Изобретение относится к биохимии и молекулярной биологии прокариотической клетки и может быть применено к анализу молекулярно-генетических механизмов формирования структуры клетки прокариот и роли белковых компонентов в их организации, что необходимо для получения дополнительной информации в разработках и построении компьютерных моделей организации генных и эпигенных сетей управления.
Известен способ получения ядерных фракций, обладающих протеиназной и ингибирующей активностью [1], в котором был описан способ получения фракций из клеточных ядер соматических клеток растений. Недостаток этого метода заключается в том, что получение надмолекулярно-генетических структур осуществляется из клеточных ядер растений, а не из безъядерной прокариотической клетки, оболочка которой не может быть снята 0,5% тритоном Х-100.
Вышеуказанный способ получения ядерных фракций, обладающих протеиназной и ингибирующей активностью был принят за основу, в котором первоначально производят консервацию растительных тканей в забуференном 80-90% глицерине с последующей трехступенчатой гомогенизацией в забуференной 20% глицериновой среде, низкоскоростное центрифугирование растительного гомогената и очистку ядер через пятислойный (50, 60, 70, 80, 90 мас./объем) забуференный глицериновый градиент с последующей очисткой ядер буфером, содержащим 0,5% тритон Х-100 с последующей экстракцией ядерных фракций возрастающими концентрациями 0,14 М, 0,35 М, 2 М хлористого натрия, 6 М гуанидин гидрохлорида с 0,1% β-меркаптоэтанолом и 0,5 н. раствором гидроксида натрия при сопутствующей аффинной хроматографии вышеперечисленных ядерных фракций на сефарозе 4В с иммобилизованным трипсином либо ингибитором трипсина.
Недостатком этого способа является то, что трехступенчатая гомогенизация неприемлема для клеток прокариот, т.к. культура уже состоит из отдельных клеток необъединенных в ткань, 0,5% тритон Х-100 не снимает клеточную оболочку прокариотической клетки, экстракция фракций 0,5 н. гидроксидом натрия невозможна, т.к. клеточный остаток целиком растворяется в 6 М гуанидин гидрохлориде с 0,1% β-меркаптоэтанолом.
Целью изобретения является повышение выхода и активности целевого продукта.
Указанная цель достигается тем, что в способе выделения надмолекулярно-генетических структур из клеток прокариот из консервированных в 80-90% глицерине клеток, 3% тритоном Х-100 снимают клеточную оболочку с последующей экстракцией ядерных фракций возрастающими концентрациями 0,14 М, 0,35 М, 2 М хлористого натрия и 6 М гуанидин гидрохлорида с 0,1% β-меркаптоэтанолом при сопутствующей аффинной хроматографии вышеперечисленных ядерных фракций на сефарозе 4В с иммобилизованным трипсином либо ингибитором трипсина.
Изобретение иллюстрируется следующим примером.
Пример. Опыты проводили на протяжении жизненного цикла клеток штамма Е. coli JC-158 (Hfr PO1, thil, serA6, lacI22, relA1) [5], предоставленного Ступак И.В. и Ступак Е.Э., выращивание культуры проводилось Тропыниной Т.С. (Институт биологии УНЦ РАН, лаборатория математической и молекулярной генетики) и штамма E.coli 5а, предоставленного Маркушевой Т.В. (Институт биологии УНЦ РАН, группа генетики микроорганизмов). В работе для выращивания использовалась богатая питательная среда LB (Луриа-Бертани). В 1 литре дистиллированной воды растворялись при помешивании (на магнитной мешалке типа MM 2A): бакто-триптон (Difco, США) - 10 г; дрожжевой экстракт (Difco, США) - 5 г; NaCl - 10 г, pH до 7,5. Среда стерилизовалась при 120-123°С, при давлении пара 1 атм в стандартном автоклаве.
Бактериальные клетки, использованные в эксперименте, первоначально находились в агаризованных столбиках LB (на 1 литр среды 1,5 г агар-агара, Difco, США) и хранились при температуре 4°С. При такой температуре и практически полном отсутствии кислорода происходит замедление всех физиологических процессов в клетках. Для того чтобы перевести клетки в "нормальное физиологическое состояние", а именно аэробное дыхание, бактериальную культуру из агаризованного столбика в стерильных условиях переносили с помощью бактериологической петли в жидкую среду LB в объеме 5 мл, находящуюся в химической пробирке объемом 20 мл, закрывали ватно-марлевой пробкой и инкубировали при 37°С, 160 об/мин на лабораторном термостатируемом встряхивателе (П5.10-Э5960) в течение 16 часов. Отдельно выросшая хорошо сформировавшаяся колония бактерий с агаризованной среды LB пересевалась с помощью бактериологической петли в жидкую среду LB в объеме 5 мл и инкубировалась при 37°С, 160 об/мин 7 часов. Затем 100 мкл подросшей культуры клеток пересевалась в свежую жидкую среду LB в объеме 5 мл и инкубировалась при 37°С, 160 об/мин 16 часов. 2 мл культуры клеток вносилось в кювету с рабочей длиной 5,075 мм, и измерялась оптическая плотность на колориметре фотоэлектрическом концентрационном (КФК-2) при длине волны 590 нм. Это значение составляло 1,0. В свежую жидкую среду LB в объеме 120 мл в 500 мл колбе высевалось 120 мкл 16-часовой культуры, и проводилось инкубирование при 37°С, 160 об/мин в течение 7 часов 10 минут.
Первая проба была взята через 50 минут после начала инкубирования. Оптическая плотность первой пробы составляла 0,005. Для дальнейшего анализа отбирались образцы в объеме 1,5 мл. Клетки осаждались центрифугированием при 12000 об/мин на центрифуге Эппендорф в течение 5 мин. Надосадочная жидкость удалялась, осадки подсушивались. К осадкам добавлялось по 50 мкл среды следующего состава: 80-90% глицерин на 0,01 М трис-HCl буфере pH 6,8 с добавлением 0,005 М MgCl2; 0,025 М KCl; 0,003 М CaCl2; 0,005 М NaCl для консервации клеток при минус 25°С. Последующие пробы отбирались через каждые 20 минут в течение 7 часов 10 минут.
Далее осадки клеток промывали 3% тритоном Х-100 в среде следующего состава: 0,02М триэтаноламин (ТЭА)·HCl pH 6,8; 0,005 М MgCl2; 0,025 М KCl; 0,003 М CaCl2; 0,005 М NaCl pH 6,8; встряхивали в течение 30 мин на микрошейкере (Micro-shaker type 326 m, Польша), с последующим центрифугированием при 4000 об/мин (К-23, ГДР) в течение 20 мин для снятия клеточной оболочки, после чего осадок дважды промывали в среде следующего состава: 0,005 М MgCl2; 0,025 М KCl; 0,003 М CaCl2; 0,005 М NaCl; 0,01 М трис-HCl pH 6,8 с последующим центрифугированием при вышеуказанных условиях. Более низкие концентрации тритона Х-100 не снимали оболочки клеток E.coli.
Цитоплазматические белки экстрагировали 0,14 М NaCl, 0,01 М трис-HCl pH 6,8 буфером. Фракцию непрочно связанную с клеточным остатком выделяли путем экстракции осадка 0,35 М NaCl, 0,01 М трис-HCl pH 6,8 буфером. Далее осадок фракционировали суспендированием в трис-HCl буфере с 2 М NaCl. В осадке оставалась фракция, содержащая клеточный остаток с клеточной оболочкой. Последующую экстракцию проводили 6 М гуанидин гидрохлоридом с 0,1% β-меркаптоэтанолом на трис-HCl буфере. В вышеуказанном буфере осадок растворялся полностью, поэтому экстракцию 0,5 н. NaOH не проводили. Ядерные фракции хранили при - 196°C в азоте.
Количество белка определяли по связыванию белка с кумасси ярко-синим G (Loba, Австрия) [1, 3]. Метод использовался в случае микронаноколичественного определения белка.
Аффинную хроматографию проводили на колонках (0,5×4 см) либо с иммобилизованным трипсином («СПОФА», ЧССР), либо ингибитором трипсина («Reanal», Венгрия), ковалентно присоединенных к CNBr-активированной агарозе (Институт химии АН Эстонии) [1].
Протеолитическую активность протеаз определяли по расщеплению низкомолекулярного белка протамина («Calbiochem», США) [1]. Количество освободившегося аргинина рассчитывали, пользуясь калибровочным графиком. В качестве стандарта использовали D,L-аргинин («Reanal», Венгрия). Активность протеаз и их ингибиторов выражали в нмоль аргинина на 1 мг/с.
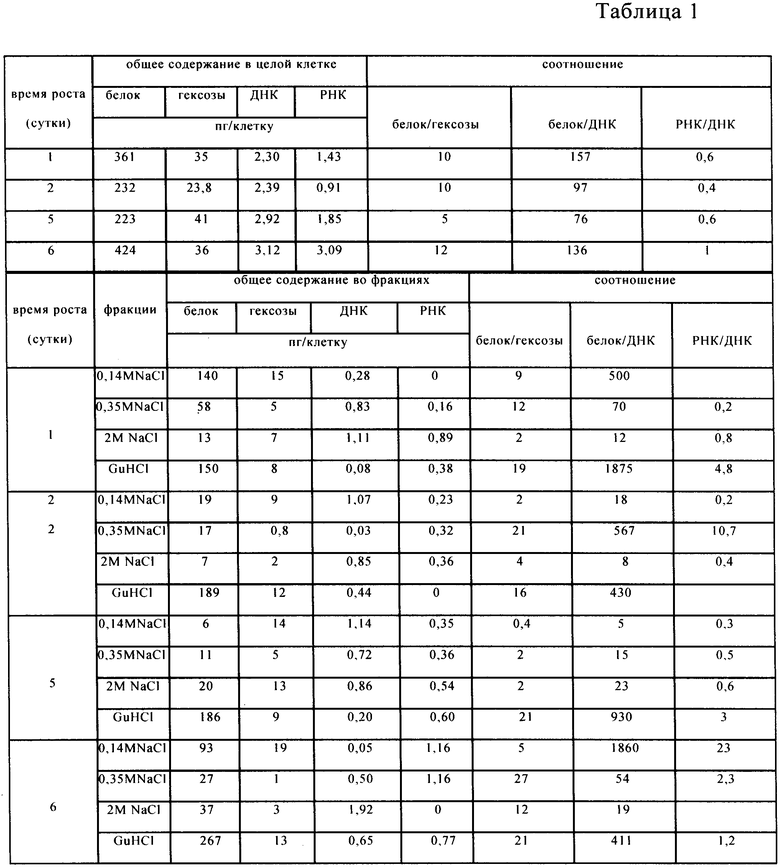
Анализ полноты выделения клеточных фракций (табл.1) и фракций, полученных после аффинной хроматографии (табл.3), показал, что все фракции представляют собой не чистый белок, а комплексы нуклеиновых кислот, белка и гексоз. Для определения ДНК и РНК использовали метод А.С.Спирина [4]. Для определения содержания гексоз использовали метод [2].
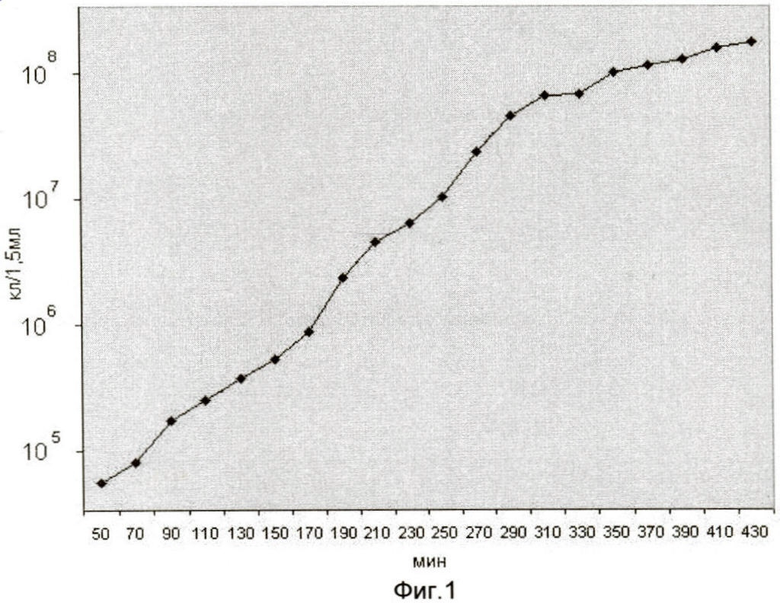
На фиг.1 представлена кривая роста штамма Е. coli JC-158. На оси ординат показана оптическая плотность периодической культуры. На оси абсцисс показан возраст периодической культуры в мин, измеряемой в течение 7 часов 10 минут в растущей культуре клеток.
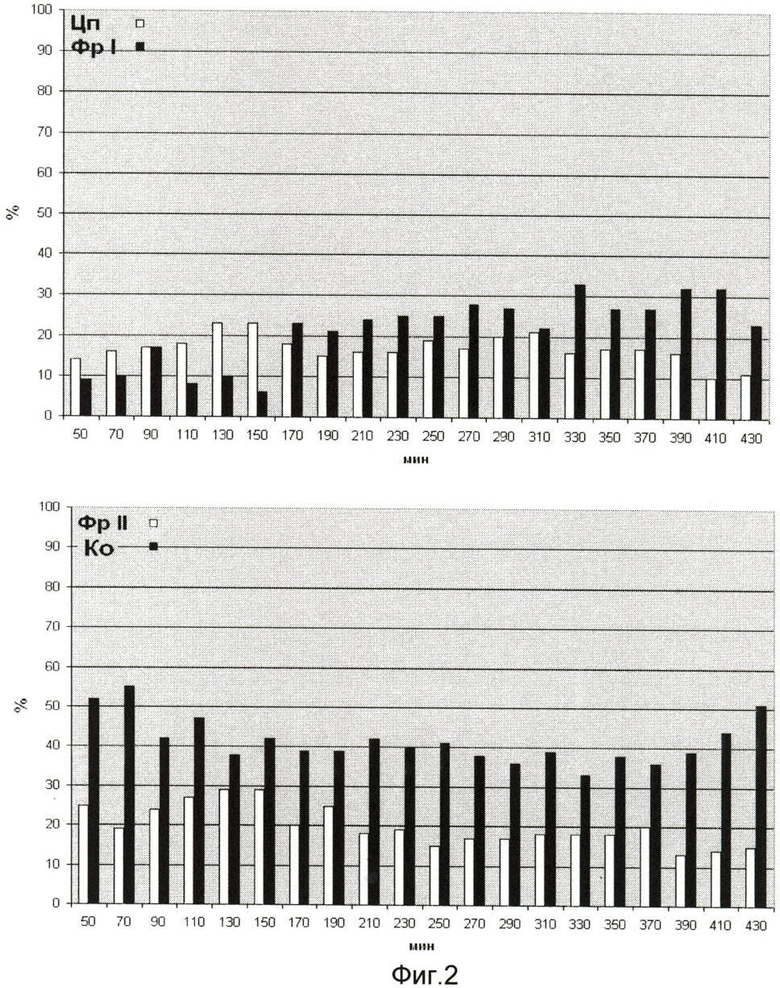
На фиг.2 представлена полнота выделения белкового компонента в надмолекулярно-генетических структурах клеток Е. coli JC-158. На оси ординат показан процент выхода белка, на оси абсцисс - возраст культуры.
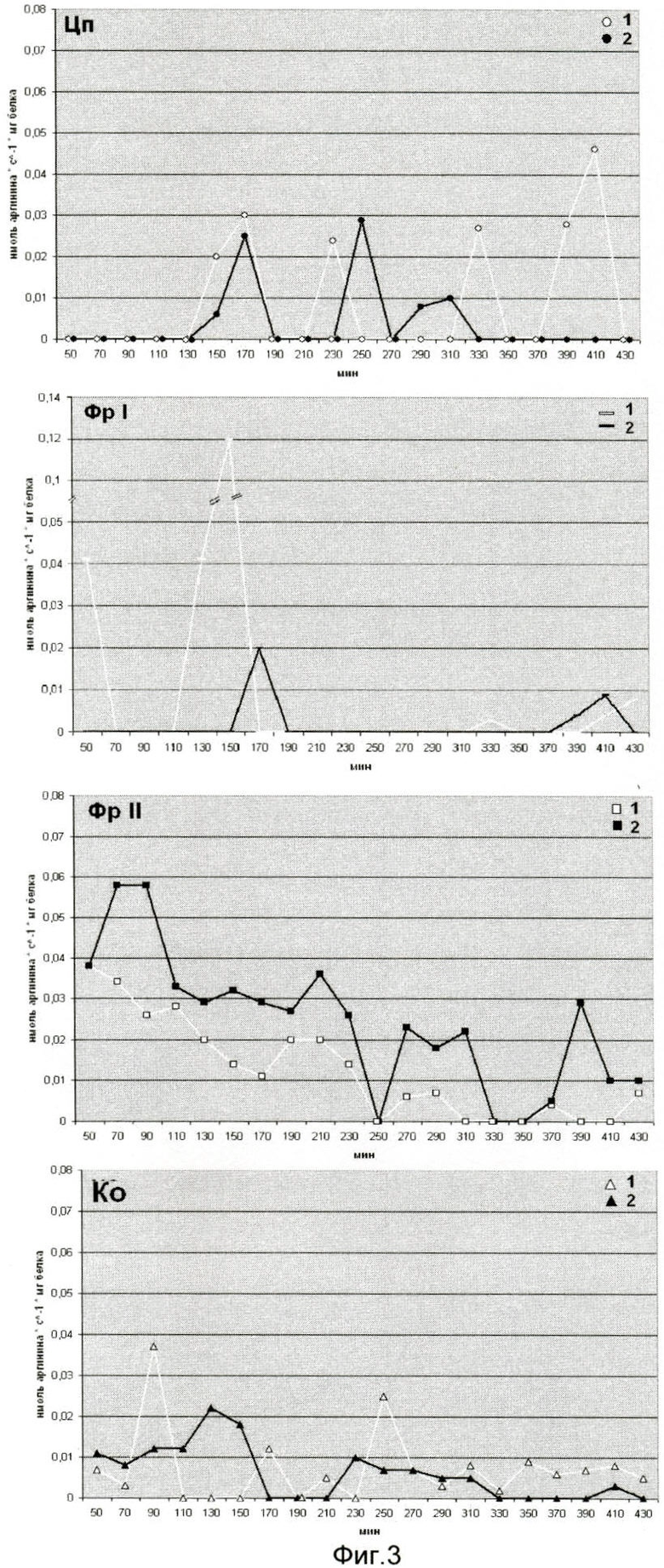
Эффективность последовательной ступенчатой экстракции протеаз и ингибиторов трипсина из клеток Е. coli JC-158 показана на фиг.3. На оси ординат (фиг.3) показана активность протеаз (1) и ингибиторов трипсина (2), выраженная в нмоль аргинина на 1 мг белка/с. На оси абсцисс указан возраст культуры в мин от 50 до 430 мин. Использованы следующие обозначения: Цп - цитоплазма; Фр-1 - фракция непрочно связанная с клеточным остатком (35 М NaCl); Фр-II - фракция прочно связанная с клеточным остатком (2 М NaCl); Ко - клеточный остаток с клеточной оболочкой;
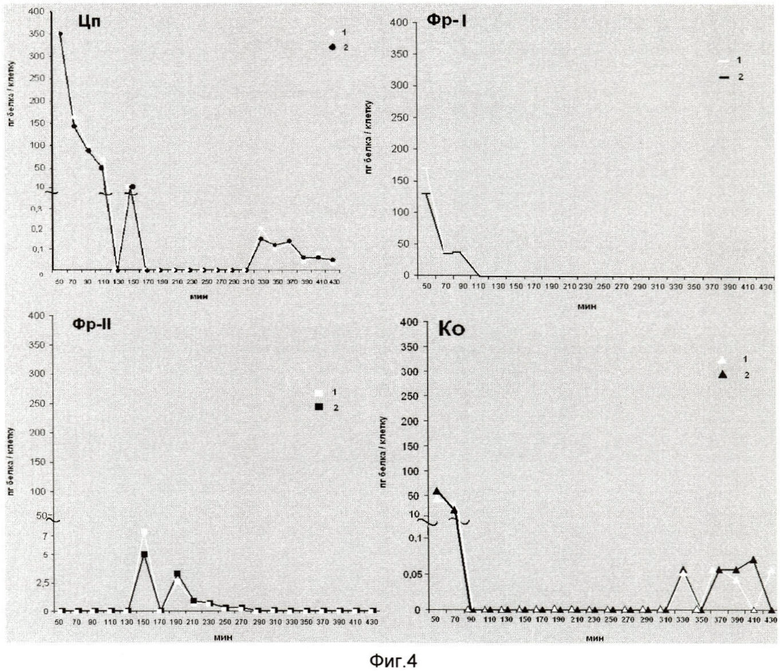
На фиг.4 показана последующая идентификация фракций, полученных из клеток Е. coli JC-158, на колонках либо с иммобилизованным ингибитором трипсина, либо с трипсином. На оси ординат показано содержание протеолитических и ингибиторных надмолекулярно-генетических комплексов в расчете на 1 клетку (пг). На оси абсцисс указан возраст культуры в мин от 50 до 430 мин. Использованы следующие обозначения: 1 - протеолитический комплекс; 2 - ингибиторный комплекс.
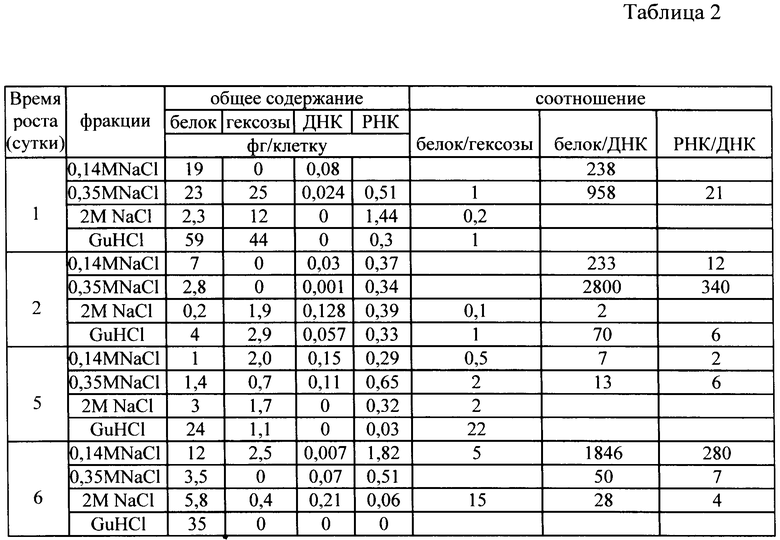
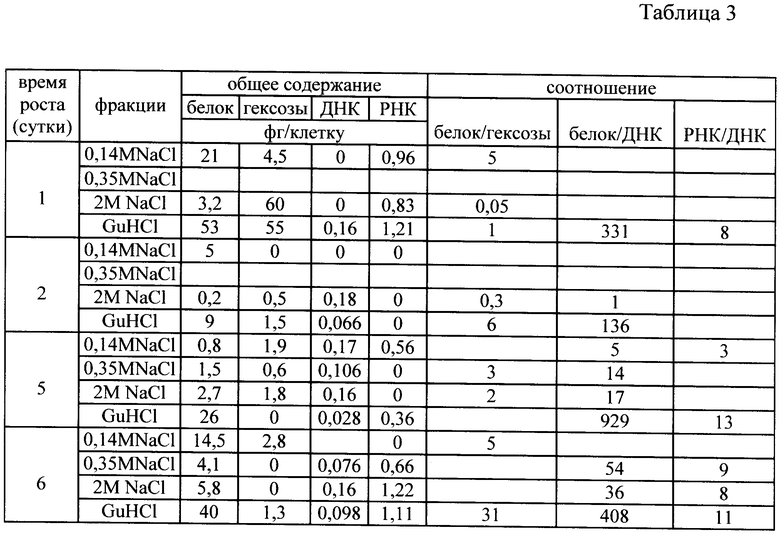
Исследование фракций, обладающих протеолитической и ингибиторной активностью включает предварительную консервацию клеток в глицериновой среде, снятие клеточной оболочки и выделение из них клеточных фракций возрастающими концентрациями солей. Табл.1 показывает, что структура клеток (штамма Е. coli 5а) не нарушена и иллюстрирует полноту выделения клеточных фракций. Табл.2 и 3 иллюстрируют компонентный состав аффинно-выделенных протеолитических (табл.2) и ингибиторных (табл.3) комплексов в течение роста клеток штамма Е. coli 5а в периодической культуре.
Предложенный способ рекомендуется в исследовании молекулярно-генетических механизмов формирования прокариотической структуры клетки и роли белковых компонентов в их организации, в частности в исследованиях Arg-X протеиназ, участвующих в ремоделировании хроматина, а также для построения компьютерных моделей организации прокариот.
Источники информации
1. Иванова Э.А., Вафина Г.Х. Способ получения ядерных фракций, обладающих протеиназной и ингибирующей активностью. Авторское свидетельство 1733471 // БИ 1992, Т. 18, С.96.
2. Иванова Э.А., Вафина Г.Х., Ремеева Р.Г. Способ определения углеводных компонентов в клеточных ядрах. Патент №2108571 // БИ №10, 10.04.98.
3. Скоупс Р. Методы очистки белков. М.: Мир, 1985, С.342.
4. Спирин А.С. Спектрофотометрическое определение суммарного количества нуклеиновых кислот // Биохимия, 1968, Т.23, 35, С.656.
5. Myrphy D.B., Pembroke J.T. Transfer of the IncJ plasmid R391 to recombination deficient E.coli K12: evidence that R391 behaves as a conjugal transposon // FEMS Microbiology Letters, 1995, V.134, P.153-158.
Изобретение относится к области биотехнологии, конкретно к получению протеолитических фракций из прокариотических клеток, и может быть использовано при анализе молекулярно-генетических механизмов формирования структуры клетки прокариот и роли белковых компонентов в их организации, что необходимо для получения дополнительной информации в разработках и построении компьютерных моделей организации генных и эпигенных сетей управления. Клетки Escherichia coli консервируют в присутствии забуференного 80-90% глицерина, затем обрабатывают 3% тритоном Х-100, с целью снятия клеточной оболочки, далее полученные цитоплазматические белки экстрагируют возрастающими концентрациями солей: 0,14 М, 0,35 М; 2 М NaCl, 6 М гуанидин гидрохлоридом с 0,1% β-меркаптоэтанолом, проводят аффинную хроматографию на сефарозе 4В с иммобилизованным ингибитором трипсина и оценивают протеолитическую активность. Данное изобретение позволяет получить фракцию из клеток прокариот. 3 табл., 4 ил.
Способ получения фракции из клеток Escherichia coli, обладающей протеолитической активностью, включающий консервацию клеток в присутствии забуференного 80-90% глицерина с последующим снятием клеточных оболочек 3% тритоном Х-100, экстракцию возрастающими концентрациями солей: 0,14 М, 0,35 М; 2 М NaCl, 6 М гуанидин гидрохлоридом с 0,1% β-меркаптоэтанолом, аффинную хроматографию на сефарозе 4 В с иммобилизованным ингибитором трипсина и последующей оценкой в элюатах протеолитической активности.
| Способ получения ядерных фракций, обладающих протеиназной и ингибирующей активностью | 1990 |
|
SU1733471A1 |
| JP 2008167756 A, 24.07.2008 | |||
| ЧЕРКАСОВ И.А | |||
| Аффинная хроматография ферментов | |||
| Успехи химии, 1972, т.41, №10, с.1911-1934. | |||

