Предлагаемое изобретение относится к области биотехнологии и медицине и может быть использовано для получения биологически активных веществ из генетически модифицированных растительных клеток.
Современные методы молекулярной биологии и генной инженерии позволяют переносить гены одних организмов в другие организмы, а также осуществлять (конечно, далеко не во всех случаях) синтез белков, кодируемых этими генами. Это позволяет создавать системы, которые способны продуцировать биологически активные вещества с высокой эффективностью.
Секреция рекомбинантных белков из клетки в культуральную жидкость всегда представляет значительный интерес для упрощения дальнейших процедур очистки целевого белка. Для растительных клеток, обладающих сложно разрушаемой стенкой, данный подход еще более актуален.
Секретируемые белки обычно синтезируются как молекулы-предшественники, включающие «сигнальный» пептид (чаще всего имеющий N-терминальную локализацию), который взаимодействует с рецепторным белком на мембране эндоплазматичекого ретикулума (ЭР) и направляет растущую полипептидную цепь через мембрану в ЭР для секреции из клетки. «Сигнальный» пептид часто отщепляется от молекулы-предшественника, превращая ее в зрелый белок.
Альтернативно, сигнальные пептиды млекопитающих могут также быть использованы для растительной секреции. Было показано, что эти сигналы узнаются в растениях и могут направлять секрецию слитых с ними полипептидов не только через плазматическую мембрану, но и через растительную клеточную стенку.
Попавшие в ЭР полипептиды могут содержать дополнительные сигналы (часто С-концевые), определяющие перенос белка в вакуолю или аппарат Гольджи. Если таких сигналов не содержится, по умолчанию идет секреция внутри клетки. Растительные сигналы, направляющие транспорт синтезируемого белка в эндоплазматический ретикулум (ЭР), описаны в целом ряде работ и патентов [1]. В отдельных случаях используют сигнальный пептид основной эндохитиназы Arabidopsis thaliana [2], сигнальный пептид экстенсина [3], сигнальный пептид альфа-амилазы риса [4].
Секретируемые в культуральную среду зрелые полипептиды могут быть выделены из нее любым подходящим способом, включая хромотографию, электрофорез, диализ и т.п.
Известно, что последовательность секреторного пептида может существенно влиять на уровень секреции целевого белка. Таким образом, для выбора оптимальной конструкции для трансгенеза требуется предварительная длительная проверка эффективности секреции целевого белка путем получения трансгенных растений и прямого сравнения уровня продукции целевого белка [5].
Известен способ секреции рекомбинантных белков из высших растений, включающий конструирование экспрессирующего вектора, трансформацию агробактерий, агроперенос ДНК в клетки растений и получение непосредственно модифицированных растений из каллусной культуры [6].
Недостатком известного способа является его длительность и трудоемкость.
Наиболее ближайшим к заявляемому способу - прототипом является способ секреции рекомбинантных белков из ряски болотной, включающий конструирование рекомбинантной плазмидной ДНК, содержащей целевой ген, передачу полученной плазмидной ДНК в штамм Agrobacterium tumefaciens штамм C58Z707, кокультивирование трансформированных клеток Agrobacterium с клетками ряски болотной с последующим выявлением трансформированных клеток [7].
Недостатком известного способа является его длительность, трудоемкость и низкая эффективность. Например, получение клеток ряски болотной, пригодных для трансформации, занимает несколько недель. Кроме этого, известный способ непригоден для трансформации клеток микроводоросли рода хлорелла.
Технической задачей изобретения является ускорение процесса и повышение эффективности секреции рекомбинантных белков из клеток микроводоросли рода хлорелла (Chlorella).
Поставленная техническая задача достигается предлагаемым способом, заключающимся в следующем.
Конструируют рекомбинантную плазмидную ДНК pAUTL-GFP, несущую гибридный ген, кодирующий зеленый флуоресцентный белок (GFP) и сигнальную последовательность аутолизина. Полученной плазмидой трансформируют штамм агробактерий. Суспензию трансформированных агробактериальных клеток в логарифмической стадии роста инкубируют с клетками микроводоросли Chlorella в течение 14-16 часов. Уровень (эффективность) секреции GFP оценивают измеряя флуоресценцию культуральной среды.
Определяющими отличиями предлагаемого способа по сравнению с прототипом являются:
- в качестве плазмиды используют рекомбинантную плазмидную ДНК pAUTL-GFP с сигнальной последовательностью аутолизина из Chlamydomonas reinhardtii, которая наиболее адаптирована к геному микроводоросли рода Chlorella.
- в качестве хозяина для агробактериальной транзиентной экспрессии используют клетки зеленой микроводоросли рода Chlorella, что является экономически выгодным, поскольку она растет быстро и в дешевой среде.
Изобретение иллюстрируется следующими примерами конкретного выполнения.
Пример 1. Конструирование плазмидной ДНК pAUTL-GFP.
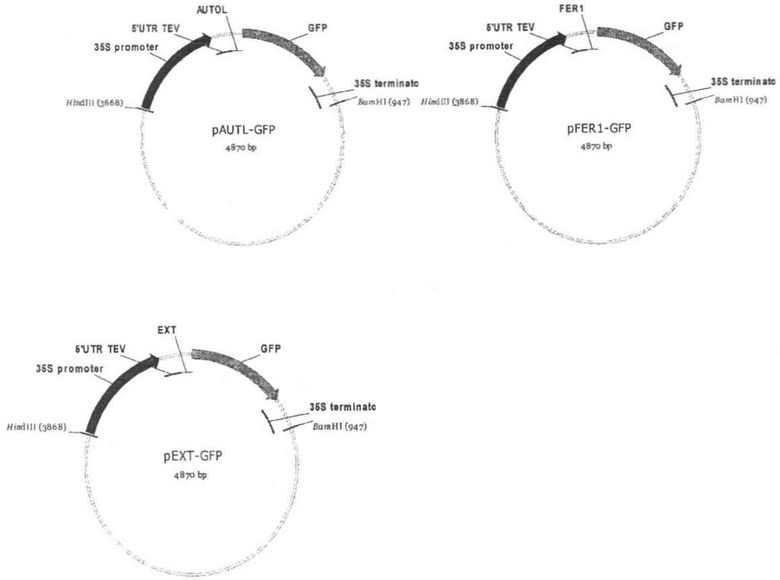
Для осуществления способа были сконструированы две плазмиды, несущие лидерные последовательности аутолизина и железотранспортного белка 1 Chlamydomonas reinhardtii и ген GFP, оптимизированный под экспрессию в растительных системах. Клонирование осуществляли путем амплификации соответствующих нуклеотидных последовательностей на геномной ДНК Chlamydomonas reinhardtii, гидролиза эндонуклеазой рестрикции NcoI и клонирования по этому же сайту в плазмиду pAWA319. Ориентацию вставки и соответствие структур рекомбинантных клонов подтверждали секвенированием. В результате были получены плазмиды pAUTL-GFP с сигнальной последовательностью аутолизина и pFER1-GFP с сигнальной последовательностью железотранспортного белка 1 Chlamydomonas reinhardtii. Плазмида pEXT-GFP, содержащая сигнальную последовательность экстенсина моркови, была получена ранее. Физические карты плазмид pAUTL-GFP, pFER1-GFP и pEXT-GFP приведены на чертеже. Экспрессионные кассеты переносили в бинарный вектор pBI101 по сайтам рестрикции BamHI и HindIII. Полученными плазмидами: pBI101-AUTL-GFP, pBI101-FER1-GFP и pBI101-EXT-GFP трансформировали клетки агробактериального штамма Agrobacterium tumifaciens EHA 105.
Пример 2. Транзиентная трансфекция клеток зеленой микроводоросли Chlorella с помощью агробактерии.
Используемые среды:
1. Среда LB (бакто-пептон 10 г/л, бакто-дрожжевой экстракт 5 г/л, NaCl 10 г/л).
2. Среда Eu (ацетат натрия 1 г/л, солянокислый гидролизат казеина 1 г/л, бакто-пептон 2 г/л, дрожжевой экстракт 2 г/л, CaCl2×2H2O 0,01 г/л).
3. Среда YEP (бакто-пептон 10 г/л, дрожжевой экстракт 10 г/л, NaCl 5 г/л).
4. Инокуляционная среда (CaCl2×2Н2О 0,01 г/л, 10 мМ MES, 10 мМ глюкоза, 200 мкМ ацетосирингон, бакто-пептон 10 г/л, дрожжевой экстракт 10 г/л, NaCl 5 г/л, рН 5,8).
5. Среда "Basal Medium" (Среда "Basal Medium" (2.0 г/л KNO3, 0.2 г/л K2HPO4, 0.2 г/л MgSO4. 7H2O, 5 мл/л раствор минералов).
Клетки микроводоросли Chlorella kessleri IPPAS C-9 выращивали в 20 мл среды Eu (в прозрачном флаконе объемом 50 мл) с ампициллином (100 мкг/мл) на шейкере при 28±2°С в режиме 18 часов свет интенсивностью около 3000 люм, 6 часов без света до достижения плотности, примерно равной 107 клеток/мл (примерно 3-5 дней).
Колонию агробактерий Agrobacterium tumifaciens EHA105, трансформированную pAUTL-GFP, pFER1-GFP или pEXT-GFP плазмидой, стерильной петлей инокулировали в 30 мл среды YEP, содержащей канамицин (50 мкг/мл) и рифампицин (25 мкг/мл), инкубировали на шейкере при 28°С до достижения оптической плотности (при 600 нм), равной 2 единицам (примерно двое суток).
По окончании инкубации отбирали 1 мл среды YEP с агробактериями, помещали в пробирку объемом 1,5 мл и центрифугировали при 3000 g в течение 3 мин. Супернатант отбрасывали, осадок ресуспендировали в 1 мл инокуляционной среды, содержащей канамицин (50 мкг/мл), переносили в пробирку объемом 15 мл и добавляли еще 3 мл инокуляционной среды с антибиотиком.
Далее пробирку с агробактериями в инокуляционной среде инкубировали при 28°С в течение 6 ч.
Затем отбирали 1 мл культуры хлореллы (107 клеток/мл), помещали в микропробирку объемом 1,5 мл и центрифугировали при 500 g в течение 10 мин. Супернатант отбрасывали, осадок ресуспендировали в 1 мл инокуляционной среды, содержащей агробактерии, объединяли с оставшимися 3 мл инокуляционной среды с агробактериями в 15 мл пробирке.
Далее коинкубировали клетки хлореллы и агробактерии на шейкере при 28°С в течение 14-16 ч.
По окончании инкубации пробирку центрифугировали при 500 g в течение 10 мин. Супернатант сливали, осаженные клетки ресуспендировали в 10 мл «Basal Medium», содержащей 60 мМ сахарозу и ампициллин (50 мкг/мл) и цефатоксим (250 мкг/мл). Инкубировали при 28±2°С в режиме 18 часов свет интенсивностью около 3000 люм, 6 часов без света в течение 2 дней.
Клетки отделяли центрифугированием, а супернатант анализировали в флюориметре при длине волны 490 /520 нм.
Результаты анализа флуоресценции супернатантов среды хлореллы, транзиентно трансфецированной агробактериями, содержащими различные секреторные варианты гена GFP, приведены в таблице.
Из таблицы 1 видно, что сигнальная последовательность из гена аутолизина Chlamydomonas reinhardtii в 4 раза более эффективна, чем общепринятая сигнальная последовательность экстенсина моркови.
Разработанный способ является оригинальным и существенно превосходит имеющиеся аналогичные технологии в мировой практике по времени проведения, 7-10 дней, тогда как получение трансформированных растений ряски болотной занимает 11-14 недель.
Использование предлагаемого способа позволит значительно упростить и повысить эффективность секреции рекомбинантных белков из микроводоросли рода хлорелла.
Источники информации
1. Kapila J, Rycke RD, Van Montagu M, Agenon G (1997) An Agrobacterium-mediated transient gene expression system for intact leaves. Plant Sci 122: 101-108.
2. Патент US 5889189 (А), МПК C12N 15/82, опубл. 30.03.1999.
3. Патент US 6815184 (B2), МПК C12N 15/82, опубл. 04.07.2002.
4. Stiefel, V., Ruiz-Avila, L., Raz, R., Valles, M.P., Ghez, J., Pages, M., Martinez-lzquierdo, J.A., Ludevid, M.D., Langdale, J.A., Nelson, Т., and Puigdoménech, P. (1990). Expression of a maize cell wall hydroxyproline-rich glycoprotein gene in early leaf and root vascular differentiation. Plant Cell 2, 785-793.
5. Zahur M, Maqbool A, Irfan M, Barozai MY, Qaiser U, Rashid B, Husnain T, Riazuddin S. Functional analysis of cotton small heat shock protein promoter region in response to abiotic stresses in tobacco using Agrobacterium-mediated transient assay. 2008 Mol Biol Rep.
6. Патент US 2003, 0097682, опубл. 20.11.2002.
7. Патент US 2009, 0282584, опубл. 16.07.2007.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pBinPLUS-ARS-EPSPS, ОБЕСПЕЧИВАЮЩАЯ ЭКСПРЕССИЮ ГЕНА АГРОБАКТЕРИАЛЬНОЙ 5-ЕНОЛПИРУВИЛ-ШИКИМАТ-3-ФОСФАТ-СИНТЕТАЗЫ В ТРАНСГЕННЫХ МИКРОВОДОРОСЛЯХ РОДА ХЛОРЕЛЛА | 2009 |
|
RU2407794C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК pBi101-IL18, КОДИРУЮЩАЯ СИНТЕЗ ИНТЕРЛЕЙКИНА-18 ЧЕЛОВЕКА В ТРАНСГЕННЫХ РАСТЕНИЯХ | 2005 |
|
RU2302460C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ ТАБАКА С ПОВЫШЕННЫМ СОДЕРЖАНИЕМ ПРОЛИНА | 2006 |
|
RU2324736C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ ТАБАКА С ПОВЫШЕННЫМ СОДЕРЖАНИЕМ ПРОЛИНА | 2006 |
|
RU2324737C1 |
| Способ получения фотодекарбоксилаз жирных кислот | 2020 |
|
RU2750455C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ МОРКОВИ, ПРОДУЦИРУЮЩИХ ИНТЕРЛЕЙКИН-10 ЧЕЛОВЕКА | 2007 |
|
RU2374321C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PGII, ОПРЕДЕЛЯЮЩАЯ СИНТЕЗ БЕТА-ИНТЕРФЕРОНА ЧЕЛОВЕКА В ТРАНСГЕННЫХ РАСТЕНИЯХ, И СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ, УСТОЙЧИВЫХ К ВИРУСНОЙ ИНФЕКЦИИ | 1994 |
|
RU2103361C1 |
| Способ получения безмаркерных трансгенных растений каланхоэ перистого, экспрессирующих ген цекропина Р1 | 2015 |
|
RU2624042C2 |
| СПОСОБ ПОЛУЧЕНИЯ ВЫСОКОПРОДУКТИВНЫХ ДРОЖЖЕВЫХ ШТАММОВ | 2020 |
|
RU2770894C1 |
| СПОСОБ УЛУЧШЕННОЙ ТРАНСФОРМАЦИИ С ИСПОЛЬЗОВАНИЕМ АГРОБАКТЕРИИ | 2012 |
|
RU2620975C2 |
Конструируют рекомбинантную плазмидную ДНК pAUTL-GFP, несущую гибридный ген, кодирующий зеленый флуоресцентный белок (GFP) и сигнальную последовательность аутолизина из Chlamydomonas reinhardtii. Полученной плазмидой трансформируют штамм Agrobacterium. Суспензию трансформированных агробактериальных клеток в логарифмической стадии роста инкубируют с клетками микроводоросли Chlorella в течение 14-16 часов. Уровень секреции белка оценивают, измеряя флуоресценцию культуральной среды. 1 з.п. ф-лы, 1 ил., 1 табл.
1. Способ секреции зеленого флуоресцентного белка (GFP) из растительных клеток, включающий конструирование рекомбинантной плазмидной ДНК, содержащей целевой ген, передачу полученной плазмиды в штамм Agrobacterium, кокультивирование трансформированных клеток Agrobacterium с растительными клетками с последующим определением эффективности секреции белка, отличающийся тем, что в качестве рекомбинантной плазмиды используют плазмиду pAUTL-GFP с сигнальной последовательностью аутолизина из Chlamydomonas reinhardtii, а в качестве растительных клеток используют клетки микроводоросли рода Chlorella.
2. Способ по п.1, отличающийся тем, что эффективность секреции белка оценивают путем измерения флуоресценции культуральной среды на флюориметре при длине волны 490/520 нм.
| СПОСОБ ПОЛУЧЕНИЯ БЕЛКОВЫХ ВЕЩЕСТВ | 2000 |
|
RU2250264C2 |
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| Дорожная спиртовая кухня | 1918 |
|
SU98A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| PROT EXPR PURIF, 1996 Mar, 7 (2): 220-8. | |||

