Представленное изобретение относится к новой β-галактозидазе с трансгалактозилирующей активностью, способной преобразовывать лактозу в смесь галактоолигосахаридов. В частности, оно относится к β-галактозидазе, выделенной из недавно открытого штамма Bifidobacterium bifidum.
Изобретение, в частности, относится к последовательностям ДНК, кодирующим выделенный новый фермент β-галактозидазы, к ферменту, кодируемому такими последовательностями ДНК, и к клеткам-хозяевам, содержащим последовательности ДНК или включающим рекомбинантный вектор, в состав которого входят такие последовательности ДНК. Изобретение также относится к применению фермента, кодируемого последовательностью ДНК, или клетки-хозяина, содержащей последовательность ДНК или рекомбинантный вектор, к производству галактоолигосахаридов.
В естественном состоянии бифидобактерии поселяются в нижних отделах желудочно-кишечного тракта, которые являются средой, бедной моно- и дисахаридами, поскольку эти сахара предпочтительно потребляются организмом-хозяином и микробами, присутствующими в верхних отделах желудочно-кишечного тракта. Для того, чтобы выжить в нижнем желудочно-кишечном тракте, бифидобактерии продуцируют различные виды экзо- и эндогликозидаз в поверхностносвязанной и/или внеклеточной форме, посредством которых они могут использовать разнообразные углеводы.
Помимо гидролазной активности, некоторые ферменты бифидобактерий демонстрируют трансферазную активность. Эта трансгликозилирующая активность гликозидаз широко применяется для ферментативного синтеза различных олигосахаридов, относительно которых было доказано, что они выступают в качестве стимуляторов роста бифидобактерий.
Известно, что представители рода бифидобактерий продуцируют ферменты β-галактозидазы, которые вовлечены в процесс бактериального метаболизма лактозы. Меллер и соавт. (Møller, P.L. et al. in Appl & Environ. Microbial, (2001), 62, (5), 2276-2283) описали выделение и исследование трех генов β-галактозидазы из штамма Bifidobacterium bifidum. Они обнаружили, что все три β-галактозидазы были способны катализировать образование бета-связанных галактоолигосахаридов посредством трансгалактозилирования.
Дюмортье и соавт. (Dumortier et al. in Carbohydrate Research. 201. (1990), 115-123) описали образование бета-связанных галактоолигосахаридов посредством реакции трансгалактозилирования во время гидролиза лактозы штаммом Bifidobacterium bifidum DSM 20456. Проведенный ими анализ структуры смеси продуцированных олигосахаридов показал, что имели место связи β-(1→3), β-(1→6) и β-(1→4)-D-галактозил. Дюмортье предположил, что соединения, продуцируемые Bifidobacterium bifidum, вовлечены в адгезию бактерии в толстом кишечнике.
В документе WO 01/90317 описана новая β-галактозидаза из Bifidobacterium bifidum, а именно укороченная версия фермента, которая обладает высокой трансгалактозилирующей активностью.
Был обнаружен штамм Bifidobacterium bifidum, способный к продуцированию фермента галактозидазы, обладающего активностью в преобразовании лактозы в новую смесь галактоолигосахаридов, которая неожиданно содержала вплоть до 35% дисахаридов, включая галабиозу (Gal (α 1-6) - Gal). Известно (см. Paton, J C and Paton, A W (1998), Clin. Microbiol. Revs., 11, 450-479; Carlsson, K A (1989), Ann. Reviews Biochem., 58, 309-350), что этот дисахарид обладает антиадгезионными свойствами и может препятствовать адгезии токсинов, например, токсина Шига, и патогенных микроорганизмов, таких как E.coli, к стенке кишки.
Данный штамм B.bifidum был депонирован под инвентарным номером NCIMB 41171 в Национальную коллекцию промышленных и морских бактерий, Абердин, Великобритания, 31 марта 2003. Он также описывается в патенте США №2412380.
На данный момент обнаружено, что данный штамм B.bifidum продуцирует несколько β-галактозидаз, включая новую β-галактозидазу. Данный фермент продуцирует ряд различных олигосахаридов, которые являются β-связанными.
В соответствии с изобретением, представлена последовательность ДНК, кодирующая белок с аминокислотной последовательностью, соответствующей представленной в SEQ ID NO: 2, или гибридизирующаяся в жестких условиях с последовательностью ДНК, кодирующей данный белок. Последовательность ДНК представлена в SEQ ID NO: 1 или может содержать фрагмент этой последовательности или вырожденную последовательность, полученную из нее.
Термин "вырожденная" подразумевает последовательность ДНК, обладающую по меньшей мере 50% гомологией в SEQ ID NO: 1, предпочтительно гомологией на уровне от 50 до 98%, и наиболее предпочтительно гомологией на уровне от 75 до 95%.
Такая последовательность ДНК может содержать нуклеотидные замены, вставки и удаления, результатом которых является изменение менее 60%, предпочтительно менее 45%, и наиболее предпочтительно - менее 25% аминокислотной последовательности, приведенной в SEQ ID NO: 2. Следствием нуклеотидных замен могут быть консервативные аминокислотные замены.
В соответствии со вторым аспектом изобретения, представлен фермент, кодируемый последовательностью ДНК, как определено выше. Данный фермент может содержать аминокислотную последовательность, представленную в SEQ ID NO: 2, или ее фрагмент.
В соответствии с третьим аспектом изобретения, представлен рекомбинантный вектор, предпочтительно, экспрессионный вектор, содержащий последовательность ДНК, как определено выше. Данный вектор может быть встроен в клетку хозяина, например, клетку бактерий, дрожжей или грибов. В качестве альтернативы, последовательность ДНК может быть встроена в подобную клетку хозяина. Подходящая клетка-хозяин может быть выбрана из группы, включающей Bifidobacterium, Lactococcus, Lactobacillus, Bacillus, например, Bacillus subtilus или Bacillus circulans, Escherichia и Aspergillus, например, Aspergillus niger.
Используя лактозу в качестве субстрата, фермент, кодируемый последовательностью ДНК, определенной выше, продуцирует смесь олигосахаридов, содержащую дисахариды, такие как Gal(β1-3)Glc, Gal(β1-3)Gal, Gal(β1-6)Gal и Gal(α1-6)Gal, трисахариды и тетрасахариды, такие как Gal(β1-6)Gal(β1-4)Glc, Gal(β1-3)Gal(β1-4)Glc и Gal(β1-6)Gal(β1-6)Gal(β1-4)Glc.
Фермент или клетка-хозяин, описанные выше, могут применяться для продуцирования смеси галактоолигосахаридов, которые могут служить составной частью продуктов, улучшающих состояние кишечника. Такие продукты могут быть выбраны из группы, состоящей из молочных продуктов (например, жидкое молоко, порошковое молоко, такое как цельное сухое молоко, обезжиренное сухое молоко, сухое молоко, насыщенное жирами, сухая молочная сыворотка, молоко для детского питания, смесь для детского питания, мороженое, йогурт, сыр, ферментированные молочные продукты), напитков, таких как фруктовый сок, детского питания, каши, хлеба, печенья, кондитерских изделий, пирожных, пищевых добавок, диетических добавок, кормов для животных, кормов для птицы или фактически из любых других продуктов питания или напитков. Присутствие галактоолигосахаридов в таких продуктах дает преимущество в виде усиления роста полезных для здоровья бифидобактерий в продуктах или в кишечной флоре потребителя после приема, или и в том, и в другом.
В качестве альтернативы, олигосахариды, продуцированные таким способом, могут применяться для приготовления медикаментов, например, в форме таблеток или капсул, для предотвращения адгезии патогенных микроорганизмов или токсинов, продуцируемых патогенными микроорганизмами, к стенке кишечника. Медикамент может вводиться пациенту, например, после курса лечения антибиотиками, который часто изменяет или даже уничтожает нормальную здоровую флору кишечника.
В соответствии с еще одним аспектом изобретения, представлен способ продуцирования вышеописанного фермента, который включает культивирование клеток-хозяев, определенных выше, в подходящей культуральной питательной среде в условиях, допускающих экспрессию фермента, и выделение полученного в результате фермента или ферментных продуктов из культуры.
Изобретение также ориентировано на способ продуцирования смеси галактоолигосахаридов, который включает соединение вышеопределенного фермента с материалом, содержащим лактозу, в условиях, приводящих к образованию смеси галактоолигосахаридов.
Подходящий материал, содержащий лактозу, может быть выбран из коммерчески доступной лактозы, цельного молока, полуобезжиренного молока, обезжиренного молока, молочной сыворотки, насыщенного жирами молока и фильтратов молочной сыворотки. Данные молочные продукты могут быть получены от коров, буйволов, овец или коз. Насыщенное жирами молоко определено как цельное молоко, подвергнутое обезжириванию с целью удаления молочных жиров, которые затем заменяются путем добавления растительных жиров или растительного масла.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На Фиг.1 показана нуклеотидная последовательность (SEQ ID NO: 1) β-галактозидазы Bifidobacterium bifidum, представленной в изобретении.
На Фиг.2 показана аминокислотная последовательность (SEQ ID NO: 2), соответствующая нуклеотидной последовательности, представленной на Фиг.1.
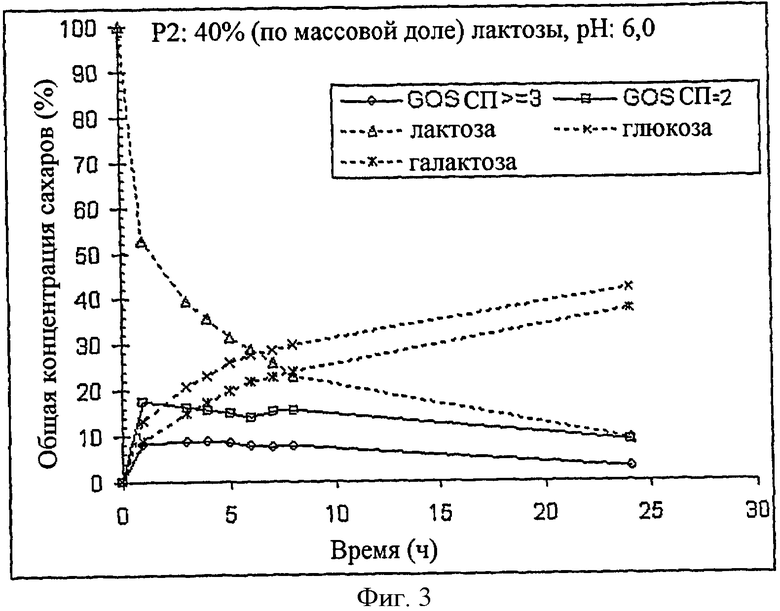
На Фиг.3 представлен график, показывающий, в зависимости от времени, реакцию, происходящую при синтезе галактоолигосахаридов с помощью β-галактозидазы в 40% (по массовой доле) растворе лактозы в 0,1М фосфатном буфере при pH 6,0, используемом в качестве субстрата.
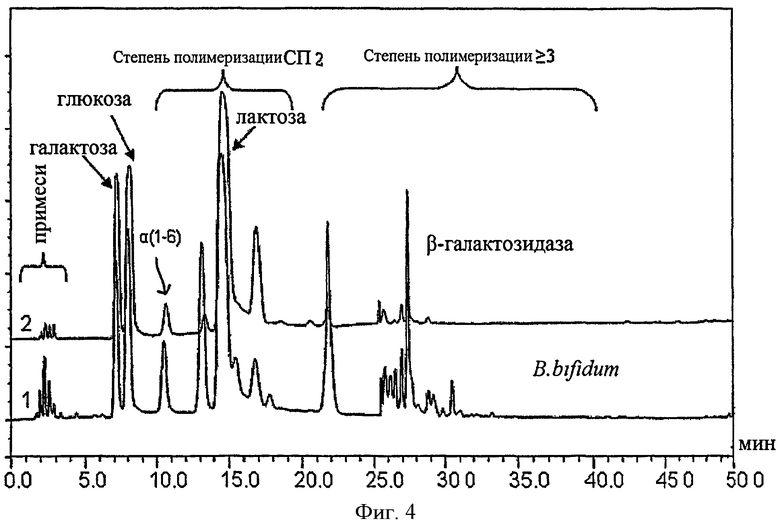
На Фиг.4 показана хроматограмма высокоэффективной анионообменной хроматографии смеси галактоолигосахаридов, синтезированных с помощью β-галактозидазы из B.bifidum NCIMB 41171 с использованием 40% (по массовой доле) раствора лактозы в 0,1М фосфатном буфере при pH 6,0 в качестве субстрата (Glc = глюкоза, Gal = галактоза, Lac = лактоза, α (1-6) = галактобиоза, DP = степень полимеризации).
Геномная ДНК была выделена из штамма Bifidobacterium bifidum (NCIMB 41171) с использованием метода Лоусона и соавт. (Lawson et al. (1989) Fems Microbiol Letters, 65, (1-2), 41-45). ДНК была расщеплена посредством рестрикционных ферментов, и фрагменты, максимальный размер которых составлял 15 т.п.н., лигировались с вектором pSP72, расщепленным посредством тех же рестрикционных ферментов. Клетки E.coli были трансформированы вектором, содержащим вставки, состоящие из хромосомной ДНК B.Bifidum, расщепленной посредством PstI, Eco RI, Bam HI, KpnI, SmaI или HindIII. Клоны, обладающие β-галактозидазной активностью, были отобраны на чашках с агаровой средой Луриа-Бертани, содержащей p-нитрофенил, X-β-Gal (5-бромо-4-хлоро-3-индолил-β-D-галактозид) и изопропил-β-D-тиогалактозид (IPTG). Смеси для лигирования хромосомной ДНК с Pst I послужили источником тринадцати положительных клонов β-галактозидазы, один из которых был идентифицирован как pP2.
Секвенирование вставленного фрагмента ДНК P2 проводилось с применением метода дидезокси-секвенирования (обрыва цепи) по Сангеру (Russel P., 2002 iGenetics, Pearson Education, Inc., San Francisco, 187-189) с использование набора для циклического секвенирования BigDye Terminator V.3.0 (Applied Biosystems, США). Последовательность ДНК P2 показана на Фиг.1 (SEQ ID NO: 1).
Открытая рамка считывания (ОРС) была обнаружена с использованием программы поиска ОРС с сайта NCBI (Национальный центр биотехнологической информации). Нуклеотидная последовательность, показанная на Фиг.1, транслировалась с помощью всех шести возможных рамок считывания, при этом была идентифицирована одна открытая рамка считывания, с которой транслировались 738 аминокислот, предположительно кодирующих β-галактозидазу. Результат трансляции показан на Фиг.2 (SEQ ID NO: 2).
Далее представленное изобретение будет описано посредством обращения к следующему примеру.
Пример 1
Материалы и Методы
Все реактивы и заготовки сред, использованные в данном исследовании, были получены от компаний Sigma (Дорсет, Великобритания), Invitrogen (Пейсли, Великобритания), Oxoid (Бейсигсток, Великобритания), Qiagen (Западный Суссекс, Великобритания) и Promega (Саутгемптон, Великобритания).
Штаммы бактерий
Штамм Bifidobacterium bifidum (NCIMB 41171) содержался в криогенных гранулах в трубках Microbank при -70°C. Для последующих экспериментов, штамм был восстановлен на агаре Уилкинсона-Чалгрена (Oxoid, Великобритания) и TPY-среде (среда с дрожжевым экстрактом фитона триптиказы) и выращивался в анаэробных условиях (содержание CO2 и N2 - 80% и 20% соответственно) при 37°C в течение 48 часов. Морфология колонии и отсутствие загрязнений проверялось окрашиванием по методу Грама.
Штаммы E.coli
Штамм Escherichia coli DH5a, использованный в данном исследовании, обычно инкубировался в аэробных условиях при 37°C на агаре или в питательной среде Луриа-Бертани (Sambrook J. и Russell W.D. (2001). Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, New York), при этом в случае необходимости добавлялись антибиотики (100 мкг/мл ампициллина и/или 15 мкг/мл хлоромицетина) и 40 мкл 2% X-β-Gal, 7 мкл 20% IPTG (изопропил-β-D-тиогалактозида), которые наносились на поверхность предварительно подготовленной 90 мм чашки с агаровой средой.
Штамм DH5a E.coli (Invitrogen, Пейсли, Великобритания) (генотип: F-φ80lacZΔMΔ(lacZYA-argF)U169 recA1 endA1 hsdR17(rk -, mk -)phoA supE44 thi-1 gyrA96 relA1λ-) является положительным штаммом для α-галактозидазы, и применялся в экспериментах по экспрессии и для других генетических манипуляций.
Извлечение геномной ДНК из Bifidobacterium bifidum
Геномная ДНК выделялась из штамма Bifidobacterium bifidum (NCIMB 41171) посредством применения следующего метода, в котором хромосомная ДНК была заготовлена из клеточного дебриса, собранного из 100 мл анаэробной питательной среды Уилкинсона-Чалгрена. Клетки ресуспендировались в 10 мл TES-буфера (10 ммоль Tris-HCl, 10 ммоль EDTA, 10 ммоль NaCl, pH=8) и обрабатывались 200 мкл смеси лизоцим/мутанолизин (4:1, лизоцим - 10 мг/мл мутанолизин 1 мг/мл) в течение 30 минут при 37°C. Затем клетки обрабатывались 200 мкл протеиназы K (при 20 мг/мл) и 200 мкл РНКазы (обе - 10 мг/мл) и инкубировались в течение одного часа при 65°C. В конце клетки обрабатывались 2 мл 10%-ного SDS и инкубировались в течение 15 минут при 65°C. Было добавлено 12 мл фенола/хлороформа, и извлечение повторялось до тех пор, пока не стало возможным легко разделить водную фазу и интерфазу. Геномная ДНК осаждалась изопропанолом и ресуспендировалась в 10 ммоль Tris-HCl - 1 ммоль EDTA (pH 8). Затем геномная ДНК расщеплялась ферментами рестрикции, лигировалась с pSP72, расщепленным теми же ферментами, и обрабатывалась щелочной фосфатазой. Расщепление геномной ДНК B.bifidum проводилось посредством EcoRI, PstI, BamHI, SmaI и KpnI. Лигированные смеси применялись для трансформации E.coli DH5a, и клоны, положительные по β-галактозидазе, идентифицировались как голубые колонии на чашках, содержащих X-Gal.
Приготовление векторной ДНК
В данном исследовании клонировался и экспрессировался вектор pSP72 (Promega, Великобритания) (Krieg, P.A. and Melton, D.A. (1987). In vitro RNA synthesis with SP6 RNA polymerase. Methods in Enzymology. 155: 397-415).
Данный вектор был выбран по причине отсутствия комплементарной активности α-фрагмента β-галактозидазы, который не закодирован в pSP72. Данный вектор не несет короткого сегмента ДНК E. coli, содержащего регуляторную последовательность, и кодирующей информации для первых 146 аминокислот β-галактозидазы, которая, в комбинации со штаммами E.coli (например, DH5a), экспрессирующими карбоксил-концевую часть данной β-галактозидазы, образует активную β-галактозидазу (α-комплементация).
Вектор расщеплялся посредством следующих рестрикционных ферментов: PstI, BamHI, HindIII, SmaI, KpnI и EcoRI в соответствии с инструкциями производителя, при десятикратном превышении количества ферментов относительно количества ДНК (единиц ферментов: мкг ДНК, что соответствует десяти единицам фермента на один мкг плазмидной ДНК или десяти единицам фермента на 0,5 пмоль плазмидной ДНК). После теплового инактивирования ферментов (20 мин при 65°C), рестрикционные карты анализировались посредством горизонтального гель-электрофореза. Наличие единственного фрагмента в геле указывало на полное расщепление вектора и его единственное расщепление посредством рестрикции.
Достаточность расщепления вектора также проверялась посредством трансформации нелигированных молекул в компетентные клетки E.coli DH5a. Число образовавшихся колоний на чашках с агаром Луриа-Бертани с добавлением ампициллина (100 мкг/мл) являлось индикатором нерасщепленных молекул и ожидаемого фонового уровня при последовательных экспериментах.
Затем векторы дефосфорилировались посредством щелочной фосфатазы из кишечника телят CIAP (Promega, Саутгемптон, Великобритания) в соответствии с инструкциями производителя. Эффективность обработки проверялась посредством лигирования (с помощью ДНК-лигазы бактериофага T4 в соответствии с инструкциями производителя) после трансформации в клетки DH5a. Число образовавшихся колоний соответствовало числу рециркулированных молекул (неклонированных векторов), и в результате вычитания данного числа из количества образовавшихся колоний без проведения обработки векторов посредством CIAP определялось число векторов, не подвергшихся дефосфорилированию.
Построение библиотеки геномной ДНК
Геномная ДНК подвергалась частичному расщеплению посредством шести ферментов рестрикции, распознающих часто появляющиеся гексануклеотидные последовательности в прокариотической ДНК. EcoRI, BamHI, PstI, KpnI, SmaI и HindIII являются рестрикционными эндонуклеазами типа II, специфически распознающими последовательности 5'G/AATTC'3, 5'G/GATCC'3, 5'CTGCA/G'3, 5'GGTAC/C3', 5'CCC/GGG3' и 5Α/AGCTT3' соответственно и создающими двухцепочечные разрывы в пределах данных последовательностей, генерируя 5'-«залипание» четырех нуклеотидов, AATT, GATC, AGCT в случае EcoRI, BamHI и Hind III соответственно, 3'-«залипания», ACGT и GTAC, в случае PstI и KpnI соответственно, и тупые концы в случае SmaI.
Все эти ферменты были активными и могли расщеплять ДНК только в присутствии двухвалентных ионов магния. Данные ионы были единственным необходимым кофактором.
Рестрикционное расщепление ДНК
Все расщепленные участки образцов геномной ДНК инкубировались в течение 2 часов при 37°C и в конце подвергались тепловому инактивированию при 65°C в течение 20 минут. Затем реагенты охлаждались при комнатной температуре, добавлялось необходимое количество загрузочного буфера, после чего производилось слабое перемешивание герметичной стеклянной капиллярной трубкой. Затем растворы были помещены в резервуар с 0,8% агарозным гелем (источник питания - 4-5 В/см в течение 16 часов), и размер расщепленной ДНК оценивался в соответствии со стандартами 1 т.п.н. ДНК (Promega, Великобритания) (Sambrook J. Molecular Cloning: A Laboratory Manual (2002)).
Очистка фрагментов, произведенных в результате рестрикционного расщепления
Очистка фрагментов из реакционных смесей и агарозных гелей проводилась с использованием набора для извлечения из геля QIAEX производства Qiagen (Западный Суссекс, Великобритания). Протоколы подробно описаны в инструкции производителя.
Лигирование и трансформация ДНК
После очистки фрагментов ДНК с помощью набора для извлечения из геля, они были подвергнуты лигированию с вектором pSP72, обработанным CIAP. Для проведения лигирования необходимое количество ДНК переносилось в стерильные 0,5 мл микроцентрифужные пробирки, как показано в Таблице 1.
До проведения лигирования фрагменты ДНК нагревались при 45°C в течение 5 минут для расплавления всех «липких» концов, которые подверглись вторичному отжигу во время подготовки фрагментов. Молярная концентрация вектора:вставленной ДНК, равная 1:1, была выбрана для всех реакций лигирования, и подготовка реакционной смеси для реакции проводилась в соответствии с инструкциями компании Promega.
В пробирки A и B был добавлен 1,0 мкл 10х буфера для лигирования и 0,5 единиц Вейса ДНК-лигазы T4 (Promega, Великобритания), и объем смеси для лигирования доводился до 10 мкл путем добавления воды для молекулярной биологии. В пробирки С был добавлен 1,0 мкл 10х буфера для лигирования, и объем смеси для лигирования доводился до 10 мкл путем добавления воды для молекулярной биологии.
Фрагменты ДНК добавлялись в пробирки вместе с водой и затем нагревались до 45°C в течение 5 минут для расплавления всех «липких» концов, которые подверглись вторичному отжигу во время подготовки. Перед добавлением остальных реагентов для лигирования ДНК охлаждалась до 0°C, после добавления реакционная смесь инкубировалась в течение ночи при 16°C (Sambrook and Russell, 2001).
После осаждения и очистки лигированных фрагментов посредством этанола (с целью удаления смеси для лигирования, которая вызывает снижение эффективности трансформации), проводились трансформации согласно инструкциям Hanahan. ~50 нг лигированной ДНК в 5 мкл раствора было добавлено к 100 мкл компетентных клеток DH5a. После тепловой обработки и экспрессии гена устойчивости к ампициллину клетки распределялись по поверхности чашек Луриа-Бертани, содержащих ампициллин (100 мкг/мл), X-β-Gal (40 мкл 2% X-β-Gal) и IPTG (7 мкл 20% IPTG).
Измерялось количество трансформантов для каждой реакции лигирования. Обычно число полученных трансформантов для пробирки C составляло 2×105-1×106 КОЕ/мкг, тогда как для пробирки A - 500-600 КОЕ/мкг. Число трансформантов в пробирке A служило показателем эффективности обработки векторной ДНК. Число трансформантов в пробирке B находилось в диапазоне 2-4×104 КОЕ/мкг.
Число трансформантов
Для смеси для лигирования хромосомной ДНК с PstI было получено 13 положительных клонов β-галактозидазы из ~2500 просканированных трансформантов, тогда как с BamHI было получено 7 положительных клонов (из ~1500 просканированных трансформантов), для EcoRI было получено 3 положительных клона (из ~1300 просканированных трансформантов), для KpnI было получено 7 положительных клонов (из ~2000 просканированных трансформантов), для SmaI было получено 3 положительных клона (из ~1600 просканированных трансформантов) и для HindIII было получено 2 положительных клона (из ~1200 просканированных трансформантов).
Расщепление положительных клонов
С целью идентификации различных генов β-галактозидазы, плазмиды, выделенные из положительных клонов, подвергались расщеплению в соответствии с таблицей 2.
Гель-электрофорезный анализ полученных фрагментов после расщепления показал, что каждая из плазмид pB1, pP1, pP2 и pP11 содержала вставку, кодирующую различные β-галактозидазы. Клоны, содержащие P2, использовались для последующего анализа.
Секвенирование ДНК
Секвенирование ДНК проводилось с применением метода дидезокси-секвенирования (обрыва цепи) по Сангеру с использованием набора для циклического секвенирования BigDye Terminator v.3.0 (Applied Biosystems, США), анализ проводился с помощью ABI Prism 3100, системы анализа ДНК на основе флуоресценции, включающей капиллярный электрофорез.
5'- и 3'-концы вставленных фрагментов ДНК секвенировались с помощью праймеров, специфичных для вектора. Дальнейшее секвенирование вставок проводилось с использованием Genome Priming System (GPS-I) (New England Biolabs, Великобритания). GPS-1 представляет собой систему in vitro, основанную на использовании транспозона TN7, в которой транспозаза TnsABC применяется для случайной вставки транспозона в целевую ДНК. В соответствии с инструкциями производителя, использовалось соотношение масс донор:целевая ДНК, составлявшее 1:4. Число выделенных плазмид для секвенирования после вставки транспозонного праймера составило 25. Это число было рассчитано в соответствии с инструкциями производителя и предполагает 5-кратную полноту покрытия.
По причине большой длины нуклеотидной последовательности плазмиды pP2, кодирующей белок P2, для секвенирования была выбрана только ее часть, содержащая ген β-галактозидазы. Инактивация фермента после вставки транспозазы в соответствующей позиции 172 н.п. отсеквенированного фрагмента показала, что старт-кодон находился в направлении против хода транскрипции относительно данной позиции. Аналогичным образом, вставка в позиции 2882 н.п. полностью ликвидировала активность фермента, что показывает наличие стоп-кодона в направлении хода транскрипции относительно данной позиции. Более того, активность фермента полностью ликвидировалась при вставке в позициях 262 н.п., 331 н.п., 375 н.п., 621 н.п., 866 н.п., 1348 н.п., 1358 н.п., 1394 н.п., 1513 н.п., 1704 н.п., 2128 н.п., 2519 н.п. относительно первого отсеквенированного нуклеотида.
Анализ N-концевого домена программами SignalIP и PSORT не выявил ни одного сигнального пептида, что является показателем того, что P2 не секретируется внеклеточно.
Реакционная смесь для секвенирования содержала приблизительно 400-600 нг плазмидной ДНК, 3,2 пмоль раствора праймера и 4 мкл раствора BigDye Terminator.
Идентификация открытой рамки считывания
Положение открытой рамки считывания P2 было выявлено посредством программы поиска ОРС на сайте NCBI по адресу http://www.ncbi.nlm.nih.gov/gorf/gorf.html. Использовался генетический код бактерии, при этом длина рамки составила 100 н.п. Нуклеотидная последовательность транслировалась по всем шести возможным рамкам, при этом была идентифицирована открытая рамка считывания для 738 аминокислот, предположительно кодирующая β-галактозидазу. (Результат трансляции показан на Фиг.2.)
Пример 2
Синтез с клонированным ферментом β-галактозидазы, выделенным из Bifidobacterium bifidum NCIMB 41171 в клетках-хозяевах E.coli (штамм DH5a)
Нижеописанный синтез, если не указано обратное, проводился в целых клетках-хозяевах E.coli DH5a после обработки биомассы E.coli (собранной центрифугированием на 10000 g) толуолом с концентрацией 2000 частей на миллион с целью увеличения проницаемости клеток и для приведения клеток в нежизнеспособное состояние путем разрушения их цитоплазматической мембраны. Биомасса E.coli приготовлялась в соответствии с описанием, приведенным в Примере 1 в разделе "Штаммы E.coli".
Синтез с клонированным ферментом
Синтез с клонированным ферментом β-галактозидазы проводился при массовой концентрации субстрата, составляющей 40% (по массовой доле) начальной концентрации лактозы. Раствор для синтеза приготовлялся в 0,1M фосфатном буфере при pH 6,0, дополнительно содержащем 1 г/л Tween 80 (полиоксиэтиленсорбитана моноолеат (20)). Синтез проводился при 40°C на водяной бане, вращающейся на 150 об/мин. Оптимальное значение pH для конкретного фермента выбиралось на основе измерений активности (с использованием o-нитрофенил-β-D-галактопиранозида в качестве субстрата) конкретной ферментной композиции при различных значениях pH.
Для синтеза галактоолигосахарида использовалось 2 мл супернатантного клеточного лизата (после разрушения клеток E.coli французским прессом) с 8 г 50%-ной (по массовой доле) лактозы с целью получения итоговой концентрации субстрата, равной 40% (по массовой доле). Ферментная композиции имела активность 735 единиц/мл.
Концентрации различных сахаров, присутствовавших в смеси в процессе синтеза, показаны на Фиг.3. Объединенные хроматограммы высокоэффективной анионообменной хроматографии и импульсного амперометрического детектирования (HPAEC-PAD) смесей галактоолигосахаридов, синтезированных посредством β-галактозидазы, клонированной из B.bifldum NCIMB 41171, показаны на Фиг.4. Концентрации сахаров в смеси галактоолигосахаридов в оптимальной временной точке синтеза показаны в таблице 3.
DP≥3
DP=2











Изобретение относится к области биохимии. Представлена молекула ДНК, выделенная из Bifidobacterium bifidum, с последовательностью, приведенной в описании, и ее вырожденная производная, кодирующие β-галактозидазу. Также представлен β-галактозидазный фермент, имеющий последовательность, приведенную в описании. Описаны экспрессионный вектор, содержащий указанную ДНК, и бактериальная клетка-хозяин, содержащая данный вектор. Предложено применение фермента или клетки-хозяина для получения смеси олигосахаридов, которые могут быть использованы для приготовления пищевых продуктов, кормов для животных и медикаментов. Описан способ продуцирования фермента, включающий культивирование указанной клетки-хозяина в подходящей культуральной среде в условиях, допускающих экспрессию указанного фермента и выделение полученного фермента из культуры. Изобретение позволяет трансформировать лактозу в смесь олигосахаридов. 9 н. и 3 з.п. ф-лы, 4 ил., 3 табл.
1. ДНК,
a) кодирующая β-галактозидазу с аминокислотной последовательностью, представленной в SEQ ID NO: 2, и имеющую β-галактозидазную активность, или
b) имеющая вырожденную последовательность по п. а), где указанная последовательность содержит замены нуклеотидов, приводящие к консервативным аминокислотным заменам.
2. ДНК по п.1, представляющая собой последовательность, представленную в SEQ ID NO: 1.
3. Фермент β-галактозидаза, кодируемая ДНК по п.1 или 2.
4. β-галактозидаза, имеющая последовательность, представленную в SEQ ID NO: 2.
5. Рекомбинантный экспрессионный вектор, содержащий ДНК по п.1 или 2.
6. Рекомбинантная бактериальная клетка-хозяин для продуцирования фермента, используемого для получения олигосахаридов, содержащая ДНК по п.1 или 2.
7. Рекомбинантная бактериальная клетка-хозяин для продуцирования фермента, используемого для получения олигосахаридов, содержащая вектор по п.5.
8. Клетка-хозяин по п.6 или 7, где клетка выбрана из группы, состоящей из Bifidobacterium и Escherichia.
9. Клетка-хозяин по п.8, где указанная клетка представляет собой Bifidobacterium bifidum.
10. Применение фермента по п.3 или 4, или клетки по любому из пп.6-9, для продуцирования смеси олигосахаридов.
11. Применение фермента по п.3 или 4, или клетки по любому из пп.6-9 для продуцирования смеси олигосахаридов, являющейся частью продукта, выбранного из группы, состоящей из молочных продуктов, таких как жидкое молоко, порошковое молоко, молоко для детского питания, смесь для детского питания, мороженое, йогурт, сыр, ферментированные молочные продукты, напитков, таких как фруктовый сок, детского питания, каши, хлеба, печенья, кондитерских изделий, пирожных, пищевых добавок, диетических добавок, пробиотических пищевых продуктов, пребиотических пищевых продуктов, кормов для животных, кормов для птицы и медикаментов.
12. Способ продуцирования фермента по п.3 или 4, включающий культивирование клетки-хозяина по любому из пп.6-9 в подходящей культуральной среде в условиях, допускающих экспрессию указанного фермента и выделение полученного фермента из культуры.
| ИЗГОТОВЛЕНИЯ ПРУЖИН РАСТЯЖЕНИЯс УШКАМИ | 0 |
|
SU190317A1 |
| WO 2005003329, 13.01.2005 | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |
| PETER L | |||
| MELLER et | |||
| al | |||
| Intra- and Extracellular β-Galactosidases from Bifidobacterium bifidum and B.infantis: Molecular Cloning, Heterologous Expression, and Comparative Characterization, Appi Environ Microbiol., 2001 May; 67(5): 2276-2283 | |||
| СПОСОБ ПОЛУЧЕНИЯ ПРОБИОТИКА, ШТАММ STREPTOCOCCUS SALIVARIUS SUBSP. THERMOPHILUS ВКПМ В-7984, ШТАММ STREPTOCOCCUS SALIVARIUS SUBSP. THERMOPHILUS ВКПМ В-7985, ИСПОЛЬЗУЕМЫЕ ДЛЯ ПОЛУЧЕНИЯ ПРОБИОТИКА | 2002 |
|
RU2260041C2 |

