Область изобретения
Настоящее изобретение относится к рекомбинантному микроорганизму, полезному для продукции 1,2-пропандиола, и к способу получения 1,2-пропандиола. Микроорганизм по изобретению модифицирован таким образом, что продукция 1,2-пропандиола улучшается посредством усиления активности НАДФН-зависимой ацетолредуктазы.
Предшествующий уровень техники
1,2-Пропандиол или пропиленгликоль - С3 диспирт с формулой C3H8O2 или НО-СН2-СНОН-СН3-представляет собой широко используемый химический реактив. Его номер CAS - 57-55-6. Он представляет собой бесцветную, почти не имеющую запаха, прозрачную, вязкую жидкость со слабосладким вкусом, гигроскопичную и смешивающуюся с водой, ацетоном и хлороформом. Он является компонентом ненасыщенных полиэфирных смол, жидких детергентов, хладагентов, антифрозов и противообледенительных жидкостей для самолетов. Пропиленгликоль все в большей степени используется с 1993-1994 гг. в качестве замены производных этилена, которые признаются более токсичными, чем производные пропилена.
1,2-Пропандиол в настоящее время производится химическими способами с использованием способа гидратации пропиленоксида, в котором потребляются большие количества воды, используются высокотоксичные вещества и генерируются побочные продукты, такие как трет-бутанол и 1-фенилэтанол.
Недостатки химических способов получения 1,2-пропандиола делают биологический синтез привлекательной альтернативой. Для ферментативной продукции 1,2-пропандиола из сахаров посредством микроорганизмов было охарактеризовано два пути.
В первом пути 6-дезоксисахара (например, L-рамноза или L-фукоза) расщепляются на дигидроксиацетонфосфат и (S)-лактальдегид, которые могут быть далее восстановлены до (S)-1,2-пропандиола (Badia et al., 1985). Данный путь является функциональным у Е. coli, но не может давать экономически оправданный способ из-за повышенной цены дезоксигексоз.
Вторым путем является метаболизм обычных сахаров (например, глюкозы или ксилозы) через путь гликолиза, с последующим метилглиоксалевым путем. Дигидроксиацетонфосфат превращается до метилглиоксаля, который может восстанавливаться либо до лактальдегида, либо до гидроксиацетона (ацетола). Эти два соединения затем превращаются в 1,2-пропандиол. Данный путь используется природными продуцентами (R)-1,2-пропандиола, такими как Clostridium sphenoides и Thermoanaerobacter thermosaccharolyticum. Однако улучшение эффективностей, полученных с данными организмами, вероятно, является ограниченным из-за недостатка доступных генетических инструментов.
Метилглиоксалевый путь является функциональным у Е. coli или других Enterobactehaceae, и было проведено несколько исследований для генетических модификаций Е. coli для того, чтобы получить продуцент 1,2-пропандиола с использованием простых источников углерода (WO 98/37204; Cameron et al., 1998; Altaras and Cameron, 1999; Huang et al., 1999; Altaras and Cameron, 2000; Berrios-Rivera et al., 2003; Jarboe, 2011). Улучшенные штаммы E. coli, продуцирующие 1,2-пропандиол, полученные комбинацией рационального конструирования и эволюции, описаны в патентных заявках WO 2005/073364, WO 2008/116848, WO 2008/116852, WO 2008/116853, WO 2010/051849, WO 2011/012693, WO 2011/012697 и WO 2011/012702, которые тем самым являются включенными в данный документ посредством ссылки.
В штаммах Е. coli, продуцирующих 1,2-пропандиол, восстановление гидроксиацетона до 1,2-пропандиола проводится глицеролдегидрогеназой - GlyDH - с использованием НАДН (восстановленный никотинамидадениндинуклеотид) в качестве кофактора и не является полностью обусловленным внутренним окислительно-восстановительным состоянием клетки при аэробных условиях. При аэробных условиях первичной ролью НАДН является дыхательная генерация АТФ (аденозинтрифосфат) посредством окислительного фосфорилирования, и, в результате этого, отношение НАДН к НАД+ сильно сдвинуто в пользу НАД+. Химически очень сходный НАДФН (никотинамидадениндинуклеотидфосфат), в отличие от этого, управлят анаболическими восстановлениями, и отношение НАДФН к НАДФ+ является более высоким (Fuhrer and Sauer, 2009).
Следовательно, целью данного изобретения является улучшение продукции 1,2-пропандиола посредством увеличения активности НАДФН-зависимой ацетолредуктазы и посредством улучшения доставки НАДФН.
Была установлена трехмерная структура GlyDH из термофильной бактерии Bacillus stearothermophilus, и был полностью охарактеризован сайт связывания НАД+ (Ruzheinikov et al., 2001). Был сконструирован и охарактеризован мутант В. stearothermophilus по глицеральдегид-3-фосфату, способный продуцировать НАДФН (Clermont et al., 1993).
НАДФН-зависимая вторичная алкогольдегидрогеназа (Sadh) из Clostridium beijerinckii катализирует в природе восстановление ацетона до изопропанола (Ismaiel et al., 1993). В патентной заявке ЕР 2546331 от Mitsui Chemicals Inc. изопопиловый спирт продуцируется в штамме Е. coli, экспрессирующем ген sadh из Clostridium beijerinckii.
Было показано, что данный фермент, а также его гомолог из Thermoanaerobacter brockii катализируют восстановление гидроксиацетона до 1,2-пропандиола, и они были сверхэкспрессированы в облигатном фотоавтотрофе цианобактерии Synechococcus elongatus с получением 1,2-пропандиола из неуглеводного CO2 в качестве источника углерода (Li and Liao, 2013).
Неожиданно авторы изобретения обнаружили то, что сверхэкспрессия гена sadh, одного или в комбинации с другими средствами для улучшения доставки НАДФН, значительно улучшает продукцию 1,2-пропандиола из углеводов в качестве источника углерода.
Краткое изложение сущности изобретения
Данное изобретение относится к рекомбинантным микроорганизмам и способам применения указанного микроорганизма для оптимизации продукции 1,2-пропандиола, где в указанном микроорганизме усиливается активность НАДФН-зависимой ацетолредуктазы (ГАР - гидроксиацетон редуктаза). Более конкретно, в указанном рекомбинантном микроорганизме сверхэкспрессируется по меньшей мере один ген, кодирующий активность НАДФН-зависимой ацетолредуктазы, или мутантный ген gldA*, кодирующий НАДФН-зависимую глицеролдегидрогназу, где указанная НАДФН-зависимая ацетолредуктаза имеет по меньшей мере 60%-ную идентичность аминокислот с белком, кодируемым геном adh из Clostridium beijerinckii или геном adh из Thermoanaerobacter brockii, или геном adh1 из Entamoeba histolytica, или геном GOX1615 из Gluconobacter oxydans, или геном gld2 из Hypocrea jecorina, или геном yhdN из Bacillus subtilis.
Рекомбинантный микроорганизм, используемый в данном изобретении, также может содержать другие генетические модификации, такие как:
- аттенуация экспрессии гена gldA, кодирующего НАДН-зависимую глицеролдегидрогеназу,
- аттенуация экспрессии гена yqhD, кодирующего альдегидредуктазу,
- сверхэкспрессия оперона гена pntAB, кодирующего никотинамиднуклеотидтрансгидрогеназу,
- аттенуация экспрессии гена pgi, кодирующего фосфоглюкозоизомеразу,
- аттенуация экспрессии гена pfkA, кодирующего фосфофруктокиназу,
- сверхэкспрессия гена zwf, кодирующего глюкозо-6-фосфатдегидрогеназу,
-сверхэкспрессия гена yjeF, кодирующего АДФ-зависимую дегидратазу,
- сверхэкспрессия гена gapN, кодирующего НАДФ-зависимую глицеральдегид-3-фосфатдегидрогеназу,
- сверхэкспрессия мутантного гена Ipd*, кодирующего НАДФ-зависимую липоамиддегидрогеназу.
Предпочтительно данный микроорганизм представляет собой Escherichia coli, Klebsiella pneumoniae, Thermoanaerobacterium thermosaccharolyticum, Clostridium sphenoides или Saccharomyces cerevisiae.
Подробное описание изобретения
Перед подробным описанием настоящего изобретения следует понимать, что данное изобретение не ограничивается конкретно проиллюстрированными способами и, естественно, может варьировать. Также следует понимать, что терминология, используемая в данном документе, служит лишь для цели описания конкретных воплощений изобретения и не предназначена для того, чтобы быть ограничивающей, и оно будет ограничиваться только приложенной формулой изобретения.
Все публикации, патенты и патентные заявки, процитированные в данном документе, выше или ниже, являются тем самым включенными посредством ссылки во всей их полноте.
Кроме того, в воплощении настоящего изобретения на практике, если не указано иное, используются традиционные микробиологические и молекулярно-биологические методики, находящиеся в пределах квалификации в данной области. Такие методики хорошо известны квалифицированному сотруднику и полностью объясняются в литературе.
Следует отметить то, что формы единственного числа в том виде, в котором они используются в данном документе и в приложенной формуле изобретения, включают ссылку на множественное число, если контекст явно не диктует иное. Таким образом, например, ссылка на «микроорганизм» включает множество таких микроорганизмов, и ссылка на «эндогенный ген» представляет собой ссылку на один или более чем один эндогенный ген и так далее. Если не определено иначе, все использованные в данном документе технические и научные термины имеют такие значения, которые обычно понятны обычному специалисту в области, к которой принадлежит данное изобретение. Несмотря на то, что в воплощении на практике или в тестировании настоящего изобретения можно использовать любые материалы и методы, аналогичные или эквивалентные описанным в данном документе, теперь описываются предпочтительные материалы и методы.
В формуле изобретения, которая следует далее, и в последующем описании изобретения, за исключением тех случаев, когда контекст требует иного из-за прямой формулировки или необходимого подтекста, слово «вмещать», «содержать», «включать» или «включает», или вариации, такие как «вмещает», «вмещающий», «содержащий», «включенный», «включает», «включающий», используются во включительном смысле, т.е. для определения присутствия утверждаемых характеристик, но не для исключения присутствия или добавления дополнительных характеристик в разных воплощениях изобретения.
Настоящее изобретение относится к способу получения 1,2-пропандиола в ферментативном процессе, включающем стадии культивирования микроорганизма, генетически модифицированного для продукции 1,2-пропандиола в подходящей культуральной среде, содержащей углевод в качестве источника углерода, и выделения 1,2-пропандиола из культуральной среды, где в указанном генетически модифицированном микроорганизме увеличивается активность НАДФН-зависимой ГАР. Особенно предпочтительные воплощения изобретения дополнительно описаны ниже.
Термины «ацетолредуктаза» и «гидроксиацетон редуктаза» или «ГАР» используются взаимозаменяемо и обозначают ферментативную активность восстановления ацетола (или гидроксиацетона) в 1,2-пропандиол. Данная активность может быть НАДФН-зависимой или НАДН-зависимой.
Термин «микроорганизм» в том виде, как он используется в данном документе, относится к бактерии, дрожжам или грибку, которые не являются искусственно модифицированными. Предпочтительно микроорганизм выбран среди Enterobacteriaceae, Bacillaceae, Closthdiaceae, Streptomycetaceae и дрожжей. Более предпочтительно микроорганизм представляет собой вид Escherichia, Klebsiella, Thermoanaerobacterium, Clostridium или Saccharomyces. Даже более предпочтительно микроорганизм выбран среди Escherichia coli, Klebsiella pneumoniae, Thermoanaerobacterium thermosaccharolyticum, Clostridium sphenoides или Saccharomyces cerevisiae. Предпочтительно данный микроорганизм представляет собой гетеротрофный микроорганизм, т.е. не способен фиксировать атмосферный углерод и использует вместо этого источники органического углерода для его роста. Даже более предпочтительно гетеротрофный микроорганизм по изобретению представляет собой Escherichia coli.
Термин «рекомбинантный микроорганизм» или «генетически модифицированный микроорганизм» в том виде, как он используется в данном документе, относится к бактерии, дрожжам или грибку, который не встречается в природе и является генетически отличным от его эквивалента, обнаруженного в природе. Это означает то, что он модифицирован либо посредством введения, либо посредством делеции, либо посредством модификации генетических элементов. Он также может быть трансформирован путем принуждения к развитию и эволюции новых метаболических путей посредством объединения направленного мутагенеза и эволюции под специфичным давлением отбора (см., например, WO 2005/073364 или WO 2008/116852).
Микроорганизм может быть модифицирован для экспрессии экзогенных генов, если данные гены вводятся в микроорганизм со всеми элементами, обеспечивающими их экспрессию в микроорганизме-хозяине. Данная модификация или «трансформация» микроорганизмов экзогенной ДНК представляет собой рутинную задачу для специалистов в данной области.
Микроорганизм может быть модифицирован для модуляции уровня экспрессии эндогенного гена.
Термин «эндогенный ген» означает то, что ген присутствовал в микроорганизме до любой генетической модификации. Эндогенные гены могут сверхэкспрессироваться посредством введения гетерологичных последовательностей, помимо или для того, чтобы заменять эндогенные регуляторные элементы, или посредством введения одной или более чем одной дополнительной копии гена в хромосому или плазмиду. Эндогенные гены также могут быть модифицированы для модуляции их экспрессии и/или активности. Например, в кодирующую последовательность могут быть введены мутации для модификации генного продукта, или могут быть введены гетерологичные последовательности, помимо или для замены эндогенных регуляторных элементов. Модуляция эндогенного гена может приводить к повышающей регуляции и/или усилению активности генного продукта, или, в качестве альтернативы, к понижающей регуляции и/или уменьшению активности продукта эндогенного гена.
Другим способом модуляции их экспрессии является замена эндогенного промотора гена (например, промотора дикого типа) на более сильный или более слабый промотор для повышающей или понижающей регуляции экспрессии эдогенного гена. Данные промоторы могут быть гомологичными или гетерологичными. Выбор подходящих промоторов находится целиком в пределах способностей специалиста в данной области.
Наоборот, термин «экзогенный ген» означает то, что ген был введен в микроорганизм способами, хорошо известными специалисту в данной области, в то время как данный ген не встречается в природе в данном микроорганизме. Экзогенные гены могут быть интегрированы в хромосому хозяина, или могут быть экспрессированы внехромосомно плазмидами или векторами. В данной области хорошо известен целый ряд плазмид, которые отличаются в отношении их репликатора и числа их копий в клетке. Данные гены могут быть гомологичными. Термин «сверхэкспрессия» или «осуществление сверхэкспрессии» также используется для обозначения экспрессии экзогенных генов в микроорганизмах.
В контексте данного изобретения термин «гомологичный ген» не ограничивается обозначенными генами, имеющими теоретического общего генетического предка, но и включает гены, которые могут не быть родственными генетически, которые, тем не менее, эволюционировали так, что они кодируют белок, который выполняет аналогичные функции и/или имеет аналогичную структуру. Следовательно, термин «функциональный гомолог» для цели настоящего изобретения относится к тому факту, что определенная ферментативная активность может быть не только предоставлена специфическим белком с определенной аминокислотной последовательностью, но также и белками с аналогичной последовательностью из других (не)родственных микроорганизмов.
С использованием ссылок, приведенных в Uniprot для известного белка или в Genbank для известных генов, специалисты в данной области могут определять последовательности белка и/или гена и определять эквивалентные гены в других организмах, бактериальных штаммах, дрожжах, грибах, млекопитающих, растениях и т.д. Эта рутинная работа преимущественно осуществляется с использованием консенсусных последовательностей, которые могут быть определены путем проведения выравниваний последовательностей с генами, происходящими из других микроорганизмов, и разработки вырожденных зондов для клонирования соответствующего гена в другом организме. Эти традиционные методы молекулярной биологии хорошо известны специалистам в данной области.
Согласно данному изобретению термины «ферментативный процесс», «ферментация» или «культура» используются взаимозаменяемо для обозначения роста микроорганизма. Этот рост обычно проводится в ферментерах с подходящей ростовой средой, адаптированной к используемому микроорганизму.
Фраза «подходящая культуральная среда» обозначает среду (например, стерильные жидкие среды), содержащую незаменимые или полезные питательные вещества для поддержания и/или роста клетки, такие как источники углерода или углеродные субстраты, источники азота, например, пептон, дрожжевые экстракты, мясные экстракты, экстракты солода, мочевина, сульфат аммония, хлорид аммония, нитрат аммония и фосфат аммония; источники фосфора, например, монокалия фосфат или дикалия фосфат; микроэлементы (например, соли металлов), например, соли магния, соли кобальта и/или соли марганца, а также факторы роста, такие как аминокислоты и витамины.
Термин «источник углерода», «углеродный источник» или «углеродный субстрат» согласно настоящему изобретению относится к любому углеродному источнику, способному к метаболизации микроорганизмом, где данный субстрат содержит по меньшей мере один атом углерода.
Термин «углевод» относится к любому углеродному источнику, способному к метаболизации микроорганизмом и содержащему по меньшей мере один атом углерода, два атома водорода и один атом кислорода. CO2 не является углеводом, так как он не содержит водород.
Углевод выбран среди группы, состоящей из моносахаридов, таких как глюкоза, фруктоза, манноза, ксилоза, арабиноза, галактоза и тому подобные, дисахаридов, таких как сахароза, целлобиоза, мальтоза, лактоза и тому подобные, олигосахаридов, таких как раффиноза, стахиоза, мальтодекстрины и тому подобные, полисахаридов, таких как целлюлоза, гемицеллюлоза, крахмал и тому подобные, метанола, формальдегида и глицерина. Особенно предпочтительными углеродными источниками являются арабиноза, фруктоза, галактоза, глюкоза, лактоза, мальтоза, сахароза, ксилоза или их смесь. Более предпочтительно, углеродным источником является сахароза.
В конкретном воплощении изобретения углеродный источник происходит из возобновляемого сырья. Возобновляемое сырье определяется как сырье, требующееся для определенных промышленных процессов, которое может быть регенерировано в пределах короткой задержки и в достаточном количестве для обеспечения его превращения в желательный продукт. Растительная биомасса, обработанная или необработанная, представляет собой интересный возобновляемый углеродный источник.
Специалисты в данной области способны определять культуральные условия для микроорганизмов согласно изобретению. В частности бактерии осуществляют ферментацию при температуре от 20°С до 55°С, предпочтительно от 25°С до 40°С и более конкретно - при примерно 30°С-37°С для Е. coli.
Данный процесс может проводиться либо в периодическом способе, в способе с подпиткой или в непрерывном способе. Он может проводиться при аэробных, микроаэробных или анаэробных условиях.
Фраза «при аэробных условиях» означает то, что в культуру предоставляется кислород посредством растворения газа в жидкой фазе. Это могло бы быть получено (1) барботированием кислородсодержащего газа (например, воздуха) в жидкую фазу или (2) встряхиванием сосуда, содержащего культуральную среду, для того, чтобы перенести кислород, содержащийся в свободном объеме над жидкостью, в жидкую фазу. Главным преимуществом ферментации при аэробных условиях является то, что присутствие кислорода в качестве акцептора электронов улучшает способность штамма продуцировать больше энергии в форме АТФ для клеточных процессов. Следовательно, данный штамм имеет улучшенный общий метаболизм.
Микроаэробные условия определяются как культуральные условия, при которых в жидкой фазе растворены низкие процентные концентрации кислорода (например, с использованием смеси газа, содержащей от 0,1 до 10% кислорода, дополненной до 100% азотом).
Анаэробные условия определяются как культуральные условия, при которых в культуральную среду не предоставляется кислород. Строго анаэробные условия получают барботированием инертного газа, подобного азоту, в культуральную среду для удаления следов другого газа. Нитрат можно использовать в качестве акцептора электронов для улучшения продукции АТФ штаммом и улучшения его метаболизма.
Фраза «выделение 1,2-пропандиола из культуральной среды» обозначает процесс очистки продуцированного 1,2-пропандиола с использованием способов, известных специалисту в данной области. Такие способы, в особенности, раскрыты в патентных заявках WO 2011/076690 и WO 2012/130316.
Термины «микроорганизм, генетически модифицированный для продукции 1,2-пропандиола» относится к микроорганизмам, модифицированным либо посредством введения, либо делеции генетических элементов, либо посредством этапа эволюции, как описано в патентной заявке WO 2005/073364. В частности, он обозначает генетически модифицированный микроорганизм, представляющий улучшенную продукцию 1,2-пропандиола, по сравнению с эндогенной продукцией соответствующего микроорганизма дикого типа, без генетических модификаций. Такой микроорганизм, например, описан в патентных заявках WO 2008/116848, WO 2008/116853, WO 2011/012693, WO 2011/012697, WO 2011/012702 или ЕР 2532751, включенных посредством ссылки. Предпочтительными генетическими модификациями являются следующие:
- сверхэкспрессия по меньшей мере одного гена, выбранного среди гена mgsA, кодирующего метилглиоксальсинтазу, генов yqhD, yafB, ycdN, yqhE, yeaE, yghZ, yajO, tas, ydjG или ydbC, кодирующих метилглиоксальредуктазу, гена gldA, кодирующего глицеролдегидрогеназу, и гена fucO, кодирующего лактальдегидредуктазу;
- делеция либо гена edd, кодирующего фосфоглюконатдегидратазу, либо гена eda, кодирующего 2-кето-3-дезоксиглюконат-6-фосфатальдолазу, либо обоих;
- аттенуация синтеза нежелательных побочных продуктов посредством делеции генов, кодиующих ферменты, участвующие в синтезе лактата из метилглиоксаля (таких как ген gloA, кодирующий глиоксалазу I, ген aldA, кодирующий альдегиддегидрогеназу А, ген aldB, кодирующий ацетальдегиддегидрогеназу), лактата из пирувата (ген IdhA, кодирующий лактатдегидрогеназу), формиата (ген pflA, кодирующий пируватформиатлиазу, ген pflB, кодирующий пируватформиатлиазу), этанола (ген adhE, кодирующий альдегидалкогольдегдрогеназу) и ацетата (ген ackA, кодирующий ацетаткиназу, ген pta, кодирующий фосфатацетилтрансферазу, ген рохВ, кодирующий пируватоксидазу);
- устранение путей, потребляющих ФЕП (фосфоенолпируват), подобных пируваткиназам (кодируемым генами pykA и pykF), и/или стимуляция синтеза ФЕП, например, посредством сверхэкспрессии гена ppsA, кодирующего ФЕП-синтазу;
- специфическая мутация гена lpd, кодирующего липоамиддегидрогеназу;
- ген arcA, кодирующий транскрипционный двойной регуятор ArcA, и ген ndh, кодирующий НАДН: убихиноноксидоредуктазу II, могут быть подвергнуты делеции,
- ген gapA, кодирующий глицеральдегид-3-фосфатдегдрогеназу, находится под контролем температуроиндуцибельного промотора,
- гены, участвующие в импорте и метаболизме сахарозы (ген cscB, кодирующий пермеазу сахарозы, ген cscA, кодирующий гидролазу сахарозы, ген cscK, кодирующий фруктокиназу, ген scrA, кодирующий Фермент II фосфоенолпируват-зависимой фосфотрансферазной системы, ген scrK, кодирующий АТФ-зависимую фруктокиназу, ген scrB, кодирующий сахарозо-6-фосфатгидролазу (инвертазу), ген scrY, кодирующей порин сахарозы), добавляются, или их экспрессия повышается.
Предпочтительной генетической модификацией является улучшение активности метилглиоксальредуктазы, полученное посредством сверхэкспрессии гена yqhD* (G149E).
Другой предпочтительной генетической модификацией является улучшение метилглиоксальсинтазной активности, полученное посредством сверхэкспрессии гена msgA*(H21Q).
Другой предпочтительной генетической модификацией является улучшение активности гидроксиацетон редуктазы, полученное посредством сверхэкспрессии гена gldA*(A160T). Данный мутант ГАР является НАДН-зависимым, в отличие от НАДФН-зависимого мутанта ГАР, раскрытого далее.
Гидроксиацетон редуктаза (ГАР) является последним ферментом, участвующим в продукции 1,2-пропандиола. ГАР катализирует следующую реакцию:
Гидроксиацетон +НАД(Ф)Н → 1,2-пропандиол + НАД(Ф)+
Штамм, продуцирующий 1,2-пропандиол, в котором ген gldA подвергнут делеции, больше не способен продуцировать 1,2-пропандиол и накапливает гидроксиацетон (WO 2008/116851). Белок GldA был очищен до гомогенности, и он использует НАДН в качестве кофактора (Kelley and Dekker, 1984). При аэробных условиях превращение гидроксиацетона до 1,2-пропандиола является не полным из-за внутреннего окислительно-восстановительного состояния клетки: при таких условиях первичной ролью НАДН является дыхательная генерация АТФ через окислительное фосфорилирование, и в результате этого отношение НАДН к НАД+ сильно сдвинуто в пользу НАД+. Химически очень сходный НАДФН, в отличие от этого, управляет анаболическими восстановлениями, и отношение НАДФН к НАДФ+ выше (Fuhrer and Sauer, 2009).
Следуя данному наблюдению, целью данного изобретения является улучшение продукции 1,2-пропандиола посредством увеличения активности НАДФН-зависимой гидроксиацетон редуктазы.
Увеличение активности можно получать улучшением каталитической эффективности белка или уменьшением обмена белка, или уменьшением обмена матричной РНК (мРНК), или усилением транскрипции гена, или усилением трансляции мРНК.
Улучшение каталитической эффективности белка означает увеличение kcat и/или снижение Km в отношении данного субстрата и/или данного кофактора, и/или увеличение Ki в отношении данного ингибитора, kcat, Km и Ki представляют собой константы Михаэлиса-Ментен, которые способен определить специалист в данной области (Segel, 1993). Уменьшение обмена белка означает стабилизирование белка. Способы улучшения каталитической эффективности белка и/или уменьшения обмена белка хорошо известны специалисту в данной области. Они включают рациональное конструирование последовательности и/или структурный анализ и направленный мутагенез, а также случайный мутагенез и скрининг. Мутации могут вводиться сайт-направленным мутагенезом обычными способами, подобными полимеразной цепной реакции (ПЦР), или посредством методик случайного мутагенеза, таких как применение мутагенных агентов (ультрафиолетовые лучи или химические агенты, подобные нитрозогуанидину (NTG, от англ. nitrosoguanidine) или этилметансульфонату (EMS, от англ. ethylmethanesulfonate)) или применением методик ПЦР (перетасовка ДНК или ПЦР, склонная к ошибкам). Стабилизирование белка также может достигаться добавлением пептидной последовательности, именуемой «метка», либо на N-конце, либо на С-конце белка. Метки хорошо известны специалисту в данной области. Например, для стабилизации белка может быть использована глутатион-S-трансфераза (GST, от англ. Glutathione-S-Transferase).
Уменьшение обмена мРНК может достигаться модифицированием последовательности 5'-нетранслируемой области (5'-UTR) и/или кодирующей области, и/или 3'-UTR гена (Carrier and Keasling, 1999).
Усиление транскрипции гена может достигаться увеличением числа копий гена и/или применением промотора, приводящего к более высокому уровню экспрессии гена. Термин «сверхэкспрессия» или «сверхэкспрессирование» также используется для обозначения усиления транскрипции гена в микроорганизмах.
Для увеличения числа копий гена в микроорганизме данный ген кодируется хромосомно или внехромосомно. При локализации гена на хромосоме несколько копий гена могут вводиться на хромосому способами рекомбинации, известными эксперту в данной области (включая замену гена). При внехромосомной локализации гена, оно может осуществляться посредством разных типов плазмид, которые отличаются в отношении их репликаторов и, таким образом, их числа копий в клетке. Данные плазмиды присутствуют в микроорганизме в 1-5 копиях или примерно в 20 копиях, или вплоть до 500 копий, в зависимости от природы плазмиды: плазмиды с малым числом копий с ограниченной репликацией (англ. tight replication) (например, pSC101, RK2 Е. coli), плазмиды с малым числом копий (например, pACYC, pRSF1010 Е. coli) или плазмиды с большим числом копий (например, pSK bluescript II Е. coli).
В том, что касается применения промотора, приводящего к высокому уровню экспрессии гена, специалист в данной области знает то, какие промоторы являются самыми удобными, например, широко используются промоторы Ptrc, Ptac, Plac или промотор cl лямбда. Данные промоторы могут быть «индуцибельными» посредством конкретного соединения или специфического внешнего условия, подобного температуре или свету. Данные помоторы могут быть гомологичными или гетерологичными.
Усиление трансляции мРНК может достигаться модифицированием сайта связывания рибосом (RBS, от англ. ribosome binding site). RBS представляет собой последовательность на мРНК, которая связывается рибосомой при инициации трансляции белка. Он может представлять собой либо 5'-кэп мРНК у эукариот, область в 6-7 нуклеотидах выше инициирующего кодона AUG у прокариот (именуемая последовательность Шайна-Дальгарно) или участок внутренней посадки рибосомы (IRES, от англ. internal ribosome entry site) у вирусов. Посредством модифицирования данной последовательности возможно изменять скорость инициации трансляции белка, пропорционально изменять его скорость продукции и контролировать его активность в клетке. Такая же последовательность RBS не будет иметь такого же влияния, согласно природе мРНК. Возможно оптимизировать силу последовательности RBS для достижения намеченной скорости инициации трансляции посредством применения программы RBS CALCULATOR (Salis, 2011).
В первом аспекте настоящего изобретения продукция 1,2-пропандиола улучшается усилением активности НАДФН-зависимой ацетолредуктазы (НАДФН ГАР) в модифицированном микроорганизме.
В первом воплощении изобретения активность НАДФН ГАР увеличивается посредством сверхэкспрессии гена, кодирующего НАДФН ГАР (ген НАДФН ГАР), в модифицированном микроорганизме.
Adh из Clostridium beijerinckii катализирует в природе восстановление ацетона до изопропанола (Ismaiel et al., 1993). Было показано, что данный фермент, а также его гомолог из Thermoanaerobacter brockii катализируют восстановление гидроксиацетона до 1,2-пропандиола, и они были сверхэкспрессированы в облигатном фотоавтотрофе цианобактерии Synechococcus elongatus с получением 1,2-пропандиола из CO2 в качестве источника углерода (Li and Liao, 2013).
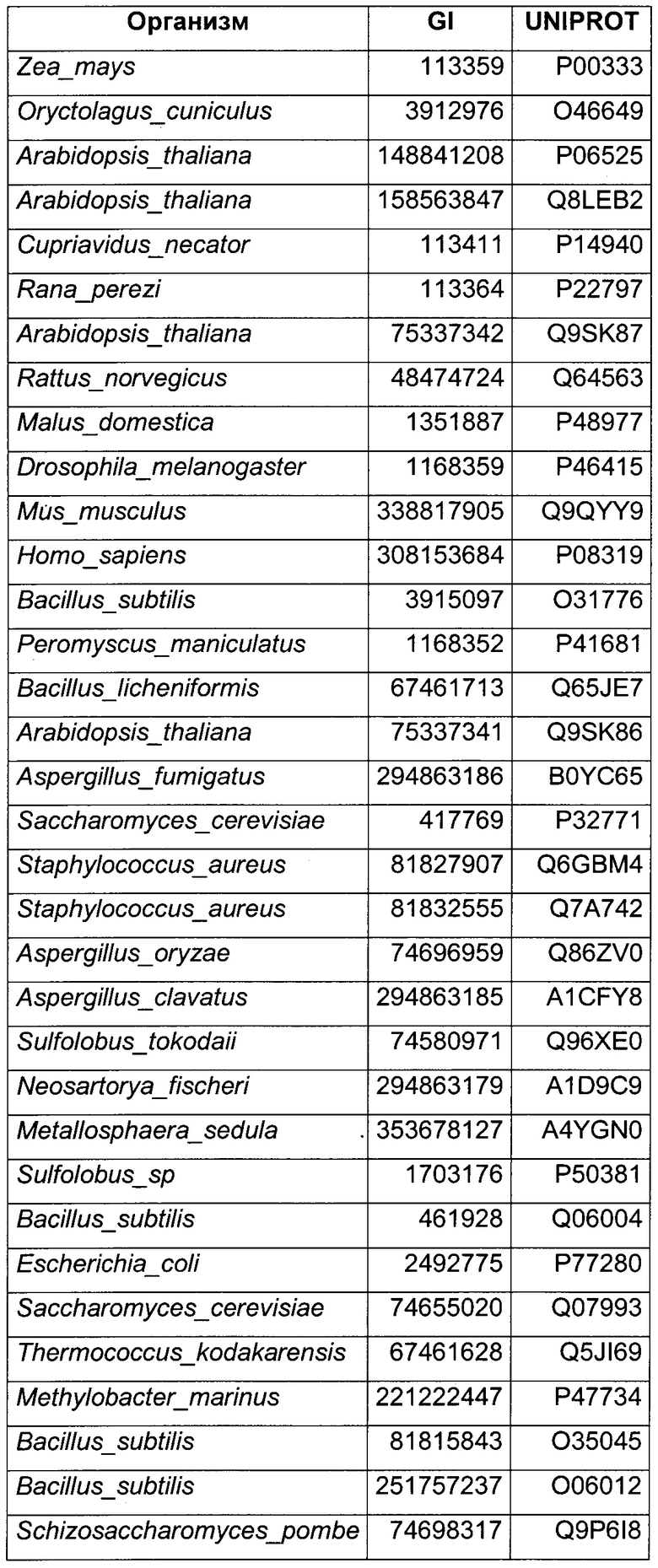
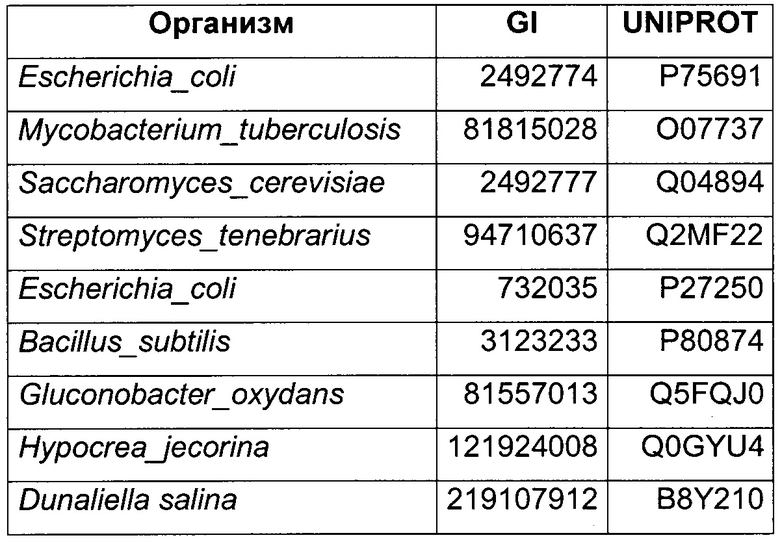
Использовали анализ данных и анализ последовательностей для определения некоторых гомологичных ферментов-кандидатов и генов, кодирующих такие ферменты. Данные кандидаты раскрыты в Таблице 1 ниже.
Средства идентификации гомологичных последовательностей и их процентные доли гомологии хорошо известны специалистам в данной области и включают, в частности, программы BLAST. Полученные последовательности можно затем использовать (например, выравнивать), используя, например, программы CLUSTALW или MULTALIN. Другой способ идентификации белковых гомологов осуществляется через конструирование филогенетческих деревьев с использованием программ биоинформатики, хорошо известных специалисту в данной области.
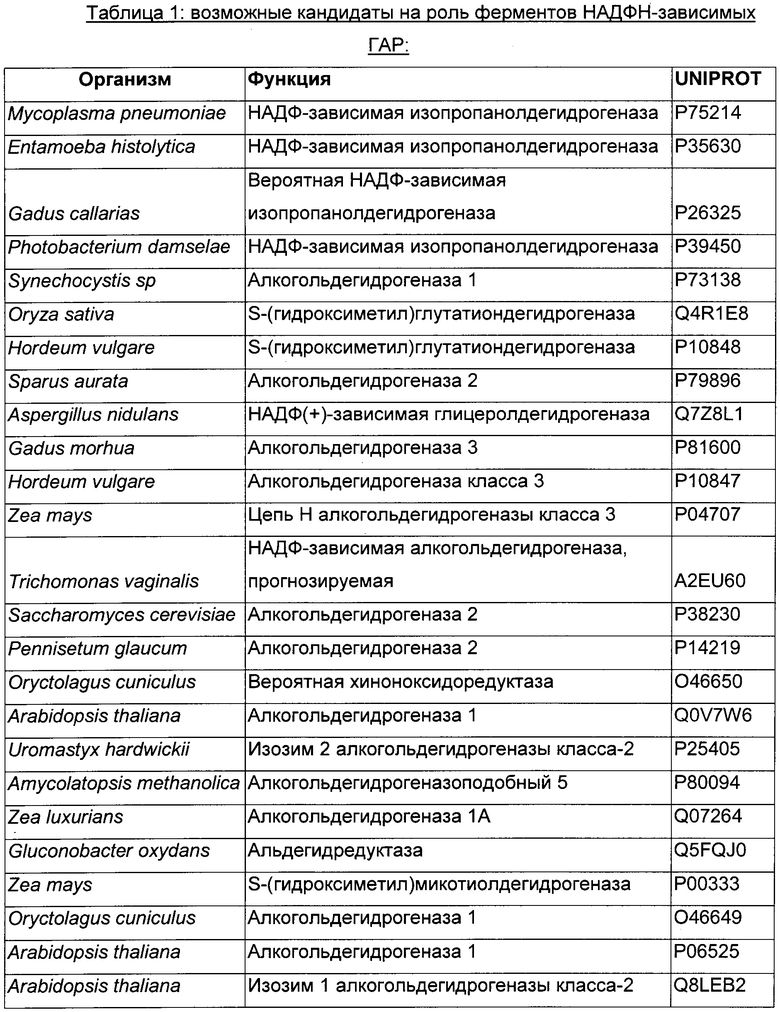
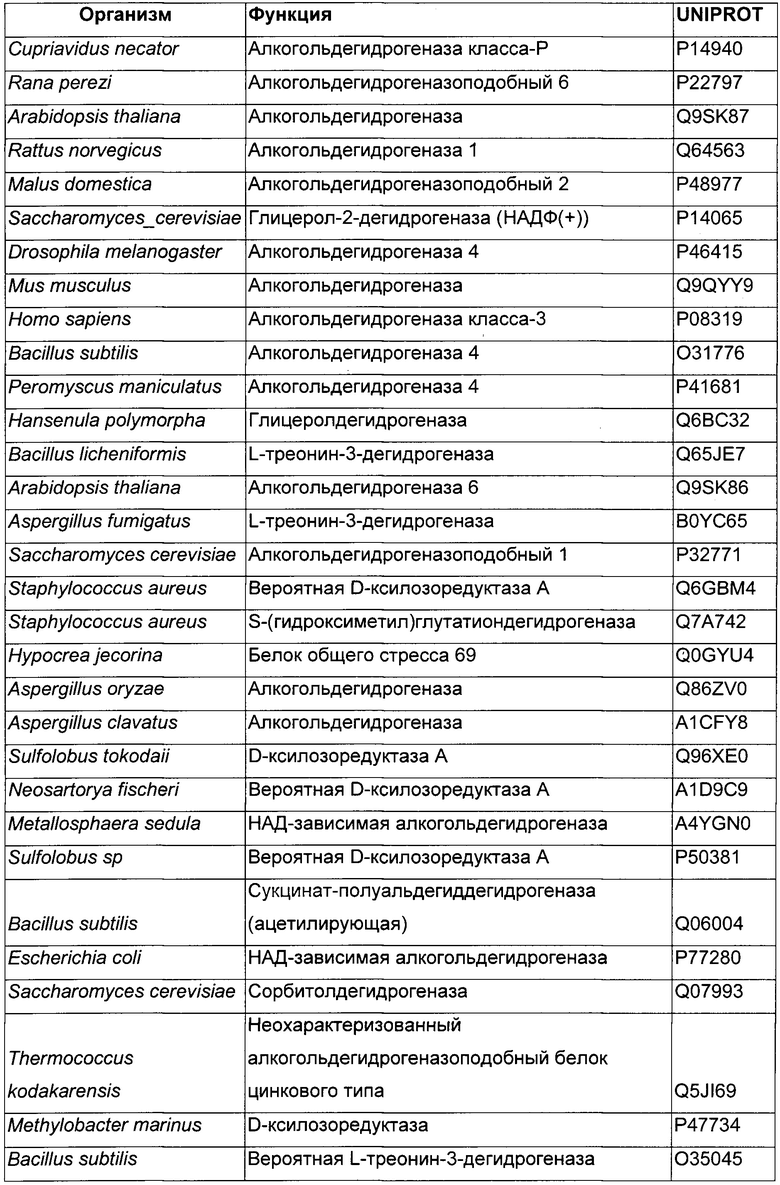
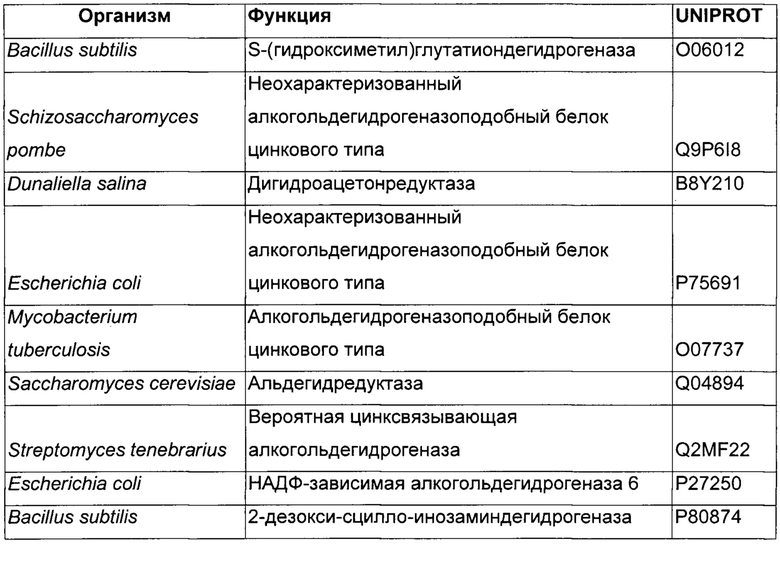
В данном воплощении изобретения ген, кодирующий НАДФН ГАР, сверхэкспрессируемую в рекомбинантном микроорганизме, кодирует НАДФН-зависимую ацетолредуктазу, выбранную среди белков, перечисленных выше. Из номера Uniprot специалист в данной области способен получить номер и последовательность соответствующего гена. Предпочтительно микроорганизм по изобретению экспрессирует по меньшей мере один ген, кодирующий НАДФН-зависимую ацетолредуктазу (ген НАДФН ГАР), имеющую по меньшей мере 60%-ную, предпочтительно по меньшей мере 70%-ную, более предпочтительно по меньшей мере 85%-ную и даже более предпочтительно по меньшей мере 90%-ную идентичность аминокислот с полным белком, кодируемым генами adh из Clostridium beijerinckii или adh из Thermoanaerobacter brockii, или adh1 из Entamoeba histolytica, или GOX1615 из Gluconobacter oxydans, или gld2 из Hypocrea jecorina, или yhdN из Bacillus subtilis. Более предпочтительно микроорганизм по изобретению экспресирует ген, выбранный среди adh из Clostridium beijerinckii или adh из Thermoanaerobacter brockii, или adh1 из Entamoeba histolytica, или GOX1615 из Gluconobacter oxydans, или gld2 из Hypocrea jecorina, или yhdN из Bacillus subtilis (т.е. 100%-ная идентичность аминокислотной последовательности с последовательностью белка, кодируемой указанными генами). Даже более предпочтительно, НАДФН-зависимая ацетолредуктаза кодируется геном adh из Clostridium beijerinckii.
Идентичность последовательности между аминокислотными последовательностями можно определять сравнением положения в каждой из последовательностей, которые могут быть выровнены для целей сравнения. Когда положение в сравниваемых последовательностях занято одинаковой аминокислотой, тогда последовательности являются идентичными в данном положении. Степень идентичности последовательности между белками является функцией числа идентичных аминокислотных остатков в аналогичных положениях, которые имеют последовательности указанных белков.
Для определения процентной доли идентичности между двумя аминокислотными последовательностями данные последовательности выравнивают для оптимального сравнения. Например, в последовательность первой аминокислотной последовательности можно вводить пробелы для оптимального выравнивания со второй аминокислотной последовательностью. Затем сравнивают аминокислотные в соответствующих положениях аминокислот. Когда положение в первой последовательности занято таким же аминокислотным остатком, что и соответствующее положение во второй последовательности, молекулы являются идентичными в данном положении.
Процентная доля идентичности между двумя последовательностями является функцией числа идентичных положений, которые имеют последовательности. Следовательно, % идентичности равен числу индентичных положений, поделенному на общее число перекрывающихся положений, с умножением на 100.
Оптимальное выравнивание последовательностей можно проводить посредством алгоритма выравнивания глобальной гомологии Needleman и Wunsch (1970), посредством компьютеризированных воплощений данного алгоритма или посредством визуальной проверки. Выбирают лучшее выравнивание (т.е. приводящее к самой высокой процентной доле идентичности между сравниваемыми последовательностями), полученное разными способами.
Другими словами, процентную долю идентичности последовательности рассчитывают посредством сравнения двух оптимально выровненных последовательностей, определяя число положений, в которых в обеих последовательностях встречаются идентичные аминокислоты, с получением числа совпадающих положений, деля число совпадающих положений на общее число положений и умножая результат на 100, с получением процентной доли идентичности последовательности.
В другом предпочтительном воплощении изобретения активность НАДФН ГАР усиливается посредством увеличения НАДФН-зависимой активности ГАР, как описано выше, и/или посредством уменьшения НАДН-зависимой активности ГАР.
Уменьшение активности фермента означает или уменьшение его удельной каталитической активности посредством мутирования гена таким образом, чтобы изменить аминокислотную последовательность, и/или уменьшение концентраций белка в клетке посредством мутирования нуклеотидной последовательности, или посредством делеции кодирующей области гена.
В настоящем изобретении уменьшение НАДН-зависимой активности ГАР осуществляли посредством делеции гена gldA или посредством осуществления инженерии кофактора на ферменте GldA, что приводило как к снижению НАДН-зависимой активности ГАР, так и к увеличению НАДФН-зависимой активности ГАР.
Следовательно, микроорганизм по изобретению предпочтительно сверхэкспрессирует мутантный ген gldA*, кодирующий НАДФН-зависимую глицеролдегидрогеназу, и/или имеет делецию эндогенного гена gldA.
Термин «инженерия кофактора» означает изменение специфичности фермента в отношении кофактора посредством замены некоторых специфических аминокислотных остатков в белке GldA. Данные полученные в результате мутанты (GldA* НАДФН) являются НАДФН-зависимыми и отличаются от мутанта GldA* (GldA* А160Т), раскрытого выше, который является НАДН-зависимым. Остатки, управляющие специфичностью в отношении кофактора в кармане связывания кофактора, известного как «складка Россмана», были тщательно исследованы (Scrutton et al., 1990; Clermont et al., 1993; Corbier et al., 1990; Wu et al., 2012), и специалист в данной области способен определять, какие аминокислотные остатки подлежат модификации для того, чтобы изменять специфичность в отношении кофактора. Инженерия кофактора недавно оказалась успешной в изменении ферментов для того, чтобы они предпочтитали НАДН в качестве кофактора, вместо НАДФН. Примером является фермент Gre2p, дегидрогеназа, предпочитающая НАДФН, обнаруженная у Saccharomyces cerevisiae, которая была модифицирована прямым мутагенезом так, чтобы иметь пониженную зависимость от НАДФН и повышенную аффинность в отношении НАДН (Katzberg et al., 2010).
В настоящем изобретении из трехмерной структуры фермента из Bacillus stearothermophilus была построена трехмерная структурная модель GldA из Е. coli (Ruzheinikov et ai, 2001). Специалист в данной области знает, как строить трехмерные модели гомологии с использованием, например, такой программы, как Discovery Studio (Accelrys). Такие модели являются полезными для определения того, какие аминокислотные остатки подлежат мутированию для того, чтобы добиваться желательного изменения спецфичности в отношении кофактора. Из трехмерной структуры GldA из Е. coli авторы изобретения идентифицировали аминокислотные остатки, подлежащие замене другим аминокислотным остатком в том же самом положении:
- D37
- F39
- V40
- F43
- Т116
- Р161
- L164
Согласно данному изобретению изменение специфичности GldA в отношении кофактора опосредуется по меньшей мере одной мутацией в положении D37. В предпочтительном воплощении аминокислотный остаток в положении D37 заменяется глицином (D37G), аланином (D37A) или валином (D37V). В наиболее предпочтительном воплощении аминокислотный остаток в положении D37 заменяется глицином (D37G).
В предпочтительном воплощении изменение специфичности GldA в отношении кофактора улучшается объединением мутации в положении D37 с по меньшей мере одной мутацией в положении Р161. Предпочтительно аминокислотный остаток в положении Р161 заменяется серином (P161S) или треонином (Р161Т). Более предпочтительно аминокислотный остаток в положении Р161 заменяется серином (P161S).
В наиболее предпочтительном воплощении изменение специфичности GldA в отношении кофактора улучшается объединением мутаций в положениях D37 и Р161 с по меньшей мере одной мутацией в положении L164. Предпочтительно аминокислотный остаток в положении L164 заменяется аланином (L164A), глицином (L164G) или валином (L164V). Более предпочтительно аминокислотный остаток в положении L164 заменяется аланином (L164A).
В конкретном воплощении микроорганизм по изобретению сверхэкспрессирует мутанта gldA*, кодирующего мутанта GldA*, содержащего по меньшей мере следующие мутации: D37G, P161S и L164A.
В предпочтительном воплощении изобретения данные мутации могут вводиться в мутанта GldA*, сконструированного ранее и содержащего мутацию А160Т.
Аминокислотная последовательность глицеролдегидроеназы, экспрессируемой в Е. coli (штамм K12) является общедоступной в базе данных UniprotKB под ссылкой P0A9S5.
Следовательно, для того, чтобы усилить НАДФН ГАР-зависимую активность, микроорганизм по изобретению предпочтительно сверхэкспрессирует по меньшей мере один ген, кодирующий НАДФН-зависимую ацетолредуктазу (НАДФН ГАР), как описано выше, или мутантный ген gldA*, кодирующий НАДФН-зависимую глицеролдегидрогеназу, как описано выше.
В других конкретных воплощениях изобретения модифицированный микроорганизм по изобретению может:
- сверхэкспрессировать по меньшей мере один ген НАДФН ГАР и может иметь делецию гена gldA,
- сверхэкспрессировать по меньшей мере один ген НАДФН ГАР и мутанта gldA* НАДФН или
- сверхэкспрессировать по меньшей мере один ген НАДФН ГАР и мутанта gldA* НАДФН, и может иметь делецию гена gldA.
В другом аспекте изобретения продукция 1,2-пропандиола дополнительно улучшается объединением увеличения активности НАДФН-зависимой ГАР, как описано выше, с увеличением доступности НАДФН в клетке.
Стратегии увеличения доступности НАДФН в клетке являются хорошо известными от специалистов в данной области (обзор дается в Lee et al., 2013). В настоящем изобретении доступность НАДФН в клетке увеличивается посредством:
- сверхэкспрессии оперона pntAB, кодирующего мембраносвязанную трансгидрогеназу (WO 2012/055798A1), и/или
- аттенуация гена pgi, кодирующего фосфоглюкозоизомеразу, и/или
- увеличения потока через пентозофосфатный путь посредством уменьшения активности фосфофруктокиназы, кодируемой геном pfkA (WO 2005/047498), и сверхэкспрессии гена zwf, кодирующего глюкозо-6-фосфатдегидрогеназу (Shin et al., 2002), и/или
- сверхэкспрессии гена gapN из Streptococcus mutans, кодирующего глицеральдегид-3-фосфат, генерирующую НАДФН (Centeno-Leija et al., 2013),
- и/или сверхэкспрессии мутантного гена Ipd*, кодирующего липоамиддегидрогеназу, способную генерировать НАДФН (Bocanegra et al., 1993),
- и/или сверхэкспресии гена yjeF, кодирующего АДФ-зависимую дегидратазу, реактивирующую НАДН(Х) и НАДФН(Х), продуцируемые ферментативной или зависимой от нагревания гидратацией (Marbaix et al., 2011).
В другом аспекте изобретения при сверхэкспрессии гена НАДФН ГАР микроорганизмом по изобретению ген yqhD, кодирующий метилглиоксальредуктазу (МГР), был подвергнут делеции. При таких условиях белок ГАР действует не только как НАДФН-зависимый фермент ГАР, но также как и НАДФН-зависимый фермент МГР, выполняя двухэтапное НАДФН-зависимое восстановление метилглиоксаля в 1,2-пропандиол. Такая активность может осуществляться любым из кандидатов на роль ферментов НАДФН-зависимых ГАР, которые были перечислены выше.
В более предпочтительном воплощении изобретения продукция 1,2-пропандиола из углевода в качестве источника углерода в ферментативном процессе, осуществляемом рекомбинантным микроорганизмом, где активность НАДФН-зависимой ацетолредуктазы усиливается согласно любой из модификаций, описанных выше, может достигаться посредством комбинации обсуждавшихся выше модификаций в указанном микроорганизме, например, следующих:
- экспрессии по меньшей мере одного гена НАДФН ГАР и генов pntAB повышаются,
- экспрессия по меньшей мере одного гена НАДФН ГАР повышается, и ген gldA подвергается делеции,
- экспрессии по меньшей мере одного гена НАДФН ГАР и pntAB повышаются, тогда как ген gldA подвергается делеции,
- экспрессии по меньшей мере одного гена НАДФН ГАР и yjeF повышаются, тогда как ген gldA подвергается делеции,
- экспрессии по меньшей мере одного гена НАДФН ГАР повышаются, pgi ослабляется, тогда как ген gldA подвергается делеции,
- экспрессии по меньшей мере одного гена НАДФН ГАР и zwf повышаются, тогда как ген gldA и ген pfkA ослабляются,
- экспрессии по меньшей мере одного гена НАДФН ГАР и gapN повышаются, тогда как ген gldA подвергается делеции,
- экспрессии по меньшей мере одного гена НАДФН ГАР и мутанта Ipd* повышаются, тогда как ген gldA подвергается делеции,
- экспрессия гена, кодирующего мутантную GldA* НАДФН, повышается, и ген gldA подвергается делеции,
- экспрессии гена, кодирующего мутантную GldA* НАДФН, и генов pntAB повышаются,
- экспрессии гена, кодирующего мутантную GldA* НАДФН, и генов pntAB повышаются, тогда как ген gldA подвергается делеции,
- экспрессия по меньшей мере одного гена НАДФН ГАР повышается, ген yqhD ослабляется, тогда как ген gldA подвергается делеции,
- экспрессии по меньшей мере одного гена НАДФН ГАР и генов pntAB повышаются, тогда как ген yqhD и ген gldA подвергаются делеции.
ПРИМЕРЫ
ПРИМЕР 1: способы
В приведенных ниже примерах используются хорошо известные в данной области способы для конструирования штаммов E. coli, содержащих реплицирующиеся векторы и/или разные хромосомные делеции и замены, с использованием гомологичной рекомбинации, хорошо описанной для Escherichia coli Datsenko & Wanner (2000). Таким же образом, применение плазмид или векторов для экспрессии или сверхэкспрессии одного или нескольких генов в рекомбинантных микроорганизмах является хорошо известным для специалиста в данной области. Примеры подходящих экспрессионных векторов Е. coli включают pTrc, pACYC184n pBR322, pUC18, pUC19, pKC30, pRep4, pHS1, pHS2, pPLc236 и т.д.
В следующих примерах использовали несколько протоколов. Протокол 1 (хромосомные модификации посредством гомологичной рекомбинации, отбор рекомбинантов), протокол 2 (трансдукция фага Р1) и протокол 3 (вырезание кассеты устойчивости к антибиотику, удаление, при необходимости, генов устойчивости), использованные в данном изобретении, были полностью описаны в патентной заявке ЕР 2532751. Хромосомные модификации были подтверждены анализом ПЦР с использованием подходящих олигонуклеотидов, которые способен сконструировать специалист в данной области.
Протокол 4: конструирование рекомбинантных плазмид
Методика генной инженерии хорошо описана в данной области. Фрагменты ДНК и выбранную плазмиду расщепляли совместимыми рестрикционными ферментами (которые способен определить специалист в данной области), затем лигировали и трансформировали в компетентные клетки. Анализировали трансформантов, и интересующие рекомбинантные плазмиды подтверждали секвенированием ДНК.



Протокол 5: оценка штаммов, продуцирующих 1,2-пропандиол
Штаммы, продуцирующие 1,2-пропандиол, культивировали в культурах в колбах, как описано в патентной заявке ЕР 2532751, за исключением того, что использовали 20 г/л глюкозы или сахарозы и 40 г/л MOPS (3-(N-морфолин)пропансульфоновая кислота). При необходимости в среду добавляли 100 мкМ IPTG (изопропилтиогалактозид). 1,2-пропандиол (PG, от англ. propylene glycol) и гидроксиацетон (НА) количественно измеряли посредством ВЭЖХ (высокоэффективная жидкостная хроматография). Продукция PG (г PG/л) и превращение НА до PG (г PG/л/(г НА/л + г PG/л)) дает оценку эффективности штамма для продукции 1,2-пропандиола.
Протокол 6: культуры в колбах для продукции рекомбинантных белков
Культуры в колбах для продукции рекомбинантных белков осуществляли, как описано в патентной заявке WO 2010/076324, за исключнием того, что бульон LB дополняли 5,0 г/л глюкозы.
ПРИМЕР 2: конструирование штаммов 1, 2, 3 и 4
Конструирование штамма 1
Для инактивации комплекса фумаратредуктазы с флавопротеином, кодируемого опероном frdABCD, и фермента глюкозофосфотрансферазы IIBC(Glc), кодируемого геном ptsG, использовали стратегию гомологичной рекомбинации (согласно Протоколам 1 и 3). Для ПЦР-амплификации кассет устойчивости использовали олигонуклеотиды DfrdABCD: SEQ ID NO 1 и 2 (перечисленные в Таблице 2) и DptsG: SEQ ID NO 3 и 4 (перечисленные в Таблице 2). Сохранившиеся штаммы обозначали MG1655 DfrdABCD::Cm и MG1655 DptsG::Km. Наконец, делеции DfrdABCD::Cm и DptsG::Km переносили поседством трансдукции фага Р1 (согласно Протоколу 2) в выработанный штамм MG1655 lpd* DtpiA DpflAB DadhE DldhA DgloA DaldA DaldB Dedd DarcA Dndh, описанный в патентной заявке WO2008/116852, с получением штамма 1.
Конструирование штамма 2
Ген gldA*(A160T) клонировали в плазмиду pME101VB06, как описано в патентной заявке ЕР 2532751. Данную плазмиду, названную pPG0078, трансформировали в штамм 1, получая штамм 2.
Конструирование штамма 3
Для экспрессии триозофосфатизомеразы, кодируемой геном tpiA, и для регуляции экспрессии глицеральдегидфосфатдегидрогеназы, кодируемой геном gapA, использовали стратегию гомологичной рекомбинации (согласно Протоколам 1 и 3). Ген tpiA вводили, как описано в патенте WO 2008/116852, в выработанный штамм MG1655 lpd* DtpiA DpflAB DadhE DldhA DgloA DaldA DaldB Dedd DarcA Dndh DfrdABCD. Затем в предыдущий штамм вводили геномную модификацию «СI857-PR01/RBS11-gapA» для регуляции экспрессии gapА, как описано в патенте ЕР 2532751, с получением штамма 3.
Конструирование штамма 4
Для обеспечения роста Escherichia coli на сахарозе, гены scrK, scrYAB и scrR из плазмиды pUR400 (Schmid et al., 1982) клонировали под контролем их природных промоторов на плазмиде pBBR1MCS3. Данную плазмиду называли pPG0231. Плазмиды pPG0078 и pPG0231 трансформировали в штамм 3 с получением штамма 4.
ПРИМЕР 3: продукция 1,2-пропандиола улучшается в штаммах, сверхэкспрессирующих дегидрогеназу вторичного спирта adh из Clostridium beijerinckii
Конструирование штамма 5
Ген adh из Clostridium beijerinckii (Hanai et al., 2007) клонировали в плазмиду pME101VB01, описанную в патентной заявке WO 2008/116853. Данную плазмиду, названную pPG0468, трансформировали в штамм 1, с получением штамма 5.
Конструирование штамма 6
Для инактивации гена gldA использовали стратегию гомологичной рекомбинации (согласно Протоколам 1 и 3). Олигонуклеотиды для DgldA: SEQ ID NO 5 и 6 (перечисленные в Таблице 2) использовали для ПЦР-амплификации кассеты устойчивости. Сохранившийся штамм обозначали MG1655 DgldA::Cm. Делецию DgldA::Cm переносили в штамм 5 посредством трансдукции фага Р1 (согласно Протоколу 2), с получением штамма 6.
Конструирование штамма 7
Плазмиды pPG0468 и pPG0231 трансформировали в штамм 3 с получением штамма 7.
Конструирование штамма 8
Для инактивации гена gldA ранее описанную делецию DgldA::Cm переносили в штамм 7 посредством трансдукции фага Р1 (согласно Протоколу 2), с получением штамма 8.
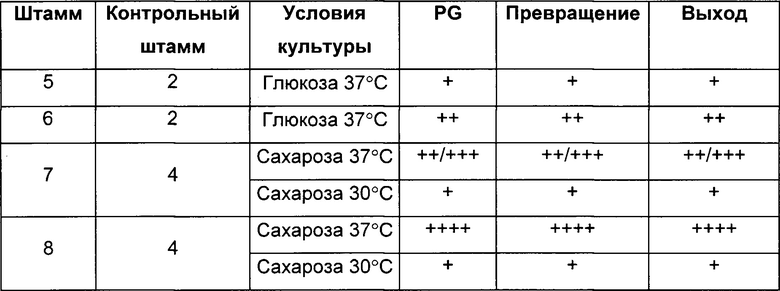
Штаммы 5, 6, 7 и 8, экспрессирующие adh из Clostridium beijerinckii, оцениваемые, как описано в Протоколе 5, продуцировали больше 1,2-пропандиола (PG) и имели лучшие показатели превращения и выходы по сравнению с соответствующими им контрольными штаммами 2 и 4 (Таблица 3).

(Символ ~ показывает то, что отсутствует значимое различие между штаммами, символ + показывает увеличение эффективности от 10 до 100% по сравнению с контрольным штаммом, символ ++ показывает увеличение от 100 до 200% по сравнению с контрольным штаммом, символ +++ показывает увеличение от 200 до 300% по сравнению с контрольным штаммом, и символ ++++ показывает увеличение более чем на 300% по сравнению с контрольным штаммом):
ПРИМЕР 4: продукция 1,2-пропандиола улучшается посредством сверхэкспрессии трансгидрогеназы пиридиновых нуклеотидов pntAB в штаммах, сверхэкспрессирующих дегидрогеназу вторичного спирта adh из Clostridium beijerinckii
Конструирование штамма 9
Природную промоторную область pntAB заменяли индуцибельным промотором trc (из плазмиды pTRC99A, Amersham Pharmacia) и определенным сайтом связывания рибосомы RBS120 (из программы RBS Calculator) (SEQ ID NO 7, перечисленная в Таблице 2), используя стратегию гомологичной рекомбинации (согласно Протоколам 1 и 3). Для хромосомной интеграции фрагмент, несущий искусственную промоторную область и маркер устойчивости, которые оба фланкированы гомологичными последовательностями ДНК к целевому локусу интеграции pntAB, ПЦР-амплифицировали посредством методики перекрывающейся ПЦР (перекрывающиеся олигонуклеотиды). Последовательности для рекомбинации в pntAB называются SEQ ID NO 8 и 9 (перечисленные в Таблице 2). Полученный ПЦР-продукт затем вводили посредством электропорации в штамм MG1655 (pKD46). Сохраненный штамм обозначали MG1655 Ptrc01/OP01/RBS120-pntAB::Cm. Затем модификацию Ptrc01/OP01/RBS120-pntAB::Cm переносили в штамм 6 посредством трансдукции фага Р1 (согласно Протоколу 2) с получением штамма 9.
Конструирование штамма 10
Ранее описанную модификацию Ptrc01/OP01/RBS120-pntAB::Cm переносили в штамм 8 посредством трансдукции фага Р1 (согласно Протоколу 2) с получением штамма 10.
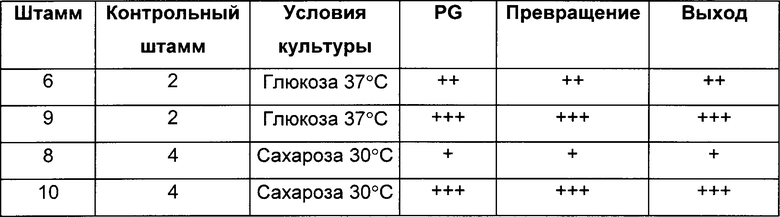
Штаммы 9 и 10, сверхэкспрессирующие pntAB, оцененные, как описано в Протоколе 5, продуцировали больше 1,2-пропандиола (PG) и имели лучшие показатели превращения и выходы по сравнению со штаммами 6 и 8 соответстенно (Таблица 4).

(Символ ~ показывает то, что отсутствует значимое различие между штаммами, символ + показывает увеличение эффективности от 10 до 100% по сравнению с контрольным штаммом, символ ++ показывает увеличение от 100 до 200% по сравнению с контрольным штаммом, символ +++ показывает увеличение от 200 до 300% по сравнению с контрольным штаммом, и символ ++++ показывает увеличение более чем на 300% по сравнению с контрольным штаммом):
ПРИМЕР 5: альтернативы сверхэкспрессии pntAB для улучшения продукции 1,2-пропандиола в штаммах, сверхэкспрессирующих дегидрогеназу вторичного спирта adh из Clostridium beijerinckii
Конструирование штамма 11
Ген yjeF из Escherichia coli клонировали в оперон с adh под определенным сайтом связывания рибосомы RBS121 (из программы RBS Calculator) (SEQ ID NO 10, перечисленная в Таблице 2) в плазмиду pPG0468, описанную в примере 3. Данную плазмиду называли pPG0518. Наконец, плазмиды pPG0518 и pPG0231 трансформировали в промежуточный штамм 8 (без плазмиды) с получением штамма 11.
Конструирование штамма 12
Для инактивации гена pgi использовали стратегию гомологичной рекомбинации (согласно Протоколам 1 и 3). Для ПЦР-амплификации кассеты устойчивости использовали олигонуклеотиды для Dpgi: SEQ ID NO 11 и 12 (перечисленные в Таблице 2). Сохранившийся штамм обозначали MG1655 Dpgi::Cm. Делецию Dpgi::Cm переносили в штамм 8 посредством трансдукции фага Р1 (согласно Протоколу 2) с получением штамма 12.
Конструирование штамма 13
Нативный ген pfkA заменяли мутировавшим геном pfkA*(L98Q) с использованием стратегии гомологичной рекомбинации (согласно Протоколам 1 и 3). Сначала ген pfkA подвергали делеции с использованием олигонуклеотидов SEQ ID NO 13 и 14 (перечисленных в Таблице 2) для ПЦР-амплификации кассеты устойчивости. Сохранившийся штамм обозначали MG1655 DpfkA::Km. Затем, для интеграции в хромосому, фрагмент, несущий мутировавшую область pfkA*(L98Q) и маркер устойчивости, которые оба фланкированы последовательностями гомологичной ДНК по отношению к целевому локусу интеграции pfkA, ПЦР-амплифицировали посредством методики перекрывающейся ПЦР (с перекрывающимися олигонуклеотидами). Последовательности для рекомбинации в pfkA называются SEQ ID NO 15 и 16 (перечисленные в Таблице 2). Полученный ПЦР-продукт затем вводили в штамм MG1655 DpfkA::Km (pKD46) посредством электропорации. Сохранившийся штамм обозначали MG1655 pfkA*(L98Q)::Cm. Модификацию pfkA*(L98Q)::Cm переносили в промежуточный штамм 8 (штамм без плазмиды) посредством трансдукции фага Р1 (согласно Протоколу 2). Ген zwf из Escherichia coli клонировали в оперон с adh под определенным сайтом связывания рибосомы RBS113 (из программы RBS Calculator) (SEQ ID NO 17, перечисленная в Таблице 2) в плазмиду pPG0468, описанную в примере 3. Данную плазмиду называли pPG0532. Наконец, плазмиды pPG0532 и pPG0231 трансформировали в предыдущий штамм с получением штамма 12.
Конструирование штамма 14
Для инактивации гена gldA описанную ранее делецию DgldA::Cm переносили посредством трансдукции фага Р1 (согласно Протоколу 2) в промежуточный выделенный штамм 3 (без регуляции gapА). Для экспрессии НАДФ+-зависимой глицеральдегид-3-фосфатдегидрогеназы из Streptococcus mutans, кодируемой геном gapN, использовали стратегию гомологичной рекомбинации (согласно Протоколам 1 и 3). Ген gapA из Escherichia coli заменяли геном gapN из Streptococcus mutans, как описано Centeno-Leija et al. (2013). Полученный ПЦР-продукт вводили в штамм MG1655 (pKD46) посредством электропорации. Сохранившийся штамм обозначали MG1655 gapN::Cm. Модификацию gapN::Cm переносили посредством трансдукции фага Р1 (согласно Протоколу 2) в предыдущий штамм. Для хромосомной экспрессии генов scrK, scrYAB и scrR использовали стратегию гомологичной рекомбинации, описанную Datsenko & Wanner, 2000 (согласно Протоколам 1 и 3). Для хромосомной интеграции фрагмент, несущий гены scrK, scrYAB и scrR, экспрессируемые под контролем их природных промоторов, связанный с маркером устойчивости, которые оба фланкированы последовательностями гомологичной ДНК по отношению к целевому локусу интеграции ykiA, ПЦР-амплифицировали посредством методики перекрывающейся ПЦР (с перекрывающимися олигонуклеотидами). Последовательности для рекомбинации в ykiA называются SEQ ID NO 18 и 19 (перечисленные в Таблице 2). Полученные ПЦР-продукты «DykiA::scrKYABR» затем вводили электропорацией в штамм MG1655 (pKD46). Сохранившийся штамм обозначали MG1655 DykiA::scrKYABR::Cm. Модификацию DykiA::scrKYABR::Cm переносили в предыдущий штамм посредством трансдукции фага Р1 (согласно Протоколу 2). Для обеспечения сверхпродукции НАДФ+-зависимой глицеральдегид-3-фосфатдегидрогеназы из Streptococcus mutans ген gapN клонировали на плазмиде pACYC184, как описано Centeno-Leija et al. (2013). Данную плазмиду называли pPG0548. Наконец, плазмиды pPG0468 и pPG0548 трансформировали в предыдущий штамм с получением штамма 13.
Конструирование штамма 15
Для инактивации НАД+-зависимой липоамиддегидрогеназы, кодируемой геном lpd, использовали стратегию гомологичной рекомбинации (согласно Протоколам 1 и 3). Таким образом, олигонуклеотиды для Dlpd: SEQ ID NO 20 и 21 (перечисленные в Таблице 2) использовали для ПЦР-амплификации кассеты устойчивости. Сохранившийся штамм обозначали MG1655 Dlpd::Cm. Наконец, делецию Dlpd::Cm переносили в промежуточный штамм 8 (штамм без плазмиды) посредством трансдукции фага Р1 (согласно Протоколу 2). Мутировавший ген lpd*(A55V/G185A/G189A/E203V/M204R/F205K/D206H/P210R) из Escherichia coli ПЦР-амплифицировали методикой перекрывающейся ПЦР (с перекрывающимися олигонуклеотидами) и клонировали в оперон с adh под определенным сайтом связывания рибосомы RBS131 (из программы RBS Calculator) (SEQ ID NO 22, перечисленная в Таблице 2) в плазмиду pPG0468, описанную в примере 3. Данную плазмиду называли pPG0523. Наконец плазмиды pPG0231 и pPG0523 трансформировали в предыдущий штамм с получением штамма 14.
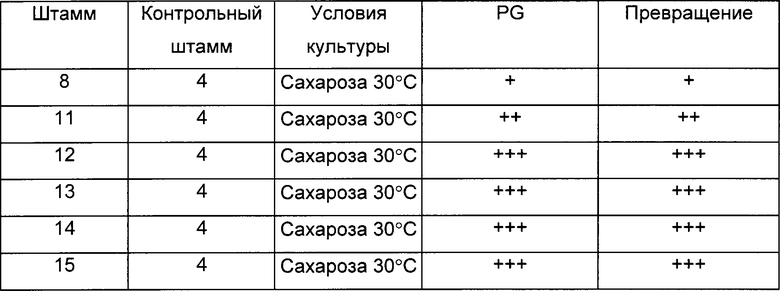
Штаммы 11, 12, 13, 14 и 15, оцененные, как описано в Протоколе 5, продуцировали больше 1,2-пропандиола (PG) и имели лучшие показатели превращения по сравнению со штаммом 8 (Таблица 5)

(Символ ~ показывает то, что отсутствует значимое различие между штаммами, символ + показывает увеличение эффективности от 10 до 100% по сравнению с контрольным штаммом, символ ++ показывает увеличение от 100 до 200% по сравнению с контрольным штаммом, символ +++ показывает увеличение от 200 до 300% по сравнению с контрольным штаммом, и символ ++++ показывает увеличение более чем на 300% по сравнению с контрольным штаммом):
ПРИМЕР 6: альтернативы adh из Clostridium beijerinckii
Идентификация гомологичных последовательностей и конструирование филогенетических деревьев
Конструирования филогенетических деревьев из набора последовательностей проводили с использованием биоинформатических программ, доступных на веб-сайте http://www.phylogeny.fr/, и данное программное обеспечение описано в Dereeper et al. (2008) и Dereeper et al. (2010).
Стадия 1: идентификация гомологичных последовательностей
Идентификацию аналогичных последовательностей проводили из семейства дегидрогеназы вторичного спирта и семейства альдегид/кетонредуктазы с использованием программы BLAST посредством установки следующих параметров: база данных (Swissprot/UNIPROT), значение е - 0,01 и фильтр для последовательностей низкой сложности.
Этап 2: конструирование филогенетических деревьев
Последовательности выравнивали с использованием программы MUSCLE, и двусмысленные области удаляли с использованием программы Gblocks. Затем филогенетическое древо реконструировали с использованием способа максимального правдоподобия, выполненного в программе PhyML. Графическое представление и редактирование филогенетического древа проводили с использованием TreeDyn.
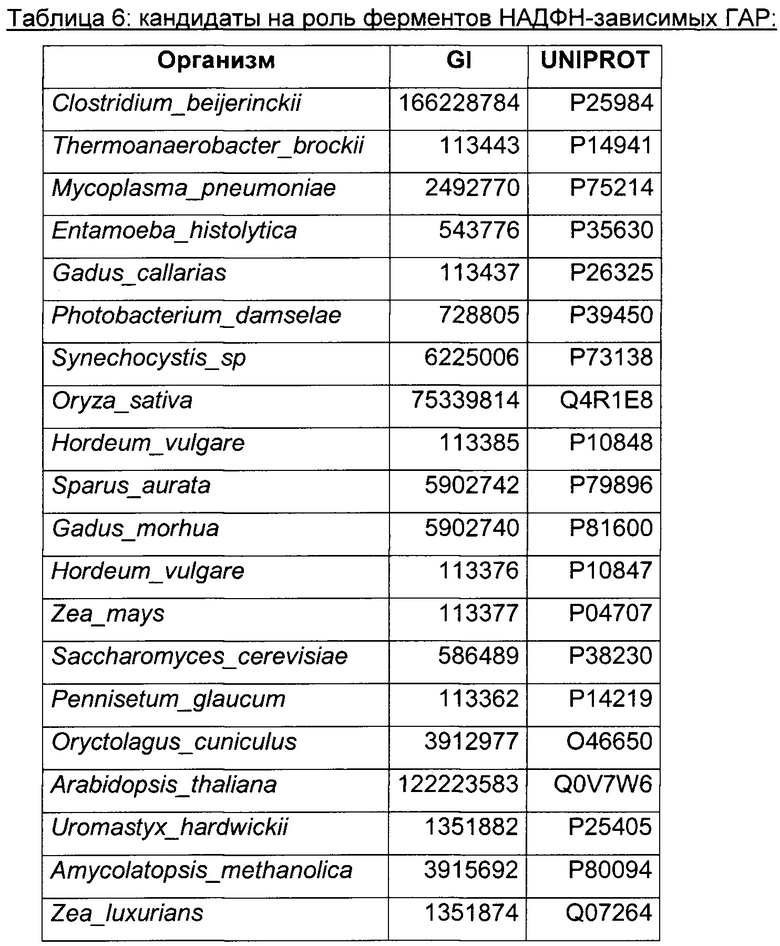
Adh из Clostridium beijerinckii была самой близкородственной ферментам Thermoanaerobacterium brockii, Entamoeba histolytica и Mycoplasma pneumoniae. Следующее наилучшее соответствие было с алкогольдегидрогеназами, найденными у эукариот. Ферменты из Gluconobacter oxydans, Hypocrea jecorina, Entamoeba histolytica и Bacillus subtilis клонировали, очищали и анализировали на активность НАДФН-зависимой гидроксиацетон редуктазы.
Конструирование штамма 16
Для инактивации гена gldA использовали стратегию гомологичной рекомбинации (согласно Протоколам 1 и 3). Для ПЦР-амплификации кассеты устойчивости использовали олигонуклеотиды для DgldA: SEQ ID NO 5 и 6 (перечисленные в Таблице 2). Сохранившийся штамм обозначали MG1655 DgldA::Km. Наконец делецию DgldA::Km переносили в штамм BL21(DE3)звездочка посредством трансдукции фага Р1 (согласно Протоколу 2). Для характеризации глицеролдегидрогеназы из Gluconobacter oxydans, синтетический ген gld, оптимизированный для Escherichia coli (SEQ ID NO 23, перечисленная в таблице 2), клонировали в экспрессионную плазмиду pPAL7 (Biorad®). Данную плазмиду называли pPG0381 и трансформировали в штамм BL21(DE3)звездочка DgldA::Km с получением штамма 16.
Конструирование штамма 17
Для характеризации глицеролдегидрогеназы из Hypocrea jecorina, синтетический ген gld2, оптимизированный для Escherichia coli (SEQ ID NO 24, перечисленная в таблице 2), клонировали в экспрессионную плазмиду pPAL7 (Biorad®). Данную плазмиду называли pPG0418 и трансформировали в штамм BL21(DE3)звездочка DgldA::Km, описанный ранее, с получением штамма 17.
Конструирование штамма 18
Для характеризации алкогольдегидрогеназы из Entamoeba histolytica, синтетический ген adh1, оптимизированный для Escherichia coli (SEQ ID NO 25, перечисленная в таблице 2), клонировали в экспрессионную плазмиду pPAL7 (Biorad®). Данную плазмиду называли pPG0539 и трансформировали в штамм BL21(DE3) кодонплюс с получением штамма 18.
Конструирование штамма 19
Для характеризации альдокеторедуктазы из Bacillus subtilis, ген yhdN из Bacillus subtilis клонировали в экспрессионную плазмиду pPAL7 (Biorad®). Данную плазмиду называли pPG0357 и трансформировали в штамм BL21(DE3) с получением штамма 19.
Очистка рекомбинантных белков
Штаммы 16, 17, 18 и 19 культивировали, как описано в Протоколе 6. Клетки (350-500 мг сухой массы) ресуспендировали в экстракционном буфере (60-90 мл) со смесью ингибиторов протеаз. Суспендированные клетки разрушали шестью циклами обработки ультразвуком по 30 с на льду (ультразвуковой дезинтегратор Branson, 70 Вт), с последующей инкубацией в течение 1 ч при комнатной температуре с 1 мМ MgCl2 и 2 ME (Международная единица)/мл ДНКазы I. Обломки клеток удаляли центрифугированием при 12000 g в течение 30 мин при 4°С. Супернатант сохраняли в виде неочищенного клеточного экстракта. Рекомбинантный белок очищали из неочищенного экстракта посредством применения аффинной хроматографии с субтилизином (PROfinity EXact Cartridge 5 мл, BIORAD) согласно инструкциям изготовителя. Фракции, содержащие белок, объединяли, концентрировали и загружали на гель-фильтрационную колонку (колонка Superdex 200 10/300 GL, GE Healthcare). Концентрацию белка определяли с использованием анализа по Бредфорд.
НАДФН-зависимая гидроксиацетон редуктаза (НАДФН ГАР) очищенных ферментов
Активность НАДФН ГАР определяли посредством измерения потребления НАДФН при 340 нм на спектрофотометре (ε340 нм равно 6290 М-1 см-1) и при 30°С.
Реакционную смесь (1 мл), содержащую буфер для анализа, 0,2 мМ НАДФН и очищенный белок, инкубировали в течение 5 мин при 30°С. Затем добавляли 30 мМ гидроксиацетон для начала реакции. Одну единицу ферментативной активности определяли как количество фермента, катализирующее уменьшение НАДФН на 1 мкмоль в минуту. Удельную ферментативную активность выражали в виде единиц ферментативной активности на мг белка. Значение активности, определенное без гидроксиацетона, вычитали.
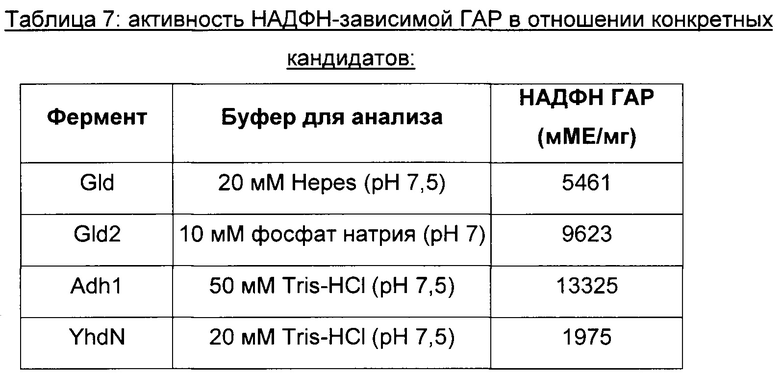
Конструирование штамма 20
Оптимизированный ген adh1 из Entamoeba histolytica клонировали в плазмиду pME101VB01, описанную в патентной заявке WO 2008/116853. Данную плазмиду называли pPG0544. Плазмиды pPG0544 и pPG0231 трансформировали в штамм 3, описанный в Примере 2, затем инактивировали ген gldA, как описано в Примере 3, и оперон pntAB сверхэкспрессировали, как описано в Примере 4, с получением штамма 19.
Штамм 20, оцененный, как описано в Протоколе 5, продуцировал больше 1,2- пропандиола (PG) и имел лучший показатель превращения по сравнению со штаммом 4 (Таблица 8).

(Символ ~ показывает то, что отсутствует значимоеразличие между штаммами, символ + показывает увеличение эффективности от 10 до 100% по сравнению с контрольным штаммом, символ ++ показывает увеличение от 100 до 200% по сравнению с контрольным штаммом, символ +++ показывает увеличение от 200 до 300% по сравнению с контрольным штаммом, и символ ++++ показывает увеличение более чем на 300% по сравнению с контрольным штаммом):


ПРИМЕР 7: замена кофактора GldA*(A160T)
Конструирование фермента и структурный анализ
Строили модели гомологии GldA*(A160T) из рентгеновской структуры глицеролдегидрогеназы Bacillus stearothermophilus. Данные модели рассчитывали с использованием программы Discovery Studio (Accelrys). Данные модели сравнивали со структурой НАД- и НАДФ-зависимых ферментов, которые имели по меньшей мере 30%-ную идентичность, и структура которых была известной.
Аминокислоты, участвующие в определении специфичности в отношении кофактора, идентифицировали посредством выравнивания последовательностей между последовательностью GldA*(A160T) и несколькими НАД- и НАДФ-зависимыми дегидрогеназами, наложения моделей гомологии GldA*(A160T) и глицеролдегидрогеназ, присутствующих в банке данных белков RCSB, и сравнения с данными, найденными в Clermont et al. (1993), Ruzheinikov et al. (2001), Corbier et al. (1990) и Wu et al. (2012).
Определили двух мутантов из анализа последовательности и структуры: D37G и D37G/P161S/L164A.
Конструирование штамма 21
Для характеризации мутантной 1,2-пропандиол: НАД+-оксидоредуктазы из Escherichia coli, сначала клонировали ген gldA в экспрессионную плазмиду pET101/D-TOPO (Lifetechnologies®). Данную плазмиду называли pPG0029. Для сверхэкспрессии мутировавшего гена gldA*(A160T) использовали сайт-направленный мутагенез на pPG0029. Данную плазмиду называли pPG0394, и трансформировали ее в штамм BL21(DE3) звездочка DgldA::Km с получением штамма 21.
Конструирование штамма 22
Для сверхэкспрессии мутировавшего гена gldA*(A160T/D37G) использовали сайт-направленный мутагенез на pETTOPO-gldA*(A160T). Данную плазмиду называли pPG0425, и ее трансформировали в штамм BL21(DE3) звездочка DgldA::Km с получением штамма 22.
Конструирование штамма 23
Для сверхэкспрессии мутировавшего гена gldA*(A160T/D37G/P161S/L164A) использовали сайт-направленный мутагенез на pPG0425. Данную плазмиду называли pPG0438, и ее трансформировали в штамм BL21(DE3) звездочка DgldA::Km с получением штамма 23.
Очистка GldA*(A160T). GldA*(A160T/D37G) и GldA*(A160T/D37G/P161S/L164A)
Штаммы 21, 22 и 23 культивировали, как описано в Протоколе 6. Клетки (500 мг сухой массы) ресуспендировали в 90 мл экстракционного буфера (100 мМ фосфат калия, рН 7,6, 20 мМ имидазол и смесь ингибиторов протеаз). Суспендированные клетки разрушали 8 циклами обработки ультразвуком по 30 с на льду (ультразвуковой дезинтегратор Branson, 70 Вт), с последующей инкубацией в течение 45 мин при комнатной температуре с 5 мМ MgCl2 и 2 МЕ/мл ДНКазы I. Обломки клеток удаляли центрифугированием при 12000 g в течение 30 мин при 4°С. Супернатант сохраняли в качестве неочищенного экстракта. Ферменты очищали из неочищенного экстракта с использованием аффинной хроматографии на никеле (HisTrapFF 1 мл, GE Healthcare) согласно инструкциям изготовителя. Ферменты элюировали с использованием линейного градиента имидазола (от 20 до 500 мМ) в 100 мМ фосфате калия (рН 7,6). После стадии обессоливания посредством гель-фильтрации (колонка Superdex200 10/300 GL, GE Healthcare, уравновешенная 100 мМ MES-KOH (рН 6,5)) определяли концентрации белка с использованием анализа по Бредфорд.
Характеризация GldA*(A160T), GldA*(A160T/D37G) и GldA*(A160T/D37G/P161S/L164A)
Активность НАДФН-зависимой гидроксиацетон редуктазы (НАДФН ГАР) определяли посредством измерения потребления НАДФН при 340 нм на спектрофотометре (ε340 равно 6290 М-1 см-1) и при 30°С. Реакционную смесь (1 мл), содержащую 100 мМ MES-KOH (рН 6,5), 0,1 мМ FeSO4, 30 мМ сульфат аммония, от 0,05 до 0,4 мМ НАДФН и очищенный фермент, инкубировали в течение 5 мин при 30°С. Затем добавляли от 0,1 до 10 мМ гидроксиацетона для начала реакции. Анализ НАДН-зависимой гидроксиацетон редуктазы (НАДН ГАР) проводили при тех же самых условиях, что и НАДФН ГАР, за исключением того, что НАДФН в реакционной смеси заменяли НАДН, и активность определяли посредством измерения потребления НАДН при 340 нм. Кинетические параметры определяли с использованием Sigmaplot посредством аппроксимации уравнения Михаэлиса- Ментен.
Ферменты GldA*(A160T/D37G) и GldA*(A160T/D37G/P161S/L164A) продемонстрировали повышенные каталитические эффективности с НАДФН и пониженные каталитические эффективности с НАДН по сравнению с GldA*(A160T) (Таблица 9).

Конструирование штамма 24
Мутировавший ген gldA*(A160T/D37G/P161S/L164A) клонировали в плазмиду, полученную из рМЕ101, как описано для конструкции pME101VB06-gldA*(A160T) в патентной заявке ЕР 2532751. Данную плазмиду называли pPG0467. Наконец, плазмиды pPG0467 и pPG0231 трансформировали в промежуточный штамм 8 (штамм без плазмиды), описанный в примере 2, с получением штамма 24.
Штамм 24, оцененный, как описано в Протоколе 5, продуцировал больше 1,2- пропандиола (PG) и имел лучший показатель превращения по сравнению со штаммом 4 (Таблица 10).
 (Символ ~ показывает то, что отсутствует значимое различие между штаммами, символ + показывает увеличение эффективности от 10 до 100% по сравнению с контрольным штаммом, символ ++ показывает увеличение от 100 до 200% по сравнению с контрольным штаммом, символ +++ показывает увеличение от 200 до 300% по сравнению с контрольным штаммом, и символ ++++ показывает увеличение более чем на 300% по сравнению с контрольным штаммом):
(Символ ~ показывает то, что отсутствует значимое различие между штаммами, символ + показывает увеличение эффективности от 10 до 100% по сравнению с контрольным штаммом, символ ++ показывает увеличение от 100 до 200% по сравнению с контрольным штаммом, символ +++ показывает увеличение от 200 до 300% по сравнению с контрольным штаммом, и символ ++++ показывает увеличение более чем на 300% по сравнению с контрольным штаммом):

ПРИМЕР 8: MG=>PG активность Adh из Clostridium beijerinckii
Конструирование штамма 25
Оптимизированный ген adh из Clostridium beijerinckii (Hanai, Atsumi and Liao, 2007) клонировали в экспрессионную плазмиду рЕТ28а. Данную плазмиду называли pPG0445, и трансформировали ее в штамм BL21(DE3) с получением штамма 25.
Очистка Adh из Clostridium beijerinckii
Штамм 25 культивировали, как описано в Протоколе 6. Клетки (сухой вес 315 мг) ресуспендировали в 50 мл экстракционного буфера (20 мМ Tris-HCl, рН 7,3, 0,1 мМ DTT (дитиотрейтол), 0,1 мМ бензамидин, 10% глицерина, 0,02% азида натрия и смесь ингибиторов протеаз). Суспендированные клетки разрушали 8 циклами обработки ультразвуком по 30 с на льду (ультразвуковой дезинтегратор Branson, 70 Вт). Обломки клеток удаляли центрифугированием при 12000 g в течение 30 мин при 4°С. Супернатант подвергали воздействию нагревания (65° в течение 5 мин) и затем повторно центрифугировали при 12000 g в течение 30 мин при 4°С. Белок очищали из супернатанта с использованием аффинной хроматографии на никеле (HisTrapFF 1 мл, GE Healthcare) согласно инструкциям изготовителя. Белок элюировали с использованием линейного градиента имидазола (от 20 до 500 мМ) в 50 мМ Tris-HCl (рН 7,4). Фракции, которые содержат белок, объединяли, концентрировали и диализировали против 50 мМ Tris-HCl (рН 7). Концентрацию белка определяли с использованием анализа по Бредфорд.
Количественное определение 1,2-пропандиола, продуцированного из метилглиоксаля
5-10 мкг очищенного фермента инкубировали в течение 30 мин при 30°С в 50 мМ Tris-HCl (рН 7,5), 10 мМ метилглиоксале и 5 мМ НАДФН. Количество 1,2-пропандиола, продуцируемого Adh из метилглиоксаля, измеряли непосредственно ЖХ-МС (Agilent Technologies).
При таких условиях продуцировалось 4,6 мМ 1,2-пропандиола, тогда как 1,2-пропандиол не продуцировался при исключении либо метилглиоксаля, либо фермента.
Конструирование штаммов 26, 27 и 28
Для инактивации гена gldA описанную ранее делецию DgldA::Km и ген tpiA совместно трансфицировали посредством трансдукции фага Р1 (согласно Протоколу 2) в штамм MG1655 lpd* DtpiA DpflAB DadhE DldhA DgloA DaldA DaldB Dedd, описанный в патентной заявке WO 2008/116852. Геномную модификацию для регуляции экспрессии gapA "CI857-PR01/RBS11-gapA" вводили в предыдущий штамм, как описано в патенте ЕР 2532751. Для инактивации альдегидредуктазы, кодируемой геном yqhD, и глиоксальредуказы, кодируемой геном yqhE, использовали стратегию гомологичной рекомбинации (согласно Протоколам 1 и 3). Для ПЦР-амплификации кассет устойчивости использовали олигонуклеотиды DyqhDE: SEQ ID NO 26 и 27 (перечисленные в Таблице 2). Сохранившийся штамм обозначали MG1655 DyqhDE::Bs. Делецию DyqhDE::Bs переносили в предыдущий штамм посредством трансдукции фага Р1 (согласно Протоколу 2), с получением штамма 26. Затем плазмиду pPG0468 трансформировали в данный штамм с получением штамма 27. Наконец, описанную ранее модификацию Ptrc01/OP01/RBS120-pntAB::Cm переносили в штамм 27 посредством трансдукции фага Р1 (согласно Протоколу 2), с получением штамма 28.
При оценке, как описано в Протоколе 5, штаммы 27 и 28 продуцировали больше 1,2-пропандиола (PG), чем штамм 26.
ССЫЛКИ
- Altaras NE and Cameron DC (2000), Biotechnol. Prog., 16: 940-946
- Altaras NE and Cameron DC (1999), Appl. Environ. Microbiol., 65: 1180-1185
- Badia J, Ros J, Aguilar J (1985), J. Bacteriol. 161: 435-437
- Berrios-Rivera SJ, San KY, Bennett GN (2003), J. Ind. Microbiol. Biotechnol., 30: 34-40
- Bocanegra J, Scrutton N, Perham R (1993) Biochemistry, 32 (11): 2737-2740
- Cameron DC, Altaras NE, Hoffman ML, Shaw AJ (1998), Biotechnol. Prog., 14: 116-125
- Carrier T & Keasling J (1999), Biotechnol Prog., 15 (1): 58-64
- Centeno-Leija S, Utrilla J, Flores N, Rodriguez A, Gosset G, Martinez A (2013) Antonie Van Leeuwenhoek., 104 (6), 913-924.
- Clermont S, Corbier C, Mely Y, Gerard D, Wonacott A, Branlant G, (1993), Biochemistry, 32 (38): 10178-10184
- Corbier C, Clermont S, Billard P, Skarzynski T, Branlant C, Wonacott A, Branlant G (1990), Biochemistry, 29 (30): 7101-7106
- Datsenko KA & Wanner BL, (2000), Proc Natl Acad Sci USA., 97: 6640-6645
- Dereeper A, Audic S, Claverie J, Blanc G (2010),BMC Evol Biol., 12: 10:8
- Dereeper A, Guignon V, Blanc G, Audic S, Buffet S, Chevenet F, Dufayard J, Guindon S, Lefort V, Lescot M, Claverie J, Gascuel О (2008) Nucleic Acids Res., 1: 36 (Web Server issue): W465-9. Epub 2008 Apr 19
- Fuhrer T and Sauer U (2009), J. Bacteriol., 191 (7): 2112-2121
- Hanai T, Atsumi S, Liao J (2007), Appl. Environ. Microbiol., 73: 7814-7818
- Huang K, Rudolph FB, Bennett GN (1999), Appl. Environ. Microbiol., 65: 3244- 3247
- Ismaiel A, Zhu C, Colby G, Chen J (1993), J Bacteriol., 175 (16): 5097-5105
- Jarboe LR (2011), Appl Microbiol Biotechnol., 89: 249-257.
- Katzberg M, Skorupa-Parachin N, Gorwa-Grauslund M, Bertau M (2010), Int. J. Mol. Sci., 11(4): 1735-1758
- Kelley J & Dekker E (1984), J Biol Chem., 259 (4): 2124-2129
- Lee S, McCormick M, Lippard S, Cho U (2013), Nature, 494: 380-384
- Li H and Liao J (2013), Microb Cell Fact., 12 (1): 4
- Marbaix A, Noel G, Detroux A, Vertommen D, Schaftingen E, Linster С (2011), J Biol Chem., 286 (48):, 41246-41252
- Needleman and Wunsch (1970), J. Mol. Biol., 48(3), 443-453
- Old SE, Sato S, Kador PF, Carper DA (1990), Proc Natl Acad Sci USA., 87: 4942-4945
- Ruzheinikov S, Burke J, Sedelnikova S, Baker P, Taylor R, Bullough P, Muir N (2001), Structure, 9 (9): 789-802
- Salis H (2011), Methods Enzymol., 498:19-42
- Schmid K, Schupfner M, Schmitt R (1982), J. Bacteriol., 151: 68-76
- Segel I (1993), Enzyme kinetics, John Wiley & Sons, pp.44-54 and 100-112
- Scrutton N, Berry An Perham R (1990), Nature, 343: 38-43
- Lim S, Jung Y, Shin H, Lee Y (2002), J Biosci Bioeng., 93 (6): 543-549
- Wu C, Hwa Y, Chen Y, Lim С (2012), J Phys Chem B, 116 (19): 5644-5652.
| название | год | авторы | номер документа |
|---|---|---|---|
| МИКРОБИОЛОГИЧЕСКИЙ СПОСОБ ПОЛУЧЕНИЯ 1,2-ПРОПАНДИОЛА | 2009 |
|
RU2521502C2 |
| БАКТЕРИЯ ESCHERICHIA COLI ДЛЯ ПОЛУЧЕНИЯ 1,2-ПРОПАНДИОЛА, СПОСОБ ЕЕ ПОЛУЧЕНИЯ, СПОСОБ ПОЛУЧЕНИЯ 1,2-ПРОПАНДИОЛА | 2005 |
|
RU2407793C2 |
| МИКРООРГАНИЗМЫ ДЛЯ ПРОИЗВОДСТВА МЕТИОНИНА С УЛУЧШЕННЫМ ВЫХОДОМ МЕТИОНИНА | 2014 |
|
RU2678757C2 |
| РЕКОМБИНАНТНЫЙ МИКРООРГАНИЗМ ДЛЯ ФЕРМЕНТАТИВНОГО ПРОИЗВОДСТВА МЕТИОНИНА | 2012 |
|
RU2629760C2 |
| СПОСОБ И МИКРООРГАНИЗМ ДЛЯ ФЕРМЕНТАТИВНОГО ПРОДУЦИРОВАНИЯ МЕТИОНИНА С УЛУЧШЕННЫМ ВЫХОДОМ МЕТИОНИНА | 2015 |
|
RU2723714C2 |
| ФЕРМЕНТАТИВНОЕ ПОЛУЧЕНИЕ ЧЕТЫРЕХУГЛЕРОДНЫХ СПИРТОВ | 2006 |
|
RU2394913C2 |
| СПОСОБ ФЕРМЕНТАТИВНОГО ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ИСПОЛЬЗОВАНИЕМ УЛУЧШЕННЫХ ШТАММОВ СЕМЕЙСТВА Enterobakteriaceae | 2013 |
|
RU2672614C2 |
| СПОСОБ ПОЛУЧЕНИЯ L-ТРЕОНИНА С ИСПОЛЬЗОВАНИЕМ БАКТЕРИИ, ПРИНАДЛЕЖАЩЕЙ К РОДУ Escherichia, В КОТОРОЙ ИНАКТИВИРОВАН ГЕН yahN | 2007 |
|
RU2392322C2 |
| СПОСОБ ПОЛУЧЕНИЯ 2,4-ДИГИДРОКСИБУТИРАТА | 2013 |
|
RU2645260C2 |
| Штамм Escherichia coli - продуцент L-треонина | 2023 |
|
RU2817252C1 |











Группа изобретений относится к гетеротрофному микроорганизму, генетически модифицированному для продукции 1,2-пропандиола, и к способу получения 1,2-пропандиола. В указанном микроорганизме сверхэкспрессируется по меньшей мере один ген, кодирующий НАДФН-зависимую ацетолредуктазу, или мутантный ген gldA*, кодирующий НАДФН-зависимую глицеролдегидрогеназу, содержащую замену аминокислотного остатка аспарагиновой кислоты в положении 37 белка GldA Escherichia coli К12 на глицин, аланин или валин. При этом указанная НАДФН-зависимая ацетолредуктаза кодируется геном adh из Clostridium beijerinckii или геном adh из Thermoanaerobacter brockii, или геном adh1 из Entamoeba histolytica, или геном GOX1615 из Gluconobacter oxydans, или геном gld2 из Hypocrea jecorina, или геном yhdN из Bacillus subtilis. Способ получения 1,2-пропандиола предусматривает культивирование указанного микроорганизма в подходящей культуральной среде, содержащей углевод в качестве источника углерода, и выделение 1,2-пропандиола из указанной культуральной среды. Группа изобретений обеспечивает улучшенную продукцию 1,2-пропандиола из углевода в качестве источника углерода. 2 н. и 15 з.п. ф-лы, 10 табл., 8 пр.
1. Способ получения 1,2-пропандиола в ферментативном процессе, включающий следующие стадии:
- культивирование гетеротрофного микроорганизма, генетически модифицированного для продукции 1,2-пропандиола, в подходящей культуральной среде, содержащей углевод в качестве источника углерода, и
- выделение 1,2-пропандиола из указанной культуральной среды,
где в указанном генетически модифицированном микроорганизме сверхэкспрессируется по меньшей мере один ген, кодирующий НАДФН-зависимую ацетолредуктазу, или мутантный ген gldA*, кодирующий НАДФН-зависимую глицеролдегидрогеназу, содержащую замену аминокислотного остатка аспарагиновой кислоты в положении 37 белка GldA Escherichia coli К12 на глицин, аланин или валин, причем указанная НАДФН-зависимая ацетолредуктаза кодируется геном adh из Clostridium beijerinckii или геном adh из Thermoanaerobacter brockii, или геном adh1 из Entamoeba histolytica, или геном GOX1615 из Gluconobacter oxydans, или геном gld2 из Hypocrea jecorina, или геном yhdN из Bacillus subtilis.
2. Способ по п. 1, где доступность НАДФН в микроорганизме увеличивается по меньшей мере посредством одной из следующих генетических модификаций:
- сверхэкспрессия оперона гена pntAB, кодирующего никотинамиднуклеотидтрансгидрогеназу, и/или
- аттенуация гена pgi, кодирующего фосфоглюкозоизомеразу, и/или
- аттенуация гена pfkA, кодирующего фосфофруктокиназу, и/или
- сверхэкспрессия гена zwf, кодирующего глюкозо-6-фосфатдегидрогеназу, и/или
- сверхэкспрессия гена yjeF, кодирующего АДФ-зависимую дегидратазу, и/или
- сверхэкспрессия гена gapN, кодирующего НАДФ-зависимую глицеральдегид-3-фосфатдегидрогеназу, и/или
- сверхэкспрессия мутантного гена lpd*, кодирующего НАДФ-зависимую липоамиддегидрогеназу, содержащую замены A55V, G185A, G189A, E203V, M204R, F205K, D206H и P210R относительно белка Lpd E. coli.
3. Способ по п. 1, где ген gldA, кодирующий НАДН-зависимую глицеролдегидрогеназу, подвергается делеции в микроорганизме.
4. Способ по п. 1, где микроорганизм дополнительно содержит делецию гена yqhD, кодирующего метилглиоксальредуктазу.
5. Способ по любому из пп. 1-4, где микроорганизм выбран из группы, состоящей из Enterobacteriaceae, Bacillaceae, Clostridiaceae, Streptomycetaceae и дрожжей.
6. Способ по п. 5, где микроорганизм выбран из группы, состоящей из Escherichia coli, Klebsiella pneumoniae, Thermoanaerobacterium thermosaccharolyticum, Clostridium sphenoides и Saccharomyces cerevisiae.
7. Способ по п. 6, где микроорганизм представляет собой E. coli.
8. Способ по п. 1, где мутантный ген gldA*, кодирующий GldA* НАДФН-зависимую глицеролдегидрогеназу, дополнительно содержит мутацию А160Т относительно последовательности белка GldA Escherichia coli К12.
9. Способ по п. 1, где мутантный ген gldA*, кодирующий GldA* НАДФН-зависимую глицеролдегидрогеназу, дополнительно содержит замену аминокислотного остатка в положении 161 относительно последовательности белка GldA Escherichia coli К12 серином или треонином.
10. Способ по п. 9, где мутантный ген gldA*, кодирующий GldA* НАДФН-зависимую глицеролдегидрогеназу, дополнительно содержит замену аминокислотного остатка в положении 164 относительно последовательности белка GldA Escherichia coli К12 аланином, глицином или валином.
11. Гетеротрофный микроорганизм, генетически модифицированный для продукции 1,2-пропандиола из углевода в качестве источника углерода, где в указанном микроорганизме сверхэкспрессируется по меньшей мере один ген, кодирующий НАДФН-зависимую ацетолредуктазу, или мутантный ген gldA*, кодирующий НАДФН-зависимую глицеролдегидрогеназу, содержащую замену аминокислотного остатка аспарагиновой кислоты в положении 37 относительно последовательности белка GldA Escherichia coli К12 на глицин, аланин или валин, причем указанная НАДФН-зависимая ацетолредуктаза кодируется геном adh из Clostridium beijerinckii или геном adh из Thermoanaerobacter brockii, или геном adh1 из Entamoeba histolytica, или геном GOX1615 из Gluconobacter oxydans, или геном gld2 из Hypocrea jecorina, или геном yhdN из Bacillus subtilis.
12. Микроорганизм по п. 11, дополнительно содержащий по меньшей мере одну из следующих генетических модификаций:
- делеция гена gldA, кодирующего НАДН-зависимую глицеролдегидрогеназу, и/или
- делеция гена yqhD, кодирующего альдегидредуктазу, и/или
- делеция гена pgi, кодирующего фосфоглюкозоизомеразу, и/или
- делеция гена pfkA, кодирующего фосфофруктокиназу, и/или
- сверхэкспрессия гена pntAB, кодирующего никотинамиднуклеотидтрансгидрогеназу, и/или
- сверхэкспрессия гена zwf, кодирующего глюкозо-6-фосфатдегидрогеназу, и/или
- сверхэкспрессия гена yjeF, кодирующего АДФ-зависимую дегидратазу, и/или
- сверхэкспрессия гена gapN, кодирующего НАДФ-зависимую глицеральдегид-3-фосфатдегидрогеназу, и/или
- сверхэкспрессия мутантного гена lpd*, кодирующего НАДФ-зависимую липоамиддегидрогеназу, содержащую замены A55V, G185A, G189A, E203V, M204R, F205K, D206H и P210R относительно белка Lpd E. coli.
13. Микроорганизм по п. 11 или 12, где он выбран из группы, состоящей из Enterobacteriaceae, Bacillaceae, Clostridiaceae, Streptomycetaceae, Corynebacteriaceae и дрожжей.
14. Микроорганизм по п. 13, где он выбран из группы, состоящей из Escherichia coli, Klebsiella pneumoniae, Thermoanaerobacterium thermosaccharolyticum, Clostridium sphenoides и Saccharomyces cerevisiae.
15. Микроорганизм по п. 11, где мутантный ген gldA*, кодирующий GldA* НАДФН-зависимую глицеролдегидрогеназу, дополнительно содержит мутацию А160Т относительно последовательности белка GldA Escherichia coli К12.
16. Микроорганизм по п. 11, где мутантный ген gldA*, кодирующий GldA* НАДФН-зависимую глицеролдегидрогеназу, дополнительно содержит замену аминокислотного остатка в положении 161 относительно последовательности белка GldA Escherichia coli К12 серином или треонином.
17. Микроорганизм по п. 16, где мутантный ген gldA*, кодирующий GldA* НАДФН-зависимую глицеролдегидрогеназу, дополнительно содержит замену аминокислотного остатка в положении 164 относительно последовательности белка GldA Escherichia coli К12 аланином, глицином или валином.
| LI et al | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| Способ приготовления лака | 1924 |
|
SU2011A1 |
| БАКТЕРИЯ ESCHERICHIA COLI ДЛЯ ПОЛУЧЕНИЯ 1,2-ПРОПАНДИОЛА, СПОСОБ ЕЕ ПОЛУЧЕНИЯ, СПОСОБ ПОЛУЧЕНИЯ 1,2-ПРОПАНДИОЛА | 2005 |
|
RU2407793C2 |
| WHISSTOCK J.C., LESK A.M | |||
| Prediction of protein function from protein sequence and structure | |||
| Q Rev Biophys | |||
| Способ и приспособление для нагревания хлебопекарных камер | 1923 |
|
SU2003A1 |

