ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к процедуре сцепления когнатных пар кодирующих последовательностей VH и VL из популяции клеток, обогащенных специфическими поверхностными антигенными маркерами. Процедура сцепления включает мультиплексную процедуру молекулярной амплификации, способную в связи с амплификацией сцеплять представляющие интерес нуклеотидные последовательности в особой полимеразной цепной реакции (мультиплексной PCR). Этот способ особенно эффективен для создания библиотек когнатных пар, а также комбинаторных библиотек кодирующих последовательностей вариабельных областей из иммуноглобулина. Изобретение также относится к способам получения химерных человеческих/отличных от человеческих антител и библиотек экспрессии, полученных такими способами.
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
Документ WO 2005/042774 (Symphogen) раскрывает способ сцепления представляющих нуклеотидных последовательностей, в частности когнатных пар кодирующих последовательностей VH и VL, с применением мультиплексной молекулярной процедуры. В предпочтительном варианте представляющие интерес последовательности амплифицируют и сцепляют из выделенных единичных клеток после их серийного разведения или разделения при помощи других методик. Указанная ссылка раскрывает различные способы обогащения клеточной популяции, содержащей лимфоциты, для получения популяции плазмацитов, которые особенно пригодны для процедуры мультиплексной молекулярной амплификации.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение направлено на способы создания библиотек кодирующих последовательностей иммуноглобулинов, происходящих из животных, не являющихся человеком, и способах создания в несколько стадий, адаптированных к высокопроизводительному клонированию и скринингу, библиотек векторов, кодирующих химерные антитела, которые содержат константные области человека и вариабельные области, отличные от человеческих.
В первом аспекте изобретение относится к способу получения библиотеки когнатных пар, содержащих сцепленные кодирующие последовательности вариабельных областей, где указанный способ включает:
a) получение от донора клеточной фракции, содержащей лимфоциты,
b) получение популяции выделенных единичных клеток, включающее распределение клеток из указанной клеточной фракции индивидуально во множество резервуаров, где, по меньшей мере, субпопуляция клеток экспрессирует антиген CD43 и CD138 или антиген MHCII и B220, и
c) амплификацию и осуществление сцепления кодирующих последовательностей вариабельных областей, содержащихся в указанной популяции выделенных единичных клеток, посредством амплификации в мультиплексной процедуре молекулярной амплификации представляющих интерес нуклеотидных последовательностей с использованием матрицы, полученной из выделенной единичной клетки или популяции изогенных клеток, и осуществления сцепления представляющих интерес нуклеотидных последовательностей.
Этот способ обеспечивает библиотеку когнатных пар антител или фрагментов антител. Еще в одном аспекте изобретение относится к способу сцепления случайным образом множества представляющих интерес несмежных нуклеотидных последовательностей, где указанный способ включает:
a) амплификацию в мультиплексной процедуре молекулярной амплификации представляющих интерес нуклеотидных последовательностей с использованием матрицы, полученной из популяции генетически различных клеток,
b) где генетически различные клетки получены из клеточной фракции, содержащей лимфоциты, от донора,
c) где, по меньшей мере, субпопуляция клеток экспрессирует антиген CD43 и CD138 или антиген MHCII и B220,
d) осуществление сцепления представляющих интерес нуклеотидных последовательностей, амплифицированных на стадии a).
Этот способ обеспечивает комбинаторную библиотеку комбинированных случайным образом вариабельных доменов тяжелой и легкой цепи.
Экспериментальные данные, представленные в настоящей заявке, доказывают, что клеточные популяции, выделенные из мышиных спленоцитов и положительные в отношении перечисленных поверхностных антигенов, обеспечивают хороший стартовый материал для клонирования кодирующих последовательностей антител с применением способа мультиплексной молекулярной амплификации. Способы, предлагаемые изобретением, легко применимы к другим биологическим видам, экспрессирующим ортологи CD43 и CD138 или MHCII и B220, и в особенности эти способы применимы к другим грызунам, например крысам.
Способ дает несколько преимуществ по сравнению с альтернативами, например, созданием гибридомы. При получении гибридомы из иммунизированной мыши установленные клеточные линии будут кодировать репертуар разных изотипов антител. Впоследствии клеточные линии гибридомы необходимо скринировать как на функцию (например, на связывание со специфическим антигеном, эффективность нейтрализации патогена), так и на изотип антитела. Таким образом, в этом случае необходима двухстадийная процедура скрининга для отбора гибридомы со специфическим изотипом антитела и со специфическим эффектом в функциональном анализе. Наконец, для того чтобы создать клеточную линию-продуцент, необходимо клонировать и секвенировать антитело, секретируемое отобранной гибридомой прежде, чем его будет можно перенести в клеточную линию-продуцент.
Используя способ, предложенный настоящим изобретением, можно детерминировать изотип антитела посредством праймеров, использованных для мультиплексной молекулярной амплификации, следовательно, необходимость в его (изотипа) установлении отпадает. Изотип антитела также можно детерминировать (и изменить) при последующем сцеплении или сплайсинге константного домена (доменов) с клонированными вариабельными последовательностями. Кроме того, способ, предложенный изобретением, обеспечивает библиотеку полинуклеотидов, которую можно легко секвенировать и/или вставить в векторы, например, такие как векторы экспрессии, переноса, дисплея или челночные векторы, таким образом, если отобрано конкретное антитело, оно уже клонировано, его последовательность уже известна и его можно легко перенести в соответствующий вектор экспрессии для получения антитела.
Ожидается, что клетки, отсортированные в соответствии с предложенным протоколом, будут служить источником высокоаффинных антител, возможно, с аффинностью в пикомолярном диапазоне. Моноклональные антитела из гибридомы не обладают аффинностью в пикомолярном диапазоне и для достижения такой аффинности нуждаются в синтетическом созревании аффинности.
В соответствии с изобретением эти способы дополнительно включают перед мультиплексной молекулярной амплификацией оценку того, содержит ли популяция клеток, содержащая лимфоциты, клетки, экспрессирующие антиген CD43 и CD138 или антиген MHCII и B220, предпочтительно CD43 и CD138. Эти способы также могут включать обогащение указанной клеточной фракции, содержащей лимфоциты, популяцией лимфоцитов, экспрессирующих антиген CD43 и CD138 или антиген MHCII и B220, перед мультиплексной молекулярной амплификацией.
Предпочтительные способы дополнительно включают выделение из указанной популяции, содержащей лимфоциты, клеток, экспрессирующих антиген CD43 и CD138 или антиген MHCII и B220, перед мультиплексной молекулярной амплификацией. В предпочтительном варианте осуществления изобретения выделенные клетки или субпопуляция клеток представляют собой CD138 High/CD43 High или CD138 Intermediate/CD43 High клеточную фракцию с лимфоцитами, в которых антиген CD43 представлен на высоком, а антиген CD138 на высоком или промежуточном уровне по сравнению с клеточной фракцией, содержащей лимфоциты. Более предпочтительно выделенные клетки или субпопуляция клеток представляют собой CD138 High/CD43 High относительно клеточной фракции, содержащей лимфоциты.
Предпочтительно, чтобы обогащение или выделение включали автоматическую процедуру сортинга клеток, например MACS или FACS.
В следующем аспекте изобретение относится к способу получения вектора, кодирующего химерное антитело с константными областями человека и вариабельными областями, отличными от человеческих, где указанный способ включает:
a) получение клеточной фракции, содержащей лимфоциты, от животного, не являющегося человеком,
b) получение популяции выделенных единичных клеток, включающее распределение клеток из указанной клеточной фракции индивидуально во множество резервуаров,
c) амплификацию и осуществление сцепления кодирующих нуклеиновых кислот вариабельных областей, содержащихся в указанной популяции выделенных клеток, посредством амплификации указанных нуклеиновых кислот в мультиплексной процедуре молекулярной амплификации с использованием матрицы, полученной из выделенной единичной клетки или популяции изогенных клеток, и осуществления сцепления амплифицированных нуклеиновых кислот, кодирующих вариабельные области тяжелой и легкой цепей,
d) осуществление сцепления амплифицированных вариабельных областей с константными областями человека,
e) инсерцию полученных нуклеиновых кислот в вектор. Предпочтительным животным, не являющимся человеком, является мышь. В той степени, до которой способы, предлагаемые изобретением, применимы к мышиным клеткам, эти способы называются: Mouse-Symplex™ или mSymplex™.
В этом аспекте изобретения предлагается новый способ создания библиотек химерных антител человека/отличных от человека животных. Это стало возможным благодаря объединению мультиплексной молекулярной амплификации и последующего клонирования в скелет вектора с лигированием и/или сплайсингом константных доменов тяжелой и легкой цепи человека. Традиционно в способе получения химерных антител человека/отличного от человека животного химеризация была последней стадией после установления и скрининга гибридомы, а также клонирования кодируемого антитела. Химеризация может повлиять на специфичность связывания и/или аффинность антитела, поэтому существует риск того, что хорошее моноклональное мышиное антитело утратит свою эффективность при химеризации в антитело человека/мыши.
Предлагая способ, который напрямую генерирует антитело из категории химерных антител, можно осуществлять скрининг на продуктах, которые не нуждаются в дополнительной модификации перед доклинической и клинической стадиями разработки.
Константные области человека могут быть обеспечены на стадии молекулярной амплификации или быть частью векторного скелета, в котором клонируют вариабельные области после молекулярной амплификации. В предпочтительном варианте осуществления изобретения способ включает дополнительную стадию амплификации, в которой полинуклеотид, кодирующий константную легкую цепь человека или ее фрагмент с перекрыванием, способным обеспечить сцепление с вариабельной легкой цепью, добавляют к смеси PCR вместе с набором праймеров, способным обеспечить амплификацию конструкции, содержащей в следующем порядке: мышиную цепь VH, линкер, мышиную цепь VL и константную легкую цепь человека.
В другом варианте осуществления изобретения способ включает дополнительную стадию амплификации, в которой полинуклеотид, кодирующий константную тяжелую цепь человека или ее фрагмент с перекрыванием, способным обеспечить сцепление с вариабельной тяжелой цепью, добавляют к смеси PCR вместе с набором праймеров, способным обеспечить амплификацию конструкции, содержащей в следующем порядке: константную тяжелую цепь человека, мышиную цепь VH, линкер и мышиную цепь VL.
Вследствие этого также предлагается библиотека векторов, кодирующих химерные антитела, где каждый член, относящийся к антителу, состоит из кодирующих последовательностей вариабельных областей иммуноглобулина, отличного от человеческого, и кодирующих последовательностей константных областей тяжелой и легкой цепи иммуноглобулина человека. Предпочтительно, чтобы векторы представляли собой векторы экспрессии, обеспечивающие экспрессию членов библиотеки, относящихся к антителу, для последующего скрининга. Более предпочтительно, чтобы вектор экспрессии был приспособлен к экспрессии в организме млекопитающего.
Векторы, входящие в состав библиотеки, могут быть получены способом, предлагаемым изобретением.
В одном из вариантов осуществления изобретения константная область легкой цепи представляет собой константную область каппа.
Последовательности, отличные от человеческих, могут быть получены от крысы, овцы, козы, кролика, морской свинки или другого подходящего животного, для которого существуют протоколы иммунизации, для которого доступна информация о последовательностях, позволяющая сконструировать подходящие праймеры, и для которого доступны подходящие методики клеточного сортинга, позволяющие отсортировать плазмациты для мультиплексной молекулярной амплификации единичной клетки с целью сцепления когнатных пар последовательностей вариабельных областей. В одном из вариантов осуществления изобретения последовательности, отличные от человеческих, могут происходить от не являющегося человеком примата, например обезьяны cynomolgous, от обезьяны резус, от шимпанзе или от макака. Предпочтительно, чтобы последовательности, отличные от человеческих, происходили от грызунов, например от мыши или крысы. Еще в одном варианте осуществления изобретения последовательности, отличные от человеческих, представляют собой последовательности кролика.
Предпочтительно, чтобы вариабельные области антител представляли собой когнатные пары.
Еще в одном аспекте изобретение относится к вспомогательной библиотеке, которая кодирует антитела, проявляющие желаемые специфичности связывания, направленные к конкретной мишени, выбранной из библиотеки в соответствии с изобретением.
ОПИСАНИЕ ФИГУР
Фиг.1. Мышиная модель - mSymplex™ PCR: Расширение мультиплексного перекрывания RT-PCR для амплификации и когнатного сцепления генов тяжелой и легкой цепи антитела из единичной клетки. Типичные смеси праймеров, использованные для RT-PCR и вложенной PCR, подробно описаны в таблице 2 и таблице 3 (или в таблице 5).
Фиг.2. Клонирование мышиного репертуара: Пул продуктов mSymplex™ PCR, кодирующих пары генов VH/VL из единичных плазматических клеток, был сплайсирован до гена, кодирующего константную легкую каппа-цепь человека, посредством сплайсинга через расширение перекрывания. Пул генов, кодирующих полноразмерные химерные антитела человека-мыши, был вставлен в вектор экспрессии с последующей вставкой двунаправленной промоторной кассеты (2×CMV). Смеси праймеров, использованные для сплайсинга каппа человека, подробно описаны в таблице 6.
Фиг.3A. Сортинг мышиных спленоцитов. (A) Для выделения плазмацитов, определенных как CD43 high (с высоким содержанием CD43), CD138 high (с высоким содержанием CD138), клетки, положительные по PI (пропидиум иодид), или мертвые клетки были исключены на нижней левой панели (не P1). Затем плазмациты были отсортированы как CD43 high, CD138 high на нижней правой панели (P2). Наконец, дубликаты были исключены на графике SSC-H по SSC-W, верхняя правая панель (P3). Клетки, положительные по всем трем выходам, были отсортированы в планшеты ELISPOT. (B) Для выделения плазмабластов, определенных как MHCII-intermediate (с промежуточным содержанием MHCII), B220-intermediate (с промежуточным содержанием B220), клетки, положительные по PI (пропидиум иодид), или мертвые клетки были исключены на нижней левой панели (не P1). Затем плазмабласты были отсортированы как MHCII-intermediate, B220-intermediate, нижняя правая панель (P2). Дубликаты были исключены на графике SSC-H по SSC-W, верхняя правая панель (P3), и, наконец, клетки были отсортированы по размеру на верхней левой панели (P4). Клетки, позитивные по всем четырем проходам, были отсортированы в планшеты ELISPOT.
Фиг.4. Сортинг мышиных спленоцитов. Прежде всего, PI-положительные или мертвые клетки были исключены на нижней левой панели (P1). На верхней правой точечной диаграмме отображены CD138 PE и CD43 FITC. Для фенотипически различных клеточных популяций были установлены четыре прохода: P2 означает CD138 intermediate, CD43 high. P3 означает CD138 high, CD43 high. P4 означает CD138 high, CD43 отрицательные. P5 означает CD138 intermediate, CD43 low. 10000 клеток, положительных по P1 и каждому из четырех проходов, были отсортированы в пробирки и заморожены для оценки в симплексной системе.
Фиг.5. Гель-электрофорез продуктов PCR, полученных в симплексной PCR при титровании клеточного лизата из 4 отсортированных фракций. P2, P3, P4 и P5 представляют собой проходы, отсортированные в соответствии с фиг.4. M - маркеры молекулярного веса.
Фиг.6. Схематическое представление вектора экспрессии 00-VP-002 полноразмерного антитела млекопитающего. Amp и Amp pro - ген устойчивости к ампициллину и его промотор, pUC origin - начало репликации pUC, CMV - промотор млекопитающих, управляющий экспрессией легкой цепи и тяжелой цепи, IGHV Leader - геномный лидер тяжелой цепи человека, H stuffer - вставка, которая заменена на кодирующую последовательность вариабельной области тяжелой цепи, IGHG1 - последовательность, кодирующая константную область тяжелой цепи изотипа G1 геномного иммуноглобулина (последовательность показана в приложении 1), Rabbit B-globin A - полиА последовательность бета-глобина кролика, IGKV Leader - мышиный каппа-лидер, L Stuffer - вставка, которая заменена на кодирующую последовательность легкой цепи, SV40 term - терминаторная последовательность вакуолизирующего обезьяньего вируса 40, FRT - целевой сайт распознавания FIp, Neo - ген устойчивости к неомицину, SV40 poly A - полиА сигнальная последовательность вакуолизирующего обезьяньего вируса 40.
Фиг.7. Анализ репертуара химерных антител anti-hEGFR. Кластерный анализ различий в поглощении на уровне 450-620 nm. Супернатанты сгруппированы по реактивности, на что указывают цифры (от 1 до 4) после номера клона. Темно-серый цвет указывает на уменьшение количества метаболически активных клеток, тогда как светло-серый цвет указывает на увеличение количества метаболически активных клеток. Черный цвет указывает на супернатанты, не оказывающие влияния на количество метаболически активных клеток.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение представляет дальнейшие возможности применения способа амплификации и сцепления, раскрытого в документе WO 2005/042774, для обеспечения коллекций векторов антител от животных, отличных от человека. Эти усовершенствования позволяют клонировать кодирующие последовательности химерных антител человека/не являющегося человеком с когнатными парами вариабельных областей для их подгонки к высокопроизводительному формату. По существу это достигается посредством доставки нового стартового материала для процессов амплификации и сцепления и обеспечения способов создания библиотек химерных антител человека/не являющегося человеком с когнатными парами вариабельных областей.
Одним из аспектов изобретения является способ сцепления вариабельных последовательностей тяжелой и легкой цепи, включающий амплификацию в мультиплексной процедуре молекулярной амплификации важных нуклеотидных последовательностей с использованием матрицы, полученной из выделенной единичной клетки, популяции изогенных клеток или популяции генетически различных клеток, и осуществление последующего сцепления амплифицированных последовательностей.
ОПРЕДЕЛЕНИЯ
Термин "когнатная пара" описывает исходную пару представляющих интерес несмежных нуклеиновых кислот, которые содержатся в единичной клетке или извлечены из единичной клетки. В предпочтительных вариантах осуществления изобретения когнатная пара содержит две кодирующие последовательности вариабельных областей, которые совместно кодируют вариабельный домен связывающего белка, причем их генные последовательности получены из одной и той же клетки. Таким образом, при экспрессии в виде полного связывающего белка или его стабильного фрагмента они сохраняют аффинность связывания и специфичность связывающего белка, изначально экспрессируемого из этой клетки. Например, когнатная пара может состоять из кодирующей последовательности вариабельной тяжелой цепи антитела, связанной с кодирующей последовательностью вариабельной легкой цепи из той же самой клетки, или из кодирующей последовательности α-цепи рецептора T-клетки, связанной с кодирующей последовательностью β-цепи из той же самой клетки. Библиотека когнатных пар представляет собой коллекцию таких когнатных пар. Термин "полимераза горячего запуска" описывает полимеразы, которые неактивны или имеют очень низкую активность при температурах, используемых для обратной транскрипции. Для того чтобы стать функциональными, такие полимеразы нуждаются в активации высокими температурами (от 90 до 95°C). Это, например, является преимуществом в процедурах одностадийной RT-PCR, поскольку исключает вмешательство полимеразы в реакцию обратной транскриптазы.
Термин "изогенная популяция клеток" описывает популяцию генетически идентичных клеток. Особый интерес в настоящем изобретении представляет изогенная популяция клеток, полученная посредством клональной экспансии выделенной единичной клетки.
Термин "выделенная единичная клетка" описывает клетку, которая была физически отделена от популяции клеток, то есть соответствует определению "единичная клетка в отдельном резервуаре". При распределении популяции клеток поодиночке во множество резервуаров получают популяцию выделенных клеток. Как указано в разделе, озаглавленном "источники матриц", для того чтобы можно было применять название совокупность единичных клеток, доля резервуаров с единичной клеткой не обязательно должна составлять 100%.
Термины "связка" или "сцепление" применительно к амплификации описывают ассоциацию амплифицированных последовательностей нуклеиновой кислоты, которая составляет представляющие интерес последовательности нуклеиновой кислоты, закодированные в одном сегменте. Применительно к когнатным парам сегмент содержит последовательности нуклеиновой кислоты, кодирующие вариабельный домен, например вариабельную область тяжелой цепи антитела, связанный с кодирующей последовательностью вариабельной области легкой цепи антитела, полученной из той же клетки. Сцепление может быть достигнуто либо одновременно с амплификацией, либо представляет собой стадию, непосредственно следующую за амплификацией. Не существует каких-либо требований к форме или функциональности сегмента: он может быть линейным, кольцевым, одноцепочечным или двухцепочечным. Сцепление также не обязательно должно быть перманентным: одну из представляющих интерес последовательностей нуклеиновой кислоты можно по желанию выделить из сегмента, например одну из кодирующих последовательностей вариабельной области можно выделить из сегмента когнатной пары. Однако до тех пор, пока исходные вариабельные области, составляющие когнатную пару, не перемешаны с другими вариабельными областями, они все еще считаются когнатной парой, хотя и не сцеплены в единый сегмент. Предпочтительное сцепление представляет собой фосфодиэфирное сцепление нуклеотидов. Однако сцепление можно также получить посредством других химических процедур перекрестного соединения.
Термин "мультиплексная молекулярная амплификация" описывает одновременную амплификацию двух или более целевых последовательностей в одной и той же реакции. Пригодные способы амплификации включают полимеразную цепную реакцию (PCR) (патент США 4683202), лигазную цепную реакцию (LCR), (Wu and Wallace, 1989, Genomics 4, 560-9), методику амплификации с замещением цепи (SDA) (Walker et al., 1992, Nucl. Acids Res. 20, 1691-6), самоподдерживающуюся амплификацию последовательности (Guatelli et al., 1990, Proc. Nat. Acad. Sci. U.S.A., 87, 1874-8) и амплификацию на основе последовательностей нуклеиновых кислот (NASBA) (Compton J., 1991, Nature 350, 91-2). Два последних способа амплификации включают изотермические реакции, основанные на изотермической транскрипции, которая продуцирует как одноцепочную РНК (ssRNA), так и двуцепочную ДНК (dsDNA).
Термин "мультиплексная PCR" описывает вариант PCR, в котором одновременно амплифицируются две или более целевые последовательности, причем в одной и той же реакции используется более одного набора праймеров, например один набор праймеров адаптирован для амплификации вариабельной области тяжелой цепи, а другой набор праймеров адаптирован для амплификации вариабельной области каппа-цепи в той же реакции PCR. Дополнительно или альтернативно набор праймеров, адаптированный для амплификации вариабельной области лямбда-цепи, может быть скомбинирован с другими наборами праймеров.
Термин "мультиплексная RT-PCR" описывает мультиплексную реакцию PCR, которая предваряется стадией обратной транскрипции (RT). Мультиплексная RT-PCR может быть проведена либо как двухстадийный процесс с отдельной стадией RT перед мультиплексной PCR, либо как одностадийный процесс, в котором все компоненты как RT, так и мультиплексной PCR комбинируются в одной пробирке. Термины "мультиплексная PCR перекрывания-удлинения" и "мультиплексная RT-PCR перекрывания-удлинения" означают, что мультиплексная PCR или мультиплексная RT-PCR проводится с применением смеси праймеров перекрывания-удлинения для амплификации целевых последовательностей, то есть для одновременного достижения амплификации и сцепления целевых последовательностей.
Термин "множество резервуаров" описывает любой объект (или коллекцию объектов), который позволяет осуществить физическое отделение единичной клетки от популяции клеток. Это могут быть пробирки, многолуночные планшеты (например, 96-луночные, 384-луночные, титрационные микропланшеты или другие многолуночные планшеты), матрицы, микроматрицы, микрочипы, гели или гелевые матрицы. Предпочтительно, чтобы объект был применим для амплификации в PCR.
Термин "поликлональный белок" или "поликлональность" при использовании здесь относится к белковой композиции, содержащей разные, но гомологичные молекулы белка, предпочтительно выбранные из суперсемейства иммуноглобулинов. Таким образом, каждая молекула белка гомологична другим молекулам композиции, но также содержит один или более участков вариабельной последовательности полипептида, которые характеризуются различиями в аминокислотной последовательности между индивидуальными элементами поликлонального белка. Известные примеры таких поликлональных белков включают молекулы антитела или иммуноглобулина, рецепторы T-клеток и рецепторы B-клеток. Поликлональный белок может состоять из определенной подгруппы молекул белка, которые были охарактеризованы таким общим признаком, как совпадающая активность связывания с желательной целью, например, с поликлональным антителом, проявляющим специфичность связывания по отношению к желательному целевому антигену.
Термин "популяция генетически различных клеток" при использовании здесь относится к такой клеточной популяции, в которой клетки, входящие в состав популяции, отличаются друг от друга на геномном уровне. Такая популяция генетически различных клеток представляет собой, например, популяцию клеток, полученных от донора, или фракцию таких клеток, например клеточную фракцию, содержащую В-лимфоциты или T-лимфоциты.
Термин "набор праймеров" употребляется взаимозаменяемо с термином "пара праймеров" и описывает два или более праймеров, которые вместе способны служить затравкой амплификации представляющей интерес нуклеотидной последовательности (например, один элемент когнатной пары). Рассматриваемый в настоящем изобретении набор праймеров можно сконструировать для затравки амплификации семейства нуклеотидных последовательностей, содержащего кодирующие последовательности вариабельной области. Примерами разных семейств являются легкие каппа-цепи антител, легкие лямбда-цепи антител, вариабельные области тяжелой цепи, а также вариабельные области α, β, γ или δ рецепторов T-клеток. Набор праймеров для амплификации семейства нуклеотидных последовательностей, содержащего кодирующие последовательности вариабельных областей, часто состоит из множества праймеров, в котором некоторые праймеры могут быть вырожденными праймерами.
Термин "идентичность последовательностей" означает процентный показатель, указывающий на степень идентичности между последовательностями нуклеиновой кислоты, исходя из длины более короткой из двух последовательностей. Идентичность можно рассчитать по формуле (Nref-Ndif)×100%/Nref, где Nref означает число остатков в более короткой из двух последовательностей, а Ndif - общее число неидентичных остатков в Nref при оптимальной выверке совпадения по длине между двумя последовательностями.
Таким образом, последовательность ДНК AGTCAGTC (Seq. no. 32) будет иметь идентичность 75% с последовательностью TAATCAATCGG (Seq. no. 33) (Ndif=2 и Nref=8) (подчеркивание показывает оптимальную выверку, а жирный шрифт выделяет два неидентичных остатка из 8).
Термины "случайно" или "случайный" применительно к сцеплению относятся к сцеплению нуклеотидных последовательностей, который не были выделены из одной и той же клетки, но поперечно сцеплены в популяции генетически различных клеток. Если представляющие интерес нуклеотидные последовательности являются кодирующими последовательностями вариабельных областей, то это приведет к комбинаторной библиотеке сцепленных последовательностей. C другой стороны, если представляющие интерес нуклеотидные последовательности кодируют не имеющий разнообразия гетеромерный белок, то случайно сцепленные последовательности окажутся подобными сцепленным последовательностям из единичной клетки.
Термин "матрица, полученная из выделенной единичной клетки", применительно к обратной транскрипции, относится к нуклеиновым кислотам, находящимся внутри такой выделенной клетки. Например, нуклеиновые кислоты могут быть в виде РНК, мРНК, ДНК или геномной ДНК. Нуклеиновые кислоты либо могут быть выделенными из клетки, либо по-прежнему сохранять связь с остальным содержимым клетки, причем клетка может быть как в интактном виде, так и в лизированном виде.
Термин "CD43" относится к поверхностному клеточному антигену мыши, известному под многими синонимическими названиями, включая антиген 3E8, A630014B01 Rik, антиген дифференциации B-клеток LP-3, Cd43, антиген CD43, Galgp, лейкоцитарный сиалогликопротеин, лейкосиалин, предшественник лейкосиалина, Ly48, Ly-48, сиалофорин, а также к ортологичным поверхностным маркерам клеток других животных.
Термин "CD138" относится к поверхностному клеточному антигену мыши, известному под многими синонимическими названиями, включая Syndecan-1, AA408134, AA409076, CD138, syn-1, Synd, Synd1, SYND1, Synd-1, предшественник Syndecan-1, а также к ортологичным поверхностным маркерам клеток других животных.
Термин "MHCII" относится к поверхностному клеточному антигену мыши, известному под многими синонимическими названиями, включая антиген CD74, CLIP, DHLAG, гамма-цепь антигена гистосовместимости класса H-2 II, HLADG, HLA-DR-GAMMA, инвариантная цепь, ассоциированная с антигеном Ia, Ia-GAMMA, Ii, инвариантная цепь, ассоциированная с MHC класса II, а также к ортологичным поверхностным маркерам клеток других животных. Термин "B220" относится к поверхностному клеточному антигену мыши, известному под многими синонимическими названиями, включая Cd45, CD45, антиген CD45, CD45R, L-CA, общий предшественник лейкоцитарного антигена, loc, Ly-5, общий лимфоцитарный антиген Ly-5, Lyt-4, T200, а также к ортологичным поверхностным маркерам клеток других животных.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Процесс амплификации и сцепления
Одной из особенностей настоящего изобретения является уменьшение числа пробирок, необходимых для амплификации представляющих интерес нуклеотидных последовательностей, с применением варианта PCR, в котором две или более целевые последовательности подвергаются одновременной амплификации в одной пробирке при участии более одного набора праймеров, например всех праймеров, необходимых для амплификации кодирующих последовательностей вариабельных областей, в одной реакции. В общем смысле этот подход известен как мультиплексная полимеразная цепная реакция (мультиплексная PCR).
Следующая особенность настоящего изобретения заключается в том, что две или более целевые последовательности, амплифицированные в мультиплексной PCR, становятся сцепленными в непосредственной близости в результате процесса амплификации. Особенно важно, что в этом процессе происходит сцепление кодирующих последовательностей вариабельных областей.
Один из вариантов осуществления настоящего изобретения использует то обстоятельство, что можно сконструировать мультиплексную смесь праймеров, работающую в процедуре PCR перекрывания-удлинения, что приводит к одновременной амплификации и сцеплению представляющих интерес нуклеотидных последовательностей. Эта методика мультиплексной PCR перекрывания-удлинения способствует уменьшению числа реакций, необходимых для выделения и сцепления представляющих интерес нуклеотидных последовательностей, в особенности когнатных пар сцепленных вариабельных областей. Другие варианты осуществления настоящего изобретения ориентированы на сцепление посредством лигирования или рекомбинации в качестве альтернативы сцеплению посредством мультиплексной PCR перекрывания-удлинения. В этих процедурах сцепление осуществляется не одновременно с мультиплексной PCR, а представляет собой самостоятельную стадию, непосредственно следующую за амплификацией. Однако сцепление по-прежнему можно осуществлять в той же пробирке, в которой проводилась мультиплексная PCR.
Мультиплексная PCR перекрывания-удлинения требует присутствия двух или более наборов праймеров (мультиплексной смеси праймеров), причем, по меньшей мере, один праймер в каждом наборе оснащен хвостом перекрывания-удлинения. Хвосты перекрывания-удлинения позволяют осуществить сцепление продуктов, генерированных каждым набором праймеров во время амплификации. Такую смесь праймеров называют мультиплексной смесью праймеров перекрывания-удлинения. Мультиплексная PCR перекрывания-удлинения отличается от обычной PCR перекрывания-удлинения тем, что последовательности, подлежащие сцеплению, генерируются одновременно в одной и той же пробирке, что обеспечивает немедленное сцепление целевых последовательностей в ходе амплификации без какой-либо промежуточной очистки.
Следующей особенностью настоящего изобретения является стадия обратной транскрипции (RT), предшествующий мультиплексной PCR или амплификации в мультиплексной PCR перекрывания-удлинения, с использованием матрицы, полученной из выделенной единичной клетки или популяции изогенных клеток.
Следующей особенностью настоящего изобретения является применение нуклеотидных последовательностей, происходящих из выделенной единичной клетки или популяции изогенных клеток, в качестве матрицы для амплификации в мультиплексной PCR. Предпочтительно, чтобы перед проведением мультиплексной PCR была осуществлена обратная транскрипция РНК в кДНК. Для амплификации некоторых представляющих интерес последовательностей нуклеиновых кислот в качестве альтернативы мРНК можно использовать геномную ДНК. Используя в качестве матрицы выделенные единичные клетки или популяцию изогенных клеток, полученную посредством клональной экспансии из выделенной единичной клетки, можно избежать перемешивания нуклеотидных последовательностей, кодирующих представляющий интерес гетеродимерный белок, с нуклеотидными последовательностями, извлеченными из разных клеток, входящих в популяцию клеток. Это важно, если задачей является получение начальной композиции представляющих интерес последовательностей. Особенно важной отличительной особенностью изобретения является применение выделенной единичной клетки или популяции изогенных клеток в качестве матрицы для генерирования когнатной пары кодирующих последовательностей вариабельных областей.
В дополнение к этому настоящее изобретение облегчает создание библиотек представляющих интерес сцепленных последовательностей нуклеиновой кислоты в виде специальных комбинаторных библиотек и библиотек когнатных пар вариабельных областей. Далее, настоящее изобретение использует нуклеиновые кислоты, извлеченные из единичных клеток, предпочтительно в виде РНК, не нуждающейся в отделении от остального содержимого клетки перед ее использованием в качестве матрицы.
Один из вариантов осуществления настоящего изобретения заключает в себе сцепление множества представляющих интерес несмежных нуклеотидных последовательностей. Этот способ включает амплификацию в мультиплексной PCR или в процедуре мультиплексной амплификации RT-PCR представляющих интерес нуклеотидных последовательностей с использованием матрицы, полученной из выделенной единичной клетки или популяции изогенных клеток, и осуществление сцепления представляющих интерес нуклеотидных последовательностей. Далее этот способ включает факультативную стадию проведения дополнительной амплификации сцепленных продуктов.
Следующий вариант осуществления настоящего изобретения представляет собой способ продуцирования библиотеки когнатных пар, содержащих кодирующие последовательности вариабельных областей. Этот способ включает получение от донора клеточной фракции, содержащей лимфоциты, причем эта фракция необязательно может быть обогащена особой популяцией лимфоцитов из указанной клеточной фракции, либо особая популяция лимфоцитов может быть выделена из указанной клеточной фракции. Далее, популяцию выделенных единичных клеток получают посредством распределения клеток из клеточной фракции, содержащей лимфоциты, или из обогащенной клеточной фракции индивидуально во множество резервуаров. Затем проводят мультиплексную молекулярную амплификацию (мультиплексную RT-PCR амплификацию) кодирующих последовательностей вариабельных областей, содержащихся в популяции выделенных единичных клеток и осуществляют сцепление пар кодирующих последовательностей вариабельных областей, причем индивидуальную пару последовательностей вариабельных областей извлекают из единичной клетки в пределах совокупности выделенных единичных клеток. Далее методика включает две необязательных стадии: на первой стадии отдельную клетку, выделенную из совокупности единичных клеток, размножают, создавая популяцию изогенных клеток, еще до проведения мультиплексной RT-PCR амплификации. Таким образом, можно получить множество резервуаров с разнообразной популяцией изогенных клеток (одна популяция изогенных клеток в одном резервуаре). Вторая необязательная стадия заключается в проведении дополнительной амплификации сцепленных кодирующих последовательностей вариабельных областей.
В предпочтительных вариантах осуществления настоящего изобретения отдельный элемент указанной библиотеки когнатных пар, состоящий из кодирующей последовательности вариабельной области легкой цепи иммуноглобулина, связан с кодирующей последовательностью вариабельной области тяжелой цепи иммуноглобулина, происходящей из той же клетки, или кодирующие последовательности связывающего домена рецептора T-клетки, состоящие из вариабельной области альфа-цепи, связаны с вариабельной областью бета-цепи или вариабельной областью гамма-цепи, связанным с вариабельной областью дельта-цепи, причем связанные вариабельные области происходят из одной и той же клетки.
Рассматриваемая в настоящем изобретении мультиплексная амплификация RT-PCR может быть проведена либо как двухстадийный процесс, когда обратная транскрипция (RT) проводится отдельно от мультиплексной амплификации PCR (или альтернативной мультиплексной молекулярной амплификации), либо как одностадийный процесс, когда стадии RT и мультиплексной амплификации PCR проводятся с одними и теми же праймерами в одной и той же пробирке.
Обратную транскрипцию (RT) проводят с ферментом, обладающим активностью обратной транскриптазы, что приводит к получению кДНК из совокупной РНК, мРНК или специфической целевой РНК из выделенной единичной клетки. Праймерами, которые можно использовать для обратной транскрипции, являются, например, олиго-dT праймеры, случайные гексамеры, случайные декамеры, другие случайные праймеры или праймеры, которые специфичны для представляющих интерес нуклеотидных последовательностей.
Двухстадийная процедура мультиплексной амплификации RT-PCR позволяет распределить кДНК, полученную на стадии RT, более чем в один резервуар, что создает возможность для хранения фракции матрицы перед проведением амплификации. Кроме того, распределение кДНК более чем в один резервуар позволяет провести несколько мультиплексных PCR с амплификацией нуклеиновой кислоты, извлеченной из одной и той же матрицы. Хотя при этом число отдельных реакций увеличивается, в то же время открывается возможность для уменьшения сложности мультиплексной смеси праймеров, если это желательно. Такой двухстадийный подход можно использовать, например, для амплификации и сцепления кодирующих последовательностей вариабельной области тяжелой цепи и вариабельной области каппа-цепи в одной пробирке, а также кодирующих последовательностей вариабельной области тяжелой цепи и вариабельной области лямбда легкой цепи в другой пробирке с использованием одной и той же матрицы. Единичная клетка обычно экспрессирует только одну из легких цепей. Однако часто бывает проще провести реакции одновременно, а не ждать результата одной реакции прежде, чем начать другую. Кроме того, амплификация как каппа, так и лямбда служит внутренним отрицательным контролем, поскольку ожидается, что из единичной клетки амплифицируются только каппа или лямбда.
В одностадийной мультиплексной процедуре RT-PCR обратную транскрипцию и мультиплексную амплификацию PCR выполняют в одном и том же резервуаре. Чтобы провести реакцию в одну стадию, все компоненты, необходимые для проведения обратной транскрипции и мультиплексной PCR, добавляют в резервуар с самого начала. Если реакция началась, то, как правило, не возникает необходимости во введении дополнительных компонентов. Преимущество одностадийной мультиплексной амплификации RT-PCR заключается в том, что она еще в большей степени уменьшает число стадий, необходимых для получения рассматриваемых в настоящем изобретении сцепленных нуклеотидных последовательностей. Это особенно полезно тогда, когда мультиплексную RT-PCR проводят с множеством единичных клеток, то есть одну и ту же реакцию необходимо выполнить во многих резервуарах. Одностадийную мультиплексную RT-PCR проводят, используя обратные праймеры, представленные в мультиплексной смеси праймеров, необходимых в мультиплексной амплификации PCR также в качестве праймеров для обратной транскрипции. Обычно композиция, необходимая для одностадийной мультиплексной RT-PCR, включает матрицу нуклеиновой кислоты, фермент с активностью обратной транскриптазы, фермент с активностью ДНК полимеразы, смесь деоксинуклеозидтрифосфатов (смесь dNTP, содержащая dATP, dCTP, dGTP и dTTP) и мультиплексную смесь праймеров. Предпочтительной матрицей нуклеиновой кислоты является совокупная РНК или мРНК, извлеченная из выделенной единичной клетки или в очищенном виде, или в виде лизата клетки или все еще находящаяся внутри интактной клетки. Обычно для точности композиции реакционной смеси требуется некоторая оптимизация каждой мультиплексной смеси праймеров, используемых в настоящем изобретении. Это применимо как для двухстадийной, так и для одностадийной процедуры мультиплексной RT-PCR.
Для некоторых одностадийных реакций мультиплексной RT-PCR может оказаться выгодным введение дополнительных компонентов во время реакции. Например, это может быть добавление полимеразы после стадии RT. Другими компонентами могут быть, например, смесь dNTP или мультиплексная смесь праймеров, возможно, с другим составом праймеров. В таком случае это можно считать мультиплексной RT-PCR в одной пробирке, которая, в общем, имеет те же преимущества, что и одностадийная мультиплексная RT-PCR, поскольку также уменьшает число пробирок, необходимых для получения желаемых сцепленных продуктов.
Представляющие интерес нуклеотидные последовательности, амплифицированные в мультиплексной RT-PCR, могут быть сцеплены друг с другом различными способами, например мультиплексной RT-PCR перекрывания-удлинения, лигированием или рекомбинацией с применением разных мультиплексных смесей праймеров. Предпочтительно, чтобы мультиплексная амплификация RT-PCR и сцепление были осуществлены в одностадийном или двухстадийном процессе. Однако процесс сцепления может быть также проведен как многостадийный процесс с применением, например, вставочного фрагмента для сцепления представляющих интерес последовательностей нуклеиновой кислоты посредством PCR, сшивания или рекомбинации. Такой вставочный фрагмент может содержать цис-элементы, промоторные элементы, или релевантную кодирующую последовательность, или последовательность распознавания. В предпочтительном варианте осуществления изобретения процесс сцепления осуществляется в том же резервуаре, что и мультиплексная амплификация RT-PCR.
В одном из вариантов осуществления настоящего изобретения сцепление множества представляющих интерес несмежных нуклеотидных последовательностей проводят в сочетании с мультиплексной амплификацией PCR, используя мультиплексную смесь праймеров перекрывания-удлинения. Это приводит к комбинированию амплификации и сцепления целевых последовательностей. Обычно композиция, необходимая для мультиплексной PCR перекрыванием-удлинением, включает матрицу нуклеиновой кислоты, фермент с активностью ДНК-полимеразы, смесь деоксинуклеозидтрифосфатов (смесь dNTP, содержащая dATP, dCTP, dGTP и dTTP) и мультиплексную смесь праймеров перекрывания-удлинения.
В особом варианте осуществления настоящего изобретения сцепление множества представляющих интерес несмежных нуклеотидных последовательностей проводят посредством мультиплексной RT-PCR перекрывания-удлинения с применением матрицы, извлеченной из выделенной единичной клетки или популяции изогенных клеток. Далее этот способ включает необязательную стадию проведения дополнительной молекулярной амплификации сцепленных продуктов. Предпочтительно, чтобы мультиплексная RT-PCR перекрывания-удлинения была проведена как одностадийная/однопробирочная реакция.
Рассматриваемая в настоящем изобретении мультиплексная смесь праймеров перекрывания-удлинения содержит, по меньшей мере, два набора праймеров, способных служить затравкой для амплификации и сцепления, по меньшей мере, двух кодирующих последовательностей вариабельных областей, например амплификацию и сцепление последовательностей из семейств вариабельной области тяжелой цепи иммуноглобулинов с семействами вариабельной области легких цепей каппа или лямбда или амплификацию и сцепление последовательностей из семейства рецепторов T-клеток α, β, γ или δ.
Еще в одном варианте осуществления настоящего изобретения множество представляющих интерес нуклеотидных последовательностей, амплифицированных посредством мультиплексной RT-PCR, сцепляются посредством сшивания. Для того чтобы достичь этого, мультиплексную смесь праймеров, используемую для мультиплексной RT-PCR, конструируют так, чтобы соответствующими рестрикционными ферментами можно было расщепить амплифицированные целевые последовательности и провести ковалентное сцепление посредством сшивания ДНК (конструирование праймеров описано в разделе "Смеси и конструирование праймеров"). После амплификации в мультиплексной RT-PCR с такой мультиплексной смесью праймеров к смеси наряду с лигазой добавляют рестрикционные ферменты, необходимые для формирования у целевых последовательностей совместимых концов. Какая-либо очистка продуктов PCR перед этой стадией не требуется, хотя такую очистку можно провести. Температура реакции для комбинированного рестрикционного расщепления и сшивания варьируется приблизительно от 0 до 40°C. Однако если в смеси еще присутствует полимераза из мультиплексной PCR, то предпочтительна температура инкубации ниже комнатной и наиболее предпочтительна температура от 4 до 16°C.
Еще в одном варианте осуществления настоящего изобретения множество представляющих интерес нуклеотидных последовательностей, амплифицированных посредством мультиплексной RT-PCR, сцепляются посредством рекомбинации. При таком подходе амплифицированные целевые последовательности можно соединить при помощи идентичных сайтов рекомбинации. Затем проводят сцепление, добавляя рекомбиназы, облегчающие рекомбинацию. Некоторыми подходящими системами рекомбиназ являются рекомбиназа Flp с множеством сайтов FRT, рекомбиназа Cre с множеством сайтов lox, интеграза ΦC31, которая выполняет рекомбинацию между сайтом attP и сайтом attB, система β-рекомбиназы-шесть, а также система Gin-gix. Сцепление посредством рекомбинации было проиллюстрировано для двух нуклеотидных последовательностей (VH в сцеплении с VL) (Chapal, N. et al. 1997 BioTechniques 23, 518-524).
В предпочтительном варианте осуществления настоящего изобретения представляющие интерес нуклеотидные последовательности содержат кодирующие последовательности вариабельной области, а сцепление создает когнатную пару кодирующих последовательностей вариабельных областей. Такая когнатная пара может в дополнение к вариабельным областям содержать одну или более кодирующих последовательностей константных областей. Предпочтительно, чтобы константные области имели происхождение от человека, а когнатная пара вариабельных областей имела иное происхождение, например от мыши, крысы или кролика.
В еще более предпочтительном варианте осуществления настоящего изобретения представляющие интерес нуклеотидные последовательности содержат кодирующие последовательности вариабельных областей иммуноглобулина, а сцепление создает когнатную пару кодирующих последовательностей вариабельной области легкой цепи и вариабельной области тяжелой цепи. Такая когнатная пара может в дополнение к вариабельным областям содержать одну или более кодирующих последовательностей константных областей. Затем такую когнатную пару можно выделить из шаблона, который был извлечен из клеток обогащенной B-лимфоцитарной линии из фракции, содержащей лимфоциты, например из цельной крови, из мононуклеарных клеток или из лейкоцитов.
Еще в одном варианте осуществления настоящего изобретения представляющие интерес нуклеотидные последовательности содержат кодирующие последовательности вариабельной области TcR, а сцепление создает когнатную пару кодирующих последовательностей вариабельной области α-цепи и вариабельной области β-цепи или кодирующих последовательностей вариабельной области γ-цепи и вариабельной области δ-цепи. Такая когнатная пара может в дополнение к вариабельным областям содержать одну или более кодирующих последовательностей константной области. Затем такую когнатную пару можно выделить из шаблона, который был извлечен из клеток обогащенной T-лимфоцитарной линии из фракции, содержащей лимфоциты, например, из цельной крови, из мононуклеарных клеток или из лейкоцитов.
Еще один аспект настоящего изобретения заключается в использовании мультиплексной RT-PCR с популяцией генетически различных клеток в качестве источника шаблона. Большинство кодирующих последовательностей гетеромерного белка не варьируются от клетки к клетке, как это происходит в случае кодирующих последовательностей вариабельной области связывающих белков. Таким образом, применяя подход, предлагаемый настоящим изобретением, для клонирования таких не изменяющихся кодирующих последовательностей гетеромерного белка, мы избавлены от необходимости изначального выделения единичных клеток.
В этом варианте осуществления настоящего изобретения множество представляющих интерес несмежных нуклеотидных последовательностей сцепляются случайным образом, причем способ сцепления включает проведение мультиплексной RT-PCR для амплификации представляющих интерес нуклеотидных последовательностей с применением шаблона, извлеченного из популяции генетически различных клеток, и результирующее сцепление представляющих интерес амплифицированных нуклеотидных последовательностей. Далее этот способ включает факультативную стадию проведения дополнительной амплификации сцепленных продуктов. Как и при подходе с единичной клеткой, сцепление может быть осуществлено с применением мультиплексной смеси праймеров перекрывания-удлинения для амплификации или альтернативными способами сшивания или рекомбинации. Предпочтительно, чтобы шаблон, извлеченный из популяции клеток, не был строго заключен в клетках. Например, популяция клеток может быть лизирована.
Применение процесса случайного сцепления к популяции клеток, экспрессирующих вариантные связывающие белки, позволяет упростить генерирование комбинаторных библиотек кодирующих последовательностей вариабельной области. Предпочтительно, чтобы клеточная популяция состояла из таких клеток, которые экспрессируют связывающие белки вариабельной области, например из B-лимфоцитов, T-лимфоцитов, клеток гибридомы, плазмацитов, плазмабластов или из смеси этих клеток.
В вышеуказанном варианте осуществления изобретения можно, например, сделать проницаемой или лизировать популяцию клеток, не прибегая к дополнительной очистке, либо можно выделить из клеток шаблонные нуклеиновые кислоты, применяя стандартные процедуры. Предпочтительна одностадийная процедура мультиплексной RT-PCR. Однако в виде варианта осуществления изобретения возможно применение и двухстадийной процедуры.
Эффективный способ увеличить специфичность, чувствительность и выход мультиплексного процесса RT-PCR-сцепления заключается в проведении дополнительной молекулярной амплификации сцепленных нуклеотидных последовательностей, полученных в мультиплексной RT-PCR с последующим сцеплением посредством сшивания, рекомбинации или на основе мультиплексной RT-PCR перекрывания-удлинения. В предпочтительном варианте эту дополнительную амплификацию проводят через PCR, используя смесь праймеров, адаптированную для амплифицирования представляющих интерес сцепленных нуклеотидных последовательностей. Используемой смесью праймеров могут быть внешние праймеры мультиплексной смеси праймеров или мультиплексная смесь праймеров перекрывания-удлинения, а это означает, что праймеры, которые отжигаются с самым удаленным 5'-концом и 3'-концом смысловой нити сцепленных кодирующих последовательностей вариабельной области, тем самым делают возможной амплификацию всего сцепленного продукта. Внешние праймеры также могут быть охарактеризованы как те праймеры в мультиплексной смеси праймеров перекрывания-удлинения, которые не содержат хвостов перекрывания-удлинения. В альтернативном варианте для дополнительной амплификации сцепленных нуклеотидных последовательностей можно применить вложенный или полувложенный набор праймеров. Такая вложенная PCR особенно полезна для повышения специфичности способа, а также для увеличения количества сцепленного продукта. В настоящем изобретении полувложенная PCR (как это описано в разделе, озаглавленном Смеси и конструирование праймеров) считается функциональной так же, как и вложенная PCR.
Так образом, в настоящем изобретении желательно, хотя и не обязательно, проводить дополнительную PCR-амплификацию сцепленных продуктов из мультиплексной RT-PCR перекрывания-удлинения или продуктов, сцепленных посредством сшивания или рекомбинации, предпочтительно с использованием вложенной PCR или полувложенной PCR. Дополнительную амплификацию можно провести либо напрямую, используя частично или полностью продукт реакции из мультиплексной RT-PCR перекрывания-удлинения, или продукт сшивания, или продукт рекомбинации, или фракцию любого из этих продуктов, либо использовать частично очищенные сцепленные продукты любой из указанных реакций, например, проведя электрофорез сцепленных продуктов в агарозном геле и вырезав фрагмент, соответствующий ожидаемому размеру сцепленных кодирующих последовательностей вариабельной области. Для продуктов, сцепленных посредством мультиплексной RT-PCR перекрывания-удлинения, предпочтительно проводить дополнительную амплификацию непосредственно на фракции из мультиплексной реакции RT-PCR перекрывания-удлинения, поскольку это должно помочь сцеплению отдельных целевых последовательностей, которые на были сцеплены в первой реакции.
ПРЕДСТАВЛЯЮЩИЕ ИНТЕРЕС ПОСЛЕДОВАТЕЛЬНОСТИ
В настоящем изобретении представляющие интерес последовательности можно выбрать из последовательностей, которые кодируют разные субъединицы или домены, которые при экспрессировании образуют белок или часть белка. Такие белки, состоящие, по меньшей мере, из двух неидентичных субъединиц, известны как гетеродимерные белки. Гетеродимерные белки распространены во всех биологических родах и видах. Некоторыми из классов веществ, к которым относятся такие белки, являются, например, ферменты, ингибиторы, структурные белки, белки суперсемейства иммуноглобулинов, транспортные белки и т.д. Нуклеотидные последовательности, кодирующие такие гетеродимерные белки, несмежны, а это означает, например, что они происходят из разных генов или разных молекул мРНК. Однако термин несмежные при его использовании в настоящем изобретении может также означать нуклеотидные последовательности, кодирующие домены одного и того же белка, если эти домены разделены нуклеотидными последовательностями, не представляющими интереса.
В одном из вариантов осуществления настоящего изобретения представляющие интерес нуклеотидные последовательности содержат кодирующие последовательности вариабельной области из суперсемейства иммуноглобулинов, например вариабельной области иммуноглобулинов (антител), рецепторов B-клеток и рецепторов T-клеток (TcR). Особый интерес представляют кодирующие последовательности вариабельной области иммуноглобулинов. Такие кодирующие последовательности вариабельной области включают кодирующие последовательности полноразмерных антител, а также Fab, Fv, scF' и комбинации фрагментов кодирующих последовательностей вариабельной области, например гипервариабельных участков (CDR), стыковочных генов, V-ген или их комбинаций. В общем, настоящее изобретение можно использовать с любыми комбинациями кодирующих последовательностей вариабельной области и их фрагментов. Настоящее изобретение делает возможным только сцепление вариабельных доменов тяжелых и легких цепей, генерирующих кодирующие последовательности Fv или scFv. Или сцепление всей легкой цепи с вариабельной областью тяжелой цепи + доменом константной области CH1 + частью шарнирной области, порождающее Fab, Fab' или F(ab)2. Кроме того, можно добавить к вариабельной тяжелой цепи любой участок из доменов константной области тяжелой цепи, посредством чего будут генерированы кодирующие последовательности полноразмерного антитела или кодирующие последовательности усеченного антитела. В одном из аспектов изобретения вариабельные последовательности, отличные от человеческих, сцепляются с константными областями человека для генерирования полных химерных антител человека/неродственного животного, предпочтительно химерных антител с константными областями человека. В следующем варианте осуществления настоящего изобретения кодирующие последовательности вариабельной области содержат один тип кодирующей последовательности легкой цепи иммуноглобулина (каппа или лямбда) и одну кодирующую последовательность вариабельной области тяжелой цепи иммуноглобулина. Это достигается за счет отбора праймеров, амплифицирующих только один изотип легкой и тяжелой цепи. Изотип также можно определить посредством сцепления или сплайсинга константных областей человека из одного или более специфических изотипов тяжелой и легкой цепи.
Также представляют интерес кодирующие последовательности вариабельной области, извлеченные из рецепторов T-клеток (TcR). Такие кодирующие последовательности TcR содержат кодирующие последовательности полноразмерных альфа-цепей и бета-цепей или гамма-цепей и дельта-цепей, а также растворимые TcR либо только вариабельные домены этих цепей или их одноцепочечные гибридные белки (например, одиночную цепь αβ или одиночную цепь γδ).
ИСТОЧНИКИ ШАБЛОНОВ
Одной из особенностей настоящего изобретения является способность осуществлять сцепление нуклеотидных последовательностей, извлеченных из выделенной единичной клетки, популяции изогенных клеток или популяции генетически различных клеток, которые не были разнесены в отдельные резервуары.
Предпочтительной особенностью настоящего изобретения является применение выделенных единичных клеток или популяции изогенных клеток в качестве источника шаблона, поскольку в этом случае можно избежать перемешивания представляющих интерес последовательностей нуклеиновых кислот, в особенности кодирующих последовательностей вариабельной области. Это важно в том случае, если желательно получить исходную пару кодирующих последовательностей, например, вариабельной области.
Еще одной предпочтительной особенностью настоящего изобретения является получение единичной клетки или совокупности единичных клеток из клеточной фракции, содержащей лимфоциты, в частности B-лимфоциты, T-лимфоциты, плазмациты и/или клетки из этих линий на разных стадиях развития. Для получения единичных клеток также можно использовать другие клеточные популяции, которые экспрессируют связывающие белки из суперсемейства иммуноглобулинов. В настоящем изобретении также применимы такие клеточные линии, как клетки гибридомы, линии B-лимфоцитов и T-лимфоцитов, клетки, иммортализованные вирусом, или донорские клетки, участвующие в иммунном ответе. Донорские клеточные фракции, содержащие лимфоциты, могут быть получены из природной ткани или жидкости, которые богаты такими клетками, например из крови, костного мозга, лимфатических узлов, ткани селезенки, ткани миндалин или из инфильтратов в опухолях, вокруг опухолей и инфильтратов в воспалительной ткани. В случае животных, не являющихся человеком, предпочтительно использовать ткань селезенки или костный мозг. В отношении желательной цели доноры могут быть интактными или гипериммунными. Для выделения антигенсвязывающих белков со специфичностью связывания, направленной на желательную цель, предпочтительны гипериммунные доноры. Такими гипериммунными донорами могут быть либо доноры, иммунизированные целевым объектом или его фрагментами, либо выздоравливающие больные, либо пораженные заболеванием лица, которые проявляют иммунный ответ, направленный против целевого объекта, например лица с аутоиммунными заболеваниями, раковые больные, больные с инфекционными заболеваниями, например больные с HIV-инфекцией, гепатитом A, B или C, больные с SARS и т.д., а также больные с хроническими заболеваниями. Однако в особенно предпочтительном варианте осуществления изобретения донором, не являющимся человеком, является животное, которое было иммунизировано аутоантигеном человека, например таким белком человека, вовлеченным в раковый процесс, как EGFR.
Для применения в настоящем изобретении доноры клеток могут принадлежать к тому же биологическому виду, что и субъект, подлежащий лечению продуктами, получаемыми из сцепленных нуклеотидных последовательностей, рассматриваемых настоящим изобретением. Предпочтительно, чтобы донором клеток были сельскохозяйственное животное, домашнее животное или человек. Специальная отличительная особенность настоящего изобретения заключается в том, что оно позволяет создавать библиотеки химерных антител человека/не являющимся человеком, животных с целью генерирования химерных антител, пригодных для лечебного применения на человеке. Такой подход предпочтителен, когда антитела направлены против так называемых аутоантигенов, то есть антигенов человека.
Донором также может быть трансгенное животное, в особенности трансгенная мышь. Трансгенные животные, несущие локусы иммуноглобулинов человека, описаны в патенте США № 6111166 и в ссылке Kuroiwa Y. et al. Nature Biotechnology; 2002; 20: 889-893. Такие трансгенные животные способны вырабатывать иммуноглобулины человека. Таким образом, посредством обычных методик иммунизации указанных трансгенных животных можно создать полностью человеческое антитело против специфического целевого объекта. Это позволяет создавать библиотеки кодирования связывающих белков со специфичностями, направленными на более трудные целевые объекты, например антигены человека, к которым не существует природного ответа антител человека или такой ответ недостаточен. Например, такие трансгенные животные могут быть выведены с целью выработки рецепторов T-клеток человека.
В следующем варианте осуществления настоящего изобретения клеточная фракция, содержащая лимфоциты, может быть представлена цельной кровью, костным мозгом, мононуклеарными клетками или лейкоцитами, полученными от донора. Мононуклеарные клетки можно выделять из крови, костного мозга, лимфатических узлов, селезенки, инфильтратов вокруг раковых клеток и воспалительных инфильтратов. Мононуклеарные клетки можно выделять, применяя методики центрифугирования в градиенте плотности, например в градиенте среды Фиколл. Если мононуклеарные клетки выделяют из тканевых образцов, то ткань измельчают перед центрифугированием в градиенте. Измельчение можно провести, например, такими механическими способами, как помол, электропорация, и/или такими химическими способами, как ферментативная обработка. Можно провести выделение лейкоцитов непосредственно от донора, применяя лейкоферез. В настоящем изобретении можно также использовать необработанные препараты, например, костного мозга или ткани, которые содержат лимфоциты. Для того чтобы облегчить распределение единичных клеток, такие препараты будут нуждаться в измельчении, например таком, как описано выше.
Следующей отличительной особенностью настоящего изобретения является обогащение клеточной фракции, содержащей лимфоциты, например, цельной крови, мононуклеарных клеток, лейкоцитов или костного мозга, особой популяцией лимфоцитов, например линией B-лимфоцитов или T-лимфоцитов. Обогащение B-лимфоцитами можно провести, например, применяя сортинг клеток на магнитных микрочастицах (MACS) или сортинг клеток с флуоресцентной активацией (FACS), используя преимущество линиеспецифических маркерных белков клеточной поверхности, таких как CD19, или других линиеспецифических маркеров B-клеток, таких как B220. Обогащение T-лимфоцитами можно провести, например, используя маркеры клеточной поверхности, такие как CD3, или другие линиеспецифические маркеры T-клеток.
Предпочтительная особенность настоящего изобретения заключается в дальнейшем сортинге обогащенных B-лимфоцитов с целью получения плазмацитов перед распределением клеток по отдельности во множество резервуаров. Выделение плазмацитов обычно проводят посредством сортинга MACS или сортинга FACS, используя такие поверхностные маркеры, как CD19. Также можно использовать другие поверхностные маркеры, специфические для плазмацитов, например CD138, CD43, CD19, MHC-II, причем точный выбор маркера зависит от источника плазмацитов, например селезенки, миндалин, крови или костного мозга. Разумеется, точный источник поверхностных маркеров также зависит от биологического вида, источника выделенных клеток.
В одном из аспектов изобретения маркерами, используемыми для сортинга и/или отбора клеток, являются CD43 и CD138 или MHCII и B220 либо их ортологи. Предпочтительно, чтобы была использована комбинация маркеров CD43 и CD138, и предпочтительно, чтобы отобранные клетки проявляли промежуточную или высокую экспрессию этих маркеров по сравнению с той клеточной популяцией, содержащей лимфоциты, откуда они были выбраны или выделены. Более предпочтительно, чтобы уровень экспрессии CD43 и CD138 был высоким по сравнению с клеточной популяцией, содержащей лимфоциты, откуда они были выбраны или выделены.
Плазмациты также можно получить из необогащенной клеточной популяции, содержащей лимфоциты, которая была получена из любого указанного выше источника. Плазмациты, выделенные из крови, иногда называют ранними плазмацитами или плазмабластами. В настоящем изобретении эти клетки также упоминаются под названием плазмациты (плазматические клетки). Плазмациты желательны для выделения когнатных пар кодирующих последовательностей иммуноглобулинов, поскольку эти клетки с большей частотой продуцируют антигенспецифические антитела, которые отражают приобретенный иммунитет к заданному антигену, причем большинство клеток прошли соматическую гипермутацию и, следовательно, кодируют высокоаффинные антитела. Далее, уровень мРНК в плазмацитах повышен по сравнению с остальной популяцией B-лимфоцитов, поэтому при использовании единичных плазмацитов процедура обратной транскрипции более эффективна. В качестве альтернативы выделению плазмацитов можно выделять из клеточной фракции, содержащей лимфоциты, B-клетки памяти, используя такие поверхностные клеточные маркеры, как CD27 и IgG.
Альтернативная особенность настоящего изобретения заключается в отборе обогащенных B-лимфоцитов для антигенной специфичности ghgl распределением клеток во множество резервуаров. Выделение антигенспецифических B-лимфоцитов осуществляют посредством контакта обогащенных B-лимфоцитов с заданным антигеном или антигенами, что делает возможным связывание антигена с выставленным на поверхности иммуноглобулином и последующее выделение связывающих компонентов. Это можно осуществить, например, посредством биотинилирования заданного антигена или антигенов и последующего применения подходящих методик сортинга клеток. Если желательно, то плазмациты, B-лимфоциты, необогащенные мононуклеарные клетки, лейкоциты, цельную кровь, костный мозг или тканевые препараты можно подвергнуть выделению на специфичность к антигенам.
Еще одна особенность настоящего изобретения заключается в сортинге обогащенных T-лимфоцитов (например, CD3-положительных клеток) с применением поверхностных маркеров, например CD27, для получения фракции T-клеток памяти. T-лимфоциты также можно отбирать на специфичность к MHC-антигену, применяя комплексы MHC-пептида (e.g. Callan M. F. et al. 1998. J. Exp. Med. 187, 1395-1402; Novak EJ. et al. 1999. J. Clin. Invest 104, R63-R67).
В качестве альтернативы сортинга клеток, экспрессирующих определенные поверхностные маркеры, то есть позитивному отбору, возможно обеднение клеточной композиции клетками, не экспрессирующими маркеры, оставляя нетронутыми клетки, которые действительно экспрессируют маркеры.
Следующей особенностью настоящего изобретения является иммортализация любой выделенной клеточной фракции из числа описанных выше (например, B-лимфоцитов, плазмацитов, клеток памяти или T-лимфоцитов). Иммортализацию можно провести, например, вирусом Эпштейна-Барра (Traggiai E. et al., 2004. Nat Med 10, 871-875) перед распределением клеток по резервуарам. В альтернативном варианте выделенные единичные клетки можно иммортализовать и размножить перед проведением обратной транскрипции. (Traggiai et al., Nat Med. 2004 Aug;10(8):871-5).
Следующей особенностью настоящего изобретения является распределение популяции заданных клеток (например, клеток гибридомы, клеточных линий по направлениям дифференцировки B-лимфоцитов или T-лимфоцитов, клеток цельной крови, клеток костного мозга, мононуклеарных клеток, лейкоцитов, B-лимфоцитов, плазмацитов, антигенспецифических B-лимфоцитов, B-клеток памяти, T-лимфоцитов, пептид/MHC-специфических T-лимфоцитов или T-клеток памяти) во множество резервуаров для того, чтобы получить совокупность выделенных единичных клеток. Такое выделение единичных клеток относится к физическому разделению клеток из клеточной популяции таким образом, что отдельный резервуар содержит единичную клетку либо микроматрица, чип или гелевая матрица загружаются таким образом, что продуцируют единичные клетки.
Клетки можно распределить непосредственно во множество резервуаров, например в ряд отдельных резервуаров посредством серийного разведения. Отдельные резервуары, используемые в настоящем изобретении, предпочтительно представляют собой инвентарь, применимый в PCR (например, пробирки для PCR, 96-луночные или 384-луночные планшеты для PCR либо более крупные матрицы с резервуарами). Однако вполне можно использовать и другие резервуары. Распределяя единичные клетки в большое количество единичных резервуаров (например, и 384-луночные планшеты), можно получить совокупность единичных клеток. Такое распределение можно осуществить, например, посредством разлива объема в отдельный резервуар при соблюдении заданной концентрации одна клетка, 0,5 или 0,3 клетки, то есть получая резервуары, которые в среднем содержат единственную клетку или менее. Поскольку распределение клеток методом серийных разведений представляет собой статистическое событие, часть резервуаров будут пустыми, значительная часть будет содержать единственную клетку, а небольшая часть будет содержать две клетки или более. Если в резервуаре представлены две или более клеток, то среди этих клеток может наблюдаться некоторое перемешивание кодирующих последовательностей вариабельной области. Однако это событие является статистически незначимым и не влияет на общую полезность настоящего изобретения. В дополнение к этому в процессе скрининга с наибольшей вероятностью не будут отобраны, то есть будут элиминированы комбинации кодирующих последовательностей вариабельной области, которые не обладают желательной аффинностью и специфичностью связывания. Таким образом, малочисленные события перемешивания не окажут значительного влияния на конечную библиотеку, соответствующую настоящему изобретению.
Существуют альтернативы распределению клеток способом серийных разведений, основанные, например, на использовании сортировщиков клеток, таких как машины FACS или автоматические аппараты, которые можно запрограммировать на точное распределение единичных клеток в отдельные резервуары. Эти альтернативы предпочтительны, поскольку они менее трудоемки и более эффективны в аспекте однородности распределения единичных клеток в отдельные резервуары. Описанные выше процедуры обогащения, сортинга и выделения осуществляются таким образом, что большинство клеток остаются интактными. Разрыв клеток во время обогащения и сортинга может привести к перемешиванию кодирующих последовательностей вариабельной области. Однако это не должно быть существенной проблемой, поскольку ожидаемая частота разрывов невелика.
Любые утечки РНК во время процесса можно удалить посредством отмывания клеток и их возможной обработки РНК-азой перед распределением по отдельным резервуарам.
Далее, рассматривая приведенные выше описания по вопросу о том, как надо распределять клетки, чтобы получить совокупность единичных клеток в совокупности отдельных резервуаров, не следует воспринимать их как абсолютное требование того, что каждый резервуар должен содержать единичную клетку. Скорее они указывают на то, что большинство резервуаров содержит по одной клетке, например доля резервуаров с двумя или более клетками составляет менее 25% от общего количества распределенных клеток, предпочтительнее менее 10%.
Следующая особенность настоящего изобретения заключается в выполнении обратной транскрипции с применением шаблона, извлеченного из клеток, которые распределены по отдельности во множество резервуаров.
В целях обратной транскрипции (RT) и в соответствии с настоящим изобретением считается, что нуклеиновые кислоты единичной клетки, служащей источником шаблона для RT, действительно, извлечены из единичной клетки, хотя не обязательно, чтобы они были отделены от остального содержимого этой единичной клетки.
После того как было проведено заключительное распределение единичных клеток по отдельным резервуарам, можно размножить единичные клетки, чтобы получить популяцию изогенных клеток перед стадией обратной транскрипции. Этот процесс дает больший выход мРНК для использования в качестве шаблона, что может оказаться важным, если необходимо осуществить амплификацию и сцепление редкого целевого объекта. Однако при размножении клетки должны оставаться генетически идентичными в отношении целевого гена. Выделенные клетки или популяцию изогенных клеток можно сохранять в интактном состоянии или лизировать, пока шаблон для обратной транскрипции не разрушен. Для упрощения последующей обратной транскрипции и амплификации в PCR предпочтительно, чтобы клетки были лизированы.
В другом варианте осуществления настоящего изобретения можно также использовать раскрытый здесь способ мультиплексной RT-PCR перекрывания-удлинения или мультиплексной RT-PCR с последующим сцеплением посредством сшивания или рекомбинации, применяя шаблон, извлеченный из генетически разнообразной популяции клеток, которые не были распределены по отдельным резервуарам, но оставлены вместе в виде клеточного пула. Этот способ можно использовать для генерирования комбинаторных библиотек. Такой подход не требует распределения единичных клеток. Однако клетки, которые могут быть использованы при таком подходе, представляют собой точно такие же клетки, как и при описанном выше подходе с единичными клетками, например это может быть популяция (пул) отсортированных B-лимфоцитов или T-лимфоцитов. При проведении одностадийной мультиплексной RT-PCR перекрывания-удлинения или одностадийной мультиплексной RT-PCR с последующим сцеплением посредством сшивания или рекомбинации на такой популяции клеток предпочтительно лизировать клетки перед реакцией и, по желанию, выделить из лизата совокупную РНК или мРНК. В настоящем изобретении чувствительность одностадийной мультиплексной RT-PCR перекрывания-удлинения делает возможным применение шаблона в очень небольшом количестве, например в таком количестве, которое соответствует лизату единичной клетки.
СМЕСИ И КОНСТРУИРОВАНИЕ ПРАЙМЕРОВ
Смеси праймеров, рассматриваемые в настоящем изобретении, содержат, по меньшей мере, четыре праймера, которые образуют набор праймеров два на два, способный амплифицировать, по меньшей мере, две разные представляющие интерес целевые последовательности. Смеси из двух или более таких наборов праймеров составляют мультиплексную смесь праймеров. Предпочтительно, чтобы мультиплексная смесь содержала, по меньшей мере, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20, 30, 40, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140 или 150 наборов праймеров (пар праймеров). Особенно в случае амплификации кодирующих последовательностей вариабельной области индивидуальный набор праймеров в пределах мультиплексной смеси праймеров может содержать более двух праймеров. Предпочтительно, чтобы индивидуальный набор праймеров содержал, по меньшей мере, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 20, 25, 30, 35, 45, 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 160, 170, 180, 190, 200, 220, 240, 260, 280 или 300 праймеров. Предпочтительно, чтобы общее число праймеров в мультиплексной смеси праймеров составляло, по меньшей мере, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 20, 25, 30, 35, 45, 50, 60, 70, 80, 90, 100, 125, 150 или 200 и, самое большее, 225, 250, 275, 300, 325, 350, 375 или 400 праймеров.
Все праймеры, рассматриваемые в настоящем изобретении, содержат геноспецифический участок, и предпочтительно все праймеры дополнительно оснащены хвостами на 5'-конце, то есть 5'-некодирующей последовательностью, которая слита с 3'-концом геноспецифической части праймера. Таким образом, хвост праймера имеет длину приблизительно от 6 до 50 нуклеотидов, но, если это желательно, он может быть и длиннее. При амплификации хвосты праймеров добавляются к целевым последовательностям.
В соответствии с настоящим изобретением хвосты праймеров представляют собой, например, хвосты клонирования и хвосты сцепления, в частности хвосты, адаптированные для сцепления посредством сшивания, хвосты, адаптированные для сцепления посредством рекомбинации, или хвосты перекрывания-удлинения.
Хвосты клонирования могут иметь длину от 6 до 20 нуклеотидов или быть длиннее, а также содержат сайты рестрикции и/или сайты рекомбинации, которые полезны для вставления сцепленного продукта в подходящий вектор.
Для того чтобы создать возможность сцепления посредством сшивания, набор праймеров или мультиплексную смесь праймеров конструируют таким образом, чтобы одна часть первого набора праймеров (прямой или обратный праймер (праймеры)) была оснащена хвостом сцепления, содержащим сайт рестрикции, который при расщеплении будет совместим с сайтом рестрикции, расположенным на хвосте сцепления одной из частей второго набора праймеров. Для сцепления более чем двух целевых последовательностей вторую часть второго набора праймеров оснащают сайтом рестрикции, который при расщеплении будет совместим с сайтом рестрикции, расположенным в первой части третьего набора праймеров. Этот второй сайт рестрикции, расположенный во втором наборе праймеров, должен быть несовместимым с сайтом рестрикции первого набора праймеров. Конструируя наборы праймеров таким способом, можно осуществить сцепление значительного числа целевых последовательностей. Следует выбирать такие сайты рестрикции, которые редко встречаются или вообще не встречаются в целевых последовательностях. Далее, предпочтительно, чтобы совместимые сайты рестрикции не были идентичными, то есть сайт сшивания был бы устойчив к расщеплению конкретными используемыми рестрикционными ферментами. Это направит реакцию в сторону сцепления первой целевой последовательности со второй целевой последовательностью, поскольку сцепление между идентичными целевыми последовательностями будет уязвимо расщеплением рестрикционными ферментами. Подходящими парами сайтов рестрикции являются, например, SpeI c XbaI (в альтернативном варианте один из них или оба могут быть замещены сайтами NheI или AvrII), NcoI c BspHI, EcoRI c MfeI или PstI с NsiI. В целях сцепления SpeI может быть расположен, например, в первой целевой последовательности, XbaI может быть расположен во второй целевой последовательности, NcoI может быть расположен на другом конце второй целевой последовательности, а BspHI в третьей целевой последовательности и так далее. Преимуществом для дальнейшего упрощения процесса является функционирование рестрикционных ферментов в одном и том же буфере.
Для того чтобы создать возможность сцепления посредством рекомбинации, наборы праймеров из мультиплексной смеси праймеров можно, например, конструировать так, как это разъясняется в статье Chapal (1997, BioTechniques 23, 518-524), которая включена сюда в качестве ссылки.
Для того чтобы создать возможность сцепления представляющих интерес нуклеотидных последовательностей на той же стадии, что и амплификация в мультиплексной PCR, хвосты, адаптированные для PCR перекрывания-удлинения, добавляют, по меньшей мере, к одному праймеру из первого набора праймеров в мультиплексной смеси праймеров, посредством чего создается мультиплексная смесь праймеров перекрывания-удлинения. Типичные хвосты перекрывания-удлинения варьируются по длине от 8 до 75 нуклеотидов и могут содержать сайты рестрикции или сайты рекомбинации, позволяющие осуществить последующую вставку таких регуляторных элементов, как промоторы, рибосомные сайты связывания, терминационные последовательности или линкерные последовательности, например, как в scFv. Если это желательно, то хвост перекрывания-удлинения может также содержать стоп-кодон. Обычно имеются три типа хвостов перекрывания-удлинения, как это проиллюстрировано на фиг.1 документа WO 2005/042774. В типе I хвосты перекрывания-удлинения двух наборов праймеров перекрываются только друг с другом. Не обязательно, чтобы все нуклеотиды двух хвостов перекрывания-удлинения были комплементарны друг другу. В одном из аспектов настоящего изобретения комплементарность нуклеотидов в хвостах перекрывания-удлинения составляет от 60 до 85%. В типе II хвостов перекрывания-удлинения от 4 до 6 нуклеотидов на 5'-конце комплементарны геноспецифическому участку прилегающей целевой последовательности. В типе III хвостов перекрывания-удлинения все перекрывание комплементарно прилегающей целевой последовательности. Типы I и II хвостов перекрывания-удлинения предпочтительны тогда, когда позднее должна быть осуществлена вставка регуляторных и т.п. элементов между сцепленными целевыми последовательностями. Хвосты перекрывания-удлинения II типа предпочтительны в том случае, если сцепление целевых последовательностей должно осуществляться посредством определенного линкера, например scFv. Хвосты перекрывания-удлинения III типа предпочтительны в том случае, если сцепление целевых последовательностей друг с другом должно осуществляться в рамке считывания. Конструирование хвостов перекрывания-удлинения зависит от таких свойств последовательности, как длина, относительное содержание GC (GC%), наличие сайтов рестрикции, палиндромы, температуры плавления, геноспецифическая часть, к которой они присоединяются и т.д. Длина хвостов перекрывания-удлинения должна составлять от 8 до 75 нуклеотидов, предпочтительно от 15 до 40 нуклеотидов. Еще более предпочтительно, чтобы длина таких хвостов составляла от 22 до 28 нуклеотидов. Применение очень длинных хвостов перекрывания-удлинения (от 50 до 75 нуклеотидов) будет благоприятствовать сцеплению продуктов, вырабатываемых каждым набором праймеров. Однако при использовании очень длинных хвостов перекрывания-удлинения соотношение между длиной хвоста перекрывания-удлинения и геноспецифическим участком, вероятно, потребует подгонки. Преимущество GC% зависит от длины хвоста перекрывания-удлинения. Поскольку более короткие хвосты имеют меньшую территорию комплементарности, они по сравнению с долее длинными хвостами нуждаются в более высоком GC% для усиления взаимодействия. Подобно этому, следует рассматривать и другие принципы конструирования праймеров, например необходимо свести к минимуму димеризацию праймеров и образование шпилек в нуклеиновой кислоте. К тому же праймеры не должны вовлекаться в ложную затравку. Кроме того, известно, что ДНК-полимераза Taq часто добавляет аденозин (A) на 3'-конце новой синтезируемой нити ДНК, и это обстоятельство можно приспособить к конструированию хвостов перекрывания-удлинения, позволяя этим хвостам воспринимать нешаблонное добавление на 3'-конце.
Выбор праймеров, несущих хвост сцепления, например хвост перекрывания-удлинения, хвост, адаптированный для сцепления посредством сшивания, или хвост, адаптированный для сцепления посредством рекомбинации, определяет порядок и направление сцепления целевых последовательностей. В настоящем изобретении не важно, оснащены ли хвостом сцепления прямой праймер (праймеры), обратный праймер (праймеры) из набора праймеров или, возможно, как прямой, так и обратный праймеры. Однако в любом случае этот вопрос заслуживает некоторого рассмотрения, поскольку порядок и направление целевых последовательностей в конечном продукте может иметь значение, например, для вставки таких регуляторных элементов, как промоторы и терминационные последовательности, или для сцепления индивидуальных целевых последовательностей в рамке считывания. Для сцепления двух представляющих интерес нуклеотидных последовательностей хвосты сцепления могут быть добавлены или к прямому праймеру (праймерам), или к прямому праймеру (праймерам) каждого набора праймеров, используемого для амплификации в PCR каждой целевой последовательности.
Настоящее изобретение иллюстрирует применение хвостов перекрывания-удлинения и хвостов, адаптированных для сцепления посредством сшивания к прямым праймерам mVH и mVK каждого набора. Это приводит к направлению сцепления продуктов 5' с 5' (голова к голове и двунаправленное). Однако хвосты сцепления также могут быть добавлены к обратному праймеру (праймерам) каждого набора. Это приводит к направлению сцепления продуктов 3' с 3' (хвост к хвосту и двунаправленное). Третий возможный вариант заключается в добавлении хвостов сцепления к обратному праймеру (праймерам) первого набора праймеров и к прямому праймеру (праймерам) второго набора праймеров или наоборот. Это приводит к ориентации сцепления 3' с 5' (голова к хвосту и одностороннее направление).
При сцеплении более чем двух представляющих интерес нуклеотидных последовательностей некоторые наборы праймеров нуждаются в хвостах сцепления как на прямом, так и на обратном праймерах, чтобы один хвост был комплементарен хвосту предыдущего набора праймеров, а другой хвост был комплементарен хвосту следующего набора праймеров. Этот принцип выдерживается для всех наборов праймеров, амплифицирующих целевые последовательности, которые должны быть сцеплены между двумя другими целевыми последовательностями. При конструировании геноспецифической части праймера, в общем, надо соблюдать известные правила конструирования праймеров, такие как сведение к минимуму димеризации праймера, образования шпилек и неспецифического отжига. Кроме того, по возможности, следует избегать множественных нуклеотидов G или C как оснований в области 3'. Предпочтительная температура плавления (Tm) геноспецифических участков в наборе праймеров должна быть приблизительно одинаковой с допустимым отклонением плюс/минус 5°C. В настоящем изобретении желательны значения Tm от 45°C до 75°C, а оптимальной для большинства приложений является температура около 60°C.
Предпочтительно, чтобы при начальном конструировании праймера были использованы вспомогательные компьютерные программы, специально разработанные для решения этой задачи. Однако плановые конструкции праймеров, как правило, нуждаются в лабораторном тестировании и стандартной оптимизации. Это может быть сделано, например, посредством анализа размера, полиморфизма длины рестрикционных фрагментов (RFLP) и секвенирования продуктов амплификации, полученных при использовании наборов праймеров. При амплифицировании последовательностей с вариабельными областями или при поиске новых членов семейства, принадлежащих к определенному классу белков полезным подходом является использование в праймерах вырожденных положений. Числа вырожденных положений также могут потребовать оптимизации.
Одной из особенностей настоящего изобретения являются смеси праймеров, составленные, по меньшей мере, из двух наборов праймеров, которые способны служить затравкой для амплификации и сцепления, по меньшей мере, двух представляющих интерес нуклеотидных последовательностей. Смеси праймеров, рассматриваемые в настоящем изобретении, способны служить затравкой амплификации, по меньшей мере, двух субъединиц или доменов гетеромерных белков, например, принадлежащих к классу ферментов, ингибиторов, структурных белков, токсинов, канальных белков, G-белков, рецепторных белков, белков из суперсемейства иммуноглобулинов, транспортных белков и т.д., предпочтительно иммуноглобулинов.
Следующей особенностью настоящего изобретения является мультиплексная смесь праймеров перекрывания-удлинения, содержащая наборы праймеров, в которой, по меньшей мере, один праймер из каждого набора праймеров содержит хвост перекрывания-удлинения, способный гибридизироваться с хвостом перекрывания-удлинения праймера из второго набора праймеров.
Хвосты перекрывания-удлинения создают возможность для непосредственного сцепления представляющих интерес нуклеотидов во время амплификации в мультиплексной PCR перекрывания-удлинения за счет оснащения каждого отдельного продукта, возникающего из набора праймеров, хвостом, который комплементарен прилегающему продукту. Однако это не означает, что во время этой первой амплификации в PCR обязательно происходит сцепление. В зависимости от настройки реакции бóльшая часть фактического сцепления может произойти во время дополнительной амплификации с внешними праймерами первой PCR-амплификации (мультиплексной PCR-амплификации).
Следующей особенностью настоящего изобретения является набор праймеров, сконструированный для амплификации семейства нуклеотидных последовательностей, содержащего кодирующие последовательности вариабельной области. Примерами таких семейств являются легкие цепи каппа (например, VK1-19 у мышей), легкие цепи лямбда (например, VL1-8 у мышей) и вариабельные тяжелые цепи (например, VH1-15 у мышей) из иммуноглобулинов, а также вариабельные области TcR α, β, γ или δ. Набор праймеров для амплификации семейства нуклеотидных последовательностей, содержащего кодирующие последовательности вариабельной области, часто состоит из множества праймеров, в котором некоторые праймеры могут быть вырожденными праймерами. Амплификацию семейств кодирующих последовательностей вариабельной области легкой цепи иммуноглобулина проводят, например, применяя набор праймеров, состоящий из множества праймеров, комплементарных 5'-концу вариабельной области цепи каппа (праймер (праймеры) mVK) или лидерной последовательности каппа и/или цепи лямбда или лидерной последовательности лямбда (прямые праймеры), наряду с константным областям каппа (праймер (праймеры) mKappa1) и/или праймеров лямбда (обратные праймеры) или множества подобных праймеров.
В альтернативном варианте можно использовать праймеры соединительного участка легкой цепи в качестве обратных праймеров вместо праймеров константной области. В альтернативном варианте можно использовать прямые праймеры, отжигающиеся в нетранслируемом участке (UTR), предшествующем лидерной последовательности вариабельной легкой цепи. В равной степени можно амплифицировать семейства кодирующих последовательностей вариабельной области тяжелой цепи иммуноглобулинов с одним набором праймеров, используя различные комбинации праймеров. Например, для этого можно использовать множество праймеров, комплементарных 5'-концу вариабельной области тяжелой цепи (праймер (праймеры) mVH) или лидерной последовательности этого области (прямые праймеры), наряду с множеством праймеров соединительного области тяжелой цепи или праймером (праймерами) константной области тяжелой цепи (обратные праймеры). Праймер mCH может быть специфичным по изотипу, и, в принципе, можно использовать любой праймер mCH, включая и такой, который приведет к амплификации полноразмерной тяжелой цепи. Все же предпочтительно использовать такой праймер mCH, который не амплифицирует полноразмерную тяжелую цепь, чтобы к продукту амплификации можно было добавить константную область тяжелой цепи человека. В альтернативном варианте можно использовать прямые праймеры, отжигающиеся в нетранслируемом участке (UTR), предшествующем лидерной последовательности вариабельной тяжелой цепи.
Использование прямых праймеров, отжигающихся в лидерной последовательности, вместо 5'-конца вариабельной области особенно полезно в том случае, если для праймеров вариабельной области наблюдается перекрестная гибридизация вследствие высокой степени сходства последовательностей. При использовании праймеров лидерной последовательности удается элиминировать из конечного белкового продукта мутации, связанные с перекрестной гибридизацией, поскольку лидерные последовательности отщепляются во время внутриклеточного процессинга белка.
Одной из особенностей настоящего изобретения являются праймеры, которые отжигаются в области 3'-конца лидерной кодирующей последовательности, предшествующей кодирующей последовательности вариабельной области, и их применение для амплификации кодирующих последовательностей вариабельной области.
В одном из вариантов осуществления настоящего изобретения мультиплексная смесь праймеров перекрывания-удлинения, используемая для мультиплексной PCR перекрывания-удлинения и, возможно, также для стадии обратной транскрипции, содержит:
a) по меньшей мере, один праймер mKappa1 или hmJK, комплементарный смысловой нити кодирующей последовательности вариабельной области легкой цепи иммуноглобулина,
b) по меньшей мере, один праймер mVK, комплементарный антисмысловой нити кодирующей последовательности вариабельной области легкой цепи иммуноглобулина или лидерной последовательности вариабельной области легкой цепи, который способен образовывать набор праймеров с праймером (праймерами) стадии a),
c) по меньшей мере, один праймер mCHrev1, mHCrev1-ext или mJH, комплементарный смысловой нити кодирующей последовательности домена тяжелой цепи иммуноглобулина, а также
d) по меньшей мере, один праймер mVH, комплементарный антисмысловой нити кодирующей последовательности вариабельной области тяжелой цепи иммуноглобулина или лидерной последовательности вариабельной области тяжелой цепи, который способен образовывать набор праймеров с праймером (праймерами) стадии c).
В одном из вариантов осуществления настоящего изобретения праймер легкой цепи адаптирован для амплификации кодирующих последовательностей вариабельной области легких цепей каппа и лямбда.
В следующем варианте осуществления настоящего изобретения иммуноглобулиновые праймеры mVK несут хвосты сцепления предпочтительно в виде комплементарных хвостов перекрывания-удлинения. Это порождает кодирующие последовательности вариабельной области, которые сцеплены в направлении голова к голове. Для сцепления кодирующих последовательностей вариабельной области в направлении голова к голове или праймеры mKappa1 и mVH должны содержать хвосты сцепления, или праймеры mVK и mCHrev1 должны содержать хвосты сцепления предпочтительно в виде комплементарных хвостов перекрывания-удлинения. Для сцепления кодирующих последовательностей вариабельной области в направлении хвост к хвосту праймеры mCH и mKappa1 должны содержать хвосты сцепления предпочтительно в виде комплементарных хвостов перекрывания-удлинения.
Предпочтительно, чтобы мультиплексные смеси праймеров, включая мультиплексные смеси праймеров перекрывания-удлинения, рассматриваемые в настоящем изобретении, содержали два набора праймеров. Таким образом, предпочтительная мультиплексная смесь праймеров содержит, по меньшей мере, четыре разных праймера. В следующем аспекте настоящего изобретения мультиплексная смесь праймеров содержит более четырех разных праймеров. Мультиплексную смесь праймеров, рассматриваемую в настоящем изобретении, применяют для амплификации целевых последовательностей в единичном резервуаре. Например, вариабельные области цепей каппа и лямбда можно амплифицировать в одном и том же резервуаре.
Настоящее изобретение также охватывает праймеры для дополнительной амплификации в PCR сцепленных продуктов, полученных посредством мультиплексной RT-PCR с их последующим сцеплением посредством сшивания или рекомбинации или посредством мультиплексной RT-PCR перекрывания-удлинения. Такую дополнительную амплификацию в PCR можно провести, используя смесь праймеров, адаптированную для амплификации сцепленных целевых последовательностей.
Такая смесь праймеров может содержать внешние праймеры из мультиплексной смеси праймеров или мультиплексную смесь праймеров перекрывания-удлинения, а это означает, что праймеры, которые отжигаются с самыми удаленными 5'-концом и 3'-концом смысловой нити сцепленных кодирующих последовательностей вариабельной области, тем самым делают возможной амплификацию всего сцепленного продукта. Этот процесс обычно применяется для увеличения количества сцепленного продукта, который был получен в результате мультиплексной RT-PCR с последующим сцеплением посредством сшивания или рекомбинации или в результате мультиплексной RT-PCR перекрывания-удлинения.
В альтернативном варианте для дополнительной амплификации сцепленных нуклеотидных последовательностей можно использовать набор праймеров, который вложен по сравнению с внешними праймерами, применяемыми в мультиплексной RT-PCR или в мультиплексной RT-PCR перекрывания-удлинения. В настоящем изобретении такой набор праймеров называется вложенным набором праймеров. Конструирование вложенных праймеров в целом подчиняется тем же правилам конструирования, как и для описанных выше геноспецифических праймеров, за тем исключением, что они служат затравкой для частичной или полной амплификации от 3' до положения отжига внешних праймеров, применяемых в мультиплексной RT-PCR или в мультиплексной RT-PCR перекрывания-удлинения. Следовательно, результирующий продукт вложенной PCR может быть короче, чем сцепленный продукт, полученный в мультиплексной RT-PCR с последующим сцеплением посредством сшивания или рекомбинации либо в мультиплексной RT-PCR перекрывания-удлинения. В дополнение к увеличению количества сцепленного продукта вложенная PCR также помогает увеличить общую специфичность, особенно специфичность технологии мультиплексной RT-PCR перекрывания-удлинения. Однако необходимо отметить, что не все мультиплексные смеси праймеров/мультиплексные смеси праймеров перекрывания-удлинения, которые были описаны выше, подходят для комбинирования с вложенным набором праймеров при проведении дополнительной амплификации. В таких случаях для дополнительной амплификации можно использовать внешние праймеры мультиплексной смеси праймеров/мультиплексной смеси праймеров перекрывания-удлинения или применить полувложенную PCR, как это будет описано ниже.
В одном из вариантов осуществления настоящего изобретения для дополнительной амплификации сцепленных кодирующих последовательностей вариабельной области иммуноглобулина в качестве вложенных праймеров используют смесь праймеров JL и JH. Вложенные наборы праймеров, рассматриваемые в настоящем изобретении, также могут состоять из обратного (или прямого) внешнего праймера (праймеров) первой мультиплексной смеси праймеров/мультиплексной смеси праймеров перекрывания-удлинения и второго вложенного праймера, который служит затравкой для амплификации отрезка последовательности от 3'-конца до положения отжига прямого (или обратного) внешнего праймера (праймеров) первой мультиплексной смеси праймеров/мультиплексной смеси праймеров перекрывания-удлинения. Применение такого набора праймеров для дополнительной амплификации в PCR, в общем, известно под названием полувложенной PCR. Полувложенную PCR можно, например, применить, если трудно сконструировать вложенный праймер в одном специфическом участке, например, для последовательностей вариабельной области, поскольку такой праймер будет отжигаться в областях определения комплементарности (CDR). Кроме того, полувложенную PCR можно использовать тогда, когда желательно сохранить один конец сцепленных последовательностей интактным, например, в целях клонирования.
ОПТИМИЗАЦИЯ МУЛЬТИПЛЕКСНОЙ PCR ПЕРЕКРЫВАНИЯ-УДЛИНЕНИЯ
Параметры стадии мультиплексной PCR перекрывания-удлинения как для двухстадийной, так и для одностадийной процедуры можно оптимизировать по нескольким параметрам (см., например, ссылки Henegariu O. et al. 1997. BioTechniques 23, 504-511; Markoulatos P. et al. 2002. J. Clin. Lab. Anal. 16, 47-51). Для мультиплексной RT-PCR обычно применимы те же параметры оптимизации, хотя соотношение между внешними и внутренними праймерами для такой реакции менее важно.
a. Концентрация праймеров
Предпочтительно, чтобы концентрация праймеров, несущих хвосты перекрывания-удлинения (например, праймеров VH и VL), была ниже, чем концентрация внешних праймеров без хвостов перекрывания-удлинения (например, праймеров JH и каппа). Если одна из целевых последовательностей амплифицируется с меньшей эффективностью, чем другие, например, в результате более высокого показателя GC%, то вполне возможно выровнять эффективность амплификации. Это можно осуществить, применяя более высокую концентрацию набора праймеров, который опосредует малоэффективную амплификацию, или понизив концентрацию других праймеров. Например, последовательности, кодирующие вариабельные области тяжелой цепи, проявляют тенденцию к более высокому показателю GC% и, следовательно, к пониженной эффективности амплификации по сравнению с последовательностями, кодирующими вариабельные области легкой цепи. Это указывает на целесообразность использования праймеров VL с пониженной концентрацией по сравнению с праймерами VH.
Далее, при использовании большого числа праймеров общая концентрация праймеров может оказаться серьезной проблемой. Верхний предел определяют экспериментально посредством титрования. Для системы PCR "AmpliTaq Gold" производства компании Applied Biosystems был установлен верхний предел общей концентрации олигонуклеотидов 1,1 мкм, однако для других систем он может быть выше и достигать 2,4 мкм. Такой верхний уровень общей концентрации олигонуклеотидов влияет на максимальную концентрацию отдельных праймеров. При слишком низкой концентрации какого-либо отдельного праймера чувствительность PCR может оказаться плохой.
Также было обнаружено, что качество олигонуклеотидных праймеров довольно важно для мультиплексной PCR перекрывания-удлинения. Лучшие результаты давали олигонуклеотиды, очищенные способом HPLC.
b. Условия циклов PCR:
Предпочтительно, чтобы циклы проводились в следующих условиях:
Все эти параметры можно оптимизировать. Особенно важна температура отжига. В связи с этим все индивидуальные наборы праймеров, которые составляют конечную смесь праймеров, необходимо протестировать по отдельности, чтобы идентифицировать оптимальные температуру отжига и время, а также время удлинения и денатурации. Это дает хорошее представление об окне, в пределах которого можно оптимизировать указанные параметры для мультиплексной смеси праймеров перекрывания-удлинения.
Проблемы, связанные с плохой чувствительностью PCR, например, вследствие недостаточной концентрации праймеров или шаблона, можно преодолеть за счет увеличения числа термических циклов. Увеличенное число термических циклов составляет от 35 до 80 предпочтительно, около 40 циклов. Кроме того, процесс мультиплексной PCR перекрывания-удлинения можно улучшить за счет увеличения времени удлинения. Увеличенное время удлинения составляет 1,5-5 минут × EPL по сравнению с обычным удлинением в течение 1 минуты.
c. Использование адъювантов
Мультиплексные реакции PCR можно существенно улучшить, применяя добавки PCR, например DMSO, глицерин, формамид или бетаин, которые расслабляют ДНК, то есть упрощают денатурацию шаблона.
d. dNTP и MgCl 2
Для мультиплексной PCR перекрывания-удлинения важны качество и концентрация деоксинуклеозидтрифосфата (dNTP). Лучшая концентрация dNTP составляет от 200 до 400 мкм в расчете на каждый dNTP (dATP, dCTP, dGTP и dTTP), а при более высоких концентрациях наступает быстрое подавление амплификации. Для достижения амплификации в PCR вполне достаточна пониженная концентрация dNTP (100 мкм для каждого dNTP). Запасы dNTP чувствительны к циклам замораживания/оттаивания. После трех-пяти таких циклов мультиплексная PCR часто теряет эффективность. Для того чтобы избавиться от таких проблем, можно приготовить маленькие аликвоты dNTP и держать их замороженными при -20°C.
Оптимизация концентрации Mg2+ критически важна, поскольку большинство ДНК-полимераз являются ферментами, зависимыми от магния. В дополнение к ДНК-полимеразе ионы Mg2+ связываются праймерами шаблона ДНК и dNTP. Следовательно, оптимальная концентрация Mg2+ будет зависеть от концентрации dNTP, от шаблона ДНК и от композиции пробного буфера. Если праймеры и/или буферы шаблона ДНК содержат такие хелатообразователи, как EDTA ИЛИ EGTA, кажущийся оптимум Mg2+ может быть изменен. Избыточная концентрация Mg2+ стабилизирует двойную нить ДНК и предупреждает полную денатурацию ДНК, которая ограничивает выход продукта. Избыток Mg2+ может также стабилизировать ложный отжиг праймера с неправильными сайтами шаблона, снижая тем самым специфичность. С другой стороны неадекватная концентрация Mg2+ уменьшает количество продукта.
Хороший баланс между dNTP и MgCl2 составляет приблизительно от 200 до 400 мкм dNTP (каждого) на 1,5-3 мкм MgCl2.
e. Концентрация буферов в PCR
Для мультиплексной PCR перекрывания-удлинения обычно достаточны буферы на основе KCl, однако для функционирования с мультиплексной RT-PCR перекрывания-удлинения также могут быть оптимизированы буферы, основанные на других компонентах, например (NH4)2SO4, MgSO4, Tris-HCl или на их комбинациях. Пары праймеров, вовлеченные в амплификацию более длинных продуктов, лучше действуют при пониженных концентрациях соли (например, от 20 до 50 mM KCl), тогда как пары праймеров, вовлеченные в амплификацию коротких продуктов, лучше действуют при повышенных концентрациях соли (например, от 80 до 100 mM KCl). Увеличение концентрации буфера до 2Х вместо 1Х может улучшить эффективность мультиплексной реакции.
f. ДНК-полимераза
Настоящее изобретение иллюстрируется на примере полимеразы Taq. В альтернативных вариантах можно использовать другие типы термостабильных ДНК-полимераз, включая, например, полимеразы Pfu, Phusion, Pwo, Tgo, Tth, Vent, Deep-vent. Полимеразы без активности экзонуклеазы от 3' до 5' или обладающие такой активностью могут использоваться как поодиночке, так и в комбинациях друг с другом.
ВЕКТОРЫ И БИБЛИОТЕКИ
В соответствии с настоящим изобретением сцепление представляющих интерес нуклеотидных последовательностей приводит к созданию нуклеотидного сегмента, содержащего сцепленные нуклеотидные последовательности, кодирующие вариабельные области иммуноглобулинов. Далее, на основе способов, описанных в настоящем изобретении, можно создать библиотеки таких сцепленных последовательностей нуклеиновой кислоты, особенно библиотеки кодирующих последовательностей вариабельной области, отличных от человеческих, сцепленных или сплайсированных с кодирующими последовательностями константной области человека (легкой и тяжелой цепи).
Одна из особенностей настоящего изобретения заключается во вставлении сегмента, содержащего представляющие интерес нуклеотидные последовательности или библиотеки представляющих интерес сцепленных нуклеотидных последовательностей, которая создана способом, предложенным настоящим изобретением, в подходящие векторы. Библиотеки могут быть комбинаторными библиотеками или более предпочтительно библиотеками когнатных пар кодирующих последовательностей вариабельной области. Предпочтительные сайты рестрикции, созданные внешними праймерами, вложенными праймерами или полувложенными праймерами, предназначены для согласования с соответствующими сайтами рестрикции выбранного вектора. Если один из полувложенных, вложенных или внешних праймеров оснащен подходящим сайтом рекомбинации, а выбранный вектор также содержит такой сайт, то представляющие интерес сцепленные последовательности нуклеиновой кислоты также можно вставлять в векторы посредством рекомбинации. По существу, нет никаких ограничений на векторы, которые можно использовать в качестве носителей продуктов, созданных каким-либо из способов мультиплексной RT-PCR-сцепления, предлагаемых настоящим изобретением. Векторами выбора могут быть векторы, пригодные для амплификации и экспрессии в клетках, включая, например, бактерии, дрожжи, другие грибки, клетки насекомых, растительные клетки или клетки млекопитающих. Такие векторы можно использовать для облегчения дальнейших стадий клонирования, челночных перемещений между векторными системами, демонстрации продукта, вставленного в вектор, экспрессии вставленного продукта и/или интеграции в геном клетки-хозяина.
Предпочтительными векторами клонирования и челночными векторами являются бактериальные векторы. Однако в процедурах клонирования и челночных перемещений можно также использовать другие типы векторов.
Векторами демонстрации могут быть, например, фаговые векторы или фагемидные векторы, происходящие класса нитчатых бактериофагов fd, M13 или f1. Такие векторы способны облегчать демонстрацию белков, включая, например, белок связывания или его фрагмент на поверхности нитчатого бактериофага. В данной области техники также известны векторы демонстрации, пригодные для демонстрации на рибосомах, ДНК, дрожжевых клетках или клетках млекопитающих. К ним относятся, например, вирусные векторы или векторы, кодирующие химерные белки.
Векторы экспрессии существуют для всех указанных биологических видов, и выбор вектора полностью зависит от экспрессируемого белка. Некоторые векторы экспрессии дополнительно способны к встраиванию в геном клетки-хозяина посредством либо случайной интеграции, либо сайт специфической интеграции с использованием соответствующих сайтов рекомбинации. Векторы экспрессии можно конструировать так, чтобы обеспечить дополнительные кодирующие последовательности, причем сцепленный продукт должен встраиваться в рамку считывания этих последовательностей, делая возможной при введении в клетку-хозяина экспрессию более крупного белка, например, полноразмерного моноклонального антитела. Такое вставление в рамку считывания может также облегчить экспрессию химерных белков, которая облегчает представление на поверхности нитчатого бактериофага или клетки. В системе бактериофагового дисплея представляющие интерес сцепленные нуклеотидные последовательности могут быть вставлены в рамку считывания последовательности, кодирующей белок оболочки, например, pIII или pVIII (Barbas CF. et al. 1991. Proc. Natl. Acad. Sci. USA 88, 7978-7982; Kang A.S. et al. 1991. Proc. Natl. Acad. Sci. USA 88, 4363-4366).
В одном из вариантов осуществления настоящего изобретения отдельные сегменты представляющих интерес сцепленных нуклеотидных последовательностей состоят из кодирующей последовательности вариабельной области тяжелой цепи иммуноглобулина, связанной с кодирующей последовательностью вариабельной области легкой цепи от одного биологического вида, вставленных в вектор, который содержит кодирующую последовательность (последовательности) одного или более константных доменов иммуноглобулина человека, предпочтительно константных областей как легкой, так и тяжелой цепи человека. Вставку конструируют таким образом, что сцепленные кодирующие последовательности вариабельной области тяжелой цепи и/или вариабельной области легкой цепи вставляются в рамку считывания кодирующих последовательностей константной области. Такая вставка может, например, привести к созданию вектора экспрессии Fab или F(ab')2, вектора экспрессии полноразмерного антитела или вектора экспрессии, кодирующего фрагмент полноразмерного антитела. Предпочтительно, чтобы такой вектор был пригоден для экспрессии (например, векторы E. coli, фагемидные векторы или векторы млекопитающих), а кодирующие последовательности константной области тяжелой цепи были выбраны из классов иммуноглобулинов человека IgGI, IgG2, IgG3, IgG4, IgM, IgA1, IgA2, IgD или IgE, создавая тем самым возможность для экспрессии Fab или полноразмерного рекомбинантного антитела. В дополнение к кодирующим последовательностям константной тяжелой цепи вектор также может содержать кодирующую последовательность константной легкой цепи, выбранной из цепей каппа и лямбда человека. Это предпочтительно при создании химерных антител, поскольку в этих случаях сцепленные нуклеотидные последовательности представлены только кодирующими последовательностями вариабельной области иммуноглобулина (Fv's) от биологических видов, не являющихся человеком. В альтернативном варианте осуществления изобретения константная область (области) человека сплайсируется или сцепляется с вариабельными областями, отличными от человеческих на той стадии, когда выполняется процедура молекулярной амплификации, причем это осуществляется посредством добавления в резервуары константной области человека, обеспечивающего перекрывание с последовательностью, отличной от человеческих, а также праймеров, служащих затравкой для амплификации как вариабельного, так и константной области (областей) в рамке считывания. Таким способом можно добавить константную цепь каппа или лямбда человека и/или константную тяжелую цепь человека. Применяя эту процедуру, можно избавиться от необходимости присутствия сайта рестрикции в пределах кодирующей последовательности, что является преимуществом.
Еще в одном варианте осуществления настоящего изобретения отдельные сегменты сцепленных нуклеотидных последовательностей состоят из кодирующей последовательности вариабельной области цепи TcR α, связанной с кодирующей последовательностью вариабельной области цепи β, или из кодирующей последовательности вариабельной области цепи γ, связанной с кодирующей последовательностью вариабельной области цепи δ.
Предпочтительно, чтобы эти сцепленные последовательности были вставлены в вектор, который содержит последовательности, кодирующие один или более константных доменов TcR. Вставку конструируют таким образом, что вставленные сцепленные кодирующие последовательности вариабельной области попадают в рамку считывания соответствующих кодирующих последовательностей константной области TcR. В следующем варианте осуществления настоящего изобретения такой вектор представляет собой химерный вектор экспрессии, содержащий последовательности, которые кодируют лейциновую застежку в рамке считывания константных областей TcR. Было показано, что такие конструкции повышают стабильность растворимых TcR (Willcox B.E. et al. 1999. Protein Sci 8, 2418-2423).
Библиотеки когнатных пар, предлагаемые настоящим изобретением, могут быть введены в векторы двумя разными способами. При первом подходе единичные когнатные пары вставляют по отдельности в подходящий вектор. Такую библиотеку векторов впоследствии можно держать по отдельности или объединить. При втором подходе все когнатные пары перед вставлением в вектор объединяют, а затем общей массой вставляют в подходящие векторы, создавая объединенную библиотеку векторов. Такая библиотека векторов содержит большое разнообразие пар кодирующих последовательностей вариабельной области.
Одним из аспектов настоящего изобретения является библиотека антител с когнатными парами сцепленных кодирующих последовательностей вариабельной области. Предпочтительно, чтобы отдельные антитела в составе библиотеки содержали кодирующую последовательность вариабельной области легкой цепи иммуноглобулина, связанную с кодирующей последовательностью вариабельной области тяжелой цепи, от одного биологического вида, а также константные области человека.
Еще одна предпочтительная библиотека когнатных пар содержит сцепленные кодирующие последовательности области TcR, причем каждая отдельная кодирующая последовательность области TcR содержит кодирующую последовательность вариабельной области цепи альфа, связанную с кодирующей последовательностью вариабельной области цепи бета, и/или кодирующую последовательность вариабельной области TcR гамма, связанную с кодирующей последовательностью вариабельной области цепи дельта.
Одним из вариантов осуществления настоящего изобретения является вспомогательная библиотека когнатных пар сцепленных кодирующих последовательностей вариабельной области, которые кодируют желательные специфичности связывания, направленные на конкретную цель. Предпочтительно, чтобы эти когнатные пары включали сцепленные кодирующие последовательности вариабельной области легкой цепи иммуноглобулина и вариабельной области тяжелой цепи иммуноглобулина, кодирующие последовательности вариабельной области цепи TcR альфа и вариабельной области цепи бета и/или кодирующие последовательности вариабельной области цепи TcR гамма и вариабельной области цепи дельта.
Еще один вариант осуществления настоящего изобретения заключается во вспомогательной библиотеке, выбранной из родительской библиотеки когнатных пар кодирующих последовательностей вариабельной области, как это описано во всем тексте заявки.
Предпочтительный вариант осуществления настоящего изобретения заключается в библиотеке или вспомогательной библиотеке, кодирующей когнатные пары полноразмерного химерного иммуноглобулина, выбранного из иммуноглобулинов человека классов IgA1, IgA2, IgD, IgE, IgG1, IgG2, IgG3, IgG4 или IgM.
Еще одной предпочтительной особенностью настоящего изобретения является библиотека или вспомогательная библиотека, кодирующая растворимые и стабильные когнатные пары TcR.
Особенностью настоящего изобретения является разнообразие указанных библиотек, которые состоят, по меньшей мере, из 5, 10, 20, 50, 100, 1000, 104, 105 или 106 различных когнатных пар антител.
В следующем варианте осуществления настоящего изобретения указанные библиотеки когнатных пар сцепленных кодирующих последовательностей вариабельной области могут быть получены способом, который включает описанные здесь стадии. Такую библиотеку также называют родительской библиотекой.
СКРИНИНГ И ОТБОР
Ожидается, что родительская библиотека пар сцепленных кодирующих последовательностей вариабельной области, выделенных от донора с применением одного из способов, описанных в настоящем изобретении, должна представлять разнообразие связывающих белков, в котором некоторые белки будут нерелевантными, то есть неспособными связываться с нужной целью, что особенно характерно для комбинаторных библиотек. Следовательно, настоящее изобретение охватывает обогащение и скрининг вспомогательной библиотеки, кодирующей подмножество различных связывающих специфичностей, направленных против конкретной цели.
Для библиотек когнатных пар ожидается, что разнообразие библиотеки будет отражать разнообразие, представленное в донорском материале, и лишь небольшое число случайно сцепленных вариабельных областей. Таким образом, перед скринингом библиотеки, состоящей из когнатных пар, на целеспецифические аффинности связывания стадия обогащения может быть необязательной.
В следующем варианте осуществления настоящего изобретения способ создания библиотеки пар сцепленных кодирующих последовательностей вариабельной области дополнительно включает создание вспомогательной библиотеки посредством отбора подмножества пар сцепленных последовательностей вариабельной области, которые кодируют связывающие белки с желательной целевой специфичностью. Такой отбор сцепленных кодирующих последовательностей вариабельной области также называется созданием библиотеки целеспецифических когнатных пар.
В предпочтительном варианте осуществления настоящего изобретения библиотеку целеспецифических когнатных пар кодирующих последовательностей вариабельной области переносят в вектор экспрессии. Вектором экспрессии может быть вектор экспрессии млекопитающего, дрожжевой вектор экспрессии, грибковый вектор экспрессии, растительный вектор экспрессии, бактериальный вектор экспрессии в зависимости от типа клетки, используемого для скрининга. Предпочтительно, чтобы вектор экспрессии был вектором млекопитающего.
Для отбора кодирующих последовательностей вариабельной области целеспецифического иммуноглобулина обычно бывают пригодными иммунологические анализы. Такие анализы хорошо известны в данной области техники и включают, например, FMAT, FLISA, ELISPOT, ELISA, мембранные анализы (например, вестерн-блоттинг), матрицы на фильтрах или FACS. Анализы можно проводить прямым образом, используя полипептиды, полученные из кодирующих последовательностей вариабельной области иммуноглобулина. В альтернативном варианте иммуноанализы можно проводить в комбинации с такими способами обогащения, как фаговый дисплей, рибосомный дисплей, представление на бактериальной поверхности, дрожжевой дисплей, эукариотический вирусный дисплей, РНК-дисплей или ковалентный дисплей (обзор см. в ссылке FitzGerald K., 2000. Drug Discov. Today 5, 253-258). Как когнатные библиотеки экспрессии Fab, так и когнатные библиотеки экспрессии полноразмерных антител можно подвергать скринингу, посредством которого можно создавать вспомогательные библиотеки позитивных клонов. Такие анализы скрининга и процедуры обогащения также пригодны для фрагментов Fv или scFv либо для комбинаторных библиотек сцепленных вариабельных областей.
В дополнение к иммунологическому скринингу отличительной особенностью изобретения является возможность применения разных типов функционального скрининга для отбора клонов, секретирующих антитела с желательными свойствами. Такие скрининговые анализы включают, но не ограничиваясь ими, анализы пролиферации, анализы вирусной инактивации, анализы на уничтожение клеток и т.д. Предпочтительно, чтобы функциональные анализы проводились в формате высокой производительности с применением супернатантов из клеток, трансфицированных векторами экспрессии, рассматриваемыми в изобретении.
В предпочтительном варианте осуществления настоящего изобретения отбор вспомогательной библиотеки целеспецифических когнатных пар или комбинаторных пар кодирующих последовательностей вариабельной области осуществляют с применением высокопроизводительного скринирующего анализа. Высокопроизводительными скринирующими анализами могут быть, но не ограничиваясь ими, анализы ELISA, выполняемые с полуавтоматическим или полностью автоматизированным оборудованием. Это также может быть мембранный анализ, в котором бактерии собраны с помощью робота и нанесены сеткой на соответствующую мембрану поверх агаровых пластинок, создавая матрицы для колоний, экспрессирующих антигенсвязывающие молекулы. Молекулы секретируются через мембрану на вторую мембрану, расположенную ниже и покрытую антигеном, которую можно разработать отдельно и использовать для идентификации клонов, секретирующих антигенсвязывающие молекулы, направленные против нужной цели (de Wildt R.M. et al. 2000. Nat. Biotechnol. 18, 989-994).
После того как вспомогательная библиотека когнатных пар или комбинаторных пар антигенсвязывающих клонов была отобрана при помощи подходящей технологии, можно провести дополнительный анализ посредством секвенирования ДНК сцепленных кодирующих последовательностей вариабельной области легких цепей и вариабельной области тяжелых цепей иммуноглобулина. Прежде всего такое секвенирование ДНК даст информацию о разнообразии библиотеки, в частности о зародышевой линии ее происхождения, о распределении по семействам и о созревании в пределах участков CDR. Такой анализ позволит отобрать клоны, которые представляют широкое разнообразие, и исключить повторяющиеся клоны. Во-вторых, секвенирование ДНК отразит мутации, внесенные в процессе выделения.
При анализе кодирующих последовательностей вариабельной области могут встретиться три типа мутаций, которые надо оценивать в аспекте приемлемости:
i) Мутации наиболее частого типа происходят в результате перекрестного примирования, когда праймер V-гена вследствие сходства последовательностей служит затравкой для последовательности зародышевой линии, с которой он не имеет полной гомологии. Внесенные изменения, главным образом, представлены замещениями природных кодонов в одном особом положении вследствие высокой степени сходства между V-генными последовательностями. Некоторые из этих изменений могут быть значительными, иногда не имея аналога в природе. Такие изменения потенциально способны повлиять на иммуногенность вариабельной области, создавая в нем новые эпитопы. Такие изменения достаточно просто поддаются идентификации и последующему исправлению с применением стандартных методик молекулярной биологии, либо мутантные клоны можно исключить из библиотеки.
ii) Ошибки, созданные ДНК-полимеразой Taq, проще всего поддаются идентификации в кодирующих последовательностях константной области и легко элиминируются. Однако мутации, индуцированные Taq, конечно, также будут присутствовать в кодирующих последовательностях вариабельной области, где они неотличимы от природных соматических мутаций, которые также являются результатом случайных мутаций в кодирующих последовательностях вариабельной области. Принимая во внимание, что мутации не являются систематическими и особым образом затрагивают только специфические пары, представляется разумным игнорировать такие изменения. В следующем варианте осуществления настоящего изобретения вспомогательная библиотека целеспецифических и, возможно, секвенированных пар сцепленных кодирующих последовательностей вариабельной области легкой цепи и вариабельной области тяжелой цепи иммуноглобулина переносят в вектор экспрессии млекопитающего. Такой перенос можно осуществить в любой из векторов, описанных в предыдущем разделе, что создает возможность для экспрессии полноразмерного рекомбинантного антитела. Такой перенос может не понадобиться в том случае, если проводится скрининг библиотеки экспрессии когнатных полноразмерных антител млекопитающего.
Еще в одном варианте осуществления настоящего изобретения создается родительская библиотека из клеточной фракции, содержащей лимфоциты, которая обогащена T-лимфоцитами. Пары сцепленных кодирующих последовательностей вариабельной области, составляющие родительскую библиотеку, могут быть отобраны для кодирования подмножества пар сцепленных последовательностей вариабельной области, состоящих из альфа и бета и/или гамма и дельта-цепей, которые кодируют связывающие белки с желательной целевой специфичностью, посредством чего создается вспомогательная библиотека когнатных пар или комбинаторных пар. Впоследствии в пуле трансфицированных клеток могут быть идентифицированы антигенспецифические рецепторы T-клеток с применением стандартной методологии, например посредством окрашивания тетрамерными комплексами MHC-пептид (e.g., Callan M. F. et al. 1998. J. Exp. Med. 187, 1395-1402; Novak EJ. et al. 1999. J. Clin. Invest 104, R63-R67), измерения клеточных ответов в виде выброса IL-2, или более сложными средствами, например методиками дрожжевого или ретровирусного дисплея.
КЛЕТКИ-ХОЗЯЕВА И ЭКСПРЕССИЯ
Библиотеки, рассматриваемые в настоящем изобретении, могут быть перенесены в векторы, пригодные для экспрессии и выработки белков, кодируемых представляющими интерес сцепленными нуклеотидными последовательностями, в особенности связывающих белков или их фрагментов, содержащих вариабельный область. Такие векторы описаны с разделе Векторы и библиотеки и предусматривают экспрессию, например, полноразмерных антител, фрагментов Fab, фрагментов Fv, scFv, связанных с мембранами или растворимых TcR или фрагментов TcR биологических видов выбора.
Одна из особенностей настоящего изобретения заключается во введении в клетку-хозяина библиотеки или вспомогательной библиотеки векторов с когнатными парами сцепленных кодирующих последовательностей вариабельной области или единичного клона, кодирующего когнатную пару сцепленных кодирующих последовательностей вариабельной области для амплификации и/или экспрессии. Клетки-хозяева могут быть выбраны из бактерий, дрожжей, других грибков, клеток насекомого, растительных клеток или клеток млекопитающего. В целях экспрессии предпочтительны такие клетки млекопитающих, как клетки яичника китайского хомячка (CHO), клетки COS, клетки BHK, клетки миеломы (например, клетки Sp2/0 или NSO), NIH 3T3, такие фибробласты или иммортализованные клетки человека, в частности клетки HeLa, клетки HEK 293 или PER.C6.
Введение векторов в клетку-хозяина можно осуществить множеством способов трансформации или трансфекции, известных компетентному специалисту в данной области техники, включая такие способы, как осаждение фосфатом кальция, электропорация, микроинъекция, липосомное слияние, слияние с пустой стромой эритроцитов, слияние протопластов, вирусная инфекция и т.д.
Достаточно хорошо известна выработка полноразмерных моноклональных антител, фрагментов Fab, фрагментов Fv и фрагментов scFv. Выработка рекомбинантных поликлональных антител для их лечебного применения является относительно новой областью. Технология производства рекомбинантных поликлональных антител была описана в заявке PCT WO 2004/061104. Вкратце, эта технология связана с созданием коллекции клеток, пригодной для выработки клеточных линий. Последующее описание методики дано для библиотеки когнатных пар, однако оно с тем же успехом применимо для комбинаторной библиотеки. Отдельные клетки в коллекции клеток способны экспрессировать индивидуальный рекомбинантный связывающий белок, например, из библиотеки когнатных пар. Для большей уверенности в том, что индивидуальные клетки экспрессируют единственную когнатную пару, а не несколько когнатных пар поликлонального связывающего белка, последовательности нуклеиновой кислоты, кодирующие когнатные пары, вводят в единичный сайт-специфический участок в геноме каждой индивидуальной клетки. Это является важным атрибутом для коллекции клеток, поскольку предупреждает перемешивание тяжелой и легкой цепей, экспрессируемых из каждой клетки, но также потому, что позволяет создать клетки, которые практически идентичны друг другу за исключением небольших различий в вариабельных областях отдельных когнатных пар. Это свойство создает возможность для несмещенного роста коллекции клеток в течение периода времени, необходимого для выработки продукта. Для того чтобы гарантировать единичную сайт-специфическую интеграцию, надо использовать линию клеток-хозяев, имеющих только один сайт интеграции (такие клетки поступают в продажу, например клетки CHO Flp-In, содержащие единственный сайт FRT). Подходящие векторы для этой клеточной линии содержат соответствующую рекомбиназу. Существует несколько других известных рекомбиназ, которые можно использовать в комбинации с их соответствующими сайтами интеграции, например Cre, бета-рекомбиназа, Gin, Pin, PinB, PinD, R/RS, лямбда-интеграза или фаговая ΦC31 интеграза. Кроме того, подходящие векторы содержат маркер отбора, который делает возможным отбор сайт-специфических интегрантов.
Размножение поликлональной производительной клеточной линии и выработку рекомбинантного поликлонального белка из такой клеточной линии можно получить на основе нескольких разных стратегий трансфекции и производственной технологии.
Один из способов заключается в использовании векторов, смешанных в единую композицию, для трансфекции линии клеток-хозяев с единственным сайтом интеграции на клетку. Этот способ называется массовой трансфекцией или трансфекцией в массе. Как правило, описанная выше конструкция вектора и клетки-хозяина обеспечивает получение в результате правильного отбора поликлональной клеточной линии, способной к несмещенному росту. Перед началом производства рекомбинантного поликлонального белка создается замороженный запас поликлональной клеточной линии.
Другой способ заключается в использовании библиотеки векторов, которая разделена на фракции, содержащие приблизительно от 5 до 50 индивидуальных векторов из этой библиотеки, в композиции для трансфекции. Предпочтительно, чтобы фракция библиотеки состояла из 10-20 индивидуальных векторов. Затем каждую композицию трансфицируют в аликвотную часть клеток-хозяев. Этот способ называется полумассовой трансфекцией. Число трансфицированных аликвотных частей зависит от размера библиотеки и от числа индивидуальных векторов в каждой фракции. Например, если библиотека состоит из 100 разных когнатных пар, которые разделены на фракции, содержащие по 20 разных пар в композиции, нужны 5 аликвот клеток-хозяев, которые будут трансфицированы композициями, составляющими отдельные фракции исходной библиотеки. Для сайт-специфической интеграции отбирают аликвоты клеток-хозяев. Предпочтительно, чтобы отбор разных аликвотных частей проводился по отдельности. Однако перед отбором их также можно объединить. Аликвоты можно проанализировать на клональное разнообразие, причем для создания запаса библиотеки поликлональных когнатных пар используются только аликвоты с достаточным разнообразием.
Для получения желательной поликлональной клеточной линии в целях выработки продукта аликвоты можно смешать перед созданием замороженного запаса сразу же после их извлечения из исходного запаса или после короткого периода пролиферации и адаптации. По выбору аликвоты клеток можно держать по отдельности в процессе получения, а композицию поликлональных антител составляют, скорее комбинируя продукты каждой аликвоты, чем перемешивая аликвоты клеток перед выработкой антител.
Третий подход заключается в использовании высокопроизводительного способа, в котором трансфекцию клеток-хозяев проводят по отдельности с применением индивидуальных векторов, составляющих библиотеку когнатных пар. Этот способ называется индивидуальной трансфекцией. При индивидуальной трансфекции предпочтительно проводить отбор клеток-хозяев для сайт-специфической интеграции по отдельности. Индивидуальные клоны клеток, полученные в результате отбора, можно проанализировать на время пролиферации, и для создания запаса библиотеки поликлональных когнатных пар предпочтительно использовать клоны со сходной скоростью роста. Для получения желательной поликлональной клеточной линии индивидуальные клоны клеток можно смешать перед созданием запаса сразу же после их извлечения из исходного запаса или после короткого периода пролиферации и адаптации. Такой подход помогает элиминировать какое-либо остаточное смещение последовательности во время трансфекции, интеграции и отбора. В альтернативном варианте индивидуально трансфицированные клетки-хозяева перемешивают перед проведением отбора, что позволяет устранять смещение последовательности вследствие трансфекции.
Общая особенность технологических концепций, охарактеризованных выше, заключается в том, что все индивидуальные когнатные пары, составляющие рекомбинантный поликлональный белок, можно получить в одном биореакторе или в немногих биореакторах. Единственное различие связано со стадией, на которой выбирают создание коллекции клеток, составляющих поликлональную производительную клеточную линию.
Один из вариантов осуществления настоящего изобретения заключается в популяции клеток-хозяев, содержащей когнатную библиотеку или вспомогательную библиотеку сцепленных пар кодирующих последовательностей вариабельной области.
В следующем варианте осуществления настоящего изобретения популяция клеток-хозяев содержит библиотеку, полученную из совокупности выделенных единичных клеток (лимфоцитов), с применением амплификации в мультиплексной RT-PCR и последующим сцеплением посредством сшивания или рекомбинации либо по технологии мультиплексной RT-PCR перекрывания-удлинения в соответствии с настоящим изобретением для сцепления когнатных пар.
Еще один вариант осуществления настоящего изобретения заключается в популяции клеток-хозяев, содержащей комбинаторную библиотеку или вспомогательную библиотеку сцепленных пар кодирующих последовательностей вариабельной области.
В соответствии с настоящим изобретением популяция клеток-хозяев охватывает разнообразную популяцию клеток, соответствующую разнообразию библиотеки, которой были трансформированы/трансфицированы эти клетки. Предпочтительно, чтобы каждая клетка в указанной популяции представляла только одну когнатную пару из всей библиотеки когнатных пар и ни один из отдельных элементов библиотеки когнатных пар не превышал 50%, предпочтительнее 25%, еще предпочтительнее 10%, от общего числа отдельных элементов, экспрессируемых из популяции клеток-хозяев.
В предпочтительном варианте осуществления настоящего изобретения популяция клеток-хозяев представлена клетками млекопитающего.
Как описано выше, популяцию клеток-хозяев можно использовать для экспрессии рекомбинантного поликлонального связывающего белка, поскольку отдельные клетки этой популяции представляют кодирующие последовательности вариабельной области с несходным разнообразием.
Одним из вариантов осуществления настоящего изобретения является рекомбинантный поликлональный белок, который экспрессируется из популяции клеток-хозяев, содержащей библиотеку векторов, кодирующих разные когнатные пары сцепленных кодирующих последовательностей вариабельной области, причем эта библиотека получена способом, предлагаемым настоящим изобретением. Типичный рекомбинантный поликлональный белок, соответствующий настоящему изобретению, состоит, по меньшей мере, из 2, 5, 10, 20, 50, 100, 1000, 104, 105 или 106 белков, происходящих из различных когнатных пар.
Предпочтительным вариантом осуществления настоящего изобретения является рекомбинантный поликлональный иммуноглобулин, который экспрессируется из популяции клеток-хозяев, содержащей библиотеку векторов, кодирующих различные когнатные пары кодирующих последовательностей вариабельной области тяжелой цепи и вариабельной области легкой цепи.
Еще одним предпочтительным вариантом осуществления настоящего изобретения является рекомбинантный поликлональный TcR, экспрессируемый из популяции клеток-хозяев, которая содержит библиотеку векторов кодирующих различные когнатные пары кодирующих последовательностей вариабельной области альфа-цепи TcR, сцепленных с кодирующими последовательностями вариабельной области бета-цепи и/или кодирующих последовательностей вариабельной области гамма-цепи TcR, сцепленных с кодирующими последовательностями вариабельной области дельта-цепи.
Еще одним вариантом осуществления настоящего изобретения является клетка-хозяин, пригодная для выработки моноклонального белка. В особенности это относится к моноклональному антителу, содержащему когнатную пару вариабельной области легкой цепи с вариабельной областью тяжелой цепи, либо к моноклональному TcR, состоящему из когнатной пары вариабельной области альфа с вариабельной областью бета или вариабельной области дельта с вариабельной областью гамма. Предпочтительно, чтобы такая моноклональная продуктивная клеточная линия не была клеточной линией гибридомы.
Такие моноклональное антитело или TcR можно создать, добавив к способу сцепления множества представляющих интерес несмежных нуклеотидных последовательностей следующие стадии:
a) вставление указанных последовательностей нуклеиновой кислоты в вектор,
b) введение указанного вектора в клетку-хозяина,
c) культивирование указанных клеток-хозяев в условиях, благоприятных для экспрессии,
d) получение белкового продукта, который экспрессирован из вектора, вставленного в указанную клетку-хозяина. Предпочтительно, чтобы вектор, введенный в клетку-хозяина, кодировал отдельную когнатную пару кодирующих последовательностей вариабельной области.
ВОЗМОЖНОСТИ ПРИМЕНЕНИЯ ИЗОБРЕТЕНИЯ
Одно из основных применений настоящего изобретения заключается в сцеплении когнатных пар кодирующих последовательностей вариабельных областей, особенно кодирующих последовательностей вариабельной области тяжелой и легкой цепи иммуноглобулина либо кодирующих последовательностей вариабельной области альфа-цепей и бета-цепей TcR или гамма-цепей и дельта-цепей TcR посредством высокопроизводительного способа генерирования библиотек когнатных пар. В дополнение к созданию библиотек когнатных пар предлагаемые настоящим изобретением методики мультиплексной RT-PCR с последующим сцеплением посредством сшивания или рекомбинации либо мультиплексной RT-PCR перекрывания-удлинения можно использовать для создания комбинаторных библиотек химерных антител человека/неродственных животных, применяя методику, предлагаемую изобретением, на популяции генетически различных клеток, на клеточных лизатах из такой клеточной популяции или на РНК, очищенной из такой клеточной популяции. Библиотеки, вспомогательные библиотеки или единичные клоны из какой-либо библиотеки такого рода облегчают экспрессию поликлональных или моноклональных белков. Из библиотек, предлагаемых настоящим изобретением, в особенности, можно получать моноклональные или поликлональные антитела. Применение рекомбинантных моноклональных антител в диагностике, лечении и профилактике известно достаточно хорошо. Рекомбинантные моноклональные и поликлональные антитела, созданные в соответствии с настоящим изобретением, будут иметь такую же сферу применения, как и продукты из категории антител, получаемые на основе существующих технологий. Посредством настоящего изобретения, в особенности, можно производить фармацевтические композиции, содержащие в качестве активного ингредиента поликлональный рекомбинантный иммуноглобулин, скомбинированный, по меньшей мере, с одним фармацевтически приемлемым наполнителем. Более предпочтительны такие фармацевтические композиции, в которых поликлональный рекомбинантный иммуноглобулин генерируется из когнатных пар кодирующих последовательностей вариабельной области. Такие фармацевтические композиции поликлональных рекомбинантных иммуноглобулинов можно применять в качестве медикаментов. Поликлональный рекомбинантный иммуноглобулин в такой композиции может быть специфичным или реактивным по отношению к заданной патологической цели, таким образом, указанную композицию можно применять для лечения, облегчения или предупреждения таких заболеваний, как рак, инфекции, воспалительные заболевания, аллергия, астма и другие респираторные заболевания, аутоиммунные заболевания, иммунологические расстройства, сердечно-сосудистые заболевания, заболевания центральной нервной системы, метаболические и эндокринные заболевания, отторжение трансплантата или нежелательная беременность у таких млекопитающих, как человек, сельскохозяйственные животные или домашние животные.
ПРИМЕРЫ
Пример 1:
Мыши Balb/c были иммунизированы подкожно 50 мкг столбнячного анатоксина (TT) в полном адъюванте Фрейнда. Иммунизацию мышей форсировали на 14-й день введением 50 мкг TT в неполном адъюванте Фрейнда. Еще через 30 дней иммунизацию мышей дополнительно форсировали введением 50 мкг TT в неполном адъюванте Фрейнда. Через три дня после заключительной стимуляции иммунного ответа мышей умерщвляли, извлекали из них селезенки и переносили материал в пробирки, содержащие 30 мл RPMI 1640 вес/10% FCS при 4°C. Ткань переносили на клеточный фильтр 74 мкм (Corning, 136350-3479) в 10-см чашку. Ткань селезенки мацерировали через фильтр задней стороной поршня шприца. Фильтр промывали 10 мл раствора RPMI 1640, 10% FCS. Затем фильтр извлекали, а чашку наполняли 20 мл холодного раствора RPMI 1640, 10% FCS. Клетки переносили в пробирку 50 мл и центрифугировали со скоростью 300×g при 2-8°C в течение 5 минут. Затем клетки ресуспендировали в 5-10 мл RPMI 1640 вес/1% FCS при 4°C и фильтровали через шприцевой фильтр калибра 50 мкм (Becton Dickinson, 340603). Клетки осаждали и ресуспендировали в FCS с 10% DMSO, чтобы получить плотность 2×107 клеток на ампулу, после чего их замораживали.
Замороженные ампулы со спленоцитами в одноклеточной суспензии от мышей Balb/c, иммунизированных столбнячным анатоксином, оттаивали при 37°C и переносили в пробирки емкостью 15 мл с нерастаявшими остатками льда. В пробирку при постоянном раскручивании добавляли по каплям 10 мл охлажденного на льду раствора RPMI, 10% FCS. После однократного отмывания в 10 мл FACS PBS, добавляли 5 мл PBS, 2% FCS перед фильтрованием клеток через фильтр Filcon калибра 50 µm (Becton Dickinson, № по каталогу 340603). Клетки осаждали, затем ресуспендировали в 1 мл PBS, 2% FCS (конечный объем) и окрашивали или FITC anti-CD43 в разведении 1:100 (BD, № по каталогу 553270) и PE anti-CD138 в разведении 1:40 (BD, № по каталогу 553714) в 1 мл PBS, 2% FCS, или APC anti-B220 (BD, № по каталогу 553092) в разведении 1:40 и FITC anti-MHCII (BD, № по каталогу 553547) в разведении 1:200. Клетки инкубировали при 4°C в течение 20 минут в темноте. Наконец, клетки дважды отмывали 2 мл PBS, 2% FCS и добавляли до объема 15 мл PBS, 2% FCS (эмбриональной телячьей сыворотки). Непосредственно перед сортингом добавляли PI в соотношении 1:100, а затем проводили сортинг клеток в счетчике событий со скоростью приблизительно 1000-2000 клеток в секунду. Пропускание для обоих вариантов окрашивания отображено на фиг.3.
Фиг.3A: PI-позитивные (мертвые) клетки были исключены на нижней левой панели (P1). Затем плазмациты были отсортированы как CD43 high, CD138 high на нижней правой панели (P2). Наконец, дубликаты были исключены на графике SSC-H по SSC-W (верхняя правая панель) (P3). Клетки, позитивные по всем трем проходам, были отсортированы в планшеты ELISPOT в соответствии с данными, представленными в таблице 1.
ELISPOT:
96-луночные планшеты с дном из нитроцеллюлозы (планшеты HA, Millipore, Bedford, MA) были предварительно увлажнены PBS (в том числе блокирующие лунки) перед покрытием 100 мкл (25 мкг/мл) столбнячного анатоксина (TT) или овечьего противомышиного IgG (Jackson Immuno Research, № по каталогу 515-005-062) в PBS. В контрольные лунки вводили такой же объем PBS. Планшеты выдерживали при температуре 4°C. На следующий день лунки троекратно отмывали PBS перед блокированием 200 мкл RPMI+2% снятого порошкового молока и оставляли при температуре 4°C. За 1 час до добавления клеток планшет помещали в инкубатор (37°C, 5% CO2, влажность 100%). В лунки, покрытые TT-, anti-IgG и только заблокированные, добавляли 100 мкл клеток в полной среде RPMI. В качестве контроля использовали питательную среду без клеток. Планшет помещали в инкубатор (37°C, 5% CO2, влажность 100%). На следующий день планшет шестикратно отмывали для удаления клеток в буфере 3х: PBS+0,01% Tween 20 и 3х в PBS. Впоследствии в лунки добавляли HRP-конъюгированный козий противомышиный IgG (Caltag M30007), разведенный 1:3000 в RPMI+2% снятого порошкового молока (100 мкл на лунку). После 2-часовой инкубации при 37°C лунки троекратно отмывали PBS+0,01% tween 20 с последующим троекратным отмыванием чистым PBS. Затем пятна разрабатывали, используя 100 мкл хромогенного субстрата, состоящего из 0,015% H2O2 и 0,3 мг/мл 3-амино-9-этилкарбазола в 0,1 M ацетате натрия, pH 5,1. Развитие окраски останавливали отмыванием проточной водой в течение 5 минут.
Судя по результатам, представленным в таблице 1, TT-специфичные плазмациты (PC, CD43 high, CD138 high, проход P3 на фиг.3A) представлены на уровне приблизительно 2%, тогда как TT-специфичные плазмабласты (PB, позитивные на B220 и MHCII, проход P4 на фиг.3B) представлены на уровне приблизительно 4%. Это иллюстрирует превосходство PC над PB в выработке специфических антител после иммунизации TT.
Пример 2:
Замороженную ампулу со спленоцитами окрашивали, как это описано в примере 1.
Четыре разных фенотипа были отсортированы четырехсторонним образом. Проходы сортинга отображены на фиг.4. Прежде всего PI-позитивные или мертвые клетки были исключены на нижней левой панели (P1). P2 означает CD138-промежуточные, CD43-высокие. P3 означает CD138-высокие, CD43-высокие. P4 означает CD138-высокие, Cd43-отрицательные. P5 означает CD138-промежуточные, CD43-низкие. 10000 клеток, позитивных по P1 и каждому из четырех проходов, были отсортированы в пробирки и заморожены для оценки в мышиной системе Symplex.
Фракции P2, P3, P4 и P5 были отсортированы в общей массе в пробирки (по 10000 клеток в каждой). Пробирки были центрифугированы с последующим ресуспендированием осадка в среде Игла с модификацией по Дульбекко, содержавшей 2 ед./мкл ингибитора РНК-азы (RNasin, Promoga, № по каталогу N2511) до концентрации 250 клеток на мкл (10 мкл на пробирку) и замораживанием при -80°C.
Амплификацию в мышиной системе Symplex проводили на серии разведений каждого из 4 наборов отсортированных лимфоцитов для сравнения по содержанию мРНК антитела IgG-каппа. Основные принципы реакций таковы.
- Сначала проводят реакцию RT, в которой затравкой для тяжелой и легкой цепи служат специфические праймеры константной области.
- Второй по очереди проводят мультиплексную реакцию с применением праймеров 5' области VH и VK, охватывающих все вариабельные области и оснащенных комплементарными выступами, облегчающими формирование зон перекрывания. 3'-праймеры расположены в константном области тяжелой и легкой цепи.
- Последней по очереди проводят сложенную реакцию, амплифицирующую только конъюгированные VH и VK с использованием праймеров JH и JK.
- Конечный продукт реакции состоит из VH и VK, конъюгированных между 5'-концом одного и 5'-концом другого и соединенных линкером. Размер должен составлять приблизительно 700 bp.
Для комбинированной мультиплексной реакции RT-PCR был использован набор праймеров, показанный в таблице 2, а также набор Qiagen OneStep RT-PCR, по существу, в соответствии с инструкциями производителя. Замороженные клетки оттаивали на льду, ресуспендировали и центрифугировали. В каждой серии разведений был использован клеточный лизат, соответствующий 100, 32, 10, 3,2, 1, 0,32, 0,1 и 0 клеток. Общий объем реакции составлял 20 мкл. Условия циклов были таковы:
Вложенную реакцию проводили с набором праймеров, показанным в таблице 3, используя полимеразу FastStart (Roche) и прилагаемые к ней реактивы, по существу, в соответствии с инструкциями производителя. Для вложенной реакции использовали 1 мкл продукта реакции RT-PCR в общем объеме 20 мкл. Условия реакции были таковы:
В конце 10 мкл каждого конечного продукта реакции анализировали на 1% агарозном геле.
Судя по результатам, полученным в системе Symplex на титрованных клеточных лизатах (фиг.5), очевидно, что мы можем проводить сцепление вариабельных областей тяжелой и легкой цепи с применением клеток из P3, понижая концентрацию лизата приблизительно до 0,1 клетки, а с клетками из P2 при снижении концентрации лизата, начиная от 3,2 клетки. Другие проходы оказались менее эффективными, то есть сцепление было возможно только при концентрации лизата, соответствующей приблизительно 32 клеткам и более. В заключение можно констатировать, что клетки CD43 high CD138 high (P3) наиболее полезны в системе Symplex™ на уровне одной клетки, тогда как клетки CD43 high CD138 intermediate можно использовать, но с меньшей эффективностью.
Конц. - конечная концентрация.
Праймеры для вложенной PCR в мышиной системе Symplex
(nM)
ПРИМЕР 3. Клонирование антител против-EGFR
Иммунизация
Для иммунизации посредством инъекций различных очищенных белков в дополнение к клеткам, избыточно экспрессирующим EGFR, были использованы самки мышей BALB/c, линии A или C57B16 (в возрасте 8-10 недель).
В некоторых случаях для иммунизации были использованы поступающие в продажу белки EGFR (R&D systems, № по каталогу 1095-ER или Sigma № E3641). В других случаях для иммунизации были использованы рекомбинантные EGFR и EGFRvIII человека, выработанные как гибридные белки, состоящие из ECD EGFR или EGFRvIII, и гормон роста человека (hGH), также включая сайт расщепления вируса гравировки табака (TEV) в дополнение к His-tag. В некоторых случаях ECD EGFR выделяли посредством расщепления TEV-протеазой с последующей очисткой в никелевой колонке.
Для клеточной иммунизации была использована линия клеток рака головы и шеи человека HN5 (Easty DM, Easty GC, Carter RL, Monaghan P, Butler LJ. Br J Cancer. 1981 Jun;43(6):772-85. Ten human carcinoma cell lines derived from squamous carcinomas of the head and neck.), экспрессирующая приблизительно 107 рецепторов на клетку. Клетки были культивированы в среде DMEM в добавками 10% FBS (эмбриональной телячьей сыворотки), 3 mM глицерина, 5 mM пирувата натрия и 1% пенициллина/стрептомицина. Перед каждой иммунизацией клетки отмывали в PBS, трипсинизировали ферментом TrypLE и ресуспендировали в питательной среде для выращивания. Затем клеточные суспензии дважды отмывали в PBS при центрифугировании на скорости 250Xg в течение 5 минут с выбиванием клеток и ресуспендированием в 15 мл стерильного PBS.
Клетки или антиген разбавляли в PBS, после чего смешивали в соотношении 1:1 с адъювантом Фрейнда. Адъювант применяют для усиления и модуляции иммунного ответа. Для первых иммунизаций использовали полный адъювант Фрейнда (CFA), а для последующих иммунизаций использовали неполный адъювант Фрейнда. IFA представляет собой эмульсию типа масло в воде, состоящую из минеральных масел, а CFA представляет собой IFA, к которому добавлены термически убитые и высушенные микобактерии. Оба адъюванта обладают депонирующим эффектом. CFA дает начало длительной персистенции иммунного ответа, и его применяют при первых иммунизациях для стимуляции иммунного ответа, а IFA применяют для последующих иммунизаций. Проверку эмульсий осуществляли, нанося каплю эмульсии на поверхность стекла с водой. Если капля остается каплей, то эмульсия стабильна и ее можно вводить посредством инъекций. Мышам вводили только стабильные эмульсии.
В зависимости от плана (см. таблицу 4) для каждой инъекции использовали 25-100 мкг антигена или 107 клеток. В общей сложности, мыши получали 4 инъекции. Всем мышам впрыскивали или 300 мкл или 200 мкл эмульсии. В зависимости от плана инъекции проводили подкожно (s.c), внутрибрюшинно (i.p.) или внутривенно (i.v.).
В конце эксперимента мышей умерщвляли, сворачивая им шею, после чего извлекали селезенки и переносили их на клеточное сито с размером ячеек 74 мкм (Corning#136350-3479). Клетки мацерировали через фильтр, ресуспендировали в холодной среде RPMI 1640 с 10% FBS и центрифугировали на скорости 300Xg в течение 5 минут. Клеточный осадок ресуспендировали в среде RPMI 1640 с 1% FBS, фильтровали через шприцевой фильтр с размером ячеек 50 мкм (BD# 340603) и накапливали центрифугированием. Клеточный осадок криоконсервировали после ресуспендирования в FCS с 10% DMSO, а замороженные клетки хранили при температуре -80°C вплоть до сортинга мот методу FACS.
Сортинг мышиных плазматических клеток методом FACS
Флаконы с замороженными спленоцитами оттаивали при 37°C и переносили в 15 мл пробирку, где все еще находился лед. В пробирку при постоянном покручивании добавляли по каплям 10 мл охлажденной на льду среды RPMI, 10% FBS (эмбриональной телячьей сыворотки). После однократного отмывания в 10 мл FACS PBS добавляли 5 мл FCS PBS перед фильтрованием клеток через Filcon с размером ячеек 50 мкм. Затем клетки осаждали центрифугированием, ресуспендировали в 1 мл PBS с 2% FBS (конечный объем) и окрашивали anti-CD43-FITC и anti-CD138-PE в соответствии со специфичным разбавлением до конечной концентрации приблизительно 5 мкг/мл. Клетки инкубировали при 4°C в течение 20 минут в темноте. Затем клетки дважды отмывали в 2 мл буфера FACS. Добавляли до 15 мл FACS PBS. Пропидиум иодид (PI) добавляли в соотношении 1:100, после чего клетки сортировали в 96-луночных планшетах PCR, содержавших реакционный буфер PCR (см. ниже), и центрифугировали в течение 2 минут на скорости 400Xg перед замораживанием планшетов при температуре -80°C. Плазмациты пропускали как CD43-позитивные/CD138-позитивные.
Сцепление когнатных пар V H и V L
Сцепление кодирующих последовательностей VH и VL проводили на одиночных клетках, отсортированных как плазмациты, что облегчало когнатное спаривание кодирующих последовательностей VH и VL. При этом была использована двухстадийная процедура PCR, основанная на одностадийной мультиплексной RT-PCR перекрывания-удлинения с последующей стадией вложенной PCR. Смеси праймеров, использованные в настоящем примере, амплифицируют только легкие каппа-цепи. Однако по желанию к мультиплексной смеси праймеров и к смеси праймеров вложенной PCR можно добавить праймеры, способные амплифицировать легкие лямбда-цепи. Если добавляют праймеры лямбда, то процедуру сортинга следует адаптировать так, чтобы не исключались лямбда-позитивные клетки. Принцип сцепления когнатных последовательностей VH и VL проиллюстрирован на фиг.1.
Полученные 96-луночные планшеты для PCR оттаивали, а отсортированные клетки служили матрицей для мультиплексной RT-PCR перекрывания-удлинения. Сортировочный буфер добавляли в каждую лунку перед реакционным буфером для сортинга одиночных клеток (OneStep RT-PCR Buffer; Qiagen), праймерами для RT-PCR (см. таблицу 2 выше) и ингибитором РНК-азы (RNasin, Promega). Это дополняли смесью ферментов для одностадийной RT-PCR (25-кратное разведение, Qiagen) и смесью dNTP (по 200 мкм каждой) для получения заданной конечной концентрации в реакционном объеме 20 мкл. Планшеты инкубировали в течение 30 минут при 55°C для получения обратной транскрипции РНК из каждой клетки. После проведения RT планшеты проводили через следующий цикл PCR: 10 минут при 94°C, 35×(40 секунд при 94°C, 40 секунд при 60°C, 5 минут при 72°C), 10 минут при 72°C.
Реакции PCR проводили в термоциклере H20BIT с корзинкой со съемным уплотнением для 24 96-луночных планшетов (ABgene) для повышения пропускной способности. После циклической обработки планшеты для PCR хранили при -20°C.
Для проведения стадии вложенной PCR 96-луночные планшеты для PCR готовили со следующей смесью в каждой лунке (реакции на 20 мкл), чтобы получить заданную конечную концентрацию: 1х буфера FastStart (Roche), смесь dNTP (200 каждого компонента), смесь вложенных праймеров (см. таблицу 5), ДНК-полимераза Phusion (0,08 U, Finnzymes) и высококачественная смесь ферментов FastStart (0,8 U, Roche). В качестве матрицы для вложенной PCR переносили 1 мкл материала из реакций мультиплексной PCR перекрывания-удлинения. Планшеты для вложенной PCR проводили через следующие термоциклы: 35×(30 секунд при 95°C, 30 секунд при 60°C, 90 секунд при 72°C), 10 минут при 72°C.
Для верификации присутствия фрагментов перекрывания-удлинения длиной приблизительно 890 пар оснований (bp) случайно отобранные реакции были проанализированы на 1% агарозном геле.
Планшеты хранили при -20°C до следующего процессинга фрагментов PCR. Репертуар сцепленных кодирующих пар VH и VL из вложенной PCR накапливали, не смешивая пары от разных доноров, и очищали электрофорезом в препаративном 1% агарозном геле. Кодирующая последовательность константной легкой каппа-цепи человека была сплайсирована посредством перекрывающегося удлинения в кодирующий область VL объединенных продуктов PCR сцепленных кодирующих пар VH и VL (фиг.2). Кодирующая последовательность константной легкой каппа-цепи человека была амплифицирована из плазмиды, содержащей кодирующую последовательность антитела человека с легкой каппа-цепью, в реакции, содержащей: фермент Phusion (2 U, Finnzymes), 1х буфера Phusion, смесь dNTP (200 мкм каждого компонента), праймер hKCforw-v2 и праймер Kappa3' (таблица 6), а также плазмидный шаблон pLL138 (10 нг/мкл) в общем объеме 50 мкл. Реакцию проводили следующими термоциклами: 25×(30 секунд при 95°C, 30 секунд при 55°C, 45 секунд при 72°C), 10 минут при 72°C. Полученный в результате PCR фрагмент очищали электрофорезом в препаративном 1% агарозном геле.
Очищенные объединенные фрагменты PCR из каждого репертуара сплайсировали в амплифицированный и очищенный фрагмент PCR константной кодирующей каппа-области человека (приложение 1) при последующем сплайсинге через PCR с перекрыванием-удлинением (общий объем 50 мкл), содержащую: фрагмент кодирующей константной каппа-области человека (1,4 нг/мкл), очищенный накопленный фрагмент PCR (1,4 нг/мкл), связующую ДНК-полимеразу (0,5 U, Finnzymes), высококачественную смесь ферментов FastStart (0,2 U, Roche), 1х буфера FastStart (Roche), смесь dNTP (200 мкм каждого компонента), праймер mhKCrev и набор праймеров mJH (см. таблицу 6). Реакцию проводили следующими термоциклами: 2 минуты при 95°C, 25×(30 секунд при 95°C, 30 секунд при 55°C, 1 минута при 72°C), 10 минут при 72°C. Полученный в результате PCR фрагмент (длиной приблизительно 1070 bp) очищали электрофорезом в препаративном 1% агарозном геле.
Инсерция когнатных кодирующих пар V H и V L в вектор скрининга
Для того чтобы идентифицировать антитела, обладающие специфичностью связывания с EGFR, полученные кодирующие последовательности VH и VL были экспрессированы в виде полноразмерных антител. Это включало инсерцию репертуара кодирующих пар VH и VL в вектор экспрессии и трансфекцию в клетку-хозяина.
Для генерирования репертуара векторов экспрессии, содержащих сцепленные кодирующие пары VH и VL, была использована двухстадийная процедура клонирования. Со статистической точки зрения, если репертуар векторов экспрессии содержит в десять раз больше рекомбинантных плазмид, чем число когнатных спаренных VH и VL, как продуктов PCR, использованных для генерирования репертуара скрининга, то существует 99% вероятность того, что представлены все уникальные пары генов. Таким образом, если было получено 400 фрагментов V-гена с перекрыванием-удлинением, то репертуар, генерированный для скрининга, составит, по меньшей мере, 4000 клонов.
Вкратце, очищенный продукт PCR из репертуаров сцепленных кодирующих пар VH и VL, сплайсированный в кодирующую константную каппа-область человека, был расщеплен ДНК-эндонуклеазами XhoI и NotI в сайтах распознавания, введенных в концы продуктов PCR. Расщепленные и очищенные фрагменты были лигированы в вектор экспрессии IgG млекопитающих, расщепленный XhoI/NotI, OO-VP-002 (фиг.6) посредством стандартных процедур лигирования. Лигирующая смесь была электропорирована в E. coli, добавлена в 2×YT планшеты, содержащие подходящий антибиотик, и инкубирована при 37°C в течение ночи. Амплифицированный репертуар векторов был очищен из клеток, восстановленных из планшетов с применением стандартных способов очистки ДНК (Qiagen). Плазмиды были подготовлены для инсерции фрагментов промотор-лидер посредством расщепления эндонуклеазами AscI и NheI. Сайты рестрикции для этих ферментов были расположены между парами генов, кодирующих VH и VL. После очистки вектора двунаправленный фрагмент промотор-лидер млекопитающих, расщепленный AscI-NheI, был вставлен в сайты рестрикции AscI и NheI посредством стандартных процедур лигирования. Лигированный вектор был амплифицирован в E. coli, а плазмида была очищена стандартными способами. Генерированный репертуар векторов скрининга был трансформирован в E. coli посредством общепринятых процедур. Полученные колонии были объединены в 384-луночные мастер-планшеты и заложены на хранение. Число выстроенных колоний превышало число входных продуктов PCR, по меньшей мере, в 3 раза, что давало 95-процентную вероятность присутствия всех полученных уникальных пар V-генов.
Плазмиду ДНК приготовили из клонов, отобранных выше, и клетки CHO-S (Invitrogen) трансфицировали в 2-мм масштабе для экспрессии антител (в соответствии с инструкциями производителя). Через 96 часов после трансфекции проводили сбор супернатантов.
СКРИНИНГ НА СВЯЗЫВАНИЕ С ВНЕКЛЕТОЧНЫМ ДОМЕНОМ EGFR
В целом скрининг был проведен как двухстадийная процедура. Библиотеки антител были скринированы на реактивность по отношению к рекомбинантному белку EGFR по методике ELISA, после чего способ FMAT (FLISA) был использован как подход на клеточной основе с клеточной линией NR6wtEGFR (Batra et al., 1995, Cell Growth Differ, 6(10):1251-9), для выявления антител против EGFR, связывающихся с EGFR, который экспрессируется на клеточной поверхности. Для библиотек 101 и 108/109 (таблица 4) анализ по методике ELISA был проведен с рекомбинантным EGFR, представляющим внеклеточный домен EGFR.
Вкратце, для проведения ELISA планшеты Nunc maxisorb (№ по каталогу 464718) были покрыты 1 мкг/мл белка (собственного производства), разбавленного в PBS и выдержанного при 4°C в течение ночи. Перед блокированием в 50 мкл 2%-MiIk-PBS-T планшеты были однократно отмыты смесью PBS+0,05% Tween 20 (PBS-T). Планшеты однократно отмывали PBS-T, затем добавляли 20 мкл 2%-milk-PBS-T и 5 мкл супернатантов из трансфектантов Freestyle CHO-S (см. выше), проводили инкубацию материала в течение 1,5 часа при комнатной температуре, после чего планшеты однократно отмывали PBS-T из расчета 20 мкл на лунку. Для выявления антител, связанных с лунками, было добавлено вторичное антитело (HRP-Goat-anti-human IgG, Jackson, № по каталогу 109-035-097), разбавленное в соотношении 1:10000 в 2% milk-PBS-T с последующей инкубацией в течение 1 часа при комнатной температуре. Шаблоны однократно отмывали в PBS-T перед добавлением 25 мкл субстрата (Kem-en-tec Diagnostics, № по каталогу 4390) и инкубацией материала в течение 5 минут. После инкубации добавляли 25 мкл 1 M серной кислоты для остановки реакции. Специфический сигнал выявляли считывающим устройством ELISA на длине волны 450 нм.
Для определения антител против EGFR на клеточной основе по методике FMAT клетки SKBR-3 (ATCC #HTB-30) или NR6wtEGFR содержали в питательной среде для роста. Клетки подсчитывали и разбавляли до концентрации 125000 клеток на миллилитр антителом Alexa-647-конъюгированный козий IgG против человека (H-L) (молекулярный зонд № A21445, партия № 34686A) при разведении 1:40000. Общее количество 20 мкл этой суспензии переносили в 384-луночные планшеты Nunc с прозрачным дном. Впоследствии к клеткам добавляли 10 мкл трансфекционного супернатанта. Сигнал FMAT из реакции измеряли после 6-10 часов инкубации.
Данные скрининга указывают на то, что 221 клон (4,8% от общего числа клонов) оказался позитивным в ELISA. 93 (2,0%) этих клонов также были позитивными в анализе FMAT. В целом 220 (4,8%) клонов были позитивными в анализе FMAT, а среди них 127 (220-93) уникально позитивными на антиген клеточной поверхности. Библиотека 111 была скринирована сходным образом, но, поскольку процедура иммунизации была направлена на выработку антител, специфичных по отношению к делеционному мутанту рецептора EGFR (EGFRvIII), скрининговые тесты ELISA включали анализы для выявления как EGFR дикого типа, так и EGFRvIII. Семь клонов были идентифицированы как специфичные на EGFRvIII в анализах ELISA, причем интересно, что именно эти клоны были негативными при окрашивании клеток, экспрессирующих wtEGFR, в анализе FMAT. 13 клонов в анализах FMAT и ELISA были идентифицированы как позитивные на wtEGFR, но не на EGFRvIII, что оказалось уникальным для этой библиотеки по сравнению с библиотеками 101 и 108/109. Все позитивные клоны по результатам ELISA были отобраны для дальнейшего анализа.
Анализ последовательности и отбор клона
Клоны, идентифицированные как EGFR-специфичные по результатам анализов ELISA, были извлечены из первоначальных мастер-шаблонов (384-луночного формата) и объединены в новых шаблонах. Из клонов выделяли ДНК и подвергали ее секвенированию на V-гены. Последовательности выверяли и отбирали все уникальные клоны. Множественные выверки полученных последовательностей отразили уникальность каждого отдельного клона и позволили идентифицировать уникальные антитела. В результате анализа последовательности 220 клонов были идентифицированы 70 генетически различных кластеров последовательностей антител. Каждый кластер родственных последовательностей, вероятно, происходил в результате соматических гипермутаций общего клона предшественника. В общем, для подтверждения последовательности и специфичности были выбраны один-два клона из каждого кластера.
Валидация последовательности и специфичности
Для того чтобы подтвердить клоны, кодирующие антитела, приготовили плазмиду ДНК и провели трансфекцию клеток FreeStyle CHO-S клетки (Invitrogen) в 2-мл масштабе для экспрессии. Через 96 часов после трансфекции проводили сбор супернатантов. Уровень экспрессии оценивали стандартным анализом ELISA на anti-IgG, а специфичность определяли EGFR- и EGFRvIII-специфическим методом ELISA. В результате было показано, что 85% клонов имеют правильную специфичность и последовательность.
Скрининг на антипролиферативные эффекты
Клеточное повреждение неизбежно приводит к утрате способности клетки поддерживать и обеспечивать энергию для метаболической функции и роста клетки. На этой предпосылке основаны анализы метаболической активности. Обычно они направлены на измерение митохондриальной активности. Реактив клеточной пролиферации WST-1 (Roche, № по каталогу 11644807001) представляет собой готовый к употреблению субстрат, который измеряет метаболическую активность жизнеспособных клеток. Далее допускается, что метаболическая активность коррелирует с количеством жизнеспособных клеток. В этом примере анализ с WST-1 был использован для измерения количества метаболически активных клеток после обработки супернатантами клеточной культуры, содержащими разные антитела anti-EGFR.
Перед проведением анализа WST-1 разные объемы 2-мл супернатантов (0, 10, 25, 50 и 150 мкл) переносили в соответствующие лунки 96-луночного планшета.
Затем клетки HN5 отмывали 1×PBS и разделяли трипсинизацией с использованием 3 мл раствора трипсина. Затем добавляли 17 мл полной среды и клетки центрифугировали на скорости 300xg (1200 rcf) в течение 5 минут. Супернатант удаляли, а клетки ресуспендировали в смеси DMEM+0,5% FBS. Клетки подсчитывали, их концентрацию корректировали и в лунки с супернатантами добавляли 1500 клеток, так, что каждая лунка содержала в общей сложности 200 мкл материала. Планшеты инкубировали 4 дня в увлажненном инкубаторе при 37°C. Затем в каждую лунку добавляли 20 мкл реактива WST-1 и планшеты инкубировали в течение 1 часа при 37°C. Далее планшеты переносили в орбитальный планшетный шейкер и оставляли еще на один час. Поглощательную способность измеряли на уровне 450 и 620 нм (опорная длина волны) считывающим устройством ELISA. Различие в уровне метаболически активных клеток (MAC) вычисляли как процентный показатель по сравнению с контрольными супернатантами исходя из следующей формулы:

Затем эти величины были использованы как основа для контролируемого иерархического кластерного анализа (кластеризация основана на реактивности в ELISA), проведенного с применением бесплатного программного обеспечения Cluster и TreeView.
Предпочтительно иметь возможности для скрининга на функциональные антитела на ранней стадии процесса отбора антител. Супернатанты культур из 83 2-мл трансфекций были использованы для скрининга на функции подавления роста в анализе пролиферации, проведенном с применением клеток HN5 в 0,5% FBS. Результаты были визуализированы посредством простого иерархического кластерного анализа. Как можно увидеть из кластерного анализа (фиг.7), было обнаружено, что многие супернатанты снижают количество метаболически активных клеток HN5 (темно-серый цвет) с эффектом зависимости от концентрации (кластер 2).
Сходным образом, некоторые супернатанты увеличивают количество метаболически активных клеток HN5 (светло-серый цвет) с эффектом зависимости от концентрации (кластеры 1, 3 и 4). Интересное наблюдение заключалось в том, что те супернатанты, которые снижали количество метаболически активных клеток HN5, имели уровень реактивности 2 (черные стрелки), тогда как супернатанты, увеличивающие количество метаболически активных клеток HN5, имели уровень реактивности 1 (серые стрелки). Супернатанты с реактивностью 2 были положительными в анализах ELISA как на wtEGFR, так и на EGFRvIII, тогда как супернатанты с реактивностью 1, были реактивными только в отношении wtEGFR. Таким образом, указанные анализы могут установить связь между реактивностью антител в ELISA и их функциональностью в клеточных анализах.
ВОССТАНОВЛЕНИЕ КЛОНОВ
При использовании мультиплексного подхода PCR ожидается некоторая степень перекрестного примирования внутри- и меж-V-генного семейства вследствие вырожденности праймеров и высокой степени гомологии. Перекрестное примирование вводит в каркас иммуноглобулина аминокислоты, не встречающиеся в природе, что влечет за собой некоторые потенциальные последствия, например структурные изменения и увеличение иммуногенности, причем все они приводят к снижению терапевтической активности.
Для того чтобы устранить эти препятствия и быть уверенным в том, что отобранные клоны воспроизводят естественный гуморальный иммунный ответ, такие перекрестно-примирующие мутации были скорректированы в процессе, называемом репарацией клонов.
На первой стадии процедуры репарации клонов последовательность VH была амплифицирована в PCR с набором праймеров, содержащим последовательность, соответствующую последовательности гена VH из представляющего интерес клона, благодаря чему можно было скорректировать любые мутации, внесенные перекрестным примированием. Фрагмент, полученный в PCR, подвергали расщеплению ферментами XhoI и AscI с последующим обратным лигированием в вектор экспрессии млекопитающих, расщепленный ферментами XhoI/AscI (фиг.6) с применением общепринятых приемов лигирования. Лигированный вектор был амплифицирован в E. coli, а плазмида была очищена стандартными способами. Последовательность VH была секвенирована для верификации исправлений, а вектор был подвергнут расщеплению ферментами NheI/NotI для его подготовки к вставлению легкой цепи.
На второй стадии полноразмерная легкая цепь была амплифицирована в PCR с набором праймеров, содержащим последовательность, соответствующую последовательности гена VL из представляющего интерес родоначального клона, благодаря чему можно было скорректировать любые мутации, внесенные перекрестным примированием. Полученный в PCR фрагмент был расщеплен ферментами NheI/NotI и лигирован в приготовленный ранее вектор, содержащий VH. Продукт лигирования был амплифицирован в E. coli, а плазмида была очищена стандартными способами. Впоследствии легкая цепь была секвенирована для верификации исправлений.
В том случае, если константная каппа-область выбранного клона содержит мутации, внесенные во время амплификации генов, ее замещают константной областью без мутаций. Это осуществляют в PCR с перекрыванием, где репарированный ген VL (амплифицированный без константной области) гибридизируют с константной областью, имеющей правильную последовательность (его получают в отдельной PCR). Всю последовательность амплифицируют и клонируют в вектор, содержащий VH, как это описано выше, после чего исправленную легкую цепь секвенируют для верификации исправлений.

Набор вложенных праймеров
Набор праймеров для сплайсинга константной области каппа
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ СВЯЗЫВАНИЯ ИНТЕРЕСУЮЩИХ ПОСЛЕДОВАТЕЛЬНОСТЕЙ | 2004 |
|
RU2392324C2 |
| ИДЕНТИФИКАЦИЯ АССОЦИИРОВАННЫХ С ОБРАЗЦОМ ПОЛИНУКЛЕОТИДОВ | 2012 |
|
RU2644681C2 |
| КОМПОЗИЦИИ РЕКОМБИНАНТНЫХ АНТИТЕЛ ПРОТИВ РЕЦЕПТОРА ЭПИДЕРМАЛЬНОГО ФАКТОРА РОСТА | 2008 |
|
RU2488596C2 |
| Способ амплификации нуклеиновых кислот тяжелой и легкой цепи человеческих иммуноглобулинов для создания рекомбинантных антител, композиции праймеров (варианты) | 2020 |
|
RU2775914C2 |
| СЕКВЕНИРОВАНИЕ ПОЛИНУКЛЕОТИДНЫХ БИБЛИОТЕК С ВЫСОКОЙ ПРОПУСКНОЙ СПОСОБНОСТЬЮ И АНАЛИЗ ТРАНСКРИПТОМОВ | 2018 |
|
RU2790291C2 |
| СПОСОБ И НАБОР ДЛЯ ВЫЯВЛЕНИЯ ГЕНА, КОДИРУЮЩЕГО МЕМБРАНОСВЯЗАННЫЙ БЕЛОК, ВЕКТОР (ВАРИАНТЫ) | 1999 |
|
RU2246538C2 |
| БЫСТРЫЙ СПОСОБ КЛОНИРОВАНИЯ И ЭКСПРЕССИИ СЕГМЕНТОВ ГЕНА РОДСТВЕННОЙ ВАРИАБЕЛЬНОЙ ОБЛАСТИ АНТИТЕЛА | 2012 |
|
RU2612910C2 |
| СИСТЕМА ИНДИКАЦИИ НА ОСНОВЕ ПОЛНОРАЗМЕРНОГО АНТИТЕЛА ДЛЯ ЭУКАРИОТИЧЕСКИХ КЛЕТОК И ЕЕ ПРИМЕНЕНИЕ | 2012 |
|
RU2625033C2 |
| МЫШИ, У КОТОРЫХ ВЫРАБАТЫВАЮТСЯ СВЯЗЫВАЮЩИЕ БЕЛКИ, СОДЕРЖАЩИЕ VL-ДОМЕНЫ | 2011 |
|
RU2612903C2 |
| АНТИТЕЛА, УЗНАЮЩИЕ УГЛЕВОДСОДЕРЖАЩИЙ ЭПИТОП НА CD43 И СЕА, ЭКСПРЕССИРУЕМЫХ НА РАКОВЫХ КЛЕТКАХ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2008 |
|
RU2528738C2 |














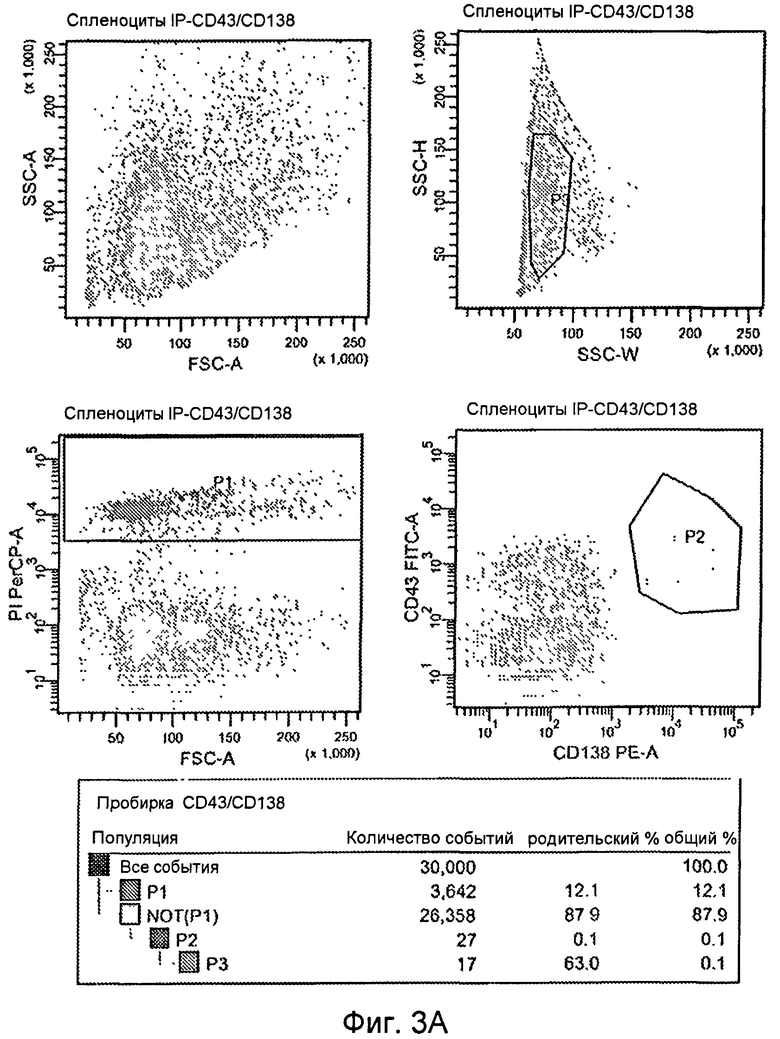


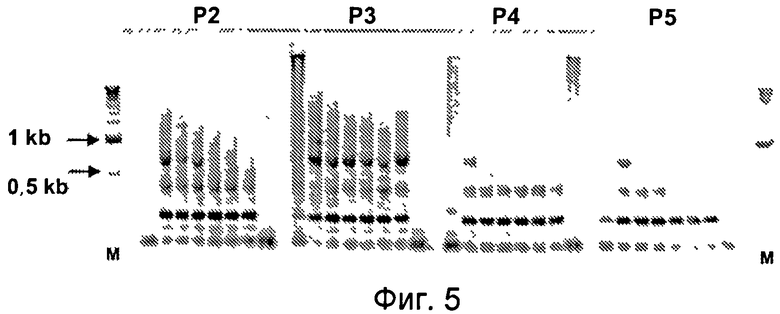


Изобретение относится к области биотехнологии и молекулярной генетики. Способ относится к процедуре сцепления когнатных пар кодирующих последовательностей VH и VL из популяции клеток, обогащенных специфическими поверхностными антигенными маркерами. Процедура сцепления включает мультиплексную процедуру молекулярной амплификации, способную сцеплять представляющие интерес нуклеотидные последовательности в особой полимеразной цепной реакции (мультиплексной PCR). Этот способ особенно эффективен для создания библиотек когнатных пар, а также комбинаторных библиотек, кодирующих последовательностей вариабельных областей из иммуноглобулинов. 24 з.п. ф-лы, 7 ил., 6 табл., 3 пр.
1. Способ получения библиотеки когнатных пар, содержащей сцепленные кодирующие последовательности вариабельной области иммуноглобулина, где указанный способ включает:
a) получение от донора, являющегося грызуном, клеточной фракции, содержащей лимфоциты,
b) получение популяции выделенных единичных клеток путем распределения клеток из указанной клеточной фракции индивидуально во множество резервуаров, где, по меньшей мере, субпопуляция клеток экспрессирует антиген CD43 и CD138 или антиген MHCII и В220,
c) амплификацию и осуществление сцепления кодирующих последовательностей вариабельных областей, содержащихся в указанной популяции выделенных единичных клеток, посредством амплификации в мультиплексной процедуре молекулярной амплификации представляющих интерес нуклеотидных последовательностей с использованием матрицы, полученной из выделенной единичной клетки; и осуществления сцепления представляющих интерес амплифицированных нуклеотидных последовательностей.
2. Способ по п.1, в котором индивидуальные выделенные единичные клетки из популяции единичных клеток размножают до популяций изогенных клеток перед проведением амплификации и сцепления (стадия с).
3. Способ по п.1, в котором клеточная фракция, содержащая лимфоциты, включает спленоциты, цельную кровь, костный мозг, мононуклеарные клетки или лейкоциты, предпочтительно спленоциты или костный мозг, более предпочтительно спленоциты.
4. Способ по п.1, в котором клеточная фракция, содержащая лимфоциты, обогащена плазмацитами или плазмабластами.
5. Способ по п.1, в котором представляющие интерес нуклеотидные последовательности содержат кодирующие последовательности вариабельной области иммуноглобулина, а сцепление создает когнатную пару кодирующей последовательности вариабельной области легкой цепи, связанной с кодирующей последовательностью вариабельной области тяжелой цепи.
6. Способ по п.1, дополнительно включающий перед мультиплексной молекулярной амплификацией оценку того, содержит ли популяция клеток, содержащая лимфоциты, клетки, экспрессирующие антиген CD43 и CD138 или антиген MHCII и В220, предпочтительно CD43 и CD138.
7. Способ по п.1, дополнительно включающий обогащение указанной клеточной фракции, содержащей лимфоциты, популяцией лимфоцитов, экспрессирующих антиген CD43 и CD138 или антиген MHCII и В220, перед мультиплексной молекулярной амплификацией.
8. Способ по п.1, дополнительно включающий выделение из указанной популяции, содержащей лимфоциты, клеток, экспрессирующих антиген CD43 и CD138 или антиген MHCII и В220, перед мультиплексной молекулярной амплификацией.
9. Способ по п.1, в котором выделенные клетки или субпопуляция клеток представляют собой клетки CD138 High/CD43 High или CD138 Intermediate/CD43 High по отношению к клеточной фракции, содержащей лимфоциты.
10. Способ по п.7, в котором обогащение или выделение включает MACS или FACS или другую процедуру автоматического сортинга.
11. Способ по п.1, в котором донором является мышь.
12. Способ по п.11, где донором является трансгенная мышь, которая экспрессирует иммуноглобулины человека.
13. Способ по п.1, в котором указанная процедура мультиплексной молекулярной амплификации представляет собой мультиплексную амплификацию RT-PCR.
14. Способ по п.13, в котором указанная мультиплексная амплификация RT-PCR представляет собой двухстадийный процесс, включающий отдельную стадию обратной транскрипции (RT) перед мультиплексной амплификацией PCR; или в котором указанную мультиплексную амплификацию RT-PCR проводят в одну стадию, включающую в начале добавление всех компонентов, необходимых для осуществления как обратной транскрипции (RT), так и мультиплексной амплификации PCR в одном резервуаре.
15. Способ по п.1, в котором указанное сцепление представляющих интерес нуклеотидных последовательностей проводят в том же самом резервуаре, что и мультиплексную молекулярную амплификацию.
16. Способ по п.15, в котором указанное сцепление представляющих интерес нуклеотидных последовательностей производится в сочетании с мультиплексной амплификацией PCR с применением мультиплексной смеси праймеров перекрывания-удлинения.
17. Способ по п.1, в котором проводится дополнительная молекулярная амплификация с использованием смеси праймеров, адаптированной для амплификации представляющих интерес сцепленных последовательностей нуклеиновой кислоты.
18. Способ по п.16 или 17, в котором мультиплексная смесь праймеров перекрывания-удлинения содержит:
a) по меньшей мере, один праймер mKappal или hmJK, комплементарный смысловой цепи кодирующей последовательности области легкой цепи иммуноглобулина,
b) по меньшей мере, один праймер mVK, комплементарный антисмысловой цепи кодирующей последовательности вариабельной области легкой цепи иммуноглобулина или лидерной последовательности вариабельной области легкой цепи, который способен образовывать набор праймеров с праймером (праймерами) со стадии а),
c) по меньшей мере, один праймер mCHrevl, mHCrevl-ext или mJH, комплементарный смысловой цепи кодирующей последовательности домена тяжелой цепи иммуноглобулина, и
d) по меньшей мере, один праймер mVH, комплементарный антисмысловой цепи кодирующей последовательности вариабельной области тяжелой цепи иммуноглобулина или лидерной последовательности вариабельной области тяжелой цепи, который способен образовывать набор праймеров с праймером (праймерами) со стадии с).
19. Способ по п.1, дополнительно включающий инсерцию сцепленных нуклеотидных последовательностей или библиотеки когнатных пар в вектор.
20. Способ по п.19, в котором сцепленные нуклеотидные последовательности или отдельные члены библиотеки когнатных пар содержат кодирующую последовательность вариабельной области тяжелой цепи иммуноглобулина, ассоциированную с кодирующей последовательностью вариабельной области легкой цепи, и указанные последовательности вставлены в рамку считывания вектора, уже содержащего кодирующие последовательности одного или нескольких доменов константных областей иммуноглобулина или их фрагментов.
21. Способ по п.19, дополнительно включающий создание вспомогательной библиотеки посредством отбора подмножества когнатных пар сцепленных последовательностей вариабельной области, которые кодируют связывающие белки с желаемой специфичностью к мишени, что позволяет создать библиотеку специфичных к мишени когнатных пар кодирующих последовательностей вариабельной области.
22. Способ по п.20 или 21, дополнительно включающий перенос указанной когнатной пары или библиотеки специфных к мишени когнатных пар кодирующих последовательностей вариабельной области в вектор экспрессии млекопитающего.
23. Способ по п.22, в котором вектор экспрессии млекопитающего кодирует один или несколько доменов константной области, которые выбраны из классов иммуноглобулинов человека IgA1, IgA2, IgD, IgE, IgGI, IgG2, IgG3, IgG4, IgM, легкой цепи каппа или легкой цепи лямбда.
24. Способ по п.19, дополнительно включающий следующие стадии:
a) введения вектора, кодирующего сегмент сцепленных нуклеотидных последовательностей, в клетку-хозяина,
b) культивирование указанных клеток-хозяев в условиях, адаптированных для экспрессии, и
c) получение белкового продукта, экспрессированного из вектора, вставленного в указанную клетку-хозяина.
25. Способ по п.24, в котором указанный белковый продукт представляет собой антитело, содержащее когнатную пару вариабельной области легкой цепи, ассоциированную с вариабельной областью тяжелой цепи.
| WO 2005042774, 12.05.2005 | |||
| WO 2007003041, 11.01.2007 | |||
| RU 2006112833, 17.09.2004 | |||
| BABCOOK JS et al., A novel strategy for generating monoclonal antibodies from single, isolated lymphocytes producing antibodies of defined specificities, Proc | |||
| Natl | |||
| Acad | |||
| Sci | |||
| Домовый номерной фонарь, служащий одновременно для указания названия улицы и номера дома и для освещения прилежащего участка улицы | 1917 |
|
SU93A1 |
| ORLANDI R et al., Cloning immunoglobulin variable | |||

