Настоящее изобретение относится к β-галактозидазе с трансгалактозилирующей активностью, способной превращать лактозу в смесь олигосахаридов, которые соединены β-связью, и неожиданно продуцировать дисахарид α1-6-галактобиозу с α-связью. В частности, изобретение относится к β-галактозидазе, выделенной из недавно открытого штамма Bifidobacterium bifidum.
В частности, изобретение относится к последовательностям ДНК, кодирующим выделенный фермент β-галактозидазу, к ферменту, кодируемому последовательностью ДНК, и к клетке-хозяину, содержащей последовательность ДНК или рекомбинантный вектор, включающий последовательность ДНК. Настоящее изобретение также относится к применению фермента, кодируемого последовательностью ДНК, или клетки-хозяина, содержащей последовательность ДНК или рекомбинантный вектор, для производства олигосахаридов.
Бифидобактерии естественным образом колонизируют нижний отдел кишечного тракта, среда которого содержит незначительное количество моно- и дисахаридов, поскольку такие сахара преимущественно потребляют хозяин и микроорганизмы, присутствующие в верхнем отделе кишечного тракта. Для того чтобы существовать в нижнем отделе кишечного тракта, бифидобактерии продуцируют различные поверхностно-связанные экзо- и эндогликозидазы и/или внеклеточные формы, с помощью которых они могут утилизовать различные углеводы.
Наряду с гидролазной активностью, некоторые ферменты бифидобактерий обладают трансферазной активностью. Эта трансгликозилирующая активность гликозидаз широко используется для ферментативного синтеза различных олигосахаридов, которые оказались способными действовать в качестве факторов, стимулирующих рост бифидобактерий.
Известно, что представители бифидобактерий продуцируют ферменты β-галактозидазы, которые участвуют в бактериальном метаболизме лактозы. Møller, P.L. et al. в Appl & Environ. Microbial, (2001), 62, (5), 2276-2283 описывают выделение и дают характеристики трех β-галактозидазных генов из штамма Bifidobacterium bifidum. Они обнаружили, что все три β-галактозидазы способны катализировать образование бета-связанных галактоолигосахаридов посредством трансгалактозилирования.
Dumortier et al. в Carbohydrate Research, 201, (1990), 115-123 описали образование бета-связанных олигосахаридов по реакции транслактозилирования при гидролизе лактозы Bifidobacterium bifidum DSM 20456. Их анализ структуры смеси продуцируемых олигосахаридов показал, что эти связи были β-(1→3)-, β-(1→6)- и β-(1→4)-D-галактозильными связями. Dumortier предположил, что соединения, продуцируемые Bifidobacterium bifidum, необходимы для адгезии бактерий в толстом кишечнике.
Был обнаружен штамм Bifidobacterium bifidum, способный продуцировать активный фермент галактозидазу, превращающий лактозу в новую смесь галактоолигосахаридов, которая, как неожиданно было обнаружено, содержит до 35% дисахаридов, включая галабиозу (Gal(α1-6)-Gal). Известно (см. Paton, J C and Paton, A W (1998), Clin. Microbiol. Revs., 11, 450-479; Carlsson, K A (1989), Ann. Reviews Biochem., 58, 309-350), что этот дисахарид является антиадгезивом, способным предотвращать адгезию токсинов, например, токсина Shiga, и патогенов, таких как E. coli, к стенкам кишечника.
Этот штамм B. bifidum 31 марта 2003 года был депонирован в Национальной коллекции промышленных и морских бактерий, Абердин, Великобритания(Eng). Он также описан в патенте UK №2412380.
Было обнаружено, что штамм B. bifidum продуцирует несколько β-галактозидаз, одна из которых неожиданно демонстрирует активность α-галактозилтрансферазы. Этот фермент продуцирует ряд различных олигосахаридов, которые связаны β-связями, но также продуцирует и α-связанный дисахарид галабиозу.
Настоящее изобретение относится к последовательности ДНК, которая кодирует белок с аминокислотной последовательностью, представленной в SEQ ID NO: 2, или в жестких условиях гибридизуется с последовательностью ДНК, которая кодирует этот белок. В SEQ ID NO: 1 дана эта последовательность ДНК, или ее фрагмент, или вырожденное производное.
Термин «вырожденное производное» означает последовательность ДНК, которая гомологична SEQ ID NO: 1 не менее чем на 50%, предпочтительно, на 50-98%, наиболее, предпочтительно, на 75-95%.
Такая последовательность ДНК может содержать нуклеотидные замены, добавления или делеции, которые в результате дают менее 60%, предпочтительно менее 45%, более предпочтительно менее 25% изменений в аминокислотной последовательности, показанной в SEQ. ID NO: 2. Нуклеотидные замены могут в результате давать консервативные замены аминокислот.
В соответствии со вторым аспектом изобретение относится к ферменту, кодируемому последовательностью ДНК, определенной выше. Такой фермент может содержать аминокислотную последовательность, приведенную в SEQ ID NO: 2, или ее фрагмент.
В соответствии с третьим аспектом изобретение относится к рекомбинантному вектору, предпочтительно, вектору экспрессии, содержащему последовательность ДНК, определенную выше. Такой вектор может быть частью клетки-хозяина, такого как бактериальная, дрожжевая или грибковая клетка. Альтернативно, последовательность ДНК может быть частью такой клетки-хозяина. Соответствующая клетка-хозяин может быть выбрана из Bifidobacterium, Lactococcus, Lactobacillus, Bacillus (например, Bacillus subtilus или Bacillus circulans), Escherichia и Aspergillus (например, Aspergillus niger).
Используя лактозу в качестве субстрата, фермент, кодируемый последовательностью ДНК, определенной выше, продуцирует смесь дисахаридов, включающую Gal(β1-3)Glc, Gal(β1-3)Gal, Gal(β1-6)Gal и Gal(α1-6)Gal. В этой смеси олигосахаридов также присутствуют трисахариды Gal(β1-6)Gal(β1-4)Glc, Gal(β1-3)Gal(β1-4)Glc, тетрасахарид Gal(β1-6)Gal(β1-6)Gal(β1-4)Glc и пентасахарид Gal(β1-6)Gal(β1-6)Gal(β1-6)Gal(β1-4)Glc.
Фермент клетки-хозяина, описанный выше, можно использовать для продуцирования смеси дисахаридов, включая Gal(α1-6)Gal (галабиозу), которая может составить часть продукта, предназначенного для улучшения состояния здоровья кишечника. Такой продукт может быть выбран из группы, состоящей из молочных продуктов (например, жидкого молока, сухого молочного порошка, такого как порошок цельного молока, порошок обезжиренного молока, молочные порошки, обогащенные жиром, порошки молочной сыворотки, молоко для детей, детские молочные смеси, мороженое, детские пищевые продукты, йогурт, сыр, ферментированные молочные продукты), напитков, таких как фруктовый сок, детское питание, крупы, хлеб, печенье, кондитерские изделия, торты, пищевые добавки, диетические добавки, симбиотические пищевые продукты, пребиотические пищевые продукты, корма для животных, корма для птиц или вообще любые другие пищевые продукты или напитки.
Альтернативно, олигосахариды, произведенные таким образом, можно использовать для приготовления медикамента (например, в форме таблетки или капсулы) для предотвращения адгезии патогенов или токсинов, продуцируемых патогенами, к стенке кишечника. Этот медикамент можно вводить пациенту, например, после курса лечения антибиотиком, который часто изменяет или даже повреждает нормальную здоровую кишечную флору.
В соответствии с дальнейшим аспектом изобретение относится к способу производства фермента, определенного выше, который включает культивирование клетки-хозяина, определенной выше, в соответствующей культуральной среде в условиях, позволяющих экспрессию этого фермента, и выделение продуцированного фермента из этой культуры.
Изобретение также относится к способу получения смеси олигосахаридов, включая дисахарид Gal(α1-6)-Gal (галабиозу), который включает контактирование фермента, определенного выше, или клетки-хозяина, определенной выше, с материалом, содержащим лактозу, в условиях, ведущих к образованию этой смеси олигосахаридов.
Подходящий материал, содержащий лактозу, может быть выбран из коммерчески доступной лактозы, цельного молока, полуобезжиренного молока, обезжиренного молока, молочной сыворотки и молока, обогащенного жиром, фильтрата молочной сыворотки. Такие молочные продукты могут быть получены от коров, буйволиц, овец или коз. Молоко, обогащенное жиром, определяют как цельное молоко, из которого удалили молочный жир, который затем заменили добавлением растительного жира или масла.
Краткое описание чертежей
На фигуре 1 показана нуклеотидная последовательность (SEQ ID NO: 1) β-галактозидазы Bifidobacterium bifidum по изобретению; и
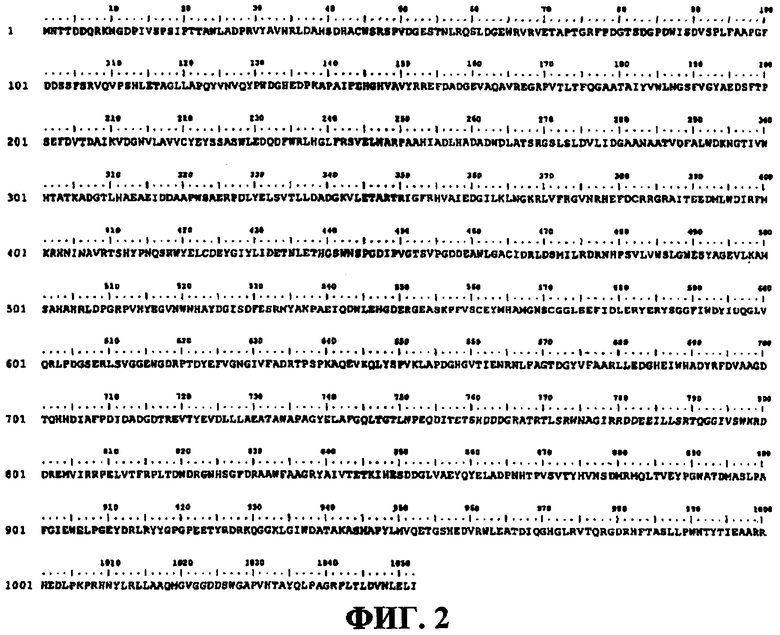
на фигуре 2 показана аминокислотная последовательность (SEQ ID NO: 2), соответствующая нуклеотидной последовательности фигуры 1;
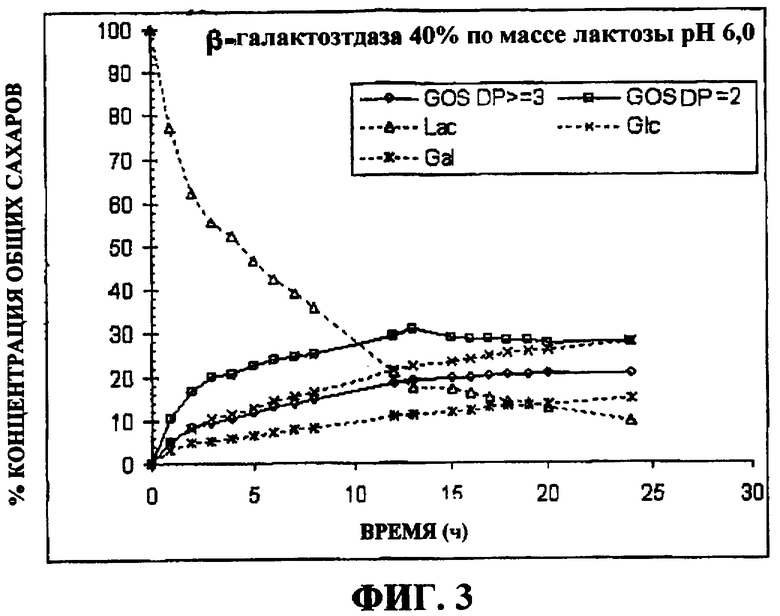
на фигуре 3 представлен график, показывающий протекание во времени реакции при синтезе галактоолигосахарида с β-галактозидазой и 40% по массе лактозы в качестве субстрата в 0,1 М фосфатном буфере при рН 6,0; и
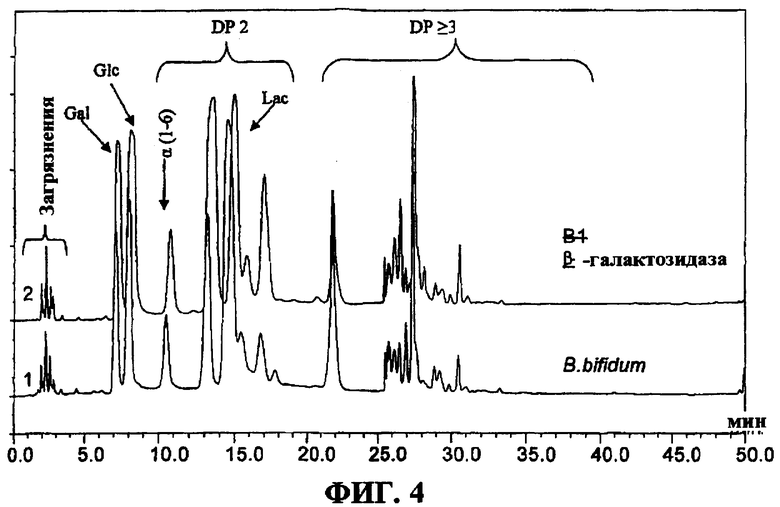
на фигуре 4 представлена высокоэффективная анионобменная хроматограмма смеси галактоолигосахаридов, синетзированных β-галактозидазой из B. bifidum NCIMB 41171 с использованием 40% по массе лактозы в качестве субстрата в 0,1 М фосфатном буфере при рН 6,0. (Glc = глюкоза, Gal = галактоза, Lac = лактоза, α(1-6)галактобиоз, DP = степень полимеризации).
Геномную ДНК выделяли из штамма Bifidobacterium bifidum (NCIMB 41171) с использованием способа Lawson et al. (1989) Fems Microbiol Letters, 65, (1-2), 41-45. ДНК обрабатывали ферментами рестрикции, и фрагменты с максимальным размером 15 т.п.н. лигировали с вектором pSP72, который подвергся действию тех ферментов рестрикции. Клетки E. coli трансформировали вектором, содержащим вставки, состоящие из хромосомной ДНК B. bifidum, обработанной (рестриктазами) PstI, Eco RI, Bam HI, KpnI, SmaI или HindIII. Клоны с β-галактозидазной активностью отобрали на агаровых средах Лурия-Бертани, содержащих п-нитрофенол, X-β-Gal (5-бром-4-хлор-3-индолил-β-D-галактозид) и изопропил-β-D-тиогалактозид (IPTG). Лигирующие смеси с хромосомной ДНК, переваренной Bam HI, дали семь клонов, положительных на β-галактозидазу, один из которых был идентифицирован как рВ1.
Секвенирование вставленного фрагмента ДНК В1 проводили с использованием дидезоксинуклеотидного метода терминации цепи по Сенгеру (Russel P., 2002 iGenetics, Pearson Education, Inc., San Francisco, 187-189) с использованием набора для циклического секвенирования BigDye Terminator V.3.0 (Applied Biosystems, USA). Последовательность ДНК B1 показана на фигуре 1 (SEQ ID NO: 1).
Открытую рамку считывания (ORF) идентифицировали с помощью программы ORF Finder из NCBI (National Center of Biotechnology Information). Нуклеотидная последовательность Фигуры 1 была транслирована во всех шести возможных рамках считывания и была идентифицирована одна открытая рамка считывания (1052 аминокислот), кодирующая предполагаемую β-галактозидазу. Трансляция показана на Фигуре 2 (SEQ ID NO: 2).
Ниже настоящее изобретение описано со ссылками на следующие примеры.
Пример 1
Материалы и методы
Все химические реагенты и препараты сред, использованные в этой работе, были получены от Sigma (Dorset, UK), Invitrogen (Paisley, UK), Oxoid (Basingstoke, UK), Qiagen (West Sussex, UK) и Promega (Southampton, UK).
Бактериальные штаммы
Штамм Bifidobacterium bifidum (NCIMB 41171) хранили на криогенных гранулах в пробирках Microbank при -70°С. Для последующих экспериментов штамм оживляли на агаре Wilkinson Chalgren (WC) (Oxoid, UK) и среде TPY (среда Триптиказа с фитоном и дрожжевым экстрактом) и выращивали анаэробно (состав CO2 и N2 - 80% и 20% соответственно) при 37°C в течение 48 часов. Морфологию колоний и отсутствие загрязнений проверяли окрашиванием по Граму.
Штаммы E. coli
Штамм Escherichia coli DH5a, использованный в этой работе, обычно инкубировали в аэробных условиях при 37°С на агаровой среде Лурия-Бертани (LB) или бульоне (Sambrook J. and Russell W. D. (2001). Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, New York) и при необходимости добавляли антибиотики (100 мкг/мл ампициллина и/или 15 мкг/мл хлорамфеникола) и 40 мкл 2% X-β-Gal, 7 мкл 20% изопропил-β-D-тиокалактозида (IPTG), которые наносили на поверхности заранее приготовленных 90-мм агаровых чашек.
Штамм E. coli DH5a (Invitrogen, Paisley, UK) (генотип: F- φ80lacZΔM Δ(lacZYA-argF)U169 recAl endAl hsdR17(rk -, mk -) phoA sιιpE44 thi-l gyrA96 relAlλ-) является штаммом, положительным по α-галактозидазе, и его использовали в экспериментах по экспрессии и для других генетических манипуляций.
Экстракция геномной ДНК из Bifidobacterium bifidum
Геномную ДНК выделяли из штамма Bifidobacterium bifidum (NCIMB 41171) с использованием следующего способа, в котором хромосомную ДНК готовили из клеточного осадка, собранного из 100 мл анаэробного бульона WC. Клетки ресуспендировали в 10 мл TES-буфера (10 мМ Tris-HCl, 10 мМ EDTA, 10 мМ NaCl, pH 8) и обработали 200 мкл смеси лизоцима с мутанолизином (4:1, лизоцима 10 мг/мл, мутанолизин 1 мг/мл) в течение 30 минут при 37°С. Затем клетки обработали 200 мкл протеиназы К (при 20 мг/мл) и 200 мкл РНКазы (обе 10 мг/мл), перемешали и инкубировали 1 час при 65°С. Окончательно клетки обработали 2 мл 10% ДДС и инкубировали 15 мин при 65°С. Добавили 12 мл фенола с хлороформом и экстракцию повторяли до тех пор, пока водная фаза не стала легко отделяться от промежуточной фазы. Геномную ДНК осадили изопропанолом и ресуспендировали в 10 мМ Tris-HCl - 1 мM EDTA (pH 8). Затем геномную ДНК переварили с ферментами рестрикции, провели лигирование в pSP72, переваренную теми же ферментами, и обработали щелочной фосфатазой. Переваривание геномной ДНК B. bifidum проводили, используя EcoRI, PstI, BamHI, SmaI и KpnI. Лигирующие смеси использовали для трансформирования E. coli DH5a; клоны, положительные по β-галактозидазе, идентифицировали как синие колонии на пластинах, содержащих X-Gal.
Приготовление векторной ДНК
В этой работе в качестве вектора для клонирования и экспрессии использовали pSP72 (Promega, UK) (Krieg, P.A. and Melton, D.A. (1987). In vitro RNA synthesis with SP6 RNA polymerase. Methods in Enzymology. 155: 397-415).
Этот вектор выбрали вследствие отсутствия комплементарной активности α-фрагмента β-галактозидазы, который не кодирован в pSP72. Этот вектор не несет короткий сегмент ДНК E. coli, содержащий регуляторную последовательность и кодирующий информацию для первых 146 аминокислот β-галактозидазы, который, в комбинации со штаммами E. coli (т.е. DH5a), экспрессирующими С-концевую часть этой β-галактозидазы, дает активную β-галактозидазу (α-комплементация).
Вектор переваривали следующими рестрикционными ферментами: PstI, BamHI, HindIII, SmaI, KpnI and EcoRI в соответствии с инструкциями изготовителя, с использованием десятикратного избытка фермента по отношению к ДНК (единицы фермента: микрограмм ДНК равен 10 единицам фермента на один мкг плазмидной ДНК или 10 единицам фермента на 0,5 пмоль плазмидной ДНК). После термоинактивации ферментов (20 мин при 65°С) рестрикционную картину анализировали с помощью электрофореза в горизонтальном геле. Присутствие единственного фрагмента в геле означало полное переваривание вектора и его единственный сайт рестрикционного переваривания.
Достаточность переваривания вектора проверяли также трансформацией нелигированных молекул в компетентные клетки E. coli DH5a. Количество образованных колоний на пластинах LB-агара с добавленным ампицилином (100 мкг/мл) служило показателем непереваренных молекул и ожидаемого фона при последующих экспериментах.
Векторы дополнительно дефосфорилировали телячьей кишечной щелочной фосфатазой CIAP (Promega, Southampton, UK) в соответствии с инструкциями производителя. Эффективность этой обработки проверяли по самолигированию (с ДНК-лигазой бактериофага Т4 в соответствии с инструкцией производителя) после трансформации в клетки DH5a. Количество образованных колоний показывало количество молекул, повторно замкнутых в кольца (неклонированный вектор), а вычитание из него числа образованных колоний без обработки вектора CIAP давало количество недефосфорилированного вектора.
Построение библиотеки геномной ДНК
Геномную ДНК частично переварили шестью ферментами рестрикции, которые узнают часто встречающиеся гексануклеотидные последовательности в прокариотической ДНК. EcoRI, BamHI, PstI, KpnI, SmaI и HindIII являются рестрикционными эндонуклеазами типа II, специфично узнающими последовательности 5'G/AATTC3', 5'G/GATCC3', 5'CTGCA/G3', 5'GGTAC/C3', 5'CCC/GGG3' и 5'Α/AGCTT3' соответственно и делают двухнитевые разрывы в этих последовательностях, образуя 5'-оверхенги из четырех нуклеотидов, AATT, GATC, AGCT для EcoRI, BamHI и HindIII соответственно, и 3'-оверхенги, ACGT, GTAC для PstI и KpnI соответственно и тупые концы для SmaI.
Все эти ферменты были активными и способными расщеплять ДНК только в присутствии ионов магния. Эти ионы были единственным необходимым кофактором.
Рестрикционное переваривание ДНК
Переваривание образцов ДНК всеми рестриктазами проводили при инкубации в течение 2 часов при 37°С с конечной термоинактивацией при 65°С в течение 20 минут. Реакционные смеси затем охладили до комнатной температуры и добавили соответствующее количество рабочего буфера с последующим осторожным перемешиванием запаянным стеклянным капилляром. Затем растворы загрузили в лунки 0,8% агарозного геля (при напряжении 4-5 вольт/см в течение 14-16 часов) и размер переваренной ДНК оценивали с помощью 1-тпо стандартов ДНК (Promega, UK) (Sambrook J. Molecular Cloning: A Laboratory Manual (2002)).
Очистка фрагментов, образованных после переваривания рестриктазами
Из реакционных смесей и агарозных гелей фрагменты очищали с помощью набора для экстракции из гелей QIAEX от Qiagen (West Sussex, UK). Протоколы подробно описаны в инструкции производителя.
Лигирование ДНК и трансформация
После очистки фрагментов ДНК с помощью набора для экстракции из гелей Qiaex их лигировали с вектором pSP72, обработанным CIAP. Для лигировния соответствующие количества ДНК переносили в стерильные микроцентрифужные пробирки, как показано в Таблице 1.
Таблица 1. Смеси лигирования. Пробирка А показывает количество самолигированной векторной ДНК, которую следует вычесть из общего количества трансформантов после трансформации. Пробирка В показывает лигирование вектора с фрагментами ДНК, а пробирка С показывает контроль для расчета эффективности трансформации.
Перед каждым лигированием фрагменты ДНК нагревали при 45°С в течение 5 минут для расплавления любых липких концов, которые подверглись повторному отжигу при приготовлении фрагмента. Молярное отношение вектор:вставка ДНК было выбрано на уровне 1:1 для всех реакций лигирования, которые проводили по инструкциям Promega.
В пробирки А и В добавили по 1,0 мкл 10-кратного буфера лигирования и 0,5 единиц Вейсса ДНК-лигазы Т4 (Promega, UK) и объем довели до 10 мкл водой молекулярно-биологической категории чистоты. В пробирку С добавили 1,0 мкл 10-кратного буфера лигирования и объем лигирования довели до 10 мкл водой молекулярно-биологической категории чистоты.
В пробирки добавили фрагменты ДНК с водой и нагрели до 45°С в течение 5 минут для расплавления всех липких концов, которые подверглись повторному отжигу при приготовлении. Перед добавлением остальных реагентов лигирования ДНК охладили до 0°С и реакционные смеси инкубировали в течение ночи при 16°С (Sambrook and Russell, 2001).
После осаждения этанолом и очистки лигированных фрагметов (для удаления лигирующей смеси, которая снижает эффектвность трансформации) проводили трансформацию по инструкциям Hanahan. ~50 нг лигированной ДНК в 5 мкл раствора добавили к 100 мкл компетентных клеток E. coli DH5a. После термообработки и экспрессии гена резистентности к ампициллину клетки распределили по поверхности чашек среды LB, содержащей ампициллин (100 мкг/мл), X-β-Gal (40 мкл 2% X-β-Gal) и IPTG (7 мкл 20% IPTG).
Определяли количество трансформантов из каждой реакции лигирования. Обычно количество трансформантов, получаемых из пробирки С, составляло 2×105-1×106 КОЕ/мкг, тогда как из пробирки А получали 500-600 КОЕ/мкг. Количество трансформантов в пробирке А было показателем эффективности векторной ДНК. Количество трансформантов в пробирке В было в диапазоне 2-4×106 КОЕ/мкг.
Количество трансформантов
Лигирующие смеси с хромосомной ДНК, переваренной Pstl, дали 13 клонов, положительных на β-галактозидазу из ~2500 проскринированных трансформантов, тогда как после переваривания BamHI получено 7 положительных клонов (~1500 проскринированных трансформантов), обработка EcoRI дала 3 положительных клона (~1300 проскринированных трансформантов), KpnI - 7 положительных клонов (~2000 проскринированных трансформантов), SmaI - 3 положительных клона (~1600 проскринированных трансформантов) и HindIII - 2 положительных клона (~1200 проскринированных трансформантов).
Переваривание положительных клонов
Для идентификации различных генов β-галактозидазы плазмиды, выделенных из положительных клонов, подвергли перевариванию в соответствии со следующей таблицей 2.
Гель-электрофоретический анализ фрагментов, образованных после переваривания, показал, что каждая плазмида pB1, pP1, pP2 и pP11 имеет вставку, которая кодирует иную β-галактозидазу. Для дальнейшего анализа использовали клоны, содержащие рВ1.
Секвенирование ДНК
ДНК секвенировали, применяя дидезоксинуклеотидную терминацию по Сэнгеру, используя набор для циклического секвенирования BigDye Terminator v.3.0 (Applied Biosystems, USA), и анализировали с помощью флуоресцентной системы анализа ДНК ABI Prism 3100, включающей капиллярный электрофорез.
5'- и 3'-концы вставленных фрагментов ДНК секвенировали с праймерами, специфичными для векторов. Для дальнейшего секвенирования вставок использовали Genome Priming System (GPS-I) (New England Biolabs, UK). GPS-I является системой, действующей in vitro, основанной на транспозоне TN7 и использующей транспозазу TnsABС для того, чтобы случайным образом вставлять транспозон в ДНК-мишень. В соответствии с инструкцией производителя использовали отношение масс донор:ДНК-мишень, равное 1:4. Количество плазмид, выделенных для секвенирования после вставок транспраймера в плазмиду-мишень, составляло 25. Это количество было рассчитано в соответствии с инструкциями производителя и предполагало 5-кратную глубину покрытия.
Для плазмиды рВ1 вставка транспозона, приблизительно равного 1699 тпо, в позиции у 973-й пары оснований ниже множественно клонированного сайта использованного вектора полностью элиминировала активность β-галактозидазы, тем самым демонстрируя, что стартовый кодон находился между вектором МCS (multiple cloning site) и сайтом транспозона, тогда как внедрение вставки в позиции 841 ниже MCS вело к образованию активной β-калактозидазы, тем самым указывая, что стартовый кодон существует между парами оснований в позициях 841 и 973 ниже MCS. Ферментативная активность полностью устранялась внедрением вставки в позиции 3586 пар оснований ниже MCS, тем самым указывая, что терминирующий кодон находится ниже этой позиции. Более того, вставки в позициях 1239 по, 1549 по, 1683 по, 1832 по, 2108 по, 2189 по, 2270 по, 2340 по, 2414 по, 2574 по, 2648 по, 2734 по, 2807 по и 3410 по полностью устраняли ферментативную активность.
Реакционная смесь секвенирования содержала приблизительно 400-600 нг плазмидной ДНК, 3,2 пмоль раствора праймера и 4 мкл раствора BigDye Terminator.
Идентификация открытой рамки считывания
Открытая рамка считывания (ОРС) была обнаружена с использованием ОРС-искателя от NCBI. Длина рамки, определенная с использованием бактериального генетического кода, составила 100 пар оснований. Нуклеотидная последовательность была транслирована в шесть возможных рамок, что позволило идентифицировать открытую рамку считывания 1052 аминокисот, кодирующую предполагаемую β-галактозидазу (эта трансляция показана на Фигуре 2).
Пример 2
Синтез с помощью клонированного фермента β-галактозидазы, выделенной из Bifidobacterium bifidum NCIMB 41171 в E. coli в качестве хозяина (штамм PH5a)
Синтез, описанный ниже, проводили, если не указано иначе, с целыми клетками E. coli DH5a в качестве хозяина после обработки биомассы E. coli (собранной центрифугированием при 10000 g) толуолом в концентрации 2000 чнм, для увеличения проницаемости клетки, чтобы сделать ее нежизнеспособной, разрушив ее цитоплазматическую мембрану. Биомассу E. coli готовили, как описано в п. «Штаммы E. coli» Примера 1.
Синтез с помощью клонированного фермента
Синтез с β-галактозидазой проводили при начальной концентрации лактозы, равной 40 мас.%, принятой в качестве концентрации субстрата. Раствор для этого синтеза готовили в 0,1 М фосфатном буфере при рН 6,8 (или 0,1 М цитратном буфере рН 6,2 или калий-фосфатном буфере рН 6,8). Синтез проводили при 40°С на водяной бане при встряхивании при 150 об/мин. Оптимум рН для этого специфического фермента выбрали, основываясь на измерениях активности (с использованием о-нитрофенил-β-D-галактопиранозида в качестве субстрата) препарата специфического фермента при различных значениях рН.
Для синтеза галактоолигосахарида 5 мл суспензии клеток E. coli DH5a (с активностью 2,2 ед./мл) центрифугировали (при 10000 g) для сбора биомассы, отбросив супернатант. Чтобы провести синтез, эту биомассу ресуспендировали с 10 г 40% по массе раствора субстрата.
Концентрации различных сахаров, присутствующих в смеси при синтезе, показаны на фигуре 3. На Фигуре 4 показаны хроматограммы, полученные при высокоэффективной анионообменной хроматографии, сопряженной с пульсирующим амперометрическим детектированием (HPAEC-PAD), для смесей галактоолигосахаридов, синтезированных клонированной β-галактозидазой из B. bifidum NCIMB 41171. Концентрации сахаров в этой смеси галактоолигосахаридов в оптимальное время синтеза показаны в Таблице 3.
Углеводный состав синтеза галактоолигосахаридов при начальной концентрации лактозы, равной 40% по массе, в момент времени, когда наблюдалась максимальная концентрация олигосахаридов.
DP>3
DP=2











Настоящее изобретение относится к биохимии. Представлена β-галактозидаза, имеющая трансгалактозилирующую активность, с последовательностью, представленной в описании, и кодирующая ее ДНК. Описан рекомбинантный вектор экспрессии, содержащий указанную ДНК, и клетка-хозяин, содержащая представленный вектор или ДНК. Предложено применение фермента, имеющего трансгалактозилирующую активность, для получения смеси олигосахаридов, состоящей из дисахаридов, трисахаридов, тетрасахаридов и пентасахарида. Описан способ получения указанного фермента, включающий культивирование клетки-хозяина в соответствующей культуральной среде в условиях, допускающих экспрессию фермента, и выделение его из культуры. Описан также способ получения смеси олигосахаридов, включающий контактирование данного фермента или клетки-хозяина с материалом, содержащим лактозу. Изобретение позволяет получить смесь олигосахаридов путем ферментативной трансформации лактозы. 11 н. и 6 з.п. ф-лы, 4 ил., 3 табл.
1. ДНК или ее фрагмент, которая кодирует белок с последовательностью SEQ ID NO: 2.
2. ДНК по п.1, где ДНК имеет последовательность SEQ ID NO: 1.
3. Фермент, обладающий трансгалактозилирующей активностью, кодируемый последовательностью ДНК по п.1 или 2.
4. Фермент, обладающий трансгалактозилирующей активностью, содержащий аминокислотную последовательность SEQ ID NO: 2.
5. β-галактозидаза с последовательностью SEQ ID NO: 2.
6. Рекомбинантный вектор экспрессии, содержащий последовательность ДНК по п.1 или 2.
7. Клетка-хозяин, содержащая последовательность ДНК по п.1 или 2, которая продуцирует фермент по любому из пп.3-5.
8. Клетка-хозяин, содержащая вектор экспрессии по п.6, которая продуцирует фермент по любому из пп.3-5.
9. Клетка-хозяин по п.7 или 8, где указанная клетка является бактериальной клеткой.
10. Клетка-хозяин по п.9, где клетка-хозяин является Escherichia coli.
11. Применение фермента по любому из пп.3-5 для получения смеси олигосахаридов, содержащей β-связанные галактоолигосахариды и α-связанные галактоолигосахариды Gal(α1-6)Gal.
12. Применение по п.11, где смесь содержит дисахариды Gal(β1-3)Glc, Gal(β1-3)Gal, Gal(β1-6)Gal и Gal(α1-6)Gal.
13. Применение по п.11 или 12, где смесь содержит трисахариды Gal(β1-6)Gal(β1-4)Glc, Gal(β1-3)Gal(β1-4)Glc, тетрасахарид Gal(β1-6)Gal(β1-6)Gal(β1-4)Glc и пентасахарид Gal(β1-6)Gal(β1-6)Gal(β1-6)Gal(β1-4)Glc.
14. Применение фермента по любому из пп.3-5 или клетки по любому из пп.7-10 для получения смеси олигосахаридов, предназначенной для того, чтобы быть частью продукта, выбранного из группы, состоящей из молочных продуктов, таких как жидкое молоко, высушенный молочный порошок, детские молочные смеси, детские питательные смеси, мороженое, йогурт, сыр, ферментированные молочные продукты, напитки, такие как фруктовый сок, детское питание, крупяные продукты, хлеб, печенье, кондитерские изделия, торты, пищевые добавки, диетические добавки, пробиотические пищевые продукты, пребиотические пищевые продукты, корма для животных, корма для птицы и медикаменты.
15. Применение по п.14, где смесь включает дисахариды Gal(β1-3)Glc, Gal(β1-3)Gal, Gal(β1-6)Gal и Gal(α1-6)Gal, трисахариды Gal(β1-6)Gal(β1-4)Glc, Gal(β1-3)Gal(β1-4)Glc, тетрасахарид Gal(β1-6)Gal(β1-6)Gal(β1-4)Glc и пентасахарид Gal(β1-6)Gal(β1-6)Gal(β1-6)Gal(β1-4)Glc.
16. Способ получения фермента по любому из пп.3-5, включающий культивирование клетки-хозяина по любому из пп.7-10 в соответствующей культуральной среде в условиях, допускающих экспрессию указанного фермента, и выделение образующегося фермента из культуры.
17. Способ получения смеси олигосахаридов, содержащей дисахариды Gal(β1-3)Glc, Gal(β1-3)Gal, Gal(β1-6)Gal и Gal(α1-6)Gal, трисахариды Gal(β1-6)Gal(β1-4)Glc, Gal(β1-3)Gal(β1-4)Glc, тетрасахарид Gal(β1-6)Gal(β1-6)Gal(β1-4)Glc и пентасахарид Gal(β1-6)Gal(β1-6)Gal(β1-6)Gal(β1-4)Glc, включающий контактирование фермента по любому из пп.3-5 или клетки-хозяина по любому из пп.7-10 с материалом, содержащим лактозу.
| WO 2005003329 A1, 13.01.2005 | |||
| RU 93045473 A, 20.04.1996 | |||
| Верхняя опора подвижного конуса конусной дробилки | 1978 |
|
SU764720A1 |
| WO 2004080200 A1, 23.09.2004. | |||

