Изобретение относится к биохимии и молекулярной биологии прокариотической клетки и может быть применено к анализу молекулярно-генетических механизмов формирования структуры клетки прокариот и роли белковых компонентов в их организации, а также особенностей ремоделирования генома, что является необходимым для раскрытия путей регулирования механизмов воздействия макро- и микроорганизмов, а также поиску новых мишеней для лекарственных средств и разработке экологически безопасных лечебных препаратов.
Известен способ препаративного выделения основных белков клеточных ядер из тимуса теленка [1], в котором был описан способ фракционирования гистонов с помощью ионообменной хроматографии на колонке с амберлитом ИРЦ-50. Недостаток этого метода заключается в том, что были использованы колонки размером 7,5×55 см и 4,5×49 см, для загрузки которых необходимо достаточно большое количество белка (2-3 г и 0,5-1 г соответственно), в то время как при работе с микроорганизмами выделение белка в таком объеме чрезвычайно трудоемко.
Известен способ фракционирования растительных гистонов на колонках с амберлитом ИРЦ-50 [2], в котором был описан способ разделения основных белков клеточных ядер проростков кукурузы. Недостаток этого метода заключается в том, что с его помощью невозможно расфракционировать микроколичества белка из клеточных супраструктур микроорганизмов (2-5 мкг).
Вышеуказанный способ выделения растительных гистонов на колонках с амберлитом ИРЦ-50 [2] был принят за основу, в котором первоначально из проростков кукурузы изолируют клеточные ядра, получают ядерный экстракт с помощью 2 М хлористого натрия и добавления 1 н. соляной кислоты, с последующим нанесением полученного белка на колонку с амберлитом ИРЦ-50 и элюированием гистонов в линейном градиенте гуанидин гидрохлорида, определением белка в элюатах турбидиметрически. Недостаток этого метода заключается в том, что гомогенизация неприемлема для клеток прокариот, т.к. культура уже состоит из отдельных клеток, необъединенных в ткань, с помощью 2 М хлористого натрия была выделена только одна фракция белков, содержание белка в которой определялось турбидиметрически, методом с низким порогом чувствительности (до 3-5 мкг белка в 0,1 мл раствора).
Цель изобретения - предлагается способ для препаративного выделения основных белков из надмолекулярных структур растущей популяции Escherichia coli с помощью ионообменной хроматографии на колонке с амберлитом ИРЦ-50.
Указанная цель достигается тем, что в способе препаративного выделения основных белков надмолекулярных структур растущей популяции Escherichia coli у консервированных в присутствии забуференного 80-90% глицерина клеток снимают клеточную оболочку 3% тритоном Х-100, затем проводят экстракцию возрастающими концентрациями солей: 0,14 М, 0,35 М; 2 М NaCl, 6 М гуанидин гидрохлоридом с 0,1% β-меркаптоэтанолом с последующим выделением из вышеперечисленных фракций основных белков с помощью ионообменной хроматографии с амберлитом ИРЦ-50 в прерывистом градиенте гуанидин гидрохлорида: 6%, 8,9%, 10,6%, 13% на 0,1 М калий-фосфатном буфере рН 6,8.
Изобретение иллюстрируется следующим примером.
Пример. Опыты проводили на протяжении жизненного цикла клеток штамма Е. coli JC-158 (Hfr PO1, thil, serA6, lacI22, relA1) [3], предоставленного Ступак И.В. и Ступак Е.Э., выращивание культуры проводилось Тропыниной Т.С. (Институт биологии УНЦ РАН, лаборатория математической и молекулярной генетики), и штамма E.coli 5а, предоставленного Маркушевой Т.В. (Институт биологии УНЦ РАН, группа генетики микроорганизмов). В работе для выращивания использовалась богатая питательная среда LB (Луриа-Бертани). В 1 литре дистиллированной воды растворялись при помешивании (на магнитной мешалке типа MM 2A): бакто-триптон (Difco, США) - 10 г; дрожжевой экстракт (Difco, США) - 5 г; NaCl - 10 г, рН до 7,5. Среда стерилизовалась при 120-123°С, при давлении пара 1 атм в стандартном автоклаве.
Бактериальные клетки, использованные в эксперименте, первоначально находились в агаризованных столбиках LB (на 1 литр среды 1,5 г агар-агара, Difco, США) и хранились при температуре 4°С. При такой температуре и практически полном отсутствии кислорода происходит замедление всех физиологических процессов в клетках. Для того чтобы перевести клетки в "нормальное физиологическое состояние", а именно аэробное дыхание, бактериальную культуру из агаризованного столбика в стерильных условиях переносили с помощью бактериологической петли в жидкую среду LB в объеме 5 мл, находящуюся в химической пробирке объемом 20 мл, закрывали ватно-марлевой пробкой и инкубировали при 37°С, 160 об/мин на лабораторном термостатируемом встряхивателе (П5.10-Э5960) в течение 16 часов. Отдельно выросшая хорошо сформировавшаяся колония бактерий с агаризованной среды LB пересевалась с помощью бактериологической петли в жидкую среду LB в объеме 5 мл и инкубировалась при 37°С, 160 об/мин 7 часов. Затем 100 мкл подросшей культуры клеток пересевалась в свежую жидкую среду LB в объеме 5 мл и инкубировалась при 37°С, 160 об/мин 16 часов. 2 мл культуры клеток вносилось в кювету с рабочей длиной 5,075 мм и измерялась оптическая плотность на колориметре фотоэлектрическом концентрационном (КФК-2) при длине волны 590 нм. Это значение составляло 1,0. В свежую жидкую среду LB в объеме 120 мл в 500 мл колбе высевалось 120 мкл 16 часовой культуры и проводилось инкубирование при 37°С, 160 об/мин в течение 7 часов 10 минут. Первая проба была взята через 50 минут после начала инкубирования. Оптическая плотность первой пробы составляла 0,005. Для дальнейшего анализа отбирались образцы в объеме 1,5 мл. Клетки осаждались центрифугированием при 12000 об/мин на центрифуге Эппендорф в течение 5 мин. Надосадочная жидкость удалялась, осадки подсушивались. К осадкам добавлялось по 50 мкл среды следующего состава: 80-90% глицерин на 0,01 М трис-HCl буфере рН 6.8 с добавлением 0.005 М MgCl2; 0.025 М KCl; 0.003 М CaCl2; 0.005 М NaCl для консервации клеток при минус 25°С. Последующие пробы отбирались через каждые 20 минут в течение 7 часов 10 минут. Далее осадки клеток промывали 3% тритоном Х-100 в среде следующего состава: 0.02М триэтаноламин (ТЭА)-HCl рН 6.8; 0.005 М MgCl2; 0.025 М KCl; 0.003 М CaCl2; 0.005 М NaCl рН 6,8; встряхивали в течение 30 мин на микрошейкере (Micro-shaker type 326 m, Польша), с последующим центрифугированием при 4000 об/мин (К-23, ГДР) в течение 20 мин для снятия клеточной оболочки, после чего осадок дважды промывали в среде следующего состава: 0.005 М MgCl2; 0.025 М KCl; 0.003 М CaCl2; 0.005 М NaCl; 0,01 М трис-HCl рН 6.8 с последующим центрифугированием при вышеуказанных условиях.
Бактериоплазматические белки экстрагировали 0,14 М NaCl, 0.01 М трис-HCl рН 6.8 буфером. Фракцию, непрочносвязанную с клеточным остатком, выделяли путем экстракции осадка 0,35 М NaCl, 0.01 М трис-HCl рН 6.8 буфером. Далее осадок фракционировали суспендированием в трис-HCl буфере с 2 М NaCl, получая фракцию, прочносвязанную с клеточным остатком. В осадке оставалась фракция, содержащая клеточный остаток с клеточной оболочкой. Последующую экстракцию проводили 6 М гуанидин гидрохлоридом с 0,1% β-меркаптоэтанолом на трис-HCl буфере. В вышеуказанном буфере осадок растворялся полностью. Клеточные фракции хранили при - 196°C в азоте.
Количество белка определяли по связыванию белка с кумасси ярко-синим G (Loba, Австрия) [4]. Метод использовался в случае микронаноколичественного определения белка. Полученные фракции пропускали через колонку с амберлитом ИРЦ-50 (полиментакриловая синтетическая смола со свободными карбоксильными группами). Смола использовалась в виде порошка, получаемого при размалывании в шаровой мельнице. Размельченная смола просеивалась через сито 200 меш, после чего многократно промывалась водой для удаления мельчайших частиц, высушивалась и промывалась ацетоном. Для достижения большей хроматографической эффективности проводили циклизацию. К 100 г смолы добавляли 500 мл 4 н. NaOH, перемешивали в течение 3 ч с последующим отмыванием на фильтре водой до нейтральной реакции и переводили в кислую форму, пропуская через нее 500 мл 4 М HCl, избыток HCl удаляли промыванием водой. Этой смолой заполняли колонку размером 0,4 х 4,5 см. На колонку наносили белок в количестве 20-50 мкг, растворенный в 6% гуанидин гидрохлориде на 0,1 М калий-фосфатном буфере рН 6,8. Скорость элюции составляла 6 мл/ч. Препаративное отделение основных белков протеома клеток Е. coli проводили в прерывистом градиенте гуанидин гидрохлорида: 6%, 8,9%, 10,6%, 13% на 0,1 М калий-фосфатном буфере рН 6,8. Содержание белка в элюатах определяли методом Бредфорд в нашей модификации [4]. В работе использовался отечественный препарат гуанидин гидрохлорида («Реахим»), который предварительно был очищен. Неперекристаллизованный препарат имеет высокое поглощение в ультрафиолете. Перекристаллизация проводилась по методу, описанному Луком [1]. Концентрацию гуанидин-гидрохлорида определяли рефрактометрически при комнатной температуре. Расчет концентрации велся исходя из рефрактометрического индекса [5], используя следующее соотношение:
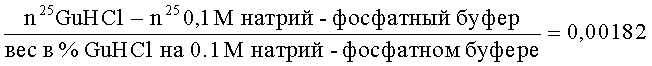
где n25Gu HCl - показатель преломления гуанидин гидрохлорида (величина, зависящая от концентрации препарата); n250,1 М натрий-фосфатный буфер - показатель преломления этого буфера (величина постоянная для данной концентрации); показатель 25 указывает на температуру, при которой проводились рефрактометрические исследования.
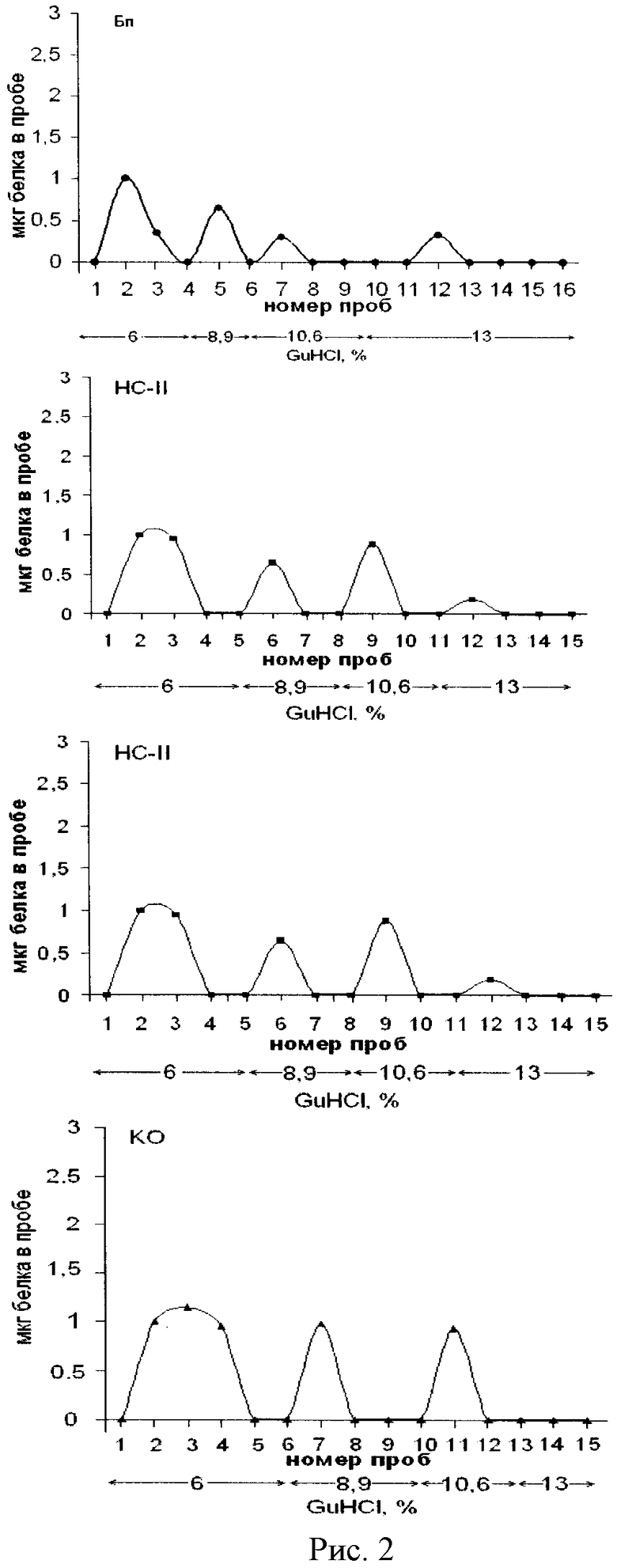
Описанным выше методом удалось отделить основные белки от «кислых» в полученных, как описано выше, надмолекулярных структурах (Бп, HC-I HC-II, КО) на 4 фракции: 0-фракция (незадержавшиеся «кислые» белки) и 3 фракции, полученные элюцией 8,9%, 10,6% и 13% концентраций гуанидингидрохлорида (положительнозаряженные основные гистононоподобные белки).
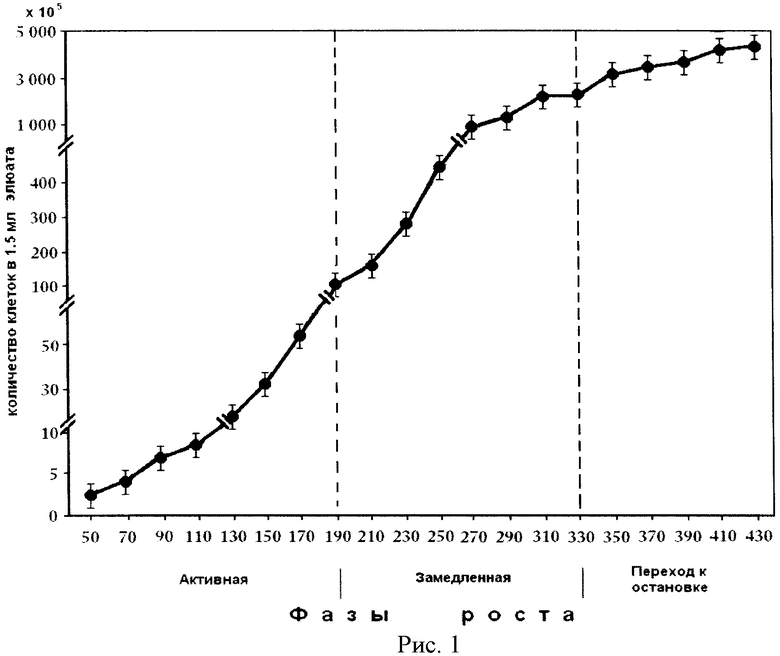
На рис.1 представлена кривая роста штамма Е. coli JC-158. На оси ординат показана оптическая плотность периодической культуры. На оси абсцисс показан возраст периодической культуры в мин, измеряемой в течение 7 часов 10 минут в растущей культуре клеток на фоне фаз роста растущей популяции E.coli.
На рис.2 показан хроматографический профиль элюции белков на колонке с ИРЦ-50 с помощью прерывистого градиента гуанидин гидрохлорида, элюированных из надмолекулярных структур растущей популяции E.coli. На оси ординат указано содержание белка в пробе, на оси абсцисс - номера проб. Использованы следующие обозначения: Бп - бактериоплазма, HC-I - надмолекулярные структуры, непрочносвязанные с клеточным остатком (КО), HC-II - надмолекулярные структуры, прочносвязанные с КО, КО - клеточный остаток. Из рисунка видно, что фракции гистоноподобных белков выделяются при следующих концентрациях гуанидин гидрохлорида: 8,9%, 10,6%, 13%, на 0,1 М калий-фосфатном буфере рН 6,8; пик полученный элюцией 6% гуонидингидрохлорида - это незадержавшиеся («кислые») белки надмолекулярных структур.
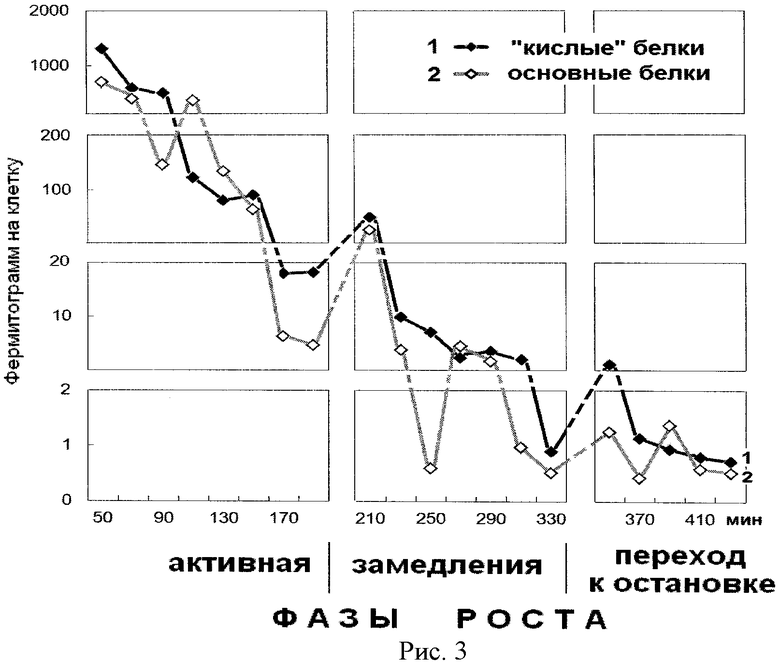
На рис.3 представлена динамика содержания суммарного белка в ступенчато-градиентных фракциях гуанидин гидрохлорида, элюированных из надмолекулярных структур растущей популяции E.coli. На оси абсцисс показано время роста популяции E.coli с обозначением фаз роста, мин. На оси ординат - содержание белка на 1 клетку, фермитограмм. Использованы следующие обозначения: 1 - динамика содержания «кислого» белка, элюированного 6% гуанидингидрохлоридом из надмолекулярных структур (незадержавшиеся на колонке с ИРЦ-50); 2 - динамика суммарного белка, элюированного 8,9%, 10,6% и 13% гуанидингидрохлоридом (сумма задержавшихся положительно заряженных основных белков на колонке с ИРЦ-50). Анализ рисунка показывает относительно одинаковую динамику содержания основных и «кислых» белков.
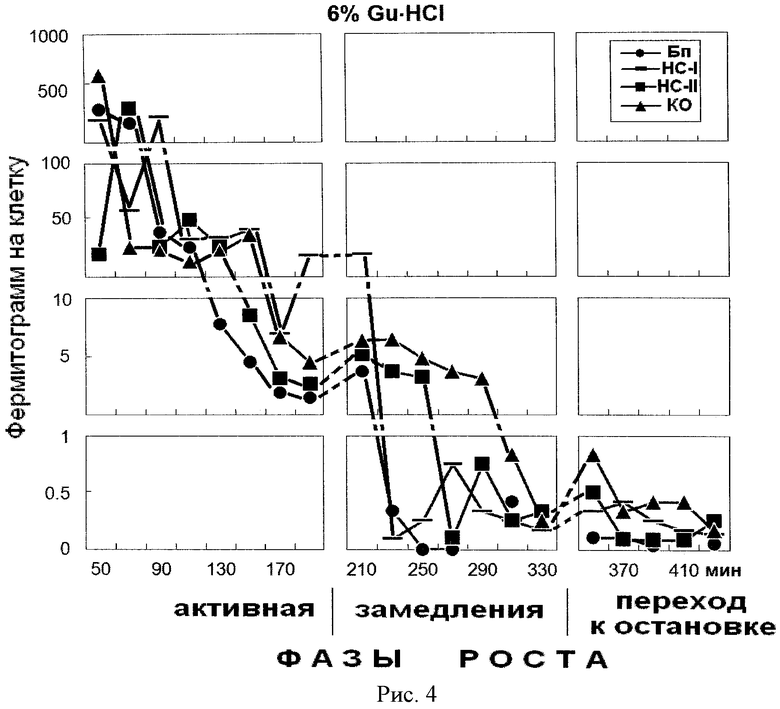
На рис.4 представлено содержание белка в 6% фракциях гуанидингидрохлорида, элюированных из надмолекулярных структур растущей популяции E.coli. На оси абсцисс показано время роста популяции E.coli с обозначением фаз роста. На оси ординат - содержание белка на 1 клетку, фермитограмм. Использованы следующие обозначения: Бп - бактериоплазма, HC-I - надмолекулярные структуры, непрочносвязанные с клеточным остатком (КО), HC-II - надмолекулярные структуры, прочносвязанные с КО, КО - клеточный остаток. На рисунке отражена динамика содержания «кислых» несвязавшихся с амберлитом ИРЦ-50 белков.
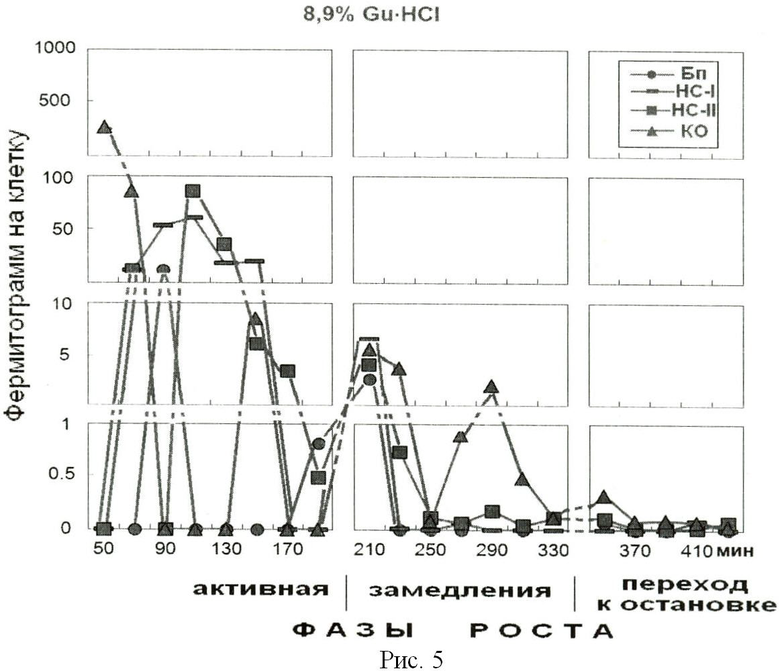
На рис.5 представлено содержание белка в 8,9% фракции гуанидингидрохлорида, элюированного из надмолекулярных структур растущей популяции E.coli. На оси абсцисс показано время роста популяции E.coli с обозначением фаз роста. На оси ординат - содержание белка на 1 клетку, фермитограмм. Использованы следующие обозначения: Бп - бактериоплазма, HC-I - надмолекулярные структуры, непрочносвязанные с клеточным остатком (КО), HC-II - надмолекулярные структуры, прочносвязанные с КО, КО - клеточный остаток. На рисунке отражена динамика содержания основных положительно заряженных белков, связавшихся с амберлитом ИРЦ-50, элюированных раствором 8,9% гуанидингидрохлорида.
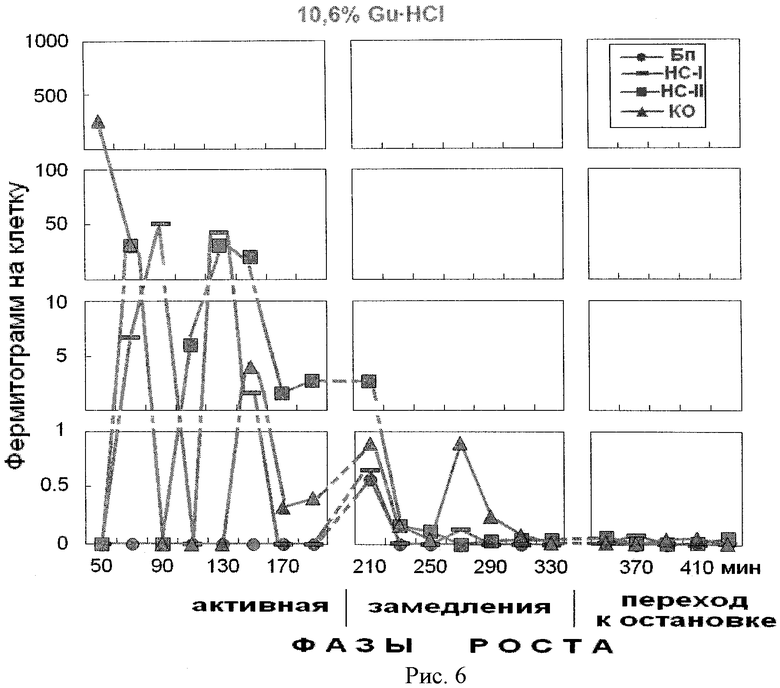
На рис.6 представлено содержание белка в 10,6% фракции гуанидингидрохлорида, элюированного из надмолекулярных структур растущей популяции Е.coli. На оси абсцисс показано время роста популяции E.coli с обозначением фаз роста. На оси ординат - содержание белка на 1 клетку, фермитограмм. Использованы следующие обозначения: Бп - бактериоплазма, HC-I - надмолекулярные структуры, непрочносвязанные с клеточным остатком (КО), HC-II - надмолекулярные структуры, прочносвязанные с КО, КО - клеточный остаток. На рисунке отражена динамика содержания основных положительно заряженных белков, связавшихся с амберлитом ИРЦ-50 и элюированных ступенчато-градиентно 10,6% гуанидингидрохлоридом.
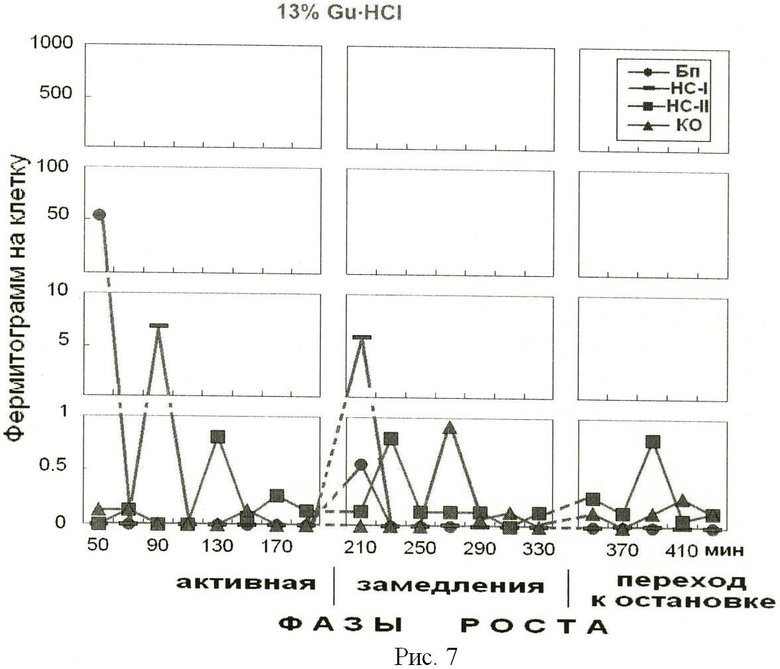
На рис.7 представлено содержание белка в 13% фракции гуанидингидрохлорида, элюированного из надмолекулярных структур растущей популяции E.coli. На оси абсцисс показано время роста популяции E.coli с обозначением фаз роста. На оси ординат - содержание белка на 1 клетку, фермитограмм. Использованы следующие обозначения: Бп - бактериоплазма, HC-I - надмолекулярные структуры, непрочносвязанные с клеточным остатком (КО), HC-II - надмолекулярные структуры, прочносвязанные с КО, КО - клеточный остаток. На рисунке отражена динамика содержания основных положительно заряженных связавшихся с амберлитом ИРЦ-50 белков, элюированных сиупенчато-градиентно 13% гуанидингидрохлоридом.
Анализ рис.5-7 показывает периодичность появления основных гистоноподобных белков, выделенных из надмолекулярных структур растущей популяции E.coli на фоне фаз роста периодической культуры. В работе, принятой за прототип [2], с помощью этого метода можно было отделить основные белки от «кислых» белков во фракции, полученной с помощью 2 М NaCl из клеточных ядер эукариот. Этот метод позволил получить фракции, обогащенные гистоном H1, гистонами Н2А и Н2В, гистонами Н3 и Н4. Мы полагаем, что заявленный подход позволит идентифицировать гистоноподобные белки в прокариотической клетки, определить их роль в метаболизме клетки, участие в укладке и структурировании нуклеоида в процессе роста бактериальной популяции и определить аминокислотный состав полученных белков. Знание биохимических процессов, происходящих на разных фазах роста популяции бактерий, позволяет раскрыть пути регулирования механизмов воздействия макро- и микроорганизмов, а также способствует поиску новых мишеней для лекарственных средств, что дает возможность управления бактериальными сообществами, населяющими человеческий организм.
Источники информации
1. Luck J.M., Rasmussen P.S., Satake K., Tsvetikov A.N. Further studies on the fractionation of calf thymus histone. // The J. of Biological Chemistry. 1958, V.233, N 6, P.1407-1414.
2. Иванова Э.А. Фракционирование растительных гистонов на колонках с амберлитом ИРЦ-50. // Материалы третьей научной конференции молодых ученых. Уфа: Башкирский филиал АН СССР, 1972, С.54-55.
3. Myrphy D.B., Pembroke J.T. Transfer of the IncJ plasmid R391 to recombination deficient E.coli K12; evidence that R391 behaves as a conjugal transposon. // FEMS Microbiology Letters, 1995, V.134, P.153-158.
4. Иванова Э.А., Вафина Г.Х. Способ получения ядерных фракций, обладающих протеиназной и ингибирующей активностью. Авторское свидетельство 1733471 // Б.И. 1992. Т.18, С.96.
5. Bonner J., Chalkley G.R., Dahmus М., Fambrough D., Fujimura F., Huang R.C., Huberman J., Jensen R., Marushige K., Ohlenbusch H., Olivera В., Widholm J. Isolation and characterization of chromosomal nucleoproteins. // Methods Enzymology. 1968, Acad. Press. New York. V.XII, part B, sec. V, ch. VII, P.25-31.
Изобретение относится к биохимии и молекулярной биологии. Предложен способ препаративного выделения основных белков из надмолекулярных структур растущей популяции Escherichia coli. Проводят консервацию клеток Escherichia coli в забуференном 80-90% глицерине при -25°С. Затем осадок клеток промывают 3% тритоном Х-100. Далее осуществляют экстракцию осадка возрастающими концентрациями солей: 0,14 М, 0,35 М; 2 М NaCl, 6 М гуанидин гидрохлоридом с 0,1% β-меркаптоэтанолом. Выделяют из полученных фракций основные белки с помощью ионообменной хроматографии с амберлитом ИРЦ-50 в прерывистом градиенте гуанидин гидрохлорида: 6%, 8,9%, 10,6%, 13% на 0,1 М калий-фосфатном буфере рН 6,8. Способ позволяет получать фракции, обогащенные основными белками, при использовании микроколичества белка клеточных супраструктур Escherichia coli. 7 ил., 1 пр.
Способ препаративного выделения основных белков из надмолекулярных структур растущей популяции Escherichia coli, включающий консервацию клеток в забуференном 80-90% глицерине при минус 25°С с последующим снятием клеточной оболочки 3% тритоном Х-100, экстракцией возрастающими концентрациями солей: 0,14 М, 0,35 М; 2 М NaCl, 6 М гуанидин гидрохлоридом с 0,1% β-меркаптоэтанолом и выделением из вышеперечисленных фракций основных белков с помощью ионообменной хроматографии с амберлитом ИРЦ-50 в прерывистом градиенте гуанидин гидрохлорида: 6%, 8,9%, 10,6%, 13% на 0,1 М калий-фосфатном буфере рН 6,8.
| Иванова Э.А | |||
| Устройство для выпрямления многофазного тока | 1923 |
|
SU50A1 |
| - Материалы третьей научной конференции молодых ученых.: Уфа, Башкирский филиал АН СССР, 1972, с.54-55 | |||
| LUCK J | |||
| MURRAY ЕТ AL | |||
| Further Studies on the Fractionation of Calf Thymus Histone // The Journal of Biological Chemistry, 1958, Vol.233, No.6, pp.1407-1414. | |||

