Изобретение относится к медицине, а именно к биотехнологии, и может быть использовано для получения недифференцированных гемопоэтических клеток-предшественников из пуповинной крови (ГСПК), для клинического использования.
Пуповинная кровь в настоящее время рассматривается как полноценный источник гемопоэтических стволовых и прогениторных клеток.
Важным преимуществом, по сравнению с костным мозгом и мобилизованной периферической кровью, является возможность неинвазивного получения образцов ГСПК в достаточно большом количестве и использования клеток с меньшим соответствием по антигенам гистосовместимости, достаточная безопасность реципиента в связи с более редкими случаями возникновения реакции «трансплантат против хозяина» [Broxmeyer, 2005; Gluckman, Rocha, 2009].
Однако абсолютное количество гемопоэтических стволовых клеток в пуповинной крови невелико, поэтому все больший интерес вызывает возможность их экспансии ex vivo для последующего клинического применения.
Обычно для экспансии ex vivo используются суспензионные монокультуры CD34+ в статическом режиме с добавлением гемопоэтиновых коктейлей (содержащих цитокины, стимулирующие гемопоэз), обеспечивающих самообновление и пролиферацию примитивных гемопоэтических предшественников [Andrade et al., 2013]. При этом требуется очень точный подбор условий культивирования и необходимо поддержание постоянной концентрации кислорода и рН среды.
Для культивирования больших объемов клеточной суспензии применяют биореакторы, в которых обеспечивается постоянное перемешивание культуральной среды, поддержание постоянной концентрации кислорода и ее рН. Применение таких систем требует больших объемов культуральных сред и высокой квалификации персонала.
В настоящее время ведутся разработки использования трехмерных матриц с применением пористых биоматериалов Cytomatrix с целью ex vivo экспансии ГСПК [Ehring et al., 2003]. Однако использование такого поддерживающего слоя является дорогостоящим и трудоемким.
Одним из направлений экспансии ex vivo ГСПК является сокультивирование со стромальными компонентами, в частности, с мультипотентными мезенхимальными клетками (МСК). Преимущество использования фидер-содержащих систем для поддержания малодифференцированных гемопоэтических предшественников в настоящее время не вызывает сомнения.
Еще в классических работах [Dexter et al., 1977, 1982] на подслое из смешанной популяции клеток костного мозга было продемонстрировано длительное, в течение нескольких месяцев, поддержание кроветворения. Кроме того, для успешной амплификации необходим непосредственный контакт гемопоэтических и стромальных клеток, что было убедительно показано Silva et al., [2010], когда прирост малодифференцированных предшественников в трансвеллах был практически в 10 раз меньшим, чем при прямом взаимодействии. Использование стромального подслоя позволяет, в какой-то мере, имитировать структурно-функциональные особенности гемопоэтической ниши. В частности, в сокультуре с клетками стромы гемопоэтические клетки могут занимать различные компартменты.
В так называемых «Декстеровских» культурах из стромальных клеток костного мозга, которые были упомянуты выше, была описана локализация гемопоэтических клеток в 3-х областях относительно стромального подслоя.
Во-первых - это клетки, адгезировавшие к строме при инокуляции гемопоэтической суспензии, во-вторых, клетки переходящие в суспензию при делении прикрепившихся, и, в-третьих, клетки, трансмигрирующие под стромальный подслой [Dexter et al., 1977; Wagner et al., 2007; Jing et al., 2010; Andreeva et al., 2015]. CD34+ гемопоэтические предшественники в трех описанных компартментах на подслое из мезенхимальных стромальных клеток (МСК) костного мозга были фенотипированы, и установлено, что в суспензии находятся наиболее активно пролиферирующие клетки, тогда как предшественники, ассоциированные со стромой, делятся медленнее и обогащены недифференцированными гемопоэтическими клетками с фенотипом CD34+/CD38- [Wagner et al., 2007; Jing et al., 2010].
Таким образом, использование стромального подслоя из мезехимальных стромальных клеток костного мозга позволяет получать популяции гемопоэтических клеток с разной степенью коммитированности.
К сожалению, в различных протоколах, использующих сокультуру МСК/ГСПК, только вновь образующиеся суспензионные ГСПК рассматриваются в качестве целевых клеток для экспансии, хотя, как было сказано выше, это популяция содержит больше коммитированых предшественников, чем строма-ассоциированные клетки.
Рутинным этапом при выделении популяции и экспансии ГСПК является их сортировка по CD34 антигену [Brandt et al., 1990]. В популяции CD34+ ГСПК присутствуют клетки, различающиеся по потенциям к дифференцировке: часть из них может коммитироваться в несколько гемопоэтических ростков, а у других эта способность ограничена [Perey et al., 1998; Dykstra et al., 2007]. К сожалению, при иммуноселекции CD34+ клеток могут быть потеряны наиболее ранние ГСПК с фенотипом CD34- [Zanjani et al., 2003]. Такие клетки экспрессируют антиген CD133, обладая свойствами стволовых кроветворных клеток [Gallacher et al., 2000; Rutella et al., 2003]. По наличию/отсутствию CD34 и CD133 антигенов
недифференцированные гемопоэтические предшественники разделяют на ранние (CD34-/CD133+), средние (CD34+/CD133+) и поздние CD34+/CD133-) [McGuckin et al., 2003].
Вне зависимости от того, какие клетки взяты для экспансии - мононуклеары из костного мозга, пуповинной или мобилизованной периферической крови, или CD34+ популяции, в ходе ex vivo экспансии вновь образующиеся ГСПК утрачивают недифференцированный фенотип в пользу коммитированных предшественников. Это снижает их ценность, как клеточного материала для восстановления кроветворения.
Особый интерес для исследователей представляют клетки, способные длительно восстанавливать кроветворение при трансплантации, в отличие от более коммитированных предшественников, обеспечивающих только короткосрочные эффекты.
Известно, что среди ГСПК присутствуют клетки, способные формировать «области булыжной мостовой» (КООБ или CAFC (Cobblestone Area Forming Cell)) под МСК в сокультуре. Для выявления таких клеток используются тесты in vitro, основанные на выявлении частоты КООБ или их коммитированных потомков (клеток, инициирующих длительные культуры (КИДК) или LTC-IC (от слов Long-term Culture Initiating Cell)) методом лимитирующих разведений [van Os et al., 2008].
Показано, что CD34+/CD133+ клетки ПК существенно обогащены КИДК, в то время как среди CD34+/CD133- КИДК значительно меньше [De Wynter et al., 1998]. При этом известно, что большая часть (до 80%) гемопоэтических клеток пкМНК представлена поздними предшественниками с фенотипом CD34+/CD133- и, в меньшей степени, клетками с экспрессией ранних маркеров.
В связи с этим задача разработки подходов для более эффективной и физиологически обоснованной экспансии гемопоэтических клеток, по-прежнему остается одной из наиболее значимых в области клеточной
терапии и регенеративной медицины.
Наиболее близким аналогом изобретения является способ экспансии мононуклеарных клеток пуповинной крови (МНК ПК) ex vivo в присутствии мультипотентных мезенхимальных стромальных клеток (МСК) человека (RU 2525143, 10.08.2014 г.). Данный способ включает культивирование МСК из стромально-васкулярной фракции жировой ткани до достижения монослоя при концентрации O2 в среде 5%, добавление суспензии МНК ПК к монослою МСК, культивирование в течение 72 часов при концентрации O2 в среде 5%, отбор неприкрепленных МНК ПК и замену среды, продолжение культивирования МСК с прикрепившимися к ним МНК ПК в течение 7 дней при концентрации O2 в среде 5%.
Показано, что МСК из жировой ткани могут эффективно поддерживать жизнеспособность гемопоэтических предшественников и сокультивирование МНК и МСК ex vivo, обогащая популяцию мононуклеаров из пуповинной крови гемопоэтическими предшественниками.
Недостатком данного метода является получение клеток ГСПК разной степени коммитированности. Кроме того, в данном методе экспансии показано обогащение популяции клетками-предшественниками, положительными по каждому отдельному маркеру CD34+ и CD133+, при этом такая позитивная сортировка CD34+ и CD133+ клеток включает популяцию клеток с фенотипами более поздней дифференцировки.
Поэтому задачей изобретения была разработка технологии, позволяющей не только увеличить кратность числа гемопоэтических предшественников в эксперименте in vitro, а также получить популяцию, обогащенную недифференцированными гемопоэтическими предшественниками пуповинной крови с комплексным фенотипом CD34+/CD133+, соответствующим именно ранним предшественникам кроветворения.
Для получения МСК-ассоциированных недифференцированных
гемопоэтических клеток-предшественников с фенотипом CD34+/CD133+ осуществляли следующие этапы:
- проводили подготовку стромального подслоя из МСК из стромально-васкулярной фракции жировой ткани, постоянно культивируя при концентрации O2 в среде 5%,
- добавляли мононуклеарную фракцию пуповинной крови (МНК ПК) к монослою МСК и совместно культивировали 72 часа,
- осуществляли селекцию из МНК ПК недифференцированных гемопоэтических предшественников за счет адгезии к МСК при 20% O2,
- культивировали МСК с прикрепившимися к ним МНК ПК в течение 96 часов при концентрации O2 в среде 20% с образованием строма-ассоциированной популяции ГСПК с фенотипом CD34+/CD133+.
Техническим результатом предлагаемого способа является получение популяции ГСПК, более чем на 90% представленной клетками-предшественниками с комплексным фенотипом CD34+/CD133+, ассоциированными со стромальным подслоем из мультипотентных стромальных клеток (МСК) жировой ткани и образующими области булыжника (КООБ).
Кроме того, получаемые после сокультивирования клеточные препараты не нуждаются в отделении МСК, поскольку аллогенные МСК способствуют приживлению гемопоэтических трансплантатов.
Этот технический результат достигается тем, что в способе получения популяции недифференцированных гемопоэтических предшественников пуповинной крови (ГСПК), содержащих ГСПК с фенотипом CD34+/CD133+, ассоциированных со стромальным подслоем из мультипотентных стромальных клеток жировой ткани (МСК) включающих подготовку стромального подслоя из МСК из стромально-васкулярной фракции жировой ткани, постоянно культивируя при концентрации O2 в среде 5%, добавление мононуклеарной фракции пуповинной крови (МНК
ПК) к монослою МСК, их совместное культивирование 72 часа с дальнейшей селекцией из МНК ПК недифференцированных гемопоэтических предшественников за счет адгезии к МСК при 20% O2, продолжение культивирования МСК с прикрепившимися к ним МНК ПК в течение 96 часов при концентрации O2 в среде 20% с образованием популяции ГСПК, включающих ГСПК с фенотипом CD34+/CD133+ ассоциированных со стромальным подслоем из МСК жировой ткани.
Краткое описание чертежей.
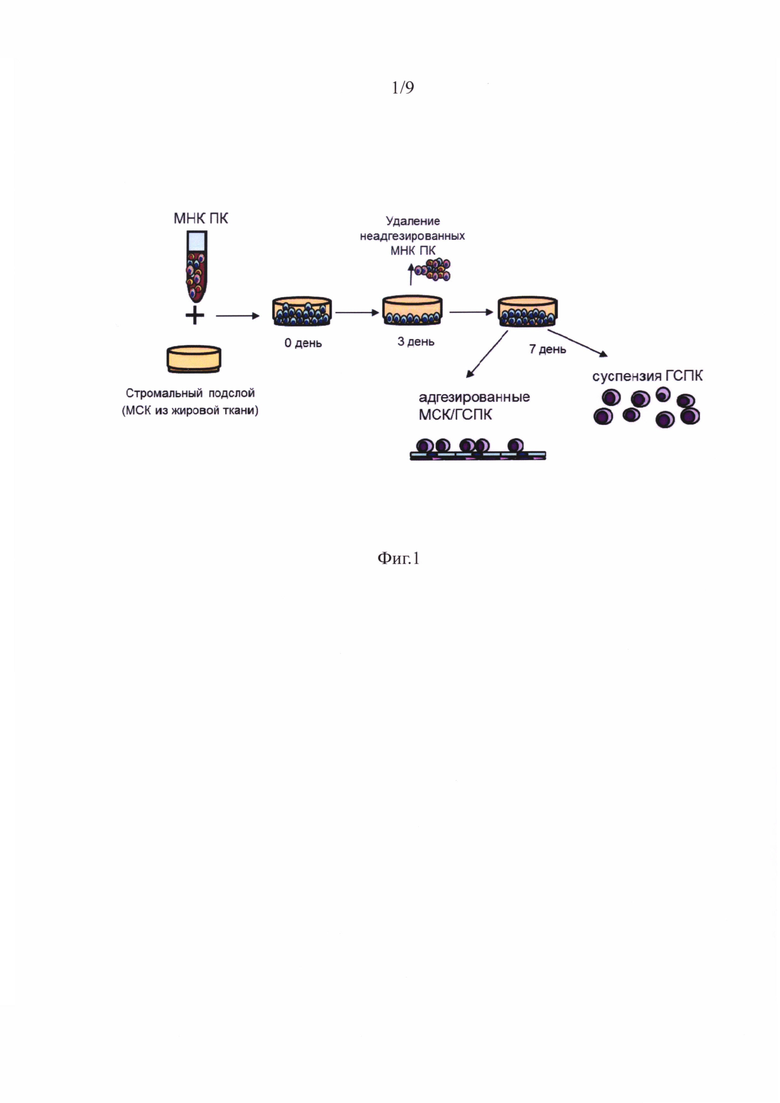
Фигура 1 - Схема получения ГСПК после сокультивирования МНК ПК с МСК.
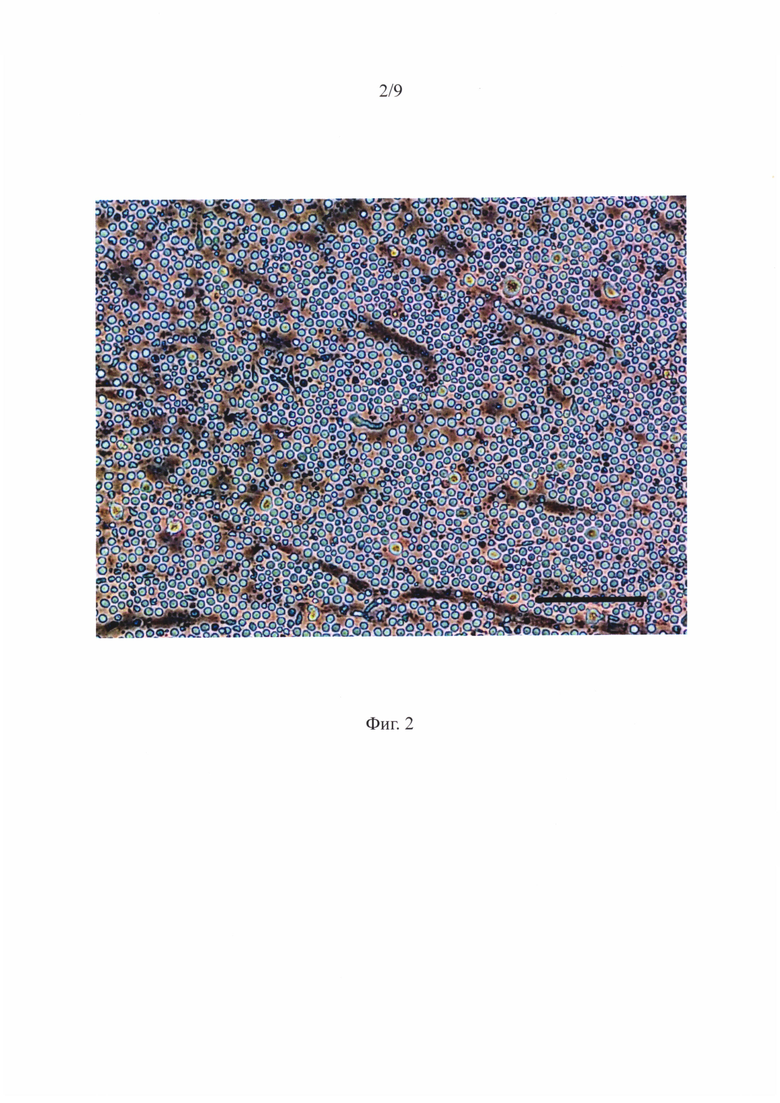
Фигура 2 - Суспензия ГСПК в сокультуре с МСК из жировой ткани после 7 суток ex vivo экспансии.
Фигура 3 - МСК-ассоциированные ГСПК на поверхности МСК и под стромальным подслоем - «клетки образующие области булыжной мостовой - (КООБ)» в сокультуре с МСК из жировой ткани после 7 суток ex vivo экспансии.
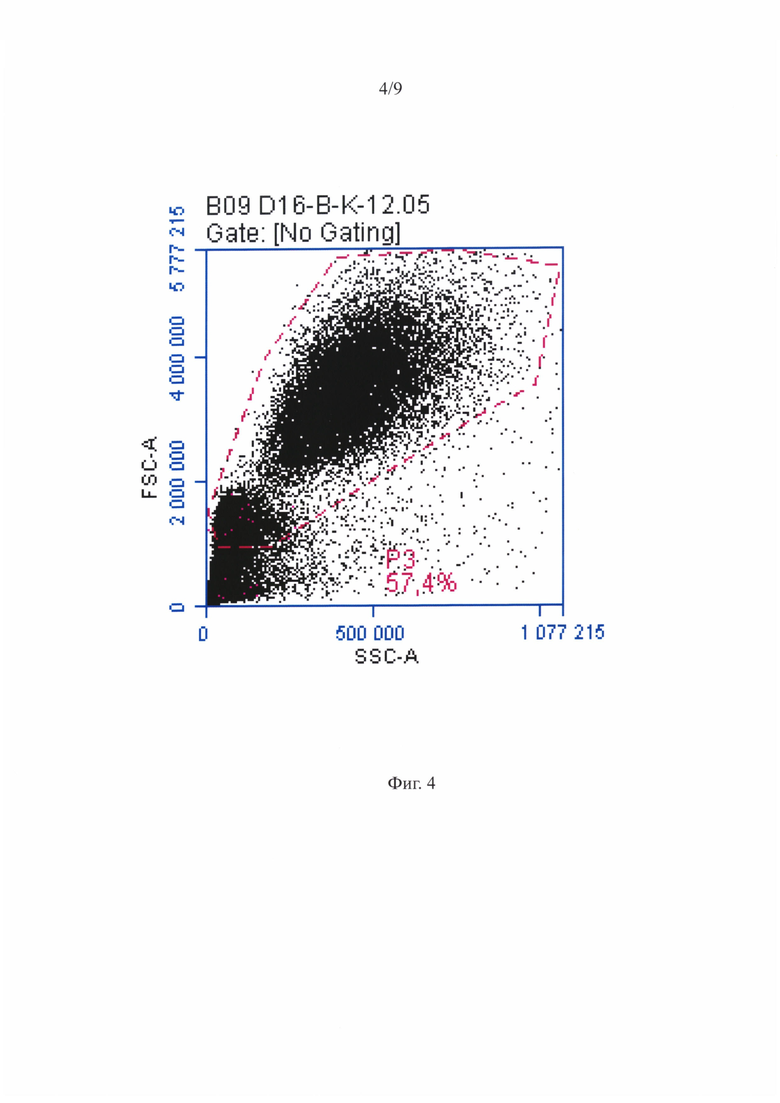
Фигура 4 - Суспензионные ГСПК при анализе на проточном цитофлуориметре - распределение по размеру и гранулярности.
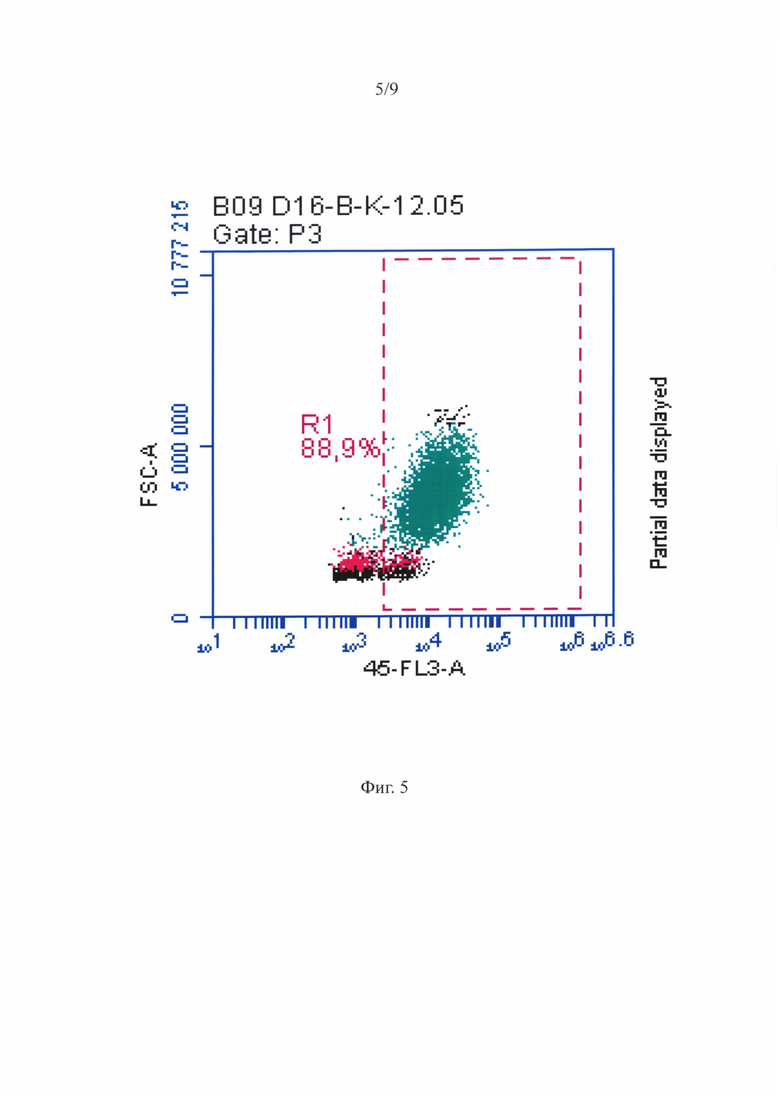
Фигура 5 - Распределение суспензионных ГСПК при анализе на проточном цитофлуориметре - гейтирование популяции ГСПК по CD45 антигену.
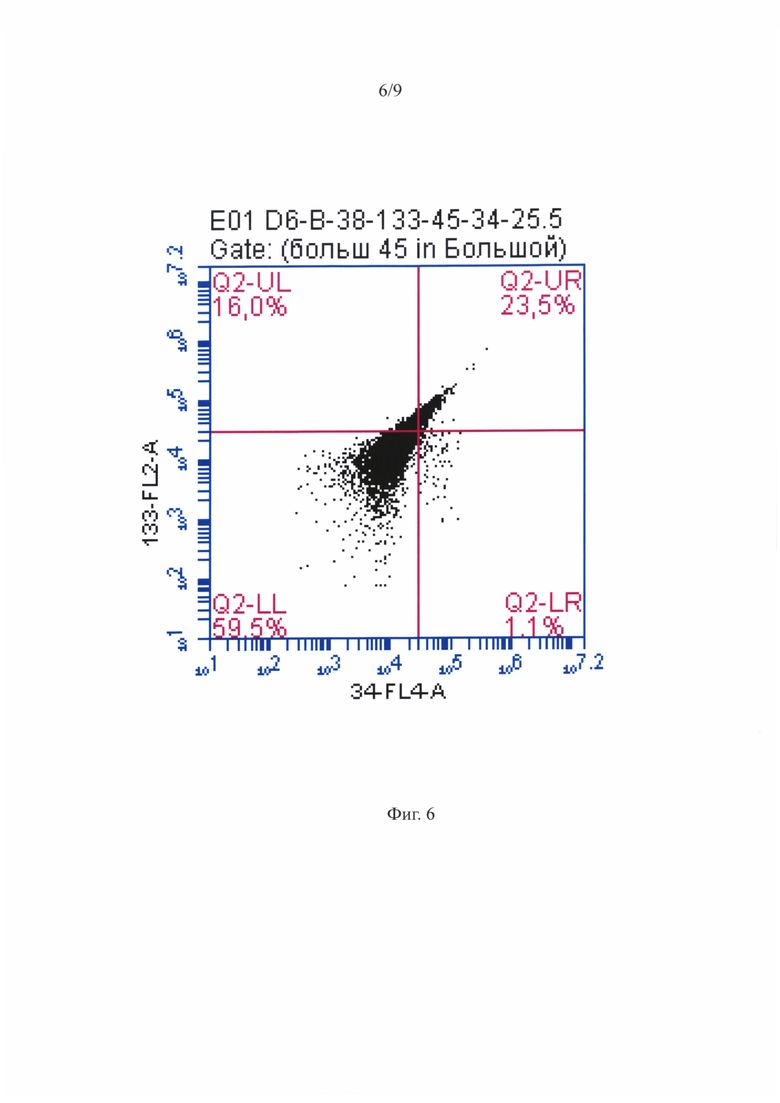
Фигура 6 - Распределение суспензионных ГСПК при анализе на проточном цитофлуориметре - CD34+ и CD133+ ГСПК среди CD45+ клеток
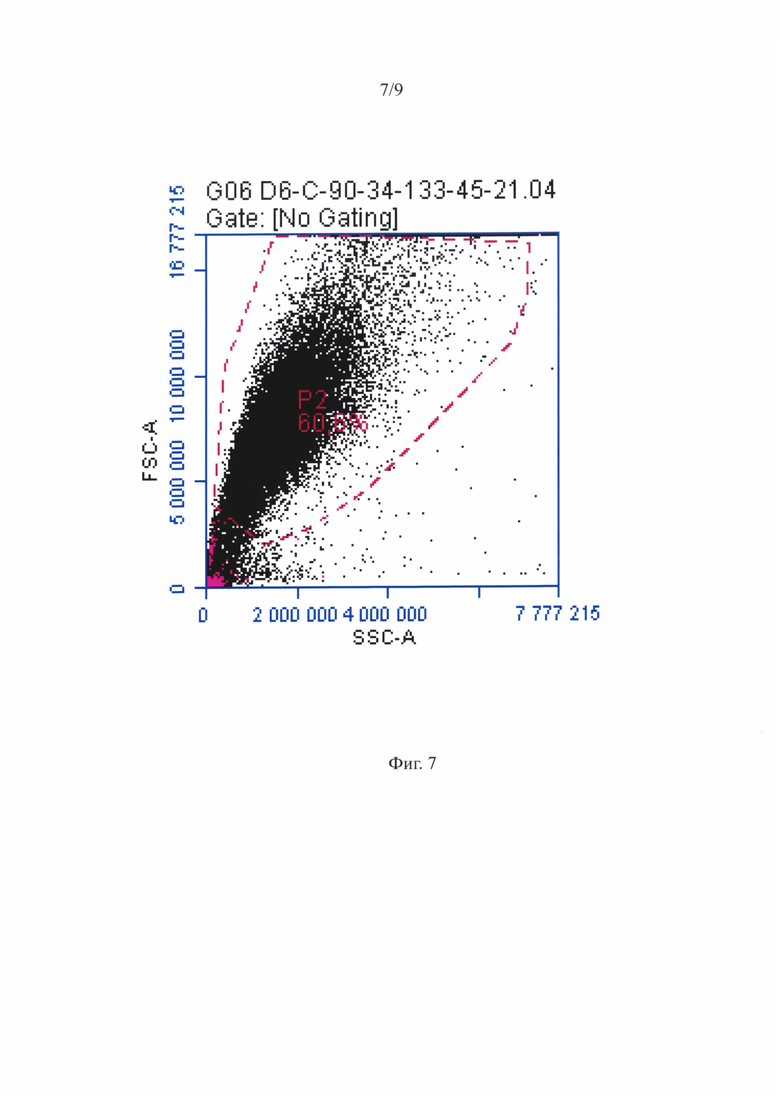
Фигура 7 - МСК-ассоциированные ГСПК при анализе на проточном цитофлуориметре - распределение по размеру и гранулярности.
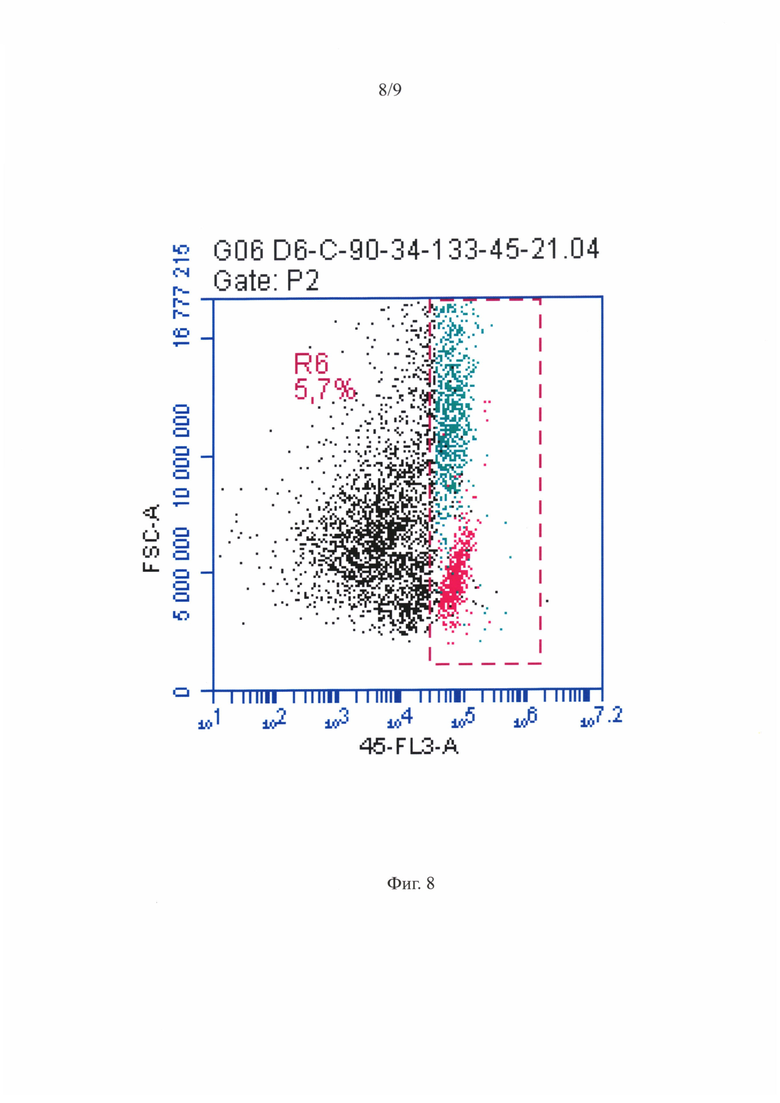
Фигура 8 - Распределение МСК-ассоциированных ГСПК при анализе на проточном цитофлуориметре. Гейтирование популяции МСК и ГСПК по CD45 антигену.
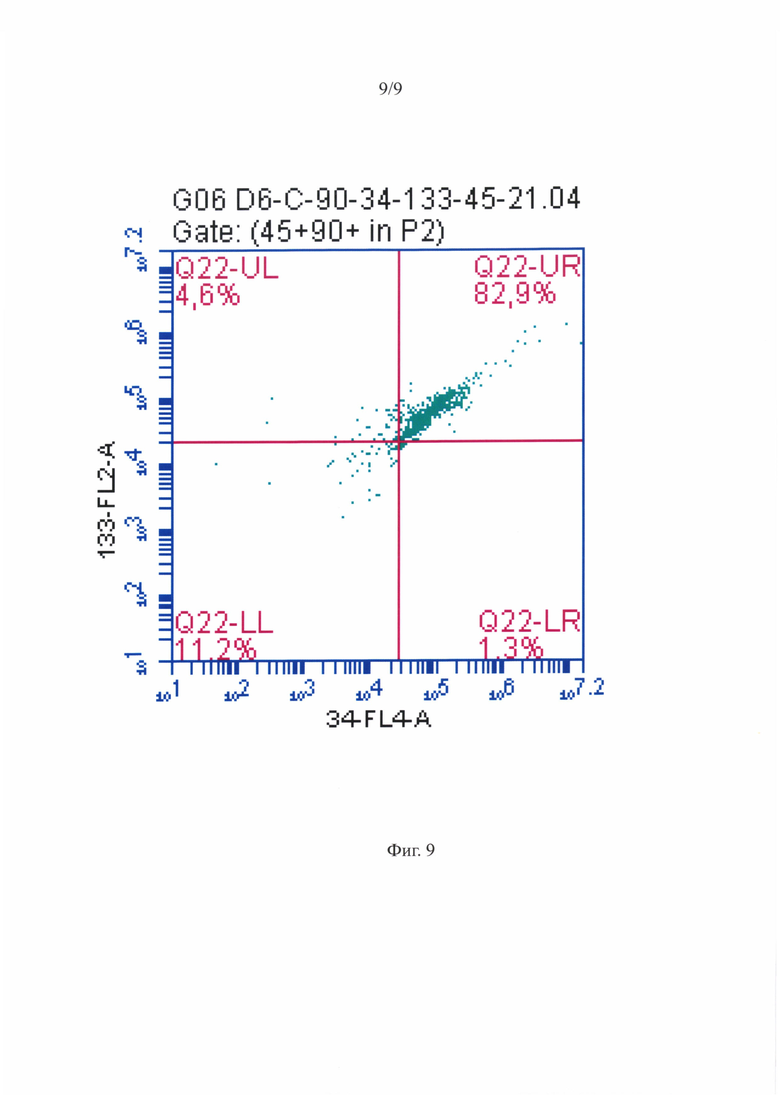
Фигура 9 - Распределение МСК-ассоциированных ГСПК при анализе на проточном цитофлуориметре - CD34+ и CD133+ ГСПК среди CD45+ клеток.
Подробное описание изобретения.
Выделение мононуклеарной фракции пуповинной крови (МНК ПК).
Заготовку ПК проводили с письменного информированного согласия обследованных здоровых рожениц в отделениях Научного центра акушерства, гинекологии и перинатологии им. акад. В.И. Кулакова. Кровь собирали в мешки донорской системы с антикоагулянтом ЦФДА-1 и обрабатывали в течение 24 часов. Получение «концентрата ядросодержащих клеток» проводили методом двойного центрифугирования в соответствии с зарегистрированной медицинской технологией (ФС2009/387 от 23.11.2009 г.). После седиментации эритроцитов и удаления избытка плазмы фракцию ядросодержащих клеток ПК ресуспендировали в аутологичной плазме с добавлением 10% диметилсульфоксида (Sigma, США) и 1% Декстрана-40, расфасовывали в криопробирки и подвергали программному замораживанию до конечной температуры -90°С в соответствии со Стандартными операционными процедурами Банка стволовых клеток. В день эксперимента клетки размораживали на водяной бане при +37°С и отмывали от криопротектора в избытке среды культивирования (см. ниже). После оценки жизнеспособности в тесте с трипановым синим концентрацию клеток доводили до 1.5-2.5×106 клеток/мл и использовали в течение 30 минут.
Выделение и культивирование МСК
Для получения МСК использовали стромально-васкулярную фракцию жировой ткани человека (жтМСК). Клетки выделяли по стандартной методике и культивировали жтМСК постоянно при 5% O2 в мультигазовом инкубаторе (Sanyo, Япония) в среде α-МЕМ (Gibco, США) с добавлением 10% инактивированной фетальной бычьей сыворотки (Hyclone, США), 100 ед./мл пенициллина и 100 мкг/мл стрептомицина (Биолот, Россия). По
достижении 70-80% конфлуентности клетки пересевали. Для экспериментов использовали МСК 2-4 пассажей.
Получение жтМСК-ассоциированных ГСПК ПК
Суспензию МНК ПК (10x106) в 5 мл среды добавляли к предмонослою МСК в чашках Петри диаметром 60 мм. Через 72 часа неадгезированные клетки отбирали и проводили замену среды культивирования на свежую. Сокультивирование приводило к адгезии части МНК ПК. Эти клетки при дальнейшем культивировании в течение 96 часов давали начало новой популяции суспензионных ГСПК, существенно обогащенных CD34+ недифференцированными гемопоэтическими предшественниками. При этом часть ГСПК оставались ассоциированными с МСК, формируя два компартмента - на поверхности и под монослоем МСК. По окончании сокультивирования вновь образованную суспензию ГСПК отбирали, а оставшийся комплекс МСК/ГСПК трипсинизировали. Затем из суспензионных и МСК-ассоциированных ГСПК готовили пробы для характеристики иммунофенотипа и жизнеспособности с помощью проточной цитофлуориметрии. Схема получения ГСПК после сокультивирования МНК ПК с МСК показана на фигуре 1.
Иммунофенотипирование ГСПК проводили среди исходных ядросодержащих клеток ПК, адгезированных ГСПК ПК и популяции клеток, вновь образующихся при дальнейшем культивировании прикрепленных клеток с помощью проточной цитометрии (Accuri С6, BD Biosciences, США). В работе использовали ФИТЦ-, фикоэритрин- и PerCP-конъюгированные антитела к CD45, CD34, CD133 а также FITC-меченые антитела к антигену МСК - CD90 (BD Pharmingen, США) в концентрации, рекомендованной изготовителем.
Статистика . Статистический анализ проводили с использованием пакета программ «Microsoft Excel 2000» и «Statistica 7.0» и критерия Манна-Уитни. Различия считали достоверными при р<0.05.
Пример 1.
К предмонослою жтМСК (70-80% конфлюэнтности) культивирование которых проводили при 5% O2 добавляли суспензию МНК ПК в количестве 10×106.
Сокультивирование мононуклеаров пуповинной крови и стромально-васкулярной фракции жировой ткани человека проводили при концентрации O2 в среде 20%. Неадгезированные в течение 72 часов МНК ПК удаляли и проводили дальнейшее культивирование оставшихся клеток в течение 96 часов при концентрации O2 в среде 20% с образованием строма-ассоциированной популяции ГСПК.
В ходе работы было показано, что после 7 суток ex vivo экспансии в сокультуре жтМСК/ГСПК можно обнаружить 2 популяции гемопоэтических клеток. На фигуре 2 приведено репрезентативное изображение суспензии ГСПК, полученное при микроскопии методом фазового контраста, масштабный отрезок соответствует 100 мкм. На фигуре 3 красными стрелками показаны МСК-ассоциированные ГСПК на поверхности МСК, желтыми стрелками - МСК-ассоциированные ГСПК под стромальным подслоем - «клетки образующие области булыжной мостовой - (КООБ)» в сокультуре с МСК из жировой ткани после 7 суток ex vivo экспансии (фазовый контраст, масштабный отрезок - 100 мкм),
суспензионные ГСПК и строма-ассоциированные ГСПК, куда были отнесены как клетки, находящиеся на поверхности МСК, так и клетки, образующие области булыжной мостовой под стромальным слоем (фигура 3).
Жизнеспособность ГСПК в обеих популяциях согласно тесту с трипановым синим составляла более 90%.
В обеих популяциях было определено соотношение ранних, средних и поздних недифференцированных гемопоэтических предшественников.
На фигуре 4 приведена репрезентативная гистограмма распределения суспензионных ГСПК по размеру и гранулярности при анализе на проточном цитофлуориметре. На точечной диаграмме, характеризующей ГСПК по размеру и структуре, можно было видеть присутствие двух субпопуляций разного размера. Клетки меньшего размера образовывали плотное облако, характерное для лимфоцитарно-моноцитарного гейта. Во второй субпопуляции клетки были крупнее с более гранулярной цитоплазмой. Количество крупных клеток было в среднем в 1, 5 раза больше, чем мелких (фигура 4). На фигуре 6 показано распределение суспензионных ГСПК при анализе на проточном цитофлуориметре - гейтирование популяции ГСПК по наличию CD45 антигена. На фигуре 7 приведено репрезентативное распределение клеток несущих CD34, CD133 и оба антигена сразу среди CD45+ ГСПК (крупные клетки). В пробах МСК-ассоциированных ГСПК, использованных для цитофлуориметрического анализа, присутствовали как ГСПК, так и МСК.
Репрезентативная диаграмма распределения МСК-ассоциированныхГСПК при анализе на проточном цитофлуориметре по размеру и гранулярности приведена на фигуре 7. Анализ по размеру/структуре не позволил выявить четко различимых субпопуляций. Также, как и для суспензии ГСПК, было проведено гейтирование популяции МСК и ГСПК по CD45 антигену, как показано на фигуре 8, после чего были выявлены CD34+ и CD133+ ГСПК среди CD45+ клеток, репрезентативное распределение которых показано на фигуре 9. Практически все CD45+ клетки положительно окрашивались антителами против CD34 и CD133 антигенов некоммитированных гемопоэтических предшественников.
В настоящей работе мы впервые проанализировали соотношение недиференцированных ГСПК разной степени зрелости после ex vivo экспансии с использованием МСК из жировой ткани, культивирование которых было проведено в условия «физиологической» гипоксии (5% O2).
В таблице 1 представлены количественные данные по содержанию ГСПК, относящихся к ранним, средним и поздним недифференцированным предшественникам.
Как видно из представленной таблицы 1 менее половины суспензионных CD45+ клеток несли антигены примитивных ГСПК. Среди них было около 20% ранних (CD34-/CD133+) и 30% средних (CD34+/CD133+) предшественников. Практически все CD45+ ГСПК, ассоциированные с МСК, были представлены примитивными гемопоэтическими предшественниками с фенотипом средних предшественников CD34+/CD133+ (более 90%). В обеих популяциях практически не было поздних CD34+/CD133- ГСПК.
Эффективность предложенного способа, позволяющего культивировать ГСПК, способствует пролиферации гемопоэтических предшественников с фенотипом CD34+/CD133+ без их дальнейшей дифференцировки в короткие сроки (7 суток).
В нашей работе среди CD34+/CD133+ ГСПК, ассоциированных с МСК, большая часть была представлена именно КООБ, что подтверждено микроскопическим анализом (фигура 3), выявившим КООБ под МСК, а также данными проточной цитофлуориметрии о присутствии крупных CD34+/CD133+ клеток в общей популяции МСК-ассоциированных ГСПК. Мы обнаружили, что ГСПК в сокультуре с жтМСК занимают те же компартменты относительно стромального слоя, что и в случае с МСК из костного мозга.
Поскольку время в сокультуре, за которое мы получили эти клетки, составило 168 часов (7 суток), то можно предположить, что в иерархии КООБ описанные клетки относятся к КООБ-7, способные коммитироваться в мульти- и бипотентные КОЕ: КОЕ-ГЭММ, КОЕ-ГМ, БОЕ-Э in vitro, а также КОЕ-С in vivo [de Haan and van Zant, 1997].
Таким образом, нами предложен эффективный метод получения и увеличения количества недифференцированных ГСПК с комплексным фенотипом CD34+/CD133+, ассоциированных со стромальным подслоем из
мультипотентных стромальных клеток (МСК) жировой ткани, при этом популяция получаемых клеток более чем на 90% представлена гемопоэтическими прогениторами с фенотипом CD34+/CD133+ и с формированием клеток, образующих области булыжника (КООБ), имеющих фенотип - примитивных предшественников.
Список литературы
- Broxmeyer HE. Biology of cord blood cells and future prospects for enhanced clinical benefit. Cytotherapy. 2005. - 7(3): 209-218.
- Gluckman E, Ruggeri A, Rocha V, et al. Family-directed umbilical cord blood banking. Haematologica. 2011. 961: 1700-1707.
- Andrade PZ, Santos FD, Cabral JM, da Silva CL. Stem cell bioengineering strategies to widen the therapeutic applications of haematopoietic stem/progenitor cells from umbilical cord blood. J Tissue Eng Regen. 2015. 9:988-1003.
- Ehring B, Biber K, Upton TM, et al. Expansion of HPCs from cord blood in a novel 3D matrix. Cytotherapy. 2003; 5(6): 490-9.
- Dexter TM, Allen TD, Lajtha L.G. Conditions controlling the proliferation of haemopoietic stem cells in vitro. J Cell Physiol. 1977. 91.3: 335-344.
- Dexter TM. Stromal cell associated haemopoiesis. J Cell Physiol Suppl. 1982. 1: 87-94.
- Da Silva CL, Goncalves R, Crapnell KB et al. A human stromal-based serum-free culture system supports the ex vivo expansion/maintenance of bone marrow and cord blood hematopoietic stem/progenitor cells. Exp Hematol. 2005. 33: 828-835.
- Wagner W, Wein F, Roderburg C, et al. Adhesion of hematopoietic progenitor cells to human mesenchymal stem cells as a model for cell-cell interaction. Exp. Hematol. 2007. 35(2): 314-325.
- Jing D, Fonseca AV, Alakel N et al. Hematopoietic stem cells in co-culture with mesenchymal stromal cells--modeling the niche compartments in vitro. Haematologica. 2010. 95(6):542-550.
- Andreeva ER, Andrianova IV, Sotnezova EV et al. Human adipose-tissue derived stromal cells in combination with hypoxia effectively support ex vivo expansion of cord blood haematopoietic progenitors. PLoS One. 2015. 10(4):e0124939.
- Brandt JE, Galy AH, Luens KM, et al. Bone marrow repopulation by human marrow stem cells after longterm expansion culture on a porcine endothelial cell line. Exp Hematol. 1998. 26: 950-961.
- Perey L, Peters R, Pampallona S et al. Extensive phenotypic analysis of CD34 subsets in successive collections of mobilized peripheral blood progenitors. Br J Haematol. 1998. 103(3):618-29.
- Dykstra B, Kent D, Bowie M, McCaffrey et al. Long-term propagation of distinct hematopoietic differentiation programs in vivo. Cell Stem Cell. 2007. 1(2):218-29.
- Zanjani ED, Almeida-Porada G, Livingston AG et al. Reversible expression of CD34 by adult human bone marrow long-term engrafting hematopoietic stem cells. Exp Hematol. 2003; 31(5): 406-412.
- Gallacher L, Murdoch B, Wu DM et al. Isolation and characterization of human CD34(-)Lin(-) and CD34(+)Lin(-) hematopoietic stem cells using cell surface markers AC133 and CD7. Blood. 2000 May 1;95(9):2813-20.
- Rutella S, Bonanno G, Marone M et al. Identification of a novel subpopulation of human cord blood CD34-CD133-CD7-CD45+lineage-cells capable of lymphoid/NK cell differentiation after in vitro exposure to IL-15. J Immunol. 2003. 171(6):2977-88.
- McGuckin CP, Pearce D, Forraz N et al. Multiparametric analysis of immature cell populations in umbilical cord blood and bone marrow. Eur J
Haematol. 2003. 71(5):341-50
- van Os RP, Dethmers-Ausema B, de Haan G. In vitro assays for cobblestone area-forming cells, LTC-IC, and CFU-C. Methods Mol Biol. 2008; 430: 143-157.
- De Wynter EA, Buck D, Hart С et al. CD34+AC133+ cells isolated from cord blood are highly enriched in long-term culture-initiating cells, NOD/SCID-repopulating cells and dendritic cell progenitors. Stem Cells. 1998.16: 387-396.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения ассоциатов гемопоэтических и стромальных клеток-предшественников, способных подавлять активацию и пролиферацию аллогенных лимфоцитов | 2019 |
|
RU2722669C1 |
| СПОСОБ ЭКСПАНСИИ МОНОНУКЛЕАРНЫХ КЛЕТОК ПУПОВИННОЙ КРОВИ (пкМНК) ex vivo В ПРИСУТСТВИИ МУЛЬТИПОТЕНТНЫХ СТРОМАЛЬНЫХ МЕЗЕНХИМАЛЬНЫХ КЛЕТОК (ММСК) | 2013 |
|
RU2525143C1 |
| Способ улучшения сбора гемопоэтических клеток при их культивировании на стромальных слоях путем предварительной магнетизации последних | 2016 |
|
RU2688321C2 |
| СПОСОБ УВЕЛИЧЕНИЯ КОЛИЧЕСТВА ГЕМОПОЭТИЧЕСКИХ НЕДИФФЕРЕНЦИРОВАННЫХ СТВОЛОВЫХ КЛЕТОК ПАЦИЕНТА ex vivo | 2007 |
|
RU2360965C1 |
| БИОМЕДИЦИНСКИЙ КЛЕТОЧНЫЙ ПРОДУКТ, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ | 2018 |
|
RU2741769C2 |
| СПОСОБ ПОЛУЧЕНИЯ КУЛЬТУР МЕЗЕНХИМАЛЬНЫХ СТРОМАЛЬНЫХ КЛЕТОК ЧЕЛОВЕКА, ВЫДЕЛЕННЫХ ИЗ ЛИПОАСПИРАТА | 2007 |
|
RU2351649C1 |
| АДГЕЗИВНЫЕ КЛЕТКИ ЖИРОВОЙ ТКАНИ ИЛИ ПЛАЦЕНТЫ И ИХ ИСПОЛЬЗОВАНИЕ В ЛЕЧЕБНЫХ ЦЕЛЯХ | 2008 |
|
RU2709780C2 |
| СПОСОБ ЭКСПАНСИИ CD34+ ГЕМАТОПОЭТИЧЕСКИХ КЛЕТОК | 2011 |
|
RU2469086C1 |
| Способ получения и ведения мезенхимальных стволовых клеток из костного материала млекопитающих | 2023 |
|
RU2821926C1 |
| Биомедицинский клеточный препарат | 2017 |
|
RU2647429C1 |

Изобретение относится к медицине, биотехнологии, конкретно к получению клеточных культур, обогащенных гемопоэтическими клетками-предшественниками с фенотипом CD34+/CD133+. Способ включает подготовку стромального подслоя, добавление фракции пуповинной крови, культивирование и селекцию. Из стромально-васкулярной фракции жировой ткани получают мультипотентные стромальные клетки жировой ткани (МСК), которые культивируют при концентрации O2 в среде 5%. К монослою МСК добавляют мононуклеарную фракцию пуповинной крови (МНК ПК). После сокультивирования осуществляют селекцию из мононуклеарной фракции ПК недифференцированных гемопоэтических предшественников за счет адгезии к МСК при 20% O2, продолжают культивировать МСК с прикрепившимися к ним МНК ПК в течение 96 часов при концентрации O2 в среде 20%. Изобретение позволяет получить популяцию, обогащенную недифференцированными гемопоэтическими предшественниками пуповинной крови с комплексным фенотипом CD34+/CD133+. 9 ил., 1 табл., 1 пр.
Способ получения популяции недифференцированных гемопоэтических предшественников пуповинной крови (ГСПК), включающих ГСПК с фенотипом CD34+/CD133+, ассоциированных со стромальным подслоем из мультипотентных стромальных клеток жировой ткани (МСК), включающий подготовку стромального подслоя из МСК из стромально-васкулярной фракции жировой ткани, постоянно культивируя при концентрации O2 в среде 5%, добавление мононуклеарной фракции пуповинной крови (МНК ПК) к монослою МСК, их совместное культивирование 72 часа с дальнейшей селекцией из МНК ПК недифференцированных гемопоэтических предшественников за счет адгезии к МСК при 20% O2, продолжение культивирования МСК с прикрепившимися к ним МНК ПК в течение 96 часов при концентрации O2 в среде 20% с образованием популяции ГСПК, включающих ГСПК с фенотипом CD34+/CD133+, ассоциированных со стромальным подслоем из МСК жировой ткани.

