Область техники
Изобретение относится к биотехнологии, а именно к технологии получения дрожжевых штаммов продуцентов рекомбинантных белков с максимальным уровнем их продукции. Оно может быть использовано для получения дрожжевых штаммов-продуцентов любых рекомбинантных белков с максимальным уровнем продукции таковых.
Уровень техники
Получение рекомбинантных белков имеет огромное значение для производства биофармацевтических препаратов. Дрожжевые системы экспрессии являются стабильными, простыми в использовании и легко масштабируемыми системами получения рекомбинантных белков и имеют ряд преимуществ перед другими системами экспрессии, такими как Escherichia coli и культуры клеток млекопитающих. Дрожжевые клетки имеют системы фолдинга и пострансляционной модификации белков, что обеспечивает корректный фолдинг и посттрансляционные модификации рекомбинантных белков, аналогичные таковым высших эукариот, которые часто экспрессируются в неактивных формах в виде телец включения у Е. coli.
Хотя дрожжи обладают большими преимуществами для продукции чужеродных белков, уровень экспрессии рекомбинантных белков в этих системах может непредсказуем. Потенциально дрожжи обладают способностью секретировать несколько граммов белка на литр культуры, однако уровень продукции сильно зависит от рекомбинантного белка. Например, сывороточный альбумин человека и человеческий желатин может секретироваться в концентрации до нескольких г/л клетками дрожжей Pichia pastoris [Kobayashi, K. et al. High-level expression of recombinant human serum albumin from the methylotrophic yeast Pichia pastoris with minimal protease production and activation. J Biosci Bioeng 89, 55-61 (2000); Werten, M.W., van den Bosch, T.J., Wind, R.D., Mooibroek, H. & de Wolf, F.A. High-yield secretion of recombinant gelatins by Pichia pastoris. Yeast 15, 1087-1096 (1999)]. Однако в то же время продукция клетками дрожжей Pichia pastoris таких фармацевтических белков, таких как интерлейкин2 человека(hIL2), гранулоцитарный колониестимулирующий фактор (GCSF) ифактор роста тромбоцитов (PDGF) чрезвычайно низка [Anzur Lasnik, М., Nemec, М., Romih, R. & Schara, M. An EPR study of the secretion of G-CSF heterologous protein from Pichia pastoris. Biotechnol Bioeng 81, 768-774 (2003); Guan, B. et al. Constitutive expression of a rhIL-2-HSA fusion protein in Pichia pastoris using glucose as carbon source. Appl Biochem Biotechnol 171, 1792-1804 (2013)., Ernst, J.F. Efficient secretion and processing of heterologous proteins in Saccharomyces cerevisiae is mediated solely by the presegment of alpha-factor precursor. DNA7, 355-360 (1988)].
Известно, что на уровень экспрессии рекомбинантных белков в дрожжах метилотрофных дрожжей Pichia pastoris в значительной мере могут влиять такие необходимые элементы генетических конструкций, как сигнальная последовательность, промоторная и терминаторная последовательности, а так же другие цис- и транс- действующие элементы. В настоящее время существует ряд работ указывающих на необходимость подбора этих элементов для максимальной экспрессии корректно процессированных рекомбинантных белков в каждом конкретном случае (HuX, ChuJ, ZhangS, ZhuangY. ComparativeperformanceofS-adenosyl-L-methioninebiosynthesisanddegradationinPichiapastorisusingdifferentpromotersandnovelconsumptionin hibitors. Enzyme Microb Technol. 2014; 55:94-9, Hohenblum H, Gasser B, Maurer M, Borth N, Mattanovich D. Effects of gene dosage, promoters, and substrates on unfolded protein stress of recombinant Pichia pastoris. Biotechnol Bioeng. 2004; 85(4):367-75; Su GD, Huang DF, Han SY, Zheng SP, Lin Y. Display of Candida antarctica lipase В on Pichia pastoris and its application to flavor ester synthesis. ApplMicrobiolBiotechnol. 2010; 86(5): 1493-501), и нет никаких данных о существовании какого либо унивесального генетического окружения гена, обеспечивающего максимально возможный уровень экспрессии любого гена рекомбинантного белка. Используя комбинаторный перебор регуляторных и других генетических элементов, с целью найти эффективную их комбинацию, максимизирующую уровень продукции рекомбинантного белка, мы ограничены лишь небольшим количеством вариантов, поскольку анализ уровня экспрессии целевого белка для каждого из этих вариантов должен быть проведен отдельно, но в тоже время в одинаковых условиях для каждого клона. В настоящее время для таких масштабных скринирований используют 96-луночные планшеты [Neha Kaushik, 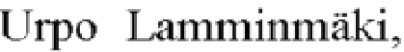 Navin Khanna, Gaurav Batra. Enhanced cell density cultivation and rapid expression-screening of recombinant Pichia pastoris clones in microscale. Sci Rep. 2020 May 4; 10(1):745]. Недостатком приведенной технологии остается трудоемкость и отсутствие масштабирования данного процесса, особенно в случае смены культуральной среды для индукции экспрессии целевого белка.
Navin Khanna, Gaurav Batra. Enhanced cell density cultivation and rapid expression-screening of recombinant Pichia pastoris clones in microscale. Sci Rep. 2020 May 4; 10(1):745]. Недостатком приведенной технологии остается трудоемкость и отсутствие масштабирования данного процесса, особенно в случае смены культуральной среды для индукции экспрессии целевого белка.
Известен подход скрининга дрожжевых клонов, продуцирующих рекомбинантные белки внутриклеточно с использованием прижизненного окрашивания клеток флуоресцентно меченными антителами к целевому белку [Yeast Intracellular Staining (yICS): Enabling High-Throughput, Quantitative Detection of Intracellular Proteins via Flow Cytometry for Pathway Engineering Brett D. Hill ACS Synth. Biol. 2020, 9, 8, 2119-2131]. Клоны, продуцирующие целевой белок на разном уровне, будут обладать разной флуоресценцией и могут быть отобраны цитофлуориметрически. Недостатком приведенной технологии является неприменимость ее для секретирующихся белков.
Известны подходы скрининга дрожжевых клонов с использованием микрофлюидной технологии, когда в каплю помещаются единичничные дрожжевые клетки-продуценты секретируемых белков, обладающих каталитической активностью разного уровня, вместе с собственным флуорогенным субстратом [Terekhov SS.Microfluidic droplet platform for ultrahigh-throughput single-cells creening of biodiversity. Proc Natl Acad Sci USA. 2017 Mar 7; 114(10):2550-2555]. Клоны, обладающие наибольшей активностью, могут быть отобраны из смеси клонов за счет максимальной флуоресценции. Это подход может быть применен аналогично для отбора клонов, обладающих лучшей продуктивностью. Недостатком приведенной технологии является то, что целевым белком в таком случае должен являться фермент, для которого известен флуорогенный субстрат или же сам целевой белок должен обладать флуоресценцией. Однако, большинство белков, востребованных для биотехнологического применения, этими свойствами не обладают.
Известен подход к эффективному отбору дрожжевых высокопродуктивных клонов из смеси клонов, содержащих избыток клонов с низкой продукцией с использованием цитофлуориметрии [RakestrawJA, BaskaranAR, WittrupKD. A flow cytometric assay for screening improved heterologous protein secretionin yeast. Biotechnol Prog. 2006 Jul-Aug; 22(4): 1200-8; J Andy Rakestraw. Directed evolution of a secretory leader for the improved expression of heterologous proteins and full-length antibodies in Saccharomyces cerevisiae. Biotechnol Bioeng. 2009 Aug 15; 103(6): 1192-201] на примере одноцепочечного антитела 4m5.3, связывающего флуоресцеин. Скрининг осуществлялся через цитофлуорметрический отбор клеток, ковалентно модифицированных флуресцеином за счет связывания с ним секретируемого клеткой антитела 4m5.3, содержащего FLAG эпитоп. Клоны, обладающие наибольшей продукцией, могут быть отобраны из смеси клонов за счет максимальной флуоресценции клеток, окрашенных FITC-мечеными анти-FLAG антителами. Недостатком приведенной технологии является необходимость получения и модификации лиганда, с которым связывается целевой белок. Далеко не всегда таковой имеется, а если и имеется, то его получение и модификация может представлять собой задачу сравнимую по сложности с получением целевого белка.
Ближайшим аналогом заявленного изобретения является публикация [Peter Kotter, Julia Е. Weigand, Britta Meyer, Karl-Dieter Entian and Beatrix Suess. A fast and efficient translational control system for conditional expression of yeast genes. Nucleic Acids Research, 2009, Vol. 37, No. 18], в которой описано уменьшение уровня экспрессии зеленого флуоресцентного белка при введении в его промоторную зону последовательностей тетрациклин-связывающего аптамера, которая была принята в качестве прототипа.
Таким образом, технической проблемой, на решение которой направлено настоящее изобретение, является способ получения клеток дрожжей с использованием простого и высокопроизводительного метода проточной цитофлуориметрии; осуществление отбора клеток дрожжей с максимальным уровнем продукции рекомбинантного целевого белка из клонов репертуара любого размера, обладающих разным уровнем его продукции.
Техническим результатом является способ получения клеток дрожжей обладающих максимальным уровнем продукции рекомбинантного целевого белка из репертуара клонов любого размера, обладающих разным уровнем его продукции методом проточной цитофлуориметрии.
Технический результат достигается за счет проведения проточного цитофлуориметрического анализа смеси клонов, в которых флуоресценция маркерного флуоресцентного белка прямо пропорциональна концентрации мРНК целевого белка, и отбора клеток, обладающих максимальным уровнем флуоресценции, и, следовательно, обладающих максимальным уровнем экспрессии целевого белка.
Для решения технической проблемы и достижения заявленного технического результата предлагается рекомбинантная плазмидная ДНК pGAP3tcGFP (SEQ ID NO 1) для экспрессии маркерного рекомбинантного белка GFP (зеленый флуоресцентный белок), имеющая длину 4307 пар оснований и состоящая из следующих ключевых генетических элементов:
a. укороченный конститутивный промотор гена глицерид-3-фосфат дегидрогеназы GAP дрожжей Pichia pastoris (указанные структурные признаки отражены во введенном обозначении «GAP»);
b. трех тандемно расположенных нуклеотидных последовательностей аптамера к тетрациклину 32sh, представленной в SEQ ID NO 2 (указанные структурные признаки отражены во введенном обозначении «3tc»);
c. нуклеотидную последовательность кДНК гена зеленого флуоресцентного белка (указанные структурные признаки отражены во введенном обозначении «GFP»);
d. нативный терминатор и сигнал полиаденилирования гена алкоголь оксидазыАОХ1 дрожжей Pichia pastoris;
e. точку начала репликации плазмид pUCori;
f. конститутивную промоторную последовательность гена фактора элонгации 1 Saccharomyces cerevisiae TEF1;
g. конститутивную синтетическую промоторную последовательность ЕМ7;
h. последовательность гена блеомицина BleoR Streptoalloteichus hindustanus, продукт которого обеспечивает устойчивость трансформантов к антибиотику зеоцину;
i. 3'-концевую терминаторную последовательность гена цитохрома C1 CYC1 Saccharomyces cerevisiae.
Другим объектом изобретения является штамм-реципиент Yst-pGAP3tcGFP метилотрофных дрожжей P.pastoris полученный путем трансформации клеток метилотрофных дрожжей P.pastoris штамма GS115 рекомбинантной плазмидной ДНК pGAP3tcGFP.
Предлагается способ получения любого целевого рекомбинантного белка, включающий в себя стадии:
1. трансформация штамма-реципиента Yst-pGAP3tcGFP рекомбинантной плазмидной ДНК, содержащей следующие обязательные генетические элементы:
- кДНК целевого гена;
- любые промоторные и терминаторные последовательности для экспрессии целевого белка;
- последовательность любого сигнального пептида для секреции целевого белка;
- лежащую в приделах транскрибируемой области гена целевого белка искусственно сконструированную нуклеотидную последовательность интрона (SEQ ID NO 6), с включенной в его состав нуклеотидной последовательностью аптамера к тетрациклину;
- любой селекционный маркер, для отбора трансформированных клеток, кроме последовательности гена блеомицина BleoR Streptoalloteichus hindustanus, продукт которого обеспечивает устойчивость трансформантов к антибиотику зеоцину.
2. Культивирование выросших клеток в культуральной среде, содержащей 55 мкМ тетрациклина;
3. Отбор 10% клеток, обладающих максимальной зеленой флуоресценцией методом проточной цитофлуориметрии.
4. Анализ уровня экспрессии отобранных клонов и выбор одного с максимальным уровнем продукции.
Раскрытие изобретения
В основе предлагаемой системы отбора (фиг. 1) лежит использование РНК-аптамера к тетерациклину (32sh) [S. Hanson, K. Berthelot, В. Fink, J.Е.G. McCarthyandB. Suess Molecular Microbiology (2003) 49 (6), 1627-1637 Tetracycline-aptamer-mediated translational regulationin yeast]. При связывании тетрациклина несколькими повторами РНК-аптамера расположенными тандемно в 5' нетранслируемой области мРНК белка приводит к значительному снижению уровня синтеза белка [P. Kotter, J.Е. Weigand, B.Meyer, K.-D. EntianandB. Suess. A fast and efficient translational control system for conditional expression of yeast genes. Nucleic Acids Research, 2009, Vol. 37, No. 18 е120]. В качестве такого маркерного белка в патентуемой системе используется зеленый флуоресцентный белок (GFP), в 5' нетранслируемой области мРНК которого находятся три тандемно расположенные последовательности аптамера к тетрациклину 32sh (фиг. 1А). При добавлении к дрожжевым клеткам тетрациклина происходит уменьшение синтеза GFP (фиг. 1Б) за счет появления стабильных структур 32sh - тетрациклин, мешающих связыванию 80S субъединицы рибосомы со сканирующей 40S субъединицей [S. Hanson, K. Berthelot, В. Fink, J.E.G. McCarthyand B. SuessMolecular Microbiology (2003) 49 (6), 1627-1637 Tetracycline-aptamer-mediated translational regulation in yeast]. При появлении в клетке пула молекул мРНК целевого белка, содержащих структуры 32sh в составе сплайсированных интронов (1В), конкуренция за связывание тетрациклина мРНК GFP и сплайсированными интронами мРНК целевого белка приводит к частичной разблокировке синтеза GFP (1Г). В клетках, где выше концентрация мРНК целевого белка за счет более высокого уровня транскрипции его гена или же за счет меньшей скорости деградации будет в большей степени накапливаться GFP (1Д). Выбор клона, клетки которого содержат максимальное количество мРНК целевого белка, осуществляется с помощью проточной цитофлуориметрии. Уровень продукции целевого белка такими клонами будет выше за счет более высокой концентрации мРНК целевого белка. С помощью предлагаемой системы можно получать высокопродуктивные дрожжевые штаммы, варьируя генетическое окружение гена рекомбинантного белка, включающее в себя промоторные, терминаторные и сигнальные сигнальные последовательности, а также изучать влияние различных цис- и транс- действующих элементов на уровень продукции целевого белка.
В процессе поэтапного выполнения патентных исследований осуществлялись:
- создание рекомбинантной плазмидной ДНК pGAP3tcGFP (SEQ ID NO: 1) на основе искусственно синтезированного фрагмента ДНК (SEQ ID NO: 2), содержащего нуклеотидную последовательность трех тандемно расположенных последовательностей аптамера к тетрациклину 32sh и кДНК зеленого флуоресцентного белка (GFP) путем их клонирования в плазмиду pGAPZαA;
- трансформация метилотрофных дрожжей P.pastoris полученной рекомбинантной плазмидной ДНК pGAP3tcGFP, с последующим получением штамма-реципиента Yst-pGAP3tcGFP и определение рабочих концентраций тетрациклина;
- апробация предлагаемого способа получения на примере красного флуоресцентного белка (RFP) в качестве целевого.
Создание генетической конструкции pGAP3tcGFP (SEQ ID NO: 1)
Искусственно синтезированный фрагмент ДНК, содержащий нуклеотидную последовательность трех тандемно расположенных последовательностей аптамера к тетрациклину 32sh (SEQ ID NO: 2) был клонирован в плазмиду pGAPZαA ("Invitrogen", США) с использованием эндонуклеаз рестрикции MfeI и BstBI. Далее в полученную конструкцию был клонирован ПЦР-продукт, содержащий последовательность кДНК зеленого флуоресцентного белка (GFP) с использованием эндонуклеаз рестрикции BstBI и NotI с получением рекомбинантной плазмидной ДНК pGAP3tcGFP (SEQ ID NO: 1) (фиг. 2,).
Обозначение «pGAP» в рекомбинантной плазмидной ДНК pGAP3tcGFP отражает содержание структурных элементов плазмидной ДHK pGAPZαA ("Invitrogen", США). Рекомбинантная плазмидная ДНК pGAP3tcGFP содержит:
- укороченный конститутивный промотор гена глицерид-3-фосфат дегидрогеназы GAP дрожжей Pichia pastoris (указанные структурные признаки отражены во введенном обозначении «GAP»),
- нуклеотидную последовательность трех тандемно расположенных последовательностей аптамера к тетрациклину 32sh (указанные структурные признаки отражены во введенном обозначении «3tc»),
- нуклеотидную последовательность кДНК гена зеленого флуоресцентного белка (указанные структурные признаки отражены во введенном обозначении «GFP»),
- нативный терминатор и сигнал полиаденилирования гена алкоголь оксидазы АОХ1 дрожжей Pichia pastoris, точку начала репликации плазмид pUCori.
- конститутивную промоторную последовательность гена фактора элонгации 1 Saccharomyces cerevisiae TEF1,
- конститутивную синтетическую промоторную последовательность ЕМ7,
- последовательность гена блеомицина BleoR Streptoalloteichus hindustanus, продукт которого обеспечивает устойчивость трансформантов к антибиотику зеоцину,
- 3'-концевую терминаторную последовательность гена цитохрома C1 CYC1 Saccharomyces cerevisiae.
Получение штамма-реципиента Yst-pGAP3tcGFP и определение рабочих концентраций тетрациклина
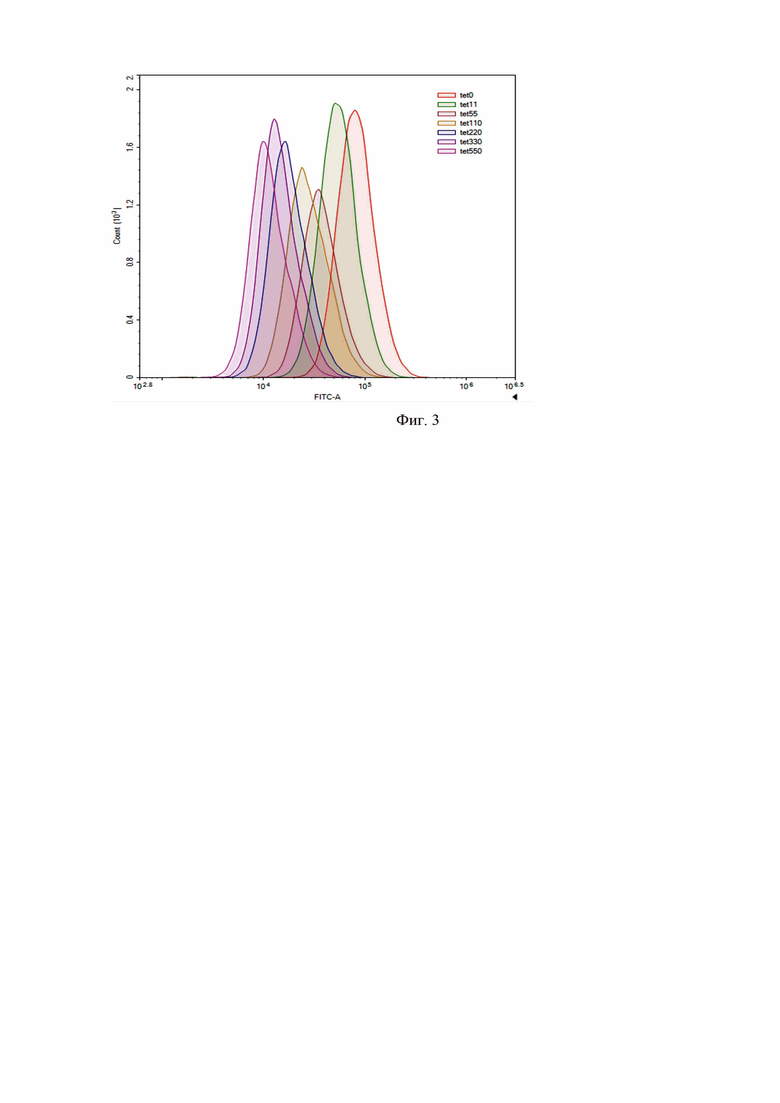
Клетки P.pastoris штамма GS115 были трансформированы рекомбинантной плазмидной ДНК pGAP3tcGFP, линеаризованной эндонуклеазой рестрикции SphI. Селекцию трансформантов проводили на агаризованной среде YPD, содержащий антибиотик зеоцин. Среди выросших трансформантов случайным образом была выбрана единичная колония - штамм реципиент Yst-pGAP3tcGFP. Клетки штамма Yst-pGAP3tcGFP выращивали 24 часа при добавлении различных концентраций тетрациклина (до конечной концентрации 0-550 мкМ). Методом цитофлуориметрии было установлено падение интенсивности флуоресценции в 10 раз (фиг. 3) в исследуемом диапазоне концентраций.
Апробация предлагаемого способа получения на примере красного флуоресцентного белка (RFP) в качестве целевого
Для апробации разработанной системы в качестве целевого белка использовали красный флуоресцентный белок (RFP), отбор проводили из смеси клеток дрожжей P.pastoris, трансформированных рекомбинантной плазмидной ДНК pPIC9Kintron-tc-alphaRFP (SEQ ID NO: 3), pPIC9Kintron-tc-alpha_shortRFP (SEQ ID NO: 4) и pPIC9Kintron-tc-hsaRFP (SEQ ID NO: 5) содержащими одну из нуклеотидных последовательностей сигнальной последовательности (препро- или пре-последовательность альфа-фактора дрожжей Saccharomyces cerevisiae, или сигнальную последовательность человеческого сывороточного альбумина соответственно) в единой рамке считывания с кДНК гена RFP. Влияние данных сигнальных последовательностей на уровень экспрессии RFP клетками дрожжей P.pastoris оценивалось нами ранее [A. Zlobin. Description of Newly Selected Catalytic Bioscavengers Against Organophosphorus Compounds Revealed Reactivation Stimulus Mediated by Histidine Residue in the Acyl-Binding Loo. Front Pharmacol. 2018 Aug 3; 9:834.QM/MM]. Установлено, что сигнальные последовательности человеческого сывороточного альбумина (hsa) и пре-последовательность альфа-фактора дрожжей Saccharomyces cerevisiae (alpha) обеспечивают значительно больший уровень экспрессии RFP чем препро-альфа-фактора дрожжей Saccharomyces cerevisiae (alpha-sort), но в то же время в случае сигнальной последовательности hsa диапазон уровня экспрессии RFP различными клонами значительно больше, чем в случае других сигнальных последовательностей. В составе рекомбинантных плазмидных ДНК в области, соответствующей 3' нетранслируемой области ДНК гена RFP находилась искусственно сконструированная последовательность интрона, с включенной в его состав нуклеотидной последовательностью аптамера к тетрациклину 32sh.
Рекомбинантные плазмидные ДНК pPIC9Kintron-tc-alphaRFP, pPIC9Kintron-tc-alpha_shortRFP (SEQ ID NO: 4) и pPIC9Kintron-tc-hsaRFP (SEQ ID NO: 5) (фиг. 4, приложение 1) получали следующим образом. Искусственно синтезированный фрагмент ДНК (SEQ ID NO: 6), содержащий нуклеотидную последовательность аптамера к тетрациклину 32sh был клонирован в плазмиду pPIC9k ("Invitrogen", США) с использованием эндонуклеазы рестрикции NotI. Далее, в полученную конструкцию были клонированы ПЦР-продукты, содержащий сигнальные последовательности, слитые с кДНК красного флуоресцентного белка (RFP), с использованием эндонуклеаз рестрикции BamHI и NotI с получением рекомбинантных плазмидных ДНК pPIC9Kintron-tc-alphaRFP, pPIC9Kintron-tc-alpha_shortRFP и pPIC9Kintron-tc-hsaRFP (SEQ ID NO: 3, SEQ ID NO: 4, SEQ ID NO: 5).
Обозначение «pPIC9» в рекомбинантных плазмидных ДНК pPIC9Kintron-tc-alphaRFP, pPIC9Kintron-tc-alpha_shortRFP и pPIC9Kintron-tc-hsaRFP отражает содержание структурных элементов плазмидной ДНК pPIC9k ("Invitrogen", США). Рекомбинантные плазмидные ДНК pPIC9Kintron-tc-alphaRFP, pPIC9Kintron-tc-alpha_shortRFP и pPIC9Kintron-tc-hsaRFP содержат:
- индуцибельный промотор гена алкогольоксидазы 1 АОХ1 дрожжей Pichia pastoris;
- одну из нуклеотидных последовательностей сигнального пептида препро- или пре-последовательность альфа-фактора дрожжей Saccharomyces cerevisiae, или сигнальную последовательность человеческого сывороточного альбумина (указанные структурные признаки отражены во введенном обозначении «alpha», «alpha_short» и «hsa» соответственно), находящихся в единой рамке считывания с кДНК гена красного флуоресцентного белка (RFP) (указанные структурные признаки отражены во введенном обозначении «RFP»);
- искусственно сконструированную нуклеотидную последовательность интрона, с включенной в его состав нуклеотидной последовательностью аптамера к тетрациклину (указанные структурные признаки отражены во введенном обозначении «intron» и «tc» соответственно);
- нативный терминатор и сигнал полиаденилирования гена алкоголь оксидазы АОХ1 дрожжей Pichia pastoris;
- нуклеотидную последовательность ДНК фрагмента, соответствующий 3' нетранслируемой области мРНК гена алкогольоксидазы 1 АОХ1 дрожжей Pichia pastoris фрагмент;
- точку начала репликации плазмид pUCori;
- последовательность Kan гена аминогликозил 3'-фосфотрансферазы тип 1 (APHI) E.coli вместе с собственной промоторной последовательностью;
- последовательность bla гена β-лактамазы, продукт которого обеспечивает устойчивость трансформантов E.coli к антибиотику ампициллин; последовательность гена HIS4, продукт которого позволяет комплементировать ауксотрофность по гистидину у трансформантов дрожжей P.pastoris штамма GS115.
Полученные рекомбинантные плазмидные ДНК pPIC9Kintron-tc-alphaRFP, pPIC9Kintron-tc-alpha_shortRFP и pPIC9Kintron-tc-hsaRFP были фрагментированы эндонуклеазой рестрикции BglII, рестриктные смеси объединяли объединенной смесью трансформировали клетки P. pastoris штамма-реципиента Yst-pGAP3tcGFP. Такой способ линеаризации генетических конструкций на основе плазмидной ДНК рРIС9k обеспечивает интеграцию плазмидной ДНК в дрожжевой геном по механизму Ω-рекомбинации, что позволяет избежать интеграции в дрожжевой геном более, чем одной копии плазмидной ДНК. Таким образом, можно утверждать, что каждый клон, полученный в результате трансформации, несет только одну из экспрессионных генетических конструкций - pPIC9Kintron-tc-alphaRFP, pPIC9Kintron-tc-alpha_shortRFP или pPIC9Kintron-tc-hsaRFP и разница в уровне продукции RFP различными клонами определяется одной из трех используемых сигнальных последовательностей. Выросшие клоны объединяли, подращивали и индуцировали синтез RFP добавлением метанола. Через 24 часа после индукции к клеточной суспензии добавляли раствор тетрациклина до концентрации 55 мкМ и культивировали еще 24 часа. Далее с помощью цитофлуориметрии пул клеток разделяли на четыре когорты (A-D) по уровню интенсивности зеленой флуоресценции (фиг. 5). У каждого из 10 случайно выбранных клонов для каждой когорты и из несортированных клеток определяли уровень продукции RFP с помощью флуориметрии культуральной среды. Для выбранных клонов наблюдали зависимость, при которой средний уровень продукции целевого белка (в данном случае RFP) клетками отобранной когорты прямо пропорционален интенсивности зеленой флуоресценции данной когорты при их отборе (фиг. 6). Кроме того, клоны, обладающие наибольшим уровнем продукции RFP находились когорте А, обладающей на момент сортировки набольшей зеленой флуоресценцией. Присутствие смеси клонов разного уровня экспрессии, сходных с таковой с пулом несортированных клеток, в когорте D, обладающей на момент сортировки наименьшей зеленой флуоресценцией, может объясняться нахождением этих клеток в тех фазах клеточного цикла, в которых не происходит или снижен синтез белка.
Таким образом, апробация заявляемого способа получения высокопродуктивных дрожжевых штаммов на примере красного флуоресцентного белка (RFP) показала, что заявляемым способом возможно эффективно отбирать клоны, обладающие наиболее высокой продукцией целевого белка из пула клеток, обладающих различным уровнем продукции. К настоящему времени в мире не существует системы отбора, которая бы позволила отбирать из смеси клеток с различным уровнем экспрессии целевого белка высокопродуктивные дрожжевые штаммы, варьируя в широком диапазоне генетическое окружение гена рекомбинантного белка, включающее в себя промоторные, терминаторные и сигнальные сигнальные последовательности, а также изучать влияние различных цис- и транс- действующих элементов на уровень продукции целевого белка.
Заявленное изобретение иллюстрируют следующими графическими материалами:
Фиг. 1. Схема патентуемой системы отбора.
Фиг. 2. Схема рекомбинантной плазмидной ДНК pGAP3tcGFP
Фиг. 3. Гистограмма распределения дрожжевых клеток штамма-реципиента Yst-pGAP3tcGFP P.pastoris, культивированных в присутствии различных концентраций тетрациклина, по уровню зеленой флуоресценции.
Фиг. 4. Схема рекомбинантной плазмидной ДНК pPIC9Kintron-tc-signalRFP (Б).
Фиг. 5. Гистограмма распределения дрожжевых клеток P.pastoris трансформированных смесью рекомбинантных плазмидных ДНК - pPIC9Kintron-tc-alphaRFP, pPIC9Kintron-tc-alpha_shortRFP или pPIC9Kintron-tc-hsaRFP, культивированных в присутствии 55 мкМ тетрациклина, по уровню зеленой флуоресценции.
Фиг. 6. Диаграмма распределения результатов флуориметрического анализа аликвот культуральной среды, отражающая уровень экспрессии RFP клонами клеток когорт A-D и несортированных клетками.
Осуществление изобретения
Осуществление заявляемого изобретения иллюстрируется приведенными ниже примерами 1-3.
ПРИМЕР 1
Создание рекомбинантной плазмидной ДНК pGAP3tcGFP.
Искусственно синтезированный фрагмент ДНК, содержащий нуклеотидную последовательность трех тандемно расположенных последовательностей аптамера к тетрациклину 328sh клонируют в плазмиду pGAPZαA («Invitrogen», США) с использованием эндонуклеаз рестрикции MfeI и BstBI. Фрагмент ДНК, содержащий последовательность кДНК зеленого флуоресцентного белка (GFP), амплифицируют методом ПЦР с плазмиды pBAV1KT5gfp, («Addgene», США), с использованием олигонуклеотидов:
(5') aatattcgaaacgatgggcggcggcagtaaagga
(5') aatagcggccgctttttgtatagttcatccatgccatg
Полученный фрагмент ДНК клонируют вполученную вышеплазмидную ДНК с использованием эндонуклеазрестрикции BstBI и NotI, с получением рекомбинантной плазмидной ДНК pGAP3tcGFP (Фиг. 2).
ПРИМЕР 2
Получение штамма-реципиента Yst-pGAP3tcGFP и определение рабочих концентраций тетрациклина.
Рекомбинантную плазмидную ДНК pGAP3tcGFP линеаризуют эндонуклеазой рестрикции SphI, и 1 мкг рестрикционной смеси фрагментов ДНК трансформируют электрокомпетентные клетки P. pastoris штамма GS115 методом электропорации. Селекцию трансформантов проводят на агаризованной среде YPD (20 г/л бактотриптона, 10 г/л дрожжевого экстракта, 50 мМ глюкоза, 2 г/л агар) содержащий антибиотик зеоцин. Среди выросших колоний случайным образом выбирают одну, получая таким образом, штамм-реципиент Yst-pGAP3tcGFP. Колонию Yst-pGAP3tcGFP подращивают в жидкой среде YPD, содержащую антибиотик зеоцин и клеточную суспензию используют для получения музейной культуры.
Единичную колонию штамма Yst-pGAP3tcGFP выращивают 24 часа в жидкой среде YPD, далее добавляют к клеточной суспензии различные концентрации тетрациклина (0-550 мкМ) и продолжают культивирование еще 24 часа. По окончании культивирования уровень содержания GFP в клетках штамма Yst-pGAP3tcGFP анализируют методом проточной цитофлуориметрии. Установлено, что увеличение концентрации тетрациклина в культуральной среде приводит к падению интенсивности флуоресценции в 10 раз в исследуемом диапазоне концентраций (фиг. 3).
ПРИМЕР 3
Апробация созданной системы отбора на примере красного флуоресцентного белка (RFP) в качестве целевого.
Рекомбинантные плазмидные ДНК pPIC9Kintron-tc-alphaRFP, pPIC9Kintron-tc-alpha_shorfRFP и pPIC9Kintron-tc-hsaRFP (фиг. 4) получают следующим образом. Искусственно синтезированный фрагмент ДНК, содержащий нуклеотидную последовательность аптамера к тетрациклину 32sh клонируют в плазмиду pPIC9k ("Invitrogen", США) с использованием эндонуклеазы рестрикции NotI. ПЦР-продукты, содержащий сигнальные последовательности, слитые с кДНК красного флуоресцентного белка (RFP), амплифицируютс матрицы плазмидных ДНК, полученных ранее [A. Zlobin Description of Newly Selected Catalytic Bioscavengers Against Organophosphorus Compounds Revealed Reactivation Stimulus Mediated by Histidine Residue in the Acyl-Binding Loop.Front Pharmacol, 2018 Aug 3; 9:834.QM/MM] с использованием прямых олигонуклеотидов:
(5') cgaaggatccaaacgatgagatttccttcaatttttactgctg
(5') cgaaggatccaaacgatgagatttccttcaatttttactgc
(5') cgaaggatccaaacgatgaagtgggttacctttatctc
в случае tc-alphaRFP, tc-alpha_shortRFP и tc-hsaRFP, соответственно, и обратного олигонуклеотида:
(5') etcatgcggccgccctagggaattcactagtgtt
и клонируют ПЦР-продуктыс использованием эндонуклеаз рестрикции BamHI и NotI в полученную ранее конструкцию, содержащую последовательность аптамера 32sh, с получением рекомбинантных плазмидных ДНК pPIC9Kintron-tc-alphaRFP, pPIC9Kintron-tc-alpha_shortRFP и pPIC9Kintron-tc-hsaRFP.
Полученные рекомбинантные плазмидные ДНК pPIC9Kintron-tc-alphaRFP, pPIC9Kintron-tc-alpha_shortRFP и pPIC9Kintron-tc-hsaRFP обрабатывают эндонуклеазой рестрикции BglII, рестриктные смеси объединяют и объединенной смесью трансформируют клетки P.pastoris штамма-реципиента Yst-pGAP3tcGFP. Селекцию трансформантов проводят на минимальной среде RDB, содержащий антибиотик зеоцин.
Выросшие клоны объединяют, подращивают 24 часа в жидкой среде YPD и индуцируют синтез RFP добавлением метанола в среде BMMY, рН=6.0 (20 г/л бактотриптона, 10 г/л дрожжевого экстракта, 100 mM калий-фосфатный буфер, рН=6.0, 13.4 г/л дрожжевой азотной основы, 1.6 мкМ биотин, 1% метанол). Через 24 часа после индукции к клеточной суспензии добавляют раствор тетрациклина до концентрации 55 мкМ и культивируют еще 24 часа. Далее с помощью цитофлуориметрии пул клеток разделяют на четыре когорты (A-D) по уровню интенсивности зеленой флуоресценции: А - 30% клеток, обладающей максимально зеленой флуоресценцией; и далее по убыванию флуоресценции В - 50%, С - 15%, D - 5% (фиг. 5). Несортированные клетки и клетки из каждого отобранного пула высевают на чашку с агаризованной средой YPD до получения единичных колоний. Каждый из 10 случайно выбранных клонов для каждой когорты и из несортированных клеток подращивают 24 часа в жидкой среде YPD и индуцируют синтез RFP добавлением метанола в среде BMMY, и культивируют еще 72 часа с добавлением метанола до 1% каждые 24 часа. По окончании культивирования клетки удаляют центрифугированием и определяют уровень продукции RFP с помощью флуориметрии осветленной культивационной среды. Полученные результаты свидетельствуют, что уровень продукции целевого белка (в данном случае RFP) прямо пропорционален интенсивности зеленой флуоресценции при их отборе (фиг. 6).
Приложение 1
SEQIDNO:1
Нуклеотидная последовательность экспрессионной генетической конструкции pGAP3tcGFP:
gatctctgctactctggtcccaagtgaaccaccttttggaccctattgaccggaccttaacttgccaaacctaaacgcttaatgcctcagacgttttaatgcctctcaacacctccaaggttgctttcttgagcatgcctactaggaactttaacgaactgtggggttgcagacagtttcaggcgtgtcccgaccaatatggcctactagactctctgaaaaatcacagttttccagtagttccgatcaaattaccatcgaaatggtcccataaacggacatttgacatccgttcctgaattatagtcttccaccgtggatcatggtgttcctttttttcccaaagaatatcagcatcccttaactacgttaggtcagtgatgacaatggaccaaattgttgcaaggtttttctttttctttcatcggcacatttcagcctcacatgcgactattatcgatcaatgaaatccatcaagattgaaatcttaaaattgcccctttcacttgacaggatccttttttgtagaaatgtcttggtgtcctcgtccaatcaggtagccatctctgaaatatctggctccgttgcaactccgaacgacctgctggcaacgtaaaattctccggggtaaaacttaaatgtggagtaatggaaccagaaacgtctcttcccttctctctccttccaccgcccgttaccgtccctaggaaattttactctgctggagagcttcttctacggcccccttgcagcaatgctcttcccagcattacgttgcgggtaaaacggaggtcgtgtacccgacctagcagcccagggatggaaaagtcccggccgtcgctggcaataatagcgggcggacgcatgtcatgagattattggaaaccaccagaatcgaatataaaaggcgaacacctttcccaattttggtttctcctgacccaaagactttaaatttaatttatttgtccctatttcaatcaattgggcctaaaacataccagatcgccacccgcgctttaatctggagaggtgaagaatacgaccacctaggccttaagaaccggtaaaacataccagatcgccacccgcgctttaatctggagaggtgaagaatacgaccacctaccggtcaacaacaacaacaacaacaacaacaactcgaggcctaaaacataccagatcgccacccgcgctttaatctggagaggtgaagaatacgaccacctaggcctcgagaacatttcgaaacgatgagtaaaggagaagaacttttcactggagttgtcccaattcttgttgaattagatggtgatgttaatgggcacaaattttctgtcagtggagagggtgaaggtgatgcaacatacggaaaacttacccttaaatttatttgcactactggaaaactacctgttccatggccaacacttgtcactactctgacgtatggtgttcaatgcttttcccgttatccggatcatatgaaacggtatgactttttcaagagtgccatgcccgaaggttatgtacaggaacgcactatatctttcaaagatgacgggaactacaagacgcgtgctgaagtcaagtttgaaggtgatacccttgttaatcgtatcgagttaaaaggtattgattttaaagaagatggaaacattctcggacacaaattggagtacaactataactcacacaatgtatacatcacggcagacaaacaaaagaatggaatcaaagctaacttcaaaattcgccacaacattgaagatggatccgttcaactagcagaccattatcaacaaaatactccaattggcgatggccctgtccttttaccagacaaccattacctgtcgacacaatctgcccttttgaaagatcccaacgaaaagcgtgaccacatggtccttcttgagtttgtaactgctgctgggattacacatggcatggatgaactatacaaaaagcggccgccagctttctagaacaaaaactcatctcagaagaggatctgaatagcgccgtcgaccatcatcatcatcatcattgagtttgtagccttagacatgactgttcctcagttcaagttgggcacttacgagaagaccggtcttgctagattctaatcaagaggatgtcagaatgccatttgcctgagagatgcaggcttcatttttgatacttttttatttgtaacctatatagtataggattttttttgtcattttgtttcttctcgtacgagcttgctcctgatcagcctatctcgcagctgatgaatatcttgtggtaggggtttgggaaaatcattcgagtttgatgtttttcttggtatttcccactcctcttcagagtacagaagattaagtgagaccttcgtttgtgcggatcccccacacaccatagcttcaaaatgtttctactccttttttactcttccagattttctcggactccgcgcatcgccgtaccacttcaaaacacccaagcacagcatactaaattttccctctttcttcctctagggtgtcgttaattacccgtactaaaggtttggaaaagaaaaaagagaccgcctcgtttctttttcttcgtcgaaaaaggcaataaaaatttttatcacgtttctttttcttgaaatttttttttttagtttttttctctttcagtgacctccattgatatttaagttaataaacggtcttcaatttctcaagtttcagtttcatttttcttgttctattacaactttttttacttcttgttcattagaaagaaagcatagcaatctaatctaaggggcggtgttgacaattaatcatcggcatagtatatcggcatagtataatacgacaaggtgaggaactaaaccatggccaagttgaccagtgccgttccggtgctcaccgcgcgcgacgtcgccggagcggtcgagttctggaccgaccggctcgggttctcccgggacttcgtggaggacgacttcgccggtgtggtccgggacgacgtgaccctgttcatcagcgcggtccaggaccaggtggtgccggacaacaccctggcctgggtgtgggtgcgcggcctggacgagctgtacgccgagtggtcggaggtcgtgtccacgaacttccgggacgcctccgggccggccatgaccgagatcggcgagcagccgtgggggcgggagttcgccctgcgcgacccggccggcaactgcgtgcacttcgtggccgaggagcaggactgacacgtccgacggcggcccacgggtcccaggcctcggagatccgtcccccttttcctttgtcgatatcatgtaattagttatgtcacgcttacattcacgccctccccccacatccgctctaaccgaaaaggaaggagttagacaacctgaagtctaggtccctatttatttttttatagttatgttagtattaagaacgttatttatatttcaaatttttcttttttttctgtacagacgcgtgtacgcatgtaacattatactgaaaaccttgcttgagaaggttttgggacgctcgaaggctttaatttgcaagctggagaccaacatgtgagcaaaaggccagcaaaaggccaggaaccgtaaaaaggccgcgttgctggcgtttttccataggctccgcccccctgacgagcatcacaaaaatcgacgctcaagtcagaggtggcgaaacccgacaggactataaagataccaggcgtttccccctggaagctccctcgtgcgctctcctgttccgaccctgccgcttaccggatacctgtccgcctttctcccttcgggaagcgtggcgctttctcaatgctcacgctgtaggtatctcagttcggtgtaggtcgttcgctccaagctgggctgtgtgcacgaaccccccgttcagcccgaccgctgcgccttatccggtaactatcgtcttgagtccaacccggtaagacacgacttatcgccactggcagcagccactggtaacaggattagcagagcgaggtatgtaggcggtgctacagagttcttgaagtggtggcctaactacggctacactagaaggacagtatttggtatctgcgctctgctgaagccagttaccttcggaaaaagagttggtagctcttgatccggcaaacaaaccaccgctggtagcggtggtttttttgtttgcaagcagcagattacgcgcagaaaaaaaggatctcaagaagatcctttgatcttttctacggggtctgacgctcagtggaacgaaaactcacgttaagggattttggtcatgagatca
SEQIDNO:2
Нуклеотидная последовательность искусственно синтезированного фрагмента ДНК, содержащегонуклеотидную последовательность трех тандемно расположенныхпоследовательностей аптамера к тетрациклину 32sh:
caattgggcctaaaacataccagatcgccacccgcgctttaatctggagaggtgaagaatacgaccacctaggccttaagaaccggtaaaacataccagatcgccacccgcgctttaatctggagaggtgaagaatacgaccacctaccggtcaacaacaacaacaacaacaacaacaactcgaggcctaaaacataccagatcgccacccgcgctttaatctggagaggtgaagaatacgaccacctaggcctcgagaacatttcgaa
SEQIDNO:3
Нуклеотидная последовательность экспрессионной генетической конструкции pPIC9Kintron-tc-alphaRFP
agatctaacatccaaagacgaaaggttgaatgaaacctttttgccatccgacatccacaggtccattctcacacataagtgccaaacgcaacaggaggggatacactagcagcagaccgttgcaaacgcaggacctccactcctcttctcctcaacacccacttttgccatcgaaaaaccagcccagttattgggcttgattggagctcgctcattccaattccttctattaggctactaacaccatgactttattagcctgtctatcctggcccccctggcgaggttcatgtttgtttatttccgaatgcaacaagctccgcattacacccgaacatcactccagatgagggctttctgagtgtggggtcaaatagtttcatgttccccaaatggcccaaaactgacagtttaaacgctgtcttggaacctaatatgacaaaagcgtgatctcatccaagatgaactaagtttggttcgttgaaatgctaacggccagttggtcaaaaagaaacttccaaaagtcgccataccgtttgtcttgtttggtattgattgacgaatgctcaaaaataatctcattaatgcttagcgcagtctctctatcgcttctgaaccccggtgcacctgtgccgaaacgcaaatggggaaacacccgctttttggatgattatgcattgtctccacattgtatgcttccaagattctggtgggaatactgctgatagcctaacgttcatgatcaaaatttaactgttctaacccctacttgacagcaatatataaacagaaggaagctgccctgtcttaaacctttttttttatcatcattattagcttactttcataattgcgactggttccaattgacaagcttttgattttaacgacttttaacgacaacttgagaagatcaaaaaacaactaattattcgaaggatccaaacgatgagatttccttcaatttttactgctgttttattcgcagcatcctccgcattagctgctccagtcaacactacaacagaagatgaaacggcacaaattccggctgaagctgtcatcggttactcagatttagaaggggatttcgatgttgctgttttgccattttccaacagcacaaataacgggttattgtttataaatactactattgccagcattgctgctaaagaagaaggggtatctctcgagaaaagacaggtgcagctgcagggcgaaacgatggtgagtaaaggtgaagagttgattaaagagaacatgcatatgaagttatacatggagggaactgtcaataatcaccactttaagtgtacatcagagggtgagggaaagccatacgagggaacccaaactatgagaatcaaagtagtggagggaggtcctcttccatttgcttttgatatactagcaacaagtttcatgtatggttccaggaccttcattaaccatactcagggaatccctgacttctttaaacagtcttttcctgaaggttttacatgggagagggttaccacttacgaggacggtggagtcttgacagcaacccaggacacttcattacaagatggatgcctaatatacaatgtgaaaattaggggtgtgaatttccctagtaacggaccagttatgcagaagaaaacactaggttgggaagctaatactgaaatgttgtaccctgccgacggaggtttagaaggtagatccgacatggcccttaagctagtcggaggtggacacttgatctgtaactttaaaaccacatataggtctaagaagccagcaaagaatctaaaaatgcctggtgtttactatgtggaccatagactagaaaggataaaagaagcagacaaagaaacttacgtggagcaacatgaggtcgccgtcgctaggtattgcgacttaccttccaagctaggtcacaaattgaacactagtgaattccctagggcggccgcatgagtaagtttggcctaaaacataccagatcgccacccgcgctttaatctggagaggtgaagaatacgaccacctaggcctttactaacaagtttctttaggcggccgcgaattaattcgccttagacatgactgttcctcagttcaagttgggcacttacgagaagaccggtcttgctagattctaatcaagaggatgtcagaatgccatttgcctgagagatgcaggcttcatttttgatacttttttatttgtaacctatatagtataggattttttttgtcattttgtttcttctcgtacgagcttgctcctgatcagcctatctcgcagctgatgaatatcttgtggtaggggtttgggaaaatcattcgagtttgatgtttttcttggtatttcccactcctcttcagagtacagaagattaagtgagaagttcgtttgtgcaagcttatcgataagctttaatgcggtagtttatcacagttaaattgctaacgcagtcaggcaccgtgtatgaaatctaacaatgcgctcatcgtcatcctcggcaccgtcaccctggatgctgtaggcataggcttggttatgccggtactgccgggcctcttgcgggatatcgtccattccgacagcatcgccagtcactatggcgtgctgctagcgctatatgcgttgatgcaatttctatgcgcacccgttctcggagcactgtccgaccgctttggccgccgcccagtcctgctcgcttcgctacttggagccactatcgactacgcgatcatggcgaccacacccgtcctgtggatctatcgaatctaaatgtaagttaaaatctctaaataattaaataagtcccagtttctccatacgaaccttaacagcattgcggtgagcatctagaccttcaacagcagccagatccatcactgcttggccaatatgtttcagtccctcaggagttacgtcttgtgaagtgatgaacttctggaaggttgcagtgttaactccgctgtattgacgggcatatccgtacgttggcaaagtgtggttggtaccggaggagtaatctccacaactctctggagagtaggcaccaacaaacacagatccagcgtgttgtacttgatcaacataagaagaagcattctcgatttgcaggatcaagtgttcaggagcgtactgattggacatttccaaagcctgctcgtaggttgcaaccgatagggttgtagagtgtgcaatacacttgcgtacaatttcaacccttggcaactgcacagcttggttgtgaacagcatcttcaattctggcaagctccttgtctgtcatatcgacagccaacagaatcacctgggaatcaataccatgttcagcttgagacagaaggtctgaggcaacgaaatctggatcagcgtatttatcagcaataactagaacttcagaaggcccagcaggcatgtcaatactacacagggctgatgtgtcattttgaaccatcatcttggcagcagtaacgaactggtttcctggaccaaatattttgtcacacttaggaacagtttctgttccgtaagccatagcagctactgcctgggcgcctcctgctagcacgatacacttagcaccaaccttgtgggcaacgtagatgacttctggggtaagggtaccatccttcttaggtggagatgcaaaaacaatttctttgcaaccagcaactttggcaggaacacccagcatcagggaagtggaaggcagaattgcggttccaccaggaatatagaggccaactttctcaataggtcttgcaaaacgagagcagactacaccagggcaagtctcaacttgcaacgtctccgttagttgagcttcatggaatttcctgacgttatctatagagagatcaatggctctcttaacgttatctggcaattgcataagttcctctgggaaaggagcttctaacacaggtgtcttcaaagcgactccatcaaacttggcagttagttctaaaagggctttgtcaccattttgacgaacattgtcgacaattggtttgactaattccataatctgttccgttttctggataggacgacgaagggcatcttcaatttcttgtgaggaggccttagaaacgtcaattttgcacaattcaatacgaccttcagaagggacttctttaggtttggattcttctttaggttgttccttggtgtatcctggcttggcatctcctttccttctagtgacctttagggacttcatatccaggtttctctccacctcgtccaacgtcacaccgtacttggcacatctaactaatgcaaaataaaataagtcagcacattcccaggctatatcttccttggatttagcttctgcaagttcatcagcttcctccctaattttagcgttcaacaaaacttcgtcgtcaaataaccgtttggtataagaaccttctggagcattgctcttacgatcccacaaggtggcttccatggctctaagaccctttgattggccaaaacaggaagtgcgttccaagtgacagaaaccaacacctgtttgttcaaccacaaatttcaagcagtctccatcacaatccaattcgatacccagcaacttttgagttgctccagatgtagcacctttataccacaaaccgtgacgacgagattggtagactccagtttgtgtccttatagcctccggaatagactttttggacgagtacaccaggcccaacgagtaattagaagagtcagccaccaaagtagtgaatagaccatcggggcggtcagtagtcaaagacgccaacaaaatttcactgacagggaactttttgacatcttcagaaagttcgtattcagtagtcaattgccgagcatcaataatggggattataccagaagcaacagtggaagtcacatctaccaactttgcggtctcagaaaaagcataaacagttctactaccgccattagtgaaacttttcaaatcgcccagtggagaagaaaaaggcacagcgatactagcattagcgggcaaggatgcaactttatcaaccagggtcctatagataaccctagcgcctgggatcatcctttggacaactctttctgccaaatctaggtccaaaatcacttcattgataccattattgtacaacttgagcaagttgtcgatcagctcctcaaattggtcctctgtaacggatgactcaacttgcacattaacttgaagctcagtcgattgagtgaacttgatcaggttgtgcagctggtcagcagcatagggaaacacggcttttcctaccaaactcaaggaattatcaaactctgcaacacttgcgtatgcaggtagcaagggaaatgtcatacttgaagtcggacagtgagtgtagtcttgagaaattctgaagccgtatttttattatcagtgagtcagtcatcaggagatcctctacgccggacgcatcgtggccgacctgcagggggggggggggcgctgaggtctgcctcgtgaagaaggtgttgctgactcataccaggcctgaatcgccccatcatccagccagaaagtgagggagccacggttgatgagagctttgttgtaggtggaccagttggtgattttgaacttttgctttgccacggaacggtctgcgttgtcgggaagatgcgtgatctgatccttcaactcagcaaaagttcgatttattcaacaaagccgccgtcccgtcaagtcagcgtaatgctctgccagtgttacaaccaattaaccaattctgattagaaaaactcatcgagcatcaaatgaaactgcaatttattcatatcaggattatcaataccatatttttgaaaaagccgtttctgtaatgaaggagaaaactcaccgaggcagttccataggatggcaagatcctggtatcggtctgcgattccgactcgtccaacatcaatacaacctattaatttcccctcgtcaaaaataaggttatcaagtgagaaatcaccatgagtgacgactgaatccggtgagaatggcaaaagcttatgcatttctttccagacttgttcaacaggccagccattacgctcgtcatcaaaatcactcgcatcaaccaaaccgttattcattcgtgattgcgcctgagcgagacgaaatacgcgatcgctgttaaaaggacaattacaaacaggaatcgaatgcaaccggcgcaggaacactgccagcgcatcaacaatattttcacctgaatcaggatattcttctaatacctggaatgctgttttcccggggatcgcagtggtgagtaaccatgcatcatcaggagtacggataaaatgcttgatggtcggaagaggcataaattccgtcagccagtttagtctgaccatctcatctgtaacatcattggcaacgctacctttgccatgtttcagaaacaactctggcgcatcgggcttcccatacaatcgatagattgtcgcacctgattgcccgacattatcgcgagcccatttatacccatataaatcagcatccatgttggaatttaatcgcggcctcgagcaagacgtttcccgttgaatatggctcataacaccccttgtattactgtttatgtaagcagacagttttattgttcatgatgatatatttttatcttgtgcaatgtaacatcagagattttgagacacaacgtggctttcccccccccccctgcaggtcggcatcaccggcgccacaggtgcggttgctggcgcctatatcgccgacatcaccgatggggaagatcgggctcgccacttcgggctcatgagcgcttgtttcggcgtgggtatggtggcaggccccgtggccgggggactgttgggcgccatctccttgcatgcaccattccttgcggcggcggtgctcaacggcctcaacctactactgggctgcttcctaatgcaggagtcgcataagggagagcgtcgagtatctatgattggaagtatgggaatggtgatacccgcattcttcagtgtcttgaggtctcctatcagattatgcccaactaaagcaaccggaggaggagatttcatggtaaatttctctgacttttggtcatcagtagactcgaactgtgagactatctcggttatgacagcagaaatgtccttcttggagacagtaaatgaagtcccaccaataaagaaatccttgttatcaggaacaaacttcttgtttcgaactttttcggtgccttgaactataaaatgtagagtggatatgtcgggtaggaatggagcgggcaaatgcttaccttctggaccttcaagaggtatgtagggtttgtagatactgatgccaacttcagtgacaacgttgctatttcgttcaaaccattccgaatccagagaaatcaaagttgtttgtctactattgatccaagccagtgcggtcttgaaactgacaatagtgtgctcgtgttttgaggtcatctttgtatgaataaatctagtctttgatctaaataatcttgacgagccaaggcgataaatacccaaatctaaaactcttttaaaacgttaaaaggacaagtatgtctgcctgtattaaaccccaaatcagctcgtagtctgatcctcatcaacttgaggggcactatcttgttttagagaaatttgcggagatgcgatatcgagaaaaaggtacgctgattttaaacgtgaaatttatctcaagatctctgcctcgcgcgtttcggtgatgacggtgaaaacctctgacacatgcagctcccggagacggtcacagcttgtctgtaagcggatgccgggagcagacaagcccgtcagggcgcgtcagcgggtgttggcgggtgtcggggcgcagccatgacccagtcacgtagcgatagcggagtgtatactggcttaactatgcggcatcagagcagattgtactgagagtgcaccatatgcggtgtgaaataccgcacagatgcgtaaggagaaaataccgcatcaggcgctcttccgcttcctcgctcactgactcgctgcgctcggtcgttcggctgcggcgagcggtatcagctcactcaaaggcggtaatacggttatccacagaatcaggggataacgcaggaaagaacatgtgagcaaaaggccagcaaaaggccaggaaccgtaaaaaggccgcgttgctggcgtttttccataggctccgcccccctgacgagcatcacaaaaatcgacgctcaagtcagaggtggcgaaacccgacaggactataaagataccaggcgtttccccctggaagctccctcgtgcgctctcctgttccgaccctgccgcttaccggatacctgtccgcctttctcccttcgggaagcgtggcgctttctcaatgctcacgctgtaggtatctcagttcggtgtaggtcgttcgctccaagctgggctgtgtgcacgaaccccccgttcagcccgaccgctgcgccttatccggtaactatcgtcttgagtccaacccggtaagacacgacttatcgccactggcagcagccactggtaacaggattagcagagcgaggtatgtaggcggtgctacagagttcttgaagtggtggcctaactacggctacactagaaggacagtatttggtatctgcgctctgctgaagccagttaccttcggaaaaagagttggtagctcttgatccggcaaacaaaccaccgctggtagcggtggtttttttgtttgcaagcagcagattacgcgcagaaaaaaaggatctcaagaagatcctttgatcttttctacggggtctgacgctcagtggaacgaaaactcacgttaagggattttggtcatgagattatcaaaaaggatcttcacctagatccttttaaattaaaaatgaagttttaaatcaatctaaagtatatatgagtaaacttggtctgacagttaccaatgcttaatcagtgaggcacctatctcagcgatctgtctatttcgttcatccatagttgcctgactccccgtcgtgtagataactacgatacgggagggcttaccatctggccccagtgctgcaatgataccgcgagacccacgctcaccggctccagatttatcagcaataaaccagccagccggaagggccgagcgcagaagtggtcctgcaactttatccgcctccatccagtctattaattgttgccgggaagctagagtaagtagttcgccagttaatagtttgcgcaacgttgttgccattgctgcaggcatcgtggtgtcacgctcgtcgtttggtatggcttcattcagctccggttcccaacgatcaaggcgagttacatgatcccccatgttgtgcaaaaaagcggttagctccttcggtcctccgatcgttgtcagaagtaagttggccgcagtgttatcactcatggttatggcagcactgcataattctcttactgtcatgccatccgtaagatgcttttctgtgactggtgagtactcaaccaagtcattctgagaatagtgtatgcggcgaccgagttgctcttgcccggcgtcaacacgggataataccgcgccacatagcagaactttaaaagtgctcatcattggaaaacgttcttcggggcgaaaactctcaaggatcttaccgctgttgagatccagttcgatgtaacccactcgtgcacccaactgatcttcagcatcttttactttcaccagcgtttctgggtgagcaaaaacaggaaggcaaaatgccgcaaaaaagggaataagggcgacacggaaatgttgaatactcatactcttcctttttcaatattattgaagcatttatcagggttattgtctcatgagcggatacatatttgaatgtatttagaaaaataaacaaataggggttccgcgcacatttccccgaaaagtgccacctgacgtctaagaaaccattattatcatgacattaacctataaaaataggcgtatcacgaggccctttcgtcttcaagaattaattctcatgtttgacagcttatcatcgataagctgactcatgttggtattgtgaaatagacgcagatcgggaacactgaaaaataacagttattattcg
SEQIDNO:4
Нуклеотидная последовательность экспрессионной генетической конструкции pPIC9Kintron-tc-alpha_shortRFP
agatctaacatccaaagacgaaaggttgaatgaaacctttttgccatccgacatccacaggtccattctcacacataagtgccaaacgcaacaggaggggatacactagcagcagaccgttgcaaacgcaggacctccactcctcttctcctcaacacccacttttgccatcgaaaaaccagcccagttattgggcttgattggagctcgctcattccaattccttctattaggctactaacaccatgactttattagcctgtctatcctggcccccctggcgaggttcatgtttgtttatttccgaatgcaacaagctccgcattacacccgaacatcactccagatgagggctttctgagtgtggggtcaaatagtttcatgttccccaaatggcccaaaactgacagtttaaacgctgtcttggaacctaatatgacaaaagcgtgatctcatccaagatgaactaagtttggttcgttgaaatgctaacggccagttggtcaaaaagaaacttccaaaagtcgccataccgtttgtcttgtttggtattgattgacgaatgctcaaaaataatctcattaatgcttagcgcagtctctctatcgcttctgaaccccggtgcacctgtgccgaaacgcaaatggggaaacacccgctttttggatgattatgcattgtctccacattgtatgcttccaagattctggtgggaatactgctgatagcctaacgttcatgatcaaaatttaactgttctaacccctacttgacagcaatatataaacagaaggaagctgccctgtcttaaacctttttttttatcatcattattagcttactttcataattgcgactggttccaattgacaagcttttgattttaacgacttttaacgacaacttgagaagatcaaaaaacaactaattattcgaaggatccaaacgatgagatttccttcaatttttactgctgttttattcgcagcatccagcgcgctagctcaggtgcagctgcagggcgaaacgatggtgagtaaaggtgaagagttgattaaagagaacatgcatatgaagttatacatggagggaactgtcaataatcaccactttaagtgtacatcagagggtgagggaaagccatacgagggaacccaaactatgagaatcaaagtagtggagggaggtcctcttccatttgcttttgatatactagcaacaagtttcatgtatggttccaggaccttcattaaccatactcagggaatccctgacttctttaaacagtcttttcctgaaggttttacatgggagagggttaccacttacgaggacggtggagtcttgacagcaacccaggacacttcattacaagatggatgcctaatatacaatgtgaaaattaggggtgtgaatttccctagtaacggaccagttatgcagaagaaaacactaggttgggaagctaatactgaaatgttgtaccctgccgacggaggtttagaaggtagatccgacatggcccttaagctagtcggaggtggacacttgatctgtaactttaaaaccacatataggtctaagaagccagcaaagaatctaaaaatgcctggtgtttactatgtggaccatagactagaaaggataaaagaagcagacaaagaaacttacgtggagcaacatgaggtcgccgtcgctaggtattgcgacttaccttccaagctaggtcacaaattgaacactagtgaattccctagggcggccgcatgagtaagtttggcctaaaacataccagatcgccacccgcgctttaatctggagaggtgaagaatacgaccacctaggcctttactaacaagtttctttaggcggccgcgaattaattcgccttagacatgactgttcctcagttcaagttgggcacttacgagaagaccggtcttgctagattctaatcaagaggatgtcagaatgccatttgcctgagagatgcaggcttcatttttgatacttttttatttgtaacctatatagtataggattttttttgtcattttgtttcttctcgtacgagcttgctcctgatcagcctatctcgcagctgatgaatatcttgtggtaggggtttgggaaaatcattcgagtttgatgtttttcttggtatttcccactcctcttcagagtacagaagattaagtgagaagttcgtttgtgcaagcttatcgataagctttaatgcggtagtttatcacagttaaattgctaacgcagtcaggcaccgtgtatgaaatctaacaatgcgctcatcgtcatcctcggcaccgtcaccctggatgctgtaggcataggcttggttatgccggtactgccgggcctcttgcgggatatcgtccattccgacagcatcgccagtcactatggcgtgctgctagcgctatatgcgttgatgcaatttctatgcgcacccgttctcggagcactgtccgaccgctttggccgccgcccagtcctgctcgcttcgctacttggagccactatcgactacgcgatcatggcgaccacacccgtcctgtggatctatcgaatctaaatgtaagttaaaatctctaaataattaaataagtcccagtttctccatacgaaccttaacagcattgcggtgagcatctagaccttcaacagcagccagatccatcactgcttggccaatatgtttcagtccctcaggagttacgtcttgtgaagtgatgaacttctggaaggttgcagtgttaactccgctgtattgacgggcatatccgtacgttggcaaagtgtggttggtaccggaggagtaatctccacaactctctggagagtaggcaccaacaaacacagatccagcgtgttgtacttgatcaacataagaagaagcattctcgatttgcaggatcaagtgttcaggagcgtactgattggacatttccaaagcctgctcgtaggttgcaaccgatagggttgtagagtgtgcaatacacttgcgtacaatttcaacccttggcaactgcacagcttggttgtgaacagcatcttcaattctggcaagctccttgtctgtcatatcgacagccaacagaatcacctgggaatcaataccatgttcagcttgagacagaaggtctgaggcaacgaaatctggatcagcgtatttatcagcaataactagaacttcagaaggcccagcaggcatgtcaatactacacagggctgatgtgtcattttgaaccatcatcttggcagcagtaacgaactggtttcctggaccaaatattttgtcacacttaggaacagtttctgttccgtaagccatagcagctactgcctgggcgcctcctgctagcacgatacacttagcaccaaccttgtgggcaacgtagatgacttctggggtaagggtaccatccttcttaggtggagatgcaaaaacaatttctttgcaaccagcaactttggcaggaacacccagcatcagggaagtggaaggcagaattgcggttccaccaggaatatagaggccaactttctcaataggtcttgcaaaacgagagcagactacaccagggcaagtctcaacttgcaacgtctccgttagttgagcttcatggaatttcctgacgttatctatagagagatcaatggctctcttaacgttatctggcaattgcataagttcctctgggaaaggagcttctaacacaggtgtcttcaaagcgactccatcaaacttggcagttagttctaaaagggctttgtcaccattttgacgaacattgtcgacaattggtttgactaattccataatctgttccgttttctggataggacgacgaagggcatcttcaatttcttgtgaggaggccttagaaacgtcaattttgcacaattcaatacgaccttcagaagggacttctttaggtttggattcttctttaggttgttccttggtgtatcctggcttggcatctcctttccttctagtgacctttagggacttcatatccaggtttctctccacctcgtccaacgtcacaccgtacttggcacatctaactaatgcaaaataaaataagtcagcacattcccaggctatatcttccttggatttagcttctgcaagttcatcagcttcctccctaattttagcgttcaacaaaacttcgtcgtcaaataaccgtttggtataagaaccttctggagcattgctcttacgatcccacaaggtggcttccatggctctaagaccctttgattggccaaaacaggaagtgcgttccaagtgacagaaaccaacacctgtttgttcaaccacaaatttcaagcagtctccatcacaatccaattcgatacccagcaacttttgagttgctccagatgtagcacctttataccacaaaccgtgacgacgagattggtagactccagtttgtgtccttatagcctccggaatagactttttggacgagtacaccaggcccaacgagtaattagaagagtcagccaccaaagtagtgaatagaccatcggggcggtcagtagtcaaagacgccaacaaaatttcactgacagggaactttttgacatcttcagaaagttcgtattcagtagtcaattgccgagcatcaataatggggattataccagaagcaacagtggaagtcacatctaccaactttgcggtctcagaaaaagcataaacagttctactaccgccattagtgaaacttttcaaatcgcccagtggagaagaaaaaggcacagcgatactagcattagcgggcaaggatgcaactttatcaaccagggtcctatagataaccctagcgcctgggatcatcctttggacaactctttctgccaaatctaggtccaaaatcacttcattgataccattattgtacaacttgagcaagttgtcgatcagctcctcaaattggtcctctgtaacggatgactcaacttgcacattaacttgaagctcagtcgattgagtgaacttgatcaggttgtgcagctggtcagcagcatagggaaacacggcttttcctaccaaactcaaggaattatcaaactctgcaacacttgcgtatgcaggtagcaagggaaatgtcatacttgaagtcggacagtgagtgtagtcttgagaaattctgaagccgtatttttattatcagtgagtcagtcatcaggagatcctctacgccggacgcatcgtggccgacctgcagggggggggggggcgctgaggtctgcctcgtgaagaaggtgttgctgactcataccaggcctgaatcgccccatcatccagccagaaagtgagggagccacggttgatgagagctttgttgtaggtggaccagttggtgattttgaacttttgctttgccacggaacggtctgcgttgtcgggaagatgcgtgatctgatccttcaactcagcaaaagttcgatttattcaacaaagccgccgtcccgtcaagtcagcgtaatgctctgccagtgttacaaccaattaaccaattctgattagaaaaactcatcgagcatcaaatgaaactgcaatttattcatatcaggattatcaataccatatttttgaaaaagccgtttctgtaatgaaggagaaaactcaccgaggcagttccataggatggcaagatcctggtatcggtctgcgattccgactcgtccaacatcaatacaacctattaatttcccctcgtcaaaaataaggttatcaagtgagaaatcaccatgagtgacgactgaatccggtgagaatggcaaaagcttatgcatttctttccagacttgttcaacaggccagccattacgctcgtcatcaaaatcactcgcatcaaccaaaccgttattcattcgtgattgcgcctgagcgagacgaaatacgcgatcgctgttaaaaggacaattacaaacaggaatcgaatgcaaccggcgcaggaacactgccagcgcatcaacaatattttcacctgaatcaggatattcttctaatacctggaatgctgttttcccggggatcgcagtggtgagtaaccatgcatcatcaggagtacggataaaatgcttgatggtcggaagaggcataaattccgtcagccagtttagtctgaccatctcatctgtaacatcattggcaacgctacctttgccatgtttcagaaacaactctggcgcatcgggcttcccatacaatcgatagattgtcgcacctgattgcccgacattatcgcgagcccatttatacccatataaatcagcatccatgttggaatttaatcgcggcctcgagcaagacgtttcccgttgaatatggctcataacaccccttgtattactgtttatgtaagcagacagttttattgttcatgatgatatatttttatcttgtgcaatgtaacatcagagattttgagacacaacgtggctttcccccccccccctgcaggtcggcatcaccggcgccacaggtgcggttgctggcgcctatatcgccgacatcaccgatggggaagatcgggctcgccacttcgggctcatgagcgcttgtttcggcgtgggtatggtggcaggccccgtggccgggggactgttgggcgccatctccttgcatgcaccattccttgcggcggcggtgctcaacggcctcaacctactactgggctgcttcctaatgcaggagtcgcataagggagagcgtcgagtatctatgattggaagtatgggaatggtgatacccgcattcttcagtgtcttgaggtctcctatcagattatgcccaactaaagcaaccggaggaggagatttcatggtaaatttctctgacttttggtcatcagtagactcgaactgtgagactatctcggttatgacagcagaaatgtccttcttggagacagtaaatgaagtcccaccaataaagaaatccttgttatcaggaacaaacttcttgtttcgaactttttcggtgccttgaactataaaatgtagagtggatatgtcgggtaggaatggagcgggcaaatgcttaccttctggaccttcaagaggtatgtagggtttgtagatactgatgccaacttcagtgacaacgttgctatttcgttcaaaccattccgaatccagagaaatcaaagttgtttgtctactattgatccaagccagtgcggtcttgaaactgacaatagtgtgctcgtgttttgaggtcatctttgtatgaataaatctagtctttgatctaaataatcttgacgagccaaggcgataaatacccaaatctaaaactcttttaaaacgttaaaaggacaagtatgtctgcctgtattaaaccccaaatcagctcgtagtctgatcctcatcaacttgaggggcactatcttgttttagagaaatttgcggagatgcgatatcgagaaaaaggtacgctgattttaaacgtgaaatttatctcaagatctctgcctcgcgcgtttcggtgatgacggtgaaaacctctgacacatgcagctcccggagacggtcacagcttgtctgtaagcggatgccgggagcagacaagcccgtcagggcgcgtcagcgggtgttggcgggtgtcggggcgcagccatgacccagtcacgtagcgatagcggagtgtatactggcttaactatgcggcatcagagcagattgtactgagagtgcaccatatgcggtgtgaaataccgcacagatgcgtaaggagaaaataccgcatcaggcgctcttccgcttcctcgctcactgactcgctgcgctcggtcgttcggctgcggcgagcggtatcagctcactcaaaggcggtaatacggttatccacagaatcaggggataacgcaggaaagaacatgtgagcaaaaggccagcaaaaggccaggaaccgtaaaaaggccgcgttgctggcgtttttccataggctccgcccccctgacgagcatcacaaaaatcgacgctcaagtcagaggtggcgaaacccgacaggactataaagataccaggcgtttccccctggaagctccctcgtgcgctctcctgttccgaccctgccgcttaccggatacctgtccgcctttctcccttcgggaagcgtggcgctttctcaatgctcacgctgtaggtatctcagttcggtgtaggtcgttcgctccaagctgggctgtgtgcacgaaccccccgttcagcccgaccgctgcgccttatccggtaactatcgtcttgagtccaacccggtaagacacgacttatcgccactggcagcagccactggtaacaggattagcagagcgaggtatgtaggcggtgctacagagttcttgaagtggtggcctaactacggctacactagaaggacagtatttggtatctgcgctctgctgaagccagttaccttcggaaaaagagttggtagctcttgatccggcaaacaaaccaccgctggtagcggtggtttttttgtttgcaagcagcagattacgcgcagaaaaaaaggatctcaagaagatcctttgatcttttctacggggtctgacgctcagtggaacgaaaactcacgttaagggattttggtcatgagattatcaaaaaggatcttcacctagatccttttaaattaaaaatgaagttttaaatcaatctaaagtatatatgagtaaacttggtctgacagttaccaatgcttaatcagtgaggcacctatctcagcgatctgtctatttcgttcatccatagttgcctgactccccgtcgtgtagataactacgatacgggagggcttaccatctggccccagtgctgcaatgataccgcgagacccacgctcaccggctccagatttatcagcaataaaccagccagccggaagggccgagcgcagaagtggtcctgcaactttatccgcctccatccagtctattaattgttgccgggaagctagagtaagtagttcgccagttaatagtttgcgcaacgttgttgccattgctgcaggcatcgtggtgtcacgctcgtcgtttggtatggcttcattcagctccggttcccaacgatcaaggcgagttacatgatcccccatgttgtgcaaaaaagcggttagctccttcggtcctccgatcgttgtcagaagtaagttggccgcagtgttatcactcatggttatggcagcactgcataattctcttactgtcatgccatccgtaagatgcttttctgtgactggtgagtactcaaccaagtcattctgagaatagtgtatgcggcgaccgagttgctcttgcccggcgtcaacacgggataataccgcgccacatagcagaactttaaaagtgctcatcattggaaaacgttcttcggggcgaaaactctcaaggatcttaccgctgttgagatccagttcgatgtaacccactcgtgcacccaactgatcttcagcatcttttactttcaccagcgtttctgggtgagcaaaaacaggaaggcaaaatgccgcaaaaaagggaataagggcgacacggaaatgttgaatactcatactcttcctttttcaatattattgaagcatttatcagggttattgtctcatgagcggatacatatttgaatgtatttagaaaaataaacaaataggggttccgcgcacatttccccgaaaagtgccacctgacgtctaagaaaccattattatcatgacattaacctataaaaataggcgtatcacgaggccctttcgtcttcaagaattaattctcatgtttgacagcttatcatcgataagctgactcatgttggtattgtgaaatagacgcagatcgggaacactgaaaaataacagttattattcg
SEQIDNO:5
Нуклеотидная последовательность экспрессионной генетической конструкции pPIC9Kintron-tc-hsaRFP
agatctaacatccaaagacgaaaggttgaatgaaacctttttgccatccgacatccacaggtccattctcacacataagtgccaaacgcaacaggaggggatacactagcagcagaccgttgcaaacgcaggacctccactcctcttctcctcaacacccacttttgccatcgaaaaaccagcccagttattgggcttgattggagctcgctcattccaattccttctattaggctactaacaccatgactttattagcctgtctatcctggcccccctggcgaggttcatgtttgtttatttccgaatgcaacaagctccgcattacacccgaacatcactccagatgagggctttctgagtgtggggtcaaatagtttcatgttccccaaatggcccaaaactgacagtttaaacgctgtcttggaacctaatatgacaaaagcgtgatctcatccaagatgaactaagtttggttcgttgaaatgctaacggccagttggtcaaaaagaaacttccaaaagtcgccataccgtttgtcttgtttggtattgattgacgaatgctcaaaaataatctcattaatgcttagcgcagtctctctatcgcttctgaaccccggtgcacctgtgccgaaacgcaaatggggaaacacccgctttttggatgattatgcattgtctccacattgtatgcttccaagattctggtgggaatactgctgatagcctaacgttcatgatcaaaatttaactgttctaacccctacttgacagcaatatataaacagaaggaagctgccctgtcttaaacctttttttttatcatcattattagcttactttcataattgcgactggttccaattgacaagcttttgattttaacgacttttaacgacaacttgagaagatcaaaaaacaactaattattcgaaggatccaaacgatgaagtgggttacctttatctctttgttgtttcttttctctagcgcttactcgcaggtgcagctgcagggcgaaacgatggtgagtaaaggtgaagagttgattaaagagaacatgcatatgaagttatacatggagggaactgtcaataatcaccactttaagtgtacatcagagggtgagggaaagccatacgagggaacccaaactatgagaatcaaagtagtggagggaggtcctcttccatttgcttttgatatactagcaacaagtttcatgtatggttccaggaccttcattaaccatactcagggaatccctgacttctttaaacagtcttttcctgaaggttttacatgggagagggttaccacttacgaggacggtggagtcttgacagcaacccaggacacttcattacaagatggatgcctaatatacaatgtgaaaattaggggtgtgaatttccctagtaacggaccagttatgcagaagaaaacactaggttgggaagctaatactgaaatgttgtaccctgccgacggaggtttagaaggtagatccgacatggcccttaagctagtcggaggtggacacttgatctgtaactttaaaaccacatataggtctaagaagccagcaaagaatctaaaaatgcctggtgtttactatgtggaccatagactagaaaggataaaagaagcagacaaagaaacttacgtggagcaacatgaggtcgccgtcgctaggtattgcgacttaccttccaagctaggtcacaaattgaacactagtgaattccctagggcggccgcatgagtaagtttggcctaaaacataccagatcgccacccgcgctttaatctggagaggtgaagaatacgaccacctaggcctttactaacaagtttctttaggcggccgcgaattaattcgccttagacatgactgttcctcagttcaagttgggcacttacgagaagaccggtcttgctagattctaatcaagaggatgtcagaatgccatttgcctgagagatgcaggcttcatttttgatacttttttatttgtaacctatatagtataggattttttttgtcattttgtttcttctcgtacgagcttgctcctgatcagcctatctcgcagctgatgaatatcttgtggtaggggtttgggaaaatcattcgagtttgatgtttttcttggtatttcccactcctcttcagagtacagaagattaagtgagaagttcgtttgtgcaagcttatcgataagctttaatgcggtagtttatcacagttaaattgctaacgcagtcaggcaccgtgtatgaaatctaacaatgcgctcatcgtcatcctcggcaccgtcaccctggatgctgtaggcataggcttggttatgccggtactgccgggcctcttgcgggatatcgtccattccgacagcatcgccagtcactatggcgtgctgctagcgctatatgcgttgatgcaatttctatgcgcacccgttctcggagcactgtccgaccgctttggccgccgcccagtcctgctcgcttcgctacttggagccactatcgactacgcgatcatggcgaccacacccgtcctgtggatctatcgaatctaaatgtaagttaaaatctctaaataattaaataagtcccagtttctccatacgaaccttaacagcattgcggtgagcatctagaccttcaacagcagccagatccatcactgcttggccaatatgtttcagtccctcaggagttacgtcttgtgaagtgatgaacttctggaaggttgcagtgttaactccgctgtattgacgggcatatccgtacgttggcaaagtgtggttggtaccggaggagtaatctccacaactctctggagagtaggcaccaacaaacacagatccagcgtgttgtacttgatcaacataagaagaagcattctcgatttgcaggatcaagtgttcaggagcgtactgattggacatttccaaagcctgctcgtaggttgcaaccgatagggttgtagagtgtgcaatacacttgcgtacaatttcaacccttggcaactgcacagcttggttgtgaacagcatcttcaattctggcaagctccttgtctgtcatatcgacagccaacagaatcacctgggaatcaataccatgttcagcttgagacagaaggtctgaggcaacgaaatctggatcagcgtatttatcagcaataactagaacttcagaaggcccagcaggcatgtcaatactacacagggctgatgtgtcattttgaaccatcatcttggcagcagtaacgaactggtttcctggaccaaatattttgtcacacttaggaacagtttctgttccgtaagccatagcagctactgcctgggcgcctcctgctagcacgatacacttagcaccaaccttgtgggcaacgtagatgacttctggggtaagggtaccatccttcttaggtggagatgcaaaaacaatttctttgcaaccagcaactttggcaggaacacccagcatcagggaagtggaaggcagaattgcggttccaccaggaatatagaggccaactttctcaataggtcttgcaaaacgagagcagactacaccagggcaagtctcaacttgcaacgtctccgttagttgagcttcatggaatttcctgacgttatctatagagagatcaatggctctcttaacgttatctggcaattgcataagttcctctgggaaaggagcttctaacacaggtgtcttcaaagcgactccatcaaacttggcagttagttctaaaagggctttgtcaccattttgacgaacattgtcgacaattggtttgactaattccataatctgttccgttttctggataggacgacgaagggcatcttcaatttcttgtgaggaggccttagaaacgtcaattttgcacaattcaatacgaccttcagaagggacttctttaggtttggattcttctttaggttgttccttggtgtatcctggcttggcatctcctttccttctagtgacctttagggacttcatatccaggtttctctccacctcgtccaacgtcacaccgtacttggcacatctaactaatgcaaaataaaataagtcagcacattcccaggctatatcttccttggatttagcttctgcaagttcatcagcttcctccctaattttagcgttcaacaaaacttcgtcgtcaaataaccgtttggtataagaaccttctggagcattgctcttacgatcccacaaggtggcttccatggctctaagaccctttgattggccaaaacaggaagtgcgttccaagtgacagaaaccaacacctgtttgttcaaccacaaatttcaagcagtctccatcacaatccaattcgatacccagcaacttttgagttgctccagatgtagcacctttataccacaaaccgtgacgacgagattggtagactccagtttgtgtccttatagcctccggaatagactttttggacgagtacaccaggcccaacgagtaattagaagagtcagccaccaaagtagtgaatagaccatcggggcggtcagtagtcaaagacgccaacaaaatttcactgacagggaactttttgacatcttcagaaagttcgtattcagtagtcaattgccgagcatcaataatggggattataccagaagcaacagtggaagtcacatctaccaactttgcggtctcagaaaaagcataaacagttctactaccgccattagtgaaacttttcaaatcgcccagtggagaagaaaaaggcacagcgatactagcattagcgggcaaggatgcaactttatcaaccagggtcctatagataaccctagcgcctgggatcatcctttggacaactctttctgccaaatctaggtccaaaatcacttcattgataccattattgtacaacttgagcaagttgtcgatcagctcctcaaattggtcctctgtaacggatgactcaacttgcacattaacttgaagctcagtcgattgagtgaacttgatcaggttgtgcagctggtcagcagcatagggaaacacggcttttcctaccaaactcaaggaattatcaaactctgcaacacttgcgtatgcaggtagcaagggaaatgtcatacttgaagtcggacagtgagtgtagtcttgagaaattctgaagccgtatttttattatcagtgagtcagtcatcaggagatcctctacgccggacgcatcgtggccgacctgcagggggggggggggcgctgaggtctgcctcgtgaagaaggtgttgctgactcataccaggcctgaatcgccccatcatccagccagaaagtgagggagccacggttgatgagagctttgttgtaggtggaccagttggtgattttgaacttttgctttgccacggaacggtctgcgttgtcgggaagatgcgtgatctgatccttcaactcagcaaaagttcgatttattcaacaaagccgccgtcccgtcaagtcagcgtaatgctctgccagtgttacaaccaattaaccaattctgattagaaaaactcatcgagcatcaaatgaaactgcaatttattcatatcaggattatcaataccatatttttgaaaaagccgtttctgtaatgaaggagaaaactcaccgaggcagttccataggatggcaagatcctggtatcggtctgcgattccgactcgtccaacatcaatacaacctattaatttcccctcgtcaaaaataaggttatcaagtgagaaatcaccatgagtgacgactgaatccggtgagaatggcaaaagcttatgcatttctttccagacttgttcaacaggccagccattacgctcgtcatcaaaatcactcgcatcaaccaaaccgttattcattcgtgattgcgcctgagcgagacgaaatacgcgatcgctgttaaaaggacaattacaaacaggaatcgaatgcaaccggcgcaggaacactgccagcgcatcaacaatattttcacctgaatcaggatattcttctaatacctggaatgctgttttcccggggatcgcagtggtgagtaaccatgcatcatcaggagtacggataaaatgcttgatggtcggaagaggcataaattccgtcagccagtttagtctgaccatctcatctgtaacatcattggcaacgctacctttgccatgtttcagaaacaactctggcgcatcgggcttcccatacaatcgatagattgtcgcacctgattgcccgacattatcgcgagcccatttatacccatataaatcagcatccatgttggaatttaatcgcggcctcgagcaagacgtttcccgttgaatatggctcataacaccccttgtattactgtttatgtaagcagacagttttattgttcatgatgatatatttttatcttgtgcaatgtaacatcagagattttgagacacaacgtggctttcccccccccccctgcaggtcggcatcaccggcgccacaggtgcggttgctggcgcctatatcgccgacatcaccgatggggaagatcgggctcgccacttcgggctcatgagcgcttgtttcggcgtgggtatggtggcaggccccgtggccgggggactgttgggcgccatctccttgcatgcaccattccttgcggcggcggtgctcaacggcctcaacctactactgggctgcttcctaatgcaggagtcgcataagggagagcgtcgagtatctatgattggaagtatgggaatggtgatacccgcattcttcagtgtcttgaggtctcctatcagattatgcccaactaaagcaaccggaggaggagatttcatggtaaatttctctgacttttggtcatcagtagactcgaactgtgagactatctcggttatgacagcagaaatgtccttcttggagacagtaaatgaagtcccaccaataaagaaatccttgttatcaggaacaaacttcttgtttcgaactttttcggtgccttgaactataaaatgtagagtggatatgtcgggtaggaatggagcgggcaaatgcttaccttctggaccttcaagaggtatgtagggtttgtagatactgatgccaacttcagtgacaacgttgctatttcgttcaaaccattccgaatccagagaaatcaaagttgtttgtctactattgatccaagccagtgcggtcttgaaactgacaatagtgtgctcgtgttttgaggtcatctttgtatgaataaatctagtctttgatctaaataatcttgacgagccaaggcgataaatacccaaatctaaaactcttttaaaacgttaaaaggacaagtatgtctgcctgtattaaaccccaaatcagctcgtagtctgatcctcatcaacttgaggggcactatcttgttttagagaaatttgcggagatgcgatatcgagaaaaaggtacgctgattttaaacgtgaaatttatctcaagatctctgcctcgcgcgtttcggtgatgacggtgaaaacctctgacacatgcagctcccggagacggtcacagcttgtctgtaagcggatgccgggagcagacaagcccgtcagggcgcgtcagcgggtgttggcgggtgtcggggcgcagccatgacccagtcacgtagcgatagcggagtgtatactggcttaactatgcggcatcagagcagattgtactgagagtgcaccatatgcggtgtgaaataccgcacagatgcgtaaggagaaaataccgcatcaggcgctcttccgcttcctcgctcactgactcgctgcgctcggtcgttcggctgcggcgagcggtatcagctcactcaaaggcggtaatacggttatccacagaatcaggggataacgcaggaaagaacatgtgagcaaaaggccagcaaaaggccaggaaccgtaaaaaggccgcgttgctggcgtttttccataggctccgcccccctgacgagcatcacaaaaatcgacgctcaagtcagaggtggcgaaacccgacaggactataaagataccaggcgtttccccctggaagctccctcgtgcgctctcctgttccgaccctgccgcttaccggatacctgtccgcctttctcccttcgggaagcgtggcgctttctcaatgctcacgctgtaggtatctcagttcggtgtaggtcgttcgctccaagctgggctgtgtgcacgaaccccccgttcagcccgaccgctgcgccttatccggtaactatcgtcttgagtccaacccggtaagacacgacttatcgccactggcagcagccactggtaacaggattagcagagcgaggtatgtaggcggtgctacagagttcttgaagtggtggcctaactacggctacactagaaggacagtatttggtatctgcgctctgctgaagccagttaccttcggaaaaagagttggtagctcttgatccggcaaacaaaccaccgctggtagcggtggtttttttgtttgcaagcagcagattacgcgcagaaaaaaaggatctcaagaagatcctttgatcttttctacggggtctgacgctcagtggaacgaaaactcacgttaagggattttggtcatgagattatcaaaaaggatcttcacctagatccttttaaattaaaaatgaagttttaaatcaatctaaagtatatatgagtaaacttggtctgacagttaccaatgcttaatcagtgaggcacctatctcagcgatctgtctatttcgttcatccatagttgcctgactccccgtcgtgtagataactacgatacgggagggcttaccatctggccccagtgctgcaatgataccgcgagacccacgctcaccggctccagatttatcagcaataaaccagccagccggaagggccgagcgcagaagtggtcctgcaactttatccgcctccatccagtctattaattgttgccgggaagctagagtaagtagttcgccagttaatagtttgcgcaacgttgttgccattgctgcaggcatcgtggtgtcacgctcgtcgtttggtatggcttcattcagctccggttcccaacgatcaaggcgagttacatgatcccccatgttgtgcaaaaaagcggttagctccttcggtcctccgatcgttgtcagaagtaagttggccgcagtgttatcactcatggttatggcagcactgcataattctcttactgtcatgccatccgtaagatgcttttctgtgactggtgagtactcaaccaagtcattctgagaatagtgtatgcggcgaccgagttgctcttgcccggcgtcaacacgggataataccgcgccacatagcagaactttaaaagtgctcatcattggaaaacgttcttcggggcgaaaactctcaaggatcttaccgctgttgagatccagttcgatgtaacccactcgtgcacccaactgatcttcagcatcttttactttcaccagcgtttctgggtgagcaaaaacaggaaggcaaaatgccgcaaaaaagggaataagggcgacacggaaatgttgaatactcatactcttcctttttcaatattattgaagcatttatcagggttattgtctcatgagcggatacatatttgaatgtatttagaaaaataaacaaataggggttccgcgcacatttccccgaaaagtgccacctgacgtctaagaaaccattattatcatgacattaacctataaaaataggcgtatcacgaggccctttcgtcttcaagaattaattctcatgtttgacagcttatcatcgataagctgactcatgttggtattgtgaaatagacgcagatcgggaacactgaaaaataacagttattattcg
SEQIDNO:6
Фрагмент ДНК, содержащий нуклеотидную последовательность аптамера к тетрациклину 32sh
gtaagtttggcctaaaacataccagatcgccacccgcgctttaatctggagaggtgaagaatacgaccacctaggcctttactaacaagtttctttag
| название | год | авторы | номер документа |
|---|---|---|---|
| Штамм метилотрофных дрожжей Pichia pastoris Yst-HSA-PDI1, продуцирующий рекомбинантный человеческий сывороточный альбумин | 2019 |
|
RU2733423C1 |
| Штамм метилотрофных дрожжей PICHIA PASTORIS Yst-pPIC9pGAPZalpha-short_hTFNG, продуцирующий рекомбинантный человеческий трансферрин, содержащий аминокислотные замены N413D и N611D | 2019 |
|
RU2723084C1 |
| РЕКОМБИНАНТНЫЙ ХИМЕРНЫЙ БЕЛОК SUMO3-apoA-I ДЛЯ ПОЛУЧЕНИЯ ЗРЕЛОГО АПОЛИПОПРОТЕИНА A-I ЧЕЛОВЕКА, ШТАММ ДРОЖЖЕЙ Pichia pastoris - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО ХИМЕРНОГО БЕЛКА SUMO3-apoA-I И СПОСОБ ПОЛУЧЕНИЯ ЗРЕЛОГО АПОЛИПОПРОТЕИНА A-I ЧЕЛОВЕКА | 2015 |
|
RU2605326C1 |
| МОЛЕКУЛА ДНК ДЛЯ ЭКСПРЕССИИ, СТИМУЛИРУЕМОЙ СОЛЯМИ ЖЕЛЧИ ЛИПАЗЫ (BSSL) | 1996 |
|
RU2157847C2 |
| СПОСОБ МИКРОБИОЛОГИЧЕСКОГО СИНТЕЗА ЦЕЛЕВОГО СЕКРЕТИРУЕМОГО БЕЛКА В ДРОЖЖАХ Saccharomyces cerevisiae | 2012 |
|
RU2502805C1 |
| ПРЕПАРАТ ЦИСТЕИНОВОЙ ПРОТЕИНАЗЫ ПШЕНИЦЫ ТРИТИКАИНА-АЛЬФА, ПОЛУЧЕННОЙ В РАСТВОРИМОЙ ФОРМЕ, И СПОСОБ ПОЛУЧЕНИЯ ПРЕПАРАТА | 2017 |
|
RU2676322C1 |
| ПРЕПАРАТ РЕКОМБИНАНТНОГО ЧЕЛОВЕЧЕСКОГО СЫВОРОТОЧНОГО АЛЬБУМИНА И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2006 |
|
RU2337966C2 |
| Способ получения штамма-продуцента фосфолипазы А2 Komagataella phaffii (Pichia pastoris) YIB Δleu2_PLA2S | 2020 |
|
RU2746817C1 |
| Рекомбинантный RBD S-белок коронавируса SARS-CoV-2 и способ его получения | 2023 |
|
RU2833839C1 |
| СРЕДСТВО ДЛЯ ИММУНОПРОФИЛАКТИКИ ТКАНЕВЫХ ГЕЛЬМИНТОЗОВ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ И ЧЕЛОВЕКА, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2023 |
|
RU2830894C1 |




Группа изобретений относится к биотехнологии, а именно к получению дрожжевых штаммов-продуцентов рекомбинантных белков. Предложена рекомбинантная плазмидная ДНК pGAP3tcGFP, представленная в SEQ ID NO 1, для экспрессии маркерного рекомбинантного белка GFP. Также предложен штамм Yst-pGAP3tcGFP метилотрофных дрожжей P.pastoris, полученный путем трансформации клеток метилотрофных дрожжей P.pastoris штамма GS115, для продукции внутри клеток маркерного рекомбинантного белка GFP, содержащий рекомбинантную указанную плазмидную ДНК. Также предложен способ получения целевого рекомбинантного белка с использованием указанного штамма. Группа изобретений обеспечивает высокий уровень продукции целевого белка. 3 н. и 3 з.п. ф-лы, 6 ил., 3 пр.
1. Рекомбинантная плазмидная ДНК pGAP3tcGFP, представленная в SEQ ID NO 1, для экспрессии маркерного рекомбинантного белка GFP (зеленый флуоресцентный белок), имеющая длину 4307 пар оснований и состоящая из следующих генетических элементов:
a) укороченный конститутивный промотор гена глицерид-3-фосфат дегидрогеназы GAP дрожжей Pichia pastoris;
b) три тандемно расположенные нуклеотидные последовательности аптамера к тетрациклину 32sh, представленные в SEQ ID NO 2;
c) нуклеотидная последовательность кДНК гена зеленого флуоресцентного белка;
d) нативный терминатор и сигнал полиаденилирования гена алкоголь оксидазы AOX1 дрожжей Pichia pastoris;
e) точка начала репликации плазмид pUCori;
f) конститутивная промоторная последовательность гена фактора элонгации 1 Saccharomyces cerevisiae TEF1;
g) конститутивная синтетическая промоторная последовательность ЕМ7;
h) последовательность гена блеомицина BleoR Streptoalloteichus hindustanus, продукт которого обеспечивает устойчивость трансформантов к антибиотику зеоцину;
i) 3'-концевая терминаторная последовательность гена цитохрома С1 CYC1 Saccharomyces cerevisiae.
2. Штамм Yst-pGAP3tcGFP метилотрофных дрожжей P.pastoris, полученный путем трансформации клеток метилотрофных дрожжей P.pastoris штамма GS115, для продукции внутри клеток маркерного рекомбинантного белка GFP, содержащий рекомбинантную плазмидную ДНК по п.1.
3. Способ получения любого целевого рекомбинантного белка, включающий в себя стадии:
a) трансформация штамма Yst-pGAP3tcGFP по п.2 рекомбинантной плазмидной ДНК, содержащей следующие обязательные генетические элементы:
- кДНК целевого гена;
- промоторные и терминаторные последовательности для экспрессии целевого белка;
- последовательность сигнального пептида для секреции целевого белка;
- лежащую в пределах транскрибируемой области гена целевого белка искусственно сконструированную нуклеотидную последовательность интрона, представленную в SEQ ID NO 6, с включенной в его состав нуклеотидной последовательностью аптамера к тетрациклину;
- селекционный маркер, для отбора трансформированных клеток, кроме последовательности гена блеомицина BleoR Streptoalloteichus hindustanus, продукт которого обеспечивает устойчивость трансформантов к антибиотику зеоцину;
b) культивирование выросших клеток в культуральной среде, содержащей 55 мкМ тетрациклина;
c) отбор клеток, составляющих 30% от общего числа анализируемых клеток, флуоресценция GFP которых превышает таковую оставшихся 70% клеток методом проточной цитофлуориметрии;
d) анализ уровня экспрессии отобранных клонов и выбор одного, обладающего наиболее высоким уровнем продукции целевого белка.
4. Способ по п.3, отличающийся тем, что рекомбинантная плазмидная ДНК, трансформирующая штамм Yst-pGAP3tcGFP, содержит любые промоторные и терминаторные последовательности для экспрессии целевого белка.
5. Способ по п.3, отличающийся тем, что рекомбинантная плазмидная ДНК, трансформирующая штамм Yst-pGAP3tcGFP, содержит последовательность любого сигнального пептида для секреции целевого белка
6. Способ по п.3, отличающийся тем, что рекомбинантная плазмидная ДНК, трансформирующая штамм Yst-pGAP3tcGFP, содержит любой селекционный маркер для отбора трансформированных клеток, кроме последовательности гена блеомицина BleoR Streptoalloteichus hindustanus.
| KОTTER P | |||
| ET AL | |||
| A fast and efficient translational control system for conditional expression of yeast genes | |||
| Nucleic Acids Res | |||
| Колосоуборка | 1923 |
|
SU2009A1 |
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| СИСТЕМА, ГЕНЕРИРУЮЩАЯ АЭРОЗОЛЬ, С НЕСКОЛЬКИМИ ИНДУКЦИОННЫМИ КАТУШКАМИ | 2018 |
|
RU2764425C2 |
| RAKESTRAW J.A | |||
| ET AL | |||
| A flow cytometric assay for | |||

