Изобретение относится к новой α-галактозидазе с трансгалактозилирующей активностью, способной преобразовывать мелибиозу в α-галактобиозные дисахариды. В частности, изобретение относится к α-галактозидазе, выделенной из недавно открытого штамма Bifidobacterium bifidum.
Изобретение, в частности, относится к последовательностям ДНК, кодирующим выделенный фермент α-галактозидазу, к ферменту, кодируемому такой последовательностью ДНК, и к клетке-хозяину, содержащей последовательность ДНК или содержащей рекомбинантный вектор, встраивающий последовательность ДНК. Изобретение также относится к применению фермента, кодируемого последовательностью ДНК, или клетке-хозяину, содержащей последовательность ДНК или рекомбинантный вектор для получения α-галактобиозы.
Бифидобактерии обычно населяют нижний отдел кишечника, и такая среда бедна моно- и дисахаридами, поскольку указанные сахара, предпочтительно, потребляются хозяевами и микробами, присутствующими в верхнем отделе кишечника. Для выживания в нижнем отделе кишечника бифидобактерии продуцируют различные виды экзо- и эндогликозидаз, поверхностно связанных и/или находящихся в виде внеклеточных форм, которые способны утилизировать различные углеводы.
Помимо гидролазной активности, некоторые ферменты бифидобактерий обладают трансферазной активностью. Такая трансгликозилирующая активность гликозидаз интенсивно используется для ферментативного синтеза различных олигосахаридов, которые, как было доказано, выполняют функции факторов, стимулирующих рост бифидобактерий.
Известно, что некоторые виды бифидобактерий продуцируют β-галактозидазные ферменты, участвующие в бактериальном метаболизме лактозы. Так в Mollet, P.L. et al. in Appl & Environ Microbial., (2001), 62, (5), 2276-2283 описано выделение и характеристики трех генов β-галактозидазы из штамма Bifidobacterium bifidum. Авторы обнаружили, что все три β-галактозидазы способны катализировать образование бета-связанных галактоолигосахаридов в результате трансгалактозиляции.
Известно, что некоторые виды бифидобактерий, в отличие от B. bifidum продуцируют α-галактозидазы, а также β-галактозидазы. α-Галактозидазы принадлежат к группе гликозилгидролаз и могут быть классифицированы на две группы по специфичности к субстрату, т.е. одна из таких групп обладает специфичностью к малым сахаридам, таким как p-нитрофенил-α-D-галактопиранозид, мелибиоза и раффиноза, а другая группа способна высвобождать галактозу из галактоманназ таких, как гуаровая камедь, кроме других малых субстратов.
Было обнаружено, что штамм Bifidobacterium bifidum способен продуцировать фермент с галактозидазной активностью, преобразующий лактозу в новую смесь галактоолигосахаридов, которая, как неожиданно было отмечено, содержит до 35% дисахаридов, включая галабиозу (Gal (α 1-6)-(Gal). Известно, что такой дисахарид (см. Paton, J.S. & Paton A.W. (1989), Clin. Microbial. Revs, 11, 450-479; Carisson, K.A. (1989), Ann. Revievs Biochem, 58, 309-350) обладает антиадгезивной способностью предотвращать адгезию токсинов, например, токсина Shiga и таких патогенов, как E. coli, на стенки пищеварительного тракта.
Штамм B. bifidum депонирован под номером доступа NCIMB 41171 в the National Collection of Industrial & Marine Bacteria, Aberdeen, UK 31 марта 2003 г. Кроме того, он также описан в патенте США No.2412 380.
Было обнаружено, что штамм B. bifidum продуцирует α-галактозидазу, способную превращать мелибиозу в α-галактобиозные дисахариды.
Настоящее изобретению относится к последовательности ДНК, кодирующей белок с аминокислотной последовательностью, приведенной в SEQ ID NO:2 или гибридизирующейся в жестких условиях с последовательностью ДНК, кодирующей рассматриваемый белок. Последовательность ДНК приведена в SEQ ID NO:1 или может содержать фрагменты или вырожденную последовательность.
Термин «вырожденная» относится к последовательности ДНК, которая гомологична последовательности SEQ ID NO:1, по меньшей мере, на 50%, более предпочтительно на 50-98%, наиболее предпочтительно, на 75-95%.
Такая последовательность ДНК может содержать нуклеотидные замены, вставки или делеции, способные изменять аминокислотную последовательность SEQ ID NO:2 менее чем на 60%, предпочтительно, менее чем на 45%, более предпочтительно, менее чем на 25%. Нуклеотидные замены могут давать консервативные замены в аминокислотных последовательностях.
Во втором аспекте настоящее изобретение относится к ферменту, кодируемому указанной выше последовательностью ДНК. Такой фермент может содержать аминокислотную последовательность SEQ ID NO:2 или ее фрагмент.
В третьем аспекте настоящее изобретение относится к рекомбинантному вектору, предпочтительно, экспрессирующему вектору, содержащему указанную выше последовательность ДНК. Такой вектор может вводиться в клетку-хозяин такую, как бактерия, дрожжи или грибки. В качестве альтернативы, последовательность ДНК может быть введена в такую клетку-хозяин. Подходящая клетка-хозин может быть выбрана из Bifidobacterium, Lactococcus, Lactobacillus, Bacillus, например, Bacillus subtilis, или Bacillus circulans, Esherichia и Aspergillus, например, Aspergillus niger.
При использовании в качестве субстрата мелибиозы, фермент, кодируемый указанной выше последовательностью ДНК, продуцирует смесь олигосахаридов, главным образом, α-галактобиозных дисахаридов.
Указанный выше фермент или клетка-хозяин могут использоваться для получения α-галактобиозных дисахаридов, которые могут являться частью продукта, предназначенного для стабилизации деятельности желудочно-кишечного тракта. Продукты такого типа могут быть выбраны из группы, состоящей из молочных продуктов (например, таких как жидкое молоко, сухое молоко, например, сухое цельное молоко, сухое обезжиренное молоко, сухое полножирное молоко, сухая сыворотка, молочные смеси для детского питания, смеси для детского питания, мороженое, йогурт, сыр, ферментированные молочные продукты), а также напитков, таких как фруктовые соки, продукты для детского питания, зерновые, хлеб, крекеры, кондитерские изделия, выпечка, пищевые добавки, диетические добавки, пищевые продукты с пробиотиками, пищевые продукты с пребиотиками, корма для животных, корма для домашней птицы, а также других пищевых продуктов или напитков.
В качестве альтернативы, полученные таким образом дисахариды могут использоваться для получения лекарственных средств в виде таблеток или капсул для предотвращения адгезии на стенках кишечного тракта патогенов или токсинов, продуцируемых патогенами. Лекарственные средства такого типа могут применяться пациентом, например, после курса лечения антибиотиками, которое часто нарушает нормальную флору пищеварительного тракта.
Также настоящее изобретение относится к описанному выше способу получения ферментов, который включает культивирование клеток-хозяев в подходящей культуральной среде в условиях, обеспечивающих экспрессию фермента и извлечение полученного в результате фермента или продуктов ферментации из культуральной среды.
Таким образом, изобретение также относится к способу получения галактобиозных дисахаридов, включающему контактирование описанных выше фермента или клетки-хозяина с материалом, содержащим мелибиозу, в условиях, приводящих к образованию дисахаридов.
Подходящие материалы, содержащие мелибиозу, могут быть выбраны из коммерчески доступных мелибиоз, раффиноз, стахиоз или соевых экстрактов.
Краткое описание фигур
На фигуре 1 приведена нуклеотидная последовательность (SEQ ID NO:1) α-галактозидазы Bifidobacterium bifidum с инициирующим или терминирующим кодоном, указанным жирным курсивом;
на фигуре 2 приведена нуклеотидная последовательность согласно фигуре 1 с аминокислотной последовательностью (SEQ ID NO:2) фермента;
на фигуре 3 приведены первые 540 аминокислот аминокислотной последовательности (SEQ.ID. NO:2) согласно фигуре 2;
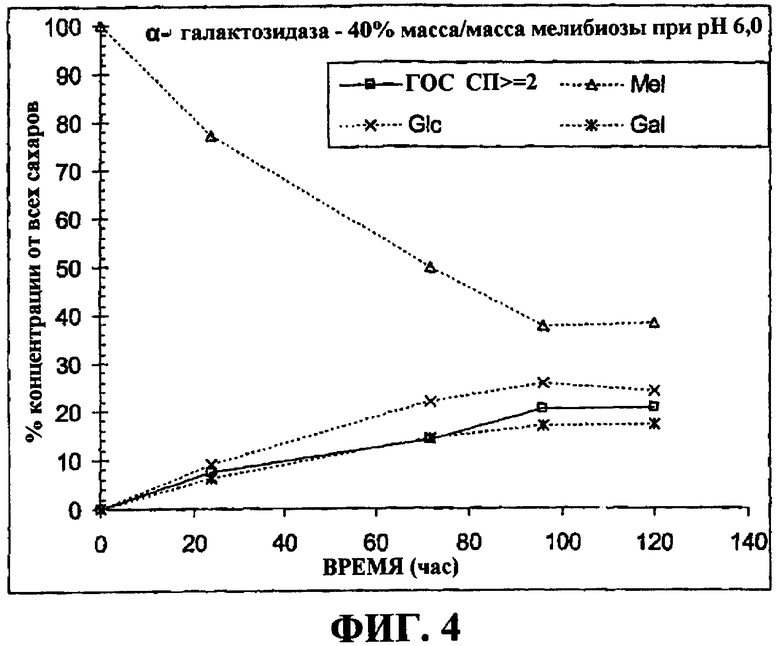
на фигуре 4 приведен график, демонстрирующий кинетику реакции во время синтеза галалактоолигосахарида α-галактозидазой с использованием 40% (мас./мас.) меллибиозы в 0,1M фосфатном буфере в качестве субстрата; и
на фигуре 5 приведена хроматограмма, полученная высокоэффективной анионообменной хроматографией смеси α-галактоолигосахаридов, синтезированной с помощью α-галактозидазы из B. bifidum NCIMB 41171 с использованием 40% (мас./мас.) мелибиозы в 0,1М фосфатном буфере в качестве субстрата при рН 6,0. (Здесь Glc=глюкоза; Gal=галактоза, Mel=мелибиоза; СП=степень полимеризации). Стрелки, обозначенные пунктирными линиями, определяют положение галактоолигосахаридных продуктов.
Геномную ДНК выделяли из штамма Bifidobacterium bifidum (NCIMB 41171) с использованием способа Lawson et al. (1989) Fems Microbiol Letters, 65, (1-2), 41-45. ДНК обрабатывали рестрикционными ферментами, и фрагменты с максимальным размером 15 т.п.н. лигировали вектором PBluescript KS(+). Клетки E.coli трансформировали вектором, содержащим инсерции, состоящие из PstI обработанной ферментами хромосомной ДНК из B. bifidum. Клоны с α-галактозидазной активностью подвергали селекции на пластинах с агаром Luria Bertani, содержащим п-нитрофенил α-D-галактопиронозид и изопропил-β-D-тиогалактозид (IPTG). Идентифицировали два α-галактозидазных положительных клона (pMeIA1 и pMeIA2).
Два полученных положительных клона, обработанные ферментами EcroRI, PstI и Bam HI, продемонстрировали аналогичные рестрикционные карты, что указывает на то, что оба они содержат одинаковые ДНК-вставки. Секвенирование вставленного фрагмента ДНК MelA1 осуществляли с помощью Sanger дидеокси метода обрыва цепи (Russel P., 2002 iGenetics, Pearson Education Inc., San Francisco, 187-189) с использованием набора для циклического секвенирования BigDye Terminator V.3.0. (Applied Biosystems, USA). Последовательность ДНК MelA1 представлена на Фигуре 1 (SEQ ID NO:1).
Открытую рамку считывания (ORF) локализовали с использованием ORF finder от NCBI (National center of Biotechnology information). Использовали генетический код бактерии и определили длину рамки считывания в 300 пн. Нуклеотидную последовательность, показанную на фигуре 1, транслировали в шесть возможных рамок и была выявлена одна открытая рамка считывания из 759 аминокислот, кодирующая предполагаемую α-галактозидазу (трансляция показана на Фигуре 2 SEQ ID NO:2).
Следующие примеры иллюстрируют настоящее изобретение.
Пример 1
Материалы и методы
Все химические реагенты и препараты сред, использованные в этой работе, были получены от Sigma (Dorset, UK), Invitrogen (Paisley, UK), Oxoid (Basingstoke, UK), Qiagen (West Sussex, UK) и Promega (Southmpton, UK).
Бактериальные штаммы
Штамм Bifidobacterium bifidum (NCIMB 41171) хранили на криогенных условиях в пробирках Microbank при -70°С. Для последующих экспериментов штамм оживляли на агаре Wilkinson Chalgren (WC) агаре (Oxoid, UK) и среде TPY (среда, Триптиказа с фитоном и дрожжевым экстрактом) и выращивали анаэробно (состав СО2 и N2 - 80% и 20% соответственно) при 37°С в течение 38 часов. Морфологию колоний и отсутствие загрязнений проверяли окрашиванием по Граму.
Штаммы E.coli
Штамм Eschericia coli RAllr и DH5a, использованный в этой работе, обычно инкубировали в аэробных условиях при 37°С на агаровой среде Лурия-Бертани (LB) или бульоне (Sambrook J. and Russel W.D. (2001) Molecular Cloning: A Laboratory Manual. Cold Spring Harbour Laborotory Press, New York) и при необходимости добавляли антибиотики (100 мкг/мл ампицилина и/или 15 мкг/мл хлорамфеникола) и 40 мкл 2% Х-α-галактопиранозида (Х-α-Gal), 7 мкл 20% раствора IPTG (изопропил-β-D-тиогалактозида), которые наносили на поверхности заранее приготовленных подготовленной 90-мм агаровых чашек.
В экспериментах по экспрессии использовали дефицитный по α-галактозидазе штамм E.coli RAllr (Hanatani et al., 1983, J.Biol. Chem,259(3) (генотип: melA-B+, recA-, lacZ-Y-), полученный из E.coli. K12. Штамм E.coli DH5a (Invitrogen, Paisley, UK) (генотип: F- φ80lacZΔM Δ (lacZYA-argF)U169 recA1 и endA1 hsdR17 (rk -; mk - ) phoA supE44 thi-1 gyrA96 relA1λ-) использовали во всех других генетических экспериментах.
Выбор штаммов E.coli RAllr для экспериментов по экспрессии проводили в соответствии с генотипом. Выбранный штамм не кодирует активную α-галактозидазу за счет melA мутации в его собственной ДНК. Однако рассматриваемый штамм содержит активный переносчик мелибиозы, который необходим для переноса сахаров (мелибиозы) в цитоплазму и, следовательно, для их метаболизма активной α-галактозидазой. На настоящем этапе исследований нельзя сделать вывод о том, протекает ли экспрессия α-галоксидазы Bifidobacterium bifidum внутриклеточно или внеклеточно. Таким образом, наличие активного переносчика мелибиозы является существенным фактором для идентификации α-gal позитивных клонов и, следовательно, для выделения генов, кодирующих α-галактозидазу.
Следует отметить, что в том случае, когда рассматриваемый штамм представляет собой мутант recA, который минимизирует рекомбинацию введенной ДНК под воздействием ДНК хозяина, наблюдается повышение стабильности инсерции.
Экстракция Геномной ДНК из Bifidobacterium bifidum.
Геномную ДНК выделяли из штамма Bifidobacterim bifiidum (NCIMB 411171) с использованием метода Lawson et al. (1989).
Согласно описанному способу клетки собирали с пластин в пробирки Эппендорфа емкостью 1,5 мл, содержащие 0,5 мл TES буфера. Вводили 10 мкл смеси лизозим/мутанолизин (4:1, лизозим 10 мг/мл; мутанолизин 1 мг/мл) и далее полученную смесь перемешивали и инкубировали в течение 30 минут при 37°С. Далее клетки обрабатывали 10 мкл протеиназы К (в концентрации 20 мг/мл) и 10 мкл рибонуклеазы (10 мг/мл), перемешивали и инкубировали в течение 1 часа при 65°С. После инкубации в систему добавляли 100 мкл 10% раствора SDS и клеточные лизаты осторожно перемешивали методом инверсии и инкубировали еще в течение 15 минут при 65°С, после чего добавляли 0,62 мл смеси фенола/хлороформа, перемешивали методом инверсии до образования эмульсии. Далее клеточные лизаты центрифугировали со скоростью 6500 об/мин в течение 10 минут, и верхний слой жидкости переносили в чистую пробирку Эппендорфа, используя для этой операции отожженные ширококанальные пипетки. Экстракцию (этап депротеинизации) повторяли до полного удаления клеточных фрагментов. ДНК осаждали добавлением 1 мл ледяного этанола, после чего проводили инкубацию, по меньшей мере, в течение 30 минут на льду или, сохраняя в течение ночи в морозильнике при температуре -20°С. Геномную ДНК выделяли центрифугированием при скорости 13000 об/мин в течение 5 минут, и после сушки ресуспендировали в 5 мкл стерильной смеси из 1 мМ Трис-CI при рН 8.
Экстрагированную ДНК анализировали методом гель-электрофореза и ее концентрацию измеряли при 260 нм. Хранили при температуре -20°С или -70°С в течение длительного периода времени, избегая многократного оттаивания и замораживания для снижения возможности разрушения.
Векторный ДНК препарат
В этом исследовании использовали pBluescript KS(+) (Stratagen, Norh Torreyv Road). Выбор этого клонирующего средства обусловлен тем, что lac промотор кодирующей плазмиды pBluescript KS(+) необходим для инициации транскрипции генов, теряющих собственный промотор.
Вектор переваривали в присутствии следующих рестрикционных ферментов: PstI, BamHI и EcoRI в соответствии с инструкциями производителя, используя десятикратный избыток фермента относительно количества ДНК (ферментативные единицы: мкг ДНК эквивалентно десяти единицам фермента на один µг плазмидной ДНК или десяти единицам фермента в расчете на 0,5 пикомолей плазмидной ДНК). После термоинактивации фермента (20 минут при 65°С) анализировали карту рестрикции методом горизонтального гель-электрофореза.
Далее векторы дефосфорилировали с использованием кишечной щелочной фосфатазы теленка CIAP (Promega, Southampton, UK) в соответствии с инструкциями производителя. Эффективность обработки тестировали само-лигированием вектора (Bacteriofag T4 ДНК лигазой в соответствии с инструкциями производителя), после чего производили трансформацию DH5a клеток.
Наличие единого фрагмента геля является свидетельством полного переваривания вектора и его единичного рестрикционного переваривания. Степень переваривания вектора тестировали трансформацией не лигированных молекул компетентных клеток E.coli DH5a. Число колоний, образовавшихся на пластинах LB агара, дополнительно содержащих ампициллин (100 мкл/мл), служило показателем числа не переваренных молекул и могло служить фоновым сигналом для последующих экспериментов.
Конструирование библиотеки геномных ДНК
Геномные ДНК частично переваривали в присутствии трех рестрикционных ферментов, способных распознавать часто встречающиеся гекса-нуклеотидные последовательности, в которые включена прокариотная ДНК. EcoRI, BamHI и PstI представляют собой рестрикционные эндонуклеазы типа II специфически распознающие последовательности типа 5'G/AATTC'3, 5'G/GATCC'3 и 5'CTGCA/G'3 соответственно, это явление сопровождается двухнитевым разрывом, приводящим к формированию 5' выступающих концов из четырех нуклеотидов AATT, GATS для EcoRI и BamHI, соответственно, а также 3' выступающих концов при использовании ACGT для PstI.
Было установлено, что все ферменты обладали активностью и были способны расщеплять ДНК только в присутствии ионов двухвалентного магния. Такие ионы служили единственным требующимся кофактором.
Рестрикционное переваривание ДНК
Все рестрикционные перевары образцов геномной ДНК инкубировали в течение 2 часов при 37°С и инактировали при 65°С в течение 20 минут. Затем реакционную смесь охлаждали до комнатной температуры и добавляли необходимое количество буфера, подвергая смесь мягкому перемешиванию с использованием герметизированного стеклянного капилляра. Далее растворы загружали в ампулы с 0,8% агарозного геля (подвод мощности 4-5 В/см в течение 14-16 часов) и размер переваренной ДНК определяли с использованием 1 т.п.н. стандартных ДНК (Promega, UK) (Sambrook, J, Molecular Cloning: A Laboratory Mannual (2002).
Очистка фрагментов, образовавшихся после процесса рестрикционного переваривания
Очистку фрагментов от реакционной смеси и агарозного геля выполняли с использованием набора для экстракции геля QIAEX от Qiagen (West Sussex, UK). Протоколы анализа подробно изложены в инструкциях производителя.
Лигирование и трансформация ДНК
После очистки фрагментов ДНК с использованием набора для экстракции геля QIAEX, материал подвергали лигированию с использованием CIAP-обработанного pBluescript KC(+) вектора. С целью лигирования определенные количества ДНК переносили в стерильные микроцентрифужные пробирки емкостью 0,5 мл, как показано в таблице 1.
А
Количество вектора (15 фемтомолей) (~29,7 нг)
(посторонние вещества в 15 фемтомолях ~69,3 нг)
Перед каждой операцией лигирования фрагменты ДНК нагревали при 45°С в течение 5 минут до плавления всех «липких» концов, которые повторно отжигались при получении фрагмента. Реакцию проводили согласно инструкции Promega для всех реакций лигирования, молярное соотношение вектор:вставка ДНК выбиралось 1:1.
В пробирки А и В вводили по 1,0 мкл 10-кратного избытка лигирующего буфера и 0,5 единиц Weiss T4 ДНК лигазы (Promega, UK), и весь объем реакционной смеси доводили до уровня 10 мкл с помощью биологически очищенной воды. В пробирки С вводили 1,0 мкл 10-кратного избытка лигирующего буфера, и весь объем реакционной смеси доводили до уровня 10 мкл с помощью биологически очищенной воды.
Фрагменты ДНК добавляли в пробирки совместно с водой, после чего смесь нагревали до 45°С в течение 5 минут для плавления всех «липких» концов, которые повторно отжигались при получении фрагмента. ДНК охлаждали до 0°С перед добавлением оставшихся реагентов лигирования и реакционную смесь инкубировали в течение ночи при 16°С (Sambrook and Rusell, 2001).
После осаждения этанолом и очистки лигированных фрагментов (с целью удаления лигирующей среды, которая может понижать эффективность трансформационных превращений) дальнейшие операции проводили, следуя инструкциям Hanan. ~50 нг лигированной ДНК в 5 мкл раствора добавляли к 100 мкл компетентных E. Coli RAllr клеток. После термообработки и экспрессии в присутствии ампициллин-устойчивого гена клетки распределяли по поверхности LB пластин, содержащих ампициллин (100 мкг/мл), X-α-Gal (40 мкл 2% Х-α-Gal) и (7 мкл 20% IPTG).
Измеряли число трансформаций в каждой реакции лигирования. Число трансформантов в пробирке С составляло 2×105-1×106 КОЕ/мкг, в то время как соответствующее количество в пробе В составляло 500-600 КОЕ/мкг.Число трансформантов в пробе А отражает эффективность обработки векторной ДНК. Число трансформантов в пробе В составляло 2-4×104 КОЕ/мкг.
Число трансформантов
При лигировании смесей в присутствии PstI хромосомной ДНК получали два α-галктозидазных положительных клона (pMelA1 и PmelA2) из около 2500 скринированных трансформантов, тогда как в присутствии хромосомной ДНК, обработанной EcoRI и BamHI, не наблюдалось образования какого-либо положительного клона из около 4000 всех скринированных трансформантов.
Переваривание положительного клона
Два положительных PstI клона переваривали рестрикционными ферментами EcroRI, PstI, BamHI, HindIII, SmaI и KpnI. Рестрикционные ферменты EcroRI, PstI и BamHI показали аналогичную рестрикционную карту, один фрагмент ~5 т.п.н. (целевой ген) и другой ~ около 3 т.п.н. (плазмидная ДНК), что свидетельствует о том, что указанные ферменты режут в аналогичных позициях. HindIII дают фрагмент размером 6,5 т.п.н. и фрагмент размером 1,5 т.п.н., тогда как ферменты SmaI и KpnI образуют лишь один фрагмент размером ~8 т.п.н., что свидетельствует о разрезании лишь в одной позиции. Аналогичные рестрикционные карты для обеих плазмид указывают на то, что обе содержат аналогичные ДНК вставки.
Секвенирование последовательности ДНК
Секвенирование ДНК осуществляли с использованием набора для циклического секвенирования BigDye Terminator V.3.0 (Applied Biosystems, USA) и анализ проводили с использованием ABI Prism 3100 системы для флуоресцентного анализа ДНК, включающей методы капиллярного электрофореза.
5'- и 3'-концы встроенных фрагментов ДНК секвенировали вектор-специфическими праймерами. Вставки дополнительно секвенировали с использованием системы геномного праймирования (Genome Priming System (GPS-I)) (New England Biolabs, Uk). GPS-1 представляет собой in vitro систему на основе транспозона TN7, который использует TnsABC транспозазу для встраивания случайным образом в целевую ДНК. Донор: была использована целевая ДНК в массовом соотношении 1:4 согласно инструкции производителя. Количество выделенных плазмид для секвенирования после вставки транспраймера в целевую плазмиду составило 25. Это количество было высчитано согласно инструкциям производителя и исходя из 5-кратной глубины охвата it assumes a 5 -fold depth of coverage.
Уникальные сайты праймирования на обоих концах транспраймерного элемента позволяют секвенировать обе нити целевой ДНК в позиции вставки.
Смесь для реакции секвенирования содержала около 400-600 нг плазмидной ДНК, 3,2 пикомоля раствора праймера и 4 мкл раствора терминатора BigDye.
Идентификация открытой рамки считывания
Открытую рамку считывания (ORF) выявляли с использованием ORF finder от NCBI. Использовали генетический код бактерии и определили длину рамки считывания в 300 пн. Нуклеотидную последовательность транслировали в шесть возможных рамок и была выявлена одна открытая рамка считывания из 759 аминокислот, кодирующая предполагаемую α-галактозидазу (трансляция показана на фигуре 2). Были подтверждены инициирующий и терминирующий кодоны.
Ген α-галактозидазы Bifidobacterium pMelAl плазмидой был экспрессирован в Е.coli при условиях роста, которые в норме подавляют экспрессию с индуцибельным Е.coli lacZ промотором, расположенным во фланкирующей области клонирующего вектора. Эти результаты указывают, что эндогенные, внутренние бифидобактериальные последовательности, направленные против гена α-галактозидазы, могут служить сигналом инициации транскрипции в Е.coli.
Начало транскрипции указанно жирным курсивом. Приведенные выше результаты указывают, что ген контролируется собственным промотором транскрипции.
Пример 2
Синтез с помощью клонированного фермента а-галактозидазы,
выделенный из Bifidobacterium bifidum NCIMB 41171 в клетке-хозяине Е.coli (штамм RA11).
Синтез, описанный ниже, проводили, если не указанно иначе, с целыми клетками Е.coli RAll в качестве хозяина после обработки биомассы E.coli (собранной центрифугированием при 10000 g) толуолом в концентрации 2000 чнм, для увеличения проницаемости клетки, чтобы сделать ее нежизнеспособной, разрушив ее цитоплазматическую мембрану. Биомассу E.coli готовили, как описано в п.«Штаммы Е coli» Примера 1.
Синтез с помощью клонированного фермента
Синтез с помощью α-галактозидазы проводили при начальной концентрации мелибиозы в субстрате 40% (мас./мас.). Синтезирующий раствор получали в 0,1 М фосфатном буфере с рН 6,0. Синтез проводили при температуре 40°С на водной бане-шейкере при 150 об/мин. Оптимальные рН для конкретного фермента выбирали, исходя из измерений активности (с использованием в качестве субстрата р-нитрофенил-α-D-галактопиранозида) специфического ферментного препарата при различных показателях рН.
Для синтеза с помощью α-галактозидазы 2 мл клеточной суспензии Е.coli RAll (с активностью 0,3 ед./мл) центрифугировали (при 10000 g) для сбора биомассы, и супернатант выгружали. Эту биомассу ресуспендировали 1 г 40% (мас./мас.) раствора субстрата мелибиозы для проведения синтеза. На Фигуре 4 показана концентрация различных сахаров, присутствующих в смеси во время синтеза. На фигуре 5 показаны хроматограммы галактоолигосахаридных смесей, синтезированных α-галактозидазой, клонированной из В.bifidum NCIMB 41171, полученные высокоэффективной анионообменной хроматографией совместно с импульсным амперометрическим детектированием (HPAEC-PAD). В Таблице 2 приведены концентрации сахаров галактоолигосахаридной смеси в момент времени оптимального синтеза.
Углеводная композиция при синтезе α-галактоолигосахаридов при 40% (мас./мас.) начальной концентрации мелибиозы в момент времени, когда наблюдается максимальная
концентрация олигосахаридов











Изобретение относится к биотехнологии и представляет собой новую альфа-галактозидазу, молекулу ДНК, ее кодирующую. Также изобретение относится к вектору экспрессии, содержащему такую молекулу ДНК, а также к клетке, трансформированной вектором. Изобретение также касается способа получения альфа-галактобиозных дисахаридов с использованием новой альфа-галактозидазы. Изобретение позволяет получать альфа-галактобиозные дисахариды с высокой степенью эффективности. 10 н. и 5 з.п. ф-лы, 5 ил., 2 табл.
1. Молекула ДНК, кодирующая α-галактозидазу, имеющая последовательность, которая
a) кодирует белок с аминокислотной последовательностью, представленной SEQ ID NO: 2, или
b) гибридизуется в жестких условиях гибридизации с последовательностью а), или
c) представляет собой вырожденную последовательность а) или b), которая на 75-95% гомологична им.
2. Молекула ДНК по п.1, где последовательность представляет собой SEQ ID NO: 1 или ее фрагмент или вырожденную последовательность, которая на 75-95% ей гомологична.
3. Молекула ДНК по п.1 или 2, где полученная из указанной последовательности вырожденная последовательность включает нуклеотидные замены, вставки или делеции, в результате которых менее чем 25% изменено в аминокислотной последовательности SEQ ID NO: 2 или ее фрагмент.
4. Молекула ДНК по п.1 или 2, где полученная из указанной последовательности вырожденная последовательность содержит нуклеотидные замены, представляющие собой консервативные аминокислотные замены.
5. α-галактозидаза, кодируемая ДНК последовательностью по любому из пп.1-4.
6. α-галактозидаза, содержащая аминокислотную последовательность SEQ ID NO: 2 или ее функциональный фрагмент.
7. Рекомбинантный вектор экспрессии, включающий последовательность ДНК по любому из пп.1-4.
8. Бактериальная клетка-хозяин для экспрессирования α-галактозидазы по п.5 или 6, содержащая последовательность ДНК по любому из пп.1-4.
9. Бактериальная клетка-хозяин для экспрессирования α-галактозидазы по п.5 или 6, содержащая вектор по п.7.
10. Клетка-хозяин по п.8 или 9, где указанная клетка выбрана из группы, состоящей из Bifidobacterium, Lactococcus, Lactobacillus, Escherichia, Bacillus и Aspergillus.
11. Клетка-хозяин по п.10, где указанная клетка выбрана из группы, состоящей из Bifidobacterium bifidum, Bacillus subtilis, Bacillus circulans и Aspergillus niger.
12. Применение α-галактозидазы по любому из пп.5 или 6 или клетки по любому из пп.8-11 для получения α-галактобиозных дисахаридов.
13. Применение α-галактозидазы по любому из пп.5 или 6 или клетки по любому из пп.8-11 для получения α-галактобиозных дисахаридов, являющихся частью продукта, выбранного из группы, состоящей из молочных продуктов, таких как жидкое молоко, сухое молоко, детские молочные смеси, смеси для детского питания, мороженое, йогурт, сыр, ферментированные молочные продукты, напитков, таких как фруктовые соки, продуктов для детского питания, зерновых, хлеба, печенья, кондитерских изделий, выпечки, пищевых добавок, диетических добавок, пищевых продуктов с пробиотиками, пищевых продуктов с пребиотиками, кормов для животных, кормов для домашней птицы и лекарственных средств.
14. Способ получения α-галактозидазы по любому из пп.5 или 6, включающий культивирование клетки-хозяина по любому из пп.8-11 в подходящей культуральной среде при условиях, позволяющих экспрессировать указанный фермент, и извлечение полученной в результате этого α-галактозидазы из культуральной среды.
15. Способ получения α-галактобиозного дисахарида, включающий приведение α-галактозидазы по любому из пп.5 или 6 или клетки-хозяина по любому из пп.8-11 в контакт с раствором мелибиозы.
| DATABASE UNIPROT, Q9XCX2, 01.11.1999, - Найдено в Интернет http://www.ebi.ac.uk./ | |||
| VAN DEN BROEK, L.A.M., et.al | |||
| Synthesis of alpha-galacto-oligosaccharides by a cloned alpha-galactosidase from Bifidobacterium adolescentis | |||
| Biotechnol | |||
| Lett | |||
| Металлический водоудерживающий щит висячей системы | 1922 |
|
SU1999A1 |
| КОНСТРУКЦИЯ УПЛОТНИТЕЛЬНОГО УЗЛА ДЛЯ ГИДРО- ИЛИ ПНЕВМОПРИВОДА | 2007 |
|
RU2412380C2 |
| ШТАММ БАКТЕРИЙ PSEUDOALTEROMONAS SP. - ПРОДУЦЕНТ АЛЬФА-ГАЛАКТОЗИДАЗЫ | 1997 |
|
RU2113479C1 |

