Стратегии генной и клеточной терапии являются перспективными подходами для лечения многочисленных заболеваний, которые в настоящее время с трудом поддаются традиционными методами лечения или вовсе не излечимы. Тем не менее эффективность этих подходов в настоящее время невелика, что связано, во-первых, с относительно низкой эффективностью введения генных терапевтических конструкций в клетки in vitro и в особенности in vivo и, во-вторых, с конкуренцией трансплантируемых клеток модифицированных клеток со стороны клеток организма. Особый интерес представляет генная терапия, связанная с использованием стволовых клеток крови (СКК), которая является многообещающим подходом для коррекции многочисленных заболеваний крови и костного мозга. Многочисленные работы посвящены использованию трансдукции СКК с помощью вирусных векторов, однако на практике большинство из этих методов остаются сложными и маловоспроизводимыми, а главное недостаточно эффективными. При современном состоянии техники одним из наиболее действенных способов повышения эффективности использования генотерапевтических подходов является селекция клеток, содержащих терапевтические конструкции. Такая селекция может проводиться in vitro при культивировании клеток после введения терапевтических конструкций. Однако наиболее перспективной представляется селекция клеток in vivo, которая могла бы значительно повысить долю клеток, несущих терапевтические конструкции. Тем не менее селекция клеток in vivo представляет собой значительно более сложную техническую задачу, поскольку процедура селекции должна быть переносима организмом и обладать минимальной токсичностью. В настоящее время имеются несколько способов проведения селекции клеток in vivo. В частности, применяют селекцию с помощью введения в клетки конструкций, экспрессирующих ген множественной лекарственной устойчивости (MDR1) (Schiedlmeier et al., 2000, Abonour et al., 2000), ген дигидрофолатредуктазы (Allay et al., 1998) или ген цитидин деаминазы (Richara et al., 2004). Наиболее перспективным подходом и наиболее близким аналогом представляемого изобретения является метод, основанный на использовании для селекции клеток in vivo гена, кодирующего фермент O6-метилгуанин ДНК-метилтрансферазу (MGMT), обеспечивающий устойчивость клеток к алкилирующим агентам, таким как темозоломид или 1,3-бис(2-хлорэтил)-1-нитрозомочевина (BCNU), формирующим O6-ДНК алкильные производные (Pegg, 2000). Для селекции используют мутантные формы фермента, устойчивые к действию определенных ингибиторов MGMT, таких как O6-бензилгуанин (BG). В литературе описано несколько мутантов MGMT, обладающих устойчивостью к действию BG за счет специфических аминокислотных замен (Davis et al., 1999). Возможность использования таких мутантов для селекции и размножения СКК, несущих маркерный ген, в комбинации с совместной обработкой реципиентов BG и алкилирующими агентами, была показана на модели мыши (Davis et al., 1997, Cai et al., 2008), ксенотрансплантатах (Pollok et al., 2003, Zielske et al., 2003) и на модели крупных животных (Neffet al., 2003, Larochelle et al., 2009).
Несмотря на то что обработка BCNU/BG приводит к эффективной in vivo амплификации клеток, несущих мутантный ген MGMT (Cai et al., 2008, Pollok et al., 2003), многочисленные исследования посвящены изучению возможных побочных эффектов и разработке оптимальных протоколов введения алкилирующих препаратов (Sawai et al., 2003, Sorg et al., 2007, Kisby et al., 2009). В частности, существует проблема токсичности применяемых алкилирующих агентов (Larochelle et al., 2009), делающая настоятельной разработку подходов, связанных с использованием препаратов, обладающих минимальной токсичностью, позволяющих снизить миелосупрессию, цитопению и риск онкогенной трансформации.
Задачей изобретения является селекция клеток in vitro и in vivo с применением алкилирующих агентов с пониженной токсичностью.
Поставленная задача решается с использованием известного и разрешенного к использованию в медицине препарата лизомустин (2-хлорэтилнитрозоуреидопроизводное аминокислоты лизин) (номер госрегистрации Р №000127/01-2000) для селекции клеток с введенными конструкциями, одна из которых экспрессирует мутантную форму фермента MGMT, устойчивую к действию ингибиторов, подавляющих природную форму фермента, в частности, устойчивую к действию ингибитора O6-бензилгуанина. После введения в организм ингибитора O6-бензилгуанина лизомустин подавляет размножение собственных клеток организма, в частности гемопоэтических клеток, в результате чего происходит селекция генетически модифицированных клеток, несущих какую-либо нуклеотидную последовательность, обладающую терапевтическим действием. Терапевтическая нуклеотидная последовательность может экспрессировать белок, обладающий терапевтическим действием, или РНК (например, антисмысловую РНК, короткую шпилечную РНК или микроРНК), подавляющую экспрессию определенного белка. Лизомустин представляет собой смесь изомеров положения нитрозогруппы: А) (S)-2-амино-6-[N-нитрозо-N-[[(2-хлорэтил)амино]карбонил]амино]гексановая кислота; Б) (S)-2-амино-6-[[[N-нитрозо-N-(2-хлорэтил)амино]карбонил]амино]гексановая кислота. Проведенные клинические исследования показали, что препарат хорошо переносится и проявляет высокую эффективность в отношении злокачественных меланомы, лимфосаркомы, а также при лечении больных раком легкого. Лизомустин обладает доказанной пониженной токсичностью при клиническом применении по сравнению с другими алкилирующими агентами, что позволяет считать целесообразным его применение у тяжелых больных, больных пожилого возраста и в амбулаторных условиях.
Предложенный способ селекции генетически модифицированных клеток может быть использован для осуществления заместительной терапии патологических состояний, связанных с нарушением функционирования клеток в организме, в частности с нарушениями функционирования гемопоэтических клеток (ГК) и системы иммунитета, а также метаболических заболеваний, связанных с клетками кровеносной системы. Предложенный способ является новым применением лизомустина.
Технический результат представляет собой расширение арсенала средств указанного действия и снижение токсичности алкилирующего агента.
Примеры применения
На Фиг.1 показаны формулы двух изомеров лизомустина.
Лизомустин - противоопухолевый препарат из группы нитрозоалкилмочевин (НАМ), который представляет собой смесь изомеров положения нитрозогруппы: (S)-2-амино-6-[N-нитрозо-N-[[(2-хлорэтил)амино]карбонил]амино]гексановая кислота (1) и (S)-2-амино-6-[[[N-нитрозо-N-(2-хлорэтил)амино]карбонил]амино]гексановая кислота (2).
Изомеры положения нитрозогруппы, 1 и 2, входящие в состав лизомустина, обладают разной противоопухолевой активностью. Изомер 2 обладает большей противоопухолевой активностью. Клинически используется смесь изомеров 1 и 2, которая также обладает значительной противоопухолевой активностью, но при этом имеет более широкий дозовый диапазон применения.
Для исследования возможности применения лизомустина и его изомеров для селекции клеток вначале были проведены эксперименты на культуре клеток, где проводилась отработка протоколов и проверка созданных экспрессионных векторов. Для этого была создана серия генетических конструктов, содержащих мутантную форму MGMT (MGMTm). кДНК MGMTwt была получена с использованием MGC клона (Opened Biosystems, BC000824) с использованием олигонуклеотидов TCATGGATCCAAGCCACCATGGACAAGGATTGTGAA и ATTAGAATTCACATACTCAATTGCGGCCAGCAGG. Мутация, кодирующая замену пролина на лизин в положении 140 (P140K), была введена методом ПЦР с использованием олигонуклеотидов: GGCAATCCTGTCAAGATCCTCATCCCG и CGGGATGAGGATCTTGACAGGATTGCC, после чего наличие мутации подтверждено секвенированием. Для создания мутантной формы MGMT, слитой на С- и N-конце с эпитопом FLAG (DYKDDDDK), эпитоп вводился методом ПЦР. Все кДНК были клонированы в вектора pLenti6V5 (Invitrogen, Carlsbad, CA), pMigRI (Pear et al., 1998) или pLeGo-G2 (Weber et al., 2008) с получением N-концевой сшивки с FLAG в векторе pMigRI (вектор FM) и С-концевой сшивки с FLAG в pLeGo-G2 (вектор MF). Для получения вектора G2AMF пептид 2А вируса FMDV был получен попарным отжигом двух пар олигонуклеотидов: CTGTCAAACAAACTCTTAACTTTGATTT и GTTTGAGTAAATCAAAGTTAAGAGTTTGTT, ACTCAAACTGGCTGGGGATGTAGAAAGCAA и CCTGGATTGCTTTCTACATCCCCAGCCA, после чего их лигировали с MGMTm-FLAG. Полученный фрагмент клонировали в вектор pEGFP-C1 (Clontech) в общую открытую рамку считывания с геном EGFP, служащим на данном этапе в качестве модели терапевтической нуклеотидной последовательности, корректирующей гипотетическую аномалию в клетке. Флуоресценция модельного белка EGFP является показателем его количества в клетке, а процент флуоресцирующих клеток в организме-реципиенте является показателем степени размножения в нем генетически модифицированных ГК. После этого кассету, содержащую EGFP-2A-MGMTm-FLAG, вырезали и вставляли в вектор pLeGo-G2 на место EGFP.
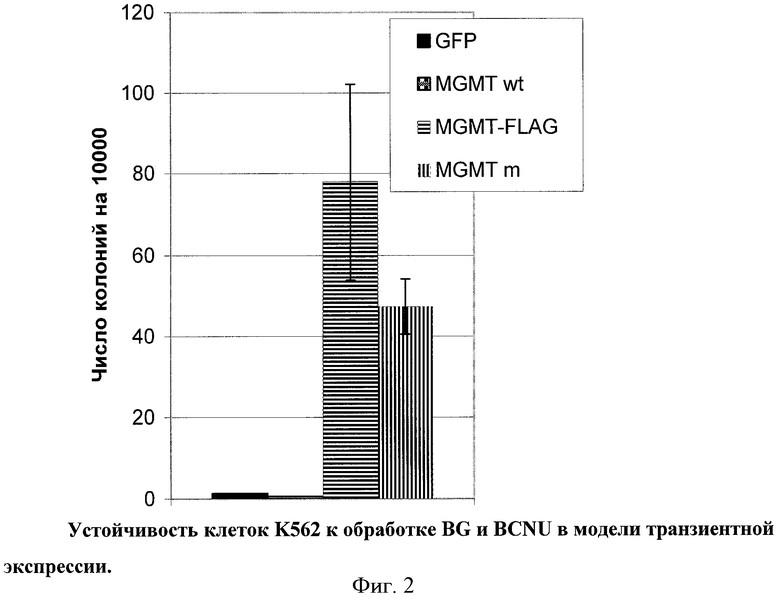
Для оценки ферментативной активности мутантных вариантов MGMT фермент дикого типа (MGMTwt), а также слитую с пептидным эпитопом FLAG мутантную форму, нечувствительную к действию BG, клонировали в вектор pLenti6V5. Далее была произведена серия экспериментов по транзиентной трансфекции клеток полученными векторами. Клетки К562, экспрессирующие EGFP или MGMTwt, не были устойчивы к совместному действию BG и BCNU, в отличие от мутантной формы (Фиг.2). Кроме того, добавление короткого эпитопа FLAG не снижало активности MGMTm, что давало возможность определять его наличие методами проточной цитометрии, используя антитела к FLAG.
Для получения лентивирусных частиц клетки HEK293FT (Invitrogen) были трансфецированы с помощью линейного полиэтиленимина упаковочными плазмидами pMDLg/pRRE, PRSV-Rev (Addgene, Cambridge, MA), плазмидой, кодирующей оболочку pVSV-G (Invitrogen), и вектором, несущим соответствующую форму MGMT. Кондиционированную среду, содержащую вирусные частицы, собирали спустя 24-72 часа после трансфекции, фильтровали и центрифуговали при 28000 оборотах ротора в течение 2 часов при +4°С. Осадок ресуспендировали в 1/100 исходного объема и хранили при -70 градусах в аликвотах. Для получения ретровирусных частиц упаковочные клетки линии 293.1 трансфецировали упаковочной плазмидой, а также вектором FM, супернатанты собирали через 36 часов.
Клетки K562 или первичную культуру костного мозга мыши ресуспензировали в бессывороточной среде и вначале инкубировали с 0 или 20 мкМ BG в течение 1.5 часов при 37°C, после чего - с различными концентрациями лизомустина или BCNU в течение еще 1.5 часов. Затем культуры отмывали от препаратов и высевали в трех повторах в метилцеллюлозу (StemCells, Vancouver, ВС, Канада), содержащую 20 нг/мл рекомбинантного интерлейкина 3 человека и 50 нг/мл рекомбинантного гранулоцитарного колониестимулирующего фактора (Peprotech, Rocky Hill, NJ). Клетки костного мозга мыши высевали в метилцеллюлозу, содержащую по 20 нг/мл рекомбинантных мышиных интерлейкиов 3 и 6, а также мышиный рекомбинантный фактор роста стволовых клеток (50 нг/мл). Колонии, содержащие более 30 клеток, подсчитывали и оценивали разницу между образцами.
BCNU и лизомустин по своей химической структуре относятся к классу нитрозомочевин. Характерной особенностью нитрозоалкилмочевин как химиотерапевтических препаратов является их способность к гидролитическому распаду в водных растворах с образованием алкилирующих частиц. В связи с этим была проверена способность мутантной формы MGMT защищать клетки К562 от действия лизомустина.
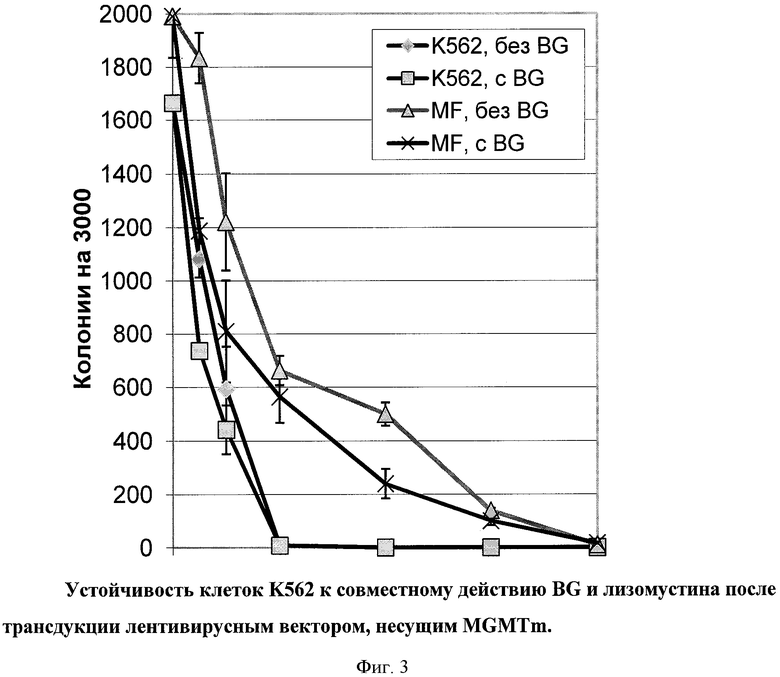
Клетки K562 инкубировали с лизомустином в концентрации 0-400 мкM отдельно или в комбинации с 20 мкМ BG. Кривые выживания клеток, основанные на способности выживших клеток формировать колонии, представлены на Фиг.3. Трансдукция вектором MF обеспечивала устойчивость клеток к обработке лизомустином вплоть до концентрации 300 мкМ. Показано выживание 21% колониеобразующих клеток в трансдуцированной MF культуре против 0.3% в контрольной популяции при концентрации лизомустина в концентрации 100 мкМ. BG в данной модельной системе усиливал токсичность лизомустина лишь в незначительной степени. При 100 мкМ лизомустина уменьшение колониеобразования при использовании BG составляло с 21 до 18.8%, очевидно, за счет ингибирования эндогенного фермента. Таким образом, препарат лизомустин может быть использован для получения популяций гемопоэтических клеток, в той или иной степени обогащенных генетическими конструкциями, несущими MGMTm.
В следующей серии была проанализирована устойчивость трансдуцированных вектором MF клеток костного мозга мыши к действию лизомустина in vitro. Обработали культуру костного мозга BCNU или лизомустином, отдельно или в комбинации с BG, после чего провели анализ выживания колониеобразующих предшественников в метилцеллюлозе. Присутствие BG в инкубационной среде драматически увеличивало чувствительность колониеобразующих клеток к обработке алкилирующими агентами, достигая 100% при 30 мкM BCNU (Фиг.4). Напротив, 26.4±2.9% колониеобразующих клеток выживало при такой же концентрации в отсутствие BG, что может объясняться высоким уровнем эндогенной MGMT.
Предварительные эксперименты показали, что летальная для 50% клеток доза (ЛД50) для лизомустина примерно в 2-3 раза выше по сравнению с BCNU. Клетки костного мозга обработали лизомустином отдельно или в комбинации с BG. Колониеобразование при 50 мкM лизомустина в присутствии 20 мкМ BG составило 46.7±4.2% и 15.9±1.1% при 100 мкМ. Цитотоксический эффект лизомустина в меньшей степени зависел от присутствия BG, по сравнению с BCNU. Учитывая определенные проблемы с системным введением BG при использовании на животных и человеке, данное свойство лизомустина дает ему преимущество перед BCNU и другими препаратами подобного класса.
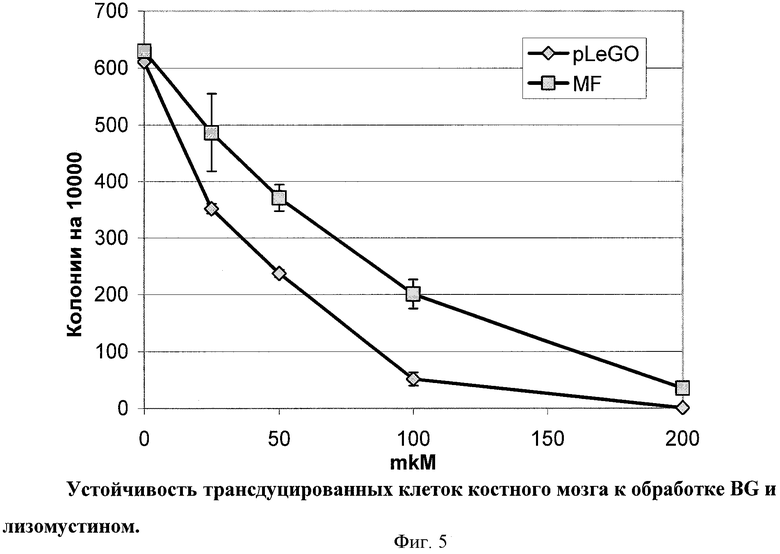
Для того чтобы выяснить, возможно ли с помощью вирусной доставки векторов, несущих MGMT, обеспечить устойчивость клеток костного мозга к действию лизомустина, клетки костного мозга трансдуцировали MF лентивирусным и FM ретровирусным векторами. Около 8% ядерных клеток экспрессировали MGMTm после трансдукции MF, по данным цитометрии с антителами против FLAG. Кривые выживания, построенные с помощью подсчета миелоидных колониеобразующих единиц, приведены на Фиг.5. Трансдуцированные вектором MF популяции обладали повышенной устойчивостью к лизомустину и BG (ЛД50 около 70 мкМ) по сравнению с трансдуцированными «пустым» вектором (ЛД50 около 35 мкМ). Обработка одним лизомустином, в отсутствие BG, не позволяла выявить различий в устойчивости двух популяций, что, возможно, связано с высоким уровнем эндогенной MGMT.
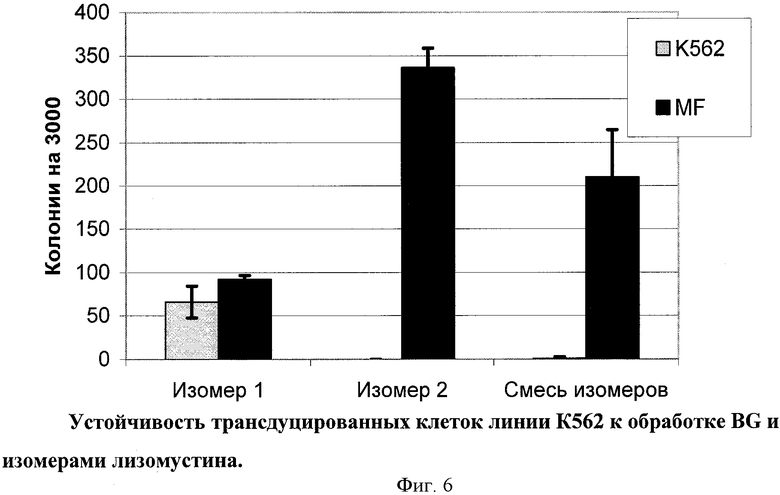
Было также исследовано действие изомеров лизомустина на трансдуцированные вектором MF клетки линии К562. Полученные результаты (Фиг.6) показывают, что изомер 1 лизомустина (не обладающий значительным антираковым воздействием) оказывает определенное токсическое воздействие на клетки К562, однако его токсическая активность не может быть подавлена действием мутантного фермента MGMT. В то же время изомер 2 лизомустина обладает мощным токсическим эффектом на клетки, и при этом данное его воздействие может быть подавлено активностью мутантной формы MGMT. Клинически используемая смесь изомеров лизомустина оказывает промежуточное воздействие, более близкое к действию изомера 2.
Таким образом, в экспериментах на культуре клеток была показана возможность применения лизомустина для in vitro обогащения ГК, несущих одновременно MGMTm и использованный в качестве модельного ген EGFP. При этом изомер 2 лизомустина является более эффективным при селекции, чем применяемая в клинической практике смесь изомеров, в то время как изомер 1 лизомустина не обладает селектирующим действием.
Для подтверждения возможности использования лизомустина для замещения собственных гемопоэтических клеток организма на клетки, несущие заданный ген в системе in vivo, были проведены серии экспериментов на мышиной модели. Из уровня техники известно, в качестве гена, служащего для in vivo обогащения трансгенных ГК, может быть использован ген MGMT. Так, на модели мыши недавно была продемонстрирована успешная клеточная терапия бета-талассемии и протопорфирии с помощью трансплантации клеток, несущих ген мутантной формы MGMT и соответствующий терапевтический ген (Persons et al., 2003, Richard et al., 2004). В этих экспериментах были использованы другие производные нитрозомочевины, отличные по структуре от лизомустина.
В экспериментах были использованы 8-12-недельные реципиентные мыши линии С57 В1/6. Вначале была проанализирована чувствительность мышей к лизомустину без трансплантации клеток. Лизомустин без BG в количествах до 160 мг/кг массы тела не влиял на выживание мышей при внутрибрюшинном способе введения. Напротив, две инъекции лизомустина в количествах по 60 мг/кг совместно с 30 мг/кг бензилгуанина приводили к гибели 40% реципиентов (2 из 5). На основании этих данных, а также данных in vitro экспериментов для дальнейшей работы по трансплантации ГК, несущих гены MGMTm и EGFP, были использованы дозы лизомустина, не превышающие 50-60 мг/кг. В качестве модели нуклеотидной последовательности, обладающей терапевтическим действием, был использован ген EGFP. Флуоресценция модельного белка EGFP отражает его количество в клетке, а процент флуоресцирующих клеток в организме-реципиенте является показателем степени селекции в нем генетически модифицированных ГК.
Облучение животных производили при 1.5-4 Гр с помощью рентгеновского аппарата, после чего реципиентам трансплантировали в хвостовую вену по 3×106 клеток костного мозга, модифицированых по генам MGMTm и EGFP, взятому в качестве маркера. Введение химических препаратов осуществляли внутрибрюшинно. BG вводили в количестве 30 мг/кг массы тела в предварительно нагретом растворе, содержащем 40% PEG400 (AppliChem, Darmstadt, Германия) и 60% PBS. Введение алкилирующих препаратов осуществляли через 2 часа после введения BG в количествах: BCNU - 10 мг/кг, лизомустин - 50 мг/кг в PBS. За 5 дней до начала эксперимента мышам давали подкисленную питьевую воду, содержащую неомицин (0.5 г/л).
Для анализа количества наработанного продукта, каковым являются генетически модифицированные ГК, эритроциты крови или костного мозга лизировали с помощью реагента OptiLyseC (Beckman Coulter). Затем ГК промывали в PBS, ресуспендировали в PBS, содержащем 1% FBS, и анализировали с помощью проточного цитоспектрофлуориметра Epics XL4, данные обсчитывали в программе WinMDI 2.9.
Кровь мышей была проанализирована непосредственно после селекции для оценки уровня модифицированных ГК. Селективное преимущество трансдуцированных FM вектором клеток наблюдалось как после обработки BG с BCNU, так и после обработки BG с лизомустином (Фиг.7).
Один раунд селекции показал следующие средние значения процента позитивных по EGFP клеток: 0.046% в крови у контрольной группы, 0.62% в группе BCNU, 0.26 и 0.79% в группах лизомустина, получавших по 25 мг/кг и 60 мг/кг соответственно. Таким образом, системная обработка лизомустином повышает уровень накопления ГК содержащих гены MGMTm и модельного белка EGFP, причем при условии применения дозы 60 мг/кг уровень накопления превышает таковой для BCNU в среднем на 30%.
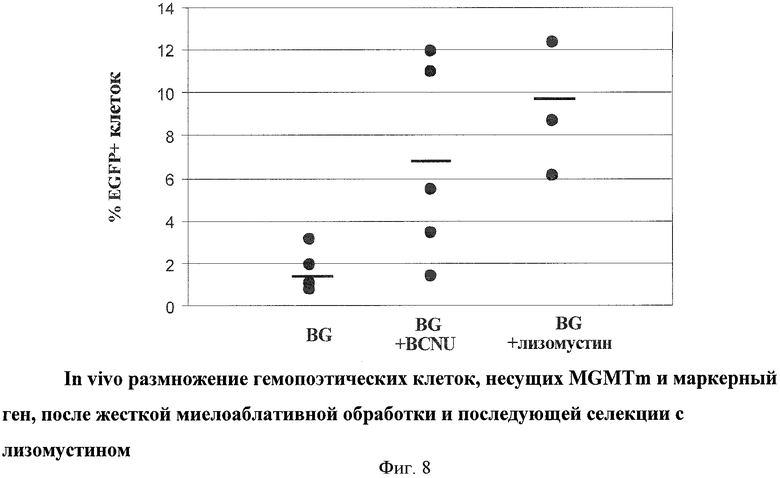
Для дальнейшего исследования возможности получения генетически модифицированных ГК с использованием лизомустина провели второй модельный эксперимент, в котором была использована более жесткая миелосупрессия реципиентных мышей, после чего спустя 2 недели произвели анализ крови. Средние значения уровня модифицированных ГК в периферической крови для трех групп составили 1.6; 6.7 и 9.1 для контрольной, BCNU и группы лизомустина соответственно. Данные приведены на Фиг.8.
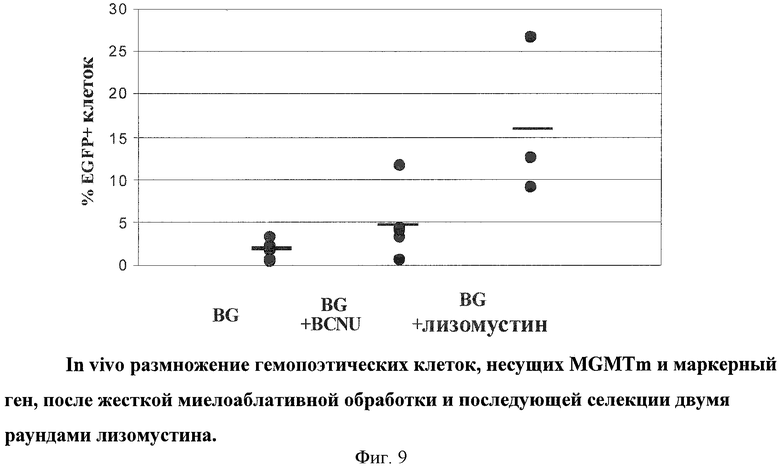
Для дальнейшего обогащения генетически модифицированных ГК провели второй раунд селекции ГК в тех же реципиентных мышах, используя те же концентрации реагентов. Показано дальнейшее обогащение содержания модифицированных ГК в группе лизомустина (16.2%), в то время как среднее значение процента в группе BCNU снизилось до 4.9%. При этом контрольная группа не претерпела существенных изменений (1.8%). Данные приведены на Фиг.9.
На основании проведенных экспериментов можно сделать вывод, что обработка лизомустином мышей как способ оценки действия лизомустина приводит к увеличению в периферической крови животных процента клеток, несущих ГК с генетическим конструктом, содержащим мутантную форму MGMT и какую-либо другую нуклеотидную последовательность, необходимую для выполнения терапевтической функции.
Полученные данные по сравнительному анализу лизомустина и BCNU показали более эффективную селекцию ГК при использовании лизомустина. Способ замещения генетически модифицированными ГК описан на модели линейной мыши, однако может быть применен для аутотрансплантации, аллогенной трансплантации или ксенотрансплантации. Кроме введения в клетки с помощью трансдукции ретровирусных и лентивирусных векторов, описанных в вышеприведенных примерах, из литературы известны также способы введения генетических конструктов в клетки с помощью аденоассоциированных векторов (Daya & Berns, 2008), транспозонов (Mates et al., 2009) и плазмидных эписомальных конструкций (Ehrhardt et al., 2008). При современном состоянии техники поэтому возможно также применение аденоассоциированных векторов, транспозонов и плазмидных эписомальных конструкций для использования в предлагаемом способе селекции клеток с помощью лизомустина.
Список источников
Экспериментальная онкология на рубеже веков. Под ред. Давыдова М.И., Барышникова А.Ю. С.147, с.161. Москва. 2003.
Abonour R, Williams DA, Einhorn L et al. Efficient retrovirus-mediated transfer of the multidrug resistance 1 gene into autologous human long-term repopulating hematopoietic stem cells. Nat. Med. 2000; 6: 652-658.
Allay JA, Persons DA, Galipeau J et al. In vivo selection of retro virally transduced hematopoietic stem cells. Nat. Med. 1998; 4: 1136-1143.
Cai S, Ernstberger A, Wang H et al. In vivo selection of hematopoietic stem cells transduced at a low multiplicity-of-infection with a foamy viral MGMT(P140K) vector. Exp Hematol. 2008 Mar; 36(3): 283-92.
Davis BM, Reese JS, Кос ON et al. Selection for G156A O6-methylguanine DNA methyltransferase gene-transduced hematopoietic progenitors and protection from lethality in mice treated with O6-benzylguanine and 1,3-bis(2-chloroethyl)-1-nitrosourea. Cancer Res. 1997; 57(22): 5093-9.
Davis BM, Roth JC, Lui L et al. Characterization of the P140K, PVP(138-140)MLK, and G156A 06-methylguanine-DNA methyltransferase mutants: implications for drug resistance gene therapy. Human Gene Ther. 1999; 10(17): 2769-78.
Daya S, Berns K.I. Gene therapy using adeno-associated virus vectors. Clin Microbiol Rev. 2008; 21(4): 583-89.
Ehrhardt A, Haase R, Schepers A, Deutsch MJ, Lipps HJ, Baiker A. Episomal vectors for gene therapy. Curr Gene Ther. 2008; 8(3): 147-61.
Kisby GE, Olivas A, Park Т et al. DNA repair modulates the vulnerability of the developing brain to alkylating agents. DNA Repair (Amst), 2009; 8(3): 400-12.
Larochelle A, Choi U, Shou Y et al. In vivo selection ofhematopoietic progenitor cells and temozolomide dose intensification in rhesus macaques through lentiviral transduction with a drug resistance gene. J Clin Invest. 2009; 119(7): 1952-1963.
Mates L, Chuah MK, Belay E, Jerchow B, Manoj N, Acosta-Sanchez A, Grzela DP, Schmitt A, Becker K, Matrai J, Ma L, Samara-Kuko E, Gysemans C, Pryputniewicz D, Miskey C, Fletcher B, Vandendriessche T, Ivies Z, Izsvak Z. Molecular evolution of a novel hyperactive Sleeping Beauty transposase enables robust stable gene transfer in vertebrates. Nat Genet. 2009; 41(6): 753-61.
Neff T, Horn PA, Peterson LJ et al. Methylguanine methyltransferase-mediated in vivo selection and chemoprotection ofallogeneic stem cells in a large-animal model. J. Clin. Invest. 2003; 112(10): 1581-1588.
Pear WS, Miller JP, Xu L et al. Efficient and rapid induction of a chronic myelogenous leukemia-like myeloproliferative disease in mice receiving P210 bcr/abl-transduced bone marrow. Blood. 1998; 92(10): 3780-92.
Pegg AE. Repair of O(6)-alkylguanine by alkyltransferases. Mutat. Res. 2000; 462: 83-100.
Persons DA, Allay ER, Sawai N et al. Successful treatment of murine beta-thalassemia using in vivo selection of genetically modified, drug-resistant hematopoietic stem cells. Hargrove PW, Brent TP, Hanawa H. Blood. 2003; 102(2): 506-13.
Pollok KE, Hartwell JR, Braber A. In vivo selection of human hematopoietic cells in a xenograft model using combined pharmacologic and genetic manipulations. Hum. Gene Ther. 2003; 14(18): 1703-14.
Rattmann I, Kleff V, Sorg UR et al. Gene transfer of cytidine deaminase protects myelopoiesis from cytidine analogs in an in vivo murine transplant model. Blood. 2006; 108(9): 2965-71.
Richara E, Robert E, Cario-Andre M et al. Hematopoietic stem cell gene therapy of murine protoporphyria by methylguanine-DNA-methyltransferase-mediated in vivo drug selection. Gene Ther. 2004; 11(22): 1638-47.
Sawai N, Persons DA, Zhou S et al. Reduction in hematopoietic stem cell numbers with in vivo drug selection can be partially abrogated by HOXB4 gene expression. Mol Ther. 2003; 8(3): 376-84.
Schiedlmeier B, Kuhlcke K, Eckert HG et al. Quantitative assessment of retroviral transfer of the human multidrug resistance 1 gene to mobilized peripheral blood progenitor cells engrafted in nonobese diabetic/severe combined immunodeficient mice. Blood. 2000; 95: 1237-1248.
Sorg UR, Kleff V, Fanaei S et al. O6-methylguamne-DNA-methyltransferase (MGMT) gene therapy targeting haematopoietic stem cells: studies addressing safety issues. DNA Repair (Amst). 2007; 6(8): 1197-209.
Weber K, Bartsch U, Stocking C, Fehse В A multicolor panel of novel lentiviral "gene onthology" (LeGO) vectors for functional gene analysis. Mol. Ther. 2008; 16(4): 698-706.
Zielske SP and Gerson SL Blood Cells Mol. Dis. 2003; 31(1): 48-50. SarCNU mediates selection ofP140K methylguanine-DNA-methyltransferase transduced human CD34(+) cells in vitro.
Zielske SP, Reese JS, Lingas KT et al. In vivo selection of MGMT(PHOK) lentivirus-transduced human NOD(SCID) repopulating cells without pretransplant irradiation conditioning. J. Clin Invest. 2003 Nov; 112(10): 1561-70.
| название | год | авторы | номер документа |
|---|---|---|---|
| Рекомбинантный вектор для создания плазмидных генетических конструкций, обладающих повышенной длительностью экспрессии целевых генов | 2018 |
|
RU2686102C1 |
| СПОСОБ КОНСТРУИРОВАНИЯ АЛЛОГЕННЫХ И УСТОЙЧИВЫХ К ЛЕКАРСТВЕННЫМ ПРЕПАРАТАМ Т-КЛЕТОК ДЛЯ ИММУНОТЕРАПИИ | 2014 |
|
RU2689558C1 |
| ВИРУСНЫЕ ВЕКТОРЫ С ЗАВИСИМОЙ ОТ УСЛОВИЙ РЕПЛИКАЦИЕЙ И ИХ ПРИМЕНЕНИЕ | 2001 |
|
RU2301260C2 |
| РЕКОМБИНАНТНЫЙ ЛЕНТИВИРУСНЫЙ ВЕКТОР, КЛЕТКА-ХОЗЯИН, ТРАНСДУЦИРОВАННАЯ ЛЕНТИВИРУСНЫМ ВЕКТОРОМ, СПОСОБ ЕЕ ТРАНСДУКЦИИ И ПРИМЕНЕНИЕ | 2002 |
|
RU2305708C2 |
| УСОВЕРШЕНСТВОВАНИЕ ГЕНЕТИЧЕСКИХ КОНСТРУКЦИЙ ДЛЯ ПОВЫШЕНИЯ ЭФФЕКТИВНОСТИ АНТИВИЧ ТЕРАПИИ | 2013 |
|
RU2533817C1 |
| Рекомбинантная клеточная линия TMDK562-15, проявляющая способность к активации и пролиферации ЕК клеток человека | 2022 |
|
RU2803178C1 |
| ВЕКТОРЫ AAV | 2018 |
|
RU2793112C2 |
| СПОСОБ СТИМУЛИРОВАНИЯ РЕГЕНЕРАЦИИ СПИННОГО МОЗГА С ПОМОЩЬЮ ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫХ КЛЕТОК КРОВИ ПУПОВИНЫ ЧЕЛОВЕКА | 2013 |
|
RU2521225C2 |
| Двухцепочечная РНК, способная снижать экспрессию мутантного аллеля с.607GA гена GNAO1 человека | 2022 |
|
RU2816137C1 |
| СПОСОБ ПОЛУЧЕНИЯ ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫХ РАСТЕНИЙ САХАРНОЙ СВЕКЛЫ С ИСПОЛЬЗОВАНИЕМ AGROBACTERIUM TUMEFACIENS | 2004 |
|
RU2278162C2 |


Изобретение относится к области биотехнологии, конкретно к клеточным технологиям, и может быть использовано для селекции генетически модифицированных клеток. В клетки вводят одну или несколько генетических конструкций, в составе которых находится ген мутантной формы фермента MGMT (О6-метилгуанин ДНК-метилтрансфераза), устойчивой к действию ингибитора О6-бензилгуанина, подавляющего действие природной формы MGMT, а также нуклеотидная последовательность, обладающая терапевтическим действием. Модифицированные таким образом клетки трансплантируют в организм, после чего в организм вводят указанный ингибитор, а затем изомер 2 лизомустина или смесь изомеров 1 и 2 лизомустина, которые подавляют размножение собственных клеток организма, в результате чего происходит селекция модифицированных клеток. Изобретение обеспечивает более избирательную селекцию модифицированных клеток и позволяет повысить безопасность и эффективность способа селекции клеток из-за использования лизомустина, характеризующегося пониженной токсичностью. 3 з.п. ф-лы, 9 ил.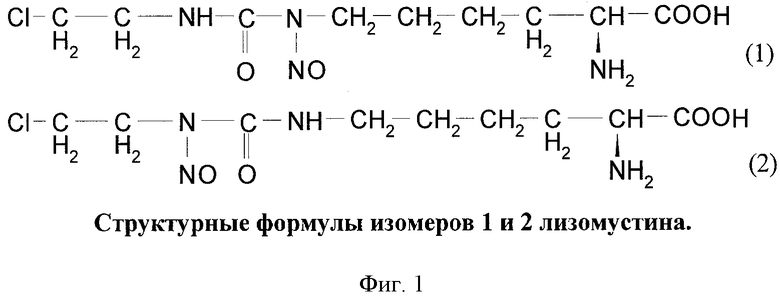
1. Способ селекции генетически модифицированных клеток человека или животных, заключающийся в том, что в клетки вводят одну или несколько генетических конструкций, в составе которых находится ген мутантной формы фермента MGMT (О6-метилгуанин ДНК-метилтранфераза), устойчивой к действию ингибитора О6-бензилгуанина, подавляющего действие природной формы MGMT, а также нуклеотидная последовательность, обладающая терапевтическим действием; модифицированные таким образом клетки трансплантируют в организм, после чего в организм вводят указанный ингибитор, а затем изомер 2 лизомустина или смесь изомеров 1 и 2 лизомустина, которые подавляют размножение собственных клеток организма, в результате чего происходит селекция модифицированных клеток.
2. Способ по п.1, характеризующийся тем, что в качестве генетических конструкций используют интегрирующие в ДНК клеток конструкции, выбранные из группы, включающей ретровирусные, лентивирусные, аденоассоциированные вирусные, транспозоновые и плазмидные эписомально поддерживающиеся конструкции.
3. Способ по п.1, характеризующийся тем, что ген мутантной формы фермента MGMT и нуклеотидная последовательность, обладающая терапевтическим действием, находятся в составе одной и той же конструкции.
4. Способ по п.1, характеризующийся тем, что перед введением модифицированных клеток в организм проводят обработку клеток в культуре ингибитором, подавляющим действие природной формы MGMT, а затем лизомустином.
| LAROCHELLE A | |||
| et al | |||
| In vivo selection of hematopoietic progenitor cells and temozolomide dose intensification in rhesus macaques through lentiviral transduction with a drug resistance gene, The Journal of Clinical Investigation, 2009, v.119, n.7, p.1952-1963 | |||
| CAI S | |||
| et al | |||
| In vivo selection of hematopoietic stem cells transduced at a low |

