Изобретение относится к области молекулярной биологии и нанофармакологии и может найти применение в разработке новых противоопухолевых препаратов направленного типа действия.
Коннексин-43 (Сх43) является интегральным мембранным белком, образующим щелевые межклеточные контакты (gap-junction) между астроцитами в нормальной нервной ткани, а также между реактивными астроцитами и глиомными клетками при первичных глиальных опухолях мозга. С помощью щелевых контактов осуществляется передача межклеточных сигналов, регулирующих пролиферацию, дифференцировку, апоптоз и миграцию клеток, как в процессе нормального онтогенеза, так и при глиомной инвазии [Prochnow N, et al. 2008]. Последнее обуславливает высокий интерес к коннексину-43 как к потенциальной молекулярной мишени в терапии глиом [Huang R., et аl., 2002, Fu CT, et al., 2004 Oliveira R., et al. 2005].
Известно, что глиомные клетки, экспрессирующие белки щелевых контактов, обладают высоким инвазивным потенциалом и большей способностью к адгезии, чем коннексин-отрицательные [Lin J.Н., 2002, Bates DC, 2007], и, таким образом, могут являться источником рецидивов после комбинированного лечения мультиформной глиобластомы.
Системы для направленного транспорта к коннексин-43 положительным глиомным клеткам диагностических агентов, противоопухолевых препаратов и генетического материала до настоящего момента не разработано.
Технической задачей предлагаемого изобретения является создание наносистемы, селективной по отношению к глиомным клеткам, экспрессирующим белки щелевых контактов, в частности коннексин-43, и разработка способа получения данной наносистемы.
Решение поставленной задачи достигается в заявленной бинарной (двухкомпонентной) иммунолипосомальной наносистеме адресной доставки к коннексин-43 положительным опухолевым клеткам на основе биотинилированных моноклональных антител к экстраклеточному фрагменту коннексина-43 (первый компонент) и наноразмерных ПЭГилированных липосомальных контейнеров диаметром 70-100 нм, ковалентно связанных со стрептавидином (второй компонент).
Введение биотинилированных антител к Сх43 в живую систему сопровождается их специфическим связыванием с экстраклеточным фрагментом Сх43 на цитолемме живых глиомных клеток. Последующее введение второго компонента наносистемы сопровождается селективным накоплением липосом на поверхности этих клеток вследствие связывания стрептавидина липосом с биотином моноклональных антител. Таким образом, данная бинарная система обеспечивает высокую селективность адресной доставки содержимого липосомальных наноконтейнеров к Сх43 положительным клеткам низкодифференцированных глиом.
Бинарная наносистема адресной доставки на основе биотинилированных антител и наноразмерных липосом со стрептавидином имеет несколько существенных преимуществ по сравнению с однокомпонентными липосомальными препаратами, описанными в литературе и представленными на рынке фармпрепаратов. Во-первых, константа диссоциации комплекса стрептавидин-биотин (10-15 М) почти в два раза ниже средней константы диссоциации комплекса антиген-антитело (не менее 10-9 М), вследствие чего применение первого комплекса должно повысить специфичность накопления транспортируемых липосомальных контейнеров. Во-вторых, в случае применения бинарной системы конъюгация стрептавидина с молекулой липосом приводит к образованию существенно меньшего комплекса, чем при конъюгации с липосомой антител (молекулярная масса стрептавидина почти в три раза меньше молекулярной массы иммуноглобулинов G). Кроме того, весьма важным является следующий методический аспект: при биотинилировании сукцинимидным методом иммунохимическая активность моноклональных антител изменяется в меньшей степени, чем при тиолировании, необходимом для пришивки антител к липосомам малеимидным методом. С учетом всех перечисленных факторов бинарные (двухкомпонентные) системы на основе биотинилированных антител и липосом со стрептавидином предпочтительнее, чем иммунолипосомы.
Моноклональные антитела для бинарной системы получают при иммунизации рекомбинантным экстраклеточным фрагментом Сх43 (Q173 - 1208); [Баклаушев В.П. и соавт. 2009]. ПЦР амплификацию последовательности фрагмента Е2 из кДНК библиотеки мозга крысы проводят с помощью праймеров: CX43_173F: 5'-GATCAGA-TCTCAGTGGTACATCTATGGGT-3'; CX43_73B: 5'-GATCAAGCTTAGATGGTTTTCT-CCGTGGGAC-3' («СибЭнзим», Новосибирск), содержащих сайты узнавания рестриктаз BgIII и HindIII соответственно. Плазмидные ДНК, не содержащие ошибок в кодирующей последовательности, клонируют в экспрессирующие векторы pHPMLQ и pCBDQ (ИБХ РАН) [Dmitriev RI et al., 2007]. Эти векторы содержат последовательности двух доменов Са-АТФазы плазматической мембраны клеток человека hРМСА4b (CBD, a.o. 1057-1205, 17 кДа и HPML, а.о. 166-371, 22,5 кДа) в качестве высокомолекулярных носителей. Для экспрессии, плазмидными ДНК pHPMLQ-Cx43 и pCBDQ-Cх43 трансформируют клетки E.coli SG13009 (Qiagen). Очищенный химерный полипептид CBD-Cx43 применяют для иммунизации мышей; a HPMLQ-Cx43 - для тестирования клонов гибридных клеток методом ELISA, чтобы исключить получения антител к полипептиду-носителю. Моноклональные антитела получают с помощью традиционной гибридомной технологии, с некоторыми модификациями [Чехонин В.П., 2007]. Отобранные с помощью ELISA клоны тестируют в иммуногистохимическом анализе со срезами мозга, чтобы отобрать моноклональные антитела, взаимодействующие с Сх43 в нативной конформации.
Очистку моноклональных антител из асцита проводят с помощью аффинной хроматографии на агарозе с иммобилизованным белком G по стандартному протоколу с элюцией градиентом рН. Очищенные антитела ковалентно связывают с биотином по свободным NH2-группам с помощью карбокси-сукцинимидного эфира биотина; несвязавшийся биотин удаляют с помощью гель-фильтрации на любом из носителей, в котором иммуноглобулины G элюируются в свободном объеме. Эффективность биотинилирования определяют с помощью аффинной хроматографии на авидин-агарозе. Пробы до и после инкубации с авидин-агарозой подвергают диск-электрофорезу в ПААГ с ДСН с последующей денситометрией и сравнением количественных показателей. Эффективность биотинилирования при применении свежего карбоксисукцинимидного эфира биотина и правильного молярного соотношения IgG:биотин (1:3-1:4) составляет 90-100%.
Синтез ПЭГилированных липосомальных наноконтейнеров проводится по общепринятой методике [Kamps et al., 2000], модифицированной в соответствии с задачами конкретного эксперимента. Основными компонентами ПЭГилированных липосом являются: лецитин из желтков куриных яиц, холестерин и дистеароилфосфатидилэтаноламин (DSPE), конъюгированный с PEG-2000 (полиэтиленгликоль 2000 Да). Для связывания с тиолированным стрептавидином в состав липосом вводят малеимидное производное DSPE-PEG-2000 (1,2-дистеароил-sn-глицеро-3-фосфоэтаноламин-N-[малеимид-(полиэтиленгликоль)-2000]). В его молекуле малеимидная группа присоединена к внешнему концу цепи ПЭГ-2000, что позволяет «выводить» белок-вектор за пределы экранирующего слоя PEG. В качестве флюоресцентной метки к липосомам добавляют Dil (1,1'-октадецил-3,3,3',3'-тетраметилиндокарбоцианин перхлорат).
Лецитин, холестерин, DSPE-PEG-2000, его малеимидное производное и Dil растворяют в смеси хлороформ-метанол (9:1) в концентрации суммарных липидов 5-10 мг/мл в молярном соотношении 23:16:1.6:1:0.4. Смесь высушивают на роторном испарителе при пониженном давлении. Сухую липидную пленку растворяют в абсолютном циклогексане, замораживают в жидком азоте, лиофилизуют, после чего смесь липидов эмульгируют в 0,1 М фосфатном буфере. Гидратированную эмульсию обрабатывают в ультразвуковом дезинтеграторе и 15-кратно пропускают через поликарбонатные мембранные фильтры с размерами пор 400, 200, 100 и 50 нм с помощью мини-экструдера.
Синтезированные ПЭГилированные липосомы ковалентно связывают со стрептавидином, предварительно модифицированным 2-иминотиоланом, который реагирует со свободными первичными аминогруппами белка (в частности, с ε-аминогруппой лизина), присоединяя к ним защищенную SH-группу. Очистку stealth-липосом со стептавидином проводят с помощью гель-фильтрации на сефадексе G100 или аналогичном сорбенте. Эффективность связывания стрептавидина с липосомами оценивается по соотношению концентрации белка во фракциях липосом и несвязавшегося стрептавидина. Этот показатель при эффективной пришивке стрептавидина составляет 90-95%.
ПРИМЕР
Моноклональные антитела к Е2 Сх43 очищают из асцита на Protein-G сефарозе (Invitrogen) и биотинилируют с помощью набора ProtOn (Vector Lab, USA) согласно рекомендациям фирмы производителя. Свободный биотин удаляют методом гель-фильтрации путем центрифугирования на микроколонках для обессоливания (Biorad).
Эффективность биотинилирования, оцененная с помощью инкубации с авидин-агарозой, входящей в состав кита, составила не менее 90%.
Все манипуляции по приготовлению липосом, кроме краткосрочных (взешивание, центрифугирование и т.п.), проводят в атмосфере аргона высокой чистоты. Все липидные компоненты получены от фирмы "Avanti Polar Lipids", Alabaster AL (USA).
Лецитин, холестерин, DSPE-PEG-2000, его малеимидное производное и Dil растворяют в смеси хлороформ-метанол (9:1) в концентрации суммарных липидов 10 мг/мл в молярном соотношении 23:16:1.6:1:0.4. Смесь высушивают на роторном испарителе при пониженном давлении. Сухую липидную пленку растворяют в абсолютном циклогексане, замораживают в жидком азоте, лиофилизуют, после чего смесь липидов эмульгируют в 0,1 М фосфатном буфере. Гидратированную эмульсию обрабатывают в ультразвуковом дезинтеграторе G112SP1 Special Ultrasonic Cleaner (Laboratory Supplies Company, Hicksviil, N.Y., USA) и 15-кратно пропускают через поликарбонатные мембранные фильтры с размерами пор 400, 200, 100 и 50 нм с помощью мини-экструдера ("Avanti Polar Lipids", Alabaster AL, USA). Рекомбинантный стрептавидин (Имтек) модифицируют 2-иминотиоланом и ковалентно пришивают к ПЭГилированным липосомам.
Полученные ПЭГилированные липосомы характеризуются по диаметру и концентрациям суммарных липидов и белка. Диаметр липосом, определенный методом фотодинамического рассеивания, находится в пределах 70-100 нм. Средняя концентрация липидов в полученных липосомальных препаратах составляет 20.5±1.1 мг/мл, концентрация стрептавидина в готовой эмульсии соответствующих липосом составляет 0,4-0,6 мг/мл.
Биотинилированные антитела стерилизуют фильтрованием (Milliex filter, 0,22 µm Millipore Bedford MA USA) и добавляют в живую культуру клеток С6 глиомы в ростовой среде (DMEM с 10% FBS) до концентрации 10 µg/ml. После инкубации в течение часа среду с антителами удаляют, клетки дважды промывают PBS, затем вносят свежую ростовую среду с добавленными в нее ПЭГилированными липосомами в возрастающей концентрации (1-50 µl липосомальной эмульсии с известной концентрацией липидов и стрептавидина). Через 30 и 60 минут инкубации регистрируют флюоресценцию живых клеток на обращенном микроскопе Leica DM 6000. В данном примере проводят три серии экспериментов с несколькими контролями. В первом контроле за 30 минут до введения биотинилированных антител к Е2 Сх43 в лунки добавляли 10-кратный избыток (100 µg) небиотинилированных MAb к Е2 фрагменту. Дальнейший эксперимент проводили так же, как в опыте. Во втором контроле применялись неспецифические биотинилированные IgG мыши и векторные stealth-липосомы. В третьем контроле применялись stealth-липосомы, неконъюгированные со стрептавидином.
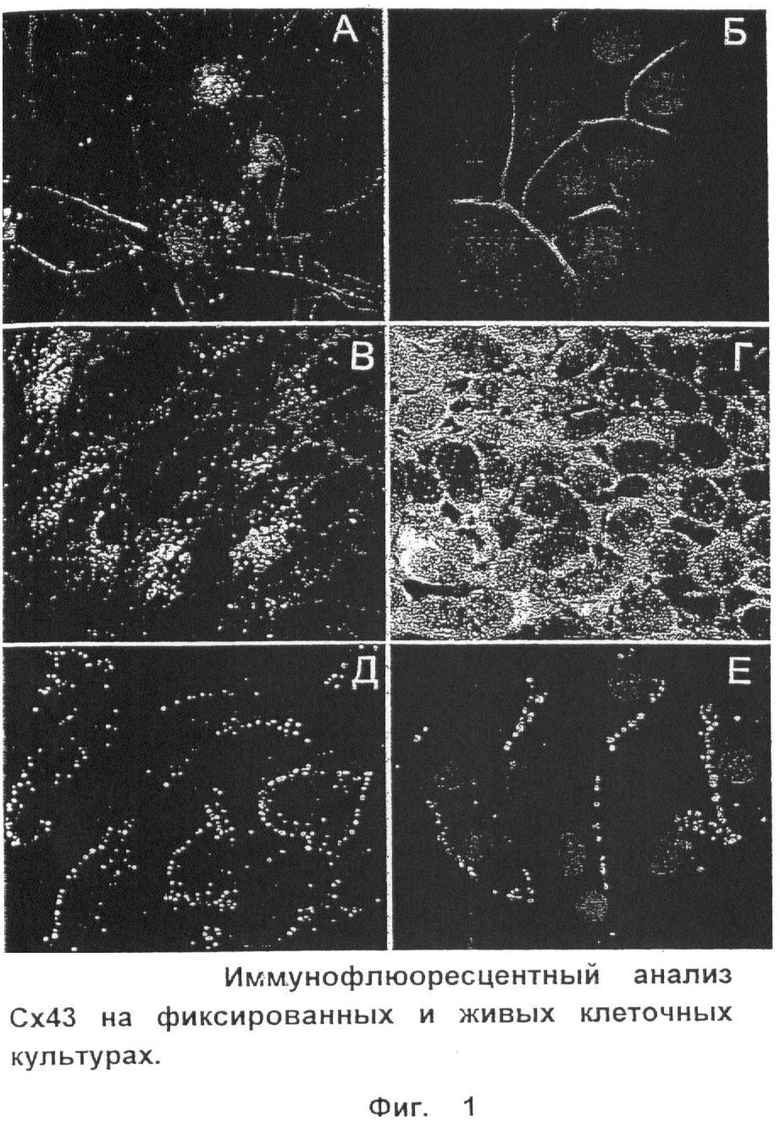
При иммунофлюоресцентном анализе полученные моноклональные антитела визуализируют плакоидные структуры коннексонов на фиксированных первичных астроцитах и клетках глиомы С6 крысы, равно как и на клетках глиобластомы U251 и 293-й линий эмбрионального почечного эпителия человека (Фиг.1, А-Е).
А - культура астроцитов из мозга новорожденных крысят: красная флюоресценция - поликлональные антитела к GFAP; зеленая флюоресценция - MAb Сх43; ядра клеток-DAPI (Invitrogen). Б - глиобластома человека (линия U251): синяя флюоресценция - МАb Сх43 + антимышиные IgG с Alexa Fluor 350 (Invitrogen); оранжево-красная флюоресценция - актиновые филаменты, окрашенные Phalloidin TRITC (Fluka); ядра клеток - ТОТО 633 (Invitrogen). В - глиома С6 крысы: красное - β-Катенин; зеленое - MAb Сх43. Г - эмбриональный почечный эпителий (линия 293): красное - пан-Кадгерины; зеленое - MAb Сх43. Д - визуализация Сх43 в живой культуре глиобластомы U251 (зеленая флюоресценция); ядра клеток докрашены DAPI. E - визуализация Сх43 в живой культуре глиомы С6 (зеленая флюоресценция); ядра клеток докрашены ТОТО 633. Увеличение ×1000.
Наиболее яркая флюоресценция наблюдается в препаратах эмбрионального почечного эпителия, что соответствует представлениям о высоком уровне экспрессии белков gap-junction в эмбриогенезе (Фиг.1, Г). У фиксированных клеточных препаратов визуализируется как мембранный, так и цитоплазматический пул Сх43.
Очищенные и биотинилированные моноклональные антитела к экстраклеточному домену Е2 хорошо визуализируют Сх43-положительные клетки низкодифференцированных глиом в живой культуре (Фиг.1, Д, E). В этом случае наблюдается флюоресценция только мембранных коннексонов. Ни в одном из препаратов не наблюдалось флюоресценции межклеточных соединений, характерной для димерных коннексонов, интегрированных в gap-junction. Это позволяет сделать вывод о том, что полученные антитела визуализируют Сх43 на этапе презентирования коннексонов на цитолемме в виде гемиканала.
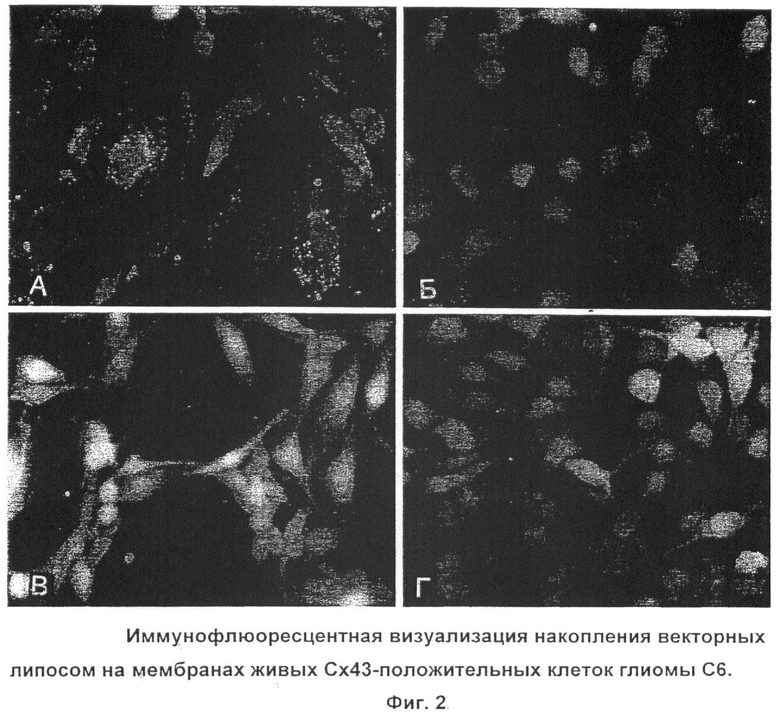
На фиг.2: А - биотинилированные антитела + липосомы со стрептавидином; Б - конкурентное ингибирование связывания биотинилированных антител с клетками-мишенями путем прединкубации с избытком небиотинилированных антител к Е2 Сх43; В - биотинилированные неспецифические иммуноглобулины мыши + липосомы со стрептавидином; Г - биотинилированные моноклональные антитела к Е2 Сх43 + липосомы без стрептавидина.
В эксперименте с бинарной системой направленного транспорта специфическая флюоресценция Dil наблюдается только в препаратах глиомных клеток последовательно инкубированных с биотинилированными антителами к Е2 Сх43 и липосомами со стрептавидином и флюоресцентной меткой. Уже через 30 минут после добавления минимального объема липосомальной эмульсии (1 µl) начинали визуализироваться характерные мембранные плакоиды коннексонов (Фиг.2, А). Этот феномен наблюдается во всем диапазоне концентраций (1, 10, 25, 50 µl/ml). Феномен флюоресценции Сх43-положительных клеток С6 глиомы был воспроизведен в трех независимых сериях экспериментов.
Прединкубация клеток с избытком небиотинилированных антител к Сх43 полностью блокирует появление флюоресценции после добавления биотинилированных антител и липосом со стрептавидином (Фиг.2, Б). В контрольных лунках с неспецифическими биотинилированными антителами и векторными липосомами (Фиг.2, В), также как при применение невекторных липосом (Фиг.2, Г), специфической флюоресценции не наблюдалось ни в одной из трех серий эксперимента.
Интенсивная красная флюоресценция Сх43-положительных клеток глиомы С6 свидетельствует о селективной адгезии векторных липосом со стрептавидином и Dil на мембранах этих клеток за счет связывания с биотинилированными антителами к Сх43, взаимодействующими с экстраклеточной петлей этого белка.
Разработанная бинарная система на основе биотинилированных антител к Сх43 и липосом со стрептавидином показала высокую селективность по отношению к Сх43-положительным клеткам глиомы. Адгезия векторных липосом на мембранах клеток может способствовать слиянию билипидных слоев и интернализации содержимого липосом. Для интенсификации процесса интернализации в состав векторных липосом могут быть введены дополнительные компоненты (в частности ТАТ пептид или другие вирусные транскрипционные факторы).
Бинарная система на основе биотинилированных антител к экстраклеточному фрагменту Сх43и ПЭГилированных липосом со стрептавидином может применяться для направленного транспорта диагностических и терапевтических агентов к Сх43-положительным клеткам глиомы in vivo.

| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОГЕННОГО РЕКОМБИНАНТНОГО ЭКСТРАКЛЕТОЧНОГО ФРАГМЕНТА КОННЕКСИНА-43 | 2009 |
|
RU2408728C1 |
| СПОСОБ ЛЕЧЕНИЯ НИЗКОДИФФЕРЕНЦИРОВАННЫХ ГЛИОМ | 2011 |
|
RU2457862C1 |
| СПОСОБ ПОЛУЧЕНИЯ СРЕДСТВА ВНУТРИКЛЕТОЧНОЙ ДОСТАВКИ БИОЛОГИЧЕСКИ АКТИВНЫХ НИЗКОМОЛЕКУЛЯРНЫХ СОЕДИНЕНИЙ, НА ОСНОВЕ НАНОЧАСТИЦ | 2014 |
|
RU2557987C1 |
| СРЕДСТВО ВНУТРИКЛЕТОЧНОЙ ДОСТАВКИ БИОЛОГИЧЕСКИ АКТИВНОГО ВЫСОКОМОЛЕКУЛЯРНОГО СОЕДИНЕНИЯ НА ОСНОВЕ НАНОЧАСТИЦ И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2014 |
|
RU2541121C1 |
| ПРОТИВООПУХОЛЕВОЕ СРЕДСТВО НА ОСНОВЕ ИММУНОЛИПОСОМАЛЬНОЙ БИОЛОГИЧЕСКОЙ КОНСТРУКЦИИ, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ВЕКТОРНОЙ ДОСТАВКИ В ЦЕНТРАЛЬНУЮ НЕРВНУЮ СИСТЕМУ ПРИ ОПУХОЛЕВОМ ПРОЦЕССЕ | 2007 |
|
RU2336901C1 |
| УЛУЧШЕННЫЕ СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЗАЖИВЛЕНИЯ РАН | 2007 |
|
RU2542471C2 |
| ИММУНОЛИПОСОМА, СВЯЗЫВАЮЩАЯСЯ С RBD S-БЕЛКА SARS-COV-2 | 2023 |
|
RU2827165C1 |
| Рекомбинантный химерный белок, состоящий из большой экстраклеточной петли человеческого рецептора CD81 и стрептавидина | 2020 |
|
RU2778251C2 |
| УСОВЕРШЕНСТВОВАННЫЕ ЛИПОСОМЫ И ИХ ПРИМЕНЕНИЕ | 2008 |
|
RU2482837C2 |
| ИММУНОКОНЪЮГАТ, СПОСОБ ДОСТАВКИ ПОЛИНУКЛЕОТИДОВ В СПЕЦИФИЧЕСКИЙ ТИП КЛЕТОК, СПОСОБ ДОСТАВКИ ЦИТОТОКСИЧЕСКОГО ГЕЛОНИНА В КЛЕТКУ | 1995 |
|
RU2164943C2 |
Изобретение относится к области молекулярной биологии. Предложена бинарная система адресной доставки липосом к коннексин-43 (Сх-43) положительным опухолевым клеткам. Система включает биотинилированные моноклональные антитела к экстраклеточному фрагменту коннексина-43 (первый компонент) и ПЭГилированные липосомальные контейнеры диаметром 70-100 нм, ковалентно связанные со стрептавидином (второй компонент). Последовательное введение в живую систему первого и второго компонентов бинарной системы приводит к селективной доставке липосом к Сх-43-положительным клеткам путем специфического связывания биотинилированных антител с экстраклеточным фрагментом Сх-43 на цитолемме глиомных клеток и последующим образованием стрептавидин-биотинового комплекса. Система может найти применение в разработке новых противоопухолевых препаратов направленного типа действия. 2 ил.
Бинарная иммунолипосомальная наносистема адресной доставки к коннексин-43 положительным опухолевым клеткам, содержащая биотинилированные моноклональные антитела к экстраклеточному фрагменту коннексина-43 и ковалентно связанные со стрептавидином наноразмерные ПЭГилированные липосомальные контейнеры диаметром 70-100 нм.
| US 4948590 А, 14.08.1990 | |||
| PAN Н, ет al.Targeting to tumor necrotic regions with biotinylated antibody and streptavidin modified liposomes.J Control Release | |||
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| Пресс для выдавливания из деревянных дисков заготовок для ниточных катушек | 1923 |
|
SU2007A1 |
| CHIH-JEN WEI ет al | |||
| Зубчатое колесо со сменным зубчатым ободом | 1922 |
|
SU43A1 |
| Прибор для промывания газов | 1922 |
|
SU20A1 |

