЖХ, 50 мг/л для колориметрического анализа и 10 мг/л для анализа методом ГХ-МС.
В случае рядового анализа 1,2-PD в супернатантах определяют колориметрическим методом, описанным Jones and Riddick {Jones, 1957}. Сначала к образцу бесклеточного супернатанта добавляют серную кислоту, перемешивают и нагревают. Затем добавляют раствор нингидрина и бисульфит натрия, перемешивают и инкубируют в течение одного часа. Добавляют еще одну аликвоту серной кислоты, и регистрируют поглощение при 595 нм, которое эквивалентно концентрации 1,2-PD.
В случае количественного анализа 1,2-PD в образцах определяют анализом методом ГХ-МС. 1,2-PD идентифицируют по идентичным временам удерживания при сравнении с аутентичным материалом и с помощью масс-фрингопринтинга.
Пример 2. Конструирование рекомбинантных организмов
2.1. Штаммы и пдазмиды, используемые в изобретении
В качестве хозяина для выработки 1,2-PD используют Е.coli MG1655 (F-, лямбда-ilvG-rfb-50 rph-1) и DH5-альфа (F-, φ80dlacZΔM15, Δ(lacZYA-argF), U169, deoR, recA1, hsdR17(rk-, mk+), phoA, supE44, λ-, thi-1, gyrA96, relA1) и их производные. Кроме того, геномная ДНК MG1655 обеспечивает источник для амплификации релевантных генов. Геномную ДНК из Citrobacter freundii (DSM30040) используют в качестве матрицы для амплификации гена dhaK.
2.2. Выделение и клонирование генов
Как первую стадию, ген Е.coli для глицеролдегидрогеназы gldA (SEQ ID NO:29) вводят в плазмиде pB2araJ (фиг.3а) или плазмиде pCR2.1 (фиг.3b). Все праймеры, используемые для амплификации генов, представляющих интерес, перечислены в таблице 1. Праймеры gldH_for1 и gldH_rev1 используют для амплификации фрагмента gldA в 1104 п.о. из Е.coli. Очищенный гель-фильтрацией фрагмент ПЦР встраивают в сайт AatII-SwaI вектора pB2araJ, и получают pDP_g. В такой плазмиде экспрессия gldA находится под контролем промотора paraBAD, создавая возможность строго регулированной индуцируемой экспрессии с помощью L-арабинозы, как описано Guzman et al. (Guzman L.M., 1995). Ген dhaK амплифицируют из С.freundii с использованием праймеров dhaK_for1 и dhaK_rev1. Полученная последовательность в 1659 нукл. показана в SEQ ID NO:27, соответствующая белковая последовательность в SEQ ID NO:30. Очищенный гель-фильтрацией фрагмент dhaK встраивают в сайт SwaI-AscI pDP_g, получая pDP_gd, которую котранскрибируют как с gldA, так и с dhaK. Праймеры fucO_for1 и fucO_rev1 используют для амплификации гена в 1152 нукл., кодирующего пропандиолоксидоредуктазу (fucO) из Е.coli (SEQ ID NO:31), который лигируют в сайт AvrII-SmaI pDP_gd, и получают плазмиду pDP_gdf, котранскрибируя gldA, dhaK и fucO. Фрагмент mgsA в 468 и.о., кодирующий метилглиоксальсинтазу Е.coli, показанный в SEQ ID NO:32, амплифицируют из Е.coli с использованием праймеров mgsA_xhoI_for и mgsA_xhoI_rev и вводят в смысловой ориентации в сайт XhoI и получают pDP_mgdf. Последовательность транскрибированных генов после индукции из плазмиды pDP_mgdf является следующей: mgsA, gldA, dhaK, fucO и lacZ-альфа, в то время как оставшийся пептид lacZ-альфа используют только для транскрипционных исследований в подходящем штамме-хозяине (например, DHA5-альфа). Нуклеотидная последовательность всей плазмиды pDP_mgdf показана в SEQ ID NO:28.
Гены dkgA и dkgB, кодирующие многофункциональную MG-редуктазу (SEQ ID NO:33) и 4-нитробензальдегидредуктазу (SEQ ID NO:34), соответственно, амплифицируют из Е.coli с использованием Taq-полимеразы и праймеров dkgB_up и dkg_dw или dkgA_up и dkgA_dw. Очищенные продукты ПЦР вводят в вектор pCR2.1 (фиг.3b) ТА-клонированием (Invitrogen).
2.3. Делеция активностей, кодированных gloA, gloB, rpoS, aldA, IdhA, в штаммах-хозяевах E.coli
Специалистам в данной области техники известен ряд методов специфической делеции генов. Такие методы включают, но не ограничиваются перечисленным, разрыв генов модифицированными интронами группы II (Karberg M., 2001), инактивацию генов, опосредуемую фагорекомбиназой, с использованием ПЦР-амплифицированной ДНК (Dastenko K.A., 2000) (Ellis E.H., 2001) (Yu D., 2000) (Marx and Lidstrom, 2002) и введение линейной двухцепочечной ДНК, гомологичной гену, представляющему интерес, в клетки-хозяев (Cunnibgham R.P. et al. (1980)).
Методом, предпочтительно используемым для введения таких генов в штамм, является электропорация, которая хорошо известна специалистам в данной области техники.
В данном изобретении используют гомологичную рекомбинацию ПЦР-амплифицированной ДНК, несущей гены селектируемых маркеров. Праймеры создают специфически для генов, представляющих интерес. Последовательности праймеров приводятся ниже. Успешную делецию гена, представляющего интерес, подтверждают ПЦР-анализом и секвенированием ДНК.
Обозначение делегированных генов (1-6) и праймеров, используемых для получения гомологичной линейной ДНК
1) Название: субъединица альдегиддегидрогеназы А
Ген: aldA
Инвентарный номер: Ecogene, EG10035
Хромосомная локализация: 1486256=>1487695
Праймер 1:
AACAATGTATTCACCGAAAACAAACATATAAATCACAGGAGTCGCCCATG
(SEQ ID NO:15)
Праймер 2:
GAGGAAAAAACCTCCGCCTCTTTCACTCATTAAGACTGTAAATAAACCAC
(SEQ ID NO:16)
2) Название: D-лактатдегидрогеназа
Ген: ldhA
Инвентарный номер: Ecogene, EG13186
Хромосомная локализация: 1440867=>1439878
Праймер 1:
CTCCCCTGGAATGCAGGGGAGCGGCAAGATTAAACCAGTTCGTTCGGGCA
(SEQ ID NO:17)
Праймер 2:
TATTTTTAGTAGCTTAAATGTGATTCAACATCACTGGAGAAAGTCTTATG
(SEQ ID NO:18)
3) Название: РНК-полимераза, фактор сигма S (сигма 38)
Ген: rpoS
Инвентарный номер: Ecogene, EG10510
Хромосомная локализация: 2865573=>2864581
Праймер 1:
TGAGACTGGCCTTTCTGACAGATGCTTACTTACTCGCGGAACAGCGCTTC
(SEQ ID NO:19)
Праймер 2:
CTTTTGCTTGAATGTTCCGTCAAGGGATCACGGGTAGGAGCCACCTTATG
(SEQ ID NO:20)
4) Название: глиоксилаза I
Ген: gloA
Инвентарный номер: Ecogene, EG13421
Хромосомная локализация: 1725861=>1726268
Праймер 1:
TACTAAAACAACATTTTGAATCTGTTAGCCATTTTGAGGATAAAAAGATG
(SEQ ID NO:21)
Праймер 2:
GGCGCGATGAGTTCACGCCCGGCAGGAGATTAGTTGCCCAGACCGCGACC
(SEQ ID NO:22)
5) Название: глиоксилаза II
Ген: gloB
Инвентарный номер: Ecogene, EG13330
Хромосомная локализация: 234782=>234027
Праймер 1:
CGAACGGAGCCGATGACAAGAAAGTTTTATCAGAACCTATCTTTCTTTGA
(SEQ ID NO:23)
Праймер 2:
CTTGCCGGTTTCATCACAACCTTCCGTTTCACACTGAGAGGTAATCTATG
(SEQ ID NO:24)
6) Название: мономер L-рибулокиназы
Ген: araB
Инвентарный номер: Ecogene, EG10053
Хромосомная локализация: 70048=>68348
Праймер 1:
AATTATCAAAAATCGTCATTATCGTGTCCTTATAGAGTCGCAACGGCCTG
(SEQ ID NO:25)
Праймер 2:
ACTCTCTACTGTTTCTCCATACCCGTTTTTTTGGATGGAGTGAAACGATG
(SEQ ID NO:26)
Пример 3. Получение 1,2-PD
3.1. Культивирование микроорганизмов в минимальной среде с неочищенным глицерином
Исследуют утилизацию различными микроорганизмами препаратов неочищенного глицерина (чистота примерно 85%) по сравнению с по существу чистыми препаратами глицерина (чистота >99%). Широкий ряд микроорганизмов, представляющих различные таксоны, вместе с Е.coli MG1655 выращивают при нерегулируемом рН в минимальной среде с добавлением различных количеств чистого и неочищенного глицерина в аэробных условиях. Табл. 2 и 3 показывают, что не наблюдается ингибирующего действия на выработку биомассы, когда единственным источником углерода и энергии вместо чистого глицерина является неочищенный препарат глицерина. Кроме того, когда неочищенный препарат глицерина служит в качестве источника углерода и энергии, количество фосфата в определенной минимальной среде можно уменьшить на 70% (табл.4).
Выработка биомассы, поддерживаемая неочищенными препаратами глицерина. Проводят сравнение выработки биомассы, когда в качестве источника углерода для выращивания в аэробных и анаэробных условиях служат чистые (чистота >99%) или неочищенные препараты глицерина (10 г/л). Штаммы, представляющие различные таксоны, выделенные из образцов окружающей среды, культивируют в микропланшетах без перемешивания в определенных атмосферных условиях.
Сравнение выработки биомассы Е.coli MG1655, полученной с помощью неочищенных или чистых препаратов глицерина. Проводят сравнение выработки биомассы, когда в качестве источника углерода для выращивания в аэробных условиях при нерегулируемом рН служат чистые (чистота >99%) или неочищенные препараты глицерина. Штаммы культивируют в культуральных пробирках в аэробных условиях, ⌀ средний; ср.-кв.откл. - среднеквадратическое отклонение.
Потенциал примесей, присутствующих в неочищенном глицерине (10 г/л) для замены макроэлементов минимальной среды. Проводят сравнение выработки биомассы Е.coli MG1655, когда в качестве источника углерода и макроэлементов для выращивания в условиях окисления при нерегулируемом рН служат чистые (чистота >99%) или неочищенные препараты глицерина. ⌀ средний; ср.-кв.откл. - среднеквадратическое отклонение.
3.2. Строение функционального пути, дающего дигидроксиацетонфосфат, в обход глиперодкиназы и глицеральдегид-3-фосфатдегидрогеназы
Гены gldA из Е.coli, кодирующий глицеролдегидратазу, dhaK из Citrobacter freundii, кодирующий дигидроксиацетонкиназу (dhaK), и fucO из Е.coli, кодирующий пропандиолоксидоредуктазу, клонируют в плазмиде pB2araJ, и получают плазмиду pDP_gdf. Плазмида pDP_mgdf дополнительно содержит метилглиоксальсинтазу из Е.coli. Согласно известным биохимическим путям, пропандиолоксидоредуктаза нерелевантна для такого примера. Однако плазмиду pDP_gdf и ее производное pDP_mgdf испытывают на комплементацию нокаута glpK, так как указанные плазмиды релевантны в отношении продуцирования рекомбинантного 1,2-PD.
Анализ показывает, что присутствие плазмиды pDP_gdf или pDP_mgdf облегчает ингибирование роста в glpK-мутанте Е.coli, когда глицерин является единственным источником углерода (табл.5). Таким образом, устанавливается индуцируемый нерегулируемый путь, независимый от эндогенной доставки для метаболизма глицерина. Окончательно полученная биомасса равна или даже больше по сравнению с ростом контрольного штамма (DH5-альфа□araB) или когда субстратом для роста является глюкоза.
Нокаут araB- Е.coli DH5-альфа и его производные культивируют в минимальной среде, содержащей в качестве источника углерода глюкозу или глицерин в концентрации 10 г/л. Различные штаммы культивируют при 37°С в культуральных пробирках в аэробных условиях в течение 5 суток. Биомассу определяют в конце эксперимента как оптическую плотность при 580 нм. Ср.-кв.откл. - среднеквадратическое отклонение.
3.3. Определение минимального набора генов, необходимых для получения рекомбинантного 1,2-РР из глицерина
Мутант araB DH5-альфа Е.coli трансформируют различными плазмидами, содержащими гены, представляющие интерес, перечисленные в таблице 6. Штамм дикого типа (контроль) и рекомбинантные штаммы культивируют в аэробных условиях в минимальной среде, содержащей 10 г/л неочищенного глицерина, в течение 3-5 суток до тех пор, пока штаммы не войдут в стационарную фазу. Супернатанты анализируют анализом ГХ-МС на содержание 1,2-PD. Результаты приводятся в таблице 6.
Содержание 1,2-PD, определенное в супернатантах после культивирования штаммов Е.coli в минимальной среде, содержащей 10 г/л неочищенного глицерина.
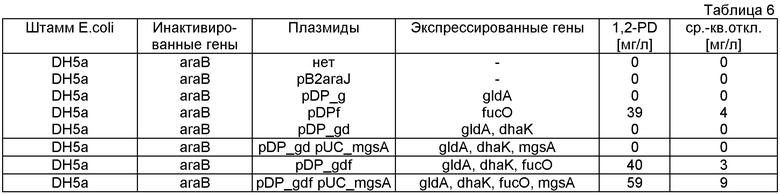
3.4. Исключительное продуцирование 1,2-изомера пропандиола рекомбинантными штаммами Е.coli, экспрессирующими глицеролдегидрогеназу, дигидроксиацетонкиназу, метилглиоксальсинтазу и пропандиолоксидоредуктазу, кодированные в синтетическом опероне
Е.coli MG1655□araB трансформируют плазмидой pDP_mgdf или без нее и культивируют в минимальной среде, содержащей 10 или 15 г/л неочищенного глицерина как источника углерода. Инкубацию проводят в аэробных условиях в культуральных пробирках или в плотно закрытых склянках без перемешивания (микроанаэробные условия). Е.coli MG1655□araB без плазмиды является контрольным штаммом. Период инкубации составляет 5 суток, температура инкубации 37°С.
В конце эксперимента определяют рост как оптическую плотность (OD580) и определяют уровни 1,2- или 1,3-PD анализом методом ГХ-МС. Анализы проводят двукратно. Результаты (табл.7) показывают успешное введение созданного методами генной инженерии пути, что позволяет Е.coli продуцировать 1,2-PD из неочищенного глицерина, из которого вырабатывается исключительно 1,2-изомер пропандиола.
Штаммы нокаута araB Е.coli MG1655 трансформируют с/без плазмиды pDP_mgdf и культивируют в минимальной среде, содержащей неочищенный глицерин как источник углерода в концентрации 10 или 15 г/л. Культивирование проводят при 37°С в аэробных или микроанаэробных условиях. В конце эксперимента (5 суток) определяют биомассу как оптическую плотность при 580 нм; определяют содержание 1,2-PD в супернатантах анализом ГХ-МС: 0 - не определяется, предел детекции 10 мг/л. ⌀ средний; ср.-кв.откл. - среднеквадратическое отклонение.
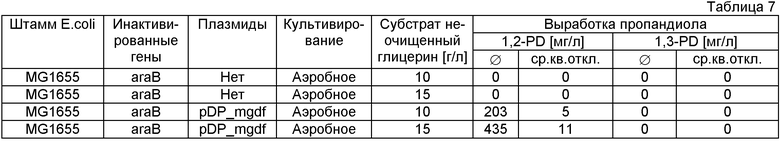
3.5. Токсичность метилглиоксаля для штамма дикого типа и мутантных штаммов Е.coli
Е.coli MG1655 дикого типа или мутанты, инактивированные в генах gloA и gloB, соответственно, культивируют в минимальной среде с чистым глицерином (10 г/л) в качестве источника углерода и энергии, содержащим различные количества метилглиоксаля. Испытания с глиоксалем показывают период химической устойчивости по меньшей мере 72 часа. Е.coli культивируют в микропланшетах без перемешивания при 37°С во влажной атмосфере. Рост определяют при OD580 через 48,5 час после инокуляции (фиг.1; сплошные столбики - Е.coli MG1655 дикого типа; серые столбики - мутант Е.coli MG1655 gloA; заштрихованные столбики - мутант Е.coli MG1655 gloB). Ингибирование роста штамма дикого типа наблюдают в случае внеклеточной концентрации метилглиоксаля, равной или превышающей 2,5 мМ. Мутантные штаммы значительно более чувствительны, в особенности мутант глиоксилазы I (gloA), что таким образом поддерживает открытие авторов, что gloA является основным путем отвода внутриклеточного метилглиоксаля (см. 4.6). Результаты также показывают, что повышенные уровни метилглиоксаля ингибируют рост Е.coli, что является основой для идентификации нечувствительных к фосфату вариантов метилглиоксальсинтаз.
3.6. Идентификация основной MG-удаляющей активности
Е.coli MG1655 инактивируют (нокаут генов) в генах, чьи генные продукты вовлечены в метаболизм метилглиоксаля (MG), например детоксикацию MG. Гены перечислены в таблице 8. Штамм дикого типа и мутантные штаммы культивируют в культуральных пробирках в аэробных условиях и при перемешивании до тех пор, пока рост не прекратится. Уровни 1,2-PD в объеме жидкости определяют анализом ГХ-MC и сравнивают с уровнями, которые обнаруживают, когда источником углерода и энергии является глюкоза (отрицательный контроль). Когда субстратом для выращивания является глицерин, обнаруживают уровень фона примерно 20 мг/мл 1,2-PD. Однако существенно повышенные уровни 1,2-PD обнаруживают только для мутанта gloA.
Штаммы Е.coli MG1655, мутированные в генах, вовлеченных в метаболизм метилглиоксаля, культивируют в минимальной среде, содержащей глицерин или глюкозу (10 г/л). Культивирование выполняют в аэробных условиях при 37°С до тех пор, пока рост не прекратится. Рост определяют как оптическую плотность (OD580); уровни пропандиола определяют анализом ГХ-МС. ⌀ средний; ср.-кв.откл. - среднеквадратическое отклонение.
3.7. Рекомбинантные организмы, продуцирующие высокие уровни 1,2-PD
Е.coli MG1655 инактивируют (нокаут генов) в генах аrаВ или в генах aldA и gloA. Соответствующих мутантов трансформируют с плазмидой pDP_mgdf. Мутантов аrаВ, укрывающих плазмиду pDP_mgdf, дополнительно трансформируютс плазмидой pCR2.1, колирующей гены, придающие альдокеторедуктазную активность, например, dkgA или dkgB Е.coli. Клетки культивируют в присутствии неочищенного глицерина (15 г/л) в аэробных и микроанаэробных условиях при 37°С в течение 2-5 суток или до тех пор, пока оптическая плотность не перестанет возрастать далее. Супернатанты анализируют на содержание 1,2-PD анализом ГХ-МС (табл.9).
Мутантные штаммы Е.coli MG1655, экспрессирующие гены, превращающие глицерин в 1,2-PD, культивируют в минимальной среде, содержащей глицерин или глюкозу (15 г/л). Культивирование выполняют в аэробных и микроанаэробных условиях при 37°С до тех пор, пока рост не прекратится. Уровни пропандиола в супернатантах определяют анализом ГХ-МС. ⌀ средний; ср.-кв.откл. - среднеквадратическое отклонение.,
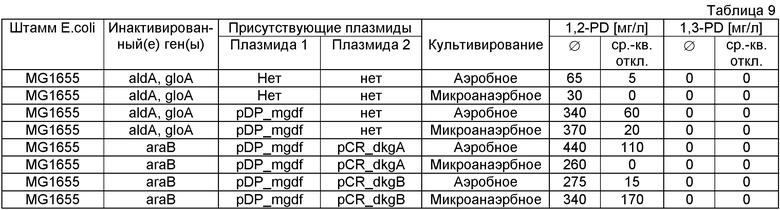
Результаты
Как видно из таблиц 2 и 3, неочищенные препараты глицерина могут использоваться без дополнительной обработки самыми разными организмами, не ограниченными Е.coli и близкими формами. Выработка биомассы, полученной за счет утилизации неочищенного глицерина, равна или выше при сравнении с чистым глицерином как углеродным субстратом. Таким образом, неочищенный глицерин заменяет чистые препараты глицерина фактически в любом процессе, который основан на ферментации глицерина.
Как видно из таблицы 4, примеси, присутствующие в неочищенном глицерине, создают источник фосфора, поддерживающего продленный рост клеток-хозяев. Таким образом, количество фосфора, добавляемого в культуральную жидкость, можно снизить на примерно 70%.
Как видно из таблицы 6, активность пропандиолоксидоредуктазы необходима для синтеза 1,2-PD из глицерина. Клетки-хозяева, которые экспрессируют глицеролдегидрогеназу, не вырабатывают 1,2-PD. Клетки, которые экспрессируют глицеролдегидрогеназу и метилглиоксальсинтазу, но с отсутствием пропандиолоксидоредуктазы, не вырабатывают заметных количеств 1,2-PD, в то время как дополнительное присутствие активности пропандиолоксидоредуктазы приводит к существенной выработке 1,2-PD.
Как видно из таблицы 7, высокие титры 1,2-PD получают из глицерина с помощью рекомбинантных штаммов, коэкспрессирующих метилглиоксальсинтазу, глицеролдегидрогеназу, дигадроксиацетонкиназу и пропандиолоксидоредуктазу в аэробных условиях. Синтез пропандиола на основе настоящего изобретения приводит к синтезу исключительно 1,2-изомера, но не 1,3-изомера пропандиола.
Как видно из таблицы 8, инактивация активности глиоксилазы I, кодированной в Е.coli gloA, приводит к существенной выработке 1,2-PD из глицерина. Таким образом, ферментативная активность, кодированная gloA, является главной конкурирующей активностью, влияющей на выработку 1,2-PD в клетках-хозяевах на высоком уровне.
Воплощения изобретения
1. Клетка-хозяин, созданная методами генной инженерии для выработки высоких уровней 1,2-пропандиола при выращивании на глицерине как единственном источнике углерода.
2. Клетка-хозяин по предшествующему воплощению, где глицерин имеет степень чистоты по меньшей мере 70%, в особенности по меньшей мере 75%, в особенности по меньшей мере 80%, в особенности по меньшей мере 85%, в особенности по меньшей мере 90%, в особенности по меньшей мере 95%, в особенности по меньшей мере 99% и до 100%.
3. Клетка-хозяин по любому из предшествующих воплощений, где глицерин имеет степень чистоты от 80% до 90%.
4. Клетка-хозяин по любому из предшествующих воплощений, где глицерин имеет степень чистоты примерно 85%.
5. Клетка-хозяин по любому из предшествующих воплощений, где глицерин представляет собой неочищенный препарат глицерина с производства биодизеля и/или биоэтанола.
6. Клетка-хозяин по любому из предшествующих воплощений, созданная методами генной инженерии рекомбинантных ДНК.
7. Клетка-хозяин по любому из предшествующих воплощений, в особенности воплощению 6, созданная методами генной инженерии путем введения гена, кодирующего активность пропандиолоксидоредуктазы (fucO).
8. Клетка-хозяин по воплощению 7, созданная методами генной инженерии путем введения по меньшей мере одного дополнительного гена, кодирующего ферментативную активность, выбранную из группы, состоящей из глицеролдегидрогеназы (gldA), дигидроксиацетонкиназы (dhaK) и метилглиоксальсинтазы (mgsA), с тем, чтобы экспрессировать указанные активности вместе с активностью пропандиолоксидоредуктазы (fucO).
9. Клетка-хозяин по воплощению 7, созданная методами генной инженерии путем введения дополнительного гена, кодирующего глицеролдегидрогеназу, с тем, чтобы экспрессировать указанную активность глицеролдегидрогеназы вместе с активностью пропандиолоксидоредуктазы.
10. Клетка-хозяин по воплощению 7, созданная методами генной инженерии путем введения дополнительного гена, кодирующего дигидроксиацетонкиназу, с тем, чтобы экспрессировать указанную активность дигидроксиацетонкиназы вместе с активностью пропандиолоксидоредуктазы.
11. Клетка-хозяин по воплощению 7, созданная методами генной инженерии путем введения дополнительного гена, кодирующего метилглиоксальсинтазу (mgsA), с тем, чтобы экспрессировать указанную активность метилглиоксальсинтазы вместе с активностью пропандиолоксидоредуктазы.
12. Клетка-хозяин по воплощению 7, созданная методами генной инженерии путем введения дополнительных генов, кодирующих глицеролдегидрогеназу и дигидроксиацетонкиназу, с тем, чтобы экспрессировать указанные активности глицеролдегидрогеназы и дигидроксиацетонкиназы вместе с активностью пропандиолоксидоредуктазы.
13. Клетка-хозяин по воплощению 7, созданная методами генной инженерии путем введения дополнительных генов, кодирующих глицеролдегидрогеназу и метилглиоксальсинтазу, с тем, чтобы экспрессировать указанные активности глицеролдегидрогеназы и метилглиоксальсинтазы вместе с активностью пропандиолоксидоредуктазы.
14. Клетка-хозяин по воплощению 7, созданная методами генной инженерии путем введения дополнительных генов, кодирующих дигидроксиацетонкиназу и метилглиоксальсинтазу, с тем, чтобы экспрессировать указанные активности дигидроксиацетонкиназы и метилглиоксальсинтазы вместе с активностью пропандиолоксидоредуктазы.
15. Клетка-хозяин по воплощению 7, созданная методами генной инженерии путем введения дополнительных генов, кодирующих глицеролдегидрогеназу, дигидроксиацетонкиназу и метилглиоксальсинтазу, с тем, чтобы экспрессировать указанные активности глицеролдегидрогеназы, дигидроксиацетонкиназы и метилглиоксальсинтазы вместе с активностью пропандиолоксидоредуктазы.
16. Клетка-хозяин по любому из предшествующих воплощений, в особенности воплощения 7, созданная методами генной инженерии путем введения дополнительных генов, кодирующих глицеролдегидратазу, с тем, чтобы экспрессировать указанную активность глицеролдегидратазы вместе с активностью пропандиолоксидоредуктазы.
17. Клетка-хозяин по любому из предшествующих воплощений, в особенности воплощения 7, созданная методами генной инженерии путем введения дополнительных генов, кодирующих альдокеторедуктазу, с тем, чтобы экспрессировать указанную активность альдокеторедуктазы вместе с активностью пропандиолоксидоредуктазы.
18. Клетка-хозяин по воплощению 17, где указанная активность альдокеторедуктазы вносится геном, выбранным из группы, состоящей из dkgA, dkgB, уеаЕ и yghZ.
19. Клетка-хозяин по любому из предшествующих воплощений, дефектная по метаболизму арабинозы.
20. Клетка-хозяин по предшествующему воплощению, где указанный дефект имеет место из-за уменьшенной или отсутствующей активности рибулозкиназы.
21. Клетка-хозяин по любому из предшествующих воплощений, дефектная по метаболизму метилглиоксаля.
22. Клетка-хозяин по предшествующему воплощению, где указанный дефект имеет место из-за уменьшенной или отсутствующей активности, выбранной из группы, состоящей из активности системы глиоксилазы I, системы глиоксилазы II, лактатдегидрогеназы А, системы глиоксилазы III, альдегиддегидрогеназы А, но предпочтительно активности системы глиоксилазы I.
23. Клетка-хозяин по любому из предшествующих воплощений, дефектная по метаболизму дигидроксиацетонфосфата.
24. Клетка-хозяин по предшествующему воплощению, где указанный дефект имеет место из-за уменьшенной или отсутствующей активности тиозофосфатизомеразы.
25. Клетка-хозяин по любому из предшествующих воплощений, вырабатывающая высокие уровни 1,2-пропандиола при выращивании на глицерине как единственном источнике углерода, но по существу, без 1,3-пропандиола.
26. Клетка-хозяин по любому из предшествующих воплощений, представляющая собой клетку-хозяина микроорганизма или гриба.
27. Клетка-хозяин микроорганизма по воплощению 26, представляющая собой Е.coli.
28. Полинуклеотид, включающий синтетический оперон под контролем индуцируемого промотора, включающий гены, кодирующие глицеролдегидрогеназу (gldA) и пропандиолоксидоредуктазу (fucO).
29. Полинуклеотид, включающий синтетический оперон под контролем индуцируемого промотора, включающий гены, кодирующие глицеролдегидрогеназу (gldA), дигидроксиацетонкиназу (dhaK) и пропандиолоксидоредуктазу (fucO).
30. Полинуклеотид по воплощению 29, где синтетический оперон расширен для того, чтобы также содержать ген, кодирующий метилглиоксальсинтазу (mgsA).
31. Полинуклеотид по воплощению 29 и 30, где гены, кодирующие глицеролдегидрогеназу (gldA), пропандиолоксидоредуктазу (fucO) и метилглиоксальсинтазу (mgsA), соответственно, доступны из Е.coli, и ген, кодирующий дигидроксиацетонкиназу (dhaK), доступен из С.freundii.
32. Полинуклеотид по любому из предшествующих воплощений, представляющий собой плазмиду.
33. Полинуклеотид по любому из предшествующих воплощений, в котором порядок генов в синтетическом опероне 5'-mgsA-gldA-dhaK-fucO-3'.
34. Полинуклеотид по любому из предшествующих воплощений, в котором индуцируемый промотор представляет собой арабинозаиндуцируемый промотор.
35. Микроорганизм по любому из предшествующих воплощений, включающий полинуклеотид по любому из воплощений 28-34.
36. Микроорганизм по любому из предшествующих воплощений, включающий нечувствительный к фосфату ген mgsA, который полностью операбелен при высоких концентрациях фосфата, в особенности при концентрациях фосфата, превышающих 0,7 нМ, в культуральной среде или превышающей 9,3е-05 в цитоплазме клетки.
37. Способ получения 1,2-пропандиола, включающий выращивание клетки-хозяина по любому из воплощений 1-27 в соответствующей среде для выращивания, содержащей простой источник углерода, в особенности неочищенный препарат глицерина, после чего выработанный 1,2-пропандиол извлекают и, необязательно, очищают.
38. Способ по воплощению 37, включающий
i) культивирование клеток-хозяев по любому из воплощений 1-27, экспрессирующих активность пропандиолоксидоредуктазы (fucO), в среде, содержащей неспособный к брожению углеродный субстрат, в связи с чем углеродный субстрат поддерживает выработку биомассы и в то же время служит в качестве субстрата для выработки 1,2-пропандиола (1,2-PD), и неспособный к брожению источник углерода метаболизируется клетками-хозяевами в 1,2-пропандиол;
ii) извлечение 1,2-пропандиола, полученного на стадии i); и, необязательно,
iii) очистку извлеченного 1,2-пропандиола.
39. Способ по любому из предшествующих воплощений, где указанный неспособный к брожению углеродный субстрат представляет собой неочищенный препарат глицерина, в особенности препарат, содержащий глицерин с чистотой по меньшей мере 70%, в особенности по меньшей мере 75%, в особенности по меньшей мере 80%, в особенности по меньшей мере 85%, в особенности по меньшей мере 90%, в особенности по меньшей мере 95%, в особенности по меньшей мере 99% и до 100%.
40. Способ по любому из предшествующих воплощений, где глицерин имеет чистоту от 80% до 90%, в особенности примерно 85%.
41. Способ по любому из предшествующих воплощений, где неспособный к брожению углеродный субстрат, в особенности неочищенный препарат глицерина селективно метаболизируется до 1,2-пропандиола.
42. Способ по любому из предшествующих воплощений, в котором используют клетку-хозяина, созданную методами генной инженерии для сверхэкспрессии пропандиолоксидоредуктазы (fucO).
43. Способ по любому из предшествующих воплощений, в котором используют клетку-хозяина, созданную методами генной инженерии для коэкспрессии по меньшей мере одного фермента, выбранного из группы, состоящей из глицеролдегидрогеназы (gldA), дигидроксиацетонкиназы (dhaK) и метилглиоксальсинтазы (mgsA), вместе с активностью пропандиолоксидоредуктазы (fucO).
44. Способ по любому из предшествующих воплощений, в котором используют клетку-хозяина, в которой инактивирована по меньшей мере одна ферментативная активность, вовлеченная в непродуктивный путь, конкурирующий с выработкой 1,2-PD.
45. Способ по любому из предшествующих воплощений, в котором используют мутант микроорганизма, в особенности мутант Е.coli, где инактивированы один или несколько генов, кодирующих системы глиоксилазы I и II (gloA и gloB), лактатгидрогеназу A (ldhA), системе глиоксилазы III (косвенно, путем инактивации основного регулятора rpoS).
46. Способ по любому из предшествующих воплощений, в котором используют мутант или штамм микроорганизма, в особенности мутант Е.coli, где ген, кодирующий активность gloA, частично или полностью инактивирован.
47. Способ по любому из предшествующих воплощений, в котором используют мутант или штамм микроорганизма, инактивированный по метаболизму арабинозы.
48. Способ по любому из предшествующих воплощений, в котором в качестве организма-хозяина используют штамм Е.coli MG1655 и DH5-альфа, соответственно.
49. Способ по любому из предшествующих воплощений, где по меньшей мере один из генов, кодирующих ферментативную активность, выбранный из группы, состоящей из глицеролдегидрогеназы (gldA), дигидроксиацетонкиназы (dhaK) и метилглиоксальсинтазы (mgsA) и пропандиолоксидоредуктазы (fucO), находится под контролем индуцируемого промотора, в особенности арабинозаиндуцируемого промотора, в особенности промотора paraBAD.
50. Способ по любому из предшествующих воплощений, в котором используют синтетический оперон для предоставления клетки-хозяина, коэкспрессирующей по меньшей мере одну ферментативную активность, выбранную из группы, состоящей из активности глицеролдегидрогеназы (gldA), дигидроксиацетонкиназы (dhaK) и метилглиоксальсинтазы (mgsA), вместе с активностью пропандиолоксидоредуктазы (fucO).
51. Способ по любому из предшествующих воплощений, где гены, кодирующие вышеуказанные активности, находятся под контролем индуцируемого промотора, в особенности арабинозаиндуцируемого промотора, но в особенности промотора paraBAD.
52. Способ по любому из предшествующих воплощений, где непрерывный ряд генов, транскрибированных после индукции из указанного оперона, является следующим: mgsA, gldA, dhaK, fucO.
53. Способ получения клетки-хозяина, которую можно использовать в способе по любому из воплощений 37-52 для получения 1,2-пропандиола, включающий трансформацию указанной клетки-хозяина полинуклеотидом по любому из воплощений 27-33.
54. Способ по воплощению 53, где указанная клетка-хозяин представляет собой клетку-хозяина микроорганизма, в особенности Е.coli.
55. Способ по воплощению 53, где трансформацию осуществляют электропорацией.
56. Клетка-хозяин, полученная способом по любому из воплощений 53-55.
Ссылки
Altaras, N.E, 2001; Biotechnology Progress (17), 52-56.
Appleyard, R.K., Genetics, 39, 440-452 (1954).
Ausubel, Current Protocols in Molecular Biology, Green Publishing Associates and Wiley Interscience, N.Y. (1994).
Ausubel, F.M., Brent, R., Kingston, R. E., Moore, D.D., Seidman, J.G., Smith, J.A. & Struhl, K. (1995). Short Protocols in Molecular Biology. A Compendium of Methods from Current Protocols in Molecular Biology, 3 edn. New York: John Wiley & Sons, Inc.
Batzer, et al., Nucleic Acid Res. 19:5081 (1991);
Cameron, D.C., 1986; Bio/Technology (4), 651-654.
Cameron, D.C, 1998; Biotechnology Progress (14), 116-125.
Cunningham, R.P., DasGupta, C., Shibata, Т., and Radding, С.М. (1980) Cell 20, 223-235).
Datsenko, K.A., 2000; PNAS (97), 6640-6645.
Ellis, E.H., 2001; PNAS (98), 6742-6746.
Gough JA, Murray NE., J. Mol. Biol., 166, 1-19 (1983).
Grandison, Alistair S., Sep. Processes Food Biotechnol. Ind. (1996), 155 177.
Guzman, L.M., 1995; Journal of Bacteriology (177), 4121-4130.
Hanahan J., et al., Mol. Biol., 166, 557-580 (1983).
Heath, B.C., 1962; The Journal of Biological Chemistry (237), 2423-2426.
Hopper, D.J. 1972; Biochemical Journal (128), 321-329.
Jones, L.R. & Riddick, J.A. (1957). Colorimetric determination of 1,2-propanediol and related compounds. Anal Chem 39, 1214-1216.
Joyner, A. 1999, Gene Targeting, A Practical Approach, Oxford University Press. ISBN-13: 978-0-19-963792-8.
Karberg, М., 2001; Nature Biotechnology (19), 1162-1167.
Kluyver and Schellen, 1937.
Le Mouellic, K. et al, Proc. Natl. Acad. Sci. USA, 87 (1990), 4712-4716.
Marx, C.J. and Lidstrom, M.E., (2002), BioTechniques (33)5, 1062-1067.
Miller, J.H. (1992). A Short Course in Bacterial Genetics: A Laboratory Manual and Handbook for Escherichia Coli and Related Bacteria, 1st edn: Cold Spring Harbor Laboratory Press.
Neidhardt et al. 1996, Escherichia coli and Salmonella: Cellular and Molecular Biology, ASM Press.
Ohtsuka, et al., J. Biol. Chem. 260:2605-2608 (1985).
Pfennig, N. & Lippert, K.D. (1966). Uber das Vitamin B12-Bedurfnis phototropher Schwefelbakterien. Archives of microbiology 55, 245-256.
Reynolds, A.B. et al. Cell 38, 275-285 (1984).
Rossolini, et al., Mol. Cell. Probes 8:91-98 (1994).
Sambrook et al., Molecular Cloning: A Laboratory Manual, Second Edition (1989) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989).
Sambrook, J. & Russell, D.W. (2001). Molecular Cloning: A Laboratory Manual, 3rd edn: Cold Spring Harbor Laboratory Press.
Tran Din, K, 1985; Archives of Microbiology (142), 87-92.
Wood, W.B, J. Mol. Biol, 16, 118-133 (1966).
Yu, D., 2000; PNAS (97), 5978-5983.
Jerpseth, В. et al. Strategies, 5, 81-83 (1992).
Young, R.A. and R.W. Davis, Science, 222, 778-782 (1983).
Патентная литература
Пат. США №5008473.
Пат. США №5356812.
Пат. США №7049109.
Пат. США №6087140.
Пат. США №6303352.
Пат. США №4683202.
Заявка на пат. США 2007/072279.
WO 2005/073364.
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЙ МИКРООРГАНИЗМ И СПОСОБ ПОЛУЧЕНИЯ 1,2-ПРОПАНДИОЛА, ОСНОВАННЫЙ НА НАДФН-ЗАВИСИМОЙ АЦЕТОЛРЕДУКТАЗЕ И УЛУЧШЕННОЙ ДОСТАВКЕ НАДФН | 2015 |
|
RU2711870C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТЫ | 2008 |
|
RU2515044C2 |
| БАКТЕРИЯ ESCHERICHIA COLI ДЛЯ ПОЛУЧЕНИЯ 1,2-ПРОПАНДИОЛА, СПОСОБ ЕЕ ПОЛУЧЕНИЯ, СПОСОБ ПОЛУЧЕНИЯ 1,2-ПРОПАНДИОЛА | 2005 |
|
RU2407793C2 |
| МУТАНТНАЯ АДЕНИЛАТЦИКЛАЗА, ДНК, КОДИРУЮЩАЯ ЕЕ, БАКТЕРИЯ СЕМЕЙСТВА ENTEROBACTERIACEAE, СОДЕРЖАЩАЯ УКАЗАННУЮ ДНК, И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ | 2010 |
|
RU2471868C2 |
| НОВЫЕ ФЕРМЕНТЫ МЕТАНОЛДЕГИДРОГЕНАЗЫ ИЗ Bacillus | 2013 |
|
RU2624017C2 |
| Способ получения 2,4-дигидроксимасляной кислоты | 2011 |
|
RU2626531C2 |
| БАКТЕРИЯ - ПРОДУЦЕНТ ПРОДУКТА РЕАКЦИИ, КАТАЛИЗИРУЕМОЙ БЕЛКОМ, ОБЛАДАЮЩИМ АКТИВНОСТЬЮ 2-ОКСОГЛУТАРАТ-ЗАВИСИМОГО ФЕРМЕНТА, И СПОСОБ ПРОДУКЦИИ УКАЗАННОГО ПРОДУКТА | 2008 |
|
RU2444568C2 |
| РЕКОМБИНАНТНЫЙ МИКРООРГАНИЗМ ДЛЯ ФЕРМЕНТАТИВНОГО ПРОИЗВОДСТВА МЕТИОНИНА | 2012 |
|
RU2629760C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-АРГИНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ СЕМЕЙСТВА Enterobacteriaceae, СОДЕРЖАЩЕЙ N-АЦЕТИЛОРНИТИНДЕАЦЕТИЛАЗУ С НАРУШЕННОЙ АКТИВНОСТЬЮ | 2012 |
|
RU2550269C2 |
| ИЗОПРЕНСИНТАЗА И КОДИРУЮЩИЙ ЕЕ ГЕН, СПОСОБ ПОЛУЧЕНИЯ МОНОМЕРА ИЗОПРЕНА | 2013 |
|
RU2612916C2 |

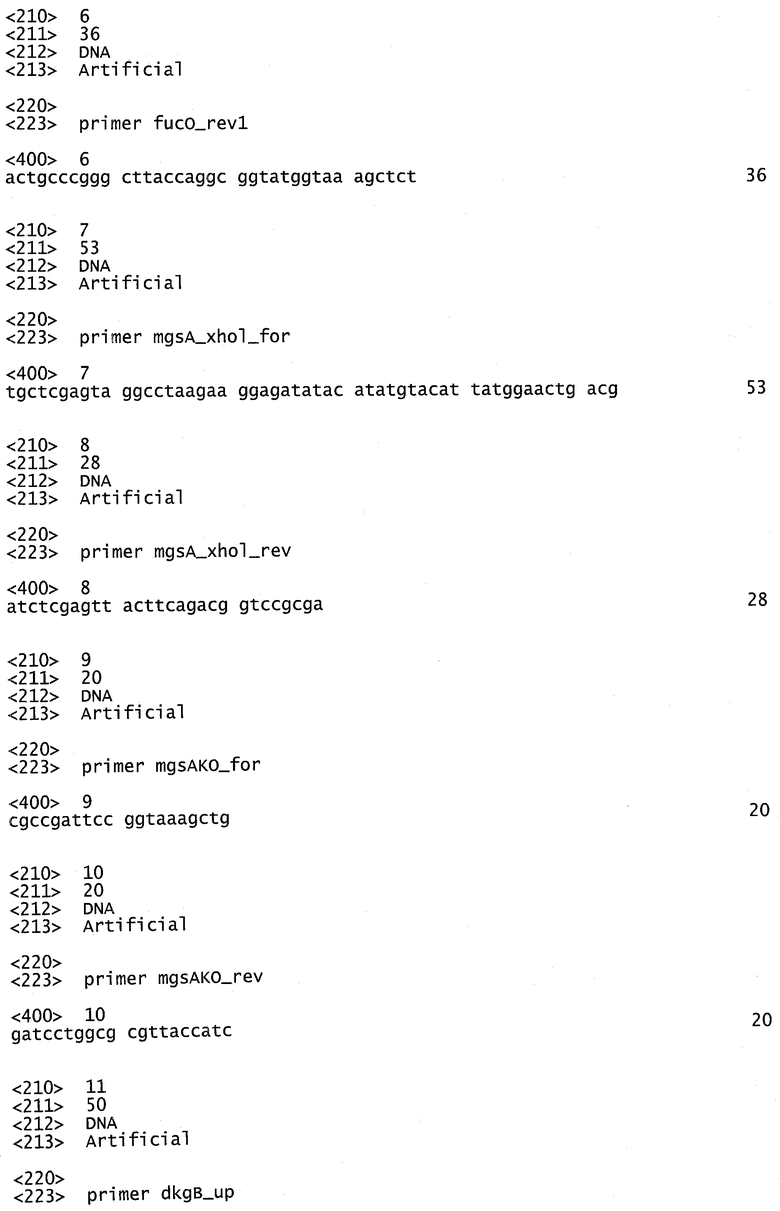
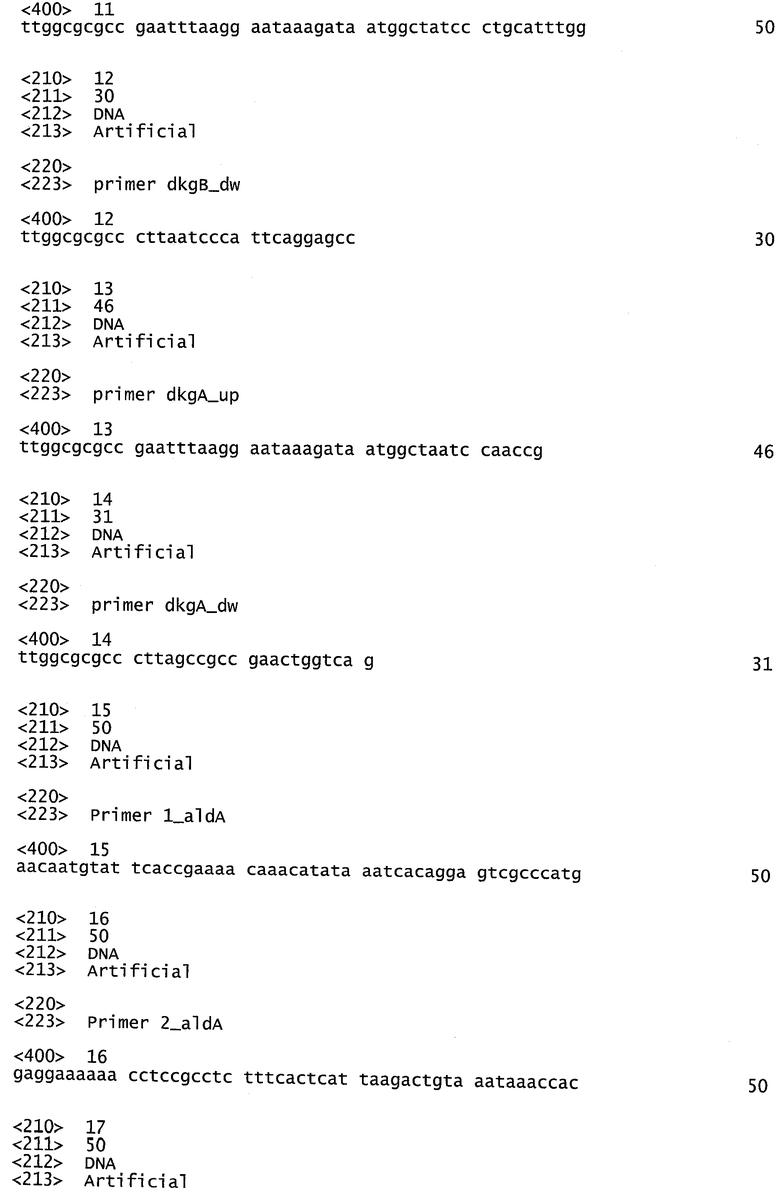















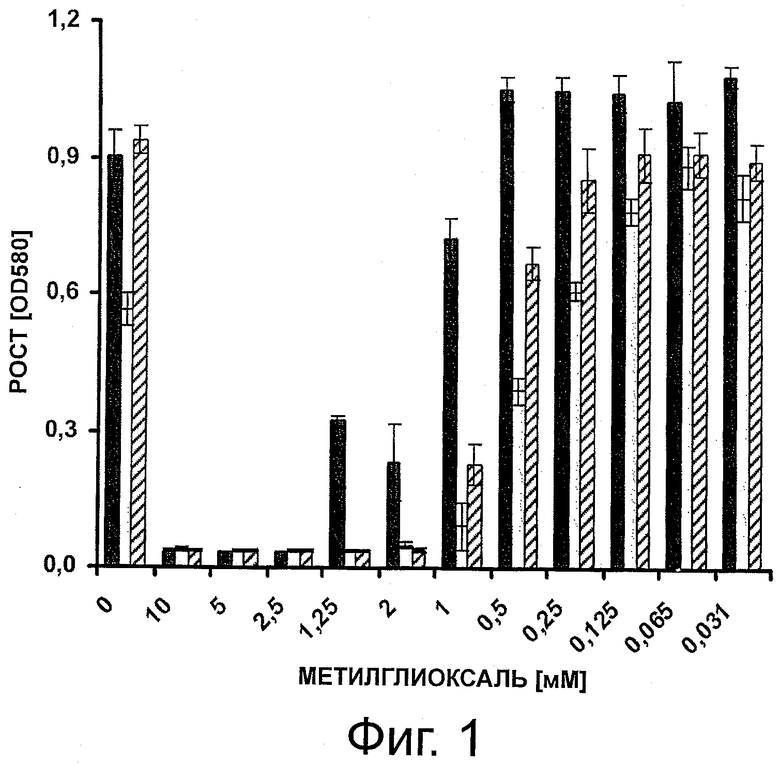
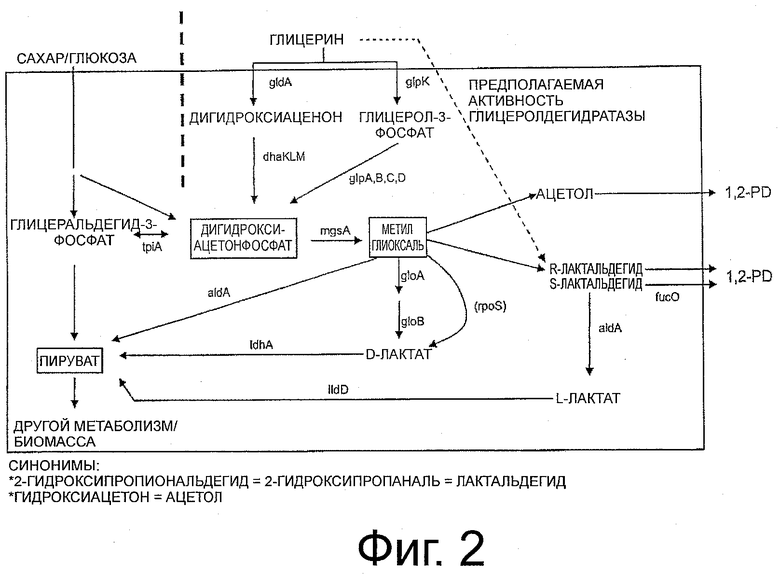
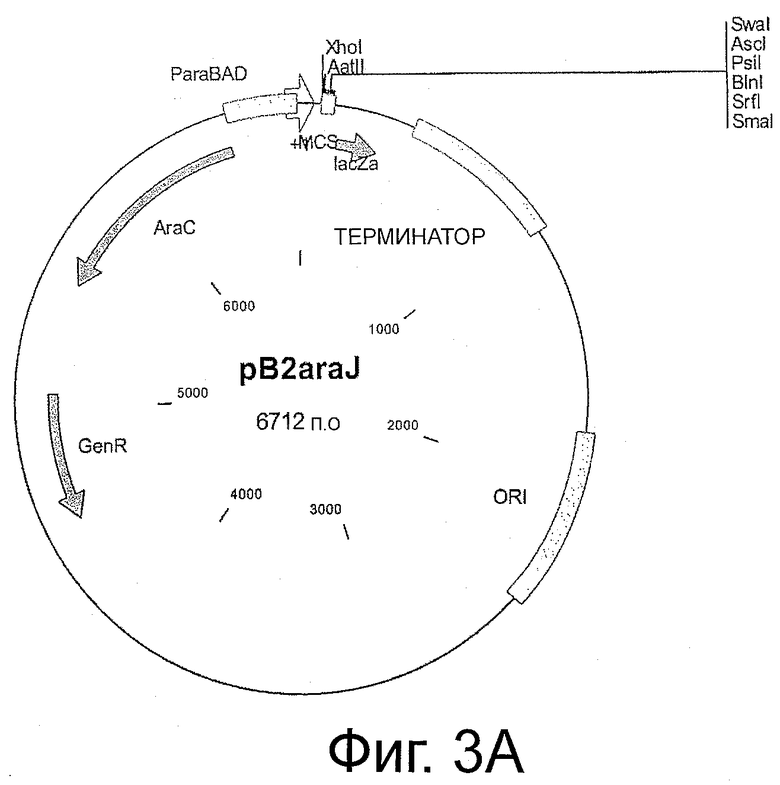
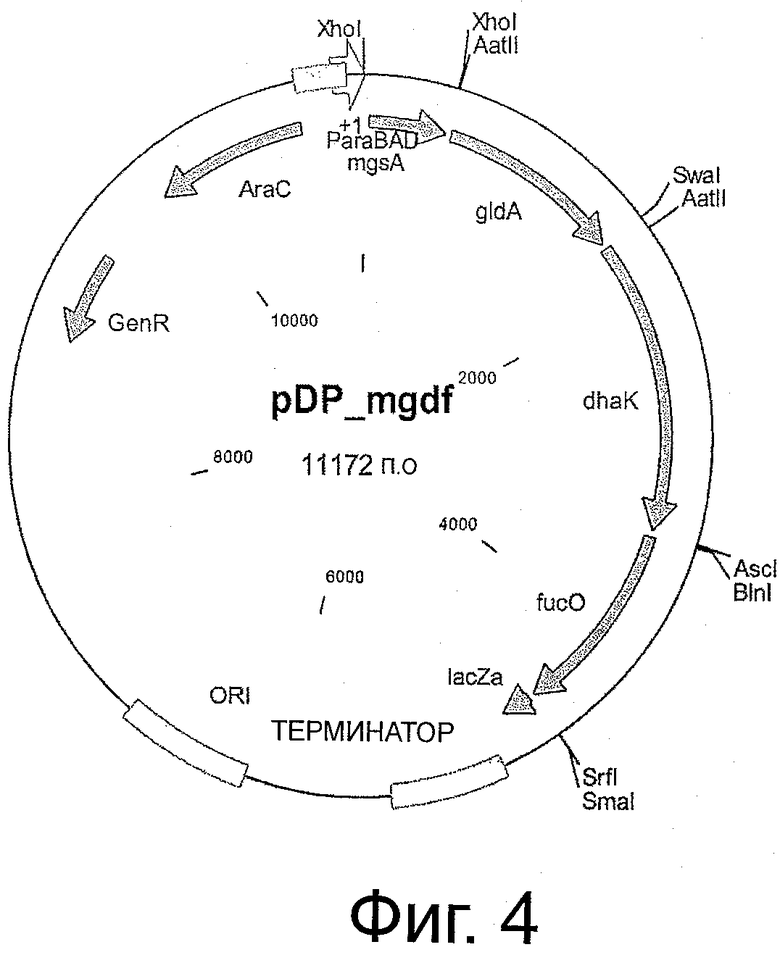
Изобретение относится к области генной инженерии и может быть использовано для рекомбинантного получения 1,2-пропандиола (1,2-PD). В клетку E.coli вводят ген, кодирующий пропандиолоксидоредуктазу, что позволяет продуцировать высокие уровни 1,2-пропандиола, по существу без 1,3-пропандиола, при выращивании на глицерине как единственном источнике углерода. Дополнительно могут быть введены гены глицеролдегидрогеназы (gldA), дигидроксиацетонкиназы (dhaK) и/или метилглиоксальсинтазы (mgsA) с тем, чтобы экспрессировать указанные ферменты вместе с пропандиолоксидоредуктазой (fucO), а также ген глицеролдегидратазы или альдокеторедуктазы, с тем, чтобы экспрессировать указанную глицеролдегидратазу или альдокеторедуктазу вместе с пропандиолоксидоредуктазой. Клетка E.coli, трансформированная геном, кодирующим пропандиолоксидоредуктазу, может быть дефектна по метаболизму арабинозы, метилглиоксаля и/или дигидроксиацетонфосфата. 8 н. и 1 з.п. ф-лы, 4 ил., 9 табл., 3 пр.
1. Клетка E.coli, созданная методами генной инженерии путем введения гена, кодирующего пропандиолоксидоредуктазу, предназначенная для выработки высоких уровней 1,2-пропандиола, но по существу, без 1,3-пропандиола, при выращивании на глицерине как единственном источнике углерода.
2. Клетка E.coli по п.1, где глицерин имеет степень чистоты от 80% до 90%.
3. Клетка E.coli, созданная методами генной инженерии путем введения гена, кодирующего пропандиолоксидоредуктазу, предназначенная для выработки высоких уровней 1,2-пропандиола, но по существу, без 1,3-пропандиола, при выращивании на глицерине как единственном источнике углерода, где указанная клетка также сконструирована путем введения по меньшей мере одного дополнительного гена, кодирующего фермент, выбранный из группы, состоящей из глицеролдегидрогеназы (gldA), дигидроксиацетонкиназы (dhaK) и метилглиоксальсинтазы (mgsA), с тем, чтобы экспрессировать указанные ферменты вместе с пропандиолоксидоредуктазой (fucO).
4. Клетка E.coli, созданная методами генной инженерии путем введения гена, кодирующего пропандиолоксидоредуктазу, предназначенная для выработки высоких уровней 1,2-пропандиола, но по существу, без 1,3-пропандиола, при выращивании на глицерине как единственном источнике углерода, где указанная клетка также сконструирована путем введения дополнительных генов, кодирующих глицеролдегидрогеназу, дигидроксиацетонкиназу и метилглиоксальсинтазу, с тем, чтобы экспрессировать указанные глицеролдегидрогеназу, дигидроксиацетонкиназу и метилглиоксальсинтазу вместе с пропандиолоксидоредуктазой.
5. Клетка E.coli, созданная методами генной инженерии путем введения гена, кодирующего пропандиолоксидоредуктазу, предназначенная для выработки высоких уровней 1,2-пропандиола, но по существу, без 1,3-пропандиола, при выращивании на глицерине как единственном источнике углерода, где указанная клетка также сконструирована путем введения дополнительных генов, кодирующих глицеролдегидратазу, с тем, чтобы экспрессировать указанную глицеролдегидратазу вместе с пропандиолоксидоредуктазой.
6. Клетка E.coli, созданная методами генной инженерии путем введения гена, кодирующего пропандиолоксидоредуктазу, предназначенная для выработки высоких уровней 1,2-пропандиола, но по существу, без 1,3-пропандиола, при выращивании на глицерине как единственном источнике углерода, где указанная клетка также сконструирована путем введения дополнительных генов, кодирующих альдокеторедуктазу, с тем, чтобы экспрессировать указанную альдокеторедуктазу вместе с пропандиолоксидоредуктазой.
7. Клетка E.coli, созданная методами генной инженерии путем введения гена, кодирующего пропандиолоксидоредуктазу, предназначенная для выработки высоких уровней 1,2-пропандиола, но по существу, без 1,3-пропандиола, при выращивании на глицерине как единственном источнике углерода, где указанная клетка дефектна по метаболизму по меньшей мере одного из соединений, выбранных из группы, состоящей из:
i) арабинозы,
ii) метилглиоксаля,
iii) дигидроксиацетонфосфата.
8. Клетка E.coli, созданная методами генной инженерии путем введения гена, кодирующего пропандиолоксидоредуктазу, предназначенная для выработки высоких уровней 1,2-пропандиола, но по существу, без 1,3-пропандиола, при выращивании на глицерине как единственном источнике углерода, где указанная клетка дефектна по метаболизму по меньшей мере одного из соединений, выбранных из группы, состоящей из:
i) арабинозы,
ii) метилглиоксаля,
iii) дигидроксиацетонфосфата,
и где глицерин имеет степень чистоты между 80% и 90%.
9. Способ получения 1,2-пропандиола, включающий выращивание клетки E.coli по любому из пп.1-8 в соответствующей среде для выращивания, содержащей глицерин как единственный источник углерода, после чего выработанный 1,2-пропандиол извлекают и, необязательно, очищают.
| WO 2008006037 A2, 10.01.2008 | |||
| ALTARAS N | |||
| E | |||
| et.al., Enhanced production of ®-1,2-propanediol by metabolically engineered Escherichia coli,Biotechnol Prog 2000, v.16, p.940-946 | |||
| ALTARAS N | |||
| E | |||
| et.al, Metabolic Engineering of a 1,2-Propanediol Pathway in Escherichia coli, Applied and environmental microbiology, 1999, p | |||
| Приспособление к лесопильной раме для распиливания коротких кряжей | 1924 |
|
SU1180A1 |
| RU 2007143867 A, | |||

