Перекрестная ссылка на родственную заявку
В настоящей заявке испрашивается приоритет заявки на патент США рег. № 60/833239, поданной 26 июля 2006 и вводимой во всей своей полноте в настоящее описание посредством ссылки.
Область, к которой относится изобретение
Настоящее изобретение относится к пептидам или пептидным композициям и к способам их применения для ослабления (или ингибирования, или снижения уровня) высвобождения медиаторов из воспалительных клеток, стимулируемого в процессе воспаления. Настоящее изобретение также относится к применению этих пептидов или пептидных композиций для модуляции механизма внутриклеточной передачи сигнала регуляции секреции медиаторов воспаления из воспалительных клеток.
Предшествующий уровень техники
Воспалительные лейкоциты синтезируют ряд медиаторов воспаления, которые находятся внутри клеток в изолированном виде и сохраняются в цитоплазматических мембраносвязанных гранулах. Примерами таких медиаторов являются, но не ограничивается ими, миелопероксидаза [MPO] в нейтрофилах (см., например, Borregaard N, Cowland J. B. Granules of the human neutrophilic polymorphonuclear leukocyte. Blood 1997; 89:3503-3521), пероксидаза эозинофилов [EPO] и основный мажорный белок [MBP] в эозинофилах (см., например, Gleich G J. Mechanisms of eosinophil-associated inflammation. J. Allergy Clin. Immunol. 2000; 105:651-663), лизоцим в моноцитах/макрофагах (см., например, Hoff T, Spencker T, Emmendoerffer A., Goppelt-Struebe M. Effects of glucocorticoids on the TPA-induced monocytic differentiation. J. Leukoc Biol. 1992; 52:173-182; and Balboa M A, Saez Y, Balsinde J. Calcium-independent phospholipase A2 is required for lysozyme secretion in U937 promonocytes. J. Immunol. 2003; 170:5276-5280) и гранзим в природных клетках-киллерах (NK) в цитотоксических лимфоцитах (см., например, Bochan MR, Goebel WS, Brahmi Z. Stably transfected antisense granzyme B and perforin constructs inhibit human granule-mediated lytic ability. Cell Immunol. 1995; 164:234-239; Gong J.H., Maki G, Klingemann H.G. Characterization of a human cell line (NK-92) with phenotypical and functional characteristics of activated natural killer cells. Leukemia 1994; 8:652-658; Maki G, Kiingemann H.G, Martinson J.A, Tarn Y.K. Factors regulating the cytotoxic activity of the human natural killer cell line, NK-92. J. Hematother Stem Cell Res. 2001; 10:369-383; and Takayama H, Trenn G, Sitkovsky M.V. A novel cytotoxic T lymphocyte activation assay. J. Immunol. Methods 1987; 104:183-190). Такие медиаторы высвобождаются в области поражения и участвуют в развитии воспаления и репарации ткани, например, в легких и в каких-либо других органах. Известно, что лейкоциты высвобождают эти гранулы по механизму экзоцитоза (см., например, Burgoyne R.D, Morgan A. Secretory granule exocytosis. Physiol. Rev. 2003; 83:581-632; and Logan M.R, Odemuyiwa S.O, Moqbel R. Understanding exocytosis in immune and inflammatory cells: the molecular basis of mediator secretion. J. Allergy Clin. Immunol. 2003; 111:923-932), однако более точное описание регуляторных молекул и специфических путей, участвующих в процессе экзоцитоза, не приводится.
Некоторые экзогенные стимуляторы могут вызывать дегрануляцию лейкоцитов по механизму, который включает активацию протеинкиназы С и последующие события фосфорилирования (см., например, Burgoyne R.D, Morgan A. Secretory granule exocytosis. Physiol. Rev. 2003; 83:581-632; Logan M.R, Odemuyiwa S.O, Moqbel R. Understanding exocytosis in immune and inflammatory cells: the molecular basis of mediator secretion. J. Allergy Clin. Immunol. 2003; 111:923-932; Smolen J.E, Sandborg R.R. Ca2+-induced secretion by electropermeabilized human neutrophils: the roles of Ca2+, nucleotides and protein kinase C. Biochim Biophys Acta 1990; 1052:133-142; Niessen H.W, Verhoeven A.J. Role of protein phosphorylation in the degranulation of electropermeabilized human neutrophils. Biochim, Biophys. Acta 1994; 1223:267-273; and Naucler C, Grinstein S, Sundler R., Tapper H. Signaling to localized degranulation in neutrophils adherent to immune complexes. J. Leukoc. Biol. 2002; 71:701-710).
Белок MARCKS (где используемое здесь обозначение MARCKS означает «миристоилированный богатый аланином субстрат киназы С» («Myristoylated Alanine-Rich C Kinase Substrate») представляет собой часто встречающуюся мишень фосфорилирования протеинкиназы С (PKC), и экспрессируется в лейкоцитах на высоком уровне (см., например, Aderem A.A., Albert K.A., Keum M.M., Wang J.K., Greengard P Cohn Z.A. Stimulus-dependent myristoylation of a major substrate for protein kinase C. Nature 1988; 332:362-364; Thelen M, Rosen A, Nairn A.C., Aderem A. Regulation by phosphorylation of reversible association of a myristoylated protein kinase C substrate with the plasma membrane. Nature 1991; 351:320-322; and Hartwig J.H., Thelen M, Rosen A, Janmey P.A, Nairn A.C, Aderem A. MARCKS is an actin filament crosslinking protein regulated by protein kinase C and calcium-calmodulin. Nature 1992; 356:618-622). Белок MARCKS механистически участвует в процессе экзоцитозной секреции муцина бокаловидными клетками, которые выстилают дыхательные пути (см., например, Li et al., J. Biol. Chem. 2001; 276:40982-40990; и Singer et al., Nat. Med. 2004; 10:193-196). MARCKS миристоилируется посредством амидной связи у N-концевой аминокислоты в аминокислотной последовательности белка MARCKS в положении альфа-аминогруппы глицина, который присутствует у N-конца (то есть в положении 1) аминокислотной последовательности. В эпителиальных клетках дыхательных путей миристоилированная N-концевая область MARCKS, очевидно, является неотъемлемой частью секреторного процесса. Термин «N-конец белка MARCKS» означает пептид MANS, содержащий последовательность миристоил-GAQFSKTAAKGEAAAERPGEAAVA (SEQ ID NO:1), которая представляет собой последовательность, состоящую из L-аминокислот. Кроме того, описанные здесь пептидные фрагменты пептида MANS также предпочтительно состоят из L-аминокислот. Очевидно, что указанный механизм заключается в связывании миристоилированного белка MARCKS, то есть с мембранами внутриклеточных гранул.
Было показано, что N-концевой миристоилированный пептид, присутствующий у N-конца MARCKS, блокирует секрецию муцина и связывание MARCKS с мембранами муциновых гранул в бокаловидных клетках (см., например, Singer et al., Nat. Med. 2004; 10:193-196). Этот пептид содержит 24 аминокислоты белка MARCKS начиная с N-концевого глицина белка MARCKS, который является миристоилированным посредством амидной связи и известен как миристоилированная альфа-N-концевая последовательность (MANS), то есть миристоил-GAQFSKTAAKGEAAAERPGEAAVA (SEQ ID NO:1). В публикации Vergeres et al., J. Biochem. 1998, 330; 5-11, сообщается, что N-концевой глициновый остаток белков MARCKS миристоилируется посредством реакции, катализируемой миристоил- CoA:протеин-N-миристоилтрансферазой (NMT).
При воспалительных заболеваниях, таких как астма, ХОБЛ и хронический бронхит; при наследственных заболеваниях, таких как кистозный фиброз; при аллергических состояниях (атопии, аллергическом воспалении); при бронхиэктазе и при различных острых инфекционных респираторных заболеваниях, таких как пневмония, ринит, грипп или насморк, а также артрите или аутоиммунных заболеваниях воспалительные клетки обычно присутствуют на участках поражения или инфицирования, ассоциированных с воспалительными патологическими состояниями, либо мигрируют в дыхательные участки, а в частности указанные клетки присутствуют в дыхательных или воздухоносных путях либо мигрируют в дыхательные пути пациентов, страдающих указанными заболеваниями. Такие воспалительные клетки могут играть значительную роль в патологии заболеваний посредством поражения ткани, вызываемого медиаторами воспаления, высвобождаемыми из этих клеток. Одним из примеров такого поражения или деструкции ткани при хроническом воспалении может служить поражение ткани у пациентов с кистозным фиброзом, где медиаторы, высвобождаемые из нейтрофилов (например, миелопероксидаза [MPO]), индуцируют слущивание эпителиальной ткани дыхательных путей.
MARCKS, то есть белок массой приблизительно 82 кД, имеет три эволюционно консервативных области (Aderem et al, Nature 1988; 332:362-364; Thelen et al., Nature 1991; 351:320-322; Hartwig et al., Nature 1992; 356:618-622; Seykora et al., J. Biol. Chem. 1996; 271:18797-18802), а именно N-конец, домен сайта фосфорилирования (или PSD) и домен множественной гомологии 2 (MH2). Последовательности человеческих кДНК белка MARCKS известны и приводятся в публикации Harlan et al., J.Biol. Chem. 1991, 266:14399 (GenBank рег. № M68956), а также в публикации Sakai et al, Genomics 1992, 14:175. Эти последовательности также описаны в заявке WO 00/50062, которая во всей своей полноте вводится в настоящее описание посредством ссылки. N-концевая альфа-аминокислотная последовательность, содержащая 24 аминокислотных остатка, вместе с молекулой миристиновой кислоты, присоединенной посредством амидной связи к N-концевому глициновому остатку, участвует в связывании MARCKS с клеточными мембранами (Seykora et al., J. Biol. Chem. 1996; 271:18797-18802) и, вероятно, с кальмодулином (Matsubara et al., J. Biol. Chem. 2003; 278:48898-48902). Указанная последовательность из 24 аминокислот известна как пептид MANS.
Описание сущности изобретения
Участие белка MARCKS в высвобождении медиаторов воспаления из гранул инфильтрующихся лейкоцитов играет важную роль в развитии воспалительных заболеваний всех тканей и органов, включая болезни легких, характеризующиеся воспалением дыхательных путей, таким как астма, ХОБЛ и кистозный фиброз. Однако воспаление и секреция слизи в дыхательных путях происходят по двум отдельным и независимым механизмам (Li et al., J. Biol. Chem. 2001; 276:40982-40990; Singer et al., Nat. Med. 2004; 10:193-196). Продуцирование и секреция слизи могут индуцироваться различными факторами, включая медиаторы, высвобождаемые воспалительными клетками, однако прямой связи между избытком слизи и развитием воспаления пока не установлено.
В одном из аспектов изобретения, пептид MANS может играть определенную роль в снижении скорости и/или уровня высвобождения медиаторов воспаления из гранул или везикул в воспалительных лейкоцитах.
В другом аспекте изобретения, пептиды, присутствующие на N-конце MARCKS, а особенно у N-концевой последовательности, состоящей из 24 аминокислот, то есть активные смежные пептидные фрагменты, присутствующие в N-концевой последовательности MARCKS, состоящей из 1-24 аминокислот и имеющей глицин в положении 1, а также N-концевые амиды таких фрагментов, такие как N-концевые амиды уксусной кислоты и/или С-концевые амиды таких фрагментов, например C-концевые амиды аммиака, могут ингибировать или снижать скорость и/или уровень высвобождения медиаторов воспаления из воспалительных лейкоцитов. Такое ингибирование или снижение уровня высвобождения включает ингибирование MARCKS-ассоциированного высвобождения медиаторов воспаления из воспалительных лейкоцитов.
В другом аспекте изобретения, пептиды, присутствующие на N-конце MARCKS, а особенно у N-концевой последовательности, состоящей из 1-24 аминокислот, то есть активные смежные пептидные фрагменты, присутствующие в N-концевой последовательности MARCKS, состоящей из 1-24 аминокислот и имеющей глицин в положении 1, а также N-концевые амиды таких фрагментов, такие как N-концевые амиды уксусной кислоты и/или С-концевые амиды таких фрагментов, например C-концевые амиды аммиака, могут ингибировать скорость и/или уровень высвобождения медиаторов воспаления, например медиаторов воспаления идентифицированных в настоящей заявке, путем подавления процесса дегрануляции воспалительных лейкоцитов.
В другом аспекте изобретения, пептид MANS и его активные фрагменты, а также активные амиды таких описанных здесь фрагментов могут конкурировать с нативным белком MARCKS за связывание с мембраной воспалительных клеток, что будет приводить к ослаблению (снижению или подавлению) MARCKS-опосредуемого высвобождения медиаторов воспаления из гранул или везикул, содержащих такие медиаторы воспаления и присутствующих в указанных воспалительных клетках.
Клетки лейкоцитарного типа и их модели, которые секретируют содержимое специфических гранул в ответ на индуцированную форболовым эфиром активацию PKC, могут быть использованы для in vitro иллюстрации эффективности пептидов согласно изобретению и замещенных пептидов (например, альфа-N-амидов, C-концевых амидов и сложных эфиров) согласно изобретению.
Снижение уровня высвобождения мембраносвязанных медиаторов воспаления под действием соединений и композиций согласно изобретению может быть продемонстрировано с использованием человеческих лейкоцитарных клеточных линий. Так, например, нейтрофилы, выделенные из человеческой крови, могут быть использованы для иллюстрации ослабления или ингибирования высвобождения миелопероксидазы (MPO). Клон 15 человеческой промиелоцитарной клеточной линии HL-60 может быть использован для иллюстрации ослабления высвобождения или ингибирования высвобождения или секреции пероксидазы эозинофилов (EPO) под действием соединений и композиций согласно изобретению (см., например, Fischkoff S.A. Graded increase in probability of eosinophilic differentiation of HL-60 promyelocyte leukemia cells induced by culture under alkaline conditions. Leuk Res 1988; 12:679-686; Rosenberg H.F., Ackerman S. J., Tenen D.G. Human eosinophil cationic protein: molecular cloning of a cytotoxin and helminthotoxin with ribonuclease activity. J. Exp. Med. 1989; 170:163-176; Tiffany H.L., Li F, Rosenberg H.F. Hyperglycosylation of eosinophil ribonucleases in a promyelocytic leukemia cell line and in differentiated peripheral blood progenitor cells. J. Leukoc. Biol. 1995; 58:49-54; and Badewa A.P., Hudson C.E., Heiman A.S. Regulatory effects of eotaxin, eotaxin-2, and eotaxin-3 on eosinophil degranulation and superoxide anion generation. Exp. Biol. Med. 2002; 227:645-651). Клеточная линия моноцитарного лейкоза U937 может быть использована для иллюстрации ослабления высвобождения или ингибирования высвобождения или секреции лизоцима под действием соединений и композиций согласно изобретению (см., например, Hoff T, Spencker T, Emmendoerffer A., Goppelt-Struebe M. Effects of glucocorticoids on the TPA- induced monocytic differentiation. J. Leukoc. Biol. 1992; 52:173-182; Balboa M. A., Saez Y, Balsinde J. Calcium-independent phospholipase A2 is required for lysozyme secretion in U937 promonocytes. J. Immunol. 2003; 170:5276-5280; and Sundstrom C, Nilsson K. Establishment and characterization of a human histiocytic lymphoma cell line (U-937). Int. J. Cancer 1976; 17:565-577). Лимфоцитарная клеточная линия клеток природных киллеров NK-92 может быть использована для иллюстрации ослабления или ингибирования высвобождения гранзима под действием соединений и композиций согласно изобретению (см., например, Gong J.H., Maki G, Klingemann H.G. Characterization of a human cell line (NK-92) with phenotypical and functional characteristics of activated natural killer cells. Leukemia 1994; 8:652-658; Maki G, Klingemann H.G., Martinson J.A., Tam Y.K. Factors regulating the cytotoxic activity of the human natural killer cell line, NK-92. J Hematother Stem Cell Res 2001; 10:369-383; and Takayama H, Trenn G, Sitkovsky M.V. A novel cytotoxic T lymphocyte activation assay. J Immunol Methods 1987; 104:183-190). В методе ингибирования или ослабления высвобождения медиатора воспаления in vitro, например, описанного в настоящей заявке, клетки каждого типа предварительно инкубируют с пептидным соединением или с пептидной композицией согласно изобретению при различных концентрациях, а затем эти клетки инкубируют со стимулятором высвобождения медиаторов воспаления, таким как форболовый эфир. Процент ингибирования высвобождения медиатора воспаления определяют по отношению к высвобождению медиатора в отсутствие пептидного соединения или пептидной композиции, например, спектрофотометрическим методом определения концентрации высвобождаемого медиатора.
Для того чтобы продемонстрировать важное значение соответствующей аминокислотной последовательности, присутствующей в пептидах согласно изобретению, относительную способность к ингибированию или снижению уровня медиатора воспаления, высвобождаемого пептидом, идентичным последовательности белка MARCKS, состоящего из 24 аминокислот в N-концевой области (то есть пептидом MANS с миристоилированной альфа-N-концевой последовательностью) сравнивают со способностью к ингибированию или снижению уровня медиатора воспаления, высвобождаемого под действием пептида, содержащего те же самые 24 аминокислотных остатка, которые присутствуют в MANS, но по сравнению с порядком расположения аминокислотных остатков в последовательности MANS, расположены в произвольном порядке (то есть пептида RNS, иначе называемого «пептидом с рандомизированной N-концевой последовательностью). В клетках каждого из оцениваемых типов пептид MANS, но не пептид RNS ослабляет высвобождение медиаторов воспаления в зависимости от концентрации в течение периода времени от 0,5 до 3,0 часа. Полученные результаты дают основание предполагать, что соответствующая аминокислотная последовательность, присутствующая в пептидах согласно изобретению, которая находится в белке MARCKS в установленном порядке, в частности в его N-концевой области, а более конкретно последовательность его 24 аминокислотных остатков в N-концевой области, участвует по меньшей мере в одном внутриклеточном пути ингибирования дегрануляции лейкоцитов.
Настоящее изобретение относится к новому применению пептидной последовательности, состоящей из 24 аминокислот, и к альфа-N-концевой ацетилированной пептидной последовательности, миристоилированного полипептида, также известного как пептид MANS и их активных фрагментов, где указанные активные фрагменты могут быть выбраны из группы пептидов, имеющих от 4 до 23 смежных аминокислотных остатков аминокислотной последовательности пептида MANS, и где указанные фрагменты могут быть миристоилированными у N-конца в том случае, если они не начинаются с N-концевого глицина в положении 1 SEQ ID NO:1, или где указанные фрагменты могут быть ацилированными у N-конца C2- C12-ацильными группами, включая группу, ацилированную у N-конца, и/или C-концевую группу, амидированную группой NH2.
Настоящее изобретение также относится к новому способу блокирования MARCKS-ассоциированных клеточных секреторных процессов, а в частности процессов, которые заключаются в MARCKS-ассоциированном высвобождении медиаторов воспаления из воспалительных клеток, в путях стимуляции которых участвует субстрат протеинкиназы С (PKC) для белка MARCKS и в которых происходит высвобождение содержимого из внутриклеточных везикул или гранул.
Настоящее изобретение относится к способу ингибирования экзоцитозного высвобождения по меньшей мере одного медиатора воспаления по меньшей мере из одной воспалительной клетки, где указанный способ включает контактирование по меньшей мере одной воспалительной клетки, содержащей по меньшей мере один медиатор воспаления, присутствующий во внутриклеточной везикуле, по меньшей мере с одним пептидом, выбранным из группы, состоящей из пептида MANS и его описанного здесь активного фрагмента в количестве, эффективном для снижения уровня высвобождения медиатора воспаления из воспалительной клетки, по сравнению с уровнем высвобождения медиатора воспаления из воспалительной клетки того же типа, наблюдаемым в отсутствие по меньшей мере одного пептида.
Настоящее изобретение также относится к способу ингибирования высвобождения по меньшей мере одного медиатора воспаления по меньшей мере из одной воспалительной клетки в ткани или физиологической жидкости индивидуума, где указанный способ включает введение в ткань и/или физиологическую жидкость индивидуума, содержащие по меньшей мере одну воспалительную клетку, включающую по меньшей мере один медиатор воспаления, присутствующий во внутриклеточной везикуле, терапевтически эффективного количества фармацевтической композиции, содержащей по меньшей мере один пептид, выбранный из группы, состоящей из пептида MANS и его описанного здесь активного фрагмента в количестве, терапевтически эффективном для снижения уровня высвобождения медиатора воспаления по меньшей мере из одной воспалительной клетки, по сравнению с уровнем высвобождения медиатора воспаления по меньшей мере из одной воспалительной клетки того же типа в отсутствие по меньшей мере одного пептида. Более конкретно, ингибирование высвобождения медиатора воспаления включает блокирование или снижение уровня высвобождения медиатора воспаления из воспалительной клетки.
Более конкретно, настоящее изобретение относится к способу снижения воспаления у индивидуума, где указанный способ включает введение терапевтически эффективного количества фармацевтической композиции, содержащей пептид MANS (то есть N-миристоил- GAQFSKTAAKGEAAAERPGEAAVA (SEQ ID NO:1)) или его активный фрагмент. Такой активный фрагмент имеет длину по меньшей мере в четыре, а предпочтительно, по меньшей мере в шесть аминокислот. Используемый здесь термин «активный фрагмент» белка MARCKS означает фрагмент, который влияет (ингибирует или ослабляет) на опосредуемое белком MARCKS высвобождение, например опосредуемое белком MARCKS высвобождение медиатора воспаления. Активный фрагмент может быть выбран из группы, состоящей из GAQFSKTAAKGEAAAERPGEAAV (SEQ ID NO:2); GAQFSKTAAKGEAAAERPGEAA (SEQ ID NO:4); GAQFSKTAAKGEAAAERPGEA (SEQ ID NO:7); GAQFSKTAAKGEAAAERPGE (SEQ ID NO:11); GAQFSKTAAKGEAAAERPG (SEQ ID NO:16); GAQFSKTAAKGEAAAERP (SEQ ID NO:22); GAQFSKTAAKGEAAAER (SEQ ID NO:29); GAQFSKTAAKGEAAAE (SEQ ID NO:37); GAQFSKTAAKGEAAA (SEQ ID NO:46); GAQFSKTAAKGEAA (SEQ ID NO:56); GAQFSKTAAKGEA (SEQ ID NO:67); GAQFSKTAAKGE (SEQ ID NO:79); GAQFSKTAAKG (SEQ ID NO:92); GAQFSKTAAK (SEQ ID NO:106); GAQFSKTAA (SEQ ID NO:121); GAQFSKTA (SEQ ID NO:137); GAQFSKT (SEQ ID NO:154); GAQFSK (SEQ ID NO:172); GAQFS (SEQ ID NO:191) и GAQF (SEQ ID NO:211). Эти пептиды не содержат миристоильной группы у N-концевой аминокислоты, то есть либо не содержат химической группы, либо содержат немиристоильную химическую группу у N-концевой аминокислоты и/или химическую группу у С-концевой аминокислоты, например описанную здесь N-концевую ацетильную группу и/или С-концевую амидную группу. Присутствие гидрофобной N-концевой миристоильной группы в пептидах MANS и в их N-концевых миристоилированных фрагментах может приводить к повышению их способности проникать в плазматические мембраны, а вероятно, и к повышению их проницаемости в плазматических мембранах, а также, возможно, к сообщению этим пептидам растворимости в клетках. После введения гидрофобной миристоильной группы в липидный бислой мембраны эти липиды могут иметь коэффициент распределения или кажущуюся константу ассоциации до 104 М-1 или унитарную свободную энергию связывания Гиббса, составляющую примерно 8 ккал/моль (см., например, Peitzsch, R. M., and McLaughlin, S. 1993, Binding of acylated peptidas and fatty acids to phospholipid vesicles: pertinence to myristoylated proteins. Biochemistry. 32:10436-10443), где указанные величины являются достаточными, по меньшей мере частично, для распределения пептида MANS и миристоилированных фрагментов пептида MANS в плазматической мембране клетки, а дополнительные функциональные группы и их взаимодействия в пептиде MANS (который является миристоилированным) и в миристоилированых фрагментах пептида MANS могут потенцировать их относительную способность к проникновению в мембрану. Каждый из этих фрагментов может иметь коэффициенты распределения и аффинности по отношению к мембранам, которые по своей структуре являются репрезентативными. Эти фрагменты могут быть получены методами пептидного синтеза, известными специалистам, такими как твердофазный пептидный синтез (см., например, методы, описанные в публикациях Chan, Weng C. and White, Peter D.Eds., Fmoc Solid Phase Peptide Synthesis: A Practical Approach, Oxford University Press, New York, New York (2000); и Lloyd-Williams, P. et al. Chemical Approaches to the Synthesis of Peptides and Proteins (1997)), и могут быть очищены методами, известными специалистам, такими как жидкостная хроматография высокого давления. Молекулярная масса каждого пептида может быть подтверждена с помощью масс-спектроскопии, где каждый пептид имеет пик с соответствующей молекулярной массой. Эффективность отдельных пептидов и их комбинаций (например, комбинаций из 2 пептидов, комбинаций из 3 пептидов, комбинаций из 4 пептидов) в способах согласно изобретению может быть легко определена без излишнего экспериментирования в соответствии с процедурами, описанными в примерах, представленных в настоящей заявке. Предпочтительная комбинация содержит два пептида, при этом предпочтительное молярное отношение указанных пептидов может составлять от 50:50 (то есть 1:1) до 99,99-0,01, где указанное отношение может быть легко определено с применением процедур, описанных в примерах, представленных в настоящей заявке.
Предпочтительно, пептид MANS или его активный фрагмент присутствуют в фармацевтической композиции, которая может быть использована для блокирования воспаления. Настоящее изобретение также относится к способам ингибирования клеточного секреторного процесса у индивидуума, где указанный способ включает введение терапевтически эффективного количества соединения, содержащего пептид MANS или его активный фрагмент, ингибирующего медиатор воспаления у индивидуума. Такое введение обычно выбирают из группы, состоящей из местного введения, парентерального введения, ректального введения, внутрилегочного введения, введения путем ингаляции и интерназального введения или перорального введения, где указанное внутрилегочное введение обычно осуществляют с помощью аэрозоля, инсуффлятора, ингалятора с дозирующим клапаном или небулайзера .
Введение композиции, содержащей ингибирующее дегрануляцию количество пептида MANS или ингибирующее дегрануляцию количество его активного фрагмента, например фармацевтической композиции пептида MANS или его активного фрагмента, человеку или животному обеспечивает контактирование пептида MANS или его активного фрагмента, присутствующего по меньшей мере на участке, или в ткани или на ткани, или в слое или на слое, содержащем физиологическую жидкость, с поверхностью ткани, в которой присутствуют воспалительные гранулоцитарные клетки или в которую они проникают, что позволяет пептиду MANS или его активному фрагменту контактировать с воспалительной гранулоцитарной клеткой. В одном из аспектов изобретения, введение такой композиции может быть осуществлено при первых признаках или при первом обнаружении воспаления или при первом ощущении воспаления человеком или животным или при первом заметном изменении степени воспаления у человека или животного в целях ослабления воспаления, которое наблюдается в отсутствие пептида MANS или его активного фрагмента. В другом аспекте изобретения, такое введение может быть осуществлено в процессе воспаления ткани у человека или животного для ослабления уровня прогрессирования воспаления, которое происходит в отсутствие пептида MANS или его активного фрагмента. Хотя количество и частота вводимой дозы могут быть определены в соответствии с клинической оценкой и зависят от типа заболевания или источника воспаления, от степени поражения ткани и от возраста и массы пациента, однако предполагается, что доза фармацевтической композиции может быть снова введена через 3-8 часов, а предпочтительно через 6-8 часов после первого введения фармацевтической композиции.
Настоящее изобретение также относится к способам ослабления воспаления у индивидуума, где указанные способы включают введение терапевтически эффективного количества соединения, которое ингибирует MARCKS-ассоциированное высвобождение медиаторов воспаления, в результате чего уровень высвобождения по меньшей мере одного медиатора воспаления у индивидуума снижается по сравнению с уровнем, который наблюдается в отсутствие такой обработки. Используемый здесь термин «ослабление», по существу, означает уменьшение степени воспаления. Предпочтительно, высвобождение медиаторов воспаления ингибируется или блокируется описанными здесь способами.
В другом своем варианте, настоящее изобретение относится к способам ослабления воспаления у индивидуума, где указанные способы включают введение терапевтически эффективного количества соединения, которое ингибирует MARCKS-ассоциированное высвобождение медиаторов воспаления, в результате чего степень воспаления у индивидуума снижается по сравнению со степенью воспаления, наблюдаемой в отсутствие такой обработки. Настоящее изобретение также относится к способам ослабления или ингибированния воспаления у индивидуума, где указанные способы включают введение терапевтически эффективного количества пептида MANS или его активного фрагмента для ингибирования медиатора воспаления в зоне воспаления. Термин «ингибирование» означает снижение уровня секреции медиаторов воспаления. Термин «полное ингибирование» означает снижение уровня секреции медиаторов воспаления до нулевого значения. Как указывалось выше, активный фрагмент имеет длину по меньшей мере четыре, а предпочтительно, по меньшей мере шесть аминокислот. Термин «процесс экзоцитоза» означает экзоцитоз, то есть процесс клеточной секреции или экскреции, при котором вещества, содержащиеся в везикуле, присутствующей внутри клетки, высвобождаются из клетки путем слияния мембраны везикулы с внешней клеточной мембраной. Термин «дегрануляция» означает высвобождение содержимого клеточных гранул. Термин «ингибирование дегрануляции» означает снижение уровня высвобождения медиаторов воспаления, содержащихся в гранулах воспалительной клетки. Таким образом, количеством пептида MANS и/или его активного фрагмента, ингибирующим дегрануляцию, является количество этих пептидов, достаточное для снижения уровня высвобождения медиаторов воспаления, содержащихся в гранулах, по сравнению с уровнем высвобождения указанных медиаторов в отсутствие такого пептида.
В эталонном пептиде GAQFSKTAAKGEAAAERPGEAAVA (SEQ ID NO:1), начиная с N-конца: в положении 1 присутствует G; в положении, смежном с положением 1, в котором присутствует G, то есть в положении 2, присутствует А; в положении, смежном с положением 2, в котором присутствует А, то есть в положении 3, присутствует Q; в положении, смежном с положением 3, в котором присутствует Q, то есть в положении 4, присутствует F; в положении, смежном с положением 4, в котором присутствует F, то есть в положении 5, присутствует S; в положении, смежном с положением 5, в котором присутствует S, то есть в положении 6, присутствует K; в положении, смежном с положением 6, в котором присутствует K, то есть в положении 7, присутствует T; в положении, смежном с положением 7, в котором присутствует T, то есть в положении 8, присутствует А; в положении, смежном с положением 8, в котором присутствует A, то есть в положении 9, присутствует А; в положении, смежном с положением 9, в котором присутствует A, то есть в положении 10, присутствует K; в положении, смежном с положением 10, в котором присутствует K, то есть в положении 11, присутствует G; в положении, смежном с положением 11, в котором присутствует G, то есть в положении 12, присутствует E; в положении, смежном с положением 12, в котором присутствует E, то есть в положении 13, присутствует А; в положении, смежном с положением 13, в котором присутствует A, то есть в положении 14, присутствует А; в положении, смежном с положением 14, в котором присутствует A, то есть в положении 15, присутствует А; в положении, смежном с положением 15, в котором присутствует A, то есть в положении 16, присутствует E; в положении, смежном с положением 16, в котором присутствует E, то есть в положении 17, присутствует R; в положении, смежном с положением 17, в котором присутствует R, то есть в положении 18, присутствует P; в положении, смежном с положением 18, в котором присутствует P, то есть в положении 19, присутствует G; в положении, смежном с положением 19, в котором присутствует G, то есть в положении 20, присутствует E; в положении, смежном с положением 20, в котором присутствует E, то есть в положении 21, присутствует А; в положении, смежном с положением 21, в котором присутствует A, то есть в положении 22, присутствует А; в положении, смежном с положением 22, в котором присутствует A, то есть в положении 23, присутствует V; и в положении, смежном с положением 23, в котором присутствует V, то есть в положении 24, присутствует А, где положением 24 является С-концевое положение эталонного пептида.
«Вариант» эталонного пептида или вариант сегмента эталонного пептида, состоящего из 4-23 аминокислот, представляет собой пептид, который имеет аминокислотную последовательность, отличающуюся от аминокислотной последовательности эталонного пептида или от аминокислотной последовательности сегмента эталонного пептида, соответственно, по меньшей мере в одном положении аминокислоты аминокислотной последовательности эталонного пептида или сегмента эталонного пептида, соответственно, но при этом сохраняет муцин- или слизь-ингибирующую активность, которая обычно в 0,1-10 раз, предпочтительно, в 0,2-6 раз, а более предпочтительно, в 0,3-5 раз превышает указанную активность эталонного пептида или его сегмента, соответственно. «Вариант» эталонной аминокислотной последовательности или вариант сегмента эталонной аминокислотной последовательности, состоящей из 4-23 аминокислот, представляет собой аминокислотную последовательность, которая отличается от эталонной аминокислотной последовательности или от сегмента эталонной аминокислотной последовательности по меньшей мере на одну аминокислоту, соответственно, имеет аминокислотную последовательность пептида, которая сохраняет муцин- или слизь-ингибирующую активность кодируемой аминокислотной последовательности указанного эталонного пептида или сегмента, соответственно, где указанная активность обычно в 0,1-10 раз, предпочтительно, в 0,2-6 раз, а более предпочтительно, в 0,3-5 раз превышает активность указанного пептида или его сегмента эталонной последовательности, соответственно. Вариант пептида с заменой или вариант аминокислотной последовательности с заменой может отличаться от аминокислотной последовательности (то есть не совпадать с данной последовательностью) эталонного пептида или от эталонной аминокислотной последовательности одной или несколькими аминокислотными заменами, введенными в эталонную аминокислотную последовательность; вариант пептида с делецией или вариант аминокислотной последовательности с делецией может отличаться от аминокислотной последовательности (то есть не совпадать с данной последовательностью) эталонного пептида или от эталонной аминокислотной последовательности одной или несколькими аминокислотными делециями, введенными в эталонную аминокислотную последовательность; вариант пептида с добавлениями или вариант аминокислотной последовательности с добавлениями может отличаться от аминокислотной последовательности (то есть не совпадать с данной последовательностью) эталонного пептида или от эталонной аминокислотной последовательности одной или несколькими аминокислотными добавлениями, введенными в эталонную последовательность. Вариант пептида или вариант аминокислотной последовательности может быть получен в результате замены одной или нескольких аминокислот (например, замены по меньшей мере 1, 2, 3, 4, 5, 6, 7 или 8 аминокислот), введенной в эталонную последовательность, либо он может быть получен в результате делеции одной или нескольких аминокислот (например, по меньшей мере 1, 2, 3, 4, 5, 6, 7 или 8 аминокислот), введенной в эталонную последовательность, либо он может быть получен в результате добавления одной или нескольких аминокислот (например, по меньшей мере 1, 2, 3, 4, 5, 6, 7 или 8 аминокислот), введенной в эталонную последовательность, или их комбинаций в любом порядке. Вариант пептидного сегмента из 4-23 аминокислот с заменой или вариант последовательности сегмента из 4-23 аминокислот с заменой может отличаться от эталонного пептидного сегмента (то есть не совпадать с ним), состоящего из 4-23 аминокислот, или эталонной последовательности, состоящей из 4-23 аминокислот одной или несколькими аминокислотными заменами, введенными в эталонную последовательность аминокислотного сегмента; вариант пептидного сегмента из 4-23 аминокислот с делецией или вариант последовательности сегмента из 4-22 аминокислот с делецией может отличаться от эталонного пептидного сегмента (то есть не совпадать с ним) из 5-23 аминокислот или от эталонной аминокислотной последовательности сегмента из 4-23 аминокислот одной или несколькими аминокислотными делециями, введенными в эталонную последовательность аминокислотного сегмента; и вариант пептидного сегмента из 4-23 аминокислот с добавлением или вариант аминокислотной последовательности сегмента из 4-23 аминокислот с добавлением может отличаться от эталонного пептидного сегмента (то есть не совпадать с ним) из 4-22 аминокислот или от эталонной последовательности из 4-22 аминокислот одной или несколькими аминокислотными добавлениями, введенными в эталонную аминокислотную последовательность. Вариант пептида из 4-23 аминокислот или вариант аминокислотной последовательности из 4-23 аминокислот может быть получен в результате замены одной или нескольких аминокислот (например, замены по меньшей мере 1, 2, 3, 4, 5, 6, 7 или 8 аминокислот), введенной в сегмент эталонной аминокислотной последовательности из 4-23 аминокислот, либо он может быть получен в результате делеции одной или нескольких аминокислот (например, делеции по меньшей мере 1, 2, 3, 4, 5, 6, 7 или 8 аминокислот), введенной в соответствующую более крупную эталонную аминокислотную последовательность, либо он может быть получен в результате добавления одной или нескольких аминокислот (например, делеции по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8 аминокислот), введенной в соответствующую меньшую по размеру эталонную аминокислотную последовательность, либо он может быть получен в результате комбинированных модификаций. Предпочтительно, вариант пептида или аминокислотной последовательности отличается от эталонного пептида или от сегмента эталонного пептида, или эталонной аминокислотной последовательности, или от сегмента эталонной аминокислотной последовательности, соответственно, менее чем на 10 аминокислотных замен, делеций и/или добавлений, более предпочтительно, менее чем на 8 аминокислотных замен, делеций и/или добавлений, еще более предпочтительно, менее чем на 6 аминокислотных замен, делеций и/или добавлений, еще более предпочтительно, менее чем на 5 аминокислотных замен, делеций и/или добавлений, а наиболее предпочтительно, менее чем на 4 аминокислотных замены, делеции и/или добавления. Наиболее предпочтительно, вариант аминокислотной последовательности отличается от аминокислотной последовательности эталонного пептида или его сегмента последовательности на одну, две или три аминокислоты.
Термин «идентичность последовательностей», если он относится к аминокислотным последовательностям двух пептидов, означает число положений с идентичными аминокислотами, деленное на число аминокислот в более короткой из двух сравниваемых последовательностей.
Термин «по существу, идентичный», если он употребляется при сравнении аминокислотных последовательностей их двух пептидов или аминокислотных последовательностей из двух пептидных сегментов (например, сегментов аминокислотной последовательности эталонного пептида) означает, что аминокислотные последовательности пептидов или сегментов пептидов имеют по меньшей мере 75% идентичность, предпочтительно, по меньшей мере 80% идентичность, более предпочтительно, по меньшей мере 90% идентичность, а наиболее предпочтительно, по меньшей мере 95% идентичность.
Используемый здесь термин «пептид» включает пептид, а также его фармацевтически приемлемые соли.
Используемый здесь термин «выделенный» пептид означает природный пептид, который был отделен или, по существу, отделен от клеточных компонентов (например, нуклеиновых кислот и других пептидов), в окружении которых он обычно присутствует в природе, путем очистки, рекомбинантного синтеза или химического синтеза, а также охватывает не природные пептиды, которые были синтезированы рекомбинантными или химическими методами и которые были очищены или, по существу, очищены от клеточных компонентов, биологических материалов, химических предшественников или других химических веществ.
В описании настоящего изобретения используются нижеследующие трехбуквенные и однобуквенные обозначения аминокислот: аланин: (Ala) A; аргинин: (Arg) R; аспарагин: (Asn) N; аспарагиновая кислота: (Asp) D; цистеин: (Cys) C; глутамин: (Gln) Q; глутаминовая кислота: (Glu) E; глицин: (Gly) G; гистидин: (His) H; изолейцин: (Ile) I: лейцин: (Leu) L; лизин (Lys) K; метионин: (Met) M; фенилаланин: (Phe) F; пролин: (Pro) P; серин: (Ser) S; треонин: (Thr) T; триптофан (Trp) W; тирозин: (Tyr) Y; валин: (Val) V. В настоящей заявке используются и другие трехбуквенные символы аминокислот, указанные в скобках, а именно (Hyp) - гидроксипролин, (Nle) - норлейцин, (Orn) - орнитин, (Pyr) - пироглутаминовая кислота и (Sar) - саркозин. В соответствии с принятым соглашением, аминоконец (или N-конец) пептида указывается с левого конца записанной аминокислотной последовательности пептида, а карбоксиконец (или С-конец) указывается с правого конца записанной аминокислотной последовательности. Аминокислотная последовательность пептида может быть записана однобуквенными символами, означающими аминокислоты, которые ковалентно связаны пептидными амидными связями.
Активные фрагменты пептида MANS могут быть использованы в целях предотвращения воспаления или снижения уровня воспаления в ткани животного, где указанное воспаление вызывается медиаторами воспаления. Активные фрагменты пептида MANS могут быть также использованы в целях предотвращения поражения или снижения уровня поражения в ткани животного, где указанное поражение вызывается медиаторами воспаления. Активный фрагмент пептида MANS состоит по меньшей мере из 4 смежных аминокислот и не более чем 23 смежных аминокислот пептида MANS (SEQ ID NO:1). В контексте настоящего изобретения, термин «активный фрагмент» означает фрагменты пептидов MANS, которые могут предотвращать высвобождение или снижать уровень высвобождения медиаторов воспаления из воспалительных клеток. Такое снижение уровня высвобождения медиаторов воспаления под действием активных фрагментов может составлять по меньшей мере от 5% до 99% по сравнению с уровнем, наблюдаемым в присутствии эталонного пептида, такого как пептид MANS.
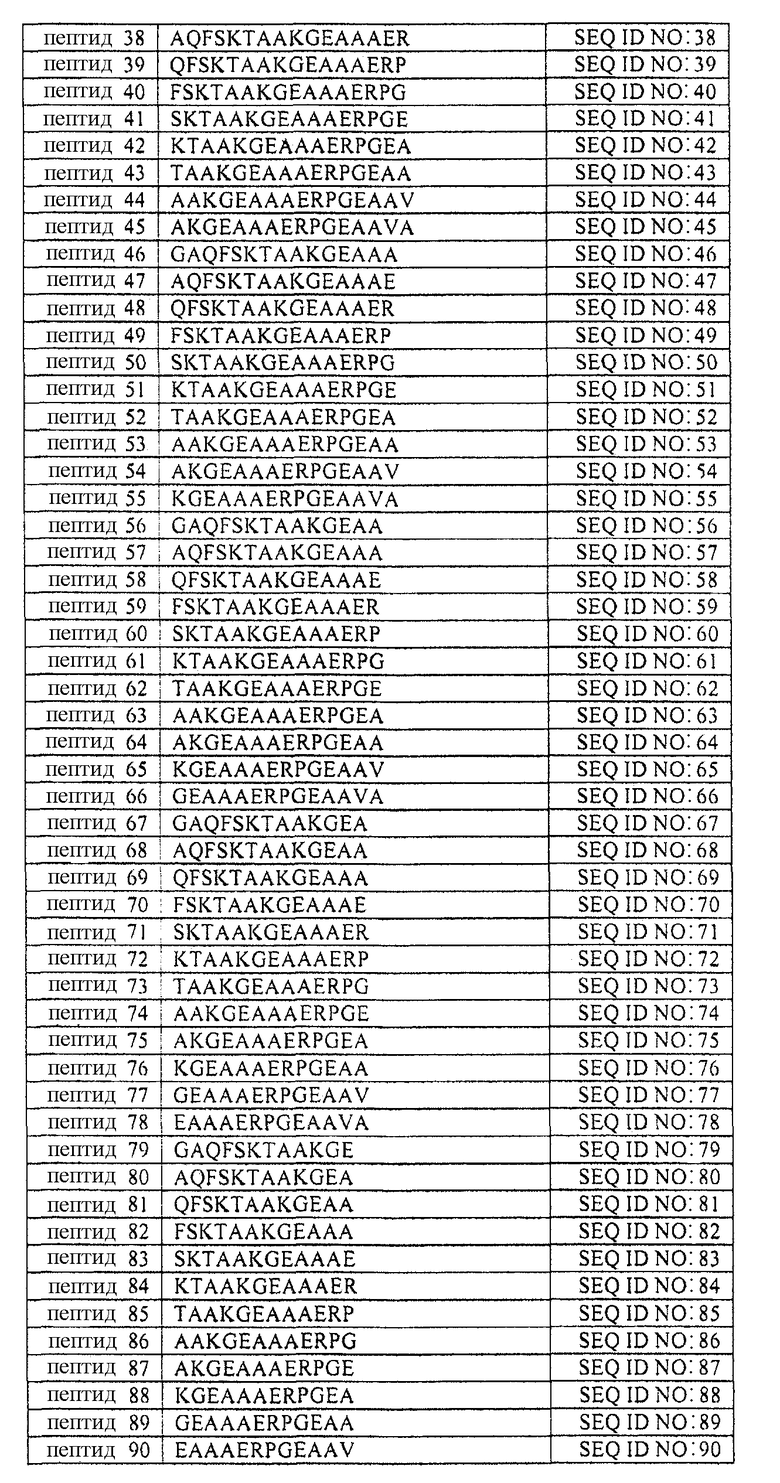
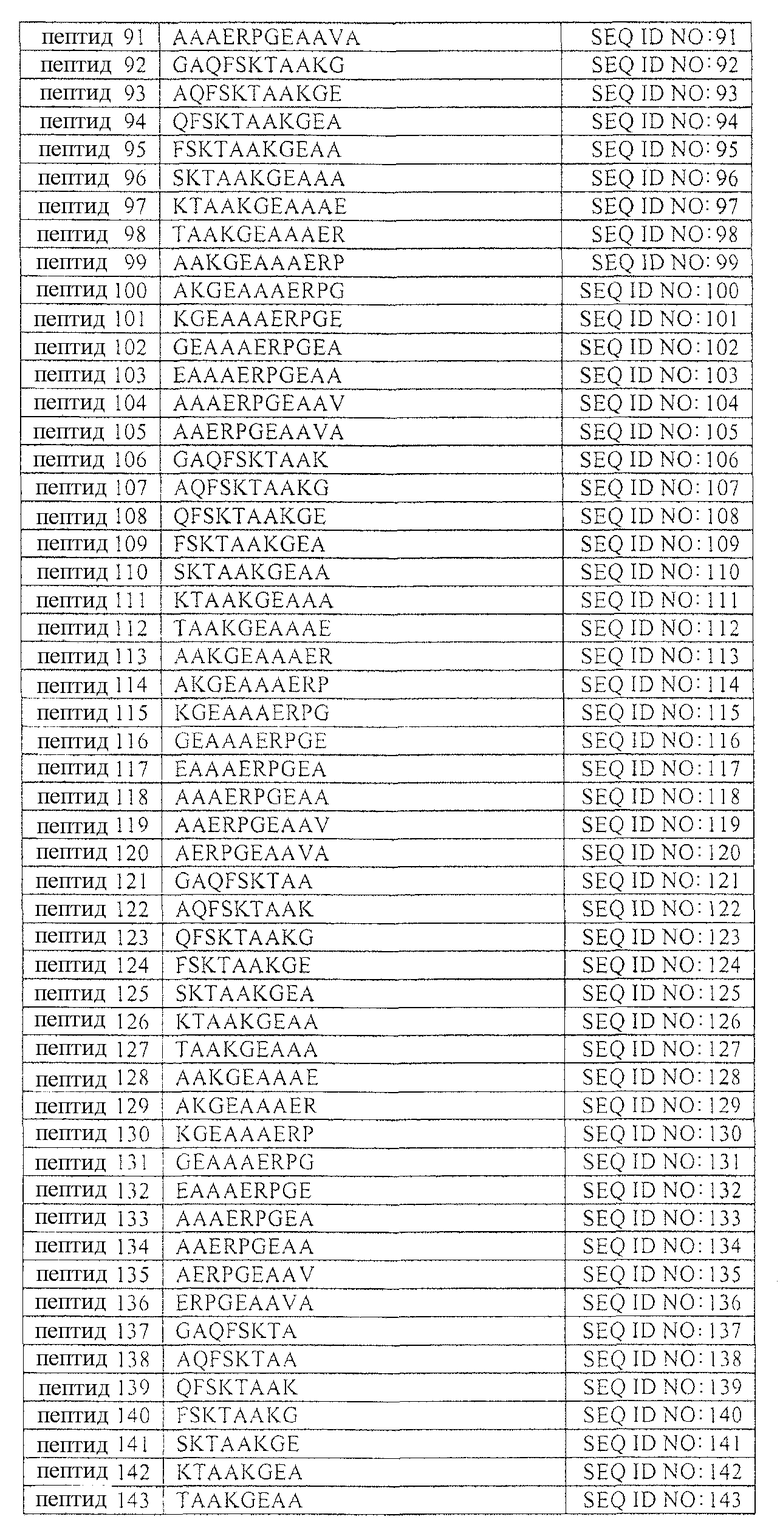
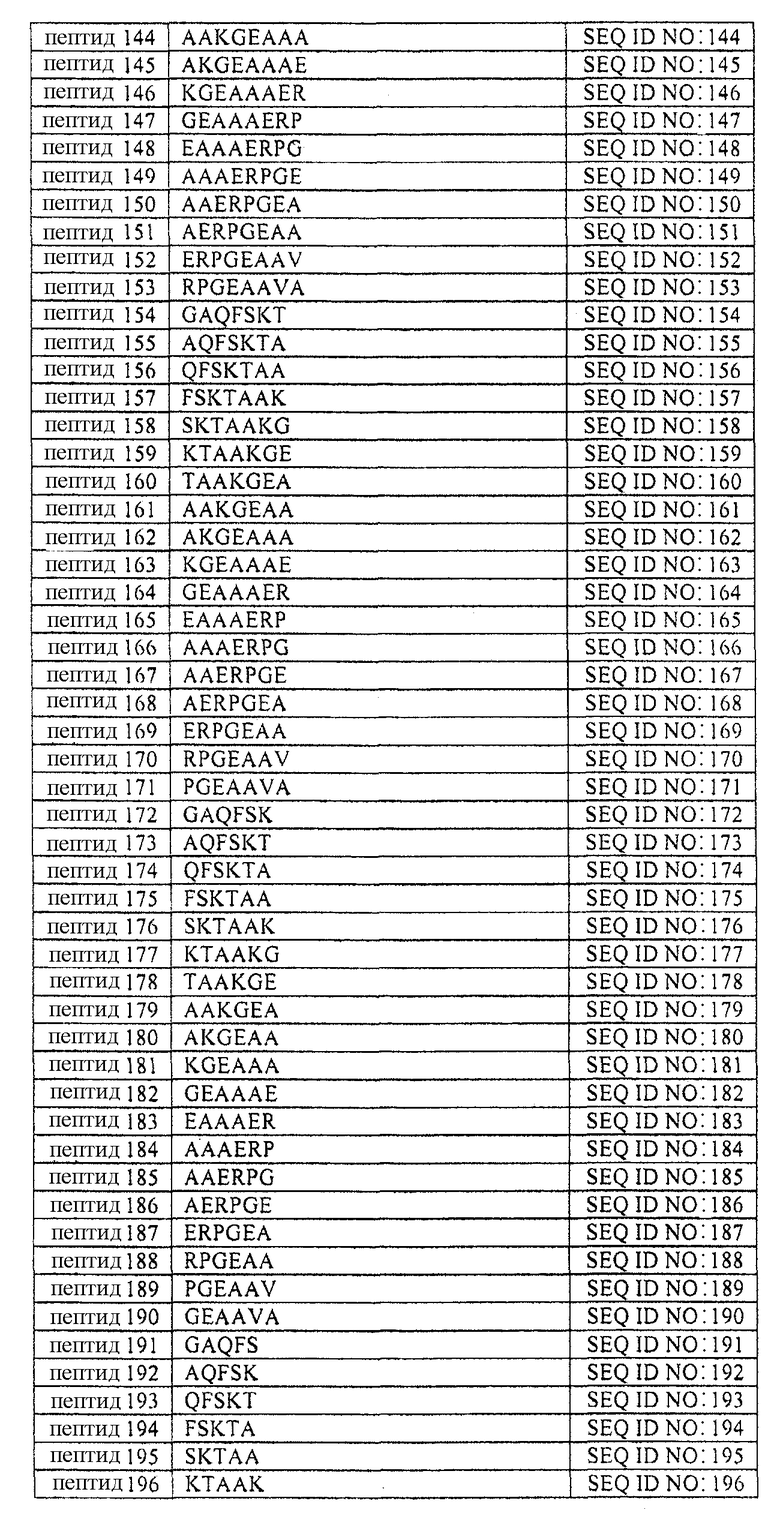
В таблице 1 приводится список аминокислотных последовательностей, представленных однобуквенным кодом, вместе с их соответствующими номерами и SEQ ID NO. Аминокислотная последовательность эталонного пептида (пептида MANS) представлена как пептид 1. Аминокислотные последовательности пептидов согласно изобретению, имеющих аминокислотную последовательность, состоящую из 4-23 смежных аминокислот эталонной аминокислотной последовательности, представлены как пептиды 2-231, вместе с аминокислотной последовательностью рандомизированной N-концевой последовательности (RNS), содержащей аминокислоты пептида MANS и представленной как пептид 232. Аминокислотные последовательности репрезентативных вариантов аминокислотных последовательностей описаны в настоящей заявке, а также пептидов согласно изобретению представлены как пептиды 233-245 и 247-251. Варианты перечисленных здесь пептидов не должны рассматриваться как ограничивающая группа пептидов и служат лишь репрезентативными примерами вариантов пептидов согласно изобретению. Также представлена репрезентативная обратная аминокислотная последовательность (пептид 246) и репрезентативная рандомизированная аминокислотная последовательность пептида (пептид 232) согласно изобретению. Обратная и рандомизированная аминокислотные последовательности, представленные в таблицах, не рассматриваются в настоящей заявке как репрезентативные последовательности.
Таблица 1 содержит список пептидов согласно изобретению и их соответствующих аминокислотных последовательностей и соответствующих SEQ ID NOS.


Аминокислотная последовательность пептида, представленного в таблице 1, может быть химически модифицирована. Так, например, если аминокислотная последовательность пептида, представленного в таблице 1, химически модифицирована у N-концевого амина с образованием амида карбоновой кислоты, то такой пептид иногда обозначается здесь в виде комбинации обозначений карбоновой кислоты, представленных в виде префикса, соединенного с номером пептида дефисом. Так, например, в случае пептида 79, N-концевой миристоилированный пептид 79 может иногда называться здесь «миристоилированным пептидом 79» или «mir-пептидом 79»; N-концевой ацетилированный пептид 79 может иногда называться здесь «ацетил-пептидом 79» или «Ac-пептидом 79». Циклический вариант пептида 79 может называться «циклическим пептидом 79» или цик-пептидом 79». Кроме того, например, если аминокислотная последовательность пептида, представленного в таблице 1, химически модифицирована у С-концевой карбоксильной группы, например, амином, таким как аммиак, с образованием С-концевого амида, то такой пептид иногда обозначают здесь в виде комбинаций обозначений аминокислотного остатка, представленных в виде суффикса, соединенного дефисом с номером пептида. Так, например, С-концевой амид пептида 79 иногда может обозначаться как «пептид-NH2». Если N-концевой амин пептида (например, пептида 79) химически модифицирован, например, миристоильной группой, а С-концевая карбоксильная группа химически модифицирована, например, аммиачной группой с образованием описанного выше амида, то такой пептид иногда может обозначаться с использованием префикса и суффикса, например «mir-пептид 79-NH2».
Настоящее изобретение относится к пептидам, имеющим аминокислотные последовательности, содержащие менее чем 24 аминокислоты, и аминокислотные последовательности, родственные аминокислотной последовательности пептида MANS (то есть пептид MANS представляет собой миристоил-пептид 1, а эталонная аминокислотная последовательность пептида MANS из 24 аминокислот представляет собой пептид 1). Пептиды согласно изобретению состоят из аминокислотных последовательностей, содержащих менее чем 24 аминокислоты, и могут состоять из 8-14, 10-12, 9-14, 9-13, 10-13, 10-14, по меньшей мере 9, по меньшей мере 10 аминокислот или т.п. Пептидами обычно являются пептиды с прямыми цепями, но ими также могут быть и циклические пептиды. Кроме того, указанными пептидами могут быть выделенные пептиды.
Что касается пептида 1 (SEQ ID NO:1), то есть эталонной последовательности из 24 аминокислот, то сегмент эталонной аминокислотной последовательности из 23 смежных аминокислот иногда называется здесь 23-мером. Аналогичным образом, сегмент эталонной последовательности из 22 смежных аминокислот иногда называется здесь 22-мером; аминокислотная последовательность из 21 аминокислоты называется 21-мером; аминокислотная последовательность из 20 аминокислот называется 20-мером; аминокислотная последовательность из 19 аминокислот называется 19-мером; аминокислотная последовательность из 18 аминокислот называется 18-мером; аминокислотная последовательность из 17 аминокислот называется 17-мером; аминокислотная последовательность из 16 аминокислот называется 16-мером; аминокислотная последовательность из 15 аминокислот называется 15-мером; аминокислотная последовательность из 14 аминокислот называется 14-мером; аминокислотная последовательность из 13 аминокислот называется 13-мером; аминокислотная последовательность из 12 аминокислот называется 12-мером; аминокислотная последовательность из 11 аминокислот называется 11-мером; аминокислотная последовательность из 10 аминокислот называется 10-мером; аминокислотная последовательность из 9 аминокислот называется 9-мером; аминокислотная последовательность из 8 аминокислот называется 8-мером; аминокислотная последовательность из 7 аминокислот называется 7-мером; аминокислотная последовательность из 6 аминокислот называется 6-мером; аминокислотная последовательность из 5 аминокислот называется 5-мером, а аминокислотная последовательность из 4 аминокислот называется 4-мером. В одном из аспектов изобретения, любая из этих «4-23-мерных» аминокислотных последовательностей, которые сами являются пептидами (иногда обозначаемыми здесь H2N-пептид-COOH), может быть независимо химически модифицирована, например, путем введения химической модификации, которая может быть выбрана из группы, состоящей из (i) образования амида у N-концевой аминогруппы (H2N-пептид-), например, под действием Cl- или предпочтительно C2 (уксусной кислоты) - C22-карбоновой кислоты; (ii) образования амида у С-концевой карбоксильной группы (пептид-СООН-), например, под действием аммиака или первичного или вторичного Cl-C22-амина; и (iii) их комбинаций.
Такие пептиды имеют аминокислотную последовательность, выбранную из группы, состоящей из (a) аминокислотной последовательности, имеющей 4-23 смежных аминокислот эталонной последовательности, пептида 1; (b) последовательности, по существу, идентичной последовательности, определенной в (a); и (c) варианта аминокислотной последовательности, определенной в (a), где указанный вариант выбран из группы, состоящей из варианта с заменами, варианта с делециями, варианта с добавлениями и их комбинаций. В некоторых вариантах изобретения, указанные пептиды имеют аминокислотную последовательность, выбранную из группы, состоящей из: (a) аминокислотной последовательности, имеющей 8-14 смежных аминокислот эталонной последовательности, пептида 1; (b) аминокислотной последовательности, по существу, идентичной последовательности, определенной в (a); и (c) варианта аминокислотной последовательности, определенной в (a), где указанный вариант выбран из группы, состоящей из варианта с заменами, варианта с делециями, варианта с добавлениями и их комбинаций. В других вариантах изобретения, указанные пептиды имеют аминокислотную последовательность, выбранную из группы, состоящей из: (a) аминокислотной последовательности, имеющей 10-12 смежных аминокислот эталонной последовательности, пептида 1; (b) аминокислотной последовательности, по существу, идентичной аминокислотной последовательности, определенной в (a); и (c) варианта аминокислотной последовательности, определенной в (a), где указанный вариант выбран из группы, состоящей из варианта с заменами, варианта с делециями, варианта с добавлениями и их комбинаций. В других вариантах изобретения, указанные пептиды имеют аминокислотную последовательность, содержащую по меньшей мере 9, по меньшей мере 10, 9-14, 9-13, 10-13, 10-14 или т.п. смежных аминокислот эталонной последовательности, пептида 1; аминокислотной последовательности, по существу, идентичной этой последовательности, или их вариантов, где указанный вариант выбран из группы, состоящей из варианта с заменами, варианта с делециями, варианта с добавлениями и их комбинаций. Как подробно объясняется ниже, одна или несколько аминокислот пептидов (например, N-концевых и/или C-концевых аминокислот) могут быть, но необязательно, независимо химически модифицированы; в некоторых вариантах изобретения, одна или несколько аминокислот пептида могут быть химически модифицированы, а в других вариантах изобретения, ни одна из аминокислот пептида не является химически модифицированной. В одном из аспектов изобретения, предпочтительная модификация может присутствовать у аминогруппы (-NH2) N-концевой аминокислоты пептида или пептидного сегмента (где указанная аминогруппа образует пептидную амидную связь в том случае, если она присутствует внутри пептидной последовательности, а не в N-концевом положении). В другом аспекте изобретения, предпочтительная модификация может присутствовать у карбоксигруппы (-СООН) С-концевой аминокислоты пептида или пептидного сегмента (где указанная карбоксигруппа образует пептидную амидную связь в том случае, если она присутствует внутри пептидной последовательности, а не в С-концевом положении). В другом аспекте изобретения, предпочтительная модификация может присутствовать как у N-концевой аминогруппы (NH2), так и у С-концевой карбоксильной группы (-СООН).
В некоторых вариантах изобретения, аминокислотная последовательность пептида начинается с N-концевой аминокислоты эталонной последовательности пептида 1. Так, например, такие пептиды могут иметь аминокислотную последовательность, выбранную из группы, состоящей из (а) аминокислотной последовательности, имеющей 4-23 смежные аминокислоты эталонной последовательности пептида 1, где указанная аминокислотная последовательность начинается с N-концевой аминокислоты эталонной последовательности (то есть пептида 2, пептида 4, пептида 7, пептида 11, пептида 16, пептида 22, пептида 29, пептида 37, пептида 46, пептида 56, пептида 67, пептида 79, пептида 92, пептида 106, пептида 121, пептида 137, пептида 154, пептида 172, пептида 191 или пептида 211); (b) последовательности, по существу, аналогичной аминокислотной последовательности, определенной в (a); и (c) варианта аминокислотной последовательности, определенной в (a). Эти пептиды не содержат химической группы или содержат химическую группу у N-концевого глицина, не являющуюся миристоильной группой. Предпочтительной химической группой является ацильная группа, представленная в форме амидной связи, такая как ацетильная группа, или алкильная группа.
В других вариантах изобретения, аминокислотная последовательность пептида заканчивается у С-концевой аминокислоты эталонной последовательности пептида 1. Так, например, такие пептиды могут иметь аминокислотную последовательность, выбранную из группы, состоящей из (а) аминокислотной последовательности, имеющей 4-23 смежных аминокислоты эталонной последовательности пептида 1, где указанная аминокислотная последовательность заканчивается у С-концевой аминокислоты эталонной последовательности (то есть пептида 3, пептида 6, пептида 10, пептида 15, пептида 21, пептида 28, пептида 36, пептида 45, пептида 55, пептида 66, пептида 78, пептида 91, пептида 105, пептида 120, пептида 136, пептида 153, пептида 171, пептида 190, пептида 210 или пептида 231); (b) последовательности, по существу, аналогичной аминокислотной последовательности, определенной в (a); и (c) варианта аминокислотной последовательности, определенной в (a).
В других вариантах изобретения, аминокислотная последовательность пептида начинается не с N-концевой аминокислоты эталонной последовательности пептида 1 (SEQ ID NO:1), а с положения аминокислоты от 2 до 21 эталонной последовательности пептида 1. Так, например, эти пептиды могут иметь аминокислотную последовательность, выбранную из группы, состоящей из (a) аминокислотной последовательности, имеющей 4-23 смежных аминокислоты эталонной последовательности пептида 1, где указанная аминокислотная последовательность начинается с любого положения аминокислоты от 2 до 21 эталонной последовательности, где указанные пептиды могут иметь длину из 4-23 смежных аминокислот и могут представлять собой пептиды, присутствующие в середине эталонной последовательности, пептида 1; (b) последовательности, по существу, аналогичной аминокислотной последовательности, определенной в (a); и (c) варианта аминокислотной последовательности, определенной в (a). Такие пептиды описаны в таблицах 1 или 2. Эти пептиды могут не содержать ковалентно связанной химической группы или содержат химическую группу у N-концевой аминокислоты, которая не является N-концевым глицином или аминокислотой, эквивалентной N-концевому глицину аминокислотной последовательности SEQ ID NO:1. Предпочтительной химической группой является ацильная группа, такая как ацетильная группа или миристоильная группа, представленная в форме амидной связи, или алкильная группа.
Пептидными аминокислотными последовательностями, которые могут быть использованы в настоящем изобретении для ингибирования гиперсекреции муцина у млекопитающего, и которые могут быть использованы для снижения уровня гиперсекреции муцина млекопитающего, а также в способах ингибирования гиперсекреции муцина и в способах снижения уровня гиперсекреции муцина, являются аминокислотные последовательности выделенных пептидов и аминокислотные последовательности пептидов, которые содержат, но необязательно, N-концевые и/или С-концевые химически модифицированные группы согласно изобретению, где указанные пептидные аминокислотные последовательности выбраны из группы, состоящей из 23-меров (то есть пептидов, имеющих последовательность из 23 аминокислот): пептида 2 и пептида 3; 22-меров (то есть пептидов, имеющих последовательность из 22 аминокислот): пептида 4, пептида 5 и пептида 6; 21-меров (то есть пептидов, имеющих последовательность из 21 аминокислоты); пептида 7, пептида 8, пептида 9 и пептида 10; 20-меров (то есть пептидов, имеющих последовательность из 20 аминокислот): пептида 11, пептида 12, пептида 13, пептида 14 и пептида 15; 19-меров (то есть пептидов, имеющих последовательность из 19 аминокислот): пептида 16, пептида 17, пептида 18, пептида 19, пептида 20 и пептида 21; 18-меров (то есть пептидов, имеющих последовательность из 18 аминокислот): пептида 22, пептида 23, пептида 25, пептида 26, пептида 27 и пептида 28; 17-меров (то есть пептидов, имеющих последовательность из 17 аминокислот): пептида 29; пептида 30, пептида 31, пептида 32, пептида 33, пептида 34, пептида 35 и пептида 36; 16-меров (то есть пептидов, имеющих последовательность из 16 аминокислот): пептида 37, пептида 38, пептида 39, пептида 40, пептида 41, пептида 42, пептида 43, пептида 44 и пептида 45; 15-меров (то есть пептидов, имеющих последовательность из 15 аминокислот): пептида 46, пептида 47, пептида 48, пептида 49, пептида 50, пептида 51, пептида 52, пептида 53, пептида 54 и пептида 55; 14-меров (то есть пептидов, имеющих последовательность из 14 аминокислот): пептида 56, пептида 57, пептида 58, пептида 59, пептида 60, пептида 61, пептида 62, пептида 63, пептида 64, пептида 65 и пептида 66; 13-меров (то есть пептидов, имеющих последовательность из 13 аминокислот): пептида 67, пептида 68, пептида 69, пептида 70, пептида 71, пептида 72, пептида 73, пептида 74, пептида 75, пептида 76, пептида 77 и пептида 78; 12-меров (то есть пептидов, имеющих последовательность из 12 аминокислот): пептида 79, пептида 80, пептида 81, пептида 82, пептида 83, пептида 84, пептида 85, пептида 86, пептида 87, пептида 88, пептида 89, пептида 90 и пептида 91; 11-меров (то есть пептидов, имеющих последовательность из 11 аминокислот): пептида 92, пептида 93, пептида 94, пептида 95, пептида 96, пептида 97, пептида 98, пептида 99, пептида 100, пептида 101, пептида 102, пептида 103, пептида 104 и пептида 105; 10-меров (то есть пептидов, имеющих последовательность из 10 аминокислот): пептида 106, пептида 107, пептида 108, пептида 109, пептида 110, пептида 111, пептида 112, пептида 113, пептида 114, пептида 115, пептида 116, пептида 117, пептида 118, пептида 119 и пептида 120; 9-меров (то есть пептидов, имеющих последовательность из 9 аминокислот): пептида 121, пептида 122, пептида 123, пептида 124, пептида 125, пептида 126, пептида 127, пептида 128, пептида 129, пептида 130, пептида 131, пептида 132, пептида 133, пептида 134, пептида 135 и пептида 136; 8-меров (то есть пептидов, имеющих последовательность из 8 аминокислот): пептида 137, пептида 138, пептида 139, пептида 140, пептида 141, пептида 142, пептида 143, пептида 144, пептида 145, пептида 146, пептида 147, пептида 148, пептида 149, пептида 150, пептида 151, пептида 152 и пептида 153; 7-меров (то есть пептидов, имеющих последовательность из 7 аминокислот): пептида 154, пептида 155, пептида 156, пептида 157, пептида 158, пептида 159, пептида 160, пептида 161, пептида 162, пептида 163, пептида 164, пептида 165, пептида 166, пептида 167, пептида 168, пептида 169, пептида 170 и пептида 171; 6-меров (то есть пептидов, имеющих последовательность из 6 аминокислот): пептида 172, пептида 173, пептида 174, пептида 175, пептида 176, пептида 177, пептида 178, пептида 179, пептида 180, пептида 181, пептида 182, пептида 183, пептида 184, пептида 185, пептида 186, пептида 187, пептида 188, пептида 189 и пептида 190; 5-меров (то есть пептидов, имеющих последовательность из 5 аминокислот): пептида 191, пептида 192, пептида 193, пептида 194, пептида 195, пептида 196, пептида 197, пептида 198, пептида 199, пептида 200, пептида 201, пептида 202, пептида 203, пептида 204, пептида 205, пептида 206, пептида 207, пептида 208, пептида 209 и пептида 210; и 4-меров (то есть пептидов, имеющих последовательность из 4 аминокислот): пептида 211, пептида 212, пептида 213, пептида 214, пептида 215, пептида 216, пептида 217, пептида 218, пептида 219, пептида 220, пептида 221, пептида 222, пептида 223, пептида 224, пептида 225, пептида 226, пептида 227, пептида 228, пептида 229, пептида 230 и пептида 231.
Предпочтительные аминокислотные последовательности выделенных пептидов и химически модифицированных по N- и/или С-концам пептидов согласно изобретению выбраны из группы, состоящей из 23-меров: пептида 2 и пептида 3; 22-меров: пептида 4, пептида 5 и пептида 6; 21-меров: пептида 7, пептида 8, пептида 9 и пептида 10; 20-меров: пептида 11, пептида 12, пептида 13, пептида 14 и пептида 15; 19-меров: пептида 16, пептида 17, пептида 18, пептида 19, пептида 20 и пептида 21; 18-меров: пептида 22, пептида 23, пептида 24, пептида 25, пептида 26, пептида 27 и пептида 28; 17-меров: пептида 29, пептида 30, пептида 31, пептида 32, пептида 33, пептида 34, пептида 35: и пептида 36; 16-меров: пептида 37, пептида 38, пептида 39, пептида 40, пептида 41, пептида 42, пептида 43, пептида 44 и пептида 45; 15-меров: пептида 46, пептида 47, пептида 48, пептида 49, пептида 50, пептида 51, пептида 52, пептида 53 и пептида 54; 14-меров: пептида 56, пептида 57, пептида 58, пептида 59, пептида 60, пептида 61, пептида 62, пептида 63 и пептида 64: 13-меров: пептида 67, пептида 68, пептида 69, пептида 70, пептида 71, пептида 72, пептида 73, пептида 74 и пептида 75; 12-меров: пептида 79, пептида 80, пептида 81, пептида 82, пептида 83, пептида 84, пептида 85, пептида 86 и пептида 87; 11-меров: пептида 92, пептида 93, пептида 94, пептида 95, пептида 96, пептида 97, пептида 98, пептида 99 и пептида 100; 10-меров: пептида 106, пептида 107, пептида 108, пептида 109, пептида 110, пептида 111, пептида 112, пептида 113 и пептида 114; 9-меров: пептида 122, пептида 123, пептида 124, пептида 125, пептида 126, пептида 127, пептида 128 и пептида 129; 8-меров: пептида 139, пептида 140, пептида 141, пептида 142, пептида 143, пептида 144 и пептида 145; 7-меров: пептида 157, пептида 158, пептида 159, пептида 160, пептида 161 и пептида 162; 6-меров: пептида 176, пептида 177, пептида 178, пептида 179 и пептида 180; 5-меров: пептида 196, пептида 197, пептида 198 и пептида 199; и 4-меров: пептида 217 и пептида 219.
Более предпочтительные аминокислотные последовательности выделенных пептидов и химически модифицированных по N- и/или С-концам пептидов согласно изобретению выбраны из группы, состоящей из 23-меров: пептида 2 и пептида 3; 22-меров: пептида 4, пептида 5 и пептида 6; 21-меров: пептида 7, пептида 8, пептида 9 и пептида 10; 20-меров: пептида 11, пептида 12, пептида 13, пептида 14 и пептида 15; 19-меров: пептида 16, пептида 17, пептида 18, пептида 19, пептида 20 и пептида 21; 18-меров: пептида 22, пептида 23, пептида 24, пептида 25, пептида 26, пептида 27 и пептида 28; 17-меров: пептида 29, пептида 30, пептида 31, пептида 32, пептида 33, пептида 34, пептида 35 и пептида 36; 16-меров: пептида 37, пептида 38, пептида 39, пептида 40: пептида 41, пептида 42, пептида 43, пептида 44 и пептида 45; 15-меров: пептида 46, пептида 47, пептида 48, пептида 49, пептида 50, пептида 51, пептида 52, пептида 53 и пептида 54; 14-меров: пептида 56, пептида 57, пептида 58, пептида 59, пептида 60, пептида 61, пептида 62, пептида 63 и пептида 64; 13-меров: пептида 67, пептида 68, пептида 69, пептида 70, пептида 71, пептида 72, пептида 73, пептида 74, пептида 80, пептида 81, пептида 82, пептида 83, пептида 84, пептида 85, пептида 86 и пептида 87; 11-меров: пептида 92, пептида 93, пептида 94, пептида 95, пептида 96, пептида 97, пептида 98, пептида 99 и пептида 100; 10-меров: пептида 106, пептида 108, пептида 109, пептида 110, пептида 111, пептида 112, пептида 113 и пептида 114; 9-меров: пептида 124, пептида 125, пептида 126, пептида 127, пептида 128 и пептида 129; 8-меров: пептида 141, пептида 142, пептида 143, пептида 144 и пептида 145; 7-меров: пептида 159, пептида 160, пептида 161 и пептида 162; 6-меров: пептида 178, пептида 179 и пептида 180; 5-меров: пептида 198 и пептида 199; и 4-мера: пептида 219.
В других вариантах изобретения, аминокислотная последовательность пептида включает смежные остатки A,K,G и E эталонной последовательности пептида 1 как в пептиде 219. Так, например, такие пептиды могут иметь аминокислотную последовательность, выбранную из группы, состоящей из: (a) аминокислотной последовательности, имеющей 4-23 смежных аминокислоты эталонной последовательности пептида 1, где указанная аминокислотная последовательность пептида включает смежные остатки A,K,G и E эталонного пептида 1 как в пептиде 219 (например, в пептиде 219, пептиде 45, пептиде 79, пептиде 67, пептиде 80 и т.п.); (b) последовательности, по существу, аналогичной аминокислотной последовательности, определенной в (a); и (c) варианта аминокислотной последовательности, определенной в (a).
Примерами пептидных сегментов, содержащих аминокислотную последовательность AKGE эталонной пептидной аминокислотной последовательности, пептида 1, являются (a): 23-меры: пептид 2 и пептид 3; 22-меры: пептид 4, пептид 5 и пептид 6; 11-меры: пептид 7, пептид 8, пептид 9 и пептид 10; 20-меры: пептид 11, пептид 12, пептид 13, пептид 14 и пептид 15; 19-меры: пептид 16, пептид 17, пептид 18, пептид 19, пептид 20 и пептид 21; 18- меры: пептид 22, пептид 23, пептид 24, пептид 25, пептид 26, пептид 27 и пептид 28; 17-меры: пептид 29, пептид 30, пептид 31, пептид 32, пептид 33, пептид 34, пептид 35 и пептид 36; 16-меры: пептид 37, пептид 38, пептид 39, пептид 40, пептид 41, пептид 42, пептид 43, пептид 44 и пептид 45; 15-меры: пептид 46, пептид 47, пептид 48, пептид 49, пептид 50, пептид 51, пептид 52, пептид 53 и пептид 54; 14-меры: пептид 56, пептид 57, пептид 58, пептид 59, пептид 60, пептид 61, пептид 62, пептид 63 и пептид 64; 13-меры: пептид 67, пептид 68, пептид 69, пептид 70, пептид 71, пептид 72, пептид 73, пептид 74 и пептид 75; 12-меры: пептид 79, пептид 80, пептид 81, пептид 82, пептид 83, пептид 84, пептид 85, пептид 86 и пептид 87; 11-меры: пептид 93, пептид 94, пептид 95, пептид 96, пептид 97, пептид 98, пептид 99 и пептид 100; 10-меры: пептид 108, пептид 109, пептид 110, пептид 111, пептид 112, пептид 113 и пептид 114; 9-меры: пептид 124, пептид 125, пептид 126, пептид 127, пептид 128 и пептид 129; 8-меры: пептид 141, пептид 142, пептид 143, пептид 144 и пептид 145; 7-меры: пептид 159, пептид 160, пептид 161 и пептид 162; 6-меры: пептид 178, пептид 179 и пептид 180; 5-меры: пептид 198 и пептид 199 и 4-мер: пептид 219, (b) последовательность, по существу, аналогичная аминокислотной последовательности, определенной в (a); и (c) вариант аминокислотной последовательности, определенной в (a), где указанный вариант выбран из группы, состоящей из варианта с заменами, варианта с делециями, варианта с добавлениями и их комбинаций, и где указанный сегмент содержит 4-23 смежных аминокислоты или состоит из этих аминокислот.
В другом варианте изобретения, предпочтительные пептидные последовательности имеют аминокислотную последовательность, выбранную из группы, состоящей из (a) аминокислотной последовательности, имеющей 10-23 смежных аминокислоты эталонной последовательности, пептида 1; (b) последовательности, по существу, аналогичной аминокислотной последовательности, определенной в (а), и (с) варианта аминокислотной последовательности, определенной в (а), где указанный вариант выбран из группы, состоящей из варианта с заменами, варианта с делециями и варианта с добавлениями и их комбинаций, где предпочтительные аминокислотные последовательности включают 23-мер: пептид 2; 22-мер: пептид 4; 21-мер: пептид 7; 20-мер: пептид 11; 19-мер: пептид 16; 18-мер: пептид 22; 17-мер: пептид 29; 16-мер: пептид 37; 15-мер: пептид 46; 14-мер: пептид 56; 13-мер: пептид 67; 12-мер: пептид 79; 11-мер: пептид 92; и 10-мер: пептид 106.
В других вариантах изобретения, аминокислотная последовательность пептида начинается с N-концевой аминокислоты эталонной последовательности пептида 1 и включает смежные остатки A, K, G и E эталонной последовательности пептида 1 как в пептиде 219, а в других вариантах изобретения, аминокислотная последовательность пептида заканчивается у С-концевой аминокислоты эталонной последовательности пептида 1 и включает смежные остатки A, K, G и E эталонной последовательности пептида 1 как в пептиде 219.
Указанные пептиды могут включать одну или несколько аминокислотных модификаций, а именно делеций, замен и/или добавлений, введенных в эталонную аминокислотную последовательность. Предпочтительными заменами могут быть консервативные аминокислотные замены, либо такими заменами могут быть неконсервативные аминокислотные замены. В некоторых вариантах изобретения, пептиды, включая пептиды, аминокислотные последовательности которых, по существу, идентичны эталонной аминокислотной последовательности или их варианты, не имеют делеций или добавлений, в отличие от соответствующих смежных аминокислот эталонной аминокислотной последовательности, но могут иметь консервативные или не консервативные замены. Аминокислотными заменами, которые могут быть введены в эталонную аминокислотную последовательность пептидов согласно изобретению, являются, но не ограничиваются ими, замены, где: аланин (A) может быть заменен лизином (K), валином (V), лейцином (L) или изолейцином (I); глутаминовая кислота (E) может быть заменена аспарагиновой кислотой (D); глицин (G) может быть заменен пролином (P); лизин (K) может быть заменен аргинином (R), глутамином (Q) или аспарагином (N); фенилаланин (F) может быть заменен лейцином (L), валином (V), изолейцином (I) или аланином (A); пролин (P) может быть заменен глицином (G); глутамин (Q) может быть заменен глутаминовой кислотой (E) или аспарагином (N); аргинин (R) может быть заменен лизином (K), глутамином (Q) или аспарагином (N); серин (S) может быть заменен треонином; треонин (T) может быть заменен серином (S), а валин (V) может быть заменен лейцином (L), изолейцином (I), метионином (M), фенилаланином (F), аланином (A) или норлейцином (Nle). Так, например, заменами, которые могут быть введены в эталонную аминокислотную последовательность пептидов согласно изобретению, являются замена фенилаланина (F) аланином (А) (например, в положении 4 аминокислоты эталонной аминокислотной последовательности), замена глутамина (Q) глутаминовой кислотой (Е) (например, в положении 3 аминокислоты эталонной аминокислотной последовательности), замена аланина (А) лизином (K) (например, в положениях 2 и/или 8 аминокислот эталонной аминокислотной последовательности), и/или замена треонина (Т) серином (S) (например, в положении 7 аминокислоты эталонной аминокислотной последовательности).
Если в аминокислотные последовательности пептидов согласно изобретению вводят замены (где пептиды являются немодифицированными, а также представляют собой пептиды, которые были химически модифицированы, например, введением N-концевой и/или С-концевой модификации посредством образования амида), то предпочтительно, чтобы при сравнении с эталонной аминокислотной последовательностью идентичность между аминокислотной последовательностью указанного пептида и эталонной аминокислотной последовательностью составляла по меньшей мере 80%. Пептиды, имеющие 5-23 аминокислоты и включающие одну аминокислотную замену, примерно на 80-96% (то есть ~95,7%) идентичны эталонной аминокислотной последовательности. Пептиды, имеющие 10-23 аминокислоты и включающие одну аминокислотную замену, примерно на 90-96% (то есть ~95,7%) идентичны эталонной аминокислотной последовательности. Пептиды, имеющие 20-23 аминокислоты и включающие одну аминокислотную замену, примерно на 95-96% (то есть ~95,7%) идентичны эталонной аминокислотной последовательности. Пептиды, имеющие 10-23 аминокислоты и включающие две аминокислотные замены, примерно на 80-92% (то есть ~91,3%) идентичны эталонной аминокислотной последовательности. Пептиды, имеющие 16-23 аминокислоты и включающие две аминокислотные замены, примерно на 87,5-92% (то есть ~91,3%) идентичны эталонной аминокислотной последовательности. Пептиды, имеющие 20-23 аминокислоты и включающие две аминокислотные замены, примерно на 90-92% (то есть ~91,3%) идентичны эталонной аминокислотной последовательности. Пептиды, имеющие 15-23 аминокислоты и включающие три аминокислотные замены, примерно на 80-87% идентичны эталонной аминокислотной последовательности. Пептиды, имеющие 20-23 аминокислоты и включающие три аминокислотные замены, примерно на 85-87% идентичны эталонной аминокислотной последовательности. Пептиды, имеющие 20-23 аминокислоты и включающие четыре аминокислотные замены, примерно на 80-83% (то есть ~82,6%) идентичны эталонной аминокислотной последовательности.
В пептидах согласно изобретению, а именно в аминокислотной последовательности, состоящей из смежных аминокислот эталонного пептида (который является 24-мером), замена одной аминокислоты в аминокислотной последовательности, состоящей из смежных 23 аминокислот (23-мера), выбранных из 24 аминокислот эталонной аминокислотной последовательности, приводит к получению пептида, аминокислотная последовательность которого на 95,65% (или ~96%) идентична аминокислотному сегменту эталонного пептида, которому идентичен указанный 23-мер. Аналогичным образом, замена двух, трех, четырех и пяти аминокислот в указанном 23-мере приводит к получению пептида с аминокислотной последовательностью, которая на 91,30% (или ~91%), 86,96% (или ~87%), 82,61% (или ~83%) и 78,27% (или ~78%) идентична, соответственно, аминокислотной последовательности эталонного пептида. Аналогичным образом, замена одной, двух, трех, четырех и пяти аминокислот в указанном 22-мере приводит к получению пептида с аминокислотной последовательностью, которая на 95,45% (или ~95%), 90,91% (или ~91%), 86,36% (или ~86%), 81,82% (или ~82%) и 77,27% (или ~77%), идентична, соответственно, аминокислотной последовательности эталонного пептида. Аналогичным образом, замена одной, двух, трех, четырех и пяти аминокислот в указанном 21-мере приводит к получению пептида с аминокислотной последовательностью, которая на 95,24% (~95%), 90,48% (~91%), 85,71% (~86%), 80,95% (~81%) и 76,19% (~76%) идентична, соответственно, аминокислотной последовательности эталонного пептида. Аналогичным образом, замена одной, двух, трех, четырех и пяти аминокислот в указанном 20-мере приводит к получению пептида с аминокислотной последовательностью, которая на 95,00% (95%), 90,00% (90%), 85,00% (85%), 80,00% (80%) и 75,00% (75%) идентична, соответственно, аминокислотной последовательности эталонного пептида. Аналогичным образом, замена одной, двух, трех и четырех аминокислот в указанном 19-мере приводит к получению пептида с аминокислотной последовательностью, которая на 94,74% (~95%), 89,47% (~89%), 84,21% (~84%) и 78,95% (~79%) идентична, соответственно, аминокислотной последовательности эталонного пептида. Аналогичным образом, замена одной, двух, трех и четырех аминокислот в указанном 18-мере приводит к получению пептида с аминокислотной последовательностью, которая на 94,44% (~94%), 88,89% (~89%), 83,33% (~83%) и 77,78% (~78%) идентична, соответственно, аминокислотной последовательности эталонного пептида. Аналогичным образом, замена одной, двух, трех и четырех аминокислот в указанном 17-мере приводит к получению пептида с аминокислотной последовательностью, которая на 94,12% (~94%), 88,23% (~88%), 82,35% (~82%) и 76,47% (~76%) идентична, соответственно, аминокислотной последовательности эталонного пептида. Аналогичным образом, замена одной, двух, трех и четырех аминокислот в указанном 16-мере приводит к получению пептида с аминокислотной последовательностью, которая на 93,75% (~94%), 87,50% (~88%), 81,25% (~81%) и 75,00% (75%) идентична, соответственно, аминокислотной последовательности эталонного пептида. Аналогичным образом, замена одной, двух и трех аминокислот в указанном 15-мере приводит к получению пептида с аминокислотной последовательностью, которая на 93,33% (~93%), 86,67% (~87%) и 80,00% (80%) идентична, соответственно, аминокислотной последовательности эталонного пептида. Аналогичным образом, замена одной, двух и трех аминокислот в указанном 14-мере приводит к получению пептида с аминокислотной последовательностью, которая на 92,86% (~93%), 85,71% (~86%), и 78,57% (79%) идентична, соответственно, аминокислотной последовательности эталонного пептида. Аналогичным образом, замена одной, двух и трех аминокислот в указанном 13-мере приводит к получению пептида с аминокислотной последовательностью, которая на 92,31% (~92%), 84,62% (~85%), и 76,92% (~77%) идентична, соответственно, аминокислотной последовательности эталонного пептида. Аналогичным образом, замена одной, двух и трех аминокислот в указанном 12-мере приводит к получению пептида с аминокислотной последовательностью, которая на 91,67% (~92%), 83,33% (~83%) и 75,00% (75%) идентична, соответственно, аминокислотной последовательности эталонного пептида. Аналогичным образом, замена одной и двух аминокислот в указанном 11-мере приводит к получению пептида с аминокислотной последовательностью, которая на 90,91% (~91%) и 81,82% (~82%) идентична, соответственно, аминокислотной последовательности эталонного пептида. Аналогичным образом, замена одной и двух аминокислот в указанном 10-мере приводит к получению пептида с аминокислотной последовательностью, которая на 90,00% (90%) и 80,00% (80%) идентична, соответственно, аминокислотной последовательности эталонного пептида. Аналогичным образом, замена одной и двух аминокислот в указанном 9-мере приводит к получению пептида с аминокислотной последовательностью, которая на 88, 89% (~89%) и 77, 78% (~78%) идентична, соответственно, аминокислотной последовательности эталонного пептида. Аналогичным образом, замена одной и двух аминокислот в указанном 8-мере приводит к получению пептида с аминокислотной последовательностью, которая на 87,50% (~88%) и 75,00% (75%) идентична, соответственно, аминокислотной последовательности эталонного пептида. Аналогичным образом, замена одной аминокислоты в указанном 7-мере, 6-мере, 5-мере и 4-мере приводит к получению пептида с аминокислотной последовательностью, которая на 85,71% (~86%), 83,33% (~83,3%), 80,00% (80%) и 75,00% (75%) идентична, соответственно, аминокислотной последовательности эталонного пептида. Аминокислотные последовательности согласно изобретению, предпочтительно, более чем на 80%, более предпочтительно на 81-96%, а еще более предпочтительно, на 80-96% идентичны аминокислотной эталонной последовательности. Предпочтительно, аминокислотные последовательности могут быть, но необязательно, химически связаны у N-концевой аминогруппы пептида с линейной C2-C22-алифатической группой карбоновой кислоты, более предпочтительно, с линейной С2-С16-алифатической группой карбоновой кислоты, а наиболее предпочтительно, с линейной С2- или С16-алифатической группой карбоновой кислоты посредством амидной связи и необязательно, химически связаны у С-концевой группы карбоновой кислоты пептида с амином, таким как аммиак, или с первичным или вторичным амином, таким как С1-С16-алифатический первичный амин с прямой цепью, посредством амидной связи.
Примерами вариантов пептида 79 с заменами, а именно 12-мера, являются, например, пептид 238, где аминокислота Q, присутствующая в положении 3 в пептиде 79, в последовательности 238 была заменена на E; пептид 233, где аминокислота A, присутствующая в положении 2 в пептиде 79, в пептиде 233 была заменена на K; пептид 234, где аминокислота A, присутствующая в положении 8 в пептиде 79, в пептиде 234 была заменена на К; пептид 235, где аминокислота A, присутствующая в положениях 2 и 8 в пептиде 79, в пептиде 235 была заменена на K; пептид 237, где аминокислота F, присутствующая в положении 4 в пептиде 79, в пептиде 237 была заменена на A; пептид 239, где аминокислота K, присутствующая в положении 10 в пептиде 79, в пептиде 239 была заменена на A; пептид 240, где аминокислота G, присутствующая в положении 11 в пептиде 79, в пептиде 240 была заменена на A; и пептид 241, где аминокислота Е, присутствующая в положении 12 в пептиде 79, в пептиде 241 была заменена на A.
Примерами вариантов пептида 106 с заменами, а именно 10-мера, являются, например, пептид 236, где аминокислота F, присутствующая в положении 4 в пептиде 106, в пептиде 236 была заменена на A; пептид 242, где аминокислота G, присутствующая в положении 1 в пептиде 106, в пептиде 237 была заменена на A; пептид 243, где аминокислота Q, присутствующая в положении 3 в пептиде 106, в пептиде 243 была заменена на A; пептид 244, где аминокислота S, присутствующая в положении 5 в пептиде 106, в пептиде 244 была заменена на A; пептид 245, где аминокислота K, присутствующая в положении 6 в пептиде 106, в пептиде 245 была заменена на A; пептид 247, где аминокислота T, присутствующая в положении 7 в пептиде 106, в пептиде 247 была заменена на A; пептид 248, где аминокислота K, присутствующая в положении 10 в пептиде 106, в пептиде 248 была заменена на A; пептид 249, где аминокислота K, присутствующая в положениях 6 и 10 в пептиде 106, в пептиде 249 была заменена на A;
Примером варианта пептида 137 с заменами, а именно 8-мера, является, например, пептид 250, где аминокислота F, присутствующая в положении 4 в пептиде 137, в пептиде 250 была заменена на A.
Примером варианта пептида 219 с заменами, а именно 4-мера, является, например, пептид 251, где аминокислота K, присутствующая в положении 2 в пептиде 219, в пептиде 251 была заменена на A.
Вариант пептида с заменами, такой как вариант, описанный в настоящей заявке, может быть получен в виде выделенного пептида или в виде химически модифицированного пептида, такого как, например, пептид с N-концевым амидом, таким как миристоил-амид, ацетил-амид и т.п., как описано в настоящей заявке, и пептид с С-концевым амидом, таким как амид, образованный аммиаком, а также пептид с N-концевым и C-концевым амидом.
Если в аминокислотные последовательности пептидов согласно изобретению вносят делеции, то при сравнении с эталонной аминокислотной последовательностью предпочтительно, чтобы идентичность между аминокислотной последовательностью указанного пептида и эталонной аминокислотной последовательностью составляла по меньшей мере 80%. Пептиды, имеющие 5-23 аминокислоты и включающие одну аминокислотную делецию, примерно на 80-96% (то есть ~95,7%) идентичны эталонной аминокислотной последовательности. Пептиды, имеющие 10-23 аминокислоты и включающие одну аминокислотную делецию, примерно на 90-96% (то есть ~95,7%) идентичны эталонной аминокислотной последовательности. Пептиды, имеющие 20-23 аминокислоты и включающие одну аминокислотную делецию, примерно на 95-96% (то есть ~95,7%) идентичны эталонной аминокислотной последовательности. Пептиды, имеющие 10-23 аминокислоты и включающие две аминокислотные делеции, примерно на 80-92% (то есть ~91,3%) идентичны эталонной аминокислотной последовательности. Пептиды, имеющие 16-23 аминокислоты и включающие две аминокислотные делеции, примерно на 87,5-92% (то есть ~91,3%) идентичны эталонной аминокислотной последовательности. Пептиды, имеющие 20-23 аминокислоты и включающие две аминокислотные делеции, примерно на 90-92% (то есть ~91,3%) идентичны эталонной аминокислотной последовательности. Пептиды, имеющие 15-23 аминокислоты и включающие три аминокислотные делеции, примерно на 80-87% идентичны эталонной аминокислотной последовательности. Пептиды, имеющие 20-23 аминокислоты и включающие три аминокислотные делеции, примерно на 85-87% идентичны эталонной аминокислотной последовательности. Пептиды, имеющие 20-23 аминокислоты и включающие четыре аминокислотные делеции, примерно на 80-83% (то есть ~82,6%) идентичны эталонной аминокислотной последовательности.
Как указывалось выше, одна или несколько аминокислот пептидов могут быть также химически модифицированными. Аминокислоты пептидов могут быть подвергнуты любым модификациям, известным специалистам, с применением любого известного метода.
В некоторых вариантах изобретения, N-концевая и/или C-концевая аминокислота может быть модифицирована. Так, например, альфа-N-концевая аминокислота пептидов может быть алкилирована, амидирована или ацилирована у альфа-N-концевой (N-концевой) амино(альфа-H2N-)-группы и, например, C-концевая аминокислота указанных пептидов может быть амидирована или этерифицирована у C-концевой карбоксильной (-COOH) группы. Так, например, N-концевая аминогруппа может быть модифицирована путем ацилирования с образованием амида, в результате чего она будет содержать любую ацильную или жирную ацильную группу, включая ацетильную группу (то есть CH3-C(=O)- или миристоильную группу, которые являются предпочтительными группами с точки зрения настоящего изобретения). В некоторых вариантах изобретения, N-концевая аминогруппа может быть модифицирована так, что она будет содержать ацильную группу, имеющую формулу -C(O)R, где R представляет собой прямую или разветвленную алкильную группу, имеющую от 1 до 15 атомов углерода, либо она может быть модифицирована так, что она будет содержать ацильную группу, имеющую формулу -C(O)R1, где R1 представляет собой прямую алкильную группу, имеющую от 1 до 15 атомов углерода. N-амид может также представлять собой формамид (R=H). C-концевая аминокислота пептидов может быть также химически модифицированной. Так, например, C-концевая карбоксильная группа C-концевой аминокислоты может быть химически модифицирована путем превращения карбоксильной группы в карбоксамидную группу (то есть она может быть амидирована). В некоторых вариантах изобретения, N-концевые и/или C-концевые аминокислоты не являются химически модифицированными. В некоторых вариантах изобретения, N-концевая группа является модифицированной, а С-концевая группа не является модифицированной. В некоторых вариантах изобретения, модифицированными являются как N-концевая, так и С-концевая группы.
Пептид может быть ацилирован у аминогруппы N-концевой аминокислоты с образованием N-концевого амида посредством взаимодействия с кислотой, выбранной из группы, состоящей из:
(i-a) C2(ацетил)-C13алифатической (насыщенной или необязательно ненасыщенной) карбоновой кислоты (например, N-концевого амида, образованного уксусной кислотой (которая является предпочтительной группой), пропановой кислотой, бутановой кислотой, гексановой кислотой, октановой кислотой, декановой кислотой, додекановой кислотой), которая может иметь прямую или разветвленную цепь (более, чем С3), либо она может содержать кольцо (более, чем С3);
(i-b) насыщенной C14алифатической карбоновой кислоты, которая может иметь прямую или разветвленную цепь, либо она может содержать кольцо;
(i-c) ненасыщенной C14алифатической карбоновой кислоты, которая может иметь прямую или разветвленную цепь, либо она может содержать кольцо;
(i-d) C15-C24алифатической (насыщенной или необязательно ненасыщенной) карбоновой кислоты, которая может иметь прямую или разветвленную цепь, либо она может содержать кольцо (например, с тетрадекановой кислотой (миристиновой кислотой, которая является предпочтительной группой), с гексадекановой кислотой, с 9-гексадеценовой кислотой, с октадекановой кислотой, 9-октадеценовой кислотой, с 11-гексадеценовой кислотой, с 9,12- октадекадиеновой кислотой, с 9,12,15-октадекатриеновой кислотой, 6,9,12- октадекатриеновой кислотой, с эйкозановой кислотой, с 9-эйкозеновой кислотой, с 5,8,11,14-эйкозатетраеновой кислотой, с 5,8,11,14,17-эйкозапентаеновой кислотой, с докозановой кислотой, с 13-докозеновой кислотой, с 4,7,10,13,16,19-докозагексаеновой кислотой, с тетракозановой кислотой и т.п.);
(ii) трифторуксусной кислоты;
(iii) бензойной кислоты; и
(iv-a) Cl-C12алифатической алкилсульфоновой кислоты, которая образует алифатический алкилсульфонамид, где структура Cl-C12алифатической алкильной цепи сульфоновой кислоты аналогична структуре алифатической алкильной цепи карбоновой кислоты в описанных выше алифатических алкилкарбоновых кислотах. Так, например, пептид может быть ацилирован с использованием группы карбоновой кислоты, представленной формулой (С1-С11)алкил-C(O)OH, посредством реакции дегидратации путем активации группы карбоновой кислоты с образованием амида, представленного формулой (С1-С11)алкил-C(O)-NH-пептид. Аналогичным образом, сульфонамид может быть получен посредством взаимодействия группы сульфоновой кислоты (представленной формулой (С1-С12)алкил-S(O2)-X, например, где X представляет собой галоген или OCH3 или другую совместимую уходящую группу) с N-концевой аминогруппой с образованием сульфонамида, представленного формулой (С1-С12)алкил-S(O2)-NH-пептид;
(iv-b) Cl4-C24алифатической алкилсульфоновой кислоты, которая образует алифатический алкилсульфонамид, где структура Cl4-C24алифатической алкильной цепи сульфоновой кислоты аналогична структуре алифатической алкильной цепи карбоновой кислоты в описанных выше алифатических алкилкарбоновых кислотах. Так, например, пептид может быть ацилирован с использованием группы карбоновой кислоты, представленной формулой (С13-С23)алкил-C(O)OH, посредством реакции дегидратации путем активации группы карбоновой кислоты с образованием амида, представленного формулой (С13-С23)алкил-C(O)-NH-пептид. Аналогичным образом, сульфонамид может быть получен посредством взаимодействия группы сульфоновой кислоты (представленной формулой (С14-С24)алкил-S(O2)-X, например, где X представляет собой галоген или OCH3 или другую совместимую уходящую группу) с N-концевой аминогруппой с образованием сульфонамида, представленного формулой (С14-С24)алкил-S(O2)-NH-пептид.
В другом примере, N-концевая аминогруппа N-концевой аминокислоты может быть алкилирована С1-С12алифатической алкильной группой, структура которой описана выше. Алкилирование может быть осуществлено, например, с использованием алифатического алкилгалогенида или алифатического алкилового эфира сульфоновой кислоты (мезилата, тозилата и т.п.), а предпочтительно, с использованием первичного алкилгалогенида или первичного алкилового эфира сульфоновой кислоты. N-концевая аминокислота может быть также модифицирована у концевой аминогруппы так, чтобы она содержала в качестве амида любую ацильную или алифатическую ацильную или жирную ацильную группу, включая ацетильную группу (то есть -C(O)CH3, которая является предпочтительной группой), миристоильную группу (которая является предпочтительной группой), бутаноильную группу, гексаноильную группу, октаноильную группу, деканоильную группу, додеканоильную группу, тетрадеканоильную группу, гексадеканоильную группу, 9-гексадеценоильную группу, октадеканоильную группу, 9-октадеценоильную группу, 11-октадеценоильную группу, 9,12-октадекадиеноильную группу, 9,12,15-октадекатриеноильную группу, 6,9,12-октадекатриеноильную группу, эйкозаноильную группу, 9-эйкозеноильную группу, 5,8,11,14-эйкозатетраеноильную группу, 5,8,11,14,17-эйкозапентаеноильную группу, докозаноильную группу, 13-докозеноильную группу, 4,7,10,13,16,19-докозагексаеноильную группу и тетракозаноильную группу, где указанные группы ковалентно связаны с концевой аминогруппой пептида амидной связью.
С-концевая группа карбоновой кислоты С-концевой аминокислоты пептидов согласно изобретению может быть также химически модифицирована. Так, например, С-концевая аминокислота может быть химически модифицирована посредством взаимодействия С-концевой группы карбоновой кислоты пептида с амином с образованием амидной группы, такой как амид аммиака, который является предпочтительной группой; амид С1-С12алифатического алкиламина, а предпочтительно, алифатического алкиламина с прямой цепью; амид гидроксил-замещенного С1-С12алифатического алкиламина; амид 2-С1-С12алифатической алкилоксиэтиламиногруппы с прямой цепью и амид омега-метокси-поли(этиленокси)n-этиламиногруппы (также называемой омега-метокси-ПЭГ-альфа-аминогруппой или омега-метокси-(полиэтиленгликоль)аминогруппы), где n равно 0-10. C-концевая группа карбоновой кислоты C-концевой аминокислоты пептида может также присутствовать в форме сложного эфира, выбранного из группы, состоящей из сложного эфира С1-С12алифатического алкилового спирта и сложного эфира 2-(омега-метокси-поли(этиленокси)n-этанола (МПЭГ), где n равно 0-10. В одном из аспектов изобретения, полиэтиленгликолевый компонент, такой как компонент, присутствующий в сложном эфире ПЭГ, в сложном эфире МПЭГ, в амиде ПЭГ, в амиде МПЭГ и т.п., предпочтительно, имеет молекулярную массу примерно от 500 до 40000 дальтон, более предпочтительно, от 1000 до 25000 дальтон, а наиболее предпочтительно, примерно от 1000 до 10000 дальтон.
C-концевая группа карбоновой кислоты на пептиде, который может быть представлен формулой пептид-C(O)OH, может быть также амидирована путем ее превращения в активированную форму, такую как галогенид карбоновой кислоты, ангидрид карбоновой кислоты, N-гидроксисукцинимидоэфир, пентафторфениловый (OPfp) эфир, сложный эфир 3-гидрокси-2,3-дигидро-4-оксо-бензотриазона (ODhbt) и т.п., для облегчения взаимодействия с аммиаком или с первичным или вторичным амином, а предпочтительно, с аммиаком или первичным амином, при этом, предпочтительно, чтобы любые другие реакционноспособные группы пептида были защищены химически синтезированными совместимыми защитными группами, хорошо известными специалистам в области пептидного синтеза, а в частности твердофазного пептидного синтеза, и такими группами являются бензиловый эфир, трет-бутиловый эфир, фениловый эфир и т.п. Амид полученного пептида может быть представлен формулой пептид-C(O)-NR3R4 (амид, присутствующий у С-конца пептида), где R3 и R4 независимо выбраны из группы, состоящей из водорода, С1-С12алкила, такого как метил, этил, бутил, изобутил, циклопропилметил, гексил, додецил и, необязательно, высшего алкила, например С14-С24алкила, такого как тетрадецил и т.п., описанные выше.
C-концевая карбоновая кислота С-концевой аминокислоты может быть также превращена в амид гидроксил-замещенного C2-C12алифатического алкиламина (где гидроксильная группа присоединена к атому углерода, а не к атому азота амина), такого как 2-гидроксиэтиламин, 4-гидроксибутиламин и 12-гидроксидодециламин и т.п.
C-концевая карбоновая кислота может быть также превращена в амид гидрокси-замещенного C2-С12алифатического алкиламина, где гидроксильная группа может быть ацилирована с образованием сложного эфира C2-С12алифатической карбоновой кислоты, описанного выше. Предпочтительно, в амиде пептида, у С-конца указанного пептида, представленного формулой пептид-C(O)NR5R6, R5 представляет собой водород, а R6 выбран из группы, состоящей из водорода, С1-С12алкила и гидроксил-замещенного C2-С12алкила.
C-концевая карбоновая кислота С-концевой аминокислоты может быть также превращена в амид 2-(C2-С12)алифатического алкилоксиэтиламина с прямой цепью. Такой амид может быть получен, например, посредством взаимодействия линейного C2-С12 алифатического спирта с гидридом калия в диглиме с 2-хлорэтанолом, с образованием C2-С12 алифатического алкилэтанола с прямой цепью, который может быть превращен в амин путем окисления альдегидом с последующим восстановительным аминированием до амина (например, с использованием аммиака) или путем превращения в алкилгалогенид (например, с использованием тионилхлорида) с последующей обработкой амином, таким как аммиак.
C-концевая карбоновая кислота С-концевой аминокислоты может быть также превращена в амид линейного ПЭГ-амина (например, омега-гидрокси-ПЭГ-альфа-амина, омега-(C2-С12)-ПЭГ-альфа-амина, такого как омега-метокси-ПЭГ-альфа-амин, то есть МПЭГ-амин). В одном из аспектов изобретения, полиэтиленгликолевый компонент или ПЭГ предпочтительно, имеют молекулярную массу примерно от 500 до 40000 дальтон, более предпочтительно, от 1000 до 25000 дальтон, а наиболее предпочтительно, примерно от 1000 до 10000 дальтон.
C-концевая карбоновая кислота С-концевой аминокислоты может быть также превращена в амид омега-метокси-поли(этиленокси)n-этиламина, где n равно 0-10, который может быть получен из соответствующего омега-метокси-поли(этиленокси)n-этанола, например, путем превращения спирта в амин, как описано выше.
В другом варианте изобретения, C-концевой карбоксил может быть превращен в амид, представленный формулой пептид-C(O)-NR7R8, где R7 представляет собой водород, а R8 представляет собой линейную 2-(C1-С12)алифатическую алкилоксиэтильную группу, где указанная C1-С12алифатическая алкильная группа определена выше и включает такие группы, как метоксиэтил (то есть CH3O-СН2СН2), 2-додецилоксиэтил и т.п., или R7 представляет собой водород, а R8 представляет собой омега-метокси-поли(этиленокси)n-этильную группу, где n поли(этиленокси)групп составляет 0-10, и где такими группами являются 2-метоксиэтил (то есть CH3O-СН2СН2), омега-метоксиэтоксиэтил (то есть CH3O-CH2CH2O-CH2CH2 - CH3O-(CH2CH2O)10-CH2CH2-).
C-концевая группа карбоновой кислоты C-концевой аминокислоты пептида может также присутствовать в форме сложного эфира C1-С12 алифатического алкилового спирта, где указанная алифатическая алкильная группа спирта определена выше. C-концевая группа карбоновой кислоты С-концевой аминокислоты пептида может также присутствовать в форме сложного эфира 2-(омега-метокси-поли(этиленокси)n)этанола, где n равно 0-10, который может быть получен посредством взаимодействия 2-метоксиэтанола, такого как 2-метоксиэтанолат натрия, со стехиометрическими количествами этиленоксида, где указанное стехиометрическое количество зависит от числа n.
Боковая цепь аминокислоты пептидов может быть также химически модифицированной. Так, например, фенильная группа фенилаланина или тирозина может быть замещена заместителем, выбранным из группы, состоящей из:
C1-С24алифатической алкильной группы (то есть прямой или разветвленной группы, и/или насыщенной или ненасыщенной группы, и/или группы, содержащей циклическую группу), такой как метил (предпочтительный), этил, пропил, изопропил, бутил, изобутил, циклопропил, 2-метилциклопропил, циклогексил, октил, децил, додецил, гексадецил, октадецил, эйкозанил, докозанил, тетракозанил, 9-гексадеценил, 9-октадеценил, 11-октадеценил, 9,12-октадекадиенил, 9,12,15-октадекатриенил, 6,9,12-окстадекатриенил, 9-эйкозенил, 5,8,11,14-эйкозатетраенил, 5,8,11,14,17-эйкозапентаенил, 13-докозенил и 4,7,10,13,16,19- докозагексаенил;
C1-С12алифатической алкильной группы, замещенной гидроксильной группой по меньшей мере у одного атома углерода, находящегося на определенном расстоянии от ненасыщенного положения, и примерами таких гидроксиалкильных групп являются гидроксиметил, гидроксиэтил, гидроксидодецил и т.п.;
C1-С12 алкильной группы, замещенной гидроксильной группой, которая является этерифицированной C2-С25алифатической карбоксильной группой кислоты, такой как уксусная кислота, бутановая кислота, гексановая кислота, октановая кислота, декановая кислота, додекановая кислота, тетрадекановая кислота, гексадекановая кислота, 9-гексадеценовая кислота, октадекановая кислота, 9-октадеценовая кислота, 11-октадеценовая кислота, 9,12-октадекадиеновая кислота, 9,12,15-октадекатриеновая кислота, 6,9,12-октадекатриеновая кислота, эйкозановая кислота, 9-эйкозеновая кислота, 5,8,11,14-эйкозатетраеновая кислота, 5,8,11,14,17-эйкозапентаеновая кислота, докозановая кислота, 13-докозеновая кислота, 4,7,10,13,16,19-докозагексаеновая кислота, тетракозановая кислота и т.п., дикарбоновая кислота, такая как янтарная кислота, или оксикислота, такая как молочная кислота, где общее число атомов углерода сложноэфирного заместителя составляет 3-25;
галогена, такого как фтор, хлор, бром и иод;
нитро;
амино, такого как NH2, метиламино, диметиламино;
трифторметила;
карбоксила (-СООН);
С1-С24алкокси (например, алкокси, который может быть образован путем алкилирования тирозина), такого как метокси, этокси, пропилокси, изопропилокси, бутилокси, изобутилокси, циклопропилокси, 2-метоксициклопропилокси, циклогексилокси, октилокси, децилокси, додецилокси, гексадецилокси, октадецилокси, эйкозанилокси, докозанилокси, тетракозанилокси, 9-гексадеценилокси, 9-октадеценилокси, 11-октадеценилокси, 9,12-октадекадиенилокси, 9,12,15-октадекатриенилокси, 6,9,12-октадекатриенилокси, 9-эйкозенилокси, 5,8,11,14-эйкозатетраенилокси, 5,8,11,14,17-эйкозапентаенилокси, 13-докозенилокси и 4,7,10,13,16,19-докозагексаенилокси; и
C2-C12гидроксиалкилокси, такого как 2-гидроксиэтилокси и его сложных эфиров карбоновых кислот, описанных выше, или сложных эфиров трифторуксусной кислоты.
Гидроксильная группа серина может быть этерифицирована заместителем, выбранным из группы, состоящей из:
C2-C12алифатической группы карбоновой кислоты, описанной выше;
группы трифторуксусной кислоты; и
группы бензойной кислоты.
Эпсилон-аминогруппа лизина может быть химически модифицирована, например, путем образования амида посредством взаимодействия с C2-C12алифатической группой карбоновой кислоты (например, посредством взаимодействия амина с химически активированной формой карбоновой кислоты, такой как хлорангидрид, ангидрид, N-гидроксисукцинимидоэфир, пентафторфениловый (OPfp) эфир, сложный эфир 3-гидрокси-2,3-дигидро-4-оксо-бензотриазона (ODhbt) и т.п.), например, описанной выше, или с группой бензойной кислоты или с аминокислотной группой. Кроме того, эпсилон-аминогруппа лизина может быть химически модифицирована путем алкилирования одной или двумя С1-С4алифатическими алкильными группами.
Группа карбоновой кислоты в глутаминовой кислоте может быть модифицирована посредством образования амида путем взаимодействия с амином, таким как аммиак, первичный C1-С12алифатический алкиламин (алкильная группа которого определена выше), включая метиламин, или с аминогруппой аминокислоты.
Группа карбоновой кислоты в глутаминовой кислоте может быть модифицирована посредством образования сложного эфира путем взаимодействия с C2-С12алифатической гидроксиалкильной группой, описанной выше, а предпочтительно, с первичным C1-С12алифатическим алкиловым спиртом, таким как метанол, этанол, пропан-1-ол, н-додеканол и т.п., описанными выше.
В своем предпочтительном варианте, настоящее изобретение относится к способу ингибирования высвобождения по меньшей мере одного медиатора воспаления из гранул по меньшей мере одной воспалительной клетки в ткани и/или в физиологической жидкости индивидуума, где указанный способ включает введение в указанную ткань и/или физиологическую жидкость терапевтически эффективного количества фармацевтической композиции, содержащей по меньшей мере один пептид, имеющий аминокислотную последовательность, выбранную из группы, состоящей из:
(a) аминокислотной последовательности, имеющей 4-23 смежных аминокислоты эталонной последовательности, GAQFSKTAAKGEAAAERPGEAAVA (SEQ ID NO:1);
(b) аминокислотной последовательности, имеющей последовательность GAQFSKTAAKGEAAAERPGEAAVA (SEQ ID NO:1); и
(c) аминокислотной последовательности, по существу, идентичной последовательности, определенной в (a), где С-концевая аминокислота пептида является, но необязательно, независимо химически модифицированной, а N-концевая аминокислота пептида либо является независимо химически модифицированной посредством ацилирования карбоновой кислотой, выбранной из группы, состоящей из C2-С13насыщенной или ненасыщенной алифатической карбоновой кислоты, С14насыщенной (миристиновой кислоты) или ненасыщенной алифатической карбоновой кислоты, C15-С24насыщенной или ненасыщенной алифатической карбоновой кислоты, и трифторуксусной кислоты, либо не является химически модифицированной; при условии, что указанный пептид может быть модифицирован путем ацилирования, если его аминокислотная последовательность начинается с последовательности GAQF эталонной последовательности, то есть путем ацилирования только карбоновой кислотой, выбранной из группы, состоящей из C2-С13насыщенной или ненасыщенной алифатической карбоновой кислоты, С14ненасыщенной алифатической карбоновой кислоты, C15-С24насыщенной или ненасыщенной алифатической карбоновой кислоты и трифторуксусной кислоты, либо кислотой, которая не является химически модифицированной, где указанный пептид объединен, но необязательно, с фармацевтически приемлемым носителем и присутствует в количестве, которое является терапевтически эффективным для ослабления высвобождения медиаторов воспаления и тем самым для снижения уровня высвобождения указанного медиатора воспаления по меньшей мере из одной воспалительной клетки по сравнению с уровнем высвобождения указанного медиатора воспаления по меньшей мере из одной воспалительной клетки того же типа, но в отсутствие по меньшей мере одного пептида.
В этом способе, предпочтительно, используется пептид, который может быть ацетилирован у альфа-N-концевой аминокислоты. Этот пептид может состоять по меньшей мере из десяти смежных аминокислотных остатков, а предпочтительно, представляет собой ацетил-пептид 106 (SEQ ID NO:106).
В этом способе также используется пептид, состоящий по меньшей мере из четырех смежных аминокислотных остатков, а более предпочтительно, по меньшей мере из шести смежных аминокислотных остатков. Кроме того, такой пептид может быть миристоилирован у альфа-N-концевой аминокислоты указанного пептида. В таком способе может быть также использован пептид, который может быть амидирован аммиаком у альфа-С-концевой аминокислоты.
В способе согласно другому варианту изобретения используется пептид, который содержит аминокислотную последовательность, выбранную из (a) аминокислотной последовательности, имеющей 4-23 смежных аминокислоты эталонной последовательности GAQFSKTAAKGEAAAERPGEAAVA (SEQ ID NO:1), где N-концевая аминокислота аминокислотной последовательности (a) выбрана из аминокислоты, присутствующей в положениях 2-21 эталонной последовательности, GAQFSKTAAKGEAAAERPGEAAVA (SEQ ID NO:1). Кроме того, эти пептиды могут быть миристоилированы у альфа-N-концевой аминокислоты и могут быть также амидированы аммиаком у альфа-С-концевой аминокислоты.
Способ введения согласно изобретению направлен на снижение уровня высвобождения медиатора воспаления по механизму блокирования или ингибирования высвобождения медиатора воспаления из воспалительной клетки у указанного индивидуума.
Указанный способ включает введение в описанные пептиды фармацевтически приемлемого носителя или смешивание этих пептидов с указанным носителем с получением фармацевтической композиции.
Способ введения согласно изобретению направлен на снижение уровня высвобождения по меньшей мере одного медиатора воспаления и тем самым на снижение уровня высвобождения указанного медиатора воспаления по меньшей мере из одной воспалительной клетки по сравнению с уровнем высвобождения указанного медиатора воспаления по меньшей мере из одной воспалительной клетки того же типа, но в отсутствие по меньшей мере одного указанного пептида. Воспалительной клеткой указанного индивидуума может быть лейкоцит, гранулоцит, базофил, эозинофил, моноцит, макрофаг или их комбинации.
Медиатор воспаления, высвобождаемый по меньшей мере из одной гранулы по меньшей мере одной воспалительной клетки, выбран из группы, состоящей из миелопероксидазы (MPO), пероксидазы эозинофилов (EPO), основного мажорного белка [MBP], лизоцима, гранзима, гистамина, протеогликана, протеазы, хемотаксического фактора, цитокина, метаболита арахидоновой кислоты, дефензина, белка, повышающего проницаемость бактерий (BPI), эластазы, катепсина G, катепсина B, катепсина D, бета-D- глюкуронидазы, альфа-маннозидазы, фосфолипазы A2, хондроитин-4-сульфата, протеиназы 3, лактоферрина, коллагеназы, активатора комплемента, рецептора комплемента, рецептора N-формилметионил-лейцил-фенилаланина (FMLP), рецептора ламинина, цитохрома b558, моноцитарного-хемотаксического фактора, гистаминазы, белка, связывающегося с витамином B12, желатиназы, активатора плазминогена, бета-D-глюкуронидазы, и их комбинаций. Предпочтительный медиатор воспаления выбран из группы, состоящей из миелопероксидазы (MPO), пероксидазы эозинофилов (EPO), основного мажорного белка (MBP), лизоцима, гранзима и их комбинаций.
Способ по п.13 формулы изобретения, где эффективное количество указанного пептида, способствующее снижению уровня высвобождения медиатора воспаления, включает ингибирующее дегрануляцию количество пептида, которое снижает уровень медиатора воспаления, высвобождаемого по меньшей мере из одной воспалительной клетки, примерно на 1%-99%, или предпочтительно, примерно на 5-50% - 99%, по сравнению с количеством, высвобождаемым по меньшей мере из одной воспалительной клетки в отсутствие указанного пептида.
Способ согласно изобретению может быть применен для лечения индивидуума, страдающего респираторным заболеванием. Таким респираторным заболеванием может быть астма, хронический бронхит, хроническая обструктивная болезнь легких (ХОБЛ) и кистозный фиброз. Предпочтительными индивидуумами, которые могут быть подвергнуты лечению в соответствии с настоящим изобретением, являются млекопитающие, такие как человек, собаки, лошади и кошки.
Способом введения пептидов согласно изобретению является местное введение, парентеральное введение, ректальное введение, внутрилегочное введение, интраназальное введение и пероральное введение. Более предпочтительно, внутрилегочное введение включает введение с помощью аэрозоля, которое может быть осуществлено с применением инсуффлятора, ингалятора с дозируемым клапаном или небулайзера. Кроме того, введение индивидууму может также включать введение второй молекулы, выбранной из группы, состоящей из антибиотика, противовирусного соединения, антипаразитарного соединения, противовоспалительного соединения и иммуномодулятора.
Указанный способ может быть также применен для лечения индивидуума, страдающего заболеванием, выбранным из группы, состоящей из кишечного заболевания, кожной болезни, аутоиммунного заболевания, болевого синдрома и их комбинаций. Более конкретно, указанное кишечное заболевание выбрано из группы, состоящей из язвенного колита, болезни Крона и синдрома раздраженного кишечника. Кожными болезнями, которые также могут быть подвергнуты лечению способом согласно изобретению, являются розацея, экзема, псориаз и угревая сыпь в тяжелой форме. Кроме того, индивидуум, страдающий ревматоидным артритом, также может быть подвергнут лечению способом согласно изобретению.
В одном из своих вариантов, настоящее изобретение включает введение пептидов, содержащих аминокислотную последовательность, по существу, идентичную аминокислотной последовательности: (a) имеющей 4-23 смежных аминокислот эталонной последовательности GAQFSKTAAKGEAAAERPGEAAVA (SEQ ID NO:1). Эти пептиды, предпочтительно, выбраны из группы, состоящей из SEQ ID NOS:233, 234, 235, 236, 237, 238, 239, 240, 241, 242, 243, 244, 245, 247, 248, 249, 250, 251 и 252. Указанные пептиды могут быть также ацетилированы по альфа-N-концевой аминокислоте или миристоилированы по альфа-N-концевой аминокислоте и необязательно амидированы аммиаком по альфа-С-концевой аминокислоте.
Способ согласно изобретению может быть также применен для снижения уровня гиперсекреции слизи у индивидуума путем введения описанных здесь пептидов согласно изобретению, а, также для снижения уровня MARCKS-ассоциированной гиперсекреции слизи по меньшей мере из одной слизь-секретирующей клетки или ткани индивидуума, так чтобы гиперсекреция слизи у указанного индивидуума снижалась по сравнению с гиперсекрецией слизи, наблюдаемой в отсутствие указанного пептида.
Настоящее изобретение относится к выделенному пептиду, имеющему аминокислотную последовательность, выбранную из группы, состоящей из:
(a) аминокислотной последовательности, имеющей 4-23 смежных аминокислот эталонной последовательности GAQFSKTAAKGEAAAERPGEAAVA (SEQ ID NO:1);
(b) аминокислотной последовательности, имеющей последовательность GAQFSKTAAKGEAAAERPGEAAVA (SEQ ID NO:1); и
(c) аминокислотной последовательности, по существу, идентичной последовательности, определенной в (a),
где С-концевая аминокислота пептида является, но необязательно, независимо химически модифицированной, а N-концевая аминокислота пептида либо является независимо химически модифицированной посредством ацилирования карбоновой кислотой, выбранной из группы, состоящей из C2-С13насыщенной или ненасыщенной алифатической карбоновой кислоты, С14насыщенной (миристиновой кислоты) или ненасыщенной алифатической карбоновой кислоты, C15-С24насыщенной или ненасыщенной алифатической карбоновой кислоты, и трифторуксусной кислоты, либо не является химически модифицированной, при условии, что указанный пептид модифицирован путем ацилирования, если его аминокислотная последовательность начинается с последовательности GAQF эталонной последовательности, то есть путем ацилирования только карбоновой кислотой, выбранной из группы, состоящей из C2-С13насыщенной или ненасыщенной алифатической карбоновой кислоты, С14ненасыщенной алифатической карбоновой кислоты, C15-С24насыщенной или ненасыщенной алифатической карбоновой кислоты и трифторуксусной кислоты, или кислотой, которая не является химически модифицированной, где указанный пептид необязательно объединен с фармацевтически приемлемым носителем и присутствует в количестве, которое является терапевтически эффективным для ослабления высвобождения медиаторов воспаления и тем самым для ослабления уровня высвобождения указанного медиатора воспаления по меньшей мере из одной воспалительной клетки по сравнению с уровнем высвобождения указанного медиатора воспаления по меньшей мере из одной воспалительной клетки того же типа, но в отсутствие по меньшей мере одного указанного пептида.
Выделенный пептид может быть ацетилирован у альфа-N-концевой аминокислоты. Этот выделенный пептид состоит по меньшей мере из десяти смежных аминокислотных остатков, а предпочтительно, представляет собой выделенный пептид, который состоит из ацетил-пептида 106 (SEQ ID NO:106).
В другом варианте изобретения, указанный пептид состоит по меньшей мере из четырех смежных аминокислотных остатков или по меньшей мере из шести смежных аминокислотных остатков.
Такой пептид может быть также миристоилирован у альфа-N-концевой аминокислоты и/или он может быть амидирован аммиаком у альфа-C-концевой аминокислоты.
Выделенный пептид может также содержать аминокислотную последовательность, описанную выше в (a) и имеющую 4-23 смежных аминокислоты эталонной последовательности GAQFSKTAAKGEAAAERPGEAAVA (SEQ ID NO:1), где N-концевая аминокислота аминокислотной последовательности (a) выбрана из аминокислот, присутствующих в положениях 2-21 эталонной последовательности, GAQFSKTAAKGEAAAERPGEAAVA (SEQ ID NO:1). Этот пептид может быть также миристоилирован или ацетилирован у альфа-N-концевой аминокислоты или, но необязательно, амидирован аммиаком у альфа-С-концевой аминокислоты.
В другом своем варианте, настоящее изобретение относится к выделенному пептиду, аминокислотная последовательность которого, по существу, идентична аминокислотной последовательности (a), имеющей 4-23 смежных аминокислот эталонной последовательности GAQFSKTAAKGEAAAERPGEAAVA (SEQ ID NO:1). Эти пептиды, предпочтительно, выбраны из группы, состоящей из SEQ ID NOS:233, 234, 235, 236, 237, 238, 239, 240, 241, 242, 243, 244, 245, 247, 248, 249, 250, 251 и 252. Указанные пептиды могут быть также ацетилированы у альфа-N-концевой аминокислоты или миристоилированы у альфа-N-концевой аминокислоты и необязательно амидированы аммиаком у альфа-С-концевой аминокислоты. Аминокислотная последовательность (с), по существу, идентичная аминокислотной последовательности (a), выбрана из группы, состоящей из SEQ ID NOS: 233, 234, 235, 236, 237, 238, 239, 240, 241, 242, 243, 244, 245, 247, 248, 249, 250, 251 и 252.
Настоящее изобретение также относится к композиции, включающей выделенный пептид, описанный в настоящей заявке, и наполнитель. Настоящее изобретение также относится к фармацевтической композиции, содержащей выделенный пептид, описанный в настоящей заявке, и фармацевтически приемлемый носитель. Фармацевтическая композиция может быть также, предпочтительно, стерильной, стерилизуемой или стерилизованной. Эти пептиды могут содержаться в наборе вместе с реагентами, подходящими для введения.
Краткое описание графического материала
На фиг.1A-1B проиллюстрировано, что PKC-зависимое фосфорилирование приводит к высвобождению MARCKS из плазматической мембраны в цитоплазму.
На фиг.2A-2C показано, что PKG индуцирует дефосфорилирование MARCKS под действием активирующего PP2A.
На фиг.3 представлены гистограммы, на которых проиллюстрировано, что PP2A является основным компонентом пути секреции муцина.
На фиг.4 показан гель, который иллюстрирует, что MARCKS ассоциирован с актином и миозином в цитоплазме.
На фиг.5 проиллюстрирован механизм передачи сигнала, регулирующего секрецию MPO в нейтрофилах.
На фиг.6 представлена гистограмма, иллюстрирующая способность пептида MANS блокировать секрецию миелопероксидазы из выделенных собачьих нейтрофилов.
На фиг.7 представлена гистограмма, иллюстрирующая способность пептида MANS блокировать секрецию миелопероксидазы из выделенных человеческих нейтрофилов.
На фиг.8 представлена гистограмма, указывающая на то, что PMA стимулирует небольшое увеличение уровня секреции MPO из ЛПС-стимулированных человеческих нейтрофилов, которое усиливается ко-стимуляцией под действием 8-Br-cGMP в зависимости от концентрации.
На фиг.9 представлена гистограмма, указывающая на то, что 8-Br-cGMP-стимуляция оказывает незначительное влияние на секрецию MPO из ЛПС-стимулированных человеческих нейтрофилов до тех пор, пока не будет достигаться костимуляция под действием PMA в зависимости от концентрации.
На фиг.10 представлена гистограмма, указывающая на то, что PMA стимулирует небольшое увеличение уровня секреции MPO из ЛПС-стимулированных собачьих нейтрофилов, которое усиливается костимуляцией под действием 8-Br-cGMP в зависимости от концентрации.
На фиг.11 представлена гистограмма, указывающая на то, что 8-Br-cGMP-стимуляция оказывает незначительное влияние на секрецию MPO из ЛПС-стимулированных собачьих нейтрофилов до тех пор, пока не будет достигаться костимуляция под действием PMA в зависимости от концентрации.
На фиг.12 представлена гистограмма, указывающая на то, что костимуляция под действием PMA+8-Br-cGMP необходима для максимальной секреции MPO из ЛПС-стимулированных собачьих нейтрофилов.
Подробное описание изобретения
Ниже приводится более подробное описание настоящего изобретения со ссылками на графический материал, на котором проиллюстрированы предпочтительные варианты осуществления изобретения. Однако настоящее изобретение может быть осуществлено в различных вариантах, которые не должны рассматриваться как ограничение объема изобретения. Напротив, эти варианты приводятся для детального и более полного описания изобретения и во всей своей полноте входят в объем настоящего изобретения.
Если это не оговорено особо, то все используемые здесь технические и научные термины имеют свои общепринятые значения, понятные среднему специалисту в области, к которой относится настоящее изобретение. Все публикации, патентные заявки, патенты и другие упомянутые здесь работы во всей своей полноте вводятся в настоящее описание посредством ссылки.
Настоящее изобретение относится к способу ингибирования экзоцитозного высвобождения по меньшей мере одного медиатора воспаления по меньшей мере из одной воспалительной клетки, где указанный способ включает контактирование по меньшей мере одной воспалительной клетки, содержащей по меньшей мере один медиатор воспаления, присутствующий во внутриклеточной везикуле, по меньшей мере с одним пептидом, выбранным из группы, состоящей из пептида MANS и его активного фрагмента в количестве, эффективном для снижения уровня высвобождения медиатора воспаления из воспалительной клетки, по сравнению с уровнем высвобождения медиатора воспаления из воспалительной клетки того же типа, наблюдаемым в отсутствие по меньшей мере одного пептида.
Настоящее изобретение также относится к способу ингибирования высвобождения по меньшей мере одного медиатора воспаления по меньшей мере из одной воспалительной клетки в ткани или физиологической жидкости индивидуума, где указанный способ включает введение в ткань и/или физиологическую жидкость индивидуума, содержащие по меньшей мере одну воспалительную клетку, включающую по меньшей мере один медиатор воспаления, присутствующий во внутриклеточной везикуле, терапевтически эффективного количества фармацевтической композиции, содержащей по меньшей мере один пептид, выбранный из группы, состоящей из пептида MANS и его активного фрагмента в количестве, терапевтически эффективном для снижения уровня высвобождения медиатора воспаления по меньшей мере из одной воспалительной клетки, по сравнению с уровнем высвобождения медиатора воспаления по меньшей мере из одной воспалительной клетки того же типа в отсутствие по меньшей мере одного пептида. Более конкретно, снижение уровня высвобождения медиатора воспаления включает блокирование или ингибирование механизма высвобождения медиатора воспаления из воспалительной клетки.
Настоящее изобретение относится к контактированию описанного в настоящей заявке пептида с любой известной воспалительной клеткой и/или к введению указанного пептида в указанную клетку, которая может присутствовать в ткани или в физиологической жидкости индивидуума, содержащих по меньшей мере один медиатор воспаления, присутствующий во внутриклеточной везикуле. Такой воспалительной клеткой, предпочтительно, является лейкоцит, а более предпочтительно, гранулоцит, который может быть также классифицирован как нейтрофил, базофил, эозинофил или их комбинации. Воспалительными клетками, подвергаемыми контакту способом согласно изобретению, могут быть также моноциты/макрофаги.
Настоящее изобретение относится к снижению уровня высвобождения медиаторов воспаления, содержащихся в везикулах воспалительных клеток, и такие медиаторы воспаления выбраны из группы, состоящей из миелопероксидазы (MPO), пероксидазы эозинофилов (EPO), основного мажорного белка (MBP), лизоцима, гранзима, гистамина, протеогликана, протеазы, хемотаксического фактора, цитокина, метаболита арахидоновой кислоты, дефензина, белка, повышающего проницаемость бактерий (BPI), эластазы, катепсина G, катепсина B, катепсина D, бета-D-глюкуронидазы, альфа-маннозидазы, фосфолипазы A2, хондроитин-4-сульфата, протеиназы 3, лактоферрина, коллагеназы, активатора компелмента, рецептора комплемента, рецептора N-формилметионил-лейцил-фенилаланина (FMLP), рецептора ламинина, цитохрома b558, моноцитарного-хемотаксического фактора, гистаминазы, белка, связывающегося с витамином B12, желатиназы, активатора плазминогена, бета-D-глюкуронидазы, и их комбинаций. Предпочтительные медиаторы воспаления выбраны из группы, состоящей из миелопероксидазы (MPO), пероксидазы эозинофилов (EPO), основного мажорного белка (MBP), лизоцима, гранзима и их комбинаций.
Настоящее изобретение относится к контактированию эффективного количества пептида с воспалительной клеткой, где указанное эффективное количество определено как ингибирующее дегрануляцию количество пептида MANS или его активного фрагмента, которое снижает уровень медиатора воспаления, высвобождаемого по меньшей мере из одной воспалительной клетки примерно на 1%-99% по сравнению с уровнем медиатора воспаления, высвобождаемого по меньшей мере из одной воспалительной клетки в отсутствие пептида MANS или его активного фрагмента. Такое количество также известно как количество, эффективное для снижения уровня высвобождения медиатора воспаления. Более предпочтительно, такое эффективное количество контактируемого пептида включает ингибирующее дегрануляцию количество пептида MANS или его активного фрагмента, которое снижает уровень медиатора воспаления, высвобождаемого по меньшей мере из одной воспалительной клетки примерно на 5-50% - 99%, по сравнению с уровнем медиатора воспаления, высвобождаемого по меньшей мере из одной воспалительной клетки в отсутствие пептида MANS или его активного фрагмента.
В одном из своих вариантов, настоящее изобретение относится к способу введения по меньшей мере одного пептида, содержащего пептид MANS и его активный фрагмент в терапевтически эффективном количестве, в ткань или в физиологическую жидкость индивидуума, страдающего респираторным заболеванием, которым, предпочтительно, является астма, хронических бронхит или ХОБЛ. В другом варианте изобретения, указанный индивидуум может страдать кишечным заболеванием, кожной болезнью, аутоиммунным заболеванием, болевым синдромом и их комбинациями. Указанным кишечным заболеванием может быть язвенный колит, болезнь Крона или синдром раздраженного кишечника. Индивидуум может страдать кожной болезнью, такой как розацея, экзема, псориаз или угревая сыпь в тяжелой форме. Индивидуум может также страдать артритом, таким как ревматоидный артрит, псориатический артрит и системная красная волчанка. Индивидуумы, страдающие кистозным фиброзом, могут быть также подвергнуты лечению способом согласно изобретению и лечению с использованием пептидов. Способ согласно изобретению, предпочтительно, может быть применен для лечения индивидуумов, таких как млекопитающие, а предпочтительно, человек, собаки, лошади и кошки.
В соответствии с настоящим изобретением, способом лечения индивидуумов является введение одного или нескольких пептидов, включая описанный здесь пептид MANS или его активный фрагмент, и такими способами введения являются местное введение, парентеральное введение, ректальное введение, внутрилегочное введение, интраназальное введение или пероральное введение. Более предпочтительно, внутрилегочное введение выбрано из группы, состоящей из введения с помощью аэрозоля, инсуффлятора, ингалятора с дозируемым клапаном и небулайзера. Кроме того, описанный здесь способ может также включать введение указанному индивидууму второй молекулы, выбранной из группы, состоящей из антибиотика, противовирусного соединения, антипаразитарного соединения, противовоспалительного соединения и иммуномодулятора.
В одном из своих аспектов, настоящее изобретение относится к способу введения фармацевтической композиции. Такая фармацевтическая композиция включает терапевтически эффективное количество известного соединения и фармацевтически приемлемый носитель. Используемый здесь термин «терапевтически эффективное» количество означает количество соединения, которое является достаточным для ослабления симптомов, наблюдаемых у индивидуума. Такое терапевтически эффективное количество может варьироваться в зависимости от возраста и физического состояния пациента, а также от тяжести состояния пациента, подвергаемого лечению, от продолжительности курса лечения, от типа любого одновременно проводимого лечения, от используемого фармацевтически приемлемого носителя и других факторов, известных или установленных специалистом в данной области. Фармацевтически приемлемыми препаратами, предпочтительно, являются твердые лекарственные формы, такие как таблетки или капсулы. Для перорального введения могут быть также использованы жидкие препараты, которые могут быть получены в форме сиропов или суспензий, например растворов, содержащих активный ингредиент, сахар и смесь этанола, воды, глицерина и пропиленгликоля. Если это необходимо, то такие жидкие препараты могут включать один или несколько из нижеследующих агентов, таких как красители, ароматизаторы и сахарин. Кроме того, могут быть также использованы загустители, такие как карбоксиметилцеллюлоза, а также другие приемлемые носители, выбор которых может быть осуществлен самим специалистом в данной области.
Как указывалось выше, настоящее изобретение относится к способам регуляции клеточных секреторных процессов, а в частности процессов высвобождения медиаторов воспаления из воспалительных клеток. Используемый здесь термин «регуляция» означает блокирование, ингибирование, снижение, ослабление, увеличение, усиление или стимуляцию. Ряд клеточных секреторных процессов включает высвобождение содержимого из мембраносвязанных везикул или гранул, находящихся в клетках. Мембраносвязанную везикулу или гранулу определяют как внутриклеточную частицу, которая является, главным образом, везикулярной (или везикулу, присутствующую в клетке) и содержит запасный материал, который может быть секретирован. Было обнаружено, что определенное содержимое этих везикул, например, присутствующих в воспалительных клетках, ответственно за развитие различных патологий в ряде тканей млекопитающих. Некоторыми эффектами такой секреции, очевидно, являются поражения ранее нормальной ткани в процессе воспаления. Настоящее изобретение относится к способу блокирования секреции из любой мембраносвязанной везикулы, включая везикулу, присутствующую в воспалительных клетках, путем нацеливания синтетического пептида на специфическую молекулу, играющую важную роль во внутриклеточном секреторном пути. Этот подход может иметь терапевтически важное значение для лечения состояний широкого ряда, ассоциированных с гиперсекрецией и воспалением у человека и животных.
Более конкретно, настоящее изобретение относится к воспалительным клеткам, которые содержат медиаторы воспаления в одной или нескольких гранулах или везикулах, присутствующих в цитоплазме клеток. Такие клетки подвергают контактированию с одним или несколькими пептидами, выбранными из пептида MANS или его активного фрагмента, каждый из которых подробно описан в настоящей заявке. Предпочтительно, контактирование такого пептида с воспалительной клеткой осуществляют путем введения этого пептида индивидууму, страдающему заболеванием, при котором такие воспалительные клетки находятся в конкретной ткани или в физиологической жидкости, присутствующей в данной ткани. После введения пептида в клетку или контактирования этого пептида с клеткой указанный пептид конкурирует за связывание или конкурентно ингибирует связывание нативного белка MARCKS с мембраной внутриклеточных гранул или везикул, содержащих медиаторы воспаления. В результате блокирования связывания белка MARCKS с везикулами в воспалительных клетках такие везикулы в этих клетках не транспортируются в плазматическую мембрану клеток, как это обычно происходит при стимуляции с последующим экзоцитозным высвобождением содержимого этих клеток, а именно медиаторов воспаления. Таким образом, способ согласно изобретению позволяет ингибировать транспорт везикул в плазматическую мембрану клеток, что, в свою очередь, приводит к снижению уровня высвобождения медиаторов воспаления из воспалительных клеток. Количество медиаторов воспаления, высвобождаемых из клеток в течение всего периода времени, снижается, поскольку скорость высвобождения и количество высвобождаемых медиаторов из воспалительных клеток зависит от концентрации пептида, вводимого в воспалительные клетки и контактируемого с этими клетками.
Преимущество настоящего изобретения заключается в том, что оно позволяет проводить комбинированную терапию, которая включает прямое блокирование секреции слизи с применением уникальной противовоспалительной терапии. Преимущество терапии согласно изобретению по сравнению с современной противовоспалительной терапией, которая приводит к общему подавлению иммунной системы, заключается в том, что пептид согласно изобретению, очевидно, блокирует секрецию только внутриклеточных компонентов, секретируемых из воспалительных клеток. Таким образом, многие аспекты иммунной системы должны функционировать даже при ингибировании медиаторов воспаления.
Соединения согласно изобретению могут регулировать, то есть блокировать высвобождение медиаторов воспаления из клеток. Такое ингибирование высвобождения медиаторов воспаления является привлекательным средством для предупреждения и лечения различных расстройств, например заболеваний и патологических состояний, ассоциированных с воспалением. Таким образом, соединения согласно изобретению могут быть использованы для лечения таких состояний. Указанными состояниями являются заболевания дыхательных путей и хронические воспалительные заболевания, включая, но не ограничиваясь ими, остеоартрит, рассеянный склероз, синдром Гийена-Барре, болезнь Крона, язвенный колит, псориаз, реакцию «трансплантат против хозяина» и системную красную волчанку. Соединения согласно изобретению могут быть также использованы для лечения других расстройств, ассоциированных с активностью повышенных уровней медиаторов провоспаления и ферментов, такой как ответы на различные инфекционные агенты, и различных аутоиммунных заболеваний, таких как ревматоидный артрит, синдром токсического шока, диабет и воспалительные кишечные заболевания.
Использование пептидов и применение способов согласно изобретению позволяет проводить лечение воспалений, а также терапию, которая основана на объединении противовоспалительной активности пептида и его способности блокировать секрецию слизи. Заболеваниями, которые могут быть подвергнуты лечению с применением способности пептида блокировать воспаление и секрецию слизи, являются, но не ограничивается ими, воспалительные кишечные заболевания, расстройства пищеварительной системы (то есть воспаление желчного пузыря, болезнь Менетье) и воспалительные заболевания дыхательных путей.
Другие провоспалительные медиаторы ассоциируются с рядом патологических состояний, при которых происходит приток нейтрофилов в области воспаления или поражения. Было продемонстрировано, что блокирующие антитела могут быть использованы для лечения ассоциированного с нейтрофилами поражения тканей при остром воспалении (Harada et al., 1996, Molecular Medicine Today 2, 482). Кроме нейтрофилов, другими клетками, которые могут высвобождать медиаторы воспаления, являются другие лейкоциты, такие как базофилы, эозинофилы, моноциты и лимфоциты, и в этом случае, терапия может быть направлена на подавление секреции медиаторов из этих клеток. Нейтрофилы, эозинофилы и базофилы принадлежат к типу гранулоцитов, то есть лейкоцитов, в цитоплазме которых присутствуют гранулы. Лейкоциты синтезируют ряд медиаторов воспаления, которые упаковываются в цитоплазматические гранулы и хранятся в этих гранулах. Такими медиаторами являются, среди прочих, например, миелопероксидаза [MPO] в нейтрофилах (Borregaard N, Cowland J.B. Granules of the human neutrophilic polymorphonuclear leukocyte. Blood 1997, 89:3503-3521), пероксидаза эозинофилов [EPO] и основный мажорный белок [MBP] в эозинофилах (Gleich G J. Mechanisms of eosinophil-associated inflammation. J Allergy Clin. Immunol. 2000, 105:651-663), лизоцим в моноцитах/макрофагах (Hoff T., Spencker T., Emmendoerffer A., Goppelt-Struebe M. Effects of glucocorticoids on the TPA-induced monocytic differentiation. J Leukoc. Biol. 1992, 52:173-182, Balboa M A, Saez Y, Balsinde J. Calcium-independent phospholipase A2 is required for lysozyme secretion in U937 promonocytes. J Immunol. 2003, 170:5276-5280), и гранзим в природных клетках-киллерах (NK) и в цитотоксических лимфоцитах (Bochan M.R., Goebel W.S., Brahmi Z. Stably transfected antisense granzyme B и perforin constructs inhibit human granule-mediated lytic ability. Cell Immunol 1995, 164:234-239, Gong J. H., Maki G., Klingemann H.G. Characterization of a human cell line (NK-92) with phenotypical и functional characteristics of activated natural killer cells. Leukemia 1994, 8:652-658, Maki G., Klingemann H.G., Martinson J.A., Tam Y.K. Factors regulating the cytotoxic activity of the human natural killer cell line, NK-92. J. Hematother Stem Cell Res. 2001, 10:369-383 и Takayama H, Trenn G, Sitkovsky M.V. A novel cytotoxic T lymphocyte activation assay. J. Immunol. Methods 1987, 104:183-1907-10). Эти медиаторы могут высвобождаться в области поражения и могут играть определенную роль в развитии воспаления и в репарации, например, в легких и в других органах, что обусловлено инфильтрацией этих клеток в область поражения или патологии ткани. Лейкоциты высвобождают эти гранулы по механизму экзоцитоза (Burgoyne R.D, Morgan A. Secretory granule exocytosis. Physiol Rev 2003, 83:581-632, Logan M.R, Odemuyiwa S.O, Moqbel R. Understanding exocytosis in immune и inflammatory cells: the molecular basis of mediator secretion. J Allergy Clin. Immunol. 2003, 111:923-932).
Тучные клетки, обычно не циркулирующие в кровотоке, и базофилы содержат секреторные цитоплазматические гранулы, которые сохраняют и могут высвобождать после активации клеток ранее образованные медиаторы воспаления (анафилаксические), такие как гистамин; протеогликаны, такие как сульфат гепарина и хондроитина; протеазы, такие как триптаза, химаза, карбоксипептидаза и катепсин G-подобная протеаза; хемотаксические факторы, цитокины и метаболиты арахидоновой кислоты, которые действуют на сосудистую систему, гладкие мышцы, соединительную ткань, слизистые железы и воспалительные клетки.
Нейтрофилы, также известные как полиморфонуклеарные лейкоциты (PMN), содержат 50-60% всех циркулирующих в кровотоке лейкоцитов. Нейтрофилы действуют против инфекционных агентов, таких как бактерии, грибы, простейшие, вирусы, инфицированные вирусом клетки, а также опухолевые клетки, которые проникают через физические барьеры организма на участках инфицирования или поражения. Созревание нейтрофилов происходит за шесть морфологических стадий: миелобласт, промиелобласт, миелоцит, метамиелоцит, несегментированный (палочкоядерный) нейтрофил и сегментированный (функционально активный) нейтрофил.
У нейтрофилов, медиаторы воспаления сохраняются в первичных (азурофильных), вторичных (специфических) и третичных (желатиназных) гранулах, а также в секреторных везикулах. Среди различных медиаторов воспаления, первичные (азурофильные) гранулы содержат миелопероксидазу (MPO), лизоцим, дефензины, белка, повышающего проницаемость бактерий (BPI), эластазу, катепсин G, катепсин B, катепсин D, бета-D-глюкуронидазу, альфа-маннозидазу, фосфолипазу A2, хондроитин-4-сульфат и протеиназу 3 (см., например, Hartwig J.H; Thelen M, Rosen A, Janmey P.A. Nairn A.C, Aderem A. MARCKS is an actin filament crosslinking protein regulated by protein kinase C and calcium-calmodulin. Nature 1992, 356:618-622); вторичные (специфические) гранулы содержат лизоцим, лактоферрин, коллагеназу, активатор комплемента, фосфолипазу А2, рецепторы комплемента, например, CR3, CR4, рецепторы N-формилметионил-лейцил-фенилаланина (FMLP), рецепторы ламинина, цитохром b558, моноцитарный-хемотаксический фактор, гистаминазу и белок, связывающийся с витамином B12; а небольшие запасающие гранулы содержат желатиназу, активатор плазминогена, катепсин В, катепсин D, бета-D-глюкуронидазу, альфа-маннозидазу и цитохром b558.
Нейтрофильные гранулы содержат противомикробные или цитотоксические вещества, нейтральные протеиназы, кислые гидролазы и пулы цитоплазматических мембранных рецепторов. Азурофильные гранулы, помимо других компонентов, содержат миелопероксидазу (MPO), которая представляет собой фермент, играющий важную роль в превращении пероксида водорода в хлорноватистую кислоту. Этот фермент вместе с пероксидом водорода и галогенидным кофактором, образуют миелопероксидазную систему, которая обеспечивает эффективное противомикробное и цитотоксическое действие лейкоцитов.
Дефензины, которые составляют 30-50% от всех белков азурофильных гранул, представляют собой небольшие (с молекулярной массой <4000) пептиды, обладающие сильным противомикробным действием, которые являются цитотоксичными для широкого ряда бактерий, грибов и некоторых вирусов. Их токсичность может быть обусловлена проницаемостью мембраны клеток-мишеней, которые являются также мишенями для других каналообразующих белков (перфоринов).
Белок, повышающий проницаемость бактерий (BPI), представляет собой член семейства перфоринов. Он является в высокой степени токсичным для грамотрицательных бактерий, но не для грамположительных бактерий или грибов, и может также нейтрализовать эндотоксин, то есть токсический липополисахаридный компонент оболочки грамотрицательных бактериальных клеток.
Лактоферриновые секвестранты не содержат железа, а поэтому они предотвращают рост перорально поступаемых микроорганизмов, которые не подвергаются цитолизу и способствуют улучшению проницаемости бактерий для лизоцимов.
Сериновые протеазы, такие как эластаза и катепсин G, гидролизуют белки оболочек бактериальных клеток. Субстратами гранулоцитарных элатаз являются перекрестно-связанные коллагены и протеогликаны, а также эластины кровеносных сосудов, связок и хряща. Катепсин D расщепляет протеогликаны хряща, тогда как коллагенезы гранулоцитов расщепляют коллаген типа 1 и в меньшей степени коллаген типа III костей, хряща и сухожилия. Продукты расщепления коллагена обладают хемотаксической активностью для нейтрофилов, моноцитов и фибробластов.
Регуляция деструктивного потенциала лизосомных протеаз в ткани опосредуется ингибиторами протеазы, такими как альфа-2-макроглобулин и альфа-1-антипротеаза. Такие антипротеазы присутствуют в сыворотке и в синовиальной жидкости. Они могут функционировать путем связывания с активными центрами протеаз и захватывания этих активных центров. Дисбаланс системы «протеаза-антипротеаза» может играть важную роль в патогенезе эмфиземы.
Азурофильные гранулы функционируют преимущественно во внутриклеточной среде (в фаголизосомных вакуолях), где они участвуют в цитолизе и разложении микроорганизмов. Нейтрофилспецифические гранулы восприимчивы к высвобождению их внеклеточного содержимого и играют важную роль в развитии воспаления. Специфические гранулы представляют собой внутриклеточный резервуар для различных компонентов плазматической мембраны, включая цитохром b (компонент NADPH-оксидазы, фермент, ответственный за продуцирование супероксида), рецепторы для фрагмента комплемента iC3b (CR3, CR4), рецепторы для ламинина и формилметионил-пептидные хемоаттрактанты. Помимо прочих компонентов, в этих гранулах присутствуют гистаминаза, которая играет определенную роль в разложении гистамина; витаминсвязывающий белок, и активатор плазминогена, который является ответственным за образование плазмина и отщепление C5a от C5.
Важная роль нейтрофильных гранул в развитии воспаления была выявлена в результате исследований, проводимых с участием нескольких пациентов с врожденными аномалиями указанных гранул. У пациентов с синдромом Чедиака-Хигаши наблюдаются серьезные нарушения в скорости вырабатывания воспалительного ответа и аномально высокое содержание лизосомных гранул. Врожденный синдром дефекта специфических гранул является исключительно редким заболеванием, характеризующимся пониженным уровнем воспалительного ответа и развитием тяжелых бактериальных инфекций кожи и глубоких тканей.
Хотя механизмы, регулирующие экзоцитозную секрецию этих гранул, изучены лишь частично, однако было идентифицировано несколько ключевых молекул, участвующих в таком процессе, включая внутриклеточный Ca2+ в переходном состоянии (Richter et al. Proc. Natl. Acad. Sci USA 1990, 87:9472-9476, Blackwood et al., Biochem J 1990, 266: 195-200), G-белки, тирозин- и протеинкиназы (РК, а в частности PKC) (Smolen et al., Biochim Biophys Acta 1990, 1052:133-142, Niessen et al., Biochim. Biophys. Acta 1994, 1223:267-273, Naucler et al., Pettersen et al., Chest 2002, 121, 142-150), Rac2 (Abdel-Latif et al., Blood 2004, 104:832-839, Lacy et al., J. Immunol. 2003, 170:2670-2679) и различные SNARE, SNAP и VAMP (Sollner et al., Nature 1993, 362:318-324, Lacy, Pharmacol. Ther 2005, 107:358-376).
Белки SNARE (растворимый рецептор белка, связывающегося с N-этилмалеимидом) принадлежат к семейству мембраносвязанных белков, характеризующихся присутствием альфа-суперспирализованного домена, называемого мотивом SNARE (Li et al., Cell. Mol. Life Sci. 60:942-960 (2003)). Эти белки подразделяются на v-SNARE и t-SNARE исходя из их локализации на везикулах или мембране клетки-мишени, а по другой схеме классификации, эти белки подразделяются на R-SNARE и Q-SNARE исходя из присутствия в них консервативного аргининового или глутаминового остатка в центре мотива SNARE. SNARE локализованы на отдельных мембранных компартментах секреторного и эндоцитозного транспортного пути и вносят свой вклад в специфичность внутриклеточных процессов слияния мембран. Домен t-SNARE состоит из пучка из 4 спиралей с суперспирализованной структурой. Мотив SNARE играет определенную роль в слиянии двух мембран. Мотивы SNARE подразделяются на четыре класса: гомологи синтаксина 1a (t-SNARE), VAMP-2 (v-SNARE) и N- и C-концевые мотивы SNARE SNAP-25. Один член каждого класса может взаимодействовать с образованием комплекса SNARE. Мотив SNARE присутствует в N-концевых доменах некоторых членов семейства синтаксина, таких как синтаксин 1a, который необходим для высвобождения нейромедиаторов (Lerman et al., Biochemistry 39: 8470-8479 (2000)), и синтаксин 6, который присутствует в везикулах эндосомного транспорта (Misura et al., Proc. Natl. Acad. Sci. U.S.A. 99:9184-9189 (2002)).
Белки SNAP-25 (ассоциированные с синаптосомой белки размером 25 кДа) представляют собой компоненты комплексов SNARE, которые могут быть ответственны за специфичность слияния мембран и за прямое слияние посредством образования плотного комплекса (SNARE или корового комплекса), который связывает синаптическую везикулу и плазматические мембраны. SNARE составляют большое семейство белков, характеризующихся последовательностями, состоящими из 60 остатков и известными как мотивы SNARE, которые имеют высокую степень предрасположенности к образованию суперспирализованных структур, и в большинстве случаев, являются предшественниками карбокси-концевых трансмембранных областей. Синаптический коровый комплекс образован четырьмя мотивами SNARE (два мотива от SNAP-25 и по одному от синаптобревина и синтаксина 1), которые по отдельности являются неструктурированными, но вместе образуют пучок из параллельных четырех спиралей. Анализ кристаллической структуры корового комплекса показал, что указанный спиральный пучок имеет высокую степень спирализации и содержит несколько солевых мостиков на своей поверхности, а также слои из внутренних гидрофобных остатков. Полярный слой в центре такого комплекса образован тремя глутаминами (двумя от SNAP-25 и одним от синтаксина 1) и одним аргинином (от синаптобревина) (Rizo et al., Nat. Rev. Neurosci 3:641-653 (2002)). Члены семейства SNAP-25 содержат кластер из цистеиновых остатков, которые могут быть пальмитоилированными для связывания с мембраной (Risinger et al., J. Biol. Chem. 268:24408-24414 (1993)).
Нейтрофилы играют важную роль в фагоцитозе и в разрушении инфекционных агентов. Они также ограничивают рост некоторых микробов перед началом вырабатывания адаптивных (специфических) иммунных ответов. Хотя нейтрофилы играют важную роль в защите хозяина, однако они также приводят к осложнению многих хронических воспалительных состояний и к реперфузионным поражениям при ишемии. Гидролитические ферменты, происходящие от нейтрофилов, и инактивированные в результате окисления ингибиторы протеазы могут быть детектированы в физиологической жидкости, выделенной из зоны воспаления. В нормальных условиях, нейтрофилы могут мигрировать на участки инфицирования, не повреждая при этом ткани хозяина. Однако иногда все же может наблюдаться нежелательное повреждение ткани хозяина. Такое повреждение может происходить по нескольким независимым механизмам. Такими механизмами являются преждевременная активация в процессе миграции, внеклеточное высвобождение токсических продуктов в процессе цитолиза некоторых микробов, удаление инфицированных или поврежденных клеток-хозяев и клеточного дебриса в качестве первой стадии ремоделирования ткани, или неспособность к блокированию острых воспалительных ответов. Реперфузионное повреждение при ишемии ассоциируется с притоком нейтрофилов в пораженную ткань и их последующей активацией. Такой процесс может запускаться под действием веществ, высвобождаемых из пораженных клеток-хозяев, либо он может быть вызван образованием супероксида под действием ксантин-оксидазы.
В нормальных условиях, кровь может содержать смесь нормальных, примированных, активированных нейтрофилов и с ослабленной функцией нейтрофилов. В зоне воспаления присутствуют, главным образом, активированные нейтрофилы и нейтрофилы с ослабленной функцией. Активированные нейтрофилы усиливают продуцирование промежуточных продуктов реакционноспособного кислорода (ROI). Субпопуляция нейтрофилов с усиленной дыхательной вспышкой была обнаружена в крови людей с острой бактериальной инфекцией и у пациентов с острым респираторным дистресс-синдромом (ARDS). В этом заключается нейтрофильный парадокс. Нейтрофилы усугубляют патологию этого состояния, что обусловлено значительным притоком этих клеток в легкие и ассоциированным с ним поражением ткани, вызываемым действием окислителей и гидролитических ферментов, высвобождаемых из активированных нейтрофилов. Нарушение противомикробной активности нейтрофилов, которое приводит к ухудшению состояния при ARDS, может служить протективным ответом на том участке организма хозяина, который локально индуцируется продуктами воспаления.
Острая фаза термического поражения также ассоциируется с активацией нейтрофилов с последующим общим нарушением различных функций нейтрофилов. Активация нейтрофилов иммунными комплексами в синовиальной жидкости вносит свой вклад в патологию ревматоидного артрита. Перманентная активация нейтрофилов может также инициировать развитие опухоли, поскольку некоторые ROI, образованные нейтрофилами, разрушают ДНК и стимулируют миграцию опухолевых клеток под действием протеаз. У пациентов, страдающих ожогами тяжелой степени, была установлена корреляция между началом развития бактериальной инфекции и уменьшением относительного и абсолютного числа нейтрофилов, позитивных по вырабатыванию антител и рецепторов комплемента. Было также обнаружено, что окислители нейтрофильной природы окисляют липопротеины низкой плотности (ЛНП), которые затем более эффективно присоединяются к плазматической мембране макрофагов посредством специфических акцепторных рецепторов. Поглощение этих окисленных ЛНП макрофагами может инициировать атеросклероз. Кроме того, примированные нейтрофилы были обнаружены у людей с эссенциальной гипертензией, болезнью Ходжкина, воспалительным заболеванием кишечника, псориазом, саркоидозом и септицемией, где такое примирование коррелирует с высокими концентрациями циркулирующего в кровотоке TNF-альфа (кахектина).
Гидролитическое повреждение ткани хозяина, а следовательно развитие хронических воспалительных состояний может происходить в том случае, если защитные слои из антиоксидантов и антипротеаз истощаются. Считается, что дефицит антипротеаз ответственен за развитие патологий при энфиземе. Многие антипротеазы являются членами семейства ингибиторов сериновых протеаз (SERPIN). Хотя кровоток обогащен антипротеазами, однако эти крупные белки могут селективно удаляться из участков воспаления, что обусловлено адгезией нейтрофилов к своим мишеням. Окислительный стресс может инициировать поражение ткани в результате снижения концентрации внеклеточных антипротеаз до значений ниже уровня, требуемого для ингибирования высвобождаемых протеаз. Хлорированные окислители и пероксид водорода могут инактивировать антипротеазы, такие как ингибитор альфа-1-протеазы и альфа-2-макроглобулин, которые являются эндогенными ингибиторами эластазы, но одновременно активируют латентные металлопротеазы, такие как коллагеназы и желатиназа, играющие определенную роль в последующей инактивации антипротеаз.
Цитоплазматические компоненты нейтрофилов могут также вызывать образование специфических антител против цитоплазматических нейтрофилов (ANCA), которые тесно связаны с развитием системных васкулитов и гломерулонефрита. ANCA представляют собой антитела, направленные против ферментов, которые присутствуют, главным образом, в азурофильных или первичных гранулах нейтрофилов. Существует три типа ANCA, которые могут различаться по характеру непрямой иммунофлуоресценции, продуцируемой на нормальных фиксированных этанолом нейтрофилах. Диффузная мелкоструктурная цитоплазматическая гранулярная флуоресценция (cANCA) обычно наблюдается при гранулематозе Вегенера, в некоторых случаях при микроскопическом полиартериите и синдроме Черга-Штрауса, а в некоторых случаях, при серповидно-клеточном и сегментарном некрозирующем гломерулонефрите. Антигеном-мишенью обычно является протеиназа 3. Перинуклеарная флуоресценция (pANCA) наблюдается во многих случаях при микроскопическом полиартериите и гломерулонефрите. Эти антитела часто направлены против миелопероксидазы, а другими их мишенями являются эластаза, катепсин G, лактоферрин, лизоцим и бета-D-глюкуронидаза. Третья группа, называемая «атипическими» ANCA, включает антитела, выявляемые по флуоресценции нейтрофильных ядер и по некоторым необычным цитоплазматическим паттернам, причем лишь немногие из антигенов-мишеней продуцируют pANCA, а другие антигены-мишени пока еще не были идентифицированы. pANCA также были обнаружены у трети пациентов с болезнью Крона. Сообщалось, что частота встречаемости ANCA при ревматоидном артрите и СКВ значительно варьируется, при этом преимущественно наблюдались pANCA и атипичные ANCA.
Эозинофилы представляют собой лейкоциты конечной стадии дифференцировки, которые присутствуют, преимущественно, в слое ткани, расположенном под слизистой оболочкой, и рекрутинг таких лейкоцитов происходит на участках специфических иммунных реакций, включая аллергические реакции. Цитоплазма эозинофилов содержит крупные эллипсоидные гранулы с электронноплотным кристаллическим ядром и частично проницаемым матриксом. Помимо этих крупных первичных кристаллоидных гранул существуют гранулы и другого типа, которые имеют меньший размер (небольшие гранулы) и не содержат кристаллического ядра. Крупные специфические гранулы эозинофилов содержат по меньшей мере четыре различных катионных белка, которые оказывают различное биологическое действие на клетки-хозяева и микробные мишени, а именно основный мажорный белок (MBP), катионный белок эозинофилов (ECP), нейротоксин, происходящий от эозинофилов (EDN), и пероксидаза эозинофилов (EPO). Базофилы содержат примерно одну четвертую всех основных мажорных белков, присутствующих в эозинофилах, вместе с детектируемыми количествами EDN, ECP и EPO. Небольшие количества EDN и ECP также обнаруживаются в нейтрофилах (Gleich G. J. Mechanisms of eosinophil-associated inflammation. J. Allergy Clin. Immunol. 2000, 105:651-663). MBP, очевидно, не обладают ферментативной активностью и представляют собой в высокой степени катионногенный полипептид, который может обладать токсической активностью, вызываемой взаимодействием с липидными мембранами, приводящим к их разрушению. MBP и EPO могут действовать как селективные аллостерические ингибиторы агониста, связывающегося с мускариновыми рецепторами M2. Эти белки могут играть определенную роль в дисфункции рецептора M2 и усиливать опосредуемый блуждающим нервом бронхостеноз при астме. EDN может специфически разрушать миелиновую оболочку нейронов. Гистаминаза и различные гидролитические лизосомные ферменты также присутствуют в большинстве специфических гранул эозинофилов. В небольших гранулах эозинофилов среди прочих ферментов присутствуют арилсульфатаза, кислая фосфатаза, металлопротеиназа размером 92 кДа и желатиназа. Эозинофилы могут вырабатывать цитокины, которые обладают потенциальной аутокринной активностью факторов роста для эозинофилов и которые играют потенциальную роль в продуцировании острых и хронических воспалительных ответов. Существует три цитокина, которые обладают активностью факторов роста для эозинофилов, а именно: гранулоцитарный-макрофагальный колониестимулирующий фактор (GM-CSF), IL-3 и IL-5. Другие цитокины, продуцируемые человеческими эозинофилами, могут обладать активностью, направленной на вырабатывание острых и хронических воспалительных ответов, и такими цитокинами являются IL-1-альфа, IL-6, IL-8, TNF-альфа и трансформирующие факторы роста, TGF-альфа и TGF-бета.
Эозинофилы содержат кристаллоидные гранулы, которые включают MBP, катионный эозинофильный белок, EPO и эозинофильный нейротоксин (Gleich, J. Allergy Clin. Immunol. 2000, 105:651-663). Клон 15 человеческой промиелоцитарной клеточной линии HL-60 может быть использован для оценки секреции EPO. Эта клеточная линия была выведена из клона HL-60, который культивировали при повышенном рН в течение двух месяцев (Fischkoff, Leuk Res 1988, 12:679-686), а затем обрабатывали масляной кислотой для обеспечения дифференцировки клеток и сообщения этим клеткам множества свойств, присущих эозинофилам периферической крови, включая экспрессию белков эозинофил-специфических гранул (Rosenberg et al., J. Exp. Med. 1989, 170:163-176, Tiffany et al., J. Leukoc. Biol. 1995, 58:49-54, Badewa et al., Exp. Biol. Med. 2002, 227:645-651).
Эозинофилы могут участвовать в реакциях гиперчувствительности, в частности, посредством двух липидных медиаторов воспаления, лейкотриена С4 (LTC4) и фактора активации тромбоцитов (PAF). Оба медиатора сокращают гладкие мышцы дыхательных путей, стимулируют секрецию слизи, изменяют сосудистую проницаемость и вызывают инфильтрацию эозинофилов и нейтрофилов. Помимо прямой активности указанных эозинофильных медиаторов, MBP может стимулировать высвобождение гистамина из базофилов и тучных клеток, а MBP может стимулировать высвобождение EPO из тучных клеток. Эозинофилы могут служить локальным источником специфических липидных медиаторов, а также могут индуцировать высвобождение медиаторов из тучных клеток и базофилов. Содержимое эозинофильных гранул высвобождается после аналогичной стимуляции нейтрофильными гранулами, например, в процессе фагоцитоза опсонизированных частиц и под действием хемотаксических факторов. Нейтрофильные лизосомные ферменты действуют, главным образом, на материал, присутствующий в фаголизосомах, тогда как содержимое эозинофильных гранул действует, главным образом, на внеклеточную структуру-мишень, такую как паразиты и медиаторы воспаления.
Моноциты и макрофаги развиваются в костном мозге и проходят следующие стадии: стволовая клетка, коммитированная стволовая клетка, монобласт, промоноцит, моноцит в костном мозге, моноцит в периферической крови и макрофаг в тканях. Дифференцировка моноцитов в костном мозге происходит достаточно быстро (за 1,5-3 дня). В процессе дифференцировки, в цитоплазме моноцитов образуются гранулы, и эти гранулы, как и в нейтрофилах, могут быть подразделены по меньшей мере на два типа. Однако они являются более малочисленными и имеют меньший размер, чем их нейтрофильные аналоги (азурофильные и специфические гранулы). Эти гранулы содержат аналогичные ферменты.
Связанными с гранулами ферментами моноцитов/макрофагов являются лизоцим, кислая фосфатаза и бета-глюкуронидаза. В качестве модели для in vivo исследований была использована секреция лизоцима из клеток U937. Такая клеточная линия происходит от человеческой гистиоцитарной лимфомы и была использована в качестве моноцитарной клеточной линии, которая может быть активирована различными агонистами, такими как PMA (Hoff et al. J. Leukoc. Biol. 1992, 52:173-182; Balboa et al., J. Immunol. 2003; 170:5276-5280, Sundstrom et al., Int. J. Cancer 1976; 17:565-577).
Природные клетки-киллеры (NK) и цитотоксические лимфоциты содержат сильнодействующие цитотоксические гранулы, включающие перфорин, порообразующий белок, гранзимы и лимфоцит-специфические сериновые протеазы. Так, например, клеточная линия NK-92 представляет собой IL-2-зависимую человеческую клеточную линию, выделенную у пациента с быстро прогрессирующей неходжкинской лимфомой (Gong J.H., Maki G, Klingemann H.G. Characterization of a human cell line (NK-92) with phenotypical и functional characteristics of activated natural killer cells. Leukemia 1994, 8:652-658). Клетки NK-92 экспрессируют высокие уровни молекул, участвующих в цитолитическом пути перфорина-гранзима, который направлен на злокачественные клетки широкого ряда (Gong et al, vide infra, and Maki G, Klingemann H.G, Martinson J.A, Tarn Y.K. Factors regulating the cytotoxic activity of the human natural killer cell line, NK-92. J Hematother Stem Cell Res 2001; 10:369-383).
Гранзимы представляют собой экзогенные сериновые протеазы, которые высвобождаются цитоплазматическими гранулами в цитотоксических Т-клетках и в природных клетках-киллерах. Гранзимы могут индуцировать апоптоз в инфицированных вирусом клетках и тем самым разрушать эти клетки.
Внеклеточная секреция медиатора воспаления (воспалительного медиатора) из гранулоцита (или лейкоцита) и внеклеточная секреция более одного медиатора воспаления (воспалительного медиатора) из гранулоцита (или лейкоцита) иногда называется здесь дегрануляцией. В предпочтительном варианте изобретения, высвобождение медиатора воспаления представляет собой высвобождение указанного медиатора из гранулы, расположенной внутри гранулоцита или лейкоцита. Высвобождение медиатора воспаления, предпочтительно, представляет собой высвобождение указанного медиатора воспаления из этих гранул.
В нейтрофилах и макрофагах после их примирования провоспалительными агентами (стимуляторами воспаления), такими как TNFα, резко усиливается синтез белка MARCKS, причем более чем 90% от всего количества новых белков, образующихся в ответ на действие TNFα или липополисахарида (ЛПС), составляет белок MARCKS (Thelen M, Rosen A, Nairn AC, Aderem. A. Tumor necrosis factor alpha modifies agonist-dependent responses in human neutrophils by inducing the synthesis и myristoylation of a specific protein kinase C substrate. Proc. Natl. Acad. Sci. USA 1990, 87:5603-5607). Таким образом, MARCKS может играть важную роль в последующем высвобождении медиаторов воспаления в случае, если содержащие гранулы клетки, такие как нейтрофилы и макрофаги, были стимулированы агонистами, а в частности агонистами, которые действуют посредством активации PKC (Burgoyne et al., Physiol Rev 2003, 83:581-632, Logan et al. J. Allergy Clin. Immunol. 2003, 111:923-932, Smolen et al., Biochim. Biophys. Acta 1990, 1052:133-142, Niessen et al., Biochim. Biophys. Acta 1994, 1223:267-273, Naucler et al., J Leukoc Biol. 2002, 71:701-710).
В одном из аспектов изобретения, введение ингибирующего дегрануляцию количества описанного здесь пептида MANS или его активного фрагмента в область воспаления у индивидуума, которая явилась очагом заболевания, патологического состояния или травмы, либо местом внедрения чужеродного тела, или их комбинаций, может приводить к снижению количества медиатора воспаления, высвобождаемого из инфильтрирующихся лейкоцитов в области воспаления, где указанными лейкоцитами, предпочтительно, являются гранулоциты. Введение пептида MANS и/или его по меньшей мере одного активного фрагмента может приводить к снижению количества медиатора воспаления, высвобождаемого из лейкоцитов, таких как гранулоциты, инфильтрирующиеся в область воспаления. Ингибирующее дегрануляцию количество пептида MANS или ингибирующее дегрануляцию количество его активного фрагмента является достаточным для снижения или ингибирования уровня экзоцитозного высвобождения медиаторов воспаления из гранул, содержащихся в воспалительных клетках, инфильтрирующихся в указанную область воспаления. Эффективность ингибирования дегрануляции определяют сразу после введения пептида MANS или его активного фрагмента путем сравнения процента ингибирования (то есть процента снижения) высвобождения медиаторов воспаления из указанных клеток (лейкоцитов или гранулоцитов или других воспалительных клеток) по отношению к уровню или количеству или концентрации указанных медиаторов воспаления, высвобождаемых или продуцируемых приблизительно в то же самое время в отсутствие пептида MANS и/или в отсутствие его активного фрагмента. Кроме того, врач-клиницист может самостоятельно определить степень снижения воспаления на участке ткани путем выявления симптомов и измерения параметров воспаления, известных как индикаторы заболевания, для того чтобы оценить, является ли количество пептида MANS и/или его активного фрагмента достаточным или терапевтически эффективным для такого ослабления воспаления. Количеством, достаточным для ингибирования дегрануляции, является количество, которое обеспечивает определенный процент снижения уровня медиатора воспаления, высвобождаемого из гранулоцитов на участке воспаления, где указанный процент составляет примерно 1% - 99%, предпочтительно, примерно 5% - 99%, более предпочтительно, примерно 10% - 99%, еще более предпочтительно, примерно 25% - 99%, а наиболее предпочтительно, примерно 50% - 99% от количества указанного медиатора воспаления, высвобождаемого из указанных гранулоцитов в отсутствие пептида MANS или его активного фрагмента, тестируемых в тех же самых условиях.
В одном из аспектов изобретения, введение ингибирующего дегрануляцию количества пептида MANS в область стимуляции воспаления у животного, где указанная область воспаления была индуцирована введением в нее стимулирующего воспаление количества стимулятора воспаления, может приводить к снижению количества медиатора воспаления, высвобождаемого из гранулоцитов, стимулируемых указанным стимулятором воспаления в указанной области стимуляции воспаления, примерно на 1% - 99%, предпочтительно, примерно на 5% - 99%, более предпочтительно, примерно на 10% - 99%, еще более предпочтительно, примерно на 25% - 99%, а наиболее предпочтительно, примерно на 50% - 99% от количества указанного медиатора воспаления, высвобождаемого из указанных гранулоцитов в отсутствие пептида MANS, но в присутствии идентичного стимулирующего воспаление количества указанного стимулятора воспаления.
В другом аспекте изобретения, введение ингибирующего дегрануляцию количества пептида MANS в область стимуляции воспаления у животного, где указанная область воспаления была индуцирована введением в нее стимулирующего воспаление количества стимулятора воспаления, может приводить к снижению количества медиатора воспаления, высвобождаемого из гранулоцитов, стимулируемых указанным стимулятором воспаления в указанной области стимуляции воспаления, на 100% от количества указанного медиатора воспаления, высвобождаемого из указанных гранулоцитов в отсутствие пептида MANS, но в присутствии идентичного стимулирующего воспаление количества указанного стимулятора воспаления.
Примером стимулятора воспаления, используемого in vitro, как описано в примерах, приведенных в настоящей заявке, является форбол-12-миристат-13-ацетат (PMA). Моноцитарный белок-хемоаттрактант (MCP-1) обладает почти такой же эффективностью, как C5a, и гораздо большей активностью, чем IL-8, в отношении дегрануляции базофилов и вызывает высвобождение гистамина. Высвобождение гистамина может происходить после стимуляции хемокинами (то есть цитокинами-хемоаттрактантами), RANTES и MIP-1.
Что касается базальной концентрации пептида MARCKS, присутствующего в области стимуляции воспаления, то в предпочтительном варианте изобретения, ингибирующее дегрануляцию количество пептида MANS, вводимого животному в область стимуляции воспаления, примерно в 1-1000000 раз, предпочтительно, примерно в 1-100000 раз, более предпочтительно, примерно в 1-10000 раз, еще более предпочтительно, примерно в 1-1000 раз, еще более предпочтительно, примерно в 1-100 раз, а наиболее предпочтительно, примерно в 1-10 раз превышает концентрацию пептида MARCKS в указанной области стимуляции воспаления.
В предпочтительном варианте изобретения, гранулоциты присутствуют на поверхности или внутри дыхательных путей животного, а предпочтительно, человека, а поэтому пептид MANS вводят путем ингаляции, например, фармацевтической композиции, содержащей указанный пептид MANS, например, фармацевтической композиции, содержащей пептид MANS и водный раствор, где указанную композицию вводят в форме аэрозоля или в форме сухого порошка, либо указанную композицию вводят с помощью инсуффлятора. Могут быть применены и другие методы и устройства для введения раствора или порошка путем ингаляции, такие, например, как капельки, спреи и небулайзеры.
В некоторых вариантах изобретения, пептид согласно изобретению может блокировать физиологически важные секреторные процессы, включая блокирование базальных секреторных функций. Не ограничиваясь какой-либо конкретной теорией, авторы настоящего изобретения считают, что механизмы регуляции такой базальной секреции отличаются от механизмов регуляции стимулируемой секреции. Альтернативно, для осуществления базальных механизмов секреции может потребоваться меньшее количество белка MARCKS, чем для стимулируемой секреции. Базальная секреция может быть сохранена, поскольку все методы терапии, направленные на блокирование MARCKS-опосредуемой секреции, могут и не подавлять все функции MARCKS.
Используемый здесь термин «нуклеотидная последовательность MARCKS» означает любую нуклеотидную последовательность, происходящую от гена, кодирующего белок MARCKS, включая, например, последовательность ДНК или РНК, последовательность ДНК гена, любую транскрибируемую последовательность РНК, последовательность РНК пре-мРНК или мРНК-транскрипт, а также ДНК или РНК, связанные с белком.
Точная доставка MARCKS-блокирующего пептида может также предотвращать любые возможные ограничения блокирования важных секреторных процессов. Доставка таких агентов в дыхательные пути может быть легко осуществлена с помощью композиций, вводимых путем ингаляции. Поскольку такие агенты могут быть использованы для лечения воспалительного заболевания кишечника, то может рассматриваться и доставка блокирующих агентов в область прямой кишки/толстой кишки/тонкой кишки с помощью клизмы или суппозиториев. Внутрисуставные инъекции или чрескожная доставка в воспаленные суставы может приносить облегчение пациентам с артритными или аутоиммунными заболеваниями путем ограничения секреции из клеток, локализованных на участке воспаления. Инъекция в области, окружающие нервные окончания, может приводить к ингибированию секреции нейромедиаторов некоторых типов, то есть к блокированию сигналов возникновения сильной боли или неконтролируемых спазмов мышц. Доставка пептида для лечения воспалительных кожных болезней может быть легко осуществлена с помощью различных препаратов для местного введения, известных специалистам.
Очевидно, что MARCKS взаимодействует с актином и миозином в цитоплазме, а поэтому он может обладать способностью связывать гранулы с клеточным сократительным аппаратом и тем самым опосредовать дальнейший транспорт гранул и экзоцитоз. Секреция медиатора воспаления МРО из нейтрофилов может быть также максимизирована посредством активации PKC и PKG. Возможно, что MARCKS служит точкой конвергенции координирующих действий этих двух протеинкиназ, которые регулируют секрецию из мембраносвязанных компартментов воспалительных клеток (то есть секрецию МРО из нейтрофилов).
В настоящем изобретении было продемонстрировано, что секреция медиатора воспаления МРО из собачьих или человеческих нейтрофилов усиливается под действием конкурентной активации PKC и PKG, однако активации одной киназы не достаточно для индуцирования максимального секреторного ответа. Усиленный секреторный ответ на один PMA наблюдался в клетках NHBE и в нейтрофилах, как описано в настоящей заявке, хотя величина такого ответа была гораздо меньше величины ответа, наблюдаемого в крысиной клеточной линии бокаловидных клеток. См., выше Abdullah et al. Кроме того, хотя ранее сообщалось, что аналог cGMP может индуцировать значительный уровень секреции муцина из культивированных эпителиальных клеток трахеи морских свинок (Fischer et al, см. выше), однако следует отметить, что этот ответ не достигал значимых уровней в течение 8 часов после начала обработки. Секреторный ответ с таким длительным лаг-периодом (периодом запаздывания) едва ли будет давать прямой эффект, а, вероятнее всего, он будет участвовать в синтезе белка de novo, а не в высвобождении уже образованных и запасных цитоплазматических гранул.
Как указывалось выше, настоящее изобретение может быть использовано для получения фармацевтической композиции. В некоторых вариантах изобретения, лекарственный продукт присутствует в твердой фармацевтической композиции, которая может быть использована для перорального введения. Твердая композиция, рассматриваемая в настоящем изобретении, может быть приготовлена вместе с наполнителем, либо она может быть смешана с наполнителем и/или разведена в этом наполнителе. Рассматриваемая твердая композиция может быть также заключена в носитель, которым может быть, например, капсула, саше, таблетка, бумага или другая емкость. Если наполнитель служит в качестве разбавителя, то он может быть твердым, полутвердым или жидким веществом, которое действует как эксципиент, носитель или среда для рассматриваемой композиции.
Различные подходящие наполнители известны специалистам в данной области, и их можно найти на страницах 2404-2406 Национального реестра лекарственных средств (National Formulary, 19:2404-2406 (2000)), которые во всей своей полноте вводятся в настоящее описание. Примерами подходящих наполнителей являются, но не ограничивается ими, крахмалы, аравийская камедь, силикат кальция, микрокристаллическая целлюлоза, метакрилаты, шеллак, поливинилпирролидон, целлюлоза, вода, сироп и метилцеллюлоза. Композиции лекарственного продукта могут дополнительно включать замасливатели, такие как, например, тальк, стеарат магния и минеральное масло; смачивающие агенты; эмульгаторы и суспендирующие агенты; консерванты, такие как метил- и пропилгидроксибензоаты; подсластители или ароматизаторы. Могут быть также использованы полиолы, буферы и инертные наполнители. Примерами полиолов являются, но не ограничивается ими, маннит, сорбит, ксилит, сахароза, мальтоза, глюкоза, лактоза, декстроза и т.п. Подходящими буферами являются, но не ограничивается ими, фосфат, цитрат, тартрат, сукцинат и т.п. Другими инертными наполнителями, которые могут быть использованы, являются наполнители, известные специалистам и подходящие для их использования в целях приготовления различных лекарственных форм. Если это необходимо, то твердые композиции могут включать и другие компоненты, такие как агенты, придающие объем и/или грануляторы и т.п. Лекарственные продукты согласно изобретению могут быть приготовлены для быстрого, пролонгированного или замедленного высвобождения активного ингредиента после введения таких продуктов пациенту методами, хорошо известными специалистам.
В целях получения таблеток для перорального введения, рассматриваемая композиция согласно изобретению может быть получена методом прямого прессования. В этом процессе, активные лекарственные ингредиенты могут быть смешаны с твердым летучим носителем, таким как, например, лактоза, сахароза, сорбит, маннит, крахмал, амилопектин, производные целлюлозы или желатин и их смеси, а также с антифрикционным агентом, таким как, например, стеарат магния, стеарат кальция и воски на основе полиэтиленгликоля. Затем смесь может быть спрессована в таблетки на машине с соответствующими перфораторами и штампами с получением таблетки нужного размера. Рабочие параметры этой машины могут быть выбраны самим специалистом. Альтернативно, таблетки для перорального введения могут быть получены методом мокрой грануляции. Активные лекарственные ингредиенты могут быть смешаны с наполнителями и/или носителями. Твердые вещества могут быть измельчены или просеяны до получения частиц нужного размера. В лекарственное средство может быть добавлен связывающий агент. Связывающий агент может быть суспендирован и гомогенизирован в подходящем растворителе. Активный ингредиент и вспомогательные агенты могут быть также смешаны с раствором связывающего агента. Полученную сухую смесь равномерно увлажняют раствором. Увлажнение обычно придает частицам некоторую степень агрегации, а поэтому полученную массу прессуют через сито из нержавеющей стали с нужным размером ячеек. После этого смесь сушат в сушильных шкафах с регулируемой температурой в течение определенного периода времени, необходимого для достижения нужного размера частиц и консистенции. Гранулы осушенной смеси просеивают для удаления любого порошка. К этой смеси могут быть добавлены дезинтегрирующие, антифрикционные и/или антиадгезивные агенты. И наконец, смесь спрессовывают в таблетки на машине с соответствующими перфораторами и штампами с получением таблетки нужного размера. Рабочие параметры этой машины могут быть выбраны самим специалистом.
Если нужно получить таблетки с покрытиями, то полученная как описано выше сердцевина таблетки может быть покрыта концентрированным раствором сахаров или целлюлозных полимеров, который может содержать аравийскую камедь, желатин, тальк и диоксид титана, или лаком, растворенным в летучем органическом растворителе или в смеси растворителей. Для идентификации таблеток с различными активными соединениями или с различными количествами присутствующего в них активного соединения к этому покрытию могут быть добавлены различные красители. В конкретном варианте изобретения, активный ингредиент может присутствовать в сердцевине, окруженной одним или несколькими слоями, включая слои энтеросолюбильного покрытия.
Могут быть получены мягкие желатиновые капсулы, содержащие смесь активного ингредиента и растительного масла. Жесткие желатиновые капсулы могут содержать гранулы активного ингредиента в комбинации с твердым летучим носителем, таким как, например, лактоза, сахароза, сорбит, маннит, картофельный крахмал, кукурузный крахмал, амилопектин, производные целлюлозы и/или желатин.
Жидкие препараты для перорального введения могут быть приготовлены в виде сиропов или суспензий, например, в виде растворов, содержащих активный ингредиент, сахар и смесь этанола, воды, глицерина и пропиленгликоля. Если это необходимо, то такие жидкие препараты могут содержать один или несколько из следующих компонентов, таких как красители, ароматизаторы и сахарин. Могут быть также использованы загустители, такие как карбоксиметилцеллюлоза.
В случае, если вышеуказанные фармацевтические препараты используются для парентерального введения, то такие препараты могут содержать стерильные водные растворы для инъекций, безводные растворы для инъекций или те и другие, включающие композицию согласно изобретению. В случае получения водных растворов для инъекций, композиция согласно изобретению может присутствовать в виде водорастворимой фармацевтически приемлемой соли. Препараты для парентерального введения могут содержать антиоксиданты, буферы, бактериостаты и растворенные вещества, которые сообщают данному препарату изотоничность с кровью данного реципиента. Водные и безводные стерильные суспензии могут содержать суспендирующие агенты и загустители. Такие препараты могут содержаться в контейнерах, предназначенных для хранения одноразовой дозы или доз для многократного приема, например, в герметично закрытых ампулах и флаконах. Растворы и суспензии, приготовленные в виде инъекций для немедленного приема, могут быть получены из стерильных порошков, гранул и таблеток описанного ранее типа.
Рассматриваемая композиция может быть также приготовлена так, чтобы она могла быть использована для местного применения (например, в виде крема для нанесения на кожу). Такие композиции могут содержать различные наполнители, известные специалистам. Подходящими наполнителями могут быть, но не ограничиваются ими, воск на основе цетиловых эфиров, цетиловый спирт, белый воск, глицерилмоностеарат, пропиленгликоль, моностеарат, метилстеарат, бензиловый спирт, лаурилсульфат натрия, глицерин, минеральное масло, вода, карбомер, этиловый спирт, акрилатные адгезивы, полиизобутиленовые адгезивы и силиконовые адгезивы.
В предпочтительном варианте изобретения, пептидные фрагменты описаны в таблице 2 и имеют длину по меньшей мере 4-23 аминокислотных остатка, и их аминокислотные последовательности идентичны аминокислотной последовательности пептида MANS, где указанные N-концевые аминокислоты данных пептидов выбраны из аминокислот, присутствующих в положениях 2-21 пептидной последовательности MANS (SEQ ID NO:1). Более предпочтительно, пептидный фрагмент имеет длину по меньшей мере от 6 аминокислот до 23 аминокислот. Эти пептиды, предпочтительно, являются ацилированными у альфа-N-концевой аминокислоты, а более предпочтительно, они являются миристоилированными в альфа-N-концевом положении аминокислоты.



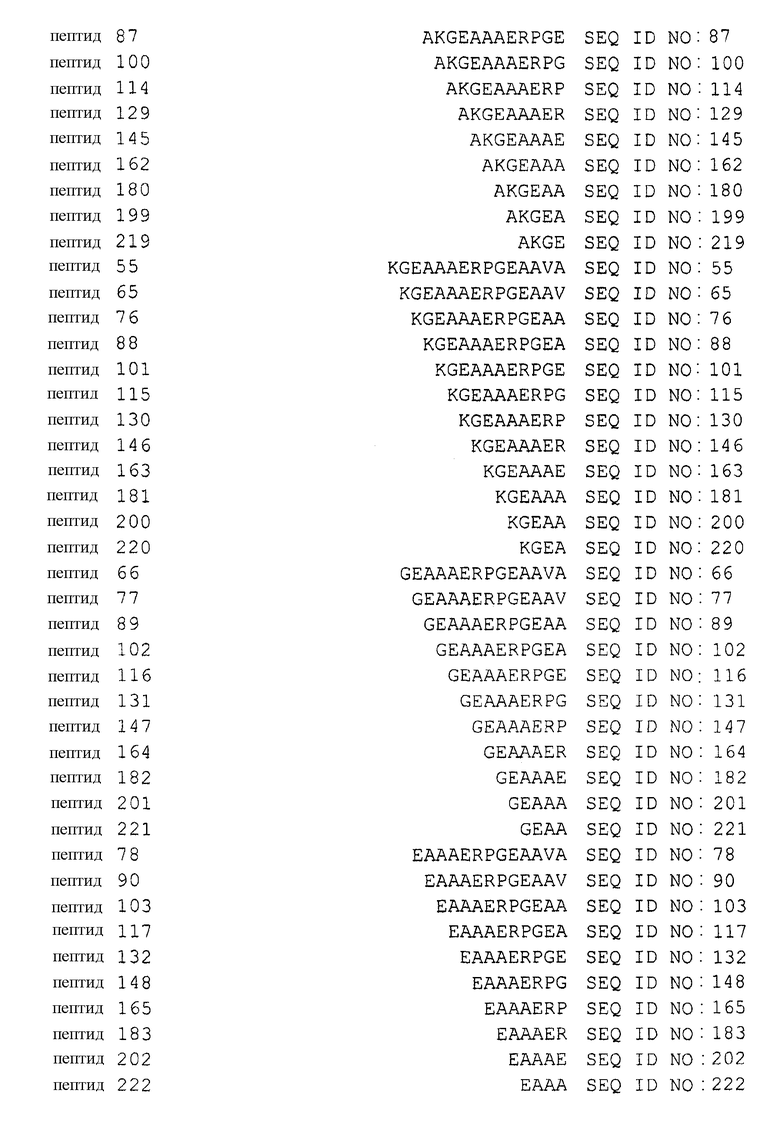
Как проиллюстровано на фиг.5, MARCKS фосфорилируется под действием протеинкиназы С (PKC), а затем транспортируется из мембраны в цитоплазму. В цитоплазме, PKG, очевидно, индуцирует дефосфорилирование MARCKS (фиг.2A, дорожка 4, и фиг.2B). Такое дефосфорилирование прекращается под действием ингибитора PKG, Rp-8-Br-PET-cGMP (фиг.2A, дорожка 5), что свидетельствует о том, что дефосфорилирование, в частности, является PKG-зависимым. На фиг.2, клетки NHBE были помечены [32P]ортофосфатом, а затем они были обработаны указанными реагентами. Фосфорилирование MARCKS в ответ на такую обработку оценивали с помощью иммунопреципитационного анализа. На фиг.2A, 8-Br-cGMP предотвращает фосфорилирование MARCKS, индуцированное PMA, и этот эффект 8-Br-cGMP может быть блокирован под действием Rp-8-Br-PET-cGMP (ингибитора PKG) или окадаиновой кислоты (ингибитора PP1/2A). На фиг.2B, PMA-индуцированное фосфорилирование MARCKS прекращается после обработки клеток 8-Br-cGMP. Дорожка 1, только среда, 8 минут; дорожка 2, 100 нМ PMA, 3 минуты; дорожка 3, 100 нМ PMA, 3 минуты, а затем 1 мкМ 8-Br-cGMP, 5 минут; дорожка 4, 100 нМ PMA, 8 минут; дорожка 5, только среда, 3 минуты, а затем 100 нМ PMA + 1 мкМ 8-Br-cGMP, 5 минут. На фиг. 2C, 8-Br-cGMP-индуцировнное дефосфорилирование MARCKS подавляется под действием фостриецина в зависимости от концентрации.
Очевидно, что PKG дефосфорилирует MARCKS посредством активации протеинфосфатазы. Как проиллюстрировано на фиг.2A (дорожка 6), окадаиновая кислота при концентрации 500 нМ, то есть при концентрации, которая может ингибировать PPl и PP2A, блокирует PKG-индуцированное дефосфорилирование MARCKS, что позволяет предположить, что PKG вызывает дефосфорилирование под действием активирующих PPl и/или PP2A. Дополнительные исследования, проводимые с использованием фостриецина и с помощью прямого анализа на фосфатазную активность, показали, что только PP2A активировался под действием PKG и был ответственным за удаление фосфатных групп из MARCKS (фиг.2C). Очевидно, что окадаиновая кислота или фостриецин, при концентрациях, которые ингибируют PKG-индуцированное дефосфорилирование MARCKS, снижают уровень секреции муцина, индуцированной PMA+8-Br-cGMP или UTP, как показано на фиг.3. Фиг.3 позволяет продемонстрировать, что PP2A является основным компонентом пути секреции муцина. Клетки NHBE предварительно инкубировали с фостриецином и с окадаиновой кислотой в указанной концентрации (500 нМ), или с одной средой в течение 15 минут, а затем стимулировали PMA (100 нМ)+8-Br-cGMP (1 мкМ) в течение 15 минут или UTP (100 мкМ) в течение 2 часов. Уровень секретированного муцина измеряли с помощью ELISA. Данные представлены как среднее ± ср.кв.ош. (n=6 в каждой точке), где * означает значимое отличие от контроля (среда) (p<0,05); † означает значимое отличие от PMA+8-Br-cGMP-стимуляции (p<0,05); а ‡ означает значимое отличие от UTP-стимуляции (p<0,05). Таким образом, очевидно, что дефосфорилирование MARCKS под действием PKG-активированного PP2A является основным компонентом пути передачи сигнала, приводящего к экзоцитозу муциновых гранул.
Для выявления молекулярных механизмов, посредством которых MARCKS является звеном во взаимосвязи активации киназы с секрецией муцина, были проведены тщательные исследования фосфорилирования MARCKS в ответ на активацию PKC/PKG. Как проиллюстрировано на фиг.1А, PMA (100 нМ), вероятно, индуцирует значительное увеличение (в 3-4 раза) уровня фосфорилирования MARCKS в клетках NHBE, и такое фосфорилирование было ослаблено под действием ингибитора PKC, кальфостина C (500 нМ). После фосфорилирования MARCKS траспортируется из плазматической мембраны в цитоплазму (фиг. 1B). Более конкретно, на фиг.1A показано, что активация PKC приводит к фосфорилированию MARCKS в клетках NHBE. Клетки метили [32P]ортофосфатом в течение 2 часов, а затем обрабатывали стимулирующими и/или ингибирующими реагентами. Фосфорилирование MARCKS в ответ на данную обработку оценивали с помощью иммунопреципитации, как описано выше. Дорожка 1, среда (контроль); дорожка 2, носитель, 0,1% Me2SO; дорожка 3, 100 нМ 4α-PMA; дорожка 4, 100 нМ PMA; дорожка 5, 100 нМ PMA+500 нМ, кальфостина C; дорожка 6, 500 нМ кальфостина C. На фиг. 1B продемонстрировано, что фосфорилированный MARCKS транспортируется из плазматической мембраны в цитоплазму. 32P-меченные клетки обрабатывали PMA (100 нМ) или одной средой в течение 5 минут, а затем выделяли мембрану и цитозольные фракции. Активация PKG под действием 8-Br-cGMP (1 мкМ), то есть другое событие активации киназы, необходимое для стимуляции секреции муцина, не приводила к фосфорилированию MARCKS, и действительно, наблюдался обратный эффект: фосфорилирование MARCKS, индуцированное PMA, прекращалось под действием 8-Br-cGMP (фиг.2A). Этот эффект 8-Br-cGMP не был обусловлен подавлением активности PKC, поскольку PMA-индуцированное фосфорилирование может быть элиминировано последующим добавлением 8-Br-cGMP в клетки (фиг.2B). Следовательно, активация PKG, вероятно, приводит к дефосфорилированию MARCKS.
Дополнительное исследование продемонстрировало, что PKG-индуцированное дефосфорилирование MARCKS блокировалось под действием 500 нМ окадаиновой кислоты, ингибитора протеинфосфатазы (типа 1 и/или 2A (PP1/2A))(фиг.2A, дорожка 6). Таким образом, очевидно, что дефосфорилирование опосредуется PPl и/или PP2A. Для определения подтипа действующей протеинфосфатазы, в дополнительных исследованиях по фосфорилированию был использован новый и более специфичный ингибитор PP2A, фостриецин (IC50=3,2 нМ). Как проиллюстрировано на фиг.2C, фостриецин ингибирует PKG-индуцированное дефосфорилирование MARCKS в зависимости от концентрации (1-500 нМ), что позволяет предположить, что PKG индуцирует дефосфорилирование посредством активации PP2A. Для подтверждения дополнительной активации PP2A под действием PKG в клетках NHBE определяли активности цитозольных PP1 и PP2A после обработки клеток реагентом 8-Br-cGMP. Активность PP2A увеличивалась приблизительно в 3 раза (от 0,1 до 0,3 нмоль/мин/мг белка, p<0,01) при концентрациях 8-Br-cGMP, составляющих до 0,1 мкМ, а активность PPl не изменялась. Полученные данные показали, что PP2A может активироваться под действием PKG и является ответственной за дефосфорилирование MARCKS. В соответствии с этим такая активность PP2A, очевидно, играет решающую роль в секреции муцина; причем, если PKG-индуцированное дефосфорилирование MARCKS блокируется под действием окадаиновой кислоты или фостриецина, то секреторный ответ на активацию PKC/PKG или UTP-стимуляцию улучшался (фиг.3).
MARCKS ассоциируется с актином и миозином в цитоплазме
На фиг.4 проиллюстрирован иммунопреципитационный анализ с радиоактивным мечением, в котором было выявлено, что MARCKS может ассоциироваться с двумя другими белками (примерно 200 и примерно 40 кДа) в цитоплазме. На фиг.4, клетки NHBE были помечены [3Н]лейцином и [3Н]пролином в течение ночи, а мембранные и цитозольные фракции были получены, как описано в разделе «Экспериментальные процедуры». Выделенные фракции предварительно очищали с использованием преиммунного контрольного антитела (6F6). Затем цитозоль разделяли на две равные фракции и использовали для иммунопреципитации, осуществляемой в присутствии 10 мкМ цитохалазина D (Biomol, Plymouth Meeting, Pa.) с использованием анти-MARCKS антитела 2Fl2 (дорожка 2) и преиммунного контрольного антитела 6F6 (дорожка 3), соответственно. Белок MARCKS в мембранной фракции также анализировали с помощью иммунопреципитации, проводимой с использованием антитела 2Fl2 (дорожка 1). Осажденный комплекс белка разделяли с помощью электрофореза в 8% полиакриламидном геле с ДСН и визуализировали с помощью авторадиографии с применением усиливающих экранов. Очевидно, что MARCKS ассоциируется с двумя цитоплазматическими белками, имеющими молекулярную массу примерно 200 и 40 кДа, соответственно. Эти два MARCKS-ассоциированных белка вырезали из геля и анализировали с помощью матричной лазерной масс-спектрометрии с десорбционной ионизацией/времяпролетной масс-спектрометрии/внутреннего секвенирования (the Protein/DNA Technology Center of Rockefeller University, N. Y.). Полученные данные массы и последовательности пептида использовали для поиска в базе данных белков с помощью программ Интернета ProFound и MS-Fit. Результаты показали, что такими белками являются миозин (тяжелая цепь, немышечный типа A) и актин, соответственно. Анализ, проводимый с помощью матричной лазерной масс-спектрометрии с десорбционной ионизацией/времяпролетной масс-спектрометрии/внутреннего секвенирования, показал, что эти два MARCKS-ассоциированных белка представляют собой миозин (тяжелая цепь, немышечный типа A) и актин, соответственно.
Эти исследования позволяют выдвинуть новую парадигму относительно механизма передачи сигнала, регулирующего экзоцитозную секрецию муциновых гранул дыхательных путей, а также указывают на то, что такой механизм, очевидно, является первым прямым свидетельством конкретной биологической функции MARCKS в физиологическом процессе. MARCKS служит в качестве ключевой молекулы-медиатора, регулирующей высвобождение муциновых гранул из человеческих эпителиальных клеток дыхательных путей. Очевидно, что для секреции муцина дыхательных путей требуется двойная активация и синергическое действие PKC и PKG. Активированный PKC фосфорилирует MARCKS, что приводит к транспорту MARCKS из внутренней поверхности плазматической мембраны в цитоплазму. Активация PKG, в свою очередь, приводит к активации PP2A, которая дефосфорилирует MARCKS в цитоплазме. Поскольку способность MARCKS связываться с мембраной зависит от его состояния фосфорилирования, то такое дефосфорилирование позволяет белку MARCKS снова приобретать способность связываться с мембраной и присоединяться к мембранам цитоплазматических муциновых гранул. Благодаря такому взаимодействию с актином и миозином в цитоплазме (фиг.4), MARCKS может обладать способностью связывать гранулы с клеточным сократительным аппаратом и тем самым опосредовать транспорт гранул на периферию клеток и последующее высвобождение посредством экзоцитоза. Широкое распространение MARCKS позволяет предположить возможность того, что этот или аналогичный механизм может регулировать секрецию мембраносвязанных гранул в клетках различных типов при нормальных или патологических состояниях.
Как показано на фиг.5, MARCKS может функционировать как молекулярный линкер посредством взаимодействия с мембранами гранул у его N-концевого домена и связывания с актиновыми филаментами в сайте PSD и тем самым связывать гранулы с сократительным цитоскелетом для последующего транспорта и экзоцитоза. На фиг.5 показан возможный механизм, указывающий на то, что секретагон муцин взаимодействует с эпителиальными (бокаловидными) клетками дыхательных путей и активирует две отдельные протеинкиназы, PKC и PKG. Активированная PKC фосфорилирует MARCKS, что приводит к транспорту MARCKS из плазматической мембраны в цитоплазму, тогда как PKG, активируемая по пути «окись азота (NO) → GC-S → cGMP → PKG», в свою очередь, активирует цитоплазматическую PP2A, которая дефосфорилирует MARCKS. Такое дефосфорилирование стабилизирует присоединение MARCKS к мембранам гранул. Кроме того, MARCKS также взаимодействует с актином и миозином и тем самым связывает гранулы с клеточным сократительным аппаратом для последующего транспорта и высвобождения медиаторов воспаления, таких как MPO, посредством экзоцитоза. Присоединение MARCKS к гранулам после его высвобождения в цитоплазму может также управляться специфическими нацеливающими белками или некоторыми другими формами белок-белковых взаимодействий, в которых участвует N-концевой домен MARCKS. В любом случае, пептид MANS или его активный фрагмент, содержащий по меньшей мере 4 аминокислоты, должен конкурентно ингибировать нацеливание MARCKS на мембраны муциновых гранул и тем самым блокировать секрецию.
Настоящее изобретение также относится к новому способу блокирования любого клеточного серекторного процесса посредством экзоцитоза, а в частности высвобождения медиаторов воспаления из гранул, содержащихся в воспалительных клетках, в путях стимуляции которых участвует белок MARCKS, который является субстратом для протеинкиназы С (PKC), и высвобождения содержимого из мембраносвязанных везикул. В частности, авторами настоящего изобретения было показано, что стимулированное высвобождение медиатора воспаления, миелопероксидазы, из человеческих (фиг.6) или собачьих (фиг.7) нейтрофилов может блокироваться пептидом MANS в зависимости от концентрации. В частности, на фиг.6 показаны выделенные нейтрофилы, которые были стимулированы 100 нМ PMA и 10 мкМ 8-Br-cGMP для секреции миелопероксидазы (MPO). 100 мкМ пептида MANS снижает уровень секреции MPO до контрольных уровней (*=p<0,05). 10 мкМ MANS вызывает небольшое снижение уровня секреции MPO. 10 или 100 мкМ контрольного пептида (RNS) не оказывали какого-либо влияния на секрецию MPO. На фиг.7 показаны выделенные нейтрофилы, которые были стимулированы 100 нМ PMA и 10 мкМ 8-Br-cGMP для секреции миелопероксидазы (MPO). 100 мкМ пептида MANS снижает уровень секреции MPO до контрольных уровней (*=p<0,05). 10 мкМ MANS вызывает небольшое снижение уровня секреции MPO. 10 или 100 мкМ контрольного пептида (RNS) не оказывали какого-либо влияния на секрецию MPO. Таким образом, указанный пептид может иметь терапевтическое применение для блокирования высвобождения медиаторов воспаления, секретируемых из инфильтрирующихся воспалительных клеток в любых тканях. Многие из этих высвобождаемых медиаторов ответственны за обширное поражение ткани, наблюдаемое при различных хронических воспалительных заболеваниях (то есть при респираторных заболеваниях, таких как астма, хронический бронхит и ХОБЛ, при воспалительных заболеваниях кишечника, включая язвенный колит и болезнь Крона, при аутоиммунных заболеваниях, при кожных болезнях, таких как розацея, экзема и угревая сыпь в тяжелой форме, и при артритных и болевых синдромах, таких как ревматоидный артрит и фибромиалгия). Настоящее изобретение может быть также применено для лечения заболеваний, таких как артрит, хронический бронхит, ХОБЛ и кистозный фиброз. В соответствии с этим настоящее изобретение может быть также применено для лечения заболеваний человека и животных, а в частности заболеваний, поражающих лошадей, собак, кошек и других домашних питомцев.
На фиг.8-12 показана секреция MPO у человека и собак. Во всех этих экспериментах выделенные нейтрофилы стимулировали ЛПС при концентрации 1×10-6 M в течение 10 минут при 37°С с последующим добавлением стимуляторов, как указано на чертежах. Клетки примировали липополисахаридами (ЛПС) так, чтобы они могли отвечать на действие секретагона.
В одном из своих вариантов, настоящее изобретение относится к способу регуляции воспаления у индивидуума, где указанный способ включает введение терапевтически эффективного количества фармацевтической композиции, содержащей пептид MANS или его активный фрагмент. В одном из аспектов указанного варианта, указанный активный фрагмент белка MANS содержит по меньшей мере четыре, а предпочтительно, шесть аминокислот. В другом аспекте изобретения, указанное воспаление вызывается респираторными заболеваниями, кишечными заболеваниями, кожными болезнями, аутоиммунными заболеваниями и болевыми синдромами. В другом аспекте изобретения, указанные респираторные заболевания выбраны из группы, состоящей из астмы, хронического бронхита и ХОБЛ. В другом аспекте изобретения, указанные кишечные заболевания выбраны из группы, состоящей из язвенного колита, болезни Крона и синдрома раздраженного кишечника. В другом аспекте изобретения, указанные кожные болезни выбраны из группы, состоящей из розацеи, экземы, псориаза и угревой сыпи в тяжелой форме. В другом аспекте изобретения, указанное воспаление вызывается артритом или кистозным фиброзом. В другом аспекте изобретения, указанным индивидуумом является млекопитающее. Кроме того, в другом аспекте изобретения, указанное млекопитающее выбрано из группы, состоящей из человека, собак, лошадей и кошек. В другом аспекте изобретения, указанная стадия введения выбрана из группы, состоящей из местного введения, парентерального введения, ректального введения, внутрилегочного введения, интраназального введения, введения путем ингаляции и перорального введения. В другом аспекте изобретения, указанное внутрилегочное введение выбрано из группы, состоящей из введения с помощью аэрозоля, инсуффлятора, ингалятора с дозируемым клапаном и небулайзера.
В другом своем варианте, настоящее изобретение относится к способу регуляции клеточного секреторного процесса у индивидуума, где указанный способ включает введение терапевтически эффективного количества фармацевтической композиции, содержащей по меньшей мере одно соединение, включающее пептид MANS или его активный фрагмент, которые регулируют высвобождение медиатора воспаления у индивидуума. В одном из аспектов указанного варианта, указанный активный фрагмент белка MANS содержит по меньшей мере четыре, а предпочтительно, шесть аминокислот. В другом аспекте изобретения, указанной регуляцией клеточного секреторного процесса является блокирование или ослабление клеточного секреторного процесса. В другом аспекте изобретения, указанное высвобождение медиатора воспаления вызывается респираторными заболеваниями, кишечными заболеваниями, кожными болезнями, аутоиммунными заболеваниями и болевыми синдромами. В другом аспекте изобретения, указанные респираторные заболевания выбраны из группы, состоящей из астмы, хронического бронхита и ХОБЛ. В другом аспекте изобретения, указанные кишечные заболевания выбраны из группы, состоящей из язвенного колита, болезни Крона и синдрома раздраженного кишечника. В другом аспекте изобретения, указанные кожные болезни выбраны из группы, состоящей из розацеи, экземы, псориаза и угревой сыпи в тяжелой форме. В другом аспекте изобретения, указанное воспаление вызывается артритом или кистозным фиброзом. В другом аспекте изобретения, указанным индивидуумом является млекопитающее. В другом аспекте изобретения, указанное млекопитающее выбрано из группы, состоящей из человека, собак, лошадей и кошек. В другом аспекте изобретения, указанная стадия введения выбрана из группы, состоящей из местного введения, парентерального введения, ректального введения, внутрилегочного введения, интраназального введения, введения путем ингаляции и перорального введения. В другом аспекте изобретения, указанное внутрилегочное введение выбрано из группы, состоящей из введения с помощью аэрозоля, инсуффлятора, ингалятора с дозируемым клапаном и небулайзера.
В другом своем варианте, настоящее изобретение относится к способу ослабления воспаления у индивидуума, где указанный способ включает введение терапевтически эффективного количества соединения, ингибирующего MARCKS-ассоциированное высвобождение медиаторов воспаления, где указанное высвобождение медиаторов воспаления у индивидуума снижается по сравнению с высвобождением, наболюдаемым в отсутствие такого введения. В одном из аспектов данного варианта, указанным соединением является по меньшей мере один активный фрагмент белка MARCKS. В другом аспекте изобретения, указанный активный фрагмент имеет длину по меньшей мере четыре, а предпочтительно, шесть аминокислот. В другом аспекте изобретения, указанным соединением является пептид MANS или его активный фрагмент. В другом аспекте изобретения, указанным соединением является антисмысловой олигонуклеотид, направленный против кодирующей последовательности белка MARCKS или его активного фрагмента. В другом аспекте изобретения, указанный активный фрагмент имеет длину по меньшей мере четыре, а предпочтительно, шесть аминокислот.
В другом своем варианте, настоящее изобретение относится к способу ослабления воспаления у индивидуума, где указанный способ включает введение терапевтически эффективного количества фармацевтически активной композиции, включающей соединение, ингибирующее MARCKS-ассоциированное высвобождение медиаторов воспаления, где указанное воспаление у индивидуума снижается по сравнению с воспалением, наблюдаемым в отсутствие такого введения. В одном из аспектов данного варианта, указанным соединением является активный фрагмент белка MARCKS. В другом аспекте изобретения, указанный активный фрагмент имеет длину по меньшей мере четыре, а предпочтительно, шесть аминокислот. В другом аспекте изобретения, указанным соединением является пептид MANS или его активный фрагмент. В другом аспекте изобретения, указанным соединением является антисмысловой олигонуклеотид, направленный против кодирующей последовательности белка MARCKS или его активного фрагмента. В другом аспекте изобретения, указанный активный фрагмент имеет длину по меньшей мере четыре, а предпочтительно, шесть аминокислот. Настоящее изобретение также относится к композиции, содержащей один или несколько пептидов MANS или его активных фрагментов, и к ее применению в лечении путем ингибирования высвобождения медиаторов воспаления из гранул или везикул воспалительных клеток.
В другом своем варианте, настоящее изобретение относится к способу ослабления или ингибирования воспаления у индивидуума, где указанный способ включает введение терапевтически эффективного количества по меньшей мере одного пептида, включающего пептид MANS или его активный фрагмент, которые являются эффективными для ингибирования или подавления высвобождения медиатора воспаления в области воспаления. В одном из аспектов указанного варианта, указанный активный фрагмент имеет длину по меньшей мере четыре, а предпочтительно, по меньшей мере шесть аминокислот. В другом аспекте изобретения, указанные медиаторы воспаления продуцируются клетками, выбранными из группы, состоящей из нейтрофилов, базофилов, эозинофилов, моноцитов и лейкоцитов. Предпочтительными клетками являются лейкоциты, более предпочтительными клетками являются гранулоциты, а еще более предпочтительными являются нейтрофилы, базофилы, эозинофилы или их комбинации. В другом аспекте изобретения, указанный агент вводят перорально, парентерально, в полость органа, ректально или через дыхательные пути. В другом аспекте изобретения, указанная композиция также содержит вторую молекулу, выбранную из группы, состоящей из антибиотика, противовирусного соединения, антипаразитарного соединения, противовоспалительного соединения и иммунодепрессанта.
Активный фрагмент пептида MANS может быть выбран из группы, состоящей из пептидов, описанных в таблице 1. Как описано в настоящей заявке, такие пептиды могут содержать, но необязательно, химические молекулы у N-концевой и/или С-концевой аминокислоты.
В другом аспекте изобретения, способы, описанные в настоящем изобретении, могут быть осуществлены путем применения или введения комбинаций пептидов, описанных в таблице 1 настоящего изобретения, то есть применения или введения одного или нескольких из указанных пептидов. В описанных здесь способах, предпочтительно, используют или вводят один пептид.
В ответ на активацию протеинкиназы С (PKC) под действием стимулятора воспаления, дегрануляция клеток, выбранных из группы, состоящей из нейтрофилов, эозинофилов, моноцитов/макрофагов и лимфоцитов, может быть ослаблена путем предварительного инкубирования и совместного инкубирования с пептидом, идентичным N-концевой области белка MARCKS, где указанный пептид выбран из группы фрагментов пептида MANS, описанного в таблице 1. Хотя время курса лечения и концентрации для клеток различных типов могут варьироваться, однако во всех случаях пептид MANS будет вызывать ослабление PKC-индуцированной дегрануляции.
Описанное здесь изобретение будет продемонстрировано ниже на некоторых примерах лишь в целях иллюстрации, и эти примеры не должны рассматриваться как ограничение объема изобретения. Примеры
Методы и материалы
Иммунопреципитационный анализ с радиоактивным мечением
При мечении [32P]фосфатом, клетки предварительно инкубировали в течение 2 часов в модифицированной по способу Дульбекко среде Игла, не содержащей фосфата и содержащей 0,2% альбумин бычьей сыворотки, а затем метили 0,1 мКи/мл [32P]ортофосфата (9000 Ки/ммоль, PerkinElmer Life Sciences) в течение 2 часов. Для мечения [3Н]миристиновой кислотой или 3Н-аминокислотами клетки инкубировали в течение ночи в среде, содержащей 50 мкКи/мл [3Н]-миристиновой кислоты (49 Ки/ммоль, PerkinElmer Life Sciences) или 0,2 мКи/мл [3Н]лейцина (159 Ки/ммоль, PerkinElmer Life Sciences) плюс 0,4 мКи/мл [3Н]пролина (100 Ки/ммоль, PerkinElmer Life Sciences). После мечения клетки обрабатывали стимулирующими реагентами в течение 5 минут. Если использовали ингибитор, то клетки предварительно инкубировали с ингибитором в течение 15 минут, а затем подвергали стимуляции. По окончании обработки, клетки подвергали лизису в буфере, содержащем 50 мМ трис-HCl (pH 7,5), 150 мМ NaCl, 1 мМ EDTA, 10% глицерина, 1% Nonidet P-40, 1 мМ фенилметилсульфонилфторида, 1 мМ бензамидина, 10 мкг/мл пепстатина A и 10 мкг/мл лейпептина. С помощью преципитации трихлоруксусной кислотой и сцинтилляционного считывания можно определить эффективность радиоактивного мечения в каждой культуре. Иммунопреципитацию белка MARCKS осуществляли методом, описанным Spizz и Blackshear, с использованием клеточных лизатов, содержащих равное количество импульсов/мин. Spizz et al., J. Biol. Chem. 271, 553-562 (1996). Осажденные белки разделяли с помощью электрофореза в 8% полиакриламидном геле с ДСН и визуализировали с помощью авторадиографии. В этом анализе использовали антитело против человеческого MARCKS (2Fl2) и преиммунное антитело в качестве контроля (6F6).
Для оценки MARCKS или MARCKS-ассоциированных белковых комплексов в различных субклеточных фракциях радиоактивно меченные и обработанные клетки соскабливали в буфер для гомогенизации (50 мМ трис-HCl (pH 7,5), 10 мМ NaCl, 1 мМ EDTA, 1 мМ фенилметилсульфонилфторида, 1 мМ бензамидина, 10 мкг/мл пепстатина A, 10 мкг/мл лейпептина), а затем разрушали путем кавитации азотом (800 фунтов/кв.дюйм, в течение 20 минут при 4°С). Клеточные лизаты центрифугировали при 600×g в течение 10 минут при 4°С для удаления ядер и неразрушенных клеток. Супернатанты, полученные после удаления ядер, разделяли на мембранные и цитозольные фракции путем ультрацентрифугирования при 400000×g в течение 30 минут при 4°С. Мембранный осадок солюбилизировали в буфере для лизиса путем обработки ультразвуком. Затем осуществляли иммунопреципитацию, как описано выше.
MARCKS-ассоциированные пептиды
Пептиды с миристоилированной N-концевой последовательностью (MANS) и рандомизированной N-концевой последовательностью (RNS) синтезировали в лаборатории Genemed Synthesis, Inc. (San Francisco, Calif.), а затем очищали с помощью жидкостной хроматографии высокого давления (чистота >95%) и подтверждали с помощью масс-спектроскопии, где было показано, что каждый отдельный пик имел соответствующую молекулярную массу. Пептид MANS состоит из последовательности, идентичной первым 24 аминокислотам MARCKS, то есть миристоилированной N-концевой области, которая опосредует встраивание MARCKS в мембраны, MA-GAQFSKTAAKGEAAAERPGEAAVA (SEQ ID NO:1 (где MA представляет собой N-концевую миристоильную цепь)). Соответствующий контрольный пептид (RNS) имеет такой же аминокислотный состав, как пептид MANS, но его последовательность содержит аминокислоты, расположенные в произвольном порядке, MA- GTAPAAEGAGAEVKRASAEAKQAF (SEQ ID NO:232). Присутствие гидрофобной миристатной молекулы в этих синтетических пептидах приводит к повышению их способности проникать через плазматические мембраны, что позволяет клеткам легко поглощать эти пептиды. Для определения влияния этих пептидов на секрецию муцина клетки предварительно инкубировали с пептидами в течение 15 минут, а затем добавляли секретагоны, после чего оценивали секрецию муцина с помощью ELISA.
Антисмысловые олигонуклеотиды
Антисмысловой олигонуклеотид MARCKS и его соответствующий контрольный олигонуклеотид синтезировали в лаборатории Biognostik GmbH (Gottingen, Germany). Клетки NHBE обрабатывали 5 мкМ антисмыслового или контрольного олигонуклеотида в течение максимум 3 дней (в присутствии 2 мкг/мл липофектина в течение первых 24 часов). Затем клетки инкубировали с секретагонами, и секрецию муцина оценивали с помощью ELISA. Из обработанных клеток выделяли полноразмерные РНК и белок. мРНК MARCKS оценивали с помощью нозерн-гибридизации в соответствии со стандартными процедурами с использованием человеческой кДНК MARCKS в качестве зонда. Уровень белка MARCKS определяли с помощью вестерн-блот-анализа с использованием очищенного анти-MARCKS IgGl (клон 2Fl2) в качестве «первого» детектирующего антитела.
Временная трансфекция
Домен сайта фосфорилирования (PSD) MARCKS содержит PKC-зависимые сайты фосфорилирования и сайт, связывающийся с актиновым филаментом. Для конструирования PSD-делетированной кДНК MARCKS генерировали два фрагмента, фланкирующие последовательность PSD (кодирующую 25 аминокислот), с помощью полимеразной цепной реакции, а затем лигировали посредством сайта Xhol, который был присоединен к 5'-концам олигонуклеотидных праймеров, сконструированных для проведения полимеразной цепной реакции. Полученные мутированную кДНК и кДНК MARCKS дикого типа встраивали в экспрессионный вектор млекопитающего pcDNA4/TO (Invitrogen, Carlsbad, Calif.). Выделенные рекомбинантные конструкции подтверждали путем гидролиза рестриктирующими ферментами и секвенирования ДНК.
HBEl представляет собой трансформированную папиломавирусом человеческую бронхиальную эпителиальную клеточную линию, способную секретировать муцин при ее культивировании на границе раздела фаз воздух/жидкость. Трансфекцию клеток HBEl проводили с использованием трансфецирующего реагента Effectene (Qiagen, Valencia, Calif.) в соответствии с инструкциями производителей. Вкратце, дифференцированные клетки HBEl, выращенные на границе раздела фаз воздух/жидкость, подвергали диссоциации трипсином/EDTA и снова засевали в 12-луночные планшеты для культивирования клеток при плотности 1×105 клеток/см2. После инкубирования в течение ночи клетки трансфецировали кДНК MARCKS дикого типа, кДНК MARCKS, не содержащей PSD, или векторной ДНК. Клетки культивировали в течение 48 часов для экспрессии гена, а затем обрабатывали секретагоном и измеряли уровень секреции муцина с помощью ELISA. Все процедуры трансфекции осуществляли в присутствии плазмиды pcDNA4/TO/lacZ (Invitrogen) (отношение ДНК 6:1, всего 1 мкг ДНК, отношение ДНК к реагенту Effectene = 1:25) для мониторинга изменения эффективности трансфекции. Полученные результаты не выявили какого-либо значимого различия в бета-галактозидазных активностях клеточных лизатов, выделенных из трансфецированных клеток, что указывает на то, что различные ДНК-конструкции обладают аналогичной эффективностью трансфекции (данные не приводятся).
Анализ на протеинфосфатазную активность
Активности PPl и PP2A измеряли с использованием системы анализов на протеинфосфатазную активность (Life Technologies, Inc.), известных специалистам, с небольшими модификациями. Huang et al. Adv. Exp. Med Biol. 396, 209-215 (1996). Вкратце, клетки NHBE обрабатывали 8-Br-cGMP или только средой в течение 5 минут. Затем клетки соскабливали в буфер для лизиса (50 мМ трис-HCl (pH 7,4), 0,1% бета-меркаптоэтанола, 0,1 мМ EDTA, 1 мМ бензамидина, 10 мкг/мл пепстатина A, 10 мкг/мл лейпептина) и подвергали дизрупции под действием ультразвука в течение 20 секунд при 4°С. Клеточные лизаты центрифугировали, и супернатанты сохраняли для проведения анализа на фосфатазную активность. Данный анализ проводили с использованием 32P-меченой фосфорилазы А в качестве субстрата. Уровень высвобождаемого 32Pi определяли путем сцинтилляционного подсчета. Концентрацию белка в каждом образце определяли с помощью анализа Брэдфорда. Активность PP2A выражали как общую фосфатазную активность образца минус остаточную активность в присутствии 1 нМ окадаиновой кислоты. Активность PPl выражали как разность между остаточными активностями в присутствии 1 нМ и 1 мкМ окадаиновой кислоты, соответственно. Протеинфосфатазные активности регистрировали как нмоль Pi, высвобождаемого в мин/мг общего белка.
Анализ на цитотоксичность
Все реагенты, используемые для обработки клеток NHBE, оценивали на цитотоксичность путем определения уровня общего высвобождения лактат-дегидрогеназы из клеток. Анализ осуществляли с использованием набора Promega Cytotox 96 в соответствии с инструкциями производителей. Все эксперименты осуществляли с использованием реагентов в нецитотоксических концентрациях.
Статистический анализ
Полученные данные анализировали на статистическую значимость с помощью одностороннего дисперсионного анализа с поправками Бонферрони, применяемыми после проведения такого анализа (post-test). Различия между группами обработки считались значимыми при p<0,05.
Выделение PMN из крови собак
Стадии выделения PMN включают сбор 10 мл антикоагулированной крови в растворе ACD. Затем 5 мл крови наносили слоями на 3,5 мл среды для выделения PMN, так чтобы указанная среда для выделения PMN (IM) поддерживалась при комнатной температуре (RT). После этого кровь центрифугировали при комнатной температуре в течение 30 минут, 550×g при 1700 об/мин. Самую нижнюю белую полосу переносили в коническую центрифужную 15 мл-пробирку (CCFT). Затем добавляли 2V HESS с 10% фетальной бычьей сывороткой (PBS) и центрифугировали при комнатной температуре в течение 10 минут, 400×g при 1400 об/мин. После этого осадок ресуспендировали в 5 мл 1-1ESS с PBS. Клеточную суспензию добавляли в 50 мл CCFT, содержащую 20 мл охлажденного льдом 0,88% NH4Cl, и пробирку переворачивали 2-3 раза. Полученный продукт центрифугировали в течение 10 минут, 800×g при 2000 об/мин, а затем отсасывали и ресуспендировали в 5 мл HBSS с FBS. Препарат анализировали путем подсчета клеток и цитоцентрифугирования, предпочтительно клеток цельной крови, число которых должно составлять 109-1011 клеток, и PMN, число которых должно составлять 2-4×107 клеток. В общих чертах, см. Wang et al., J. Immunol, «Neutrophil- induced changes in the biomechanical properties of endothelial cells: roles of ICAM-I and reactive oxygen species» 6487-94 (2000).
Колориметрический ферментный анализ на MPO-активность.
Образцы анализировали на MPO-активность в 96-луночных круглодонных микротитрационных планшетах с использованием набора для «сэндвич»-ELISA (R & D Systems, Minneapolis, Minn.). Вкратце, 20 микролитров образца смешивали с 180 микролитрами субстратной смеси, содержащей 33 мМ фосфата калия, pH 6,0, 0,56% тритона X-100, 0,11 мМ пероксида водорода и 0,36 мМ дигидрохлорида O-дианнизидина в одной лунке микротитрационного планшета. Конечные концентрации аналитической смеси составляли: 30 мМ фосфата калия, pH 6,0, 0,05% тритона X-100, 0,1 мМ пероксида водорода и 0,32 мМ дигидрохлорида O-дианнизидина. После смешивания аналитическую смесь инкубировали при комнатной температуре в течение 5 минут, и активность фермента MPO определяли на спектрофотометре на длине волны 550 нанометров. Образцы анализировали в дубликате.
Пример 1
Исследования секреции медиаторов воспаления
Были использованы лейкоциты четырех различных типов или модели, которые секретируют специфическое содержимое гранул в ответ на индуцируемую форболовым эфиром активацию PKC. Нейтрофилы выделяли из человеческой крови и оценивали in vitro высвобождение MPO этими клетками. Также оценивали высвобождение мембраносвязанных медиаторов воспаления из коммерчески доступных человеческих лейкоцитарных клеточных линий. Для оценки секреции ЕРО использовали клон 15 человеческой промиелоцитарной клеточной линии HL-60 (Fischkoff S.A. Graded increase in probability of eosinophilic differentiation of HL-60 promyelocytic leukemia cells induced by culture under alkaline conditions. Leuk Res 1988, 12:679-686, Rosenberg H.F., Ackerman S. J., Tenen D.G. Human eosinophil cationic protein: molecular cloning of a cytotoxin и helminthotoxin with ribonuclease activity. J. Exp. Med. 1989, 170:163-176, Tiffany H.L., Li F, Rosenberg H.F. Hyperglycosylation of eosinophil ribonucleases in a promyelocytic leukemia cell line и in differentiated peripheral blood progenitor cells. J. Leukoc. Biol. 1995, 58:49-54, Badewa A.P., Hudson C.E., Heiman A.S. Regulatory effects of eotaxin, eotaxin-2, and eotaxin-3 on eosinophil degranulation and superoxide anion generation. Exp. Biol. Med. 2002, 227:645-651). Для оценки секреции лизоцима использовали клеточную линию моноцитарного лейкоза U937 (Hoff T., Spencker T., Emmendoerffer A., Goppelt-Struebe M. Effects of glucocorticoids on the TPA-induced monocytic differentiation. J. Leukoc. Biol. 1992, 52: 173-182, Balboa M.A., Saez Y., Balsinde J. Calcium-independent phospholipase A2 is required for lysozyme secretion in U937 promonocytes. J. Immunol. 2003, 170:5276-5280, Sundstrom C, Nilsson K. Establishment and characterization of a human histiocytic lymphoma cell line (U-937). Int. J. Cancer 1976, 17:565-577). Для оценки высвобождения гранзима использовали лимфоцитарную клеточную линию природных киллеров NK-92 (Gong J.H., Maki G, Klingeniann H.G. Characterization of a human cell line (NK-92) with phenotypical and functional characteristics of activated natural killer cells. Leukemia 1994, 8:652-658, Maki G, Klingemann H.G, Martinson J.A, Tam Y.K. Factors regulating the cytotoxic activity of the human natural killer cell line, NK-92. J. Hematother Stem Cell Res 2001, 10:369-383, Takayama H, Trenn G, Sitkovsky M.V. A novel cytotoxic T lymphocyte activation assay. J Immunol Methods 1987, 104:183-190). Во всех случаях, клетки предварительно инкубировали с различными концентрациями синтетического пептида, идентичного 24 аминокислотам N-конца MARCKS (пептида MANS с миристоилированной N-концевой последовательностью; MA-GAQFSKTAAKGEAAAERPGEAAVA (SEQ ID NO:1), где MA представляет собой миристоил, присоединенный к N-концевому амину пептида посредством амидной связи), или контрольного миссенс-пептида (RNS: пептида с рандомизированной N-концевой последовательностью, MA-GTAPAAEGAGAEVKRASAEAKQAF, SEQ ID NO:232), последовательность которого состоит из тех же самых 24 аминокислот, но расположенных в произвольном порядке, где указанная последовательность менее чем на 13% идентична последовательности пептида MANS. Альтернативно, клетки предварительно обрабатывали одним из синтетических усеченных пептидов, перечисленных ниже в таблице 3.
В клетках каждого типа MANS, но не RNS ослабляет высвобождение медиаторов воспаления в зависимости от концентрации. Время наблюдения такого высвобождения может составлять 0,5-3,0 часа. Полученные результаты соответствуют результатам, полученным для N-концевой области белка MARCKS, участвующей во внутриклеточных путях, приводящих к дегрануляции лейкоцитов.
Выделение человеческих нейтрофилов
Эти исследования были проведены с разрешения Регуляторных органов, осуществляющих контроль за исследованиями с участием человека (IRB). Человеческие нейтрофилы выделяли, как описано выше (см. Takashi S, OkuboY, Horie S. Contribution of CD54 to human eosinophil and neutrophil superoxide production. J. Appl. Physiol. 2001, 91:613-622) с небольшими модификациями. Вкратце, гепаринизированную венозную кровь получали от нормальных здоровых добровольцев, разводили средой RPMI-1640 (Cellgro, Mediatech, Inc., Herndon, VA) в отношении 1:1, наносили на гистопак (с плотностью 1,077 г/мл, Sigma-Aldrich Co., St. Louis, MO) и центрифугировали при 400×g в течение 20 минут при 4°С. Супернатант и мононуклеарные клетки на границе раздела фаз осторожно удаляли, и эритроциты, находящиеся в осадке, подвергали лизису в охлажденной дистиллированной воде. Выделенные гранулоциты два раза промывали сбалансированным солевым раствором Хэнкса (HBSS) и ресуспендировали в HBSS на льду. Нейтрофилы, используемые в экспериментах, имели чистоту >98%, где примеси эозинофилов составляли <2%, а жизнеспособность нейтрофилов составляла >99%, как было определено методом исключения трипановым синим.
Измерение активности MPO, высвобождаемой нейтрофилами
Для измерения уровня высвобождения MPO, очищенные человеческие нейтрофилы, суспендированные в HBSS, разделяли на аликвоты при плотности 4×106 клеток/мл в 15 мл пробирках и предварительно инкубировали с 50 или со 100 мкМ MANS, RNS или одного из пептидов согласно изобретению в течение 10 минут при 37°С. Затем клетки стимулировали 100 нМ форбол-12-миристат-13-ацетата (PMA) в течение периода времени до 3 часов. Контрольный эталон (PMA-эталон) создавали с использованием очищенных человеческих нейтрофилов, суспендированных в HBSS и разделенных на аликвоты при плотности 4×106 клеток/мл в 15 мл-пробирках, где указанные клетки стимулировали 100 нМ форбол-12-миристат-13-ацетата (PMA) в отсутствие тестируемого пептида в течение того же самого периода времени. Реакцию завершали путем выдерживания пробирок на льду и центрифугирования при 400×g в течение 5 минут при 4°С.
Активность MPO в клеточном супернатанте анализировали с использованием тетраметилбензидина (TMB) в соответствии с разработанной ранее технологией (Abdel-Latif D, Steward M, Macdonald DL, Francis GA., Dinauer MC, Lacy P. Rac2 is critical for neutrophil primary granule exocytosis. Blood 2004, 104:832-839). Вкратце, 100 мкл раствора субстрата TMB добавляли к 50 мкл клеточных супернатантов или стандартной человеческой MPO (EMD Biosciences, Inc., San Diego, CA) в 96-луночном микропланшете, а затем инкубировали при комнатной температуре в течение 15 минут. Реакцию завершали путем добавления 50 мкл 1M H2SO4 и оптическую плотность считывали при 450 нм на спектрофотометрическом микропланшет-ридере (VERSA max, Molecular Devices, Sunnyvale, CA).
Исследования культуры лейкоцитов
Клеточные линии человеческих лейкоцитов трех типов, а в частности клон 15 человеческой промиелоцитарной клеточной линии HL-60, моноцитарную клеточную линию U937 и лимфоцитарную клеточную линию природных киллеров NK-92 закупали в Американской коллекции типовых культур (ATCC, Rockville, MD). Клетки клона 15 HL-60 (ATCC CRL-1964) поддерживали в среде, состоящей из среды RPMI 1640 с L-глутамином, в которую были добавлены 10% термоинактивированная фетальная бычья сыворотка (Gibco, Invitrogen Co., Carlsbad, CA), 50 МЕ/мл пенициллина, 50 мкг/мл стрептомицина и 25 мМ буфера HEPES, pH 7,8, при 37°С в атмосфере, содержащей 5% CO2. Конечная дифференцировка в клетки эозинофил-подобного фенотипа была иницирована путем культивирования клеток при плотности 5×105 клеток/мл в вышеуказанной среде, содержащей 0,5 мМ масляную кислоту (Sigma-Aldrich Co.), в течение 5 дней как описано ранее (Tiffany HL, Li F, Rosenberg HF. Hyperglycosylation of eosinophil ribonucleases in a promyelocytic leukemia cell line and in differentiated peripheral blood progenitor cells. J Leukoc. Biol. 1995, 58:49-54, Tiffany H.L, Alkhatib G, Combadiere C, Berger E.A, Murphy P.M. CC chemokine receptors 1 and 3 are differentially regulated by IL-5 during maturation of eosinophilic HL-60 cells. J Immunol. 1998, 160:1385-1392). Клетки U937 (ATCC CRL-1593.2) культивировали при 37°С в атмосфере с 5% CO2 в полной среде, состоящей из среды RPMI 1640 с L-глутамином, в которую были добавлены 10% FBS, 50 МЕ/мл пенициллина и 50 мкг/мл стрептомицина. Клетки NK-92 (ATCC CRL-2407) поддерживали в среде альфа-MEM (Sigma-Aldrich Co.), в которую было добавлено 20% FBS, 100 ед/мл интерлейкина-2 (IL-2) (Chemicon International, Inc., Temecula, CA), 5×10-5M 2- меркаптоэтанола, 50 МЕ/мл пенициллина и 50 мкг/мл стрептомицина при 37°С в атмосфере, содержащей 5% CO2. Морфологию клеток оценивали методом окрашивания по Райту-Гимзе. Жизнеспособность собранных для эксперимента клеток оценивали методом исключения трипановым синим и использовали популяцию клеток с жизнеспособностью >95%.
Инкубирование клеток для анализов на дегрануляцию
Клон 15 клеток HL-60, клетки U937 и клетки NK-92 промывали и ресуспендировали при плотности 2,5×106 клеток/мл в среде RPMI-1640, не содержащей фенолового красного (Cellgro, Mediatech, Inc.), для проведения всех анализов на дегрануляцию. Аликвоты клеток в 15 мл-пробирках предварительно инкубировали с указанными концентрациями MANS, RNS или тестируемого пептида в течение 10 минут при 37°С. Затем клетки стимулировали PMA в течение периода времени до 2 часов. Контрольный эталон (PMA-эталон) создавали для каждого типа клеток, а именно клона 15 клеток HL-60, клеток U937 и клеток NK-92, соответственно, которые промывали и ресуспендировали при плотности 2,5×106 клеток/мл в среде RPMI-1640, не содержащей фенолового красного, а затем стимулировали PMA в отсутствие MANS, RNS или тестируемого пептида в течение того же самого периода времени. Реакцию завершали путем выдерживания пробирок на льду и центрифугирования клеток при 400×g в течение 5 минут при 4°С.
Для измерения уровня высвобождаемой MPO из нейтрофилов и высвобождаемого лизоцима из клеток U937 авторами настоящего изобретения была проведена количественная оценка уровня секреции с использованием человеческой MPO и овальбумина яичного белка в качестве стандартов, соответственно. Для количественной оценки уровня высвобождаемой EPO из клона 15 клеток HL-60 и высвобождаемого гранзима из клеток NK-92, стандарты не использовали. А поэтому измеряли уровни высвобождения EPO и гранзима и их внутриклеточные уровни (лизированных клеток), и эти уровни секретированных EPO и гранзима выражали как процент от общего уровня каждого фермента (уровня внутриклеточного и секретированного фермента). Для измерения уровня внутриклеточного EPO в клоне 15 клеток HL-60 и внутриклеточного гранзима в клетках NK-92 брали соответствующие аликвоты клеток, лизированных 0,1% тритоном X-100, в целях проведения количественной оценки внутриклеточных белков гранул, описанных ниже. Для минимизации вариабельности между культурами, каждую обработку выражали как процент от контроля.
Измерение уровня высвобождения EPO клетками HL-60
Активность EPO, высвобождаемой клоном 15 клеток HL-60, анализировали с использованием TMB в соответствии с разработанной ранее технологией (Lacy P, Mahmudi-Azer S, Bablitz B, Hagen S.C, Velazquez J.R, Man S.F, Moqbel R. Rapid mobilization of intracellular stored RANTES in response to interferon-gamma in human eosinophils. Blood 1999, 94:23-32). Таким образом, 100 мкл раствора субстрата TMB добавляли к 50 мкл (мкл = микролитров) образца в 96-луночном микропланшете, а затем инкубировали при комнатной температуре в течение 15 минут. Реакцию завершали путем добавления 50 мкл 1,0M H2SO4 и оптическую плотность считывали при 450 нм на спектрофотометрическом микропланшет-ридере. Количество секретируемой ЕРО выражали как процент от общего количества фермента, присутствующего в том же самом количестве клеток, лизированных тритоном X-100.
Измерение уровня секреции лизоцима моноцитами
Уровень лизоцима, секретируемого клетками U937, измеряли с помощью спектрофотометрического анализа как описано ранее (Balboa MA, Saez Y, Balsinde J. Calcium-independent phospholipase A2 is required for lysozyme secretion in U937 promonocytes. J. Immunol 2003, 170:5276-5280) с небольшими модификациями. Таким образом, 100 мкл образца смешивали со 100 мкл суспензии Micrococcus lysodeikticus (Sigma-Aldrich Co.) (0,3 мг/мл в 0,1 M фосфатно-натриевого буфера, pH 7,0) в 96-луночном микропланшете. Снижение оптической плотности при 450 нм измеряли при комнатной температуре. Калибровочную кривую строили с использованием лизоцима куриного яичного белка (EMD Biosciences, Inc.) в качестве стандарта.
Измерение уровня секреции гранзима NK-клетками
Уровень гранзима, секретируемого клетками NK-92, измеряли путем оценки гидролиза тиобензилового эфира Nα-бензилоксикарбонил-L-лизина (BLT), в основном, как описано ранее (Takayama H, Trenn G, Sitkovsky M.V. A novel cytotoxic T lymphocyte activation assay. J. Immunol Methods 1987, 104:183-190). Вкратце, 50 мкл супернатанта переносили в 96-луночный планшет, и к этому супернатанту добавляли 150 мкл раствора BLT (0,2 мМ BLT, EMD Biosciences, Inc., и 0,22 мМ DTNB, Sigma-Aldrich Co.) в забуференном фосфатом физиологическом растворе (PBS, pH 7,2). Оптическую плотность при 410 нм считывали после инкубирования в течение 30 минут при комнатной температуре. Результаты выражали как процент от общего количества клеточного фермента, присутствующего в том же самом количестве клеток, лизированных тритоном X-100.
Статистический анализ
Статистическую значимость различий между различными группами обработки оценивали с помощью одностороннего ANOVA. Величины P<0,05 считали значимыми.
Ингибирование высвобождения MPO человеческими нейтрофилами
Было обнаружено, что через 30 минут, РМА в концентрации 100 нМ (стимулятор высвобождения медиаторов воспаления) повышал уровень высвобождения MPO человеческими нейтрофилами приблизительно в три раза по сравнению с контрольным уровнем, а через 3 часа, уровень высвобождения повышался приблизительно в 5-6 раз. Через 30 минут, измеряли активность MPO, где указанная активность МРО в отсутствие PMA, и в отсутствие PMA + MANS, RNS или тестируемого пептида по отношению к контролю принимали за 100%, а активность MPO в присутствии РМА-эталона составляла приблизительно 275%, активность MPO в присутствии PMA + 50 мкМ MANS составляла приблизительно 275%, и активность MPO в присутствии 100 мкМ MANS составляла приблизительно 305%. Таким образом, пептид MANS не обнаруживал детектируемого эффекта через 30 минут. Однако через 1 час, MANS при более высокой концентрации (100 мкМ) обнаруживал значительный ингибирующий эффект (примерно 260% от контроля) или вызывал примерно 25%-ное снижение уровня высвобождения MPO по сравнению с уровнем в присутствии PMA-эталона (который по измерениям составлял примерно 340% от контроля). Как было измерено, 50 мкМ образец MANS обнаруживал активность МРО примерно 290% от контроля или примерно 15%-ное снижение по отношению к активности в присутствии PMA-эталона. Через 2 часа и непрерывно в течение 3 часов, пептид MANS значительно снижал активность MPO в зависимости от концентрации. Через 2 часа, активность МРО в присутствии контрольного PMA-эталона составляла примерно 540% от контроля; при этом 50 мкМ MANS (по измерениям примерно 375% от контроля) вызывал приблизительно 30%-ное снижение уровня высвобождения MPO по сравнению с уровнем высвобождения в присутствии контрольного PMA-эталона, а 100 мкМ MANS (по измерениям примерно 295% от контроля) вызывал приблизительно 45%-ное снижение высвобождения MPO по сравнению с высвобождением в присутствии контрольного PMA-эталона. Через 3 часа активность МРО в присутствии контрольного PMA-эталона составляла примерно 560% от контроля; при этом 50 мкМ MANS (по измерениям примерно 375% от контроля) вызывал приблизительно 33%-ное снижение высвобождения MPO по сравнению с контролем в присутствии контрольного PMA-эталона, а 100 мкМ MANS (по измерениям примерно 320% от контроля) вызывал приблизительно 40%-ное снижение высвобождения MPO по сравнению с контролем в присутствии контрольного PMA-эталона. Пептид RNS не влиял на PMA-индуцированное высвобождение MPO в любые периоды времени или при любых тестируемых концентрациях. Данные, приведенные в нижеследующей таблице, были получены для концентрации 100 мкМ тестируемых пептидов, которые были инкубированы в течение двух часов со 100 нМ PMA.
Ингибирование высвобождения EPO из клеток HL-60
Активность EPO в супернатанте клона 15 клеток HL-60 значительно увеличивалась через 1 и 2 часа после PMA-стимуляции. Через 1 час, активность EPO по отношению к контролю принималась за 100%; активность в присутствии PMA-эталона составляла примерно 110%; образец, содержащий 10 мкМ MANS, по измерениям, примерно 95%, давал примерно 15%-ное снижение EPO-активности по сравнению с контролем в присутствии PMA-эталона; образец, содержащий 50 мкМ MANS, по измерениям, примерно 78%, давал примерно 30%-ное снижение EPO-активности по сравнению с контролем в присутствии PMA-эталона; и образец, содержащий 100 мкМ MANS, по измерениям, примерно 65%, давал примерно 40%-ное снижение EPO-активности по сравнению с контролем в присутствии PMA-эталона. Через 2 часа, активность EPO по отношению контролю принималась за 100%; активность в присутствии контрольного PMA-эталона составляла примерно 145%; образец, содержащий 10 мкМ MANS, по измерениям, примерно 130%, давал примерно 10%-ное снижение EPO-активности по сравнению с контролем в присутствии PMA-эталона; образец, содержащий 50 мкМ MANS, по измерениям, примерно 70%, давал примерно 50%-ное снижение EPO-активности по сравнению с контролем в присутствии PMA-эталона; и образец, содержащий 100 мкМ MANS, по измерениям, примерно 72%, давал примерно 50%-ное снижение EPO-активности по сравнению с контролем в присутствии PMA-эталона. Таким образом, через 1 и 2 часа, MANS при 50 или 100 мкМ в значительной степени ослаблял высвобождение EPO. Пептид RNS не влиял на PMA-усиленное высвобождение EPO в любое время или при любых тестируемых концентрациях. Данные, представленные в нижеследующей таблице, были получены для концентрации 50 мкМ тестируемых пептидов, которые были инкубированы в течение двух часов со 100 нМ PMA.
Ингибирование высвобождения лизоцима из клеток U937
Секреция лизоцима клетками U937 усиливалась при PMA-стимуляции через 1 час после инкубирования, а через 2 часа после стимуляции, она увеличивалась еще больше. Через 1 час, секреция лизоцима клетками U937 по отношению к контролю принималась за 100%; секреция лизоцима клетками U937 в присутствии контрольного PMA-эталона составляла примерно 210%; образец, содержащий 10 мкМ MANS, по измерениям, примерно 170%, давал примерно 20%-ное снижение лизоцима клетками U937 по сравнению с секрецией в присутствии контрольного PMA-эталона; образец, содержащий 50 мкМ MANS, по измерениям, примерно 170%, давал примерно 20%-ное снижение лизоцима клетками U937 по сравнению с секрецией в присутствии контрольного PMA-эталона; и образец, содержащий 100 мкМ MANS по измерениям, 115%, давал примерно 45%-ное снижение лизоцима клетками U937 по сравнению с секрецией в присутствии контрольного PMA-эталона. Через 2 часа, секреция лизоцима клетками U937 по отношению к контролю принималась за 100%; секреция лизоцима клетками U937 в присутствии контрольного PMA-эталона составляла примерно 240%; образец, содержащий 10 мкМ MANS, по измерениям, примерно 195%, давал примерно 20%-ное снижение лизоцима клетками U937 по сравнению с секрецией в присутствии контрольного PMA-эталона; образец, содержащий 50 мкМ MANS, по измерениям, примерно 185%, давал примерно 25%-ное снижение лизоцима клетками U937 по сравнению с секрецией в присутствии контрольного PMA-эталона; и образец, содержащий 100 мкМ MANS по измерениям, примерно 140%, давал примерно 40%-ное снижение лизоцима клетками U937 по сравнению с секрецией в присутствии контрольного PMA-эталона. Таким образом, секреция лизоцима значительно ослаблялась через 1 и 2 часа после стимуляции под действием 100 мкМ MANS, но не под действием 50 или 10 мкМ MANS. RNS пептид не влиял на PMA-усиленную секрецию лизоцима в любое время или при любых тестируемых концентрациях. Данные, представленные в нижеследующей таблице, были получены для концентрации 50 мкМ тестируемых пептидов, которые были инкубированы в течение двух часов со 100 нМ PMA.
Ингибирование высвобождения гранзима из клеток NK-92
Для оценки высвобождения гранзима использовали лимфоцитарную клеточную линию природных клеток-киллеров NK-92 (Gong J.H, Maki G, Klingemann H.G. Characterization of a human cell line (NK-92) with phenotypical и functional characteristics of activated natural killer cells. Leukemia 8:652-658, 1994, Maki G, Klingemann H.G, Martinson J.A, Tarn Y.K. Factors regulating the cytotoxic activity of the human natural killer cell line, NK-92. J. Hematother. Stem Cell Res., 10:369-383, 2001, Takayama H, Trenn G, Sitkovsky M.V. A novel cytotoxic T lymphocyte activation assay. J. Immunol. Methods 104:183-190, 1987).
Измерение уровня секреции гранзима NK-клетками: уровень гранзима, секретируемого клетками NK-92, измеряли путем оценки гидролиза тиобензилового эфира Nα-бензилоксикарбонил-L-лизина (BLT, EMD Bioscience, Inc.) в основном, как описано ранее (Takayama H, Trenn G, Sitkovsky M.V. A novel cytotoxic T lymphocyte activation assay. J. Immunol. Methods 104:183-190, 1987). Аликвоту 50 мкл супернатанта переносили в 96-луночный планшет, и к этому супернатанту добавляли 150 мкл 0,2 мМ раствора BLT и 0,22 мМ DTNB (Sigma-Aldrich Co.) в забуференном фосфатом физиологическом растворе (PBS, pH 7,2). Оптическую плотность при 410 нм измеряли после инкубирования в течение 30 минут при комнатной температуре. Результаты выражали как процент от общего количества клеточного фермента, присутствующего в том же самом количестве клеток, лизированных тритоном X-100.
Поскольку для количественной оценки уровня высвобождения гранзима клетками NK-92 не использовали стандарт, то авторами настоящего изобретения были измерены уровни высвобождения гранзима и его внутриклеточные уровни (лизированных клеток), и эти уровни секретированного гранзима выражали как процент от общего уровня каждого фермента (уровня внутриклеточного и секретированного фермента). Для измерения уровня внутриклеточного гранзима, секретированного клетками NK-92, брали соответствующие аликвоты клеток, лизированных 0,1% тритоном X-100, в целях проведения количественной оценки фермента, описанного выше. Для минимизации вариабельности между культурами, все данные выражали как процент от контроля. Данные, представленные в нижеследующей таблице, были получены для концентрации 50 мкМ тестируемых пептидов, которые инкубировали в течение двух часов со 100 нМ PMA.
Цитотоксичность
Поскольку ни в одном из экспериментов генерирования токсического ответа в клетках не использовался стандарт, то проводили оценку удерживания/высвобождения LDH (данные не приводятся) (см. также Park J-A, He F, Martin LD, Li Y, Adler KB. Human neutrophil elastase induces hypersecretion of mucin from human bronchial epithelial cells in vitro via a PKC δ - mediated mechanism. Am. J. Pathol. 2005, 167:651-661).
В предварительных экспериментах: пептиды, представленные в нижеследующей таблице, продемонстрировали соответствующий процент ингибирования высвобождения MPO из человеческих нейтрофилов, EPO из клона 15 клеток HL-60, лизоцима из клеток U937 и гранзима из клеток NK-92, где MA означает присутствие миристоильной группы-заместителя в альфа-N-концевом положении пептида; Ac означает присутствие ацетильной группы-заместителя а альфа-N-концевом положении пептида; Н означает отсутствие группы, присоединенной к пептиду, а NH2 означает присутствие амида в С-концевом положении. Данные ингибирования для нескольких экспериментов усредняли. Качественная растворимость пептидов в 0,5н физиологическом растворе, pH 6,5, выраженная в мг/мл, представлена ниже в таблице 3. Замена миристоильной группы другой N-концевой химической группой может приводить к изменению растворимости описанных здесь пептидов в водной среде. Так, например, замена миристоильной группы на ацетильную группу приводит к повышению уровня растворимости этих пептидов в воде, как показано в таблице 3.
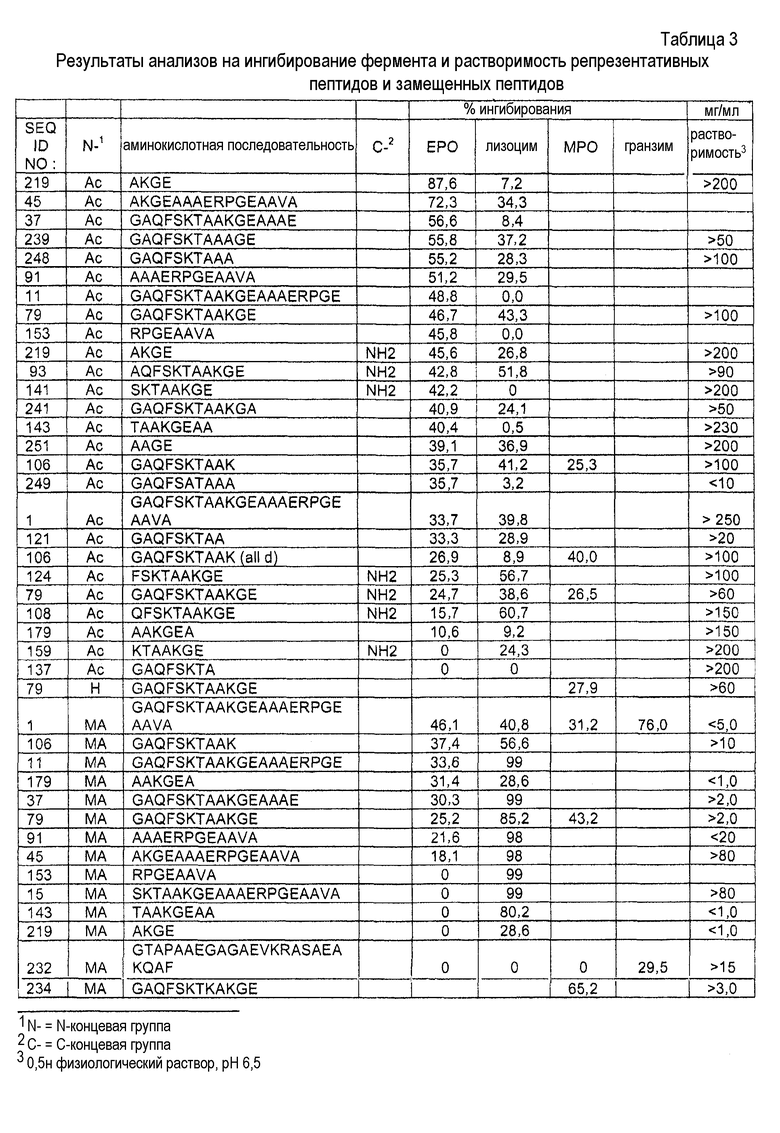
Пример 2
In vivo подавление индуцированного липополисахаридом (ЛПС) воспаления легких под действием пептида MANS и родственных пептидов
Эксперимент, проиллюстрированный в этом примере, в основном, осуществляли методами, описанными в публикациях Cox, G, Crossley, J., and Xing, Z., Macrophage engulfment of apoptotic neutrophils contributes to the resolution of acute pulmonary inflammation in vivo, Am. J. Respir. Cell Mol. Biol. 12:232-237, 1995, Hirano S., Quantitative time-course profiles of bronchoalveolar lavage cells following intratracheal instillation of lipopolysaccharide in mice, Ind. Health 35:353-358, 1997; и Ulich T.R, Watson L.R, Yin SM, Guo K.Z, Wang P, Thang H, and del Castillo, J. Am. J. Pathol. 138:1485-1496, 1991.
Таким образом, 6-7-недельных самок мышей CDl весом 15-20 граммов получали из лаборатории Charles River и содержали в клетках по 5 мышей на каждую клетку. Животным давали стандартный корм для грызунов и отфильтрованную воду ad libitum. Этих животных содержали в условиях, рекомендованных Национальным институтом здравоохранения (NIH), при стандартной температуре (64°-79°F) и относительной влажности 30-70%.
Пять экспериментальных групп мышей, по 5 животных в каждой группе, обрабатывали PBS с последующей обработкой PBS; PBS с последующей обработкой ЛПС; (миристоилированным) пептидом MANS с последующей обработкой ЛПС; ацетилированным пептидом SEQ ID NO:1 с последующей обработкой ЛПС, или ацетилированным пептидом SEQ ID NO:106 с последующей обработкой ЛПС.
Предварительная обработка путем интраназальной инстилляции пептида: пептид согласно изобретению, оцениваемый in vivo на его способность ингибировать или ослаблять ЛПС-индуцированное воспаление легких, растворяли в PBS при концентрации 1 мМ. Животных, анестезированных 0,8% изофлуораном путем ингаляции, предварительно обрабатывали ударной дозой 2×10 мкл пептидного раствора, интраназально вводимого в одну ноздрю за 30 минут до последующей инстилляции ЛПС.
Интраназальная инстилляция ЛПС: эндотоксин липополисахарид (ЛПС) (эндотоксин, происходящий от Escherichia coli серотипа 011:B4, Sigma, St Louis, MO, см. стр.L4130, где приводится информация о продукте Sigma под заголовком «Липополисахариды Escherichia coli 011:B4» (Lipopolysaccharides from Escherichia coli 011:B4)) растворяли в забуференном фосфатом физиологическом растворе (PBS) при 2500 мкг/мл. Для обработки животных эндотоксином, указанным животным, которые были анестезированы 0,8% изофлуораном путем ингаляции, интраназально вводили ударную дозу 10 мкл 2500 мкг/мл раствора эндотоксина. Ударную дозу в 10 мкл вводили в одну ноздрю. После инстилляции эндотоксина, животных наблюдали на наличие затрудненного дыхания, летаргию и снижение потребления воды/корма.
Бронхоальвеолярный лаваж (БАЛ): через шесть часов после последней инстилляции, животных анестезировали (90 мг/кг нембутала) и умерщвляли путем обескровливания. Легкие 2 раза последовательно промывали 1,0 мл-аликвотами PBS. Собранную БАЛ-жидкость центрифугировали в целях удаления клеток для их последующего подсчета и определения лейкоцитарной формулы. Собранную лаважную жидкость использовали для анализа на общий белок, миелопероксидазу (МРО), LDH и гемоглобин.
Анализы: Аликвоты БАЛ-жидкости сразу использовали для анализа на уровни LDH, общего белка или гемоглобина, проводимого на анализаторе COBAS Fara (автоматическом анализаторе COBAS FARA II, Roche Diagnostic Systems Inc., Montclair, NJ). Аликвоту БАЛ-жидкости замораживали при -80°С для последующей количественной оценки миелопероксидазы (MPO), проводимой с помощью мышь-специфического ELISA-анализа (Cell Sciences, Inc., Canton, Mass). БАЛ-данные анализировали стандартными методами для оценки различий между контролем и группами обработки. Результаты, иллюстрирующие ингибирование или ослабление воспаления под действием тестируемого пептида, представлены в нижеследующих таблицах.
Средние величины маркеров воспаления в присутствии пептида MANS, MA-GAQFSKTAAKGEAAAERPGEAAVA, SEQ ID NO:1
Средние величины маркеров воспаления в присутствии N-концевого ацетилированного аналога пептида MANS, Ac-GAQFSKT AAKGEAAAERPGEAAVA, SEQ ID NO:1
Средние величины маркеров воспаления в присутствии ацетилированного пептида Ac-GAQFSKTAAK, SEQ ID NO:106
Данные ингибирования маркеров воспаления под действием пептида MANS (Myr-SEQ ID NO:1), тестируемых пептидов (Ac-GAQFSKTAAKGEAAAERPGEAAVA), SEQ ID NO:1 и Ac-GAQFSKTAAK, SEQ ID NO: 106, по сравнению с данными, полученными при PBS/ЛПС-обработке
PBS/PBS означает введение только PBS в качестве контроля без добавления эндотоксина ЛПС для стимуляции миграции хемотаксических нейтрофилов; PBS/ЛПС означает добавление ЛПС (эндотоксина) для стимуляции миграции хемотаксических нейтрофилов; MANS/ЛПС означает предварительную обработку пептидом MANS в PBS с последующей ЛПС-стимуляцией для индуцирования миграции нейтрофилов. Процент нейтрофилов от общего числа клеток в группах ЛПС-обработки снижался от 41,7% до 30,9% при обработке пептидом MANS; от 65,5% до 41,47% при обработке пептидом Ac-GAQFSKTAAKGEAAAERPGEAAVA, SEQ ID NO:1, и от 24,4% до 3,0% при обработке пептидом Ac-GAQFSKTAAK, SEQ ID NO:106. Измеренные уровни MPO в группах ЛПС-обработки снижались от 28,98 нг/мл до 9,49 нг/мл при обработке пептидом MANS, от 37,9 нг/мл до 30,79 нг/мл при обработке пептидом с ацетилированной SEQ ID NO:1 и от 7,19 нг/мл до 1,50 нг/мл при обработке пептидом с ацетилированной SEQ ID NO:106.
Пример 3
Мышь с моделью ХОБЛ, индуцированной озоном
Окислительный стресс, вызванный химическими раздражителями, такими как озон, является широко известным признаком хронической обструктивной болезни легких (ХОБЛ). См. публикации Repine JE, Bast A, Lankhorst I, и the Oxidative Stress Study Group, Am. J. Respir. Crit. Care Med. 156:341-357, 1997, а также Harkema J.R и Hotchkiss J.A, Toxicology Letters, 68:251-263, 1993.
10-недельных самок мышей Balb/C получали из лаборатории Charles River и содержали в клетках по 5 мышей на каждую клетку в условиях, рекомендованных Национальным институтом здравоохранения (NIH). Животным давали стандартный корм для грызунов и отфильтрованную воду ad libitum. Мышей в трех группах обработки, содержащих по 5 животных в каждой, анестезировали путем внутрибрюшинной инъекции кетамина (100 мг/кг) и ксилазина (20 мг/кг), а затем предварительно обрабатывали путем внутритрахеального введения 25 мкл одного PBS, или раствора 1,0 мМ пептида MANS в PBS, или раствора 1,0 мМ фрагмента ацетилированного пептида MANS Ac-GAQFSKTAAK, называемого здесь ацетилированной SEQ ID NO:106, в PBS. Через 30 минут, животных помещали в соответствующую специально оборудованную камеру для подачи озона и нагнетаемого воздуха. Животных обрабатывали озоном в течение 2 часов (при концентрациях озона 1-10 м.д.) слегка модифицированным методом, описанным Haddad et al., 1995 (Haddad E-B, Salmon M, Sun J, Liu S, Das A, Adcock I, Barnes P.J., и Chung KF, FEBS Letters, 363:285-288, 1995). Озон генерировали с использованием генератора озона модели OL80F/B, OzoneLab, Burton, British Columbia, Canada. Последующий непрерывный мониторинг концентрации озона проводили с использованием анализатора Teledyne Photometric 03 (модель 400E, Teledyne Instruments, City of Industry, CA). Мышей двух дополнительных групп, которых ранее не подвергали какой-либо обработке, обрабатывали озоном в тех же самых условиях, или нагнетаемым воздухом в условиях, аналогичных условиям обработки озоном, но в отсутствие озона. После обработки, животных умерщвляли путем обескровливания, и легкие 2 раза последовательно промывали 1,0 мл-аликвотами PBS. Собранный бронхоальвеолярный лаваж (БАЛ) центрифугировали в целях удаления клеток для их последующего подсчета и определения лейкоцитарной формулы. Собранную лаважную жидкость использовали для анализа на белок и для дополнительного анализа на IL-6, IFNγ и КС (мышиный аналог IL-8), проводимого с помощью ELISA (с использованием аналитических наборов, полученных от R&D Systems, Minneapolis, MN).
Процент ингибирования миграции нейтрофилов в БАЛ-жидкость для групп обработки по отношению к данным, полученным для контрольной группы, обработанной только PBS, представлен в нижеследующей таблице.
Ингибирование индуцированной озоном миграции нейтрофилов пептидом MANS и пептидом с ацетилированной SEQ ID NO:106, Ac-GAQFSKTAAK
Концентрации IL-6 в пг/мл в БАЛ-жидкости в зависимости от предварительной внутритрахеальной инъекции и последующей обработки озоном получали следующим образом. Были получены следующие уровни IL-6: приблизительно 364,5 пг/мл в группе мышей, предварительно обработанных пептидом MANS, а затем озоном; приблизительно 130,4 пг/мл в группе мышей, предварительно обработанных ацетилированным фрагментом пептида MANS, Ac-GAQFSKTAAK (SEQ ID NO:106), а затем озоном; приблизительно 1041,3 пг/мл в группе мышей, предварительно обработанных PBS и озоном, приблизительно 43,2 пг/мл в группе мышей, непосредственно обработанных нагнетаемым воздухом без какой-либо предварительной обработки.
Концентрации KC в пг/мл в БАЛ-жидкости в зависимости от предварительной внутритрахеальной инъекции и последующей обработки озоном получали следующим образом. Были получены следующие уровни КС: приблизительно 183,6 пг/мл в группе мышей, предварительно обработанных пептидом MANS, а затем озоном; приблизительно 159,7 пг/мл в группе мышей, предварительно обработанных ацетилированным фрагментом пептида MANS, Ac-GAQFSKTAAK (SEQ ID NO:106), а затем озоном; приблизительно 466,6 пг/мл в группе мышей, предварительно обработанных PBS и озоном, приблизительно 36,3 пг/мл в группе мышей, непосредственно обработанных нагнетаемым воздухом без какой-либо предварительной обработки.
Концентрации IFNγ в пг/мл в БАЛ-жидкости в зависимости от предварительной внутритрахеальной инъекции и последующей обработки озоном получали следующим образом. Были получены следующие уровни IFNγ: приблизительно 7,4 пг/мл в группе мышей, предварительно обработанных пептидом MANS, а затем озоном; приблизительно 3,6 пг/мл в группе мышей, предварительно обработанных ацетилированным фрагментом пептида MANS, Ac-GAQFSKTAAK (SEQ ID NO:106), а затем озоном; приблизительно 8,6 пг/мл в группе мышей, предварительно обработанных PBS и озоном, приблизительно 5,0 пг/мл в группе мышей, непосредственно обработанных нагнетаемым воздухом.
Введение озона мышам приводило к значительному увеличению числа инфильтрированных нейтрофилов, а также уровней IL-6 и KC в БАЛ. По сравнению с контрольной группой мышей, которых предварительно обрабатывали PBS, каждая мышь в группах, предварительно обработанных пептидом MANS, ацетилированным пептидом Ac-GAQFSKTAAK, то есть ацетилированной SEQ ID NO:106, обнаруживала снижение уровня инфильтрации нейтрофилов в БАЛ-жидкость после обработки озоном (например, 93% ± 10% и 81% ± 10%, соответственно, по сравнению с PBS-контролем). Параллельно, пептид MANS и ацетилированный пептид, а именно ацетилированная SEQ ID NO:106 после обработки озоном также заметно снижают концентрации KC (например, 65,8% ± 10% и 71,3% ± 10%, соответственно, по сравнению с PBS-контролем), и уровни IL-6 (например, 67,8% ± 15%, MANS и 91,3% ± 15%, ацетилированная SEQ ID NO:106, по сравнению с PBS-контролем), но оказывают незначительное влияние на уровни интерферона-γ. В целом, эти данные показали, что пептид MANS и ацетилированный пептид SEQ ID NO:106 значительно снижают или ингибируют индуцированную озоном миграцию нейтрофилов в дыхательные пути, а также селективно снижают уровни хемокинов и цитокинов. Уровни IL-6 в БАЛ-жидкостях, взятых у животных, предварительно обработанных пептидом MANS или ацетилированным пептидом SEQ ID NO:106, ингибировались приблизительно на 68% и 91%, соответственно, по сравнению с уровнями у животных, предварительно обработанных PBS. Кроме того, уровни KC в БАЛ-жидкостях, взятых у животных, предварительно обработанных пептидом MANS или ацетилированным пептидом SEQ ID NO:106, ингибировались приблизительно на 65% и 71%, соответственно, по сравнению с уровнями у животных, предварительно обработанных PBS.
Пример 4
Модель хронического бронхита
Данную процедуру, описанную в публикациях Voynow J.A, Fischer B.M, Malarkey D.E, Burch LH, Wong T, Longphre M, Ho S.B, Foster W.M, Neutrophil Elastase induces mucus cell metaplasia in mouse lung, Am. J. Physiol. Lung Cell Mol. Physiol. 287:L1293-L1302, 2004, проводили для создания модели хронического бронхита у мышей. В частности, гиперплазию бокаловидных клеток, которая является отличительным признаком хронического бронхита, индуцировали путем перманентной обработки мышей человеческой эластазой нейтрофилов (NE), вводимой в дыхательные пути посредством инстилляции.
Человеческую NE интратрахеально отсасывали у самцов мышей Balb/c. Всего было закуплено 30 мышей (весом примерно 25-30 г) у поставщика, такого как Jackson Laboratories, Bar Harbor, ME. Мышей содержали в 12-часового цикле смены дня и ночи со свободным доступом к корму и воде. Животным вводили NE путем ротоглоточной аспирации на дни 1, 4 и 7. Сразу после анестезии путем ингаляции изофлуораном (IsoFlo от Abbott Laboratories) и системы анестезии газом по открытому контуру от Stoelting, животных подвешивали за их верхние резцы (передние зубы) на наклонной плоскости под углом 60°, и объем жидкости, содержащей человеческую NE [50 мкг (43,75 единиц)/40 мкл PBS (Elastin Products, Owensville, MO)], вводили путем ее нанесения на вытянутый язык животного, простирающийся до дистального отдела ротоглотки. При вытянутом языке животное не может проглотить жидкость, а поэтому объем жидкости попадает в дыхательные пути.
Через 7 дней после последней NE-обработки, в том случае, если гиперплазия бокаловидных клеток, смоделированная в дыхательных путях при хроническом бронхите, достигала своего максимума (см. Voynow et al, 2004), мышам (5 животных на группу) путем интратрахеальной инстилляции вводили 50 мкл либо PBS (в качестве контроля), либо 100 мкМ раствора пептида MANS, раствора пептида RNS или раствора пептида, такого как ацетилированный пептид SEQ ID NO:106, растворенный в PBS. Через 15 минут, секрецию слизи стимулировали путем введения метахолина с использованием системы аэрозольных ингаляторов Buxco, в результате чего получали тонкодисперсный аэрозоль метахолина, поступающего в концентрации приблизительно 60 мМ в течение 3 минут. Через 15 минут после введения метахолина, мышей умерщвляли путем ингаляционной обработки 100% газом CO2.
Гистохимический анализ
После обработки, описанной выше, легкие животных продували для удаления крови, а затем вдували среду OCT (среду с оптимально низкой температурой) (Sakura Finetck, Torrance, CA), наполовину разведенную в физиологическом растворе. Легкие погружали на ночь в 10% формальдегид в PBS при 4°С и заливали в парафин. 5 мкм-срезы обрабатывали периодной кислотой Шиффа/гематоксилином для окрашивания на муцины дыхательных путей, например, как описано в публикациях Singer M, Vargaftig B.B., Martin L.D., Park J.J., Gruber A.D., Li Y, Adler K.B., A MARCKS-related peptide blocks mucus hypersecretion in a murine model of asthma., Nature Medicine 10:193-196, 2004.
Гистологический индекс слизи
Гистологический индекс слизи (Whittaker L, Niu N, Temann U-A, Sloddard A. Flavell R.A, Ray A, Homer R.J, and Cohn L, Interleukin-13 mediates a fundamental pathway for airway epithelial mucus induced by CD4 T cells and interleukin-9, Am. J. Respir. Cell Mol. Biol. 27:593-602, 2002) определяли на AB/PAS-окрашенных срезах, которые охватывают центральные и периферические дыхательные пути. Предметные стекла наблюдали через 10X объектив и изображение снимали на цифровую видиокамеру. Для каждого животного получали минимум четыре репрезентативных снимка поперечных или сагиттальных срезов дыхательных путей. Визуализировать можно было только те дыхательные пути, которые имели полную окружность, и изображение этих срезов анализировали. Дыхательные пути, которые были открыты непосредственно в альвеолярное пространство, не анализировали. Степень PAS-позитивного окрашивания в каждом изображении дыхательных путей полуколичественно оценивалась исследователем, который не знал об условиях обработки каждого среза, в соответствии с нижеследующей 5-балльной системой оценки: балл 0, PAS-окрашивание отсутствует; балл 1, PAS-окрашивание наблюдалось на 25% или менее эпителия дыхательных путей; балл 2, PAS-окрашивание наблюдалось на 26-50% эпителия дыхательных путей; балл 3, PAS-окрашивание наблюдалось на 51-75% эпителия дыхательных путей; балл 4, PAS-окрашивание наблюдалось на >75% эпителия дыхательных путей. Такая система оценок используется для вычисления индекса слизи в каждой группе, и результаты такой оценки представлены как среднее ± ср.кв.ош.
Все результаты представлены как среднее ± ср.кв.ош (n=5 животных, 10-20 срезов для каждого животного). Уровни значимости вычисляли с помощью одностороннего дисперсионного анализа ANOVA, а затем с помощью критерия Шеффе с использованием компьютерной программы SPSS 6.1 (* = значимость данных по сравнению с пороговой величиной p<0,05).
Пример 5
In vivo анализы
Целью проведения нижеследующей серии экспериментов является определение влияния пептидов согласно изобретению после их доставки in vivo либо путем локальной инстилляции на участок воспаления, либо внутривенной инъекции на развитие воспаления по сравнению с контрольными пептидами, такими как RNS. Для этой цели использовали две модели: (i) мышиную модель с воспалением воздушного кармана и (ii) мышиную модель перитонита, индуцированного тиогликолятом. Обе эти модели представляют собой хорошо охарактеризованные модели воспаления, в развитии которого важную роль играют нейтрофилы. Модель воздушного кармана позволяет определить влияние пептидов на быстрое течение воспаления (приблизительно 4 часа), а модель перитонита может быть использована для оценки влияния пептидов на более длительное течение воспаления (приблизительно 24 часа).
Общий протокол эксперимента
В целях определения действия пептидов, описанных в настоящей заявке, для каждой из двух моделей, а именно модели i.v. доставки пептидов и модели локальной доставки пептидов, проводили четыре исследования. В каждом исследовании участвовали 2 экспериментальные группы, а именно контрольная группа без воспаления (обработанная носителем) и группа с индуцированным воспалением (то есть обработанная стимулятором воспаления). Каждую группу разделяли на 5 и необязательно на 6 подгрупп обработки, n=6 для каждой подгруппы. Эти подгруппы были обработаны, например носителем, MANS, RNS, тестируемым пептидом, необязательно пептидом, имеющим «перетасованную» последовательность тестируемого пептида, где указанная «перетасованная» последовательность является двухкомпонентной последовательностью «пептид-SCR», и дексаметазоном. Дексаметазон служит в качестве эталонного противовоспалительного средства. Выбор соответствующих доз для i.v. инъекции или локальной инстилляции осуществляли исходя из предварительных экспериментов по определению зависимости «доза-ответ». Экспериментальные дозы, определенные исходя из ингибирующей активности MANS в человеческих нейтрофилах, составляют 1 мг/кг для i.v. одноразовой доставки, или конечная концентрация 50 мкМ, вводимая локально (в воздушный карман или i.p.). Доза для i.v. доставки была выбрана с учетом объема распределения 2 л/кг.
Модель воспаления воздушного кармана:
Анализы на инфильтрацию нейтрофилов и воспаление в мышином воздушном кармане осуществляли, как описано в публикации Clish C.B., O'Brien J.A., Gronert K., Stahl G.L., Petasis N.A., Serhan C.N. Local and systemic delivery of a stable aspirin-triggered lipoxin prevents neutrophil recruitment in vivo. Proc. Natl. Acad. Sci USA. 1999 Jul 6; 96(14):8247-52. Таким образом, самцов белых мышей BALB/c (6-8-недельных) анестезировали изофлуораном, и объем дорсальных воздушных карманов увеличивали путем подкожной инъекции 3 мл стерильного воздуха на дни 0 и 3. На 6-й день мышей анестезировали изофлураном, а затем вводили носитель, MANS, RNS, тестируемый пептид или, но необязательно, пептид-SCR, либо путем i.v. инъекции ударной дозы в хвостовую вену в 100 мкл стерильного 0,9% физиологического раствора, либо местно в воздушный карман в 900 мкл PBS-/- (забуференного фосфатом физиологического раствора Дульбекко, не содержащего ионов магния или кальция, BioWhittaker). Дексаметазон (Sigma), вводимый либо i.v. в дозе 0,1 мг/кг в 100 мкл стерильного 0,9% физиологического раствора, либо местно в дозе 10 мкг в 900 мкл of PBS-/-, служит в качестве эталонного противовоспалительного средства. Воспаление в воздушном кармане индуцировали путем местной инъекции рекомбинантного мышиного фактора некроза опухоли α (TNF-α, 20 нг) (Boehringer Mannheim), растворенного в 100 мкл стерильного PBS. Через 4 часа после первой инъекции TNF-α, воздушные карманы мышей, анестезированных изофлуораном, два раза промывали 3 мл стерильного PBS. Аспираты центрифугировали при 2000 об/мин в течение 15 минут при 23°С. Супернатанты удаляли, и клетки суспендировали в 500 мкл PBS. Аликвоты супернатанта анализировали на концентрации медиаторов воспаления (необязательно, за исключением TNFα), активность MPO и перокисление липидов.
Общее число лейкоцитов в клеточной суспензии подсчитывали под оптическим микроскопом с использованием гемоцитометра. Ресуспендированные клетки аспирата (50 мкл) на предметных стеклах микроскопа добавляли к 150 мкл 30% BSA и центрифугировали при 2200 об/мин в течение 4 минут на цитоцентрифуге. Лейкоцитарную формулу клеток, центрифугированных в цитоцентрифуге и окрашенных по Райту-Гимзе, использовали для подсчета абсолютного числа лейкоцитов каждого типа на эксудат воздушного кармана. Для проведения анализа под микроскопом ткани толщиной в 6 мм, полученные с помощью пробойника для биопсии ткани (Acu-Punch, Acuderm), фиксировали в 10% забуференном формальдегиде. Затем образцы заливали в парафин, приготавливали срезы, и эти срезы окрашивали гематоксилином-эозином. Число нейтрофилов в гистологических срезах определяли путем подсчета числа клеток/hpf (в поле зрения при большом увеличении микроскопа). В качестве контроля для дермы воспаленного воздушного кармана служила дистальная область дермы.
Данные были представлены как общее число нейтрофилов, моноцитов, эозинофилов, базофилов и лимфоцитов на эксудат или число нейтрофилов на ткань в поле зрения при большом увеличении микроскопа. Полученные величины выражали как среднее ± ср.кв.ош. (n=6). Значимость влияния какой-либо обработки на миграцию определяли с помощью ANOVA. Величина P<0,05 рассматривалась как значимая.
Пример 6
Модель воспаленной брюшины
Самцов мышей BALB/c (6-8 недель) использовали для создания модели индуцированного тиогликолятом перитонита, как описано в публикации Tedder T.F, Steeber D.A, Pizcueta P. L-selectin-deficient mice have impaired leukocyte recruitment into inflammatory sites. J. Exp. Med. 1995 Jun 1, 181(6):2259-64. Носитель, MANS, RNS, тестируемый пептид и, необязательно, пептид-SCR вводили путем инъекции ударной дозы либо в хвостовую вену в 100 мкл стерильного 0,9% физиологического раствора, либо местно в брюшину в 900 мкл PBS-/- непосредственно перед i.p.-инъекцией тиогликолята. Дексаметазон, вводимый либо i.v. в дозе 0,1 мг/кг в 100 мкл стерильного 0,9% физиологического раствора, либо местно в дозе 10 мкг в 900 мкл PBS-/-, служит в качестве эталонного противовоспалительного средства. Воспаление индуцировали путем внутрибрюшинной инъекции мышам 1 мл раствора тиогликолята (3% мас./об, Sigma Immunochemicals). Через 24 часа после индуцирования воспаления, мышей подвергали гуманной эвтаназии и в брюшину инъецировали 5 мл теплой среды (среды при ~37°С (RPMI 1640, 2% FCS, и 2 мМ EDTA)), а затем делали легкий массаж брюшной полости. Аспираты лаважной жидкости брюшной полости центрифугировали при 2000 об/мин в течение 15 минут при 23°С. Супернатанты удаляли, и клетки суспендировали в 500 мкл PBS. Аликвоты супернатанта анализировали на активность MPO, концентрации медиаторов воспаления и перокисление липидов.
Общее число лейкоцитов в клеточной суспензии подсчитывали под оптическим микроскопом с использованием гемоцитометра. Ресуспендированные клетки аспирата (50 мкл) на предметных стеклах микроскопа добавляли к 150 мкл 30% BSA и центрифугировали при 2200 об/мин в течение 4 минут на цитоцентрифуге. Лейкоцитарную формулу клеток, центрифугированных в цитоцентрифуге и окрашенных по Райту-Гимзе, использовали для подсчета абсолютного числа лейкоцитов каждого типа на аспират воздушного кармана.
Данные были представлены как общее число нейтрофилов, моноцитов, эозинофилов, базофилов и лимфоцитов на эксудат. Полученные величины выражали как среднее ± ср.кв.ош. (n=6). Значимость влияния какой-либо обработки на миграцию определяли с помощью ANOVA. Величина P<0,05 рассматривалась как значимая.
Дегрануляция
В качестве маркера дегрануляции использовали миелопероксидазу. Миелопероксидазную активность в клеточном супернатанте, полученном из лаважной жидкости воздушного кармана или брюшины, оценивали и анализировали, как описано выше с применением TMB-метода.
Концентрации медиаторов воспаления
Концентрации ключевых провоспалительных медиаторов TNF-α, IL-1β, IL-10, IL-6, KC и PGE2 в лаважной жидкости воздушного кармана и брюшины определяли с использованием коммерчески доступных наборов для ELISA (R&D Systems) в соответствии с инструкциями производителей.
Перокисление липидов
Концентрация F2-изопростанов является чувствительным и специфическим показателем окислительного повреждения, вызываемого высвобождением промежуточных продуктов реакционноспособного кислорода из нейтрофилов и других клеток (Milne G.L, Musiek E.S, Morrow J.D. F2-isoprostanes as markers of oxidative stress in vivo: an overview. Biomarkers. 2005 Nov; 10 Suppl 1:S10-23). Концентрацию F2-изопростанов определяли в супернатантах эксудата воздушного кармана и брюшины с помощью коммерчески доступного ELISA-анализа (8-Isoprostane EIA, Cayman Chemical) в соответствии с инструкциями производителей.
Результаты
Эксперимент считался успешным, если местная или системная доставка тестируемых пептидов, проводимая одним или несколькими описанными выше способами ингибирования высвобождения медиаторов воспаления, приводила к ослаблению воспаления.
Активные фрагменты пептидов согласно изобретению ингибируют приток нейтрофилов в воспаленный воздушный карман или в воспаленную брюшину и ингибируют дегрануляцию нейтрофилов в этих органах, что приводит к снижению активности МРО, уровня перокисления липидов и продуцирования медиаторов воспаления.
Вышеописанные примеры приводятся лишь в иллюстративных целях и не должны рассматриваться как ограничение объема изобретения. Объем настоящего изобретения определен нижеследующей формулой изобретения, в которую включены описанные здесь эквиваленты.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ РЕГУЛЯЦИИ МЕДИАТОРОВ ВОСПАЛЕНИЯ И ПЕПТИДЫ, ИСПОЛЬЗУЕМЫЕ В НИХ | 2007 |
|
RU2429004C2 |
| ИНГИБИТОРЫ ГИПЕРСЕКРЕЦИИ МУЦИНА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2006 |
|
RU2423379C2 |
| ИНГИБИТОРЫ МЕТАСТАЗИРОВАНИЯ | 2014 |
|
RU2681213C2 |
| ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ ДЛЯ ИНГИБИРОВАНИЯ ВОСПАЛИТЕЛЬНЫХ ЦИТОКИНОВ | 2019 |
|
RU2816868C2 |
| АНТИБАКТЕРИАЛЬНЫЕ ПЕПТИДЫ | 2007 |
|
RU2468033C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ПРОФИЛАКТИКИ ИЛИ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ, СОСТОЯНИЙ ИЛИ ПРОЦЕССОВ, ХАРАКТЕРИЗУЕМЫХ АБЕРРАНТНОЙ ПРОЛИФЕРАЦИЕЙ ФИБРОБЛАСТОВ И ОТЛОЖЕНИЕМ ВНЕКЛЕТОЧНОГО МАТРИКСА | 2012 |
|
RU2620066C2 |
| Применение пептида для лечения сердечно-сосудистых заболеваний | 2013 |
|
RU2672341C2 |
| ЛЕЧЕНИЕ ВОСПАЛИТЕЛЬНЫХ, АУТОИММУННЫХ И НЕЙРОДЕГЕНЕРАТИВНЫХ НАРУШЕНИЙ ИММУННОДЕПРЕССИВНЫМИ ПОЛИПЕТИДАМИ, ЯВЛЯЮЩИМИСЯ ПРОИЗВОДНЫМИ ТАТ | 2013 |
|
RU2653754C2 |
| ПЕПТИДЫ, ВЫДЕЛЕННЫЕ ИЗ ЧЕЛОВЕЧЕСКОГО ЛАКТОФЕРРИНА, И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2593757C2 |
| АНТИТЕЛА ПРОТИВ G-CSFR И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2605595C2 |










































































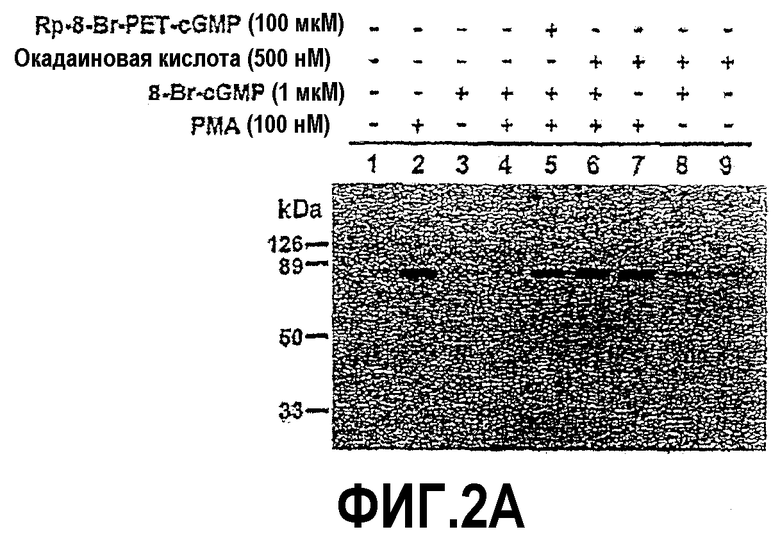
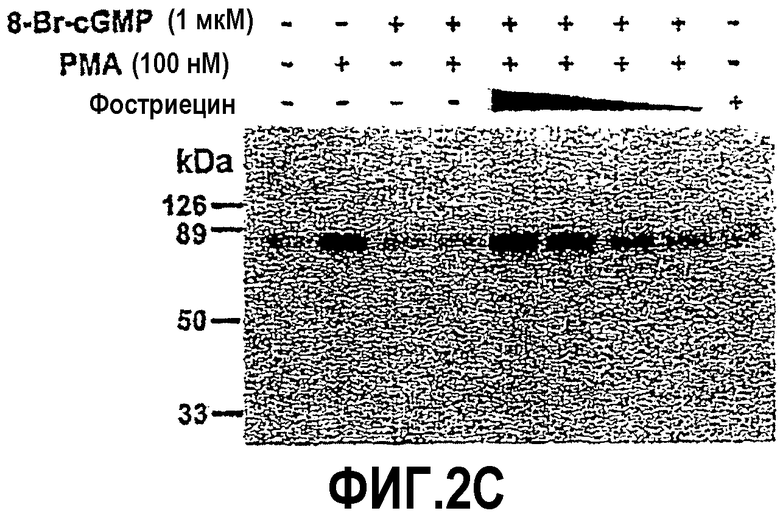
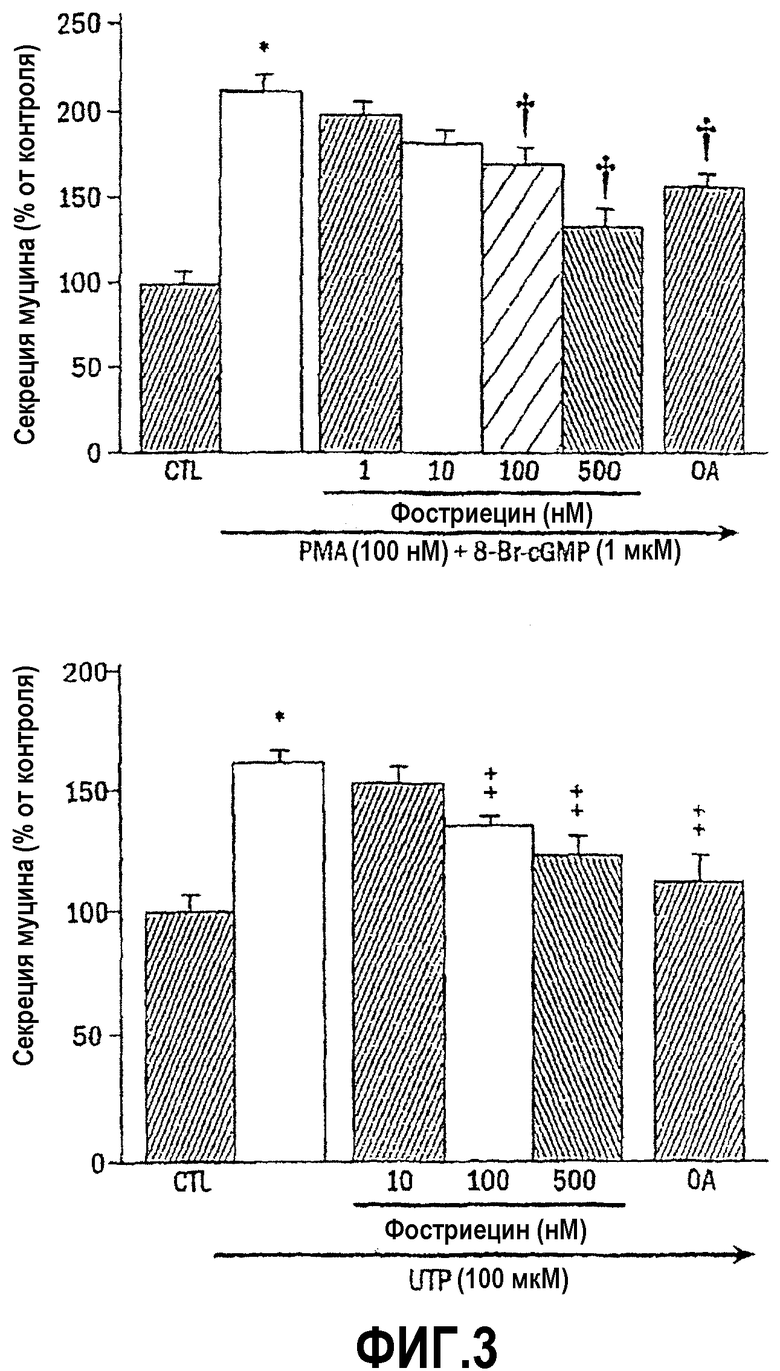
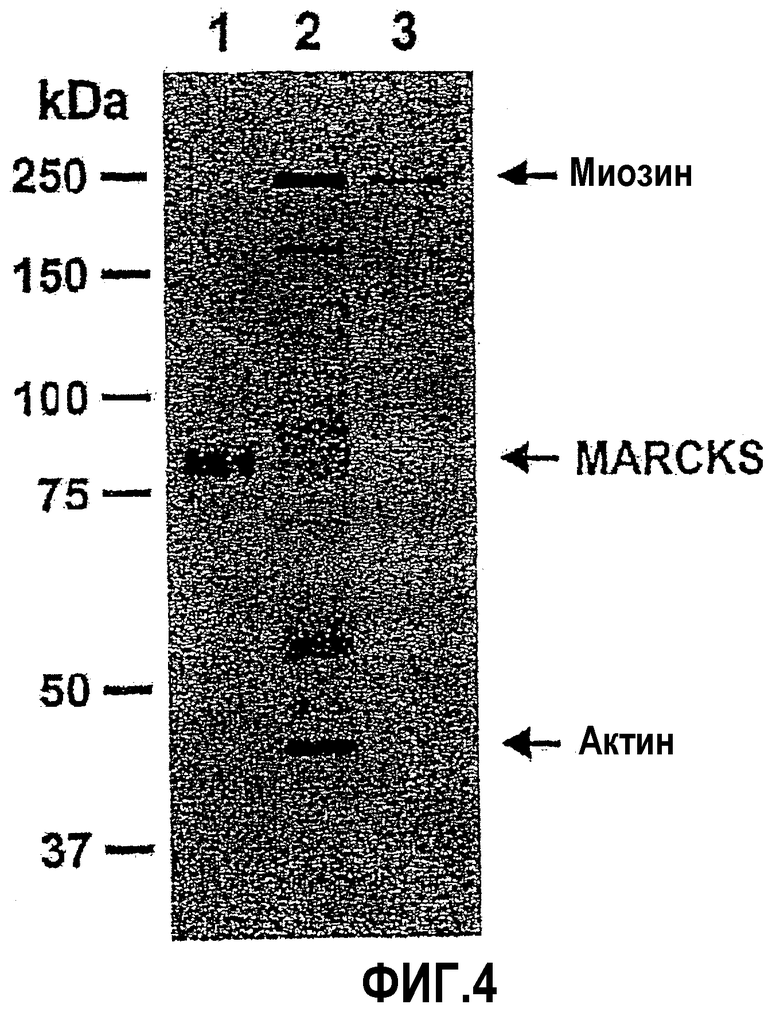
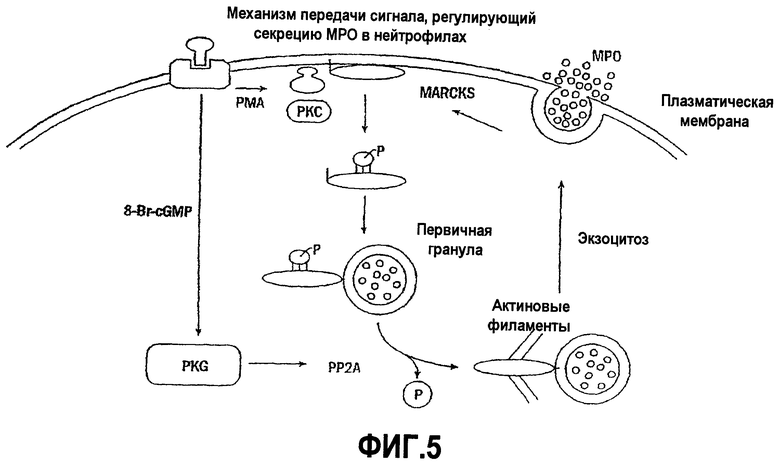
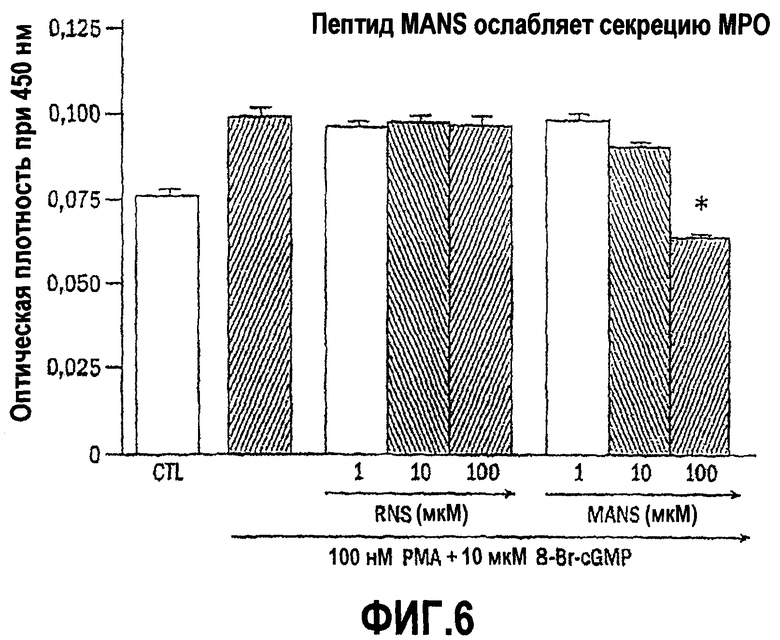
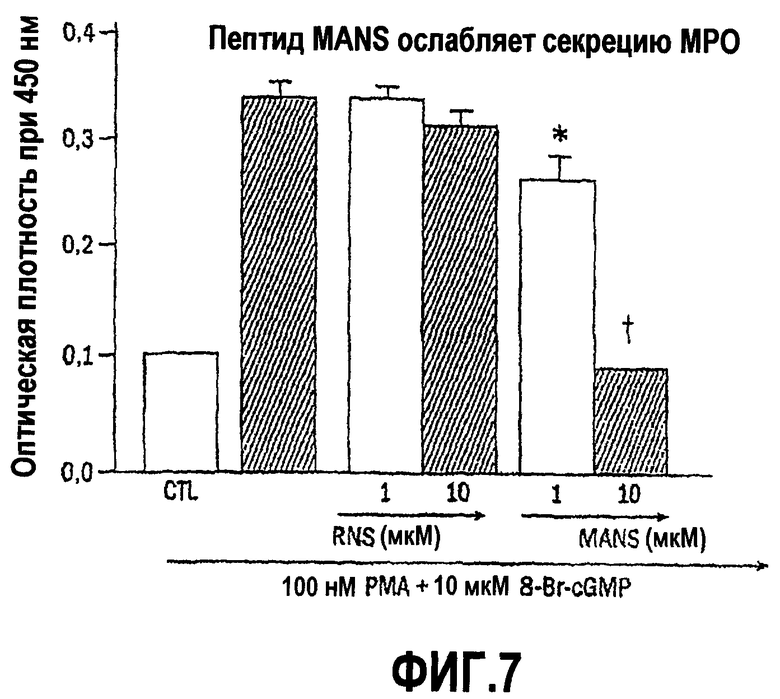
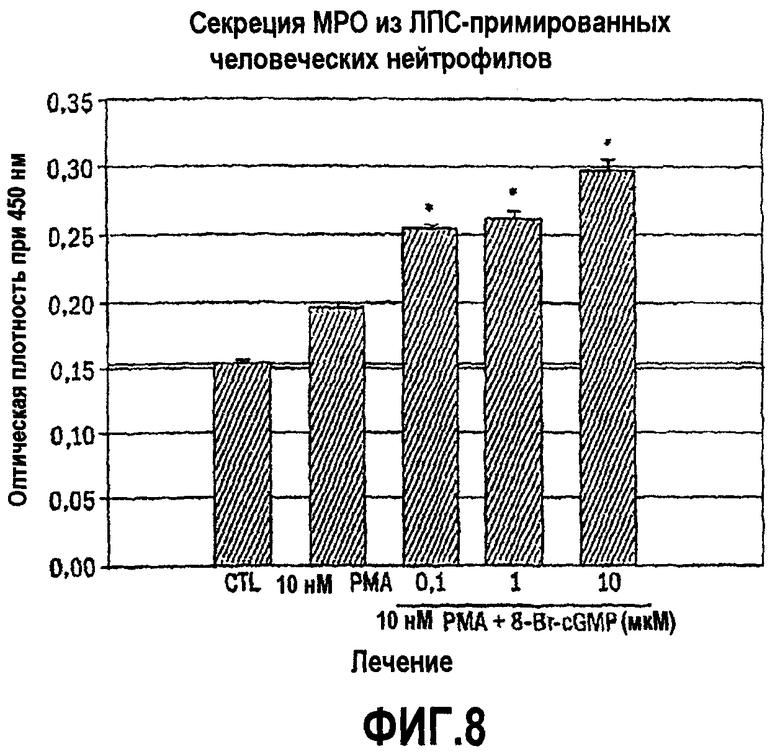
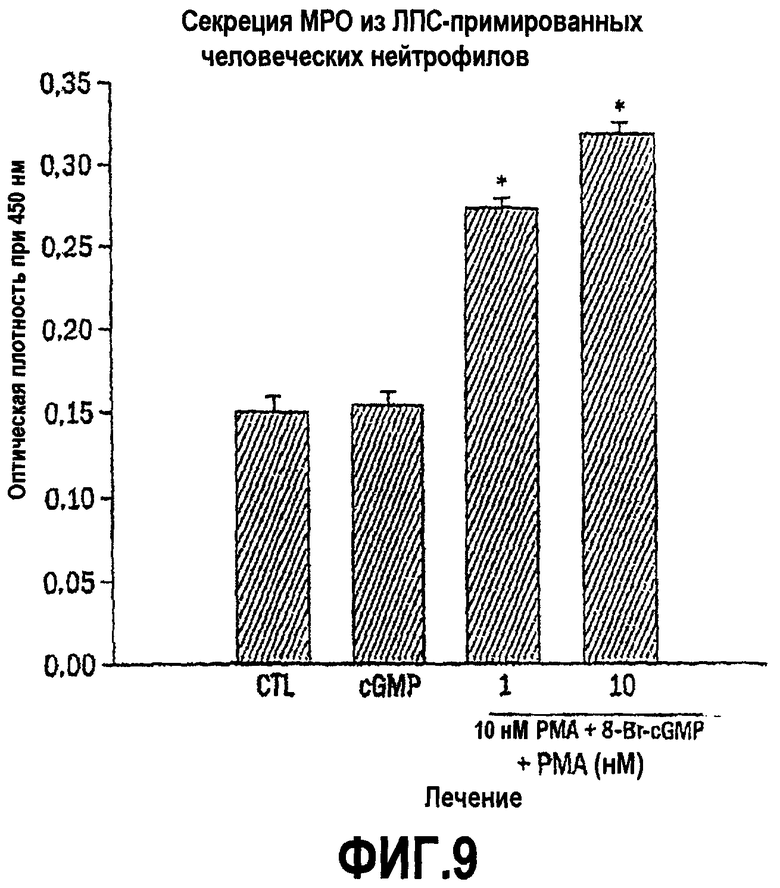
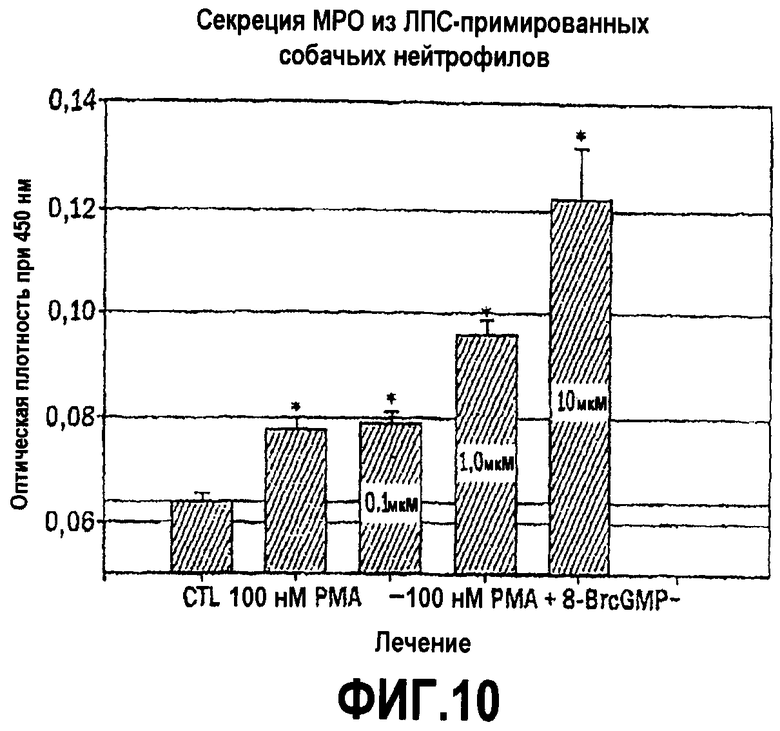
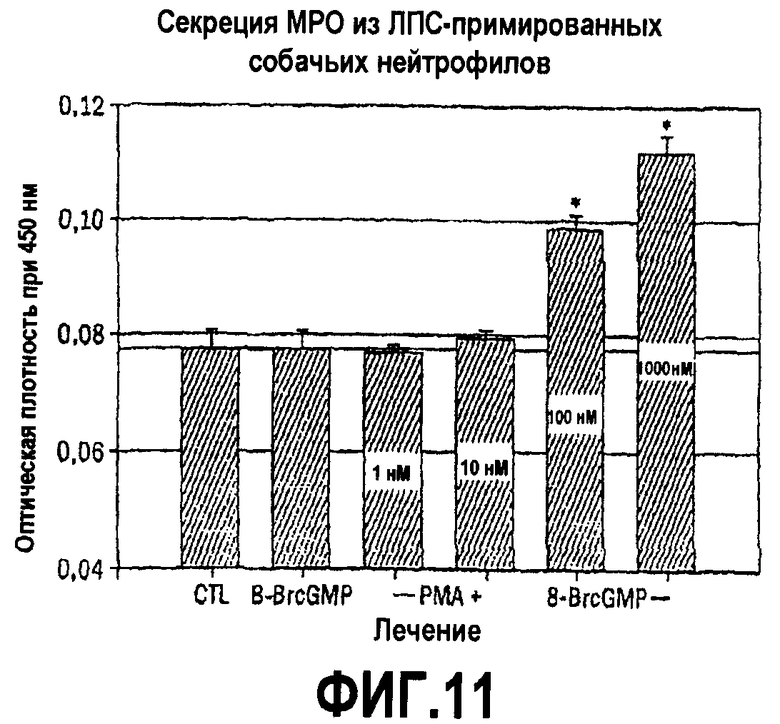
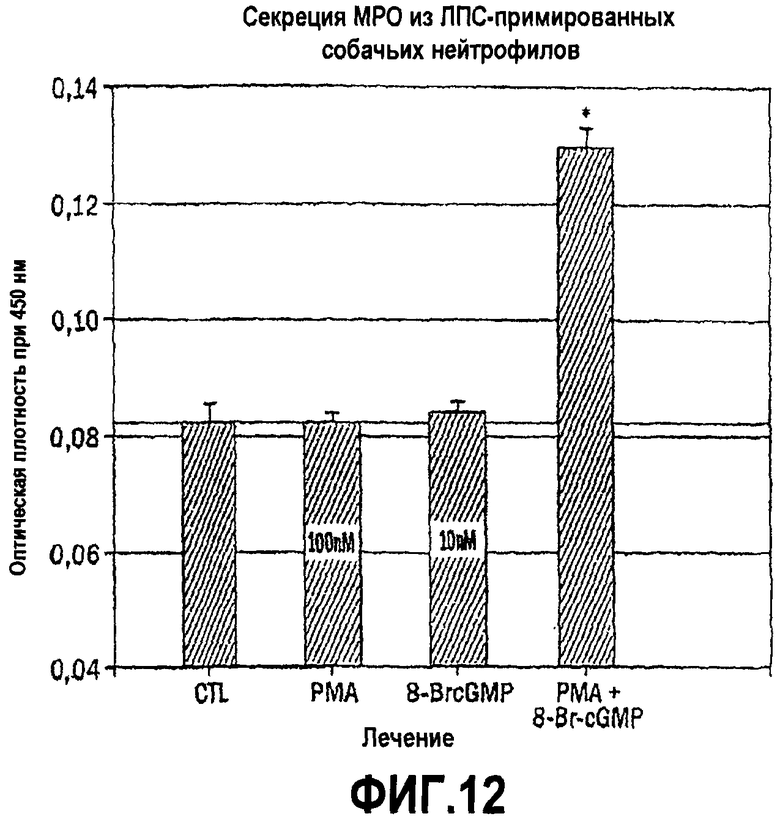
Изобретение относится к ингибированию или снижению уровня высвобождения медиаторов воспаления из воспалительных клеток путем подавления механизма, ассоциированного с высвобождением медиаторов воспаления из гранул воспалительных клеток путем использования вариантов пептида MANS. В соответствии с этим настоящее изобретение относится к внутриклеточному механизму передачи сигнала, который позволяет обнаруживать несколько новых внутриклеточных мишеней для фармакологического лечения расстройств, ассоциированных с секрецией медиаторов воспаления из везикул воспалительных клеток. 24 з.п. ф-лы, 8 табл., 15 ил.
1. Способ ингибирования MARCKS-ассоциированного высвобождения по меньшей мере одного медиатора воспаления из гранул по меньшей мере одной воспалительной клетки в ткани и/или жидкости индивида, включающий:
введение в указанную ткань и/или жидкость терапевтически эффективного количества фармацевтической композиции, содержащей по меньшей мере один пептид, ацетилированный по N-концевой альфа-аминогруппе, с аминокислотной последовательностью, выбранной из группы, состоящей из:
(a) последовательности из 24 аминокислот, GAQFSKTAAKGEAAAERPGEAAVA (SEQ ID NO:1);
(b) аминокислотной последовательности размером от 4, но не более 23, последовательных аминокислот эталонной последовательности GAQFSKTAAKGEAAAERPGEAAVA (SEQ ID NO:1);
(c) последовательности с одной аминокислотной заменой, отличающейся одной аминокислотой от последовательности, определенной в подпункте (b), где единичная замена выбрана из группы, состоящей из G в положении 1 заменен А, А в положении 2 заменен К, Q в положении 3 заменен А или Е, F в положении 4 заменен A, S в положении 5 заменен А, К в положении 6 заменен А, Т в положении 7 заменен А, А в положении 8 заменен К, К в положении 10 заменен A, G в положении 11 заменен А и Е в положении 12 заменен А; и
(d) последовательности с двумя аминокислотными заменами, отличающейся двумя аминокислотами от последовательности, определенной в подпункте (b), где две замены выбраны из группы, состоящей из А в положениях 2 и 8 заменены К, а К в положениях 6 и 10 заменены А;
где С-концевая аминокислота пептида с аминокислотной последовательностью, определенной в подпункте (а), или (b), или (с), или (d), химически модифицирована, например, амидом аммиака или немодифицирована на группе карбоновой кислоты С-конца; и
где указанный пептид необязательно объединен с фармацевтически приемлемым носителем, и в количестве, терапевтически эффективном для ослабления высвобождения медиаторов воспаления, снижает уровень высвобождения медиатора воспаления по меньшей мере из одной воспалительной клетки по сравнению с уровнем высвобождения указанного медиатора воспаления по меньшей мере из одной воспалительной клетки того же типа, в отсутствие указанного пептида.
2. Способ по п.1, где указанный пептид, ацетилированный по N-концевой альфа-аминогруппе, выбран из группы, состоящей из ацетил-GAQFSKTAAK (SEQ ID NO:106), ацетил-GAQFSKTAAKGEAAAERPGEAAVA (SEQ ID NO:1), ацетил-GAQFSKTAAKGEAAAERPGE (SEQ ID NO:11), ацетил-GAQFSKTAAKGEAAAE (SEQ ID NO:37), ацетил-AKGEAAAERPGEAAVA (SEQ ID NO:45), ацетил-GAQFSKTAAKGE (SEQ ID NO:79), ацетил-AAAERPGEAAVA (SEQ ID NO:91), ацетил-GAQFSKTAA (SEQ ID NO:121), ацетил-TAAKGEAA (SEQ ID NO:143), ацетил-RPGEAAVA (SEQ ID NO:153), ацетил-AAKGEA (SEQ ID NO:179), ацетил-AKGE (SEQ ID NO:219), ацетил-GAQFSKTAAKGE-NH2 (SEQ ID NO:79), ацетил-AQFSKTAAKGE-NH2 (SEQ ID NO:93), ацетил-QFSKTAAKGE-NH2 (SEQ ID NO:108), ацетил-FSKTAAKGE-NH2 (SEQ ID NO:124), ацетил-SKTAAKGE-NH2 (SEQ ID NO:141), ацетил-KTAAKGE-NH2 (SEQ ID NO:159) и ацетил-AKGE-NH2 (SEQ ID NO:219).
3. Способ по п.1 или 2, где указанный пептид состоит по меньшей мере из десяти смежных аминокислотных остатков.
4. Способ по п.3, где указанный пептид состоит из ацетил-пептида 106 (SEQ ID NO:106).
5. Способ по п.1 или 2, где аминокислотная последовательность пептида включает в себя смежные аминокислотные остатки AKGE.
6. Способ по п.1, где указанный пептид состоит по меньшей мере из шести смежных аминокислотных остатков.
7. Способ по п.1 или 2, где указанный пептид амидирован аммиаком у С-концевой альфа-аминокислоты.
8. Способ по п.1 или 6, где указанный пептид включает в себя аминокислотную последовательность (b) и не начинается с N-концевой аминокислоты эталонной последовательности GAQFSKTAAKGEAAAERPGEAAVA (SEQ ID NO:1).
9. Способ по п.8, где указанный пептид амидирован аммиаком у С-концевой альфа-аминокислоты.
10. Способ по п.1, где аминокислотная последовательность с единичной заменой (с) выбрана из группы, состоящей из GKQFSKTAAKGE (SEQ ID NO:233), GAQFSKTKAKGE (SEQ ID NO:234), GAQASKTAAK (SEQ ID NO:236), GAQASKTAAKGE (SEQ ID NO:237), GAEFSKTAAKGE (SEQ ID NO:238), GAQFSKTAAAGE (SEQ ID NO:239), GAQFSKTAAKAE (SEQ ID NO:240), GAQFSKTAAKGA (SEQ ID NO:241), AAQFSKTAAK (SEQ ID NO:242), GAAFSKTAAK (SEQ ID NO:243), GAQFAKTAAK (SEQ ID NO:244), GAQFSATAAK (SEQ ID NO:245), GAQFSKAAAK (SEQ ID NO:247), GAQFSKTAAA (SEQ ID NO:248), GAQASKTA (SEQ ID NO:250) и AAGE (SEQ ID NO:251).
11. Способ по п.1, где аминокислотная последовательность с двумя заменами выбрана из группы, состоящей из GKQFSKTKAKGE (SEQ ID NO:235) и GAQFSATAAA (SEQ ID NO:249).
12. Способ по п.10 или 11, где указанный пептид амидирован аммиаком на С-концевой альфа-аминокислоте.
13. Способ по п.1, где указанная воспалительная клетка выбрана из группы, состоящей из лейкоцита, гранулоцита, нейтрофила, базофила, эозинофила, моноцита, макрофага и их сочетаний.
14. Способ по п.1 или 13, где указанный медиатор воспаления выбран из группы, состоящей из миелопероксидазы (МРО), пероксидазы эозинофилов (ЕРО), основного мажорного белка [МБР], лизоцима, гранзима, гистамина, протеогликана, протеазы, хемотаксического фактора, цитокина, метаболита арахидоновой кислоты, дефензина, белка, повышающего проницаемость бактерий (BPI), эластазы, катепсина G, катепсина В, катепсина D, бета-D-глюкуронидазы, альфа-маннозидазы, фосфолипазы А2, хондроитин-4-сульфата, протеиназы 3, лактоферрина, коллагеназы, активатора комплемента, рецептора комплемента, рецептора N-формилметионил-лейцил-фенилаланина (FMLP), рецептора ламинина, цитохрома b558, моноцитарного хемотаксического фактора, гистаминазы, белка, связывающегося с витамином В12, желатиназы, активатора плазминогена, бета-D-глюкуронидазы и их комбинаций.
15. Способ по п.1, где указанное эффективное количество указанного пептида, способствующее снижению уровня высвобождения медиатора воспаления, включает количество пептида, ингибирующее дегрануляцию, которое снижает уровень медиатора воспаления, высвобождаемого по меньшей мере из одной воспалительной клетки, (i) от 1% примерно до 99% или (ii) от 5-50% до 99% по сравнению с уровнем, высвобождаемым по меньшей мере из одной воспалительной клетки в отсутствие указанного пептида.
16. Способ по п.1, где указанный медиатор воспаления связан с респираторным заболеванием.
17. Способ по п.16, где указанное респираторное заболевание выбрано из группы, состоящей из астмы, хронического бронхита, ХОБЛ и кистозного фиброза.
18. Способ по п.1, где указанное введение выбрано из группы, состоящей из местного применения, парентерального введения, ректального введения, легочного введения, интраназальноого введения и перорального приема.
19. Способ по п.18, где указанное легочное введение включает введение с помощью аэрозоля.
20. Способ по п.19, где указанный аэрозоль получен из ингалятора сухого порошка, ингалятора с дозирующим клапаном или небулайзером.
21. Способ по п.1, дополнительно включающий введение указанному индивиду второй молекулы, выбранной из группы, состоящей из антибиотика, противовирусного соединения, антипаразитарного соединения, противовоспалительного соединения и иммуномодулятора.
22. Способ по п.1, где указанный медиатор воспаления связан с заболеванием, выбранным из группы, состоящей из кишечного заболевания, кожного заболевания, аутоиммунного заболевания, болевого синдрома и их комбинаций.
23. Способ по п.22, где указанное кишечное заболевание выбрано из группы, состоящей из язвенного колита, болезни Крона и синдрома раздраженного кишечника.
24. Способ по п.22, где указанное кожное заболевание выбрано из группы, состоящей из розацеи, экземы, псориаза и угревой сыпи в тяжелой форме.
25. Способ по п.1, где указанный медиатор воспаления связан с артритом.
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| WO 00/50062 A2, 31.08.2000. | |||

