Изобретение относится к биотехнологии, в частности к генной и белковой инженерии, и может быть использовано для получения рекомбинантного соматотропина.
Соматотропин (синонимы: соматропин, гормон роста человека, соматотропный гормон, соматотропный гормон роста) относится к наиболее изученным гормонам гипофиза и принадлежит к белковому семейству, включающему гормон роста, пролактин, ХС. Соматотропин синтезируется в передней доле гипофиза в соматотрофах, которые составляют подкласс ацидофильных клеток гипофиза и являются наиболее многочисленной группой в этой железе. Соматотропин у всех видов млекопитающих представляет собой одиночный пептид с молекулярной массой около 22000. Несмотря на высокую степень гомологии последовательностей гормонов роста различных млекопитающих, в клетках человека активен только собственный гормон роста человека или гормон роста высших приматов. Молекула соматотропина состоит из 191 аминокислотного остатка и содержит два внутримолекулярных дисульфидных мостика. Соматотропин - анаболический гормон, который необходим для постнатального роста, нормализации углеводного, липидного, азотного и минерального обмена, регенерации тканей и выживанию клеток. Ростовые эффекты соматотропина опосредуются главным образом инсулинподобным фактором роста 1 (ИФР-1), ген которого относится к семейству инсулиноподобных генов. Таким образом, соматотропин стимулирует секрецию ИФР-1, дифференцировку и пролиферацию миобластов, усвоение аминокислот и синтез белка в мышцах и других тканях [Roskam W.G., Rougeon F.// Molecular cloning and nucleotide sequence of the human growth hormone structural gene. Nucl.Acids Res., 1979, v. 7, p. 305-320.; Martial J.A., Hallewell R.A., Baxter J.D., Goodman H.M.// Human growth hormone: complementary DNA cloning and expression in bacteria. Science, 1979, v. 205, No 4406, p. 602-607].
Соматотропин, как и другие гормоны, осуществляет свое действие через связывание со специфическим рецептором [Herington AC.// Identification and characterization of growth hormone receptors on isolated rat adipocytes. J Recept Res 1981-82; 2(4):299-316].
Лекарственные формы соматотропина (синонимы - нутропин или протропин (Genentech), нордитропин (Novo Nordisk), генотропин (Pharmacia Upjohn), гуматроп (Eli Lilly), саизен или серостим (Serono)) широко используются в медицине для терапии дефицитности гормона роста и синдрома Тернера.
Известен способ получения соматотропина, состоящий в его выделении из тканей гипофиза человека [Simionescu L., Dimitriu V., Zamfir-Grigorescu D., Aman E., Terbancea M.// The simultaneous isolation of human pituitary hormones. I. Human growth hormone. Endocrinologie 1982 Oct.-Dec.; 20(4):273-83]. Недостатками способа являются сложность с получением исходного материала и риск заражения инфекцией [Rappaport Е.В.// Iatrogenic Creutzfeldt-Jakob disease. Neurology, 1987, Sep.; 37(9):1520-2].
В настоящее время известно два основных способа получения рекомбинантного соматотропина.
Способ, основанный на получении рекомбинантного соматотропина в трансформированных клетках мыши линии С 127 [Carter M.J., Facklam T.J., Long P.C., Scotland R.A.// Are continuous cell lines safe as substrates for human drugs and biologies? A case study with human growth hormone. Dev Biol Stand 1989; 70:101-7]. При таком подходе удается получить гормон роста человека в правильной конформации с хорошей физиологической активностью. Недостатками этого способа являются чрезвычайно низкий выход и длительное время культивации продуцирующих клеток.
А также способ получения соматотропина, включающий экспрессию в клетках Escherichia coli, заключающийся в достаточно быстром биосинтезе клетками бактерий рекомбинантного соматотропина в виде нерастворимых “телец включения” [Olson К.С., Fenno J., Lin N., Harkins R.N., Snider C., Kohr W.H., Ross M.J., Fodge D., Prender G., Stebbing N.// Purified human growth hormone from E. coli is biologically active. Nature 1981 Oct. 1; 293(5831):408-11]. При использовании данного общего способа получения соматотропина уровень биосинтеза целевого белка может колебаться в широких пределах.
Наиболее близким по технической сущности к предлагаемому изобретению является продуцент соматотропина на основе плазмиды pHGH 107 (патент США №5795745, МКИ С 12 В 21/02), несущей под контролем двух последовательных Lac-промоторов ген соматотропина, состоящий из синтетической части (соответствующей аминокислотам 1-24) и фрагмента природного гена (соответствующего аминокислотам 25-191). Использование Lac-промоторов, а также почти полностью (на 88%) природной кДНК, включающей редко встречающиеся в генах Escherichia coli (минорные) кодоны, приводит к существенному снижению уровня биосинтеза белка относительно теоретически возможного.
Изобретение решает задачу конструирования плазмиды, детерминирующей синтез рекомбинантного белка, и создания высокопродуктивного бактериального штамма-продуцента, позволяющего получать рекомбинантный соматотропин с высоким выходом и по упрощенной технологии.
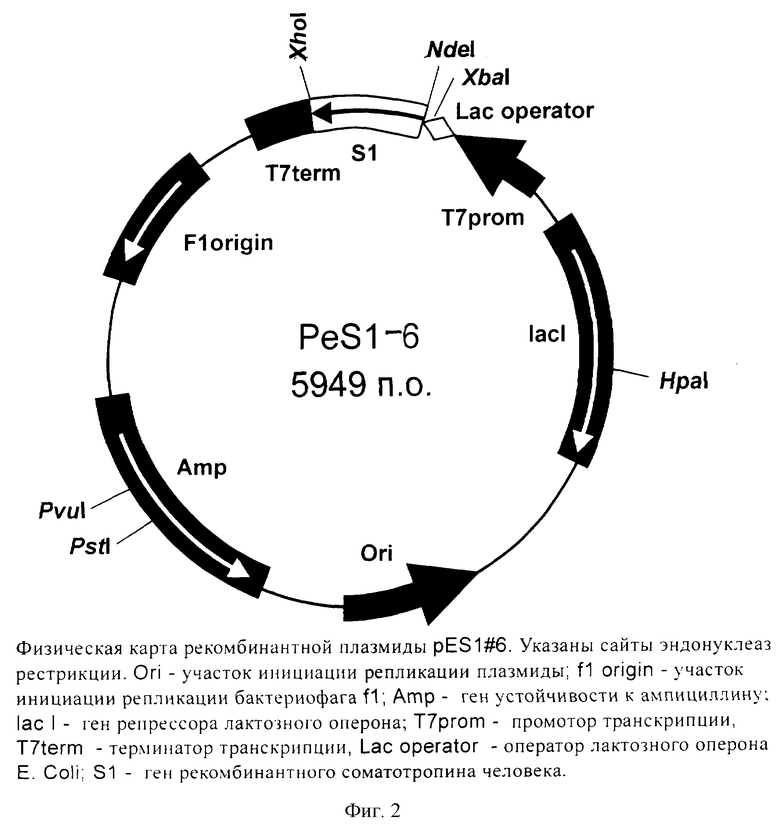
Поставленная задача решается за счет конструирования рекомбинантной плазмидной ДНК pES1-6, кодирующей полипептид с последовательностью соматотропина, имеющей молекулярную массу 3,66 Мда (5949 п.о.); состоящей из: NdeI/XhoII-фрагмента ДНК плазмиды pет 22b(+), содержащего промотор и терминатор транскрипции Т7-РНК-полимеразы, усилитель трансляции гена 10 фага Т7, ген β-лактамазы, и NdeI/XhoI-фрагмента ДНК, содержащего последовательность искусственного гена рекомбинантного соматотропина; содержащую: в качестве генетического маркера ген β-лактамазы, детерминирующей устойчивость трансформированных плазмидой pES1-6 клеток Е.coli к ампициллину; уникальные сайты узнавания эндонуклеаз рестрикции, расположенные на следующем расстоянии вправо от сайта NdeI: XbaI - 38 п.о., НраI - 1332 п.о., PstI - 4065 п.о., PvuI - 4190 п.о., XhoI - 5363 п.о., а также штамма-продуцента Escherichia coli BL21(DE3)/pES1-6, содержащего рекомбинантную плазмидную ДНК pES1-6 - продуцент рекомбинантного соматотропина, обеспечивающего синтез рекомбинантного соматотропина в количестве 60-70% от суммарного содержания белка клеток.
Преимущества предлагаемого изобретения заключаются, во-первых, в использовании при химико-ферментативном синтезе гена соматотропина максимально широкого набора кодонов, являющихся оптимальными для продукции белка в Escherichia coli, расположение которых в синтетическом гене устраняет возможность образования на синтезируемой мРНК протяженных "шпилек", потенциально ингибирующих трансляцию. На фиг. 1 приведена аминокислотная последовательность соматотропина и соответствующая ей нуклеотидная последовательность гена: верхняя строка - природная кДНК соматотропина, нижняя строка - синтезированный искусственный ген. Во-вторых, - применение для биосинтеза рекомбинантного белка оптимальных регуляторных элементов, контролирующих его экспрессию: Т7-lас промотора для предотвращения базальной экспрессии гена до момента начала индукции и высокого уровня транскрипции соответствующей мРНК при индукции, высокоэффективного терминатора транскрипции Т7 и блока различных стоп-кодонов, исключающих биосинтез удлиненных вариантов рекомбинантного соматотропина. Преимущество предлагаемого штамма E.coli заключается в использовании бактерий с фенотипом Lon OmpT, что исключает возможность протеолитического расщепления синтезируемого de novo рекомбинантного соматотропина и загрязнения выделяемого белка наиболее активными протеазами E.coli.
Конструирование нового гена, кодирующего соматотропин человека, осуществляют на основе плазмиды рЕТ22b(+). Искусственный ген, кодирующий соматотропин, фланкированный сайтами рестриктаз NdeI и XhoI, получают химико-ферментативным синтезом набора олигонуклеотидных фрагментов с последующей их сборкой и амплификацией при помощи полимеразной цепной реакции (ПЦР). Перед лигированием для генерации липких концов амплификат и векторную плазмиду обрабатывают рестриктазами NdeI и XhoI. Лигазную смесь используют для трансформации компетентных клеток Е.coli DH5α. Отбор положительных клонов проводят при помощи ПЦР с использованием специфических праймеров, с последующим рестриктным анализом выделенной плазмидной ДНК рестриктазами NaeI и XhoI. Структуру гена, кодирующего рекомбинантный соматотропин, определяют секвенированием по методу Сенгера.
Она должна полностью соответствовать нуклеотидной последовательности исходного искусственного гена соматотропина (фиг.1).
Рекомбинантная плазмидная ДНК pES1-6, кодирующая полипептид соматотропин, характеризуется следующими признаками:
- имеет молекулярную массу 3,66 МДа;
- кодирует полипептид соматотропин;
- состоит из: NdеI/XhoI-фрагмента ДНК плазмиды рЕТ22b(+), содержащей промотор и терминатор транскрипции Т7-РНК-полимеразы, усилитель трансляции гена 10 фага Т7, ген β-лактамазы и NdеI/XhoI-фрагмента ДНК, включающего искусственный ген соматотропина;
- имеет уникальную совокупность признаков: промотор и терминатор транскрипции РНК-полимеразы бактериофага Т7, усилитель трансляции гена 10 бактериофага Т7; искусственный ген, кодирующий соматотропин; ген β-лактамазы, детерминирующей устойчивость трансформированных плазмидой pES1-6 клеток Е.coli к ампициллину; уникальные сайты узнавания рестрикционных эндонуклеаз, расположенные на следующем расстоянии вправо от сайта NdеI:XhoI - 38 п.о., НраI - 1332 п.о., PstI - 4065 п.о., PvuI - 4190 п.о., XhoI - 5363 п.о.
Для получения штамма-продуцента рекомбинантного соматотропина плазмидную ДНК pES1-6 используют для трансформации компетентных клеток Escherichia coli BL21(DE3) и проводят отбор клонов, сохраняющих уровень биосинтеза рекомбинантного полипептида не ниже 60-70% от суммарного клеточного белка в течение по крайней мере шести последовательных пассирований. Для этого клоны трансформированных плазмидой pES1-6 клеток Е.coli BL21(DE3) выращивают в богатой среде (YT-, LB-бульон и др.) с добавлением ампициллина до 100 мкг/мл и раствора глюкозы до 1% в течение 12-14 часов, инокулируют новую порцию питательной среды в соотношении 1:100, растят культуру до достижения оптической плотности 1 О.Е., индуцируют изопропилтио-β-D-галактозидом и растят еще 3-6 часов. Получение из клеток продуцента рекомбинантного соматотропина включает следующие стадии: отделение бактерий от культуральной среды, их разрушение одним из обычно применяемых способов; отмывку буферными растворами телец включения от водорастворимых компонентов клетки; солюбилизацию и восстановление целевого белка, его рефолдинг и окончательную очистку.
Полученный штамм-продуцент Escherichia coli BL21(DE3)/pES1-6 характеризуется следующими признаками.
Морфологические признаки: клетки палочковидной формы, грамотрицательные, неспороносные.
Культуральные признаки: при росте на агаризованной среде LB колонии круглые, гладкие, мутные, блестящие серые, край ровный. При росте на жидких средах (на минимальной среде с глюкозой или YT-бульоне) образуют интенсивную ровную муть.
Физико-биологические признаки: клетки растут при температуре от 4°С до 40°С при оптимуме рН от 6,8 до 7,5. В качестве источника азота используют как минеральные соли в аммонийной форме, так и органические соединения в виде пептона, триптона, дрожжевого экстракта, аминокислот и т.д. В качестве источника углерода используют аминокислоты, глицерин, углеводы.
Устойчивость к антибиотикам: клетки проявляют устойчивость к ампициллину (до 500 мкг/мл), обусловленную наличием в плазмиде гена β-лактамазы(blа).
Преимущество предлагаемого штамма-продуцента заключается в использовании бактерий с фенотипом Lon OmpT, что исключает возможность протеолитического расщепления синтезируемого de novo рекомбинантного гормона роста и загрязнения выделяемого белка наиболее активными протеазами E.coli.
Клетки Е.coli BL21(DE3)/pES1-6 являются суперпродуцентом. При индукции изопропилтио-β-D-галактозидом происходит эффективный биосинтез рекомбинантного соматотропина, который накапливается в клетках в количестве более 60% суммарного белка бактерий.
На фиг.1 представлена нуклеотидная последовательность и кодируемая ею аминокислотная последовательность NdеI/XhoI-фрагмента плазмиды pES1-6; на фиг.2 - физическая карта плазмиды pES1-6.
Изобретение иллюстрируют примеры.
Пример 1.
Конструирование рекомбинантной плазм идной ДНК pES1-6.
Нуклеотидную последовательность, соответствующую гену соматотропина, получают химико-ферментативным синтезом. Для этого теоретически рассчитанную последовательность ДНК разбивают на перекрывающиеся фрагменты размером около 50 п.о. Химический синтез олигонуклеотидов, соответствующих этим фрагментам, выполняют твердофазным фосфоамидитным методом при помощи, например, ДНК-синтезатора ASM-102U (БИОССЕТ, Новосибирск) с наращиванием олигонуклеотидной цепи в направлении от 3’-конца к 5’-концу с помощью защищенных фосфамидитов - 5’-диметокситритил-N-ацил-2’-дезоксинуклеозид-3’-O-(β-цианэтил-диизопропиламино)-фосфитов, активированных тетразолом. Синтез проводят в масштабе 0,5-0,7 мкмоль, используя в качестве носителя пористое стекло (размер пор 500 А), к которому через 3’-сукцинатную связь присоединяют первое нуклеозидное звено (нагрузка 20-30 мкмоль/г). Полученные олигонуклеотиды подвергают 5’-концевому фосфорилированию с использованием Т4 полинуклеотидкиназы (Fermentas, Литва). Для этого олигонуклеотиды в количестве 20 пмоль смешивают с ферментом в количестве 10 ед. в буферном растворе, содержащем 50 мМ Tris-HCl (pH 7,6 при 25°С), 10 мМ MgCl2, 5 мМ дитиотреита, 1 мМ спермидина, 0,1 мМ АТФ и 0,1 мМ ЭДТА. Реакцию ведут 30 минут, полинуклеотид киназу инактивируют нагреванием до 65°С в течение 10 мин.
Фософрилированные олигонуклеотиды смешивают в эквимолярном соотношении в 50 мкл буфера, содержащего 20 мМ трис-HCl, pH 7,56, 10 мМ MgCl2, 0,2 мМ rАТР, 10 мМ дитиотреита, прогревают до 65°С, медленно охлаждают до 37°С в течение часа и добавляют 10 ед.ак. Т4-ДНК-лигазы. Реакцию лигирования ДНК проводят 4 ч при 37°С. 0,1 мкл полученного раствора используют в качестве матрицы в полимеразной цепной реакции (ПЦР) в присутствии термостабильной ДНК-полимеразы Pfu и специфических праймеров
5’ATA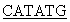 TTCCCGACCATCCCACTGTC3’ и
TTCCCGACCATCCCACTGTC3’ и
5’CCG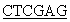 TCATTAGAAGCCACAGCTGCCC3’.
TCATTAGAAGCCACAGCTGCCC3’.
Проводят 25 циклов амплификации (95°С, 20 с; 62°С, 40 с; 72°С, 60 с) для синтеза полноразмерного фрагмента ДНК, содержащего последовательность гена соматотропина, фланкированного сайтами узнавания рестриктаз NdeI и XhoI. Продукт амплификации гидролизуют рестриктазами NdeI и XhoI, очищают электрофорезом в 5% акриламидном геле, полосу ДНК величиной около 600 п.о. выделяют из геля методом электроэлюции и осаждают ДНК из раствора этанолом.
Для приготовления вектора ДНК плазмиды рЕТ-22b(+) (3 мкг, 1 пмоль) обрабатывают в 40 мкл буфера Y (33 мМ трис-ацетат, pH 7,9, 10 мМ Mg-ацетат,66 мМ К-ацетат 1, 0,5 мМ DTT, 0,1 мг/мл BSA) рестриктазой NdeI (10 ед. акт.), осаждают ДНК этанолом, растворяют в 40 мкл буфера R (10 мМ трис-HCl, рН 8,5, 10 мМ MgCl2, 100 мМ КСl, 0,1 мг/мл BSA) и обрабатывают рестриктазой XhoI (10 ед. акт.) в течение 1 ч при 37°С. Полученный фрагмент ДНК величиной 5,4 т.п.о. после электрофоретического разделения в 1% агарозном геле выделяют из геля методом электроэлюции и осаждают ДНК из раствора этанолом.
1 мкг полученного векторного фрагмента лигируют с 2 пмоль NdeI/XhoI-фрагмента размером 600 п.о., содержащего синтетический ген рекомбинантного соматотропина, в 10 мкл буфера (20 мМ трис-HCl, рН 7,56, 10 мМ MgCl2, 0,2 мМ rATP, 10 мМ дитиотреита) с помощью 10 ед. акт. Т4-ДНК-лигазы в течение 12 ч при 10°С.
1 мкл полученной лигазной смеси используют для электротрансформации компетентных клеток Е.coli BL21(DE3), которую проводят, например, при помощи аппарата для электротрансформации ВТХ 600 при зазоре между пластинами электропорационной кюветы 1 мм и напряжении разряда 1,4 кВ. После трансформации суспензию бактерий смешивают с питательной средой SOC, растят 1 час на +37°С и высевают на чашки Петри с LB-агаром, содержащим 50 мкг/мл ампициллина.
Первичный отбор клонов, содержащих нужную плазмиду, проводят методом "ПЦР с клонов" с использованием специфических праймеров
5’ATA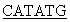 TTCCCGACCATCCCACTGTC3’ и
TTCCCGACCATCCCACTGTC3’ и
5’CCG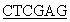 TCATTAGAAGCCACAGCTGCCC3’.
TCATTAGAAGCCACAGCTGCCC3’.
Провоят 25 циклов амплификации (95°С, 20 с; 62°С, 40 с; 72°С, 60 с), с последующим электрофоретическим анализом ПЦР продуктов в 1% агарозном геле на наличие ПЦР-продукта длиной около 600 п.н. Отобранные клоны используют для подроста в жидкой среде и выделения плазмидной ДНК плазмиды, которую анализируют на наличие вставки с помощью эндонуклеаз рестрикции NdeI и XhoI с последующим разделением продуктов гидролиза в 5% полиакриламидном геле. Окончательное строение плазмид, содержащих NdeI/XhoI-фрагмент около 600 п.о., подтверждают определением нуклеотидной последовательности методом секвенирования по Сенгеру. По данным секвенирования отбирают ту плазмиду, нуклеотидная и соответствующая ей аминокислотная последовательности NdeI/XhoI-фрагмента которой полностью идентичны первоначально запланированной (фиг.1). Проводят трансформацию клеток Е.coli BL21(DE3) выбранной плазмидой, как описано выше, петлей переносят 5-10 колоний в 5 мл жидкой среды 2xYT, содержащей 50 мкг/мл ампициллина, в течение 2 ч на качалке со скоростью вращения 190 об/мин до мутности А550 0,7-0,8, отбирают аликвоту культуры для последующего анализа, прибавляют индуктор - изопропилтио-β-D-галактозид до концентрации 0,2 мМ и продолжают рост еще 2 ч. Равные аликвоты суспензии клеток, отобранных до внесения индуктора и после завершения роста центрифугируют, отделяют супернатант и анализируют осадок клеток электрофорезом в ППАГ, как описано в примере 2. Появление отчетливо видимой полосы белка в районе 20 КДа в образце пробы, отобранной после индукции, свидетельствует о способности штамма синтезировать рекомбинантный соматотропин при индукции IPTG и полностью подтверждает корректность сборки плазмиды.
Пример 2.
Получение штамма-продуцента Е.coli BL21(DE3)/pes1-6 с рекомбинантным гормоном роста и определение его продуктивности.
Штамм-продуцент Е.coli BL21(DE3)/pES1-6 получают трансформацией компетентных клеток Е.coli BL21(DE3) плазмидой pES1-6, как описано в примере 1.
Штамм продуцента Е.coli BL21(DE3)/pES1-6 выращивают при 37°С в 100 мл YT-бульона (рН 7,0) с 50 мкг/мл ампициллина в течение 2 ч на качалке со скоростью вращения 190 об/мин до мутности А550 0,7-0,8, прибавляют изопропилтио-β-D-галактозид до концентрации 0,2 мМ и продолжают процесс еще 6 ч или продолжают выращивание в отсутствие индуктора в течение 6 ч. Каждый час отбирают пробу по 2 мл, определяют А550 и количество культуры, соответствующее 1 мл с А550 1,0, центрифугируют 5 мин при 6000 об/мин. Осажденные клетки в 100 мкл лизирующего буфера с красителем бромфеноловым синим обрабатывают 20 с ультразвуком, нагревают 3 мин при 100°С и пробы по 1 мкл используют для электрофореза в 15% SDS-ПААГ. Гель прокрашивают кумасси R-250 по стандартной методике и сканируют для определения относительного количества белка в полосе целевого белка. По данным сканирования содержание рекомбинантного соматотропина составляет 60-70% от всех клеточных белков.


Изобретение относится к биотехнологии, в частности к генной инженерии, и может быть использовано для получения рекомбинантного гормона роста человека. Рекомбинантная плазмидная ДНК pES1-6, кодирующая полипептид с последовательностью соматотропина, имеющей молекулярную массу 3,66 Мда (5949 п.о.); состоит из: NdeI/XhoI-фрагмента ДНК плазмиды рЕТ22b(+), содержащего промотор и терминатор транскрипции Т7-РНК-полимеразы, усилитель трансляции гена 10 фага Т7, ген β-лактамазы, и NdeI/XhoI-фрагмента ДНК, содержащего последовательность искусственного гена рекомбинантного соматотропина; содержащую: в качестве генетического маркера ген β-лактамазы, детерминирующей устойчивость трансформированных плазмидой pESl-6 клеток Е. coli к ампициллину; уникальные сайты узнавания эндонуклеаз рестрикции, расположенные на следующем расстоянии вправо от сайта NdeI: XbaI - 38 п.о., HpaI - 1332 п.о., PstI - 4065 п.о., PvuI - 4190 п.о., XhoI - 5363 п.о. Получают штамм Escherichia coli BL21(DE3)/pESl-6, содержащий рекомбинантную плазмидную ДНК pESl-6 - продуцент рекомбинантного соматотропина. Изобретение позволяет получить рекомбинантный соматотропин с высоким выходом и по упрощенной технологии, 2 н.з.п. ф-лы, 2 ил.
| US 5795745, 18.08.1998 | |||
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК PSTH 2191, КОДИРУЮЩАЯ СИНТЕЗ СОМАТОТРОПИНА ЧЕЛОВЕКА, И ШТАММ ESCHERICHIA COLI - ПРОДУЦЕНТ СОМАТОТРОПИНА ЧЕЛОВЕКА | 1986 |
|
SU1387414A1 |
| РЕКОМБИНАНТНЫЕ ПЛАЗМИДНЫЕ ДНК, КОДИРУЮЩИЕ СИНТЕЗ ПРОИЗВОДНЫХ СОМАТОТРОПНОГО ГОРМОНА ЧЕЛОВЕКА, СПОСОБ ИХ КОНСТРУИРОВАНИЯ, ШТАММЫ ESCHERICHIA COLI, СОДЕРЖАЩИЕ ЭТИ ПЛАЗМИДЫ - ПРОДУЦЕНТЫ ПРОИЗВОДНЫХ СОМАТОТРОПНОГО ГОРМОНА ЧЕЛОВЕКА (ИХ ВАРИАНТЫ) | 1983 |
|
RU1248280C |
| СПОСОБ ВЫДЕЛЕНИЯ РЕКОМБИНАНТНОГО СОМАТОТРОПИНА | 1990 |
|
RU2075509C1 |

