Перекрестные ссылки на связанные заявки
Настоящая заявка заявляет приоритет патентной заявки США № 11/367449, поданной 6 марта 2006, которая является частичным продолжением патентной заявки США № 10/802644, поданной 17 марта 2004, которая является продолженной заявкой для патентной заявки США № 10/180753, поданной 26 июня 2002, в настоящее время отозванной, которая заявляет приоритет предварительной заявки США № 60/300933, поданной 26 июня 2001, описание которых включено в настоящее описание полностью в виде ссылки.
Заявление о федеральной поддержке
Настоящее изобретение было осуществлено при поддержке Федерального правительства Соединенных Штатов в рамках гранта номер R01 HL36982 от Национального Института Здоровья. Правительство Соединенных Штатов может иметь определенные права на это изобретение.
Область изобретения
Настоящее изобретение относится к способам модуляции клеточных секреторных процессов. В частности, настоящее изобретение относится к модуляции высвобождения медиаторов воспаления. Настоящее изобретение также относится к внутриклеточному сигнальному механизму, регулирующему секрецию медиаторов воспаления из связанных с мембраной везикул или гранул в воспалительных клетках.
Предшествующий уровень техники
Повышенная секреция слизи участвует в патогенезе большого количества воспалительных заболеваний дыхательных путей и у людей и у животных, не принадлежащих к человеческому роду. Повышенную секрецию слизи наблюдают при хронических патологических состояниях, таких как астма, ХОБЛ (COPD) и хронический бронхит; при генетических заболеваниях, таких как муковисцидоз; при аллергических состояниях (атопия, аллергическое воспаление); при бронхоэктазах; при ряде острых инфекционных заболеваний дыхательных путей, таких как пневмония, ринит, грипп или простуда.
Повышенной секреции слизи при множестве таких заболеваний дыхательных путей сопутствует постоянное присутствие в дыхательных путях воспалительных клеток. Такие клетки участвуют в патогенезе указанных заболеваний преимущественно посредством повреждения тканей, осуществляемого медиаторами воспаления, высвобождаемыми из таких клеток. Одним примером такого разрушения посредством хронического воспаления являются пациенты с муковисцидозом, когда медиаторы, высвобождаемые из нейтрофилов (например, миелопероксидаза), вызывают десквамацию эпителиальной ткани дыхательных путей.
Пониженная секреция слизи также обладает повреждающим действием. Слизь дыхательных путей действует как физический барьер против вдыхаемых биологически активных частиц и может помочь предотвратить бактериальную колонизацию дыхательных путей и инактивировать цитотоксические продукты, высвобождаемые из лейкоцитов. King et al., Respir. Physiol. 62: 47-59 (1985); Vishwanath and Ramphal, Infect. Immun. 45: 197 (1984); Cross et al., Lancet 1: 1328 (1984). В глазу слизь поддерживает слезную пленку и важна для здоровья и комфорта глаза. Секреция слизи в желудочно-кишечном тракте также обладает цитопротективным действием. Роль слизи в качестве химического, биологического и механического барьера обозначает, что ненормально низкая секреция слизи слизистыми мембранами является нежелательной.
Дыхательные пути млекопитающих выстланы тонким слоем слизи, продуцируемой и секретируемой эпителиальными клетками дыхательных путей (бокаловидными) и подслизистыми железами. При заболеваниях дыхательных путей, таких как астма, хронический бронхит и муковисцидоз, повышенная секреция слизи является частым симптомом. Избыток слизи может вносить вклад в обструкцию и подверженность инфекции. Основным компонентом слизи являются гликопротеины муцина, синтезируемые секреторными клетками и хранящиеся в цитоплазматических гранулах. Муцины представляют собой гликопротеины, секретируемые эпителиальными клетками, включая таковые в дыхательном, желудочно-кишечном и женском репродуктивном тракте. Муцины отвечают за вязко-эластические свойства слизи и известны, по меньшей мере, восемь генов муцинов. Thornton, et al., J.Biol.Chem. 272, 9561-9566 (1997). Повреждение реснитчатого эпителия, вызванное повышенной секрецией муцина и/или гиперплазией слизистых клеток, приводит к закупориванию слизью дыхательных путей, что способствует хронической инфекции, обструкции тока воздуха и иногда смерти. Множество заболеваний дыхательных путей, такие как хронический бронхит, хронические обструктивные болезни легких, бронхоэктазы, астма, муковисцидоз и бактериальные инфекции характеризуются повышенной продукцией муцина. E. Prescott, et al. Eur. Respir.J., 8: 1333-1338 (1995); K.C. Kim, et al., Eur. Respir.J., 10: 1438 (1997); D. Steiger, et al., Am. J.Respir. Cell Mol. Biol., 12:307-314 (1995). При соответствующей стимуляции, гранулы муцина высвобождаются посредством процесса экзоцитоза, при котором гранулы переносятся на периферию клетки, где мембраны гранул соединяются с плазматической мембраной, позволяя секретировать содержимое в просвет.
Несмотря на очевидную патофизиологическую важность такого процесса, внутриклеточные сигнальные механизмы, связывающие стимуляцию на поверхности клеток с высвобождением гранул муцина, были описаны только недавно. См. Li et al., Journal of Biological Chemistry, 276: 40982-40990 (2001). Известно, что большое множество веществ и медиаторов воспаления/гуморальных медиаторов способствуют секреции слизи. Они включают холинергические вещества, липидные медиаторы, оксиданты, цитокины, нейропептиды, АТФ и УТФ, бактериальные продукты, эластазу нейтрофилов и ингалируемые загрязнители. См. Adler et al., Res. Immunol. 149, 245-248 (1998). Интересно, что также известно, что множество таких стимуляторов секреции муцина активируют некоторые протеинкиназы, и исследования, изучающие регуляцию избыточной секреции муцина различными видами эпителиальных клеток дыхательных путей, постоянно включают участие в секреторном процессе или протеинкиназы С (РКС) или цГМФ-зависимой протеинкиназы (PKG). См., например, Ko et al., Am. J.Respir. Cell Mol. Biol. 16, 194-198 (1997); Abdullah et al., Am. J. Physiol. 273, L201-L210 (1997); Abdullah et al., Biochem. J. 316, 943-954 (1996); Larivee et al., Am.J.Respir. Cell Mol. Biol.11, 199-205 (1994); и Fisher et al., Am.J.Respir. Cell Mol. Biol. 20, 413-422 (1999). Только недавно было продемонстрировано, что координированные взаимодействия или «взаимное влияние» между такими двумя протеинкиназами в регуляции секреции муцина включают белки MARCKS. См. Li et al., Journal of Biological Chemistry, 276: 40982-40990 (2001). Однако сигнальные события после совместного действия таких протеинкиназ, которое, в конечном счете, приводит к экзоцитозному высвобождению гранул муцина, полностью не освещены.
MARCKS, белок, приблизительно 82 кДа, обладает тремя эволюционно-консервативными участками (Aderem et al., Nature 1998; 332: 362-364; Thelen et al., Nature 1991; 351:320-322; Hartwig et al., Nature 1992; 356: 618-622; Seykora et al., J. Biol. Chem. 1996; 271: 18797-18802): N-конец, домен сайта фосфорилирования (PSD) и домен множественной гомологии 2 (МН2). N-конец, последовательность 24 аминокислот с компонентом миристиновой кислоты, прикрепленным к концевому остатку глицина, участвует в связывании MARCKS с мембранами (Seykora et al., J. Biol. Chem. 1996; 271: 18797-18802) и возможно с кальмодулином (Matsubara et al., J. Biol. Chem. 2003; 278: 48898-48902). Эта последовательность из 24 аминокислот известна как MANS-пептид. MANS-пептид и его активные фрагменты могут конкурировать с нативным MARCKS в клетках в отношении связывания с мембраной. Участие белка MARCKS в высвобождении медиаторов воспаления из гранул инфильтрирующих лейкоцитов имеет отношение к воспалению при заболеваниях во всех тканях и органах, включая заболевания легких, характеризуемые воспалением дыхательных путей, такие как астма, ХОБЛ и муковисцидоз. Однако воспаление и секреция слизи в дыхательных путях представляют собой два отдельных и независимых процесса (Li et al., J.Biol. Chem. 2001; 276: 40982-40990; Singer et al., Nat.Med 2004; 10: 193-196). Тогда как продукция и секреция слизи могут быть вызваны рядом факторов, включая медиаторы, высвобождаемые из воспалительных клеток, прямая связь между избытком слизи и воспалением неизвестна.
Сущность изобретения
Изобретение относится к новому применению 24-аминокислотного, миристоилированного полипептида, также известного как MANS-пептид. Изобретение также относится к новому способу блокирования любого клеточного секреторного процесса, особенно такого, который включает высвобождение медиаторов воспаления из воспалительных клеток, чьи стимулирующие пути включают субстрат протеинкиназы С (РКС)-белок MARCKS, и высвобождение содержимого из внутриклеточных везикул или гранул.
Настоящее изобретение относится к способу ингибирования экзоцитозного высвобождения, по меньшей мере, одного медиатора воспаления из, по меньшей мере, одной воспалительной клетки, включающему контакт, по меньшей мере, одной воспалительной клетки, которая содержит, по меньшей мере, один медиатор воспаления, содержащийся в везикуле в клетке, с, по меньшей мере, одним пептидом, выбираемым из группы, состоящей из MANS-пептида и его активного фрагмента в эффективном количестве для уменьшения высвобождения медиатора воспаления из воспалительной клетки по сравнению с высвобождением медиатора воспаления из воспалительной клетки такого же типа, которое возникает в отсутствие, по меньшей мере, одного пептида.
Настоящее изобретение, кроме того, относится к способу ингибирования высвобождения, по меньшей мере, одного медиатора воспаления из, по меньшей мере, одной воспалительной клетки в ткани или жидкости пациента, включающему введение в ткани и/или жидкости пациента, которые содержат, по меньшей мере, одну воспалительную клетку, содержащую, по меньшей мере, один медиатор воспаления, содержащийся в везикуле в клетке, терапевтически эффективного количества фармацевтической композиции, включающей, по меньшей мере, один пептид, выбираемый из группы, состоящей из MANS-пептида и его активного фрагмента, в терапевтически эффективном количестве для снижения высвобождения медиатора воспаления из, по меньшей мере, одной воспалительной клетки, по сравнению с высвобождением медиатора воспаления из, по меньшей мере, одной воспалительной клетки того же типа, которое возникает в отсутствие, по меньшей мере, одного пептида. В частности, ингибирование высвобождения медиатора воспаления включает блокаду или уменьшение высвобождения медиатора воспаления из воспалительной клетки.
В частности, настоящее изобретение включает способ уменьшения воспаления у пациента, включающий введение терапевтически эффективного количества фармацевтической композиции, содержащей MANS-пептид [т.е. N-миристоил-GAQFSKTAAKGEAAAERPGEAAV (SEQ.ID.NO:20)] или его активный фрагмент. Активный фрагмент представляет собой, по меньшей мере, шесть аминокислот в длину. Как используется в настоящем описании, «активный фрагмент» MARCKS-белка представляет собой таковой, который влияет на (ингибирует или усиливает) высвобождение, опосредованное MARCKS белком. Активный фрагмент может быть выбран из группы, состоящей из N-миристоил-GAQFSKTAAKGEAAAERPGEAA (SEQ.ID.NO:3); N-миристоил-GAQFSKTAAKGEAAAERPGEA (SEQ.ID.NO:4); N-миристоил-GAQFSKTAAKGEAAAERPGE (SEQ.ID.NO:5); N-миристоил-GAQFSKTAAKGEAAAERPG (SEQ.ID.NO:6); N-миристоил-GAQFSKTAAKGEAAAERP (SEQ.ID.NO:7); N-миристоил-GAQFSKTAAKGEAAAER (SEQ.ID.NO:8); N-миристоил-GAQFSKTAAKGEAAAE (SEQ.ID.NO:9); N-миристоил-GAQFSKTAAKGEAAA (SEQ.ID.NO:10); N-миристоил-GAQFSKTAAKGEAA (SEQ.ID.NO:11); N-миристоил-GAQFSKTAAKGEA (SEQ.ID.NO:12); N-миристоил-GAQFSKTAAKGE (SEQ.ID.NO:13); N-миристоил-GAQFSKTAAKG (SEQ.ID.NO:14); N-миристоил-GAQFSKTAAK (SEQ.ID.NO:15); N-миристоил-GAQFSKTAA (SEQ.ID.NO:16); N-миристоил-GAQFSKTA (SEQ.ID.NO:17); N-миристоил-GAQFSKT (SEQ.ID.NO:18); и N-миристоил-GAQFSK (SEQ.ID.NO:19). Присутствие гидрофобного N-концевого миристатного компонента в таких пептидах может усиливать их совместимость с и, вероятно, их проницаемость через плазматические мембраны и потенциально делает возможным поглощение пептидов клетками. Гидрофобная вставка миристата в бислой может обеспечивать коэффициент распределения или наблюдаемую константу ассоциации с липидами до 104 М-1 или свободную энергию связывания Гиббса около 8 ккал/моль (см., например, Peitzsch, R.M. и McLaughlin, S. 1993, Binding of acylated peptides and fatty acids to phospholipid vesicles: pertinence to myristoylated proteins. Biochemistry. 32: 10436-10443), что является достаточным, по меньшей мере, частично для возможности распределения MANS-пептида и пептидных фрагментов MANS-пептида в плазматической мембране клеток, как описано в настоящем описании, тогда как дополнительные функциональные группы и их взаимодействия в MANS-пептиде (который является миристоилированным) и в пептидных фрагментах миристоилированного MANS-пептида могут потенциировать их относительную мембранную проницаемость. Каждый фрагмент может проявлять коэффициенты распределения и сродство к мембране, которые характерны для их соответствующей структуры. Фрагменты могут быть получены методами пептидного синтеза, известными в области техники, такими как твердофазный пептидный синтез (см., например, методы, описанные в Chan, Wеng C. and White, Peter D. Eds., Fmoc Solid Phase Peptide Synthesis: A Practical Approach, Oxford University Press, New York, New York (2000); и Lloyd-Williams, P. Et al. Chemical Approaches to the Synthesis of Peptides and Proteins (1997)) и очищены методами, известными в области техники, такими как высокоэффективная жидкостная хроматография. Молекулярная масса каждого пептида может быть подтверждена масс-спектроскопией, с каждым показанным пиком с соответствующей молекулярной массой. Эффективность отдельных пептидов и комбинаций отдельных пептидов (например, комбинации 2 пептидов, комбинации 3 пептидов, комбинации 4 пептидов) в способах по настоящему описанию могут быть легко определены без ненужных экспериментов с использованием методик, описанных в примерах, представленных в настоящем описании. Предпочтительная комбинация включает два пептида; молярное соотношение пептидов может составлять предпочтительно от 50:50 до 99,99:0,01, такое соотношение может быть легко определено с использованием методик, описанных в примерах, представленных в настоящем описании.
Предпочтительно MANS-пептид или его активный фрагмент содержится в фармацевтической композиции, которую применяют для блокады воспаления. Настоящее изобретение также включает способы контроля клеточного секреторного процесса у пациента, включающие введение терапевтически эффективного количества соединения, содержащего MANS-пептид или его активный фрагмент, которое регулирует медиаторы воспаления у пациента. Введение обычно выбирают из группы, состоящей из местного введения, парентерального введения, ректального введения, легочного введения, ингаляции и назального или перорального введения, где легочное введение обычно включает или аэрозоль, или введения с помощью ингалятора сухого порошка, ингалятора с дозирующим клапаном или небулайзера.
Введение композиции, содержащей количество MANS-пептида, ингибирующее дегрануляцию, или количество его активного фрагмента, ингибирующее дегрануляцию, так что фармацевтическая композиция MANS-пептида или его активного фрагмента для применения у человека или животного обеспечивает MANS-пептид или его активный фрагмент, по меньшей мере, в месте или в ткани, или в жидкостьсодержащем, или слизьсодержащем слое в контакте с поверхностью ткани, где находятся воспалительные гранулоцитарные клетки или в которые воспалительные гранулоцитарные клетки могут внедряться, таким образом, позволяя MANS-пептиду или его фрагменту контактировать с воспалительной гранулоцитарной клеткой. В одном аспекте введение такой композиции может быть осуществлено при первичном начале, или первичном определении воспаления, или первом ощущении воспаления человеком или животным, или при первом ощутимом изменении степени воспаления у человека или животного для уменьшения выраженности воспаления, которое возникает в отсутствие MANS-пептида или его активного фрагмента. В другом аспекте введение может быть осуществлено во время текущего воспаления ткани у человека или животного для уменьшения выраженности дополнительного воспаления, которое иным образом возникает в отсутствие MANS-пептида или его активного фрагмента. Тогда как количество и частота дозирования могут быть определены путем клинического обследования и являются функцией заболевания или источника воспаления и степени вовлечения ткани, и возраста, и размера пациента, считают, что дозирование фармацевтической композиции может быть повторено через 3-8 часов, предпочтительно через 6-8 часов после первого введения фармацевтической композиции.
Настоящее изобретение также включает способы уменьшения воспаления у пациента, включающие введение терапевтически эффективного количества соединения, которое ингибирует MARCKS-связанное высвобождение медиаторов воспаления, вследствие чего высвобождение, по меньшей мере, одного медиатора воспаления у пациента снижается по сравнению с таковым, которое возникает в отсутствие указанного лечения. Как используется в настоящем описании, «снижение» обычно обозначает уменьшение эффектов воспаления. Предпочтительно, медиаторы воспаления ингибируются или блокируются описанными способами.
Другой вариант осуществления настоящего изобретения включает способы уменьшения воспаления у пациента, включающие введение терапевтически эффективного количества соединения, которое ингибирует MARCKS-связанное высвобождение медиаторов воспаления, вследствие чего воспаление у пациента уменьшается по сравнению с таковым, которое возникает в отсутствие указанного лечения. Настоящее изобретение также описывает способы снижения или ингибирования воспаления у пациента, включающие введение терапевтически эффективного количества MANS-пептида или его активного фрагмента, эффективного для модуляции медиатора воспаления в месте воспаления. Термин «ингибирование» обозначает снижение количества секреции медиатора воспаления. Термин «полное ингибирование» обозначает снижение степени секреции медиатора воспаления до нуля. Также, как указано выше, активный фрагмент имеет по меньшей мере шесть аминокислот в длину. Термин «экзоцитозный процесс» обозначает экзоцитоз, т.е. процесс клеточной секреции или экскреции, при которой вещества, содержащиеся в везикулах, которые находятся внутри клетки, выделяются из клетки путем сшивки мембраны везикул с наружной клеточной мембраной. «Дегрануляция» обозначает высвобождение содержимого клеточных гранул. Термин «ингибирование дегрануляции» обозначает снижение высвобождения медиаторов воспаления, содержащихся в гранулах воспалительных клеток. Следовательно, количество MANS-пептида и/или его активного фрагмента, ингибирующее дегрануляцию, представляет собой количество таких пептидов, которое достаточно для снижения высвобождения медиаторов воспаления, содержащихся в гранулах, по сравнению с высвобождением в отсутствие того же пептида.
MANS-пептид и его активные фрагменты могут быть применимыми при профилактике или уменьшении выраженности воспаления, вызванного медиаторами воспаления в ткани животного. MANS-пептид и его активные фрагменты могут быть применимы при профилактике или уменьшении степени повреждения ткани у животного, вызываемого или вызванного медиаторами воспаления.
Краткое описание чертежей
Фиг. 1А-1D представляют собой гистограммы, иллюстрирующие, что гиперсекреция муцина клетками NHBE доводится до максимума путем активации и РКС, и PKG.
На фиг. 2А-2В продемонстрировано, что MARCKS-белок является ключевым компонентом секреторного пути муцина.
На фиг. 3А-3С изображен гель, иллюстрирующий, что антисмысловой нуклеотид, направленный против MARCKS, подавляет экспрессию MARCKS и ослабляет гиперсекрецию муцина.
На фиг. 4А-4В проиллюстрировано, что РКС-зависимое фосфорилирование высвобождает MARCKS из плазматической мембраны в цитоплазму.
На фиг. 5А-5С показано, что PKG индуцирует дефосфорилирование MARCKS путем активации РР2А.
На фиг. 6 изображены гистограммы, которые демонстрируют, что РР2А является обязательным компонентом секреторного пути муцина.
Фиг. 7 представляет собой гель, который иллюстрирует, что MARCKS ассоциируется с актином и миозином в цитоплазме.
На фиг. 8 изображен сигнальный механизм, регулирующий секрецию муцина эпителиальными клетками дыхательных путей человека.
Фиг. 9 представляет собой гистограмму, изображающую способность MANS-пептида блокировать секрецию миелопероксидазы из выделенных нейтрофилов собак.
Фиг. 10 представляет собой гистограмму, изображающую способность MANS-пептида блокировать секрецию миелопероксидазы из выделенных человеческих нейтрофилов.
Фиг. 11 представляет собой гистограмму, показывающую, что РМА стимулирует небольшое увеличение секреции МРО из человеческих нейтрофилов, стимулированных LPS, которая усиливается в зависимости от концентрации путем совместной стимуляции с помощью 8-Br-cGMP.
Фиг. 12 представляет собой гистограмму, показывающую, что стимуляция 8-Br-cGMP обладает небольшим эффектом на секрецию МРО из человеческих нейтрофилов, стимулированных LPS, до возникновения совместной стимуляции с РМА в зависимости от концентрации.
Фиг. 13 представляет собой гистограмму, показывающую, что РМА стимулирует небольшое увеличение секреции МРО из собачьих нейтрофилов, стимулированных LPS, которая усиливается в зависимости от концентрации путем совместной стимуляции с помощью 8-Br-cGMP.
Фиг. 14 представляет собой гистограмму, показывающую, что стимуляция 8-Br-cGMP обладает небольшим эффектом на секрецию МРО из собачьих нейтрофилов, стимулированных LPS, до возникновения совместной стимуляции с РМА в зависимости от концентрации.
Фиг.15 представляет собой гистограмму, показывающую, что для максимальной секреции МРО из нейтрофилов собак, стимулированных LPS, требуется совместная стимуляция PMA + 8-Br-cGMP.
Подробное описание изобретения
Далее настоящее описание будет описано более полно со ссылками на сопутствующие чертежи, в которых проиллюстрированы предпочтительные варианты осуществления изобретения. Настоящее изобретение, однако, может быть воплощено в различных формах и не должно расцениваться как ограниченное вариантами осуществления изобретения, указанными в настоящем описании. Более того, обеспечены такие варианты осуществления изобретения, посредством которых настоящее описание является полным и основательным и полностью покрывает спектр изобретения для специалиста в области техники.
Если не определено иначе, все технические и научные термины, используемые в настоящем описании, имеют такое же значение, как обычно понимает обычный специалист в области техники, к которой принадлежит настоящее изобретение. Все публикации, патентные заявки, патенты и другие ссылки, упомянутые в настоящем описании, включены полностью в виде ссылки. Применение единственного числа в настоящем описании для описания любого аспекта настоящего изобретения должно интерпретироваться как указывающее на один или более.
Настоящее изобретение относится к способу ингибирования экзоцитозного высвобождения, по меньшей мере, одного медиатора воспаления из, по меньшей мере, одной воспалительной клетки, включающему контакт, по меньшей мере, одной воспалительной клетки, которая содержит, по меньшей мере, один медиатор воспаления, содержащийся в везикулах внутри клетки, с, по меньшей мере, одним пептидом, выбираемым из группы, состоящей из MANS-пептида и его активного фрагмента, в эффективном количестве для снижения высвобождения медиатора воспаления из воспалительной клетки по сравнению с высвобождением медиатора воспаления из воспалительной клетки такого же типа, которое возникает в отсутствие, по меньшей мере, одного пептида.
Настоящее изобретение, кроме того, относится к способу ингибирования высвобождения, по меньшей мере, одного медиатора воспаления из, по меньшей мере, одной воспалительной клетки в ткани или жидкости пациента, включающему введение в ткань и/или жидкость пациента, которые содержат, по меньшей мере, одну воспалительную клетку, содержащую, по меньшей мере, один медиатор воспаления, содержащийся в везикулах в клетке, терапевтически эффективного количества фармацевтической композиции, содержащей, по меньшей мере, один пептид, выбираемый из группы, состоящей из MANS-пептида и его активного фрагмента, в терапевтически эффективном количестве для снижения высвобождения медиатора воспаления из, по меньшей мере, одной воспалительной клетки по сравнению с высвобождением медиатора воспаления из, по меньшей мере, одной воспалительной клетки такого же типа, которое возникает в отсутствие, по меньшей мере, одного пептида. В частности, снижение высвобождения медиатора воспаления включает блокаду или ингибирование механизма, который высвобождает медиатор воспаления из воспалительной клетки.
MANS-пептид, используемый в настоящих способах, описанных выше, включает SEQ ID NO:1. Активный фрагмент, применимый в настоящем изобретении, включает, по меньшей мере, один миристоилированный N-концевой фрагмент SEQ ID NO:1, который содержит, по меньшей мере, шесть аминокислот, где первая аминокислота указанного фрагмента начинается с N-концевого глицина SEQ ID NO:1. В частности, активный фрагмент может быть выбран из группы, состоящей из N-миристоил-GAQFSKTAAKGEAAAERPGEAA (SEQ ID NO:3); N-миристоил-GAQFSKTAAKGEAAAERPGEA (SEQ ID NO:4); N-миристоил-GAQFSKTAAKGEAAAERPGE (SEQ ID NO:5); N-миристоил-GAQFSKTAAKGEAAAERPG (SEQ ID NO:6); N-миристоил-GAQFSKTAAKGEAAAERP (SEQ ID NO:7); N-миристоил-GAQFSKTAAKGEAAAER (SEQ ID NO:8); N-миристоил-GAQFSKTAAKGEAAAE (SEQ ID NO:9); N-миристоил-GAQFSKTAAKGEAAA (SEQ ID NO:10); N-миристоил-GAQFSKTAAKGEAA (SEQ ID NO:11); N-миристоил-GAQFSKTAAKGEA (SEQ ID NO:12); N-миристоил-GAQFSKTAAKGE (SEQ ID NO:13); N-миристоил-GAQFSKTAAKG (SEQ ID NO:14); N-миристоил-GAQFSKTAAK (SEQ ID NO:15); N-миристоил-GAQFSKTAA (SEQ ID NO:16); N-миристоил-GAQFSKTA (SEQ ID NO:17); N-миристоил-GAQFSKT (SEQ ID NO:18); и N-миристоил-GAQFSK (SEQ ID NO:19).
Настоящее изобретение относится к контакту и/или введению пептида, описанного выше и на всем протяжении описания, с любой известной воспалительной клеткой, которая может содержаться в ткани или жидкости пациента, которая содержит по меньшей мере один медиатор воспаления, содержащийся в везикулах внутри клетки. Воспалительной клеткой предпочтительно является лейкоцит, более предпочтительно - гранулоцит, который может быть классифицирован далее как нейтрофил, базофил, эозинофил или их комбинация. Воспалительными клетками, контактирующими в настоящем способе, также могут быть моноцит/макрофаг.
Настоящее изобретение относится к уменьшению высвобождения медиаторов воспаления, содержащихся в везикулах воспалительных клеток, и такие медиаторы воспаления выбирают из группы, состоящей из миелопероксидазы (МРО), эозинофильной пероксидазы (ЕРО), главного основного белка (МВР), лизозима, гранзима, гистамина, протеогликана, протеазы, хемотаксического фактора, цитокина, метаболита арахидоновой кислоты, дефензина, бактерицидного белка, повышающего проницаемость (BPI), эластазы, катепсина G, катепсина В, катепсина D, бета-D-глюкуронидазы, альфа-маннозидазы, фосфолипазы А2, хондроитин-4-сульфата, протеиназы 3, лактоферрина, коллагеназы, активатора комплемента, рецептора комплемента, рецептора N-формилметионил-лейцил-фенилаланина (FMLP), рецептора ламинина, цитохрома b558, моноцитарного хемотаксического фактора, гистаминазы, белка, связывающего витамин В12, желатиназы, активатора плазминогена, бета-D-глюкуронидазы и их комбинации. Предпочтительно такие медиаторы воспаления выбирают из группы, состоящей из миелопероксидазы (МРО), эозинофильной пероксидазы (ЕРО), главного основного белка (МВР), лизозима, гранзима и их комбинации.
Настоящее изобретение приводит в соприкосновение эффективное количество пептида с воспалительной клеткой, где эффективное количество определяют как ингибирующее дегрануляцию количество MANS-пептида или его активного фрагмента, которое снижает количество медиатора воспаления, высвобождаемое из, по меньшей мере, одной воспалительной клетки от около 1% до около 99% по сравнению с количеством, высвобождаемым из, по меньшей мере, одной воспалительной клетки в отсутствие MANS-пептида или его активного фрагмента. Более предпочтительно такое эффективное количество контактирующего пептида включает ингибирующее дегрануляцию количество MANS-пептида или его активного фрагмента, которое снижает количество медиатора воспаления, высвобождаемое из, по меньшей мере, одной воспалительной клетки между около 5-50% до около 99% по сравнению с количеством, высвобождаемым из, по меньшей мере, одной воспалительной клетки в отсутствие MANS-пептида или его активного фрагмента.
Настоящее изобретение в одном варианте осуществления направлено на введение, по меньшей мере, одного пептида, включающего MANS-пептид или его активный фрагмент, в терапевтически эффективном количестве в ткань или жидкость пациента, где пациент страдает от респираторного заболевания, которым предпочтительно является астма, хронический бронхит или ХОБЛ. В следующем варианте осуществления изобретения пациент может быть поражен заболеванием кишечника, заболеванием кожи, аутоиммунным заболеванием, болевым синдромом и их комбинацией. Болезнью кишечника может быть язвенный колит, болезнь Крона или синдром раздраженной кишки. Пациент может страдать от заболевания кожи, такого как розовые угри, экзема, псориаз или тяжелое акне. Пациент также может страдать от артрита, такого как ревматоидный артрит, псориатический артрит, системная красная волчанка. Пациенты, страдающие от муковисцидоза, также могут получать лечение посредством настоящего способа и пептидов. Настоящий способ предпочтительно применим для лечения пациентов, таких как млекопитающие, и предпочтительно людей, собак, лошадей и кошек.
Настоящий способ лечения пациентов представляет собой введение одного или более пептидов, включающих MANS-пептид или его активный фрагмент, описанный в настоящем описании, включая местное введение, парентеральное введение, ректальное введение, легочное введение, назальное введение или пероральное введение. В частности, легочное введение выбирают из группы аэрозольного введения, введения с помощью ингалятора сухого порошка, ингалятора с дозирующим клапаном и небулайзера. Дополнительно, описанный способ может кроме того включать введение пациенту второй молекулы, выбираемой из группы, состоящей из антибиотика, противовирусного соединения, противопаразитарного соединения, противовоспалительного соединения и иммунодепрессанта.
В одном аспекте, изобретение относится к способу введения фармацевтической композиции. Фармацевтическая композиция включает терапевтически эффективное количество известного соединения и фармацевтически приемлемый носитель. «Терапевтически эффективным» количеством, как используется в настоящем описании, является количество соединения, достаточное для облегчения имеющихся у пациента симптомов. Терапевтически эффективное количество варьируется в зависимости от возраста и физического состояния пациента, тяжести состояния пациента, получающего лечение, длительности лечения, природы любого сопутствующего лечения, используемого фармацевтически приемлемого носителя и подобных факторов в рамках знаний и опыта специалиста в данной области техники. Фармацевтически приемлемыми носителями являются предпочтительно твердые лекарственные формы, такие как таблетки или капсулы. Также могут быть использованы жидкие препараты для перорального введения, и могут быть получены в форме сиропов или суспензий, например, растворов, содержащих активный ингредиент, сахар и смесь этанола, воды, глицерина и пропиленгликоля. При желании, такие жидкие препараты могут включать одно или более из следующего: красители, ароматизаторы и сахарин. Кроме того, также могут быть использованы загустители, такие как карбоксиметилцеллюлоза, а также другие приемлемые носители, выбор которых очевиден в области техники.
Как указано выше, настоящее изобретение относится к способам регуляции клеточных секреторных процессов, особенно высвобождения медиаторов воспаления из воспалительных клеток. Как используется в настоящем описании, термин «регуляция» обозначает блокаду, ингибирование, снижение, понижение, увеличение, усиление или стимуляцию. Ряд клеточных секреторных процессов включает высвобождение содержимого из везикул или гранул, связанных с мембраной клеток. Везикулы или гранулы, связанные с мембраной, определяют как внутриклеточные частицы, которые являются преимущественно пузырькообразной (или везикулы внутри клетки) и которые содержат хранящееся вещество, которое может секретироваться. Было обнаружено, что некоторое из содержимого таких везикул, таких как содержащихся в воспалительных клетках, отвечает за множество патологических процессов во множестве тканей млекопитающих. Некоторые эффекты такой секреции видимо включают повреждение ранее здоровых тканей при воспалении. Настоящее изобретение обеспечивает средство блокады секреции из любых везикул, связанных с мембраной, включая таковые, обнаруживаемые в воспалительных клетках, путем нацеливания синтетического пептида на специфическую молекулу, важную во внутриклеточном секреторном пути. Такой подход может иметь терапевтическую пользу для лечения широкого множества гиперсекреторных и воспалительных состояний у людей и животных.
В частности, настоящее изобретение имеет мишенью воспалительные клетки, которые содержат медиаторы воспаления в одной или более гранул или везикул в цитоплазме клеток. Клетки контактируют с одним или более пептидами, которые выбирают из MANS-пептида или его активного фрагмента, все из которых ниже описаны детально. Предпочтительно контакт пептида с воспалительной клеткой происходит посредством введения пациенту, пораженному или страдающему от заболевания, при котором такие воспалительные клетки присутствуют в специфической ткани или жидкости в ткани. При введении или контакте пептида с клеткой пептид конкурентно конкурирует за и конкурентно ингибирует связывание естественного MARCKS-белка с мембраной внутриклеточных гранул или везикул, которые содержат медиаторы воспаления. В результате блокады связывания MARCKS-белка с везикулами в воспалительных клетках, такие везикулы в таких клетках не передвигаются к плазматической мембране клетки, как они должны в норме при стимуляции для экзоцитозного высвобождения из клетки содержащихся в них медиаторов воспаления. Следовательно, способ по настоящему изобретению ингибирует движение везикул к плазматической мембране клетки, что в свою очередь снижает высвобождение медиаторов воспаления из воспалительных клеток. Количество медиаторов воспаления, высвобождаемых из клеток, с течением времени снижается, так как и скорость высвобождения, и количество высвобождаемых медиаторов из воспалительных клеток зависит от концентрации вводимого и контактирующего с воспалительными клетками пептида.
Одним преимуществом настоящего изобретения является то, что оно может комбинировать терапию, которая включает непосредственную блокаду секреции слизи с уникальной противовоспалительной терапией. Преимуществом настоящего изобретения над существующим противовоспалительным лечением, которое вызывает общее подавление иммунной системы, является то, что, вероятно, пептид блокирует секрецию только внутриклеточных компонентов, секретируемых из воспалительных клеток. Следовательно, множество аспектов иммунной системы все еще функционируют даже при ингибировании медиаторов воспаления.
Соединения по настоящему изобретению могут регулировать, т.е. блокировать, высвобождение медиатора воспаления из клеток. Такое ингибирование высвобождения медиаторов воспаления является привлекательным свойством для предотвращения и лечения множества расстройств, например болезней и патологических состояний, включающих воспаление. Следовательно, соединения по изобретению могут быть применимы для лечения таких состояний. Они включают заболевания дыхательных путей и хронические воспалительные заболевания, включая, но, не ограничиваясь, остеоартрит, рассеянный склероз, синдром Гийена-Барре, болезнь Крона, язвенный колит, псориаз, болезнь трансплантат-против-хозяина, системную красную волчанку и инсулинозависимый сахарный диабет. Соединения по изобретению также могут быть использованы для лечения других расстройств, ассоциированных с активностью повышенного уровня провоспалительных медиаторов и ферментов, таких как ответ на различные инфекционные агенты и ряд заболеваний, таких как ревматоидный артрит, синдром токсического шока и воспалительные заболевания кишечника.
Применение пептидов и способы по изобретению включают терапию для борьбы с воспалением вместе с терапией, которая сочетает противовоспалительную активность пептида со способностью блокировать секрецию слизи. Заболевания, которые можно лечить посредством способности блокировать и воспаление, и секрецию слизи, включают, но не ограничиваются, воспалительные заболевания кишечника, расстройства пищеварения (воспаление желчного пузыря, болезнь Менетрие) и воспалительные заболевания дыхательных путей. Пептид также может быть использован для блокады высвобождения избытка инсулина из островковых клеток поджелудочной железы.
Другие провоспалительные медиаторы коррелируют со множеством патологических состояний, которые связаны с притоком нейтрофилов в места воспаления или повреждения. Было продемонстрировано, что блокирующие антитела являются применимым лечением повреждения ткани, ассоциированного с нейтрофилами при остром воспалении (Haradr et al., 1996, Molecular Medicine Today 2, 482). Клетки, иные чем нейтрофилы, которые могут высвобождать медиаторы воспаления, включают другие лейкоциты, такие как базофилы, эозинофилы, моноциты и лимфоциты, и лечение может быть направлено на секрецию из таких клеток. Нейтрофилы, эозинофилы и базофилы, каждый, являются типом гранулоцитов, т.е. лейкоцитов, которые имеют гранулы в цитоплазме. Лейкоциты синтезируют ряд медиаторов воспаления, которые упаковываются и хранятся в цитоплазматических гранулах. Среди таких медиаторов находятся, например, миелопероксидаза [МРО] в нейтрофилах (Borregaard N, Cowland JB. Granules of the human neitrophilic polymorphonuclear leukocyte. Blood 1997; 89: 3503-3521), эозинофильная пероксидаза [ЕРО] и главный основной белок [МВР] в эозинофилах (Gleich G.I. Mechanisms of eosinophil-associated inflammation. J Allergy Clin Immunol 2000; 105:651-663), лизозим в моноцитах/макрофагах (Hoff T., Spencker T., Emmendoerffer A., Goppelt-Struebe M. Effects of glucocorticoids on the TPA-induced monocytic differentiation. J. Leukoc Biol 1992; 52: 173-182; Balboa M A, Saez Y, Balsinde J.Calcium-independent phospholipase A2 is required for lysozyme secretion in U937 promonocytes. J. Immunol 2003; 170: 5276-5280) и гранзим в NK-клетках и цитотоксических лимфоцитах (Bochan MR, Goebel WS, Brahmi Z. Stably transfected antisense granzyme B and perforin constructs inhibit human granule-mediated lytic ability. Cell Immunol 1995; 164: 234-239; Gong J H., Maki G, Кlingemann HG. Characterization of a human cell line (NK-92) with phenotypical and functional characteristics of activated natural killer cells. Leukemia 1994; 8:652-658; Maki G., Klingemann HG, Martinson JA, Tam YK. Factors regulating the cytotoxic activity of the human natural killer cell line, NK-92. J. Hematother Stem Cell Res 2001; 10: 369-383; и Takayama H, Trenn G, Sitkovsky MV. A novel cytotoxic T lymphocyte activation assay. J Immunol Methods 1987; 104:183-1907-10). Такие медиаторы могут высвобождаться в месте повреждения и могут вносить вклад в воспаление и заживление, например, в легких и в других местах, в результате инфильтрации такими клетками поврежденной или больной ткани. Лейкоциты высвобождают такие гранулы посредством механизма экзоцитора (Burgoyne RD, Morgan A. Secretory granule exocytosis. Physiol Rev 2003; 83: 581-632; Logan MR, Odemuyiwa SO, Moqbel R. Understanding exocytosis in immune and inflammatory cells: the molecular basis of mediator secretion. J Allergy Clin Immunol 2003; 111:923-932).
Тучные клетки, которые обычно не циркулируют в кровотоке, и базофилы содержат секреторные цитоплазматические гранулы, которые хранят и могут высвобождать при активации клетки подготовленные воспалительные (анафилактические) медиаторы, такие как гистамин; протеогликаны, такие как гепарин и хондроитинсульфат; протеазы, такие как триптаза, химаза, карбоксипептидаза и катепсин G-подобная протеаза; хемотактические факторы, цитокины и метаболиты арахидоновой кислоты, которые действуют на сосудистое русло, гладкую мускулатуру, соединительную ткань, слизистые железы и воспалительные клетки.
Нейтрофилы, также известные как полиморфоядерные лейкоциты (PMN), составляют 50-60% всех циркулирующих лейкоцитов. Нейтрофилы действуют против инфекционных агентов, таких как бактерии, грибы, простейшие, вирусы, клетки, инфицированные вирусами, а также опухолевые клетки, которые проникают через физические барьеры организма в местах инфекции или повреждения. Нейтрофилы проходят через шесть морфологических стадий: миелобласт, промиелобласт, миелоцит, метамиелоцит, не-сегментированный (ленточный) нейтрофил и сегментированный (функционально активный) нейтрофил.
В нейтрофилах, воспалительные медиаторы хранятся в первичных (азурофильных), вторичных (специфических) и третичных (желатиназных) гранулах, а также в секреторных везикулах. Среди множества медиаторов воспаления первичные (азурофильные) гранулы содержат миелопероксидазу (МРО), лизозим, дефензины, бактерицидный белок, повышающий проницаемость (BPI), эластазу, катепсин G, катепсин В, катепсин D, бета-D-глюкуронидазу, альфа-маннозидазу, фосфолипазу А2, хондроитин-4-сульфат и протеиназу 3 (см., например, Hartwig JH, Thelen M, Rosen A, Janmey PA, Nairn AC, Aderem A. MARCKS is an actin filament crosslinking protein regulated by protein kinase C and calcium-calmodulin. Nature 1992; 356: 618-622); вторичные (специфические) гранулы содержат лизозим, лактоферрин, коллагеназу, активатор комплемента, фосфолипазу А2, рецепторы комплемента, например, CR3, CR4, рецепторы N-формилметионил-лейцил-фенилаланина (FMLP), рецепторы ламинина, цитохром b558, моноцитарный-хемотактический фактор, гистаминазу и белок, связывающий витамин В12; и мелкие гранулы хранения содержат желатиназу, активатор плазминогена, катепсин В, катепсин D, бета-D-глюкуронидазу, альфа-маннозидазу и цитохром b558.
Гранулы нейтрофилов содержат противомикробные или цитотоксические вещества, нейтральные протеиназы, кислые гидролазы и депо цитоплазматических мембранных рецепторов. Среди составляющих азурофильных гранул миелопероксидаза (МРО) является критическим ферментом в преобразовании перекиси водорода в хлористоводородную кислоту. Вместе с перекисью водорода и галоидным кофактором она образует эффективный микробный и цитотоксический механизм лейкоцитов - миелопероксидазную систему.
Дефензины, которые составляют от 30 до 50% белка азурофильных гранул, представляют собой малые (молекулярная масса <4000) сильные противомикробные пептиды, которые являются цитотоксичными для широкого диапазона бактерий, грибов и некоторых вирусов. Их токсичность может осуществляться за счет повышения проницаемости мембран клеток-мишеней, что подобно другим белкам, образующим каналы (перфорины).
Белок, увеличивающий проницаемость бактерий (BPI) является членом семейства перфоринов. Он высокотоксичен в отношении грамотрицательных бактерий, но не грамположительных бактерий или грибов, и также может нейтрализовать эндотоксин, токсический липополисахаридный компонент клеточной оболочки грамотрицательных бактерий.
Лактоферрин отщепляет свободное железо, таким образом предотвращая рост потребленных микроорганизмов, которые переживают процесс умерщвления, и увеличивает проницаемость бактерий для лизозима.
Сериновые протеазы, такие как эластаза и катепсин G, гидролизуют белки в оболочке бактериальной клетки. Субстраты эластазы гранулоцитов включают коллагеновые сшивки и протеогликаны, а также эластиновые компоненты кровеносных сосудов, связок и хрящей. Катепсин D отщепляет протеогликаны хряща, тогда как коллагеназы гранулоцитов являются активными в отщеплении коллагена типа I и, в меньшей степени, типа III из кости, хряща и сухожилий. Продукты распада коллагена обладают хемотаксической активностью для нейтрофилов, моноцитов и фибробластов.
Регуляция разрушения тканей посредством лизосомальных протеаз опосредуется ингибиторами протеаз, такими как альфа2-макроглобулин и альфа1-антипротеаза. Такие антипротеазы присутствуют в сыворотке и синовиальной жидкости. Они могут функционировать путем связывания с и покрытия активных сайтов протеаз. Дисбаланс протеаз-антипротеаз может быть важным в патогенезе эмфиземы.
Азурофильные гранулы действуют преимущественно в внутриклеточной среде (в фаголизосомальной вакуоли), где они вовлечены в умерщвление и разрушение микроорганизмов. Нейтрофильные специфические гранулы чувствительны к высвобождению внеклеточно их содержимого и играют важную роль в развитии воспаления. Специфические гранулы представляют собой внутриклеточный резервуар различных компонентов плазматической мембраны, включая цитохром b (компонент НАДФН оксидазы, фермента, ответственного за продукцию супероксида), рецепторы фрагментов комплемента iC3b (CR3, CR4), ламинина и формилметионилпептидные хемоаттрактанты. В добавление к остальным, существует гистаминаза, которая важна для разрушения гистамина, белок, связывающий витамины, и активатор плазминогена, который отвечает за образование плазмина и отщепление С5а от С5.
Важность гранул нейтрофилов в воспалении очевидна из исследований нескольких пациентов с врожденными изменениями гранул. Пациенты с синдромом Чедиака-Хигаси имеют серьезные нарушения скорости развития воспалительного ответа и ненормально крупные лизосомальные гранулы. Врожденный синдром дефицита специфических гранул является чрезвычайно редким расстройством, характеризуемым сниженным воспалительным ответом и тяжелыми бактериальными инфекциями кожи и подлежащих тканей.
Хотя механизмы, регулирующие экзоцитозную секрецию таких гранул, известны только частично, в процессе были идентифицированы несколько ключевых молекул, включая внутриклеточные переносчики Са2+ (Richter et al. Proc Natl Acad Sci USA 1990; 87:9472-9476; Blackwood et al., Biochem J 1990; 266:195-200), G белки, тирозин и протеинкиназы (РК, особенно, РКС) (Smolen et al., Biochim Biophys Acta 1990; 1052: 133-142; Niessen et al., Biochim. Biophys. Acta 1994; 1223: 267-273; Naucler et al., Pettersen et al., Chest 2002; 121; 142-150), Rac2 (Abdel-Latif et al., Blood 2004; 104: 832-839; Lacy et al., J Immunol 2003; 170: 2670-2679) и различные SNARE, SNAP и VAMP (Sollner et al., Nature 1993; 362: 318-324; Lacy, Pharmacol Ther 2005; 107: 358-376).
Белки SNARE (растворимый рецептор белка прикрепления N-этилмалеимида) представляют собой семейство белков, ассоциированных с мембраной, характеризующихся альфа-спиральным биспиральным доменом, называемым SNARE-мотив (Li et al., Cell. Mol. Life Sci. 60: 942-960 (2003)). Такие белки классифицируют как v-SNARE и t-SNARE на основании их расположения на везикуле или мембране мишени; другая схема классификации определяет R-SNARE и Q-SNARE на основании консервативного остатка аргинина или глутамина в центре SNARE-мотива. SNARE располагаются на определенных местах мембраны секреторных и экзоцитозных путей и вносят вклад в специфичность процессов сшивки внутриклеточной мембраны. T-SNARE домен состоит из 4-спирального узла с биспиральным изгибом. SNARE-мотив вносит вклад в сшивку двух мембран. SNARE-мотивы делятся на четыре класса: гомологи синтаксина 1а (t-SNARE), VAМP-2 (v-SNARE) и N- и С-концевые SNARE-мотивы SNAP-25. Один член из каждого класса может взаимодействовать с образованием SNARE-комплекса. SNARE-мотив обнаруживают в N-концевом домене определенных членов семейства синтаксина, таких как синтаксин 1а, который требуется для высвобождения нейромедиаторов (Lerman et al., Biochemistry 39: 8470-8479 (2000)), и синтаксин 6, который обнаруживают в эндосомальном транспорте везикул (Misura et al., Proc. Natl. Acad. Sci.U.S.A. 99: 9184-9189 (2002)).
Белки SNAP-25 (белок, ассоциированный с синаптосомами 25 кДа) являются компонентами SNARE-комплексов, которые могут отвечать за специфичность мембранной сшивки и за непосредственное выполнение сшивки путем образования тесного комплекса (SNARE или стержневого комплекса), который соединяет синаптические везикулы и плазматические мембраны. SNARE составляют большое семейство белков, которые характеризуются последовательностью из 60 остатков, известной как SNARE-мотив, который обладает выраженной предрасположенностью к образованию биспиралей и часто предшествует карбокси-концевым трансмембранным участкам. Синаптический стержневой комплекс образуется из четырех SNARE-мотивов (два из SNAP-25 и по одному из синаптобревина и синтаксина 1), которые являются бесструктурными при выделении, но образуют параллельный узел из четырех спиралей при соединении. Кристаллическая структура стержневого комплекса выявила, что спиральный узел является сильно скрученным и содержит несколько солевых мостиков на поверхности, а также слои внутренних гидрофобных остатков. Полярный слой в центре комплекса образуется тремя глютаминами (два из SNAP-25 и один из синтаксина 1) и одним аргинином (из синаптобревина) (Rizo et al., Nat Rev Neurosci 3: 641-653 (2002)). Члены семейства SNAP-25 содержат кластер цистеиновых остатков, которые могут быть пальмитоированы для прикрепления к мембране (Risinger et al., J.Biol.Chem. 268: 24408-24414 (1993)).
Основной ролью нейтрофилов является фагоцитоз и разрушение инфекционных агентов. Также они ограничивают рост некоторых микроорганизмов до начала адаптивных (специфических) иммунологических ответов. Хотя нейтрофилы необходимы для защиты хозяина, они также вовлечены в патогенез множества хронических воспалительных состояний и в ишемически-реперфузионное повреждение. Гидролитические ферменты, происходящие из нейтрофилов, и окислительно инактивированные ингибиторы протеаз могут быть определены в жидкости, выделенной из участка воспаления. При обычных условиях нейтрофилы могут мигрировать в места инфекции без повреждения тканей организма-хозяина. Однако иногда может возникать нежелательное повреждение тканей организма-хозяина. Это повреждение может возникать посредством нескольких независимых механизмов. Они включают преждевременную активацию во время миграции, внеклеточное высвобождение токсических продуктов во время умерщвления некоторых микроорганизмов, удаление инфицированных или поврежденных клеток организма-хозяина и инородных веществ в качестве первой стадии ремоделирования ткани или невозможность терминировать острые воспалительные ответы. Ишемически-реперфузионное повреждение ассоциировано с притоком нейтрофилов в пораженную ткань и последующей активацией. Это может запускаться веществами, высвобождаемыми из поврежденных клеток организма-хозяина, или как последствие образования супероксида через ксантиноксидазу.
При нормальных условиях кровь может содержать смесь нормальных, примированных, активированных и использованных нейтрофилов. В месте воспаления, присутствуют преимущественно активированные и использованные нейтрофилы. Активированные нейтрофилы обладают усиленной продукцией реакционных промежуточных форм кислорода (ROI). Субпопуляцию нейтрофилов с усиленным дыхательным взрывом определяют в крови людей с острой бактериальной инфекцией и пациентов с респираторным дистресс синдромом взрослых (ARDS). Это является примером нейтрофильного парадокса. Нейтрофилы вовлечены в патогенез такого состояния из-за значительного тока таких клеток в легкие и ассоциированного повреждения тканей, вызванного окислителями и гидролитическими ферментами, высвобождаемыми из активированных нейтрофилов. Нарушение противомикробной активности нейтрофилов, которое возникает при ухудшении ARDS, может быть защитным ответом части организма-хозяина, что индуцируется местно продуктами воспаления.
Острая фаза термического повреждения также ассоциирована с активацией нейтрофилов, и за этим следует общее нарушение различных функций нейтрофилов. Активация нейтрофилов иммунными комплексами в синовиальной жидкости участвует в патогенезе ревматоидного артрита. Хроническая активация нейтрофилов может также способствовать развитию опухолей, так как некоторое количество ROI, образующееся нейтрофилами, повреждает ДНК и протеазы запускают миграцию опухолевых клеток. У пациентов, страдающих от тяжелых ожогов, была установлена корреляция между развитием бактериальной инфекции и снижением доли и абсолютного количества нейтрофилов, положительных в отношении антител и рецепторов комплемента. Также было показано, что окислители, происходящие из нейтрофилов, окисляют липопротеины низкой плотности (ЛНП), которые затем более эффективно связываются с плазматической мембраной макрофагов посредством специфических фагоцитарных рецепторов. Потребление таких окисленных ЛНП макрофагами может способствовать развитию атеросклероза. Кроме того, примированные нейтрофилы были обнаружены у людей с эссенциальной гипертензией, болезнью Ходжкина, воспалительными заболеваниями кишечника, псориазом, саркоидозом и септицемией, где примирование коррелирует с высокой концентрацией циркулирующего TNF-альфа (кахектин).
Гидролитическое повреждение тканей организма-хозяина и, следовательно, хронические воспалительные состояния могут возникать, когда антиоксидантные и антипротеазные сита перегружены. Считают, что антипротеазный дефицит отвечает за патогенез эмфиземы. Множество антипротеаз являются членами семейства ингибитора сериновых протеаз (SERPIN). Хотя циркуляция богата антипротеазами, такие крупные белки могут быть селективно не допущены в места воспаления, т.к. нейтрофилы прилипают к своим мишеням. Окислительный стресс может запускать повреждение ткани посредством снижения концентрации внеклеточных антипротеаз ниже уровня, требуемого для ингибирования высвобожденных протеаз. Хлорированные окислители и перекись водорода могут инактивировать антипротеазы, такие как ингибитор альфа1-протеазы и альфа2-макроглобулин, которые являются эндогенными ингибиторами эластазы, а одновременно активировать латентные металлопротеазы, такие как коллагеназы и желатиназа, что вносит вклад в последующую инактивацию антипротеаз.
Цитоплазматические составляющие нейтрофилов могут также быть причиной образования специфических антинейтрофильных цитоплазматических антител (ANCA), которые тесно связаны с развитием системного васкулита и гломерулонефрита. ANCA представляют собой антитела, направленные против ферментов, которые обнаруживаются преимущественно в азурофильных или первичных гранулах нейтрофилов. Существует три типа ANCA, которые могут быть различены по структуре, которую они дают при непрямой иммунофлуоресценции на нормальных нейтрофилах, фиксированных этанолом. Диффузная мелкозернистая цитоплазматическая флуоресценция (cANCA) обычно обнаруживается при гранулематозе Вегенера, в некоторых случаях микроскопического полиартериита и синдроме Чург-Штраусса, и в некоторых случаях полулунного или сегментарного некротизирующего гломерулонефрита. Антигеном-мишенью обычно является протеиназа 3. Перинуклеарная флуоресценция (pANCA) обнаруживается во множестве случаев микроскопического полиартериита и гломерулонефрита. Такие антитела обычно направлены против миелопероксидазы, но другие мишени включают эластазу, катепсин G, лактоферрин, лизозим и бета-D-глюкуронидазу. Третья группа, обозначенная «атипичной» ANCA, включает флуоресценцию ядер нейтрофилов и некоторые необычные цитоплазматические рисунки, и тогда как некоторые антигены мишени определяются pANCA, другие еще не идентифицированы. pANCA также обнаруживают у трети пациентов с болезнью Крона. Сообщаемая частота ANCA при ревматоидном артрите и SLE значительно варьируется, но структурой являются преимущественно pANCA и атипичная ANCA.
Эозинофил является окончательно дифференцированным лейкоцитом конечной стадии, который находится преимущественно в подслизистой ткани и привлекается в места специфических иммунных реакций, включая аллергические заболевания. Цитоплазма эозинофилов содержит крупные эллипсоидные гранулы с электронно-плотным кристаллическим ядром и частично проницаемым матриксом. В добавление к таким первичным кристаллоидным гранулам, существует другой тип гранул, которые меньше (мелкие гранулы) и не имеют кристаллического ядра. Крупные специфические гранулы эозинофилов содержат, по меньшей мере, четыре отдельных катионных белка, которые оказывают ряд биологических эффектов на клетки организма-хозяина и микробные мишени: главный основной белок (МВР), эозинофильный катионный белок (ЕСР), нейтротоксин эозинофилов (EDN) и эозинофильная пероксидаза (ЕРО). Базофилы содержат около одной четвертой содержания главного основного белка эозинофилов вместе с определяемыми количествами EDN, ЕСР и ЕРО. Небольшие количества EDN и ЕСР также обнаруживают в нейтрофилах (Gleich GJ. Mechanisms of eosinophil-associated inflammation. J Allergy Clin Immunol 2000; 105: 651-663). Вероятно МВР не обладает ферментной активностью, но является высококатионным полипептидом, который может проявлять свои токсические свойства путем взаимодействия с липидными мембранами, приводя к их дисфункции. И МВР и ЕРО могут действовать как селективные аллостерические ингибиторы агонистического связывания с М2 мускариновыми рецепторами. Такие белки могут участвовать в дисфункции М2 рецептора и усиливать опосредованную вагусом бронхоконстрикцию при астме. EDN может специфически повреждать миелиновую оболочку нейронов. Гистаминаза и множество гидролитических лизосомальных ферментов также присутствуют в крупных специфических гранулах эозинофилов. Среди ферментов в мелких гранулах эозинофилов представлены арилсульфатаза, кислая фосфатаза и 92 кДа металлопротеиназа, желатиназа. Эозинофилы могут производить цитокины, которые включают таковые с сильной аутокринной активностью эозинофильных факторов роста и таковые с возможным участием в остром и хроническом воспалительных ответах. Три цитокина обладают активностью эозинофильных факторов роста: гранулоцитарно-макрофагальный колониестимулирующий фактор (GM-CSF), IL-3 и IL-5. Другие цитокины, продуцируемые человеческими эозинофилами, которые могут обладать активностью при остром и хроническом воспалительных ответах, включают IL-1-альфа, IL-6, IL-8, TNF-альфа и оба трансформирующих фактора роста TGF-альфа и TGF-бета.
Эозинофилы содержат кристаллоидные гранулы, которые содержат МВР, эозинофильный катионный белок, ЕРО и эозинофильный нейротоксин (Gleich, J Allergy Clin Immunol 2000; 105:651-663). Клон 15 человеческой промиелоцитарной клеточной линии HL-60 может быть использован для изучения секреции ЕРО. Такая клеточная линия была получена из клона HL-60, которых выращивали при повышенном рН в течение двух месяцев (Fischkoff, Leuk Res 1988; 12: 679-686) и затем обрабатывали масляной кислотой для возможности клеток дифференцироваться с проявлением множества признаков эозинофилов периферической крови, включая экспрессию белков эозинофильных специфических гранул (Rosenberg et al., J Exp Med 1989; 170: 163-176; Tiffany et al., J Leukoc Biol 1995; 58:49-54; Badewa et al., Exp Biol Med 2002; 227: 645-651).
Эозинофилы могут принимать участие в реакциях гиперчувствительности, особенно посредством двух липидных медиаторов воспаления, лейкотриена С4 (LTC4) и фактора, активирующего тромбоциты (PAF). Оба медиатора сокращают гладкие мышцы дыхательных путей, запускают секрецию слизи, изменяют проницаемость сосудов и вызывают инфильтрацию эозинофилов и нейтрофилов. В добавление к непосредственному действию таких медиаторов из эозинофилов, МВР может стимулировать высвобождение гистамина из базофилов и тучных клеток, и МВР может стимулировать высвобождение ЕРО из тучных клеток. Эозинофилы могут служить местным источником специфических липидных медиаторов, а также индуцировать высвобождение медиаторов из тучных клеток и базофилов. Содержимое гранул эозинофилов высвобождается после стимула, сходного с таковым для гранул нейтрофилов, например, во время фагоцитоза опсонизированных частиц и хемотаксических факторов. Лизосомальные ферменты нейтрофилов действуют преимущественно на вещество, заключенное в фаголизосомах, тогда как содержимое гранул эозинофилов действует главным образом на внеклеточные структуры-мишени, такие как паразиты и медиаторы воспаления.
Развитие моноцитов и макрофагов имеет место в костном мозге и проходит через следующие стадии: стволовая клетка; заинтересованная стволовая клетка; монобласт; промоноцит; моноцит в костном мозге; моноцит в периферической крови; и макрофаг в тканях. Дифференцировка моноцитов в костном мозге происходит быстро (1,5-3 дня). Во время дифференцировки гранулы образуются в цитоплазме моноцитов, и они могут быть разделены, как в нейтрофилах, на, по меньшей мере, два типа. Однако их меньше по количеству и по размеру, чем их аналогов в нейтрофилах (азурофильные и специфические гранулы). Содержание в них ферментов является сходным.
Ферменты, связанные с гранулами моноцитов/макрофагов, включают лизозим, кислую фосфатазу и бета-глюкуронидазу. В качестве модели для in vivo исследований использовали секрецию лизозима из клеток U937. Такую клеточную линию получали из человеческой гистиоцитарной лимфомы и использовали в качестве клеточной линии моноцитов, которая может быть активирована множеством агонистов, такими как РМА (Hoff et al., J Leukoc Biol 1992; 52: 173-182; Balboa et al., J Immunol 2003; 170:5276-5280; Sundstrom et al., Int J Cancer 1976; 17: 565-577).
Естественные клетки-киллеры (NK-клетки) и цитотоксические лимфоциты содержат сильные цитотоксические гранулы, включающие перфорин, белок, образующий поры, и гранзим, лимфоцит-специфические сериновые протеазы. Например, клеточная линия NK-92 представляет собой IL-2-зависимую человеческую линию, полученную от пациента с быстропрогрессирующей не-Ходжкинской лимфомой (Gong JH., Maki G, Klingemann HG. Characterization of a human cell line (NK-92) with phenotypical and functional characteristics of activated natural killer cells. Leukemia 1994; 8: 652-658). NK-92 клетки экспрессируют высокий уровень молекул, вовлеченных в перфорин-гранзим цитолитический путь, который имеет мишенью широкий диапазон злокачественных клеток (Gong et al, см. ниже, и Maki G, Klingemann HG, Martinson JA, Tam YK. Factors regulating the cytotoxic activity of the human natural killer cell line, NK-92. J Hematother Stem Cell Res 2001; 10: 369-383).
Гранзимы являются экзогенными сериновыми протеазами, которые высвобождаются цитоплазматическими гранулами в цитотоксических Т-клетках и NK-клетках. Гранзимы могут вызывать апоптоз в клетках, инфицированных вирусами, таким образом разрушая их.
Внеклеточное высвобождение медиатора воспаления (воспалительного медиатора) из гранулоцита (или лейкоцита) и внеклеточное высвобождение более чем одного медиатора воспаления (воспалительного медиатора) из гранулоцита (или лейкоцита) иногда в настоящем описании называют дегрануляцией. В предпочтительном варианте осуществления изобретения высвобождение медиатора воспаления включает высвобождение указанного медиатора из гранулы, расположенной внутри гранулоцита или лейкоцита. Высвобождение медиатора воспаления представляет собой предпочтительно высвобождение медиатора воспаления из таких гранул.
Нейтрофилы и макрофаги при примировании провоспалительными агентами (стимуляторами воспаления), такими как TNFα, в значительной степени увеличивают синтез MARCKS-белка: до 90% нового белка, образующегося нейтрофилами в ответ на или TNFα, или липополисахарид (LPS), представляет собой MARCKS (Thelen M, Rosen A, Nairn AC, Aderem A. Tumor necrosis factor aloha modifies agonist-dependent responses in human neutrophils by inducing the synthesis and myristoylation of a specific protein kinase C substrate. Proc Natl Acad Sci USA 1990; 87: 5603-5607). Следовательно, MARCKS может играть важную роль в последующем высвобождении медиаторов воспаления, когда клетки, содержащие гранулы, такие как нейтрофилы и макрофаги, стимулируются агонистами, особенно такими, которые действуют путем активации РКС (Burgoyne et al., Physiol Rev 2003; 83:581-632; Logan et al. J Allergy Clin Immunol 2003; 111: 923-932; Smolen et al., Biochim Biophys Acta 1990; 1052: 133-142; Niessen et al., Biochim. Biophys Acta 1994; 1223: 267-273; Naucler et al., J Leukoc Biol 2002; 71: 701-710).
В одном аспекте настоящего изобретения введение ингибирующего дегрануляцию количества MANS-белка или его активного фрагмента, как описано в настоящем описании, в место воспаления у пациента, которое возникает в результате начала развития болезни, состояния, травмы, инородного тела или их комбинации в месте воспаления у пациента, может снижать количество медиатора воспаления, высвобождаемого из инфильтрирующих лейкоцитов в месте воспаления, где лейкоциты представляют собой преимущественно гранулоциты. Введение MANS-пептида и/или, по меньшей мере, одного его активного фрагмента может снижать количество медиатора воспаления, высвобождаемого из лейкоцитов, таких как гранулоциты, инфильтрирующих место воспаления. Ингибирующее дегрануляцию количество MANS-пептида или ингибирующее дегрануляцию количество его активного фрагмента достаточно для снижения или ингибирования экзоцитозного высвобождения медиаторов воспаления из гранул, содержащихся в воспалительных клетках, инфильтрирующих участок. Эффективность ингибирования дегрануляции измеряют в момент времени после введения MANS-пептида или его фрагмента путем сравнения процента ингибирования (т.е. процента снижения) высвобождения медиаторов воспаления из указанных клеток (лейкоцитов или гранулоцитов, или других воспалительных клеток) относительно уровня или количества, или концентрации указанных медиаторов воспаления, высвобождаемых или продуцируемых приблизительно за то же время в отсутствие MANS-пептида и/или в отсутствие его активного фрагмента. Кроме того, опытный специалист может определить, уменьшается ли воспаление в ткани путем оценки симптомов и параметров воспаления, известных как индикаторы заболевания, для определения вводится ли достаточное или терапевтически эффективное количество MANS-пептида и/или его активного фрагмента. Достаточное ингибирующее дегрануляцию количество представляет собой процент снижения медиатора воспаления, высвобождаемого из гранулоцита в месте воспаления, составляющий от около 1% до около 99%, предпочтительно от 5% до около 99%, более предпочтительно от около 10% до около 99%, еще более предпочтительно от около 25% до 99% и еще более предпочтительно от около 50% до около 99% количества указанного медиатора воспаления, высвобождаемого из указанного гранулоцита в отсутствие MANS-пептида или его активного фрагмента, исследуемое в таких же условиях.
В одном аспекте настоящего изобретения введение ингибирующего дегрануляцию количества MANS-пептида в место развития воспаления у животного, которое создается введением стимулирующего воспаление количества стимулятора воспаления в указанное место, может снижать количество медиатора воспаления, высвобождаемое из гранулоцита, который стимулируется указанным стимулятором воспаления в указанном месте стимуляции воспаления, от около 1% до около 99%, предпочтительно от 5% до около 99%, более предпочтительно от около 10% до около 99%, еще более предпочтительно от около 25% до около 99%, и еще более предпочтительно от около 50% до около 99% количества указанного медиатора воспаления, высвобождаемого из указанного гранулоцита в отсутствие MANS-пептида в присутствии такого же стимулирующего воспаление количества указанного стимулятора воспаления.
В другом аспекте настоящего изобретения введение ингибирующего дегрануляцию количества MANS-пептида в место воспалительной стимуляции у животного, которое было получено введением стимулирующего воспаление количества стимулятора воспаления в указанное место, может снизить количество медиатора воспаления, высвобождаемое из гранулоцита, который стимулируется указанным стимулятором воспаления в указанном месте воспалительной стимуляции, на 100% количества указанного медиатора воспаления, высвобождаемого из указанного гранулоцита в отсутствии MANS-пептида в присутствии такого же стимулирующего воспаление количества указанного стимулятора воспаления.
Примером стимулятора воспаления, используемого в примерах in vitro в настоящем описании, является 12-миристат 13-ацетат форбола (РМА). Моноцитарный белок-хемоаттрактант (МСР-1) является почти таким же эффективным как С5а, и гораздо более эффективным чем IL-8 для дегрануляции базофилов, приводя к высвобождению гистамина. Высвобождение гистамина может возникать после стимуляции хемокинами (т.е. хемоаттрактантными цитокинами), RANTES и MIP-1.
В предпочтительном варианте осуществления изобретения относительно базальной концентрации MARCKS-пептида, присутствующей в месте стимуляции воспаления, ингибирующее дегрануляцию количество MANS-пептида, вводимое в место стимуляции воспаления у животного, составляет от около 1 до около 1000000 концентраций MARCKS-пептида в указанном месте стимуляции воспаления, предпочтительно от около 1 раза до около 100000 раз больше концентрации MARCKS-пептида в указанном месте стимуляции воспаления, более предпочтительно от около 1 раза до около 10000 раз концентрации MARCKS-пептида в указанном месте стимуляции воспаления, еще более предпочтительно от около 1 раза до около 1000 раз концентрации MARCKS-пептида в указанном месте стимуляции воспаления, еще более предпочтительно от около 1 раза до около 100 раз концентрации MARCKS-пептида в указанном месте стимуляции воспаления, и даже более предпочтительно от около 1 раза до около 10 раз концентрации MARCKS-пептида в указанном месте стимуляции воспаления.
В предпочтительном варианте осуществления изобретения гранулоциты находятся на или в дыхательных путях животного, предпочтительно человека, MANS-пептид вводят путем ингаляции, например, путем ингаляции фармацевтической композиции, включающей MANS-пептид, например, фармацевтической композиции включающей MANS-пептид и водный раствор, такую композицию вводят в форме аэрозоля, или фармацевтической композиции, содержащей MANS-пептид в форме сухого порошка, такую композицию вводят с использованием ингалятора сухого порошка. Могут быть применимы другие способы и устройства, известные в области техники для введения раствора или порошка путем ингаляции, такие как, например, капли, спреи и небулайзеры.
В некоторых вариантах осуществления изобретения возможно, чтобы пептид по настоящему изобретению блокировал секреторные процессы, которые являются физиологически важными, включая базальные секреторные функции. Хотя авторы не хотят быть связанными с какой-либо определенной теорией изобретения, считают, что механизмы, регулирующие такую базальную секрецию, отличаются от таковых, регулирующих стимулированную секрецию. Альтернативно, базальные секреторные механизмы могут требовать меньше MARCKS-белка, чем стимулированная секреция. Базальная секреция может быть сохранена, т.к. любое лечение с блокадой секреции, опосредованной MARCKS, может не устранять все функции MARCKS.
Как используется в настоящем описании, термин «нуклеотидная последовательность MARCKS» относится к любой последовательности нуклеотидов, полученной из гена, кодирующего белок MARCKS, включая, например, последовательность ДНК или РНК, ДНК-последовательность гена, любую транскрибированную последовательность РНК, РНК-последовательность пре-мРНК или транскрипт мРНК, и ДНК или РНК, связанные с белком.
Точная доставка пептида, блокирующего MARCKS, также может преодолевать какие-либо возможные ограничения блокады важных секреторных процессов. Доставка таких агентов в дыхательный тракт должна быть легко осуществляемой с помощью ингаляционных композиций. Так как такие агенты могут быть применимыми в лечении воспалительных заболеваний кишечника, можно предусмотреть доставку блокирующих агентов в прямую кишку/толстую кишку/тонкую кишку посредством клизм или суппозиториев. Инъекции или трансдермальная доставка в воспаленные суставы могут давать облегчение пациентам с артритами или аутоиммунными заболеваниями путем ограничения секреции из локализованных воспалительных клеток. Инъекции в области, окружающие нервные окончания, могут ингибировать секрецию некоторых типов нейромедиаторов, блокируя передачу тяжелой боли или неконтролируемых мышечных спазмов. Доставка пептида для лечения воспалительных заболеваний кожи должна легко осуществляться с использованием различных местных композиций, известных в области техники.
Было показано, что миристоилированный богатый аланином С-киназный субстрат (MARCKS), широко распространенный субстрат РКС, может быть ключевой регуляторной молекулой, опосредующей высвобождение гранул муцина нормальными клетками человеческого эпителия бронхов (NHBE). Секреция муцина из таких клеток может быть доведена до максимума путем активации и РКС, и PKG. Считают, что MARCKS служит в качестве точки слияния для координации действия этих двух протеинкиназ для регуляции высвобождения гранул муцина. Вероятно, механизм включает РКС-зависимое фосфорилирование MARCKS, что высвобождает MARCKS из плазматической мембраны в цитоплазму, где он, в свою очередь дефосфорилируется протеинфосфатазой 2А (РР2А), которая активируется PKG. Такое дефосфорилирование может позволять MARCKS снова приобрести способность связываться с мембраной, допуская его прикрепление к мембранам цитоплазматических гранул муцина. Кроме того, MARCKS взаимодействует с актином и миозином в цитоплазме и, следовательно, может быть способен прикрепить гранулы к сократительному аппарату клетки, таким образом опосредуя последующее движение и экзоцитоз гранул. Секреция воспалительного медиаторного МРО из нейтрофилов также может быть доведена до максимума путем активации и РКС, и PKG (как проиллюстрировано на фиг. 11-15). Возможно, что MARCKS служит в качестве точки схождения для координирующего действия этих двух протеинкиназ, которые контролируют секрецию из отделов воспалительных клеток, связанных с мембраной (т.е. секрецию МРО из нейтрофилов).
Настоящее изобретение демонстрирует, что секреция воспалительного медиатора МРО из нейтрофилов собак или человека, усиливается сопутствующей активацией и РКС, и PKG, тогда как активация каждой киназы отдельно была недостаточной для индукции максимального секреторного ответа. Усиленный секреторный ответ на отдельно РМА был задокументирован в клетках NHBE (фиг. 1, колонка 4) и в нейтрофилах (фиг.11), хотя величина ответа была существенно меньшей, чем наблюдаемая другими в клеточной линии крысиных бокаловидных клеток. См. Abdullah et al. выше. Кроме того, хотя ранее сообщали, что аналог цГМФ может индуцировать существенную секрецию муцина из культивируемых клеток эпителия трахеи морских свинок (Fisher et al., выше), должно быть отмечено, что такой ответ не достигает значительного уровня до 8 ч воздействия. Секреторный ответ с таким длительным периодом последействия вероятно не является непосредственным эффектом и, возможно, включает синтез белка de novo в отличие от высвобождения заранее образованных и хранящихся цитоплазматических гранул. Тем не менее, очевидный синергический эффект, включающий совместную активацию и РКС, и PKG, может предполагать комплексный и точный сигнальный механизм, опосредующий секрецию муцина и/или воспалительных медиаторов. Заявители отмечают, что путь, описанный ниже, использовали для изучения высвобождения воспалительных медиаторов из нейтрофилов и, вероятно, таких же путей, как используемые для изучения секреции бокаловидных клеток.
Как указано выше, настоящее изобретение может быть использовано в фармацевтической композиции. В определенных вариантах осуществления изобретения лекарственный продукт присутствует в твердой фармацевтической композиции, которая может подходить для перорального введения. Может быть получена твердая смесь химически связанных веществ в соответствии с настоящим изобретением и она может быть смешана и/или разведена с эксципиентом. Твердая смесь химически связанных веществ также может быть заключена в носитель, который может быть, например, в форме капсулы, саше, таблетки, бумаги или другого контейнера. Когда эксципиент служит в качестве разбавителя, оно может быть твердым, полутвердым или жидким веществом, которое действует в качестве основы, носителя или среды для настоящей композиции.
Различные подходящие вспомогательные вещества понимаются специалистом в области техники и могут быть обнаружены в National Formulary, 19: 2404-2406 (2000), описание страниц 2404-2406 включено в настоящее описание полностью. Примеры подходящих эксципиентов включают, но не ограничиваются, крахмалы, гуммиарабик, силикат кальция, микрокристаллическую целлюлозу, метакрилаты, шеллак, поливинилпирролидон, целлюлозу, воду, патоку и метилцеллюлозу. Композиции лекарственного продукта, кроме того, могут содержать лубриканты, такие как, например, тальк, стеарат магния и минеральное масло; увлажняющие вещества; эмульгирующие и суспендирующие вещества; консерванты, такие как метил- и пропилгидроксибензоаты; подслащающие вещества; или ароматизаторы. Также могут быть использованы полиолы, буферы и инертные наполнители. Примеры полиолов включают, но не ограничиваются, маннит, сорбит, ксилит, сахарозу, мальтозу, глюкозу, лактозу, декстрозу и подобные. Подходящие буферы включают, но не ограничиваются, фосфат, цитрат, тартрат, сукцинат и подобные. Другие инертные наполнители, которые могут быть использованы, включают таковые, которые известны в области техники и являются применимыми в производстве различных лекарственных форм. При желании твердые композиции могут включать другие компоненты, такие как наполнители и/или гранулирующие агенты, и подобные. Лекарственные продукты по изобретению могут быть приготовлены в виде форм, таких, чтобы обеспечить быстрое, длительное или отложенное высвобождение активного ингредиента после введения пациенту с использованием методик, хорошо известных в области техники.
Для получения таблеток для перорального введения смесь химически связанных веществ по настоящему изобретению может быть получена методом прямого прессования. В таком способе активные лекарственные ингредиенты могут быть смешаны с твердым, порошкообразным носителем, таким как, например, лактоза, сахароза, сорбит, маннит, крахмал, амилопектин, производные целлюлозы или желатин и их смеси, а также с антифрикционным агентом, таким как, например, стеарат магния, стеарат кальция и воска полиэтиленгликоля. Смесь затем может быть спрессована в таблетки с использованием машины с соответствующими пуансонами и матрицами для получения желаемого размера таблеток. Рабочие параметры могут быть выбраны специалистом в данной области техники. Альтернативно таблетки для перорального введения могут быть получены способом влажной грануляции. Активные лекарственные ингредиенты могут быть смешаны с вспомогательными веществами и/или разбавителями. Твердые вещества могут быть измельчены или просеяны до желаемого размера частиц. К лекарственному средству может быть добавлено связующее вещество. Связующее вещество может быть суспендировано и гомогенизировано в подходящем растворителе. Активный ингредиент и вспомогательные вещества также могут быть смешаны с раствором связующего вещества. Полученную сухую смесь равномерно увлажняют раствором. Увлажнение обычно вызывает легкую агрегацию частиц, и полученную массу прессуют через сито из нержавеющей стали, имеющего желаемый размер. Затем смесь сушат в приборе для регулируемой сушки в течение определенного периода времени, необходимого для достижения желаемого размера и консистенции частиц. Гранулы сухой смеси просеивают для удаления любого порошка. К такой смеси могут быть добавлены дезинтегрирующие, антифрикционные и/или антиадгезивные вещества. Наконец, смесь прессуют в таблетки с использованием прибора с соответствующими пуансонами и матрицами для получения таблеток желаемого размера. Рабочие параметры прибора могут быть выбраны специалистом в данной области техники.
Если желательны таблетки, покрытые оболочкой, полученное выше ядро может быть покрыто оболочкой из концентрированного раствора сахара или полимеров целлюлозы, которые могут содержать гуммиарабик, желатин, тальк, диоксид титана, или лаком, растворенным в летучем органическом растворителе, или смесью растворителей. К таким различным оболочкам могут быть добавлены красители с целью различения таблеток с различными активными соединениями или с различными количествами присутствующего активного соединения. В определенном варианте осуществления изобретения активный ингредиент может присутствовать в ядре, окруженном одним или более слоями, включая слои кишечно-растворимой оболочки.
Могут быть получены мягкие желатиновые капсулы, в которых содержится смесь активного ингредиента и растительного масла. Твердые желатиновые капсулы могут содержать гранулы активного ингредиента в комбинации с твердым, порошкообразным носителем, таким как, например, лактоза, сахароза, сорбит, манит, картофельный крахмал, кукурузный крахмал, амилопектин, производные целлюлозы и/или желатин.
Жидкие препараты для перорального введения могут быть получены в форме сиропов или суспензий, например, растворов, содержащих активный ингредиент, сахар и смесь этанола, воды, глицерина и пропиленгликоля. При желании такие жидкие препараты могут включать одно или более из следующего: красящие вещества, ароматизаторы и сахарин. Также могут быть использованы сгущающие вещества, такие как карбоксиметилцеллюлоза.
В случае, когда вышеуказанные фармацевтические вещества должны использоваться для парентерального введения, такая композиция может включать стерильные водные растворы для инъекций, неводные растворы для инъекций или оба, содержащие смесь химически связанных веществ по настоящему изобретению. Когда получают водные растворы для инъекций, смесь химически связанных веществ может присутствовать в виде растворимой в воде фармацевтически приемлемой соли. Парентеральные препараты могут содержать антиоксиданты, буферы, бактериостатики и растворы, которые делают композицию изотоничной с кровью предназначенного реципиента. Водные и неводные стерильные суспензии могут включать суспендирующие вещества и загустители. Композиции могут быть представлены в однодозовых или многодозовых контейнерах, например герметичных ампулах или флаконах. Свежеприготовленные растворы и суспензии для инъекций могут быть получены из стерильных порошков, гранул и таблеток описанного ранее типа.
Смесь химически связанных веществ также может быть рецептирована, так что она может подходить для местного введения (например, кожный крем). Такие композиции могут содержать различные вспомогательные вещества, известные специалисту в данной области техники. Подходящие вспомогательные вещества могут включать, но не ограничиваются, воски цетиловых эфиров, цетиловый спирт, белый воск, моностеарат глицерина, пропиленгликоль, моностеарат, метилстеарат, бензиловый спирт, лаурилсульфат натрия, глицерин, минеральное масло, воду, карбомер, этиловый спирт, адгезивы акрилата, адгезивы полиизобутилена и адгезивы диоксида кремния.
После описания изобретения то же будет проиллюстрировано со ссылками на определенные примеры, которые включены в настоящее описание только с целью иллюстрации и не должны рассматриваться как ограничивающие изобретение.
ПРИМЕРЫ
Гиперсекреция муцина из клеток NHBE включает активацию как РКС, так и PKG
Для определения потенциальной роли РКС и/или PKG в процессе секреции муцина, клетки подвергали воздействию следующих двух специфических активаторов протеинкиназ: форболового эфира, форбол 12-миристата 13-ацетата (РМА), для активации РКС и моногидролизуемого аналога цГМФ, 8-Br-cGMP, для активации PKG. Предварительные исследования по изучению секреции муцина в ответ на стимуляцию РМА в различных концентрациях в течение различного периода времени (до 1 мкг в течение 2 ч) показали, что активация отдельно РКС не вызывает существенной секреции муцина из клеток NHBE, хотя умеренный секреторный ответ повторно наблюдали при концентрации РМА выше, чем 100 нМ (0,05<p<0,1). Также клетки не отвечали на аналоги цГМФ в концентрациях, таких как 500 мкМ, в течение до 2 ч воздействия. Однако комбинация РМА + 8-Br-cGMP, вызывающая двойную активацию РКС и PKG, способствовала быстрому увеличению секреции, приблизительно удваивая ее в течение 15 мин воздействия (фиг. 1А). Такой секреторный ответ, вызванный РМА + 8-Br-cGMP зависел от концентрации, с максимальной стимуляцией при 100 нМ РМА + 1 мкМ 8-Br-cGMP (фиг. 1В и 1С). На фиг. 1А, 1В и 1С клетки NHBE подвергали воздействию указанного реагента(ов) или отдельно среды (CTL) в течение 15 мин. На фиг. 1D клетки NHBE предварительно инкубировали с указанным ингибитором в течение 15 мин и затем стимулировали 100 мкM УТФ в течение 2 ч. Секретируемый в ответ на обработку муцин собирали и анализировали с помощью ELISA. Данные представлены как среднее ± стандартная ошибка (SE) (n=6 в каждой точке). * обозначает статистическое отличие от контрольной среды (p<0,05), # обозначает различие с контрольной средой (0,05<p<0,1); и ‡ обозначает статистическое отличие от стимуляции УТФ CD<0,05).
УТФ представляет собой хорошо известный физиологически важный стимулятор секреции муцина. Lethem et al., Am. J.Respir.Cell Mol.Biol 9, (315-322 (1993). Настоящее изобретение, кроме того, демонстрирует, что УТФ, в различных концентрациях, предпочтительно от 40 до 140 мкМ, может индуцировать существенное увеличение секреции муцина из клеток NHBE после 2 ч воздействия. Для определения, вовлечены ли РКС, и PKG в регуляцию секреции муцина в ответ на физиологические стимулы, изучали эффекты ингибиторов РКС/PKG на секрецию муцина, индуцированную УТФ. Клетки NHBE предварительно инкубировали с различными ингибиторами в течение 15 мин и затем подвергали воздействию УТФ (100 мкМ) плюс ингибитор в течение 2 ч. Секретируемый муцин измеряли посредством ELISA. Результаты показывают, что секреция муцина, вызванная УТФ, требует воздействия и РКС, и PKG, так как секреторный ответ снижался независимо ингибитором РКС кальфостином С (500 нМ), ингибитором PKG Rp-8-Br-PET-cGMP (10 мкМ) или растворимым ингибитором гуанилилциклазы (GC-S) LY83583 (50 мкМ), но, вероятно, не ингибитором протеинкиназы А (РКА) KT5720 (500 нМ) (фиг. 1D). Вероятно секреция муцина в клетках NHBE может регулироваться сигнальным механизмом, включающим и РКС, и PKG.
Для изучения участия PKG в секреторном процессе в таких исследованиях использовали 8-Br-cGMP. Хотя первичный физиологический эффект 8-Br-cGMP представляет собой активацию PKG, также сообщают, что он действует как агонист для цГМФ-зависимых ионных каналов в некоторых клетках и, в высоких концентрациях, перекрестным образом активирует РКА. Для исключения возможности, что цГМФ-зависимые ионные каналы и/или РКА могут играть роль в секреции муцина клетками NHBE, Rp-8-Br- cGMP, уникальный аналог цГМФ, который может активировать цГМФ-зависимые ионные каналы сходно с 8-Br-cGMP, но ингибировать активность PKG, использовали в качестве агониста для различения эффектов PKG и цГМФ-зависимых ионных каналов на высвобождение муцина. Как проиллюстрировано на чертежах, особенно, фиг. 1А (колонка 11), Rp-8-Br-cGMP не усиливал секрецию муцина при добавлении к клеткам с РМА. Подобным образом, специфический ингибитор РКА, КТ5720 (500 нМ), не влиял на секрецию муцина, вызванную или РМА + 8-Br-cGMP или УТФ (фиг. 1D, колонка 4). Такие исследования могут опровергать возможность, что цГМФ-зависимые ионные каналы или РКА ассоциированы с секрецией муцина, указывая, что активация PKG в клетках NHBE представляет собой механизм, посредством которого 8-Br-cGMP влияет на усиление секреции. Более того, так как УТФ-индуцированная гиперсекреция муцина может быть снижена растворимым ингибитором гуанилил циклазы (GC-S) LY83583, вероятно, что активация PKG происходит посредством сигнального пути оксида азота (NO) → GC-S → cGMP → PKG, как проиллюстрировано ранее в дифференцированных клетках эпителия трахеи морских свинок in vitro.
Предполагая участие и РКС, и PKG в процессе секреции муцина, настоящее изобретение изучает потенциальные внутриклеточные субстраты этих ферментов, которые могут играть роль в сигнальных событиях по ходу активации киназы. Множество внутриклеточных субстратов могут быть фосфорилированы РКС или PKG, и фосфорилирование РКС одного такого субстрата, MARCKS-белка, вероятно является особо интересным. Наблюдали, что фосфорилирование MARCKS коррелирует с рядом клеточных процессов, включающих сигнал РКС и сокращение цитоскелета, таких как движение клетки, митогенез и высвобождение нейромедиаторов. Так как динамический процесс секреции требует и активации киназы и транслокации внутриклеточных гранул на периферию клетки, вероятно, MARCKS является кандидатом в медиаторные молекулы, связывающие активацию РКС/PKG и экзоцитоз гранул муцина.
MARCKS является ключевой молекулой, связывающей активацию РКС/PKG и секрецию муцина в клетках NHBE
Для определения сигнального механизма по ходу активации протеинкиназы, изучали MARCKS-белок, специфический клеточный субстрат РКС, который может играть роль в связи активации киназы с высвобождением гранул. Во-первых, подтверждали присутствие MARCKS в клетках NHBE путем иммунопреципитации меченной [Н3] миристиновой кислотой. Как проиллюстрировано на фиг. 2А, MARCKS экспрессировался в клетках NHBE и большая часть этого белка в нестимулированном состоянии была ассоциирована с мембраной. На фиг. 2А клетки метили [3Н] миристиновой кислотой в течение ночи и затем дифференциальным центрифугированием выделяли мембранные (дорожка 1) и цитозольные (дорожка 2) фракции. Роль MARCKS в качестве ключевого регуляторного компонента в секреторном пути муцина может быть продемонстрирована тремя различными способами.
Как указано выше, непосредственное участие MARCKS в секреции муцина клетками NHBE может быть продемонстрировано тремя различными группами доказательств. Во-первых, секреция муцина в ответ на стимуляцию РМА + 8-Br-cGMP или УТФ ингибировалась в зависимости от концентрации MANS-пептида, который имеет аминокислотную последовательность, идентичную N-концевому участку MARCKS, тогда как соответствующий контрольный пептид (RNS), содержащий такую же аминокислотную композицию, но расположенную в случайном порядке, не влиял на секрецию. Известно, что N-концевой миристоилированный домен MARCKS опосредует ассоциацию MARCKS-мембрана. Как показано на фиг. 8, MARCKS может действовать как молекулярный линкер путем взаимодействия с мембранами гранул на его N-концевом домене и связывания с актиновыми волокнами в его PSD участке, таким образом прикрепляя гранулы к контрактильному цитоскелету для движения и экзоцитоза. На фиг.8 показан возможный механизм, изображающий как средства, стимулирующие секрецию муцина, взаимодействуют с клетками эпителия дыхательных путей (бокаловидными) и активируют две отдельные протеинкиназы, РКС и PKG. Активированная РКС фосфорилирует MARCKS, вызывая транслокацию MARCKS с плазматической мембраны в цитоплазму, тогда как PKG, активированная посредством пути оксида азота (NO)→GC-S→cGMP→PKG, в свою очередь активирует цитоплазматический РР2А, который дефосфорилирует MARCKS. Такое дефосфорилирование стабилизирует прикрепление MARCKS к мембранам гранул. Кроме того, MARCKS также взаимодействует с актином и миозином, таким образом связывая гранулы с клеточным контрактильным цитоскелетом для последующего движения и экзоцитозного высвобождения. Прикрепление MARCKS к гранулам после его высвобождения в цитоплазму также может быть осуществлено специфическими нацеливающими белками или некоторыми другими формами взаимодействий белок-белок, в которые вовлечен N-концевой домен MARCKS. В любом случае MANS-пептид или его активный фрагмент, включающий по меньшей мере 6 аминокислот, будет действовать, ингибируя полное нацеливание MARCKS на мембраны гранул муцина, таким образом блокируя секрецию.
Второй тест демонстрирует ингибирующий эффект MARCKS-специфического антисмыслового олигонуклеотида на секрецию муцина. Как показано на фиг. 3А-3С, антисмысловой олигонуклеотид подавлял уровень мРНК и белка MARCKS в клетках NHBE и существенно ослаблял секрецию муцина, индуцированную активацией РКС/PKG. Ингибирование не было настолько сильным, как наблюдаемое для MANS-пептида, что может быть из-за высокого уровня эндогенного MARCKS-белка в клетках NHBE и относительно длинного периода полужизни мРНК MARCKS (t1/2=4-6 ч). На фиг. 3А-3С, клетки NHBE обрабатывали антисмысловым или контрольным олигонуклеотидом в течение 3 дней и затем стимулировали РМА (100 нМ) + 8-Br-cGMP (1 мкМ) в течение 15 мин. Секрецию муцина анализировали ELISA. Всю РНК и белок выделяли из обработанных клеток. мРНК MARCKS оценивали посредством Нозерн гибридизации и белок оценивали Вестерн блоттингом. При РМА (100 нМ)+8-Br-cGMP (1 мкМ) фиг. 3А представляет собой Нозерн блоттинг, который показал снижение на около 15% мРНК MARCKS по сравнению с контролем в приложенной шкале; фиг. 3В представляет собой Вестерн блоттинг, который показывает снижение около 30% белка MARCKS на приложенном графике; и фиг. 3С показывает, что гиперсекреция муцина существенно снижалась антисмысловым нуклеотидом, тогда как контрольный олигонуклеотид не оказывал эффекта. Данные представлены как среднее ± стандартная ошибка (SE) (n=6 в каждой точке), где * представляет собой статистическое отличие от среднего контроля (p<0,05); и † представляет собой статистическое отличие от стимуляции РМА + 8-Br-cGMP (p<0,05). Кроме того, отмечают, что термин СТО представляет собой контрольный олигонуклеотид, тогда как термин ASO представляет собой антисмысловой олигонуклеотид.
Было продемонстрировано, что антисмысловые олигонуклеотиды, которые являются комплементарными к специфической РНК, могут ингибировать экспрессию клеточных генов как белков. См. Erickson and Izant, Gene Regulation: Biology of Antisense RNA and DNA, Vol. 1, Raven Press, N.Y., 1992. Например, сообщают о селективном ингибировании гена р21, который отличается от нормального гена единственным нуклеотидом. Chang et al., Biochemistry (1991), 30: 8283-8286. Было предложено множество гипотез для объяснения механизмов, посредством которых антисмысловые олигонуклеотиды ингибируют экспрессию генов, однако специфический участвующий механизм может зависеть от исследуемого типа клеток, РНК-мишени, специфического участка на целевой РНК и химической природы олигонуклеотида. Chiang et al., J.Biol.Chem. 1991, 266: 18162-18171; Stein and Cohen, Cancer Res. 1988, 48: 2659-2668.
Третий эксперимент показал, что трансфекция клеток HBE1 мутантным MARCKS с делецией PSD приводила к существенному подавлению секреции муцина, индуцированной активацией РКС/PKG. Делеция PSD устраняет способность MARCKS связываться с актином. Как показано на фиг. 8, путем конкуренции с естественным MARCKS за связывание с мембраной гранул, MARCKS, усеченный на PSD, может таким образом ингибировать высвобождение гранул, так что они не способны взаимодействовать с актиновыми волокнами. Трансфекция таких клеток кДНК MARCKS дикого типа дополнительно не увеличивала секрецию муцина. Вестерн блоттинг анализ показал, что уровень экспрессии эндогенного MARCKS в клетках HBE1 был достаточно высоким, сравнимым с таковым в клетках NHBE, и трансфекция кДНК MARCKS дикого типа не приводила к заметному увеличению общего уровня MARCKS-белка в таких клетках. Это может объяснить, почему трансфекция MARCKS дикого типа дополнительно не увеличивала секрецию и также почему трансфекция MARCKS с PSD делецией только частично препятствовала секреции муцина.
Исследования блокады пептида - клетки NHBE - предварительно инкубировали или с MANS или с RNS пептидом (1-100 мкМ) в течение 15 мин, и затем добавляли РМА (100 нМ) + 8-Br-cGMP (1 мкМ) или УТФ (100 мкМ) и клетки инкубировали в течение дополнительных 15 мин или 2 ч, соответственно. Секрецию муцина измеряли ELISA. Как показано на фиг. 2В, инкубация клеток NHBE с MANS-пептидом приводила к зависимой от концентрации супрессии секреции муцина в ответ на активацию РКС/PKG или стимуляцию УТФ, тогда как контрольный пептид (RNS) может не влиять на секрецию в таких же концентрациях. На фиг. 2В, MANS-пептид блокирует гиперсекрецию муцина, индуцированную РМА + 8-Br-cGMP или УТФ в зависимости от концентрации. Клетки NHBE предварительно инкубировали с указанным пептидом в течение 15 мин и затем подвергали воздействию РМА (100 нМ) + 8-Br-cGMP (1 мкМ) в течение 15 мин или УТФ (100 мкМ) в течение 2 ч. Секрецию муцина измеряли ELISA. Данные представлены как среднее ± стандартная ошибка (SE) (n=6 в каждой точке), где * представляет собой статистическое отличие от среднего контроля (p<0,05); † представляет собой статистическое отличие от стимуляции РМА + 8-Br-cGMP (p<0,05); и ‡ представляет собой статистическое отличие от стимуляции УТФ (<0,05). Эффекты MANS-пептида, вероятно, не были связаны с цитотоксичностью или общим подавлением клеточной метаболической активности, так как ни MANS ни RNS-пептид не влияли на высвобождение лактатдегидрогеназы или потребление [3Н]деоксиглюкозы клетками.
Исследования антисмысловых олигонуклеотидов - чтобы дополнительно продемонстрировать MARCKS в качестве ключевого сигнального компонента секреторного пути муцина, изучали эффект антисмыслового олигонуклеотида к MARCKS на секрецию муцина. Как проиллюстрировано на фиг. 3, такой антисмысловой олигонуклеотид подавлял уровень и мРНК, и белка MARCKS в клетках NHBE и существенно снижал секрецию муцина, индуцированную РМА + 8-Br-cGMP, тогда как контрольный олигонуклеотид не оказывал эффекта.
MARCKS служит конвергентной сигнальной молекулой, опосредующей взаимное влияние путей РКС и PKG
В общем, вышеуказанные результаты демонстрируют, что MARCKS в целом вовлечен в процесс секреции муцина. Далее авторы настоящего изобретения определяли, как MARCKS действует в качестве ключевой регуляторной молекулы, на которой сходятся РКС и PKG для регуляции секреции муцина. Как проиллюстрировано на фиг. 5, MARCKS фосфорилировался РКС и впоследствии переносился с мембраны в цитоплазму. Так, вероятно PKG индуцирует дефосфорилирование MARCKS (фиг. 5А, дорожка 4 и фиг. 5В). Такое дефосфорилирование обращалось ингибитором PKG Rp-8-Br-PET-cGMP (фиг. 5А, дорожка 5), указывая, что дефосфорилирование было специфически PKG-зависимым. На фиг. 5 клетки NHBE метили [32Р]ортофосфатом и затем подвергали воздействию указанных реагентов. Фосфорилирование MARCKS в ответ на обработку оценивали анализом иммунопреципитации. На фиг. 5А, 8-Br-сGMP обращал фосфорилирование MARCKS, индуцированное РАМ, и такой эффект 8-Br-сGMP мог быть заблокирован Rp-8-Br-PET-cGMP (ингибитор PKG) или окадаевой кислотой (ингибитор РР1/2А). Для фиг. 5В, РМА-индуцированное фосфорилирование MARCKS обращали последующим воздействием на клетки 8-Br-cGMP. Дорожка 1, только среда в течение 8 мин; дорожка 2, 100 нМ РМА в течение 3 мин; дорожка 3, 100 нМ РМА в течение 3 мин и затем с 1 мкМ 8-Br-cGMP в течение 5 мин; дорожка 4, 100 нМ РМА в течение 8 мин; дорожка 5, среда отдельно в течение 3 мин и затем 100 нМ РМА + 1 мкМ 8-Br-cGMP в течение 5 мин. На фиг. 5С дефосфорилирование MARCKS, индуцированное 8-Br-cGMP, уменьшали фостриецином в зависимости от концентрации.
Считают, что PKG действует, фосфорилируя MARCKS посредством активации протеинфосфатазы. Как проиллюстрировано на фиг. 5А (дорожка 6), окадаевая кислота в концентрации 500 нМ, концентрации, которая может ингибировать и РР1, и РР2А, блокировала PKG-индуцированное дефосфорилирование MARCKS, предполагая, что PKG вызывает дефосфорилирование путем активации РР1 и/или РР2А. Последующие исследования с фостриецином и прямой анализ активности фосфатазы показал, что только РР2А активировался PKG и отвечал за удаление фосфатных групп с MARCKS (фиг. 5С). Вероятно, что или окадаевая кислота или фостриецин, в концентрациях, которые ингибируют PKG-индуцированное дефосфорилирование MARCKS, снижали секрецию муцина, индуцированную РМА +8-Br-cGMP или УТФ, как показано на фиг.6. Фиг. 6 помогает продемонстрировать, что РР2А является важным компонентом секреторного пути муцина. Клетки NHBE предварительно инкубировали с указанной концентрацией фостриецина, окадаевой кислоты (500 нМ) или отдельно средой в течение 15 мин и затем стимулировали РМА (100 нМ) + 8-Br-cGMP (1 мкМ) в течение 15 мин или УТФ (100 мкМ) в течение 2 ч. Секретируемый муцин измеряли ELISA. Данные представлены как среднее ± стандартная ошибка (SE) (n=6 в каждой точке), где * представляет собой статистическое отличие от среднего контроля (p<0,05); † представляет собой статистическое отличие от стимуляции РМА + 8-Br-cGMP (p<0,05); и ‡ представляет собой статистическое отличие от стимуляции УТФ (р<0,05). Следовательно, дефосфорилирование MARCKS посредством PKG-активированного РР2А, вероятно является важным компонентом сигнального пути, приводящего к экзоцитозу гранул муцина.
Для выявления молекулярных событий, посредством которых MARCKS связывает активацию киназы с секрецией муцина, подробно исследовали фосфорилирование MARCKS в ответ на активацию РКС/PKG. Как проиллюстрировано на фиг. 4А, РМА (100 нМ), вероятно, вызывает существенное увеличение (3-4 кратное) фосфорилирования MARCKS в клетках NHBE и такое фосфорилирование уменьшается ингибитором РКС кальфостином С (500 нМ). При фосфорилировании MARCKS транслоцировался с плазматической мембраны в цитоплазму (фиг. 4В). В частности, на фиг. 4А показано, что активация РКС приводит к фосфорилированию MARCKS в клетках NHBE. Клетки метили [32Р]ортофосфатом в течение 2 ч, затем подвергали воздействию стимулирующего и/или ингибирующего реагента. Фосфорилирование MARCKS в ответ на обработку оценивали иммунопреципитацией, как описано. Дорожка 1, контрольная среда; дорожка 2 носитель, 0,1% Me2SO; дорожка 3, 100 нМ 4α-РМА; дорожка 4, 100 нМ РМА; дорожка 5, 100 нМ РМА + 500 нМ кальфостина С; дорожка 6, 500 нМ кальфостина С. Фиг. 4В демонстрирует, что фосфорилированный MARCKS транслоцируется из плазматической мембраны в цитоплазму. Клетки, меченные 32Р, подвергали воздействию РМА (100 нМ) или среды отдельно в течение 5 мин, и затем выделяли мембранную и цитозольную фракции. Активация PKG 8-Br-cGMP (1 мкМ), активация другой киназы необходима для вызывания секреции муцина, не приводит к фосфорилированию MARCKS, но, в действительности, наблюдали противоположный эффект: фосфорилирование MARCKS, индуцированное РМА, обращали 8-Br-cGMP (фиг. 5А). Такой эффект 8-Br-cGMP происходит не из-за супрессии активности РКС, так как фосфорилирование, индуцированное РМА, может быть обращено последующим добавлением 8-Br-cGMP к клеткам (фиг. 5В). Следовательно, вероятно, активация PKG приводит к дефосфорилированию MARCKS.
Последующие исследования продемонстрировали, что дефосфорилирование MARCKS, индуцированное PKG, блокировалось 500 нМ окадаевой кислоты, ингибитором протеинфосфатазы (тип 1 и/или 2А (РР1/2А)) (фиг. 5А, дорожка 6). Следовательно, вероятно, что дефосфорилирование было опосредовано РР1 и/или РР2А. Для определения вовлеченного подтипа протеинфосфатазы в дополнительных исследованиях фосфорилирования использовали новый и более специфический ингибитор РР2А, фостриецин (IC50=3,2 нМ). Как проиллюстрировано на фиг. 5С, фостриецин ингибировал PKG индуцированное дефосфорилирование MARCKS в зависимости от концентрации (1-500 нМ), предполагая, что PKG индуцирует дефосфорилирование посредством активации РР2А. Для подтверждения дальнейшей активации РР2А PKG в клетках NHBE определяли активность цитозольных РР1 и РР2А после воздействия на клетки 8-Br-cGMP. Активность РР2А увеличивалась приблизительно в 3 раза (от 0,1 до 0,3 нмоль/мин/мг белка, р<0,01) в концентрации 8-Br-cGMP, такой низкой как 0,1 мкМ, тогда как активность РР1 оставалась неизменной. Эти данные указывают, что РР2А может быть активирован PKG и является ответственным за дефосфорилирование MARCKS. Соответственно, такая активность РР2А, вероятно, является критической для возникновения секреции муцина; когда PKG-индуцированное дефосфорилирование MARCKS блокируется окадаевой кислотой или фостриецином, секреторный ответ на активацию РКС/PKG или стимуляцию УТФ уменьшается (фиг. 6).
MARCKS связывается с актином и миозином в цитоплазме
На фиг. 7 изображен анализ иммунопреципитации с радиоактивной меткой, который выявил, что MARCKS может ассоциироваться в цитоплазме с двумя другими белками (около 200 и около 40 кДа). На фиг. 7 клетки NHBE метили [3Н]лейцином и [3Н]пролином в течение ночи, и получали мембранную и цитозольную фракции, как описано в «Методиках экспериментов». Выделенные фракции предварительно очищали неиммунным контрольным антителом (6F6). Затем цитозоль делили поровну на две фракции и использовали для иммунопреципитации, проводимой в присутствии 10 мкМ цитохалазина D (Biomol, Plymouth Meeting, Pa.) с анти-MARCKS антителом 2F12 (дорожка 2) и неиммунным контрольным антителом 6F6 (дорожка 3), соответственно. MARCKS-белок в мембранной фракции также оценивали иммунопреципитацией с использованием антитела 2F12 (дорожка 1). Осажденный белковый комплекс разделяли путем электрофореза в 8% SDS-полиакриламидном геле и визуализировали с помощью усиленной авторентгенографии. Вероятно, MARCKS ассоциируется с двумя цитоплазматическими белками с молекулярной массой около 200 и около 40 кДа, соответственно. Полученные два белка, ассоциированные с MARCKS, выделяли из геля и анализировали с помощью ионизации с лазерной десорбцией с использованием матрицы/время-пролетной масс-спектрометрии/внутреннего секвенирования (the Protein/DNA Technology Center of Rockefeller University, N.Y.). Полученные данные о массе и последовательности пептида использовали для поиска в базах данных белков посредством программ Интернет ProFound и MS-Fit. Результаты показали, что это был миозин (тяжелая цепь, немышечный тип А), и актин, соответственно. Анализ с помощью ионизации с лазерной десорбцией с использованием матрицы/время-пролетной масс-спектрометрии/внутреннего секвенирования показал, что эти два белка, ассоциированные с MARCKS, представляли собой миозин (тяжелая цепь, немышечный тип А), и актин, соответственно.
Эти исследования предполагают новую модель сигнального механизма, регулирующего экзоцитозную секрецию гранул муцина дыхательных путей, а также обеспечивают то, что является первым непосредственным доказательством, демонстрирующим специфическую биологическую функцию MARCKS в физиологическом процессе. MARCKS служит в качестве ключевой медиаторной молекулы, регулирующей высвобождение гранул муцина в человеческих эпителиальных клетках дыхательных путей. Считают, что выявление секреции муцина дыхательных путей требует двойной активации и синергического действия РКС и PKG. Активированная РКС фосфорилирует MARCKS, приводя к транслокации MARCKS с внутренней поверхности плазматической мембраны в цитоплазму. Активация PKG в свою очередь активирует РР2А, который дефосфорилирует MARCKS в цитоплазме. Так как способность к ассоциации с мембраной MARCKS зависит от его состояния фосфорилирования, такое дефосфорилирование может позволить MARCKS снова приобрести его способность связываться с мембраной и может сделать возможным прикрепление MARCKS к мембранам цитоплазматических гранул муцина. Путем такого взаимодействия с актином и миозином в цитоплазме (фиг. 7) MARCKS будет способен привязывать гранулы к сократительному аппарату клетки, опосредуя движение гранул к периферии клетки и последующее экзоцитозное высвобождение. Широкое распределение MARCKS предполагает возможность, что такой или подобный механизм может регулировать секрецию связанных с мембраной гранул в различных типах клеток при нормальных или патологических состояниях.
Изобретение также относится к новому способу блокады любого клеточного экзоцитозного секреторного процесса, особенно процесса высвобождения воспалительных медиаторов из гранул, содержащихся в воспалительных клетках, где такие секреторные пути включают субстрат протеинкиназы С (РКС) MARCKS-белок и высвобождение содержимого из везикул, связанных с мембраной. В частности, авторы изобретения показали, что стимулированное высвобождение воспалительного медиатора миелопероксидазы из человеческих (фиг. 9) или собачьих (фиг. 10) нейтрофилов может быть заблокировано MANS-пептидом в зависимости от концентрации. В частности, на фиг. 9 показаны выделенные нейтрофилы, которые стимулировали для секреции миелопероксидазы (МРО) с помощью 100 нМ РМА и 10 мкМ 8-Br-cGMP. 100 мкМ MANS-пептида снижали секрецию МРО до контрольного уровня (*=p<0,05). 10 мкМ MANS вызывало небольшое снижение секреции МРО. 10 или 100 мкМ контрольного пептида (RNS) не оказывало эффекта на секрецию МРО. На фиг. 10, выделенные нейтрофилы стимулировали к секреции миелопероксидазы (МРО) с помощью 100 нМ РМА и 10 мкМ 8-Br-cGMP. 100 мкМ MANS-пептида снижали секрецию МРО до контрольного уровня (*=p<0,05). 10 мкМ MANS вызывало небольшое снижение секреции МРО. 10 или 100 мкМ контрольного пептида (RNS) не оказывало эффекта на секрецию МРО. Следовательно, пептид может быть терапевтически использован для блокады высвобождения медиаторов воспаления, секретируемых из инфильтрирующих воспалительных клеток в любых тканях. Множество из таких высвобождаемых медиаторов ответственны за выраженное повреждение тканей, наблюдаемое при множестве хронических воспалительных заболеваний (т.е. респираторных заболеваниях, таких как астма, хронический бронхит и ХОБЛ, воспалительных заболеваниях кишечника, включая язвенный колит и болезнь Крона, аутоиммунных заболеваниях, заболеваниях кожи, таких как розовые угри, экзема; тяжелые акне, артритические и болевые синдромы, такие как ревматоидный артрит и фибромиалгия). Настоящее изобретение может быть применимым для лечения заболеваний, таких как артрит, хронический бронхит, ХОБЛ и муковисцидоз. Настоящее изобретение является соответственно применимым для лечения заболеваний и людей, и животных, особенно таких, поражающих лошадей, собак, кошек и других домашних животных.
На фиг. 11-15 показана секреция МРО для людей и собак. Во всех этих экспериментах выделенные нейтрофилы стимулировали LPS в концентрации 1×10-6 М в течение 10 минут при 37°С, перед добавлением стимуляторов, как показано на чертежах. LPS примирует клетки так, что они могут отвечать на стимуляторы секреции.
В одном варианте осуществления настоящее изобретение описывает способ регуляции воспаления у пациента, включающий введение терапевтически эффективного количества фармацевтической композиции, содержащей MANS-пептид или его активный фрагмент. В одном аспекте настоящего изобретения указанный активный фрагмент MANS-белка включает, по меньшей мере, шесть аминокислот. В другом аспекте указанное воспаление вызывают респираторные заболевания, заболевания кишечника, заболевания кожи, аутоиммунные заболевания и болевые синдромы. В другом аспекте указанные респираторные заболевания выбирают из группы, состоящей из астмы, хронического бронхита и ХОБЛ. В другом аспекте указанные заболевания кишечника выбирают из группы, состоящей из язвенного колита, болезни Крона и синдрома раздраженной толстой кишки. В другом аспекте указанные заболевания кожи выбирают из группы, состоящей из розовых угрей, экземы, псориаза и тяжелого акне. В другом аспекте указанное воспаление вызывает артрит или муковисцидоз. В другом аспекте указанным пациентом является млекопитающее. Кроме того, в другом аспекте указанное млекопитающее выбирают из группы, состоящей из людей, собак, лошадей и кошек. В другом аспекте указанную стадию введения выбирают из группы, состоящей из местного введения, парентерального введения, ректального введения, легочного введения, назального введения, ингаляции и перорального введения. В другом аспекте указанное легочное введение выбирают из группы введения в виде аэрозоля, с помощью порошкового ингалятора, ингалятора с дозирующим клапаном и небулайзера.
В другом варианте осуществления настоящее изобретение описывает способ регуляции клеточного секреторного процесса у пациента, включающий введение терапевтически эффективного количества фармацевтической композиции, включающей, по меньшей мере, одно соединение, включающее MANS-пептид или его активный фрагмент, которые регулируют медиатор воспаления у пациента. В одном аспекте настоящего изобретения указанный активный фрагмент MANS-белка содержит, по меньшей мере, шесть аминокислот. В другом аспекте указанная регуляция клеточного секреторного процесса представляет собой блокаду или снижение клеточного секреторного процесса. В другом аспекте указанный воспалительный медиатор возникает в результате респираторных заболеваний, заболеваний кишечника, заболеваний кожи, аутоиммунных заболеваний и болевых синдромов. В другом аспекте указанные респираторные заболевания выбирают из группы, состоящей из астмы, хронического бронхита и ХОБЛ. В другом аспекте указанные заболевания кишечника выбирают из группы, состоящей из язвенного колита, болезни Крона и синдрома раздраженной кишки. В другом аспекте указанные кожные заболевания выбирают из группы, состоящей из розовых угрей, экземы, псориаза и тяжелого акне. В другом аспекте указанный воспалительный медиатор выделяется в результате артрита или муковисцидоза. В другом аспекте указанным пациентом является млекопитающее. В другом аспекте указанное млекопитающее выбирают из группы, состоящей из людей, собак, лошадей и кошек. В другом аспекте указанную стадию введения выбирают из группы, состоящей из местного введения, парентерального введения, ректального введения, легочного введения, назального введения, ингаляции и перорального введения. В другом аспекте указанное легочное введение выбирают из группы введения в виде аэрозоля, при помощи порошкового ингалятора, ингалятора с дозирующим клапаном и небулайзера.
В другом варианте осуществления настоящее изобретение описывает способ уменьшения воспаления у пациента, включающий введение терапевтически эффективного количества соединения, которое ингибирует MARCKS-опосредованное высвобождение медиаторов воспаления, посредством чего высвобождение медиаторов воспаления у пациента снижается по сравнению с таковым, которое возникает в отсутствие указанного лечения. В одном аспекте настоящего изобретения указанное соединении представляет собой, по меньшей мере, один активный фрагмент MARCKS-белка. В другом аспекте указанный активный фрагмент имеет по меньшей мере шесть аминокислот в длину. В другом аспекте указанным соединением является MANS-пептид или его активный фрагмент. В другом аспекте указанным соединением является антисмысловой олигонуклеотид, направленный против кодирующей последовательности MARCKS-белка или его активного фрагмента. В другом аспекте указанный активный фрагмент имеет по меньшей мере шесть аминокислот в длину.
В другом варианте осуществления настоящее изобретение описывает способ уменьшения воспаления у пациента, включающий введение терапевтически эффективного количества фармацевтически активной композиции, содержащей соединение, которое ингибирует MARCKS-связанное высвобождение медиаторов воспаления, посредством чего воспаление у пациента снижается по сравнению с таковым, которое возникает в отсутствие указанного лечения. В одном аспекте настоящего изобретения указанное соединение представляет собой активный фрагмент MARCKS-белка. В другом аспекте указанный активный фрагмент имеет по меньшей мере шесть аминокислот в длину. В другом аспекте указанным соединением является MANS-пептид или его активный фрагмент. В другом аспекте указанным соединением является антисмысловой олигонуклеотид, направленный против кодирующей последовательности MARCKS-белка или его активного фрагмента. В другом аспекте указанный активный фрагмент имеет по меньшей мере шесть аминокислот в длину. Настоящее изобретение включает композицию, которая содержит один или более из MANS-пептида или его активных фрагментов и их применение в лечении ингибирования высвобождения медиаторов воспаления из гранул или везикул воспалительных клеток.
В другом варианте осуществления настоящее изобретение описывает способ регуляции высвобождения гранул муцина у пациента, включающий введение соединения, которое регулирует высвобождение гранул муцина, посредством чего гранулы муцина уменьшаются по сравнению с тем, что возникает в отсутствие указанных гранул муцина. В одном аспекте настоящего изобретения указанное соединение представляет собой активный фрагмент MARCKS-белка. В другом аспекте указанное соединение представляет собой MANS-пептид.
В другом варианте осуществления настоящее изобретение описывает способ регуляции экзоцитозной секреции гранул муцина в дыхательных путях пациента, включающий: введение соединения, которое регулирует высвобождение гранул муцина, посредством чего гранулы муцина снижаются по сравнению с тем, что возникает в отсутствие указанных гранул муцина. В одном аспекте настоящего варианта осуществления указанное соединение представляет собой активный фрагмент MARCKS-белка. В другом аспекте указанное соединение представляет собой MANS-пептид.
В другом варианте осуществления настоящее изобретение описывает способ модуляции секреции слизи у пациента, включающий: введение терапевтического количества антисмысловой последовательности, которая является комплементарной последовательностям, кодирующим MARCKS-белок или его активный фрагмент, где секреция слизи указанной клеткой ингибируется по сравнению с таковой, которая возникает в отсутствие такой стимуляции. В одном аспекте настоящего изобретения указанная последовательность имеет по меньшей мере восемнадцать нуклеиновых кислот в длину. В другом аспекте указанное соединение комплементарно последовательностям, кодирующим MANS-пептид или его активный фрагмент. В другом аспекте, указанная модуляцией секреции слизи является блокада или уменьшение секреции слизи.
В другом варианте осуществления настоящее изобретение описывает способ уменьшения или ингибирования воспаления у пациента, включающий введение терапевтически эффективного количества, по меньшей мере, одного пептида, включающего MANS-пептид или его активный фрагмент, эффективно модулирующие медиатор воспаления в месте воспаления. В одном аспекте настоящего изобретения указанный активный фрагмент имеет в длину, по меньшей мере, шесть аминокислот. В другом аспекте указанные медиаторы воспаления продуцируются клетками, выбираемыми из группы, состоящей из нейтрофилов, базофилов, эозинофилов, моноцитов и лейкоцитов. Предпочтительно клетками являются лейкоциты, более предпочтительно - гранулоциты, и еще более предпочтительно - нейтрофилы, базофилы, эозинофилы или из комбинация. В другом аспекте агент вводят перорально, парентерально, в полости, ректально или посредством пассажа воздуха. В другом аспекте указанная композиция, кроме того, включает вторую молекулу, выбираемую из группы, состоящей из антибиотика, противовирусного соединения, антипаразитарного соединения и противовоспалительного соединения и иммуносупрессивного средства.
Активный фрагмент MANS-пептида может быть выбран из группы, состоящей из миристоилированных пептидов SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:7, SEQ ID NO:8, SEQ ID NO:9, SEQ ID NO:10, SEQ ID NO:11, SEQ ID NO:12, SEQ ID NO:13, SEQ ID NO: 14, SEQ ID NO:15, SEQ ID NO:16, SEQ ID NO:17, SEQ ID NO:18 и SEQ ID NO:19.
В другом аспекте настоящего изобретения способы, описанные в настоящем изобретении, могут быть осуществлены путем применения или введения комбинации пептидов, описанных в изобретении, т.е. путем применения или введения пептида, выбираемого из группы, состоящей из SEQ ID NO:1, SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:5, SEQ ID NO:6, SEQ ID NO:7, SEQ ID NO:8, SEQ ID NO:9, SEQ ID NO:10, SEQ ID NO:11, SEQ ID NO:12, SEQ ID NO:13, SEQ ID NO: 14, SEQ ID NO:15, SEQ ID NO:16, SEQ ID NO:17, SEQ ID NO:18, SEQ ID NO:19 и их комбинации. Предпочтительно в способах, описанных в настоящем описании, используют или вводят один пептид.
В ответ на активацию протеинкиназы С (РКС) стимулятором воспаления, дегрануляция клеток, выбираемых из группы, состоящей из нейтрофилов, эозинофилов, моноцитов/макрофагов и лимфоцитов, может быть уменьшена путем предварительной инкубации и совместной инкубации с пептидом, идентичным N-концевому участку MARCKS-белка, где пептид выбирают из группы, состоящей из MANS-пептида (SEQ ID NO:1) и его миристоилированных N-концевых фрагментов (SEQ ID NO:3-19). Хотя временные курсы и концентрации могут варьироваться среди различных типов клеток, во всех случаях MANS-пептид уменьшает дегрануляцию, индуцированную РКС.
Материалы и методы
Культура клеток NHBE - Рост, криоконсервацию и культивирование клеток NHBE на границе фаз воздух/жидкость проводили, как описано ранее. См. Krunkosky et al. Коротко, клетки NHBE (Clonetics, San Diego, Calif.) сеяли в вентилируемые колбы для культивирования тканей Т75 (500 клеток/см2) и культивировали, пока клетки не достигали 75-80% слияния. Затем клетки диссоциировали трипсином/ЭДТА и замораживали как пассаж-2. Культуру на границе раздела воздух/жидкость начинали путем посева клеток пассажа-2 (2×104 клеток/см2) в чистые культуральные вставки TRANSWELL® (Costar, Cambridge, Mass.), которые тонко покрывали слоем коллагена крысиных хвостов, тип I (Collaborative Biomedical, Bedford, Mass.). Клетки культивировали полупогруженными в среду в увлажненном 95% воздухе, окружении 5% СО2 в течение 5-7 дней почти до слияния. В этот момент создавали границу фаз воздуха/жидкости путем удаления верхушечной среды и питающих клеток базалатерально. После этого среду обновляли ежедневно. Клетки культивировали в течение дополнительных 14 дней для возможности полной дифференцировки.
Измерение секреции муцина при помощи ELISA - Перед сбором «исходных» и «тестируемых» образцов муцина накопившуюся слизь на апикальной поверхности клеток удаляли путем промывания фосфатным буферным раствором, рН 7,2. Для получения исходной секреции клетки инкубировали только со средой и секретируемый в апикальной среде муцин собирали и хранили. Клеткам давали отдохнуть в течение 24 ч и затем подвергали воздействию среды, содержащей выбранный стимулирующий и/или ингибирующий реагенты (или соответствующий контроль), после чего секретируемый муцин собирали и хранили в качестве тестируемого образца. Время инкубации для исходного и теста были одинаковыми, но варьировались в зависимости от используемого тестируемого реагента. И исходную, и тестируемую секрецию анализировали ELISA с использованием метода захвата антител, как известно в области техники. См., например, Harlow et al., Antibodies: A Laboratory Manual, pp. 570-573, Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. (1988). Первичным антителом для этого анализа было 17Q2 (Babco, Richmond, Calif.), моноклональное антитело, которое специфически реагирует с углеводородным эпитопом на муцинах дыхательных путей человека. Соотношение тестируемого/исходного муцина, подобное «секреторному индексу», использовали для количественного определения секреции муцина, позволяя каждой культуральной чашке служить ее собственным контролем и, следовательно, минимизировать отклонение, вызванное вариабельностью между культуральными ячейками. Wright et al., Am.J.Physiol. 271, L854-L861 (1996). Уровень секреции муцина указывали как процент от контроля среды.
Анализ иммунопреципитации с радиоактивной меткой - После метки [32Р]фосфатом, клетки предварительно инкубировали в течение 2 ч в бесфосфатной среде Eagle's, модифицированной Dulbecco, содержащей 0,2% бычьего альбумина сыворотки и затем метили 0,1 мКи/мл [32Р]ортофосфата (9000 Ки/ммоль, PerkinElmer Life Sciences) в течение 2 ч. Для метки [3Н]миристиновой кислотой или 3Н-аминокислотами, клетки инкубировали в среде, содержащей 50 мкКи/мл [3Н]миристиновой кислоты (49 Ки/ммоль, PerkinElmer Life Sciences) или 0,2 мКи/мл [3Н]лейцина (159 Ки/ммоль, PerkinElmer Life Sciences) плюс 0,4 мКи/мл [3Н]пролина (100 Ки/ммоль, PerkinElmer Life Sciences). После метки клетки подвергали воздействию стимулирующих реагентов в течение 5 мин. Когда использовали ингибитор, клетки предварительно инкубировали с ингибитором в течение 15 мин перед стимуляцией. В конце обработки клетки лизировали в буфере, содержащем 50 мМ Tris-HCl (рН 7,5), 150 мМ NaCl, 1 мМ ЭДТА, 10% глицерина, 1% Nonidet Р-40, 1 мМ фенилметилсульфонилфторида, 1 мМ бензамидина, 10 мкг/мл пепстатина А и 10 мкг/мл лейпептина. Путем осаждения трифторуксусной кислоты и счета сцинтилляции можно определить эффективность радиоактивной метки в каждой культуре. Иммунопреципитацию MARCKS-белка проводили в соответствии с Spizz and Blackshear с использованием лизатов клеток, содержащих равное количество импульсов/мин. Spizz et al., J.Biol.Chem. 271, 553-562 (1996). Осажденные белки разгоняли электрофорезом в 8% SDS-полиакриламидном геле и визуализировали авторадиографией. Для этого анализа использовали античеловеческое антитело к MARCKS (2F12) и неиммунное контрольное антитело (6F6).
Для оценки MARCKS или комплексов, ассоциированных с MARCKS-белком в различных субклеточных фракциях, меченные радиоактивной меткой и обработанные клетки соскабливали в буфере для гомогенизации (50 мМ Tris-HCl (рН 7,5), 10 мМ NaCl, 1 мМ ЭДТА, 1 мМ фенилметилсульфонилфторида, 1 мМ бензамидина, 10 мкг/мл пепстатина А, 10 мкг/мл лейпептина) и затем разрушали кавитацией азотом (800 фунтов на кв. дюйм в течение 20 мин при 4°С). Лизаты клеток центрифугировали при 600 х g в течение 10 мин при 4°С для удаления ядер и неразрушившихся клеток. Послеядерную надосадочную жидкость разделяли на мембранную и цитозольную фракции путем ультрацентрифугирования при 400000 х g в течение 30 мин при 4°С. Мембранный осадок растворяли в лизатном буфере путем обработки ультразвуком. Затем проводили иммунопреципитацию, как описано выше.
MARCKS-связанные пептиды - И миристоилированную N-концевую последовательность (MANS), и случайную N-концевую последовательность (RNS) пептидов синтезировали в Genemed Synthesis, Inc. (San Francisco, Calif.), затем очищали путем высокоэффективной жидкостной хроматографии (>95% чистоты) и для каждой подтверждали масс-спектроскопией, показывающей один пик с соответствующей молекулярной массой. MANS-пептид состоял из последовательности, идентичной первым 24 аминокислотам MARCKS, т.е. миристоилированного N-концевого участка, который опосредует вставку MARCKS в мембраны, МА-GAQFSKTAAKGEAAAERPGEAAVA (SEQ ID NO:1 (где МА = N-концевая миристатная цепь). Соответствующий контрольный пептид (RNS) содержал такую же аминокислотную композицию, как MANS, но расположенную в случайном порядке, MA-GTAPAAEGAGAEVKRASAEAKQAF (SEQ ID NO:2). Присутствие гидрофобного миристатного компонента в таких синтетических пептидах усиливает их проницаемость в плазматические мембраны, позволяя пептидам легко потребляться клетками. Для определения эффектов этих пептидов на секрецию муцина клетки предварительно инкубировали с пептидами в течение 15 мин перед добавлением стимуляторов секреции и затем секрецию муцина измеряли ELISA.
Антисмысловые нуклеотиды - MARCKS антисмысловой олигонуклеотид и его соответствующий контрольный олигонуклеотид синтезировали в Biognostik GmbH (Gottingen, Germany). Клетки NHBE обрабатывали 5 мкМ антисмыслового или контрольного олигонуклеотида апикально в течение 3 дней (в присутствии 2 мкг/мл липофектина в течение первых 24 ч). Затем клетки инкубировали стимуляторами секреции и секрецию муцина измеряли ELISA. Всю РНК и белок выделяли из обработанных клеток. мРНК MARCKS оценивали Нозерн гибридизацией в соответствии с обычными методиками с использованием человеческой кДНК MARCKS в качестве зонда. Уровень белка MARCKS определяли Вестерн блоттингом с использованием очищенного анти-MARCKS IgG1 (клон 2F12) в качестве первичного антитела определения.
Переходная трансфекция - Домен сайта фосфорилирования (PSD) MARCKS содержит РКС-зависимые сайты фосфорилирования и сайты связывания актиновых волокон. Для создания кДНК MARCKS без PSD полимеразной цепной реакцией создавали два фрагмента, ограничивающие последовательность PSD (кодирующие 25 аминокислот), и затем сшивали через XhоI участок, который прикрепляли к 5'-концам праймеров олигонуклеотидов, созданных для полимеразной цепной реакции. Полученную мутантную кДНК и кДНК MARCKS дикого типа каждую вставляли в вектор экспрессии млекопитающих pcDNA4/TO (Invitrogen, Carlsbad, Calif.). Выделенные рекомбинантные конструкции подтверждали рестрикционным картированием и секвенированием ДНК.
HBE1 представляет собой клеточную линию эпителия бронхов человека, трансформированного папиллома-вирусом, способную к секреции муцина при культивировании на границе фаз воздух/жидкость. Трансфекцию клеток HBE1 проводили с использованием реагента трансфекции Effectene (Qiagen, Valencia, Calif.) в соответствии с инструкциями производителя. Коротко, дифференцированные клетки HBE1, выращенные на границе фаз воздух/жидкость, диссоциировали трипсином/ЭДТА и заново сеяли в 12-луночные культуральные планшеты по 1×105 клеток/см2. После инкубации в течение ночи клетки трансфицировали кДНК MARCKS дикого типа, кДНК MARCKS без PSD или векторной ДНК. Клетки культивировали в течение 48 ч для возможности экспрессии гена и затем подвергали воздействию стимуляторов секреции, и секрецию муцина измеряли ELISA. Все трансфекции проводили в присутствии плазмиды pcDNA4/TO/lacZ (Invitrogen) (соотношение ДНК 6:1, всего 1 мкг ДНК, соотношение ДНК и реагента Effectene = 1:25) для отслеживания вариантов эффективности трансфекции. Результаты показали отсутствие достоверных различий в бета-галактозидазной активности в лизатах клеток, выделенных из трансфицированных клеток, показывая сходную эффективность трансфекции между различными конструкциями ДНК (данные не показаны).
Анализ активности протеинфосфатазы - Активность РР1 и РР2А измеряли с использованием системы анализа протеинфосфатазы (Life Technologies, Inc.), как известно в области техники с небольшими модификациями. Huang et al., Adv.Exp. Med. Biol. 396, 209-215 (1996). Коротко, клетки NHBE обрабатывали 8-Br-cGMP или средой отдельно в течение 5 мин. Затем клетки соскребали в лизатный буфер (50 мМ Tris-HСl (рН 7,4), 0,1% бета-меркаптоэтанола, 0,1 мМ ЭДТА, 1 мМ бенаамидина, 10 мкг/мл пепстатина А, 10 мкг/мл лейпептина) и разрушали путем обработки ультразвуком в течение 20 с при 4°С. Лизаты клеток центрифугировали и надосадочную жидкость сохраняли для анализа активности фосфатазы. Анализ проводили с использованием 32Р-меченной фосфорилазы А в качестве субстрата. Высвобожденный 32Рi подсчитывали сцинтилляцией. Концентрацию белка в каждом образце определяли анализом Bradford. Активность РР2А выражали как общую фосфатазную активность образца минус активность, остающуюся в присутствии 1 нМ окадаевой кислоты. Активность РР1 выражали как разницу между активностями, оставшимися в присутствии 1 нМ и 1 мкМ окадаевой кислоты, соответственно. Активность протеинфосфатазы указывали как нмоль Рi высвобожденного в мин/мг общего белка.
Анализ цитотоксичности - Все реагенты, используемые в обработке клеток NHBE, изучали в отношении цитотоксичности путем измерения общего высвобождения лактатдегидрогеназы из клеток. Анализ проводили с использованием набора Promega Cytotox 96 в соответствии с инструкциями производителя. Все эксперименты проводили с реагентами в нетоксичных концентрациях.
Статистический анализ - Данные анализировали в отношении статистической значимости с использованием однофакторного дисперсионного анализа с поправкой Бонферрони. Различия между обработками расценивали статистически значимыми при р<0,05.
Выделение PMN из крови собак - Стадии, вовлеченные в выделение PMN, включают получение 10 мл ACD антикоагулированной крови. Затем проводили наслаивание 5 мл на 3,5 мл среды для выделения PMN, при этом обеспечивая комнатную температуру (RI) среды для выделения PMN (IM). Затем кровь цетнтрифугировали при комнатной температуре в течение 30', 550 х g при 1700 об/мин. Самую нижнюю белую полоску переносили в 15 мл коническую пробирку для центрифугирования (CCFT). Затем добавляли 2 объема HESS с 10% эмбриональной телячьей сывороткой (PBS) и центрифугировали при комнатной температуре в течение 10', 400 х g при 1400 об/мин. Затем осадок ресуспендировали в 5 мл 1-1ESS с PBS. Суспензию клеток добавляли к 50 мл CCFT, содержащей 20 мл ледяной 0,88% NH4Cl и переворачивали два-три раза. Полученный продукт центрифугировали в течение 10', 800 х g при 2000 об/мин, затем аспирировали и ресуспендировали в 5 мл HBSS c FBS. Препарат исследовали подсчетом и цитоцентрифугированием, и предпочтительно для цельной крови, количество клеток должно составлять между 109-1011 клеток, и для PMN количество клеток должно составлять между 2-4 × 107 клеток. См. в общем, Wang et al., J.Immunol., «Neutrophil-induced changes in the biomechanical properties of endothelial cells: roles of ICAM-1 and reactive oxygen species”, 6487-94 (2000).
Колориметрический ферментный анализ МРО - Образцы оценивали в отношении активности МРО в 96-луночных круглодонных микротитрационных планшетах с использованием набора для сэндвич ELISA (R&D Systems, Minneapolis, Minn.). Коротко, 20 микролитров образца смешивали с 180 микролитрами субстратной смеси, содержащей 33 мМ фосфата калия, рН 6,0, 0,56% Triton X-100, 0,11 мМ перекиси водорода и 0,36 мМ дигидрохлорида О-дианнизидина в отдельной микротитрационной ячейке. Конечные концентрации в анализируемой смеси были: 30 мМ фосфата калия, рН 6,0, 0,05% Triton X-100, 0,1 мМ перекиси водорода и 0,32 мМ дигидрохлорида О-дианнизидина. После смешивания анализируемую смесь инкубировали при комнатной температуре в течение 5 минут, и ферментную активность МРО определяли спектрофотометрически при 550 нанометрах. Образцы оценивали в двойной повторности.
Исследования секреции медиатора воспаления
Использовали четыре различных типа лейкоцитов или моделей, которые секретируют специфическое содержимое гранул в ответ на индуцированную форболовым эфиром активацию РКС. Нейтрофилы изолировали из человеческой крови и in vitro оценивали высвобождение такими клетками МРО. Также оценивали высвобождение связанных с мембраной медиаторов воспаления на клеточных линиях коммерчески доступных человеческих лейкоцитов. Клеточную линию человеческих промиелоцитов HL-60 клон 15 использовали для оценки секреции ЕРО (Fischkoff SA. Graded increase in probability of eosinophilic differentiation of HL-60 promyelocytic leukemia cells induced by culture under alkaline conditions. Leuk Res 1988; 12: 679-686; Rosenberg HF., Ackerman SJ, Tenen DG. Human eosinophil cationic protein: molecular cloning of a cytotoxin and helminthotoxin with ribonuclease activity. J Exp Med 1989; 170: 163-176; Tiffany HL, Li F., Rosenberg HF. Hyperglycosylation of eosinophil ribonucleases in a promielocytic leukemia cell line and in differentiated peripheral blood progenitor cells. J Leukoc Biol 1995; 58:49-54; Badewa AP, Hudson CE, Heiman AS. Regulatory effects of eotaxin, eotaxin-2, and eotaxin-3 on eosinophil degranulation and superoxide anion generation. Exp Biol Med 2002; 227: 645-651). Клеточную линию моноцитарного лейкоза U937 использовали для оценки секреции лизозима (Hoff T, Spencker T, Emmendoerffer A., Goppelt-Struebe M. Effects of glucocorticoids on the TPA-induced monocytic differentiation J Leukoc Biol 1992; 52: 173-182; Balboa MA, Saez Y, Balsinde J. Calcium-independent phospholipase A2 is required for lysozyme secretion in U937 promonocytes. J Immunol 2003; 170: 5276-5280; Sundstrom C, Nilsson K. Establishment and characterization of a human histiocytic lymphoma cell line (U937). Int J Cancer 1976; 17: 565-577). Клеточную линию лимфоцитарных NK-клеток NK-92 использовали для оценки высвобождения гранзима (Gong JH., Klingemann HG. Characterization of a human cell line (NK-92) with phenotypical and functional characteristics of activated natural killer cells. Leukemia 1994; 8:652-658; Maki G, Klingemann HG, Martinson JA, Tam YK. Factors regulating the cytotoxic activity of the human natural killer cell line, NK-92. J Hematother Stem Cell Res 2001; 10: 369-383; Takayama H, Trenn G, Sitkovsky MV. A novel cytotoxic T lymphocyte activation assay. J Immunol Methods 1987; 104: 183-190). Во всех случаях клетки предварительно инкубировали с рядом концентраций синтетического пептида, идентичного 24 аминокислотам N-конца MARCKS [MANS-миристоилированный пептид N-концевой последовательности; MA-GAQFSKTAAKGEAAAERPGEAAVA (SEQ ID NO: 1)], где МА представляет собой миристоил, прикрепленный к N-концевому амину пептида амидной связью), или миссенс-контрольным пептидом (RNS: случайный пептид N-концевой последовательности; MA-GTAPAAEGAGAEVKRASAEAKQAF (SEQ ID NO: 2)], который состоит из таких же 24 аминокислот, но расположенных в порядке случайной последовательности, которая обладает менее чем 13% сходства последовательности с последовательностью MANS-пептида.
В каждом из типов клеток, MANS, но не RNS уменьшал высвобождение воспалительных медиаторов в зависимости от концентрации. Полезный период времени наблюдения составил 0,5-3,0 часа. Результаты согласуются с вовлечением N-концевого участка MARCKS-белка во внутриклеточные пути, ведущие к дегрануляции лейкоцитов.
Выделение человеческих нейтрофилов - Эти исследования были одобрены NCSU human studies Institutional Review Board (IRB). Человеческие нейтрофилы выделяли, как описано ранее (см. Takashi S, Okubo Y, Horie S. Contribution of CD54 to human eosinophil and neutrophil superoxide production. J Appl Physiol 2001; 91:613-622) с небольшими модификациями. Коротко, гепаринизированную венозную кровь получали от нормальных здоровых добровольцев, разводя RPMI-1640 (Cellgro; Mediatech, Inc., Herndon, VA) в соотношении 1:1, наслоенную на Histopaque (плотность, 1,077 г/мл; Sigma-Aldrich Co., St. Louis, MO) и центрифугировали при 400 g в течение 20 мин при 4°С. Надосадочную жидкость и мононуклеарные клетки в промежуточной фазе аккуратно удаляли, и эритроциты в осадке лизировали в охлажденной дистиллированной воде. Выделенные гранулоциты промывали два раза сбалансированным солевым раствором Хэнкса (HBSS) и ресуспендировали в HBSS на льду. Нейтрофилы, используемые для экспериментов, имели >98% чистоты с <2% загрязнением эозинофилами и выживаемость составила >99%, что определяли исключением красителя трипана синего.
Измерение активности высвобожденной нейтрофильной МРО - Для измерения высвобождения МРО очищенные человеческие нейтрофилы суспендированные в HBSS, разделяли на аликвоты по 4×106 клеток/мл в 15 мл пробирки и предварительно инкубировали или с 50 или с 100 мкМ MANS или RNS пептида в течение 10 мин при 37°С. Затем клетки стимулировали 100 нМ 12-миристат 13-ацетата форбола (РМА) в течение до 3 часов. Контрольный образец (РМА контрольный образец) получали с использованием очищенных человеческих нейтрофилов, суспендированных в HBSS, разделенных на аликвоты по 4×106 клеток/мл в 15 мл пробирках и стимулированных 100 нМ 12-миристат 13-ацетатом форбола (РМА) в отсутствие MANS или RNS пептида в течение таких же периодов времени. Реакцию терминировали путем помещения пробирок на лед и центрифугированием при 400 g в течение 5 мин при 4°С.
Активность МРО в надосадочной жидкости клеток оценивали с использованием тетраметилбензидина (ТМВ) на основании ранее установленной методики (Abdel-Latif D, Steward M, Macdonald DL, Francis GA, Dinauer MC, Lacy P. Rac2 is critical for neutrophil primary granule exocytosis. Blood 2004; 104:832-839). Коротко, 100 мкл ТМВ субстратного раствора добавляли к 50 мкл надосадочной жидкости клеток или стандартной человеческой МРО (EMD Biosciences, Inc., San Diego, CA) в 96-луночном микропланшете с последующей инкубацией при комнатной температуре в течение 15 мин. Реакцию терминировали добавлением 50 мкл 1М H2SO4 и поглощение считывали при 450 нм в спектрофотометрическом счетчике микропланшетов (VERSA max, Molecular Devices, Sunnyvale, CA).
Исследования культур лейкоцитов
Три типа клеточных линий человеческих лейкоцитов, в частности клеточную линию промиелоцитов HL-60 клон 15, клеточную линию моноцитов U937 и клеточную линию лимфоцитарных NK-клеток NK-92 покупали у American Type Culture Colleation (ATCC; Rockville, MD). Клетки HL-60 клон 15 (ATCC CRL-1964) выдерживали в среде, состоящей из RPMI1640 с L-глютамином, дополненной 10% теплоинактивированной бычьей эмбриональной сыворотки (FBS; Gibco; Invitrogen Co., Carlsbad, CA), 50 МЕ/мл пенициллина, 50 мкг/мл стрептомицина и 25 мМ буфера HEPES, рН 7,8 при 37°С в атмосфере, содержащей 5% СО2. Окончательную дифференцировку в эозинофил-подобный фенотип начинали путем культивирования клеток по 5×105 клеток/мл в вышеуказанной среде, содержащей 0,5 мМ масляной кислоты (Sigma-Aldrich Co.) в течение 5 дней как описано ранее (Tiffany HL, Li F, Rosenberg HF. Hyperglycosylation of eosinophil ribonucleases in promyelocytic leukemia cell line and in differentiated peripheral blood progenitor cells. J Leukoc Biol 1995; 58:49-54; Tiffany HL, Alkhatib G, Combadiere C, Berger EA, Murphy PM. CC chemokine receptors 1 and 3 are differentially regulated by IL-5 during maturation of eosinophilic HL-60 cells. J Immunol 1998; 160: 1385-1392). Клетки U937 (ATCC CRL-1593.2) выращивали при 37°С в атмосфере 5% СО2 в полной среде, состоящей из RPMI 1640 с L-глютамином, дополненной 10% FBS, 50 МЕ/мл пенициллина и 50 мкг/мл стрептомицина. NK-92 клетки (ATCC CRL-2407) поддерживали в альфа-МЕМ среде (Sigma-Aldrich Co.), дополненной 20% FBS, 100 Ед/мл интерлейкина-2 (IL-2) (Chemicon International, Inc., Temecula, CA), 5×10-5 М 2-меркаптоэтанола, 50 МЕ/мл пенициллина, и 50 мкг/мл стрептомицина при 37°С в атмосфере, содержащей 5% СО2. Клеточную морфологию изучали путем оценки клеток, окрашенных по Райт-Гимсу. Вариабельность клеток, полученных для экспериментов, оценивали по исключению трипана синего и использовали популяции клеток с выживаемостью >95%.
Инкубация клеток для анализа дегрануляции
Клетки HL-60 клон 15, U937 и NK-92 промывали и ресуспендировали по 2,5×106 клеток/мл в феноле красном-свободном RPMI-1640 (Cellgro; Mediatech, Inc.) для всех анализов дегрануляции. Аликвоты клеток в 15 мл пробирках предварительно инкубировали с указанными концентрациями MANS или RNS пептида в течение 10 мин при 37°С. Затем клетки стимулировали РМА в течение до 2 ч. Контрольный образец (РМА контрольный образец) устанавливали для каждого типа клеток с использованием клеток HL-60 клон 15, U937 и NK-92, соответственно, которые промывали и ресуспендировали по 2,5×106 клеток/мл в RPMI-1640 без фенола красного и стимулировали РМА, но в отсутствие MANS или RNS пептида в течение таких же периодов времени. Реакцию терминировали путем помещения пробирок на лед и центрифугирования клеток при 400 g в течение 5 мин при 4°С.
Для измерения высвобожденной МРО из нейтрофилов и высвобожденного лизозима из клеток U937, авторы смогли количественно определить секрецию с использованием в качестве стандартов человеческой МРО и овальбумина яичного белка, соответственно. Для высвобожденной ЕРО из клеток HL-60 клон 15 и для высвобожденного гранзима из NK-92 клеток для использования при количественном определения не было доступных стандартов. Следовательно, измеряли уровень и высвобожденных и внутриклеточных (из лизированных клеток) ЕРО и гранзима и высвобожденный ЕРО и гранзим выражали как процент общего (внутриклеточного и высвобожденного) для каждого. Для измерения внутриклеточной ЕРО в клетках HL-60 клон 15 и внутриклеточного гранзима в NK-92 клетках, для количественного определения белков внутриклеточных гранул получали соответствующие аликвоты клеток, лизированных 0,1% тритон Х-100, как описано ниже. Все обработки выражали как процент от контроля для минимизации вариабельности между культурами.
Измерение ЕРО высвобождения HL-60
Активность ЕРО, высвобожденной клетками HL-60 клон 15, оценивали с использованием ТМВ в соответствии с ранее указанной методикой (Lacy P, Mahmudi-Azer S, Bablitz B, Hagen SC, Velazquez JR, Man SF, Moqbel R. Rapid mobilization of intercellularly stored RANTES in response to interferon-gamma in human eosinophils. Blood 1999; 94:23-32). Таким образом, 100 мкл субстратного раствора ТМВ добавляли к 50 мкл (мкл = микролитры) образца в 96-луночный микропланшет и инкубировали при комнатной температуре в течение 15 мин (мин = минуты). Реакцию прекращали добавлением 50 мкл 1,0 М H2SO4 и поглощение считывали при 450 нм (нм = нанометры) в спектрофотометрическом счетчике микропланшетов. Количестве секретируемой ЕРО выражали как процент от общего содержания с использованием количества, полученного в таком же количестве клеток, лизированных тритон Х-100.
Измерение секреции лизозима моноцитами
Лизозим, секретируемый клетками U937, измеряли с использованием спектрофотометрического анализа, как описано ранее (Balboa MA, Saez Y, Balsinde J. Calcium-independent phospholipase A2 is required for lysozyme secretion in U937 promonocytes. J Immunol 2003; 170: 5276-5280) с небольшими модификациями. Таким образом, 100 мкл образца смешивали с 100 мкл суспензии Micrococcus lysodeikticus (Sigma-Aldrich Co) (0,3 мг/мл в 0,1 М натриевого фосфатного буфера, рН 7,0) в 96-луночном микропланшете. Снижение поглощения при 450 нм измеряли при комнатной температуре. Калибровочную кривую создавали с использованием лизозима белка куриного яйца (EMD Biosciences, Inc.) в качестве стандарта.
Измерение секреции гранзима NK-клетками
Гранзим, секретируемый NK-92 клетками оценивали путем измерения гидролиза Nα-бензилоксикарбонил-L-лизинтиобензилового эфира (BLT) по существу как описано ранее (Takayama H, Trenn G, Sitkovsky MV. A novel cytotoxic T lymphocyte activation assay. J Immunol Methods 1987; 104: 183-190). 50 мкл надосадочной жидкости переносили в 96-луночный планшет и к надосадочной жидкости добавляли 150 мкл раствора BLT (0,2 мМ BLT; EMD Biosciences, Inc., и 0,22 мМ DTNB; Sigma-Aldrich Co.) (мМ = миллимолярный) в фосфатном буферном растворе (PBS, рН 7,2). Поглощение при 410 нм считывали после инкубации в течение 30 мин при комнатной температуре. Результаты выражали как процент от общего содержания фермента в клетке с использованием количества, полученного в таком же количестве клеток, лизированных тритон Х-100.
Статистический анализ
Статистическую значимость различий между различными группами обработки оценивали с помощью одностороннего ANOVA. Значения Р<0,05 брали как статистически значимые.
Высвобождение МРО нейтрофилами
Было обнаружено, что 100 нМ РМА (в качестве стимулятора высвобождения медиаторов воспаления) увеличивало высвобождение МРО человеческими нейтрофилами приблизительно в три раза по сравнению с контрольным уровнем через 30 мин в контрольном образце РМА, высвобождение МРО увеличивалось приблизительно в 5-6 раз через 3 часа. Через 30 минут относительно контрольной активности МРО в качестве 100% при отсутствии РМА и отсутствии РМА плюс MANS или RNS активность МРО РМА контрольного образца составила около 275%, РМА плюс 50 мкМ MANS составило около 275% и 100 мкМ MANS составило около 305%. Следовательно, MANS-пептид не оказывал определяемого эффекта через 30 мин. Однако, через 1 ч наивысшая концентрация MANS (100 мкМ) оказывала значительный ингибирующий эффект (измеряемый как около 260% контроля) или около 25% снижения высвобождения МРО по сравнению с уровнем РМА контрольного образца (который измеряли как около 340% контроля). 50 мкМ образец MANS измеряли около 290% контроля или около 15% снижения относительно РМА контрольного образца. Через 2 ч и продолжаясь через 3 ч, MANS-пептид существенно снижал активность МРО в зависимости от концентрации. Через 2 часа, МРО активность РМА контрольного образца составила около 540% контроля, 50 мкМ MANS (измеряемого около 375% контроля) вызывала приблизительно 30% снижение высвобождения МРО по сравнению с РМА контрольным образцом; и 100 мкМ MANS (измеряя около 295% контроля) вызывали приблизительно 45% снижение высвобождения МРО по сравнению с РМА контрольным образцом. Через 3 часа, активность МРО РМА контрольного образца составила около 560% контроля, 50 мкМ MANS (измеряя около 375% контроля) вызывало приблизительно 33% снижения высвобождения МРО по сравнению с РМА контрольным образцом; 100 мкМ MANS (измеряя около 320% контроля) вызывало приблизительно 40% снижения высвобождения МРО по сравнению с РМА контрольным образом. RNS-пептид не влиял на высвобождение МРО, индуцированное РМА, в любую из временных точек или тестируемых концентраций.
Высвобождение ЕРО HL-60
Активность ЕРО в надосадочной жидкости клеток HL-60 клон 15 была значительно усилена через 1 и 2 часа после стимуляции РМА. Через 1 час относительно активности ЕРО-контроля в качестве 100%, РМА контрольного образца, измеряли около 110%; образца, содержащего 10 мкМ MANS, измеряли около 95% с получением около 15% снижения активности ЕРО относительно РМА контрольного образца: образца, содержащего 50 мкМ MANS, измеряли около 78% с получением снижения на около 30% активности ЕРО относительно РМА контрольного образца; и образца, содержащего 100 мкМ MANS, измеряли как около 65% с получением около 40% снижения ЕРО-активности относительно РМА контрольного образца. Через 2 часа, относительно EPO-активности контроля как 100%, РМА контрольный образец измеряли как около 145%; образец, содержащий около 10 мкМ MANS, измеряли как 130%, получая около 10% снижения EPO-активности относительно РМА контрольного образца; образец, содержащий 50 мкМ MANS, измеряли около 70% с получением около 50% снижения EPO-активности относительно РМА контрольного образца; и образец, содержащий 100 мкМ MANS, измеряли около 72% с получением около 50% снижения EPO-активности относительно РМА контрольного образца. Следовательно, и через 1, и через 2 часа MANS по 50 или 100 мкМ существенно снижал высвобождение ЕРО. RNS пептид не влиял на высвобождение ЕРО, усиленное РМА, в любой период времени или при любой исследуемой концентрации.
Высвобождение лизозима U937
Секреция лизозима U937 клетками увеличивалась стимуляцией РМА через 1 час после инкубации и увеличивалась еще больше через 2 ч. Через 1 час, относительно секреции лизозима U937 клетками контроля как 100%, РМА контрольный образец измеряли как около 210%; образец, содержащий 10 мкМ MANS, измеряли как около 170% с получением около 20% снижения секреции лизозима клетками U937 относительно РМА контрольного образца; образец, содержащий 50 мкМ MANS, измеряли как около 170% с получением около 20% снижения секреции лизозима клетками U937 относительно РМА контрольного образца; образец, содержащий 100 мкМ MANS, измеряли как около 115% с получением около 45% снижения секреции лизозима клетками U937 относительно РМА контрольного образца. Через 2 часа, относительно секреции лизозима клетками U937 контроля как 100%, РМА контрольный образец измеряли как около 240%; образец, содержащий 10 мкМ MANS, измеряли как около 195% с получением около 20% снижения секреции лизозима клетками U937 относительно РМА контрольного образца; образец, содержащий 50 мкМ MANS, измеряли как 185% с получением около 25% снижения секреции лизозима клетками U937 относительно РМА контрольного образца; образец, содержащий 100 мкМ MANS, измеряли около 140% с получением около 40% снижения секреции лизозима клетками U937 относительно РМА контрольного образца. Следовательно, секреция лизозима значительно снижалась через и 1, и 2 часа после стимуляции 100 мкМ MANS, но не настолько как 50 млм 10 мкМ MANS. RNS пептид не влиял на секрецию лизозима, усиленную РМА, в любой период времени или при любой исследуемой концентрации.
Высвобождение гранзима NK-клетками
Через 1 час, относительно секреции гранзима клетками NK-92 контроля в качестве 100%, РМА контрольный образец измеряли как около 125%; образец, содержащий 10 мкМ MANS, измеряли как около 115% с получением около 10% снижения секреции гранзима клетками NK-92 относительно РМА контрольного образца; образец, содержащий 100 мкМ MANS, измеряли около 85% относительно РМА контрольного образца с получением около 30% снижения секреции гранзима клетками NK-92 относительно РМА контрольного образца. Через 2 часа, относительно секреции гранзима клетками NK-92 контроля в качестве 100%, РМА контрольный образец измеряли как около 220%; образец, содержащий 10 мкМ MANS, измеряли как около 200% с получением около 10% снижения секреции гранзима клетками NK-92 относительно РМА контрольного образца; образец, содержащий 100 мкМ MANS, измеряли как около 80% с получением около 60% снижения секреции гранзима клетками NK-92 относительно РМА контрольного образца. Следовательно, секреция гранзима клетками NK-92 существенно не увеличивалась РМА через 1 ч, но увеличивалась более двух раз через 2 часа. 100 мкМ MANS, но не 10 мкМ MANS снижали секрецию гранзима через 1 и 2 часа после инкубации. RNS-пептид не влиял на секрецию гранзима, усиленную РМА, в любой период времени или при любой исследуемой концентрации.
Цитотоксичность
Ни одна из обработок не создавала токсического ответа в клетках, что оценивали путем удержания/высвобождения ЛДГ (данные не показаны) (см. также Park J-A, He F, Martin LD, Li Y, Adler KB. Human neutrophil elastase induces hypersecretion of mucin from human bronchial epithelial cells in vitro via a PKCδ - mediated mechanism. Am.J Pathol 2005; 167:651-661).
Вышеуказанные примеры являются иллюстрирующими настоящее изобретение и не должны рассматриваться как ограничивающие его. Изобретение определено следующей формулой изобретения с включением в нее эквивалентов формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ ОСЛАБЛЕНИЯ ВЫСВОБОЖДЕНИЯ МЕДИАТОРОВ ВОСПАЛЕНИЯ И ПЕПТИДЫ, ПРИМЕНЯЕМЫЕ ДЛЯ ЭТОЙ ЦЕЛИ | 2007 |
|
RU2423376C2 |
| ИНГИБИТОРЫ МЕТАСТАЗИРОВАНИЯ | 2014 |
|
RU2681213C2 |
| ИНГИБИТОРЫ ГИПЕРСЕКРЕЦИИ МУЦИНА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2006 |
|
RU2423379C2 |
| ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ ДЛЯ ИНГИБИРОВАНИЯ ВОСПАЛИТЕЛЬНЫХ ЦИТОКИНОВ | 2019 |
|
RU2816868C2 |
| АНТИБАКТЕРИАЛЬНЫЕ ПЕПТИДЫ | 2007 |
|
RU2468033C2 |
| УЛУЧШЕННЫЕ СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЗАЖИВЛЕНИЯ РАН | 2007 |
|
RU2542471C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ПЕПТИД APL-ТИПА | 2018 |
|
RU2778402C2 |
| ПРОДУКТ И СПОСОБ ДЛЯ НОРМАЛИЗАЦИИ ВЯЗКОСТИ СЛИЗИ | 2014 |
|
RU2750211C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ПРОФИЛАКТИКИ ИЛИ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ, СОСТОЯНИЙ ИЛИ ПРОЦЕССОВ, ХАРАКТЕРИЗУЕМЫХ АБЕРРАНТНОЙ ПРОЛИФЕРАЦИЕЙ ФИБРОБЛАСТОВ И ОТЛОЖЕНИЕМ ВНЕКЛЕТОЧНОГО МАТРИКСА | 2012 |
|
RU2620066C2 |
| ПРИМЕНЕНИЕ ГЕПАРИНСВЯЗЫВАЮЩИХ АНТАГОНИСТОВ В ИНГИБИРОВАНИИ ВЫСВОБОЖДЕНИЯ БРАДИКИНИНА | 2000 |
|
RU2251433C2 |










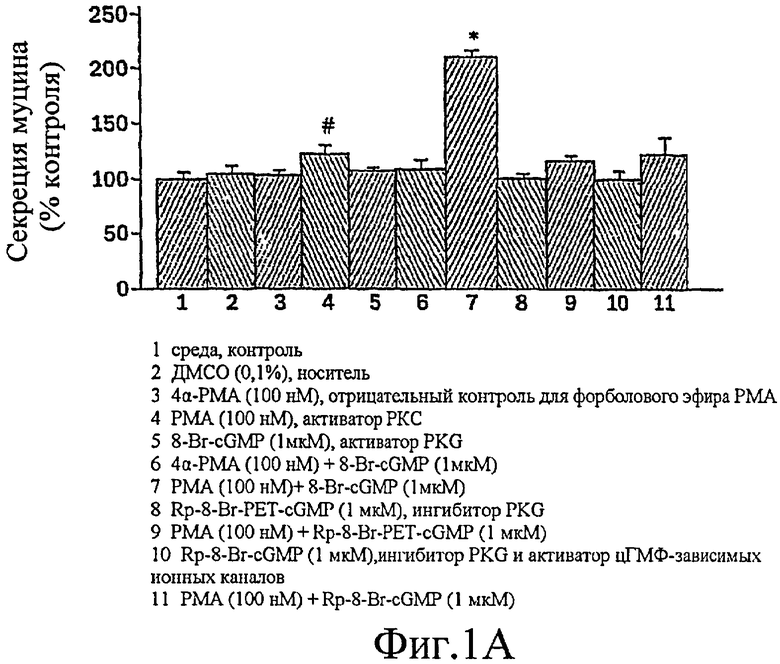
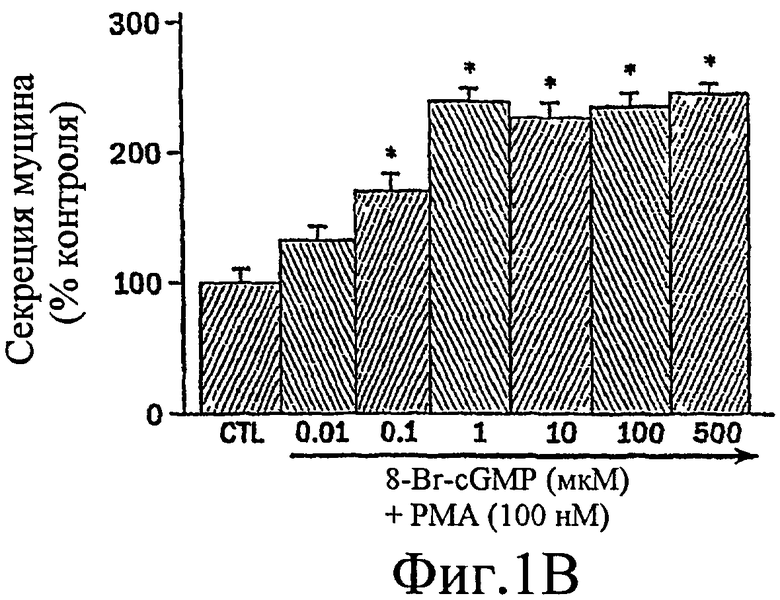
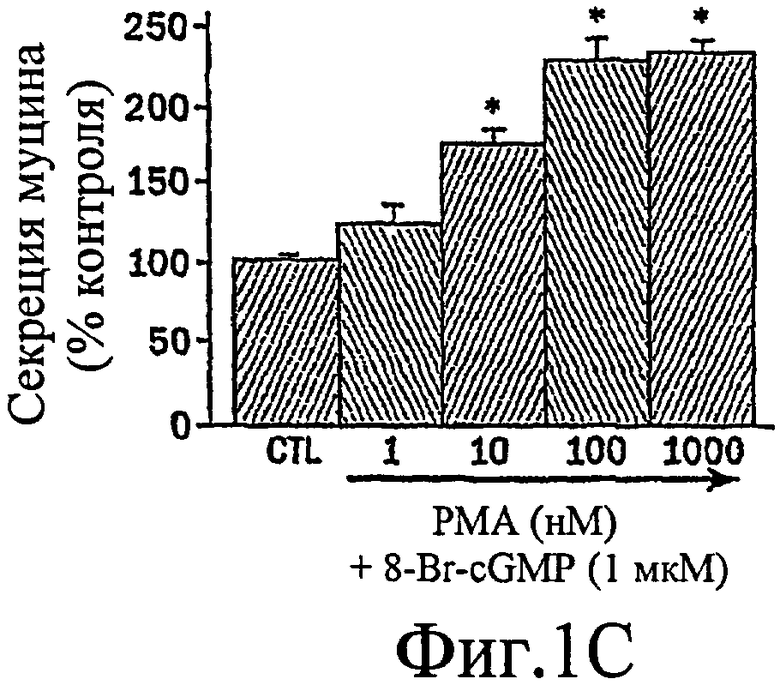
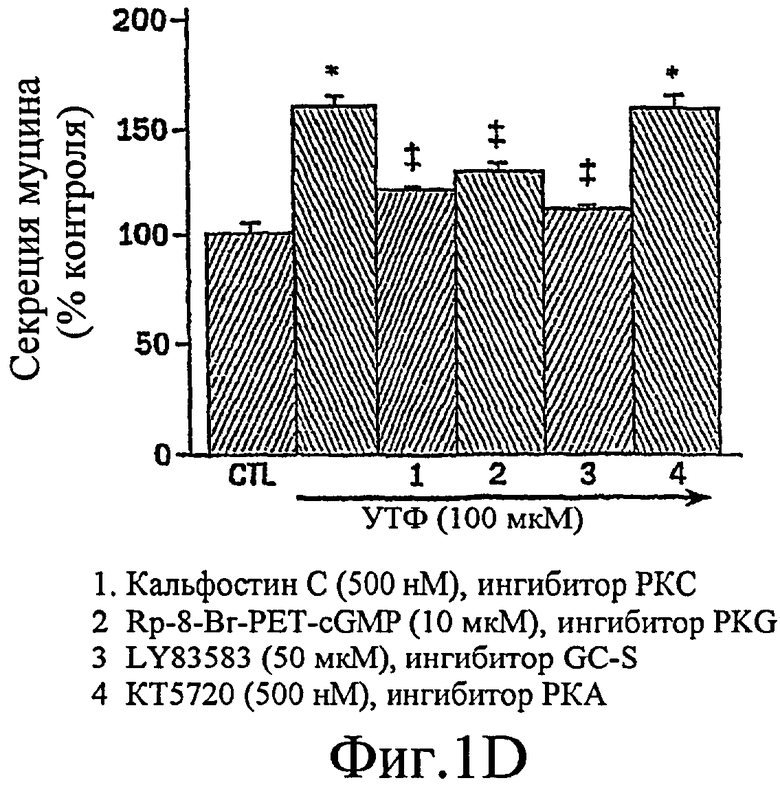

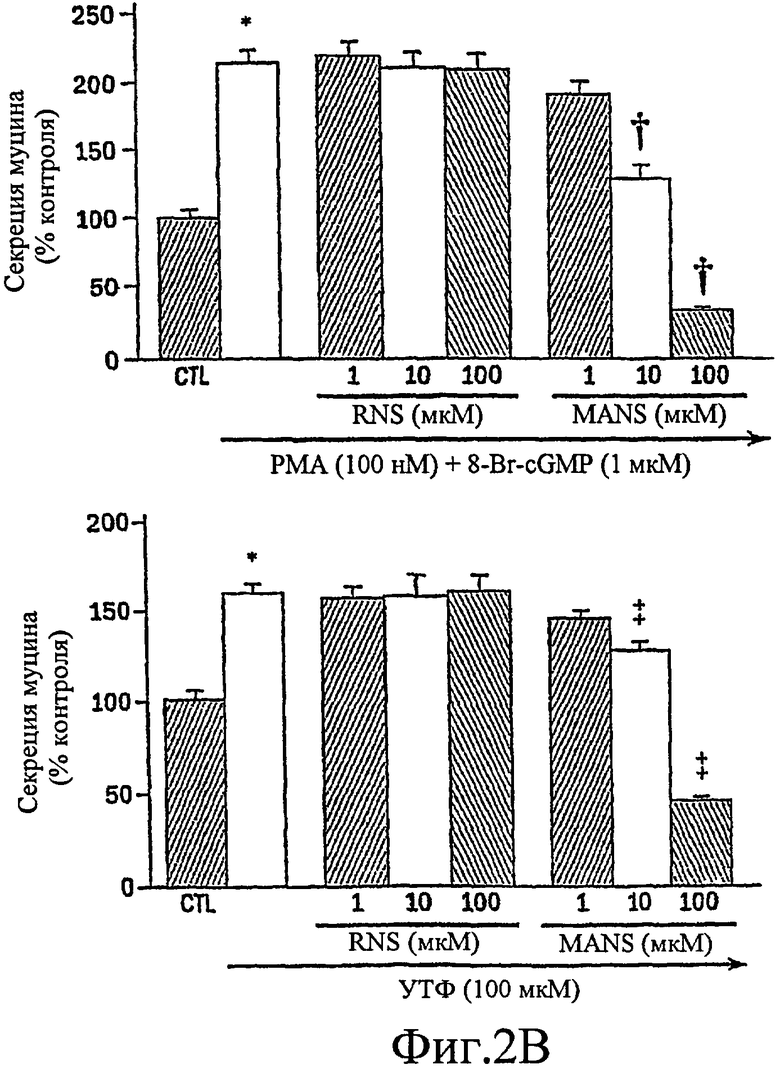
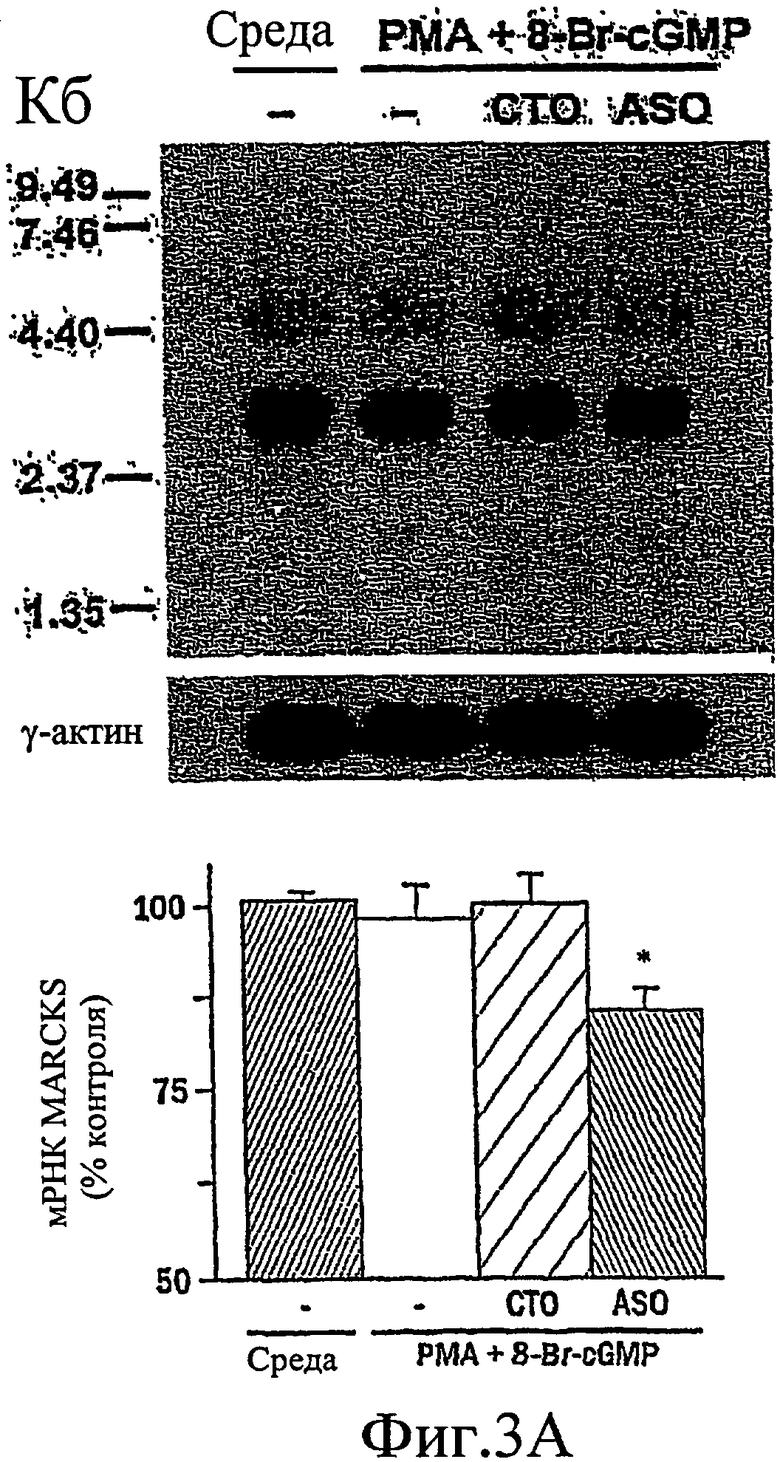
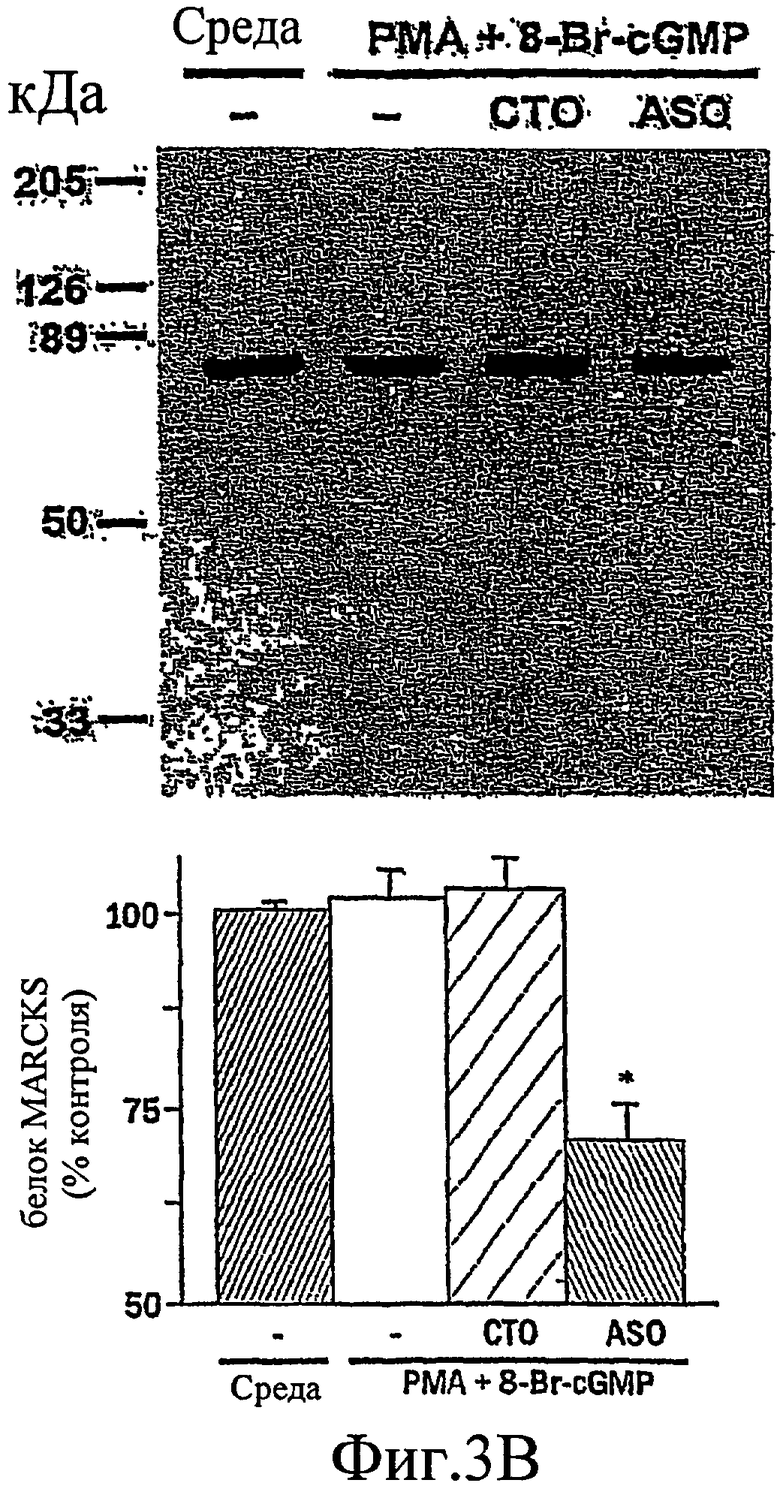
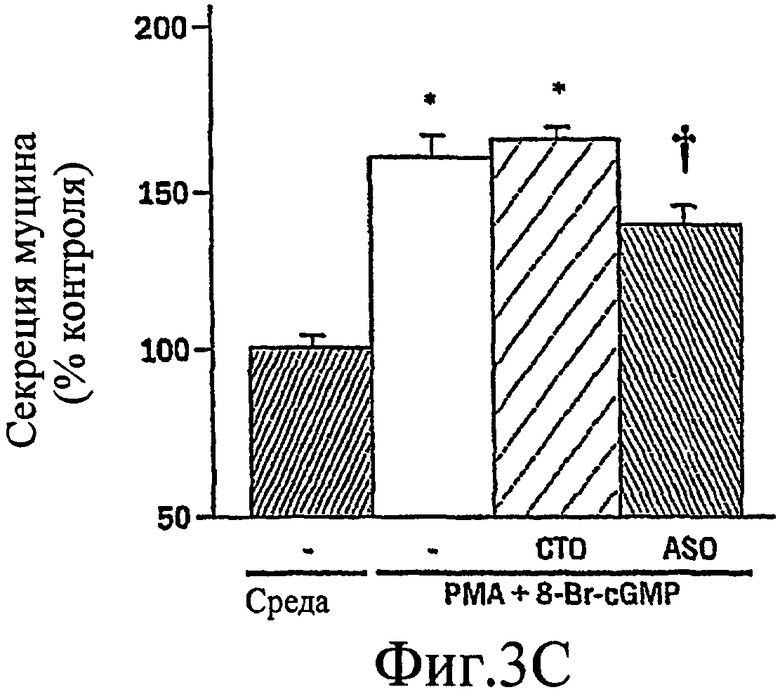
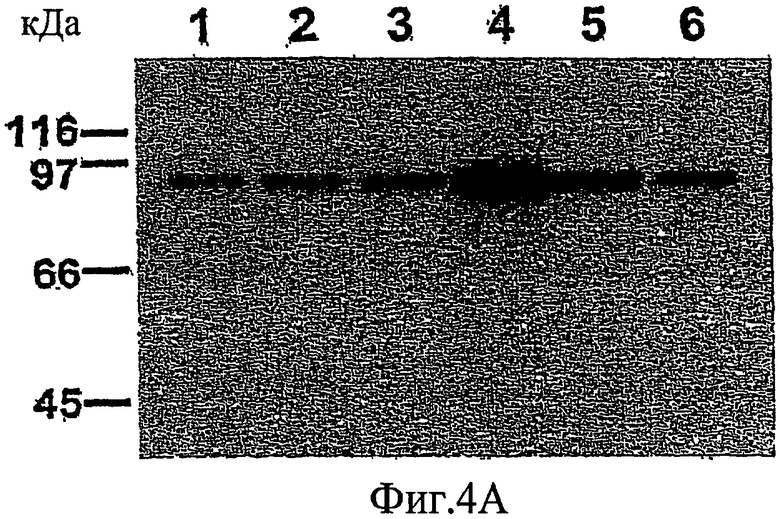
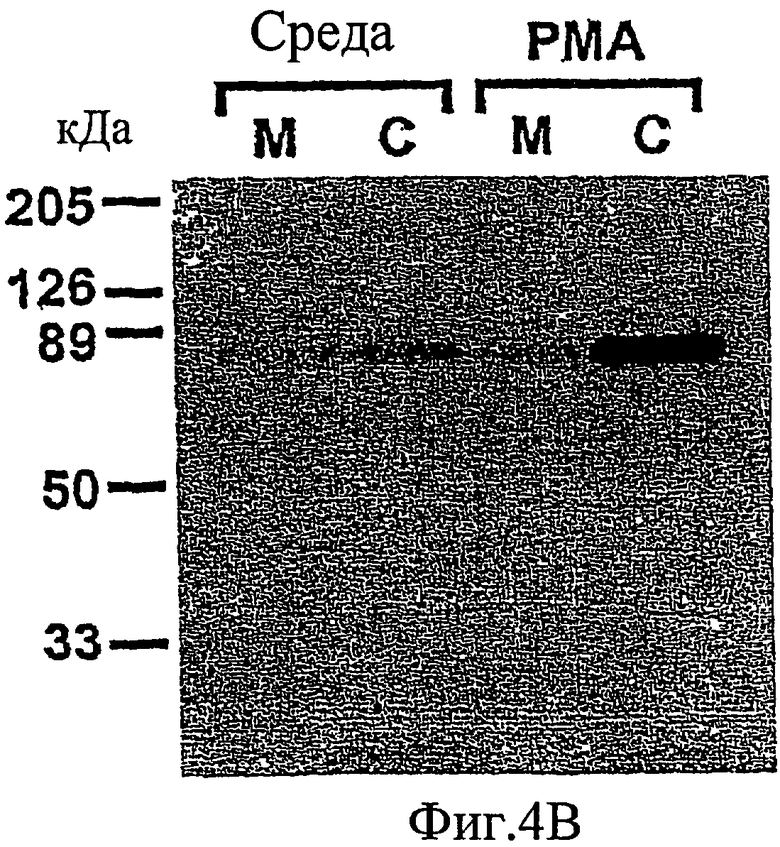

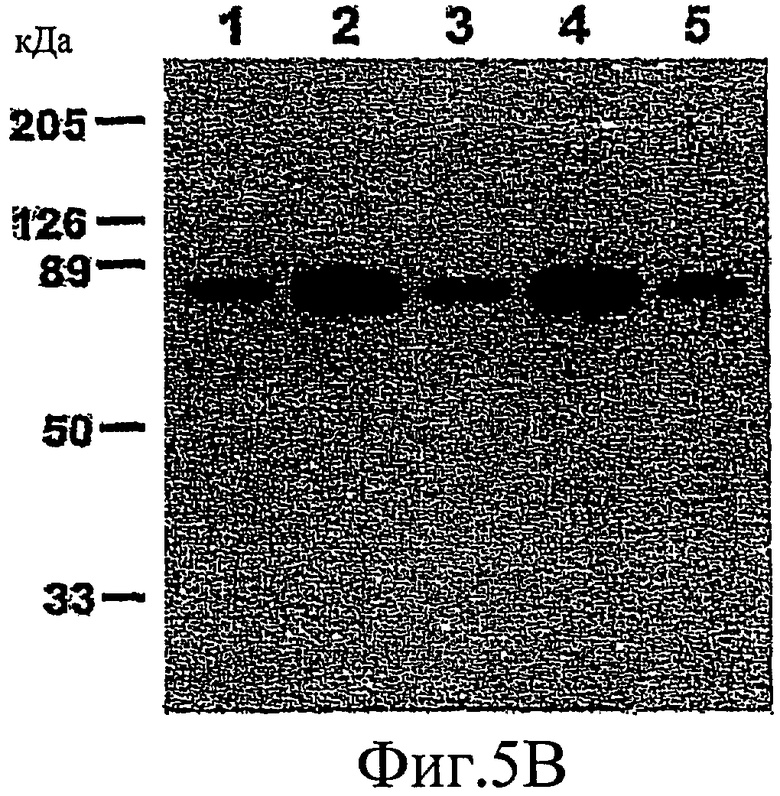
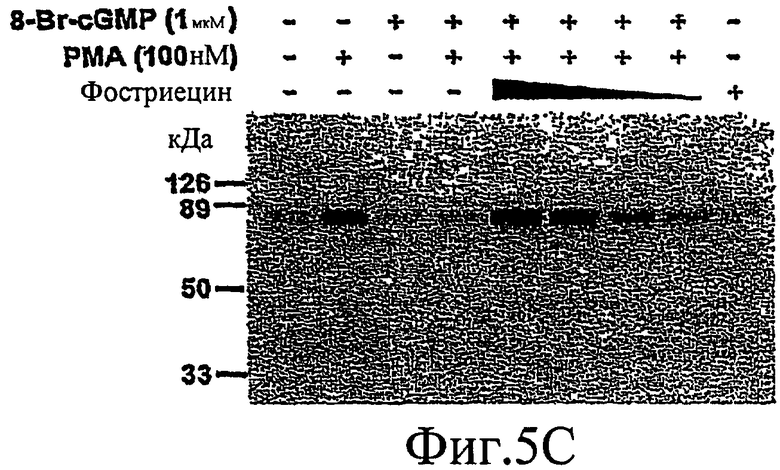

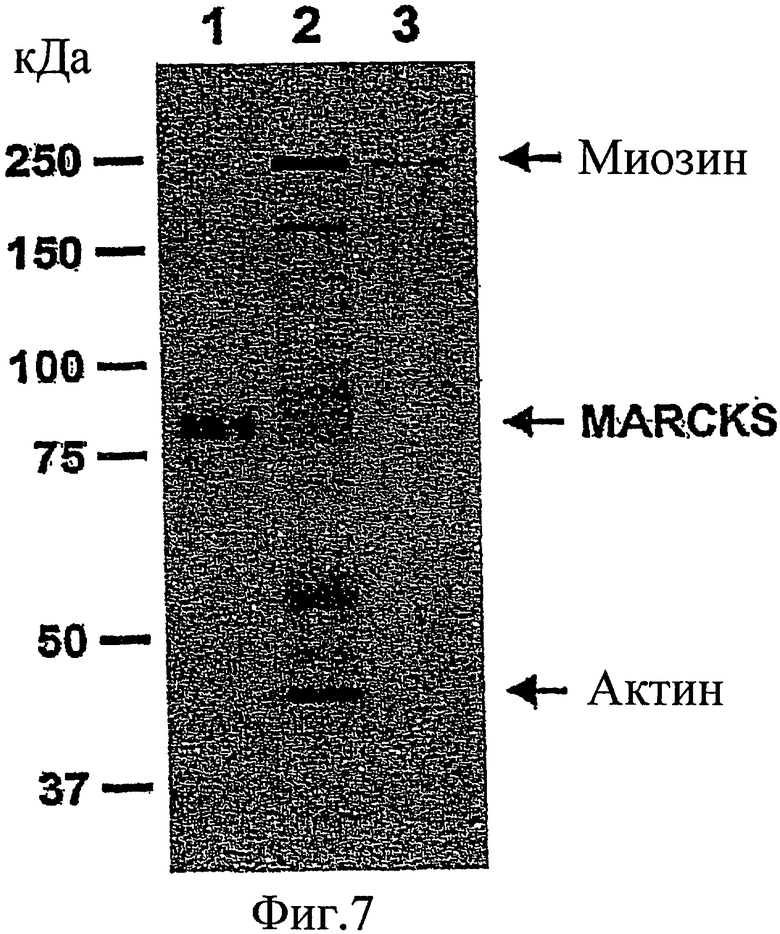
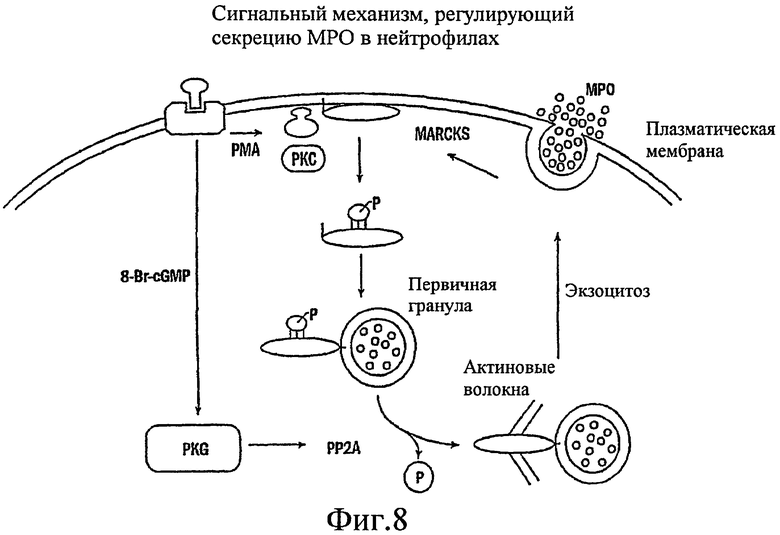
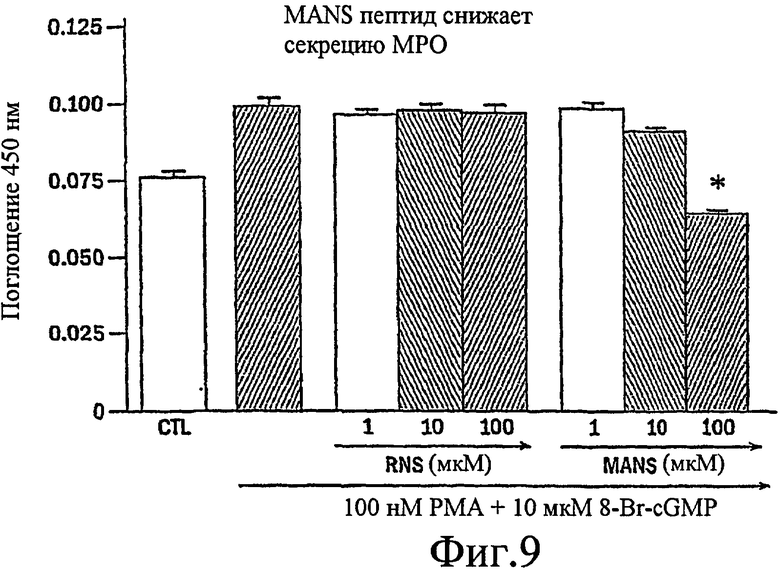

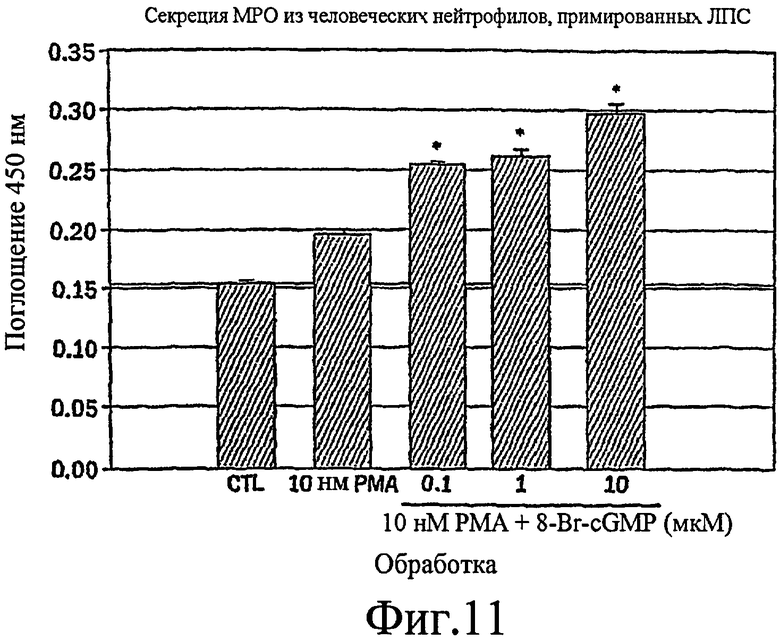
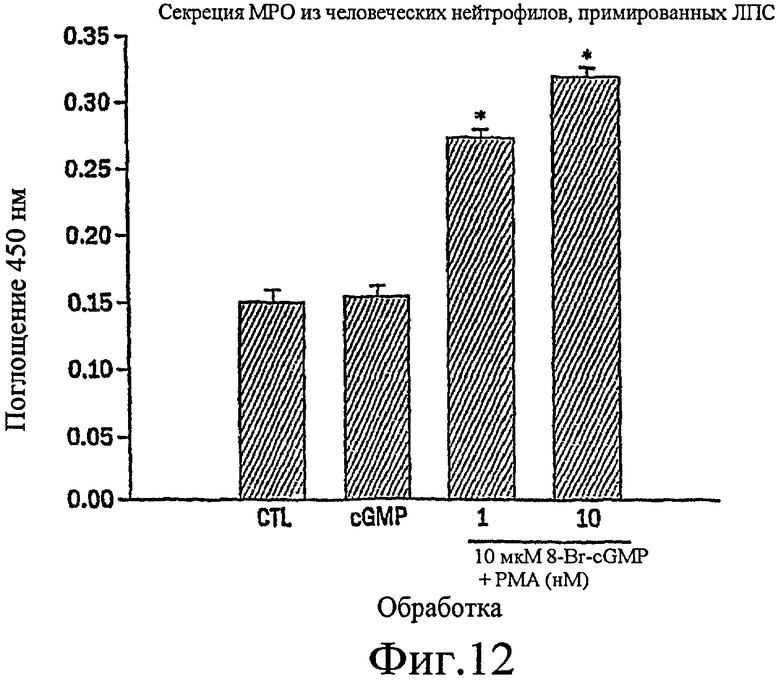
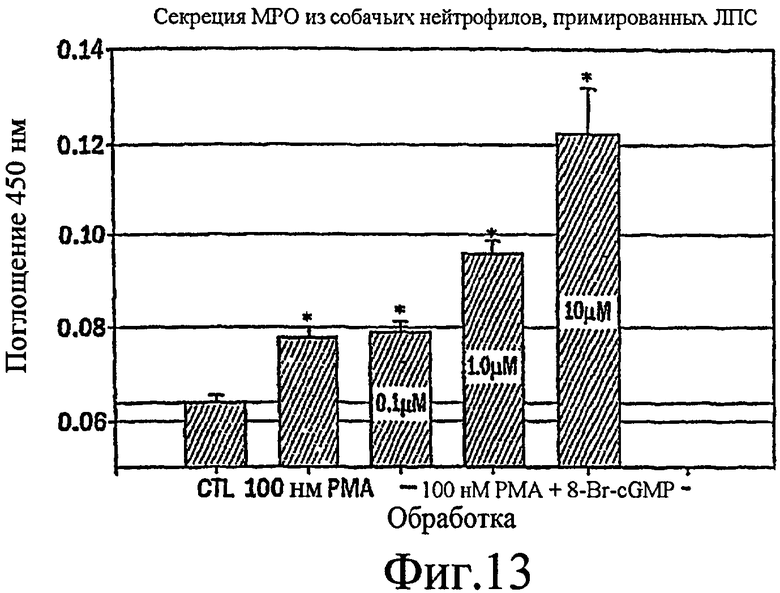
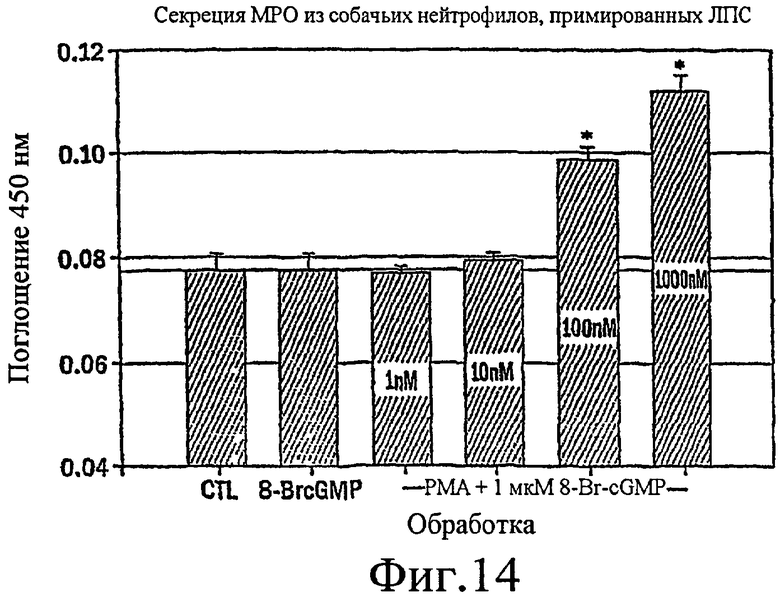
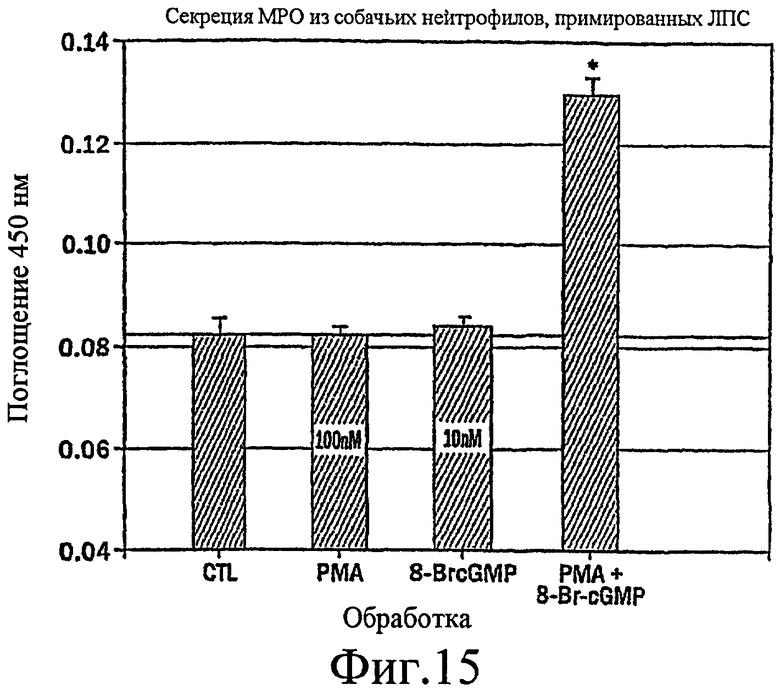
Изобретение относится к медицине и может быть использовано для снижения MARCKS-связанного высвобождения, по меньшей мере, одного медиатора воспаления, по меньшей мере, из одной воспалительной клетки, содержащегося в грануле внутри указанной клетки. Для этого вводят в указанную клетку эффективное ингибирующее дегрануляцию количество композиции, содержащей смесь двух или более миристоилированных N-концевых пептидов. Эффективное количество композиции снижает высвобождение медиатора воспаления из воспалительной клетки от 5 до 99% по сравнению с высвобождением указанного медиатора воспаления в отсутствие композиции. Изобретение позволяет одновременно снижать воспаление и блокировать секрецию слизи при воспалении, исключая при этом воздействие на другие механизмы воспалительного процесса. 8 з.п. ф-лы, 15 ил.
1. Способ снижения MARCKS-связанного высвобождения, по меньшей мере, одного медиатора воспаления, по меньшей мере, из одной воспалительной клетки, содержащегося в грануле внутри указанной клетки, где способ предусматривает
введение в указанную клетку эффективного ингибирующего дегрануляцию количества композиции, содержащей смесь двух или более миристоилированных N-концевых пептидов, которые выбраны из группы, состоящей из N-миристоил-GAQFSKTAAKGEAAAERPGEAAVA (SEQ ID NO:1); N-миристоил-САОЕЗКТААК (SEQ ID NO:15); N-миристоил-GAQFSKTAAKGEAAAERPGEAAV (SEQ ID NO:20); N-миристоил-GAQFSKTAAKGEAAAERPGEAA (SEQ ID NO:3); N-миристоил-GAQFSKTAAKGEAAAERPGEA (SEQ ID NO:4); N-миристоил-GAQFSKTAAKGEAAAERPGE (SEQ ID NO:5); N-миристоил-GAQFSKTAAKGEAAAERPG (SEQ ID NO:6); N-миристоил-GAQFSKTAAKGEAAAERP (SEQ ID NO:7); N-миристоил-GAQFSKTAAKGEAAAER (SEQ ID NO:8); N-миристоил-GAQFSKTAAKGEAAAE (SEQ ID NO:9); N-миристоил-GAQFSKTAAKGEAAA (SEQ ID NO:10); N-миристоил-GAQFSKTAAKGEAA (SEQ ID NO:11); N-миристоил-GAQFSKTAAKGEA (SEQ ID NO:12); N-миристоил-GAQFSKTAAKGE (SEQ ID NO:13); N-миристоил-GAQFSKTAAKG (SEQ ID NO:14); N-миристоил-GAQFSKTAA (SEQ ID NO:16); N-миристоил-GAQFSKTA (SEQ ID NO:17); N-миристоил-GAQFSKT (SEQ ID NO:18) и N-миристоил-GAQFSK (SEQ ID NO:19), где указанное эффективное количество снижает высвобождение указанного медиатора воспаления из указанной воспалительной клетки от 5 до 99% по сравнению с высвобождением указанного медиатора воспаления из, по меньшей мере, одной воспалительной клетки такого же типа, которое имело место в отсутствие указанной композиции.
2. Способ по п.1, где смесь двух или более миристоилированных N-концевых пептидов присутствует в молярном соотношении от 50:50 до 99,99:0,01.
3. Способ по п.1 или 2, где указанная смесь двух или более миристоилированных N-концевых пептидов присутствует в эффективном количестве, которое включает ингибирующее дегрануляцию количество, которое снижает количество медиатора воспаления, высвобождаемого из указанной воспалительной клетки: (i) от 1 до 99% или (ii) от 5-50 до 99%, по сравнению с количеством, высвобождаемым из указанной воспалительной клетки в отсутствие указанной смеси двух или более миристоилированных N-концевых пептидов.
4. Способ по п.1 или 2, где указанной воспалительной клеткой является лейкоцит, гранулоцит, нейтрофил, базофил, эозинофил, моноцит, макрофаг или их комбинация.
5. Способ по п.1, где указанный воспалительный медиатор выбирают из группы, состоящей из миелопероксидазы (МРО), эозинофильной пероксидазы (ЕРО), главного основного белка (МБР), лизозима, гранзима, гистамина, протеогликана, протеазы, цитокина, дефензина, бактерицидного белка, повышающего проницаемость (BPI), эластазы, катепсина G, катепсина В, катепсина D, бета-O-глюкуронидазы, альфа-маннозидазы, фосфолипазы А2, хондроитин-4-сульфата, протеиназы 3, лактоферрина, коллагеназы, активатора комплемента, рецептора комплемента, рецептора N-формиметиониллейцилфенилаланина (FMLP), рецептора ламинина, цитохрома b558, хемотаксического фактора моноцитов, гистаминазы, белка, связывающего витамин В12, желатиназы, активатора плазминогена и их комбинации.
6. Способ по п.1, где указанный медиатор воспаления выбирают из группы, состоящей из эозинофильной пероксидазы (ЕРО), лизозима, гранзима и их комбинации.
7. Способ по п.1 или 2, где указанная композиция представляет собой фармацевтическую композицию, пригодную для легочного введения, назального введения, перорального введения, местного введения, парентерального введения или ректального введения.
8. Способ по п.6, где указанное легочное введение выбирают из группы, состоящей из аэрозоля, ингалятора сухого порошка, ингалятора и небулайзера.
9. Способ по п.1, где фармацевтическая композиция дополнительно содержит вторую молекулу, выбираемую из группы, состоящей из антибиотика, противовирусного соединения, антипаразитарного соединения, противовоспалительного соединения и иммунодепрессанта.
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| ИБУПРОФЕНОВЫЙ ТИОЭФИР В КАЧЕСТВЕ ИНГИБИТОРА, ЗАВИСЯЩЕГО ОТ Nf-kB ОБРАЗОВАНИЯ МЕДИАТОРОВ ВОСПАЛЕНИЯ И БОЛИ | 1998 |
|
RU2204388C2 |
| TAKASHI S | |||
| et al | |||
| A peptide against the N-terminus of myristoylated alanine-rich С kinase substrate inhibits degranulation of human leukocytes in vitro // Am J Respir Cell Mol Biol | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| LI Y | |||
| et al | |||
| MARCKS protein is a key molecule regulating | |||

