Изобретение относится к биотехнологии и может быть использовано при получении культур прогениторных клеток для различных целей.
С учетом возрастания области применения прогениторных клеток перед цитологами достаточно остро стоит проблема получения в короткий временной период требуемого количества мультипотентных мезенхимальных стромальных клеток с низкой гетерогенностью и высокой жизнеспособностью, поскольку именно такие клетки необходимы для решения практических задач регенеративной медицины.
Мезенхимальные стромальные клетки-предшественники (МСК) могут быть выделены из различных тканей. Они представляют собой малочисленную популяцию клеток, характеризующихся большим пролиферативным потенциалом, способных к самоподдержанию с сохранением недифференцированного состояния, а также обладающих возможностью дифференцироваться в различные клеточные типы под действием определенных стимулов. Способность МСК дифференцироваться, по крайней мере, в клетки тканей мезенхимального происхождения лежит в основе их репаративного потенциала.
До настоящего времени костный мозг рассматривался как главный источник стволовых клеток взрослого организма. Костный мозг содержит гемопоэтические стволовые клетки и их более коммитированные потомки, строму, а также так называемые мезенхимальные стромальные клетки или клетки-предшественники взрослого организма (МСК) (Caplan AI. Mesenchymal stem cells. J Orthop Res. 1991 #9 Vol.5, p.641-650; Friedenstein AJ. Precursor cells of mechanocytes. Int. Rev. Cytol. 1976 #47, p.327-359).
В настоящее время доказано существование МСК не только в костном мозге, но и практически во всех тканях организма, например в коже, жировой ткани, эпителии тонкого кишечника и др. (Zuk P.A., Zhu M., Mizuno H., et al. Multiliniage cells from human adipose tissue: implications for cell-based therapies. // Tissue Eng. - 2001. - Vol.7. - P.211-226; Zuk P.A., Zhu M., Mizuno H. et al. Human adipose tissue is a source of multipotent stem cells. // Molecular biology of the cell. - 2002. - Vol.13. - P.4279-4295 и др.).
Жировая ткань рассматривается как одна из альтернатив костному мозгу для получения МСК и последующего их применения в терапевтических целях. Подкожная жировая клетчатка, как и костный мозг, является производным мезенхимы и содержит строму, которая может быть легко изолирована. К тому же процедура взятия жировой ткани является значительно менее травматичной и переносится пациентами значительно легче, чем пункция костного мозга. Многими исследователями, независимо друг от друга, показано, что клетки, выделяемые при ферментативной обработке жировой ткани и последующем культивировании in vitro, способны дифференцироваться в различные клеточные типы под воздействием соответствующих индукторов (Zuk P.A., Zhu M., Mizuno H. et al. Human adipose tissue is a source of multipotent stem cells. // Molecular biology of the cell. - 2002. - Vol.13. - P.4279-4295; Katz A.J., Tholpady A, Tholpady S.S., Shang H., Ogle R.C. Cell surface and transcriptional characterization of human adipose-derived adherent stromal (hADAS) cells. Stem Cells. 2005 # 23 Vol.3, p.412-423). Полученные данные свидетельствуют о том, что МСК, выделенные из жировой ткани и культивируемые in vitro, могут быть использованы в регенеративной медицине. В то же время методы выделения МСК во многих лабораториях различаются. Большинство исследователей для выделения МСК из жировой ткани используют методику, предложенную в работе Ryden (Rydén M., Dicker A., Götherström C., Aström G., Tammik C., Amer P., Le Blanc K. Functional characterization of human mesenchymal stem cell-derived adipocytes. Biochem Biophys Res Commun. 2003 #311, Vol.2. p391-397), которая заключается в измельчении ткани, обработке ее коллагеназой, нескольких последовательных центрифугированиях и адгезии полученного клеточного осадка на пластике. Получаемые при выделении из жира клетки называют мультипотентными мезенхимальными стромальными клетками-предшественниками, выделенными из липоаспирата или жировой ткани (далее - лМСК).
Возможное использование лМСК в регенеративной медицине ставит перед исследователями серьезные проблемы - как за меньший период времени культивирования МСК вне организма человека нарастить достаточную для использования клеточную массу с высоким уровнем прогениторных свойств.
Целью изобретения является ускорение получения низкодифференцированных лМСК путем модификации пролиферативной активности и дифференцировочной потенции этих клеток за счет предкультивирования лМСК в условиях гипоксии.
Состав газовой среды является одним из наиболее значимых факторов, определяющих жизнедеятельность клеток. В современной исследовательской практике культивирование клеток проводится обычно с использованием среды, содержание кислорода в которой соответствует содержанию кислорода в воздухе. Снижение уровня кислорода в среде позволяет приблизить условия культивирования к физиологическим.
Техническим результатом является ускорение наращивания массы лМСК за счет более высокой пролиферативной активности, без потери жизнеспособности клеток.
Способ осуществлялся следующим образом.
МСК выделяли из липоаспирата человека. Липоаспират можно получить после процедуры липосакции у пациентов, обратившихся в специализированную клинику. Материал до выделения хранили в холодильнике при 4°С.
Выделение лМСК из липоаспирата и получение первичной культуры проводили с помощью следующих средств.
Материалы:
Пробирки стерильные, 50 мл (Nunc, Дания)
Пипетки стерильные, 10 и 25 мл (Nunc, Дания)
Клеточный фильтр, стерильный, 100 нм (Nunc, Дания)
Чашки Петри, 60 и 90 мм, стерильные, (Nunc, Дания)
Флаконы культуральные, 25 см2 и 75 см2 (Nunc, Дания)
Пробирки нестерильные для проточного цитофлуориметра
Наконечники стерильные на 200-1000 мкл (Eppendorf, Германия)
Ростовая среда: DMEM с низким содержанием глюкозы, пенициллин 100 ед/мл, стрептомицин 100 мкг/мл
Амфотерицин В 50 мкг/мл, L-глютамин 2 мМ, натрия бикарбонат 1 г/л
Сыворотка фетальная телячья (FBS) (Hyclone, США)
Трипсин-EDTA 0,25-0,04% (Gibco, UK)
D-PBS (Gibco, UK)
Коллагеназа IА (Sigma-Aldrich, США)
Полная среда: ростовая среда +10% FBS
Оборудование:
Центрифуга Eppendorf 5804R
Ламинарный шкаф (Сампо, Россия)
Водяная баня (Elmi, Латвия)
Весы (Ohaus, Германия)
Электрическая пипетка
Пипетки автоматические, набор (Eppendorf, Германия)
Микроскоп инвертированный, фазово-контрастный (Leica, Германия)
Проточный цитофлуориметр BeckmanCoulter Epix XL (BeckmanCoulter, США)
СО2-инкубатор (Sanyo, Япония)
Стандартные условия культивирования: 5% CO2+95% воздуха, 37°С, 100% влажность.
В 50 мл пробирку помещали липоаспират (примерно 1/3 от объема пробирки) и, долив до 50 мл D-PBS, аккуратно встряхивали 5 раз.
Центрифугировали 5 минут при 1500 оборотов в минуту, при 18°С. На дне пробирки находится осадок из эритроцитов, над осадком - слой буфера и затем липоаспират. Липоаспират сверху покрыт слоем жира из разрушенных адипоцитов.
Осторожно переносили липоаспират в чистую стерильную пробирку (50 мл) и повторно отмывали ткань при следующих условиях - 10 минут, 1000 об/мин, 18°С.
Осторожно переносили липоаспират в предварительно взвешенную стерильную пробирку (50 мл), взвешивали и добавляли раствор коллагеназы IA. Приготовляли 0,15% раствор коллагеназы.
В пробирку с предварительно взвешенным липоаспиратом добавляли раствор фермента до конечной концентрации 0,075%.
Инкубировали на водяной бане 30 минут при 37°С, периодически встряхивали - 1 раз в 5 минут.
Инактивировали коллагеназу IA добавлением полной среды до 50 мл.
Центрифугировали 5 минут, 1500 об/мин, при 18°С.
Супернатант сливали и осадок ресуспендировали в 10 мл полной среды, доведя до 50 мл ростовой средой.
Центрифугировали 5 минут, 1500 об/мин, при 18°С.
Супернатант сливали и осадок ресуспендировали в 10 мл полной среды.
Клеточный фильтр помещали на пробирку 50 мл и суспензию клеток и остатков ткани пропускали через него. В пробирку добавляли ростовую среду до 50 мл.
Центрифугировали 10 минут, 1000 об/мин, при 18°С.
Супернатант сливали, осадок ресуспендировали в 10 мл ростовой среды.
Отбирали аликвоту среды с клетками и подсчитывали количество ядросодержащих клеток в гемацитометре.
Разводили клеточную суспензию из расчета, чтобы плотность посадки составляла 2×105-3×105 см2.
Размещали клетки в культуральные флаконы, оставляли в СО2-инкубаторе на 24 часа.
Отбирали надосадочную жидкость с неприкрепившимися клетками и 2 раза промывали D-PBS.
Добавляли нужное количество полной среды.
Меняли среду через 2 дня на 3.
Контролировали рост клеток под микроскопом.
Пассировали клетки при достижении 70-80% конфлуентности.
Создание условий культивирования с пониженным содержанием кислорода в среде проводили следующим образом. Часть клеток сразу после выделения помещали в мультигазовый инкубатор Sanyo (Япония), где поддерживалась соответствующая концентрация кислорода (5%). Культура клеток постоянно находилась в условиях пониженного содержания кислорода, и период нормоксии составлял не более 30 минут при замене среды.
Сразу после выделения все клетки были разделены на 2 части и помещены в условия нормоксии (95% воздуха, 5% СО2, N-клетки, N-лМСК) или гипоксии (5% O2, 5% CO2, 90% N2, Hyp-клетки, Нур-лМСК). Адгезию клеток после выделения, культивирование и последующее субкультивирование постоянно проводили в указанных газовых средах.
Далее было проведено сравнительное изучение пролиферативной активности, жизнеспособности и иммунофенотипа лМСК, перемещенных в среду культивирования с содержанием кислорода, пониженным до 5%, 3%, 1% и 3%,1% после постоянного предкультивирования клеток при 20% и 5% O2 соответственно. Сравнительную оценку пролиферативной активности проводили в течение 4-х пассажей (с 1-го по 4-й).
Предкультивирование проводится для обогащения клеточной популяции малодифференцированными клетками-предшественниками сразу после выделения МСК до достижения клетками монослоя.
Пример 1.
лМСК выделяли согласно описанной выше методике. Выделенные клетки предкультивировали в соответствующих условиях (20% или 5% О2) и получали соответственно N-лМСК и Нур-лМСК. На первом пассаже часть клеток оставляли в исходных условиях (20% или 5% О2), а часть перемещали в условия пониженного содержания О2 согласно табл.1.
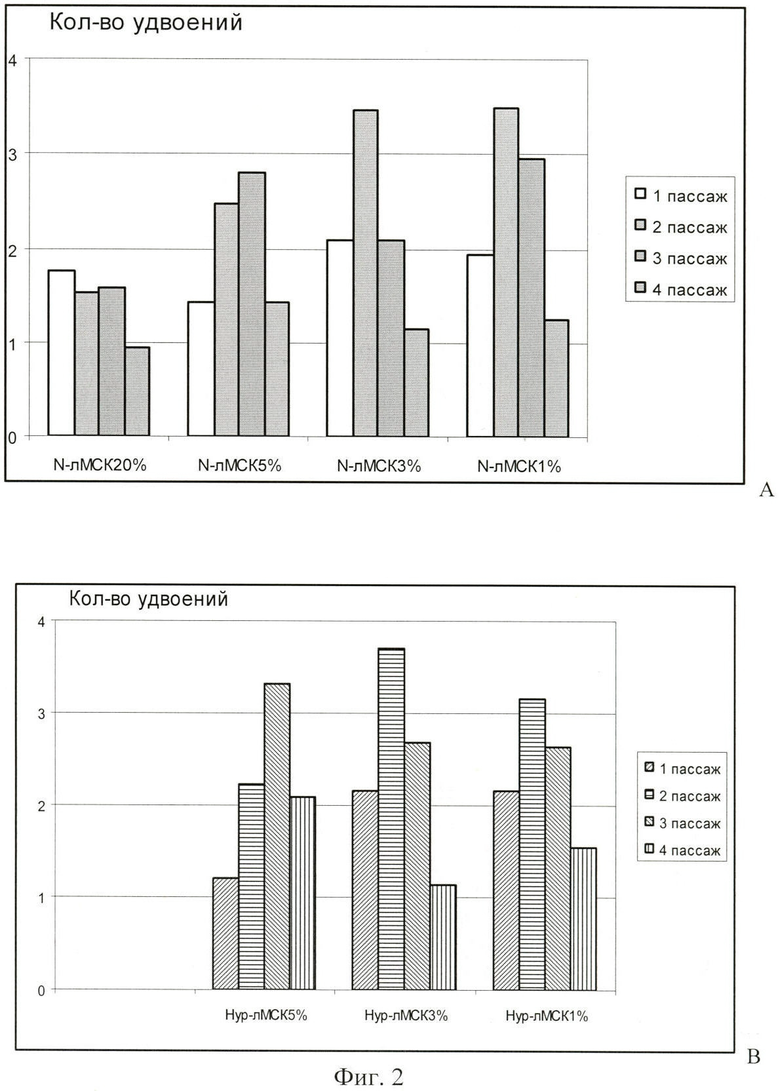
В течение 4-х пассажей при всех вариантах культивирования лМСК определяли клеточный прирост, рассчитывая количество клеточных удвоений в течение каждого пассажа и затем суммировали эти данные для определения количества клеточных удвоений за все время культивирования.
При сравнении клеточного прироста в культурах М-лМСК 20% и культурах этих же клеток, помещенных в гипоксические условия (фиг.1А), оказалось, что при понижении содержания О2 пролиферативная активность лМСК увеличивалась в 1,5-2 раза по сравнению с лМСК в нормоксии. Клеточный прирост Нур-лМСК 5%, Нур-лМСК 3% и Нур-лМСК 1% был примерно одинаков и превышал таковой для N-лМСК 20% более чем в 2 раза (фиг.1В). В целом клеточный прирост в Нур-лМСК был выше, чем в N-лМСК, переведенных в гипоксические условия.
Количество клеточных удвоений определяли по окончании каждого пассажа и затем данные суммировали. На фиг.2 приведены результаты сравнения клеточных приростов М-лМСК 20%, Нур-лМСК 5% и этих же клеток, переставленных в условия пониженного О2, в течение каждого из исследованных пассажей.
Как можно видеть на диаграммах, приведенных на фиг.2, прирост N-лМСК 20% был невелик и не отличался на 1-3-м пассажах. На 2-3-м пассажах в культурах Нур-лМСК 5%, а также в культурах N-лМСК 20% и Нур-лМСК 5%, перемещенных в условия пониженного O2, наблюдалось значительное увеличение клеточного прироста. При 3% и 1% O2 на 1-3-м пассажах количество клеточных удвоений превышало этот показатель в культурах лМСК при 5% O2, а на 4-м пассаже - было в 1,5-2 раза меньше, чем при 5% O2.
Результаты оценки жизнеспособности лМСК, культивируемых при различных концентрациях О2 в газовой фазе, приведены в табл.2, 3.
Заметное снижение доли живых клеток было отмечено после первого пассажа в N-лМСК 1% и после второго пассажа в N-лМСК 3% и N-лМСК 1%.
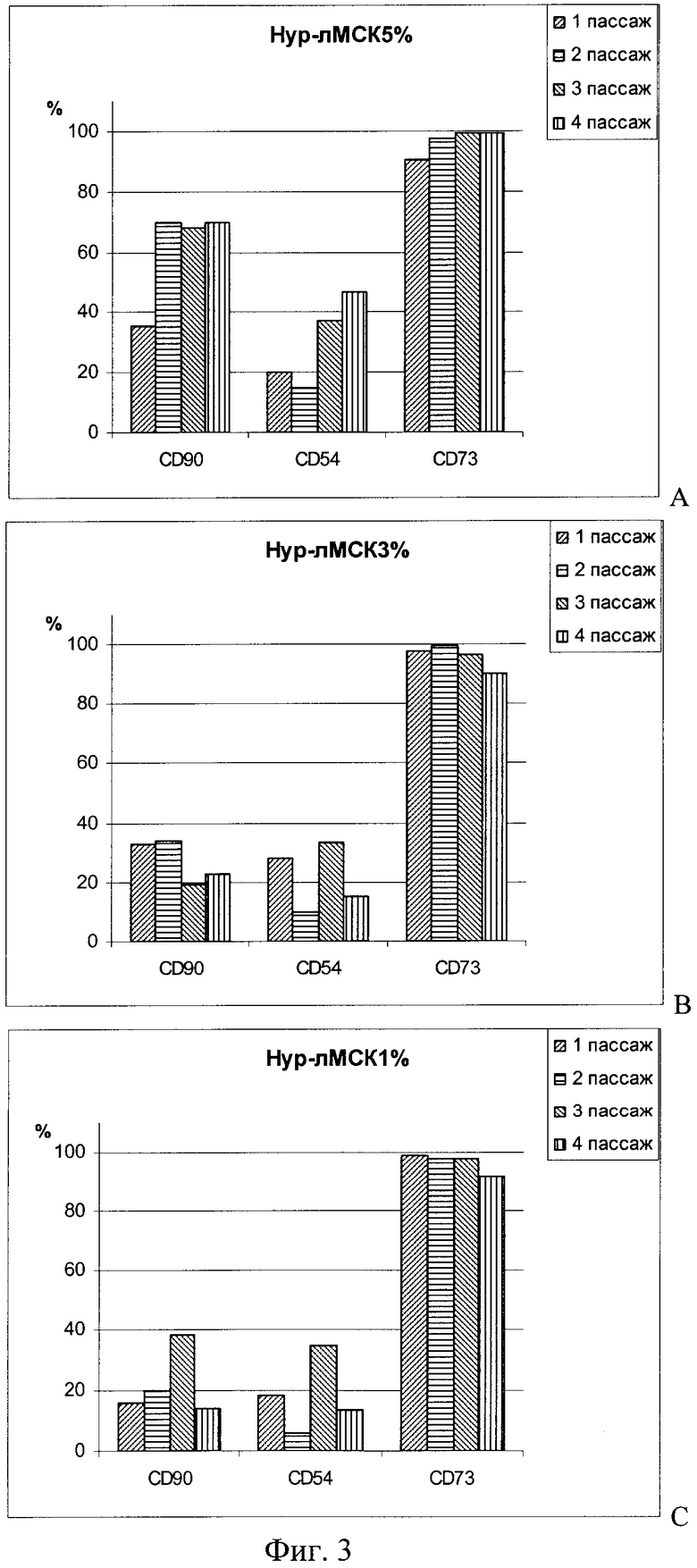
Иммунофенотипирование лМСК, культивируемых при различной концентрации О2 в среде, не выявило существенных различий по экспрессии CD73 антигена МСК (фиг.3), во всех случаях от 90 до 100% клеток были CD73-положительные клетки. Экспрессия CD54-антигена значительно варьировала на лМСК, и какой-либо зависимости от концентрации О2 или номера пассажа обнаружить не удалось. Доля СD90-положительных клеток среди лМСК, культивируемых при 5% О2 в течение 2-4-го пассажей, была достоверно выше, чем среди Нур-лМСК 3% и Нур-лМСК 1%.
Выводы.
Сравнение морфо-функциональных особенностей лМСК, культивируемых при 3% и 1% O2 после предкультивирования при 20% и 5% O2, показало следующее:
- При культивировании в условиях пониженного до 5%, 3% и 1% О2 пролиферативная активность лМСК превышает таковую при 20% О2. Наиболее значительный прирост наблюдается на 2-3-м пассажах культивирования. Пролиферативная активность лМСК, предкультивированных в нормоксии, при перемещении в гипоксию (1-3-5% О2) ниже, чем пролиферативная активность лМСК, предкультивированных в гипоксии.
- Сравнение прироста лМСК при культивировании их при различных пониженных концентрациях О2 показало, что количество клеточных удвоений в течение 4-х пассажей при 1-3-5% О2 практически одинаково, но, при 1% и 3% O2 после всплеска пролиферативной активности на 1-3-м пассажах, клеточный прирост на 4-м пассаже резко уменьшался, тогда как при 5% O2 лМСК сохраняли достаточно высокую пролиферативную активность и на последующих пассажах.
- лМСК, перемещенные в условия пониженного содержания О2, имели высокую жизнеспособность (90% и более). Заметное снижение доли живых клеток было отмечено при N-лМСК 3% и N-лМСК 1%. Можно предположить, что лМСК, которые предкультивировали в условиях нормоксии, оказались менее устойчивы к значительному снижению концентрации О2 в среде, чем лМСК, предкультивированные в условиях гипоксии.
- Большая часть лМСК в исследованных клеточных популяциях экспрессировала маркер мезенхимальных стромальных клеток CD73, а доля СD90-положительных клеток была самой большой среди Нур-лМСК 5%.
Таким образом, снижение концентрации кислорода в среде культивирования лМСК ниже 5% не оказывает повреждающего действия на них. Однако полученные данные позволяют предположить, что эффекты понижения О2 до 5% и 1-3% несколько отличаются. лМСК реагировали на снижение концентрации О2 в среде культивирования до 5% плавным увеличением клеточного прироста в течение 1-3-го пассажей с сохранением достаточно высокой пролиферативной активности в дальнейшем. При 5% О2 больше всего лМСК демонстрировали мезенхимальный маркер CD90. Кроме того, при общей высокой жизнеспособности лМСК, при перемещении N-лМСК в 3% и 1% O2 значительно увеличивалась доля мертвых клеток в популяции.
На основании полученных результатов можно предложить две возможные схемы культивирования.
Для создания условий культивирования, близких к физиологическим, которые бы обеспечивали устойчивый прирост лМСК с выраженным мезенхимальным фенотипом и высокой жизнеспособностью, необходимо использовать мягкое гипоксическое воздействие за счет снижения концентрации О2 до 5%.
В тех случаях, когда необходимо быстрое наращивание клеточной массы, в течение 1-2 пассажей могут быть использованы более жесткие гипоксические условия - снижение О2 до 3% и 1%. При этом необходимо учитывать, что лМСК, предкультивированные в нормоксических условиях перед помещением в гипоксию, имеют пониженную жизнеспособность, поэтому предпочтительно использовать лМСК, постоянно культивируемые при 5% О2.
Исследования показали, что лМСК, культивируемые при стандартном (20% O2) и пониженном до 5% содержании кислорода в среде, отличаются по своим морфо-функциональным характеристикам. В частности, лМСК, постоянно культивируемые в среде с пониженным содержанием O2, обладают более высокой пролиферативной активностью, что делает методику выращивания клеток при пониженном О2 весьма привлекательной с точки зрения подготовки значительного количества клеток для нужд клеточной терапии и регенеративной медицины.
Описание чертежей.
На Фиг.1 показан Прирост N-лМСК и Нур-лМСК, перемещенных для культивирования в среду с пониженным содержанием О2, где
А. Количество клеточных удвоений в течение 4-х пассажей в культурах N-лМСК 20% и N-лМСК при пониженном содержании О2.
В. Количество клеточных удвоений в течение 4-х пассажей в культурах Нур-лМСК 5% и Нур-лМСК при пониженном содержании О2.
Количество клеточных удвоений определяли по окончании каждого пассажа и затем данные суммировали.
На Фиг.2 показан клеточный прирост лМСК, культивируемых в газовой среде различного состава, на 1-4-м пассажах.
На Фиг.3 показан иммунофенотип лМСК, культивируемых при пониженном содержании кислорода в среде.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ КУЛЬТУР МЕЗЕНХИМАЛЬНЫХ СТРОМАЛЬНЫХ КЛЕТОК ЧЕЛОВЕКА, ВЫДЕЛЕННЫХ ИЗ ЛИПОАСПИРАТА | 2007 |
|
RU2351649C1 |
| СПОСОБ УСКОРЕННОГО ФОРМИРОВАНИЯ КОСТНОЙ МОЗОЛИ У МЛЕКОПИТАЮЩИХ | 2009 |
|
RU2404242C1 |
| СПОСОБ УСКОРЕННОГО ПОЛУЧЕНИЯ КУЛЬТУР МЕЗЕНХИМАЛЬНЫХ СТВОЛОВЫХ КЛЕТОК (МСК) МЛЕКОПИТАЮЩИХ, ИСКЛЮЧАЯ ЧЕЛОВЕКА, С НИЗКОЙ ГЕТЕРОГЕННОСТЬЮ И ВЫСОКОЙ ЖИЗНЕСПОСОБНОСТЬЮ | 2007 |
|
RU2331670C1 |
| СПОСОБ СТИМУЛЯЦИИ ФОРМИРОВАНИЯ ФИБРОЗНО-ХРЯЩЕВОГО РЕГЕНЕРАТА КОСТНОЙ МОЗОЛИ У МЛЕКОПИТАЮЩИХ | 2011 |
|
RU2461621C1 |
| СПОСОБ ЭКСПАНСИИ МОНОНУКЛЕАРНЫХ КЛЕТОК ПУПОВИННОЙ КРОВИ (пкМНК) ex vivo В ПРИСУТСТВИИ МУЛЬТИПОТЕНТНЫХ СТРОМАЛЬНЫХ МЕЗЕНХИМАЛЬНЫХ КЛЕТОК (ММСК) | 2013 |
|
RU2525143C1 |
| Ex vivo СПОСОБ ПОВЫШЕНИЯ КАЧЕСТВА КЛЕТОК-ПРЕДШЕСТВЕННИКОВ ОТ ВОЗРАСТНЫХ ДОНОРОВ | 2012 |
|
RU2484133C1 |
| СПОСОБ ОЦЕНКИ И МОНИТОРИНГА IN VITRO ЧИСТОТЫ ПРЕПАРАТА КЛЕТОК, ПРИМЕНЕНИЕ ФАКТОРА РОСТА, НАБОР | 2016 |
|
RU2738969C2 |
| Способ получения ассоциатов гемопоэтических и стромальных клеток-предшественников, способных подавлять активацию и пролиферацию аллогенных лимфоцитов | 2019 |
|
RU2722669C1 |
| КОМПОЗИЦИЯ ДЛЯ ПРОФИЛАКТИЧЕСКОГО ИЛИ ТЕРАПЕВТИЧЕСКОГО ЛЕЧЕНИЯ СОСТОЯНИЙ, СВЯЗАННЫХ С ИЗМЕНЕНИЕМ КОЛИЧЕСТВА ВОЛОС | 2018 |
|
RU2716159C2 |
| Способ получения МСК-ассоциированных недифференцированных гемопоэтических клеток-предшественников с фенотипов CD34+/CD133+ | 2016 |
|
RU2628092C1 |
Изобретение относится к биотехнологии. лМСК предкультивируют в условиях гипоксии 5% O2. Затем культивируют в условиях гипоксии: 1-3% при необходимости получить большое количество клеток за 1-2 пассажей или при 5% O2 при возможности более длительного культивирования. Изобретение позволяет получить большое количество МСК за короткий промежуток времени. 2 н.п. ф-лы, 3 ил., 3 табл.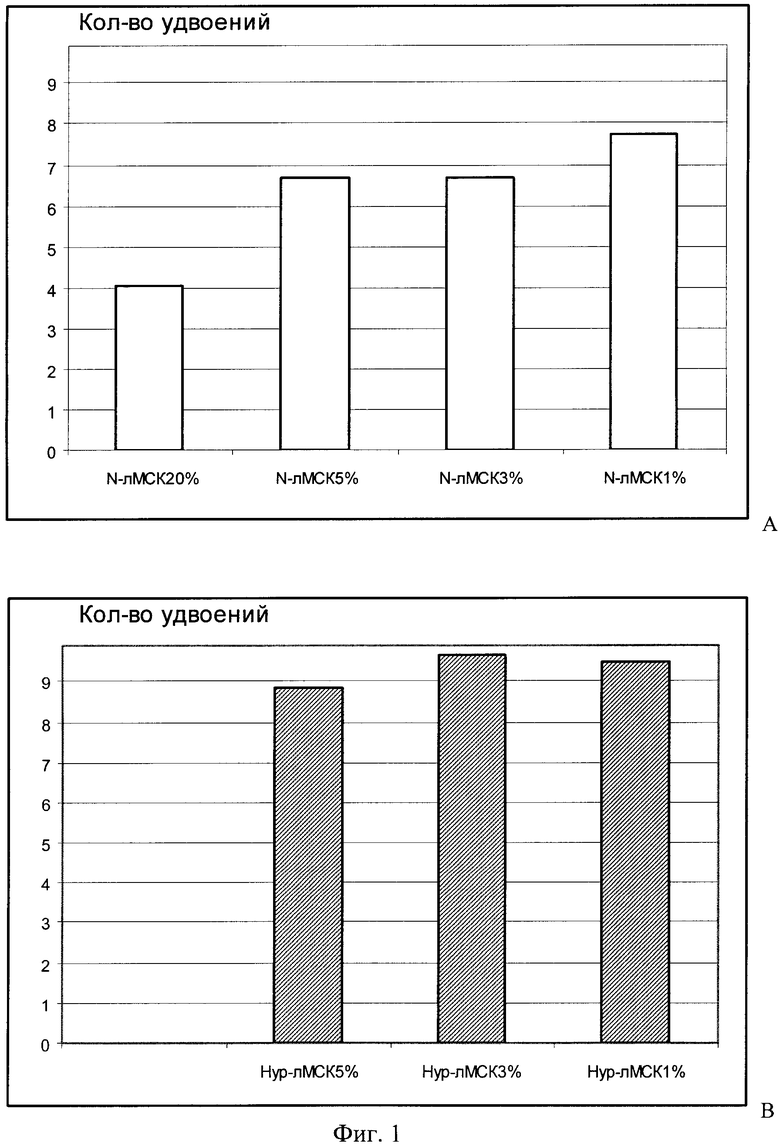
1. Способ модификации пролиферативной активности и дифференцировочной потенции мультипотентных мезенхимальных стромальных клеток in vitro, включающий:
предкультивирование лМСК в условиях гипоксии с содержанием О2 в количестве 5%;
культивирование лМСК в условиях гипоксии с содержанием O2 в количестве 1-3% в течение 1-2 пассажей.
2. Способ модификации пролиферативной активности и дифференцировочной потенции мультипотентных мезенхимальных стромальных клеток in vitro, включающий:
предкультивирование лМСК в условиях гипоксии с содержанием О2 в количестве 5%;
культивирование лМСК в условиях гипоксии с содержанием О2 в количестве 5% в течение 3 и более пассажей.
| ЖАМБАЛОВА А.П | |||
| и др | |||
| Влияние пониженного содержания кислорода на дифференцировку мультипотентных мезенхимальных стромальных клеток костного мозга человека in vitro, оригинальные исследования | |||
| - Клеточная трансплантология, 2009, т.4, №3, с.47-51 | |||
| СПОСОБ СТИМУЛЯЦИИ ПРОЛИФЕРАТИВНОЙ АКТИВНОСТИ ЛИМФОЦИТОВ | 2000 |
|
RU2167196C1 |

