Область техники
Настоящее изобретение относится к применению «готовых к применению» аллогенных терапевтических клеток для иммунотерапии в сочетании с химиотерапией для лечения пациентов с раком. В частности, авторы настоящего изобретения разработали способ конструирования аллогенных Т-клеток, устойчивых к химиотерапевтическим агентам. Терапевтическая полезность, достигаемая благодаря такой стратегии, должна усиливаться синергичным действием между химиотерапией и иммунотерапией. В частности, настоящее изобретение относится к способу модификации Т-клеток путем инактивации по меньшей мере одного гена, кодирующего компонент рецептора Т-клеток и путем модификации указанных Т-клеток для придания им устойчивости к лекарственным препаратам. Настоящее изобретение открывает возможности для стандартных и доступных стратегий адоптивной иммунотерапии для лечения рака.
Уровень техники
Адоптивная иммунотерапия, которая включает перенос аутологичных антиген-специфичных Т-клеток, полученных ex vivo, является многообещающей стратегией для лечения рака. Т-клетки, применяемые для адоптивной иммунотерапии, можно создать либо путем размножения антиген-специфичных Т-клеток, либо путем перенаправления Т-клеток способами генной инженерии (Park, Rosenberg et al. 2011). Перенос специфичных по вирусному антигену Т-клеток является общепризнанным способом, применяемым для лечения ассоциированных с трансплантатами вирусных инфекций и редких злокачественных опухолей, связанных с вирусами. Аналогично, было показано, что выделение и перенос опухоль-специфичных Т-клеток может быть успешным при лечении меланомы. Новые виды специфичности у Т-клеток были успешно сгенерированы путем генетического переноса рецепторов трансгенных Т-клеток или химерных антигенных рецепторов (CAR). CAR представляют собой синтетические рецепторы, состоящие из компонента направленного действия, который связан с одним или более сигнальными доменами в единой гибридной молекуле. CAR позволили успешно перенаправлять Т-лимфоциты на антигены, экспрессируемых на поверхности опухолевых клеток из разных типов злокачественных опухолей, включая лимфомы и солидные опухоли (Jena, Dotti et at. 2010).
Текущий протокол лечения пациентов с применением адоптивной иммунотерапии основан на переносе аутологичных клеток. Согласно данному подходу Т-лимфоциты отбирают у пациента, генетически модифицируют или подвергают селекции ex vivo, культивируют in vitro с целью увеличения числа клеток, при необходимости, и, в конце их вводят путем инфузии пациенту. При способах лечения аутологичными клетками возникают технические и логистические трудности при конкретных приложениях, их создание требует дорогостоящих специализированных объектов и опытных сотрудников, они должны быть созданы в короткое время после постановки диагноза пациенту, и во многих случаях, предварительное лечения пациента приводит к нарушению иммунной функции, например, лимфоциты пациента могут плохо функционировать и присутствовать в очень малом количестве. Из-за указанных трудностей каждый препарат аутологичных клеток пациента по существу является новым продуктом, что приводит к значительным отклонениям в эффективности и безопасности.
В идеальном случае было бы желательным применять стандартизированное средство лечения, при котором аллогенные терапевтические клетки могли бы быть получены заранее, подробно охарактеризованы, и доступны для немедленного введения пациентам. Однако аллогенные Т-клетки получают от индивидуумов, принадлежащих к тому же виду, но генетически отличных. Таким образом, специфичные эндогенных рецепторы Т-клеток (РТЛ) аллогенных клеток распознают ткань хозяина как чужую, что ведет к реакции «трансплантат против хозяина» (ТПХ), которая может приводить к серьезному повреждению ткани и гибели. Рецепторы Т-клеток (РТЛ) являются рецепторами поверхности клетки, которые участвуют в активации Т-клеток в ответ на презентирование антигена. Также как и у молекул иммуноглобулина, вариабельная область альфа- и бета-цепей генерируется посредством V(D)J-рекомбинации, что ведет к созданию большого разнообразия специфичностей по отношению к антигенам в пределах указанной популяции Т-клеток. Однако в отличие от иммуноглобулинов, которые распознают интактный антиген, Т-лимфоциты активируются процессированными пептидными фрагментами в ассоциации с молекулой главного комплекса гистосовместимости (МНС), вводя дополнительный параметр в распознавание антигена Т-клетками, известный как МНС-рестрикция. Распознавание отличий МСН между донором и реципиентом при помощи рецептора Т-клеток приводит к пролиферации Т-клеток и потенциальному развитию ТПХ. Чтобы эффективно применять аллогенные клетки, авторы изобретения инактивировали ген РТЛ альфа или РТЛ бета, что приводит к элиминации РТЛ с поверхности Т-клеток и, таким образом, препятствует распознаванию аллоантигена и, как следствие, развитию ТПХ.
Хотя в областях определения рака и биологии опухолевых клеток был достигнут заметный прогресс, лечение рака на поздних стадиях и метастатического рака остается большой нерешенной проблемой. Цитотоксические хемотерапевтические агенты по-прежнему остаются одним из наиболее широко применяемых и успешно внедряемых противораковых средств. Для уничтожения раковых клеток было разработано несколько цитотоксических агентов, таких как антиметаболиты, алкилирующие агенты, антрациклины, ингибиторы ДНК-метилтрансферазы, соединения платины и веретенные яды. Однако они не демонстрируют одинаково высокой эффективности, и введение указанных агентов с новыми лечебными средствами, такими как средства иммунотерапии, являются проблематичными. Например, химиотерапевтические агенты могут быть вредны для формирования устойчивых противоопухолевых иммунокомпетентных клеток из-за неспецифического токсического профиля данных агентов. Средства лечения на основе малых молекул, нацеленные на пути пролиферации клеток, также могут затруднять формирование противоопухолевого иммунитета. Однако, если химиотерапевтические режимы, которые обладают временной эффективностью, можно было бы сочетать с новыми средствами на основе иммунокомпетентных клеток, можно было бы достичь значительного улучшения противоопухолевой терапии (для обзора (Dasgupta, McCarty et al. 2011)).
Таким образом, для использования «готовых к применению» аллогенных терапевтических клеток в сочетании с химиотерапией, авторы настоящего изобретения разрабатывают способ конструирования аллогенной Т-клетки, устойчивой к химиотерапевтическим агентам. Терапевтическая полезность, достигаемая благодаря такой стратегии, должна усиливаться синергичным действием между химиотерапией и иммунотерапией. Кроме того, устойчивость к лекарственным веществам также может быть полезна вследствие способности к селективному размножению указанной сконструированной Т-клетки, что позволит избежать проблем с неэффективным переносом генов в указанные клетки.
Сущность изобретения
Согласно одному аспекту настоящего изобретения предложены способы конструирования иммунных клеток с приданием им устойчивости к химиотерапевтических препаратам из класса аналогов пуриновых нуклеотидов (АПН), таким как клофарабин и флударабин, так, чтобы их можно было применять при иммунотерапии рака у пациентов, предварительно подвергшихся традиционной химиотерапии. Иммунные клетки могут быть получены от пациента, как в случае TIL (проникающих в опухоль клеток), с целью проведения аутологичного лечения, или от доноров с целью получения аллогенных клеток, которые можно применять для аллогенного лечения.
В последнем случае, когда иммунные клетки являются Т-клетками, согласно настоящему изобретению также предложены способы конструирования Т-клеток, которые получают и устойчивыми к химиотерапевтическим препаратам, и аллогенными. Такие способы включают этап инактивации по меньшей мере одного гена, кодирующего компонент рецептора Т-клеток (РТЛ), в частности гены РТЛ альфа, РТЛ бета, наряду с инактивацией гена чувствительности к лекарственным препаратам, такого как гены dcK и HPRT.
Согласно другому аспекту устойчивость к лекарственным препаратам может быть придана Т-клеткам путем экспрессии гена лекарственной устойчивости. Было выявлено, что устойчивость к лекарственным препаратам клеткам согласно настоящему изобретению придают вариантные аллели нескольких генов, таких как дигидрофолатредуктаза (ДГФР), инозинмонофосфатдегидрогеназа 2 (IMPDH2), кальциневрин- или метилгуанинтрансфераза (MGMT).
Настоящее изобретение включает изолированные клетки или линии клеток, получаемые при помощи способа согласно настоящему изобретению, более конкретно, изолированные иммунные клетки, содержащие любые белки, полипептиды, аллельные варианты, измененные или делетированные гены или векторы, описанные в настоящей заявке.
Иммунные клетки согласно настоящему изобретению или линии клеток могут дополнительно содержать экзогенные рекомбинантные полинуклеотиды, в частности CAR или гены «самоубийства», или они могут содержать измененные или делетированные гены, кодирующие узловые белки или их лиганды, которые вносят вклад в их эффективность, в качестве терапевтического продукта, в идеальном случае, в качестве «готового продукта». Согласно другому аспекту настоящее изобретение относится к способу лечения или профилактики рака у пациента путем введения сконструированной иммунной клетки, получаемой при помощи описанных выше способов.
- На Фигуре 1 показана схема метаболических путей и клеточной токсичности аналогов пуриновых нуклеотидов (АПН); инактивация фермента дезоксицитидинкиназы (dCK) придает устойчивость к препаратам клофарабину и флударабину;
- На Фигуре 2 показано, что инактивация фермента гипоксантин- гуанинфосфорибозилтрансферазы (ГГФТ) придает устойчивость к препаратам 6-мекраптопурин (6МР) и 6-тиогуанин (6TG);
- На Фигуре 3А изображена общая архитектура гена dCK в отношении экзонов и интронов, и
- На Фигуре 3В показаны последовательности целевых сайтов TALE-нуклеазы, расположенных для пар TALE-нуклеаз в экзоне 2 dCK;
- На Фигуре 4 показана последовательность действий, выполняемая при генерировании и описании свойств Т-лимфцоитов с нокаутом гена ГГФТ; D0 обозначает день 0, Dn обозначает день n; Т7 обозначает эндо Т7;
- На Фигуре 5 показаны результаты, полученные при анализе с эндо Т7 для проверки процессинга гена dCK; верхняя полоса соответствует непроцессированному гену dCK дикого типа, а 2 нижних полосы соответствуют процессированному гена dCK;
- На Фигуре 6 показано наращивание Т-клеток с нокаутом гена dCK, обработанных 5 мкг и 10 мкг мРНК, кодирующей TALE-нуклеазу dCK2, и контрольных Т-клеток дикого типа 1 и 2 в течение периода 14 дней после электропорации.
- На Фигуре 7 показан анализ с эндо Т7, проводимый на 8-ой день (D8) для проверки инактивации dCK в Т-клетках (путем применения 5 мкг пары TALE-нуклеазы dCK 2) в присутствии 1 мкМ клофарабина (+) или в отсутствии клофарабина (-);
- На Фигуре 8 показан процент жизнеспособности клеток дикого типа и с нокаутом гена dCK (обработанных 5 мкг и 10 мкг мРНК, кодирующей пару dCK2-TALE-нуклеаза), культивируемых в течение двух дней в присутствии возрастающего количества клофарабина (10 нМ - 10 мкМ). Данный график позволяет определить IC50 (концентрацию полумаксимального ингибирования) клофарабина для обеих популяций клеток;
- На Фигуре 9 показаны 2 последовательности действий, применяемые при генерировании и описании свойств устойчивых к клофарабину аллогенных Т-клеток; на верхней панели показан случай, когда проводился отбор по чувствительности к лекарственному препарату, в отличие от нижней панели, где отбор по чувствительности к лекарственному препарату не проводился; день 0 (D0) - это день, когда была проведена двойная электропорация TALE-нуклеазой TRAC и dCK;
- На Фигуре 10 показан анализ с эндо Т7 для генетической проверки эффективности двойного нокаута генов dCK/TRAC в Т-клеток в разные времена после электропорации (D1, D3 и D6). Праймеры, применяемые для каждого локуса, приведены в примере, для Т-клеток с простым нокаутом dCK (+-) и Т-клеток с простым нокаутом TRAC (-+), Т-клеток с двойным нокаутом dCK/TRAC (++) и Т-клеток дикого типа (-); нижние полосы означают правильный процессинг генов dCK и TRAC;
- На Фигуре 11 показан анализ с эндо Т7 и данные глубокого секвенирования для проверки эффективности инактивации dCK в присутствии (+) или отсутствии (-) клофарабина, с (+) или без (-) инактивации TRAC, легенда такая же, как и на Фигуре 10; для оценки процента вставок/делеций в локусе dCK проводили оценку частоты вставок;
- На Фигуре 12А показан эксперимент для контроля мечения, проведенный с Т-клетками в присутствии (меченные Т-клетки) или отсутствии анти-РТЛ mAb-РЕ (немеченые Т-клетки); на Фигуре 12В отслеживаются TCAR-негативные клетки, отобранные после инкубации с присутствии или в отсутствии клофарабина, до и после очистки Т-клеток с нокаутом TRAC. Указанные клетки также инактивировали по гену dCK;
- На Фигуре 13 показана скорость роста для Т-клеток с простым нокаутом dCK и TRAC и Т-клеток с двойным нокаутом dCK/TRAC по сравнению с Т-клетками дикого типа в отсутствии клофарабина в течение 12 дней после электропорации;
- На Фигуре 14 показаны кривые скорости роста для Т-клеток с двойным нокаутом CAR dCK/TRAC в среде, содержащей разные дозы клофарабина (от 0,1 до 10 мкМ) по сравнению с Т-клетками CAR (с клофарабином или без него) в течение 11 дней;
- На Фигуре 15 показан процент жизнеспособности для Т-клеток с простым нокаутом dCK или TRAC, Т-клеток с двойным нокаутом dCK/TRAC по сравнению с Т-клетками дикого типа в среде, содержащей разные дозы клофарабина (от 0,1 до 100 мкМ); данный график позволяет определить IC50 для клофарабина для каждой популяции Т-клеток;
- На Фигуре 16 показан процент специфической цитотоксичности для Т-клеток с двойным нокаутом CAR dCK/TRAC по сравнению с Т-клетками CAR FMC63 (и те, и другие экспрессируют CD19-антиген) относительно Т-клеток с двойным нокаутом dCK/TRAC (без CAR, то есть не экспрессируют CD19-антиген) и Т-клеток дикого типа (без нокаута и без CAR);
- На Фигуре 17 показан процент жизнеспособности для Т-клеток с CAR с двойным нокаутом dCK/TCAR по сравнению с контрольными Т-клетками с CAR, когда указанные клетки инкубировали при возрастающих дозах клофарабина (10 нг - 10 мкг, верхний график), и флударабина (10 мкМ - 100 мкМ, нижний график). Данные графики позволяют определить IC50 и для клофарабина, и для флударабина;
- На Фигуре 18 показан анализ с эндо Т7 в день 2 (D2) с целью генетической проверки эффективности инактивации dCK в клетках Дауди (+) (применяли 5 мкг мРНК, кодирующей TALE-нуклеазу dCK) в сравнении с клетками дикого типа (-). Верхняя полоса соответствует непроцессированному гену dCK, тогда как 2 нижних полосы - продуктам инактивации dCK;
- На Фигуре 19 показана скорость роста (выраженная как ×106 клеток) в течение 7 дней у клеток Дауди с простым нокаутом dCK и для клеток Дауди дикого типа в отсутствии или присутствии возрастающих количеств клофарабина (0,1-1 мкМ);
- На Фигуре 20 показана архитектура целого гена ГГФТ (в отношении экзонов и интронов) и локализация разных целевых сайтов TALE-нуклеазы (все из них в экзоне 2);
- На Фигуре 21 показана последовательность действий, которая применяется при генерировании и описании свойств Т-клеток с нокаутом гена ГГФТ;
- На Фигуре 22 показан анализ с эндо Т7 с целью проверки инактивации гена ГГФТ в Т-клетках парами TALE-нуклеаза-ГГФТ №1 и Т-парой №2 (исследовались 2 дозы: 5 мкг и 10 мкг), День 4 (D4);
- На Фигуре 23 показана скорость роста (выраженная как ×106 клеток) в течение 13 дней у Т-клеток с простым нокаутом ГГФТ, с применением 5 или 10 мкг пары TALE-нуклеаза- ГГФТ1 (ГГФТ1) или пары TALE-нуклеаз ГГФТ2 в сравнении с Т-клетками дикого типа - контроль 1 и контроль 2;
- На Фигуре 24 показан анализ с эндо Т7 с целью проверки инактивации гена ГГФТ в Т- клетках с применением 5 или 10 мкг пары TALE-нуклеаза-ГГФТ №1 (TALE-нуклеаза ГГФТ 1) в сравнении с Т-клетками дикого типа [обозначены как (-)] в Д8/ и Д18, когда указанные клетки инкубировались в 1 мкМ препарата 6 TG;
- На Фигуре 25 показан анализ с эндо Т7 с целью проверки инактивации гена ГГФТ в Т- клетках в присутствии или отсутствии 4G7 CAR, данный анализ проводился без селекции на 6TG;
- На Фигуре 26 показан процент специфической цитотоксичности в отношении Т-клеток CAR с нокаутом гена ГГФТ по сравнению с Т-клетками CAR 4G7 (оба типа экспрессируют С019-антиген) и Т-клетки дикого типа (без нокаута и без CAR);
- На Фигуре 27 показан процент жизнеспособности для Т-клеток CAR с нокаутом гена ГГФТ по сравнению с Т-клетками дикого типа при возрастающих дозах препарата 6TG (10 нг-50 мкМ).
Подробное описание изобретения
Если в настоящей заявке не указано другое, все применяемые технические и научные термины имеют те же значения, что и обычно под ними понимают специалисты в области генной терапии, биохимии, генетики и молекулярной биологии.
Все способы и материалы, сходные или эквивалентные способам и материалам, описываемым в настоящей заявке, можно применять при реализации или проверке настоящего изобретения, вместе с подходящими способами и материалами, описанными в настоящей заявке. Все публикации, заявки на патенты, патенты и другие литературные источники, упоминаемые в настоящей заявке, включены посредством ссылки на их полное содержание. В случае конфликта настоящее описание, включая определения, будет являться приоритетным. Кроме того, материалы, способы и примеры являются лишь иллюстративными, и не предназначены для ограничения, если не указано другое.
Реализация настоящего изобретения должна достигаться, если не указано другое, традиционными способами клеточной биологии, культур клеток, молекулярной биологии, трансгенной биологии, микробиологии, рекомбинантных ДНК и иммунологии, которые находятся в компетенции специалистов данной области техники. См., например, Current Protocols in Molecular Biology (Frederick M. AUSUBEL, 2000, Wiley and son Inc, Library of Congress, USA); Molecular Cloning: A Laboratory Manual, Third Edition, (Sambrook et al, 2001, Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press); Oligonucleotide Synthesis (M. J. Gait ed., 1984); Mullis et al. U.S. Pat. No. 4,683,195; Nucleic Acid Hybridization (B.D. Harries & S.J. Higgins eds. 1984); Transcription And Translation (B.D. Hames & S.J. Higgins eds. 1984); Culture Of Animal Cells (R.I. Freshney, Alan R. Liss, Inc., 1987); Immobilized Cells And Enzymes (IRL Press, 1986); B. Perbal, A Practical Guide To Molecular Cloning (1984); the series, Methods In ENZYMOLOGY (J. Abelson and M. Simon, eds.-in-chief, Academic Press, Inc., New York), в частности, Vols.154 and 155 (Wu et al. eds.) и Vol.185, "Gene Expression Technology" (D. Goeddel, ed.); Gene Transfer Vectors For Mammalian Cells (J.H. Miller and M.P. Calos eds., 1987, Cold Spring Harbor Laboratory); Immunochemical Methods In Cell And Molecular Biology (Mayer and Walker, eds., Academic Press, London, 1987); Handbook Of Experimental Immunology, Volumes I-IV (D.M. Weir and С.C. Blackwell, eds., 1986); и Manipulating the Mouse Embryo, (Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1986).
- Т-клетки, устойчивые к лекарственным препаратам
В настоящей заявке термины «терапевтический агент», «химиотерапевтический агент» или «лекарственный препарат» относятся к соединению или его производному, которое может взаимодействовать с раковой клеткой, и как следствие снижать пролиферативный статус клетки и/или уничтожать клетку. Примеры химиотерапевтических агентов включают без ограничений алкилирующие агенты (например, циклофосфамид, ифосамид), метаболические антагонисты (например, антиметаболиты пуриновых нуклеозидов, такие как клофарабин, флударабин или 2'-дезоксиаденозин, метотрексат (МТХ), 5-фторурацил или его производные), противоопухолевые антибиотики (например, митомицин, адриамицин), получаемые из растений противоопухолевые агенты (например, винкристин, виндезин, таксол), цисплатин, карбоплатин, этопоксид и подобные. Такие агенты могут также включать без ограничений противоопухолевые агенты ТРИМЕТОТРИКСАТ™ (ТМТХ), ТЕМОЗОЛОМИД™, РАЛТРИТРЕКСЕД™, S-(4-нитробензил)-6-тиоинозин (NBMPR), 6-бензил-гуанидин (6-BG), бис-хлорнитрозмочевина (BCNU) и КАМПТОТЕЦИН™, или терапевтическое производное любого из них.
В настоящей заявке под клеткой, которая «устойчива или толерантна» к агенту, понимают клетку, которая была генетически модифицирована так, что указанная клетка пролиферирует в присутствии некоторого количества агента, который ингибирует или предупреждает пролиферацию клетки без модификации.
Экспрессия генов лекарственной устойчивости
Согласно конкретному варианту реализации указанную устойчивость к лекарственному препарату можно придать Т-клетке путем экспрессии по меньшей мере одного гена лекарственной устойчивости. Под указанным геном лекарственной устойчивости понимают нуклеотидную последовательность, которая кодирует «устойчивость» к агенту, такому как химиотерапевтический агент (например, метотрексат). Иными словами, экспрессия гена лекарственной устойчивости в клетке позволяет клеткам пролиферировать в присутствии указанного агента в большей степени, чем пролиферирует соответствующая клетка без гена лекарственной устойчивости. Ген лекарственной устойчивости согласно настоящему изобретению может кодировать устойчивость к антиметаболиту, метотрексату, винбластину, цисплатину, алкилирующим агентам, антрациклинам, цитотоксическим антибиотикам, антииммунофилинам, их аналогам или производным и т.п.
Было обнаружено несколько генов лекарственной устойчивости, которые потенциально можно применять для придания лекарственной устойчивости целевых клеток (Takebe, Zhao et al. 2001; Sugimoto, Tsukahara et al. 2003; Zielske, Reese et al. 2003; Nivens, Felder et al. 2004; Bardenheuer, Lehmberg et al. 2005; Kushman, Kabler et al. 2007).
Одним из- примеров гена лекарственной устойчивости также может быть мутант или модифицированная форма дигидрофолат-редуктазы (ДГФР). ДГФР является ферментом, который участвует в регулировании количества тетрагидрофолата в клетке и важен для синтеза ДНК. В клинике в качестве противоопухолевых агентов применяются такие аналоги фолата как метотрексат (МТХ), которые ингибируют ДГФР. Были описаны разные мутантные формы ДГФР, которые обладают повышенной устойчивостью к ингибированию антифолатами, применяемыми в терапии. Согласно конкретному варианту реализации ген лекарственной устойчивости согласно настоящему изобретению может быть нуклеотидной последовательностью, кодирующей мутантную форму ДГФР человека дикого типа (SEQ ID №: 14, GenBank: ААН71996.1), которая содержит по меньшей мере одну мутацию, придающую устойчивость к антифолатным препаратам, таким как метотрексат. Согласно конкретному варианту реализации мутантная форма ДГФР содержит по меньшей мере одну мутированную аминокислоту в положении G15, L22, F31 или F34, предпочтительно, в положении L22 или F31 ((Schweitzer, Dicker et al. 1990); заявка на международный патент WO 94/24277; патент США US 6,642,043). Согласно конкретному варианту реализации указанная мутантная форма ДГФР содержит две мутированные аминокислоты в положении L22 и F31. Соответствие положений аминокислот, описываемых в настоящей заявке, часто выражают для положений аминокислот в форме полипептида ДГФР дикого типа, последовательность которого указана в SEQ ID №: 14. Согласно конкретному варианту реализации остаток серина в положении 15 предпочтительно заменен остатком триптофана. Согласно другому конкретному варианту реализации остаток лейцина в положении 22 предпочтительно заменен аминокислотой, которая будет нарушать связывание мутантной ДГФР с антифолатами, предпочтительно остатками незаряженных аминокислот, таких как фенилаланин или тирозин. Согласно другому конкретному варианту реализации остаток фенилаланина в положениях 31 или 34 предпочтительно заменен малой гидрофильной аминокислотой, такой как аланин, серии или глицин.
В настоящей заявке термин «антифолатный агент» или «аналоги фолата» относится к молекуле, которая направленно нарушает метаболический путь фолата на определенном уровне. Примеры антифолатных агентов включают, например, метотрексат (МТХ); аминоптерин; триметрексат (Неутрексин™); эдатрексат; N10-пропаргил-5,8-дидеазофолиевую кислоту (СВ3717); ZD1694 (Тумодекс), 5,8-дидеазаизофолиевую кислоту (IAHQ); 5,10- дидеазатетрагидрофолиевую кислоту (DDATHF); 5-деазафолиевую кислоту; РТ523 (N альфа-(4- амино-4-дезоксипретроил)-N дельта-гемифталоил-L-орнитин); 10-этил-10-деазааминоптерин (DDATHF, ломатрексол); пиритрексим; 10-EDAM; ZD1694; GW1843; пеметрексат и PDX (10- пропаргил-10-деазааминоптерин).
Другим примером гена лекарственной устойчивости может также послужить мутантная или модифицированная форма ионизин-5'-монофосфат-дегидрогеназы II (IMPDH2), лимитирующего скорость фермента в синтезе de novo гуанозин-нуклеотидов. Мутантной или модифицированной формой является ген IMPDH, устойчивый к ингибиторам. Ингибиторами IMPDH могут быть микофенольная кислота (МФК) или ее пролекартсво - микофенолят мотефил (ММФ). Мутантная IMPDH2 может содержать по меньшей мере одну, предпочтительно две мутации в сайте связывания MAP IMPDH2 человека дикого типа (SEQ ID №: 15; NP_000875.2), которые приводят к значимому увеличению резистентности к ингибитору IMPDH. Мутации предпочтительно присутствуют в положениях Т333 и/или S351 (Yam, Jensen et al. 2006; Sangiolo, Lesnikova et al. 2007; Jonnalagadda, Brown et al. 2013). Согласно конкретному варианту реализации остаток треонина в положении 333 заменен на остаток изолейцина и остаток серина в положении 351 заменен на остаток тирозина. Соответствие положений аминокислот, описываемых в настоящей заявке, часто выражают относительно положений аминокислот в форме полипептида IMPDH2 человека дикого типа, последовательность которого приведена в SEQ ID №: 15.
Еще одним геном лекарственной устойчивости является мутантная форма калциневрина. Кальциневрин (РР2В) представляет собой обильно экспрессируемую фосфатазу серина/треонина белков, которая участвует во многих биологических процессах, и которая играет центральную роль в активации Т-клеток. Кальциневрин - гетеродимер, состоящий из каталитической субъединицы (CnA; три изоформы) и регуляторной субъединицы (CnB; две изоформы). После контактирования с рецептором Т-клеток кальциневрин дефосфорилирует фактор транскрипции NFAT, позволяя ему транслоцироваться в ядро и активировать ключевой целевой ген, такой как ИЛ2. FK506 в комплексе с FKBP12 или циклоспорин A (CsA) в комплексе с CyPA блокируют доступ NFAT к активному центру кальциневрина, препятствуя его дефосфорилированию и тем самым ингибируя активацию Т-клеток (Brewin, Mancao et al. 2009). Геном лекарственной устойчивости согласно настоящему изобретению может быть нуклеотидная последовательность, кодирующая мутантную форму кальциневрина, устойчивую к ингибитору кальциневрина, такому как FK506 и/или CsA. Согласно конкретному варианту реализации указанная мутантная форма может содержать по меньшей мере одну мутированную аминокислоту в гетеродимере кальциневрина дикого типа в положениях V314, Y341, М347, Т351, W352, L354, K360, предпочтительно двойные мутации в положениях Т351 и L354 или V314 и Y341. Согласно конкретному варианту реализации остаток валина в положении 341 может быть заменен на остаток лизина или аргинина, остаток тирозина в положении 341 может быть заменен остатком фенилаланина; метионин в положении 347 может быть заменен остатком глутаминовой кислоты, аргинина или триптофана; треонин в положении 352 может быть заменен остатком цистеина, глутаминовой кислоты или аланина; остаток серина в положении 353 может быть заменен остатком гистидина или аспарагина, лейцин в положении 354 может быть заменен остатком аланина; лизин в положении 360 может быть заменен остатком аланина или фенилаланина в последовательности SEQ ID №: 16. Соответствие положений аминокислот, описываемых в настоящей заявке, часто выражают относительно положений аминокислот в форме гетеротримера кальциневрина человека дикого типа - полипептида, последовательность которого приведена в SEQ ID №: 16 (GenBank: АСХ34092.1).
Согласно конкретному варианту реализации указанная мутантная форма может содержать по меньшей мере одну мутированную аминокислоту в гетеродимере кальциневрина b дикого типа в положениях: V120, N123, L124 или K125, предпочтительно двойные мутации в положениях L124 и K125. Согласно конкретному варианту реализации валин в положении 120 может быть заменен на остаток серина, аспарагиновой кислоты, фенилаланина или лейцина; аспарагин в положении 123 может быть заменен на остаток триптофана, лизина, фенилаланина, аргинина, гистидина или серина; лейцин в положении 124 может быть заменен на остаток треонина; лизин в положении 125 может быть заменен на остаток аланина, глутаминовой кислоты, триптофана, или после лизина в положении 125 в аминокислотной последовательности SEQ ID №: 17 могут быть добавлены два остатка, такие как лейцин-аргинин или изолейцин-глутаминовая кислота. Соответствие положений аминокислот, описываемых в настоящей заявке, часто выражают относительно положений аминокислот в форме гетеротдимера кальциневрина b человека дикого типа - полипептида, последовательность которого приведена в SEQ ID №: 17 (GenBank: АСХ34095.1).
Еще одним геном лекарственной устойчивости является ген O-(6)-метилгуанин- метилтрансферазы (MGMT), кодирующий алкилгуанин-трансферазу человека (hAGT). AGT является белком репарации ДНК, который придает устойчивость к цитотоксическому действию алкилирующих агентов, таких как нитрозмочевина и темозоломид (TMZ). 6-бензилгуанин (6- BFG) является ингибитором AGT, который потенцирует токсичность нитрозомочевины, и его вводят совместно с TMZ для потенциирования цитотоксического действия данного агента. Несколько мутантных форм MGMT, которые кодируют варианты AGT, обладают высокой устойчивостью к инактивации под действием 6-BG, но сохраняют свою способность к репарации ДНК (Maze, Kurpad et al. 1999). Согласно конкретному варианту реализации мутантная форма AGT может содержать мутированную аминокислоту в AGT дикого типа в положении Р140, в аминокислотной последовательности SEQ ID №: 18 (UniProtKB: Р16455). Согласно предпочтительным вариантам реализации указанный пролин в положении 140 заменен остатком лизина.
Еще одним геном лекарственной устойчивости может быть ген белка 1 множественной лекарственной устойчивости (MDR1). Указанный ген кодирует мембранный гликопротеин, известный под названием Р-гликопротеина (P-GP), участвующего в транспорте метаболических побочных продуктов через мембрану клетки. Белок P-GP проявляет широкую специфичность в отношении нескольких структурно не родственных химиотерапевтических агентов. Таким образом, клеткам можно придать устойчивость к лекарственным препаратам путем экспрессии нуклеотидной последовательности, которая кодирует MDR-1 (NP_000918).
Геном лекарственной устойчивости могут быть гены цитотоксических антибиотиков, таких как ген ble или ген mcrA. Эктопическая экспрессия гена ble или mcrA в иммунной клетке дает селективное преимущество, когда на клетку воздействует химиотерапевтический агент, соответственно, блеомицин или митомицин С.
Самым практичным подходом в генной терапии является добавление гена в сконструированную Т-клетку путем эффективной доставки при помощи векторов, предпочтительно вирусного вектора. Таким образом, согласно конкретному варианту реализации указанный ген лекарственной устойчивости может экспрессироваться в клетке путем введения трансгена, предпочтительно кодируемого по меньшей мере одним вектором, в клетку.
Случайное встраивание генов в геном может приводить к неадекватной экспрессии встроенного гена или гена, расположенного близко к сайту встраивания. Специфическая генная терапия с применением гомологичной рекомбинации экзогенной нуклеиновой кислоты, содержащей эндогенные последовательности, с генами-мишенями в специфические сайты в геноме, позволяет достичь конструирования устойчивых Т-клеток. Как было описано выше, этап генетической модификации в указанном способе может включать этап введения в клетки экзогенной нуклеиновой кислоты, содержащей по меньшей мере одну последовательность, кодирующую ген лекарственной устойчивости и часть эндогенного гена, такой как гомологичная рекомбинация, которая возникает между эндогенным геном и экзогенной нуклеиновой кислотой. Согласно конкретному варианту реализации указанный эндогенный ген может быть геном «лекарственной устойчивости» дикого типа, например, как после гомологичной рекомбинации, ген дикого типа заменяется мутантной формой гена, который придает устойчивость к лекарственному препарату.
Известно, что разрывы связи между нуклеотидами увеличивают частоту гомологичной рекомбинации. Таким образом, согласно конкретному варианту реализации способ согласно настоящему изобретению также включает этап экспрессии в клетке редкощепящей эндонуклеазы, которая способна расщеплять целевую последовательность в эндогенном гене. Указанный эндогенный ген может кодировать, например, DHFR, IMPDH2, кальциневрин или AGT. Указанной редкощепящей эндонуклеазой может быть TALE-эндонуклеаза, нуклеаза с цинковыми пальцами, эндонуклеаза CRISPR/Cas9, нуклеаза MBBBD или мегануклеаза.
Инактивация генов лекарственной чувствительности
Согласно еще одному конкретному варианту реализации указанная устойчивость к лекарственным препаратам может быть придана Т-клетке путем инактивации генов лекарственной чувствительности. Впервые автор настоящего изобретения попытался инактивировать потенциальный ген лекарственной чувствительности с целью конструирования Т-клетки для иммунотерапии.
Предполагается, что при инактивации рассматриваемого гена, он не будет экспрессироваться с образованием функционального белка. Согласно конкретному варианту реализации генетическая модификация, лежащая в основе способа, основана на экспрессии, в предложенных для конструирования клетках, одной редкощепящей эндонуклеазы, такой как указанная редкощепящая эндонуклеаза, специфически катализирующая расщепление в одном целевом гене и, как следствие, инактивирующая указанный целевой ген. Согласно конкретному варианту реализации этап инактивации по меньшей мере одного гена лекарственной чувствительности включает введение в клетку редкощепящей эндонуклеаз, которая способна расщепить по меньшей мере один ген лекарственной чувствительности. Согласно более конкретному варианту реализации указанные клетки трансформируют нуклеиновой кислотой, кодирующей редкощепящую эндонуклеазу, которая способна расщепить ген лекарственной чувствительности, и указная редкощепящая эндонуклеаза экспрессируется в указанных клетках. Указанной редкощепящей эндонуклеазой может быть мегануклеаза, нуклеаза с цинковыми пальцами, нуклеаза CRISPR/Cas9, MBBBD-нуклеаза или TALE-нуклеаза.
Согласно предпочтительному варианту реализации геном лекарственной чувствительности, который можно инактивировать для придания устойчивости к лекарственному препарату Т-клетке, является ген дезоксицитидинкиназы (dCK) человека. Данный фермент нужен для фосфорилирования дезоксирибонуклезидов дезоксицитидина (dC), дезоксигуанозина (dG) и дезоксиаденозина (dA). Аналоги пуриновых нуклеотидов (PNA) метаболизируются при участии dCK до моно-, ди- и трифосфат-PNA. Их трифосфатные формы и в частности клофарабин трифосфат конкурируют с АТФ для синтеза ДНК, действуют как проапоптотический агент и являются мощными ингибиторами рибонуклеотидредуктазы (RNR), которая участвует в выработке тринуклеотидов (cf-предполагаемый механизм действия на Фигуре 1).
Предпочтительно инактивация dCK в Т-клетках опосредуется TALE-нуклеазой. Для выполнения данной задачи были разработаны несколько пар TALE-нуклеаз dCK, собирающиеся на уровне полинуклеотида и проверенные путем секвенирования. Примеры пар TALE-нуклеаз, которые можно применять согласно настоящему исследованию, отображены в последовательностях SEQ ID №63 и SEQ ID №64. Когда применяется пара данных TALE-нуклеаз, целевая последовательность dCK соответствует SEQ ID №62.
Как показано в примерах, такая инактивация dCK в Т-клетках придает устойчивость к аналогам пуриновых нуклеозидов (PNA), таким как клофарабин и флударабин.
Согласно другому предпочтительному варианту реализации инактивацию dCK в Т-клетках сочетают с инактивацией генов TRAC, и получают Т-клетки с нокаутом двух указанных генов устойчивые к такому препарату, как клофарабин, и одновременно аллогенные. Такие двойные характеристики особенно полезны для терапевтических целей и позволяют применять «готовые» аллогенные клетки для иммунотерапии в сочетании с химиотерапией для лечения пациентов с раком. Такую двойную инактивацию путем осуществления нокаута генов dCK/TRAC можно проводить одновременно или последовательно. Одним примером пары TALE-нуклеазы dCK/TRAC, которая приводит к успешному результату согласно настоящему изобретению, является применение SEQ ID №63 и SEQ ID №64, а также SEQ ID №66 и №67, соответственно. Целевые последовательности в 2 локусах (dCK и TRAC) отображены в SEQ ID №62 и SEQ ID №65, соответственно.
Еще одним примером фермента, который можно инактивировать, является ген гипоксантин-гуанинфосфорибозилтрансферазы (ГГФТ) человека (Genbank: М26434.1). В частности ГГФТ можно инактивировать при конструировании Т-клеток для придания им устойчивости к цитотоксическому метаболиту 6-тиогуанину (6TG), который превращается при участии ГГФТ в цитотоксический тиогуанин нуклеотид, и который в настоящее время применяют для лечения пациентов с раком, в частности лейкемиями (Hacke, Treger et al. 2013). Аналоги гуанинов метаболизируются ферментом ГГФТ, которая катализирует присоединение фосфорибозильной группы и позволяет образоваться тиоГМФ (Фигура 2). Аналоги гуанина, включая 6-меркаптопурин (6МР) и 6-тиогуанин (6TG) обычно применяют в качестве противолимфомных препаратов для лечения ОЛЛ (острый лимфобластный лейкоз). Они метаболизируются ГФТ (гипоксантинфосфорибозилтрансферазой), которая катализирует присоединение фосфорибозильной группы и позволяет образоваться тиоГМФ. Их последующее фосфорилирование приводит к образованию их фосфорилированных форм, которые в итоге встраиваются в ДНК. Будучи включенными в ДНК тио-ГТФ, нарушает точность воспроизведения репликации ДНК за счет своих тиоловых групп и генерирует случайную точечную мутацию, которая в значительной степени вредна для целостности клетки.
Согласно еще одному варианту реализации инактивация CD3, в норме экспрессирующегося на поверхности Т-клетки, может придавать устойчивость к анти-CD3-антителам, таким как реплизумаб.
Устойчивые к лекарственным препаратам клетки Дауди CD19+/luc+ для оценки цитотоксичности устойчивых к лекарственным препаратам аллогенных Т-клеток с CAR
Также настоящее изобретение включает способ получения клеток-мишеней, которые экспрессируют и поверхностный рецептор, специфичный для Т-клеток с CAR, и ген устойчивости. Указанные клетки мишени особенно полезны при оценке цитотоксичности T-клеток с CAR. Указанные клетки обладают высокой устойчивостью к клинически значимой дозе клофарабина и обладают люциферазной активностью. Такое сочетание свойств позволяет прослеживать их in vivo в модельных мышах. Более конкретно, их можно применять для оценки цитотоксических свойств для устойчивых к лекарственным препаратам Т-клеток у мышей в присутствии клофарабина или других PNA. Устойчивые к клофарабину клетки Дауди моделируют физиологическое состояние пациентов с острым лимфобластным лейкозом (ОЛЛ), у которых возникают рецидивы при индукции терапии, у которых отмечаются опухоли из В-клеток, устойчивых к лекарственным препаратам. Таким образом, указанные клетки представляют большой интерес для оценки надежности и цитотоксичности устойчивых к лекарственным препаратам Т-клеток с CAR. Предпочтительно, указанные клетки-мишени являются клетками Дауди CD19+ люцифераза+.
Изолированные (выделенные) клетки
Настоящее изобретение также относится к изолированных клеткам, которые можно получить при помощи способа, описанного выше. В частности, настоящее изобретение относится к изолированной Т-клетке, устойчивой к лекарственным препаратам, которая содержит по меньшей мере один нарушенный ген, кодирующий компонент рецептора Т-клеток. Согласно конкретному варианту реализации указанная T-клетка экспрессирует по меньшей мере один ген лекарственной устойчивости, предпочтительно ген ble или ген mcrA, или ген, кодирующий мутантную DHFR, мутантную IMPDH2, мутантную AGT или мутантный кальциневрин. Согласно еще одному конкретному варианту реализации указанная T-клетка содержит по меньшей мере один нарушенный ген лекарственной чувствительности, такой как ген dCK или ГГФТ. Согласно более конкретному варианту реализации указанная изолированная T-клетка содержит нарушенный ген ГГФТ и экспрессирует мутантную DHFR; указанная изолированная Т-клетка содержит нарушенный ген ГГФТ и экспрессирует мутантную IMPDH2; указанная изолированная T-клетка содержит нарушенный ген ГГФТ и экспрессирует мутантный кальциневрин; указанная
изолированная Т-клетка содержит нарушенный ген ГГФТ и экспрессирует мутантную AGT. Согласно предпочтительному варианту реализации указанная изолированная клетка экспрессирует химерный рецептор антигена (CAR), который может быть CD19 или CD123.
Аллогенная Т-клетка, устойчивая к лекарственному препарату
В частности, настоящее изобретение относится к аллогенной Т-клетке, устойчивой к лекарственному препарату, особенно подходящей для иммунотерапии. Устойчивость лекарственного препарата может быть придана путем инактивации генов лекарственной чувствительности или путем экспрессии генов лекарственной устойчивости, которые были описаны выше. Некоторые примеры лекарственных препаратов, которые подходят для реализации настоящего изобретения, включают аналоги пуриновых нуклеодидов (PNA), такие как клофарабин или флударабин, или другие лекарственные препараты, такие как 6- меркаптопурин (6МР) и 6-тиогуанин (6TG).
Под клеткой согласно настоящему изобретению понимают клетку гемопоэтического происхождения, функционально вовлеченную в инициацию и/или приведение в действие врожденного и/или приобретенного иммунного ответа. Клетка согласно настоящему изобретению предпочтительно является Т-лимфоцитом, полученным от донора. Указанный Т- лимфоцит согласно настоящему изобретению может быть получен из стволовой клетки. Указанные стволовые клетки могут быть стволовыми клетками взрослого организма, эмбриональными стволовыми клетками, более конкретно стволовыми клетками не человека, стволовыми клетками из пуповинной крови, клетками-предшественниками, стволовыми клетками костного мозга, тотипотентными стволовыми клетками или тема поэтическими стволовыми клетками. Репрезентативные стволовые клетки человека включают CD34+-клетки. Указанная изолированная клетка может также быть дендритной клеткой, дендритной клеткой- киллером, тучной клеткой, клеткой NK (естественным киллером), В-лимфоцитом или Т-клеток, выбранным из группы, состоящей из воспалительных Т-клеток, цитотоксических Т-клеток, регуляторных Т-клеток или Т-клеток-хелперов. Согласно еще одному варианту реализации указанная клетка может быть получена из группы, состоящей из CD4+ Т-клеток и CD8+ Т-клеток. Перед размножением и генетической модификацией клеток согласно настоящему изобретению источник клеток можно получить от субъекта при помощи разных неограничивающих способов. Клетки можно получить из целого ряда источников, включая без ограничений мононуклеарные клетки периферической крови, костный мозг, ткань лимфатических узлов, пуповинная кровь, ткань тимуса, ткань из очага инфекции, асцит, плевральный выпот, ткань селезенки и опухоли. Согласно определенным вариантам реализации настоящего изобретения можно применять любое количество линий Т-клеток, доступных и известных специалистам в данной области техники. Согласно еще одному варианту реализации указанную клетку предпочтительно получают от здорового донора. Согласно еще одному варианту реализации указанная клетка является частью смешанной популяции клеток, которые проявляют разные фенотипические характеристики.
Множественная лекарственная устойчивость
Согласно еще одному конкретному варианту реализации авторы настоящего изобретения пытались разработать «стандартную» стратегию иммунотерапии, с применением аллогенных Т-клеток, устойчивых к нескольким лекарственным препаратам для облегчения селекции сконструированных Т-клеток, когда пациент получает разные лекарственные препараты. Лечебную эффективность можно существенно повысить путем генетического конструирования аллогенных Т-клеток с множественной лекарственной устойчивостью. Такая стратегия может быть особенно эффективна при лечении опухолей, которые отвечают на сочетания препаратов, оказывающих синергичное действие. Кроме того, сконструированные Т-клетки с множественной лекарственной устойчивостью могут размножаться и проходить селекцию при минимальной дозе лекарственных препаратов.
Таким образом, способ согласно настоящему изобретению может включать осуществление модификации Т-клетки для придания множественной лекарственной устойчивости указанной Т-клетке. Указанная множественная лекарственная устойчивость может быть придана либо за счет экспрессии более чем одного гена лекарственной устойчивости, либо за счет инактивации более чем одного гена лекарственной чувствительности. Согласно еще одному конкретному варианту реализации множественная лекарственная устойчивость может быть придана указанной Т-клетке за счет экспрессии по меньшей мере одного гена лекарственной устойчивости и за счет инактивации по меньшей мере одного гена лекарственной чувствительности. В частности, множественная лекарственная устойчивость может быть придана указанной Т-клетке за счет экспрессии по меньшей мере одного гена лекарственной устойчивости, такого как мутантная форма ДГФР, мутантная форма IMPDH2, мутантная форма кальциневрина, мутантная форма MGMT, гена ble и гена mcrA, и за счет инактивации по меньшей мере одного гена лекарственной чувствительности, такого как ген ГГФР. Согласно предпочтительному варианту реализации множественная лекарственная устойчивость может быть придана за счет инактивации гена ГГФТ и за счет экспрессии мутантной формы ДГФР; или за счет инактивации гена ГГФТ и за счет экспрессии мутантной формы IMPDH2; или за счет инактивации гена ГГФТ и за счет экспрессии мутантной формы кальциневрина; за счет инактивации гена ГГФТ и за счет экспрессии мутантной формы MGMT; за счет инактивации гена ГГФТ и за счет экспрессии гена ble; за счет инактивации гена ГГФТ и за счет экспрессии гена mcrA.
Способ конструирования аллогенных Т-клеток, устойчивых к лекарственным препаратам:
Для улучшения противораковой терапии и селективного приживления аллогенных Т-клеток указанным клеткам придают устойчивость к лекарственным препаратам, чтобы защитить их от токсических побочных эффектов химиотерапевтического агента. Устойчивые к лекарственным препаратам Т-клетки также позволяют обогащать их пул in vivo или ex vivo, поскольку Т-клетки, которые экспрессируют ген лекарственной устойчивости, будут выживать и воспроизводиться по сравнению с клетками, чувствительными к лекарственному препарату. В частности, настоящее изобретение относится к конструированию аллогенных и устойчивых к лекарственному препарату Т-клеток для иммунотерапии, включающий:
(а) Обеспечение Т-клеток;
(b) Осуществление селекции по меньшей мере с одним лекарственным препаратом;
(c) Осуществление модификации указанных Т-клеток путем инактивации по меньшей мере одного гена, кодирующего компонент рецептора Т-клеток (РТЛ);
(d) Осуществление модификации Т-клеток для придания им устойчивости к лекарственному препарату;
(e) Обеспечение размножения указанных сконструированных Т-клеток в присутствии указанного лекарственного препарата.
- Аллогенные Т-клетки
Настоящее изобретение относится к аллогенной иммунотерапии. Приживление аллогенных Т-клеток возможно при инактивации по меньшей мере одного гена, кодирующего компонент РТЛ. РТЛ делают нефункциональным в клетках путем инактивации гена РТЛ-альфа и/или гена РТЛ-бета. Инактивация РТЛ позволяет исключить ТПХ. Предполагается, что при инактивации гена рассматриваемый ген не экспрессируется с образованием функциональной формы белка. Согласно конкретным вариантам реализации генетическая модификация согласно данному способу основана на экспрессии, в полученных для конструирования клетках, одной редкощепящей эндонуклеазы, так что указанная редкощепящая эндонуклеаза специфически катализирует расщепление в одном целевом гене, тем самым инактивируя указанный целевой ген. Разрывы в нуклеотидной цепи, вызванные редкощепящей эндонуклеазой, обычно восстанавливаются по разным механизмам гомологичной рекомбинации или соединения негомологичных концов (NHEJ). Однако NHEJ является несовершенным процессом репарации, который часто приводит к изменениям в последовательности ДНК в сайте расщепления. Механизмы подразумевают повторное соединение того, что осталось на двух концах ДНК, посредством прямого повторного лигирования или посредством так называемого соединения концов, опосредованного микрогомологией (Betts, Brenchley et al. 2003; Ma, Kim et al. 2003). Репарация посредством соединения негомологичных концов (NHEJ) часто приводит к маленьким вставкам или делециям и может применяться для получения направленного нокаута генов. Указанная модификация может быть заменой, делецией или добавлением по меньшей мере одного нуклеотида. Клетки, в которых произошло событие мутагенеза, т.е. событие мутагенеза с последующим событием NHEJ, может быть идентифицирована и пройти селекцию при помощи способа, хорошо известного в технике. Согласно конкретному варианту реализации этап инактивации по меньшей мере одного гена, кодирующего компонент рецептора Т-клеток (РТЛ) в клетке пробы каждого индивидуума, включает введение в указанную клетку редкощепящей эндонуклеазы, способной нарушить по меньшей мере один ген, кодирующий компонент рецептора Т-клеток (РТЛ). Согласно более конкретному варианту реализации указанные клетки пробы каждого индивидуума трансформируют нуклеиновой кислотой, кодирующей компонент рецептора Т-клеток (РТЛ), и указанная редкощепящая эндонуклеаза экспрессируется в указанных клетках.
Указанная редкощепящая эндонуклеаза может быть мегануклеазой, нуклеазой с цинковыми пальцами, нуклеазой CRISPR/Cas9, TALE-нуклеазой или MBBBD-нуклеазой. Согласно предпочтительному варианту реализации указанная редкощепящая эндонуклеаза является TALE-нуклеазой. Предполагается, что под TALE-нуклеазой понимают гибридный белок, состоящий из ДНК-связывающего домена, полученного из подобного активатору транскрипции эффектора (TALE), и одного нуклеазного каталитического домена для расщепления целевой нуклеотидной последовательности (Boch, Scholze et al. 2009; Moscou and Bogdanove 2009; Christian, Cermak et al. 2010; Cermak, Doyle et al. 2011; Geissler, Scholze et al. 2011; Huang, Xiao et al. 2011; Li, Huang et al. 2011; Mahfouz, Li et al. 2011; Miller, Tan et al. 2011; Morbitzer, Romer et al. 2011; Mussolino, Morbitzer et al. 2011; Sander, Cade et al. 2011; Tesson, Usal et al. 2011; Weber, Gruetzner et al. 2011; Zhang, Cong et al. 2011; Deng, Yan et al. 2012; Li, Piatek et al. 2012; Mahfouz, Li et al. 2012; Mak, Bradley et al. 2012). Согласно настоящему изобретению были разработаны новые TALE-нуклеазы точного направленного действия на соответствующие гены для стратегий адоптивной иммунотерапии.
Предпочтительными TALE-нуклеазами согласно настоящему изобретению являются нуклеазы, распознающие и расщепляющие целевую последовательность, выбранную из группы, состоящей из: SEQ ID №: 1-5 (РТЛ-альфа), SEQ ID №: 6 и 7 (РТЛ-бета). Указанные TALE-нуклеазы предпочтительно содержат полипептидную последовательность, выбранную из SEQ. ID №: 8 - SEQ ID №: 13. Согласно еще одному варианту реализации в клетку вместе с указанной редкощепящей эндонуклеазой можно вводить дополнительный каталитический домен, чтобы усилить мутагенез для увеличения его способности к инактивации целевых генов. В частности, указанным дополнительным каталитическим доменом является фермент процессинга концов ДНК. Примеры ферментов процессинга концов ДНК включают 5-3'-экзонуклеазы, 3-5' экзонуклеазы, 5-3' щелочные экзонуклеазы, 5'-флэп-эндонуклеазы, геликазы, хоспазы, гидролазы и независимые от матрицы ДНК-полимеразы. Неограничивающие примеры таких каталитических доменов включают белковый домен или каталитически активное производное белкового домена, выбранные из группы, состоящей из hExol (EXO1_HUMAN), Exol дрожжей (EXO1_YEAST), Exol E.coli, TREX2 человека, TREX1 мыши, TREX1 человека, TREX1 быка, TREX1 крысы, TdT (концевая дезоксинклеотидилтрансфераза) DNA2 человека, DNA2 дрожжей (DNA2_YEAST). Согласно предпочтительному варианту реализации указанный дополнительный каталитический домен обладает 3'-5'-эндонуклеазной активностью, и согласно более предпочтительному варианту реализации указанным дополнительным каталитическим доменом является TREX, более предпочтительно каталитический домен TREX2 (WO 2012/058458). Согласно еще одному предпочтительному варианту реализации указанный каталитический домен кодируется одноцепочечным полипептидом TREX2. Указанный дополнительный каталитический домен может быть встроен в гибридный белок или химерный белок нуклеазу согласно настоящему изобретению, необязательно при помощи белкового линкера.
Известно, что разрывы между нуклеотидами повышают процент гомологичной рекомбинации. Таким образом, согласно еще одному варианту реализации этап генетической модификации указанного способа также включает этап введения в клетки экзогенной нуклеиновой кислоты, содержащей последовательность, гомологичную по меньшей мере части целевой нуклеотидной последовательности, так чтобы между целевой нуклеотидной последовательностью и экзогенной нуклеиновой кислотой возникала гомологичная рекомбинация. Согласно конкретным вариантам реализации указанная экзогенная нуклеиновая кислота включает первую и вторую части, которые гомологичны области 5' и 3' целевой нуклеотидной последовательности, соответственно. Указанная экзогенная нуклеиновая кислота согласно данным вариантам реализации также содержит третью часть, расположенную между первой и второй частями, которая не обладает гомологией с областями 5' и 3' целевой нуклеотидной последовательности. После расщепления целевой нуклеотидной последовательности стимулируют событие гомологичной рекомбинации между целевой нуклеотидной последовательностью и экзогенной нуклеиновой кислотой. Предпочтительно в пределах указанной донорной матрицы применяют гомологичные последовательности длиной по меньшей мере 50 по, предпочтительно более 100 по и более предпочтительно более 200 по. Согласно конкретному варианту реализации гомологчиная последовательность может быть от 200 по до 6000 по, более предпочтительно от 1000 по до 2000 по. Фактически, разделяемая нуклеиновыми кислотами гомология расположена в областях, фланкирующих сайт расщепления выше и ниже в последовательности, и нуклеотидная последовательность, которую требуется ввести, должна быть расположена между двумя плечами.
Согласно конкретному варианту реализации указанная экзогенная нуклеиновая кислота может содержать трансген, кодирующий ген лекарственной устойчивости согласно настоящему изобретению.
Конструирование дополнительных возможных свойств Т-клеток
Иммунные клетки согласно настоящему изобретению можно также сконструировать так, чтобы они приобрели дополнительные свойства, которые способствовали бы их более специфическому или более эффективному применению в терапии.
- Химерные рецепторы антигенов (CAR)
Химерные рецепторы антигенов (CAR) способны перенаправлять специфичность и реактивность иммунных клеток и нацеливать их на избранную мишень, на основе свойств лиганд-связывающих доменов. Так, согласно еще одному варианту реализации указанный способ также включает этап введения в указанные лимфоциты химерного рецептора антигена. Указанный химерный рецептор антигена сочетает в себе связывающий домен для компонента, присутствующего на клетке-мишени, например, основанная на антителах специфичность в отношении желаемого антигена (например, антигена опухоли) с внутриклеточным доменом, активирующим рецептор Т-клеток, для генерирования химерного белка, который проявляет специфическую иммунную активность в отношении клетки-мишени. Как правило, CAR состоит из внеклеточного одноцеопчечного антитела (scFv), соединенного с доменом внеклеточной сигнализации дзета-цепи комплекса Т-клеточного рецептора для распознавания антигенов  и обладает способностью, когда экспрессируется в Т-клетках, перенаправлять распознавание, основанное на специфичности моноклонального антитела. Одним примером CAR, применяемого согласно настоящему изобретению, является CAR, направленный на антиген CD19, который может содержать в качестве неограничивающих примеров аминокислотную последовательность: SEQ ID №: 19 или 20.
и обладает способностью, когда экспрессируется в Т-клетках, перенаправлять распознавание, основанное на специфичности моноклонального антитела. Одним примером CAR, применяемого согласно настоящему изобретению, является CAR, направленный на антиген CD19, который может содержать в качестве неограничивающих примеров аминокислотную последовательность: SEQ ID №: 19 или 20.
- Инактивация генов иммунных контрольных точек
Опосредуемый Т-клетками иммунитет включает несколько последовательных этапов, включая селекцию клонов антиген-специфичных клеток, их активацию и пролиферацию во вторичной лимфоидной ткани, их направленную миграцию к очагам присутствия антигена и воспаления, исполнение прямой эффекторной функции и оказание помощи (через цитокины и мембранные лиганды) большому числу иммунных эффекторных клеток. Каждый из указанных этапов регулируется уравновешивающими друг друга стимулирующими и подавляющими сигналами, которые производят тонкую корректировку ответа. Специалисты в данной области техники должны понимать, что термин «иммунные контрольные точки» обозначает группу молекул, экспрессируемых Т-клетками. Указанные молекулы эффективно выполняют функцию «стопоров» для отрицательной модуляции или подавления иммунного ответа. Молекулы иммунных контрольных точек включают без ограничений молекулы программированной гибели 1 (PD-1, также называемые PDCD1 или CD279, номер доступа: NM_005018), антиген 4 цитотоксических Т-клеток (CTLA-4, также называемый CD152, GenBank номер доступа AF414120.1), LAG3 (также называемый CD223, номер доступа: NM_002286.5), Tim3 (также называемый HAVCR2, номер доступа в GenBank: JX049979.1), BTLA (также называемый CD272, номер доступа: NM_181780.3), BY55 (также называемый CD160, номер доступа в GenBank: CR541888.1), TIG IT (также называемый VSTM3, номер доступа: NM_173799), LAIR1 (также называемый CD305, номер доступа в GenBank: CR542051.1, (Meyaard, Adema et al. 1997)), SIGLEC10 (номер доступа в GeneBank: AY358337.1), 2B4 (также называемый CD244, номер доступа: NM_001166664.1), РРР2СА, РРР2СВ, PTPN6, PTPN22, CD96, CRTAM, SIGLEC7 (Nicoll, Ni et al. 1999), SIGLEC9 (Zhang, Nicoll et al. 2000; Ikehara, Ikehara et al. 2004), TNFRSF10B, TNFRSF10A, CASP8, CASP10, CASP3, CASP6, CASP7, FADD, FAS, TGFBRII, TGFRBRI, SMAD2, SMAD3, SMAD4, SMAD10, SKI, SKIL, TGIF1, IL10RA, IL10RB, HMOX2, IL6R, IL6ST, EIF2AK4, CSK, PAG1, SIT1, FOXP3, PRDM1, BATF (Quigley, Pereyra et al. 2010), GUCY1A2, GUCY1A3, GUCY1B2, GUCY1B3, которые напрямую подавляют иммунный ответ. Например, CTLA-4 является поверхностным антигеном клетки, экспрессируемым на определенных CD4 и С08-Т-клетках; когда он соединяется со своим лигандом (В7-1 или В7-2) на антиген-представляющих клетках, активация Т-клеток и их эффекторная функция подавляется. Таким образом, настоящее изобретение относится к способу конструирования аллогенной Т-клетки, устойчивой к лекарственному препарату, также включающему модифицирование Т-клеток путем инактивации по меньшей мере одного белка, вовлеченного в иммунную контрольную точку, в частности PD1 и/или CTLA-4. Согласно предпочтительному варианту реализации, стадию осуществления инактивации по меньшей мере одного белка, вовлеченного в иммунную контрольную точку, реализуют путем экспрессии редкощепящей эндонуклеазы, которая может специфично расщеплять целевую последовательность в гене иммунной контрольной точки. Согласно предпочтительному варианту реализации указанной редкощепящей эндонуклеазой является TALE-эндонуклеаза. Например, указанная TALE-нуклеаза может специфически расщеплять целевую последовательность, выбираемую из группы, состоящей из: SEQ ID №: 21-23 (CTLA-4) и SEQ ID №: 24 и SEQ ID №: 25 (PDCD1), и согласно более предпочтительному варианту реализации указанная TALE-нуклеаза содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID №: 26 - SEQ ID №: 35.
- Т-клетки, устойчивые к иммуносупрессии
Аллогенные клетки быстро отбраковываются иммунной системой хозяина. Было продемонстрировано, что аллогенные лейкоциты в необлученных препаратах крови могут сохраняться не более 5-6 дней (Boni, Muranski et al. 2008). Таким образом, чтобы предупредить отторжение аллогенных клеток, иммунную систему хозяина следует как правило подавлять до определенной степени. Однако в случае адоптивной иммунотерапии применении иммуносупрессорных препаратов также оказывает вредное действие на введенные терапевтические Т-клетки. Следовательно, для эффективного применения подхода адоптивной иммунотерапии в указанных условиях введенные клетки должны быть также устойчивы к иммуносупрессорному лечению. Таким образом, согласно конкретному варианту реализации, способ согласно настоящему изобретению дополнительно включает этап модификации Т-клеток для придания им устойчивости к иммуносупрессорному агенту, предпочтительно путем инактивации по меньшей мере одного гена, кодирующего мишень для иммуносупрессорного агента. Под иммуносупрессорным агентом понимают агент, который подавляет иммунную функцию по одному из нескольких механизмов действия. Иными словами иммуносупрессорный агент выполняет роль соединения, которое проявляет способность уменьшать степень иммунного ответа. Способ согласно настоящему изобретению позволяет придать Т-клеткам для иммунотерапии устойчивость к иммуносупрессору путем инактивации мишени для указанного иммуносупрессорного агента в Т-клетках. В качестве неограничивающих примеров мишенями для иммуносупрессорного агента может быть рецептор для иммуносупрессорного агента, такой как: CD52, рецептор глюкокортикоида (GR), член семейства генов FKBP и член семейства генов циклофиллина. Согласно конкретному варианту реализации генетическая модификация согласно настоящему способу основана на экспрессии, в клетках, предложенных для конструирования, одной редкощепящей эндонуклеазы, такой как указанная редкощепящая эндонуклеаза, специфически катализирующая расщепление в одном целевом гене, таки образом инактивируя указанный целевой ген. Указанной редкощепящей эндонуклеазой может быть мегануклеаза, нуклеаза с цинковыми пальцами или TALE-эндонуклеаза. Предпочтительными TALE-нуклеазами согласно настоящему изобретению являются эндонуклеазы, распознающие и расщепляющие целевую последовательность, выбранную из группы, состоящей из: SEQ ID №: 36-41 (GR), и SEQ ID №: 54-59 (CD52). Указанные TALE-нуклеазы предпочтительно включают полипептидную последовательность, выбранную из группы, состоящей из SEQ ID №: 42 - SEQ ID №: 53 и SEQ ID №:60-SEQ ID №:61.
- Гены самоубийства
Согласно еще одному аспекту, поскольку сконструированные Т-клетки размножаются и сохраняются в течение нескольких лет после введения, желательно включить механизм безопасности, который бы позволил селективно удалять введенные Т-клетки. Так, согласно некоторым вариантам реализации способ согласно настоящему изобретению включает трансформацию указанных Т-клеток рекомбинантным геном самоубийства. Указанный рекомбинантный ген самоубийства применяют для снижения риска прямой токсичности и/или неконтролируемой пролиферации указанных Т-клеток, когда они уже введены субъекту (Quintarelli С, Vera F, blood 2007; Теу SK, Dotti G., Rooney CM, boil blood marrow transplant 2007). Гены самоубийства позволяют селективно удалять трансформированные клетки in vivo. В частности, ген самоубийства обладает способностью превращаться из нетоксичного пролекарства в цитотоксический препарат или экспрессироваться с образованием токсичного продукта. Иными словами, «ген самоубийства» представляет собой нуклеиновую кислоту, кодирующую продукт, причем указанный продукт вызывает гибель клетки сам по себе или в присутствии другого соединения. Типичным примером такого гена самоубийства является ген, кодирующий киназу вируса простого герпеса. Дополнительные примеры включают тимидинкиназу вируса ветряной оспы и бактериальный ген цитозиндезаминазы, который может превращать 5-фторцитозин в высокотоксичное соединение 5-фторурацил. Гены самоубийства также включают в качестве неограничивающих примеров каспазу-9 или каспазу-8, или цитозиндезаминазу. Каспаза-9 может быть активирована при помощи специфического химического индуктора димеризации (CID). Также генами самоубийства могут быть полипептиды, которые экспрессируются на поверхности клетки и могут придавать клетке чувствительность к терапевтическим моноклональным антителам. В настоящей заявке термин «пролекарство» относится к любому соединению, полезному для реализации способов согласно настоящему изобретению, которое может превращаться в токсичный продукт. Пролекарство превращается в токсичный продукт под действием продукта гена самоубийства при реализации способа согласно настоящему изобретению. Типичным примером такого пролекарства является ганцикловир, который превращается in vivo в токсичное соединение под действием HSV-тимидинкиназы. Следовательно, производное ганцикловира токсично для клеток опухоли. Другие типичные примеры пролекарств включают ацикловир, FIAU [1-(2- дезокси-2-фтор-β-D-арабинофуранозил)-5-йодурацил], 6-метоксипурин-арабинозид для VZV-TK и 5-фторцитозин для цитозиндезаминазы.
- Способы доставки
Разные способы, описанные выше, подразумевают экспрессию рассматриваемого белка, такого как ген лекарственной устойчивости, редкощепящей эндонуклеазы, химерного рецептора антигена (CAR), гена самоубийства, в клетке. В качестве неограничивающего примера указанный рассматриваемый белок может экспрессироваться в клетке посредством его введения в качестве трансгена, предпочтительно кодируемого по меньшей мере одним плазмидным вектором. Полипептиды могут экспрессироваться в клетке как следствие введения полинуклеотидов, кодирующих указанные полипептиды, в клетку. В качестве альтернативы указанные полипептиды можно получить за пределами клетки, а затем ввести их в нее. Способы введения полинуклеотидного конструкта в клетки известны в технике и включают в качестве неограничивающих примеров способы стабильной трансформации, при которых полинуклеотидный конструкт интегрируется в геном указанной клетки, способы временной трансформации, при которых полинуклеотидный конструкт не интегрируется в геном указанной клетки, и способы, опосредуемые вирусами. Указанные полинуклеотиды можно вводить в клетку, например, при помощи рекомбинантных вирусных векторов (например, ретровирусов, аденовирусов), липосом и т.п. Например, способы временной трансформации включают, например, микроинъекцию, электропорацию или бомбардировку частицами. Указанные полинуклеотиды можно включить в векторы, более конкретно в плазмиды или вирус, чтобы они экспрессировались в клетках. Указанный плазмидный вектор может содержать маркер селекции, который позволяет проводить идентификацию и/или селекцию клеток, которые получили указанный вектор. В один вектор можно включать разные трансгены. Указанный вектор может содержать нуклеотидную последовательность, кодирующую последовательность скачков рибосом, такую как последовательность, кодирующую пептид 2А. Пептиды 2А, которые были обнаружены у подгруппы пикорнавирусов - автовирусов, вызывают «скачок» рибосомы с одного кодона на следующий без образования пептидной связи между двумя аминокислотами, кодируемыми указанными кодонами (см. Donnelly et al., J. of General Virology 82: 1013-1025 (2001); Donnelly et al., J. of Gen. Virology 78: 13-21 (1997); Doronina et al., Mol. And. Cell. Biology 28(13): 4227-4239 (2008); Atkins et al., RNA 13: 803-810 (2007)). Под «кодоном» понимают три нуклеотида в мРНК (или в смысловой нити молекулы ДНК), которые транслируются рибосомой в остаток аминокислоты. Таким образом, когда полипептиды разделяются олигопептидной последовательностью 2А, которые находятся в рамке, могут синтезироваться два полипептида с одной, смежной открытой рамки считывания в пределах мРНК. Такие механизмы скачков рибосом хорошо известны в технике, и известно, что они применяются при помощи нескольких векторов для экспрессии нескольких белков, кодируемых одной матричной РНК.
Согласно более конкретному варианту реализации полинуклеотиды, кодирующие полипептиды согласно настоящему изобретению, могут быть мРНК, которая вводится прямо в клетки, например, путем электропорации. Авторы настоящего изобретения определили оптимальные условия для электропорации мРНК в Т-клетку. Автор настоящего изобретения применял технологию cytoPulse, которая позволяет путем применения импульсных электрических полей временно пермеабилизировать живые клетки для доставки материала в клетки. Указанная технология, основанная на применении форм импульсов электропорации PulseAgile (ВТХ Havard Apparatus, 84 October Hill Road, Holliston, MA 01746, USA), позволяет осуществлять точный контроль длительности, интенсивности импульса, а также интервалов между импульсами (патент США 6010613 и международная заявка РСТ WO 2004083379). Все указанные параметры можно модифицировать, чтобы достигать наилучших условий для высокой эффективности трансфекции при минимальной смертности. По существу, первые высокие импульсы электрического поля позволяют образовать пору, а последующие более низкие импульсы электрического поля позволяют переместить полинуклеотид в клетку.
- Активация и размножение Т-клеток
Или до, или после генетической модификации Т-клетки можно активировать и размножить как правило при помощи способов, описанных, например, в патентах США 6352694; 6534055; 6905680; 6692964; 5858358; 6887466; 6905681; 7144575; 7067318; 7172869; 7232566; 7175843; 5883223; 6905874; 6797514; 6867041; и публикациях заявок на патент США №20060121005. Можно обеспечить увеличение числа (размножение) Т-клеток in vitro или in vivo. Как правило, Т-клетки согласно настоящему изобретению размножают путем осуществления контакта с агентом, который стимулирует комплекс CD3-PT/1, и костимулирующей молекулой на поверхности Т-клеток. Например, химические вещества, такие как кальций-ионофор А23187, форбол 12-миристат 13-ацетат (РМА), или митогенетические лектины, такие как фитогемагглютинин (ФГА) можно применять для создания активирующего сигнала для Т-клетки. В качестве неограничивающего примера популяции Т-клеток можно стимулировать in vitro, например путем контакта с анти-CD3-антителом, или его антиген-связывающим фрагментом, или с анти-CD2-антителом, иммобилизованным на поверхности, или путем контакта с активатором протеинкиназы С (например, бриостатином) в сочетании с кальций- ионофором. Для совместной стимуляции вспомогательной молекулы на поверхности Т-клеток применяют лиганд, который связывает вспомогательную молекулу. Например, популяция Т-клеток может контактировать с анти-CD3-антителом и анти-CD28-антителом, при условиях, подходящих для стимуляции пролиферации Т-клеток. Для стимулирования пролиферации либо CD4+ Т-клеток, либо CD8+ Т-клеток, применяют анти-CD3-антитело и анти-CD28-антитело. Например, агенты, обеспечивающие каждый сигнал, могут быть в растворе или связаны с поверхностью. Как могут легко понять специалисты в данной области техники, отношение количества частиц к количеству клеток может зависеть от размера частиц относительно клетки- мишени.
Условия, подходящие для культивирования Т-клеток, включают соответствующую среду (например, минимальную поддерживающую среду или среду RPMI 1640 или X-vivo 5, (Lonza)), которая может содержать факторы, необходимые для пролиферации и жизнеспособности, включая сыворотку (например, фетальную коровью сыворотку или сыворотку человека), интерлейкин-2 (IL-2), IFN-g, 1L-4, 1L-7, GM-CSF, -10, - 2, 1L-15, TGFp, IL-21 и ФНО или любые другие добавки для роста клеток, известные специалистам в данной области техники. Другие добавки для роста клеток включают, без ограничений, поверхностно-активное вещество, плазманат и восстановители, такие как N-ацетилцистеин и 2-меркаптоэтанол. Среда может включать RPMI 1640, A1M-V, DMEM, MEM, а-МЕМ, F-12, X-Vivo 1 и X-Vivo 20, Optimizer, с добавлением аминокислот, пирувата натрия и витаминов, либо без сыворотки, либо с добавлением соответствующего количества сыворотки (или плазмы), или заданного набора гормонов, и/или количества цитокина(ов), достаточного для роста и размножения Т-клеток. Антибиотики, например, пенициллин и стрептомицин включают только в экспериментальные культуры, но не в культуры клеток, которые будут вводиться путем инфузии субъекту. Целевые клетки хранят при условиях, необходимых для поддержания роста, например, при соответствующей температуре (например, 37°C) и атмосфере (например, воздух плюс 5% СO2). Т-клетки, которые подверглись стимуляции в течение разного времени, могут проявлять разные свойства.
Терапевтические приложения
Согласно еще одному варианту реализации указанные изолированные Т-клетки, полученные, как было описано выше, можно применять для аллогенной адоптивной клеточной иммунотерапии. В частности, указанные Т-клетки согласно настоящему изобретению можно применять для лечения рака, инфекций или аутоиммунного заболевания у пациентов, нуждающихся в таком лечении. Согласно еще одному аспекту настоящее изобретение основано на способах для лечения пациентов, нуждающихся в таком лечении, причем указанный способ включает по меньшей мере один из следующих этапов:
(a) Обеспечение изолированной Т-клетки, которую можно получить любым из способов, описанных ранее;
(b) Введение указанных клеток пациенту.
Согласно одному варианту реализации указанные Т-клетки согласно настоящему изобретению могут подвергнуться надежному размножению in vivo, и могут сохраняться в течение длительного периода времени.
Указанное лечение может быть облегчающим, терапевтическим или профилактическим. Настоящее изобретение особенно подходит для аллогенной иммунотерапии, в той мере, в которой оно позволяет проводить трансформацию Т-клеток, обычно получаемых от доноров, с получением не-аллореактивных клеток. Это можно сделать в соответствии со стандартными протоколами и воспроизвести столько раз, сколько потребуется. Образующиеся в результате Т- клетки вводят одному или нескольким пациентам, и они становятся доступны как «готовый» терапевтический препарат.
Клетки, которые можно применять при реализации раскрываемых способов, описаны в предыдущем разделе. Указанное средство можно применять при лечении пациентов, у которых диагностирован рак, вирусная инфекция, аутоиммунные расстройства. Рак, который можно лечить, включает опухоли, которые не васкуляризованы, или еще существенно не васкуляризованы, а также васкуляризованные опухоли. Типы рака могут включать несолидные опухоли (такие как гематологические опухоли, например, лейкемии и лимфомы) или могут включать солидные опухоли. Типы рака, которые можно лечить аллогенными Т-клетками, устойчивыми к лекарственным препаратам, согласно настоящему изобретению, включают без ограничений карциному, бластому и саркому, и определенные лейкемии и лимфоидные злокачественные новообразования, доброкачественные и злокачественные опухоли, а также онкологические заболевания, например, саркомы, карциномы и меланомы. Также включены опухоли/типы рака взрослых и опухоли/типы рака детей. Согласно одному варианту реализации настоящего изобретения обычно аллогенными устойчивыми к лекарственным препаратам Т-клетками согласно настоящему изобретению лечат детский острый лимфобластный лейкоз (ОЛЛ) и амиотрофический миелоидный лейкоз (АМЛ). Этого можно достичь при применении устойчивых к лекарственным препаратам CD19+ Т-клеток с CAR с нокаутом гена TRAC и устойчивых к лекарственным препаратам CD123+ Т-клеток с CAR с нокаутом гена TRAC.
Это может быть лечение в сочетании с одним или более лечебными средствами против рака, выбранными из группы средств на основе антител, химиотерапии, средств на основе цитокинов, терапии дендритными клетками, генной терапии, гормонотерапии, терапии лазерным излучением и лучевой терапии.
Согласно предпочтительному варианту реализации настоящего изобретения указанное средство вводят пациентам, подвергающимся иммуносупрессорной терапии. Настоящее изобретение предпочтительно предполагает применение клеток или популяций клеток, которым была придана устойчивость по меньшей мере к одному лекарственному препарату согласно настоящему изобретению либо за счет экспрессии гена лекарственной устойчивости, либо за счет инактивации гена лекарственной чувствительности. Согласно данному аспекту медикаментозная терапия должна способствовать селекции и экспрессии Т-клеток согласно настоящему изобретению в организме пациента.
Введение клеток или популяций клеток согласно настоящему изобретению можно проводить любым традиционным способом, включая ингаляции аэрозолей, инъекцию, употребление внутрь, трансфузию, имплантацию или трансплантацию. Композиции, описываемые в настоящей заявке, можно вводить пациенту подкожно, внутрикожно, в опухоль, в лимфатические узлы, интрамедулярно, внутримышечно, внутричерепно, путем внутривенной или внутрилимфатической инъекции, или внутрибрюшинно. Согласно одному варианту реализации композиции клеток согласно настоящему изобретению предпочтительно вводят путем внутривенной инъекции.
Введение клеток или популяций клеток может состоять во введении 103-1010 клеток на кг массы тела, предпочтительно 105-106 клеток/кг массы тела, включая все целые значения количеств клеток в пределах данного диапазона. Клетки или популяции клеток можно вводить в одной или нескольких дозах. Согласно еще одному варианту реализации указанное эффективное количество клеток вводят в одной дозе. Согласно еще одному варианту реализации указанное эффективное количество клеток вводят в более чем одной дозе в течение некоторого периода времени. Время введения остается на усмотрение лечащего врача и зависит от клинического состояния пациента. Клетки или популяции клеток можно получить из любого источника, такого как банк крови или донор. Хотя индивидуальные потребности варьируют, определение оптимальных диапазонов эффективного количества клеток конкретного типа для конкретного заболевания или патологического состояния лежит в пределах компетенции специалистов в данной области техники. Под эффективным количеством понимают количество, которое обеспечивает терапевтическую или профилактическую пользу. Вводимая доза будет зависеть от возраста, состояния здоровья и массы получателя, типа сопутствующего заболевания, если таковое есть, частоты лечения и природы желаемого действия.
Согласно еще одному варианту реализации указанное эффективное количество клеток или фармацевтической композиции, содержащей указанные клетки, вводят парентерально. Указанное введение может быть внутривенным введением. Указанное введение может быть проведено напрямую путем инъекции в опухоль. Согласно определенным вариантам реализации настоящего изобретения клетки вводят пациенту в сочетании (например, до, одновременно или после) с любым числом соответствующих лечебных модальностей, включая без ограничений лечение агентами, такими как противовирусная терапия, цидофовир и интерлейкин-2, цитарабин (также известный под названием ARA-C) или натализиимаб для пациентов с множественным склерозом или эфализимаб для пациентов с псориазом или другие препараты для пациентов с промиелоцитарной лейкемией. Согласно дополнительному варианту реализации Т-клетки согласно настоящему изобретению могут применяться в сочетании с химиотерапией, облучением, иммуносупресосрами, такими как циклоспорин, азатиоприн, метотрексат, микофенолят и FK506, антителами или другими иммунодеструктивными агентами, такими как САМРАТН, анти-CD3 антитела или другие средства на основе антител, цитоксин, флударабин, циклоспорин, FK506, рапамицин, микофеноловая кислота, стероиды, FR901228, цитокинами и облучением. Указанные лекарственные средства либо ингибируют кальций-зависимую фосфатазу - кальциневрин (циклоспорин и FK506), либо ингибируют киназу p70S6, которая важна для индуцируемой фактором роста сигнализации (рапамицин) (Liu et al., Cell 66: 807-815, 1 1; Henderson et al., Immun. 73:316-321, 1991; Bierer et al., Citrr. Opin. mm n. 5:763-773, 93). Согласно дополнительному варианту реализации композиции клеток согласно настоящему изобретению вводят пациенту в сочетании (например, до, одновременно или после) с трансплантацией костного мозга, аблативной терапии в отношении Т-клеток с применением таких химиотерапевтических агентов как флударабин, облучением внешними лучами (ОВЛ), введением циклофосфамида или других антител, таких как ОКТ3 или САМРАТН. Согласно еще одному варианту реализации композиции клеток согласно настоящему изобретению вводят после аблативной терапии в отношении В-клеток, такой как агенты, которые реагируют с CD20, например, ритуксан. Например, согласно одному варианту реализации субъекты могут подвергнуться стандартному лечению высокой дозой химиотерапевтического агента, а затем трансплантации стволовых клеток периферической крови. Согласно определенным вариантам реализации после трансплантации субъектам проводят инфузию размноженных иммунных клеток согласно настоящему изобретению. Согласно одному дополнительному варианту реализации размноженные клетки вводят до или после операции.
Фармацевтическая композиция
Изолированные Т-клетки согласно настоящему изобретению можно вводить одни или в форме фармацевтической композиции в сочетании с растворителями и/или с другими компонентами, такими как IL-2 или другие цитокины, или с популяциями клеток. Если кратко, фармацевтические композиции согласно настоящему изобретению могут содержать Т-клетки, которые описаны в настоящей заявке, в сочетании с одним или более фармацевтически или физиологически приемлемыми носителями, растворителями или вспомогательными веществами. Такие композиции могут содержать буферы, такие как нейтральный солевой буферный раствор, фосфатно-солевой буфер и подобные; углеводы, такие как глюкоза, сахароза или декстраны, маннитол; белки; полипептиды или аминокислоты, такие как глицин; антиоксиданты; хелатирующие агенты, такие как этилендиаминтетрауксусная кислота (ЭДТК) или глутатион; адъюванты (например, гидроксид алюминия); и консерванты. Композиции согласно настоящему изобретению предпочтительно преобразуют в формы для внутривенного введения. Фармацевтические композиции согласно настоящему изобретению можно вводить способом, соответствующим заболеванию, которое требуется лечить (или предупреждать). Объем и частота введения должны определяться такими факторами, как состояние пациента, а также тип и тяжесть заболевания пациента, хотя соответствующие дозировки можно определить путем клинических исследований.
Способ оценки цитотоксичности изолированных Т-клеток CAR и набор для его применения
Еще один вариант реализации настоящего изобретения включает способ оценки цитотоксичности изолированных Т-клеток с химерным рецептором антигена (CAR), таких, как описанные выше целевые клетки, устойчивые к лекарственным препаратам; и указанных изолированных Т-клеток с CAR, экспрессирующих химерный рецептор антигена (CAR), и клеток- мишеней, экспрессирующих по меньшей мере один конкретный поверхностный антиген (и необязательно маркерный ген, такой как люцифераза), способ, включающий:
(а) Обеспечение популяций и указанных Т-клеток, и клеток-мишеней;
(b) Осуществление инкубации популяции указанных Т-клеток по меньшей мере с одной популяцией указанных клеток-мишеней;
(c) Определение процента жизнеспособности указанных специфичных клеток- мишеней.
Ген устойчивости может быть выбран из генов, перечисленных в предыдущем разделе.
Предпочтительно указанным геном устойчивости является dCK. Поверхностный антиген, который должен быть выбран согласно настоящему изобретению, включает один из тех антигенов, которые могут экспрессироваться в Т-клетках посредством химерных рецепторов антигена (CAR), и зависит от клетки, на которую должно быть направлено действие, и он как правило специфичен в отношении раковых клеток. Предпочтительно, поверхностный антиген, который требуется применить в Т- клетках с CAR, - это CD19, поскольку данный антиген, по-видимому, экспрессируется специфично при определенных лимфомах и лейкозах, таких как острый лимфоцитарный лейкоз (ОЛЛ).
И последнее, настоящее изобретение относится к набору для реализации способа для оценки цитотоксичности Т-клеток CAR в отношении клеток-мишеней, включающему:
(d) Популяцию указанных Т-клеток, наделенных CAR, специфичных в отношении определенного антигена;
(e) Указанные клетки-мишени, экспрессирующие указанный антиген;
(f) Необязательно, питательную среду;
причем и Т-клеткам, и клеткам-мишеням была придана устойчивость к химиотерапевтическим препаратам согласно настоящему изобретению.
В настоящей заявке испрашивается патентная защита не только для общего способа конструирования Т-клеток, устойчивых к аналогам пуриновых нуклеотидов (PNA) и 6GT. В более широком смысле она относится к способам получения Т-клеток, которые устойчивы к химиотерапии и аллогенны, включая по меньшей мере следующие объекты:
1) Способ конструирования аллогенных и устойчивых к лекарственным препаратам Т-клеток для иммунотерапии, включающий:
(a) Обеспечение Т-клетки;
(b) Осуществление селекции по меньшей мере с одним лекарственным препаратом, к которому указанная Т-клетка чувствительна;
(c) Осуществление модификации указанной Т-клетки путем инактивации по меньшей мере одного гена, кодирующего компонент рецептора Т-клеток (РТЛ);
(d) Осуществление модификации указанной Т-клетки для придания ей устойчивости к лекарственному препарату;
(e) Осуществление размножения указанной модифицированной Т-клетки, необязательно, в присутствии указанного лекарственного препарата.
2) Способ по п. 1, отличающийся тем, что по меньшей мере один ген, кодирующий компонент РТЛ инактивируют путем экспрессии редкощепящей эндонуклеазы, которая способна расщеплять целевую последовательность в пределах по меньшей мере одного гена, кодирующего компонент РТЛ.
3) Способ по п. 1 или 2, отличающийся тем, что указанную устойчивость к лекарственному препарату придают указанной Т-клетке путем инактивации по меньшей мере одного гена лекарственной чувствительности.
4) Способ по п. 3, отличающийся тем, что указанный ген лекарственной чувствительности инактивируют путем экспрессии редкощепящей эндонуклеазы, которая способна расщеплять целевую последовательность в пределах указанного гена лекарственной чувствительности.
5) Способ по п. 4, отличающийся тем, что указанной редкощепящей эндонуклеазой является TALE-нуклеаза.
6) Способ по п. 3-5, отличающийся тем, что указанным геном лекарственной чувствительности является dCK.
7) Способ по п. 6, отличающийся тем, что ген dCK инактивируют TALE-нуклеазами.
8) Способ по п. 7, отличающийся тем, что инактивацию гена dCK TALE-нуклеазами осуществляют с применением TALE-нуклеаз с последовательностью SEQ ID №63 и SEQ ID №64, а целевая последовательность dCK - SEQ ID №62.
9) Способ по п. 3-5, отличающийся тем, что указанным геном лекарственной чувствительности является ГГФР.
10) Способ по п. 1, отличающийся тем, что указанную устойчивость к лекарственному препарату придают Т-клетке путем экспрессии по меньшей мере одного гена лекарственной устойчивости.
11) Способ по п. 10, отличающийся тем, что указанным геном лекарственной устойчивости является мутированная дигидрофолатредуктаза (ДГФР), которая придает устойчивость к лечению антифолатами, предпочтительно метотрексатом (МТХ).
12) Способ по п. 11, отличающийся тем, что указанная мутированная DHFR содержит по меньшей мере одну аминокислотную мутацию, выбранную из группы, состоящей из: G15, L22, F31, or F34 in the SEQ ID №: 14.
13) Способ по п. 12, отличающийся тем, что указанная мутированная DHFR содержит две аминокислотных мутации в положении L22 и F31 в последовательности SEQ ID №: 14.
14) Способ по п. 10, отличающийся тем, что указанным геном лекарственной устойчивости является мутированная инозин-5'-монофосфат-дегидрогеназа II (IMPDH2), которая придает устойчивость к ингибитору IMPDH, предпочтительно к микофеноляту мотефилу (MMF).
15) Способ по п. 14, отличающийся тем, что указанная мутированная IMPDH2 содержит по меньшей мере одну аминокислотную мутацию в положении Т333 и/или S351 в SEQ ID №: 15.
16) Способ по п. 10, отличающийся тем, что указанным геном лекарственной устойчивости является гетеродимер мутированного кальциневрина (KN) a и/или b, который придает устойчивость к ингибитору кальиценврина, предпочтительно FK506 и/или CsA.
17) Способ по п. 16, отличающийся тем, что указанный гетеродимер мутированного кальциневрина содержит по меньшей мере одну аминокислотную мутацию в положении, выбранном из группы, состоящей из: V314, Y341, М347, Т351, W352, L354 и K360 в SEQ ID №: 16.
18) Способ по п. 17, отличающийся тем, что указанный гетеродимер мутированного кальциневрина содержит аминокислотные мутации в положениях: Т351 и L354 в SEQ ID №: 16.
19) Способ по п. 17, отличающийся тем, что указанный гетеродимер мутированного кальциневрина содержит аминокислотные мутации в положениях: V314 и Y341 в SEQ ID №: 17.
20) Способ по п. 16, отличающийся тем, что указанный гетеродимер мутированного кальциневрина b содержит по меньшей мере одну аминокислотную мутацию в положении, выбранном из группы, состоящей из: V120, N123, L124 и K125 в SEQ ID №: 17.
21) Способ по п. 20, отличающийся тем, что указанный гетеродимер мутированного кальциневрина b содержит аминокислотные мутации в положениях: L124 и K125 в SEQ ID №: 17.
22) Способ по любому из пп. 10-21, отличающийся тем, что указанный ген лекарственной устойчивости экспрессируется в Т-клетке в результате введения в указанную Т-клетку трансгена, кодирующего указанный ген лекарственной устойчивости.
23) Способ по любому из пп. 10-21, отличающийся тем, что указанный ген лекарственной устойчивости экспрессируется в Т-клетке в результате введения в указанную Т-клетку донорной матрицы, которая содержит по меньшей мере одну последовательность, гомологичную эндогенному гену, и последовательность, кодирующую ген лекарственной устойчивости, так что между эндогенным геном и указанной донорной матрицей происходит гомологичная рекомбинация.
24) Способ по п. 23, дополнительно включающий введение в указанную Т-клетку редкощепящей эндонуклеазы, которая способна селективно расщеплять целевую последовательность в пределах указанного эндогенного гена, так что стимулируется скорость гомологичной рекомбинации.
25) Способ по п. 24, отличающийся тем, что указанной редкощепящей эндонуклеазой является TALE-нуклеаза.
26) Способ по любому из пп. 1-25, дополнительно включающий осуществление в Т-клетках экспрессии химерного рецептора антигена.
27) Способ по любому из пп. 1-26, в котором указанным химерным рецептором антигена является CD19+ или CD123+.
28) Способ по любому из пп. 1-27, дополнительно включающий осуществление инактивации гена иммунной контрольной точки.
29) Способ по любому из пп. 1-28, отличающийся тем, что указанные сконструированные Т-клетки размножаются в крови у пациента.
30) Способ по любому из пп. 1-28, отличающийся тем, что указанные сконструированные Т-клетки размножаются in-vitro.
31) Способ по любому из пп. 1-30, отличающийся тем, что указанные сконструированные Т-клетки размножаются в присутствии указанного лекарственного препарата.
32) Изолированная Т-клетка или линия клеток, которые можно получить при помощи способа по любому из пп. 1-31.
33) Изолированная Т-клетка, устойчивая к лекарственному препарату, которая содержит по меньшей мере один нарушенный ген, кодирующий компонент рецептора Т-клеток.
34) Изолированная Т-клетка по п. 33, экспрессирующая по меньшей мере один ген лекарственной устойчивости.
35) Изолированная Т-клетка по п. 33, отличающаяся тем, что указанный ген лекарственной устойчивости выбран из группы, состоящей из: гена ble, гена mcrA и генов, кодирующих мутантную ДГФР, мутантную IMPDH2, мутантный кальциневрин и мутантную AGT.
36) Изолированная Т-клетка по п. 33, содержащая по меньшей мере один нарушенный ген лекарственной чувствительности, предпочтительно ген ГГФТ.
37) Изолированная Т-клетка по любому из пп. 32-36, отличающаяся тем, что указанная изолированная Т-клетка наделена химерным рецептором антигена (CAR), специфичным в отношении конкретного антигена.
38) Изолированная Т-клетка по п. 37, отличающаяся тем, что указанный CAR нацелен на CD19+-клетки или CD123+-клетки;
39) Изолированная Т-клетка по любому из пп. 32-38 для применения в качестве лекарственного препарата.
40) Изолированная Т-клетка по любому из пп. 32-39 для лечения рака, аутоиммунного заболевания или инфекции, вызванной патогеном.
41) Изолированная Т-клетка по п. 40 для применения при лечении острого лимфобластного лейкоза (ОЛЛ) или амиотрофического миелоидного лейкоза (АМЛ).
42) Фармацевтическая композиция, содержащая по меньшей мере одну изолированную Т- клетку по любому из пп. 32-41.
43) Способ лечения пациента, нуждающегося в таком лечении, включающий:
(a) Получение популяции Т-клеток согласно способу по любому из пп. 1-27;
(b) Введение указанных трансформированных Т-клеток указанному пациенту.
44) Способ по п. 36, отличающийся тем, что указанного пациента лечат указанным препаратом, применяемым согласно способу по пп. 1-31.
45) Способ оценки цитотоксичности изолированных Т-клеток с химерным рецептором антигена (CAR) по любому из пп. 32-41 в отношении устойчивых к лекарственным препаратам клеток-мишеней; причем изолированные Т-клетки с CAR экспрессируют химерный рецептор антигена (CAR), а клетки-мишени экспрессируют по меньшей мере один конкретный поверхностный антиген (и не обязательно маркерный ген, такой как люцифераза), включающий:
(a) Получение указанных популяций Т-клеток и клеток-мишеней;
(b) Инкубацию популяции указанных Т-клеток по меньшей мере с указанными специфичными клетками-мишенями;
(c) Определение процента жизнеспособности указанных специфичных клеток-мишеней.
46) Способ по п. 45, отличающийся тем, что указанным геном лекарственной устойчивости является dCK.
47) Способ по п. 44 или п. 45, отличающийся тем, что указанным поверхностным антигеном является CD19.
48) Способ по п. 47, отличающийся тем, что указанной мишенью являются клетки Дауди CD19+ люцифераза+.
49) Набор для реализации способа оценки цитотоксичности Т-клеток с CAR в отношении клетки-мишени, включающий:
(a) Популяцию Т-клеток, наделенных CAR, специфичным в отношении конкретного антигена;
(b) Клетки-мишени, экспрессирующие указанный антиген;
причем и указанным Т-клеткам, и указанным клеткам-мишеням была придана устойчивость к химиотерапевтическому препарату.
ОПРЕДЕЛЕНИЯ
В приведенном выше описании широко применяются некоторые термины. Чтобы облегчить понимание вариантов реализации настоящего изобретения, ниже даны определения некоторых терминов.
- Аминокислотные остатки в полипептидной последовательности обозначены в настоящей заявке согласно однобуквенному коду, по которому, например, Q означает Gln или остаток глутамина, R означает Arg или остаток аргинина, a D означает Asp или остаток аспарагиновой кислоты.
- Нуклеотиды обозначены следующим образом: для обозначения основания нуклеотида применяется однобуквенный код; а означает аденин, t означает тимин, с означает цитозин, a g - гуанин. Для вырожденных нуклеотидов г означает g или а (пуриновые нуклеотиды), k означает g или t, s соответствует g или с, w соответствует а или t, m соответствует а или с, y соответствует t или с (пиримидиновые нуклеотиды), d соответствует g, а или t, v соответствует g, а или с, b соответствует g, t или с, h соответствует a, t или с, а n соответствует g, a, t или с.
- В настоящей заявке термины «нуклеиновая кислота» или «нуклеотид нуклеиновой кислоты» относится к нуклеотидам и/или полинуклеотидам, таким как дезоксирибонуклеиновая кислота (ДНК) или рибонуклеиновая кислота (РНК), олигонуклеотиды, фрагменты, сгенерированные при полимеразной цепной реакции (ПЦР), и фрагменты, сгенерированные при лигировании, расщеплении, действии эндонуклеаз или действии экзонуклеаз. Молекулы нуклеиновых кислот могут состоять из мономеров, которые являются природными нуклеотидами (такие как ДНК и РНК) или аналогами природных нуклеотидов (например, энантиомерные формы природных нуклеотидов), или из сочетания их обоих. Нуклеиновые кислоты могут быть одноцепочечными или двухцепочечными.
- Под «геном» понимают основную единицу наследования, состоящую из сегмента ДНК, выстроенного линейно вдоль хромосомы, который кодирует специфический белок или сегмент белка, малую РНК или т.п. Обычно ген включает промотор, 5'-нетранслируемую область, одну или более кодирующих последовательностей (экзоны), необязательно интроны, 3'- нетранслируемую область. Ген может также содержать терминатор, энхансеры и/или сайленеры.
- Термин "трансген» означает нуклеотидную последовательность (кодирующую, например, один или более полипептидов), которая частично или полностью гетерологична, т.е. чужеродна клетке-хозяину, в которую ее вводят, или гомологична эндогенному гену клетки-хозяина, в которую ее вводят, но которая может быть сконструирована для ее встраивания, или может встраиваться в геном клетки таким образом, что геном клетки, в который она встроилась, изменяется (например, она встраивается в положении, которое отличается от положения в природном гене, или ее встраивание приводит к инактивации (нокауту) гена). Трансген может содержать одну или более последовательностей для регуляции транскрипции и любую другую нуклеиновую кислоту, такую как интроны, которые могут быть необходимы для оптимальной экспрессии избранной нуклеиновой кислоты, кодирующей полипептид. Полипептид кодируемый трансгеном, может либо не экспрессироваться, либо экспрессироваться, но не быть биологически активным, в клетках, в которые введен указанный трансген.
- Под термином «геном» понимают весь генетический материал, содержащийся в клетке, такой как ядерный геном, геном хлоропластов, геном митохондрий.
- Под термином «мутация» понимают замену, делецию, вставку одного или более нуклеотидов/аминокислот в полинуклеотидной (кДНК, ген) или полипептидной последовательности. Указанная мутация может влиять на кодирующую последовательность гена или на его регуляторную последовательность. Также она может влиять на структуру или геномной последовательности или структуру/стабильность кодируемой мРНК.
- Термин «редкощепящая эндонуклеаза» относится к ферменту дикого типа или варианту фермента, способным катализировать гидролиз (расщепление) связей между нуклеиновыми кислотами в молекуле ДНК или РНК, предпочтительно в молекуле ДНК. В частности, указанная нуклеаза может быть эндонуклеазой, более предпочтительно редкощепящей эндонуклеазой, которая обладает высокой специфичностью, распознает целевые сайты нуклеиновой кислоты длиной от 10 до 45 пар оснований (по), обычно длиной от 10 до 35 пар оснований. Эндонуклеаза согласно настоящему изобретению распознает и расщепляет нуклеиновую кислоту в специфических полинуклеотидных последовательностях, также называемых «целевой последовательностью». Редкощепящая эндонуклеаза может распознавать и генерировать одно- или двухцепочечный разрыв а специфических полинуклеотидных последовательностях.
Согласно конкретному варианту реализации указанной редкощепящей эндонуклеазой согласно настоящему изобретению может быть эндонуклеаза Cas9. Фактически недавно было разработано новое средство конструирования генома на основе РНК-управляемой нуклеазы Cas9 из прокариотических CRISPR II типа (короткие полиндромные повторы, регулярно расположенные группами) в системе приобретенного иммунитета (для обзора см. (Sorek, Lawrence et al. 2013)). Ассоциированная с CRISPR система (CAS) была впервые открыта у бактерии, и она функционирует как защита от чужеродной ДНК, вирусной или плазмидной. Для CRISPR-опосредуемого конструирования генома сначала проводят выбор целевой последовательности, часто фланкированной коротким мотивом, называемым мотивом, прилежащим к протоспейсеру (РАМ). После выбора целевой последовательности конструируют специфическую crРНК, комплементарную данной целевой последовательности. Трансактивирующая crРНК (tracrPHK), которая требуется в системах CRISPR II типа, спарена с crРНК и связана с заданным белком Cas9. Cas9 выступает в роли молекулярного якоря, облегчающего спаривание оснований tracPHK с кРНК (Deltcheva, Chylinski et al. 2011). В указанном тройном комплексе двойная структура tracrPHK:crPHK действует как РНК- проводник, которая направляет эндонуклеазу Cas9 при распознавании целевой последовательности. Распознавание мишени комплексом Cas9-tracrPHK:crPHK начинается со сканирования целевой последовательности на гомологию между целевой последовательностью и crРНК. Помимо комплементарности целевой последовательности-crРНК, наведение ДНК требует присутствия короткого мотива, прилежащего к протоспейсеру (мотив, прилежащий к протоспейсеру - РАМ). После спаривания между двойной-РНК и целевой последовательностью Cas9 последовательно производит двухцепоченый разрыв с «липкими» концами на 3 основания выше, чем мотив РАМ (Garneau, Dupuis et al. 2010). Согласно настоящему изобретению РНК-проводник можно сконструировать, например, для специфического направленного действия на ген, кодирующий компонент РТЛ. После спаривания между РНК-проводником и целевой последовательностью Cas9 вызывает расщепление в гене РТЛ.
Редкощепящей эндонуклеазой может быть также хоуминг-эндонуклеаза, также известная под названием мегануклеазы. Такие хоминг-эндонуклеазы хорошо известны в технике (Stoddard 2005). Хоминг-эндонуклеазы обладают высокой специфичностью, распознавая сайты-мишени ДНК длиной от 12 до 45 пар оснований (по), обычно длиной от 14 до 40 по. Хоминг-эндонуклеаза согласно настоящему изобретению может, например, соответствовать эндонуклеазе LAGLIDADG, эндонуклеазе HNH или эндонуклеазе GIY-YIG. Предпочтительной эндонуклеазой согласно настоящему изобретению может быть вариант I-Crel. «Вариант» эндонуклеазы, т.е. эндонуклеаза, которая не существует в природе и которую получают посредством генной инженерии или случайного мутагенеза, может связываться с последовательностями ДНК, отличными от тех, которые распознает эндонуклеаза дикого типа (см. заявку на международный патент WO 2006/097854).
Указанной редкощепящей эндонуклеазой может быть блочная нуклеаза, связывающая ДНК. Под блочной эндонуклеазой, связывающей ДНК, понимают любые химерные белки, содержащие по меньшей мере один каталитический домен эндонуклеазы и по меньшей мере один ДНК-связывающий домен, образованный независимо скручивающимся полипептидом или белковым доменом, который содержит по меньшей мере один мотив, который распознает двух- или одноцепочечные полинуклеотиды. В технике было описано много таких полипептидов, обладающих способностью связываться со специфическими нуклеотидными последовательностями. Такие связывающие домены часто содержат, в качестве неограничивающих примеров, домены лейциновой «молнии», домены «крылатая спираль», домены спираль-петля-спираль, домены HMG-бокс, домены иммуноглобулинов, домен В3 или сконструированный домен цинкового пальца.
Согласно предпочтительному варианту реализации настоящего изобретения НК-связывающий домен получают из подобного активатору транскрипции эффектора (TALE), причем специфичность в отношении последовательности обусловливается серией из 33-35 аминокислотных повторов, происходящих из белков бактерий Xanthomonas или Ralstonia. Указанные повторы существенно отличаются по двум аминокислотным положениям, которые определяют специфичность взаимодействия с парой оснований (Boch, Scholze et al. 2009; Moscou and Bogdanove 2009). Каждая пара оснований в целевой последовательности ДНК контактирует с одиночным повтором, и специфичность возникает вследствие двух вариантов аминокислот в повторах (так называемый дипептид с вариабельными повторами, RDV). TALE- связывающие домены могут дополнительно содержать N-концевой домен транслокации, ответственный за требование первого основания тимина (Т0), в целевой последовательности и С-концевой домен, который содержит клеточные сигналы внутриядерной локализации (NLS).
Домен TALE, связывающий нуклеиновую кислоту, соответствует сконструированному центральному каркасу TALE, содержащему множество повторенных последовательностей TALE, причем каждый повтор содержит RDV, специфичный к каждому нуклеотидному основанию сайта распознавания TALE. Согласно настоящему изобретению каждая повторенная последовательность TALE указанного центрального каркаса выполнена из 30-42 аминокислот, более предпочтительно из 33 или 34, причем две критические аминокислоты (так называемый дипептид с вариабельными повторами, RDV), расположенные в положениях 12 и 13, опосредуют распознавание одного нуклеотида указанного сайта связывания TALE; эквивалентные две критические аминокислоты могут быть расположенные в положениях, отличных от 12 и 13, особенно в повторенной последовательности TALE, которая длиннее чем 33 или 34 аминокислоты. Предпочтительно, RVD, ассоциированные с распознаванием разных нуклеотидов являются HD для распознавания С, NG для распознавания Т, NI для распознавания А, NN для распознавания G или А. Согласно еще одному варианту реализации критические аминокислоты 12 и 13 могут быть мутированы и заменены другими аминокислотными остатками с целью модулирования их специфичности в отношении нуклеотидов А, Т, С и G, и в частности для увеличения такой специфичности. Связывающий нуклеиновую кислоту домен TALE обычно содержит от 8 до 30 повторенных последовательностей TALE. Более предпочтительно, указанный центральный каркас согласно настоящему изобретению содержит от 8 до 20 повторенных последовательностей TALE; еще более предпочтительно 15 повторенных последовательностей TALE. Также он может содержать дополнительную одиночную процессированную повторенную последовательность TALE, образованную 20 аминокислотами, расположенными на С-конце указанного набора из повторенных последовательностей TALE, т.е. дополнительную С-концевую повторенную последовательность полу-TALE.
Другими сконструированными ДНК-связывающими доменами являются блочные специфичные по каждому основанию связывающие нуклеиновую кислоту домены (MBBBD) (PCT/US2013/051783). Указанный MBBBD можно сконструировать, например, из вновь обнаруженных белков, а именно белков EAV36_BURRH, E5AW43_BURRH, E5AW45_BURRH и E5AW46_BURRH из недавно секвенированного генома эндосимбионтного гриба Burkholderia Rhizoxinica (Lackner, Moebius et al. 2011). Белки MBBBD содержат блоки приблизительно из 31-33 аминокислот, которые специфичны к основаниям. Указанные блоки проявляют менее 40% идентичности последовательностей с частыми повторами TALE Xanthomonas, но они проявляют более выраженную вариабельность по полипептидной последовательности. Когда они собираются вместе, такие блочные полипептиды могут, однако, быть нацелены на специфические нуклеотидные последовательности в достаточной степени похоже на TALE- нуклеазы Xanthomonas. Согласно предпочтительному варианту реализации настоящего изобретения указанный ДНК-связывающий домен является сконструированным связывающим доменом MBBBD, содержащим от 16 до 20 модулей.
Разные домены из перечисленных выше белков (блоки, N- и С-концы) из Burkholderia и Xanthomonas полезны при конструировании новых белков или каркасов, обладающих свойствами связывания со специфическими нуклеотидными последовательностями. В частности, дополнительные N-концевые и С-концевые домены конструируемых MBBBD можно получить из природного TALE, такого как AvrBs3, PthXo1, AvrHah1, PthA, Tal1c в качестве неограничивающих примеров.
- Термины «TALE-нуклеаза» или «MBBBD-нуклеаза» относятся к белкам, получаемым при соединении ДНК-связывающего домена, обычно получаемого из белков - подобных активатору транскрипции эффекторов (TALE) или связывающего домена MBBBD, с каталитическим доменом эндонуклеазы. Таким каталитическим доменом предпочтительно является домен нуклеазы и более предпочтительно домен с эндонуклеазной активностью, например, такой как I-Tevl, ColE7, NucA и Fok-I. Согласно конкретному варианту реализации указанной нуклеазой является мономерная TALE-нуклеаза или MBBBD-нуклеаза. Мономерная нуклеаза представляет собой нуклеазу, которой не требуется димеризоваться для специфичного распознавания и расщепления, например, как химеры из сконструированного ДНК-связывающего домена с каталитическим доменом I-Tevl, описанным в WO 2012138927. Согласно еще одному конкретному варианту реализации указанной редкощепящей эндонуклеазой является димерная TALE-нуклеаза или MBBBD-нуклеаза, предпочтительно содержащая ДНК-связывающий домен, соединенный с Fokl. TALE-нуклеаза уже была описана и применена для стимуляции нацеливания на ген и модификаций генов (Boch, Scholze et al. 2009; Moscou and Bogdanove 2009; Christian, Cermak et al. 2010). Такие сконструированные TALE-нуклеазы имеются в продаже под торговым названием TALEN™ (Cellectis, 8 rue de la Croix Jarry, 75013 Paris, France).
- Термин «расщепление» относится к расщеплению ковалентного остова полинуклеотида. Расщепление можно инициировать при помощи целого ряда способов, включая без ограничений ферментативный или химический гидролиз фосфодиэфирной связи. Возможно расщепление как одноцепочечного, так и двухцепочечного полинуклеотида, и расщепление двойной цепи может возникать вследствие двух разных событий расщепления одиночных цепей. Расщепление двухцепочечной ДНК, РНК или гибрида ДНК/РНК может приводить к образованию «липких» концов или ступенчатых концов.
- Под «химерным рецептором антигена» (CAR) понимают химерный рецептор, который содержит внеклеточный лиганд-связывающий домен, трансмембранный домен и домен сигнальной трансдукции.
- В настоящей заявке под термином «внеклеточный лиганд-связывающий домен» понимают олиго- или полипептид, который способен связываться с лигандом. Предпочтительно, указанный домен может быть способен взаимодействовать с молекулой на поверхности клетки. Например, внеклеточный лиганд-связывающий домен можно выбрать так, чтобы он распознавал лиганд, который выступает в роли поверхностного маркера клетки для клеток- мишеней, ассоциированных с конкретным патологическим состоянием.
Согласно предпочтительному варианту реализации указанный внеклеточный домен содержит фрагмент одноцепочечного антитела (scFv), содержащий вариабельный фрагмент легкой (VL) и тяжелой цепи (VH) специфичного моноклонального антитела на целевой антиген, присоединенный гибким линкером. Согласно предпочтительному варианту реализации указанный scFv получают из антитела на CD19 или CD123. Предпочтительно указанный scFV согласно настоящему изобретению включает scFV, полученный из моноклонального антитела на CD19 - 4G7 (Peipp, Saul et al. 2004).
- Домен сигнальной трансдукции или внутриклеточный домен сигнальной трансдукции CAR согласно настоящему изобретению отвечает за внутриклеточную передачу сигнала после связывания внеклеточного лиганд-связывающего домена с мишенью, приводящую к активации иммунной клетки и иммунному ответу. Предпочтительными примерами домена сигнальной трансдукции для применения в составе CAR могут быть цитоплазматические последовательности рецептора Т-клеток и ко-рецепторы, которые действуют согласованно, инициируя передачу сигнала после включения рецептора антигена. Домен сигнальной трансдукции содержит два разных класса цитоплазматических сигнальных последовательностей - последовательности, которые инициируют антиген-зависимую первичную активацию, и последовательности, которые действуют антиген-независимым путем, обеспечивая вторичный или костимулирующий сигнал. Первичная цитоплазматическая сигнальная последовательность может содержать сигнальные мотивы, которые известны под названием иммунорецепторные тирозиновые активирующие мотивы или ITAM. Согласно конкретному варианту реализации домен сигнальной трансдукции CAR согласно настоящему изобретению содержит костимулирующую сигнальную молекулу. Костимулирующщая молекула представляет собой молекулу на поверхности клетки, отличную от антигенного рецептора или их лигандов, которая нужна для эффективного иммунного ответа. Костимулирующие молекулы включают без ограничений молекулу МНС I класса, BTLA и рецептор лиганда Toll. Примеры костимулирующих молекул включают CD27, CD28, CD8, 4-1 ВВ (CD137), ОХ40, CD30, CD40, PD-1, ICOS, лимфоцитарный функциональный антиген-1 (LFA-1), CD2, CD7, LIGHT, NKG2C, В7-Н3 и лиганд, который специфично связывается с CD38, и подобные.
CAR согласно настоящему изобретению экспрессируется на поверхностной мембране клетки. Таким образом, CAR может содержать трансмембранный домен. Отличительные особенности соответствующих трансмембранных доменов включают способность экспрессироваться на поверхности клетки, предпочтительно согласно настоящему изобретению на поверхности иммунной клетки, в частности клеток или естественных киллеров (NK) и взаимодействовать вместе для направления ответа иммунной клетки на заданную клетку-мишень.
Трансмембранный домен может дополнительно содержать область «стебля» между указанным внеклеточным лиганд-связывающим доменом и указанным трансмембранным доменом. В настоящей заявке термин «область стебля» как правило относится к олиго- или полипептиду, который функционирует, связывая трансмембранный домен с внеклеточным лиганд-связывающим доменом. В частности, область «стебля» применяют для достижения большей гибкости доступности внеклеточного лиганд-связывающего домена. Область стебля может содержать до 300 аминокислот, предпочтительно от 10 до 100 аминокислот и наиболее предпочтительно от 25 до 50 аминокислот. Область «стебля» можно получить из целой или части существующей в природе молекулы, например, из целой или части внеклеточной области CD8, CD4 или CD28, или из целой или части константной области антитела. В качестве альтернативы область «стебля» может быть синтетической последовательностью, которая соответствует природной последовательности «стебля», или может быть полностью синтетическая последовательность стебля.
В раковых клетках обычно наблюдается отрицательная регуляция или мутация в целевых антигенах, которая приводит к образованию «ускользающих» вариантов с утраченным антигеном. Таким образом, чтобы скомпенсировать «ускользание» опухоли и сделать иммунные клетки более специфичными в отношении мишени, CD19-специфичный CAR может содержать другие внеклеточные лиганда-связывающие домены, для одновременного связывания разных элементов в мишени, тем самым усиливая активацию и функцию иммунных клеток. Примеры CD19-специфичного CAR включают ScFv FMC63 (Kochenderfer JN, Wilson WH, Janik JE, et al. Eradication of B-lineage cells and regression of lymphoma in a patient treated with autologous T cells genetically engineered to recognize CD19. Blood 2010; 116(20): 4099- 410) или ScFv 4G7 CAR (описан в заявке, поданной под номером РСТ/ЕР2014/059662). Согласно одному варианту реализации внеклеточные лиганд-связывающие домены можно располагать в тандеме в одном трансмембранном полипептиде, и необязательно можно разделять их линкером. Согласно еще одному варианту реализации указанные другие внеклеточные лиганд- связывающие домены могут быть расположены на разных трансмембранных полипептидах, содержащих CAR. Согласно еще одному варианту реализации настоящее изобретение относится к популяции CAR, каждый из которых содержит разные внеклеточные лиганд- связывающие домены. В частности, настоящее изобретение относится к способу конструирования иммунных клеток, включающий получение иммунной клетки и экспрессию на поверхности указанной клетки популяции CAR, каждый из которых содержит разные внеклеточные лиганд-связывающие домены. Согласно еще одному варианту реализации настоящее изобретение относится к способу конструирования иммунных клеток, включающий получение иммунной клетки и введение в указанную иммунную клетку полинуклеотидов, кодирующих полипептиды, содержащие популяции CAR, каждый из которых содержит разные внеклеточные лиганд-связывающие домены. Под популяцией CAR понимают по меньшей мере два, три, четыре, пять, шесть или более CAR, каждый из которых содержит разные внеклеточные лиганд-связывающие домены. Разные внеклеточные лиганд-связывающие домены согласно настоящему изобретению могут предпочтительно одновременно связываться с разными элементами мишени, что усиливает активацию и функцию иммунных клеток. Также настоящее изобретение относится к изолированной иммунной клетке, которая содержит популяцию CAR, каждый из которых содержит разные внеклеточные лиганд-связывающие домены.
- Термин «вектор» относится к молекуле нуклеиновой кислоты, способной транспортировать другую нуклеиновую кислоту, с которой она связана. Вектор согласно настоящему изобретению включает без ограничений вирусный вектор, плазмиду, РНК-вектор или линейный или кольцевой вектор на основе ДНК, или молекулу РНК, которая может содержать хромосомные, нехромосомные, полусинтетические или синтетические нуклеиновые кислоты. Предпочтительными векторами являются вектора, которые способны к автономной репликации (эписомный вектор) и/или экспрессии нуклеиновых кислот, с которыми они соединены (векторы экспрессии). Специалистам в данной области техники известно большое количество подходящих векторов, и они есть в продаже.
- Под «вектором доставки» понимают любой вектор доставки, который можно применять согласно настоящему изобретению для установления контакта с клеткой (т.е., «контактирование») или доставки внутрь клеток или субклеточные компартменты (т.е., «введение») агентов/химических веществ и молекул (белков или нуклеиновых кислот), нужных в рамках настоящего изобретения. Они включают без ограничений липосомальные векторы доставки, вирусные векторы доставки, векторы доставки лекарственных препаратов, химические носители, полимерные носители, липоплексы, полиплексы, дендримеры, микропузырьки (контрастные вещества для ультразвукового исследования), наночастицы, эмульсии или другие подходящие средства переноса.
- Вирусные векторы включают ретровирусы, аденовирусы, парвовирусы (например, аденоассоциированные вирусы), короновирусы, вирусы отрицательными цепями РНК, такие как ортомиксовирусы (например, вирус гриппа) рабдовирус (например, вирус бешенства или вирус везикулярного стоматита), парамиксовирус (например, вирус кори и вирус Сендай), вирусы с положительными цепями РНК, такие как пикорнавирус и альфавирус, и вирусы с двухцепочечной ДНК, включая аденовирус, вирус герпеса (например, вирус простого герпеса 1 и 2 типов, вирус Эпштейна-Барр, цитомегаловирус) и поксивирус (например, вирус коровьей оспы, фоулпокс и канарипокс). Другие вирусы включают, например, норовирус, тогавирус, лавивирус, реовирусы, паповавирус, гепаднавирус и вирус гепатита. Примеры ретровирусов включают: вирус лейкоза-саркомы птиц, вирусы млекопитающих типа С, типа В, типа D, группу HTLV-BLV, лентивирус, спумавирус (Coffin, J.М., Retroviridae: The viruses and their replication, In Fundamental Virology, Third Edition, B.N. Fields, et al., Eds., Lippincott-Raven Publishers, Philadelphia, 1996).
Под «лентивирусным вектором» понимают лентивирусные векторы на основе ВИЧ, которые очень многообещающие в плане доставки генов из-за их относительно высокой упаковочной способности, сниженной иммуногенности и их способности к стабильной трансдукции с высокой эффективностью широкого спектра типов клеток. Обычно лентивирусные векторы генерируют как результат временной трансфекции трех (упаковка, оболочка и перенос) или более плазмид в клетки-продуценты. Подобно ВИЧ лентивирусные векторы входят в клетку- мишень посредством взаимодействия гликопротеинов на поверхности вируса с рецепторами на поверхности клетки. После входа вируса вирусная РНК подвергается обратной транскрипции, которая опосредуется вирусным комплексом обратной транскриптазы. Продуктом обратной транскрипции является двухцепочечная линейная вирусная ДНК, которая подходит для интеграции вируса в ДНК инфицированной клетки. Под «интегративными лентивирусными векторами (или LV)» понимают такие вектора, которые, в качестве неограничивающего примера, способны интегрироваться в геном клетки-мишени. Наоборот, под «неинтегративными лентивирусными векторами (или NLV)» понимают векторы эффективной доставки генов, которые не интегрируются в геном клетки-мишени под действием вирусной интегразы.
- Под клеткой или клетками понимают любые живые клетки эукариот, первичные клетки и линии клеток, полученные из указанных организмов в культурах in vitro.
- Под «первичной клеткой» или «первичными клетками» понимают клетки, взятые прямо из живой ткани (т.е. материал биопсии) и предназначенные для роста in vitro, которые подверглись малому числу удвоения популяции и, следовательно, являются более типичными в отношении основных функциональных компонентов и характеристик тканей, из которых они получены, по сравнению с непрерывными туморогенными или искусственно иммобилизованнми линиями клеток. В качестве неограничивающих примеров линии клеток могут быть выбраны из группы, состоящей из: клеток СНО-K1; клеток HEK293; клеток Сасо2; клеток U2-OS; клеток NIH 3Т3; клеток NSO; клеток SP2; клеток CHO-S; клеток DG44; клеток K-562, клеток U-93; клеток MRC5; клеток IMR90; клеток Юркат; клеток HepG2; клеток HeLa; клеток НТ-1080; клеток НСТ-116; Н клеток u-h7; клеток Huvec; клеток Molt 4.
- Поскольку может возникать некоторая вариабельность на основании геномных данных, на основании которых получают указанные полипептиды, а также чтобы учесть возможность замены некоторых аминокислот, присутствующих в указанных полипептидах без значимой потери активности (функциональные варианты), изобретение включает варианты полипептидов, перечисленных выше, которые обладают по меньшей мере 70%, предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 90% и даже более предпочтительно по меньшей мере 95% идентичностью с последовательностью, предложенной в настоящей заявке на патент.
Таким образом, настоящее изобретение относится к полипептидам, содержащим аминокислотную последовательность, которая обладает по меньшей мере 70%, предпочтительно по меньшей мере 80%, более предпочтительно по меньшей мере 90%, 95% 97% или 99% идентичностью с аминокислотной последовательностью, выбранной из группы, состоящей из SEQ ID №: 8 - SEQ ID №: 20 и SEQ ID №: 26 - SEQ ID №: 35.
- термин «идентичность» относится к идентичности последовательностей между двумя аминокислотными молекулами или полипептидами. Идентичность можно определить путем сравнения положения в каждой последовательности, которые можно выравнивать в целях сравнения. Когда положение в сравниваемой последовательности занято тем же основанием, то молекулы идентичны по данному положению. Степень сходства между нуклеотидными или аминокислотными последовательностями является функцией от числа идентичных или соответствующих нуклеотидов в положениях, общих для указанных нуклеотидных последовательностей. Для расчета идентичности между двумя последовательностями можно применять разные алгоритмы выравнивания, включая FASTA или BLAST, которые доступны как часть пакета для анализа последовательностей GCG (Университет Висконсина, Мэдисон, Висконсин), и могут применяться, например, с параметрами по умолчанию. Например, в изобретение включены полипептиды, обладающие идентичностью с конкретными полипептидами, описанными в настоящей заявке, по меньшей мере 70%, 85%, 90%, 95%, 98% или 99%, а также полинуклеотид, кодирующий такой полипептид;
- «нокаут гена» означает, что ген мутирован в такой степени, что он не может экспрессироваться;
- Термин "TRAC" относится к «константному альфа-рецептору Т-клеток» и соответствует субъединице TCRa константного гена.
Помимо описанных выше свойств настоящее изобретение включает дополнительные свойства, которые станут понятны из следующих примеров, иллюстрирующих способ конструирования аллогенных и устойчивых Т-клеток для иммунотерапии, а также из прилагаемых чертежей.
Пример 1: Создание и описание свойств Т-клеток, устойчивых к клофарабину
Опосредуемая TALE-нуклеазой инактивация dCK
Для инактивации dCK были разработаны, собраны и проверены путем секвенирования две пары TALE-нуклеаз dCK; последующую работу проводили только с парой, названной TALE-нуклеаза dCK2, с последовательностью SEQ ID NO:63 и SEQ ID NO:64. Подробные сведения об общей архитектуре гена dCK (экзоны и интроны) и последовательностях целевых сайтов TALE- нуклеазы, расположенных в экзоне 2, приведены на Фигуре 3.
Целевая последовательность dCK для пары TALE-нуклеаз dCK2 соответствует SEQ ID №62.
После верификации мРНК, кодирующую две TALE-нуклеазы получали, полиаденилировали и применяли для электропорации Т-клеток с применением технологии с быстрым изменением импульсов (применялись 5 или 10 мкг мРНК TALE-нуклеазы слева и справа), как описано в заявке WO 2013/176915. Холодовой температурный шок производили путем инкубации Т-клеток при 30°С сразу после электропорации и в течение 24 часов. Реактивацию (12,5 мкл гранул/106 клеток) проводили в день 8 (8 дней после электропорации).
Образовавшимся Т-клеткам давали вырасти и в итоге описывали их генотипические свойства (посредством анализа с эндо Т7 и глубокого секвенирования в локусах dCK и TRAC), а также фенотипические свойства. Описание их фенотипических свойств включало (i) - проверку способности к росту в присутствии или отсутствии лекарственного препарата, (ii) - определение IC50 PNA, клофарабина и флударабина, в отношении Т-клеток и (III) определение степени инактивации TRAC посредством анализа FACS, когда проводили двойной нокаут генов.
Описание генотипических свойств Т-клеток с нокаутом гена dCK
Чтобы оценить эффективность инактивации гена dCK, клетки, трансфецированные 5 или 10 мкг мРНК TALE-нуклеазы, выращивали в течение 4 дней (D4, 4 дня после электропорации) и отбирали для проведения анализа с Т7 в локусе dCK (Фигура 5).
Последовательности праймеров, применяемых в данном анализе с Т7, соответствуют SEQ ID №68 и SEQ ID №69. Протокол анализа с Т7 описан у Reyon, D., Tsai, S.Q., Khayter, С, Foden, J.A., Sander, J.D., and Joung, J.K. (2012) FLASH assembly of TALE-nucleases for high-throughput genome editing. Nat Biotechnologies.
Результаты данного анализа с эндо Т7 показывают, что когда было трансфецировано 5 и 10 мкг левой и правой dCK2 TALE-нуклеазы, значительный процессинг гена показал, что dCK эффективно инактивируется.
Определение скорости роста Т-клеток с нокаутом dCK
Как показано на Фигуре 6, клетки с нокаутом dCK проявляют скорость роста, аналогичную скорости роста клеток дикого типа. Кроме того, они могут быть повторно активированы на день 8 с той же эффективностью, что и Т-клетки дикого типа.
Селекция Т-клеток с нокаутом dCK в присутствии клофарабина
Т-клеткам с нокаутом dCK или дикого типа давали расти с 8 дня до 13 дня, а затем инкубировали с 1 мкМ клофарабина или без него до 18 дня. Клетки отбирали на 8 день (перед добавлением препарата) и в 18 день (после инкубации с препаратом) и применяли их для проведения анализа с эндо Т7.
Результаты, приведенные на Фигуре 7, показывают, что присутствие 1 мкМ клофарабина в среде к 18 дню селективно обогащало культуру Т-клетками с нокаутом dCK по сравнению с Т-клетками дикого типа (2 полосы с более низкой молекулярной массой для Т-клеток с нокаутом dCK по сравнению с одной полосой более высокой молекулярной массы для Т-клеток дикого типа). Это указывает на то, что TALE-нуклеаза-опосредованная инактивация dCK позволяет проводить селекцию устойчивых к лекарственному препарату Т-клеток относительно Т-клеток дикого типа. Таким образом, Т-клетки с нокаутом dCK способны выдерживать присутствие 1 мкМ клофарабина, которое соответствует клинически значимой дозе для лечения острого лимфобластного лейкоза (ОЛЛ) в соответствии с Cmax, сообщаемой Европейским агентством по лекарственным средствам (ЕМА).
Определение IC50 для клофарабина в отношении Т-клеток с нокаутом dCK по сравнению с Т- клетками дикого типа
Чтобы более подробно исследовать способность Т-клеток выдерживать присутствие клофарабина, IC50 для данного препарата определяли при действии на Т-клетки с нокаутом dCK и Т-клетки дикого типа. Клетки отбирали через 3 дня после трансфекции и инкубировали в течение 2 дней в присутствии возрастающей концентрации клофарабина (0-10 мкМ). В конце инкубации с клофарабином жизнеспособность Т-клеток определяли посредством анализа FACS2.
Результаты, представленные на Фигуре 8, явно показывают, что процессинг гена dCK, опосредуемый TALE-нуклеазами, эффективно инактивирует активность dCK в Т-клетках. Такая инактивация коррелирует с устойчивостью к клоффарабину, в отличие от чувствительности к нему Т-клеток дикого типа. Значения IC50 (количество препарата, которое требуется добавить в среду, чтобы уменьшить жизнеспособность клеток до 50%) соответствуют приблизительно 100 нМ и 10 мкМ для Т-клеток дикого типа и Т-клеток с нокаутом dCK.
В совокупности указанный первый набор данных позволяет заключить, что опосредуемая TALE- нуклеазой инактивация гена dCK является достаточной. Инактивация dCK не нарушает рост сконструированных Т-клеток, но позволяет им выдерживать клинически значимую дозу клофарабина.
Пример 2. Создание и описание свойств аллогенных Т-клеток, устойчивых к клофарабину
Для разработки и получения устойчивых к клофарабину аллогенных Т-клеток с CAR, одновременно инактивировали гены dCK и TRAC. После демонстрации в примере 1, что инактивация dCK была успешной, генерировали Т-клетки с двойным нокаутом TRAC/dCK и описывали их свойства. Параллельно выполнялись две последовательности действий, представленные на Фигуре 9. Один из них соответствует периоду инкубации клеток в присутствии клофарабина - 5 дней.
Описание генотипа
Чтобы сначала оценить эффективность, а также кинетику инактивации генов TRAC и/или dCK, трансфецированные клетки выращивали в течение 6 дней и отбирали их в день 1, день 3 и день 6 для проведения анализа с Т7 в локусах dCK и TRAC. Чтобы достигнуть этого, 2 пары праймеров с последовательностями SEQ ID №68 и №69; и SEQ ID №70 и №71, соответственно, применяли в анализе с Т7 для локусов dCK и TRAC.
Применяемый протокол был описан у Reyon, D., Tsai, S.Q., Khayter, С, Foden, J.A., Sander, J.D., and Joung, J.K. (2012) FLASH assembly of TALE-nucleases for high-throughput genome editing. Nat Biotechnol
Результаты, представленные на Фигуре 10, показывают, что опосредуемое TALE-нуклеазой одиночный нокаут TRAC и dCK высоко эффективно уже в день 1. Даже несмотря на то, что клетки с инактивированными генами нельзя охарактеризовать как однородную популяцию, представляется, что двойной нокаут TRAC/dCK также высоко эффективен.
Клетки затем выращивали в присутствии или отсутствии 1 мкМ клофарабина. В день 6 (шесть дней после трансфекции) и после 3-дневного культивирования в присутствии или отсутствии клофарабина клетки отбирали, и эффективность нокаута гена dCK определяли посредством анализа с Т7 и высокоэффективного секвенирования ДНК.
Протокол, применяемый для глубокого секвенирования, описан у Shendure, J., & Ji, Н. (2008). Next-generation DNA sequencing. Nature biotechnology, 26(10), 1135-1145.
Результаты, представленные на Фигуре 11, показывают, что частота инсерционно-делеционных мутаций, генерируемых в локусе dCK, во всех экспериментах составляет около 80-90%. Это опять же указывает на то, что опосредуемая TALE-нуклеазой инактивация dCK высоко эффективна, даже когда ее сочетают с одновременной инактивацией TRAC. Присутствие 1 мкМ клофарабина в питательной среде в течение 5 дней не приводит к увеличению полосы Т7, которая специфична для нокаута по dCK, как видно в первой серии экспериментов. Это свидетельствует о том, что в данном конкретном эксперименте инактивация dCK была достаточно успешной, чтобы сконструированные Т-клетки могли расти в присутствии клофарабина. Интересно, что это указывает на то, что если нокаут dCK достаточно эффективен, нет надобности проводить селекцию Т-клеток в присутствии клофарабина для получения устойчивых к лекарственному препарату Т-клеток. Следовательно, данное свойство представляет собой явное преимущество при получении устойчивых к лекарственному препарату аллогенных Т-клеток.
Фенотипическая оценка эффективности нокаута TCAR
Т-клетки с нокаутом TRAC, отобранные для проведения эксперимента с двойным нокаутом, анализировали и очищали посредством FACS (сортировка флуоресцентно-активированных клеток) (CliniMACS). Результаты, представленные на Фигуре 12В, показывают эксперимент с мечением Т-клеток с анти-РТЛ mAb-PE (моноклональное антитело - полиэтилен) или без него. Также Фигура 12В относится к мечению mAb-PE Т-клетки в среде с клофарабином или без него, перед и после очистки Т-клеток с нокаутом TRAC.
Результаты показывают, что эффективность нокаута РТЛ в Т-клетках, обработанных мРНК TRAC и dCK (двойной нокаут dCK/TRAC), высока (приблизительно 85%). Способ очистки позволяет проводить эффективную селекцию/очистку РТЛ-негативных клеток с получением чистоты до 99,3%.
Описание фенотипа Т-клеток с нокаутом TRAC/dCK
На Фигуре 13 показана скорость роста Т-клеток в отсутствие клофарабина. Даже если Т-клетки с нокаутом dCK проявляют небольшой дефект роста, их можно повторно активировать в день 10 с той же эффективностью, что и Т-клетки дикого типа.
На Фигуре 14 показана скорость роста Т-клеток в присутствии клофарабина. Данный эксперимент был проведен на Т-клетках с CAR с двойным нокаутом dCK/TCAR (FMC63, которые описаны в заявке на патент под регистрационным номером РСТ/ЕР2014/059662) путем культивирования данных клеток в течение 11 дней в среде, содержащей разные концентрации клофарабина (от 0,1 мкМ до 10 мкМ). Результаты, представленные на Фигуре 14, явно показывают, что увеличение количества клеток для Т-клеток с CAR с двойным нокаутом dCK/TCAR происходит до концентрации клофарабина 1 мкМ (что соответствует Сmax), даже если рост менее выражен, чем рост данных клеток в отсутствие препарата.
Определение IC50 для клофарабина в отношении сконструированных Т-клеток по сравнению с Т-клетками дикого типа
Чтобы более подробно исследовать способность Т-клеток с двойным нокаутом выдерживать присутствие клофарабина, определяли IC50 для данного препарата. Т-клетки выращивали с клофарабином или без него с дня 3 до дня 8 (см. последовательность действий 2 на Фигуре 9), затем их инкубировали в течение 2 дней (день 15 - день 17) в среде с разными концентрациями клофарабина. Жизнеспособность Т-клеток определяли посредством анализа FACS с применением набора для количественной оценки яркости.
Результаты, представленные на Фигуре 15, показывают, что Т-клетки с нокаутом dCK и dCK/TRAC проявляют значительную способность переносить клофарабин по сравнению с Т- клетками отрицательного контроля и с Т-клетками с нокаутом только TRAC. Примечательно, что селекция клеток с применением 1 мкМ клофарабина в течение 5 дней с дня 3 по день 8 (см. последовательность действий 2 на Фигуре 9) не улучшала их способность переносить клофарабин. Это свидетельствует о том, что инактивация dCK достаточно эффективна, и что 5- дневная инкубация для селекции с препаратом для получения устойчивых к клофарабину аллогенных Т-клеток с CAR не требуется.
Цитотоксичность получения устойчивых к лекарственному препарату аллогенных Т-клеток с CAR
Анализ цитотоксичности проводили следующим образом: 10 Т-клеток с CAR (FMC63, для справки см. выше) инкубировали с клетками Дауди (специфические мишени) и клетками К562 (не специфические мишени) в течение 5 часов. Затем клетки отбирали и жизнеспособность клеток Дауди и К562 определяли путем подсчета частоты направленного лизиса клеток.
Результаты, представленные на Фигуре 16, показывают, что Т-клетки с CAR с двойным нокаутом dCK/TRAC проявляют сходную направленную цитотоксичность, что и Т-клетки с CAR дикого типа (35% направленной цитотоксичности). Это указывает на то, что инактивация генов dCK и TRAC не влияет на цитотоксичность Т-клеток с CAR FMC63.
Затем указанные клетки применяли для определения их чувствительности к клофарабину и флударабину, как проводилось ранее. Результаты, представленные на Фигуре 17, показывают, что Т-клетки с CAR с нокаутом dCK/TRAC обладают значительной способностью переносить клофарабин по сравнению с Т-клетками с CAR отрицательного контроля (IC50=500 нМ и 0,1 нМ, соответственно). Аналогичные результаты были получены с флударабином (IC50=400 мкМ и 10 мкМ для Т-клеток с CAR с двойным нокаутом и Т-клеток с CAR, соответсвенно).
Выводы
В совокупности данные эксперименты показывают, что одновременная инактивация генов dCK и TRAC высоко эффективна и позволяет генерировать более 70% Т-клеток с двойным нокаутом в одном цикле электропорации. Интересно, что в связи с такой высокой эффективностью нет необходимости в длительном этапе селекции. Сконструированные Т-клетки проявляют выраженную способность переносить клофарабин и остаются на своем максимуме жизнеспособности под давлением клинически значимой дозы клофарабина.
Пример 3- Создание устойчивых к клофарабину клеток Дауди
Цель состоит в получении устойчивых к лекарственному препарату клеток-мишеней Дауди CD19+/Luc+ для оценки цитотоксичности устойчивых к клофарабину аллогенных Т-клеток с CAR.
Описание генотипа клеток Дауди с нокаутом dCK
Готовили мРНК TALE-нуклеазы dCK, и проводили электропорацию клеток Дауди мРНК TALE-нуклеазы dCK в соответствии с протоколами, описанными в WO 2013/176915.
Для оценки эффективности анализа нокаута dCK проводили анализ с Т7, как в Примере 1. Анализ проводили через 2 дня после трансфекции. Последовательности праймеров соответствовали SEQ ID №68 и №69.
Результаты, представленные на Фигуре 18, показывают высокую степень инактивации гена dCK.
Описание фенотипа клеток Дауди с нокаутом dCK
Клетки Дауди культивировали в среде, содержащей разные концентрации клофарабина (0; 0,1; 0,25; 0,5 и 1 мкМ) в течение нескольких дней и подсчитывали клетки в каждом пассаже.
Результаты, представленные на Фигуре 19, показывают, что клетки Дауди с нокаутом dCK способны расти в присутствии клофарабина в концентрации до 1 мкМ. Их скорость роста была сходна со скоростью роста Т-клеток дикого типа в отсутствии клофарабина, что свидетельствует о том, что инактивация dCK не ухудшает способность клеток Дауди к росту. Как ожидалось, рост клеток Дауди дикого типа был явно нарушен. Данные результаты демонстрируют, что клетки CD19+-Luc+-GFP+ c нокаутом dCK были сгенерированы успешно.
Пример 4. Создание описание свойств Т-клеток, устойчивых к 6TG
Для получения Т-клеток, устойчивых к 6МР и 6TG (Т-клетки с нокаутом ГГФТ), ген ГГФТ инактивировали при посредстве TALE-нуклеазы следующим образом. Общая архитектура гена ГГФТ (экзоны и интроны) и положение разных целевых сайтов TALE-нуклеазы приведены на Фигуре 20.
Опосредуемая TALE-нуклеазой инактивация гена ГГФТ
На Фигуре 21 показана последовательность действий, применяемая в данном эксперименте для генерирования и описания свойств Т-клеток с нокаутом ГГФТ. Для инактивации гена ГГФТ были разработаны 2 пары TALE-нуклеазы для ГГФТ, они были собраны и проверены посредством секвенирования (для ГГФТ 1: SEQ ID №74 и SEQ ID №75; для ГГФТ2: SEQ ID №77 и SEQ ID №78). Подробные сведения об общей архитектуре гена ГГФТ (экзоны и интроны) и расположении целевых сайтов TALE-нуклеазы приведены на Фигуре 20. Целевые последовательности пар TALE-нуклеаз ГГФТ1 и ГГФТ2 соответствуют SEQ ID №76 и SEQ ID №79, соответственно.
Описание генотипа Т-клеток с нокаутом ГГФТ
Генотип Т-клеток с нокаутом ГГФТ был охарактеризован в день 47 посредством анализа с Т7, и была показана инактивация гена ГГФТ в Т-клетках. Пара праймеров в данном анализе обладала последовательностями SEQ ID №72 и SEQ ID №73. Результаты, представленные на Фигуре 22, показывают, что пара TALE-нуклеаз ГГФТ была способна с высокой эффективностью процессировать ген ГГФТ.
Скорость роста Т-клеток с нокаутом ГГФТ
Согласно результатам, представленным на Фигуре 23, клетки с нокаутом ГГФТ демонстрируют скорость роста, аналогичную скорости роста Т-клеток дикого типа, несмотря на то, что рост несколько снижен для пары TALE-нуклеазы ГГФТ2 (эксперимент проводили с 10 мкг TALE-нуклеазы). Тем не менее Т-клетки, инактивированные 10 мкг TALE-нуклеазы ГГФТ2, были повторно активированы на 10-й день с той же эффективностью, что о Т-клетки дикого типа, что указывает на то, что инактивация ГГФТ не нарушает значительно роста Т-лимофцитов. В следующих экспериментах была выбрана пара TALE-нуклеазы ГГФТ1.
Селекция Т-клеток с нокаутом ГГФТ в присутствии 6TG
Т-клеткам с нокаутом ГГФТ или дикого типа позволяли расти с дня 8 по день 13, а затем инкубировали в присутствии или в отсутствии 1 мкМ 6TG до дня 18 (последовательность действий показана на Фигуре 22). Клетки отбирали в день 8 (перед добавлением лекарственного препарата) и вдень 18 (после инкубации с лекарственным препаратом) и применяли для проведения анализа с эндо Т7. Пара праймеров в данном анализе обладала последовательностями SEQ ID №72 и SEQ ID №73. Результаты, представленные на Фигуре 24, показывают, что присутствие 1 мкМ 6TG в среде позволяет культуре селективно обогащаться Т-клетками с нокаутом ГГФТ (как видно по меньшей плотности полосы дикого типа в присутствии 6TG в день 18).
Генерирование Т-клеток с CAR с нокаутом ГГФТ
Чтобы изучить влияние инактивации ГГФТ на цитотоксическую активность Т-клеток с CAR, проводили трансдукцию Т-клеток лентивирусным вектором с CAR 4G7 (таким как описан в заявке, поданной под регистрационным номером РСТ/ЕР2014/059662), а затем их электропорацию мРНК, кодирующей TALE-нуклеазу ГГФТ1. Все эксперименты, описанные ниже, проводили с сконструированными Т-клетками, сгенерированными без какой-либо селекции на 6TG. Эффективность процессинга ГГФТ оценивали путем анализа с эндо Т7. Пара праймеров, примененная в данном анализе, обладала последовательностями SEQ ID №72 и SEQ ID №73. Результаты, представленные на Фигуре 25, показывают, что ген ГГФТ был успешно инактивирован в присутствии или отсутствии CAR 4G7. В Т-клетках была достигнута более выраженная инактивация ГГФТ, чем в Т-клетках с CAR.
Цитотоксические свойства в Т-клеток с CAR с нокаутом ГГФТ в отношении клеток ДАУДИ
Анализ цитотоксичности проводился, как схематически показано на фигуре 27. Набор из 10 Т-клеток с CAR инкубировали в течение 5 часов с клетками Дауди (специфические мишени) и клетками K562 (неспецифические мишени). Затем клетки отбирали, и жизнеспособность клеток определяли путем подсчета частоты направленного лизиса клеток. Результаты, представленные на Фигуре 26, показывают, что Т-клетки с CAR с нокаутом ГГФТ проявляют сходную направленную цитотоксичность, что и Т-клетки с CAR дикого типа. Это свидетельствует о том, что инактивация гена ГГФТ не влияет не цитотоксичность Т-клеток с CAR 4G7
Определение IC50 для 6TG в отношении сконструированных Т-клеток по сравнению с Т- клетками дикого типа
Результаты, представленные на Фигуре 27, показывают, что процессинг гена ГГФТ (как было видно ранее при анализе с Т7) эффективно подавляет активность ГГФТ в Т-клетках. Такая инактивация придает устойчивость к 6TG, в отличие от чувствительности Т-клеток дикого типа к данному препарату. IC50 можно приблизительно определить как 10 нМ и >100 мкМ для Т-клеток дикого типа с нокаутом ГГФТ, соответственно.
Выводы
В совокупности данные результаты показывают, что инактивация гена ГГФТ эффективна. Такая инактивация позволяет Т-клеткам переносить высокую дозу 6TG без необходимости очистки, которая является времязатратным процессом. Также показано, что инактивацию ГГФТ можно проводить в Т-клетках с CAR в несколько меньшей степени. Такая инактивация не ухудшает цитотоксических свойств Т-клеток с CAR в отношении клеток Дауди.
ссылки
Bardenheuer, W., K. Lehmberg, et al. (2005). "Resistance to cytarabine and gemcitabine and in vitro selection of transduced cells after retroviral expression of cytidine deaminase in human hematopoietic progenitor cells." Leukemia 19(12): 2281-8.
Betts, M.R., J.M. Brenchley, et al. (2003). "Sensitive and viable identification of antigen-specific CD8+ T cells by a flow cytometric assay for degranulation." J Immunol Methods 281(1-2): 65-78.
Boch, J., H. Scholze, et al. (2009). "Breaking the code of DNA binding specificity of TAL-type III effectors." Science 326(5959): 1509-12.
Brewin, J., C. Mancao, et al. (2009). "Generation of EBV-specific cytotoxic T cells that are resistant to calcineurin inhibitors for the treatment of posttransplantation lymphoproliferative disease." Blood 114(23): 4792-803.
Cermak, Т., E.L. Doyle, et al. (2011). "Efficient design and assembly of custom TALEN and other TAL effector-based constructs for DNA targeting." Nucleic Acids Res 39(12): e82.
Christian, M., T. Cermak, et al. (2010). "Targeting DNA double-strand breaks with TAL effector nucleases." Genetics 186(2): 757-61.
Cong, L., F.A. Ran, et al. (2013). "Multiplex genome engineering using CRISPR/Cas systems." Science 339(6121): 819-23.
Critchlow, S.E. and S.P. Jackson (1998). "DNA end-joining: from yeast to man." Trends Biochem Sci 23(10): 394-8.
Dasgupta, A., D. McCarty, et al. (2011). "Engineered drug-resistant immunocompetent cells enhance tumor cell killing during a chemotherapy challenge." Biochem Biophys Res Commun 391(1): 170-5.
Deltcheva, E., K. Chylinski, et al. (2011). "CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III." Nature 471(7340): 602-7.
Deng, D., C. Yan, et al. (2012). "Structural basis for sequence-specific recognition of DNA by TAL effectors." Science 335(6069): 720-3.
Garneau, J.E., M.E. Dupuis, et al. (2010). "The CRISPR/Cas bacterial immune system cleaves bacteriophage and plasmid DNA." Nature 468(7320): 67-71.
Gasiunas, G., R. Barrangou, et al. (2012). "Cas9-crRNA ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria." Proc Natl Acad Sci USA 109(39): E2579-86.
Geissler, R., H. Scholze, et al. (2011). "Transcriptional activators of human genes with programmable DNA-specificity." PLoS One 6(5): e19509.
Hacke, K., J.A. Treger, et al. (2013). "Genetic modification of mouse bone marrow by lentiviral vector-mediated delivery of hypoxanthine-Guanine phosphoribosyltransferase short hairpin RNA confers chemoprotection against 6-thioguanine cytotoxicity." Transplant Proc 45(5): 2040-4.
Huang, P., A. Xiao, et al. (2011). "Heritable gene targeting in zebrafish using customized TALENs." Nat Biotechnol 29(8): 699-700.
Ikehara, Y., S.K. Ikehara, et al. (2004). "Negative regulation of T cell receptor signaling by Siglec-7 (p70/AIRM) and Siglec-9." J Biol Chem 279(41): 43117-25.
Jena, В., G. Dotti, et al. (2010). "Redirecting T-cell specificity by introducing a tumor-specific chimeric antigen receptor." Blood 116(7): 1035-44.
Jinek, M., K. Chylinski, et al. (2012). "A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity." Science 337(6096): 816-21.
Jonnalagadda, M., С.E. Brown, et al. (2013). "Engineering human T cells for resistance to methotrexate and mycophenolate mofetil as an in vivo cell selection strategy." PLoS One 8(6): e65519.
Kushman, M.E., S.L. Kabler, et al. (2007). "Expression of human glutathione S-transferase P1 confers resistance to benzo[a]pyrene or benzo[a]pyrene-7,8-dihydrodiol mutagenesis, macromolecular alkylation and formation of stable N2-Gua-BPDE adducts in stably transfected V79MZ cells co- expressing hCYP1A1." Carcinogenesis 28(1): 207-14.
Lackner, G., N. Moebius, et al. (2011). "Complete genome sequence of Burkholderia rhizoxinica, an Endosymbiont of Rhizopus microsporus." J Bacteriol 193(3): 783-4.
Li, L., M.J. Piatek, et al. (2012). "Rapid and highly efficient construction of TALE-based transcriptional regulators and nucleases for genome modification." Plant Mol Biol 78(4-5): 407-16.
Li, Т., S. Huang, et al. (2011). "Modularly assembled designer TAL effector nucleases for targeted gene knockout and gene replacement in eukarvotes." Nucleic Acids Res 39(14): 6315-25.
Ma, J.L, E.M. Kim, et al. (2003). "Yeast Mrell and Radl proteins define a Ku-independent mechanism to repair double-strand breaks lacking overlapping end sequences." Mol Cell Biol 23(23): 8820-8.
Mahfouz, M.M., L. Li, et al. (2012). "Targeted transcriptional repression using a chimeric TALE-SRDX repressor protein." Plant Mol Biol 78(3): 311-21.
Mahfouz, M.M., L. Li, et al. (2011). "De novo-engineered transcription activator-like effector (TALE) hybrid nuclease with novel DNA binding specificity creates double-strand breaks." Proc Natl Acad Sci USA 108(6): 2623-8.
Mak, A.N., P. Bradley, et al. (2012). "The crystal structure of TAL effector PthXo1 bound to its DNA target." Science 335(6069): 716-9.
Mali, P., L. Yang, et al. (2013). "RNA-guided human genome engineering via Cas9." Science 339(6121): 823-6.
Maze, R., C. Kurpad, et al. (1999). "Retroviral-mediated expression of the P140A, but not P140A/G156A, mutant form of O6-methylguanine DNA methyltransferase protects hematopoietic cells against O6-benzylguanine sensitization to chloroethylnitrosourea treatment." J Pharmacol Exp Ther 290(3): 1467-74.
Meyaard, L., G.J. Adema, et al. (1997). "LAIR-1, a novel inhibitory receptor expressed on human mononuclear leukocytes." Immunity 7(2): 283-90.
Miller, J.C, S. Tan, et al. (2011). "A TALE nuclease architecture for efficient genome editing." Nat Biotechnol 29(2): 143-8.
Morbitzer, R., P. Romer, et al. (2011). "Regulation of selected genome loci using de novo-engineered transcription activator-like effector (TALE)-type transcription factors." Proc Natl Acad Sci U S A 107(50): 21617-22.
Moscou, M.J. and A.J. Bogdanove (2009). "A simple cipher governs DNA recognition by TAL effectors." Science 326(5959): 1501.
Mussolino, C, R. Morbitzer, et al. (2011). "A novel TALE nuclease scaffold enables high genome editing activity in combination with low toxicity." Nucleic Acids Res 39(21): 9283-93.
Nicoll, G., J. Ni, et al. (1999). "Identification and characterization of a novel siglec, siglec-7, expressed by human natural killer cells and monocytes." J Biol Chem 274(48): 34089-95.
Nivens, M.С, Т. Felder, et al. (2004). "Engineered resistance to camptothecin and antifolates by retroviral coexpression of tyrosyl DNA phosphodiesterase-I and thymidylate synthase." Cancer Chemother Pharmacol 53(2): 107-15.
Park, T.S., S.A. Rosenberg, et al. (2011). "Treating cancer with genetically engineered T cells." Trends Biotechnol 29(11): 550-7.
Quigley, M., F. Pereyra, et al. (2010). "Transcriptional analysis of HIV-specific CD8+ T cells shows that PD-1 inhibits T cell function by upregulating BATF." Nat Med 16(10): 1147-51.
Sander, J.D., L. Cade, et al. (2011). "Targeted gene disruption in somatic zebrafish cells using engineered TALENs." Nat Biotechnol 29(8): 697-8.
Sangiolo, D., M. Lesnikova, et al. (2007). "Lentiviral vector conferring resistance to mycophenolate mofetil and sensitivity to ganciclovir for in vivo T-cell selection." Gene Ther 14(21): 1549-54.
Schweitzer, В.I., A.P. Dicker, et al. (1990). "Dihydrofolate reductase as a therapeutic target." Faseb J 4(8): 2441-52.
Sorek, R., С.M. Lawrence, et al. (2013). "CRISPR-mediated Adaptive Immune Systems in Bacteria and Archaea." Annu Rev Biochem.
Stoddard, B.L. (2005). "Homing endonuclease structure and function." Q Rev Biophys 38(1): 49-95.
Sugimoto, Y., S. Tsukahara, et al. (2003). "Drug-selected co-expression of P-glycoprotein and gp91 in vivo from an MDRl-bicistronic retrovirus vector Ha-MDR-IRES-gp91." J Gene Med 5(5): 366-76.
Takebe, N., S.C. Zhao, et al. (2001). "Generation of dual resistance to 4- hydroperoxycyclophosphamide and methotrexate by retroviral transfer of the human aldehyde dehydrogenase class 1 gene and a mutated dihydrofolate reductase gene." Mol Ther 3(1): 88-96.
Tesson, L., C. Usal, et al. (2011). "Knockout rats generated by embryo microinjection of TALENs." Nat Biotechnol 29(8): 695-6.
Weber, E., R. Gruetzner, et al. (2011). "Assembly of designer TAL effectors by Golden Gate cloning." PLoS One 6(5): e19722.
Yam, P., M. Jensen, et al. (2006). "Ex vivo selection and expansion of cells based on expression of a mutated inosine monophosphate dehydrogenase 2 after HIV vector transduction: effects on lymphocytes, monocytes, and CD34+ stem cells." Mol Ther 14(2): 236-44.
Zhang, F., L. Cong, et al. (2011). "Efficient construction of sequence-specific TAL effectors for modulating mammalian transcription." Nat Biotechnol 29(2): 149-53.
Zhang, J.Q., G. Nicoll, et al. (2000). "Siglec-9, a novel sialic acid binding member of the immunoglobulin superfamily expressed broadly on human blood leukocytes." J Biol Chem 275(29): 22121-6.
Zielske, S.P., J.S. Reese, et al. (2003). "In vivo selection of MGMT(P140K) lentivirus-transduced human NOD/SCID repopulating cells without pretransplant irradiation conditioning." J Clin Invest 112(10): 1561-70.
| название | год | авторы | номер документа |
|---|---|---|---|
| МНОГОЦЕПОЧЕЧНЫЙ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР И ЕГО ПРИМЕНЕНИЯ | 2013 |
|
RU2663725C2 |
| Химерные антигенные рецепторы, нацеленные на антиген созревания B-клеток | 2016 |
|
RU2706582C2 |
| СПОСОБ СОЗДАНИЯ Т-КЛЕТОК, ПРИГОДНЫХ ДЛЯ АЛЛОГЕННОЙ ТРАНСПЛАНТАЦИИ | 2015 |
|
RU2752933C2 |
| НАЦЕЛИВАНИЕ НА КЛЕТКИ, ОПОСРЕДУЕМОЕ ХИМЕРНЫМ АНТИГЕННЫМ РЕЦЕПТОРОМ | 2018 |
|
RU2783316C2 |
| ИНСЕРЦИЯ ТАРГЕТНОГО ГЕНА ДЛЯ УЛУЧШЕННОЙ КЛЕТОЧНОЙ ИММУНОТЕРАПИИ | 2017 |
|
RU2824204C2 |
| СПОСОБЫ КОНСТРУИРОВАНИЯ АЛЛОГЕННЫХ И ВЫСОКОАКТИВНЫХ Т-КЛЕТОК ДЛЯ ИММУНОТЕРАПИИ | 2014 |
|
RU2736616C2 |
| СПОСОБЫ КОНСТРУИРОВАНИЯ ВЫСОКОАКТИВНЫХ Т-КЛЕТОК ДЛЯ ИММУНОТЕРАПИИ | 2014 |
|
RU2725542C2 |
| ЦЕЛЕВАЯ ГЕННАЯ ИНТЕГРАЦИЯ ГЕНОВ-ИНГИБИТОРОВ NK ДЛЯ УЛУЧШЕННОЙ ИММУННОЙ КЛЕТОЧНОЙ ТЕРАПИИ | 2018 |
|
RU2797304C2 |
| CD123-СПЕЦИФИЧЕСКИЕ ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ ДЛЯ ИММУНОТЕРАПИИ РАКА | 2015 |
|
RU2727290C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ, НАЦЕЛЕННЫЕ НА FLT3 | 2018 |
|
RU2820859C2 |





























































































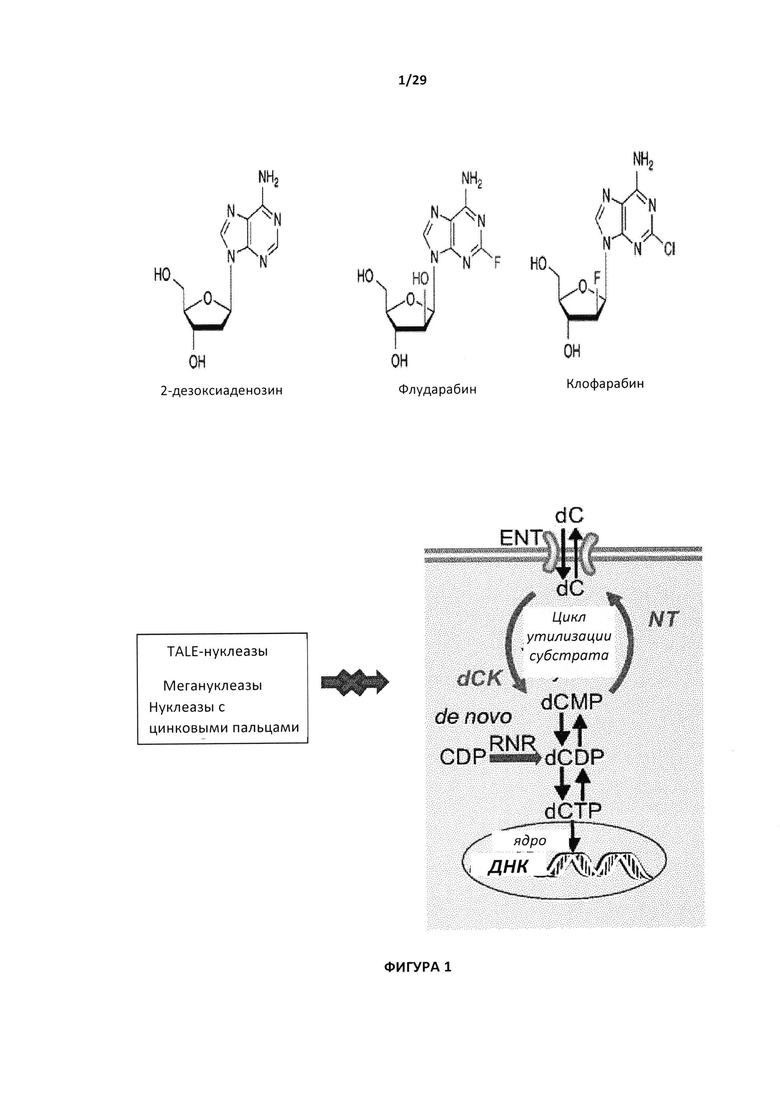
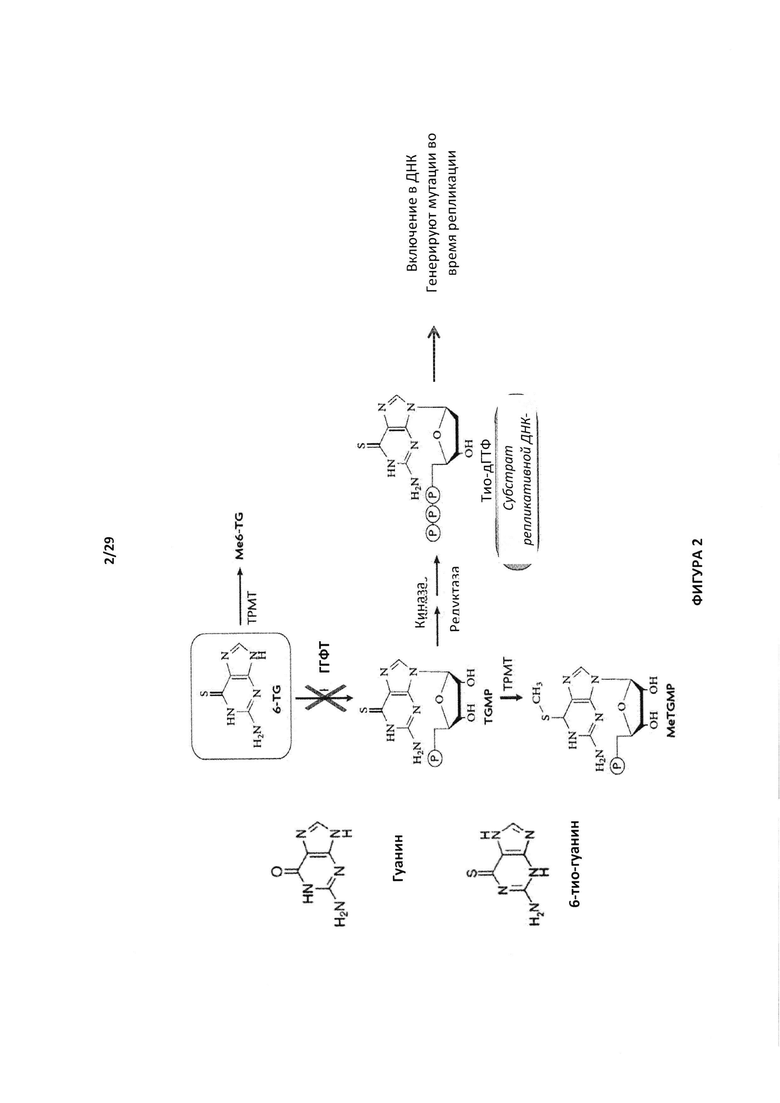
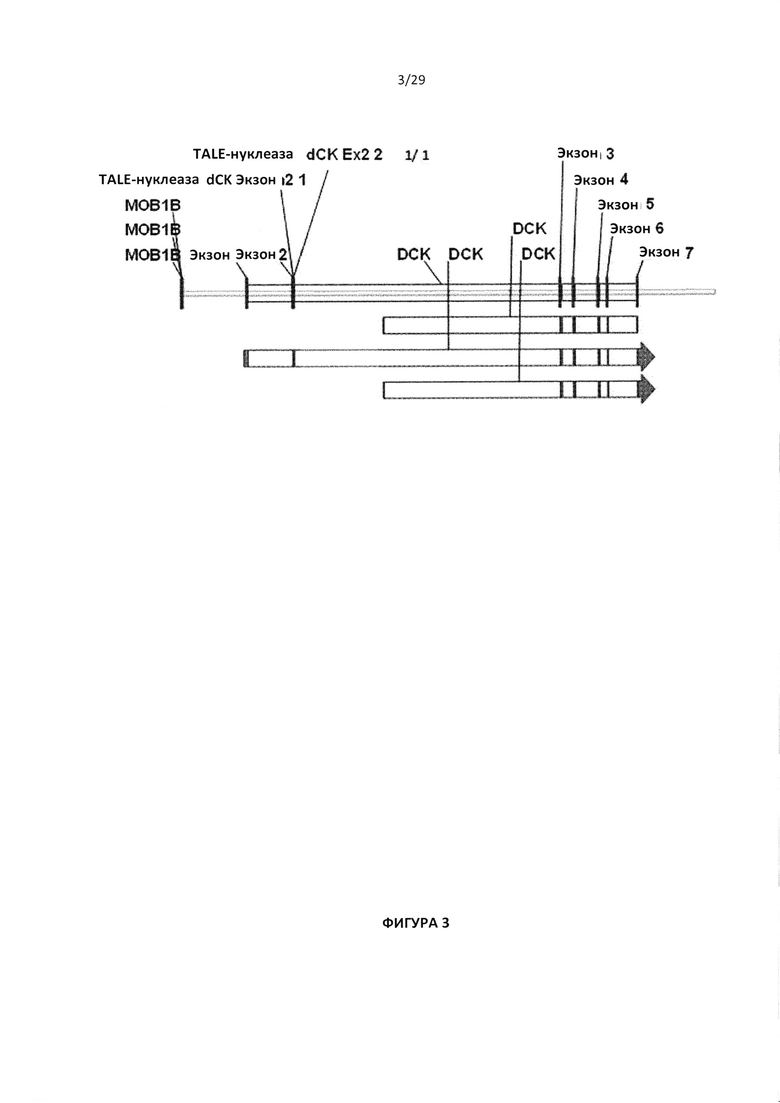

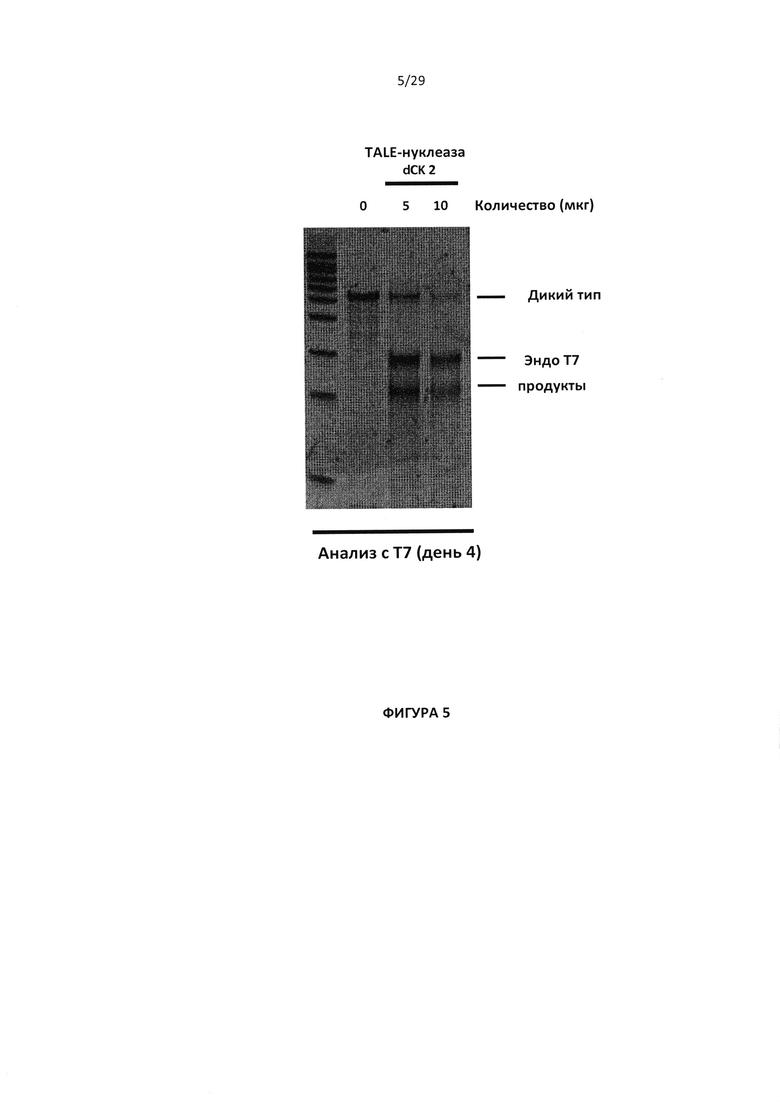
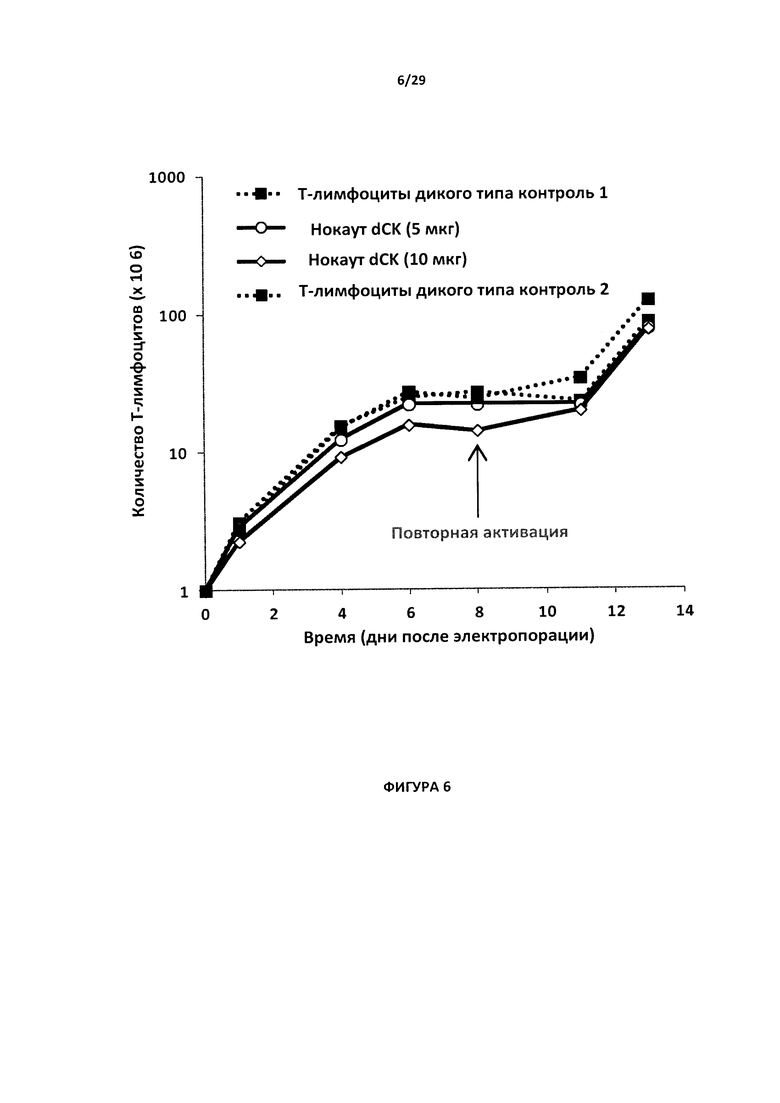
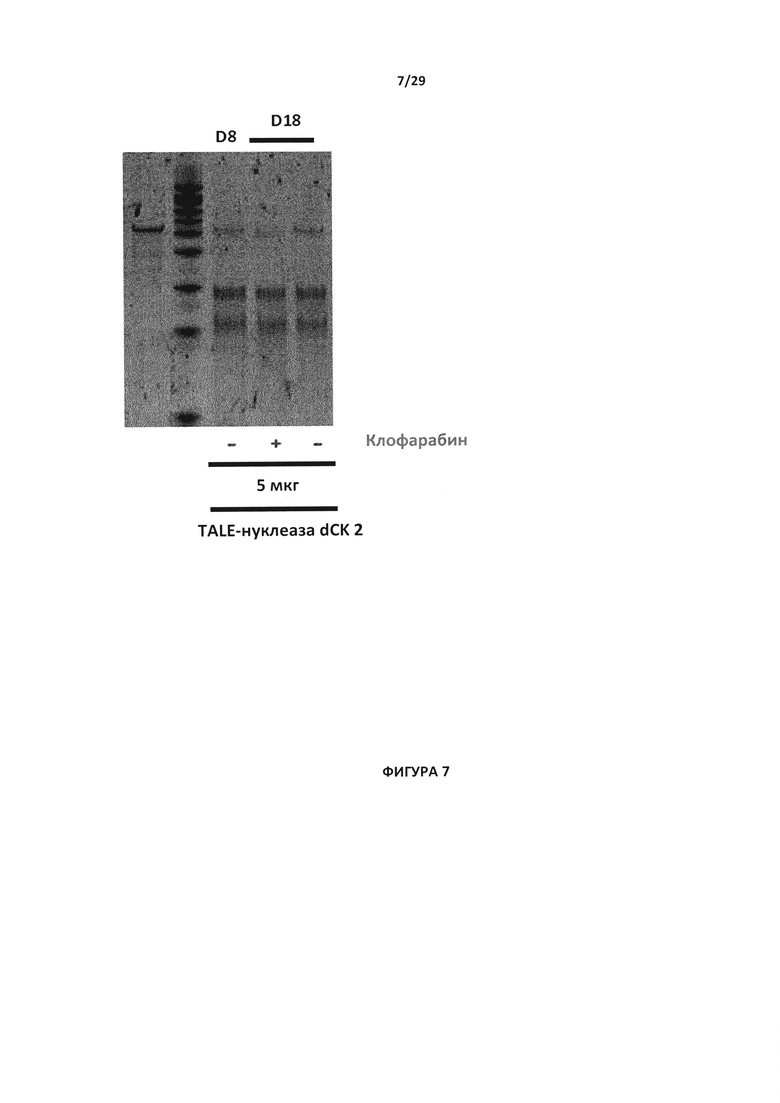
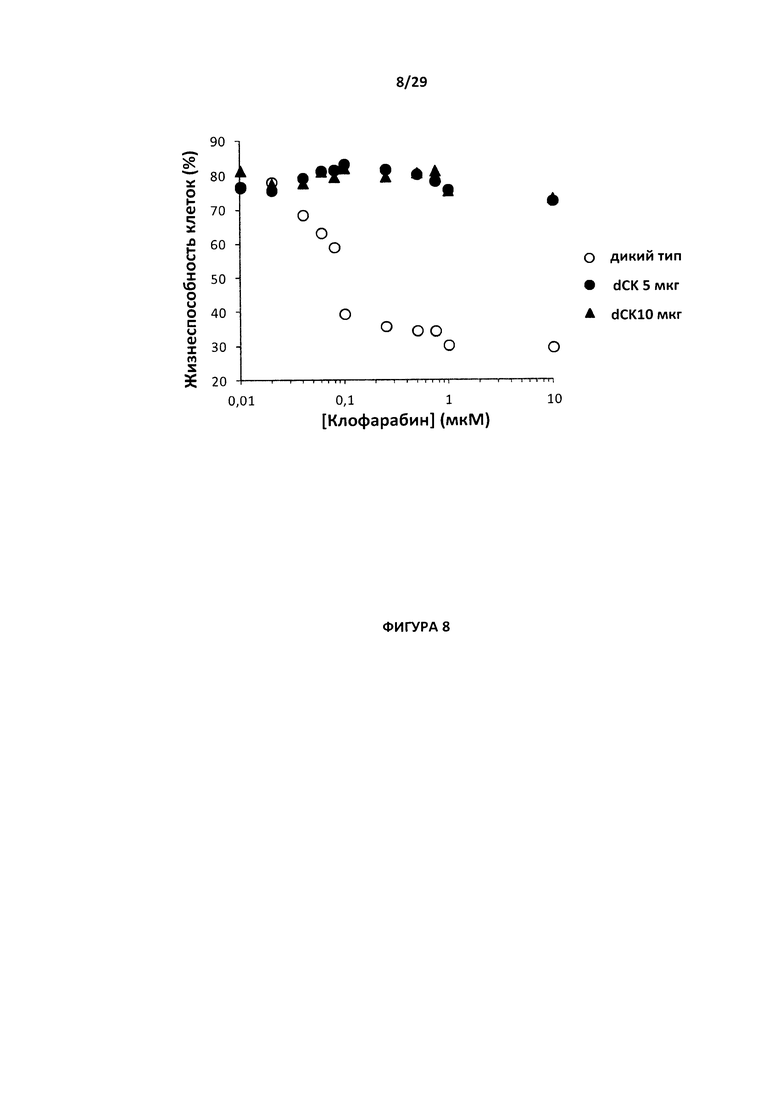

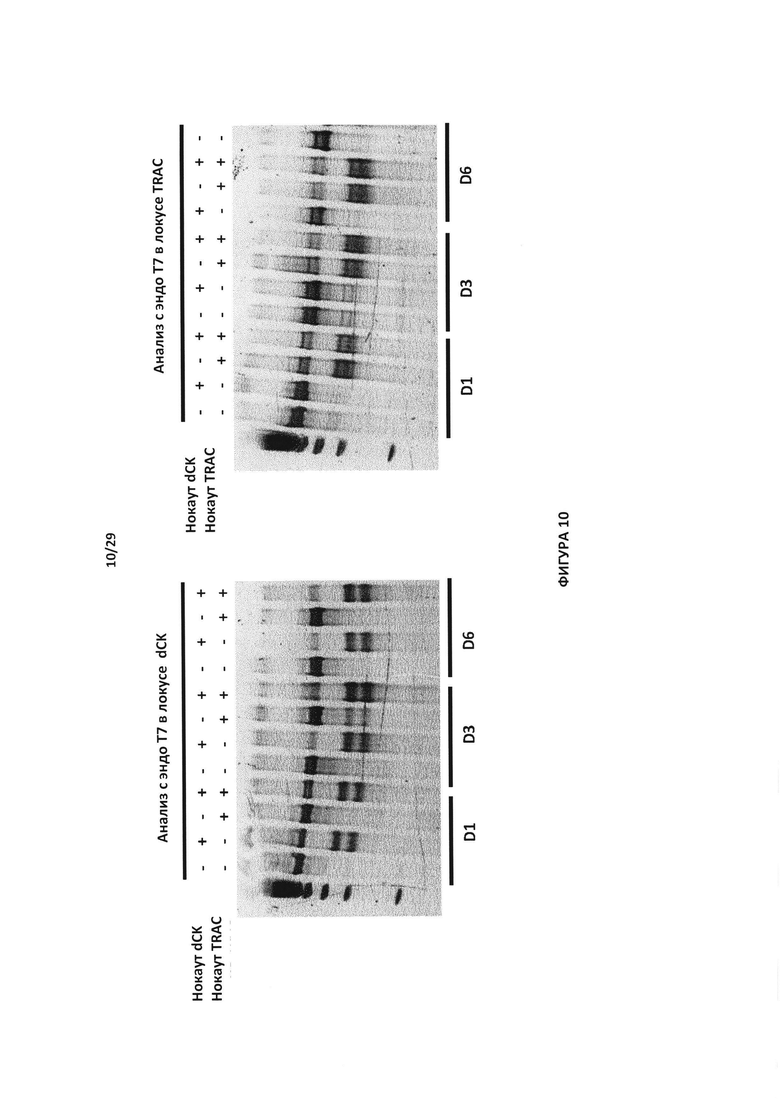
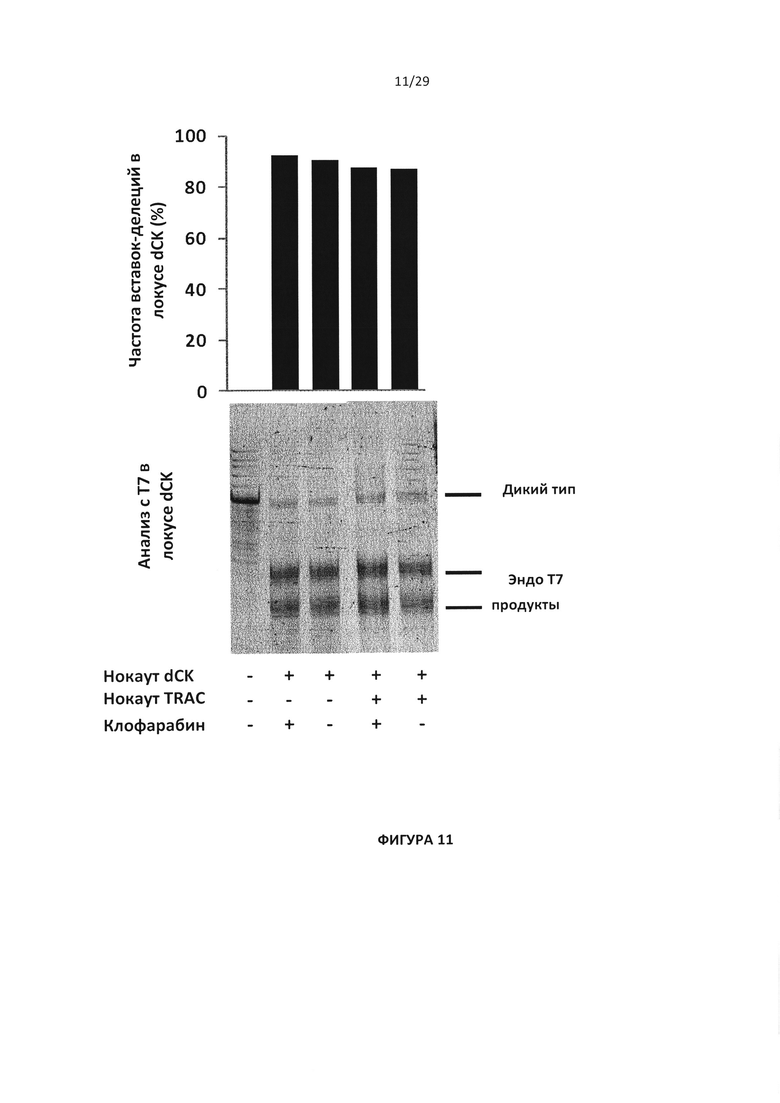
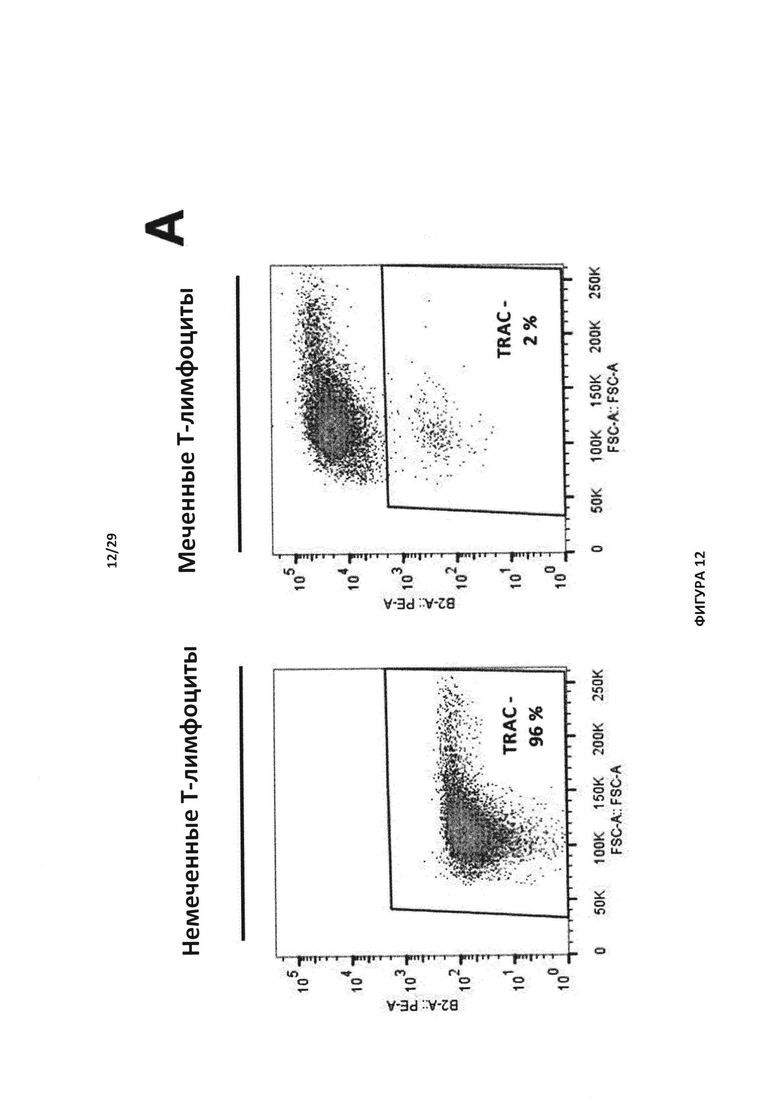
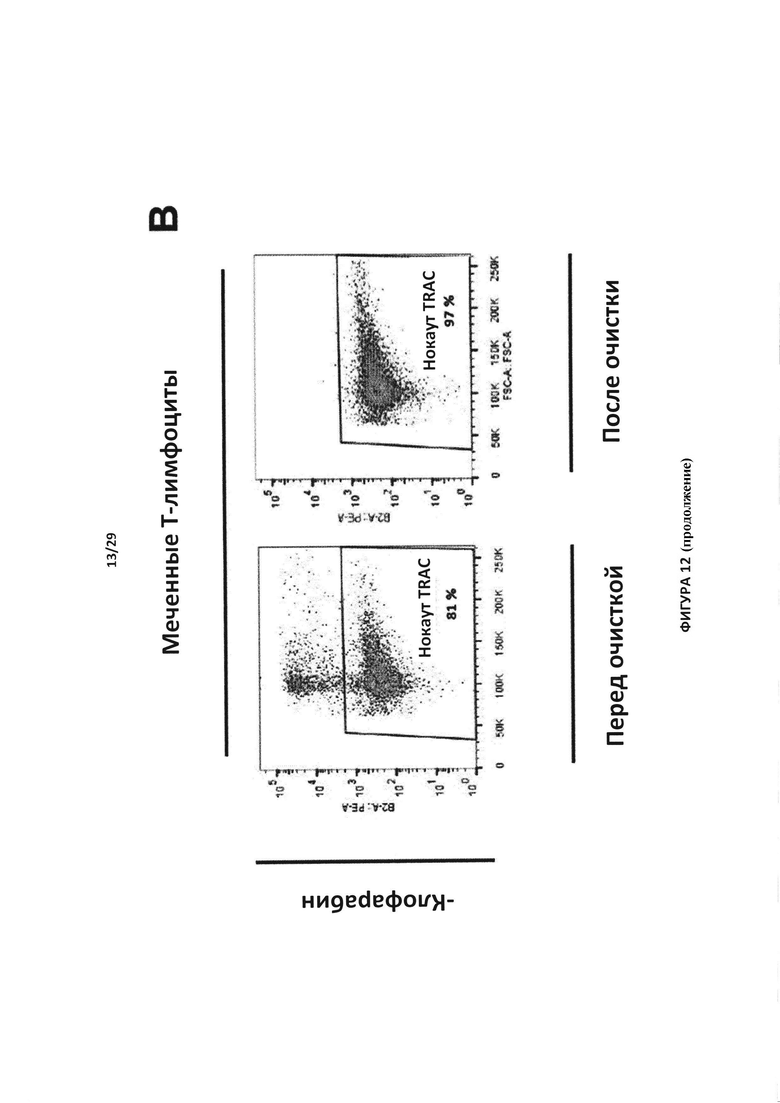
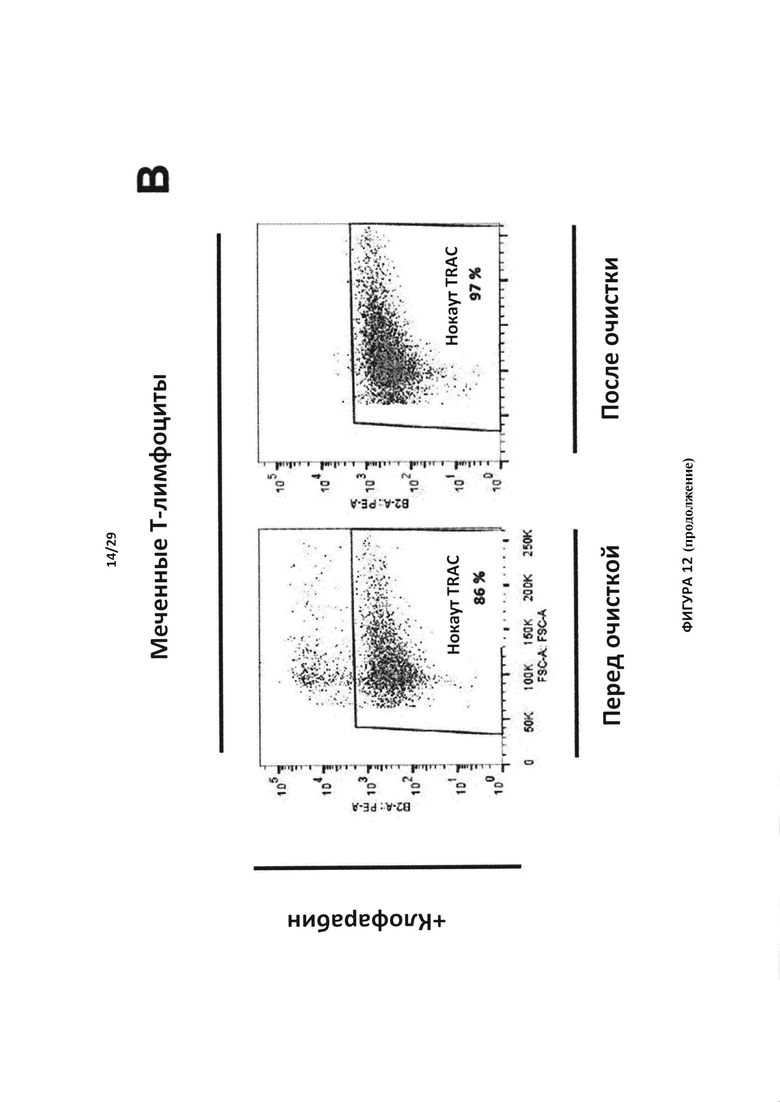
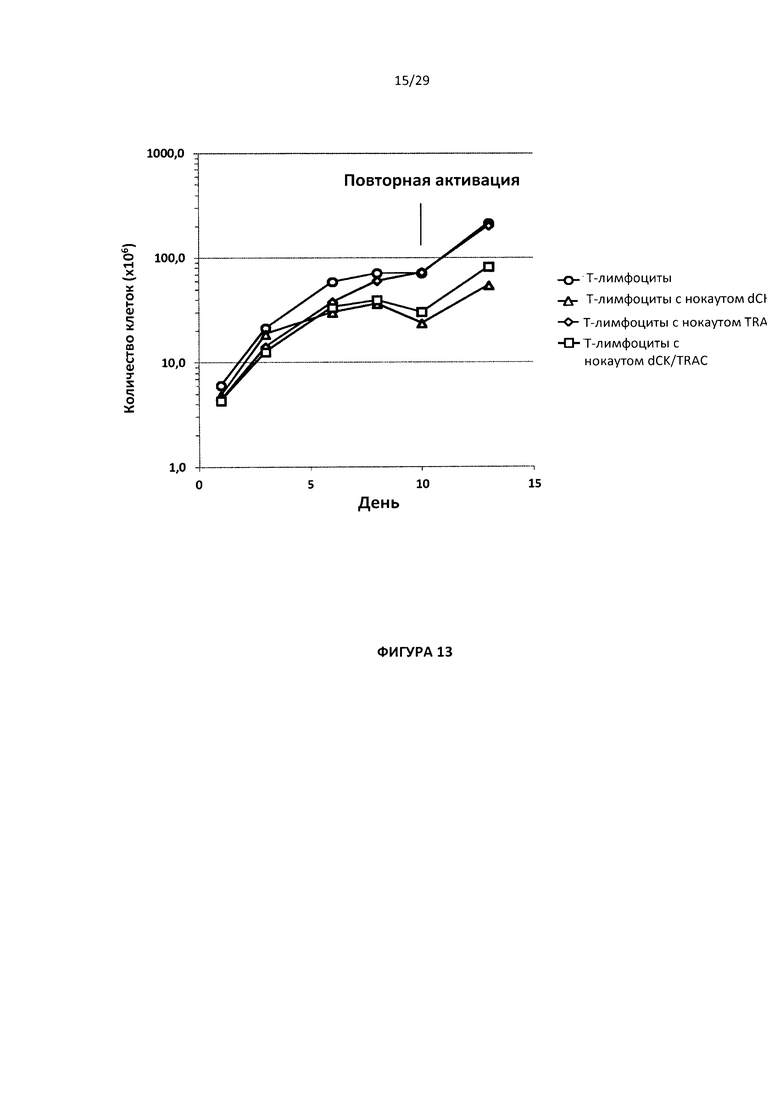
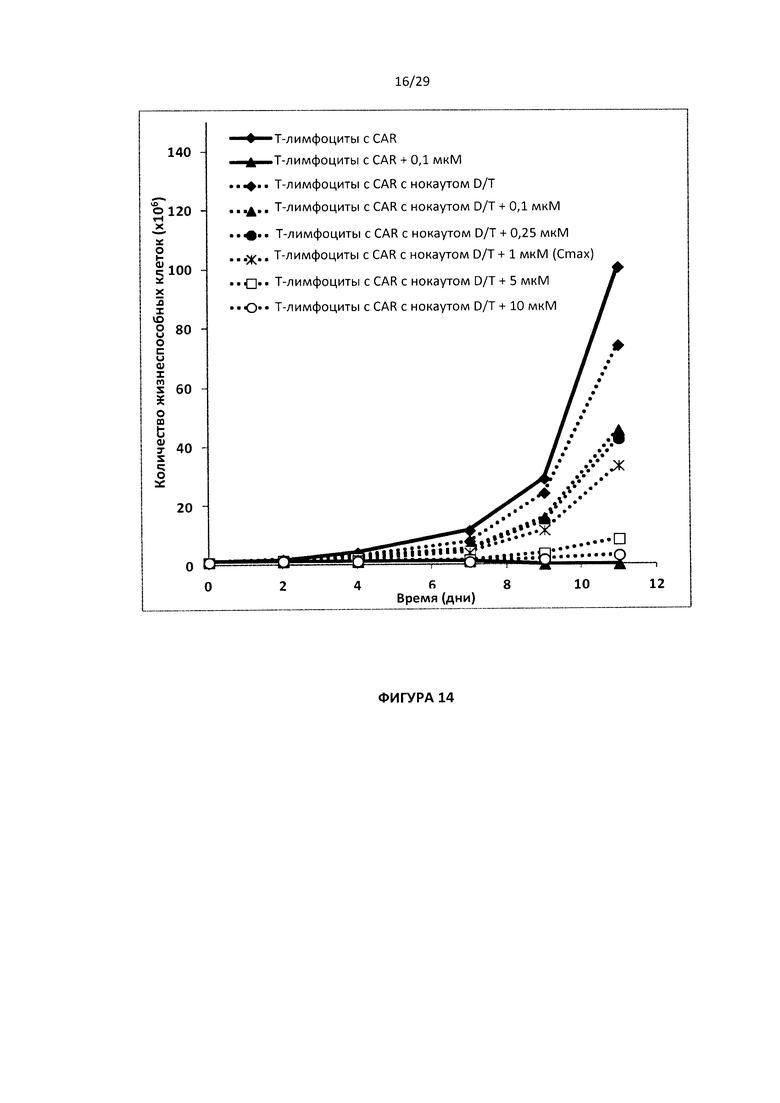
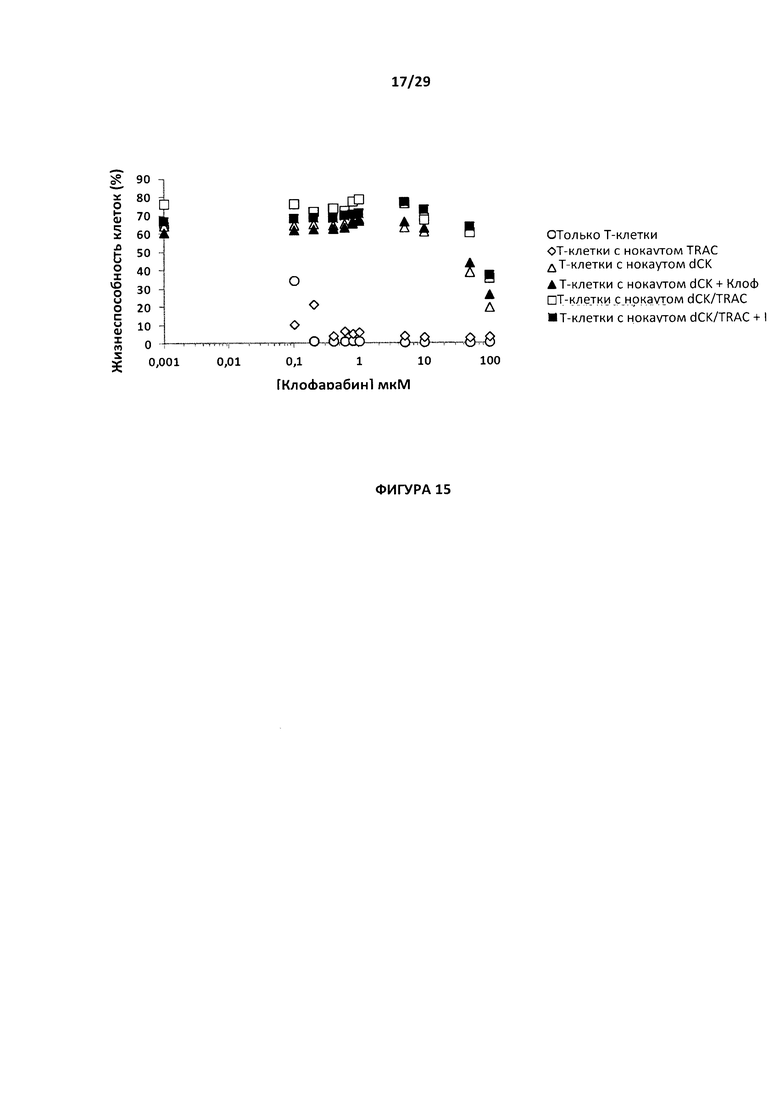
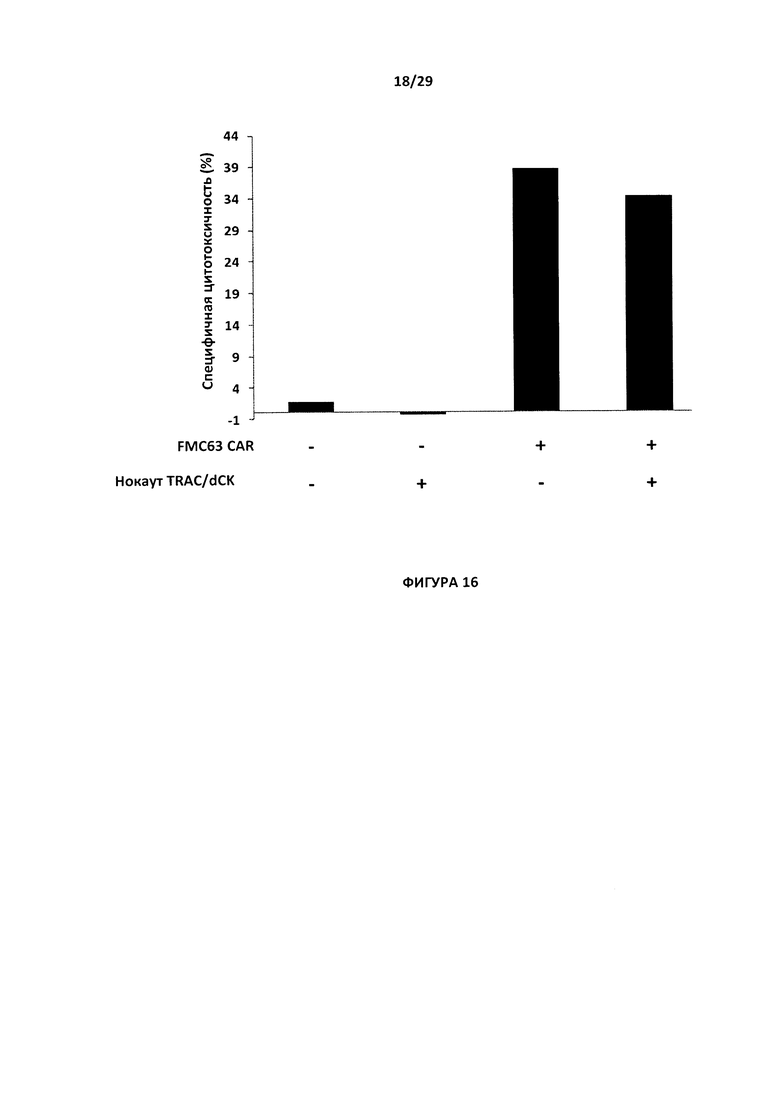
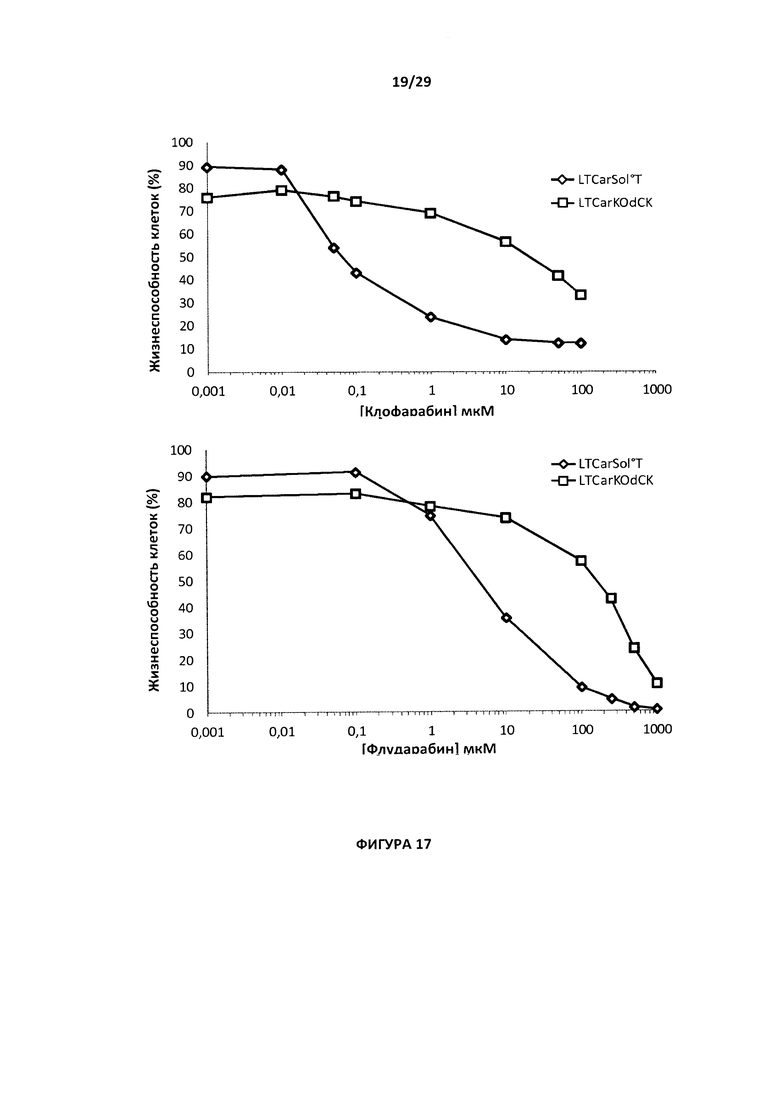



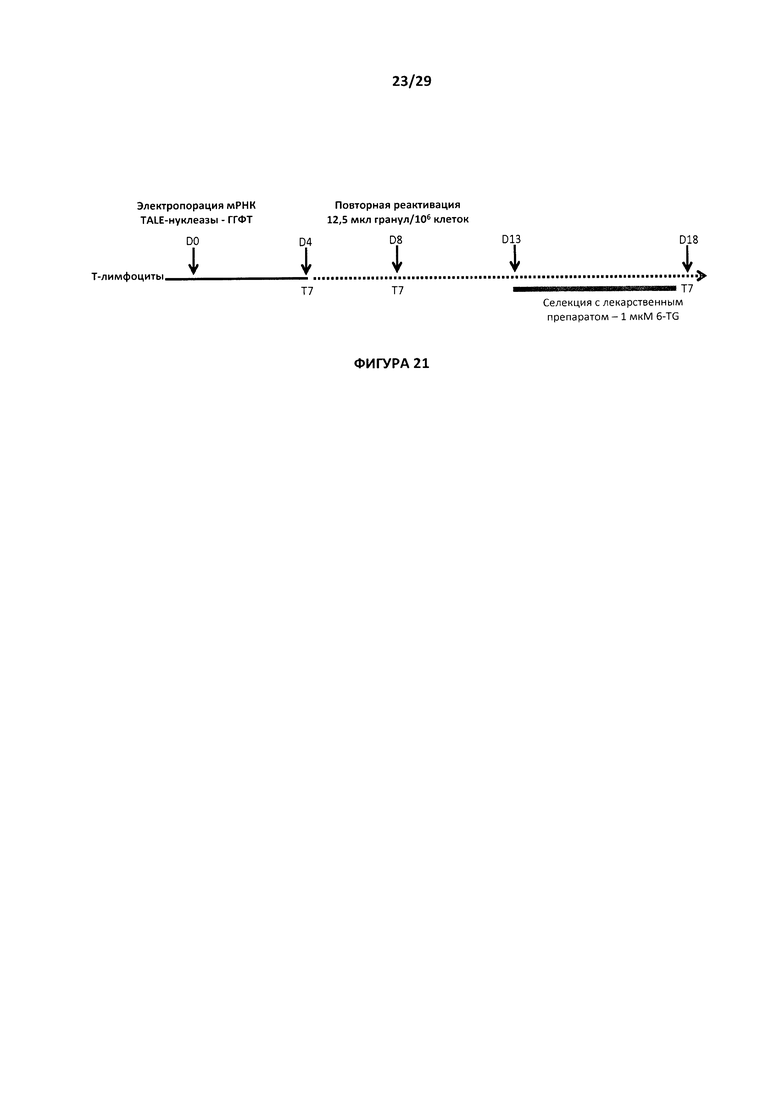
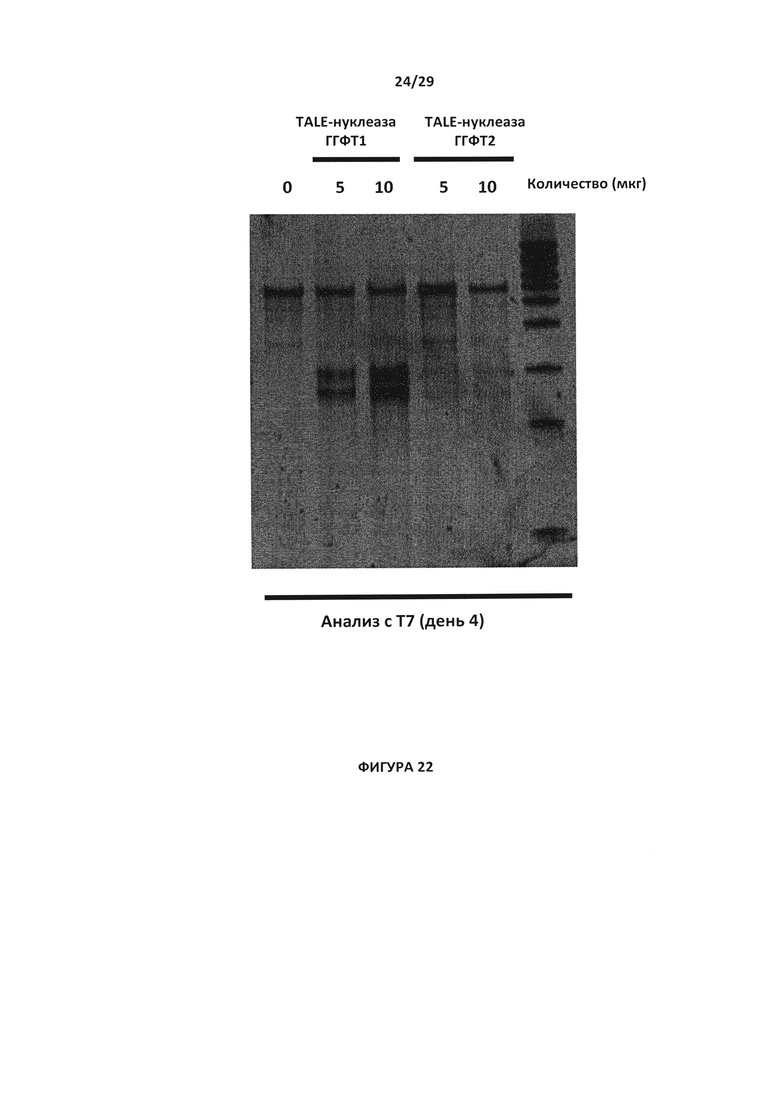
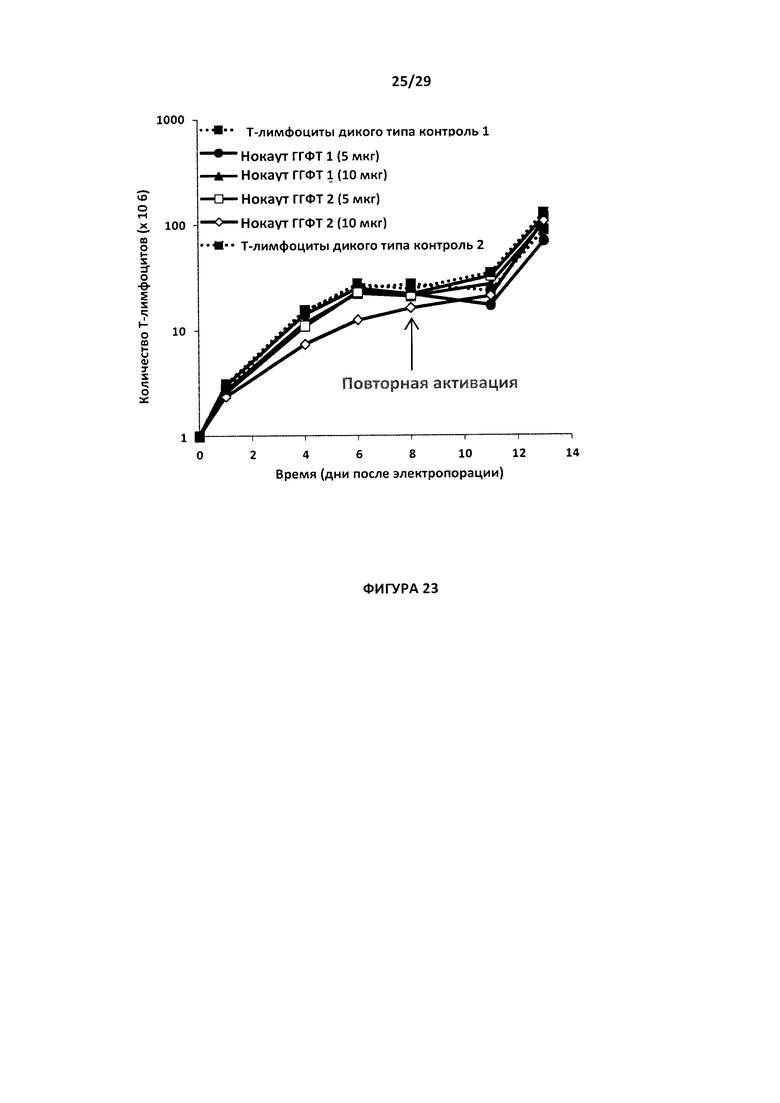
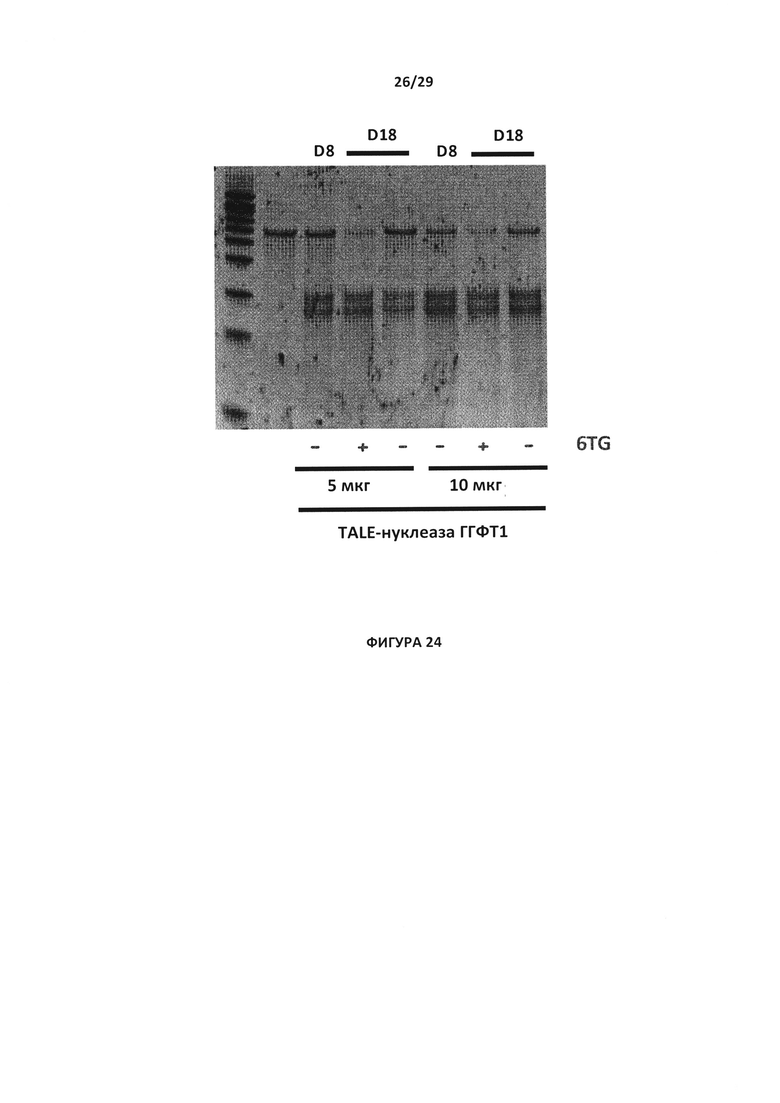
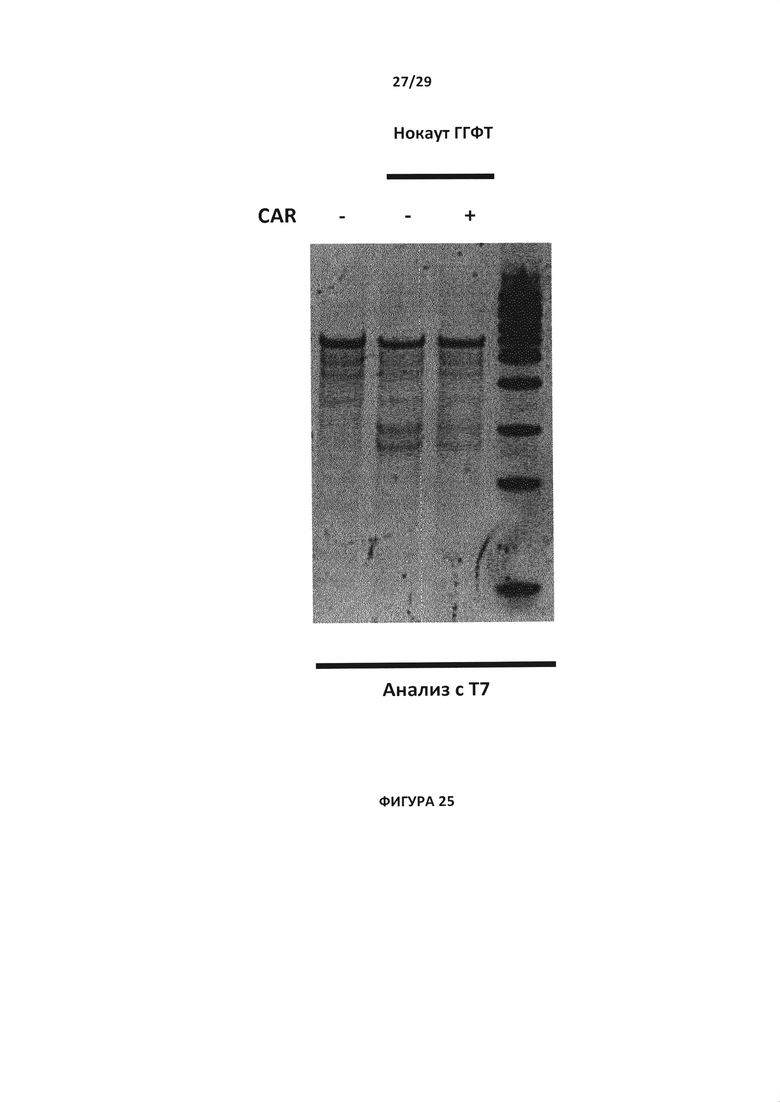
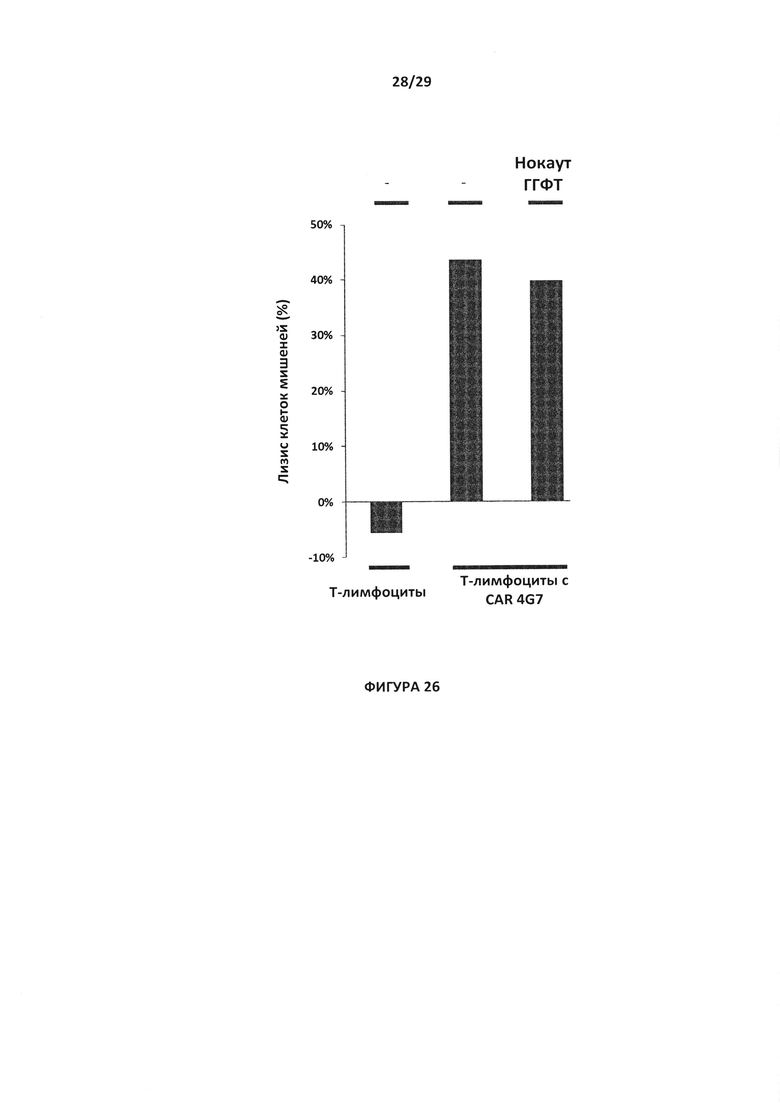
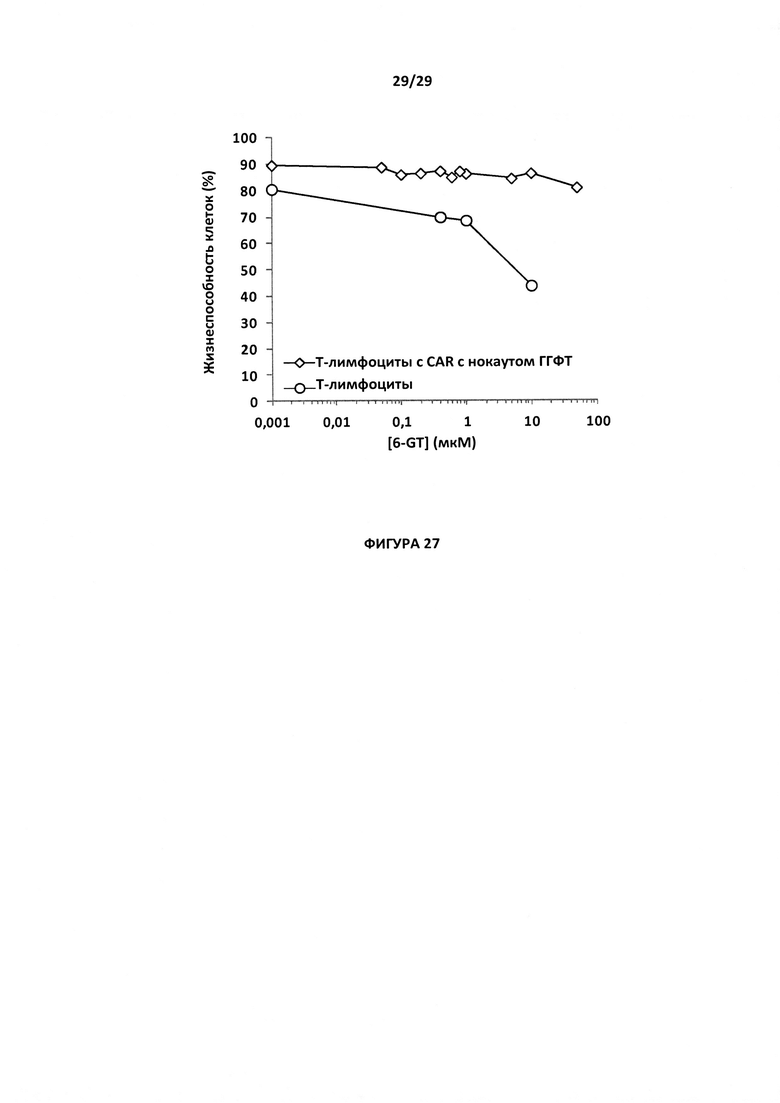
Настоящее изобретение относится к области биотехнологии, конкретно к клеточным технологиям, и может быть применено в медицине для иммунотерапии рака. Способ ex-vivo получения Т-клеток, устойчивых к клофарабину и/или флударабину, включает стадии: (а) трансфекции Т-клеток нуклеотидной последовательностью, кодирующей редкощепящую эндонуклеазу, специфически направленную на ген, экспрессирующий фермент, обладающий активностью дезоксицитидинкиназы (dcK – EC 2.7.1.74); (b) экспрессии указанной эндонуклеазы в указанных Т-клетках для достижения направленной инактивации указанного гена dcK; (c) размножения указанных сконструированных Т-клеток, полученных на этапе (b), необязательно в присутствии клофарабина и/или флударабина. При этом указанные Т-клетки представляют собой аллогенные Т-клетки от донора, в которых инактивирован ген, кодирующий компонент Т-клеточного рецептора (РТЛ), выбранный из РТЛ альфа и РТЛ бета, или представляют собой аутологичные Т-клетки от пациента, у которого диагностирован рак; и при этом указанные клетки экспрессируют химерный рецептор антигена (CAR), специфичный к раковому антигену. Изобретение позволяет повысить эффективность противоопухолевой иммунотерапии. 8 н. и 10 з.п. ф-лы, 27 ил., 4 пр.
1. Способ ex-vivo получения Т-клеток для применения при терапии рака, при этом указанные клетки устойчивы к клофарабину и/или флударабину, включающий стадии:
(а) осуществления трансфекции Т-клеток нуклеотидной последовательностью, кодирующей редкощепящую эндонуклеазу, специфически направленную на ген, экспрессирующий фермент, обладающий активностью дезоксицитидинкиназы (dcK – EC 2.7.1.74);
(b) осуществления экспрессии указанной эндонуклеазы в указанных Т-клетках для достижения направленной инактивации указанного гена dcK;
(c) осуществления размножения указанных сконструированных Т-клеток, полученных на этапе (b), необязательно в присутствии клофарабина и/или флударабина,
при этом указанные Т-клетки представляют собой аллогенные Т-клетки от донора, в которых инактивирован ген, кодирующий компонент Т-клеточного рецептора (РТЛ), выбранный из РТЛ альфа и РТЛ бета, или представляют собой аутологичные Т-клетки от пациента, у которого диагностирован рак; и при этом указанные клетки экспрессируют химерный рецептор антигена (CAR), специфичный к раковому антигену.
2. Способ по п. 1, отличающийся тем, что указанные Т-клетки представляют собой первичные клетки.
3. Способ по п. 1, отличающийся тем, что указанные Т-клетки представляют собой CD8+-клетки.
4. Способ по п. 1, отличающийся тем, что указанные Т-клетки представляют собой TIL (лимфоциты, инфильтрующие опухоль).
5. Способ по п. 1, отличающийся тем, что указанные Т-клетки происходят из организма пациента, которому диагностировали рак.
6. Способ по п. 1, отличающийся тем, что указанные Т-клетки происходят из организма донора.
7. Способ по п. 1, отличающийся тем, что указанная редкощепящая эндонуклеаза представляет собой TALE-нуклеазу.
8. Способ по п. 7, отличающийся тем, что инактивацию гена dСК TALE-нуклеазами проводят с применением TALE-нуклеаз с последовательностью SEQ ID №63 или SEQ ID №64 и целевая последовательность dCK представляет собой SEQ ID №62.
9. Способ по п. 1, отличающийся тем, что указанный химерный рецептор антигена представляет собой CD19+ или CD123+.
10. Выделенная аутологичная Т-клетка для применения при иммунотерапии рака, полученная посредством способа по любому из пп. 1 - 9, при этом указанная клетка устойчива к клофарабину и/или флударабину и в указанной клетке ген лекарственной чувствительности dCK инактивирован при помощи редкощепящей эндонуклеазы.
11. Выделенная аллогенная Т-клетка для применения при иммунотерапии рака, полученная посредством способа по любому из пп. 1 – 9, при этом указанная клетка устойчива к клофарабину и/или флударабину, и в указанной клетке ген лекарственной чувствительности dCK инактивирован при помощи редкощепящей эндонуклеазы.
12. Фармацевтическая композиция для применения при лечении рака, содержащая эффективное количество Т-клеток по любому из пп. 10 или 11.
13. Способ лечения пациента, у которого диагностирован рак, включающий введение эффективного количества Т-клеток по любому из пп. 10, 11 указанному пациенту.
14. Способ по п. 13, отличающийся тем, что указанный рак представляет собой острый лимфобластный лейкоз (ОЛЛ) или амиотрофический миелоидный лейкоз (АМЛ).
15. Способ лечения пациента, у которого диагностирован рак, включающий введение эффективного количества Т-клеток по любому из пп. 10 или 11 указанному пациенту, при этом указанные Т-клетки вводят в сочетании с введением лекарственного средства, представляющего собой аналог пурина.
16. Способ по п. 15, отличающийся тем, что указанный рак представляет собой острый лимфобластный лейкоз (ОЛЛ) или амиотрофический миелоидный лейкоз (АМЛ).
17. Популяция аутологичных Т-клеток для применения при иммунотерапии рака, состоящая из Т-клеток по п. 10.
18. Популяция аллогенных Т- клеток для применения при иммунотерапии рака, состоящая из Т-клеток по п. 11.
| FALTA M.T | |||
| ET AL., Azathioprine Associated T-Cell Mutations in Insulin-Dependent Diabetes Mellitus, Scandinavian Journal of Immunology, 2000, v.51, n.6, p.626 - 633 | |||
| JENSEN M.C | |||
| ET AL., Antitransgene Rejection Responses Contribute to Attenuated Persistence of Adoptively Transferred CD20/CD19-Specific Chimeric Antigen Receptor Redirected T Cells in Humans, BIOLOGY OF BLOOD AND MARROW TRANSPLANTATION, 2010, v | |||
| Устройство для электрической сигнализации | 1918 |
|
SU16A1 |
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| POIROT L | |||
| ET AL., T-Cell Engineering For "off-The-shelf" Adoptive Immunotherapy, Blood Journal, 2013, v.122, Iss | |||
| Выбрасывающий ячеистый аппарат для рядовых сеялок | 1922 |
|
SU21A1 |
| MANSSON E.ET AL., Down-regulation of deoxycytidine kinase in human leukemic cell lines resistant to cladribine and clofarabine and increased ribonucleotide reductase activity contributes to fludarabine resistance, BIOCHEMICAL PHARMACOLOGY, 2003, v.65, n.2, p.237 - 247 | |||
| ЛЕЧЕНИЕ РАКА, КОМБИНИРУЮЩЕЕ ЛИМФОИСТОЩАЮЩЕЕ ВЕЩЕСТВО С ЦТЛ И ЦИТОКИНАМИ | 2007 |
|
RU2447900C2 |

