ОБЛАСТЬ ТЕХНИКИ
Изобретение относится к области молекулярной биологии и биотехнологии, а именно, к препаративной биохимии РНК.
УРОВЕНЬ ТЕХНИКИ
Известно большое разнообразие ферментов нуклеаз, которые осуществляют деградацию нуклеиновых кислот - ДНК и РНК. Известны примеры контролируемой деградации ДНК. В частности, существуют эндонуклеазы рестрикции, которые расщепляют двуспиральную ДНК в участках (сайтах рестрикции) с определенной последовательностью нуклеотидов [1]. С помощью эндонуклеаз рестрикции можно получать индивидуальные фрагменты ДНК, расщепляя более длинную ДНК по сайтам рестрикции, а затем разделяя фрагменты ДНК по размеру, например, с помощью электрофореза в агарозе или в полиакриламидном геле. Получение индивидуальных фрагментов нуклеиновых кислот с заданной первичной структурой необходимо для решения многих научных и прикладных задач.
В отличие от ДНК, для РНК подобные эндонуклеазы не известны. Известен ряд белковых сайт-специфических эндорибонуклеаз, например альфа-сарцин, расщепляющий межнуклеотидную связь в РНК большой субчастицы рибосом [2], или эндонуклеазы, участвующие в процессинге тРНК эукариот и архей [3]. Однако такие рибонуклеазы расщепляют лишь определенные межнуклеотидные связи в определенных видах РНК и не могут быть использованы для расщепления любой заданной РНК. Аналогичной узкой специфичностью обладают также эндонуклеазы, построенные из молекул РНК (рибозимы) [4]. Можно искусственно (путем конструирования или селекции) получить рибозим (или ДНКзим, построенный из ДНК), расщепляющий заданную последовательность в заданном месте (см., например, [5]), но при этом для другого места той же РНК или для другой РНК необходимо получать новый рибозим.
В технике известны так называемые искусственные эндорибонуклеазы, представляющие собой олигонуклеотиды, несущие химические группы, способные расщеплять межнуклеотидные связи РНК [6-10]. Их сайт-специфичность определяется специфичностью гибридизации олигонуклеотидов: молекула РНК расщепляется в том месте, где связался олигонуклеотид. Синтез таких олигонуклеотидов значительно дешевле и проще, чем получение искусственных рибозимов. Однако, в отличие от белковых сайт-специфических эндорибонуклеаз и рибозимов, искусственные рибонуклеазы обладают относительно низкой каталитической активностью и недостаточной точностью расщепления РНК: как правило, расщепляется не одна межнуклеотидная связь, а ряд связей, находящихся вблизи реактивной группы.
Еще один известный способ сайт-специфического расщепления РНК включает использование РНКазы Н, которая способна расщеплять участок РНК, гибридизованный с олигодезоксирибонуклеотидом [11, 12]. Однако здесь, как и в случае искусственных рибонуклеаз, трудно добиться расщепления РНК в точно заданном месте.
Для получения заданных фрагментов РНК часто используют транскрипцию ДНК, содержащую промотор ДНК-зависимой РНК-полимеразы (например, РНК полимеразы фага Т7) и фрагментированную с помощью эндонуклеазы рестрикции [13]. Получаемый РНК-транскрипт с 5'-конца ограничен промотором, а с 3'-конца - сайтом рестрикции. Этот метод позволяет получать большие количества индивидуальной РНК, но его ограничением является необходимость наличия в ДНК подходящего сайта рестрикции.
Были попытки осуществлять контролируемую деградацию РНК с помощью экзонуклеаз, которые, в отличие от эндонуклеаз, разрушают РНК, последовательно отщепляя нуклеотиды с 5'- или 3'-конца. Было показано, что 5'→3' экзонуклеаза (отщепляющая нуклеотиды с 5'-конца) из ядер клеток HeLa разрушает РНК, лишенную «кэпа» на 5'-конце (кэп - 7-метилгуанозин, соединенный с 5'-концевым нуклеотидом эукариотических мРНК через 5'-трифосфат-5'-мостик), но не разрушает кэпированную РНК [14]. Это позволяет, в частности, селективно разрушать некэпированые РНК и обогащать препараты кэпированных РНК. Также было показано, что 3'→5' экзорибонуклеаза полинуклеотндфосфорилаза (ПНФаза) из клеток Escherichia coli разрушает поли(А)-последовательность на 3'-конце эукариотических мРНК, но не разрушает остальную часть мРНК, если реакцию осуществляют при 0°С в присутствии 1 M NaCl, когда вся РНК, за исключением поли(А)-последовательности, сохраняет вторичную структуру [15].
В то же время до сих пор экзонуклеазы не удавалось применить для контролируемой деградации большинства РНК. Для ПНФазы Е. coli было показано, что, расщепляя РНК с 3'-конца, этот фермент останавливается на расстоянии 6-9 нуклеотидов от стабильного черешка шпильки - элемента вторичной структуры РНК [16]. В принципе, это могло бы быть использовано для получения фрагментов РНК, оканчивающихся такими шпильками, но остановка фермента никогда не бывает полной. Кроме того, скорость, глубина, полнота деградации и однородность получаемых фрагментов зависит от стабильности шпилек и широко варьирует среди разных видов РНК [17]. Повышение температуры среды приводит к дестабилизации структуры РНК и, как следствие, повышает скорость деградации и однородность продуктов [18]. Однако ПНФаза Е. coli не стабильна при температуре выше 55°С и быстро и необратимо инактивируется при 65°С [17].
Последнюю проблему можно было бы попытаться решить, используя экзонуклеазы из термофильных организмов, живущих при высокой температуре. Было опубликовано несколько работ по исследованию свойств термофильных ПНФаз. В одной из них сообщалось, что ПНФазы из термофильных бактерий Bacillus stearothermophilus и Thermus aquaticus способны полимеризовать нуклеозид-5'-дифосфаты (то есть, катализировать реакцию, обратную фосфоролизу РНК) при 69°С и 80°С соответственно [19]. Однако способность этих ферментов осуществлять деградацию РНК не была исследована. Более того, очищенная ПНФаза из В. stearothermophilus состояла из субъединиц с необычно малой молекулярной массой - 51 кДа, тогда как все другие известные ПНФазы из разных организмов, от бактерий до человека, имеют молекулярную массу в диапазоне от 75 до 100 кДа [20-25]. ПНФаза же из Т. aquaticus не была очищена вовсе. В другой статье, Tth-ПНФаза (ПНФаза из Thermus thermophilus - термофильной бактерии, близкородственной Т. aquaticus) была очищена до гомогенного состояния, судя по тестам, осуществленным в неденатурирующих условиях. Однако очищенный фермент был полностью лишен фосфорилазной активности (то есть, не мог разрушать РНК), вероятно, в результате его протеолитической деградации в процессе выделения. Действительно, электрофорез в присутствии SDS (додецилсульфата натрия) выявил в препарате очищенной Tth-ПНФазы три вида полипептидов с кажущейся молекулярной массой 92, 73 и 35 кДа [26].
Таким образом, не было выделено термофильных ПНФаз, обладающих способностью к деградации РНК. Кроме того, не было найдено способов избирательной защиты заданной РНК или ее фрагмента от деградации ПНФазами при повышенной температуре. Наконец, повышение температуры существенно ускоряет эндонуклеотический гидролиз РНК, катализируемый ионами Mg2+ [27], которые необходимы для работы ПНФаз в относительно высокой концентрации [17]. Например, в случае ПНФазы из Micrococcus luteus кажущееся значение константы Михаэлиса (КМ) для Mg2+ равно 0,8 мМ [28]. Это может дополнительно увеличивать гетерогенность получаемых фрагментов РНК.
Наиболее близким аналогом (прототипом) данного изобретения является вышеупомянутый способ удаления поли(А)-последовательности с 3'-конца эукариотических мРНК с использованием ПНФазы Е.coli [15] - пример контролируемой деградации РНК с помощью ПНФазы.
Из вышеизложенного следует, что существует потребность в усовершенствованных способах контролируемой деградации РНК. Такой способ раскрывается данным изобретением.
Настоящее изобретение направлено на решение задачи получения индивидуальных РНК посредством контролируемой деградации РНК термофильными ПНФазами.
Другой задачей изобретения является препарат термостабильной полинуклеотидфосфорилазы, способной осуществлять фосфоролиз структурированных полирибонуклеотидов при температуре выше 55°С.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Указанные технические задачи реализуются разработкой нового способа деградации структурированных полирибонуклеотидов на основе препарата ПНФазы, практически свободного от эндонуклеазной и гидролазной активностей.
Одним из аспектов изобретения является способ деградации структурированных полирибонуклеотидов, который включает инкубацию образца, содержащего, по меньшей мере, один вид полирибонуклеотидов, в растворе, содержащем полинуклеотидфосфорилазу (ПНФазу), ее косубстрат и кофактор, в условиях, при которых данная ПНФаза обладает фосфоролитической активностью, причем в качестве ПНФазы используют термостабильную ПНФазу, а инкубацию осуществляют при таких значениях температуры, времени инкубации, концентрации косубстрата и кофактора, которые в отсутствие ПНФазы обеспечивают существенно полную сохранность, а при достаточно высокой концентрации ПНФазы - существенно полную деградацию хотя бы одного из полирибонуклеотидов образца в отсутствие каких-либо способов защиты, повышающих устойчивость данного полирибонуклеотида к данной ПНФазе по сравнению с немодифицированным полирибонуклеотидом, обладающим такой же первичной структурой.
Другой аспект изобретения состоит в том, что, по меньшей мере, один из полирибонуклеотидов защищают от действия экзорибонуклеазы путем химической модификации 3'-концевого нуклеотида, а в качестве модифицирующей группы используют фосфат или биотин.
Следующий аспект изобретения состоит в том, что полирибонуклеотид защищают путем гибридизации с олигонуклеотидом или аналогом олигонуклеотида, способным образовать с данным полирибонуклеотидом гибрид, имеющий существенную стабильность при температуре инкубации, причем используют олигонуклеотид или аналог олигонуклеотида, комплементарный 3'-концевому участку полирибонуклеотида, или используют олигонуклеотид или аналог олигонуклеотида, комплементарный внутреннему участку полирибонуклеотида.
Следующий аспект изобретения состоит в том, что в качестве 3'→5' экзонуклеазы используют полинуклеотидфосфорилазу Thermus thermophilus.
КРАТКОЕ ОПИСАНИЕ ФИГУР
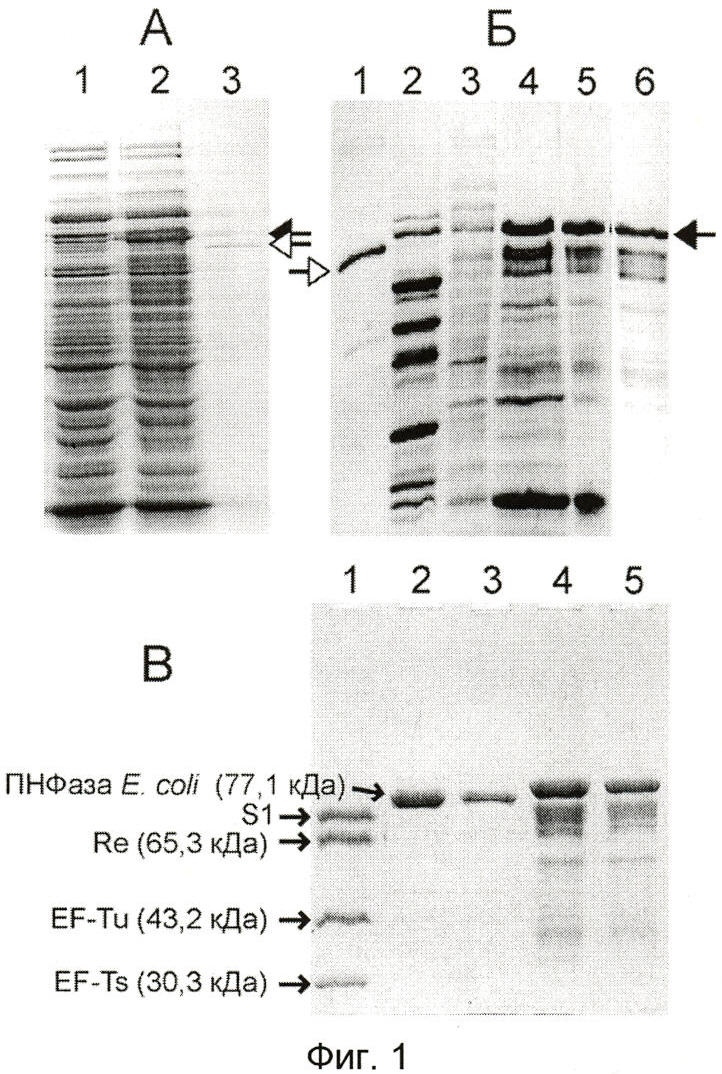
Фиг.1 демонстрирует экспрессию гена Tth-ПНФазы в клетках Е. coli и выделение продукта экспрессии.
Фиг.2 демонстрирует зависимость фосфоролиза РНК Tth-ПНФазой от концентрации ионов Mg2+.
Фиг.3 демонстрирует зависимость фосфоролиза РНК Tth-ПНФазой от температуры.
Фиг.4 демонстрирует взаимодействие Tth-ПНФазы и РНК.
Фиг.5 демонстрирует селективность и полноту деградации РНК Tth-ПНФазой.
Фиг.6 демонстрирует эффект 3'-концевых модификаций РНК на ее деградацию Tth-ПНФазой.
Фиг.7 демонстрирует сравнение свойств ПНФаз Т. thermophilus и Е. coli.
Фиг.8 демонстрирует неспособность ПНФазы Е. coli полностью разрушить 3'-фрагмент RQ135 РНК.
Фиг.9 демонстрирует эффект олигонуклеотида, гибридизованного к внутреннему участку РНК, на ее деградацию Tth-ПНФазой.
Фиг.10 демонстрирует последовательность, защищаемую от Tth-ПНФазы олигонуклеотидом, гибридизованным к внутреннему участку РНК.
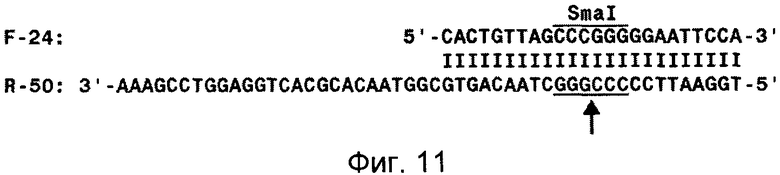
Фиг.11 демонстрирует схему гибрида двух олигонуклеотидов, расщеплением которого по сайту рестрикции SmaI получали олигонуклеотид с точным 5'-концом.
ОПИСАНИЕ ТЕРМИНОВ
Термин «полинуклеотидфосфорилаза» («ПНФаза») используется здесь для обозначения 3'→5' экзорибонуклеазы, способной катализировать реакцию фосфоролиза полирибонуклеотида, отщепляя нуклеотиды от его 3'-конца, независимо от того, способен ли этот фермент также катализировать реакцию фосфоролиза поли-2'-дезоксирибонуклеотида или иного типа нуклеотидной последовательности.
Термин «структурированный полирибонуклеотид» используется здесь для обозначения полирибонуклеотида, в том числе синтетического олигорибонуклеотида и природной РНК, способного формировать стабильную в физиологических условиях внутримолекулярную вторичную и третичную структуру, в том числе за счет спаривания нуклеотидов по Уотсону и Крику.
ОСУЩЕСТВЛЕНИЕ ИЗОБРЕТЕНИЯ
Полинуклеотидфосфорилаза (ПНФаза) - это 3'→5' экзорибонуклеаза, осуществляющая обратимый фосфоролиз полирибонуклеотида (pN)n с 3'-конца в соответствии с уравнением: (pN)n+Pi↔(pN)n-1+ppN, где Pi - неорганический ортофосфат (косубстрат фосфоролиза), a ppN - нуклеозид-5'-дифосфат [29]. Для функционирования ПНФазы необходимы кофакторы - ионы Mg2+, которые могут быть заменены другими двухвалентными катионами, такими как Со2+, Ni2+, Cd2+, Cu2+ и Zn2+, но не Са2+ [17]. Вместо Pi в качестве косубстрата ПНФаза может использовать арсенат. В этом случае реакция становится практически необратимой, так как образующиеся нуклеозид-5'-фосфориларсенаты спонтанно гидролизуются до нуклеозидмонофосфатов и арсената [17].
ПНФаза была обнаружена во всех типах бактерий [17], за исключением микоплазмы [20]. Хотя последовательностей, кодирующих ПНФазу, нет в геномах архей и дрожжей, изученных к настоящему времени (эти геномы, тем не менее, содержат гены похожего фермента - РНКазы РН), они присутствуют в высших эукариотах [21-23]. ПНФазы растений и животных кодируются ядерными генами, но локализуются, главным образом, в органеллах [30]. В растениях есть два вида ПНФазы, один из которых локализован в хлоропластах, а другой - в митохондриях [21, 31].
Ген ПНФазы (pnp) кодирует полипептид массой 75-100 кДа, состоящий из пяти эволюционно консервативных доменов: двух N-концевых «коревых» доменов, гомологичных РНКазе РН Е.coli, разделенных α-спиральным доменом, и двух С-концевых РНК-связывающих доменов - KH и S1 [20-25]. РНКаза РН, как и ПНФаза, представляет собой фосфоролитическую 3'→5' экзорибонуклеазу. В клетках Е.coli РНКаза РН ответственна за процессинг 3'-концов молекул предшественников тРНК [32]. Домен KH («K-homology») был исходно идентифицирован в РНК-связывающем белке К человека [33], а домен S1 назван так из-за гомологии с РНК-связывающим белком S1 рибосом Е.coli [34]. Молекулы ПНФаз бактерий Е.coli и Streptomyces antibioticus состоят из трех идентичных субъединиц, которые образуют структуру в виде бублика с отверстием в центре, где может помещаться цепь РНК [25, 35], а ПНФаза хлоропластов состоит из двух таких бубликов, положенных один на другой [21, 36].
В дрожжах и археях функциональными и структурными аналогами ПНФазы являются экзосомы. Экзосомы состоят из субъединиц, являющихся гомологами доменов ПНФазы, и выполняют такие же функции, катализируя обратимый фосфоролиз полирибонуклеотидов. Поэтому экзосома может быть рассмотрена как разновидность ПНФазы. В пользу этого говорит также то, что трехмерные структуры бактериальной ПНФазы [25] и РНКазы РН, составляющей «кор» (основу) архейной экзосомы [37], высокогомологичны [38].
Настоящее изобретение основано на результатах работы по клонированию гена Tth-ПНФазы, его экспрессии в клетках Е.coli и изучению биохимических свойств выделенного фермента [39]. В процессе исследования биохимических свойств выделенного фермента неожиданно было установлено, что выделенная Tth-ПНФаза явилась первой очищенной термостабильной ПНФазой, способной к осуществлению реакции фосфоролиза полирибонуклеотидов. Оказалось, что при оптимальной температуре фосфоролиза (65°С) Tth-ПНФаза способна полностью разрушить РНК даже с очень стабильной вторичной структурой.
В рамках настоящего изобретения было установлено, что РНК можно защитить от фосфоролиза Tth-ПНФазой путем модификации ее 3'-конца либо путем гибридизации ее 3'-концевого участка с комплементарным олигонуклеотидом. Если смесь защищенной и незащищенной РНК обработать Tth-ПНФазой, то разрушается только незащищенная РНК.
Далее было установлено, что олигонуклеотид, гибридизованный с внутренним участком РНК, защищает от Tth-ПНФазы этот участок; часть РНК по 5'-сторону от этого участка, а также последовательность длиной 8 нт (нуклеотидов) по 3'-сторону от этого участка. Важным обнаруженным свойством Tth-ПНФазы является ее способность к эффективному фосфоролизу в присутствии очень низкой концентрации ионов Mg2+. Это позволяет минимизировать катализируемый ионами Mg2+ неэнзиматический гидролиз РНК при осуществлении высокотемпературного фосфоролиза. Обнаруженные свойства Tth-ПНФазы позволяют впервые осуществить полностью контролируемую деградацию практически любой произвольно взятой РНК или смесей РНК. Есть основания полагать, что аналогичными свойствами могут обладать другие термофильные фосфоролитические экзорибонуклеазы, например ПНФаза Т. aquaticus и экзосомы архей.
Ген Tth-ПНФазы клонируют, используя в качестве исходного генетического материала хромосомную ДНК Т. thermophilus, как описано в примере 1, а также используя информацию о первичной структуре участка ДНК, предположительно содержащего этот ген, опубликованную в базе данных ГенБанк в 1997 под номером Z84207. Обнаружено, что данный участок ДНК содержит открытую рамку считывания, способную кодировать полипептид массой 78,2 кДа (что близко к молекулярной массе ПНФазы Е.coli - 77,1 кДа [40]), в котором присутствуют все структурные домены, присущие типичным ПНФазам. Кодируемая аминокислотная последовательность и анализ этой открытой рамки считывания приведены в белковой базе данных NCBI под номером САВ06341.
Ген Tth-ПНФазы размножают с помощью полимеразной цепной реакции (ПЦР), клонируют его в составе плазмиды под контролем Т7-промотора и экспрессируют в клетках Е.coli, продуцирующих РНК-полимеразу фага Т7. Продукт экспрессии мигрировал при денатурирующем электрофорезе в присутствии SDS несколько медленнее, чем собственная ПНФаза Е.coli. Это позволило установить, что конечный препарат Tth-ПНФазы, выделенный из клеток Е.coli, не содержал субъединиц собственной ПНФазы хозяина (фиг.1). Процедура выделения Tth-ПНФазы включает: прогревание клеточного лизата при 70°С, что приводит к денатурации и осаждению большинства клеточных белков, включая ПНФазу Е.coli; ионно-обменную хроматографию на колонке DEAE-сефарозы; инкубацию в присутствии Pi и Mg2+ («автолиз», при котором примесные эндогенные РНК деградируют под действием ПНФазы; включение в смесь фосфатазы с целью удаления с молекул РНК 3'-фосфата, который может препятствовать фосфоролизу [41]); а также проведение гель-фильтрации через колонку суперозы-6.
Полноразмерный полипептид Tth-ПНФазы с массой около 78 кДа составляет >50% белка конечного препарата, а остальная часть белка представлена двумя группами многочисленных более коротких полипептидов с молекулярной массой в диапазонах 60-70 и 30-40 кДа. Эти полипептиды со-очищаются с Tth-ПНФазой, и похожие полипептиды накапливаются в клетках Е.coli, трансформированных плазмидой, несущей ген Tth-ПНФазы, в результате индукции синтеза Т7 РНК-полимеразы (фиг.1). Вероятно, они представляют собой фрагменты полипептида с массой 78 кДа, который весьма подвержен протеолизу [26]. Поскольку, как будет видно ниже, препарат фермента не содержал примесных активностей, не предпринималось дальнейших попыток его очистки. Выход фермента (по содержанию полипептида с массой 78 кДа) составил 3,3 мг из 9 г клеток Е.coli, продуцирующих Tth-ПНФазу. АДФ-полимеризующая активность конечного препарата была ≈200 ед/мг, что выше, чем активность ПНФазы, выделенной из клеток Т. thermophilus (≈70 ед/мг [26]), и, в отличие от того препарата, выделенная Tth-ПНФаза обладала фосфоролитической активностью (см. ниже).
Очищенная Tth-ПНФаза разрушала незащищенную РНК, но не разрушала РНК, 3'-конец которой гибридизован с комплементарным олигодезоксирибонуклеотидом (фиг.2). Отсюда следует, что полученный препарат действительно обладает 3'→5'-экзонуклеазной активностью и свободен от эндорибонуклеаз. Более того, РНК оставалась целой, если ее инкубировали с ферментом в отсутствие Pi (см., например, фиг.4Б). Следовательно, имеет место именно фосфоролиз, а не гидролиз РНК.
Хотя РНК оставалась целой в присутствии 56 мкМ EDTA (этилендиаминтетраацетата), который прочно связывает следы двухвалентных катионов, полную деградацию незащищенной РНК наблюдали при добавлении эквимолярной концентрации Mg2+ (фиг.2). Принимая во внимание величину константы диссоциации (Kd) комплекса Mg2+: EDTA (≈10-9 M [42]), можно сделать вывод, что потребность Tth-ПНФазы в свободных ионах Mg2+ очень низка, ≤1 мкМ. Это значительно ниже, чем потребность в Mg2+ мезофильной ПНФазы [28]. В последующих экспериментах концентрацию ионов Mg2+ поддерживали на уровне 40 мкМ. Это позволяло осуществлять ферментативный фосфоролиз РНК при высокой температуре, избегая Mg2+-катализируемого гидролиза. В этих условиях, оптимальная температура фосфоролиза составляет 65°С, тогда как в отсутствие ПНФазы РНК остается практически целой даже при 85°С (фиг.3). Фиг.3 также демонстрирует, что рабочий диапазон температур, в котором Tth-ПНФаза проявляет существенную фосфорилазную активность, составляет 60-75°С. Это выше, чем максимальная температура, при которой проявляют фосфорилазную активность мезофильные ПНФазы, такие как ПНФаза E.coli - 55°С [17].
Фиг.4А показывает, что при насыщающей концентрации Tth-ПНФазы, когда происходит одновременный фосфоролиз всех цепей РНК, 109-нт 3'-фрагмент высокоструктурированной RQ135 РНК [43, 44] разрушается практически полностью в пределах 2 мин. Таким образом, скорость реакции фосфоролиза составляет ≈50 нт/мин при 65°С. Это гораздо выше, чем скорость фосфоролиза примерно столь же высокоструктурированного тРНК-подобного 3'-концевого домена РНК вируса мозаики табака, осуществляемого ПНФазой Е.coli при оптимальной для нее температуре (37°С) - 3,5 нт/мин [17]. С другой стороны, в отличие от ПНФазы Е.coli, которая насыщает РНК при молярном соотношении 1:1, для достижения насыщения требуется более высокая концентрация Tth-ПНФазы. При концентрации РНК, использованной в примере 7 (50 nM), насыщение достигается при 4-кратной концентрации Tth-ПНФазы (Фиг.4A). Концентрацию Tth-ПНФазы рассчитывают исходя из допущения, что одна активная в фосфоролизе молекула состоит, как и в случае ПНФазы Е.coli, из трех полноразмерных (78 кДа) субъединиц [41]. Такое же соотношение получили, когда за образованием комплекса фермента и РНК в присутствии Mg2+ (но в отсутствие Pi для предотвращения фосфоролиза) следили по сдвигу 32Р-меченой полосы РНК при электрофорезе (фиг.4Б). Полученные результаты позволяют оценить значение Kd для взаимодействия между ПНФазой и РНК: ≈100 nM. Следовательно, сродство к РНК у Tth-ПНФазы приблизительно в 10 раз ниже, чем у ПНФазы Е.coli (Kd=10-20 nM [45]). Более низкое сродство не означает, что в реакции следует использовать больше Tth-ПНФазы. Вместо этого, полной деградации РНК можно достичь, либо используя более высокие концентрации реагентов в меньшем реакционном объеме, либо увеличивая время реакции.
С помощью ОТ-ПЦР (обратной транскрипции и последующей ПЦР) установлено, что в оптимальных условиях (температура, концентрации Mg2+ и EDTA, соотношение фермента к РНК, время реакции) Tth-ПНФаза разрушает более 99% высокоструктурированной РНК, не повреждая РНК, 3'-конец которой защищен комплементарным олигодезоксирибонуклеотидом (фиг.5А). Этот результат указывает на то, что олигонуклеотид, гибридизованный с 3'-концом РНК, мог бы селективно защищать ее от Tth-ПНФазы. Следующий эксперимент (фиг.5Б) демонстрирует, что это действительно так. В этом эксперименте смесь двух видов РНК - 5'- и 3'-фрагментов RQ135 РНК [43, 44], отожгли с олигодезоксирибонуклеотидом, комплементарным 3'-концу 3'-фрагмента. Последующий фосфоролиз с Tth-ПНФазой привел к полной деградации 5'-фрагмента и получению практически чистого 3'-фрагмента (если не считать сам олигонуклеотид, который можно удалить разными способами - например, с помощью хроматографии, электрофореза или обработки ДНКазой).
Как видно из фиг.6, 3'-концевая модификация РНК делает эту РНК более устойчивой к фосфоролизу Tth-ПНФазой, дополнительно подтверждая, что деградация РНК является результатом 3'→5'-экзонуклеазной активности. Окисление РНК перйодатом, в результате которого происходит раскрытие концевого рибозного кольца и образование диальдегида [46], приводит к селективной защите модифицированных молекул (фиг.6, А и Б). Эта защита является умеренной, но она усиливается после обработки РНК анилином (фиг.6В), которая удаляет окисленный нуклеозид и дает РНК, 3'-конец которой фосфорилирован [47]. Высокая устойчивость 3'-фосфорилированной РНК к Tth-ПНФазе не связана с химической модификацией полирибонуклеотидного остова в процессе обработки перйодатом или анилином, так как РНК становится вновь подверженной фосфоролизу после удаления 3'-фосфата фосфатазой (фиг.6В). Аналогичное увеличение защиты наблюдается в результате обработки окисленной РНК гидразидом биотина, которая приводит к добавлению биотиновой группы к 3'-концу РНК (фиг.6Г).
На фиг.7 приведено сравнение некоторых свойств Tth-ПНФазы и ПНФазы Е.coli, очищенной, как описано в примере 12. Хотя оба эти фермента эффективно разрушают неструктурированный полирибонуклеотид поли(А), они по-разному ведут себя в случае 3'-фрагмента RQ135 РНК, обладающего прочной вторичной структурой [43, 44]. В отличие от Tth-ПНФазы, ПНФаза Е.coli разрушает при оптимальной температуре лишь часть 3'-фрагмента, а другая его часть остается без видимых повреждений. Это напоминает деградацию ПНФазой Е.coli другой структурированной РНК - тРНК [18], которая существует в двух конформациях, лишь одна из которых подвержена фосфоролизу при 37°С [48]. Однако, в отличие от тРНК [18, 48], ПНФаза Е.coli не способна полностью разрушить 3'-фрагмент RQ135 РНК даже при повышении температуры до 55°С и концентрации неорганического фосфата до 10 мМ (фиг.8). Другое различие между этими двумя ферментами состоит в эффекте 3'-концевого фосфата. В то время как эта группа защищает РНК от Tth-ПНФазы, такой эффект не заметен в случае ПНФазы Е.coli, возможно потому, что он маскируется неспособностью этого фермента полностью разрушить незащищенную РНК (фиг.7 и 8).
На фиг.9 представлены результаты деградации трех РНК, имеющих одинаковую 5'-концевую часть, но в разной мере удлиненных на 3'-конце. Все три РНК подвергли фосфоролизу Tth-ПНФазой после отжига в присутствии или в отсутствие 38-нт олигодезоксирибонуклеотида, комплементарного 3'-концевой последовательности наиболее короткой РНК («SmaI») и внутренним последовательностям более длинных РНК («EcoRI» и «PvuII»). Видно, что олигонуклеотид полностью защищает самую короткую РНК и частично защищает более длинные РНК. Деградация последних дает устойчивый продукт, подвижность которого при неденатурирующем электрофорезе несколько меньше, чем подвижность гибрида олигонуклеотида с РНК SmaI (фиг.9, слева). Такой продукт не образуется в отсутствие олигонуклеотида (фиг.9, справа).
Тонкую структуру продуктов фосфоролиза анализировали с помощью электрофореза высокого разрешения в денатурирующих условиях (фиг.10). Перед фосфоролизом каждую из трех РНК дефосфорилировали и затем метили на 5'-конце с помощью [γ-32P] ATP и полинуклеотидкиназы. Из фиг.10А видно, что устойчивый продукт деградации более длинных РНК имеет довольно гетерогенный 3'-конец и до 8 нт длиннее, чем РНК с защищенной концевой последовательностью. Последняя также имеет гетерогенный 3'-конец, но это обычно для РНК, полученных с помощью транскрипции обрезанных ДНК («run-off transcription») [49]. Наблюдаемая гетерогенность продуктов фосфоролиза по большей части является следствием гетерогенности защитного олигонуклеотида, так как ее удается существенно уменьшить при использовании олигонуклеотида с точным 5'-концом. Такой защитный олигонуклеотид получали, обрезая с помощью эндонуклеазы рестрикции олигонуклеотид, удлиненный на 5'-конце, как описано в примере 15. Фосфоролиз EcoRI- и PvuII-PHK, гибридизованных с таким 5'-обрезанным олигонуклеотидом, давал продукт, чья 3'-концевая гетерогенность была сравнима с гетерогенностью продукта транскрипции, и который был на 8 нт длиннее, чем РНК SmaI с защищенной концевой последовательностью (фиг.10Б). Отсюда следует, что Tth-ПНФаза останавливается на разрушаемой РНК на расстоянии 8 нт от 5'-конца олигонуклеотида, гибридизованного с внутренней последовательностью РНК.
Это согласуется с наблюдениями, что ПНФаза Е.coli останавливается на разрушаемой РНК, не доходя 6-9 нуклеотидов до основания черешка ближайшей шпильки [16]. Интересно, что «кор» экзосомы гипертермофильной археи S. solfataricus, состоящий из субъединиц РНКазы РН, также останавливается на расстоянии 8-9 нт от основания черешка ближайшей шпильки. Из структурных данных следует, что это обусловлено неспособностью двойной спирали черешка проникнуть в центральное отверстие бубликообразной структуры, образуемой шестью субъединицами РНКазы РН [50]. Однако, в отличие от ПНФазы Е.coli и РНКазы РН S. solfataricus, элементы внутримолекулярной вторичной структуры РНК не могут остановить Tth-ПНФазу. При оптимальной температуре реакции Tth-ПНФаза полностью разрушает структурированную РНК и остановить ее можно, только сформировав достаточной длинный двуспиральный участок посредством гибридизации РНК с комплементарным олигонуклеотидом.
Присутствие фосфатной группы на 5'-конце олигонуклеотида не влияло на длину защищаемого участка РНК (фиг.10Б; сравни дорожки 3 и 4 с дорожками 7 и 8).
Таким образом, с помощью описанной в примере 1 процедуры была впервые выделена термофильная ПНФаза, субъединицы которой имеют молекулярную массу, типичную для ПНФаз из разных организмов, и которая обладает фосфорилазной активностью в отношении структурированных РНК. Биохимическое исследование выделенного фермента выявило несколько важных свойств, которые не наблюдались у ПНФаз из других источников.
Во-первых, Tth-ПНФаза имеет максимальную фосфорилазную активность при температуре, при которой расплавляются даже очень стабильные шпильки. Поэтому она полностью разрушает РНК, которые не удается разрушить мезофильным ПНФазам. Во-вторых, для эффективного фосфоролиза Tth-ПНФазе достаточно очень низкой концентрации свободных ионов Mg2+ Это дает то преимущество, что позволяет осуществлять направленный 3'→5'-фосфоролиз при высокой температуре без риска разрыва цепей РНК вследствие Mg2+-катализируемого гидролиза. Именно благодаря этому свойству было установлено, что РНК может оставаться целой в условиях высокотемпературного фосфоролиза, если ее 3'-конец защищен. Защитить 3'-конец РНК можно либо гибридизацией с комплементарным олигонуклеотидом (учитывая, что стабильность гибрида должна обеспечивать его сохранение при температуре реакции), либо модификацией 3'-концевого нуклеотида.
Результаты однозначно демонстрируют, что 3'-концевые модификации защищают РНК от Tth-ПНФазы. Ранее было показано, что фосфатная группа в 3'- или 2'-положении концевой рибозы, как и 2',3'-циклофосфат, предотвращает фосфоролиз коротких (до 6 нт) олигорибонуклеотидов ПНФазами Е.coli [51] и М luteus [52]. Однако были опубликованы противоречивые сообщения о фосфоролизе ПНФазаой Е.coli более длинных (до 20 нт) 3'-фосфорилированных фрагментов, составляющих основную массу коммерческих препаратов РНК. По данным одних авторов, деградации подвергалось не более 20% такой РНК [53], по данным других - деградировали все 100% [54]. Противоречие результатов можно было бы объяснить либо разным средним размером фрагментов в исследованных препаратах РНК, либо разным содержанием примесных ферментов в использованных препаратах ПНФазы. Например, загрязнение фосфатазой могло приводить к удалению 3'-концевого фосфата.
Защита 3'-конца позволяет выделять определенный вид РНК из гетерогенной популяции, разрушая с помощью Tth-ПНФазы все остальные (незащищенные) цепи. Обнаружение, что биотиновая группа на 3'-конце эффективно защищает РНК от фосфоролиза, раскрывает способ получения 100% чистого препарата биотинилированной РНК. Очевидно, что могут быть предложены и другие приложения такого рода.
Наконец, продемонстрировано, что заданный 5'-концевой фрагмент РНК может быть получен посредством контролируемого фосфоролиза с точностью, сопоставимой с точностью транскрипции на обрезанной ДНК, но без необходимости наличия сайта рестрикции в данном месте. Это может быть осуществлено путем защиты цепи РНК олигонуклеотидом, комплементарным внутреннему сегменту РНК, который расположен на расстоянии 8 нт от заданного 3'-конца искомого фрагмента.
Полученные результаты демонстрируют, что защитное действие олигонуклеотида достигается именно благодаря гибридизации олигонуклеотида с 3'-концевым или с внутренним участком РНК (см. фиг.2, 5, 6, 9, 10). Хотя в приведенных примерах для защиты РНК использован олигодезоксирибонуклеотид, такую же защиту могут обеспечить олигорибонуклеотиды, смешанные олигонуклеотиды, в которых присутствуют дезоксирибо- и рибонуклеотидные звенья, а также аналоги олигонуклеотидов, в которых природный сахарофосфатный остов заменен на неприродный полимер, например, содержащий модифицированные фосфодиэфирные связи, неприродный сахарный компонент, химические аналоги фосфодиэфирной связи, полностью измененный остов, остов с 2'-5'-межнуклеотидными связями или составленный из α-нуклеотидных аномеров. Многочисленные примеры такого рода представлены в Патенте США №6329144 [55]. Примеры неприродных олигомеров, которые способны к правильному спариванию по Уотсону и Крику с комплементарным участком нуклеиновой кислоты-мишени, включают, но не исчерпываются такими химическими аналогами, как фосфоротиоатные, метилфосфонатные, боранфосфонатные производные нуклеиновых кислот [56, 57], пептидные нуклеиновые кислоты, имеющие полиамидный остов [58, 59], и морфолиновые олигонуклеотиды [60, 61]. Для защитного действия олигонуклеотида или его аналога необходимо и достаточно, чтобы он был способен гибридизоваться с РНК посредством спаривания азотистых оснований по Уотсону и Крику, и чтобы образующийся гибрид имел существенную стабильность при температуре реакции фосфоролиза.
Полученные результаты демонстрируют, что Tth-ПНФаза позволяет осуществлять полностью контролируемую деградацию РНК, что может быть использовано в различных научных и прикладных целях.
Очевидно, с теми же целями могут быть использованы другие термостабильные ПНФазы, например, ПНФаза Т. aquaticus. Более того, учитывая большое структурное и функциональное сходство бактериальных ПНФаз [25] и экзосом архей S. solfataricus [37] и Archaeoglobus fulgidus [62], можно ожидать, что архейные экзосомы обладают биохимическими свойствами, подобными свойствам Tth-ПНФазы, и также могут быть использованы для тех или иных из перечисленных выше целей. Было показано, что РНКаза РН S. solfataricus и A. fulgidus, формирующая «кор» экзосомы, сохраняет фосфорилазную активность при 60°С [37, 62] и даже при 70°С [50]. Однако фосфорилазная активность была продемонстрирована только в отношении неструктурированных олиго(А) последовательностей, в то время как РНКаза РН S. solfataricus оказалась неспособной разрушить шпильку с черешком из 8 G:C-пар даже при инкубации в течение 30 мин при 70°С [50]. Возможно, неспособность разрушать структурированные РНК является результатом того, что в «кор» экзосомы отсутствуют субъединицы, гомологичные РНК-связывающим доменам KH и S1 ПНФазы. Можно надеяться, что полная архейная экзосома, содержащая также KH- и S1-подобные субъединицы, будет эффективно разрушать структурированные РНК при высокой температуре.
Для применения предложенного способа важно использовать термостабильную ПНФазу и осуществлять реакцию в условиях [температура, концентрация ПНФазы, ее косубстратов (помимо РНК, таких как Pi) и кофакторов (таких как Mg2+), продолжительность инкубации], при которых достигается полная деградация незащищенной РНК, подобно тому, как это описано в примерах 5, 6 и 7 и показано на фиг.2, 3 и 4А. Несущественно, обладает ли термостабильностью ПНФаза, выделенная из природного источника, или ее термостабильность повышена с помощью методов генетической инженерии, в результате селекции, химической модификации или иными способами, известными в технике.
Для использования настоящего изобретения с целью селективной деградации одних видов в гетерогенных популяциях РНК при сохранении других видов, а также с целью получения укороченных вариантов РНК важно, чтобы целевые РНК или выбранные фрагменты РНК можно было защитить от деградации данной ПНФазой путем отжига с комплементарным олигонуклеотидом или путем химической модификации концевого нуклеотида. Хотя в приведенных здесь примерах защиты РНК или ее фрагмента стадия защиты предшествует стадии инкубации РНК с ПНФазой, возможно объединение этих стадий. Например, защитный олигонуклеотид можно вводить в реакционную смесь вместе с ПНФазой. Так как условия реакции (в частности, температура) близки к условиям, обеспечивающим гибридизацию защитного олигонуклеотида и РНК (в примере 5 оба процесса осуществляют при 65°С), значительная часть РНК будет в этом случае защищена.
Для практических целей предпочтительно использовать ПНФазу, которая способна эффективно расщеплять РНК при такой концентрации Mg2+ (или иного двух- или трехвалентного катиона, являющегося кофактором выбранной ПНФазы), при которой в условиях реакции, обеспечивающих эффективную энзиматическую деградацию РНК, не наблюдается существенного неэнзиматического гидролиза полирибонуклеотидов, как это показано на фиг.2 и 3.
Приведенные здесь результаты показывают, что всеми указанными свойствами обладает Tth-ПНФаза, поэтому в настоящее время ее использование является предпочтительным.
Предложенный способ поясняется, но не ограничивается, следующими конкретными примерами его осуществления.
Пример 1. Клонирование и экспрессия гена Tth-ПНФазы
Ген pnp размножают в ПЦР, используя хромосомную ДНК Т. thermophilus VK1 в качестве матрицы и праймеры 5'-GACGTCGACATATG GAAGGCACACCCAATG-3' (SEQ ID NO:1), перекрывающийся с началом белок-кодирующей последовательности гена (нуклеотиды 977-995 последовательности, депонированной в ГенБанке под № Z84207, подчеркнуты) и добавляющий в размножаемый фрагмент сайт рестрикции NdeI (выделен жирным шрифтом), и 5'-TTCGAATTCTTACTTGCGCCGCCTGGG-3' (SEQ ID NO:2), перекрывающийся с концом белок-кодирующей последовательности гена (нуклеотиды 3101-3117 последовательности, депонированной в ГенБанке под № Z84207, подчеркнуты) и добавляющий в размножаемый фрагмент сайт рестрикции EcoRI (выделен жирным шрифтом). Продукт ПЦР расщепляют по сайтам NdeI и EcoRI и лигируют в плазмидный вектор рТ7-7 [63] между этими сайтами. Полученную плазмиду pT7-PNP клонируют [64] и экспрессируют в клетках Е.coli cells B834(DE3)/pLysS [65], индуцируя синтез Т7 РНК-полимеразы в присутствии 1 мМ изопропил-β-D-тиогалактозида в течение 4 ч при 37°С.
Фиг.1А демонстрирует набор белков клеток Е.coli B834(DE3)/pLysS, не трансформированных (дорожка 1) и трансформированных (дорожка 2) плазмидой pT7-PNP, после 4-часовой индукции. Дорожка 3 содержит 0.4 мкг очищенной ПНФазы Е.coli, а также материал, вытекший из соседней дорожки 2. Анализ осуществляют следующим образом. После инкубации в SDS-содержащем буфере в течение 1 мин в кипящей водяной бане образцы подвергают электрофорезу через полиакриламидный гель в присутствии SDS [66] и окрашивают 0,04% кумасси синим G-250 в 3,5% хлорной кислоте. Черные стрелки указывают на полноразмерный полипептид Tth-ПНФазы, а белые стрелки - на полипептид ПНФазы Е.coli.
Пример 2. Выделение Tth-ПНФазы из клеток Е.coli
Все процедуры осуществляют при 4°С. Девять граммов содержащих Tth-ПНФазу клеток Е.coli, осажденных из 6 л культуры, суспендируют в 40 мл холодного буфера А [20 мМ Hepes-NaOH (рН 8,2), 100 мМ NaCl, 0,1 мМ Na-EDTA, 2 мМ β-меркаптоэтанол, 1 мМ фенилметилсульфонилфторид] и разрушают в гомогенизаторе Gaulin Micron Lab 40 (APV Thermotech GmbH, Германия) при 1400 бар. Лизат разбавляют в 10 раз буфером А и инкубируют в течение 60 мин при 70°С при перемешивании. Клеточный дебрис и свернувшиеся белки осаждают в течение 40 мин при 13000 об/мин в роторе JA-14 центрифуги J2-21 (Beckman, Австрия). Лизат наносят на 11-мл колонку DEAE-Sepharose FF (Amersham Biosciences Trading GmbH, Австрия), уравновешенную буфером Б [20 мМ Tris-HCl (pH8,0), 1 мМ Na-EDTA, 0,1 M NaCl, 2 мМ β-меркаптоэтанол, 5% глицерин]. После промывки колонки 5 объемами буфера Б фермент элюируют линейным градиентом NaCl (0.1-0.5 М) в 110 мл буфера Б. Фракции с высоким содержанием активности ПНФазы объединяют, добавляли ДНКазу I (до 1 мкг/мл) и щелочную фосфатазу из кишечника теленка (до 1 ед/мл), и диализуют в течение ночи против буфера Б, содержащего 3 мМ MgCl2 и 5 мМ Na-фосфат. После дополнительной инкубации в течение 30 мин при 37°С и затем в течение 60 мин при 66°С препарат охлаждают на льду и свернувшиеся белки осаждают, как указано выше. Надосадочную жидкость концентрируют до объема 0,25 мл на фильтре Centricon-30 (Amicon, США), осветляют в микроцентрифуге при 12000 об/мин и подвергают гель-фильтрации через 24-мл колонку Superose 6 HR (Amersham Biosciences) в буфере Б. Фракции с наивысшей активностью ПНФазы объединяют, диализуют в течение ночи против буфера для хранения [50 мМ Tris-HCl (pH 8,0), 100 мМ NaCl, 10% глицерин, 0,1% Nonidet P40, 2 мМ дитиотрейтол], осветляют, добавляют глицерин до 50% (по весу) и хранят при -70°С.
Активность ПНФазы определяют колориметрически как Pi [67], высвобождаемый из ADP при синтезе поли(А) при 70°С в растворе, содержащем 50 мМ Tris-HCl (рН 8,2), 20 мМ ADP (аденозин-5'-дифосфат), 0,1 М NaCl, 1 мМ MgCl2. Единица активности соответствует количеству фермента, полимеризующему 1 мкмоль ADP за 1 ч. Количество белка измеряют по методу Лоури и др. [68]. Электрофоретический анализ образцов осуществляют, как описано в примере 1. Содержание полноразмерного полипептида ПНФазы оценивают, анализируя гели, окрашенные кумасси синим G-250, с помощью программы ImageJ (http://rsb.info.nih.gov/ij/).
Фиг.1Б демонстрирует препарат Tth-ПНФазы на разных стадиях процедуры выделения: клеточный лизат (дорожка 3), надосадочная жидкость после осаждения клеточного дебриса и белков, свернувшихся в результате прогревания лизата в течение 60 мин при 70°С (дорожка 4), после колонки DEAE-Sepharose (дорожка 5) и после колонки Superose 6 HR (дорожка 6). Дорожка 1 содержит очищенную ПНФазу Е.coli (пример 3), а дорожка 2 - ПНФазу, выделенную из клеток Т. thermophilus HB8 [69] по методу [26], любезно предоставленную Е.А.Седельниковой (Институт биофизики РАН, Пущино).
Фиг.1В демонстрирует конечный препарат Tth-ПНФазы (дорожки 4 и 5, содержащие приблизительно 4 и 2 мкг белка, соответственно), подвергнутый электрофорезу в присутствии SDS вместе с ПНФазой Е.coli (дорожки 2 и 3, содержащие 2 и 1 мкг белка) и Qβ-репликазой (дорожка 1, 2 мкг белка), которые использовали в качестве стандартов размера. Qβ-репликаза содержит 4 субъединицы [70], одна из которых - белок S1 рибосом Е.coli (61.2 кДа) - имеет аномально низкую подвижность при SDS-электрофорезе [71].
Пример 3. Выделение ПНФазы Е.coli
Все процедуры осуществляют при 4°С. Двести граммов клеток Е.coli В суспендируют в 200 мл буфера В [50 мМ Tris-HCl (pH 8,0), 1 мМ Na-EDTA, 10 мМ MgCl2, 0,2 M KCl, 5 мМ β-меркаптоэтанол, 5% глицерин, 1 мМ фенилметилсульфонилфторид] и разрушают в гомогенизаторе 15М-8ТА (APV Gaulin, Inc., США) при 600 бар. После удаления клеточного дебриса (в течение 40 мин при 14,000 об/мин в роторе JA-14) компоненты лизата разделяют между жидкими полимерными фазами полиэтиленгликоля 6000 и декстрана Т500 с последующей экстракцией РНК-связывающих белков солью, как описано для процедуры выделения Qβ-репликазы [70]. Солевой экстракт диализуют против буфера Г [10 мМ Tris-HCl (pH 7,5), 1 мМ Na-EDTA, 5 мМ MgCl2, 5 мМ β-меркаптоэтанол, 5% глицерин], пока проводимость экстракта не снижалась до проводимости буфера Г100 (буфер Г + 100 мМ NaCl), осветляют в течение 40 мин при 14,000 об/мин в роторе JA-14 и наносят на 200-мл колонку DE-52 (Whatman, США), уравновешенную в буфере Г100. Колонку промывают 2,5 объемами буфера Г100, затем 2,5 объемами буфера Г150 (буфер Г + 150 мМ NaCl) и элюируют линейным градиентом NaCl (150-400 мМ) в 2 л буфера Г. Фракции с высокой активностью ПНФазы объединяют, белки высаливают сульфатом аммония (40 г на 100 мл элюата), осадок растворяют в 20 мл буфера Г100 и диализуют против того же буфера в течение ночи. После диализа препарат осветляют, добавляют 50 мкг ДНКазы I, 50 ед. щелочной фосфатазы из кишечника теленка, ZnCl2 и фенилметилсульфонилфторид (до 1 мМ каждого) и инкубируют в течение 60 мин при 37°С. Затем добавляют 1 М K-фосфат (pH 8,0) до концентрации 100 мМ и инкубируют еще в течение 30 мин. После автолиза препарат осветляют и подвергают гель-фильтрации через колонку Sepharose CL-6B (2,6×100 см), уравновешенную в буфере Г100. Фракции с высокой активностью ПНФазы объединяют и хроматографируют в несколько порций на колонке Mono Q HR 5/5 column (Amersham Biosciences), уравновешенной в буфере Г100, используя градиент NaCl (100-1000 мМ) в буфере Г. Активные фракции объединяют, концентрируют на фильтре Centricon-30 (Amicon) и подвергают гель-фильтрации через 24-мл колонку Superose 6 HR (Amersham Biosciences). После анализа с помощью SDS-электрофореза объединяют фракции, содержащие наименьшее количество примесей, добавляют глицерин до 50% (по весу) и хранят при -70°С.
Конечный препарат (2,2 мг белка) содержал >90% полипептида ПНФазы (фиг.1В, дорожки 2 и 3) и был свободен от эндонуклеазной активности. Удельная активность препарата составляла ≈450 ед/мг. За единицу активности принимают количество фермента, полимеризующего 1 мкмоль ADP в течение 1 ч при 37°С в растворе, содержащем 100 мМ Tris-HCl (pH 8,0), 10 мМ ADP, 0.5 мМ Na-EDTA, 10 мМ MgCl2.
Пример 4. Получение препаратов РНК
В данной работе использовали следующие препараты РНК:
(1) CT1n1 РНК, вариант RQ135-1 РНК [43], несущий 43-нт вставку (подчеркнута), выделенный из продуктов межмолекулярной рекомбинации РНК:

 Препарат этой РНК получают транскрипцией плазмиды, кодирующей 3'-фрагмент [72], расщепленной по сайту PvuII.
Препарат этой РНК получают транскрипцией плазмиды, кодирующей 3'-фрагмент [72], расщепленной по сайту PvuII.
Все препараты РНК получают с помощью Т7 РНК-полимеразы и очищают с помощью электрофореза в полиакриламидном геле, как описано ранее [72].
Пример 5. Исследование зависимости деградации РНК от концентрации ионов Mg2+
(1) Расплавление образцов. Непосредственно перед экспериментом, РНК [в 0,1 мМ Na-EDTA (pH 8,0)] и олигодезоксирибонуклеотид [в 10 мМ Tris-HCl (pH 9,0), 0,01 мМ Na-EDTA] раздельно расплавляют в кипящей водяной бане в течение 2 мин, после чего быстро охлаждают на льду.
(2) Гибридизация РНК с олигонуклеотидом. Расплавленные РНК (0,5 пмоль) и олигодезоксирибонуклеотид (2,5 пмоль) смешивают в 5 мкл раствора, также содержащего 80 мМ Hepes-KOH (pH 8,1), 150 мМ KCl, 0,2 мМ Na-EDTA и инкубируют в течение 15 мин при 65°С.
(3) Реакция фосфоролиза. Реакционная смесь объемом 10 мкл имела следующий состав: 50 мМ Hepes-KOH (pH 8,1), 75 мМ KCl, 1 мМ дитиотрейтол, 1 мМ Na-фосфат, 0,5 пмоль 3'-фрагмента RQ135 РНК, 2 пмоль Tth-ПНФазы, где указано - 2,5 pmol олигодезоксирибонуклеотида R-38 [5'-GGGCTAACAGTGCGGTAACACGCACTGGAGGTCCGAAA-3' (SEQ ID NO:8), синтезирован ЗАО "Синтол", Россия], комплементарного 3'-концевой последовательности 3'-фрагмента, и указанные на фиг.2 концентрации MgCl2. Сверх того, в каждый образец вместе с препаратами ПНФазы и РНК вносят 0,056 мМ EDTA. Реакцию осуществляют в течение 15 мин при 65°С и останавливают добавлением 10 мкл 10 мМ Na-EDTA.
(4) Анализ продуктов реакции. Нуклеиновые кислоты выделяют с помощью фенольной экстракции [64] и разделяют с помощью электрофореза в полиакриламидном геле в неденатурирующих условиях, в присутствии буфера ТВЕ: 100 мМ Tris-OH, 100 мМ борная кислота, 2 мМ Na-EDTA. После электрофореза гель окрашивают серебром [73]. Фиг.2 демонстрирует результат эксперимента.
Пример 6. Исследование зависимости деградации РНК от температуры
Реакцию фосфоролиза осуществляют при указанных на фиг.3 температурах в присутствии или в отсутствие 0,5 пмоль Tth-ПНФазы, как описано в примере 5, за исключением того, что олигонуклеотид не добавляют, гибридизацию не проводят, используют постоянную концентрацию MgCl2 (0,1 мМ), а продукты разделяют с помощью электрофореза в полиакриламидном геле в денатурирующих условиях: в присутствии буфера ТВЕ, содержащего 7М мочевину, и окрашивают серебром [73]. Результат показан на фиг.3.
Пример 7. Исследование зависимости деградации РНК от концентрации ПНФазы
Реакцию фосфоролиза осуществляют при постоянной температуре (65°С) в течение указанного на фиг.4А времени при указанном на фиг.4А отношении ПНФаза: РНК. Концентрация РНК составляла 50 нМ. Остальные условия эксперимента, как в примере 6. Результат показан на фиг.4А.
Пример 8. Исследование зависимости образования комплекса ПНФаза-РНК от концентрации ПНФазы
0.5 пмоль [α-32Р]-меченого 3'-фрагмента RQ135 РНК, полученного с помощью транскрипции в присутствии [α-32P]CTP, инкубируют, в отсутствие или в присутствии Tth-ПНФазы, взятой в указанном на фиг.4Б отношении к РНК, в течение 15 мин при 65°С, в 10 мкл буфера 50 мМ Hepes-KOH (рН 8,1), 75 мМ KCl, 1 мМ дитиотрейтол, 0,1 мМ MgCl2. После электрофореза в неденатурирующих условиях [пример 5 (4)] гель экспонируют в течение ночи на экран MS и получают изображение радиоактивных полос, сканируя экран на фосфоримиджере Cyclone™ (Packard Instrument, США). Результат показан на фиг.4Б.
Пример 9. Определение полноты деградации РНК
CT1n1 РНК либо ее гибрид с олигонуклеотидом R-38 (SEQ ID NO:8), полученный, как описано в примере 5(2), инкубируют в присутствии или в отсутствие Tth-ПНФазы в условиях реакции фосфоролиза [пример 5(3)] в присутствии 0,1 мМ MgCl2. Затем образцы экстрагируют фенолом [64], водную фазу смешивают с 6 объемами смеси этанол: ацетон (1:1 по объему) в присутствии 20 мкг линейного полиакриламида [74] и 300 мМ KCl, нуклеиновые кислоты осаждают в микроцентрифуге при 12000 g, осадок дважды промывают 80% этанолом и растворяют в 6 мкл воды. Остаточное количество CT1n1 РНК определяют с помощью ОТ-ПЦР следующим образом.
(1) Обратная транскрипция (ОТ). Реакционная смесь объемом 10 мкл содержала 50 мМ Tris-HCl (рН 8,3), 79 мМ KCl, 3 мМ MgCl2, 10 мМ дитиотрейтол, 0,2 мМ каждого из dATP, dGTP, dTTP и dCTP, 0.5 мкМ праймера R-38 (SEQ ID NO:8), 50 нг поли(А), 50 ед. обратной транскриптазы M-MLV (Amersham Biosciences) и анализируемый образец РНК. После инкубации в течение 60 мин при 45°С образцы осаждали смесью этанол: ацетон, как описано выше.
(2) ПЦР. Реакции осуществляли в ДНК-амплификаторе RapidCycler™ (Idaho Technology, США) в капиллярах объемом 10 мкл, содержащих 50 мМ Tris-HCl (рН 8,7), 2,5 mM MgCl2, 0,2 mM каждого из dATP, dGTP, dTTP и dCTP, 0,2 мкМ каждого из праймеров R-38 (SEQ ID NO:8) и F-39
[5'-AAGCTTAATACGACTCACTATAGGGGTTCCAACCGGAAG-3' (SEQ ID NO:9), подчеркнут участок, совпадающий с 5'-концевой последовательностью RQ135 кДНК], 10 мкг казеина, 12 нг His6-модифицированной Taq ДНК-полимеразы [75] и часть полученного на стадии ОТ препарата кДНК, соответствующая указанному на фиг.5А стартовому числу молекул CT1n1 РНК. Препарат кДНК разводили в буфере 10 мМ Tris-HCl (рН 9,0), 0,1 мМ Na-EDTA, 0,1% полиэтиленгликоль 6000, 10 нг/мкл поли(А). После пяти начальных циклов ПЦР (расплавление при 94°С в течение 10 с, отжиг при 60°С в течение 10 с, элонгация при 70°С в течение 30 с) выполняют 35 циклов, в которых температуру расплавления понижали до 88°С, а температуру отжига повышали до 65°С.
Продукты ОТ-ПЦР разделяли в неденатурирующих условиях [пример 5 (4)] и окрашивали бромистым этидием. Результаты представлены на фиг.5А. ПНФаза присутствовала во всех образцах, кроме помеченных «-ПНФаза».
Пример 10. Селективная деградация РНК, не защищенной комплементарным олигонуклеотидом
Смесь 0,5 пмоль 3'-фрагмента и 0,5 пмоль 5'-фрагмента RQ135 РНК гибридизуют с олигонуклеотидом R-38 (SEQ ID NO:8) [пример 5(2)], инкубируют в присутствии или в отсутствие Tth-ПНФазы в условиях реакции фосфоролиза в присутствии 0,1 мМ MgCl2 [пример 5(3)] и анализируют в неденатурирующих условиях, как описано в примере 5(4). Результат представлен на фиг.5Б.
Пример 11. Защита РНК с помощью химической модификации ее 3'-конца
Для защиты РНК от деградации Tth-ПНФазой применяют следующие способы модификации 3'-концевых нуклеотидов на стадии, предшествующей реакции фосфоролиза.
(1) Уничтожение гидроксильных групп 3'-концевой рибозы. Этого достигают путем окисления РНК перйодатом натрия. Процедуру осуществляют, как описано в работе [72].
(2) Получение РНК с фосфорилированным 3'-гидроксилом 3'-концевой рибозы. Такую РНК получают путем обработки аналином РНК, предварительно окисленной перйодатом натрия. Процедура описана в работе [72]. Также обрабатывают смесь окисленной и неокисленной РНК; в этом случае в реакцию с анилином вступала только окисленная РНК. Для снятия защиты РНК дефосфорилируют, как описано в той же работе, используя фосфатазу креветок (Amersham Biosciences).
(3) Получение 3'-биотинилированной РНК. РНК, предварительно окисленную перйодатом натрия, инкубируют в течение 20 ч при 4°C с 5 мМ гидразидом биотина (Sigma, США) в присутствии 0,1% SDS и 0,2 М Na-ацетата (рН 4,8). Невключенный гидразид биотина удаляют, пропуская образец через колонку Sephadex G-25 (Amersham Biosciences) с помощью центрифугирования.
РНК, модифицированные способами (1) или (2) или не модифицированные, подвергают фосфоролизу в течение 15 мин при 65°С и анализируют в денатурирующих условиях, как описано в примере 6. Результаты представлены на фиг.6, части А, Б и В. Показан результат фосфоролиза смеси 0,5 пмоль 3'-фрагмента (верхняя полоса) и 0,5 пмоль 5'-фрагмента RQ135 РНК (нижняя полоса), из которых либо 5'-фрагмент (А и В, «исходная смесь»), либо 3'-фрагмент (Б) был окислен перйодатом натрия («3'oxi»), a другой фрагмент не был модифицирован («3'ОН»). Смесь либо не обрабатывали («н/о»), либо обрабатывали указанным количеством (в пмоль) Tth-ПНФазы, либо сразу (А, Б и В, «исходная смесь»), либо после обработки смеси анилином, которая превращала окисленный фрагмент в 3'-фосфорилированный (В, «после анилина»), либо после обработки смеси анилином и затем фосфатазой, которая удаляла 3'-фосфат (В, «после фосфатазы»).
Фиг.6Г демонстрирует результат электрофореза в неденатурирующих условиях [пример 5(4)] 3'-фрагмента RQ135 РНК, либо модифицированного биотином на 3'-конце («3'bio»), либо не модифицированного («3'ОН»), который был либо не обработан («-»), либо обработан 2 пмоль Tth-ПНФазы («+»), либо был смешан с 5-кратным молярным избытком стрептавидина перед электрофорезом («Стр»). По полосе РНК, подвижность которой при электрофорезе уменьшается в результате образования комплекса со стрептавидином, можно судить о том, какая доля молекул 3'-фрагмента модифицирована биотином.
Пример 12. Сравнение свойств Tth-ПНФазы и ПНФазы Е.coli
Фиг.7 демонстрирует результаты эксперимента, в котором 20 нг поли(А) или 0,5 пмоль 3'-фрагмента RQ135 РНК, 3'-конец которого был либо не модифицирован («3'ОН»), либо фосфорилирован («3'Р») в результате последовательных реакций с перйодатом натрия и анилином [пример 11, (1) и (2)], были либо не обработаны («-»), либо обработаны («+») в присутствии 1 мМ Pi и 0.1 мМ Mg2+ с помощью 4 пмоль ПНФазы Е.coli при 37°С (Е) или 1 пмоль Tth-ПНФазы при 65°С (Т). После реакции образцы обрабатывали и анализировали, как описано в примере 6.
Пример 13. Влияние температуры и концентрации неорганического фосфата на деградацию структурированной РНК ПНФазой Е.coli
Фиг.8 демонстрирует результаты эксперимента, в котором 0,5 пмоль 3'-фрагмента RQ135 РНК обрабатывают с помощью 2 пмоль ПНФазы Е.coli PNPase в течение 15 мин при указанных значениях температуры и концентрации неорганического фосфата («Pi»). После реакции образцы обрабатывают и анализируют, как описано в примере 6.
Пример 14. Применение Tth-ПНФазы для укорачивания РНК с 3'-конца
Фиг.9 демонстрирует результаты эксперимента, в котором РНК-транскрипты одной и той же плазмиды, расщепленной по сайтам рестрикции SmaI (длина транскрипта 109 нт, SEQ ID NO:5), EcoRI (127 нт, SEQ ID NO:6) или PvuII (303 нт, SEQ ID NO:7) (каждый более длинный фрагмент целиком содержит в себе последовательности более коротких фрагментов, то есть 1-109 нуклеотиды во всех фрагментах одинаковы, пример 4), либо обрабатывали («+»), либо не обрабатывали («-») с помощью Tth-ПНФазы в присутствии 0,1 мМ MgCl2 после отжига этих транскриптов, поодиночке или в присутствии олигонуклеотида R-38 (SEQ ID NO:8), комплементарного нуклеотидам 72-109. Для осуществления эксперимента использовали условия и процедуры, изложенные в примере 5.
С целью определения точной длины укороченных фрагментов используют транскрипты, [32P]-меченные на 5'-конце, и продукты деградации разделяют с помощью электрофореза высокого разрешения в денатурирующих условиях, применяемого для секвенирования нуклеиновых кислот [64, 76]. Для приготовления 5'-меченых транскриптов, немеченые транскрипты (пример 4) обрабатывают фосфатазой с целью удаления 5'-трифосфата, а затем фосфорилируют, используя полинуклеотидкиназу и [γ-32P] ATP [64]. После электрофореза гель экспонируют в течение ночи на экран MS и получают изображение радиоактивных полос, сканируя экран на фосфоримиджере Cyclone™ (Packard Instrument, США). Результаты приведены на фиг.10.
Часть А демонстрирует результаты, полученные с использованием олигонуклеотида R-38, химически синтезированного ЗАО "Синтол" (Россия) без каких-либо дополнительных обработок. Дорожки 1 и 2 содержат полноразмерные транскрипты SmaI и EcoRI, а дорожки 3 и 4 - обработанные Tth-ПНФазой транскрипты EcoRI и PvuII соответственно.
Часть Б демонстрирует результаты, полученные с использованием олигонуклеотида R-38 с точным 5'-концом, полученного из более длинного олигонуклеотида с помощью эндонуклеазы рестрикции SmaI, как описано в примере 15. Дорожка 1 содержит полноразмерный транскрипт SmaI, дорожка 5 - транскрипт EcoRI, частично деградированный слабой щелочью с целью генерации
1-нуклеотидной «лесенки» [76]. Дорожки 2-4 содержат продукты деградации транскриптов SmaI, EcoRI и PvuII соответственно, гибридизованных с олигонуклеотидом R-38, 5'-конец которого фосфорилирован, так как эндонуклеаза рестрикции расщепляет межнуклеотидную связь между фосфатом и 3'-гидроксилом [64]. Дорожки 6-8 содержат продукты деградации тех же транскриптов, гибридизованных с тем же олигонуклеотидом, дополнительно обработанным фосфатазой [64] для удаления 5'-фосфата.
Пример 15. Получение олигодезоксирибонуклеотида с точным 5'-концом
Используют 2 олигодезоксирибонуклеотида - R-50 [5'-TGGAATTCCCCC-GGGCTAACAGTGCGGTAACACGCACTGGAGGTCCGAAA-3' (SEQ ID NO:10)], целиком включающий последовательность олигонуклеотида R-38 (SEQ ID NO:8) (подчеркнута), удлиненную на 5'-конце с целью генерации сайта рестрикции SmaI, и F-24 [5'-CACTGTTAGCCCGGGGGAATTCCA-3' (SEQ ID NO:11)], комплементарный олигонуклеотиду R-50 в области сайта рестрикции SmaI. Оба олигонуклеотида были синтезированы ЗАО "Синтол", Россия. Олигонуклеотиды гибридизуют между собой, как описано в примере 5(2). Это приводило к образованию структуры, изображенной на фиг.11. Гибрид расщепляют с помощью эндонуклеазы рестрикции SmaI (место расщепления указано стрелкой на фиг.11) и олигонуклеотид R-38, наибольший из образующихся фрагментов, выделяют с помощью электрофореза в полиакриламидном геле в денатурирующих условиях.
ПРОМЫШЛЕННАЯ ПРИМЕНИМОСЬ
Осуществление изобретения в соответствии с настоящими описанием, примерами и формулой позволяет, в частности:
(1) Достигать полной деградации незащищенных РНК благодаря осуществлению реакции при высокой температуре, когда расплавлены даже самые стабильные элементы вторичной структуры РНК.
(2) Выделять индивидуальные РНК из гетерогенных популяций путем защиты 3'-конца целевой РНК с помощью химической модификации или путем гибридизации с комплементарным олигонуклеотидом, и последующей деградации незащищенных РНК.
(3) Обогащать препарат РНК, модифицированной по 3'-концу, путем деградации немодифицированных молекул.
(4) Укорачивать РНК на заданную величину с 3'-конца, защищая РНК посредством гибридизации с олигонуклеотидом, комплементарным участку РНК, лежащему на 8 нт в 5'-направлении от запланированного 3'-конца продукта.
Настоящее изобретение может быть также использовано с целью деградации всех РНК, содержащихся в образце. Это может быть полезным, например, когда необходимо удалить примеси РНК из препарата ДНК.
Список использованных источников




















Изобретение относится к области молекулярной биологии и биотехнологии и может быть использовано в препаративной биохимии РНК. Согласно предложенному способу осуществляют деградацию структурированных полирибонуклеотидов путем инкубирования содержащего их образца с термостабильной полинуклеотидфосфорилазой (ПНФазой) Thermus thermophilus в присутствии косубстрата и кофактора. Изобретение позволяет осуществлять ферментативный фосфоролиз РНК при высокой температуре, избегая Mg2+-катализируемого гидролиза. 6 з.п. ф-лы, 11 ил.
1. Способ деградации структурированных полирибонуклеотидов, включающий инкубацию образца, содержащего, по меньшей мере, один вид полирибонуклеотидов, в растворе, содержащем 3'→5'экзорибонуклеазу, ее косубстрат и кофактор, отличающийся тем, что в качестве 3'→5'экзорибонуклеазы используют полинуклеотидфосфорилазу Thermus thermophilus, а инкубацию образца с ферментом осуществляют в диапазоне температур 60-75°С, при концентрации ионов Mg2+ в диапазоне от 1 мкМ до 44 мкМ, что обеспечивает существенно полную деградацию тех полирибонуклеотидов образца, в отношении которых не предприняты какие-либо способы защиты, повышающие устойчивость полирибонуклеотидов к данной экзорибонуклеазе.
2. Способ по п.1, отличающийся тем, что, по меньшей мере, один из полирибонуклеотидов защищают от действия экзорибонуклеазы.
3. Способ по п.2, отличающийся тем, что полирибонуклеотид защищают путем химической модификации 3'-концевого нуклеотида.
4. Способ по п.3, отличающийся тем, что в качестве модифицирующей группы используют фосфат или биотин.
5. Способ по п.2, отличающийся тем, что полирибонуклеотид защищают путем гибридизации с олигонуклеотидом или аналогом олигонуклеотида, способным образовать с данным полирибонуклеотидом гибрид, имеющий существенную стабильность при температуре инкубации.
6. Способ по п.5, отличающийся тем, что используют олигонуклеотид или аналог олигонуклеотида, комплементарный 3'-концевому участку полирибонуклеотида.
7. Способ по п.5, отличающийся тем, что используют олигонуклеотид или аналог олигонуклеотида, комплементарный внутреннему участку полирибонуклеотида.
| FALALEEVA M.V | |||
| et al., Polynucleotide phosphorylase from Thermus thermophilus as a tool for studies on RNA recombination", сборник тезисов к международной конференции "Protein biosynthesis, structure and function", 2007, June 9-13 2007, p.31 | |||
| FALALEEVA M.V | |||
| et al., Factors influencing RNA degradation by Thermus thermophilus polynucleotide |

