Изобретение относится к области биотехнологии, иммунологии и клеточной биологии. Предлагаемая клеточная линия может быть использована для активации и экспансии клеток естественных киллеров (ЕК-клеток) человека при их совместном культивировании. ЕК-клетки человека будут полезны при разработке новых подходов в иммунотерапии злокачественных новообразований.
В настоящее время ведутся активные исследования по использованию иммунных клеток, таких как активированные ЕК-клетки, ЕК- и Т-клетки с химерными антигенными рецепторами (CAR-EK, CAR-T) и другие, для адоптивной иммунотерапии различных вирусных и онкологических заболеваний [1,2]. Основной сложностью при исследовании иммунных клеток, а также при их применении для адоптивной иммунотерапии является трудоемкость их получения в достаточном количестве ex vivo. На данный момент основным способом экспансии иммунных клеток остается их культивирование в среде с добавлением различных цитокинов и ростовых факторов [3,4], однако это не приводит к значительному увеличению количества иммунных клеток и устойчивой активации лимфоцитов. В частности известно, что важную роль в активации и пролиферации иммунных клеток играет интерлейкин 15 (IL-15) цитокин общей гамма-цепи [1,5,6]. Также известно, что белок TNFRSF9L, из надсемейства факторов некроза опухоли, взаимодействуя с собственным рецептором CD 137 способен усиливать пролиферацию, повышать выживаемость и цитолитическую активность иммунных клеток, тем самым стимулируя иммунный ответ против клеток опухолей [7]. Рецептор CD215 (IL-15R alpha) представляет собой широко экспрессируемый трансмембранный гликопротеин массой 60 кДа. Высокоаффинный рецептор к IL-15 может быть использован для усиления сигнала оказываемого IL-15 на активацию и экспансию иммунных клеток [8].
Таким образом, развитие новых иммунотерапевтических подходов для лечения онкологических больных посредством элиминации опухолевых клеток за счет ЕК-клеток является крайне актуальной задачей [9]. Для некоторых типов иммунных клеток, таких как ЕК-клетки, одним из подходов к эффективной генерации и экспансии может быть обеспечение непосредственного контакта с некоторыми чувствительными типами опухолевых клеток, несущими на своей поверхности лиганды к активирующим рецепторам ЕК-клеток [10]. Такие клеточные линии могут служить основой для получения новых рекомбинантных клеточных линий, способных к экспрессии вышеописанных молекул для повышения эффективности экспансии и активации иммунных клеток. Дополнительным способом усиления пролиферативной активности ЕК-клеток с сохранением активированного фенотипа и отсутствием истощения у результирующей популяции, является ко-культивирование ЕК-клеток с рекомбинантными клеточными линиями, в которых стимулирующие цитокины, внесенные с помощью генетической модификации (ГМ), экспрессируются в виде собственных мембран-связанных форм (mbIL15 и др.). Использование такого подхода не требует добавления аналогичных цитокинов в культуральную среду, что упрощает процесс культивирования и делает его более эффективным [1,7,11-14].
Известна [15] искусственная антиген презентирующая клеточная линия (иАПК), в которой на поверхности клеточной мембраны экспрессированы CD137L, CD86, CD64, мембран-связанный IL-15 и CD 19, предназначенная для активации и экспансии CAR-T-клеток с химерным антигенным рецептором. Описываемая клеточная линия получается путем последовательной лентивирусной трансдукции клеток линии К562 генетическими конструкциями (ГК), содержащими CD137L, CD86, CD64, CD 19 и mbIL15. Однако, использование лентивирусной трансдукции не позволяет обеспечить стабильную долговременную экспрессию вводимых рекомбинантных белков на клеточной мембране, так как уровень экспрессии снижается после каждого клеточного деления, а наличие пяти дополнительных ГК негативно сказывается на функциональной активности клеток и их жизнеспособности.
Известна [16] генетически модифицированная клеточная линия К562-OX40L-mbIL 18-mbIL21 предназначенная для активации и амплификации ЕК-клеток. Описанная клеточная линия способна к экспрессии связанного с мембраной IL-18 (mbIL-18), связанного с мембраной IL-21 (mbIL-21) и лиганда рецептора ОХ40. Ген mbIL-18 и ген mbIL-21 клонировали в лентивирусный вектор pCDH-CMV-RFP для создания вектора с целью получения рекомби-нантного лентивируса. После этого полученную плазмиду (pCDH-CMV-RFP-mbIL-1821) трансфицировали в клетки 293FT вместе с упаковочной плазмидой для сборки вирусных частиц. Ген OX40L в свою очередь, клонировали в лентивирусный вектор pLVX-IRES-ZsGreen. Полученную плазмиду (OX40L-pLVX-IRES-ZsGreen) процессировали и проводили последовательную ленти-вирусную трансдукцию клеток линии К562. С помощью флуоресцентного клеточного сортинга отбирали клетки, несущие одновременно красную (RFP) и зеленую (ZsGreen) метки. Недостатком данной клеточной линии является использование лентивирусной трансдукции, которая не позволяет поддерживать экспрессию введенной ГК в течение длительного времени при многократном клеточном делении. Кроме того, использование флуоресцентных белков (RFP и ZsGreen) снижает эффективность трансдукции и увеличивает размер используемой конструкции. Клеточный сортинг, в свою очередь, приводит к нарушению стерильности клеточной культуры и делает ее непригодной для клинического использования.
Известна [17] рекомбинантная стимуляторная клеточная линия, предназначенная для активации и экспансии ЕК-клеток ex vivo. Для получения описанной клеточной линии клетки K562-mb15-41BBL, полученные аналогично иАПК из патента US7435596B2 подвергают последовательной ретровирусной трансдукцией с помощью ретровируса MSCV псевдотипа RD144, содержащего кДНК мембрансвязанного интерлейкина IL-12 и/или IL-18. Недостатком описываемой клеточной линии является отсутствие возможности адекватной селекции клеток, прошедших трансдукцию, а также большее количество итоговых ГМ, что существенно снижает жизнеспособность клеточной линии, а также ее способность к пролиферации и устойчивость к процедуре криоконсервации.
Наиболее близким аналогом, прототипом, выступает известная [18] рекомбинантная клеточная линия иАПК, полученная путем последовательной ретровирусной трансдукции клеток линии К562 ГК, несущими мембран-связанную форму IL- 15 с зеленым флуоресцентным белком (GFP) и белок-лиганд 4-1BBL. Для получения псевдоретровирусных частиц клетки НЕК293Т трансфицировали плазмидой pMSCV-IRES-GFP с клонированными целевыми конструкциями, а также вспомогательными плазмидами pEQ-PAM3(-E) и pRDF. Недостатками данной клеточной линии является использование GFP, что приводит к увеличению размера используемой конструкции и существенно снижает эффективность трансдукции. Также в данном случае для отбора трансдуцированных клеток применяют метод клеточного сортинга, при котором клетки теряют свою стерильность, что делает их непригодными для клинического использования.
Задачей изобретения является получение новой стабильной рекомбинантной клеточной линии хронической миелогенной лейкемии TMDK562-15, которая обладает высокой поверхностной экспрессией мембран-связанной формы химерного белка, состоящего из интерлейкина 15, sushi-домена CD215 и TNFRSF9L, проявляющая способность к активации и пролиферации ЕК-клеток при их совместном культивировании. Это позволит расширить коллекцию рекомбинантных клеточных линий для целей длительного культивирования, стимуляции активации и пролиферации ЕК-клеток человека с высокой жизнеспособностью. Новая рекомбинантная клеточная линия TMDK562-15 на протяжении нескольких месяцев культивирования стабильно экспрессирует в высоких концентрациях белок-лиганд рецептора CD137 (TNFRSF9L) более чем у 95% клеток и мембран-связанный химерный белок, состоящий из IL-15 и части CD215, более чем у 75% клеток. Использование фрагмента sCD215 в составе химерного белка mbIL15/sCD215 позволяет избежать критического увеличения размера ГК, при сохранении влияния на способность получаемой клеточной линии к активации и пролиферации ЕК-клеток.
Технический результат заявляемого изобретения: расширение коллекции рекомбинантных клеточных линий за счет устойчивой экспрессии мембран-связанной формы химерного белка IL15/sCD215 (mbIL15/sCD215) и белка-лиганда TNFRSF9L, которые могут быть использованы для активации и пролиферации ЕК-клеток при их совместном культивировании.
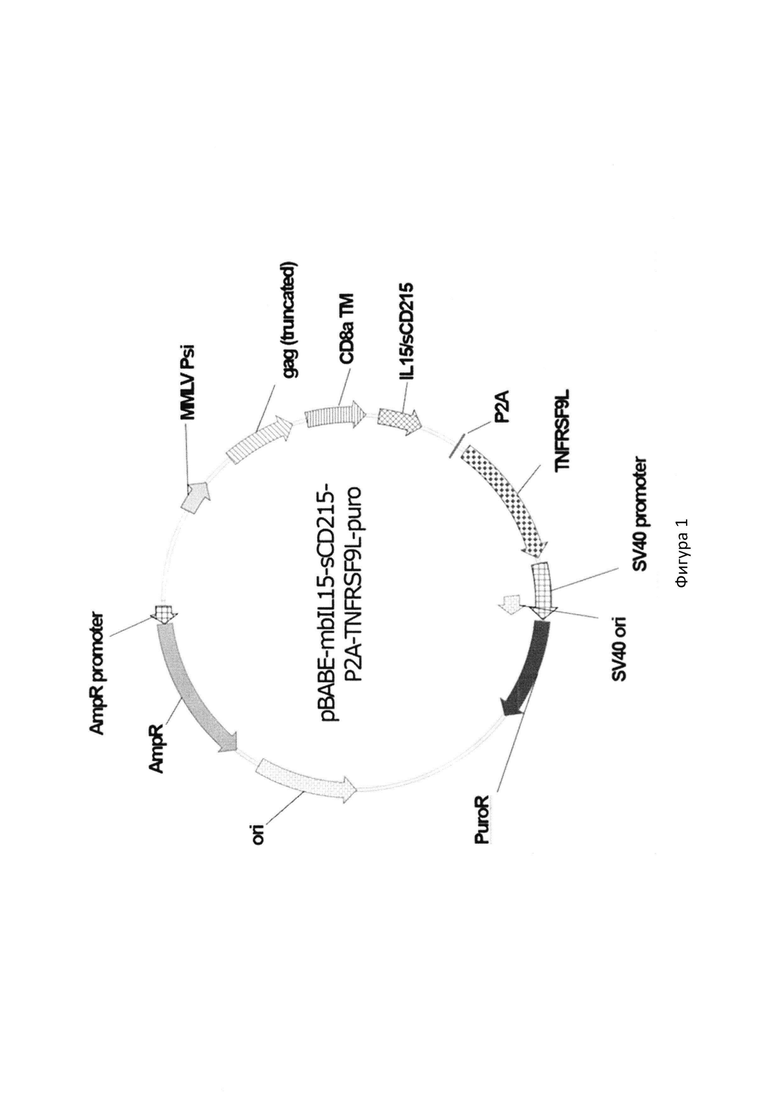
Указанный технический результат достигается получением новой рекомбинантной клеточной линией TMDK562-15, которую получают на основе культуры клеток хронической миелогенной лейкемии К562 [хранится в Российской коллекции клеточных культур позвоночных (РККК П) ИНЦ РАН] путем ретровирусной трансдукции. Для этого при интеграции кДНК кассеты, кодирующей mbIL15/sCD215-TNFRSF9L клетки К562 обрабатывали псевдо-ретровирусными частицами. Их сборку проводили, используя стандартный протокол кальций-фосфатной трансфекции с помощью псевдотипированных клеток линии НЕК293Т за счет поверхностного белка G вируса везикулярного стоматита (VSV). В качестве векторной плазмиды использовали рекомбинантную ретровирусную плазмиду pBABE-mbIL15-sCD215-P2A-TNFRSF9L-puro, кодирующую мембран-связанную форму химерного белка IL15/sCD215 и белок-лиганд TNFRSF9L, с использованием вспомогательных плазмид pUMVC и PCMV-VSV-G. Белок mbIL15/sCD215 получали путем комбинации последовательностей человеческого IL-15, sushi-домена CD215 с трансмембранным доменом CD8a.
Сконструированная рекомбинантная ретровирусная плазмида рВАВЕ-mbIL15-sCD215-P2A-TNFRSF9L-puro имеет следующие характеристики:
- кодирует мембран-связанную форму химерного белка IL15/sCD215 (mbIL15/sCD215)
- кодирует человеческий трансмембранный белок TNFRSF9L;
- состоит из следующих элементов:
а) последовательности, кодирующей основные функционально-структурные элементы костимуляторного домена трансмембранного химерного белка IL15/sCD215/TNFRSF9L;
б) фрагмента ДНК, кодирующего домен, содержащий трансмембранную область CD8 альфа;
в) саморасщепляющейся последовательности Р2А;
г) генетических маркеров:
AmpR - ген ампициллин-резистентности (bla), определяющий устойчивость к ампициллину при трансформации Escherichia coli;
PuroR - селективный ген устойчивости к пуромицину (ген рас Streptomy-ces alboniger);
д) модифицированных генетических элементов (MMLV psi, gag), необходимых для корректного процессинга вирусной РНК и сборки псевдоретровирусных частиц;
е) промотора полиомавируса sv40, обеспечивающего инициацию трансляции маркера PuroR.
Изобретение иллюстрируется следующими фигурами:
На фиг. 1 показана карта генетической конструкции рекомбинантной плазмиды pBABE-mbIL15-sCD215-P2A-TNFRSF9L-puro, где IL15/sCD215, TNFRSF9L, CD8a- последовательности ДНК, кодирующие основные функционально-структурные элементы костимуляторного домена химерного белка mbIL15/sCD215/TNFRSF9L, Р2А - саморасщепляющаяся последовательность, SV40 promoter - промотор полиомавируса sv40, SV40 ori - точка начала репликации для гена PuroR, PuroR - ген устойчивости к пуромицину, ori - последовательность начала общей репликации, AmpR - ген устойчивости к ампициллину, AmpR promoter - промотор гена устойчивости к пенициллину, MMLV Psi - сигнал для паковки ретровируса, являющийся частью вируса мышиного лейкоза, gag (truncated) - укороченная последовательность гена gag ВИЧ-1.
На фиг. 2 показана морфология клеточных линий, полученная с помощью светового инвертированного микроскопа, где:
A. Клетки исходной линии К562;
B. Клетки рекомбинантной линии TMDK562-15
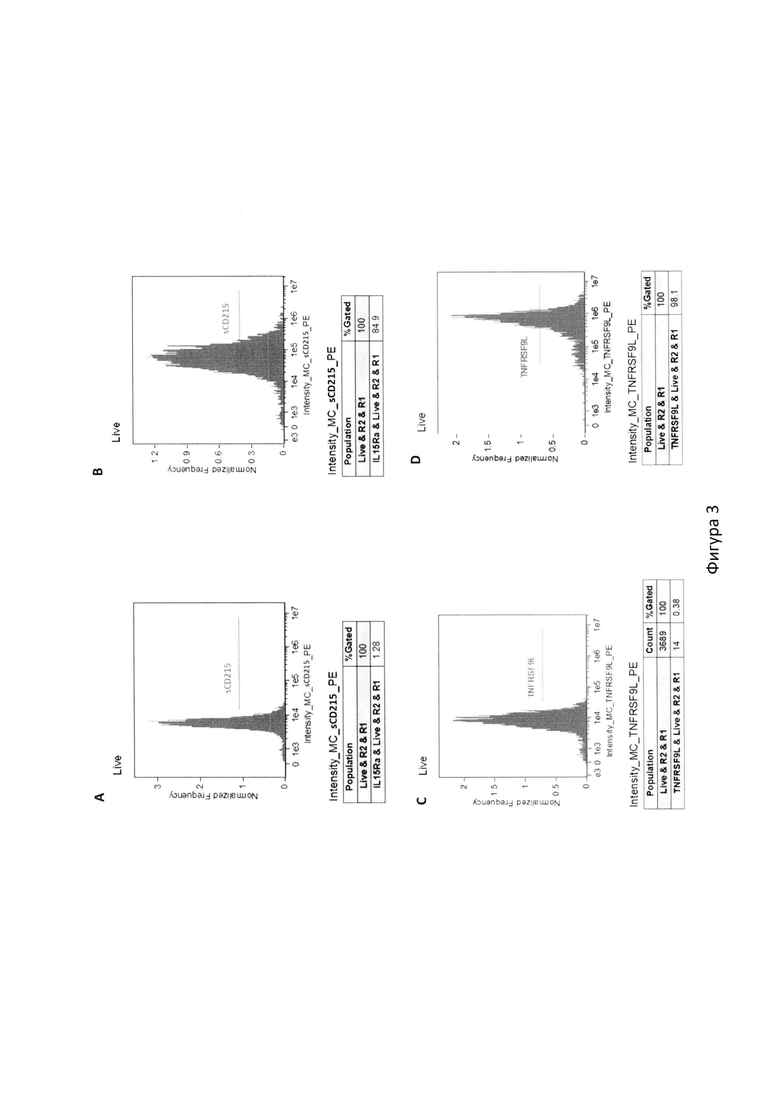
На фиг. 3 с помощью цитометрического анализа показано сравнение экспрессии поверхностных маркеров исходной клеточной линии К562 и рекомбинантной клеточной линии TMDK562-15, где:
A. Доля клеток, экспрессирующих sCD215 в исходной культуре К562;
B. Доля клеток, экспрессирующих sCD215 в культуре рекомбинантных клеток TMDK562-15;
C. Доля клеток, экспрессирующих TNFRS9FL в исходной культуре К562;
D. Доля клеток, экспрессирующих TNFRS9FL в культуре рекомбинантных клеток TMDK562-15;
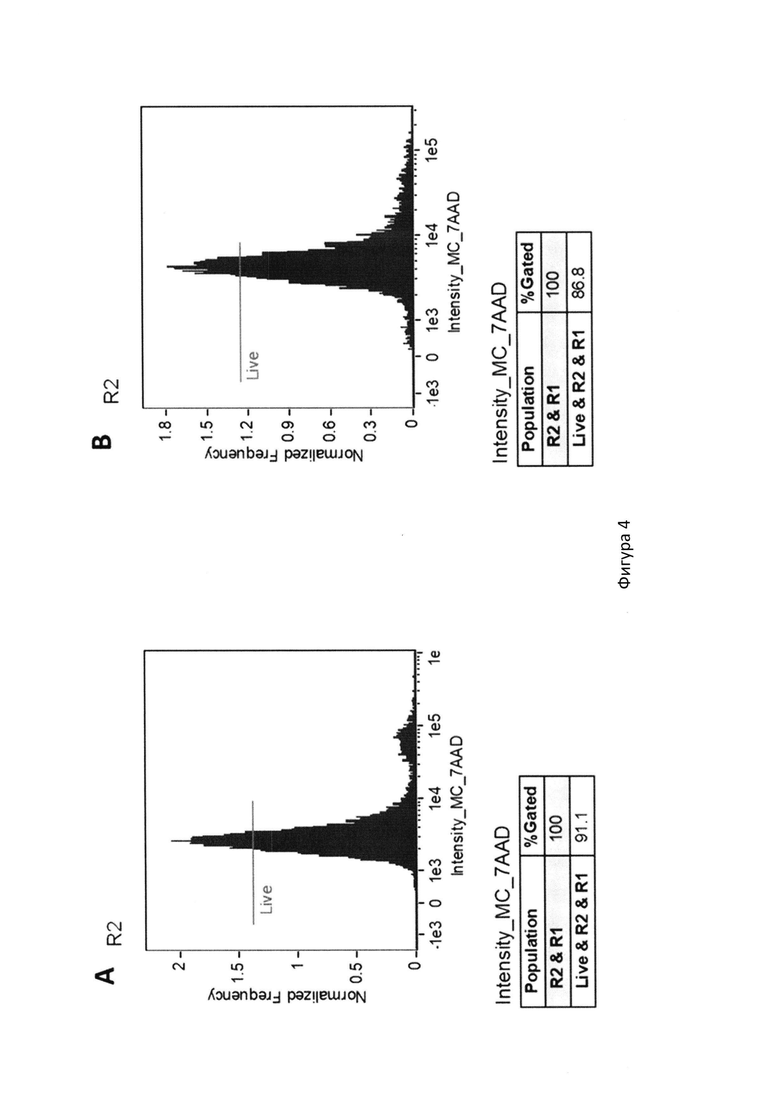
На фиг. 4 с помощью цитометрического анализа с окрашиванием витальным красителем 7AAD показана жизнеспособность клеточной линии TMDK562-15,где:
A. Количество жизнеспособных клеток до облучения;
B. Количество жизнеспособных клеток после облучения;
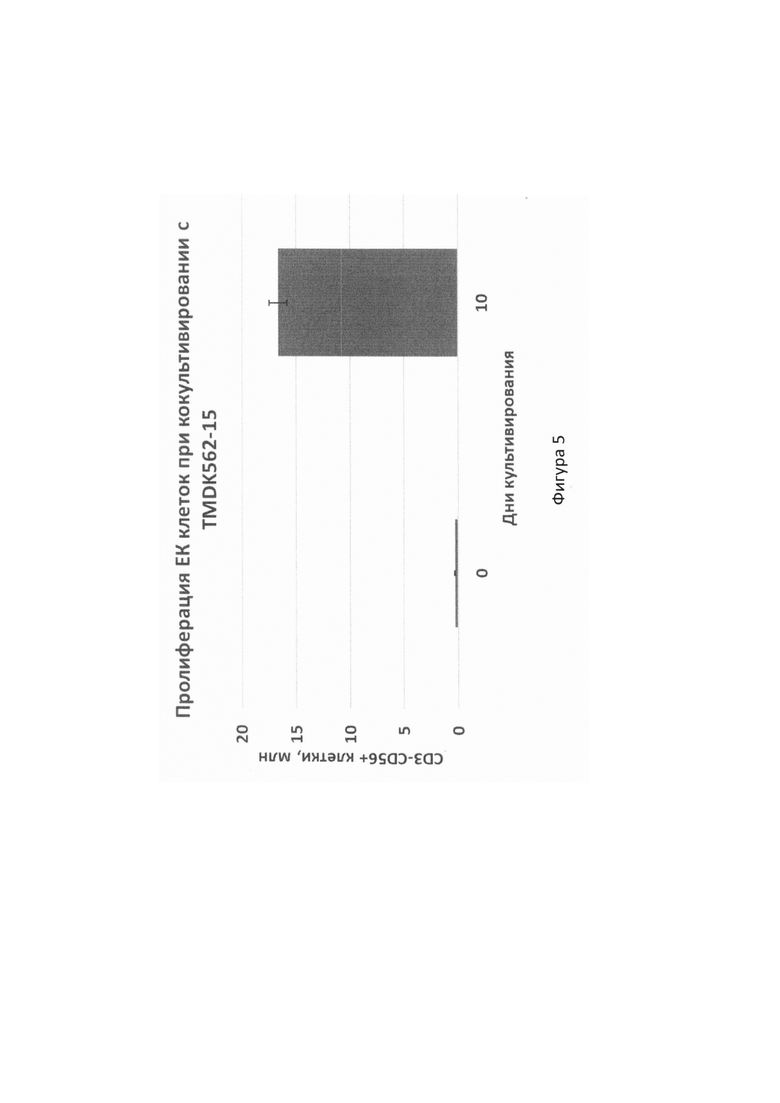
На фиг. 5 с помощью цитометрического анализа поверхностной экспрессии маркеров CD3 и CD56 показан средний уровень пролиферации ЕК-клеток при ко-культивировании мононуклеаров периферической крови доноров с линией TMDK562-15;
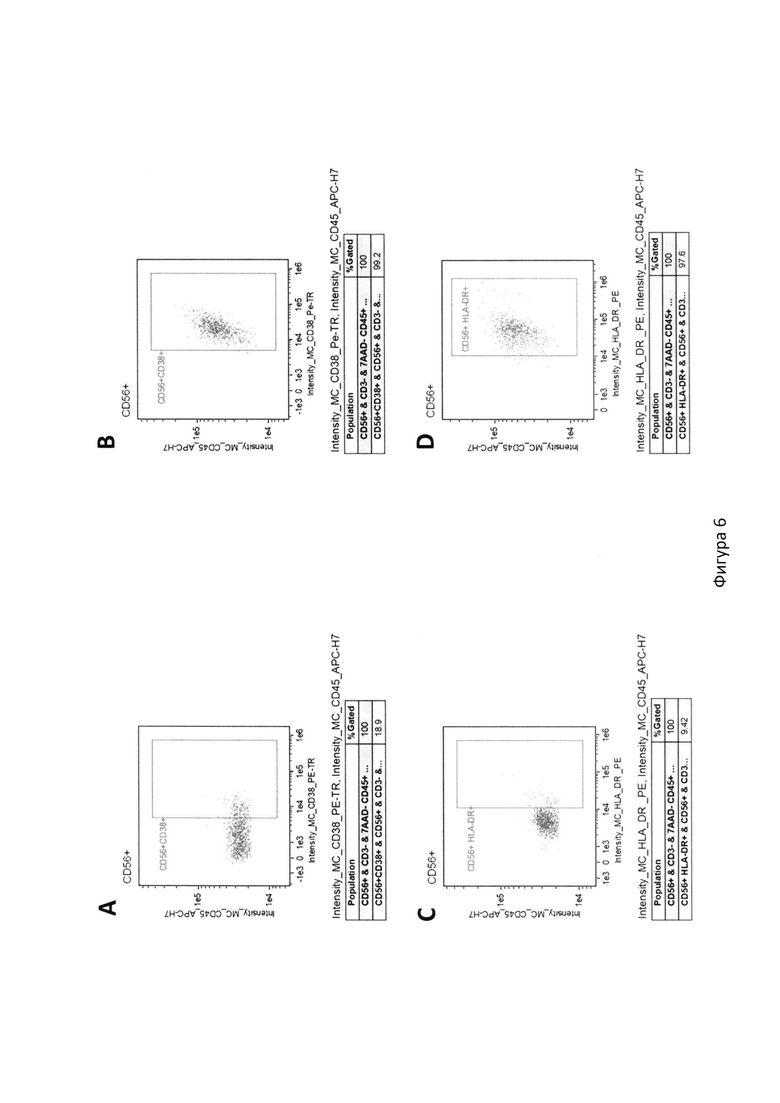
На фиг. 6 с помощью цитометрического анализа показана доля активированных ЕК-клеток, экспрессирующих CD38 и HLA-DR, где:
А. Доля ЕК-клеток, экспрессирующих поверхностный маркер CD38, до совместного культивирования с клеточной линией TMDK562-15;
B. Доля ЕК-клеток, экспрессирующих поверхностный маркер HLA-DR, до совместного культивирования с клеточной линией TMDK562-15;
C. Доля ЕК-клеток, экспрессирующих поверхностный маркер CD38, на 10-ый день совместного культивирования с клеточной линией TMDK562-15;
D. Доля ЕК-клеток, экспрессирующих поверхностный маркер HLA-DR на 10-ый день совместного культивирования с клеточной линией TMDK562-15.
Трансдукцию проводили методом спинокуляции с различными MOI (multiplicity of infection, множественность инфекции) в присутствии 8 мкг/мл полибрена (Sigma, США). Через 48 часов после трансдукции клетки подвергали селекции на среде с добавлением пуромицина (3 мкг/мл) в течение 5 дней. Клетки TMDK562-15 окрашивали антителами к TNFRSF9L (клон REA254, Miltenyi Biotec, Германия), конъюгированными с фикоэритрином (РЕ). Далее, для получения гомогенной mbIL15/sCD215-TNFRSF9L-эксперессирующей линии, клетки клонировали на сортере Sony SH800 (Sony, Япония).
Показано, что полученная клеточная линия миелогенной лейкемии человека TMDK562-15 обладает стабильными морфологическими признаками при культивировании. Она представлена клетками округлой формы, с морфологией больших гранулярных клеток с крупным ядром, отличающимися по величине до 30%, растущих одиночно или образующих скопления (фиг. 2).
Маркерные признаки клеточной линии TMDK562-15 следующие:
- поверхностная экспрессия IL-15, слитого с sushi-доменом белка CD215 наблюдается у более чем 75% клеток;
- поверхностная экспрессия белка TNFRSF9L наблюдается более чем у 95% клеток.
Клеточная линия TMDK562-15 культивируется в питательной среде RPMI1640 (ПанЭко, Россия), в присутствии 10% эмбриональной сыворотки телят (FBS) (GIBCO, США), 100 ед/мл пенициллина и 100 мкг/мл стрептомицина (Capricorn-Scientific, Germany), в атмосфере 5% СО2 при 37°С. Культура имеет суспензионный тип роста. При субкультивировании клетки пересаживают каждые 2-3 дня до концентрации 0,3-0,5 × 106 клеток/мл.
Условия криоконсервации следующие.
Среда КриоМед-М (ПанЭко, Россия). Стандартный режим замораживания 3 × 107 клеток/ампулу при снижении температуры на один градус Цельсия в минуту в программном замораживателе Biofreeze BV-65 (Biofreeze, Франция). Хранение в жидком азоте при температуре -196°С. Размораживание быстрое при 37°С. Для этого 1,5 мл клеточной суспензии разводят в 10 мл среды RPMI1640 или в фосфатно-солевом буфере и осаждают центрифугированием при 800 об/мин, клетки ресуспендируют в ростовой среде RPMI1640 и рассаживают в культуральные флаконы. Жизнеспособность клеток после криоконсервации ≥75% (окраска трипановым синим).
Анализ контаминации.
При длительном культивировании бактерии и грибы в культуре не обнаружены. Анализ уреаплазмы и микоплазмы выполнен методом ОТ ПЦР в режиме реального времени с использованием пары праймеров: 5'GGCGAATGGGTAAGTAACACG3' и 5'CGATAACGCTTGCGACCTAT3' позволяющих выявлять Mycoplasma hyorhinis, fermentans, arginini, orale, genitalium, hominis, pirum, pneumoniae, salivarium, Acholeplasma laidlawii и Ureaplasma urealyticum. Тест на микоплазму и уреаплазму отрицательный.
Пример 1. Получение рекомбинантной клеточной линии TMDK562-15
Клеточную линию TMDK562-15 получали на основе клеточной линии К562 (переданной из Российской коллекции клеточных культур позвоночных (РККК П) ИНЦ РАН) путем ее трансдукции (заражения) псевдоретровирусными частицами. Для получения псевдоретровирусных частиц, ДНК рекомбинантной ретровирусной плазмиды pBABE-mbIL15-sCD215-P2A-TNFRSF9L-puro смешивали с ДНК вспомогательных плазмид pUMVC и PCMV-VSV-G, необходимых для упаковки псевдоретровирусных частиц, в соотношении 4:1:3 и трансформировали клетки НЕК293Т методом кальций-фосфатной трансфекции, после чего культивировали 6 ч в среде IMDM с добавлением 10% FBS, 100 мкг/мл стрептомицина и 100 ед./мл пенициллина в атмосфере 5% СО2 при 37°С. Далее среду в чашках заменяли на новую и инкубировали клетки 48 часов. Затем супернатанты кондиционированной среды фильтровали через 0.45 мкм PES-фильтры и использовали для заражения клеточной линии К562 методом спинокуляции в присутствии 8 мкг/мл полибрена. После заражения проводили селекцию трансдуктантов на среде с добавлением 3 мкг/мл пуромицина в течение 5 дней. Наконец, клетки TMDK562-15 окрашивали антителами к TNFRSF9L (Miltenyi Biotec, Германия), конъюгированными с фикоэритрином (РЕ) и для получения гомогенной mbIL15/sCD215-TNFRSF9L-экспрессирующей линии клетки клонировали на сортере Sony SH800 (Sony, Япония). Поверхностная экспрессия IL15, слитого с sushi-доменом белка CD215, анализированная с помощью метода проточной цитометрии (окрашивание антителами к CD215, клон JM7A4, BioLegend, США), наблюдается у 84,9% клеток, а поверхностная экспрессия белка TNFRSF9L наблюдается у 98,1%) клеток (фиг. 3).
Пример 2. Ограничение пролиферации рекомбинантной клеточной линии TMDK562-15
Клетки TMDK562-15 были культивированы до достижения общего количества в 100 миллионов. Жизнеспособность клеток в суспензии составляла более 90%. Полученная суспензия была перенесена в пробирки объемом 25 мл (Eppendorf, Германия) и подвергнута гамма-облучению с помощью источника на базе кобальта-60 в дозе 90-100 Гр. После облучения, количество жизнеспособных клеток оставляло более 86%. Для оценки уровня пролиферации после облучения, было проведено культивирование облученных клеток в течение 3-х суток, пролиферативная активность отсутствовала, жизнеспособность сохранялась на прежнем уровне (фиг. 4).
Пример 3. Использование генетически модифицированных клеток для экспансии ЕК-клеток
Для оценки влияния TMDK562-15 на экспансию и функциональную активность иммунных клеток, проводили ко-культивирование полученной клеточной культуры с мононуклеарами периферической крови (РВМС). Для этого по стандартной методике на градиенте плотности Hystopague-1077 (Sigma Aldrich, США) из гепаринизированной крови здорового донора выделяли РВМС. Культивирование проводили в питательной среде RPMI1640, содержащей 10% FBS с добавлением IL-2 в концентрации 200 МЕ/мл в CO2-инкуба-торе во влажной атмосфере при 37°С. Клетки TMDK562-15 после гамма-облучения добавляли к культивируемым РВМС на 0 день и 7 день. Доля ЕК-клеток с фенотипом CD3-CD56+ в смеси культивированных РВМС к 10 дню составила в среднем 32,4% и у большинства доноров возрастала в среднем более чем в 55,2 раза (фиг. 5). В результате культивирования доля активированных ЕК-клеток, экспрессирующих маркеры CD38 и HLA-DR увеличивалась до 99,2% и 97,6% соответственно (фиг. 6). Данные клетки могут быть использованы для адоптивной иммунотерапии онкобольных, а также для изучения цитотоксической активности ЕК-клеток.
Источники информации:
1. Denman CJ, Senyukov VV, Somanchi SS, et al. Membrane-bound IL-21 promotes sustained ex vivo proliferation of human natural killer cells. PLoS One. 2012; 7(1): e30264.
2. Abakushina EV, Popova LI, Zamyatnin AA Jr, et al. The Advantages and Challenges of Anticancer Dendritic Cell Vaccines and NK Cells in Adoptive Cell Immunotherapy. Vaccines. 2021; 9(11): 1363.
3. Moseman JE, Foltz JA, Sorathia K, et al. Evaluation of serum-free media formulations in feeder cell-stimulated expansion of natural killer cells. Cytother-apy. 2020;22(6):322-328.
4. Гельм Ю.В., Кузьмина Е.Г., Абакушина E.B. Функциональная активность лимфоцитов здоровых доноров и онкологических больных при культивировании в присутствии ИЛ-2 и ИЛ-15. Бюллетень экспериментальной биологии и медицины. 2019; Т.167(4):471-477
5. Fang F, Xie S, Chen M, et al. Advances in NK cell production. Cell Mol Immunol. 2022; 19(4):460-481.
6. Kweon S, Phan MT, Chun S, et al. Expansion of Human NK Cells Using K562 Cells Expressing OX40 Ligand and Short Exposure to IL-21. Front Immunol. 2019; 10:879.
7. Min В, Yang В, Kim YS, et al. Harnessing novel engineered feeder cells expressing activating molecules for optimal expansion of NK cells with potent antitumor activity. Cell Mol Immunol. 2022; 19(2):296-298.
8. Van den Bergh J, Willemen Y, Lion E, et al. Transpresentation of interleukin-15 by IL-15/IL-15Rα mRNA-engineered human dendritic cells boosts antitumoral natural killer cell activity. Oncotarget. 2015;6(42):44123-44133.
9. Рыбачук B.A., Попова Л.И., Борисов M.A., и др. Функциональная активность NK-клеток при разработке способа адоптивной иммунотерапии онкологических больных. Вопросы онкологии. 2022. Т. 68. №S3. С.151
10. Becker PS, Suck G, Nowakowska P, et al. Selection and expansion of natural killer cells for NK cell-based immunotherapy. Cancer Immunol Immunother. 2016;65(4):477-484.
11. Cho D, Campana D. Expansion and activation of natural killer cells for cancer immunotherapy. Korean J Lab Med. 2009 Apr;29(2):89-96.
12. Klöß S, Oberschmidt O, Morgan M, et al. Optimization of Human NK Cell Manufacturing: Fully Automated Separation, Improved Ex Vivo Expansion Using IL-21 with Autologous Feeder Cells, and Generation of Anti-CD123-CAR-Ex-pressing Effector Cells. Hum Gene Ther. 2017; 28(10):897-913.
13. Kim EK, Ahn YO, Kim S, et al. Ex vivo activation and expansion of natural killer cells from patients with advanced cancer with feeder cells from healthy volunteers. Cytotherapy. 2013; 15(2):231-241.el.
14. Гельм Ю.В., Пасова И.А., Гривцова Л.Ю., и др. Опыт культивирования NK-клеток человека с фидерными клетками in vitro. Медицинская иммунология. 2022;24(3):481-490.
15. Wenju W, Zongliu Н, Benshan X, et al. Artificial antigen presenting cell, construction method thereof and application thereof in chimeric antigen receptor T cell amplification. CN108251376B.
16.  Genetically-modified cell line for NK cell activation and amplification, and use thereof. WO2021251707A1.
Genetically-modified cell line for NK cell activation and amplification, and use thereof. WO2021251707A1.
17. Campana D, Imai C. Modified cell line and method for expansion of NK cell. US7435596B2.
18. Campana D, Takahiro K. Stimulatory cell lines for ex vivo expansion and activation of natural killer cells. WO2018182511A1.
| название | год | авторы | номер документа |
|---|---|---|---|
| Клеточная линия PSCA-CAR-YT, обладающая поверхностной экспрессией химерных антигенных рецепторов и проявляющая цитотоксическую активность по отношению к PSCA-позитивным раковым клеткам человека | 2018 |
|
RU2712901C1 |
| ЕСТЕСТВЕННЫЕ КЛЕТКИ-КИЛЛЕРЫ, РЕЗИСТЕНТНЫЕ К ТРАНСФОРМИРУЮЩЕМУ РОСТОВОМУ ФАКТОРУ БЕТА | 2019 |
|
RU2830178C2 |
| Способ долгосрочного культивирования и экспансии NK-клеток с высокой жизнеспособностью и функциональной активностью | 2021 |
|
RU2794770C1 |
| ОБРАЗОВАНИЕ НОКАУТНЫХ ПЕРВИЧНЫХ И РАЗМНОЖЕННЫХ NK-КЛЕТОК ЧЕЛОВЕКА С ИСПОЛЬЗОВАНИЕМ РИБОНУКЛЕОПРОТЕИНОВ CAS9 | 2019 |
|
RU2808035C2 |
| КОМПОЗИЦИИ Т-КЛЕТОК С НЕДОСТАТОЧНОСТЬЮ РЕЦЕПТОРОВ Т-КЛЕТОК | 2013 |
|
RU2653761C2 |
| СПОСОБЫ EX VIVO ЭКСПАНСИИ ЕСТЕСТВЕННЫХ КЛЕТОК-КИЛЛЕРОВ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2814083C2 |
| Биомедицинский клеточный продукт для терапии злокачественных новообразований | 2023 |
|
RU2813531C1 |
| СПОСОБ ЭКСПАНСИИ NK-КЛЕТОК ЧЕЛОВЕКА С ПОМОЩЬЮ ФИДЕРНЫХ КЛЕТОК | 2021 |
|
RU2781777C2 |
| Рекомбинантная клеточная линия CYTO-CAR-YT-Lact, проявляющая повышенную цитотоксическую активность по отношению к PSCA-позитивным раковым клеткам человека | 2019 |
|
RU2724431C1 |
| УНИВЕРСАЛЬНЫЕ АНТИГЕНПРЕЗЕНТИРУЮЩИЕ КЛЕТКИ И ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2809113C2 |

Настоящее изобретение относится к области биотехнологии, а именно к рекомбинантной клеточной линии миелогенной лейкемии человека TMDK562-15, обладающей поверхностной экспрессией мембран-связанной формы химерного белка, состоящего из интерлейкина 15, комбинации сигнальных молекул sushi-домена CD215 и TNFRSF9L (mbIL15sCD215TNFRSF9L). Изобретение эффективно для активации и пролиферации клеток естественных киллеров (ЕК-клеток) при их совместном культивировании. 6 ил., 3 пр.
Рекомбинантная клеточная линия миелогенной лейкемии человека TMDK562-15, обладающая поверхностной экспрессией мембран-связанной формы химерного белка, состоящего из интерлейкина 15, комбинации сигнальных молекул sushi-домена CD215 и TNFRSF9L, проявляющая способность к активации и пролиферации ЕК-клеток при их совместном культивировании, полученная путем трансдукции клеточной линии К562 псевдоретровирусными частицами на основе рекомбинантной плазмиды pBABE-mbIL15-sCD215-P2A-TNFRSF9L-puro, характеризующейся в соответствии с графической картой, представленной на фиг. 1, следующими признаками: кодирует мембран-связанную форму химерного белка IL15/sCD215 (mbIL15/sCD215) и человеческий трансмембранный белок TNFRSF9L, которая состоит из следующих элементов:
последовательности, кодирующей основные функционально-структурные элементы костимуляторного домена трансмембранного химерного белка IL15/sCD215/TNFRSF9L;
фрагмента ДНК, кодирующего домен, содержащий трансмембранную область CD8 альфа;
саморасщепляющейся последовательности Р2А для посттрансляционного отделения трансмембранных белковых фрагментов;
генетических маркеров AmpR - ген ампициллин-резистентности (bla) и PuroR - селективный ген устойчивости к пуромицину (рас);
модифицированных генетических элементов MMLV psi, gag; промотора sv40 для обеспечения высокого уровня экспрессии используемых последовательностей и корректного процессинга вирусной РНК.
| CN113388042 А, 14.09.2021 | |||
| CN103232973 A, 07.08.2013 | |||
| WO2021251708 A1, 07.06.2021 | |||
| GRANZIN M | |||
| et al | |||
| Shaping of Natural Killer Cell Antitumor Activity by Ex Vivo Cultivation, Front | |||
| Immunol., Sec | |||
| Alloimmunity and Transplantation, 2017, vol | |||
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| АВАКУШИНА Е.В и др | |||
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
