Область техники, к которой относится изобретение
Настоящее изобретение относится к области белковой инженерии, биотехнологии и медицины, и касается получения иммунолипосомы, связывающейся с RBD S-белка SARS-CoV-2 и обладающей способностью нейтрализовать вирус SARS-CoV-2 в псевдовирусной системе. Изобретение может найти применение в биотехнологии и медицине при создании лекарств против КОВИД-19.
Уровень техники
В 2020 г человечество пережило пандемию короновирусной инфекции SARS-CoV-2. Однако говорить о том, что инфекция побеждена не приходится: в популяции по-прежнему происходит возникновение новых более агрессивных штаммов короновируса, приводящих не только к инфицированию, но и к летальным исходам. Так, по состоянию на сентябрь 2023 года коронавирус SARS-CoV-2 инфицировал более 770 млн человек, число погибших от COVID-19 во всем мире превысило 6.9 млн человек [1].
SARS-CoV-2 является членом семейства Coronavirinae и относится к роду бета-коронавирусов, который включает также MERS-CoV и SARS-CoV. Геном вируса SARS-CoV-2 на 75-80% гомологичен геному SARS-CoV-2 [2-4] Вирусные частицы имеют сферическую или плеоморфную форму диаметром около- 60-140 нм.
Бета-коронавирусы имеют три важных белка оболочки: белок шипа (S), мембранный (М) белок и белок оболочки (Е). Белок S опосредует прикрепление вируса к рецептору клеточной мембраныи проникновение вируса в клетку-хозяина. Белки М и Е отвечают за структуру мембраны коронавируса. Еще одним компонентом бета-коронавируса является белок N, который представляет собой белковый компонент спиралевидного нуклеокапсида, включающего геномную РНК [4].
Установлено, что SARS-CoV-2 использует рецептор ангиотензин-превращающего фермента 2 (ACE-2) для проникновения в клетки [5]. После проникновения в клетку геном вирусной +РНК высвобождается в цитоплазму и подвергается трансляции и репликации, в результате чего образуются новые +РНК и субгеномные мРНК. После синтеза вирусных белков происходит упаковка вириона и его высвобождение экзоцитозом [4, 6].
Чувствительность генома РНК-содержащих вирусов (к которым относится SARS-CoV-2) определяет привлекательность использование в качестве терапевтических агентов микробных рибонуклеаз. Наиболее известными микробными РНКазами являются барназа, продуцируемая бактериями вида Bacillus amyloliquefaciens, и биназа - бактериями вида Bacillus pumilis. В силу высокой стабильности и отсутствии восприимчивости к ингибиторам рибонуклеаз эукариот, оба фермента проявляют высокую каталитическую активность к молекулам РНК в любых клетках эукариот [7, 8]
Известен способ подавления размножения РНК-содержащих вирусов при помощи микробных РНКаз. Биназа проявила высокую активность против РНК-содержащего вируса бешенства [9], и вируса ящура О типа [10], вирусов гриппа типов А и В [11], а также против пандемического вируса гриппа A/Hamburg/04/09 (H1N1).
Известен способ подавления биназой вирусной инфекции MERS-CoV и HCoV-229E, вирусов близкородственных SARS-CoV-2. В нетоксичных для клеток концентрациях биназа снижала существенно титры этих вирусов [12]. К недостаткам этого способа следует отнести очень высокие действующие концентрации РНКазы.
Известен способ подавления вирусной инфекции SARS-CoV-2 нейтрализующими моноклональными антителами [13-16]. К настоящему времени одобрены для использования и продолжают проходить клинические испытания большое количество препаратов на основе нейтрализующих моноклональных антител (nAb), действие которых обусловлено связыванием с эпитопами рецептор-связывающего домена S-белка SARS-CoV-2, блокированием его взаимодействия с ангиотензин-превращающим ферментом 2 (ACE-2) и подавлением, таким образом, инфицирования клеток хозяина. Рекомендуемые для терапии COVID-19 препараты содержат значительные дозы рекомбинантных антител (400-2400 мг при однократном введении) [17], что может провоцировать нежелательные явления в некоторых случаях. Снижение дозы терапевтических антител, таким образом, было бы оправдано как с точки зрения биобезопасности, так и с экономической точки зрения. В этой связи особое значение приобретает подход, связанный с адресной доставкой препарата в зараженную вирусом клетку. Одним из подходов, позволяющих снизить концентрации терапевтического антитела, является применение конъюгатов антител с цитотоксическим агентом. В частности, в качестве цитотоксического агента используются липосомы, содержащие во внутреннем объеме токсины различной природы (органической и неорганической) - иммунолипосомы. Для этих целей липосомы получают таким образом, чтобы они содержали необходимый объект. Процесс включения желаемого объекта в липосому обычно обозначают термином «нагрузка». Включение объектов в липосомы также обозначают термином инкапсулирование, и оба указанных термина используются в настоящем описании взаимозаменяемо и имеют одинаковое значение. При инкапсулировании вещества в липосому обычно преследуется цель защитить указанное вещество от ферментативной деградации и быстрого клиренса in vivo, снизить иммуногенность включенных в липосому терапевтических агентов, снизить общую токсическую нагрузку от инкапсулированного токсина на организм. Направляющие молекулы, коньюгированные с нагруженными липосомами, обеспечивают доставку - возможность проявления активности инкапсулированным токсином в том месте, где данная активность наиболее предпочтительна, и избежать ее реализации там, где это бесполезно или нежелательно. Липосомы, содержащие на своей поверхности направляющие молекулы, обычно обозначаются термином «таргетированные». Данный подход был успешно реализован в ряде работ [18,19], демонстрирующих улучшение терапевтического индекса препарата.
Целью данного изобретения служила разработка способа получения иммунолипосомы, связывающейся с RBD S-белка SARS-CoV-2 и обладающей усиленной нейтрализующей активностью, обусловленной синергичным эффектом, вследствие наличия в ее составе как RBD-узнающего нейтрализующего антитела P4A1-2A, блокирующего проникновение в клетки вирусных частиц, так и инкапсулированной рибонуклеазы барназы, осуществляющей деградацию клеточной и вирусной РНК.
Прототипом данного изобретения может служить работа [16] которая описывает получение рекомбинантного нейтрализующего антитела P4A1-2A. Данное антитело специфично и эффективно (KD=1.02*10-10М) связывало рецептор-связывающий домен S-белка SARS-CoV-2 (RBD), и блокировало SARS-CoV-2 инфекцию клеток HEK293, экспрессирующих ACE2, со значением ингибирующей концентрации (IC50) 2 нМ.
Технической проблемой, решаемой настоящим изобретением является увеличение нейтрализующей активности антитела P4A1-2A.
Для технического решения проблемы увеличения нейтрализующей активности антитела P4A1-2A была избрана стратегия ковалентного присоединения антитела P4A1-2A к липосоме, содержащей во внутреннем объеме рибонуклеазу барназу.
В процессе поэтапного выполнения патентных исследований осуществлялись:
- культивирование клеток рекомбинантного штамма Escherichia coli BL21(DE3)/pET-STII-Ba с последующим получением целевого рекомбинантного белка барназы;
- культивирование клеток линии CHO K1 (ATCC, No. CCL 61), трансфецированных плазмидной ДНК, содержащей последовательности легкой и тяжелой цепи антитела P4A1-2A под контролем CMV промоторов [16], с последующим получением целевого рекомбинантного белка антитела P4A1-2A;
- получение препарата липосом - содержащих и не содержащих во внутреннем объеме рекомбинантный белок рибонуклеазу барназу;
- ковалентная модификация внешней поверхности пустых и нагруженных барназой липосом рекомбинантным антителом P4A1-2A, а также в качестве контроля ковалентная модификация нагруженных липосом полипептидом G3, полученного в бактериальной системе в результате экспрессии гена, кодирующего этот полипептид.
- исследование вируснейтрализующей активности полученных иммунолипосом в псевдовирусной системе, моделирующей проникновение вирусных частиц SARS-CoV-2 в клетки.
Настоящее изобретение решает техническую проблему получения иммунолипосомы, связывающейся с RBD S-белка SARS-CoV-2, способной блокировать связывание с клетками, имеющими на поверхности рецептор ACE-2 и/или препятствовать размножению вируса SARS-CoV-2 и имеющую увеличенную нейтрализующую активность в сравнении с антителом P4A1-2A [16], на основе липосомы:
- ковалентно модифицированной рекомбинантным антителом P4A1-2A, имеющим последовательности тяжелой SEQ ID NO:2 и легкой SEQ ID NO:3 цепей,
- содержащией во внутреннем пространстве рекомбинантный белок рибонуклеазу Bacillus amyloliquefaciens, имеющую последовательность SEQ ID NO: 1.
Заявленная техническая проблема решается за счет:
1) Иммунолипосомы, связывающейся с RBD S-белка SARS-CoV-2, представляющей собой иммунолипосомальную биологическую конструкцию, включающую липосому, содержащую во внутреннем объеме цитотоксический агент рибонуклеазу барназу, имеющую последовательность SEQ ID NO: 1, а внешняя поверхность которой ковалентно модифицирована моноклональным антителом P4A1-2A, имеющим последовательности тяжелой SEQ ID NO:2 и легкой SEQ ID NO:3 цепей и обладающим сродством к рецептор-связывающему домену S-белка SARS-CoV-2, характеризующейся увеличенной вируснейтрализующей активностью по сравнению с нейтрализующим антителом P4A1-2A.
2) Способа получения из биомассы рекомбинантного штамма Escherichia coli BL21(DE3)/pET-STII-Ba рекомбинантного белка барназы.
3) Способа получения из культуральной жидкости после культивирования клеток линии CHO K1 (ATCC, No. CCL 61), трансфецированных плазмидной ДНК, содержащей последовательности легкой и тяжелой цепи антитела P4A1-2A под контролем CMV промоторов [16], целевого рекомбинантного белка антитела P4A1-2A.
4) Способа получения иммунолипосомы, связывающейся с RBD S-белка SARS-CoV-2, включающего в себя формирование липосомальных частиц посредством пропускания липидно-белковой суспензии через экструдер с использованием поликарбонатных фильтров с размером пор 100 нм с одновременным инкапсулированием во внутренний объем рекомбинантного белка барназы и последующую ковалентную модификацию внешней поверхности липосомы рекомбинантным антителом P4A1-2A методом клик-химии с использованием гидрофильного бифункционального кросс-сшивающего агента (Sulfo-EMCS, N-ε-maleimidocaproyl-oxysulfosuccinimide ester) и 2-иминотиолана.
В результате решения заявленной технической проблемы получают следующий технический результат:
1) вируснейтрализующую активность иммунолипосомы, связывающейся с RBD S-белка SARS-CoV-2;
2) вируснейтрализующую активность иммунолипосомы, связывающейся с RBD S-белка SARS-CoV-2, более чем в 40 раз превышающую вируснейтрализующую активность антитела P4A1-2A в псевдовирусной системе.
Раскрытие изобретения
Получена иммунолипосома, связывающаяся с RBD S-белка SARS-CoV-2, представляющая собой липосому, содержащую во внутреннем объеме цитотоксический агент, а внешняя поверхность которой ковалентно модифицирована направляющими молекулами белковой природы, обеспечивающими таргетную доставку иммунолипосомы. Для проявления вируснейтрализующей активности, выражающейся в деградации клеточной и вирусной РНК, иммунолипосома содержит цитотоксический агент рибонуклеазу барназу, имеющую последовательность SEQ ID NO: 1. Для проявления вируснейтрализующей активности, выражающейся в блокировании проникновения в клетки вирусных частиц, иммунолипосома в качестве направляющих молекул содержит моноклональное рекомбинантное антитело P4A1-2A, имеющего последовательность SEQ ID NO:2 и SEQ ID NO: 3 и обладающего сродством к рецептор-связывающему домену S-белка SARS-CoV-2. Иммунолипосома обладает способностью специфически связываться с лентивирусами, псевдотипированными S-белком SARS-CoV-2, и блокировать связывание лентивирусов, псевдотипированных S-белком SARS-CoV-2, с рецептором ACE-2, представленным на поверхности клеток, чувствительных к заражению вирусом. При интернализации в клетки вместе с вирусными частицами иммунолипосома обладает вирус-статическим действием за счет гидролиза вирусной РНК и последующего прекращения биосинтеза белка, необходимого для синтеза новых вирионов.
Показано применение иммунолипосомы, связывающейся с RBD S-белка SARS-CoV-2, в качестве вируснейтрализующего средства в отношении SARS-CoV-2 в псевдовирусной системе.
Показано более чем 40 раз увеличение вируснейтрализующей активности иммунолипосомы, связывающейся с RBD S-белка SARS-CoV-2, по сравнению с антителом P4A1-2A с использованием псевдовирусной системы.
Таким образом, настоящее изобретение включает 2 объекта:
Первый объект - иммунолипосома, связывающаяся с RBD S-белка SARS-CoV-2,
Второй объект - способ получения иммунолипосомы, связывающейся с RBD S-белка SARS-CoV-2.
Изобретение иллюстрируется следующими фигурами:
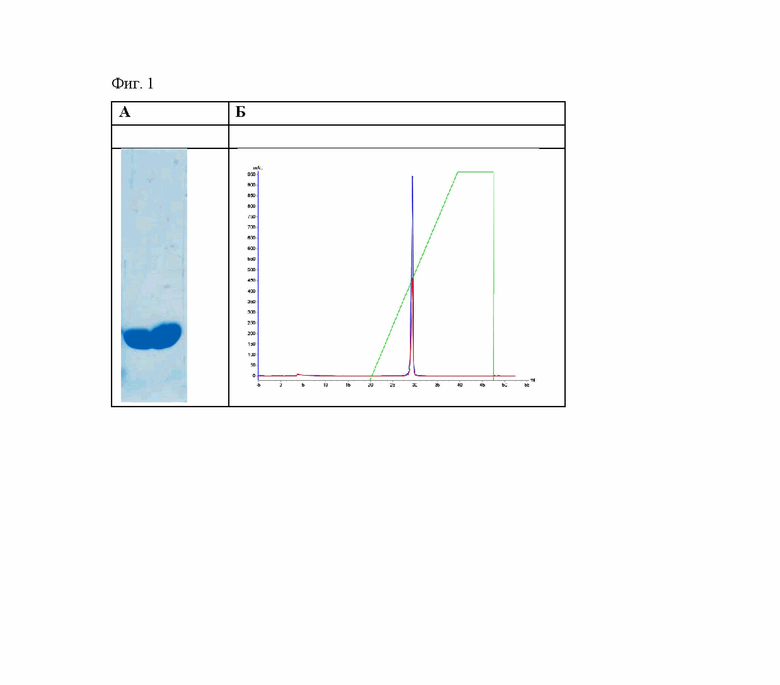
Фиг. 1 - Хроматография на MonoS 10/100 GL.
А. Электрофоретический анализ очищенной барназы;
Б. Профиль элюции.
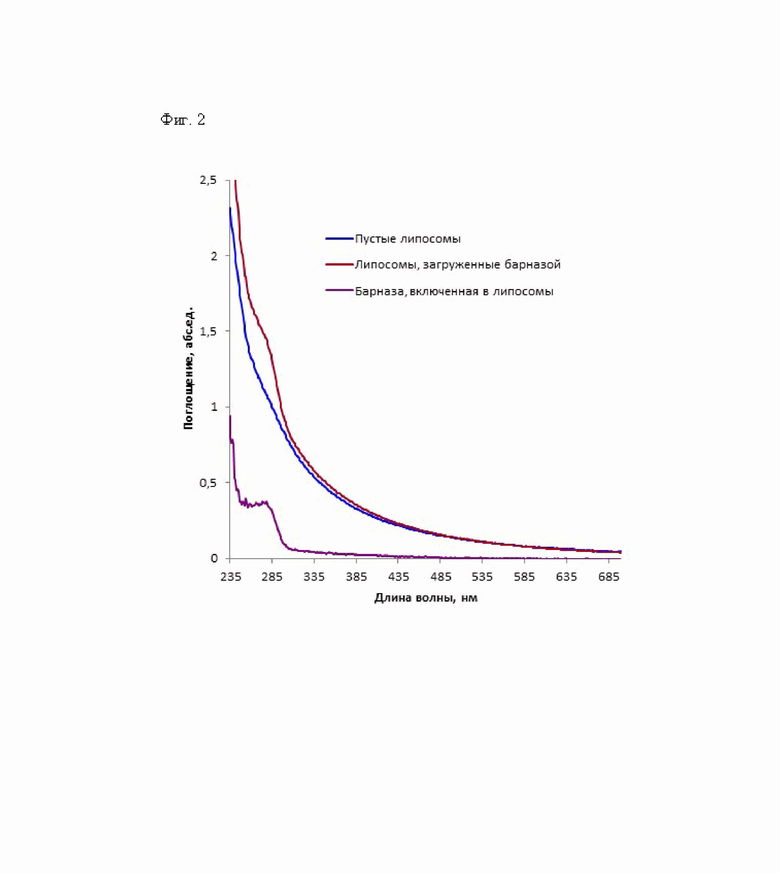
Фиг. 2 - Спектры поглощения липосом. Красная и синяя кривые - спектры поглощения липосом, загруженных барназой, и пустых липосом, соответственно. Сиреневая кривая - спектр поглощения барназы, инкапсулированной в липосомы, который получен вычитанием спектра поглощения пустых липосом из спектра поглощения протеолипосом.
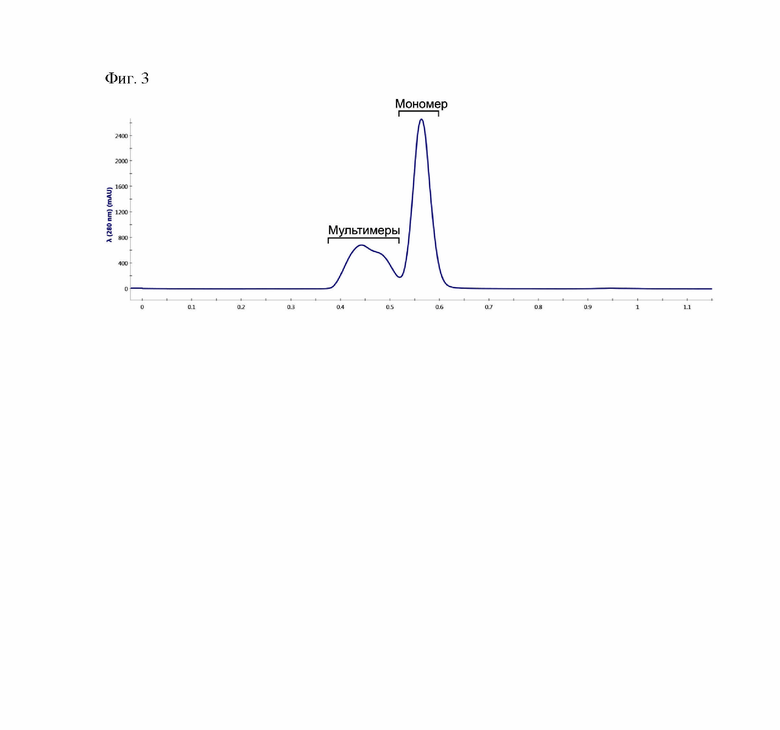
Фиг. 3 - Хроматографический профиль антитела P4A1-2А на колонке Superdex 200.
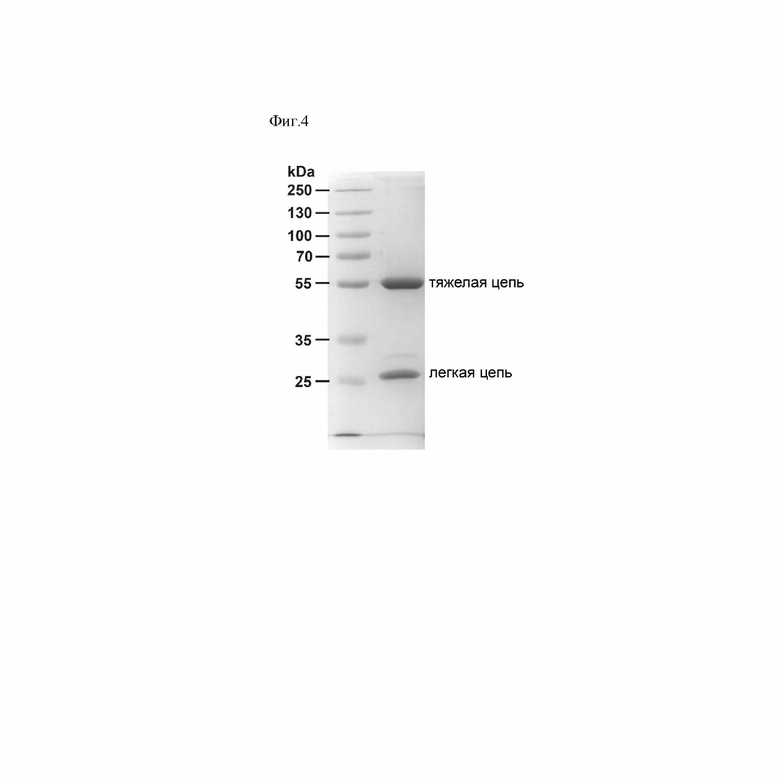
Фиг. 4 - Электрофоретический анализ очищенного препарата антитела P4A1-2А в восстанавливающих условиях.
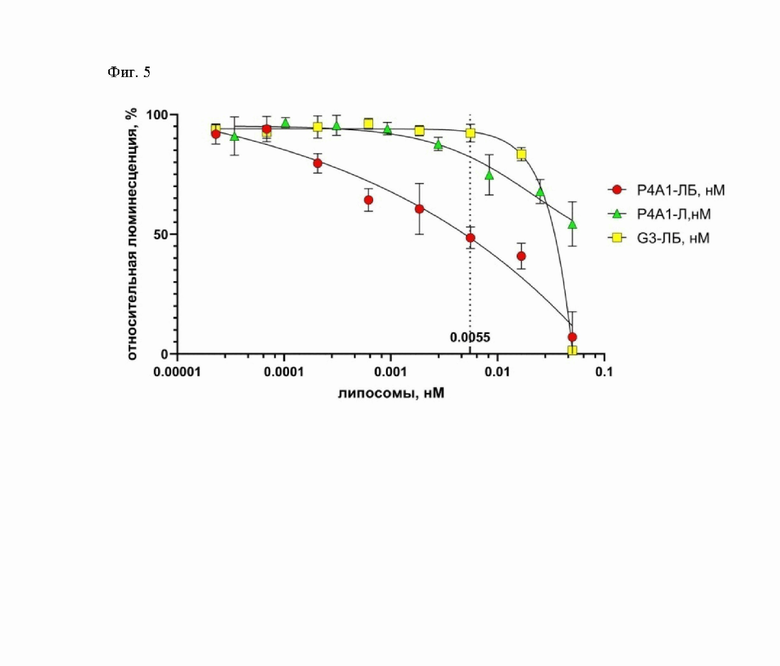
Фиг. 5 - Зависимость интенсивности люминисценции клеток линии HEK293T-ACE-2, экспрессирующих на поверхность рецептор ACE-2 человека, от концентрации липосомальных препаратов в псевдовирусной системе.
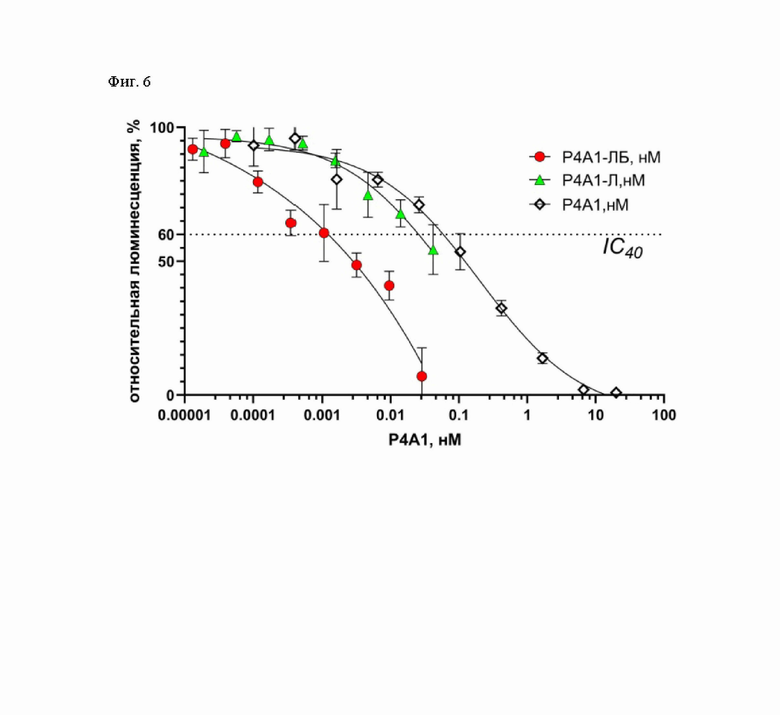
Фиг. 6 - Зависимость интенсивности люминисценции клеток линии HEK293T-ACE-2, экспрессирующих на поверхности рецептор ACE-2 человека, от концентрации антитела Р4А1-2A в составе липосомальных препаратов и буферного раствора в псевдовирусной системе.
Осуществление изобретения.
Пример 1. Получение рекомбинантного белка барназы.
Получение рекомбинантного белка барназы осуществляли как описано [20]. Для этого клетки штамма Escherichia coli BL21 (DE3)/pET-STII-Ba, полученных трансформацией клеток штамма E. coli BL21(DE3) [F-, ompT, hsdSB (rB-, mB-), dcm, gal, λ(DE3)] методом [21] плазмидной ДНК pET-STII-Ba, из глицеринового стока, хранящегося в морозильнике, рассевали на поверхности чашки с питательной агаризованной средой YT (8 г бакто-триптона, 5 г дрожжевого экстракта, 5 г NaCl на 1 л воды) и канамицином (25 мг/л). Инкубировали ночь при 37°С. Для получения посевного материала собирали стеклянной палочкой около 50 колоний с чашек Петри, засеянных трансформантами, и суспендировали в 200 мкл среды YT. Выращивали посевной материал на качалке при 250 об/мин и 37°С в течение нескольких часов до достижения оптической плотности прим. 0.5 о.е. при 260 нм. Посевной материал переносили в колбу объемом 2 л, содержащей 200 мл жидкой питательной среды ZYM-5052 [22] с 50 мг/л канамицина, и культивировали при температуре 25°С и 250 об/мин в течение суток. Биомассу осаждали центрифугированием при 6000 g в течение 10 мин. Суспендировали биомассу, собранную с 100 мл культуры, в 20 мл 200 мМ Трис-HCl, 0.5 М сахароза, 1 мМ ЕДТА (рН 8.0). Добавляли лизоцим до конечной концентрации 60 мкг/мл. Разбавляли суспензию в два раза водой и инкубировали при комнатной температуре в течение 30 мин. Центрифугировали при 6000 g, 4°С, в течение 20 мин. Супернатант разбавляли в 5 раз 20 мМ NaOAc, рН 4.5 и подтитровывали до рН 5.0 ледяной уксусной кислотой. Образующийся преципитат осаждали центрифугированием при 6000 g, 4°С, в течение 20 мин. Супернатант фильтровали через фильтр Millex-GP 0.22 μm (33 мм) (MILLIPORE, США) и наносили на колонку XK16/20, набитую фосфоцеллюлозой (Sigma, США) и уравновешенную 20 мМ NaOAc, рН 4.5. Сорбент последовательно промывали 20 мМ NaOAc, рН 4.5, затем 20 мМ NaPi, рН 7.0 до достижения стабильных значений базовой линии (примерно 10 объемов колонки). Белок элюировали 50 мМ NaPi, 1.7 М (NH4)Cl, рН 7.0; собирали все светопоглощающие фракции.
Элюат наносили на колонку HiPrep 26/10 Desalting Column (GE Healthcare, США), уравновешенную 20 мМ NaOAc, рН 4.5, и собирали все светопоглощающие фракции вплоть до выхода с колонки соли.
Объединенную фракцию наносили на колонку MonoS 10/100 GL (GE Healthcare, США), уравновешенную 20 мМ NaOAc, рН 5.0. Колонку промывали тем же буфером. Белок элюировали в линейном градиенте NaCl (Фиг. 1). Собранные фракции анализировали при помощи денатурирующего гель-электрофореза по Лэмли (Фиг. 1). Фракции, содержащие барназу, объединяли и концентрировали при помощи Amicon Ultra-15 (c MWCO 5000) (Merck Millipore, США)
Пример 2. Определение ферментативной (РНКазной) активности барназы.
Для определения ферментативной активности барназы использовали модифицированный метод кислоторастворимого осадка. В качестве субстрата использовали дрожжевую РНК.
Исследуемую пробу белка растворяли в концентрации 1,25 мкМ в буферном растворе 0.125 М Трис-HCl, pH 8.5 и затем получали серию последовательных 5-кратных разведений образца в том же буфере. В качестве отрицательного контрольного образца использовали буферный раствор 0.125 М Трис-HCl, pH 8.5. В качестве положительного контрольного образца использовали стандартный образец рекомбинантной барназы. К 40 мкл каждого образца добавляли 160 мкл раствора дрожжевой РНК (в концентрации 2 г/л) в 0.125 М Трис-HCl, pH 8.5 на льду. Реакционную смесь инкубировали 30 минут при +37°С. Реакцию останавливали добавлением 200 мкл 6% хлорной кислоты и инкубировали смесь 15 мин при 0°С. После чего центрифугировали при 16000 g в течение 10 минут. Супернатанты разбавляли в 10 раз и измеряли оптическую плотность при 260 нм (OD260) относительно контрольного образца. Бóльшим значениям оптической плотности соответствует бóльшая РНКазная активность. 1 ед. акт. РНКазы в исследуемой пробе соответствует приращению OD260 на 0.05 единиц.
Пример 3. Загрузка барназы в липосомы.
Загрузку барназы в липосомы осуществляли на основе электростатического взаимодействия между положительно заряженным белком (при нейтральных значениях рН) и отрицательно заряженной внутренней поверхностью липосом. Липосомы формировали смешивая 15 мкМ раствора барназы с суспензией фосфолипидов (Soy PC, 40%, Avanti, США), содержащих 20% фосфатидилэтаноламина, до конечной концентрации фосфолипидов в смеси 2 г/л, с последующим 7-кратным замораживанием/оттаиванием и дальнейшем продавливание через эксрудер с использованием поликарбонатных фильтров с диаметром пор 100 нм. Инкубация суспензии фосфолипидов с небольшой, гидрофильной и положительно заряженной (при нейтральных значениях рН) барназой, при низкой ионной силе в результате продавливания через поликарбонатный фильтр с размером пор 100 нм, приводит к образованию липосом с диаметром около 90-100 нм. Загрузку барназы в липосомы оценивали спектрофотометрически. Для этого из спектра поглощения протеолипосом вычитали спектр пустых липосом (Фиг. 2). Пустые липосомы, т.е. липосомы, внутренняя среда которых не содержит белка, получали продавливанием суспензии фосфолипидов (Soy PC, 40%, Avanti, США), содержащих 20% фосфатидилэтаноламина, через экструдер с использованием поликарбонатных фильтров с размером пор 100 нм. Нагруженные барназой липосомы отделяли от свободной барназы с помощью гель-проникающей хроматографии на сорбенте сефароза CL-2B (Cytiva) в 20 мМ натрий-фосфатном буфере, рН 7.5 Концентрацию пустых липосом оценивали спектрофотометрически с использованием гидрофильного мембранонепроницаемого красителя фталоцианин-3,4',4',4''-тетрасульфоната меди [23]. Концентрацию белка рассчитывали, используя молярный коэффициент экстинции ε280 = 26930 М-1см-1. Концентрация барназы в липосомах составила 13.3 мкМ. Концентрацию липосом, загруженных барназой, оценивали спектрофотометрически, сравнивая спектр пустых и протеолипосом. Поскольку спектр загруженных барназой липосом совпадает со спектром пустых липосом с концентрацией 1.1 мг/мл, концентрация загруженных белком липосом составлла 1.3 нМ.
Пример 4. Получение рекомбинантного антитела Р4А1
Получение рекомбинантного нейтрализующего антитела P4A1-2A осуществляли как описано [16]. Для этого клетки линии CHO K1 (ATCC, No. CCL 61), трансфецированные плазмидной ДНК, содержащей последовательности легкой и тяжелой цепи антитела P4A1-2A под контролем CMV промоторов культивировали в колбах Эрленмейера объемом 500 мл, содержащих 120 мл среды ExpiCHO cell (Gibco, No. A29133), при постоянном перемешивании со скоростью 125 оборотов в минуту, в атмосфере, содержащей 8% CO2. По завершении 7-дневного периода культивирования клетки осаждали последовательным центрифугированием при 200, 1000 и 1300g в течение 10 мин. Супернатант с секретированными антителами P4A1-2А наносили на колонку HiTrap Protein G объемом 1 мл, набитую Protein G (Sigma, США) и уравновешенную 20 мМ натрий-фосфатный буфер рН 7.5, 150 мМ NaCl. Сорбент последовательно промывали 20 мМ натрий-фосфатный буфер рН 7.5, 150 мМ NaCl до достижения стабильных значений базовой линии (примерно 10 объемов колонки). Белок элюировали 100 мМ глицин-HCl, рН 2.7; собирали все светопоглощающие фракции и доводили рН в объединенных фракциях до значения 7.5 с помощью 1М Трис. Далее очищенное антитело подвергали дополнительной очистке с помощью гель-проникающей хроматографии для удаления агрегатов и денатурированных форм. Объединённые фракции наносили на колонку Superdex 200 (Sigma, США) и уравновешенную 20 мМ натрий-фосфатный буфер рН 7.5, 150 мМ NaCl; собирали все светопоглощающие фракции, соответствующие мономерной форме антитела (Фиг. 3).
После очистки проводили анализ концентрации и чистоты антитела P4A1-2А с использованием спектрофотометрии, денатурирующего гель-электрофореза по Лэмли (Фиг. 4) и ВЭЖХ. Концентрацию белка оценивали спектрофотометрически, используя коэффициент экстинкции антитела E0.1%=13.7
Пример 5. Получение коньюгированных иммунолипосом
Модификацию внешней поверхности липосом, содержащих инкапсулированную барназу, осуществляли по аминогруппам фосфотидилэтаноламина с использованием 2-иминотолана (реагент Траута, позволяющий ввести SH-группу по первичным аминам фосфолипида, финальная концентрация в реакционной смеси 6 мМ) как описано в [23]. Тиолированные липосомы, содержащие инкапсулированную барназу, получали модификацией внешней поверхности липосом, содержащих инкапсулированную барназу, 2-иминотиоланом (конечная концентрация в смеси 6 мМ) в течение 40 мин при комнатной температуре с дальнейшим удалением избытка непрореагировавшего 2-иминотиолана на сорбенте сефадекс G-25, уравновешенном буфером 20 мМ натрий-фосфат, рН 7.5. Рекомбинантное антитело P4A1-2A или продуцированный в бактериальной системе полипептид G3, который использовали в качестве отрицательного контроля, смешивали со 100-кратным молярным избытком sulfo-EMCS (N-ε-maleimidocaproyl-oxysulfosuccinimideester, билинкер, позволяющий ввести в белок через оксисульфосукцинимидмалеимидную группу), инкубировали 40 мин при комнатной температуре с дальнейшим удалением избытка непрореагировавшего Sulfo-EMCS на сорбенте сефадекс G-25, уравновешенном буфером 20 мМ натрий-фосфат, рН 7.5 Тиолированные липосомы, содержащие барназу, смешивали с антителом P4A1-2A или полипептидом G3, содержащих малеимидные группы, инкубировали 1 ч при комнатной температуре с получением таргетированных липосом P4A1-ЛБ и G3-ЛБ соответственно. Адресные липосомы P4A1-ЛБ и G3-ЛБ отделяли от непрореагировавших с липосомами белковых с использованием гель-проникающей хроматографии на сорбенте сефароза CL-2B, уравновешенном 20 мМ натрий-фосфатным буфером. Таргетированные липосомы P4A1-ЛБ и G3-ЛБ при этом сходят с колонки в исключенном объеме, в то время как P4A1-2A и полипептид G3 - в полном объеме колонки. Препарат P4A1-Л получали коньюгированием пустых липосом с антителом P4A1-2A аналогичным образом. Под пустыми липосомами понимается препарат липосом, внутренняя водная фаза которого не содержит цитотоксического компонента.
Пример 6. Количественное определение P4A1-2A антитела в составе иммунолипосом методом ИФА
В лунки 96-луночных планшетов MaxiSorp (Nunc, Дания) вносили по 100 мкл раствора рекомбинантного рецептор-связывающего домена S-белка SARS-CoV-2 (RBD) в фосфатно-солевом буфере (PBS) в концентрации 1 мкг/мл и инкубировали планшет в течение ночи при 2-8°C. После блокировки свободных сайтов связывания блокирующим буфером (PBS, 0.05% Tween-20, 0.1% БСА) вносили образцы анализируемых липосомальных препаратов P4A1-Л и P4A1-ЛБ и раствор антитела P4A1-2A известной концентрации в блокирующем буфере в разведениях и инкубировали в течение 30 мин при 37°С. После 30-минутной инкубации при 37°C и промывки в лунки планшета добавляли раствор антител к Fc-фрагменту антител человека, конъюгированных с пероксидазой хрена (SigmaAldrich, США, кат. № AP113P), в разведении 1:10 000 в блокирующем буфере, планшет инкубировали в течение 30 мин. По окончании инкубации и промывки в лунки планшета добавляли по 100 мкл субстратного раствора TMB и инкубировали в темноте в течение 15 мин. Ферментативную реакцию останавливали добавлением 10% раствора фосфорной кислоты, измеряли величины ОП 450 нм в лунках планшета на планшетном спектрофотометре. Строили кривую зависимости величин ОП450 от концентрации антитела P4A1-2A и использовали ее для вычисления концентраций антитела P4A1-2A в образцах липосомальных препаратов.
Пример 7. Вируснейтрализующее действие иммунолипосом в псевдовирусной системе
Вируснейтрализующее действие иммунолипосом изучали в псевдовирусной системе, моделирующей проникновение вирусных частиц SARS-CoV-2 в клетки, несущие на своей поверхности рецептор АСЕ-2. Псевдовирусная система включает в себя линию клеток HEK293T-ACE-2, несущую на своей поверхности рецептор ACE-2 человека и рекомбинантные лентивирусные частицы, псевдотипированные S-белком SARS-CoV-2 и содержащие в качестве репортерного гена ген люциферазы светлячка [24]. Детекция вируснейтрализующей активности происходит за счет изменения люминесценции клетками, несущими на своей поверхности АСЕ-2, при добавлении к ним псевдовирусных частиц, прединкубированных с испытуемыми веществами. При этом, чем больше вируснейтрализующая активность испытуемого вещества, тем меньше уровень детектируемой люминесценции.
Линию клеток HEK293T-ACE-2, гиперэкспрессирующую на поверхности рецептор ACE-2 человека, получали путем трансдукции HEK293T псевдовирусными частицами с мембранным гликопротеином VSV-g (гликопротеин G вируса везикулярного стоматита), содержащими в геноме гены рецептора ACE-2 человека и зеленого флуоресцентного белка (GFP). После трех последовательных обогащений трансдуцированных клеток, чередующихся пассажированием, методом проточной цитофлуориметрии были отобраны клетки, стабильно экспрессирующие GFP. Присутствие ACE-2 на поверхности пула клеток подтверждали проточной цитофлуориметрией. Клетки инкубировали с рекомбинантным RBD-Fc с дальнейшей визуализацией поликлональными антителами козы, кроссреактивными с Fc-доменом человека, коньюгированными с флуоресцентной меткой DL650, испускание которой детектируется в канале APC. Согласно полученным с помощью проточной цитофлуориметрии данным 96% клеток несут на своей поверхности АСЕ-2. Полученные клетки HEK293T-ACE-2 используют далее в тестах.
Получение рекомбинантных лентивирусных частиц, псевдотипированных S-белком SARS-CoV-2 и содержащих в качестве репортерного гена ген люциферазы светлячка, проводили как описано в работе [24]. Псевдовирус способен проникать в клетки HEK293T-ACE-2, несущие на своей поверхности АСЕ-2, где на рекомбинантном РНК-геноме синтезируется ДНК провируса, содержащая маркерный ген люциферазы светлячка, встраивание которого в геном клетки-мишени придает ей способность люминесцировать.
В тесте на вируснейтрализующую активность тестировали иммунолипосому, связывающуюся с RBD S-белка SARS-CoV-2 (P4A1-ЛБ), а в качестве контролей выступали липосома без инкапсулированной барназы, модифицированная полипептидом G3 (G3-ЛБ), неспецифичным к RBD-домену S-белка, и липосома без инкапсулированной барназы, модифицированная антителом P4A1-2A(P4A1-Л).
Тест на нейтрализующую активность проводили следующим образом. Клетки HEK293T-ACE-2 рассевали в лунки 96-луночной планшета в количестве 5000 клеток на 100 мкл среды DMEM с 10% фетальной бычьей сывороткой FBS (Gibco) и инкубировали в течение 24 часов при 37°C, 5% CO2. Псевдовирусы SARS-CoV-2 вносили по 50 мкл в лунки пустой 96-луночной плашки, добавляли к ним тестируемые вещества в концентрациях 46, 4,6, 0,46, 0,046, 0,0046, 0,00046 нМ и инкубировали при 37°C, 5% CO2 в течение 1 часа. Из лунок 96-луночной плашки с клетками HEK293T-ACE-2 удаляли по 50 мкл культуральной среды, добавляли прединкубированные смеси псевдовирусных частиц SARS-CoV-2 с указанными концентрациями для каждого из тестируемых веществ и инкубировали при 37°C, 5% CO2. Через 24 часа проводили смену среды, и еще через 48 часов культуральную среду удаляли, клетки лизировали с помощью GLO lysis buffer (кат. E2661, Promega), смешивали с субстратом Bright-Glo™ LuciferaseAssay System (кат. E2620, Promega), измеряли уровень люминесценции и строили график уровня люминисценции в зависимости от концентрации тестируемого вещества. Обработку данных и расчет значений ингибирующей концентрации (IC) проводили с помощью программного обеспечения GraphPadPrism 8.
Все три исследуемых липосомы демонстрируют нейтрализующую активность, обусловленную, по-видимому, разной природой (Фиг. 5). В случае липосомы G3-ЛБ, имеющей в качестве направляющей молекулы полипептид G3, который не обладает способностью связываться с S-белком SARS-CoV-2, нейтрализующая активность проявляется при концентрации липосом более 0.0055нМ. Этот эффект, возможно, реализуется за счет неспецифического связывания липосом с поверхностью псевдовирусов и/или клеток и дальнейшего слияния с ними. Барназа, высвобождающаяся после слияния, проявляет РНКазную активность в отношении вирусного генома в составе псевдовирусов и/или цитоплазматической РНК зараженных клеток, что приводит к снижению синтеза маркерного белка люциферазы. В случае липосомы P4A1-Л, не содержащей барназу, нейтрализующая активность обусловлена концентрационно-зависимым блокированием антителом P4A1-2A S-белка SARS-CoV-2 на поверхности псевдовирусов, что в свою очередь препятствует взаимодействию с рецептором ACE-2 и дальнейшему проникновению псевдовирусов в клетки. В случае P4A1-ЛБ - иммунолипосомы, связывающейся с RBD S-белка SARS-CoV-2, мы предполагаем, что нейтрализующая активность достигается за счет синергического эффекта, обусловленного концентрационно-зависимым блокированием проникновения псевдовирусов, как в случае липосомы P4A1-Л, так и РНК-азной активности барназы, как в случае липосомы G3-ЛБ. Для корректного сравнения нейтрализующей активности полученных липосомы P4A1-Л и P4A1-ЛБ - иммунолипосомы, связывающейся с RBD S-белка SARS-CoV-2, определяли концентрацию коньюгированного антитела P4A1-2A в составе этих липосом методом ИФА. Поскольку для P4A1-ЛБ - иммунолипосомы, связывающейся с RBD S-белка SARS-CoV-2, значения люминисценции для концентраций P4A1-2A выше 0.0028 нМ, соответствующих концентрациям иммунолипосом более 0.0055 нМ, могут быть занижены вследствие влияния неспецифической интернализации барназы, то сравнение нейтрализующей активности проводили, определяя ингибирующую концентрацию 40% (IC40). Согласно полученным данным (Фиг. 6) нейтрализующая активность липосомы P4A1-Л и иммунолипосомы P4A1-ЛБ (IC40 17±9 и 1.2±0.9 рМ соответственно) выше таковой свободного антитела P4A1-2A (IC40 55±6 рМ). Эти данные позволяют сделать вывод о том, что липосомы, имеющие на поверхности антитело P4A1-2A, вносят значительный вклад в нейтрализацию взаимодействия S-белка псевдовируса с рецептором ACE2. Данный эффект может быть объяснен несколькими механизмами. Возможно, липосомы P4A1-Л и иммунолипосомы P4A1-ЛБ, имеющие на поверхности антитело P4A1-2A, эффективно экранируют взаимодействие свободных молекул S-белка с рецептором ACE2, предупреждая инфекцию клеток. Также возможен описанный ранее эффект «экстракции» молекул S-белка с поверхности псевдовирусов липосомальными частицами [25]. Значительное увеличение нейтрализующей активности P4A1-ЛБ - иммунолипосомы, связывающейся с RBD S-белка SARS-CoV-2, (IC400.12±0.09 рМ) по сравнению с липосомой P4A1-Л (IC401.7±0.9 рМ), не содержащей инкапсулированную барназу, обусловлено деградацией клеточной и вирусной РНК вследствие высвобождения барназы из иммунолипосомы.
Таким образом, показано, что иммунолипосома, связывающейся с RBD S-белка SARS-CoV-2 за счет рекомбинантного нейтрализующего антитела P4A1-2A (P4A1-ЛБ), обладает нейтрализующей активностью более чем в 40 раз превышающей активность нейтрализующего антитела P4A1-2A в псевдовирусной системе.
Литература
1. WHO Coronavirus (COVID-19) Dashboardhttps://covid19. who.int
2. Rehman S.U., Shafique L., Ihsan A. et al. Evolutionary trajectory for the emergence of novel coronavirus SARS-CoV-2 // Pathogens. 2020. V. 9. P. 240.
3. Velavan T.P., Meyer C.G. The COVID-19 Epidemic // Trop. Med. Int. Health. 2020. V. 25. 278-280.
4. Masters P.S. The molecular biology of coronaviruses // Adv. Virus Res. 2006. V. 66. P. 193-292.
5. Zhou P. Yang X.L., Wang X.G. et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin // Nature. 2020. V. 579. P. 270-273.
6. Li X. Geng M., Peng Y. et al. Molecular immune pathogenesis and diagnosis of COVID-19// J. Pharm. Anal. 2020. V. 10. P. 102-108.
7. Ulyanova V., Vershinina V., Ilinskaya O. Barnase and binase: twins with distinct fates // FEBS J. 2011. V. 278 (19). P. 3633-3643.
8. Makarov A.A., Ilinskaya O.N. Cytotoxic ribonucleases: Molecular weapons and their targets // FEBS Lett. 2003. V. 540(1-3). P. 15-20).
9. Gribencha S.V., Potselueva L.A., Barinskii I.F. et al. The antiviral activity of RNAse Bacillus intermedius in experiments with mice preinfected with street rabies virus // Vopr. Virusol. 2006. V. 51. P. 41-43.
10. Alekseeva I.I., Kurinenko B.M., Kleiner G.I. et al. Comparative analysis of the antivital activity of pancreatic and micronial RHases // Antibiot. Med. Tekhnol. 1981. V. 26. P. 527-532.
11. Shneider M.A., Shtil’bans E.B., Kupriyanov-Ashin E.G. et al. Anti-influenza effect of bacterial RNase in experiment and a pharmacokinetic rationale for the method of its application // Antibiot. Khimioterap. 1990. V. 35. P.
12. Müller C., Ulyanova V., Ilinskaya O. et al. A Novel Antiviral Strategy against MERS-CoV and HCoV-229E Using Binase to Target Viral Genome Replication // Bionanoscience. 2017. V. 7(2). P. 294-299.
13. Shi R., Shan C., Duan X. et al. A human neutralizing antibody targets the receptor-binding site of SARS-CoV-2 // Nature 2020. V. 584. P. 120-124.
14. Chen X., Li R., Pan Z.; et al. Human Monoclonal Antibodies Block the Binding of SARS-CoV-2 Spike Protein to Angiotensin Converting Enzyme 2 Receptor // Cell. Mol. Immunol. 2020. V. 17. P. 647-649.
15. Liang, K.H., Chiang, P.Y., Ko, S.H. et al. Antibody cocktail effective against variants of SARS-CoV-2 // J Biomed Sci 2021. V. 28. P. 80.
16. Guo Y., Huang L., Zhang G., et al. A SARS-CoV-2 neutralizing antibody with extensive Spike binding coverage and modified for optimal therapeutic outcomes // Nat. Commun. 2021. V. 12. № 1. Р. 2623.
17. Widyasari K., Kim J. A review of the currently available antibody therapy for the treatment of coronavirus disease 2019 (COVID-19) // Antibodies. 2023. V. 12. № 1. Р. 5.
18. Park J.W., Kirpotin D.B., Hong K. et al.Tumor targeting using anti-her2 immunoliposomes // Journal of Controlled Release. 2001. V. 74. P. 95-113.
19. Di J., Xie F., Xu Y. When liposomes met antibodies: Drug delivery and beyond //Adv. Drug Deliv. 2020. V. 154-155. P. 151-162.
20. Патент RU260871C1. Рекомбинантная плазмида, обеспечивающая синтез барназы в клетках Escherichia coli, штамм Escherichia coli - продуцент барназы и способ получения барназы. Изобретатели: Деев С.М., Шульга А.А., Лукьянова Т.И., Коновалова Е.В.
21. Studier F.W., Moffatt B.A. Use of bacteriophage T7 RNA polymerase to direct selective high-level expression of cloned genes. J Mol Biol. 1986;189(1):113-130.
22. Studier F.W. Protein production by auto-induction in high density shaking cultures. Protein Expr Purif. 2005;41(1):207-234.
23. Deyev S., Proshkina G., Baryshnikova O., et al. Selective staining and eradication of cancer cells by protein-carrying DARPin-functionalized liposomes // Eur. J. Pharm. Biopharm. 2018. V. 130. P.296-305.
24. Kruglova N., Siniavin A., Gushchin V., et al. Different neutralization sensitivity of SARS-CoV-2 cell-to-cell and cell-free modes of infection to convalescent sera // Viruses. 2021. V. 13. P. 1133.
25. Zhang Z., King M.R. Neutralization of the new coronavirus by extracting their spikes using engineered liposomes // Nanomedicine. 2023. V. 50. P. 102674.
Перечень последовательностей
Sequence Listing Information:
DTD Version: V1_3
File Name: Liposome.xml
Software Name: WIPO Sequence
Software Version: 2.3.0
Production Date: 2023-12-25
General Information:
Current application / IP Office: RU
Current application / Application number: 11
Current application / Filing date: 2023-12-20
Current application / Applicant file reference: Liposome
Earliest priority application / IP Office: RU
Earliest priority application / Application number: 11
Earliest priority application / Filing date: 2023-12-20
Applicant name: ИБХ РАН
Applicant name / Language: ru
Applicant name / Name Latin: IBCh RAS
Invention title: ИММУНОЛИПОСОМА, СВЯЗЫВАЮЩАЯСЯ С RBD S-БЕЛКА SARS-COV-2 ( ru )
Sequence Total Quantity: 3
Sequences:
Sequence Number (ID): 1
Length: 110
Molecule Type: AA
Features Location/Qualifiers:
- source, 1..110
> mol_type, protein
> organism, Escherichia coli
Residues:
AQVINTFDGV ADYLQTYHKL PDNYITKSEA QALGWVASKG NLADVAPGKS IGGDIFSNRE 60
GKLPGKSGRT WREADINYTS GFRNSDRILY SSDWLIYKTT DHYCTFTKIR 110
Sequence Number (ID): 2
Length: 464
Molecule Type: AA
Features Location/Qualifiers:
- source, 1..464
> mol_type, protein
> organism, human
Residues:
MGWSCIILFL VATATGVHSE VQLVESGGGL IQPGGSLRLS CAASGFIVSS NYMSWVRQAP 60
GKGLEWVSII YSGGSTFYAD SVKGRFTISR DNSKNTLYLQ MNSLRVEDTA VYYCARDLQE 120
LGSLDYWGQG TLVTVSSAST KGPSVFPLAP CSRSTSESTA ALGCLVKDYF PEPVTVSWNS 180
GALTSGVHTF PAVLQSSGLY SLSSVVTVPS SSLGTKTYTC NVDHKPSNTK VDKRVESKYG 240
PPCPPCPAPE FLGGPSVFLF PPKPKDTLYI TREPEVTCVV VDVSQEDPEV QFNWYVDGVE 300
VHNAKTKPRE EQFNSTYRVV SVLTVLHQDW LNGKEYKCKV SNKGLPSSIE KTISKAKGQP 360
REPQVYTLPP SQEEMTKNQV SLTCLVKGFY PSDIAVEWES NGQPENNYKT TPPVLDSDGS 420
FFLYSRLTVD KSRWQEGNVF SCSVMHEALH NHYTQKSLSL SLGK 464
Sequence Number (ID): 3
Length: 234
Molecule Type: AA
Features Location/Qualifiers:
- source, 1..234
> mol_type, protein
> organism, human
Residues:
METDTLLLWV LLLWVPGSTG DIQMTQSPSS VSASVGDRVT ITCRASQGIS SWLAWYQQKP 60
GKAPKLLIYA ASSLQSGVPS RFSGSGSGTD FTLTISSLQP EDFATYYCQE ANSFPYTFGQ 120
GTKLEIKRTV AAPSVFIFPP SDEQLKSGTA SVVCLLNNFY PREAKVQWKV DNALQSGNSQ 180
ESVTEQDSKD STYSLSSTLT LSKADYEKHK VYACEVTHQG LSSPVTKSFN RGEC 234
END
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing originalFreeTextLanguageCode="ru"
dtdVersion="V1_3" fileName="Liposome.xml" softwareName="WIPO
Sequence" softwareVersion="2.3.0" productionDate="2023-12-25">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>11</ApplicationNumberText>
<FilingDate>2023-12-20</FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>Liposome</ApplicantFileReference>
<EarliestPriorityApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>11</ApplicationNumberText>
<FilingDate>2023-12-20</FilingDate>
</EarliestPriorityApplicationIdentification>
<ApplicantName languageCode="ru">ИБХ РАН</ApplicantName>
<ApplicantNameLatin>IBCh RAS</ApplicantNameLatin>
<InventionTitle languageCode="ru">ИММУНОЛИПОСОМА, СВЯЗЫВАЮЩАЯСЯ С
RBD S-БЕЛКА SARS-COV-2</InventionTitle>
<SequenceTotalQuantity>3</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>110</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..110</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Escherichia coli</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>AQVINTFDGVADYLQTYHKLPDNYITKSEAQALGWVASKGNLADVAPGK
SIGGDIFSNREGKLPGKSGRTWREADINYTSGFRNSDRILYSSDWLIYKTTDHYCTFTKIR</INSDSeq
_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>464</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..464</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q7">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>human</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>MGWSCIILFLVATATGVHSEVQLVESGGGLIQPGGSLRLSCAASGFIVS
SNYMSWVRQAPGKGLEWVSIIYSGGSTFYADSVKGRFTISRDNSKNTLYLQMNSLRVEDTAVYYCARDLQ
ELGSLDYWGQGTLVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHT
FPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEFLGGPSVFL
FPPKPKDTLYITREPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQD
WLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWE
SNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK</INS
DSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>234</INSDSeq_length>
<INSDSeq_moltype>AA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..234</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>protein</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q6">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>human</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>METDTLLLWVLLLWVPGSTGDIQMTQSPSSVSASVGDRVTITCRASQGI
SSWLAWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCQEANSFPYTFG
QGTKLEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSK
DSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Рекомбинантный белок, связывающийся с RBD S-белка SARS-CoV-2 | 2022 |
|
RU2778942C1 |
| ПОЛИПЕПТИД С АНТИ-SARS-CoV-2 АКТИВНОСТЬЮ И ПРОЛОНГИРОВАННЫМ ВРЕМЕНЕМ ЦИРКУЛЯЦИИ В КРОВОТОКЕ | 2023 |
|
RU2822355C1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ АКТИВНОСТИ НЕЙТРАЛИЗУЮЩИХ АНТИТЕЛ К SARS-CoV-2 В СЫВОРОТКЕ ИЛИ ПЛАЗМЕ КРОВИ ЛЮДЕЙ, ПЕРЕНЕСШИХ COVID-19 ИЛИ ПРИВИТЫХ ВАКЦИНАМИ ДЛЯ ПРОФИЛАКТИКИ НОВОЙ КОРОНАВИРУСНОЙ ИНФЕКЦИИ COVID-19, С ИСПОЛЬЗОВАНИЕМ НАБОРА РЕАГЕНТОВ ДЛЯ ИММУНОФЕРМЕНТНОГО АНАЛИЗА, СОДЕРЖАЩЕГО РЕКОМБИНАНТНЫЙ РЕЦЕПТОР-СВЯЗЫВАЮЩИЙ ДОМЕН (RBD) ПОВЕРХНОСТНОГО ГЛИКОПРОТЕИНА S КОРОНАВИРУСА SARS-COV-2 И РЕКОМБИНАНТНЫЙ ЧЕЛОВЕЧЕСКИЙ РЕЦЕПТОР АСЕ2 | 2021 |
|
RU2784655C1 |
| Широко нейтрализующее антитело против SARS-CoV-2 | 2022 |
|
RU2810476C1 |
| Однодоменное антитело ламы Н5 и его производное H5-Fc, специфически связывающие RBD-домен S-белка вируса SARS-CoV-2, обладающие вируснейтрализующей активностью | 2022 |
|
RU2793967C1 |
| Моноклональное антитело iC1 и его антигенсвязывающий фрагмент, селективно связывающие рецептор-связывающий домен Spike-белка вируса SARS-CoV-2, обладающие вируснейтрализующей активностью | 2023 |
|
RU2817696C1 |
| Моноклональное антитело iC2 и его антигенсвязывающий фрагмент, селективно связывающие рецептор-связывающий домен Spike-белка вируса SARS-CoV-2, обладающие вируснейтрализующей активностью | 2023 |
|
RU2817697C1 |
| Универсальный интеграционный вектор SB7G_HIGH и рекомбинантная плазмида SB7G_HIGH_DEV_57, обеспечивающая синтез и секрецию рекомбинантного человеческого антитела к рецептор-связывающему домену (RBD) коронавируса SARS-CoV-2 в клетках млекопитающих и полученная с использованием универсального вектора SB7G_HIGH, и рекомбинантное моноклональное антитело DEV_K57, обладающее вируснейтрализующей активностью в отношении SARS-CoV-2 | 2023 |
|
RU2829359C1 |
| Моноклональное антитело к RBD фрагменту в составе S белка вируса SARS-CoV-2 | 2020 |
|
RU2744274C1 |
| Средство и способ терапии и экстренной профилактики заболеваний, вызываемых вирусом SARS-CoV-2 на основе рекомбинантного антитела и гуманизированного моноклонального антитела | 2021 |
|
RU2769223C1 |
Изобретение относится к области биотехнологии и медицины. Описана иммунолипосома, связывающаяся с RBD S-белка SARS-CoV-2, содержащая во внутреннем объеме барназу, внешняя поверхность которой ковалентно модифицирована моноклональным нейтрализующим антителом P4A1-2A. Также представлен способ получения иммунолипосомы, включающий: смешивание раствора барназы с суспензией фосфолипидов, 7-кратное замораживание/оттаивание, продавливание через эксрудер с использованием поликарбонатных фильтров с диаметром пор 100 нм, гельпроникающую хроматографию на колонке с сорбентом сефароза CL-2B с последующей модификацией в течение 40 мин при комнатной температуре внешней поверхности липосомы 2-иминотиоланом с получением тиолированной липосомы, введение малеимидной группы в антитело P4A1-2A путем смешивания со 100-кратным молярным избытком реагента sulfo-EMCS и инкубирования 40 мин при комнатной температуре, смешивание тиолированной липосомы, содержащей во внутреннем объеме барназу, с антителом P4A1- 2A, содержащим малеимидные группы, и инкубацию 1 ч при комнатной температуре с получением иммунолипосомы, связывающейся с RBD S-белка SARS-CoV-2. Техническим результатом изобретения является увеличение нейтрализующей активности антитела P4A1-2A. 2 н. и 2 з.п. ф-лы, 6 ил., 7 пр.
1. Иммунолипосома, связывающаяся с RBD S-белка SARS-CoV-2, содержащая во внутреннем объеме барназу, внешняя поверхность которой ковалентно модифицирована моноклональным нейтрализующим антителом P4A1-2A.
2. Иммунолипосома по п.1, где барназа представляет собой рекомбинантную барназу из Bacillus amyloliquefaciens.
3. Иммунолипосома по п.1, где моноклональное нейтрализующее антитело представляет собой рекомбинантное моноклональное антитело P4A1-2A, обладающее сродством к рецептор-связывающему домену S-белка SARS-CoV-2.
4. Способ получения иммунолипосомы по п.1, включающий: смешивание раствора барназы с суспензией фосфолипидов, 7-кратное замораживание/оттаивание, продавливание через эксрудер с использованием поликарбонатных фильтров с диаметром пор 100 нм, гельпроникающую хроматографию на колонке с сорбентом сефароза CL-2B с последующей модификацией в течение 40 мин при комнатной температуре внешней поверхности липосомы 2-иминотиоланом с получением тиолированной липосомы, введение малеимидной группы в антитело P4A1-2A путем смешивания со 100-кратным молярным избытком реагента sulfo-EMCS и инкубирования 40 мин при комнатной температуре, смешивание тиолированной липосомы, содержащей во внутреннем объеме барназу, с антителом P4A1-2A, содержащим малеимидные группы, и инкубацию 1 ч при комнатной температуре с получением иммунолипосомы, связывающейся с RBD S-белка SARS-CoV-2.
| AU 2008275713 B2, 07.08.2014 | |||
| WO 2009009054 A1, 15.01.2009 | |||
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ИММУНОГЛОБУЛИНА IGA2m1-ИЗОТИПА В КЛЕТКАХ МЛЕКОПИТАЮЩИХ | 2022 |
|
RU2801178C1 |
| WO 2021214204 A1, 28.10.2021 | |||
| RU 2021117171 A, 12.12.2022. | |||

