ПРИТЯЗАНИЕ НА ПРИОРИТЕТ
По настоящей заявке испрашивается приоритет временной заявки на патент США с серийным номером 62/522564, поданной 20 июня 2017 г., которая полностью включена в настоящее описание в качестве ссылки.
УРОВЕНЬ ТЕХНИКИ
1. Область техники
Настоящее изобретение, в целом, относится к областям медицины, онкологии и иммунотерапии. Более конкретно, оно относится к разработке и применению иммунореагентов для лечения рака.
2. Уровень техники
Считается, что интерфероны I типа (IFN) непосредственно подавляют пролиферацию опухолевых клеток. Они нашли успешное применение в лечении нескольких типов рака, включая гематологические опухоли (хронический миелоидный лейкоз, волосатоклеточный лейкоз, множественную миелому и неходжкинские лимфомы), а также солидные опухоли (меланому, рак почек и саркому Капоши) (Ferrantini et al., 2007, Moschos & Kirkwood, 2007, Zitvogel et al., 2015, и Antonelli et al., 2015). Действительно, согласно накопившимся данным, эндогенный IFN I типа играет ключевую роль в «обучении» дендритных клеток (DC) для повышения эффективности кросс-праймирования против опухолевых антигенов для усиления противоопухолевой активности при химиотерапии, радиотерапии, таргетной терапии и иммунотерапии (Schiavoni et al., 2011, Burnette et al. al., 2011; Stagg et al., 2011 и Woo et al., 2014).
Особое преимущество лечения с использованием IFN I типа обусловлено его способностью включаться во многих точках генерации противоопухолевых иммунных ответов, включая стимуляцию популяций врожденных и адаптивных цитотоксических лимфоцитов, негативную регуляцию клеток супрессорного типа, влияние на опухолевые клетки путем ингибирования пролиферации и модуляции апоптоза, дифференцировки, миграции и экспрессии антигена на клеточной поверхности (Parker et al., 2016). Важным является то, что некоторые из этих действий могут представлять потенциальную стратегию по использованию IFN I типа для преодоления устойчивости рака к иммунотерапии. Одним из механизмов рецидива у больных раком является неспособность Т-клеток к распознаванию из-за подавления экспрессии молекул МНС класса I и генов транспортеров пептидов для презентации опухолевого антигена (Sharma et al., 2017). IFN I типа можно использовать для индукции экспрессии MHC класса I, такой как экспрессия LMP2/7 и TAPА-1/2 в опухолевых клетках, что в совокупности является идеальной стратегией комбинированной терапии в борьбе с устойчивостью к иммунотерапии (Khanna, 1998). Кроме того, IFN I типа можно использовать для подавления Treg и MDSC путем негативной регуляции пролиферации Treg (Pace et al., 2010 и Srivastava et al., 2014) и уменьшения накопления MDSC и снижения их подавляющей функции (Zoglmeier et al., 2011), причем и те и другие непосредственно подавляют активность цитотоксических Т-лимфоцитов (Schmidt et al., 2012 и Gabrilovich et al., 2009). Эти многочисленные противоопухолевые эффекты вызывают интерес к IFN I типа как противораковым лекарственным средствам как в случае монотерапии, так и в случае их применения в комбинации с другими видами терапии.
Тем не менее, одним из самых больших препятствий для использования IFN I типа в клинике являются серьезные побочные эффекты, связанные с таким лечением. Наиболее часто встречающимися побочными эффектами являются гриппоподобные симптомы, гематологическая токсичность, повышение уровня трансаминаз, тошнота, усталость и психические последствия. Эти побочные эффекты препятствуют достижению доз, необходимых для получения максимального терапевтического эффекта, и их поддержания, поскольку появление этих побочных эффектов может полностью обесценить клиническую пользу от лечения IFN I типа (Kreutzer et al., 2004, Sleijfer et al., 2005 и Lotrich, 2009). Следовательно, возможность обеспечения специфической доставки IFN I типа в микроокружение опухоли является необходимым условием для продолжительного клинического применения IFN I типа. Необходимы стратегии модификации IFN I типа для получения новых форм лекарственного средства, активность которых проявлялась бы только внутри опухоли, а также для предотвращения серьезных побочных эффектов за пределами опухоли.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Таким образом, настоящее изобретение относится к пролекарству интерферона, содержащему (а) домен рецепторов интерферонов альфа и бета (IFNAR), который сохраняет IFN-связывающую активность; (b) домен интерферона (IFN) 1 типа, который сохраняет активность интерферона 1 типа, когда не связан с указанным доменом IFNAR; (c) Fc-домен иммуноглобулина (Ig), (d) первый линкер, слитый на одном конце с N-концом указанного IFN и слитый на другом конце с указанным IFNAR, причем указанный первый линкер является расщепляемым протеазой; и (e) второй линкер, слитый на одном конце с C-концом указанного IFN и слитый на другом конце с N-концом указанного Fc-домена Ig. Ig может представлять собой IgG, такой как IgG1 или IgG2. Пролекарство интерферона может содержать две копии указанного домена IFN 1 типа. Пролекарство интерферона может содержать более двух копий указанного домена IFN 1 типа. Первый линкер может быть расщепляемым одной или более матриксными металлопротеиназами, такими как MMP1, MMP3, MMP9, MMP10, MMP11, MMP12, MMP13 или MMP14. Первый линкер может быть расщепляемым UPA, FAPa и/или катепсином B. Линкер может представлять собой G4S-SUB1-G4S-SUB2-G4S-SUB3-G4S, где SUB1-3 представляют собой разные сайты расщепления ферментами. IFNAR может представлять собой IFNAR1 или IFNAR2. IFN может представлять собой IFN-α, IFN-β, IFN-κ, IFN-δ, IFN-ε, IFN-τ, IFN-ω или IFN-ζ.
В другом варианте осуществления изобретение относится к конструкции нуклеиновой кислоты, кодирующей пролекарство интерферона, содержащей (а) домен рецепторов интерферонов альфа и бета (IFNAR), который сохраняет IFN-связывающую активность; (b) домен интерферона 1 типа (IFN), который сохраняет активность интерферона 1 типа, когда не связан с указанным доменом IFNAR; (c) Fc-домен иммуноглобулина (Ig), (d) первый линкер, слитый на одном конце с N-концом указанного IFN и слитый на другом конце с указанным IFNAR, причем указанный первый линкер является расщепляемым протеазой; (e) второй линкер, слитый на одном конце с C-концом указанного IFN и слитый на другом конце с N-концом указанного Fc-домена Ig; и (f) промотор, расположенный 5’-концом к 5'-концу указанного домена IFNα. Ig может представлять собой IgG, такой как IgG1 или IgG2. Пролекарство интерферона содержит две копии указанного домена IFN 1 типа. Пролекарство интерферона может содержать более двух копий указанного домена IFN 1 типа. Первый линкер может быть расщепляемым матриксной металлопротеиназой, такой как MMP1, MMP3, MMP9, MMP10, MMP11, MMP12, MMP13 и/или MMP14. Первый линкер может быть расщепляемым UPA, FAPa и/или катепсином B. Линкер может представлять собой G4S-SUB1-G4S-SUB2-G4S-SUB3-G4S, где SUB1-3 представляют собой разные сайты расщепления ферментами. IFNAR может представлять собой IFNAR1 или IFNAR2. IFN может представлять собой IFN-α, IFN-β, IFN-κ, IFN-δ, IFN-ε, IFN-τ, IFN-ω или IFN-ζ.
Изобретение также относится к рекомбинантной клетке, экспрессирующей определенное выше пролекарство интерферона; рекомбинантной клетке, содержащей определенную выше конструкцию нуклеиновой кислоты; способу экспрессии пролекарства интерферона, включающему культивирование определенной выше клетки; способу экспрессии пролекарства интерферона, включающему культивирование определенной выше клетки; и применению определенного выше пролекарства интерферона (а) в получении лекарственного средства для лечения рака; или (b) для лечения рака.
В другом варианте осуществления изобретение относится к способу лечения рака, включающему введение пациенту определенного выше пролекарства интерферона. Способ может дополнительно включать этап оценки экспрессии протеазы в раковой клетке, полученной от указанного пациента. Раковая клетка может быть получена из биопсии или может быть циркулирующей опухолевой клеткой. Рак может представлять собой рак легких, рак молочной железы, рак головного мозга, рак ротовой полости, рак пищевода, рак головы и шеи, рак кожи, рак желудка, рак печени, рак поджелудочной железы, рак почки, рак яичников, рак предстательной железы, рак мочевого пузыря, рак толстой кишки, рак яичка, рак матки, рак шейки матки, лимфому или лейкоз. Рак может быть первичным, рецидивирующим, метастатическим или устойчивым ко многим лекарственным веществам. Пациент мог ранее пройти хирургическое лечение, химиотерапию, лучевую терапию, гормональную терапию или иммунотерапию.
Способ может дополнительно включать получение указанным пациентом второй терапии рака, такой как хирургическая терапия, химиотерапия, лучевая терапия, гормональная терапия или иммунотерапия. Пациент может представлять собой человека или млекопитающее, не являющееся человеком. Способ может дополнительно включать введение указанного пролекарства интерферона более одного раза, например ежедневно, через день, еженедельно, через неделю или ежемесячно. Пролекарство можно вводить системно или внутрь опухоли, локально в опухоль или регионарно по отношению к опухоли. Лечение может включать одно или более из: замедления роста опухоли, остановки роста опухоли, уменьшения размера или объема опухоли, увеличения выживаемости по сравнению с пациентом, не принимавшим лечение, индукции ремиссии рака, индукции апоптоза опухолевых клеток или индукции некроза опухоли.
Предполагается, что любой способ или композиция, описанные в настоящей заявке, могут быть реализованы по отношению к любому другому способу или композиции, описанной в настоящей заявке.
Использование единственного числа в сочетании с термином «содержащий» в формуле изобретения и/или описании может означать «один», но также подразумевает «один или более», «по меньшей мере один» и «один или более одного». Слово «примерно» означает плюс или минус 5% от указанного числа.
Другие объекты изобретения, признаки и преимущества настоящего изобретения станут очевидными из приведенного ниже подробного описания. Однако следует понимать, что подробное описание и конкретные примеры, хотя и указывают на конкретные варианты осуществления изобретения, приведены исключительно в качестве иллюстрации, поскольку из подробного описания специалистам в данной области техники будут очевидны различные изменения и модификации, находящиеся в пределах сущности и объема изобретения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Приведенные ниже чертежи являются частью настоящего описания и включены с целью дополнительной демонстрации некоторых аспектов настоящего раскрытия. Настоящее изобретение может быть лучше понято со ссылкой на один или более из этих чертежей в сочетании с подробным описанием конкретных вариантов осуществления, представленных в настоящем описании.
Фиг. 1. Схематическая структура IFN-пролекарства. IFNAR-ECD (синий), линкер, являющийся субстратом (красный), IFN (желтый), опухолеспецифический фермент (зеленый), Fc IgG (серый). Левое N-концевое плечо представляет собой интактную форму IFN-пролекарства, слитую с IFNAR-ECD и связанную с IFN, тогда как правое N-концевое плечо представляет собой активированную форму IFN-пролекарства, от которого отщеплен IFNAR-ECD.
Фиг. 2А-С. Активированная форма IFN-пролекарства демонстрирует более высокую активность IFN, чем восстановленная. (Фиг.2А) Выполняли 5-кратное серийное разведение 20 нМ hIg, IFNa4-Fc, IFN-пролекарства на основе IFNAR1 и IFN-пролекарства на основе IFNAR2. (Фиг. 2B-C) IFN-пролекарство разбавляли до 1 мкМ в буфере для анализа (50 мМ Трис, 10 мМ CaCl2, 150 мМ NaCl, 0,05% Brij-35 (мас./об.), pH 7,5 (TCNB)). Добавляли rmMMP-9 до конечной концентрации 1 нг/мкл и инкубировали при 37°C в течение 6 часов. (Фиг.2B) Выполняли 5-кратное серийное разведение 20 нМ IFNa4-Fc, R1-NSUB без или с обработкой rmMMP-9 и R1-SUB (субстрат MMP-2/9) без или с обработкой rmMMP-9. (Фиг.2C) Выполняли 5-кратное серийное разведение 20 нМ IFNa4-Fc, R2-NSUB (субстрат MMP-2/9) без или с обработкой rmMMP-9 и R2-SUB без или с обработкой rmMMP-9. Разведенные растворы слитых белков добавляли в репортерные клетки RAW-Lucia-ISG для стимуляции секреции люциферазы. Кондиционированные супернатанты собирали через 24 ч после стимуляции для анализа активности люциферазы. Планки погрешности представляют средние значения ± стандартная ошибка среднего (SEM) для трех повторов.
Фиг. 3. IFN-пролекарство на основе IFNAR2 имеет более сильный противоопухолевый эффект по сравнению с IFN-пролекарством на основе IFNAR1. Мышам C57BL/6 (n=5 на группу) инъецировали подкожно 5×105 клеток B16 и вводили внутрибрюшинно 1 нмоль hIg, IFNa4-Fc, R1-SUB (субстрат MMP2/9) или R2-SUB (субстрат MMP2/9) в дни 11, 15 и 21. Рост опухоли отслеживали два раза в неделю и регистрировали в виде среднего размера опухоли ± стандартная ошибка среднего в зависимости от времени.
Фиг. 4A-D. Экспрессия ферментов в мышиной нормальной и опухолевой тканях. Мышам C57BL/6 (n=4) инъецировали подкожно 1×106 клеток МС38 или 5×105 клеток В16. Указанные нормальные ткани и опухоли собирали в день 11. Внутриклеточную РНК экстрагировали для анализа методом ОТ-кПЦР (RT-qPCR) для определения количества копий мРНК (фиг. 4A) uPA, (фиг. 4B) MMP-2, (фиг. 4C) MMP-9 и (фиг. 4D) ММР-14. Результаты представлены в процентах относительно 18sr. Планки погрешности представляют средние значения ± стандартная ошибка среднего для трех повторов.
Фиг. 5A-E. IFN-пролекарство повышает безопасность без ущерба эффективности. Мышам C57BL/6 (n=3 на группу) инъецировали подкожно 5×105 клеток B16 и вводили внутрибрюшинно 1 мМ hIg, IFNa4-Fc или R2-SUB (субстрат ММР-14) в дни 10, 13 и 16. У мышей брали кровь и собирали сыворотку в день 17 (фиг. 5А). Концентрацию воспалительных цитокинов IL-6, TNF, MCP-1 и IFN-g в сыворотке крови у мышей с воспалением измеряли с помощью цитометрической матрицы гранул (CBA). (Фиг. 5В) Активность ALT в сыворотке измеряли с помощью системы Reflotron Plus®. (Фиг. 5C) Рост опухоли и (фиг. 5D) массу тела регистрировали два раза в неделю. (Фиг. 5Е) Строили кривую выживания, обусловленную токсичностью. Мыши с потерей массы тела более 30% считались мертвыми. Планки погрешности представляют средние значения ± стандартная ошибка среднего.
Фиг. 6A-Н. Уровень экспрессии человеческих протеаз между опухолью и соседними нормальными тканями. Модуль DiffExp TIMER (Tumor IMmune Esribution Resource Resource) на веб-сайте для онлайн анализа сравнения уровней экспрессии генов для всех образцов из TCGA (Атлас генома рака). (Фиг. 6A) MMP-1, (фиг. 6B) MMP-3, (фиг. 6C) MMP-9, (фиг. 6D) MMP-10, (фиг. 6E) MMP-11, (фиг. 6F) MMP-12, (фиг. 6G) MMP-13 и (фиг. 6H) MMP-14.
Фиг. 7А-С. Активация человеческого ProIFN in vitro. (Фиг.7A) Выполняли 10-кратное серийное разведение 50 мкМ hIFNa2-Fc, ProIFN на основе hIFNAR1 и ProIFN на основе hIFNAR2. Активность IFN измеряли с помощью репортерных клеток 293T-DualТМ hSTING-R232. (Фиг. 7В) Человеческий ProIFN на основе hIFNAR2 с субстратом (hProIFN-Sub) или без него (hProIFN-Nsub) разбавляли до 1 мкМ в буфере для анализа (50 мМ Трис, 10 мМ CaCl2, 150 мМ NaCl, 0,05% Brij-35 (мас./об.), рН 7,5 (TCNB)). Добавляли rmMMP-9 до конечной концентрации 1 нг/мкл и инкубировали при 37°C в течение 6 часов. Активность IFN измеряли с помощью репортерных клеток 293T-DualТМ hSTING-R232. (Фиг.7C) Человеческий hProIFN-Sub на основе hIFNAR2 или hProIFN-Nsub разбавляли до 1 мкМ в буфере для анализа (50 мМ Трис, 10 мМ CaCl2, 150 мМ NaCl, 0,05% Brij-35 (мас./об.), pH 7,5 (TCNB)). Добавляли rmMMP-9 до конечной концентрации 1 нг/мкл и инкубировали при 37°С в течение 0, 0,5, 2 или 6 часов. Расщепленные образцы разделяли на геле Stain FreeТМ. Гель визуализировали с помощью устройства для визуализации без красителя. Планки погрешности представляют средние значения ± стандартная ошибка среднего для трех повторов.
ОПИСАНИЕ ИЛЛЮСТРАТИВНЫХ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
Пролекарства представляют собой фармакологически неактивные химические производные лекарственных веществ, которые должны пройти трансформацию в организме для перехода в активное состояние. Они предназначены для преодоления фармацевтических и/или фармакокинетических проблем, связанных с исходным лекарственным веществом, которые в противном случае ограничивали бы клиническую применимость этого лекарственного вещества (Stella et al., 1985). Недавно было сконструировано активируемое протеазой антитело (про-антитело), нацеленное на молекулу адгезии сосудистого эндотелия 1 типа (VCAM-1), маркер атеросклеротических бляшек, путем присоединения к антителу пептида, маскирующего связывающий участок, через линкер, восприимчивый к матричной металлопротеазе (ММР). Активность таких ассоциированных с заболеванием протеаз можно использовать для сайт-специфического нацеливания активности антител in vivo (Erster et al., 2012). В одном из примеров терапии рака идентифицированный с помощью скрининга пептид, который может блокировать связывание анти-EGFR с EGFR мишенью, связывали с анти-EGFR антителом. Полученное про-антитело, направленное на рецептор эпидермального фактора роста (EGFR), имело заметно улучшенную безопасность и увеличенный период полураспада у приматов, не являющихся человеком, что обеспечивало безопасное введение гораздо более высоких доз этого вещества по сравнению с цетуксимабом (Desnoyers et al., 2013). Однако такие пептиды часто имеют слабое сродство, что приводит к неполной блокаде лекарственного белка, а также к высокой иммуногенности, которая препятствует проведению более длительного лечения. Поэтому авторы изобретения разработали новую стратегию, основанную на природном рецепторе с надлежащим сродством, без иммуногенности у хозяина, которая позволяет подавлять связывание лекарственного вещества со своим рецептором в неопухолевых тканях. В частности, упор сделан на интерферонах I типа в связи с их важной ролью в противоопухолевом иммунном ответе на многие виды рака8.
Авторами изобретения сконструировано IFN-пролекарство с использованием константных областей иммуноглобулина (IgG) и внеклеточных доменов IFNAR (IFNAR1 или IFNAR2) в качестве блокатора активности IFN, слитых на N-конце с IFN. IFNAR связан со специфическим линкером, который селективно расщепляется протеазой, сверхэкспрессируемой в микроокружении опухоли. Это позволяет маскировать токсическую активность домена IFN до тех пор, пока пролекарство не достигнет опухоли, в то время как фрагмент Fc, слитый с C-терминальным концом IFN, улучшает его период полувыведения in vivo. Такое IFN-пролекарство обладает сильно уменьшенной активностью IFN до момента отщепления линкера, при этом указанная активность восстанавливается после ферментативного расщепления. Авторы изобретения также продемонстрировали эффективность и повышенную безопасность IFN-пролекарства in vivo на мышиной модели меланомы B16-OVA.
Таким образом, преимуществом применения этого IFN-пролекарства в качестве противоракового терапевтического средства является: 1) низкая токсичность; 2) неожиданно более высокая активность IFN в активированной форме по сравнению с исходным IFN-Fc; 3) простота получения и очистки с высоким выходом; 4) специфическое нацеливание на опухолевые ткани; 5) использование неиммуногенного блокирующего реагента; и 6) персонализированный дизайн IFN-пролекарства в зависимости от уровня экспрессии опухолевого фермента, который может различаться. Таким образом, такое IFN-пролекарство может улучшить профиль безопасности интерферонов без ущерба для их эффективности и, таким образом, может обеспечить более широкое применение современных форматов интерфероновой терапии, таких как биспецифические слитые белки антитело-цитокин. Эти и другие аспекты изобретения более подробно описаны ниже.
I. Интерфероны типа 1
А. Типы интерферона
Интерфероны I типа (IFN) человека представляют собой большую подгруппу белков, интерферонов, которые помогают регулировать активность иммунной системы. Интерфероны связываются с рецепторами интерферонов. Все IFN I типа связываются со специфическим рецепторным комплексом на клеточной поверхности, известным как рецептор IFN-α (IFNAR), который состоит из цепей IFNAR1 и IFNAR2. IFN I типа обнаружены у всех млекопитающих, и гомологичные (сходные) молекулы обнаружены у видов птиц, рептилий, земноводных и рыб. Типы у млекопитающих обозначены как: IFN-α (альфа), IFN-β (бета), IFN-κ (каппа), IFN-δ (дельта), IFN-ε (эпсилон), IFN-τ (тау), IFN-ω (омега) и IFN-ζ (дзета, также известный как лимитин).
Белки IFN-α продуцируются лейкоцитами. Они в основном участвуют во врожденном иммунном ответе против вирусной инфекции. Гены, ответственные за их синтез, входят в 13 подтипов, которые известны как IFNA1, IFNA2, IFNA4, IFNA5, IFNA6, IFNA7, IFNA8, IFNA10, IFNA13, IFNA14, IFNA16, IFNA17, IFNA21. Эти гены находятся вместе в виде кластера на хромосоме 9.
IFN-α также получают синтетически в качестве лекарственного средства для лечения волосатоклеточного лейкоза. Международное непатентованное наименование (МНН, INN) продукта - интерферон альфа. Рекомбинантный тип - интерферон альфакон-1. Пегилированные типы представляют собой пегилированный интерферон альфа-2а и пегилированный интерферон альфа-2b.
Белки IFN-β продуцируются в больших количествах фибробластами. Они обладают противовирусной активностью, которая задействована в основном во врожденном иммунном ответе. Описаны два типа IFN-β: IFN-β (IFNB1) и IFN-β3 (IFNB3) (ген, обозначенный IFN-β2, фактически является IL-6). IFN-β1 используется для лечения рассеянного склероза, поскольку он снижает частоту рецидивов. IFN-β1 не подходит для лечения пациентов с прогрессирующими нерецидивными формами рассеянного склероза.
IFN-ε, -κ, -τ и -ζ у человека, по-видимому, составляют одну изоформу, IFNK. Только у жвачных животных кодируется IFN-τ, вариант IFN-ω. К настоящему времени IFN-ζ обнаружен только у мышей, в то время как структурный гомолог IFN-δ обнаружен у широкого круга плацентарных млекопитающих, не являющихся приматами и грызунами. У большинства, но не у всех плацентарных млекопитающих, происходит кодирование функциональных генов IFN-ε и IFN-κ.
IFN-ω, хотя и имеет только одну функциональную форму, описанную на сегодняшний день (IFNW1), у людей имеется несколько псевдогенов: IFNWP2, IFNWP4, IFNWP5, IFNWP9, IFNWP15, IFNWP18 и IFNWP19. У многих плацентарных млекопитающих, не являющихся приматами, экспрессируется множество подтипов IFN-ω.
IFN-ν недавно был описан как псевдоген у человека, но потенциально является функциональным в геноме домашней кошки. Во всех других геномах некошачьих плацентарных млекопитающих IFN-ν является псевдогеном; у некоторых видов этот псевдоген хорошо сохранился, в то время как у других он подвергся сильной трансформации или является недетектируемым. Более того, в геноме кошки промотор IFN-ν несет вредные мутации. Вполне вероятно, что семейство генов IFN-ν стало бесполезным до появления разнообразных видов млекопитающих. Его присутствие на краю локуса IFN I типа у млекопитающих, возможно, защитило его от уничтожения, что и привело к возможности его обнаружения.
IFN-α и IFN-β секретируются многими типами клеток, включая лимфоциты (NK-клетки, B-клетки и T-клетки), макрофаги, фибробласты, эндотелиальные клетки, остеобласты и другие. Они обеспечивают стимуляцию как макрофагов, так и NK-клеток, которые вызывают противовирусный ответ, а также проявляют противоопухолевую активность. Плазмоцитоидные дендритные клетки были идентифицированы как наиболее мощные продуценты IFN I типа в ответ на действие антигена, и, таким образом, были названы природными клетками, продуцирующими IFN. Согласно результатам современных исследований предполагается, что путем усиления экспрессии IFN-α в инфильтрирующих опухоль макрофагах можно вызвать более эффективную активацию дендритных клеток и цитотоксичность иммунных эффекторных клеток.
IFN-ω высвобождается лейкоцитами в месте появления вирусной инфекции или опухолей. IFN-α действует как пирогенный фактор, изменяя активность термочувствительных нейронов в гипоталамусе, вызывая тем самым жар. Это происходит через связывание с опиоидными рецепторами, вызывая высвобождение простагландина-E2 (PGE2). Подобный механизм используется IFN-α для уменьшения боли; IFN-α взаимодействует с μ-опиоидным рецептором, действуя как анальгетик.
У мышей IFN-β ингибирует выработку факторов роста в иммунных клетках, тем самым замедляя рост опухоли, и препятствует другим клеткам продуцировать факторы роста, активирующие формирование сосудов, тем самым блокируя ангиогенез опухоли и препятствуя соединению опухоли с системой кровеносных сосудов.
В. Рецепторы интерферона
Рецептор интерферона-α/β (IFNAR) представляет собой рецептор, который связывает интерфероны I типа, включая интерферон-α и -β. Это гетеромерный рецептор клеточной поверхности, состоящий из одной цепи с двумя субъединицами, называемыми IFNAR1 и IFNAR2. После связывания IFN I типа IFNAR активирует сигнальный путь JAK-STAT. Стимуляция интерферонов традиционно приводит к стимуляции противовирусного иммунного ответа.
Его структура была получена с помощью ЯМР. Первоначально было вычислено 35 конформеров, количество которых с учетом критерия низкой энергии было уменьшено до 22. Это была первая спиральная структура цитокинового рецептора, определенная в растворе. Молекула имеет один полимер. Структура раскрывает природу связывания. Модель IFNAR2 позволила выявить преимущественно гидрофобный участок на рецепторе, который взаимодействует с соответствующей гидрофобной поверхностью на IFN-альфа. Смежный мотив заряженных боковых цепей направляет белки на формирование прочного комплекса. Связывающая поверхность позволяет объяснить перекрестную реактивность и специфичность лиганда рецептора. Источником для экспериментов, был человек, но экспрессию осуществляли в Escherichia coli.
IFNAR относят к группе бета-белков, исходя из его вторичной структуры. Изгибы цепи IFNAR обнаруживают четкую эволюционную связь с бета-сэндвичем иммуноглобулина. С учетом структурных и функциональных сходств, которые свидетельствуют о вероятной общей эволюции, он включен в суперсемейство и семейство фибронектинов типа III. Домен, или эволюционная единица, наблюдаемая в природе, представляет собой бета-цепь рецептора интерферона-альфа/бета. Вид представлял собой человека.
II. Пролекарства интерферона
А. Общие сведения
Пролекарства обычно определяют как молекулы, которые способны быть терапевтическими агентами, но находятся в форме, требующей некоторой модификации, чтобы стать действительно терапевтически значимыми. Такие реагенты особенно полезны, когда доставка активной лекарственной формы имеет некоторые присущие им ограничения, такие как токсичность или отсутствие стабильности, путем создания формы пролекарства, позволяющей избежать этих недостатков и обеспечить эффективную активацию in vivo в подходящее время и/или подходящем участке.
Пролекарства по настоящему изобретению имеют пять разных компонентов. Первым компонентом является интерферон. Вторым компонентом является маскирующий домен, который блокирует способность интерферона проявлять свою обычную активность пока находится в связанном состоянии. Третьим компонентом является стабилизирующий агент, например, константный домен антитела. Эти три компонента соединены линкерами, причем линкер, расположенный между интерфероном и маскирующим доменом, подвергается селективному расщеплению, например, протеазой, которая экспрессируется раковой клеткой или в окружении опухоли.
Как обсуждалось выше, интерфероны 1 типа представляют собой IFN-α (альфа), IFN-β (бета), IFN-κ (каппа), IFN-δ (дельта), IFN-ε (эпсилон), IFN-τ (тау), IFN-ω (омега) и IFN-ζ (дзета). Любая из этих молекул может быть включена в конструкции, описанные в настоящей заявке. Также возможно получение гетеродимерных конструкций с двумя или даже четырьмя различными интерферонами 1 типа. Интерфероны, используемые в настоящей заявке, для получения раскрытых в настоящем описании данных представляют собой полноразмерные белки, но не содержащие сигнальные пептиды.
Важной частью конструкции, конечно же, является область маскирования. Для этой цели авторы изобретения использовали природные рецепторы для интерферонов 1 типа, а не неприродные последовательности. Преимущество использования маскирующего домена, основанного на структуре природных рецепторов, включает как (а) высокое сродство к интерферону 1 типа, так и (b) более низкую вероятность иммунного ответа против последовательностей. Последовательности:
Мышиный IFNAR1-ECD:
1 mlavvgaaal vlvagapwvl psaaggenlk ppenidvyii ddnytlkwss hgesmgsvtf
61 saeyrtkdea kwlkvpecqh ttttkcefsl ldtnvyiktq frvraeegns tsswnevdpf
121 ipfytahmsp pevrleaedk ailvhisppg qdgnmwalek psfsytiriw qksssdkkti
181 nstyyvekip ellpettycl evkaihpslk khsnystvqc isttvankmp vpgnlqvdaq
241 gksyvlkwdy iasadvlfra qwlpgyskss sgsrsdkwkp iptcanvqtt hcvfsqdtvy
301 tgtfflhvqa segnhtsfws eekfidsqkh ilppppvitv tamsdtllvy vncqdstcdg
361 lnyeiifwen tsntkismek dgpeftlknl qpltvycvqa rvlfrallnk tsnfseklce
421 ktrpgsfst (SEQ ID NO: 1)
Мышиный IFNAR2-ECD:
1 mrsrctvsav gllslclvvs asletitpsa fdgypdepct initirnsrl ilswelenks
61 gppanytlwy tvmskdenlt kvkncsdttk sscdvtdkwl egmesyvvai vivhrgdltv
121 crcsdyivpa napleppefe ivgftdhinv tmefppvtsk iiqekmkttp fvikeqigds
181 vrkkhepkvn nvtgnftfvl rdllpktnyc vslyfdddpa iksplkcivl qpgqesglse
241 sa (SEQ ID NO: 2)
Человеческий IFNAR1-ECD:
1 mmvvllgatt lvlvavapwv lsaaaggknl kspqkvevdi iddnfilrwn rsdesvgnvt
61 fsfdyqktgm dnwiklsgcq nitstkcnfs slklnvyeei klriraeken tsswyevdsf
121 tpfrkaqigp pevhleaedk aivihispgt kdsvmwaldg lsftyslviw knssgveeri
181 eniysrhkiy klspettycl kvkaalltsw kigvyspvhc ikttvenelp ppenievsvq
241 nqnyvlkwdy tyanmtfqvq wlhaflkrnp gnhlykwkqi pdcenvkttq cvfpqnvfqk
301 giyllrvqas dgnntsfwse eikfdteiqa fllppvfnir slsdsfhiyi gapkqsgntp
361 viqdypliye iifwentsna erkiiekktd vtvpnlkplt vycvkaraht mdeklnkssv
421 fsdavcektk pgntsk (SEQ ID NO: 3)
Человеческий IFNAR2-ECD:
1 mllsqnafif rslnlvlmvy islvfgisyd spdytdesct fkislrnfrs ilswelknhs
61 ivpthytlly timskpedlk vvkncanttr sfcdltdewr stheayvtvl egfsgnttlf
121 scshnfwlai dmsfeppefe ivgftnhinv mvkfpsivee elqfdlslvi eeqsegivkk
181 hkpeikgnms gnftyiidkl ipntnycvsv ylehsdeqav iksplkctll ppgqesesae
241 sak (SEQ ID NO: 4)
Далее, конструкции включают домены стабилизации, которые, например, будут увеличивать период полувыведения in vivo. Авторы изобретения решили использовать константный домен Ig. Хотя был использован Fc-домен IgG1, также можно использовать другие Fc-домены. Особое преимущество от использования Fc-доменов IgA заключается в их способности димеризоваться, что означает, что в такую конструкцию может быть включено до четырех различных интерферонов. Одним из примеров создания конструкций с множеством интерферонов является использование гетеродимера FcA-FcB IgG1 с одним блокирующим рецептором и одним интерфероном, слитым с FcA или FcB, с использованием отдельных расщепляемых субстратов, специфических для разных ферментов, встроенных в линкеры FcA и FcB, соответственно. Можно также использовать другие стабилизирующие белки, включая человеческий сывороточный альбумин и трансферрин.
Наконец, указанные выше домены связаны друг с другом короткими пептидными участками или «линкерами». Один из этих линкеров, расположенный между молекулой интерферона и маскирующим доменом, подвергается расщеплению, когда пролекарство достигает или приближается к своей цели - опухоли или раковой клетке. В конкретном варианте осуществления линкер содержит участок, представляющий собой мишень для протеазы, в котором, когда пролекарство подвергается воздействию среды, содержащей протеазу, линкер расщепляется, высвобождая маскирующий агент из пролекарства, тем самым активируя молекулу интерферона. В идеале, линкер может быть выбран для протеаз, сверхэкспрессируемых в раковых клетках или в месте образования опухоли, и даже может быть адаптирован к профилю протеазы, экспрессируемой в раковых клетках/опухоли у конкретного индивидуума с помощью предварительного тестирования. Ниже приведены несколько примеров ферментов, селективно экспрессируемых или сверхэкспрессируемых в раковых или опухолевых средах.
ТАБЛИЦА 1 - СУБСТРАТЫ ПРОТЕАЗЫ
Примером формы линкера является GGGGS-субстрат-GGGGS (GGGGS=SEQ ID NO: 13). Другими двумя примерами являются (GGGGS)n-субстрат-(GGGGS)n и Gn-субстрат-Gn (n может быть любым числом) (GGGGS=SEQ ID NO: 13).
В. Разработка и экспрессия конструкций нуклеиновых кислот.
Доступны различные генетические конструкции, которые содержат описанные выше компоненты пролекарства интерферона, и которые могут быть введены в векторы экспрессии. Нуклеиновые кислоты по настоящему изобретению, которые кодируют молекулы пролекарства, необязательно могут быть связаны с другими белковыми последовательностями. В контексте настоящей заявки термин «экспрессирующая конструкция» предназначен для обозначения любого типа генетической конструкции, содержащей нуклеиновую кислоту, кодирующую генный продукт, в которой часть или вся последовательность, кодирующая нуклеиновую кислоту, способна транскрибироваться. Транскрипт может быть транслирован в белок, но это не является обязательным. В некоторых вариантах осуществления экспрессия включает как транскрипцию гена, так и трансляцию мРНК в продукт гена. В других вариантах осуществления экспрессия включает только транскрипцию нуклеиновой кислоты, кодирующей представляющий интерес ген.
Термин «вектор» используется для обозначения молекулы нуклеиновой кислоты-носителя, предназначенной для введения в клетку, где она может быть реплицирована, в которую может быть вставлена последовательность нуклеиновой кислоты. Последовательность нуклеиновой кислоты может быть «экзогенной», что означает, что эта последовательность является чужеродной для клетки, в которую вводится вектор, или что эта последовательность является гомологичной последовательности, находящейся в клетке, но находится внутри нуклеиновой кислоты клетки-хозяина в положении, в котором обычно эта последовательность отсутствует. Векторы включают плазмиды, космиды, вирусы (бактериофаг, вирусы животных и вирусы растений) и искусственные хромосомы (например, YAC). Специалист в данной области обладает необходимыми навыками для конструирования вектора с помощью стандартных рекомбинантных способов, которые описаны в работах Sambrook et al. (1989) и Ausubel et al. (1994), обе из которых включены в настоящее описание в качестве ссылки.
Термин «вектор экспрессии» относится к вектору, содержащему последовательность нуклеиновой кислоты, кодирующую по меньшей мере часть генного продукта, способного транскрибироваться. В некоторых случаях молекулы РНК затем транслируются в белок, полипептид или пептид. В других случаях эти последовательности не транслируются, например, при продуцировании антисмысловых молекул или рибозимов. Векторы экспрессии могут содержать множество «контрольных последовательностей», которые относятся к последовательностям нуклеиновых кислот, необходимым для транскрипции и, возможно, трансляции функционально связанной кодирующей последовательности в конкретном организме-хозяине. В дополнение к контрольным последовательностям, которые управляют транскрипцией и трансляцией, векторы и векторы экспрессии могут содержать последовательности нуклеиновых кислот, которые также выполняют другие функции и которые описаны ниже.
1. Регуляторные элементы
«Промотор» является контрольной последовательностью, которая представляет собой область последовательности нуклеиновой кислоты, осуществляющей контроль инициации и скорости транскрипции. Он может содержать генетические элементы, с которыми могут связываться регуляторные белки и молекулы, такие как РНК-полимераза и другие факторы транскрипции. Фразы «функционально расположенные», «функционально связанные», «под контролем» и «под транскрипционным контролем» означают, что промотор находится в правильном функциональном положении и/или ориентации относительно последовательности нуклеиновой кислоты для осуществления контроля инициации транскрипции и/или экспрессии этой последовательности. Промотор может использоваться или может не использоваться вместе с «энхансером», который относится к цис-действующей регуляторной последовательности, участвующей в активации транскрипции последовательности нуклеиновой кислоты.
Промотор может представлять собой промотор, который в природных условиях связан с геном или последовательностью, который может быть получен путем выделения 5’-некодирующих последовательностей, расположенных выше кодирующего сегмента и/или экзона. Такой промотор может упоминаться как «эндогенный». Аналогичным образом, энхансер может представлять собой энхансер, который в природных условиях связан с последовательностью нуклеиновой кислоты, расположенной ниже или выше этой последовательности. Альтернативно, определенные преимущества могут быть получены при размещении сегмента кодирующей нуклеиновой кислоты под контролем рекомбинантного или гетерологичного промотора, относящегося к промотору, который в природных условиях не связан с последовательностью нуклеиновой кислоты в естественной среде.
Рекомбинантный или гетерологичный энхансер относится также к энхансеру, который в природных условиях не связан с последовательностью нуклеиновой кислоты в естественной среде. Такие промоторы или энхансеры могут включать промоторы или энхансеры других генов, а также промоторы или энхансеры, выделенные из любой другой прокариотической, вирусной или эукариотической клетки, и промоторы или энхансеры, которые не «встречаются в природе», т.е. содержащие другие элементы других транскрипционных регуляторных областей и/или мутации, которые изменяют экспрессию. В дополнение к синтетически полученным последовательностям нуклеиновых кислот промоторов и энхансеров, последовательности могут быть получены с помощью метода рекомбинантного клонирования и/или амплификации нуклеиновых кислот, включая ПЦРТМ, применительно к раскрытыми в настоящем описании композициям (см. патент США 4683202, патент США 5928906, каждый из которых включен в настоящее описание в качестве ссылки). Кроме того, предполагается, что контрольные последовательности также могут использоваться для прямой транскрипции и/или экспрессии последовательностей внутри неядерных органелл, таких как митохондрии, хлоропласты и т.п.
Естественно, важно отметить применение промотора и/или энхансера, эффективно направляющего экспрессию сегмента ДНК в определенном типе клеток, органелле и организме, которые выбраны для экспрессии. Специалистам в области молекулярной биологии обычно известно использование комбинаций промоторов, энхансеров и типов клеток для экспрессии белка, например, см. работу Sambrook et al. (1989), включенную в настоящее описание в качестве ссылки. Используемые промоторы могут быть конститутивными, тканеспецифичными, индуцибельными и/или полезными в соответствующих условиях для получения высокого уровня экспрессии введенного сегмента ДНК, что является преимуществом при крупномасштабном производстве рекомбинантных белков и/или пептидов. Промотор может быть гетерологичным или эндогенным.
Идентичность тканеспецифичных промоторов или элементов, а также анализы для характеристики их активности хорошо известны специалистам в данной области. Примеры таких областей включают ген LIMK2 человека (Nomoto et al. 1999), ген рецептора соматостатина 2 (Kraus et al., 1998), мышиный эпидидимальный связывающий ретиноевую кислоту ген (Lareyre et al., 1999), человеческий CD4 (Zhao-Emonet et al., 1998), мышиный альфа2 (XI) коллаген (Tsumaki, et al., 1998), ген рецептора дофамина D1A (Lee, et al., 1997), инсулиноподобный фактор роста II (Wu et al., 1997), молекулу адгезии эндотелиальных клеток тромбоцитов-1 человека (Almendro et al., 1996). Специфичные для опухоли промоторы также могут применяться в настоящем изобретении.
Для эффективной трансляции кодирующих последовательностей также может потребоваться конкретный сигнал инициации. Эти сигналы включают кодон инициации ATG или смежные последовательности. Может возникнуть необходимость в предоставлении экзогенных сигналов регуляции трансляции, включая кодон инициации ATG. Специалист в данной области техники легко сможет это определить и предоставить необходимые сигналы. Хорошо известно, что для обеспечения трансляции всей вставки кодон инициации должен находиться «внутри рамки считывания» с рамкой считывания требуемой кодирующей последовательности. Экзогенные сигналы регуляции трансляции и кодоны инициации могут быть естественными или синтетическими. Эффективность экспрессии может быть увеличена путем включения соответствующих энхансерных элементов транскрипции.
2. IRES
В некоторых вариантах осуществления настоящего изобретения для создания мультигенных или полицистронных сообщений используются такие элементы, как участки внутренней посадки рибосомы (IRES). Элементы IRES способны обойти модель сканирования рибосомой 5'-метилированного Cap и начать трансляцию на внутренних участках (Pelletier and Sonenberg, 1988). Были описаны элементы IRES, полученные от двух членов семейства пикорнавирусов (полиомиелита и энцефаломиокардита) (Pelletier and Sonenberg, 1988), а также IRES из мРНК млекопитающих (Macejak and Sarnow, 1991). Элементы IRES могут быть связаны с гетерологичными открытыми рамками считывания. Несколько открытых рамок считывания могут подвергаться совместной транскрипции, при этом каждая рамка считывания разделена IRES, обеспечивая таким образом создание полицистронных мРНК. Благодаря элементу IRES каждая открытая рамка считывания доступна рибосомам, что обеспечивает эффективную трансляцию. С помощью одного промотора/энхансера возможна эффективная экспрессия множества генов с обеспечением транскрипции одной мРНК (см. патенты США 5925565 и 5935819, включенные в настоящее описание в качестве ссылки).
3. Многоцелевые сайты клонирования
Векторы могут включать множественный сайт клонирования (MCS), который представляет собой область нуклеиновой кислоты, содержащую множество сайтов рестрикционных ферментов, любой из которых можно использовать в сочетании со стандартным рекомбинантным методом для расщепления вектора. См. Carbonelli et al., 1999, Levenson et al., 1998 и Cocea, 1997, включенные в настоящее описание в качестве ссылки. «Расщепление рестрикционными ферментами» относится к каталитическому расщеплению молекулы нуклеиновой кислоты ферментом, который функционирует только в определенных местах в молекуле нуклеиновой кислоты. Многие из этих рестрикционных ферментов имеются в продаже. Использование таких ферментов хорошо известно специалистам в данной области. Часто вектор линеаризуют или фрагментируют, используя рестрикционный фермент, который разрезает MCS внутри, обеспечивая возможность лигирования экзогенных последовательностей с вектором. «Лигирование» относится к процессу образования фосфодиэфирных связей между двумя фрагментами нуклеиновой кислоты, которые могут быть или могут не быть смежными друг с другом. Методы, включающие рестрикционные ферменты и реакции лигирования, хорошо известны специалистам в области рекомбинантных технологий.
4. Сплайсинг сайтов
Большинство транскрибируемых эукариотических молекул РНК подвергают сплайсингу РНК для удаления интронов из первичных транскриптов. Для векторов, содержащих геномные эукариотические последовательности, могут потребоваться сайты для сплайсинга доноров и/или акцепторов для обеспечения надлежащего процессинга транскрипта для экспрессии белка (см. Chandler et al., 1997, включенный в настоящее описание в качестве ссылки).
5. Сигналы терминации
Векторы или конструкции по настоящему изобретению, как правило, содержат по меньшей мере один сигнал терминации. «Сигнал терминации» или «терминатор» состоит из ДНК-последовательностей, участвующих в специфической терминации РНК-транскрипта РНК-полимеразой. Таким образом, в определенных вариантах осуществления рассматривается сигнал терминации, который завершает продуцирование РНК-транскрипта. Для получения мРНК нужного уровня может понадобиться терминатор in vivo.
В эукариотических системах область терминатора также может содержать специфические последовательности ДНК, которые обеспечивают сайт-специфическое расщепление нового транскрипта, обнажая сайт полиаденилирования. Это является сигналом для специализированной эндогенной полимеразы к добавлению участка длиной примерно 200 А остатков (полиА) к 3'-концу транскрипта. Молекулы РНК, модифицированные таким полиА хвостом, оказываются более стабильными и транслируются более эффективно. Таким образом, в других вариантах осуществления, в которых используются эукариоты, предпочтительно, чтобы этот терминатор содержал сигнал для расщепления РНК, и более предпочтительно, чтобы сигнал терминатора стимулировал полиаденилирование мРНК. Элементы терминатора и/или сайта полиаденилирования могут служить для повышения уровней мРНК и/или для минимизации считывания кассеты в другие последовательности.
Терминаторы, предполагаемые для использования в настоящем изобретении, включают любой известный терминатор транскрипции, описанный в настоящей заявке или известный специалисту в данной области, включая, без ограничения, например, последовательности терминации генов, такие как, например, терминатор бычьего гормона роста, или вирусные последовательности терминации, такие как, например, терминатор SV40. В некоторых вариантах осуществления сигнал терминации может представлять собой отсутствие транскрибируемой или транслируемой последовательности, например, в связи с усечением последовательности.
6. Сигналы полиаденилирования
При экспрессии, особенно при экспрессии в эукариотической клетке, обычно включают сигнал полиаденилирования для обеспечения правильного полиаденилирования транскрипта. Природа сигнала полиаденилирования, как полагают, не имеет решающего значения для успешной реализации изобретения, и/или может быть использована любая такая последовательность. Предпочтительные варианты осуществления включают сигнал полиаденилирования SV40 и/или сигнал полиаденилирования бычьего гормона роста, удобный и/или известный как хорошо функционирующий в различных клетках-мишенях. Полиаденилирование может увеличить стабильность транскрипта или может способствовать цитоплазматическому транспорту.
7. Точка начала репликации
Для размножения вектора в клетке-хозяине, он может содержать одну или более точек начала репликации (часто называемых «ori»), которые представляют собой специфическую последовательность нуклеиновой кислоты, в которой инициируется репликация. Альтернативно, можно использовать автономно реплицируемую последовательность (ARS), если клетка-хозяин представляет собой дрожжи.
8. Селектируемые и скринируемые маркеры
В некоторых вариантах осуществления изобретения клетки, содержащие конструкцию нуклеиновой кислоты по настоящему изобретению, могут быть идентифицированы in vitro или in vivo путем включения маркера в вектор экспрессии. Благодаря таким маркерам обеспечивается идентифицируемое изменение клетки, позволяющее легко идентифицировать клетки, содержащие вектор экспрессии. Как правило, селектируемым маркером является маркер, который придает свойство, позволяющее выполнить отбор. В случае положительного селектируемого маркера, выбирают его присутствие, в то время как в случае отрицательного селектируемого маркера, его присутствие исключает выбор. Примером положительного селектируемого маркера является маркер устойчивости к лекарственным веществам.
Обычно включение маркера отбора по чувствительности к лекарственному веществу помогает в клонировании и идентификации трансформантов; например, гены, которые придают устойчивость к неомицину, пуромицину, гигромицину, DHFR, GPT, зеоцину и гистидинолу, являются полезными селектируемыми маркерами. В дополнение к маркерам, придающим фенотип, который позволяет отличить трансформантов, исходя из реализации условий, также предполагаются другие типы маркеров, включая скринируемые маркеры, такие как GFP, основой которых является колориметрический анализ. Альтернативно, можно использовать скринируемые ферменты, такие как тимидинкиназа (tk) вируса простого герпеса или хлорамфеникол-ацетилтрансфераза (CAT). Специалист в данной области также осведомлен, как использовать иммунологические маркеры, возможно, в сочетании с анализом FACS. Считается, что используемый маркер не имеет важного значения при условии, что он способен экспрессироваться одновременно с нуклеиновой кислотой, кодирующей продукт гена. Дополнительные примеры селектируемых и скринируемых маркеров хорошо известны специалисту в данной области.
9. Вирусные векторы
Способность некоторых вирусных векторов эффективно инфицировать или проникать в клетки, интегрироваться в геном клетки-хозяина и стабильно экспрессировать вирусные гены, привела к разработке и применению ряда различных вирусных векторных систем (Robbins et al., 1998). В настоящее время вирусные системы разрабатываются для использования в качестве векторов для переноса генов ex vivo и in vivo. Например, в настоящее время оценивают использование векторов на основе аденовируса, вируса простого герпеса, ретровируса и аденоассоциированных вирусов для лечения таких заболеваний, как рак, муковисцидоз, болезнь Гоше, почечная недостаточность и артрит (Robbins and Ghivizzani, 1998; Imai et al., 1998; патент США 5670488). Различные вирусные векторы, описанные ниже, имеют определенные преимущества и недостатки в зависимости от конкретного применения генной терапии.
10. Невирусная трансформация
Считается, что подходящие способы доставки нуклеиновой кислоты для трансформации органеллы, клетки, ткани или организма, подходящие для применения в настоящем изобретении, включают практически любой способ, с помощью которого нуклеиновая кислота (например, ДНК) может быть введена в органеллу, клетку, ткань или организм, описанные в настоящей заявке или известные специалисту в данной области. Такие способы включают, без ограничения, прямую доставку ДНК, такую как инъекция (патенты США 5994624, 5981274, 5945100, 5780448, 5736524, 5702932, 5656610, 5589466 и 5580859, каждый из которых включен в настоящее описание в качестве ссылки), включая микроинъекцию (Harland и Weintraub, 1985; патент США 5789215, включенные в настоящее описание в качестве ссылки); путем электропорации (патент США 5384253, включенный в настоящее описание в качестве ссылки); осаждения фосфатом кальция (Graham and Van Der Eb, 1973; Chen и Okayama, 1987; Rippe et al., 1990); с помощью DEAE декстрана, и затем полиэтиленгликоля (Gopal, 1985); с помощью прямой ультразвуковой обработки (Fechheimer et al., 1987); опосредованной липосомами трансфекции (Nicolau and Sene, 1982; Fraley et al., 1979; Nicolau et al., 1987; Wong et al., 1980; Kaneda et al., 1989; Kato et al., 1991); бомбардировки микрочастицами (заявки PCT №№ WO 94/09699 и 95/06128; патенты США 5610042; 5322783, 5563055, 5550318, 5538877 и 5538880, каждый из которых включен в настоящее описание в качестве ссылки); путем перемешивания с волокнами карбида кремния (Kaeppler et al., 1990; патенты США 5302523 и 5464765, каждый из которых включен в настоящее описание в качестве ссылки); или с помощью опосредованной PEG трансформации протопластов (Omirulleh et al., 1993; патенты США 4684611 и 4952500, каждый из которых включен в настоящее описание в качестве ссылки); путем поглощения ДНК, опосредованного высушиванием/ингибированием (Potrykus et al., 1985). Благодаря применению таких технологий, обеспечивается стабильная или временная трансформация органеллы(органелл), клетки(клеток), ткани(ей) или организма(ов).
Инъекция. В некоторых вариантах осуществления нуклеиновая кислота может доставляться в органеллу, клетку, ткань или организм посредством одной или нескольких инъекций (т.е. инъекции с помощью иглы), таких как, например, либо подкожных, внутрикожных, внутримышечных, внутривенных, либо внутрибрюшинных инъекций. Способы введения вакцин хорошо известны специалистам в данной области (например, инъекция композиции, содержащей солевой раствор). Дополнительные варианты осуществления настоящего изобретения включают введение нуклеиновой кислоты путем прямой микроинъекции. Прямую микроинъекцию использовали для введения конструкций нуклеиновых кислот в ооциты Xenopus (Harland and Weintraub, 1985).
Электропорация. В некоторых вариантах осуществления настоящего изобретения нуклеиновую кислоту вводят в органеллу, клетку, ткань или организм посредством электропорации. Электропорация включает воздействие на суспензию клеток и ДНК высоковольтным электрическим разрядом. В некоторых вариантах этого способа используются определенные ферменты, разрушающие клеточную стенку, такие как ферменты, разрушающие пектин, делая реципиентные клетки-мишени более восприимчивыми к трансформации с помощью электропорации по сравнению с необработанными клетками (патент США 5384253, включенный в настоящее описание в качестве ссылки). Альтернативно, реципиентные клетки можно сделать более восприимчивыми к трансформации путем механического повреждения.
Трансфекция эукариотических клеток с помощью электропорации является довольно успешной. Мышиные пре-B лимфоциты были трансфицированы генами человеческого иммуноглобулина (Potter et al., 1984), а гепатоциты крысы были трансфицированы таким образом геном хлорамфеникол-ацетилтрансферазы (Tur Kaspa et al., 1986).
Для осуществления трансформации в клетках, таких как, например, растительные клетки, методом электропорации можно использовать либо рыхлые ткани, такие как суспензионная культура клеток или эмбриогенный каллус, либо альтернативно можно осуществить прямую трансформацию незрелых зародышей или других организованных тканей. В этом методе можно частично разрушить клеточные стенки выбранных клеток, подвергая их воздействию ферментов, расщепляющих пектин (пектолиазы), или нанести контролируемые механически повреждения. Примеры некоторых видов, которые были трансформированы с помощью электропорации интактных клеток, включают кукурузу (патент США 5384253; Rhodes et al., 1995; D'Halluin et al., 1992), пшеницу (Zhou et al., 1993), томат (Hou и Lin, 1996), сою (Christou et al., 1987) и табак (Lee et al., 1989).
Для трансформации растительных клеток с помощью электропорации также можно использовать протопласты (Bates, 1994; Lazzeri, 1995). Например, получение трансгенных растений сои путем электропорации протопластов, полученных из семядолей, описано Dhir и Widholm в международной заявке на патент № WO 92/17598, включенной в настоящее описание в качестве ссылки. Другие примеры видов, для которых описана трансформация протопластов, включают ячмень (Lazerri, 1995), сорго (Battraw et al., 1991), кукурузу (Bhattacharjee et al., 1997), пшеницу (He et al., 1994) и томат (Tsukada, 1989).
Фосфат кальция. В других вариантах осуществления настоящего изобретения нуклеиновую кислоту вводят в клетки, используя осаждение фосфатом кальция. Человеческие клетки KB были трансфицированы ДНК аденовируса 5 (Graham and Van Der Eb, 1973) с помощью этого метода. Таким же образом были трансфицированы мышиные клетки L (A9), мышиные клетки C127, клетки CHO, CV 1, BHK, NIH3T3 и HeLa маркерным геном неомицина (Chen and Okayama, 1987), и гепатоциты крысы, которые были трансфицированы различными генами-маркерами (Rippe et al., 1990).
DEAE декстран. В другом варианте осуществления нуклеиновую кислоту доставляют в клетку, используя DEAE декстран, и затем полиэтиленгликоль. Таким образом вводили репортерные плазмиды в клетки мышиной миеломы и эритролейкемии (Gopal, 1985).
Обработка ультразвуком. Дополнительные варианты осуществления настоящего изобретения включают введение нуклеиновой кислоты путем прямой обработки ультразвуком. LTK-фибробласты были трансфицированы геном тимидинкиназы с помощью ультразвуковой обработки (Fechheimer et al., 1987).
Опосредованная липосомами трансфекция. В другом варианте осуществления изобретения нуклеиновая кислота может быть захвачена липидным комплексом, таким как, например, липосома. Липосомы представляют собой везикулярные структуры, характеризующиеся двухслойной фосфолипидной мембраной и внутренней водной средой. Мультиламеллярные липосомы имеют несколько липидных слоев, разделенных водной средой. Они образуются спонтанно при суспендировании фосфолипидов в избытке водного раствора. Перед образованием замкнутых структур происходит самоорганизация липидных компонентов и захват воды и растворенных компонентов между липидными бислоями (Ghosh and Bachhawat, 1991). Также рассматривается нуклеиновая кислота, образующая комплекс с липофектамином (Gibco BRL) или суперфектом (Qiagen).
Опосредованная липосомами доставка нуклеиновых кислот и экспрессия чужеродной ДНК in vitro оказалась очень успешной (Nicolau and Sene, 1982; Fraley et al., 1979; Nicolau et al., 1987). Также продемонстрирована возможность опосредованной липосомами доставки и экспрессии чужеродной ДНК в культивируемых эмбрионах цыплят, клетках HeLa и гепатомы (Wong et al., 1980).
В некоторых вариантах осуществления настоящего изобретения липосома может образовывать комплекс с гемагглютинирующим вирусом (HVJ). Было показано, что это облегчает слияние с клеточной мембраной и способствует проникновению в клетку инкапсулированной в липосомы ДНК (Kaneda et al., 1989). В других вариантах осуществления липосома может образовывать комплекс или использоваться вместе с ядерными негистоновыми хромосомными белками (HMG 1) (Kato et al., 1991). В других вариантах осуществления липосома может образовывать комплекс или использоваться вместе с HVJ и HMG 1. В других вариантах осуществления носитель для доставки может содержать лиганд и липосому.
Рецептор-опосредованная трансфекция. Нуклеиновую кислоту также можно доставлять в клетку-мишень с помощью носителей для рецептор-опосредованной доставки. Их преимущество заключается в селективном поглощении макромолекул посредством опосредованного рецептором эндоцитоза, который происходит в клетке-мишени. Ввиду специфического распределения различных рецепторов в зависимости от типа клеток этот способ доставки добавляет настоящему изобретению еще одну степень специфичности.
Некоторые рецептор-опосредованные носители, нацеленные на гены, содержат специфический по отношению к клеточному рецептору лиганд и агент, связывающий нуклеиновую кислоту. Другие содержат специфический по отношению к клеточному рецептору лиганд, к которому оперативно присоединяют предназначенную для доставки нуклеиновую кислоту. Для рецептор-опосредованного переноса генов было использовано несколько лигандов (Wu и Wu, 1987; Wagner et al., 1990; Perales et al., 1994; Myers, EPO 0273085), что свидетельствует об эксплуатационной пригодности этого метода. Описана специфическая доставка в контексте другого типа клеток млекопитающих (Wu and Wu, 1993; включен в настоящее описание в качестве ссылки). В некоторых аспектах настоящего изобретения выбирают такой лиганд, который соответствует рецептору, специфически экспрессируемому в популяции клеток-мишеней.
В других вариантах осуществления компонент-носитель, предназначенный для доставки нуклеиновой кислоты, носителя, нацеленного на специфическую клеточную нуклеиновую кислоту клетки, может содержать специфический связывающий лиганд в комбинации с липосомой. Нуклеиновая(ые) кислота(ы), предназначенная(ые) для доставки, содержится(атся) в липосоме, а специфический связывающий лиганд функционально включен в липосомную мембрану. Таким образом, липосома будет специфически связываться с рецептором(ами) клетки-мишени и доставлять содержимое в клетку. Было показано, что такие системы функционируют с помощью систем, в которых, например, в случае рецептор-опосредованной доставки нуклеиновой кислоты в клетки, которые демонстрируют активацию рецептора EGF, используется эпидермальный фактор роста (EGF).
В других вариантах осуществления компонент-носитель, предназначенный для доставки нуклеиновой кислоты, носителя целевой доставки может представлять собой саму липосому, которая предпочтительно будет содержать один или более липидов или гликопротеинов, которые направляют связывание с определенными клетками. Например, лактозилцерамид, галактозатерминальный асиалганглиозид, вводили в липосомы и наблюдал увеличение поглощения гена инсулина гепатоцитами (Nicolau et al., 1987). Предполагается, что тканеспецифичные трансформирующие конструкции по настоящему изобретению могут быть специфически доставлены в клетку-мишень аналогичным образом.
11. Системы экспрессии
Существуют многочисленные системы экспрессии, которые включают, по меньшей мере, часть или все раскрытые выше композиции. В настоящем изобретении для получения последовательностей нуклеиновых кислот или их родственных полипептидов, белков и пептидов могут использоваться прокариотческие и/или эукариотические системы. Многие такие системы являются широко распространенными и коммерчески доступными.
Система на основе клеток насекомых/бакуловируса может обеспечить высокий уровень экспрессии белка в сегменте гетерологичной нуклеиновой кислоты, как описано в патентах США № 5871986 и 4879236, включенные в настоящее описание в качестве ссылки, которую можно приобрести, например, под названием MaxBac® 2.0 у компании Invitrogen® и бакуловирусная система экспрессии BacPackТМ у компании Clontech®.
Другие примеры систем экспрессии включают систему Stratagene® Complete ControlТМ индуцибельной экспрессии у млекопитающих, которая включает синтетический рецептор, индуцируемый экдизоном, или систему его экспрессии pET, экспрессируемую в E. coli. Другой пример системы индуцибельной экспрессии доступен от Invitrogen®, которая несет систему T-RexТМ (регулируемую тетрациклином экспрессию), представляющую собой систему индуцибельной экспрессии у млекопитающих, в которой используется полноразмерный промотор CMV. Invitrogen® также предоставляет дрожжевую систему экспрессии, которая называется Pichia methanolica Expression System и предназначена для высоко эффективного продуцирования рекомбинантных белков в метилотрофных дрожжах Pichia methanolica. Для экспрессии раскрытых пролекарственных конструкций также можно использовать векторы pGEM-T Easy, плазмидные векторы pCon VectorsTM, Lonza pConIgG1 или pConK2 и клетки 293 Freestyle или клетки Lonza CHO.
Первичные культуры клеток млекопитающих могут быть получены различными способами. Чтобы клетки были жизнеспособными в условиях in vitro и в контакте с экспрессирующей конструкцией, необходимо обеспечить содержание клеток в условиях правильного соотношения кислорода и углекислого газа и питательных веществ и одновременно защитить их от микробного загрязнения. Методы культивирования клеток хорошо задокументированы.
Один из вариантов осуществления вышеизложенного включает использование переноса генов для иммортализации клеток с целью продуцирования белков. Ген представляющего интерес белка может быть перенесен описанным выше методом в соответствующие клетки-хозяева с последующим культивированием клеток в соответствующих условиях. Таким образом можно использовать ген практически любого полипептида. Создание рекомбинантных векторов экспрессии и включенных в них элементов обсуждалось выше. Альтернативно, продуцируемый белок может быть эндогенным белком, обычно синтезируемым рассматриваемой клеткой.
Примеры полезных линий клеток-хозяев млекопитающих представляют собой клетки Vero и HeLa и линии клеток яичника китайского хомяка, W138, BHK, COS-7, 293, HepG2, NIH3T3, RIN и MDCK. Кроме того, может быть выбран штамм клетки-хозяина, который модулирует экспрессию введенных последовательностей или модифицирует и процессирует продукт гена требуемым способом. Такие модификации (например, гликозилирование) и процессинг (например, расщепление) белковых продуктов могут быть важными для функционирования белка. Различные клетки-хозяева имеют характерные и специфические механизмы пост-трансляционного процессинга и модификации белков. Для обеспечения правильной модификации и процессинга экспрессированного чужеродного белка могут быть выбраны подходящие клеточные линии или системы-хозяева.
Можно использовать ряд систем отбора, включая, без ограничения, тимидинкиназу HSV, гены гипоксантин-гуанинфосфорибозилтрансферазы и аденин-фосфорибозилтрансферазы в tk-, hgprt- или aprt-клетках, соответственно. Кроме того, устойчивость к метаболитам может быть использована в качестве основы для выбора dhfr, обеспечивающего устойчивость к; gpt, обеспечивающего устойчивость к микофенольной кислоте; neo, обеспечивающего устойчивость к аминогликозиду G418; и hygro, обеспечивающего устойчивость к гигромицину.
Е. Очистка
В некоторых вариантах осуществления пролекарства интерферона по настоящему изобретению могут быть очищены. В контексте настоящего изобретения термин «очищенный» обозначает композицию, изолированную от других компонентов, в которой белок очищен до любой степени по сравнению с его состоянием в случае получения естественным образом. Следовательно, очищенный белок также относится к белку, свободному от среды, в которой он может встречаться в природе. Когда используется термин «по существу очищенный», это обозначение относится к композиции, в которой белок или пептид образует основной компонент композиции, например, белки в композиции составляют примерно 50%, примерно 60%, примерно 70%, примерно 80%, примерно 90%, примерно 95% или более.
Методы очистки белков хорошо известны специалистам в данной области. Эти методы включают, на одном уровне, грубое фракционирование клеточной среды с получением полипептидной и неполипептидной фракций. После отделения полипептида от других белков представляющий интерес полипептид может быть дополнительно очищен методами хроматографии и электрофореза для достижения частичной или полной очистки (или очистки до гомогенного состояния). Аналитические методы, особенно подходящие для получения чистого пептида, включают ионообменную хроматографию, эксклюзионную хроматографию; электрофорез в полиакриламидном геле; изоэлектрическое фокусирование. Другие способы очистки белка включают осаждение сульфатом аммония, ПЭГ, антителами и т.п. или термическую денатурацию с последующим центрифугированием; гель-фильтрацию, обращенно-фазовую, гидроксилапатитную и аффинную хроматографию; и комбинацию этих и других методов.
При очистке пролекарства интерферона по настоящему изобретению может потребоваться экспрессия полипептида в прокариотической или эукариотической системе экспрессии и экстракция белка в денатурирующих условиях. Полипептид может быть очищен от других клеточных компонентов с использованием аффинной колонки, которая связывает меченную часть полипептида. В данной области техники хорошо известно, что изменение порядка проведения различных стадий очистки или пропуск некоторых стадий также обеспечит подходящий способ получения по существу очищенного белка или пептида. Обычно антитела фракционируют с помощью агентов (т.е. белка А), которые связываются с Fc-частью антитела. В тех случаях, когда пролекарство интерферона содержит такой домен, может быть использован этот подход.
Специалистам в данной области техники известны различные способы количественного определения степени очистки белка или пептида, используемые в контексте настоящего изобретения. Они включают, например, определение удельной активности активной фракции или оценку количества полипептидов во фракции с помощью анализа, основанного на методах SDS/PAGE. Другой метод оценки чистоты фракции заключается в вычислении удельной активности фракции, сравнении ее с удельной активностью исходного экстракта и, таким образом, вычисления степени чистоты. Фактические единицы, используемые для представления количества активности, конечно, зависят от конкретного метода анализа, выбранного для последующей очистки, и от того, проявляет ли экспрессируемый белок или пептид детектируемую активность.
Известно, что миграция полипептида может изменяться, иногда значительно, в разных условиях SDS/PAGE (Capaldi et al., 1977). Поэтому следует понимать, что в разных условиях электрофореза кажущаяся молекулярная масса очищенных или частично очищенных продуктов экспрессии может варьировать.
III. Фармацевтические составы и лечение рака
А. Раковые заболевания
Рак возникает в результате роста клональной популяции клеток из ткани. Развитие рака, называемого канцерогенезом, можно смоделировать и охарактеризовать несколькими способами. Уже давно установлена связь между развитием рака и воспалением. Воспалительный ответ участвует в защите хозяина от микробной инфекции, а также способствует восстановлению и регенерации тканей. Имеются многочисленные свидетельства, указывающие на связь между воспалением и риском развития рака, т.е. хроническое воспаление может привести к дисплазии.
Раковые клетки, к которым могут применяться способы по настоящему изобретению, обычно включают любую клетку, которая селективно экспрессирует протеазы, и, в частности, которая сверхэкспрессирует такие протеазы по сравнению с нормальными клетками. Подходящей раковой клеткой может быть рак молочной железы, рак легкого, рак толстой кишки, рак поджелудочной железы, рак почки, рак желудка, рак печени, рак кости, гематологический рак (например, лейкоз или лимфома), рак нервной ткани, меланома, рак яичника, рак яичка, рак предстательной железы, рак шейки матки, рак влагалища или рак мочевого пузыря. Кроме того, способы по изобретению могут быть применены к широкому кругу видов, например людям, приматам, не являющимся человеком (например, обезьянам, павианам или шимпанзе), лошадям, крупному рогатому скоту, свиньям, овцам, козам, собакам, кошкам, кроликам, морским свинкам, песчанкам, хомякам, крысам и мышам. Рак также может быть рецидивирующим, метастатическим и/или устойчивым ко многим лекарственным средствам, и способы по настоящему изобретению могут быть в частности применимы к таким раковым заболеваниям для обеспечения возможности осуществления резекции, пролонгирования или реиндуцирования ремиссии, подавления ангиогенеза, предотвращения или ограничения метастазирования и/или для лечения рака, устойчивого ко многим лекарственным средствам. На клеточном уровне это может привести к уничтожению раковых клеток, ингибированию роста раковых клеток или реверсии или уменьшению злокачественного фенотипа опухолевых клеток.
B. Состав и введение
Настоящее изобретение относится к фармацевтическим композициям, содержащим пролекарства интерферона. В конкретном варианте осуществления термин «фармацевтически приемлемый» означает одобренный регулирующим органом федерального правительства или правительства штата или внесенный в список фармакопеи США или другой общепризнанной фармакопеи для применения на животных и, более конкретно, на людях. Термин «носитель» относится к разбавителю, наполнителю или несущей среде, с которой вводится терапевтическое средство. Такими фармацевтическими носителями могут быть стерильные жидкости, такие как вода и масла, включая масла нефтяного, животного, растительного или синтетического происхождения, такие как арахисовое масло, соевое масло, минеральное масло, кунжутное масло и т.п. Другие подходящие фармацевтические наполнители включают крахмал, глюкозу, лактозу, сахарозу, физиологический раствор, декстрозу, желатин, солод, рис, муку, мел, силикагель, стеарат натрия, моностеарат глицерина, тальк, хлорид натрия, сухое обезжиренное молоко, глицерин, пропиленгликоль, воду, этанол и т.п.
Композиции могут быть приготовлены в виде нейтральных или солевых форм. Фармацевтически приемлемые соли включают соли, образованные с анионами, такими как соли, полученные из соляной, фосфорной, уксусной, щавелевой и винной кислот и т.д., а также соли, образованные с катионами, такими как соли, полученные из гидроксидов натрия, калия, аммония, кальция, трехвалентного железа, изопропиламина, триэтиламина, 2-этиламиноэтанола, гистидина, прокаина и др.
Антитела по настоящему изобретению могут включать классические фармацевтические препараты. Введение этих композиций по настоящему изобретению может осуществляться любым обычным путем при условии, что этот путь обеспечит доступ к целевой ткани. Такой путь включает пероральный, назальный, буккальный, ректальный, вагинальный или топический. Альтернативно, введение может быть внутрикожным, подкожным, внутримышечным, внутрибрюшинным или внутривенным введением. Такие композиции обычно вводят в виде фармацевтически приемлемых композиций, описанных выше. Особый интерес представляет введение непосредственно внутрь опухоли, перфузия опухоли или введение локально или регионарно относительно опухоли, например, в локальную или региональную сосудистую сеть или лимфатическую систему или в ложе резецированной опухоли.
Активные соединения также можно вводить парентерально или внутрибрюшинно. Растворы активных соединений в виде свободного основания или фармакологически приемлемых солей могут быть приготовлены в воде, подходящим образом смешанной с поверхностно-активным веществом, таким как гидроксипропилцеллюлоза. Также могут быть приготовлены дисперсии в глицерине, жидких полиэтиленгликолях и их смесях и в маслах. В обычных условиях хранения и использования эти препараты содержат консервант для предотвращения роста микроорганизмов.
C. Комбинированная терапия
В контексте настоящего изобретения также предполагается, что описанные в настоящей заявке пролекарства интерферона могут быть использованы аналогично в сочетании с иммуно-, химио- или радиотерапевтическим вмешательством или другими видами лечения. Также может оказаться эффективным, в частности, комбинирование пролекарства интерферона с другими методами лечения, которые нацелены на разные аспекты функционирования раковых клеток.
Для уничтожения клеток, ингибирования роста клеток, ингибирования метастазирования, ингибирования ангиогенеза или, с другой стороны, достижения реверсии или уменьшения злокачественного фенотипа опухолевых клеток с помощью способов и композиций по настоящему изобретению обычно клетку-мишень необходимо привести в контакт с пролекарствами интерферона по настоящему изобретению и, по меньшей мере, одним другим агентом. Эти композиции могут быть предоставлены в комбинированном количестве, эффективном для уничтожения или подавления пролиферации клетки. Этот процесс может включать приведение в контакт клеток с пролекарствами интерферона по настоящему изобретению и другим агентом(ами) или фактором(ами) одновременно. Это может быть достигнуто путем приведения в контакт клетки с одной композицией или фармакологическим составом, который включает оба агента, или путем приведения клетки в контакт одновременно с двумя разными композициями или составами, причем одна композиция включает пролекарства интерферона по настоящему изобретению, а другая включает другой агент.
Альтернативно, терапия пролекарством интерферона может предшествовать или следовать за лечением другим агентом с интервалами в пределах от минут до недель. В вариантах осуществления, в которых другой агент и пролекарства интерферона вводят в клетку по отдельности, обычно предусмотрено, чтобы доставки не были разделены значительным промежутком времени, что обеспечивает возможность комбинированного влияния обоих агентов на клетку. В таких случаях предполагается, что контактирование клетки с обоими модальностями осуществляется с промежутком между введениями в пределах примерно 12-24 и, более предпочтительно, в пределах примерно 6-12 часов, причем наиболее предпочтительно, когда время задержки составляет всего примерно 12 часов. В некоторых ситуациях может потребоваться значительное продление периода лечения, однако в этом случае интервал между соответствующими введениями составляет от нескольких дней (2, 3, 4, 5, 6 или 7) до нескольких недель (1, 2, 3, 4, 5, 6, 7 или 8).
Также может потребоваться более одного введения пролекарств интерферона или другого агента. Могут быть использованы различные комбинации, где пролекарство интерферона по настоящему изобретению представляет собой «А», а другой вид терапии представляет собой «В», как показано ниже:
A/B/A B/A/B B/B/A A/A/B B/A/A A/B/B B/B/B/A B/B/A/B
A/A/B/B A/B/A/B A/B/B/A B/B/A/A B/A/B/A B/A/A/B B/B/B/A
A/A/A/B B/A/A/A A/B/A/A A/A/B/A A/B/B/B B/A/B/B B/B/A/B
Предусмотрены и другие комбинации. И в этом случае следует отметить, что для уничтожения клеток оба агента доставляют в клетку в комбинированном количестве, эффективном для уничтожения клетки.
Агенты или факторы, подходящие для лечения рака, включают любое химическое соединение или способ лечения, который вызывает повреждение ДНК при введении в клетку. Такие агенты и факторы включают излучение и волны, вызывающие повреждение ДНК, такие как облучение, микроволны, электронное излучение и т.п. Можно использовать различные химические соединения, также называемые «химиотерапевтическими» или «генотоксическими агентами». Это может быть достигнуто путем облучения локализованного участка с опухолью; альтернативно, опухолевые клетки можно приводить в контакт с агентом путем введения пациенту терапевтически эффективного количества фармацевтической композиции.
Для использования в настоящем изобретении предлагаются различные классы химиотерапевтических агентов. Например, селективные антагонисты рецептора эстрогена («SERM»), такие как тамоксифен, 4-гидрокситамоксифен (афимоксфен), фальсодекс, ралоксифен, базедоксифен, кломифен, фемарель, лазофоксифен, ормелоксифен и торемифен.
Химиотерапевтические агенты, предлагаемые для использования, включают, например, камптотецин, актиномицин D, митомицин С. Изобретение также включает применение комбинации одного или более агентов, повреждающих ДНК, либо радиоактивные, либо фактические соединения, такие как применение рентгеновского излучения с цисплатином или применение цисплатина с этопозидом. Агент может быть приготовлен и использован в виде комбинированной терапевтической композиции.
Белок теплового шока 90 является регуляторным белком, обнаруженным во многих эукариотических клетках. Было показано, что ингибиторы HSP90 полезны при лечении рака. Такие ингибиторы включают гелданамицин, 17-(аллиламино)-17-деметоксигельданамицин, PU-H71 и рифабутин.
Также предусмотрены агенты, которые непосредственно осуществляют поперечную сшивку ДНК или образуют аддукты. Могут быть использованы такие агенты, как цисплатин и другие ДНК-алкилирующие агенты. Цисплатин широко используется для лечения рака, при этом дозы, эффективные для клинического применения, составляют 20 мг/м2 в течение 5 дней каждые три недели в течение всех трех курсов. Цисплатин не всасывается перорально и поэтому должен вводиться путем внутривенной, подкожной, внутриопухолевой или внутрибрюшинной инъекции.
Агенты, повреждающие ДНК, также включают соединения, которые препятствуют репликации ДНК, митозу и сегрегации хромосом. Такие химиотерапевтические соединения включают адриамицин, также известный как доксорубицин, этопозид, верапамил, подофиллотоксин и т.п. Широко используемые в клинических условиях для лечения новообразований, эти соединения вводят внутривенно в виде болюсной дозы от 25-75 мг/м2 с интервалами 21 день для доксорубицина, до 35-50 мг/м2 для этопозида, вводимого внутривенно или перорально в дозе, двукратно превышающей внутривенную дозу. Также предусмотрено введение ингибиторов микротрубочек, таких как таксаны. Эти молекулы представляют собой дитерпены, продуцируемые растениями рода Taxus, и включают паклитаксел и доцетаксел.
Ингибиторы рецептора эпидермального фактора роста, такие как иресса, mTOR, мишень рапамицина у млекопитающих, также известная как комплекс FK506-связывающего белка 12 с рапамицин-ассоциированным белком 1 (FRAP1), представляющая собой серин/треонин-протеинкиназу, регулируют рост клеток, пролиферацию клеток, подвижность клеток, выживание клеток, синтез белка и транскрипцию. Следовательно, рапамицин и его аналоги («рапалоги») также предусмотрены для применения в терапии рака в соответствии с настоящим изобретением.
Другим возможным терапевтическим агентом является TNF-α (фактор некроза опухоли-альфа), цитокин, участвующий в системном воспалении, и член группы цитокинов, которые стимулируют реакцию острой фазы. Основная роль TNF заключается в регуляции иммунных клеток. TNF также способен вызывать апоптотическую гибель клеток, воспаление и ингибировать онкогенез и репликацию вируса.
Агенты, которые нарушают синтез и точность вспроизведения предшественников нуклеиновых кислот и субъединиц, также приводят к повреждению ДНК. В качестве такового был разработан ряд предшественников нуклеиновых кислот. Особенно полезными являются агенты, которые прошли всевозможное тестирование и являются легко доступными. По существу, такие агенты, как 5-фторурацил (5-FU), предпочтительно используются опухолевой тканью, что делает этот агент особенно полезным для нацеливания на опухолевые клетки. Несмотря на то, что 5-FU является достаточно токсичным, его применяют для широкого круга носителей, включая топические, однако обычно используется внутривенное введение в дозах от 3 до 15 мг/кг/сутки.
Другие широко используемые факторы, вызывающие повреждение ДНК, включают факторы, обычно называемые γ-лучами, рентгеновскими лучами и/или направленной доставкой радиоизотопов в опухолевые клетки. Также предусмотрены другие формы факторов, повреждающих ДНК, такие как микроволны и УФ-излучение. Наиболее вероятно, что все эти факторы обеспечивают широкий спектр повреждений ДНК, предшественников ДНК, репликации и репарации ДНК, а также повреждения при сборке и поддержании хромосом. Диапазон доз используемых рентгеновских лучей варьирует от суточных доз от 50 до 200 рентген в течение продолжительных периодов времени (от 3 до 4 недель) до разовых доз от 2000 до 6000 рентген. Диапазоны доз для радиоизотопов варьируют в широких пределах и зависят от периода полураспада изотопа, силы и типа испускаемого излучения и поглощения опухолевыми клетками.
Кроме того, также предусмотрено возможное применение иммунотерапии, гормонотерапии, токсинотерапии и хирургического вмешательства. В частности, можно использовать препараты таргетной терапии, такие как авастин, эрбитукс, гливек, герцептин и ритуксан.
Специалист в данной области может использовать руководство «Remington's Pharmaceutical Sciences», 15-е издание, глава 33, в частности, стр. 624-652. Как правило, дозу корректируют в зависимости от состояния пациента, получающего лечение. Лицо, ответственное за введение, в любом случае, способно подобрать подходящую дозу для индивидуального пациента. Кроме того, для введения человеку препараты должны соответствовать стандартам стерильности, пирогенности, общей безопасности и чистоты, которые определенны стандартами департамента FDA по биологическим препаратам.
IV. Методы детектирования протеаз
В других вариантах осуществления может возникнуть необходимость в определении природы и количества экспрессируемых протеаз в целевой раковой клетке или опухоли, что позволит подобрать конкретное пролекарство интерферона, которое будет активировано в опухоли пациента с высокой вероятностью. В целом, существует три подхода к такому детектированию: иммунологическое детектирование, например, с помощью протеаз-специфических антител, детектирование с помощью мРНК и анализ активности протеазы.
А. Иммунодетектирование
Методы иммунодетектирования для идентификации и количественного определения протеаз могут включать, среди прочего, иммуноферментный анализ (ELISA), радиоиммуноанализ (RIA), иммунорадиометрический анализ, флюороиммуноанализ, хемилюминесцентный анализ, биолюминесцентный анализ и вестерн-блот анализ. Этапы различных полезных методов иммунодетектирования описаны в научной литературе, см., например, Doolittle and Ben-Zeev (1999), Gulbis and Galand (1993), De Jager et al. (1993) и Nakamura et al. (1987). Методы иммуноблоттинга, как правило, включают получение образца и приведение образца в контакт с первым антителом в соответствии с вариантами осуществления, обсуждаемыми в настоящем описании, в условиях, эффективных для образования иммунокомплексов.
Контактирование выбранного биологического образца с антителом в эффективных условиях и в течение периода времени, достаточного для образования иммунных комплексов (первичных иммунных комплексов), обычно заключается в простом добавлении композиции антитела к образцу и инкубации смеси в течение некоторого периода времени, достаточного для того, чтобы антитела образовали иммунные комплексы, т.е. связались с присутствующей протеазой. По истечении этого времени композицию образец-антитело, такую как на срезе ткани, в планшете для ELISA, дот-блоттинга или вестерн-блоттинга, обычно промывают для удаления любых неспецифически связанных видов антител, что обеспечивает обнаружение только тех антител, которые специфически связаны в первичных иммунных комплексах.
Обнаружение образования иммунокомплекса обычно хорошо известно в данной области и может быть выполнено путем применения многочисленных подходов. Эти методы обычно основаны на обнаружении метки или маркера, например, любой из радиоактивных, флуоресцентных, биологических и ферментных меток. Патенты, касающиеся применения таких меток, включают патенты США 3817837, 3850752, 3939350, 3996345, 4277437, 4275149 и 4366241. Конечно, использования вторичного связывающего лиганда, такого как второе антитело, и/или биотин/авидин-связывающего лиганда, известного в данной области, может обеспечить дополнительные преимущества.
Антитело, используемое для обнаружения, само может быть связано с детектируемой меткой, которая затем может быть легко обнаружена, тем самым позволяя определить количество первичных иммунных комплексов в композиции. Альтернативно, первое антитело, которое становится связанным в первичных иммунных комплексах, может быть обнаружено с помощью второго связывающего лиганда, который имеет сродство связывания с антителом. В этих случаях второй связывающий лиганд может быть связан с детектируемой меткой. Второй связывающий лиганд сам по себе часто является антителом, которое соотвественно можно назвать «вторичным» антителом. Первичные иммунные комплексы контактируют с меченым, вторичным связывающим лигандом или антителом в эффективных условиях и в течение периода времени, достаточного для образования вторичных иммунных комплексов. Вторичные иммунные комплексы затем обычно промывают для удаления любых неспецифически связанных меченых вторичных антител или лигандов, и затем выполняют детектирование оставшейся метки во вторичных иммунных комплексах.
Дополнительные методы включают обнаружение первичных иммунных комплексов с помощью двухэтапного подхода. Второй связывающий лиганд, такой как антитело, которое имеет сродство связывания с антителом, используется для образования описанных выше вторичных иммунных комплексов. После промывки вторичные иммунные комплексы приводят в контакт с третьим связывающим лигандом или антителом, которое обладает сродством связывания со вторым антителом, снова в эффективных условиях и в течение периода времени, достаточного для образования иммунных комплексов (третичных иммунных комплексов). Третий лиганд или антитело связывается с детектируемой меткой, обеспечивая возможность детектирования образованных таким образом третичных иммунных комплексов. При необходимости, эта система может обеспечивать усиление сигнала.
В одном из методов иммунодетектирования используются два разных антитела. Первое биотинилированное антитело используется для обнаружения антигена-мишени, а затем используется второе антитело для обнаружения биотина, связанного с биотином, находящемся в комплексе. В этом методе тестируемый образец сначала инкубируют в растворе, содержащем антитело первого этапа. Если целевой антиген присутствует, часть антитела связывается с антигеном с образованием биотинилированного комплекса антитело/антиген. Затем комплекс антитело/антиген амплифицируют путем инкубации в последовательных растворах стрептавидина (или авидина), биотинилированной ДНК и/или комплементарной биотинилированной ДНК, при этом каждый этап добавляет дополнительные сайты биотина к комплексу антитело/антиген. Стадии амплификации повторяют до тех пор, пока не будет достигнут подходящий уровень амплификации, после чего образец инкубируют в растворе, содержащем антитело к биотину второго этапа. Это антитело второго этапа метят, например, ферментом, который можно использовать для обнаружения комплекса антитело/антиген гистоферментативным методом с помощью хромогенного субстрата. При подходящей амплификации может быть получен макроскопически видимый конъюгат.
В другом известном методе иммунодетектирования используются преимущества метода иммуно-ПЦР (полимеразной цепной реакции). Метод ПЦР аналогичен методу Кантора вплоть до инкубации с биотинилированной ДНК, однако вместо использования нескольких циклов инкубации со стрептавидином и биотинилированной ДНК комплекс ДНК/биотин/стрептавидин/антитело вымывается буфером с низким pH или высоким содержанием соли с высвобождением антитела. Затем полученный промывочный раствор используют для проведения реакции ПЦР с подходящими праймерами с соответствующими контролями. По меньшей мере, теоретически, мощная амплификационная способность и специфичность ПЦР позволяют обнаружить одну молекулу антигена.
В одном из примеров ELISA антитела по изобретению иммобилизуют на выбранной поверхности, проявляющей сродство к белку, такой как лунка в полистирольном микротитровальном планшете. Затем тестируемую композицию, предположительно содержащую протеазу, добавляют в лунки. После связывания и промывки для удаления неспецифически связанных иммунных комплексов, может быть обнаружен связанный антиген. Обнаружение может быть достигнуто путем добавления другого антитела к протеазе, которое связывается с детектируемой меткой. Этот тип ELISA представляет собой простой «сэндвич-ELISA». Обнаружение также может быть достигнуто путем добавления второго антитела к протеазе с последующим добавлением третьего антитела, которое имеет сродство связывания со вторым антителом, причем третье антитело связано с детектируемой меткой.
В другом приведенном в качестве примера методе ELISA образцы, предположительно содержащие протеазу, иммобилизуют на поверхности лунки и затем приводят в контакт с антителом к протеазе. После связывания и промывки для удаления неспецифически связанных иммунных комплексов выполняют обнаружение связанных антител к протеазе. Если исходные антитела к протеазе связаны с детектируемой меткой, это позволяет непосредственное обнаружение иммунных комплексов. И в этом случае, иммунные комплексы могут быть обнаружены с помощью второго антитела, которое имеет сродство связывания с первым антителом к протеазе, причем второе антитело связывается с детектируемой меткой.
Независимо от используемого формата, методы ELISA имеют определенные общие черты, такие как покрытие, инкубация и связывание, промывка для удаления неспецифически связанных видов и детектирование связанных иммунных комплексов. Они описаны ниже.
При покрытии планшета антигеном или антителом, как правило, лунки планшета инкубируют с раствором антигена или антитела либо в течение ночи, либо в течение определенного периода времени. Затем лунки планшета промывают для удаления не полностью адсорбированного материала. Затем любые оставшиеся доступные поверхности лунок «покрывают» неспецифическим белком, который является антигенно нейтральным по отношению к тестируемой антисыворотке. К ним относятся бычий сывороточный альбумин (БСА), казеин или растворы сухого молока. Покрытие позволяет блокировать сайты неспецифической адсорбции на иммобилизующей поверхности и, таким образом, уменьшает фон, вызванный неспецифическим связыванием антисыворотки на поверхности.
В методах ELISA, по всей видимости, привычнее использовать вторичные или третичные средства обнаружения, а не прямую процедуру. Таким образом, после связывания белка или антитела с лункой, покрытия нереакционноспособным материалом для уменьшения фона и промывки для удаления несвязанного материала, иммобилизующую поверхность приводят в контакт с тестируемым биологическим образцом в условиях, эффективных для образования иммунного комплекса (антиген/антитело). Затем для обнаружения иммунного комплекса требуется меченый вторичный связывающий лиганд или антитело и вторичный связывающий лиганд или антитело в сочетании с меченым третичным антителом или третьим связывающим лигандом.
«В условиях, эффективных для образования иммунного комплекса (антиген/антитело)» означает, что условия предпочтительно включают разбавление антигенов и/или антител растворами, такими как BSA, бычий гамма-глобулин (BGG) или физиологический раствор с фосфатным буфером (PBS)/твин. Эти добавленные агенты также способствуют уменьшению неспецифического фона.
«Подходящие» условия также означают, что инкубация проводится при температуре или в течение периода времени, достаточного для обеспечения эффективного связывания. Стадии инкубации обычно длятся от примерно 1-2 до 4 часов или около того, при температурах, предпочтительно, от 25 до 27°C, или могут длиться в течение ночи при температуре примерно 4°C или около того.
После всех этапов инкубации в методе ELISA контактирующую поверхность промывают для удаления не находящегося в комплексе материала. Предпочтительная процедура промывки включает промывку раствором, таким как PBS/твин или боратным буфером. После образования специфических иммунных комплексов между тестируемым образцом и первоначально связанным материалом и последующей промывки может быть определено появление даже незначительных количеств иммунных комплексов.
Для обеспечения средства обнаружения второе или третье антитело обычно содержит ассоциированную метку, позволяющую выполнить детектирование. Предпочтительно, такая метка, как правило, представляет собой фермент, который обеспечивает развитие окраски при инкубации с соответствующим хромогенным субстратом. Таким образом, например, необходим контакт или инкубация первого и второго иммунного комплекса с антителами, конъюгированными с уреазой, глюкозооксидазой, щелочной фосфатазой или пероксидазой водорода, в течение периода времени и в условиях, которые способствуют развитию дальнейшего образования иммунного комплекса (например, инкубация в течение 2 часов при комнатной температуре в PBS-содержащем растворе, таком как PBS-твин).
После инкубации с меченым антителом и после промывки для удаления несвязанного материала выполняют количественное определение метки, например, путем инкубации с хромогенным субстратом, таким как мочевина или бромкрезоловый пурпурный, или 2,2'-азино-ди-(3-этилбензтиазолин-6-сульфоновая кислота (ABTS) или H2O2, в случае пероксидазы, в качестве ферментной метки. Количественное определение достигается путем измерения степени развития цвета, например, с помощью спектрофотометра в видимом диапазоне спектра.
B. Детектирование мРНК
Для оценки активности протеазы в раковой клетке или опухоли может быть использован метод детектирования мРНК. Как правило, детектирование мРНК основано на гибридизации одной нуклеиновой кислоты (зонда или праймера) с другой нуклеиновой кислотой (мишенью).
Использование зонда или праймера длиной от 13 до 100 нуклеотидов, предпочтительно длиной от 17 до 100 нуклеотидов или, в некоторых аспектах, длиной до 1-2 килобаз или более, позволяет образовывать дуплексную молекулу, которая является одновременно стабильной и селективной. Молекулы, имеющие комплементарные последовательности на непрерывных участках длиной более 20 оснований, обычно являются предпочтительными с точки зрения увеличения стабильности и/или селективности полученных гибридных молекул. Обычно предпочтительнее конструировать молекулы нуклеиновой кислоты для гибридизации, имеющие одну или более комплементарных последовательностей от 20 до 30 нуклеотидов или даже больше, где это желательно. Такие фрагменты могут быть легко получены, например, путем прямого синтеза фрагмента химическими средствами или путем введения выбранных последовательностей в рекомбинантные векторы для продуцирования рекомбинантными методами.
Соответственно, можно использовать нуклеотидные последовательности, позволяющие селективно формировать дуплексные молекулы с комплементарными мРНК-участками или обеспечивать праймеры для амплификации мРНК из образцов. В зависимости от предполагаемого применения можно использовать различные условия гибридизации для достижения различных степеней селективности зонда или праймеров для последовательности-мишени.
Для применений, требующих высокой селективности, обычно желательно использовать относительно высоко жесткие условия для образования гибридов. Например, условия с относительно низким содержанием солей и/или высокой температурой, например обеспечиваемые содержанием NaCl от примерно 0,02 М до примерно 0,10 М при температурах от примерно 50°С до примерно 70°С. Такие условия высокой жесткости допускают незначительное, если таковое имеется, несоответствие между зондом или праймерами и матрицей или нитью-мишенью и могут быть особенно подходящими для выделения специфических генов или для обнаружения специфических транскриптов мРНК. Как правило, следует понимать, что могут быть обеспечены более жесткие условия путем добавления увеличивающихся количеств формамида.
Понятно, что в некоторых случаях предпочтительны условия более низкой жесткости. В этих условиях может происходить гибридизация, даже если последовательности гибридизующихся цепей не являются точно комплементарными и не совпадают в одном или более положениях. Менее жесткие условия могут быть обеспечены за счет увеличения концентрации соли и/или уменьшения температуры. Например, условия средней жесткости могут быть обеспечены содержанием NaCl примерно от 0,1 до 0,25 М при температурах от примерно 37°С до примерно 55°С, в то время как условия низкой жесткости могут быть обеспечены содержанием соли от примерно 0,15 М до примерно 0,9 М при температурах в пределах от примерно 20°С до примерно 55°С. Условиями гибридизации можно легко манипулировать в зависимости от требуемых результатов.
В других вариантах осуществления гибридизация может быть достигнута в условиях, например, 50 мМ Трис-HCl (рН 8,3), 75 мМ KCl, 3 мМ MgCl2, 1,0 мМ дитиотреитола, при температурах от приблизительно 20°С до примерно 37°С. Другие используемые условия гибридизации могут включать приблизительно 10 мМ Трис-HCl (рН 8,3), 50 мМ KCl, 1,5 мМ MgCl2 при температурах в диапазоне от приблизительно 40°С до примерно 72°С.
В некоторых вариантах осуществления для определения гибридизации обычно выгодно использовать нуклеиновые кислоты определенных последовательностей по настоящему изобретению в сочетании с подходящими средствами, такими как метка. В данной области техники известно большое количество подходящих индикаторных средств, включая флуоресцентные, радиоактивные, ферментные или другие лиганды, такие как детектируемый авидин/биотин. В предпочтительных вариантах осуществления можно использовать флуоресцентную метку или ферментную метку, такую как уреаза, щелочная фосфатаза или пероксидаза, вместо радиоактивных или других нежелательных для окружающей среды реагентов. В случае ферментных меток известны колориметрические индикаторные субстраты, используемые для идентификации специфической гибридизации с комплементарными образцами, содержащими нуклеиновую кислоту, которые можно использовать в качестве средств, обеспечивающих визуальное или спектрофотометрическое обнаружение.
В общем случае, предполагается, что зонды или праймеры, описанные в настоящей заявке, полезны в качестве реагентов при гибридизации растворов, как в случае ПЦРТМ, для обнаружения экспрессии соответствующих генов, а также в вариантах осуществления, в которых используется твердая фаза. В вариантах осуществления, в которых используется твердая фаза, тестируемую мРНК адсорбируют или иным образом прикрепляют к выбранной матрице или поверхности. Эту фиксированную одноцепочечную нуклеиновую кислоту затем подвергают гибридизации с выбранными зондами в требуемых условиях. Выбранные условия будут зависеть от конкретных обстоятельств (в зависимости, например, от содержания G+C, типа нуклеиновой кислоты-мишени, источника нуклеиновой кислоты, размера зонда для гибридизации и т.д.). Оптимизация условий гибридизации для конкретного представляющего интерес применения хорошо известна специалистам в данной области. После промывки гибридизованных молекул для удаления неспецифически связанных молекул зонда выполняют обнаружение и/или количественное определение гибридизации путем определения количества связанной метки. Типичные твердофазные способы гибридизации раскрыты в патентах США 5843663, 5900481 и 5919626. Другие способы гибридизации, которые можно использовать при осуществлении настоящего изобретения, раскрыты в патентах США № 5849481, 5849486 и 5851772 и публикации патента США 2008/0009439. Соответствующие части этих и других ссылок, указанных в этом разделе описания, включены в него в качестве ссылки.
Нуклеиновые кислоты-мишени могут быть выделены из клеток, тканей или других образцов стандартными методами (Sambrook et al., 2001). В некоторых вариантах осуществления проводят анализ на гомогенатах цельных клеток или тканей или проб биологической жидкости без существенной очистки матричной нуклеиновой кислоты. В других случаях может потребоваться очистка и/или амплификация.
Амплификация обычно включает пары праймеров, предназначенных для селективной гибридизации с нуклеиновыми кислотами, которые соответствуют мишени, в условиях, обеспечивающих селективную гибридизацию. В зависимости от требуемого применения могут быть выбраны условия гибридизации высокой жесткости, которые обеспечивают гибридизацию только с последовательностями, которые полностью комплементарны праймерам. В других вариантах осуществления гибридизация может происходить в менее жестких условиях, чтобы обеспечить амплификацию нуклеиновых кислот, содержащих одно или более несовпадений с последовательностями праймеров. После гибридизации комплекс матрица-праймер связывают с одним или более ферментами, которые облегчают зависимый от матрицы синтез нуклеиновой кислоты. Несколько циклов амплификации, также называемых «циклами», проводят до тех пор, пока не будет получено достаточное количество продукта амплификации.
Продукт амплификации может быть обнаружен или количественно определен. В некоторых случаях обнаружение может быть выполнено с помощью визуальных средств. Альтернативно, обнаружение может включать опосредованную идентификацию продукта с помощью хемилюминесценции, радиоактивной сцинтиграфии встроенной радиоактивной метки или флуоресцентной метки или даже с помощью системы, в которой используются электрические и/или тепловые импульсные сигналы (Bellus, 1994).
Известно несколько зависимых от матрицы процессов амплификации олигонуклеотидных последовательностей, присутствующих в заданном образце матрицы. Одним из наиболее известных методов амплификации является полимеразная цепная реакция (называемая ПЦРTM), которая подробно описана в патентах США 4683195, 4683202 и 4800159, а также в Innis et al., 1988, каждый из которых включен в настоящее описание во всей своей полноте в качестве ссылки.
Для количественного определения амплифицированной мРНК может быть выполнена процедура амплификации ПЦРTM с обратной транскриптазой. Методы обратной транскрипции РНК в кДНК хорошо известны (см. Sambrook et al., 2001). В альтернативных методах обратной транскрипции используются термостабильные ДНК-полимеразы. Эти способы описаны в WO 90/07641. Методы полимеразной цепной реакции хорошо известны в данной области. Типичные методы ОТ-ПЦР описаны в патенте США № 5882864.
Для определения относительных концентраций конкретных видов мРНК, выделенных из клетки, можно использовать обратную транскрипцию (ОТ) РНК в кДНК с последующей количественной ПЦР (ОТ-ПЦР). Путем определения того, что концентрация конкретных видов мРНК варьирует, показано, что ген, кодирующий конкретные виды мРНК, экспрессируется дифференциально. При построении графика, на котором номер цикла находится на оси X, а логарифм концентрации амплифицированной ДНК-мишени находится на оси Y, путем соединения точек, нанесенных на график, получают изогнутую линию характерной формы. Начиная с первого цикла, наклон линии является положительным и постоянным. Это называется линейной частью кривой. После того, как реагент становится ограничивающим фактором, наклон линии начинает уменьшаться и в конечном итоге становится равным нулю. В этот момент концентрация амплифицированной ДНК-мишени становится асимптотической до некоторого фиксированного значения. Эта часть кривой называется плато.
Концентрация ДНК-мишени в линейной части амплификации ПЦР прямо пропорциональна начальной концентрации мишени до начала реакции. Определяя концентрацию амплифицированных продуктов ДНК-мишени в реакциях ПЦР, которые прошли одинаковое количество циклов и находятся в линейных диапазонах, можно определить относительные концентрации конкретной последовательности-мишени в исходной смеси ДНК. Если смеси ДНК представляют собой кДНК, синтезированные из РНК, выделенных из разных тканей или клеток, то для соответствующих тканей или клеток могут быть определены относительные количества копий конкретных мРНК, из которых была получена последовательность-мишень. Эта прямая пропорциональность между концентрацией продуктов ПЦР и относительным содержанием мРНК верна только в линейном диапазоне реакции ПЦР.
Конечная концентрация целевой ДНК в части кривой, представленной плато, определяется наличием реагентов в реакционной смеси и не зависит от исходной концентрации целевой ДНК. Таким образом, первое условие, которое должно быть выполнено до того, как методом ОТ-ПЦР можно будет определить относительное количество копий видов мРНК для коллекции популяций РНК, заключается в том, что забор образцов для определения концентрации амплифицированных продуктов ПЦР должен быть выполнен, когда реакции ПЦР находятся в линейной части соответствующей кривой.
Вторым условием эксперимента ОТ-ПЦР является определение относительного содержания конкретных видов мРНК. Как правило, относительные концентрации амплифицируемых кДНК нормируют относительно некоторого независимого стандарта. Цель эксперимента методом ОТ-ПЦР заключается в определении количества конкретных видов мРНК относительно среднего количества всех видов мРНК в образце.
В большинстве протоколов для конкурентной ПЦР используют внутренние стандарты ПЦР, которых имеют примерно такую же копийность как и мишени. Эти стратегии эффективны, если образцы продуктов амплификации ПЦР отбирают во время их линейных фаз. Если продукты отбирают, когда реакции приближаются к фазе плато, то появляется избыточное количество менее распространенного продукта. Это приводит к искажению результатов сравнения относительного содержания, сделанного для многих различных образцов РНК, например в случае изучения образцов РНК на предмет дифференциальной экспрессии, такому как уменьшение различий в относительном содержании РНК, что не соответствует действительности. Это не является серьезной проблемой, если внутренний стандарт имеет гораздо более высокую копийность по сравнению с мишенью. Если внутренний стандарт имеет более высокую копийность по сравнению с мишенью, то можно выполнить прямые линейные сравнения между образцами РНК.
Метод ОТ-ПЦР может быть выполнен в виде относительной количественной ОТ-ПЦР с внутренним стандартом, где внутренний стандарт представляет собой амплифицируемый фрагмент кДНК, который длиннее целевого фрагмента кДНК, и где копийность мРНК, кодирующей внутренний стандарт примерно 5-100 раз выше, чем мРНК, кодирующей мишень. Этот анализ позволяет измерять относительное содержание, а не абсолютное количество соответствующих видов мРНК.
Другим способом амплификации является лигазная цепная реакция («ЛЦР»), раскрытая в европейской заявке № 320308, полностью включенная в настоящее описание в качестве ссылки. В патенте США 4883750 описан способ, аналогичный ЛЦР, используемый для связывания пар зондов с последовательностью-мишенью. Также может быть использован способ, основанный на ПЦРTM и олигонуклеотид-лигазном анализе (ОЛА), раскрытый в патенте США № 5912148.
Альтернативные способы амплификации целевых последовательностей нуклеиновых кислот, которые можно использовать для осуществления настоящего изобретения, раскрыты в патентах США 5843650, 5846709, 5,846783, 5849,546, 5849497, 5849547, 5858652, 5866366, 5916776, 5922574, 5928905, 5928906, 5932451, 5935825, 5939291 и 5942391, в заявке Великобритании № 2202328 и в заявке PCT № PCT/US89/01025, каждая из которых включена в настоящее описание во всей полноте в качестве ссылки.
Q-бета репликаза, описанная в заявке PCT № PCT/US87/00880, также может быть использована в качестве способа амплификации в настоящем изобретении. В этом методе репликативную последовательность РНК, которая имеет область, комплементарную области-мишени, добавляют к образцу в присутствии РНК-полимеразы. Полимераза копирует репликативную последовательность, которая затем может быть обнаружена.
Метод изотермической амплификации, в котором используют рестрикционные эндонуклеазы и лигазы для достижения амплификации молекул-мишеней, содержащих нуклеотид-5'-[альфа-тио]-трифосфаты на одной нити сайта расщепления, также может быть полезным для амплификации нуклеиновых кислот по настоящему изобретению (Walker et al., 1992). Амплификация смещением цепи (SDA), раскрытая в патенте США 5916779, представляет собой другой способ выполнения изотермической амплификации нуклеиновых кислот, который включает многочисленные циклы смещения и синтеза цепи, т.е. ник-трансляции.
Другие процедуры амплификации нуклеиновых кислот включают системы амплификации на основе транскрипции (TAS), включая амплификацию на основе последовательностей нуклеиновых кислот (NASBA) и 3SR (Kwoh et al., 1989; заявка PCT WO 88/10315, полностью включенные в настоящее описание в качестве ссылки). В европейской заявке № 329822 раскрыт процесс амплификации нуклеиновой кислоты, включающий циклический синтез одноцепочечной РНК («оцРНК»), оцДНК и двухцепочечной ДНК (дцДНК), который можно использовать в соответствии с настоящим изобретением.
В заявке PCT WO 89/06700 (полностью включенной в настоящее описание в качестве ссылки) раскрыта схема амплификации последовательности нуклеиновой кислоты, основанная на гибридизации последовательности промоторной области/праймера с одноцепочечной ДНК-мишенью («оцДНК») с последующей транскрипцией многих РНК-копии последовательности. Эта схема не является циклической, т.е. новые матрицы не создаются из полученных в результате РНК-транскриптов. Другие способы амплификации включают «RACE» и «одностороннюю ПЦР» (Frohman, 1990; Ohara et al., 1989).
После любой амплификации может возникнуть необходимость в отделении продукта амплификации от матрицы и/или избытка праймера. В одном из вариантов осуществления продукты амплификации разделяют электрофорезом в агарозном, агарозо-акриламидном или полиакриламидном геле стандартными методами (Sambrook et al., 2001). Отделенные продукты амплификации могут быть вырезаны и элюированы из геля для дальнейшей манипуляции. В случае использования агарозных гелей с низкой температурой плавления, отделенную полосу можно удалить путем нагревания геля с последующей экстракцией нуклеиновой кислоты.
Разделение нуклеиновых кислот также можно осуществлять методами хроматографии, известными в данной области. Существует много видов хроматографии, которые могут быть использованы для реализации настоящего изобретения, включая адсорбционную, разделительную, ионообменную, гидроксилапатитную хроматографию, хроматографию на молекулярных ситах, обращенно-фазовую, колоночную, бумажную, тонкослойную и газовую хроматографию, а также ВЭЖХ.
В некоторых вариантах осуществления продукты амплификации визуализируют. Типичный метод визуализации включает окрашивание геля бромидом этидия и визуализацию полос в УФ-свете. Альтернативно, если выполняют целочисленное мечение продуктов амплификации радиоактивными или флуорометрически меченными нуклеотидами, то отдельные продукты амплификации могут быть экспонированы на рентгеновской пленке или визуализированы в соответствующих возбуждающих спектрах.
В одном из вариантов осуществления после разделения продуктов амплификации меченый зонд нуклеиновой кислоты приводят в контакт с амплифицированной маркерной последовательностью. Зонд предпочтительно конъюгируют с хромофором, но можно выполнить мечение радиоактивным изотопом. В другом варианте осуществления зонд конъюгируют с партнером по связыванию, таким как антитело или биотин, или другим партнером по связыванию, несущим детектируемый фрагмент.
В конкретных вариантах осуществления обнаружение осуществляется с помощью саузерн-блоттинга и гибридизации с меченым зондом. Методы саузерн-блоттинга хорошо известны специалистам в данной области (см. Sambrook et al., 2001). Один из примеров вышеизложенного описан в патенте США 5279721, включенном в настоящее описание в качестве ссылки, в котором описано устройство и способ для осуществления автоматического электрофореза и переноса нуклеиновых кислот. Устройство позволяет выполнять электрофорез и блоттинг без внешних манипуляций с гелем и идеально подходит для осуществления способов в соответствии с настоящим изобретением.
Различные методы обнаружения нуклеиновых кислот, известные в данной области техники, раскрыты в патентах США 5840873, 5843640, 5843651, 5846708, 5846717, 5846726, 5846729, 5849487, 5853990, 5853992, 5853993, 5856092, 5861244, 5863732, 5863753, 5866331, 5905024, 5910407, 5912124, 5912145, 5919630, 5925517, 5928862, 5928869, 5929227, 5932413 и 5935791, каждый из которых включен в настоящее описание в качестве ссылки.
Обнаружение мРНК протеазы может включать использование массивов или данных, сгенерированных из массива. Массив обычно относится к упорядоченным макрочипам или микрочипам молекул нуклеиновых кислот (зондов), которые полностью или почти комплементарны или идентичны множеству молекул мРНК или молекул кДНК и которые расположены на материале-носителе пространственно разделенным образом. Макрочипы, как правило, представляют собой листы нитроцеллюлозы или нейлона, на которые нанесены зонды. На микрочипах зонды нуклеиновой кислоты расположены более плотно, таким образом, что область размером обычно от 1 до 4 квадратных сантиметров может содержать до 10000 молекул нуклеиновой кислоты. Микрочипы могут быть изготовлены путем нанесения молекул нуклеиновой кислоты, например генов, олигонуклеотидов и т.д., на субстраты или путем изготовления олигонуклеотидных последовательностей in situ на субстрате. Нанесенные в виде пятен или изготовленные молекулы нуклеиновой кислоты можно наносить в виде матрицы высокой плотности, содержащей до 30 неидентичных молекул нуклеиновой кислоты на квадратный сантиметр или более, например до примерно 100 или даже 1000 на квадратный сантиметр. В микрочипах в качестве твердой подложки обычно используют стекло с покрытием, в отличие от материала на основе нитроцеллюлозы для фильтрующих матриц. Имея упорядоченный массив образцов комплементарных нуклеиновых кислот, положение каждого образца можно отслеживать и связывать с исходным образцом. Специалистам в данной области известны разнообразные матричные устройства, в которых множество различных зондов нуклеиновых кислот стабильно связаны с поверхностью твердой подложки. Подходящие подложки для массивов включают нейлон, стекло и кремний. Такие матрицы можно менять различными способами, включая среднюю длину зонда, последовательность или типы зондов, характер связи между зондом и поверхностью матрицы, например ковалентный или нековалентный, и т.п. Применение способов мечения и скрининга по настоящему изобретению и матриц не ограничено каким-либо параметром, за исключением того, что зонды должны позволять обнаружение уровней экспрессии; следовательно, способы и композиции могут быть использованы с множеством различных типов генов.
Типичные способы и устройства для изготовления микрочипов описаны, например, в патентах США №№ 5143854; 5202231; 5242974; 5288644; 5324633; 5384261; 5405783; 5412087; 5424186; 5429807; 5432049; 5436327; 5445934; 5468613; 5470710; 5472672; 5492806; 5503980; 5510270; 5525464; 5527681; 5529756; 5532128; 5545531; 5547839; 5554501; 5556752; 5561071; 5571639; 5580726; 5580732; 5593839; 5599695; 5599672; 5610287; 5624711; 5631134; 5639603; 5654413; 5658734; 5661028; 5665547; 5667972; 5695940; 5700637; 5744305; 5800992; 5807522; 5830645; 5837196; 5871928; 5847219; 5876932; 5919626; 6004755; 6087102; 6368799; 6383749; 6617112; 6638717; 6720138, а также WO 93/17126; WO 95/11995; WO 95/21265; WO 95/21944; WO 95/35505; WO 96/31622; WO 97/10365; WO 97/27317; WO 99/35505; WO 09923256; WO 09936760; WO 0138580; WO 0168255; WO 03020898; WO 03040410; WO 03053586; WO 03087297; WO 03091426; WO 03100012; WO 04020085; WO 04027093; ЕР 373203; ЕР 785280; EP 799897 и UK 8803000; раскрытие которых включено в настоящее описание в качестве ссылки.
Предполагается, что массивы могут представлять собой массивы высокой плотности, которые могут содержать 100 или более различных зондов. Предполагается, что они могут содержать 1000, 16000, 65000, 250000 или 1000000 или более различных зондов. Зонды могут быть направлены на мишени, находящиеся в одном или нескольких различных организмах. В некоторых вариантах осуществления длина олигонуклеотидных зондов составляет от 5 до 50, от 5 до 45, от 10 до 40 или от 15 до 40 нуклеотидов. В некоторых вариантах осуществления олигонуклеотидные зонды имеют длину от 20 до 25 нуклеотидов.
Расположение и последовательность каждой другой последовательности зонда в массиве общеизвестны. Кроме того, большое количество различных зондов может занимать относительно небольшую площадь, обеспечивая массив высокой плотности с плотностью зондов, как правило, более чем примерно 60, 100, 600, 1000, 5000, 10000, 40000, 100000 или 400000 различных олигонуклеотидных зондов на см2. Площадь поверхности массива может составлять примерно или менее чем примерно 1, 1,6, 2, 3, 4, 5, 6, 7, 8, 9 или 10 см2.
Кроме того, специалист в данной области может легко проанализировать данные, сгенерированные с помощью массива. Такие протоколы раскрыты выше и включают информацию, которую можно найти в заявках на патент №№ WO 9743450; WO 03023058; WO 03022421; WO 03029485; WO 03067217; WO 03066906; WO 03076928; WO 03093810; WO 03100448A1, каждая из которых специально включена в настоящее описание в качестве ссылки.
V. Примеры
Приведенные ниже примеры включены для демонстрации предпочтительных вариантов осуществления. Специалистам в данной области должно быть понятно, что методы, раскрытые в приведенных ниже примерах, являются методами, раскрытыми авторами изобретения для эффективного осуществления на практике вариантов осуществления изобретения, и, таким образом, могут рассматриваться как составляющие предпочтительные режимы его реализации. Однако специалистам в данной области техники в свете настоящего раскрытия должно быть понятно, что в конкретных раскрытых вариантах осуществления может быть сделано множество изменений, обеспечивающих такой же или аналогичный результат, без отступления от сущности и объема изобретения.
Пример 1
IFN-пролекарство на основе рецептора интерферона альфа и бета (IFNAR). Авторы изобретения разработали IFN-пролекарство на основе структуры человеческого иммуноглобулина G1 (IgG1) (фиг.1). Блокирующий реагент представляет собой внеклеточный домен (ECD) природного IFNAR1 или IFNAR2 и связан с N-концом интерферона альфа через расщепляемый линкер, являющийся субстратом. В этой форме IFNAR блокирует связывание с гетеродимером IFNAR1/IFNAR2 клеточной поверхности. Для увеличения стабильности и периода полураспада авторы изобретения выполнили слияние Fc-домена IgG1 с С-концом альфа-интерферона (фиг. 1).
Для определения, способен ли ECD либо IFNAR1, либо IFNAR2 блокировать активность альфа-интерферона, авторы изобретения очистили IFNa4-Fc, R1-IFNa4-Fc и R2-IFNa4-Fc. Линкер R1-IFNa4-Fc или R2-IFNa4-Fc представляет собой тройной пептид Gly-Gly-Gly-Gly-Ser, состоящий из 15 аминокислот, который является гибким аналогом, допускающим взаимодействие между доменами23. Были использованы клетки RAW-Lucia ISG, которые экспрессируют ген люциферазы Lucia под контролем минимального промотора ISG54 в сочетании с пятью элементами IFN-стимулированного ответа. Клетки RAW-Lucia ISG реагируют на мышиный IFN-α и IFN-β. Выполняли 5-кратное серийное разведение 20 нМ hIg, IFNa4-Fc, R1-IFNa4-Fc и R2-IFNa4-Fc для анализа активности люциферазы. Как R1-IFNa4-Fc, так и R2-IFNa4-Fc показали значимое снижение активности IFN по меньшей мере в 125 раз по сравнению с активностью IFNa4-Fc (фиг. 2A).
Для определения, была ли активность активированного IFN-пролекарства сопоставимой с активностью IFNa4-Fc, выполняли очистку R1-NSUB, R1-SUB, R2-NSUB и R2-SUB. Линкер R1-SUB или R2-SUB представляет собой пептид, состоящий из 16 аминокислот, несущий состоящий из 6 остатков субстрат (PVGLIG)24, расщепляемый протеазой, который может быть разрезан с помощью MMP-2 или MMP-9, фланкированный с двух сторон гибкими пептидами Gly-Gly-Gly-Gly-Ser. R1-NSUB и R2-NSUB без расщепляемого протеазой субстрата использовали в качестве контрольных конструкций. R1-NSUB, R1-SUB, R2-NSUB или R2-SUB инкубировали с rmMMP-9 при 37°C в течение 6 часов для активации. Активированный R1-SUB показал увеличение активности IFN в 25 раз по сравнению с активностью IFNa4-Fc, тогда как активность R1-NSUB не изменилась после введения фермента (фиг.2B). Аналогичный результат наблюдали с R2-NSUB и R2-SUB, при этом активированный R2-SUB показал еще более сильное увеличение активности IFN, более чем в 25 раз, по сравнению с IFNa4-Fc (фиг.2C).
Таким образом, биологическая активность активированного протеазой IFN-пролекарства увеличивалась в обеих формах на основе IFNA1 и на основе IFNAR2, а не при ожидаемом восстановлении до эквивалентного родительского IFNa4-Fc. Более того, до расщепления протеазой IFN-пролекарство, по-видимому, имело заметно сниженную биологическую активность, что позволяет предположить, что по своему действию IFN-пролекарство будет безопасным in vivo до расщепления, но после расщепления в локальном микроокружении опухоли терапевтическая активность увеличится.
IFN-пролекарство на основе IFNAR1 и IFNAR2 демонстрирует различные противоопухолевые эффекты in vivo. Затем на мышиной модели меланомы В16 была изучена возможность преобразования локальной активации IFN-пролекарства в противоопухолевую эффективность in vivo. Без скрининга экспрессии фермента в мышиных моделях опухолей сначала изучали IFN-пролекарство, несущее расщепляемый субстрат ММР-2 и ММР-9. Мышей с установленными опухолями меланомы В16 лечили R1-SUB, R2-SUB, IFNa4-Fc или контрольным hIg. При дозе 1 нмоль (три инъекции каждые три дня) R1-SUB, R2-SUB и IFNa4-Fc имели разную эффективность в отношении подавления роста опухоли в этой модели. Через 10 дней после лечения R1-SUB ингибировал рост опухоли на 24%, R2-SUB на 57% и IFNa4-Fc на 72% относительно контрольной группы, получавшей hIg (фиг.3).
Следовательно, IFN-пролекарство на основе IFNAR2 показало более высокие противоопухолевые эффекты in vivo чем IFN-пролекарство на основе IFNAR1, что дало основание сосредоточить основное внимание в следующем исследовании на R2-SUB. Однако R2-SUB, несущий расщепляемый субстрат ММР-2 и ММР-9 имел более низкую эффективность в отношении подавления роста опухоли по сравнению с IFNa4-Fc. Было выдвинуто предположение, что улучшение эффективности IFN-пролекарства в мышиных моделях опухолей может быть достигнуто путем выбора подходящего расщепляемого линкера, являющегося субстратом.
IFN-пролекарство на основе IFNAR2 имеет улучшенную эффективность и уменьшенные побочные эффекты. Для выбора более выгодного расщепляемого линкера, являющегося субстратом, на мышиных моделях опухолей был выполнен скрининг наиболее изученных протеаз, которые, как известно, являются активированными во многих различных раковых заболеваниях человека. Этот процесс выбора субстрата включал контр-отбор в отношении протеаз, экспрессируемых в нормальных здоровых тканях, для снижения вероятности системной (неспецифической) активации IFN-пролекарства. Мышей с установленными опухолями меланомы B16 или опухолями толстой кишки MC38 умерщвляли, а опухолевые ткани и нормальные ткани, включая селезенку, сердце, печень, легкое и почку, использовали для определения уровня экспрессии мРНК следующих генов протеаз: uPA, MMP-2, MMP-9 и MMP-14. uPA имел высокие уровни экспрессии только в почках, но не в двух опухолевых тканях (фиг.4А). ММР-2 имел высокие уровни экспрессии в опухолях МС38, но не в опухолях В16; однако он имел высокий фон в сердце и легких (фиг. 4B). Уровень экспрессии ММР-9 был очень низким во всех нормальных и опухолевых тканях (фиг.4С). Наконец, опухоли MC38 и B16 имели высокий уровень экспрессии MMP-14, тогда как, напротив, экспрессия MMP-14 в нормальных тканях оказалась минимальной (фиг.4D). Данные скрининга на мышиных моделях опухолей показали, что расщепляемый линкер, являющийся субстратом, чувствительным к ММР-14, можно использовать для получения потенциального IFN-пролекарства in vivo.
Также был проанализирован уровень экспрессии человеческих протеаз между опухолью и соседними нормальными тканями. Модуль DiffExp в TIMER (ресурсе для оценки опухолевого иммунитета) обеспечивает сравнение уровней экспрессии генов для всех образцов из TCGA (Атлас генома рака). Была проведена оценка большинства ассоциированных с опухолью ферментов, и было обнаружено, что ММР-1, ММР-3, ММР-9, ММР-10, ММР-11, ММР-12, ММР-13 и ММР-14 являются протеазами, которые являются сильно активированными в большинстве опухолей, но имеют относительно низкий фон в нормальных тканях (фиг. 6A-H). Эти протеазы являются отличными кандидатами для разработки пролекарства человеческого IFN для достижения максимального противоопухолевого эффекта и минимального побочного воздействия на нормальные ткани. Интересно, что согласованные данные об уровне экспрессии ММР-14 в образцах как у мышей, так и у людей увеличивают надежность выбора субстрата ММР-14 в получении IFN-пролекарства.
Поэтому было разработано и очищено IFN-пролекарство на основе IFNAR2 и субстрата MMP-14 (SGRSENIRTA)25. Было проведено изучение потенциальных преимуществ в отношении безопасности и противоопухолевой эффективности in vivo. Мышей с установленными опухолями меланомы B16-OVA лечили R2-SUB, IFNa4-Fc или контрольным hIg, которые вводили в виде трех инъекций в дозе 1 мМ в течение каждых трех дней. Было проведено сравнение относительной безопасности IFN-пролекарства и IFNa4-Fc. Высокие уровни воспалительных цитокинов в сыворотке, включая IL-6, TNF, MCP-1 и IFN-g, наблюдались в группе IFNa4-Fc на следующий день после третьего введения. Напротив, мыши, получавшие R2-SUB, показали очень низкие уровни этих воспалительных цитокинов (фиг. 5А). Уровень ALT, указывающий на токсичность по отношению к печени, также показал аналогичные результаты (фиг.5B).
Продолжали выполнять мониторинг роста опухоли и массы тела. Через 12 дней после лечения R2-SUB ингибировал рост опухоли на 76%, а IFNa4-Fc на 67% по сравнению с контрольной группой, получавшей лечение hIg (фиг.5C). Однако в этот момент мыши из группы, получавшей IFN-4-Fc, выглядели больными (отстраненными, неактивными и худыми), вероятно, из-за серьезных побочных эффектов, вызванных IFN. Затем было проведено сравнение кинетики веса тела. Данные показали, что мыши из группы IFNa4-Fc начали быстро терять вес после третьего введения, однако масса тела мышей из группы R2-SUB значительно не изменилась после трех введений (фиг.5D). Наконец, на кривой выживаемости, основанной на токсичности, было показано, что группа, получавшая R2-SUB, имела выживаемость 100%, в то время как группа, получавшая IFNa4-Fc, продемонстрировала выживаемость 33% (фиг.5E).
Пролекарство человеческого IFN основе рецептора интерферона альфа и бета (IFNAR).
Для определения, способен ли ECD человеческого IFNAR1 или IFNAR2 блокировать активность человеческого интерферона альфа 2, была выполнена очистка человеческих IFNa2-Fc, R1-IFNa2-Fc и R2-IFNa2-Fc. Линкер R1-IFNa2-Fc или R2-IFNa2-Fc представлял собой тройной пептид Gly-Gly-Gly-Gly-Ser, состоящий из 15 аминокислот (SEQ ID NO: 13), который представляет собой гибкий линкер, допускающий взаимодействие между доменами23. Были использованы репортерные клетки 293T-DualТМ hSTING-R232, экспрессирующие репортерный ген SEAP (секретируемая эмбриональная щелочная фосфатаза) под контролем минимального промотора IFN-β, слитого с пятью связывающими участками NF-κB и AP-1. Клетки 293T-DualТМ hSTING-R232 чувствительны к человеческому IFN-α и IFN-β. Выполняли 10-кратное серийное разведение 50 нМ человеческих IFNa2-Fc, R1-IFNa2-Fc и R2-IFNa2-Fc для анализа гена-репортера SEAP. По сравнению с человеческим IFNa2-Fc, R1-IFNa2-Fc показал уменьшение активности IFN в 10 раз, тогда как R2-IFNa2-Fc показал уменьшение активности IFN в 100 раз (фиг.7A). Таким образом, IFNAR2 является более эффективным блокирующим агентом для человеческого IFN.
Для определения, является ли активность активированного человеческого IFN-пролекарства сопоставимой с активностью IFNa2-Fc, выполняли очистку человеческих R2-NSUB и R2-SUB. Линкер R2-SUB представляет собой пептид, состоящий из 16 аминокислот, несущий состоящий из 6 остатков субстрат (PVGLIG)24, расщепляемый протеазой, который можно разрезать с помощью MMP-2 или MMP-9, фланкированный с обеих сторон гибким пептидом Gly-Gly-Gly-Gly-Сер. R2-NSUB без расщепляемого протеазой субстрата использовали в качестве контрольных конструкций. R2-NSUB или R2-SUB инкубировали с rmMMP-9 при 37°C в течение 0, 0,5, 2 или 6 часов. Человеческий R2-SUB расщепляли rmMMP-9 в зависимости от времени. Через 6 часов эффективность расщепления составила 100% (фиг. 7C). Активированный человеческий R2-SUB показал сопоставимую активность IFN по сравнению с человеческим IFNa2-Fc, тогда как активность R2-NSUB не изменилась после введения фермента (фиг.7B).
Таким образом, биологическая активность активируемого протеазой пролекарства человеческого IFN полностью восстанавливалась в форме на основе IFNAR2 по сравнению с родительским IFNa4-Fc. Кроме того, пролекарство человеческого IFN на основе IFNAR2, по-видимому, имело заметно уменьшенную биологическую активность до расщепления протеазой, что свидетельствует о том, что пролекарство человеческого IFN будет до расщепления безопасным in vivo, но после расщепления в локальном микроокружении опухоли у человека его терапевтическая активность увеличится.
Таким образом, изученное в настоящей заявке IFN-пролекарство обеспечивает улучшенное нацеливание интерферона I типа на опухолевую ткань за счет уменьшения токсичности в нормальных тканях. Противоопухолевая эффективность IFN-пролекарства может быть адаптирована с учетом активности протеазы в каждой опухоли, что в определенных ситуациях делает IFN-пролекарство персонализированным лекарственным средством с выбранным расщепляемым линкером, являющимся субстратом, обеспечивающим наилучший результат. Кроме того, IFN-пролекарство может быть разработано для переноса нескольких доменов IFN с использованием разных линкеров, которые чувствительны к разным протеазам, тем самым значительно улучшая его терапевтический эффект.
Все композиции и способы, раскрытые и заявленные в настоящем описании, могут быть изготовлены и выполнены с учетом настоящего описания без чрезмерного экспериментирования. Хотя композиции и способы по изобретению описаны в терминах предпочтительных вариантов осуществления, специалистам в данной области техники должно быть очевидно, что могут быть введены изменения в композиции и способы, а также в этапы способов и в последовательность выполнения этапов без отступления от концепции, сущности и объема изобретения. Более конкретно, будет очевидно, что определенные агенты, которые являются как химически, так и физиологически родственными, могут заменить агенты, описанные в настоящей заявке, с достижением таких же или аналогичных результатов. Все подобные аналогичные заместители и модификации, очевидные специалистам в данной области техники, считаются находящимися в пределах сущности, объема и концепции изобретения, определенного в прилагаемой формуле изобретения.
VII. ЛИТЕРАТУРА
Приведенные ниже документы в той мере, в которой они предоставляют примеры выполнения процедур или иные подробности, дополняющие изложенную в настоящем описании информацию, специально включены в настоящее описание в качестве ссылки.
Almendro et al., J. Immunol., 157(12):5411-5421, 1996.
Antonelli et al., Cytokine Growth Factor Rev 26, 121-131, 2015.
Ausubel et al., Current Protocols in Molecular Biology, Greene Publishing Associates and Wiley Interscience, N.Y., 1994.
Bates, Mol. Biotechnol., 2(2):135-145, 1994.
Battraw and Hall, Theor. App. Genet., 82(2):161-168, 1991.
Bellus, J. Macromol. Sci. Pure Appl. Chem., A31(1): 1355-1376, 1994.
Bhattacharjee et al., J. Plant Bioch. Biotech., 6(2):69-73. 1997.
Burnette et al., Cancer Res 71, 2488-2496, 2011.
Capaldi et al., Biochem. Biophys. Res. Comm., 74(2):425-433, 1977.
Carbonelli et al., FEMS Microbiol. Lett., 177(1):75-82, 1999.
Chen and Okayama, Mol. Cell Biol., 7(8):2745-2752, 1987.
Christou et al., Proc. Natl. Acad. Sci. USA, 84(12):3962-3966, 1987.
Cocea, Biotechniques, 23(5):814-816, 1997.
D’Halluin et al., Plant Cell, 4(12):1495-1505, 1992.
De Jager et al., Semin. Nucl. Med., 23(2):165-179, 1993.
Desnoyers et al., Sci Transl Med 5, 207ra144, 2013.
Doolittle and Ben-Zeev, Methods Mol. Biol., 109, :215-237, 1999.
EPO 0273085
Erster, et al., J Control Release 161, 804-812, 2012.
Fechheimer et al., Proc Natl. Acad. Sci. USA, 84:8463-8467, 1987.
Ferrantini et al., Biochimie 89, 884-893, 2007.
Fraley et al., Proc. Natl. Acad. Sci. USA, 76:3348-3352, 1979.
Frohman, In: PCR Protocols: A Guide To Methods And Applications, Academic Press, N.Y., 1990.
Gabrilovich & Nagaraj, Nat Rev Immunol 9, 162-174, 2009.
GB Application No. 2 202 328
Ghosh and Bachhawat, In: Liver Diseases, Targeted Diagnosis and Therapy Using Specific Receptors and Ligands, Wu et al. (Eds.), Marcel Dekker, NY, 87-104, 1991.
Gopal, Mol. Cell Biol., 5:1188-1190, 1985.
Graham and Van Der Eb, Virology, 52:456-467, 1973.
Greene et al., Immunology Today, 10:272, 1989.
Harland and Weintraub, J. Cell Biol., 101(3):1094-1099, 1985.
He et al., Plant Cell Reports, 14 (2-3):192-196, 1994.
Hou and Lin, Plant Physiology, 111:166, 1996.
Imai et al., Nephrologie, 19(7):397-402, 1998.
Kaeppler et al., Plant Cell Rep., 8:415-418, 1990.
Kaneda et al., Science, 243:375-378, 1989.
Kato et al., J. Biol. Chem., 266:3361 3364, 1991.
Kato et al., N Engl J Med, 350:1713-21, 2004.
Khanna, Immunol Cell Biol 76, 20-26, 1998.
Kraus et al., FEBS Lett., 428(3):165-170, 1998.
Kreutzer et al., J Dtsch Dermatol Ges 2, 689-694, 2004.
Kwoh et al., Proc. Natl. Acad. Sci. USA, 86:1173, 1989.
Lareyre et al., J. Biol. Chem., 274(12):8282-8290, 1999.
Lazzeri, Methods Mol. Biol., 49:95-106, 1995.
Lee et al., Clin Cancer Res, 14:7397-404, 2008.
Lee et al., Environ. Mol. Mutagen., 13(1):54-59, 1989.
Levenson et al., Hum. Gene Ther., 9(8):1233-1236, 1998.
Lotrich, Dialogues Clin Neurosci 11, 417-425, 2009.
Macejak and Sarnow, Nature, 353:90-94, 1991.
Moschos & Kirkwood, Cytokine Growth Factor Rev 18, 451-458, 2007.
Nakamura et al., In: Handbook of Experimental Immunology (4th Ed.), Weir et al. (Eds), 1:27, Blackwell Scientific Publ., Oxford, 1987.
Nicolau and Sene, Biochim. Biophys. Acta, 721:185-190, 1982.
Nicolau et al., Methods Enzymol., 149:157-176, 1987.
Nomoto et al., Gene, 236(2):259-271, 1999.
Ohara et al., Proc. Natl. Acad. Sci. USA, 86:5673-5677, 1989.
Olaussen et al., Curr Opin Pulm Med, 13:284-9, 2007.
Olaussen et al., N Engl J Med, 355:983-91, 2006.
Omirulleh et al., Plant Mol. Biol., 21(3):415-428, 1993.
Pace et al., J Immunol 184, 5969-5979, 2010.
Parker et al., Nat Rev Cancer 16, 131-144, 2016.
PCT Appln. WO 92/17598
PCT Appln. WO 94/09699
Pelletier and Sonenberg, Nature, 334(6180):320-325, 1988.
Perales et al., Proc. Natl. Acad. Sci. USA, 91:4086 4090, 1994.
Potrykus et al., Mol. Gen. Genet., 199(2):169-177, 1985.
Potter et al., Proc. Natl. Acad. Sci. USA, 81:7161-7165, 1984.
Remington’s Pharmaceutical Sciences, 15th Ed., 33:624-652, 1990.
Rhodes et al., Methods Mol. Biol., 55:121 131, 1995.
Rippe, et al., Mol. Cell Biol., 10:689-695, 1990.
Roepman et al., Clin Cancer Res, 15:284-90, 2009.
Sambrook et al., In: Molecular Cloning: A Laboratory Manual, 2nd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989.
Schiavoni et al., Cancer Res 71, 768-778, 2011.
Schmidt et al., Front Immunol 3, 51, 2012.
Sharma et al., Cell 168, 707-723, 2017.
Sleijfer et al., Pharm World Sci 27, 423-431, 2005.
Srivastava et al., J Exp Med 211, 961-974, 2014.
Stagg et al., Proc Nat’l Acad Sci USA 108, 7142-7147, 2011.
Stella et al., Drugs 29, 455-473, 1985.
Tsukada et al., Plant Cell Physiol., 30(4)599-604, 1989.
Tsumaki et al., J. Biol. Chem., 273(36):22861-22864, 1998.
Tur-Kaspa et al., Mol. Cell Biol., 6:716-718, 1986.
Патент США 3817837
Патент США 3850752
Патент США 3939350
Патент США 3939350
Патент США 3996345
Патент США 4275,149
Патент США 4277437
Патент США 4366241
Патент США 4415723
Патент США 4458066
Патент США 4683195
Патент США 4683202
Патент США 4684611
Патент США 4800159
Патент США 4879236
Патент США 4883750
Патент США 4952500
Патент США 5143854
Патент США 5202231
Патент США 5242974
Патент США 5279721
Патент США 5288644
Патент США 5302523
Патент США 5322783
Патент США 5324633
Патент США 5384253
Патент США 5384261
Патент США 5405783
Патент США 5412087
Патент США 5424186
Патент США 5429807
Патент США 5432049
Патент США 5436327
Патент США 5445934
Патент США 5464765
Патент США 5468613
Патент США 5470710
Патент США 5472672
Патент США 5492806
Патент США 5503980
Патент США 5510270
Патент США 5525464
Патент США 5527681
Патент США 5529756
Патент США 5532128
Патент США 5538877
Патент США 5538880
Патент США 5545531
Патент США 5547839
Патент США 5550318
Патент США 5554501
Патент США 5556752
Патент США 5561071
Патент США 5563055
Патент США 5571639
Патент США 5580726
Патент США 5580732
Патент США 5580859
Патент США 5589466
Патент США 5593839
Патент США 5599672
Патент США 5599695
Патент США 5610042
Патент США 5610287
Патент США 5624711
Патент США 5631134
Патент США 5639603
Патент США 5654413
Патент США 5656610
Патент США 5658734
Патент США 5661028
Патент США 5665547
Патент США 5667972
Патент США 5670488
Патент США 5695940
Патент США 5700637
Патент США 5702932
Патент США 5736524
Патент США 5739018
Патент США 5739169
Патент США 5744305
Патент США 5780448
Патент США 5789215
Патент США 5795715
Патент США 5800992
Патент США 5801005
Патент США 5807522
Патент США 5824311
Патент США 5830645
Патент США 5830880
Патент США 5837196
Патент США 5840873
Патент США 5843640
Патент США 5843650
Патент США 5843651
Патент США 5843663
Патент США 5846708
Патент США 5846709
Патент США 5846717
Патент США 5846726
Патент США 5846729
Патент США 5846783
Патент США 5846945
Патент США 5847219
Патент США 5849481
Патент США 5849486
Патент США 5849487
Патент США 5849497
Патент США 5849546
Патент США 5849547
Патент США 5851772
Патент США 5853990
Патент США 5853992
Патент США 5853993
Патент США 5856092
Патент США 5858652
Патент США 5861244
Патент США 5863732
Патент США 5863753
Патент США 5866331
Патент США 5866366
Патент США 5871928
Патент США 5871986
Патент США 5876932
Патент США 5882864
Патент США 5889136
Патент США 5900481
Патент США 5905024
Патент США 5910407
Патент США 5912124
Патент США 5912145
Патент США 5912148
Патент США 5916776
Патент США 5916779
Патент США 5919626
Патент США 5919630
Патент США 5922574
Патент США 5925517
Патент США 5925565
Патент США 5928862
Патент США 5928869
Патент США 5928905
Патент США 5928906
Патент США 5928906
Патент США 5929227
Патент США 5932413
Патент США 5932451
Патент США 5935791
Патент США 5935819
Патент США 5935825
Патент США 5939291
Патент США 5942391
Патент США 5945100
Патент США 5981274
Патент США 5994624
Патент США 6004755
Патент США 6087102
Патент США 6368799
Патент США 6383749
Патент США 6506559
Патент США 6573099
Патент США 6617112
Патент США 6638717
Патент США 6720138
Публикация заявки на патент США 2002/0168707
Публикация заявки на патент США 2003/0051263
Публикация заявки на патент США 2003/0055020
Публикация заявки на патент США 2003/0159161
Публикация заявки на патент США 2004/0064842
Публикация заявки на патент США 2004/0265839
Публикация заявки на патент США 2008/0009439
Wagner et al., Proc. Natl. Acad. Sci. USA 87(9):3410-3414, 1990.
Walker et al., Nucleic Acids Res. 20(7):1691-1696, 1992.
Weir et al., Nature, 450:893-8, 2007.
Woo et al., Immunity 41, 830-842, 2014.
Wu and Wu, Adv. Drug Delivery Rev., 12:159 167, 1993.
Wu and Wu, J. Biol. Chem., 262:4429 4432, 1987.
Wu et al., Biochem. Biophys. Res. Commun., 233(1):221-226, 1997.
Wu et al., Cancer Res., 58(8): 1605-8, 1998.
Zhao-Emonet et al., Biochim. Biophys. Acta, 1442(2-3):109-119, 1998.
Zhou et al., Nature, 361(6412):543-547 , 1993.
Zitvogel et al., Nat Rev Immunol 15, 405-414, 2015.
Zoglmeier et al., Clin Cancer Res 17, 1765-1775, 2011.
--->
СПИСКИ ПОСЛЕДОВАТЕЛЬНСТЕЙ
<110> The Board of Regents of the University of Texas System
<120> ПРОЛЕКАРСТВО ИНТЕРФЕРОНА ДЛЯ ЛЕЧЕНИЯ РАКА
<130> UTFD.P3214WO
<140> PCT/US2018/037982
<141> 2018-06-18
<150> US 62/522,564
<151> 2017-06-20
<160> 13
<170> PatentIn version 3.5
<210> 1
<211> 429
<212> Белок
<213> Mus musculus
<400> 1
Met Leu Ala Val Val Gly Ala Ala Ala Leu Val Leu Val Ala Gly Ala
1 5 10 15
Pro Trp Val Leu Pro Ser Ala Ala Gly Gly Glu Asn Leu Lys Pro Pro
20 25 30
Glu Asn Ile Asp Val Tyr Ile Ile Asp Asp Asn Tyr Thr Leu Lys Trp
35 40 45
Ser Ser His Gly Glu Ser Met Gly Ser Val Thr Phe Ser Ala Glu Tyr
50 55 60
Arg Thr Lys Asp Glu Ala Lys Trp Leu Lys Val Pro Glu Cys Gln His
65 70 75 80
Thr Thr Thr Thr Lys Cys Glu Phe Ser Leu Leu Asp Thr Asn Val Tyr
85 90 95
Ile Lys Thr Gln Phe Arg Val Arg Ala Glu Glu Gly Asn Ser Thr Ser
100 105 110
Ser Trp Asn Glu Val Asp Pro Phe Ile Pro Phe Tyr Thr Ala His Met
115 120 125
Ser Pro Pro Glu Val Arg Leu Glu Ala Glu Asp Lys Ala Ile Leu Val
130 135 140
His Ile Ser Pro Pro Gly Gln Asp Gly Asn Met Trp Ala Leu Glu Lys
145 150 155 160
Pro Ser Phe Ser Tyr Thr Ile Arg Ile Trp Gln Lys Ser Ser Ser Asp
165 170 175
Lys Lys Thr Ile Asn Ser Thr Tyr Tyr Val Glu Lys Ile Pro Glu Leu
180 185 190
Leu Pro Glu Thr Thr Tyr Cys Leu Glu Val Lys Ala Ile His Pro Ser
195 200 205
Leu Lys Lys His Ser Asn Tyr Ser Thr Val Gln Cys Ile Ser Thr Thr
210 215 220
Val Ala Asn Lys Met Pro Val Pro Gly Asn Leu Gln Val Asp Ala Gln
225 230 235 240
Gly Lys Ser Tyr Val Leu Lys Trp Asp Tyr Ile Ala Ser Ala Asp Val
245 250 255
Leu Phe Arg Ala Gln Trp Leu Pro Gly Tyr Ser Lys Ser Ser Ser Gly
260 265 270
Ser Arg Ser Asp Lys Trp Lys Pro Ile Pro Thr Cys Ala Asn Val Gln
275 280 285
Thr Thr His Cys Val Phe Ser Gln Asp Thr Val Tyr Thr Gly Thr Phe
290 295 300
Phe Leu His Val Gln Ala Ser Glu Gly Asn His Thr Ser Phe Trp Ser
305 310 315 320
Glu Glu Lys Phe Ile Asp Ser Gln Lys His Ile Leu Pro Pro Pro Pro
325 330 335
Val Ile Thr Val Thr Ala Met Ser Asp Thr Leu Leu Val Tyr Val Asn
340 345 350
Cys Gln Asp Ser Thr Cys Asp Gly Leu Asn Tyr Glu Ile Ile Phe Trp
355 360 365
Glu Asn Thr Ser Asn Thr Lys Ile Ser Met Glu Lys Asp Gly Pro Glu
370 375 380
Phe Thr Leu Lys Asn Leu Gln Pro Leu Thr Val Tyr Cys Val Gln Ala
385 390 395 400
Arg Val Leu Phe Arg Ala Leu Leu Asn Lys Thr Ser Asn Phe Ser Glu
405 410 415
Lys Leu Cys Glu Lys Thr Arg Pro Gly Ser Phe Ser Thr
420 425
<210> 2
<211> 242
<212> Белок
<213> Mus musculus
<400> 2
Met Arg Ser Arg Cys Thr Val Ser Ala Val Gly Leu Leu Ser Leu Cys
1 5 10 15
Leu Val Val Ser Ala Ser Leu Glu Thr Ile Thr Pro Ser Ala Phe Asp
20 25 30
Gly Tyr Pro Asp Glu Pro Cys Thr Ile Asn Ile Thr Ile Arg Asn Ser
35 40 45
Arg Leu Ile Leu Ser Trp Glu Leu Glu Asn Lys Ser Gly Pro Pro Ala
50 55 60
Asn Tyr Thr Leu Trp Tyr Thr Val Met Ser Lys Asp Glu Asn Leu Thr
65 70 75 80
Lys Val Lys Asn Cys Ser Asp Thr Thr Lys Ser Ser Cys Asp Val Thr
85 90 95
Asp Lys Trp Leu Glu Gly Met Glu Ser Tyr Val Val Ala Ile Val Ile
100 105 110
Val His Arg Gly Asp Leu Thr Val Cys Arg Cys Ser Asp Tyr Ile Val
115 120 125
Pro Ala Asn Ala Pro Leu Glu Pro Pro Glu Phe Glu Ile Val Gly Phe
130 135 140
Thr Asp His Ile Asn Val Thr Met Glu Phe Pro Pro Val Thr Ser Lys
145 150 155 160
Ile Ile Gln Glu Lys Met Lys Thr Thr Pro Phe Val Ile Lys Glu Gln
165 170 175
Ile Gly Asp Ser Val Arg Lys Lys His Glu Pro Lys Val Asn Asn Val
180 185 190
Thr Gly Asn Phe Thr Phe Val Leu Arg Asp Leu Leu Pro Lys Thr Asn
195 200 205
Tyr Cys Val Ser Leu Tyr Phe Asp Asp Asp Pro Ala Ile Lys Ser Pro
210 215 220
Leu Lys Cys Ile Val Leu Gln Pro Gly Gln Glu Ser Gly Leu Ser Glu
225 230 235 240
Ser Ala
<210> 3
<211> 436
<212> Белок
<213> Homo sapiens
<400> 3
Met Met Val Val Leu Leu Gly Ala Thr Thr Leu Val Leu Val Ala Val
1 5 10 15
Ala Pro Trp Val Leu Ser Ala Ala Ala Gly Gly Lys Asn Leu Lys Ser
20 25 30
Pro Gln Lys Val Glu Val Asp Ile Ile Asp Asp Asn Phe Ile Leu Arg
35 40 45
Trp Asn Arg Ser Asp Glu Ser Val Gly Asn Val Thr Phe Ser Phe Asp
50 55 60
Tyr Gln Lys Thr Gly Met Asp Asn Trp Ile Lys Leu Ser Gly Cys Gln
65 70 75 80
Asn Ile Thr Ser Thr Lys Cys Asn Phe Ser Ser Leu Lys Leu Asn Val
85 90 95
Tyr Glu Glu Ile Lys Leu Arg Ile Arg Ala Glu Lys Glu Asn Thr Ser
100 105 110
Ser Trp Tyr Glu Val Asp Ser Phe Thr Pro Phe Arg Lys Ala Gln Ile
115 120 125
Gly Pro Pro Glu Val His Leu Glu Ala Glu Asp Lys Ala Ile Val Ile
130 135 140
His Ile Ser Pro Gly Thr Lys Asp Ser Val Met Trp Ala Leu Asp Gly
145 150 155 160
Leu Ser Phe Thr Tyr Ser Leu Val Ile Trp Lys Asn Ser Ser Gly Val
165 170 175
Glu Glu Arg Ile Glu Asn Ile Tyr Ser Arg His Lys Ile Tyr Lys Leu
180 185 190
Ser Pro Glu Thr Thr Tyr Cys Leu Lys Val Lys Ala Ala Leu Leu Thr
195 200 205
Ser Trp Lys Ile Gly Val Tyr Ser Pro Val His Cys Ile Lys Thr Thr
210 215 220
Val Glu Asn Glu Leu Pro Pro Pro Glu Asn Ile Glu Val Ser Val Gln
225 230 235 240
Asn Gln Asn Tyr Val Leu Lys Trp Asp Tyr Thr Tyr Ala Asn Met Thr
245 250 255
Phe Gln Val Gln Trp Leu His Ala Phe Leu Lys Arg Asn Pro Gly Asn
260 265 270
His Leu Tyr Lys Trp Lys Gln Ile Pro Asp Cys Glu Asn Val Lys Thr
275 280 285
Thr Gln Cys Val Phe Pro Gln Asn Val Phe Gln Lys Gly Ile Tyr Leu
290 295 300
Leu Arg Val Gln Ala Ser Asp Gly Asn Asn Thr Ser Phe Trp Ser Glu
305 310 315 320
Glu Ile Lys Phe Asp Thr Glu Ile Gln Ala Phe Leu Leu Pro Pro Val
325 330 335
Phe Asn Ile Arg Ser Leu Ser Asp Ser Phe His Ile Tyr Ile Gly Ala
340 345 350
Pro Lys Gln Ser Gly Asn Thr Pro Val Ile Gln Asp Tyr Pro Leu Ile
355 360 365
Tyr Glu Ile Ile Phe Trp Glu Asn Thr Ser Asn Ala Glu Arg Lys Ile
370 375 380
Ile Glu Lys Lys Thr Asp Val Thr Val Pro Asn Leu Lys Pro Leu Thr
385 390 395 400
Val Tyr Cys Val Lys Ala Arg Ala His Thr Met Asp Glu Lys Leu Asn
405 410 415
Lys Ser Ser Val Phe Ser Asp Ala Val Cys Glu Lys Thr Lys Pro Gly
420 425 430
Asn Thr Ser Lys
435
<210> 4
<211> 243
<212> Белок
<213> Homo sapiens
<400> 4
Met Leu Leu Ser Gln Asn Ala Phe Ile Phe Arg Ser Leu Asn Leu Val
1 5 10 15
Leu Met Val Tyr Ile Ser Leu Val Phe Gly Ile Ser Tyr Asp Ser Pro
20 25 30
Asp Tyr Thr Asp Glu Ser Cys Thr Phe Lys Ile Ser Leu Arg Asn Phe
35 40 45
Arg Ser Ile Leu Ser Trp Glu Leu Lys Asn His Ser Ile Val Pro Thr
50 55 60
His Tyr Thr Leu Leu Tyr Thr Ile Met Ser Lys Pro Glu Asp Leu Lys
65 70 75 80
Val Val Lys Asn Cys Ala Asn Thr Thr Arg Ser Phe Cys Asp Leu Thr
85 90 95
Asp Glu Trp Arg Ser Thr His Glu Ala Tyr Val Thr Val Leu Glu Gly
100 105 110
Phe Ser Gly Asn Thr Thr Leu Phe Ser Cys Ser His Asn Phe Trp Leu
115 120 125
Ala Ile Asp Met Ser Phe Glu Pro Pro Glu Phe Glu Ile Val Gly Phe
130 135 140
Thr Asn His Ile Asn Val Met Val Lys Phe Pro Ser Ile Val Glu Glu
145 150 155 160
Glu Leu Gln Phe Asp Leu Ser Leu Val Ile Glu Glu Gln Ser Glu Gly
165 170 175
Ile Val Lys Lys His Lys Pro Glu Ile Lys Gly Asn Met Ser Gly Asn
180 185 190
Phe Thr Tyr Ile Ile Asp Lys Leu Ile Pro Asn Thr Asn Tyr Cys Val
195 200 205
Ser Val Tyr Leu Glu His Ser Asp Glu Gln Ala Val Ile Lys Ser Pro
210 215 220
Leu Lys Cys Thr Leu Leu Pro Pro Gly Gln Glu Ser Glu Ser Ala Glu
225 230 235 240
Ser Ala Lys
<210> 5
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 5
Pro Val Gly Leu Ile Gly
1 5
<210> 6
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 6
Gly Gly Tyr Ala Glu Leu Arg Met Gly Gly
1 5 10
<210> 7
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 7
Gly Pro Arg Pro Phe Asn Tyr Leu
1 5
<210> 8
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 8
Gly Gly Ala Leu Gly Leu Ser Leu
1 5
<210> 9
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 9
Gly Pro Met Ser Tyr Asn Ala Leu
1 5
<210> 10
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 10
Ser Gly Arg Ser Glu Asn Ile Arg Thr Ala
1 5 10
<210> 11
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 11
Thr Ser Gly Pro Asn Gln Glu Gln Lys
1 5
<210> 12
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 12
Gly Phe Leu Gly
1
<210> 13
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 13
Gly Gly Gly Gly Ser
1 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ГЛИКАНЗАВИСИМЫЕ ИММУНОТЕРАПЕВТИЧЕСКИЕ МОЛЕКУЛЫ | 2016 |
|
RU2754661C2 |
| ЛЕЧЕНИЕ РАКА С ИСПОЛЬЗОВАНИЕМ ХИМЕРНОГО АНТИГЕНСПЕЦИФИЧЕСКОГО РЕЦЕПТОРА НА ОСНОВЕ ГУМАНИЗИРОВАННОГО АНТИТЕЛА ПРОТИВ EGFRvIII | 2014 |
|
RU2831418C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ M971 | 2018 |
|
RU2770411C1 |
| СПОСОБЫ ПОЛУЧЕНИЯ ЭКСПРЕССИРУЮЩИХ ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР КЛЕТОК | 2015 |
|
RU2751362C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ, НАЦЕЛЕННЫЕ НА АНТИГЕН СОЗРЕВАНИЯ B-КЛЕТОК | 2013 |
|
RU2766608C2 |
| ТЕРАПЕВТИЧЕСКИЕ СРЕДСТВА И СПОСОБЫ ДЛЯ ЛЕЧЕНИЯ TLR2-ОПОСРЕДОВАННЫХ ЗАБОЛЕВАНИЙ И НАРУШЕНИЙ | 2019 |
|
RU2791022C2 |
| ВАКЦИНА И СПОСОБЫ ОБНАРУЖЕНИЯ И ПРОФИЛАКТИКИ ФИЛЯРИОЗА | 2021 |
|
RU2832185C1 |
| EGFL6-СПЕЦИФИЧЕСКИЕ МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2779902C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЕГРАДАЦИИ НЕПРАВИЛЬНО УПАКОВАННЫХ БЕЛКОВ | 2016 |
|
RU2761564C2 |
| КОМПОЗИЦИИ Т-КЛЕТОК С НЕДОСТАТОЧНОСТЬЮ РЕЦЕПТОРОВ Т-КЛЕТОК | 2013 |
|
RU2781979C2 |
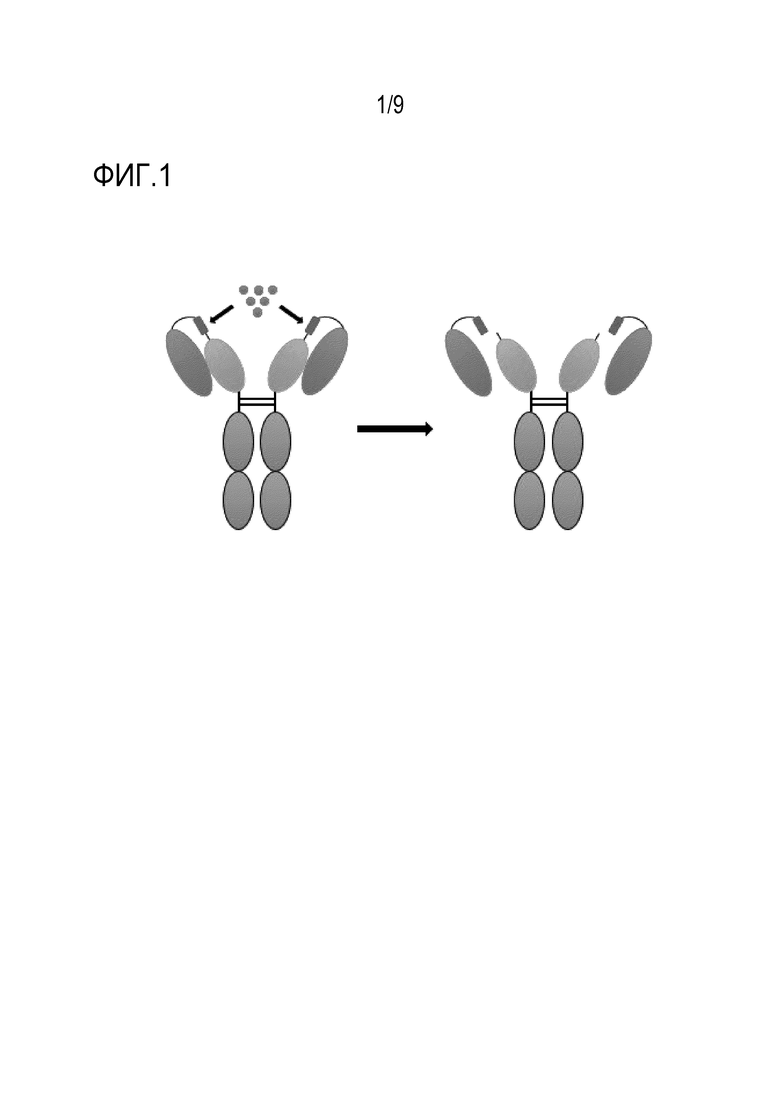
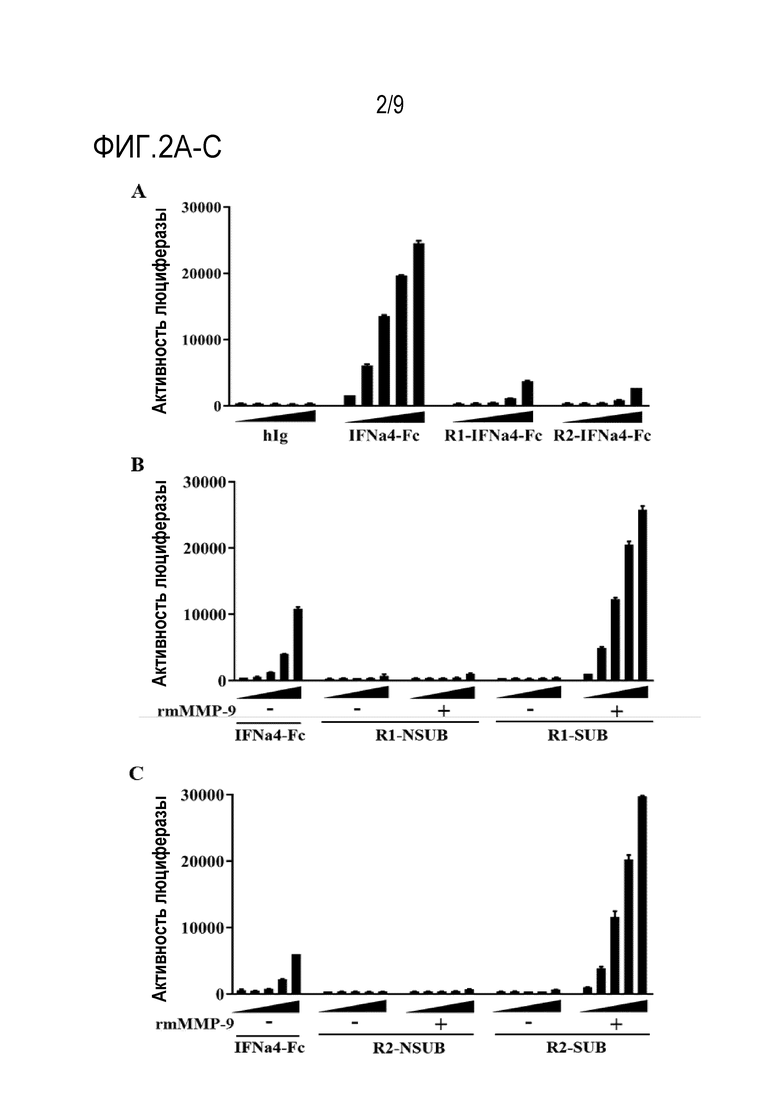

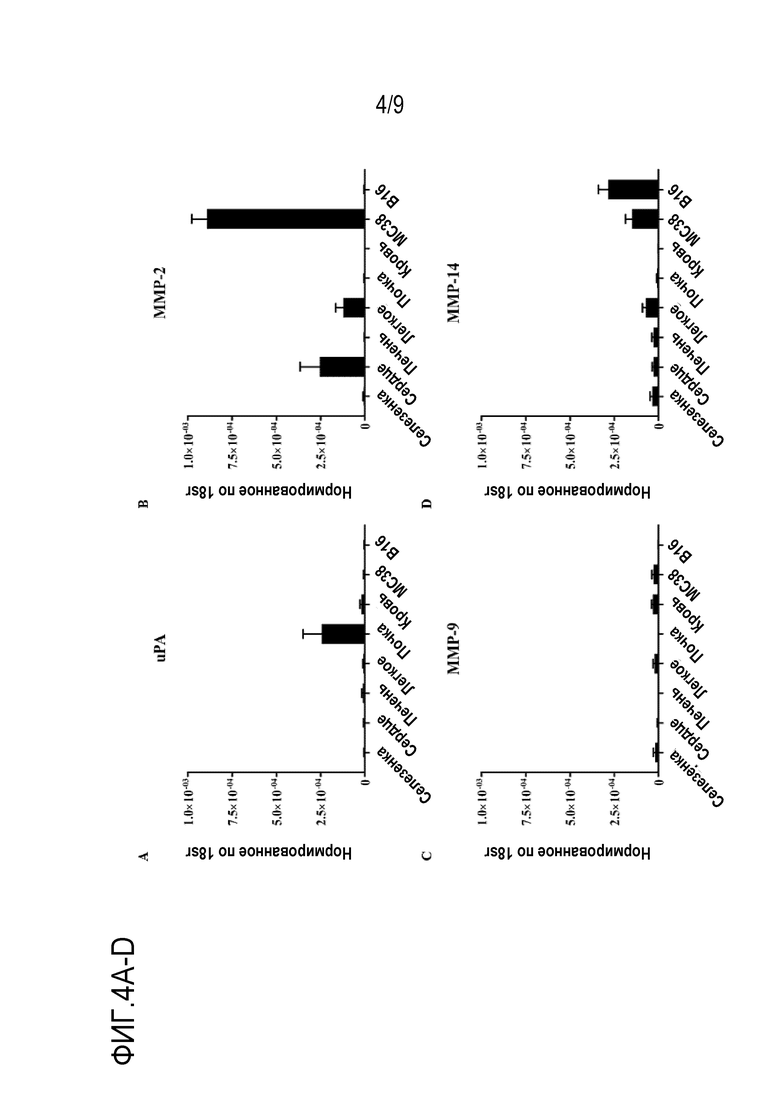
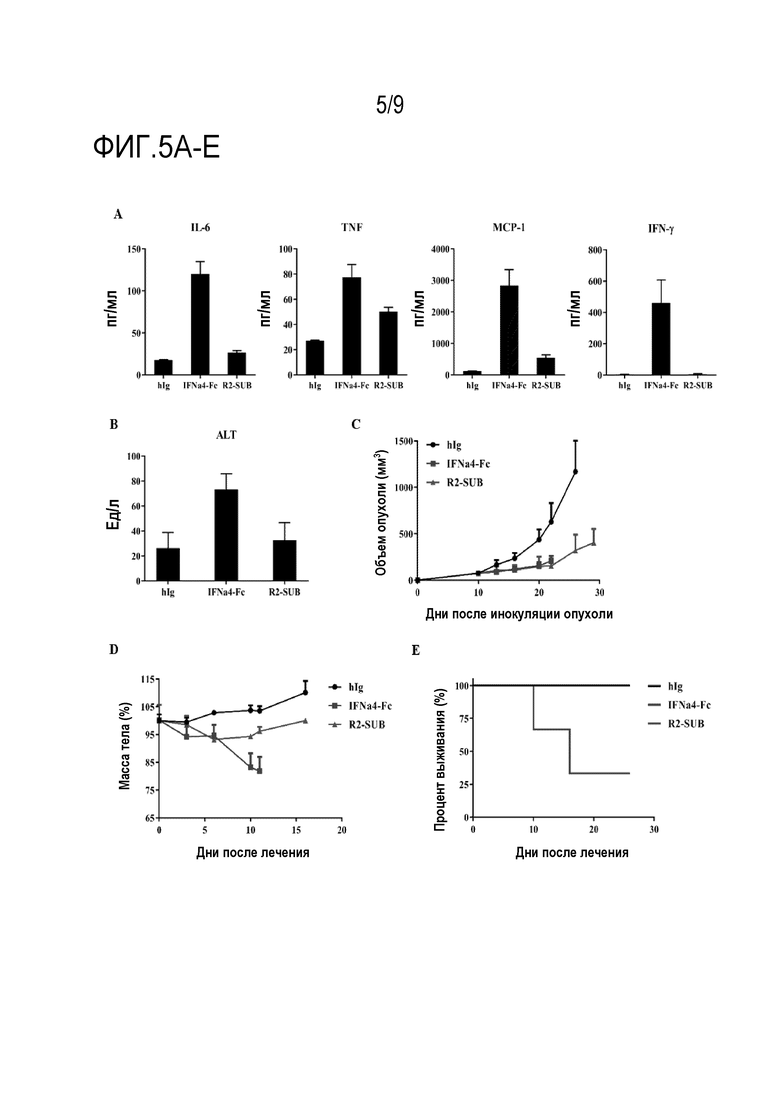
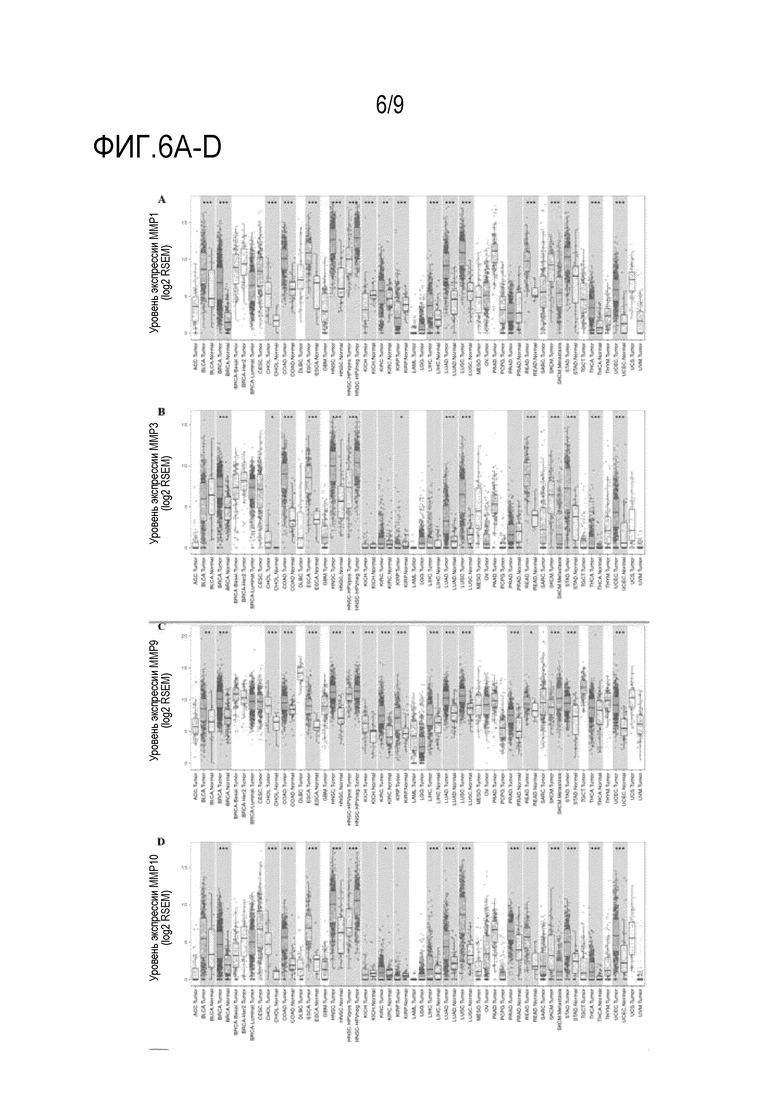
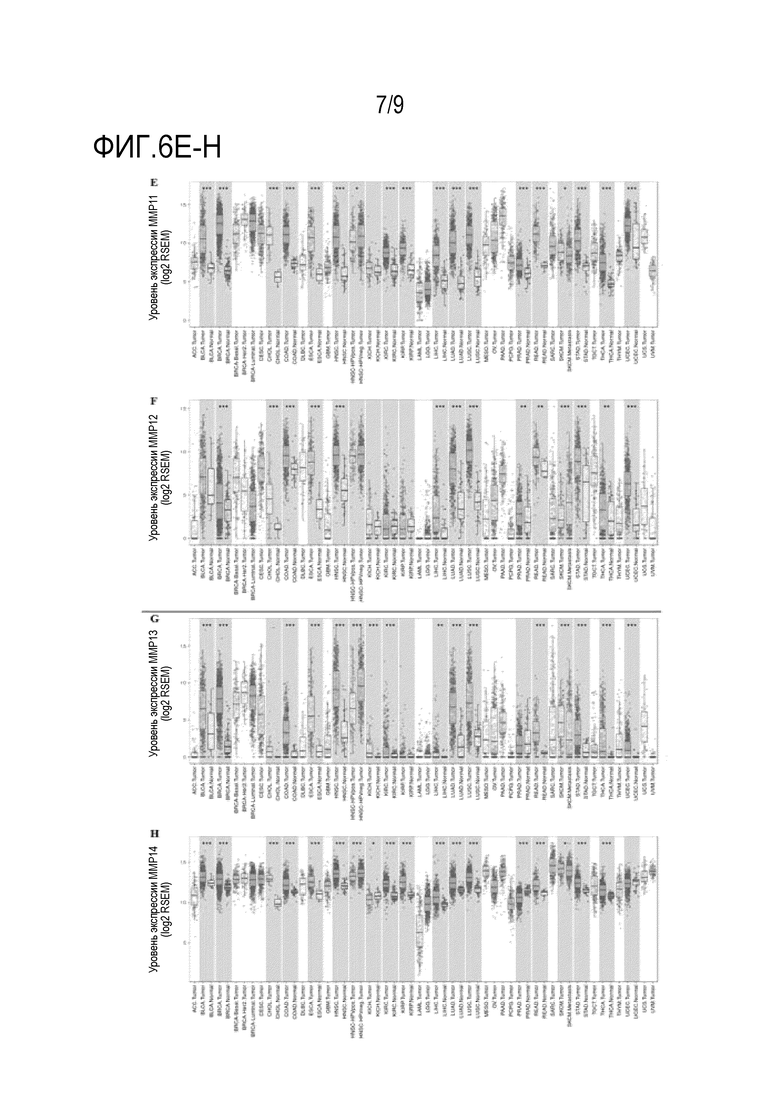
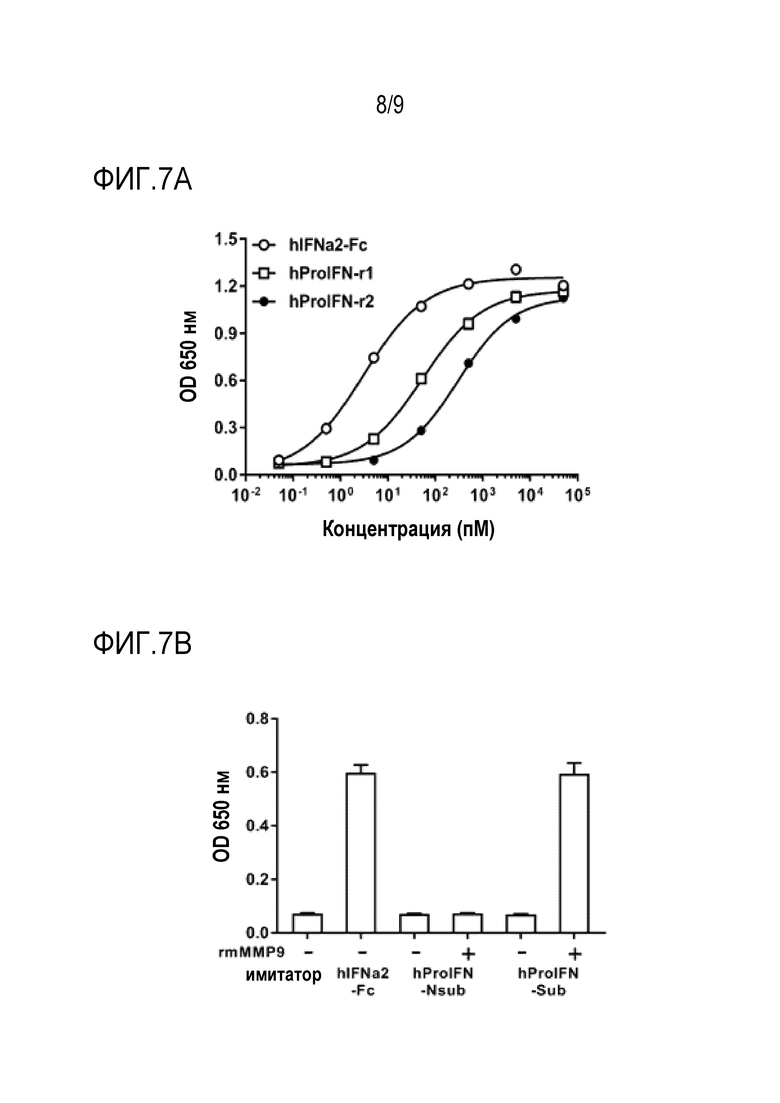
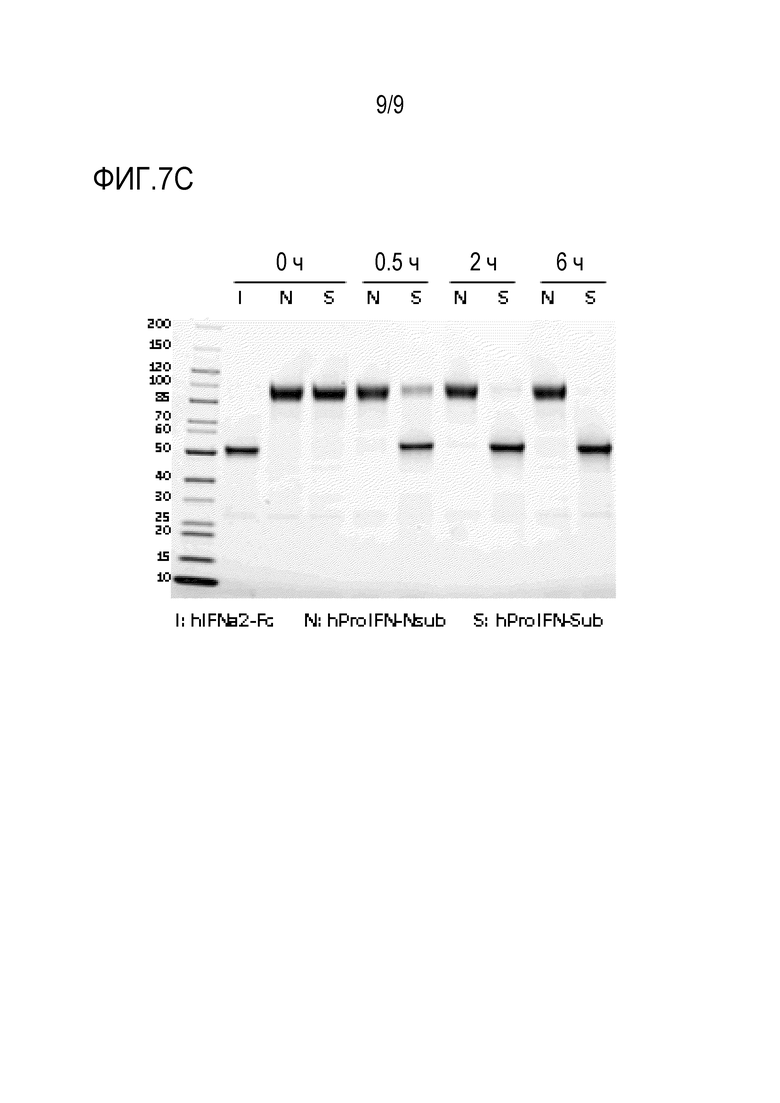
Группа изобретений относится к области фармацевтики и медицины. 1 объект представляет собой пролекарство интерферона для лечения рака, содержащее: (а) домен рецептора 2 интерферона альфа и бета (IFNAR2), который сохраняет IFN-связывающую активность; (b) домен интерферона 1 типа (IFN), который сохраняет активность интерферона 1 типа, когда он не связан указанным доменом IFNAR2; (c) Fc-домен иммуноглобулина (Ig), (d) первый линкер, слитый на одном конце с N-концом указанного IFN и слитый на другом конце с указанным IFNAR2, причем указанный первый линкер представляет собой G4S-PVGLIG-G4S; и (e) второй линкер, слитый на одном конце с C-концом указанного IFN и слитый на другом конце с N-концом указанного Fc-домена Ig. 2-4 объекты – конструкцию нуклеиновой кислоты, кодирующую пролекарство интерферона, рекомбинантную клетку, содержащую конструкцию нуклеиновой кислоты, и способ экспрессии пролекарства интерферона. 5-7 объекты – применение пролекарства интерферона для лечения рака и в получении лекарственного средства, способ лечения рака, включающий введение пациенту пролекарства интерферона. Технический результат заключается в эффективности и уменьшенных побочных эффектах IFN-пролекарства на основе IFNAR2. 7 н. и 19 з.п. ф-лы, 7 ил., 1 табл., 1 пр.
1. Пролекарство интерферона для лечения рака, содержащее:
(а) домен рецептора 2 интерферона альфа и бета (IFNAR2), который сохраняет IFN-связывающую активность;
(b) домен интерферона 1 типа (IFN), который сохраняет активность интерферона 1 типа, когда он не связан указанным доменом IFNAR2;
(c) Fc-домен иммуноглобулина (Ig),
(d) первый линкер, слитый на одном конце с N-концом указанного IFN и слитый на другом конце с указанным IFNAR2, причем указанный первый линкер является расщепляемым протеазой, указанный первый линкер является расщепляемым матриксной металлопротеиназой, указанная матриксная металлопротеиназа представляет собой MMP2 или MMP9, указанный первый линкер представляет собой G4S-PVGLIG-G4S; и
(e) второй линкер, слитый на одном конце с C-концом указанного IFN и слитый на другом конце с N-концом указанного Fc-домена Ig.
2. Пролекарство интерферона по п. 1, в котором указанный Ig представляет собой IgG, такой как IgG1 или IgG2.
3. Пролекарство интерферона по пп. 1, 2, в котором указанное пролекарство интерферона содержит две копии указанного домена IFN 1 типа.
4. Пролекарство интерферона по пп. 1-3, в котором указанное пролекарство интерферона содержит более двух копий указанного домена IFN 1 типа.
5. Пролекарство интерферона по пп. 1-4, в котором указанный IFN представляет собой IFN-α, IFN-β, IFN-κ, IFN-δ, IFN-ε, IFN-τ, IFN-ω или IFN-ζ.
6. Конструкция нуклеиновой кислоты, кодирующая пролекарство интерферона, содержащее:
(а) домен рецептора 2 интерферона альфа и бета (IFNAR2), который сохраняет IFN-связывающую активность;
(b) домен интерферона 1 типа (IFN), который сохраняет активность интерферона 1 типа, когда он не связан указанным доменом IFNAR2;
(c) Fc-домен иммуноглобулина (Ig),
(d) первый линкер, слитый на одном конце с N-концом указанного IFN и слитый на другом конце с указанным IFNAR2, причем указанный первый линкер является расщепляемым протеазой, указанный первый линкер является расщепляемым матриксной металлопротеиназой, указанная матриксная металлопротеиназа представляет собой MMP2 или MMP9, указанный первый линкер представляет собой G4S-PVGLIG-G4S;
(e) второй линкер, слитый на одном конце с C-концом указанного IFN и слитый на другом конце с N-концом указанного Fc-домена Ig; и
причем конструкция содержит промотор, расположенный 5'-концом к 5'-концу указанного домена IFN.
7. Конструкция нуклеиновой кислоты по п. 6, в которой указанный Ig представляет собой IgG, такой как IgG1 или IgG2.
8. Конструкция нуклеиновой кислоты по пп. 6, 7, в которой указанное пролекарство интерферона содержит две копии указанного домена IFN 1 типа.
9. Конструкция нуклеиновой кислоты по пп. 6-8, в которой указанное пролекарство интерферона содержит более двух копий указанного домена IFN 1 типа.
10. Конструкция нуклеиновой кислоты по пп. 6-9, в которой указанный IFN представляет собой IFN-α, IFN-β, IFN-κ, IFN-δ, IFN-ε, IFN-τ, IFN-ω или IFN-ζ.
11. Рекомбинантная клетка, содержащая конструкцию нуклеиновой кислоты по пп. 6-10, причем клетка предназначена для экспрессии пролекарства интерферона по пп. 1-5.
12. Способ экспрессии пролекарства интерферона, включающий культивирование клетки по п. 11.
13. Применение пролекарства интерферона по пп. 1-5 в получении лекарственного средства для лечения рака.
14. Применение пролекарства интерферона по пп. 1-5 для лечения рака.
15. Способ лечения рака, включающий введение пациенту пролекарства интерферона по пп. 1-5.
16. Способ по п. 15, дополнительно включающий стадию оценки экспрессии протеазы в раковой клетке, полученной от указанного пациента.
17. Способ по п. 16, в котором указанная раковая клетка получена из биопсии.
18. Способ по п. 16, в котором указанная раковая клетка представляет собой циркулирующую опухолевую клетку.
19. Способ по пп. 16-18, в котором указанный рак представляет собой рак легкого, рак молочной железы, рак головного мозга, рак ротовой полости, рак пищевода, рак головы и шеи, рак кожи, рак желудка, рак печени, рак поджелудочной железы, рак почки, рак яичника, рак предстательной железы, рак мочевого пузыря, рак толстой кишки, рак яичка, рак матки, рак шейки матки, лимфому или лейкоз.
20. Способ по пп. 15-19, в котором указанный рак является первичным, рецидивирующим, метастатическим или устойчивым ко многим лекарственным веществам.
21. Способ по пп. 15-20, в котором указанный пациент представляет собой человека или млекопитающее, не являющееся человеком.
22. Способ по пп. 15-21, дополнительно включающий введение указанного пролекарства интерферона более одного раза.
23. Способ по п. 22, в котором указанное пролекарство интерферона вводят ежедневно, через день, еженедельно, через неделю или ежемесячно.
24. Способ по пп. 15-23, в котором указанное пролекарство интерферона вводят системно.
25. Способ по пп. 15-23, в котором указанное пролекарство интерферона вводят внутрь опухоли, локально по отношению к опухоли или регионарно по отношению к опухоли.
26. Способ по пп. 15-25, в котором лечение включает одно или более из: замедления роста опухоли, остановки роста опухоли, уменьшения размера или объема опухоли, увеличения выживаемости по сравнению с пациентом, не получавшим лечение, индукции ремиссии рака, индукции апоптоза опухолевых клеток или индукции некроза опухоли.
| WO 2010096838 A2, 26.08.2010 | |||
| НАСОС ДЛЯ ПОДАЧИ ГОРЮЧЕЙ ЖИДКОСТИ В ДВИГАТЕЛЯХ ВНУТРЕННЕГО ГОРЕНИЯ | 1925 |
|
SU3635A1 |
| US 9623115 B2, 18.04.2017 | |||
| US 7906118 B2, 15.03.2011 | |||
| US 2016289324 A1, 06.10.2016 | |||
| WO 2015013671 A1, 29.01.2015 | |||
| BOSSMANN S | |||
| H | |||
| et al | |||
| Point-of-care routine rapid screening: the future of cancer diagnosis? // Expert review of molecular diagnostics | |||
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| Насос | 1917 |
|
SU13A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Счетный сектор | 1919 |
|
SU107A1 |

