[0001] Настоящая заявка испрашивает приоритет на китайскую патентную заявку №202110250342.9, поданную 08 марта 2021 года, которая включена в настоящий документ посредством ссылки во всей своей полноте.
ОБЛАСТЬ ТЕХНИКИ
[0002] Настоящее раскрытие относится к области фармацевтики и, в частности, к антителу анти-PVRIG протеин или фрагменту антитела и его применению в терапии рака.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
[0003] Опухолевые клетки уклоняются от наблюдения иммунной системы с помощью различных механизмов. Путь иммунных контрольных точек используется для поддержания самотолерантности и контроля эффекторной функции активированных лимфоцитов, но раковые клетки могут использовать этот путь, чтобы избежать нарушения. Последние сообщения показывают, что PVRIG является очень важной иммунной контрольной точкой. PVRIG в основном выражается на активированных Т-клетках и NK-клетках и подавляет эффекторную функцию Т-клеток и NK-клеток, взаимодействуя с лигандом CD112, выраженным на клетках-мишенях или DC-клетках. CD112 высоко выражается на поверхности многих видов опухолевых клеток, что позволяет подавлять функцию иммунной системы через сигнальный путь PVRIG и осуществлять иммунный побег.
[0004] В исследованиях было показано, что удаление PVRIG может привести к усилению эффекторной функции CD8+ Т-клеток. Кроме того, использование антител, блокирующих связывание PVRIG с его лигандом CD112, также может быть эффективным для восстановления функции инфильтрирующих опухоль CD8+ Т-клеток и подавления роста опухоли. Антитело, нацеленное на PVRIG человека, участвовало в клинических испытаниях, показав хороший терапевтический эффект.
[0005] Необходимо дальнейшее совершенствование антител, нацеленных на PVRIG.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0006] Целью настоящего раскрытия является создание антитела или фрагмента антитела с улучшенной противоопухолевой функцией и их применение. Представленное антитело или фрагмент антитела способны эффективно связываться с PVRIG и блокировать связывание PVRIG с его лигандом CD112. Было подтверждено, что представленное антитело или фрагмент антитела обладает высокой аффинностью к PVRIG человека и эффективно блокирует взаимодействие между PVRIG человека и CD112.
[0007] В первом аспекте настоящее раскрытие предоставляет антитело или фрагмент антитела. Антитело или фрагмент антитела включает комплементарные определяющие области тяжелой цепи HCDR1, HCDR2 и HCDR3 и комплементарные определяющие области легкой цепи LCDR1, LCDR2 и LCDR3. HCDR1 выбирается из последовательности, представленной в GYTFSSFS, GYTFSTFA или GYSFTAYT. HCDR2 выбирается из последовательности, представленной в ILPGSNST, ILPGINNT, ILPGGNNT или INPYNGGT. HCDR3 выбирается из последовательности, представленной в SSYWFAY, STYWFAY или AREGNYYGSRGDFDY. Легкая цепь включает комплементарные определяющие области легкой цепи LCDR1, LCDR2 и LCDR3. LCDR1 выбирается из последовательности, представленной в QSLLNSGNQKNY или QTIVTN. LCDR2 выбирается из последовательности, представленной в GAS или YAS. LCDR3 выбирается из последовательности, представленной в QNAHSYPPT, QNAHSYPPA или QQSHSWPFT.
[0008] Во втором аспекте настоящее раскрытие предоставляет антитело или фрагмент антитела. Антитело или фрагмент антитела выбран из одного из следующих:
[0009] (1) антитело или фрагмент антитела, включающий комплементарные определяющие области тяжелой цепи и комплементарные определяющие области легкой цепи, комплементарные определяющие области тяжелой цепи включают GYTFSSFS, ILPGSNST и SSYWFAY, а комплементарные определяющие области легкой цепи включают QSLLNSGNQKNY, GAS и QNAHSYPPT;
[0010] (2) антитело или фрагмент антитела, включающий комплементарные определяющие области тяжелой цепи и комплементарные определяющие области легкой цепи, комплементарные определяющие области тяжелой цепи включают GYTFSTFA, ILPGINNT и STYWFAY, а комплементарные определяющие области легкой цепи включают QSLLNSGNQKNY, GAS и QNAHSYPPT;
[0011] (3) антитело или фрагмент антитела, включающий комплементарные определяющие области тяжелой цепи и комплементарные определяющие области легкой цепи, комплементарные определяющие области тяжелой цепи включают GYTFSTFA, ILPGGNNT и STYWFAY, а комплементарные определяющие области легкой цепи включают QSLLNSGNQKNY, GAS и QNAHSYPPA;
[0012] (4) антитело или фрагмент антитела, включающий комплементарные определяющие области тяжелой цепи и комплементарные определяющие области легкой цепи, комплементарные определяющие области тяжелой цепи включают GYSFTAYT, INPYNGGT и AREGNYYGSRGDFDY, а комплементарные определяющие области легкой цепи включают QTIVTN, YAS и QQSHSWPFT.
[0013] В третьем аспекте настоящее раскрытие предоставляет антитело или фрагмент антитела, выбранные по меньшей мере из одного из следующих:
[0014] (1) антитело или фрагмент антитела, включающий вариабельную область тяжелой цепи и вариабельную область легкой цепи, причем вариабельная область тяжелой цепи представляет собой последовательность, представленную в SEQ ID NO: 1, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 1, а вариабельная область легкой цепи представляет собой последовательность, представленную в SEQ ID NO: 2, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 2;
[0015] (2) антитело или фрагмент антитела, включающий вариабельную область тяжелой цепи и вариабельную область легкой цепи, причем вариабельная область тяжелой цепи выбрана из группы, состоящей из последовательностей, представленных в SEQ ID NO: 3, SEQ ID NO: 6, SEQ ID NO: 9, SEQ ID NO: 12, SEQ ID NO: 15, SEQ ID NO: 18, SEQ ID NO: 21, SEQ ID NO: 24, SEQ ID NO: 27, SEQ ID NO: 30 и SEQ ID NO: 33 или последовательности, имеющие по меньшей мере одну аминокислотную замену по сравнению с последовательностями, представленными в SEQ ID NO: 3, SEQ ID NO: 6, SEQ ID NO: 9, SEQ ID NO: 12, SEQ ID NO: 15, SEQ ID NO: 18, SEQ ID NO: 21, SEQ ID NO: 24, SEQ ID NO: 27, SEQ ID NO: 30 и SEQ ID NO: 33, и вариабельная область легкой цепи представляет собой последовательность, представленную в SEQ ID NO: 36, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ IDNO: 36;
[0016] (3) антитело или фрагмент антитела, включающий вариабельную область тяжелой цепи и вариабельную область легкой цепи, причем вариабельная область тяжелой цепи представляет собой последовательность, представленную в SEQ ID NO: 39, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 39, а вариабельная область легкой цепи представляет собой последовательность, представленную в SEQ ID NO: 40, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 40;
[0017] (4) антитело или фрагмент антитела, включающий вариабельную область тяжелой цепи и вариабельную область легкой цепи, причем вариабельная область тяжелой цепи представляет собой последовательность, представленную в SEQ ID NO: 41, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 41, а вариабельная область легкой цепи представляет собой последовательность, представленную в SEQ ID NO: 42, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 42;
[0018] (5) антитело или фрагмент антитела, включающий вариабельную область тяжелой цепи и вариабельную область легкой цепи, причем вариабельная область тяжелой цепи представляет собой последовательность, представленную в SEQ ID NO: 43, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 43, а вариабельная область легкой цепи представляет собой последовательность, представленную в SEQ ID NO: 44, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 44.
[0019] В четвертом аспекте настоящее раскрытие предоставляет изолируемую нуклеиновую кислоту, кодирующую любое из антител или фрагментов антител, как описано выше.
[0020] В пятом аспекте настоящее раскрытие обеспечивает вектор экспрессии, включающий описанную выше нуклеиновую кислоту.
[0021] В шестом аспекте настоящее раскрытие обеспечивает рекомбинантную клетку, выражающую любое из антител или фрагментов антител, как описано выше.
[0022] В седьмом аспекте настоящее раскрытие обеспечивает фармацевтическую композицию, включающую антитело или фрагмент антитела, как описано выше, и фармацевтически приемлемый носитель.
[0023] В восьмом аспекте настоящее раскрытие обеспечивает использование антитела или фрагмента антитела, как описано выше, или фармацевтической композиции, как описано выше, в производстве лекарственного средства для лечения рака.
[0024] В девятом аспекте настоящее раскрытие обеспечивает фармацевтическую комбинацию, включающую: (1) антитело или фрагмент антитела, как описано выше, или фармацевтическую композицию, как описано выше; и (2) антитело для лечения рака, отличное от (1).
[0025] В десятом аспекте настоящее раскрытие обеспечивает способ профилактики или лечения рака, включающий введение субъекту эффективного количества антитела или фрагмента антитела, как описано выше, или фармацевтической композиции, как описано выше.
[0026] В одиннадцатом аспекте настоящее раскрытие предоставляет способ ингибирования активности PVRIG протеина в образце in vitro, включая контакт образца с любым из антител или фрагментов антител, как описано выше, или фармацевтической композицией, как описано выше. Согласно одному из вариантов реализации настоящего раскрытия, образец может представлять собой образец клетки.
[0027] Антитело или фрагмент антитела, представленные в настоящем раскрытии, способны специфически связываться с антигеном PVRIG и значительно усиливать убивающий эффект человеческих РВМС, а также демонстрируют хороший противоопухолевый эффект в мышиных моделях. Дополнительные особенности и преимущества настоящего раскрытия будут изложены в последующем подробном описании.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0028] ФИГ. 1 представляет собой график, показывающий результаты анализа ELISA связывания мышиного антитела с PVRIG человека в соответствии с вариантом реализации настоящего раскрытия;
[0029] ФИГ. 2 представляет собой график, показывающий результаты анализа связывания мышиного антитела к PVRIG человека на поверхности клеточной мембраны в соответствии с вариантом реализации настоящего раскрытия;
[0030] ФИГ. 3 представляет собой график, показывающий результаты анализа блокирования связывания PVRIG человека с CD112 человека мышиным антителом в соответствии с вариантом реализации настоящего раскрытия;
[0031] ФИГ. 4 представляет собой график, показывающий результаты анализа конкурентного связывания мышиного антитела с PVRIG человека на поверхности клеточной мембраны в соответствии с вариантом реализации настоящего раскрытия;
[0032] ФИГ. 5 представляет собой график, показывающий результаты активности связывания мышиных антител к PVRIG Масаса fascicularis и PVRIG мыши на поверхности клеточной мембраны в соответствии с вариантом реализации настоящего раскрытия;
[0033] ФИГ. 6 представляет собой график, показывающий результаты анализа ELISA связывания мышиного антитела с PVRIG Масаса fascicularis в соответствии с вариантом реализации настоящего раскрытия;
[0034] ФИГ. 7 представляет собой график, показывающий результаты активности мышиного антитела в обнаружении PVRIG на поверхности мембраны клеточной линии NKG в соответствии с вариантом реализации настоящего раскрытия;
[0035] ФИГ. 8 представляет собой график, показывающий результаты активности мышиного антитела в обнаружении PVRIG на поверхности лимфоцитов в соответствии с вариантом реализации настоящего раскрытия;
[0036] ФИГ. 9 представляет собой график, показывающий результаты выражения CD112 на поверхности линии опухолевых клеток человека в соответствии с вариантом реализации настоящего раскрытия;
[0037] ФИГ. 10 представляет собой график, показывающий результаты продвижения цитотоксичности NK-клеток мышиным антителом в соответствии с вариантом реализации настоящего раскрытия;
[0038] ФИГ. 11 представляет собой график, показывающий результаты продвижения цитотоксичности РВМС человека мышиным антителом в соответствии с вариантом реализации настоящего раскрытия;
[0039] ФИГ. 12 представляет собой график, показывающий результаты стимулирования выражения эффекторных молекул NK-клеток мышиным антителом в соответствии с вариантом реализации настоящего раскрытия;
[0040] ФИГ. 13 представляет собой график, показывающий результаты анализа ELISA по стимулированию секреции IFN-γ из NK-клеток мышиным антителом в соответствии с вариантом реализации настоящего раскрытия;
[0041] ФИГ. 14 - график результатов анализа ингибирования роста опухоли in vivo мышиным антителом согласно варианту реализации настоящего раскрытия;
[0042] ФИГ. 15 представляет собой график, показывающий результаты анализа ELISA связывания гуманизированного антитела с PVRIG человека в соответствии с вариантом реализации настоящего раскрытия;
[0043] ФИГ. 16 представляет собой график, показывающий результаты анализа связывания гуманизированного антитела к PVRIG человека на поверхности клеточной мембраны в соответствии с вариантом реализации настоящего раскрытия;
[0044] ФИГ. 17 представляет собой график, показывающий результаты анализа связывания гуманизированного антитела к PVRIG Масаса fascicularis на поверхности клеточной мембраны в соответствии с вариантом реализации настоящего раскрытия;
[0045] ФИГ. 18 представляет собой график показывающий результаты анализа блокирования связывания PVRIG человека с CD112 человека гуманизированным антителом в соответствии с вариантом реализации настоящего раскрытия; и
[0046] ФИГ. 19 представляет собой график, показывающий результаты стимулирования цитотоксичности РВМСчеловека гуманизированным антителом в соответствии с вариантом реализации настоящего раскрытия.
ОПИСАНИЕ ВАРИАНТОВ РЕАЛИЗАЦИИ ИЗОБРЕТЕНИЯ
[0047] Ниже будут подробно описаны варианты реализации настоящего раскрытия, которые предназначены для пояснения настоящего раскрытия и не должны толковаться как ограничивающие настоящее раскрытие. Кроме того, некоторые термины здесь описаны и интерпретированы для облегчения понимания специалистами в данной области техники и не должны толковаться как ограничивающие объем настоящего раскрытия.
[0048] Как используется в настоящем документе, термин "антитело" относится к протеину, который способен взаимодействовать с антигеном (например, путем связывания, стерического препятствия, стабилизации пространственного распределения) и включает две тяжелые цепи и две легкие цепи, соединенные дисульфидными связями. Каждая тяжелая цепь включает вариабельную область тяжелой цепи (сокращенно VH) и константную область тяжелой цепи. Константная область тяжелой цепи включает три домена CHI, СН2 и СН3. Каждая легкая цепь включает вариабельную область легкой цепи (сокращенно VL general) и константную область легкой цепи. Константная область легкой цепи включает один домен CL. Вариабельная область тяжелой цепи и вариабельная область легкой цепи могут быть дополнительно разделены на комплементарные определяющие области (CDR). Каждая вариабельная область тяжелой цепи и каждая вариабельная область легкой цепи имеют три области CDR, CDR1, CDR2 и CDR3 от аминоконцевой части до карбоксильной части. Как используется в данном документе, три области CDR тяжелой цепи обозначены как HCDR1, HCDR2 и HCDR3; три области CDR легкой цепи обозначены как LCDR1, LCDR2 и LCDR3.
[0049] Антитело в выражении "антитело или фрагмент антитела", упомянутом здесь, включает, но не ограничивается моноклональными антителами, человеческими антителами, гуманизированными антителами, верблюжьими антителами и химерными антителами. Антитело может быть любого изотипа, например, IgG, IgE, IgM, IgD, IgA, IgY и т.д.
[0050] Термин "фрагмент антитела" относится к одному или нескольким участкам антитела, которые сохраняют способность специфически взаимодействовать с антигеном (например, путем связывания, стерического препятствия, стабилизации пространственного распределения). Примеры фрагмента антитела включают, но не ограничиваются Fab-фрагментом, моновалентным фрагментом, состоящим из доменов VL, VH, CL и СН1; F(ab)2 фрагментом, бивалентным фрагментом, включающим два Fab-фрагмента, соединенных дисульфидным мостиком в области петли; Fd-фрагментом, состоящим из доменов VH и СН1; Fv-фрагментом, состоящим из доменов VL и VH одного плеча антитела; и изолированные CDR. Кроме того, хотя два домена фрагмента Fv, VL и VH, кодируются отдельными генами, они могут быть соединены рекомбинантными способами с помощью синтетического связывающего вещества, которое позволяет им образовывать одну протеиновую цепь, где области VL и VH объединяются в пару, образуя моновалентную молекулу (называемую одноцепочечным Fv (scFv); см., например, Bird et al., (1988) Science 242:423-426; и Huston et al., (1988) Proc. Natl. Acad. Sci. 85:5879-5883). Такие одноцепочечные антитела также охватываются термином "фрагмент антитела". Эти фрагменты антител получают с помощью обычных способов, известных специалистам в данной области, и выбирают фрагменты, применимые таким же образом, как и полное антитело.
[0051] "Гуманизированное антитело" относится к: (i) антителу, полученному не из человеческого источника (например, трансгенной мыши, несущей гетерологичную иммунную систему) на основе последовательностей зародышевой линии человека, или (ii) антителу, привитому с помощью CDR, CDR вариабельного домена которого получен не из человеческого источника, а один или более каркасов вариабельного домена которого получены из человеческого источника, и константный домен которого, если присутствует, получен из человеческого источника.
[0052] Термин "химерное антитело" относится к антителу, имеющему константную область антитела, полученную из или соответствующую последовательности, обнаруженной у одного вида, и вариабельную область антитела, полученную у другого вида. Предпочтительно, константные области антитела получены из или соответствуют последовательностям, обнаруженным у человека, например, в зародышевых клетках человека или соматических клетках, а вариабельные области антитела (например, VH, VL, CDR или FR области) получены из последовательностей, обнаруженных у нечеловеческих животных, например, мышей, крыс, кроликов или хомяков.
[0053] Как используется в настоящем документе, термин "рекомбинантное антитело" включает все антитела, приготовленные, выраженные, произведенные или выделенные путем рекомбинации, например, антитела, выделенные из трансгенных или трансхромосомных животных с человеческим геном иммуноглобулина (например, антитела, выделенные из клеток хозяина, трансформированных для выражения антител, антитела, выбранные и выделенные из рекомбинантных, комбинаторных библиотек человеческих антител, и антитела, полученные, выраженные, произведенные или выделенные любым другим способом, включающим сплайсинг части или всего гена и последовательности иммуноглобулина человека с другими последовательностями ДНК. Предпочтительно, эти рекомбинантные антитела имеют вариабельные области, в которых каркасные и CDR области получены из последовательностей иммуноглобулина зародышевой линии человека.
[0054] Как используется в настоящем документе, термин "моноклональное антитело" относится к препарату молекулы антитела, состоящему из одной молекулы. Моноклональные антитела демонстрируют уникальные области связывания, которые обладают уникальной специфичностью связывания и сродством к определенному эпитопу.
[0055] С этой целью, в одном аспекте, настоящее раскрытие предоставляет антитело или фрагмент антитела, включающий комплементарные определяющие участки тяжелой цепи HCDR1, HCDR2 и HCDR3, и комплементарные определяющие участки легкой цепи LCDR1, LCDR2 и LCDR3. HCDR1 выбирается из последовательности, представленной в GYTFSSFS, GYTFSTFA или GYSFTAYT; HCDR2 выбирается из последовательности, представленной в ILPGSNST, ILPGINNT, ILPGGNNT или INPYNGGT; и HCDR3 выбирается из последовательности, представленной в SSYWFAY, STYWFAY или AREGNYYGSRGDFDY. LCDR1 выбирается из последовательности, представленной в QSLLNSGNQKNY или QTIVTN; LCDR2 выбирается из последовательности, представленной в GAS или YAS; и LCDR3 выбирается из последовательности, представленной в QNAHSYPPT, QNAHSYPPA или QQSHSWPFT. Антитело или фрагмент антитела, предусмотренные настоящим раскрытием, могут связываться с PVRIG протеином с высокой аффинностью.
[0056] По меньшей мере в некоторых вариантах реализации настоящее раскрытие обеспечивает антитело или фрагмент антитела, включающий комплементарные определяющие области тяжелой цепи и комплементарные определяющие области легкой цепи. Определяющие области комплементарности тяжелой цепи включают GYTFSSFS, ILPGSNST и SSYWFAY. Области, определяющие комплементарность легкой цепи, включают QSLLNSGNQKNY, GAS и QNAHSYPPT.
[0057] По меньшей мере в некоторых вариантах реализации настоящее раскрытие обеспечивает антитело или фрагмент антитела, включающий вариабельную область тяжелой цепи и вариабельную область легкой цепи. Вариабельная область тяжелой цепи представляет собой последовательность, представленную в SEQ ID NO: 1, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 1. Вариабельная область легкой цепи представляет собой последовательность, представленную в SEQ ID NO: 2, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 2.
[0058] Вариабельная область тяжелой цепи, т.е. последовательность, представленная в SEQ ID NO: 1, представлена следующим образом:
[0059]
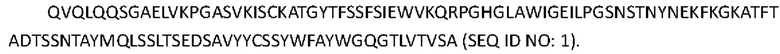
[0060] Вариабельная область легкой цепи, т.е. последовательность, представленная в SEQ ID NO: 2, представлена следующим образом:
[0061]

[0062] Последовательности предоставленного антитела или фрагмента антитела были подвергнуты модификации гуманизации, и гуманизированные антитела или фрагменты антител показаны ниже.
[0063] По меньшей мере в некоторых вариантах реализации антитело или фрагмент антитела включает вариабельную область тяжелой цепи и вариабельную область легкой цепи, вариабельная область тяжелой цепи представляет собой последовательность, представленную в SEQ ID NO: 3, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 3, а вариабельная область легкой цепи представляет собой последовательность, представленную в SEQ ID NO: 36, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 36.
[0064] По меньшей мере в некоторых вариантах реализации антитело представляет собой Антитело 4-hu1. Вариабельная область тяжелой цепи Антитела 4-hu1 представляет собой последовательность, представленную в SEQ ID NO: 3, а вариабельная область легкой цепи Антитела 4-hu1 представляет собой последовательность, представленную в SEQ ID NO: 36. По меньшей мере в некоторых предпочтительных вариантах реализации аминокислотная последовательность тяжелой цепи Антитела 4-hu1 представляет собой SEQ ID NO: 4, а соответствующая нуклеотидная последовательность, кодирующая аминокислотную последовательность тяжелой цепи, представлена в SEQ ID NO: 5; аминокислотная последовательность легкой цепи Антитела 4-hu1 представлена в SEQ ID NO: 37, а соответствующая нуклеотидная последовательность, кодирующая аминокислотную последовательность легкой цепи, представлена в SEQ ID NO: 38.
[0065] По меньшей мере в некоторых вариантах реализации антитело или фрагмент антитела включает вариабельную область тяжелой цепи и вариабельную область легкой цепи, вариабельная область тяжелой цепи представляет собой последовательность, представленную в SEQ ID NO: 6 или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 6, а вариабельная область легкой цепи представляет собой последовательность, представленную в SEQ ID NO: 36 или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 36.
[0066] По меньшей мере в некоторых предпочтительных вариантах реализации антитело представляет собой Антитело 4-hu2. Вариабельная область тяжелой цепи Антитела 4-hu2 представляет собой последовательность, представленную в SEQ ID NO: 6, а вариабельная область легкой цепи Антитела 4-hu2 представляет собой последовательность, представленную в SEQ ID NO: 36. По меньшей мере в некоторых предпочтительных вариантах реализации аминокислотная последовательность тяжелой цепи Антитела 4-hu2 представлена в SEQ ID NO: 7, а соответствующая нуклеотидная последовательность, кодирующая аминокислотную последовательность тяжелой цепи, представлена в SEQ ID NO: 8; аминокислотная последовательность легкой цепи Антитела 4-hu2 представлена в SEQ ID NO: 37, а соответствующая нуклеотидная последовательность, кодирующая аминокислотную последовательность легкой цепи, представлена в SEQ ID NO: 38.
[0067] По меньшей мере в некоторых вариантах реализации антитело или фрагмент антитела включает вариабельную область тяжелой цепи и вариабельную область легкой цепи, вариабельная область тяжелой цепи представляет собой последовательность, представленную в SEQ ID NO: 9, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 9, а вариабельная область легкой цепи представляет собой последовательность, представленную в SEQ ID NO: 36, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 36.
[0068] По меньшей мере в некоторых предпочтительных вариантах реализации антитело представляет собой Антитело 4-hu3. Вариабельная область тяжелой цепи Антитела 4-hu3 представляет собой последовательность, представленную в SEQ ID NO: 9, а вариабельная область легкой цепи Антитела 4-hu3 представляет собой последовательность, представленную в SEQ ID NO: 36. По меньшей мере в некоторых предпочтительных вариантах реализации аминокислотная последовательность тяжелой цепи Антитела 4-hu3 представлена в SEQ ID NO: 10, а соответствующая нуклеотидная последовательность, кодирующая аминокислотную последовательность тяжелой цепи, представлена в SEQ ID NO: 11; аминокислотная последовательность легкой цепи Антитела 4-hu3 представлена в SEQ ID NO: 37, а соответствующая нуклеотидная последовательность, кодирующая аминокислотную последовательность легкой цепи, представлена в SEQ ID NO: 38.
[0069] По меньшей мере в некоторых вариантах реализации антитело или фрагмент антитела включает вариабельную область тяжелой цепи и вариабельную область легкой цепи, вариабельная область тяжелой цепи представляет собой последовательность, представленную в SEQ ID NO: 12 или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 12, а вариабельная область легкой цепи представляет собой последовательность, представленную в SEQ ID NO: 36 или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 36.
[0070] По меньшей мере в некоторых предпочтительных вариантах реализации антитело представляет собой Антитело 4-hu4. Вариабельная область тяжелой цепи Антитела 4-hu4 представляет собой последовательность, представленную в SEQ ID NO: 12, а вариабельная область легкой цепи Антитела 4-hu4 представляет собой последовательность, представленную в SEQ ID NO: 36. По меньшей мере в некоторых предпочтительных вариантах реализации аминокислотная последовательность тяжелой цепи Антитела 4-hu4 представлена в SEQ ID NO: 13, а соответствующая нуклеотидная последовательность, кодирующая аминокислотную последовательность тяжелой цепи, представлена в SEQ ID NO: 14; аминокислотная последовательность легкой цепи Антитела 4-hu4 представлена в SEQ ID NO: 37, а соответствующая нуклеотидная последовательность, кодирующая аминокислотную последовательность легкой цепи, представлена в SEQ ID NO: 38.
[0071] По меньшей мере в некоторых вариантах реализации антитело или фрагмент антитела включает вариабельную область тяжелой цепи и вариабельную область легкой цепи, вариабельная область тяжелой цепи представляет собой последовательность, представленную в SEQ ID NO: 15 или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 15, а вариабельная область легкой цепи представляет собой последовательность, представленную в SEQ ID NO: 36 или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 36.
[0072] По меньшей мере в некоторых предпочтительных вариантах реализации антитело представляет собой Антитело 4-hu5. Вариабельная область тяжелой цепи Антитела 4-hu5 представляет собой последовательность, представленную в SEQ ID NO: 15, а вариабельная область легкой цепи Антитела 4-hu5 представляет собой последовательность, представленную в SEQ ID NO: 36. По меньшей мере в некоторых предпочтительных вариантах реализации аминокислотная последовательность тяжелой цепи Антитела 4-hu5 представлена в SEQ ID NO: 16, а соответствующая нуклеотидная последовательность, кодирующая аминокислотную последовательность тяжелой цепи, представлена в SEQ ID NO: 17; аминокислотная последовательность легкой цепи Антитела 4-hu5 представлена в SEQ ID NO: 37, а соответствующая нуклеотидная последовательность, кодирующая аминокислотную последовательность легкой цепи, представлена в SEQ ID NO: 38.
[0073] По меньшей мере в некоторых вариантах реализации антитело или фрагмент антитела включает вариабельную область тяжелой цепи и вариабельную область легкой цепи, вариабельная область тяжелой цепи представляет собой последовательность, представленную в SEQ ID NO: 18 или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 18, а вариабельная область легкой цепи представляет собой последовательность, представленную в SEQ ID NO: 36 или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 36.
[0074] По меньшей мере в некоторых предпочтительных вариантах реализации антитело представляет собой Антитело 4-hu6. Вариабельная область тяжелой цепи Антитела 4-hu6 представляет собой последовательность, представленную в SEQ ID NO: 18, а вариабельная область легкой цепи Антитела 4-hu6 представляет собой последовательность, представленную в SEQ ID NO: 36. По меньшей мере в некоторых предпочтительных вариантах реализации аминокислотная последовательность тяжелой цепи Антитела 4-hu6 представлена в SEQ ID NO: 19, а соответствующая нуклеотидная последовательность, кодирующая аминокислотную последовательность тяжелой цепи, представлена в SEQ ID NO: 20; аминокислотная последовательность легкой цепи Антитела 4-hu6 представлена в SEQ ID NO: 37, а соответствующая нуклеотидная последовательность, кодирующая аминокислотную последовательность легкой цепи, представлена в SEQ ID NO: 38.
[0075] По меньшей мере в некоторых вариантах реализации антитело или фрагмент антитела включает вариабельную область тяжелой цепи и вариабельную область легкой цепи, вариабельная область тяжелой цепи представляет собой последовательность, представленную в SEQ ID NO: 21 или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 21, а вариабельная область легкой цепи представляет собой последовательность, представленную в SEQ ID NO: 36 или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 36.
[0076] По меньшей мере в некоторых предпочтительных вариантах реализации антитело представляет собой Антитело 4-hu7. Вариабельная область тяжелой цепи Антитела 4-hu7 представляет собой последовательность, представленную в SEQ ID NO: 21, а вариабельная область легкой цепи Антитела 4-hu7 представляет собой последовательность, представленную в SEQ ID NO: 36. По меньшей мере в некоторых предпочтительных вариантах реализации аминокислотная последовательность тяжелой цепи Антитела 4-hu7 представлена в SEQ ID NO: 22, а соответствующая нуклеотидная последовательность, кодирующая аминокислотную последовательность тяжелой цепи, представлена в SEQ ID NO: 23; аминокислотная последовательность легкой цепи Антитела 4-hu7 представлена в SEQ ID NO: 37, а соответствующая нуклеотидная последовательность, кодирующая аминокислотную последовательность легкой цепи, представлена в SEQ ID NO: 38.
[0077] По меньшей мере в некоторых вариантах реализации антитело или фрагмент антитела включает вариабельную область тяжелой цепи и вариабельную область легкой цепи, вариабельная область тяжелой цепи представляет собой последовательность, представленную в SEQ ID NO: 24, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 24, а вариабельная область легкой цепи представляет собой последовательность, представленную в SEQ ID NO: 36, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ IDNO: 36.
[0078] По меньшей мере в некоторых предпочтительных вариантах реализации антитело или фрагмент антитела представляет собой Антитело 4-hu8. Вариабельная область тяжелой цепи Антитела 4-hu8 представляет собой последовательность, представленную в SEQ ID NO: 24, а вариабельная область легкой цепи Антитела 4-hu8 представляет собой последовательность, представленную в SEQ ID NO: 36. По меньшей мере в некоторых предпочтительных вариантах реализации аминокислотная последовательность тяжелой цепи Антитела 4-hu8 представлена в SEQ ID NO: 25, а соответствующая нуклеотидная последовательность, кодирующая аминокислотную последовательность тяжелой цепи, представлена в SEQ ID NO: 26; аминокислотная последовательность легкой цепи Антитела 4-hu8 представлена в SEQ ID N0: 37, а соответствующая нуклеотидная последовательность, кодирующая аминокислотную последовательность легкой цепи, представлена в SEQ ID NO: 38.
[0079] По меньшей мере в некоторых вариантах реализации антитело или фрагмент антитела включает вариабельную область тяжелой цепи и вариабельную область легкой цепи, вариабельная область тяжелой цепи представляет собой последовательность, представленную в SEQ ID NO: 27, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 27, а вариабельная область легкой цепи представляет собой последовательность, представленную в SEQ ID NO: 36, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 36.
[0080] По меньшей мере в некоторых предпочтительных вариантах реализации антитело представляет собой Антитело 4-hu9. Вариабельная область тяжелой цепи Антитела 4-hu9 представляет собой последовательность, представленную в SEQ ID NO: 27, а вариабельная область легкой цепи Антитела 4-hu9 представляет собой последовательность, представленную в SEQ ID NO: 36. По меньшей мере в некоторых предпочтительных вариантах реализации аминокислотная последовательность тяжелой цепи Антитела 4-hu9 представлена в SEQ ID NO: 28, а соответствующая нуклеотидная последовательность, кодирующая аминокислотную последовательность тяжелой цепи, представлена в SEQ ID NO: 29; аминокислотная последовательность легкой цепи Антитела 4-hu9 представлена в SEQ ID NO: 37, а соответствующая нуклеотидная последовательность, кодирующая аминокислотную последовательность легкой цепи, представлена в SEQ ID NO: 38.
[0081] По меньшей мере в некоторых вариантах реализации антитело или фрагмент антитела включает вариабельную область тяжелой цепи и вариабельную область легкой цепи, вариабельная область тяжелой цепи представляет собой последовательность, представленную в SEQ ID NO: 30 или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 30, а вариабельная область легкой цепи представляет собой последовательность, представленную в SEQ ID NO: 36 или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 36.
[0082] По меньшей мере в некоторых предпочтительных вариантах реализации антитело представляет собой Антитело 4-hu10. Вариабельная область тяжелой цепи Антитела 4-hu10 представляет собой последовательность, представленную в SEQ ID NO: 30, а вариабельная область легкой цепи Антитела 4-hu10 представляет собой последовательность, представленную в SEQ ID NO: 36. По меньшей мере в некоторых предпочтительных вариантах реализации аминокислотная последовательность тяжелой цепи Антитела 4-hu10 представлена в SEQ ID NO: 31, а соответствующая нуклеотидная последовательность, кодирующая аминокислотную последовательность тяжелой цепи, представлена в SEQ ID NO: 32; аминокислотная последовательность легкой цепи Антитела 4-hu10 представлена в SEQ ID NO: 37, а соответствующая нуклеотидная последовательность, кодирующая аминокислотную последовательность легкой цепи, представлена в SEQ ID NO: 38.
[0083] По меньшей мере в некоторых вариантах реализации антитело или фрагмент антитела включает вариабельную область тяжелой цепи и вариабельную область легкой цепи, вариабельная область тяжелой цепи представляет собой последовательность, представленную в SEQ ID NO: 33, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 33, а вариабельная область легкой цепи представляет собой последовательность, представленную в SEQ ID NO: 36, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 36.
[0084] По меньшей мере в некоторых предпочтительных вариантах реализации антитело представляет собой Антитело 4-hu11, вариабельная область тяжелой цепи Антитела 4-hu11 представляет собой последовательность, представленную в SEQ ID NO: 33, а вариабельная область легкой цепи Антитела 4-hu11 представляет собой последовательность, представленную в SEQ ID NO: 36. По меньшей мере в некоторых предпочтительных вариантах реализации аминокислотная последовательность тяжелой цепи Антитела 4-hu11 представлена в SEQ ID NO: 34, а соответствующая нуклеотидная последовательность, кодирующая аминокислотную последовательность тяжелой цепи, представлена в SEQ ID NO: 35; аминокислотная последовательность легкой цепи Антитела 4-hu11 представлена в SEQ ID NO: 37, а соответствующая нуклеотидная последовательность, кодирующая аминокислотную последовательность легкой цепи, представлена в SEQ ID NO: 38.
[0085] Вариабельная область тяжелой цепи Антитела 4-Hu1, т.е. последовательность, представленная в SEQ ID NO: 3, выглядит следующим образом:

[0086] Аминокислотная последовательность тяжелой цепи Антитела 4-hu1, т.е. последовательность, представленная в SEQ. ID NO: 4, выглядит следующим образом (мутация IgG4S228P):

Мутация S228P происходит в шарнирной области и способна предотвратить образование полуантитела. Так называемое полуантитело относится к феномену динамического обмена полумолекул в антителе IgG4.
[0087] Последовательность нуклеиновой кислоты, кодирующая аминокислотную последовательность тяжелой цепи Антитела 4-hu1, представлена в SEQ ID NO: 5:
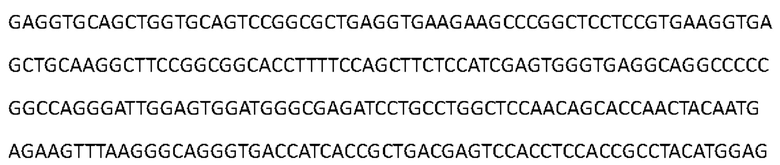

[0088] Аминокислотная последовательность вариабельной области тяжелой цепи Антитела 4-hu2 представлена в SEQ ID NO: 6:

[0089] Аминокислотная последовательность тяжелой цепи Антитела 4-hu2 представлена в SEO ID NO: 7 (мутация IgG4 S228P):


[0090] Последовательность нуклеиновой кислоты, кодирующая аминокислотную последовательность тяжелой цепи Антитела 4-hu2, представлена в SEQ ID NO: 8:

[0091] Аминокислотная последовательность вариабельной области тяжелой цепи Антитела 4-hu3 представлена в SEQ ID NO: 9:
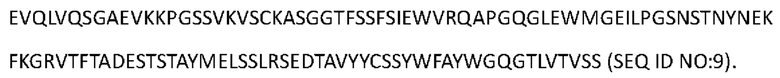
[0092] Аминокислотная последовательность тяжелой цепи Антитела 4-hu3 представлена в SEQ ID NO: 10 (мутация IgG4 S228P):

[0093] Последовательность нуклеиновой кислоты, кодирующая аминокислотную последовательность тяжелой цепи Антитела 4-hu3, представлена в SEO ID NO: 11:


[0094] Аминокислотная последовательность вариабельной области тяжелой цепи Антитела 4-hu4 представлена в SEQ ID NO: 12:
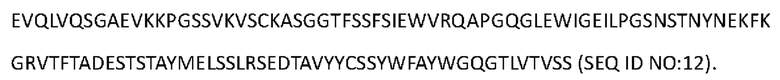
[0095] Аминокислотная последовательность тяжелой цепи Антитела 4-hu4 представлена в SEO ID NO: 13 (мутация IgG4 S228P):

[0096] Последовательность нуклеиновой кислоты, кодирующая аминокислотную последовательность тяжелой цепи Антитела 4-hu4, представлена в SEO ID NO: 14:

[0097] Аминокислотная последовательность вариабельной области тяжелой цепи Антитела 4-hu5 представлена в SEQ ID NO: 15:
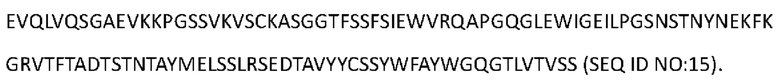
[0098] Аминокислотная последовательность тяжелой цепи Антитела 4-hu5 представлена в SEO ID NO: 16 (мутация IgG4 S228P):

[0099] Последовательность нуклеиновой кислоты, кодирующая аминокислотную последовательность тяжелой цепи Антитела 4-hu5, представлена в SEQ ID NO: 17:

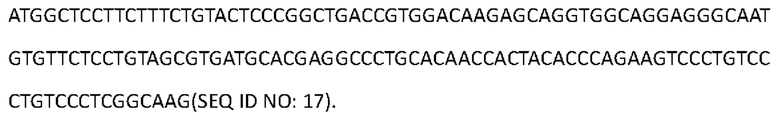
[0100] Аминокислотная последовательность вариабельной области тяжелой цепи Антитела 4-hu6 представлена в SEQ ID NO: 18:
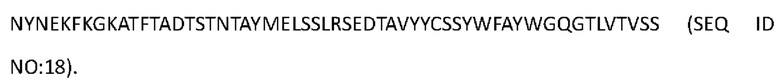
[0101] Аминокислотная последовательность тяжелой цепи Антитела 4-hu6 представлена в SEQ ID NO: 19 (мутация IgG4 S228P):

[0102] Последовательность нуклеиновой кислоты, кодирующая аминокислотную последовательность тяжелой цепи Антитела 4-hu6, представлена в SEO ID NO: 20:


[0103] Аминокислотная последовательность вариабельной области тяжелой цепи Антитела 4-hu7 представлена в SEQ ID NO: 21:
EVQLVOSGAEVKKPGSSVKVSCKASGYTFSSFSIEWVROAPGQGLEWMGEILPGSNSTNYNEKF KGRVTITADESTSTAYMΕLSSLRSEDTAVYYCSSYWFAYWGQGTLVTVSS (SEQ ID N0:21).
[0104] Аминокислотная последовательность тяжелой цепи Антитела 4-hu7 представлена в SEQ ID NO: 22 (мутация IgG4 S228P):

[0105] Последовательность нуклеиновой кислоты, кодирующая аминокислотную последовательность тяжелой цепи Антитела 4-hu7, представлена в SEQ ID NO: 23:

[0106] Аминокислотная последовательность вариабельной области тяжелой цепи Антитела 4-hu8 представлена в SEQ ID NO: 24:

[0107] Аминокислотная последовательность тяжелой цепи Антитела 4-hu8 представлена в SEQ ID NO: 25 (мутация IgG4 S228P):

[0108] Последовательность нуклеиновой кислоты, кодирующая аминокислотную последовательность тяжелой цепи Антитела 4-hu8, представлена в SEQ ID NO: 26:

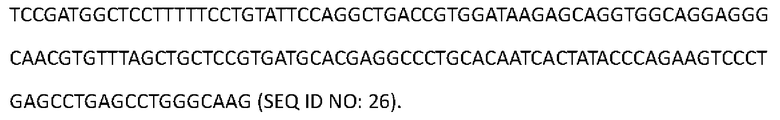
[0109] Аминокислотная последовательность вариабельной области тяжелой цепи Антитела 4-hu9 представлена в SEQ ID NO: 27:

[0110] Аминокислотная последовательность тяжелой цепи Антитела 4-hu9 представлена в SEQ ID NO: 28 (мутация IgG4 S228P):

[0111] Последовательность нуклеиновой кислоты, кодирующая аминокислотную последовательность тяжелой цепи Антитела 4-hu9, представлена в SEQ ID NO: 29:


[0112] Аминокислотная последовательность вариабельной области тяжелой цепи Антитела 4-hu10 представлена в SEQ. ID NO: 30:

[0113] Аминокислотная последовательность тяжелой цепи Антитела 4-hu10 представлена в SEQ ID NO: 31 (мутация IgG4 S228P):

[0114] Последовательность нуклеиновой кислоты, кодирующая аминокислотную последовательность тяжелой цепи Антитела 4-hu10, представлена в SEQ ID NO: 32:

[0115] Аминокислотная последовательность вариабельной области тяжелой цепи Антитела 4-hu11 представлена в SEQ ID NO: 33:
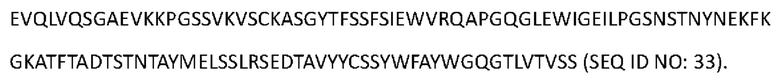
[0116] Аминокислотная последовательность тяжелой цепи Антитела 4-hu11 представлена в SEQ ID NO: 34 (мутация IgG4 S228P):

[0117] Последовательность нуклеиновой кислоты, кодирующая аминокислотную последовательность тяжелой цепи Антитела 4-hu11, представлена в SEQ ID NO: 35:


[0118] Аминокислотная последовательность вариабельной области легкой цепи гуманизированного Антитела 4 представлена в SEQ ID NO: 36:

[0119] Аминокислотная последовательность легкой цепи гуманизированного Антитела 4 изложена в SEQ ID NO: 37:

[0120] Последовательность нуклеиновой кислоты, кодирующая аминокислотную последовательность легкой цепи гуманизированного Антитела 4, изложена в SEQ ID NO: 38:

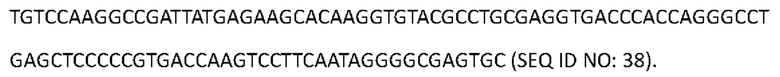
[0121] По меньшей мере в некоторых других вариантах реализации настоящее раскрытие предоставляет антитело или фрагмент антитела, включающий комплементарные определяющие области тяжелой цепи и комплементарные определяющие области легкой цепи. Комплементарные определяющие области тяжелой цепи включают GYTFSTFA, ILPGINNT и STYWFAY; и комплементарные определяющие области легкой цепи включают QSLLNSGNQKNY, GAS и QNAHSYPPT.
[0122] По меньшей мере в некоторых вариантах реализации настоящее раскрытие предоставляет антитело или фрагмент антитела, включающий вариабельную область тяжелой цепи и вариабельную область легкой цепи. Вариабельная область тяжелой цепи представляет собой последовательность, представленную в SEQ ID NO: 39, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 39; и вариабельная область легкой цепи представляет собой последовательность, представленную в SEQ ID NO: 40, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 40.
[0123] Вариабельная область тяжелой цепи, т.е. последовательность, представленная в SEQ ID NO: 39, имеет следующий вид:

[0124] Вариабельная область легкой цепи, т.е. последовательность, представленная в SEQ ID NO: 40, выглядит следующим образом:

[0125] По меньшей мере в некоторых других вариантах реализации настоящее раскрытие предоставляет антитело или фрагмент антитела, включающий комплементарные определяющие области тяжелой цепи и комплементарные определяющие области легкой цепи. Комплементарные определяющие области тяжелой цепи включают GYTFSTFA, ILPGGNNT и STYWFAY; и комплементарные определяющие области легкой цепи включают QSLLNSGNQKNY, GAS и QNAHSYPPA.
[0126] По меньшей мере в некоторых вариантах реализации настоящее раскрытие предоставляет антитело или фрагмент антитела, включающий вариабельную область тяжелой цепи и вариабельную область легкой цепи. Вариабельная область тяжелой цепи представляет собой последовательность, представленную в SEQ ID NO: 41, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 41; и вариабельная область легкой цепи представляет собой последовательность, представленную в SEQ ID NO: 42, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 42.
[0127] Вариабельная область тяжелой цепи, т.е. последовательность, представленная в SEQ ID NO: 41, имеет следующий вид:

[0128] Вариабельная область легкой цепи, т.е. последовательность, представленная в SEQ ID NO: 42, выглядит следующим образом:

[0129] По меньшей мере в некоторых других вариантах реализации настоящее раскрытие предоставляет антитело или фрагмент антитела, включающий комплементарные определяющие области тяжелой цепи и комплементарные определяющие области легкой цепи. Комплементарные определяющие области тяжелой цепи включают GYSFTAYT, INPYNGGT и AREGNYYGSRGDFDY; и комплементарные определяющие области легкой цепи включают QTIVTN, YAS и QQSHSWPFT.
[0130] По меньшей мере в некоторых вариантах реализации настоящее раскрытие предоставляет антитело или фрагмент антитела, включающий вариабельную область тяжелой цепи и вариабельную область легкой цепи. Вариабельная область тяжелой цепи представляет собой последовательность, представленную в SEQ ID NO: 43, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 43; и вариабельная область легкой цепи представляет собой последовательность, представленную в SEQ ID NO: 44, или последовательность, имеющую по меньшей мере одну аминокислотную замену по сравнению с последовательностью, представленной в SEQ ID NO: 44.
[0131] Вариабельная область тяжелой цепи, т.е. последовательность, представленная в SEQ ID NO: 43, имеет следующий вид:

[0132] Вариабельная область легкой цепи, т.е. последовательность, представленная в SEQ ID NO: 44, выглядит следующим образом:

[0133] Вышеуказанное антитело или фрагмент антитела является изолируемым. Термин "изолируемый" означает состояние, которое по существу свободно от антител или фрагментов антител, имеющих другую антигенную специфичность, чем представленное антитело. Более того, изолируемое антитело или фрагмент антитела может быть по существу свободным от других клеточных материалов и/или химических веществ. Таким образом, в некоторых аспектах предоставленное антитело представляет собой изолированное антитело, которое было выделено из антител, имеющих другую специфичность. Выделенное антитело может быть моноклональным антителом. Выделенное антитело может быть рекомбинантным моноклональным антителом. Однако выделенное антитело, которое специфически связывается с целевым эпитопом, изотипом или вариантом, может быть перекрестно реактивным с другими соответствующими антигенами, например, полученными от других видов (например, гомологов видов).
[0134] Как используется в данном документе, по меньшей мере одна аминокислотная замена может представлять собой одну аминокислотную замену, две аминокислотные замены, три аминокислотные замены, четыре аминокислотные замены, или пять аминокислотных замен, или даже больше. Согласно предпочтительному варианту реализации настоящего раскрытия, аминокислотная замена предпочтительно является консервативной аминокислотной заменой. Консервативная аминокислотная замена представляет собой аминокислотную замену, которая не ухудшает связывание антитела с PVRIG антигеном.
[0135] Консервативные аминокислотные замены включают замену аминокислоты одного класса на аминокислоту того же класса, где класс определяется общими физико-химическими свойствами боковой цепи аминокислоты и высокой частотой замен в гомологичных протеинах, встречающихся в природе, что определяется, например, стандартными матрицами частотного обмена Дейхоффа или матрицами BLOSUM. Боковые цепи аминокислот были классифицированы на шесть основных классов, включая: класс I (Cys); класс II (Ser, Thr, Pro, Ala, Gly); класс III (Asn, Asp, Gin, Glu); класс IV (His, Arg, Lys); класс V (lie, Leu, Val, Met) и класс VI (Phe, Tyr, Trp). Например, замена Asp на другой остаток класса III, такой как Asn, Gin или Glu, является консервативной заменой. Таким образом, предсказанный несущественный аминокислотный остаток в анти-EGFR антителе предпочтительно заменяется другим аминокислотным остатком из того же класса. Способы идентификации консервативных замен нуклеотидов и аминокислот, которые не устраняют связывание антигена, хорошо известны в данной области.
[0136] Настоящее раскрытие также предоставляет изолируемую нуклеиновую кислоту, кодирующую антитело или фрагмент антитела, как описано выше. Упомянутая изолируемая нуклеиновая кислота означает, что нуклеиновая кислота может быть по существу изолирована от других материалов и по существу не содержит материалов или химических веществ. Нуклеиновая кислота может присутствовать в интактных клетках, в клеточных лизатах или в частично очищенной или по существу чистой форме. Нуклеиновая кислота является "изолированной" или "по существу чистой", когда она очищена от других клеточных компонентов или других примесей, таких как другие клеточные нуклеиновые кислоты или протеин, с помощью стандартных способов, включая обработку щелочью/SDS, нанесение полос CsCl, колоночную хроматографию, агарозный гель-электрофорез и другие способы, хорошо известные в данной области.
[0137] Настоящее раскрытие также предоставляет вектор выражения, включающий нуклеиновую кислоту. Для выражения полинуклеотидов, кодирующих антитело или связывающий фрагмент, упомянутый в настоящем документе, можно использовать широкий спектр векторов выражения. Вирусные и невирусные векторы выражения могут быть использованы для получения антител в клетках-хозяевах млекопитающих. Невирусные векторы и системы включают плазмиды, эписомальные векторы, обычно с кассетами выражения для выражения протеина или РНК, и искусственные хромосомы человека (см., например, Harrington et al., (1997) Nat Genet 15:345). Например, невирусные векторы, полезные для выражения полинуклеотидов и полипептидов в клетках млекопитающих (например, человека), включают pThioHis А,В&С, pcDNA3.1/His, pEBVHis А,В&С (Invitrogen, San Diego, CA), векторы MPSV и ряд других векторов, известных в данной области для выражения другого протеина. Полезные вирусные векторы включают ретровирусы, аденовирусы, адено-ассоциированные вирусы и векторы на основе вируса герпеса, а также векторы на основе SV40, папилломавируса, НВР вируса Эпштейна Барр, вируса коровьей оспы и вируса леса Семлики (SFV). См. Brent et.al, (1995) Annu.Rev.Microbiol.49:807; и Rosenfeld et.al, (1992) Cell 68:143.
[0138] Вектор выражения содержит промотор, операбельно связанный с полинуклеотидом, кодирующим антитело или фрагмент антитела, и другие регуляторные последовательности (например, энхансеры). В некоторых вариантах реализации могут использоваться индуцибельные промоторы для предотвращения выражения вставленных последовательностей вне индуцирующих условий. Индуцибельные промоторы включают, но не ограничиваются промотором арабинозы, IacZ, промотором металлотионеина или промотором теплового шока. В дополнение к промотору для эффективного выражения антитела или фрагмента антитела могут потребоваться или необходимы дополнительные регуляторные элементы. Эти элементы обычно включают стартовый кодон ATG и прилегающий области связывания рибосомы или другую последовательность. Кроме того, экспрессия в клетках-хозяевах млекопитающих может быть повышена путем включения энхансеров, подходящих для используемой клеточной системы, таких как энхансер SV40 или энхансер CMV.
[0139] Настоящее раскрытие также предоставляет клетку-хозяина, экспрессирующую антитело или фрагмент антитела, как описано выше. Согласно одному из вариантов реализации настоящего раскрытия, клетка-хозяин содержит вектор экспрессии, как описано выше, и может быть использована для экспрессии антитела, упомянутого в настоящем раскрытии. Вектор экспрессии может быть интегрирован в геном клетки-хозяина. Клетка-хозяин может быть клеткой млекопитающего, включая, но не ограничиваясь клетками СНО или HEK293.
[0140] Клетка-хозяин для переноса и экспрессии антитела или фрагмента антитела может быть прокариоцитом или эукариоцитом. Например, прокариотической клеткой-хозяином является Е. coli. Другие подходящие микробные хозяева включают Bacillus, такие как Bacillus subtilis, и другие Enterobacteriaceae, такие как Salmonella, Serratia и различные виды Pseudomonas. В этих прокариотических хозяевах также могут быть получены векторы экспрессии, которые обычно содержат последовательности управления экспрессией, совместимые с клеткой-хозяином. Кроме того, эти клетки-хозяева также содержат промоторы, такие как система промоторов лактозы, система промоторов триптофана (trp), система промоторов β-лактамазы или система промоторов, полученная из бактериофага λ. Обычно промоторы по желанию взаимодействуют с операторными последовательностями для контроля экспрессии и имеют последовательность области связывания рибосомы или т.п.для инициации и завершения транскрипции и трансляции. Другие микроорганизмы, такие как дрожжи, также могут быть использованы для экспрессии полипептидов настоящего раскрытия. Клетки насекомых также могут быть использованы в сочетании с бакуловирусными векторами.
[0141] В некоторых предпочтительных вариантах реализации для экспрессии и получения анти-PVRIG антитела или фрагмента антитела настоящего раскрытия используются клетки-хозяева млекопитающих. Например, это могут быть линии клеток гибридомы, экспрессирующие гены эндогенных иммуноглобулинов. Предпочтительно используют линии клеток млекопитающих, несущие экзогенные векторы экспрессии, включая любые нормальные летальные или нормальные или аномальные иммортализованные клетки животных или человека, например, клетки миеломы SP2/0), клетки СНО, клетки HeLa, PER. С6 клетки, COS клетки, HKB11 клетки, NSO клетки. Например, было разработано множество подходящих линий клеток-хозяев, способных секретировать интактные иммуноглобулины, включая линии клеток СНО, различные линии клеток Cos, клетки HeLa, линии клеток миеломы, трансформированные В-клетки и гибридомы. Подходящие промоторы, содержащиеся в клетках-хозяевах, могут быть конститутивными, специфичными для типа клеток, специфичными для стадии и/или регулируемыми или настраиваемыми. Полезные промоторы включают, но не ограничиваются промотором металлотионеина, конститутивным основным поздним промотором аденовируса, дексаметазон-индуцибельным промотором MMTV, промотором SV40, промотором MRΡ poIIII, конститутивным промотором MPSV, конститутивным промотором CMV и комбинацией промотор-энхансером, известными в данной области техники.
[0142] Вектор, содержащий представляющую интерес полинуклеотидную последовательность (например, нуклеиновую кислоту, кодирующую полипептид антитела, и последовательности контроля экспрессии), может быть трансформирован в клетку-хозяина известными способами, в зависимости от типа клетки-хозяина. Например, трансфекция хлоридом кальция часто используется для прокариотических клеток, тогда как обработка фосфатом кальция или электропорация могут использоваться для других клеточных хозяев. Другими способами могут быть, например, электропорация, обработка фосфатом кальция, трансформация, опосредованная липосомами, инъекции и микроинъекции, виросомы, иммунолипосомы, поликатионные конъюгаты нуклеиновых кислот, голая ДНК, искусственные виросомы и т.д.
[0143] Настоящее раскрытие также предоставляет фармацевтическую композицию, включающую антитело, как описано выше, и фармацевтически приемлемый носитель.
[0144] Фармацевтически приемлемый носитель включает любые и все физиологически совместимые растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые агенты, изотонизирующие и задерживающие всасывание агенты и тому подобное. Предпочтительно, носитель подходит для внутривенного, внутримышечного, подкожного, парентерального, спинального или эпидермального введения (например, введение путем инъекции или инфузии). В зависимости от способа введения, активное соединение, т.е. антитело, может быть заключено в материал для защиты соединения от действия кислот и других природных условий, которые могут инактивировать соединение.
[0145] представленная фармацевтическая композиция может вводиться различными сопсобами, известными в данной области. Антитело может быть приготовлено с использованием подходящих носителей, таких как составы с контролируемым высвобождением, включая имплантаты, трансдермальные пластыри и микрокапсулированные системы доставки. Можно использовать биоразлагаемые и биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, поли(ортоэфиры) и полимолочная кислота. Многие сопосбы приготовления таких составов запатентованы или общеизвестны специалистам в данной области.
[0146] Согласно вариантам реализации настоящего раскрытия, представленное антитело может быть покрыто или совместно вводиться с материалом для предотвращения его инактивации. Например, композиция может быть введена субъекту в подходящем носителе, таком как липосома или разбавитель. Приемлемый разбавитель включает физраствор и водный буфер. Липосома включает эмульсию CGF "вода в масле в воде" и обычные липосомы.
[0147] Согласно вариантам реализации настоящего раскрытия, фармацевтически приемлемый носитель включает стерильный водный раствор или дисперсию и тому подобное, приготовленное в виде стерильного раствора для инъекций или стерильного порошка. Фармацевтическая композиция, представленная в настоящем документе, может быть сформирована в фармацевтически приемлемую лекарственную форму обычным способом, известным специалистам в данной области.
[0148] Настоящее раскрытие также предусматривает использование описанного выше антитела или описанной выше фармацевтической композиции в приготовлении лекарственного средства для лечения рака.
[0149] Настоящее раскрытие также предоставляет способ профилактики или лечения рака, включающий введение субъекту эффективного количества описанного выше антитела или описанной выше фармацевтической композиции.
[0150] "Терапевтически эффективное количество" или "эффективное количество" относится к количеству PVRIG антитела, необходимому для того, чтобы вызвать желаемый биологический ответ. Согласно настоящему раскрытию, терапевтически эффективное количество относится к количеству PVRIG антитела, необходимому для лечения и/или профилактики заболевания.
[0151] Терапевтически эффективное количество антитела, представленного в настоящем документе, зависит от относительной активности антитела и комбинации (например, активности в ингибировании роста клеток), варьируется в зависимости от субъекта и заболевания, которое лечат, веса и возраста субъекта, тяжести заболевания, способа введения и т.п., и может быть легко определено специалистом в данной области. Дозировки для введения могут находиться в следующих диапазонах: например, от примерно 1 нг до примерно 6000 мг, от примерно 5 нг до примерно 5500 мг, от примерно 10 нг до примерно 5000 мг, от примерно 20 нг до примерно 4500 мг, от примерно 30 нг до примерно 5000 мг, от примерно 300 нг до примерно 4500 мг, от примерно 500 нг до примерно 4000 мг, от примерно 1 мкг до примерно 3500 мг, от примерно 5 мкг до примерно 3000 мг, от примерно 10 мкг до примерно 2, 600 мг, от примерно 20 мкг до примерно 2,575 мг, от примерно 30 мкг до примерно 2,550 мг, от примерно 40 мкг до примерно 2,500 мг, от примерно 50 мкг до примерно 2,475 мг, от примерно 100 мкг до примерно 2,450 мг, от примерно 200 мкг до примерно 2,425 мг, от примерно 300 мкг до примерно 2,000, от примерно 400 мкг до примерно 1,175 мг, от примерно 500 мкг до примерно 1,150 мг, от примерно 0,5 мг до примерно 1125 мг, от примерно 1 мг до примерно 1100 мг, от примерно 1,25 мг до примерно 1075 мг, от примерно 1,5 мг до примерно 1050 мг, от примерно 2,0 мг до примерно 1025 мг, от примерно 2,5 мг до примерно 1000 мг, от примерно 3,0 мг до примерно 975 мг, от примерно 3,5 мг до примерно 950 мг, от примерно 4,0 мг до примерно 925 мг, от примерно 4,5 мг до примерно 900 мг, от примерно 5 мг до примерно 875 мг, от примерно 10 мг до примерно 850 мг, от примерно 20 мг до примерно 825 мг, от примерно 30 мг до примерно 800 мг, от примерно 40 мг до примерно 775 мг, от примерно 50 мг до примерно 750 мг, от примерно 100 мг до примерно 725 мг, от примерно 200 мг до примерно 700 мг, от примерно 300 мг до примерно 675 мг, от примерно 400 мг до примерно 650 мг, от примерно 500 мг или примерно 525 мг до примерно 625 мг антитела.
[0152] Как используется в данном документе, "субъект" относится к любому животному, включая грызунов, таких как мыши или крысы, приматов, таких как Масаса fascicularis, Масаса mulatta, или Homo sapiens. Предпочтительно, субъектом является примат, более предпочтительно - человек. Как используется в настоящем документе, "предотвращение" относится к способу, направленному на предотвращение или отсрочку начала заболевания.
[0153] Как используется в настоящем документе, примеры рака включают, но не ограничиваются карциномой, лимфомой, бластомой, саркомой и лейкемией. Более конкретные примеры включают: плоскоклеточный рак, мелкоклеточный рак легких, немелкоклеточный рак легких, рак желудка, рак поджелудочной железы, опухоли глиальных клеток, такие как глиобластома и нейрофиброматоз, рак шейки матки, рак яичников, рак печени, рак мочевого пузыря, гепатома, рак молочной железы, рак толстой кишки, меланома, колоректальный рак, рак эндометрия, рак слюнных желез, рак почки, почечная карцинома, рак простаты, рак вульвы, рак щитовидной железы, рак печени, а также различные виды рака головы и шеи. В конкретных воплощениях рак, который лечат или диагностируют с помощью способов, раскрытых в настоящем документе, выбран из меланомы, рака молочной железы, рака яичников, карциномы почек, рака желудочно-кишечного тракта/колонии, рака легких и рака простаты. Согласно предпочтительному варианту реализации настоящего раскрытия, рак включает, но не ограничивается меланомой, раком легкого, раком головы и шеи, колоректальным раком, раком поджелудочной железы, раком желудка, карциномой почек, раком мочевого пузыря, раком простаты, раком молочной железы, раком яичников или раком печени.
[0154] Настоящее раскрытие также предоставляет фармацевтическую комбинацию, которая может содержать другие препараты для лечения рака, в дополнение к антителу или фрагменту антитела, упомянутому выше. Композицию можно вводить отдельно или в составе комбинированной терапии, т.е. в сочетании с другими агентами. Например, комбинированная терапия может включать композицию, представленную в настоящем документе, и по меньшей мере один или более дополнительных терапевтических агентов, таких как противораковые агенты, описанные в настоящем раскрытии. Композицию можно вводить в сочетании с лучевой терапией и/или хирургическим вмешательством. Конкретная комбинация анти-EGFR антител может также вводиться отдельно или последовательно, с дополнительными терапевтическими агентами или без них.
[0155] Например, дополнительные препараты для лечения рака могут представлять собой антитела, такие как анти-CTLA-4 антитело, анти-PD-LI антитело, анти-LAG-3 антитело, анти-TIM-3 антитело, анти-BTLA антитело или другие комбинации противоопухолевых антител.
[0156] Примерные антитела, представленные в настоящем документе, включая мышиные или гуманизированные антитела, способны связываться с PVRIG человека протеином с высокой аффинностью и способствовать гибели опухолевых клеток РВМС человека. Кроме того, представленные антитела также проявляют ингибирующую рост опухоли активность in vivo.
[0157] Специалистам в данной области техники будет понятно, что следующие примеры являются лишь иллюстрацией настоящего раскрытия и не должны рассматриваться как ограничивающие объем настоящего раскрытия. Примеры, в которых конкретные методики или условия не указаны, осуществляются в соответствии с методиками или условиями, описанными в литературе по данному направлению или в соответствии с инструкциями к продукту. Все использованные агенты или инструменты, производители которых не указаны, являются обычными коммерчески доступными продуктами.
[0158] Пример 1
[0159] (I) Приготовление моноклональных антител против PVRIG человека
[0160] Моноклональные антитела против PVRIG были приготовлены по обычной технологии слияния клеток гибридомы, которая была слегка скорректирована, следующим образом:
[0161] (1) Приготовление рекомбинантного слитого протеина
[0162] Полноразмерный ген, кодирующий PVRIG человека протеин (как указано в SEQ. ID NO: 45), был искусственно синтезирован, фрагмент протеина, указанный в SEQ ID NO: 46, был амплифицирован с помощью PCR, фрагмент протеина константной области IgG1 человека (как указано в SEQ ID NO: 47) вводили на С-конце представляющей интерес последовательности и клонировали в вектор pLVX-IRES-zsGreen, клеточную линию HEK293T инфицировали упаковочным лентивирусом для получения сверхэкспрессированного слитого PVRIG человека IgG1 Fc протеина, затем супернатант клеточной культуры собирали после обычной культуры клеток и очищали способом аффинной хроматографии Protein G (GE Healthcare) для получения рекомбинантного слияния PVRIG человека IgG1 Fc протеина с чистотой более 90%. Масс-спектрометрическая идентификация показала, что пептидный фрагмент интересующего протеина соответствовал PVRIG последовательности, что подтвердило, что рекомбинантный протеин был рекомбинантным слитым PVRIG человека IgG1 Fc протеином.
[0163] Полная аминокислотная последовательность PVRIG человека: 

[0164] Аминокислотная последовательность внеклеточного сегмента PVRIG человека: (41-172)

[0165] Аминокислотная последовательность константной области IgG1 человека:

[0166] Аналогичным образом был получен рекомбинантный слитый Fc CD112 человека IgG1 Fc протеин. Полноразмерная аминокислотная последовательность CD112 человека была представлена в SEQ ID NO: 48, а аминокислотная последовательность внеклеточного сегмента CD112 человека была представлена в SEQ ID NO: 49.
[0167] Полноразмерная аминокислотная последовательность CD112 человека: 

[0168] Аминокислотная последовательность внеклеточного сегмента CD112 человека: (32-360)

[0169] Аналогичным образом был получен слитый Масаса fascicularis PVRIG человека IgG1 Fc протеин. Аминокислотная последовательность PVRIG Масаса fascicularis была представлена в SEQ ID NO: 50, а последовательность внеклеточного сегмента PVRIG Масаса fascicularis была представлена в SEQ ID NO: 51.
[0170] Аминокислотная последовательность PVRIG Масаса fascicularis:

(SEQ ID NO: 50)
[0171] Последовательность внеклеточного сегмента PVRIG Масаса fascicularis: (39-171)

[0172] (2) Иммунизация животных, слияние клеток гибридомы и скрининг клонов
[0173] Полученный выше рекомбинантный слитый PVRIG человека IgG1 Fc протеин смешивали в достаточном количестве с равным объемом полного адъюванта Фрейнда (CFA). Смесь использовали для иммунизации самок мышей BALB/c в возрасте 8-10 недель (доступных от Shanghai SLAC Laboratory Animal Co., Ltd., с массой тела около 20 г) в первый раз путем внутрибрюшинного введения 40-60 мкг рекомбинантного слияния PVRIG человека IgG1 Fc протеина на мышь; иммунизация проводилась каждые 2 недели, и такая же доза рекомбинантного слияния PVRIG человека IgG1 Fc протеина смешивалась с равным объемом неполного адъюванта Фрейнда; после иммунизации в общей сложности 5 раз, когда титр сыворотки мыши, измеренный с помощью ELISA, был не менее 1:105, для усиления иммунизации вводили 40-60 мкг рекомбинантного слитого PVRIG человека IgG1 Fc протеина. На третий день после бустинга спленоциты были выделены с помощью стандартных способов и слиты с клетками мышиной миеломы SP2/0 (номер доступа ATCC CRL-1581).
[0174] Четыре линии клеток гибридомы были получены путем проведения скрининга субклонов на клетках гибридомы, которые показали положительные сигналы, определенные как с помощью ИФА, так и проточной цитометрии.
[0175] (3) Экспрессия и очистка моноклональных антител
[0176] Моноклональные антитела были получены путем внутрибрюшинной инокуляции мышей; самки мышей BALB/c в возрасте 8-10 недель были иммунизированы внутрибрюшинно 500 мкл стерильного жидкого парафина; через неделю 1×106 клеток гибридомы были введены внутрибрюшинно; примерно через 7-10 дней асцит был собран, и супернатант был собран высокоскоростным центрифугированием. Супернатант, полученный вышеуказанным способом, очищали с помощью аффинной хроматографии на Протеине G для получения моноклональных антител с чистотой более 95%.
[0177] (II) Последовательности вариабельной области анти-PVRIG моноклональных антител
[0178] Клетки кандидатной гибридомы культивировали до общего числа 106, и клетки собирали центрифугированием при 800 об/мин в течение 10 мин. Тотальную РНК выделяли с помощью набора Trizol (Invitrogen) и использовали в качестве шаблона для синтеза библиотеки кДНК (Invitrogen) способом обратной транскрипции. Последовательность нуклеиновой кислоты вариабельной области, соответствующая клеткам гибридомы, была амплифицирована способом PCR с использованием кДНК в качестве шаблона. Затем аминокислотные последовательности вариабельной области тяжелой и легкой цепи (включая CDR-последовательности) клеток гибридомы были получены путем секвенирования.
[0179] Аминокислотные последовательности были идентифицированы, и идентифицированные последовательности антител были соответственно следующими:
[0180] Последовательность вариабельной области тяжелой цепи Антитела 4 была представлена в SEQ ID NO: 1, а последовательность вариабельной области легкой цепи Антитела 4 была представлена в SEQ ID NO: 2; последовательность вариабельной области тяжелой цепи Антитела 1 была представлена в SEQ ID NO: 39, а последовательность вариабельной области легкой цепи Антитела 1 была представлена в SEQ ID NO: 40; последовательность вариабельной области тяжелой цепи Антитела 2 была представлена в SEQ ID NO: 41, а последовательность вариабельной области легкой цепи Антитела 2 была представлена в SEQ ID NO: 42; последовательность вариабельной области тяжелой цепи Антитела 3 была представлена в SEQ ID NO: 43, а последовательность вариабельной области легкой цепи Антитела 3 была представлена в SEQ ID NO: 44.
[0181] Вышеупомянутые антитела, обозначенные как Антитело 1, Антитело 2, Антитело 3 и Антитело 4, все были мышиными антителами.
[0182] (III) Экспрессия и очистка гуманизированных антител:
[0183] Различные антитела, описанные выше, могут быть гуманизированы. С Антителом 4 в качестве примера, полученные гуманизированные антитела включают, но не ограничиваются Антителами 4-hu1 - 4-hu11, и могут быть приготовлены и очищены в общем следующим образом. Вектор, содержащий последовательности нуклеиновых кислот НС и LC, кодирующие гуманизированные антитела, получали путем искусственного генного синтеза, использовали подходящие клетки-хозяева, такие как HEK293 или СНО, и проводили транзиторную или стабильную трансфекцию с использованием системы экспрессии, способной выделять антитела, с оптимальным заданным соотношением векторов НС: LC (например, 1:1 или 1:2). Затем супернатант клеточной культуры собирали и очищали с помощью аффинной хроматографии белка А для получения гуманизированных антител с чистотой более 95%.
[0184] Пример 2
[0185] В Примере 2 различные антитела, приготовленные в Примере 1, оценивались на активность и специфичность связывания с антигеном. В качестве изотипного контроля использовали мышиный IgG1, к (доступен от Biolegend).
[0186] Активность мышиных антител из разных клонов в связывании с слитым PVRIG-Fc человека протеином оценивали способом ELISA. Экспериментальная процедура была следующей:
[0187] Рекомбинантный слитый PVRIG человека IgG1 Fc протеин был разведен до 1,0 мкг/мл в 1×PBS (фосфатный буфер), разведение было нанесено на 96-луночный ELISA планшет на ночь при 4°С по 100 мкл/лунку, планшет был промыт 3 раза PBST (0,05% Tween20-PBS) и блокирован 1% BSA, затем инкубирован при 37°С в течение 2 ч. Планшет промывали 3 раза PBST, и добавляли мышиные антитела (Антитело 1, Антитело 2, Антитело 3 или Антитело 4), разведенные в нескольких соотношениях, соответственно, до 8 концентраций, и смесь инкубировали при 37°С в течение 1 ч, используя мышиный IgG1 (доступен от Biolegend) в качестве отрицательного контроля при 100 мкл/лунку. Планшет промывали 3 раза PBST, затем в каждую лунку добавляли 100 мкл меченного пероксидазой хрена (HRP) козьего анти-мышиного IgG антитела (доступно от BOSTER, разведение 1:10000), и смесь инкубировали при 37°С в течение 1 часа. После промывки в каждую лунку добавляли 100 мкл раствора субстрата ТМБ для развития в течение 10-15 мин в темноте, добавляли 100 мкл стоп-раствора (2М⋅H2SO4) в каждую лунку, измеряли раствор с помощью микропланшетного ридера сразу после остановки реакции и считывали абсорбцию при 450 нм (OD450). Результаты показаны на ФИГ. 1.
[0188] Как видно из ФИГ. 1, все мышиные антитела из разных клонов были способны специфически связываться со слитым PVRIG-Fc человека протеином.
[0189] Активность мышиных антител из разных клонов в связывании с PVRIG человека протеином на поверхности клеточной мембраны оценивали с помощью проточной цитометрии. Экспериментальная процедура была следующей:
[0190] Клеточную линию 293Т человека PVRIG собирали, промывали 1×PBS и ресуспендировали, клетки подсчитывали. Затем 2×105 клеток помещали в каждую 1,5 мл пробирку Ер, добавляли соответствующее мышиное антитело (Антитело 1, Антитело 2, Антитело 3 или Антитело 4) в различных концентрациях, смесь инкубировали при 4°С в течение 30 мин, затем промывали 1×PBS и центрифугировали при 3500 об/мин в течение 5 мин для сбора клеток. Клеточные гранулы ресуспендировали в 100 мкл 1×PBS, добавляли меченное AF647 козье антимышиное IgG антитело (доступно от Biolegend), смесь инкубировали при 4°С в течение 30 мин в темноте, затем дважды промывали 1×PBS и собирали клетки центрифугированием при 3500 об/мин в течение 5 мин. Клеточную гранулу ресуспендировали в 200 мкл I×PBS и измеряли с помощью проточного цитометра (BD LSR II).
[0191] Как видно из результатов, представленных на ФИГ. 2, мышиные антитела из разных клонов были способны специфически связываться с PVRIG человека протеином на поверхности клеточной мембраны.
[0192] Активность мышиных антител из разных клонов в блокировании связывания PVRIG человека с его лигандом CD112 оценивали с помощью проточной цитометрии. Экспериментальная процедура была следующей:
[0193] Линию клеток 293Т-человека CD112 собирали и промывали ресуспензией в 1×PBS для подсчета клеток. Затем 2×105 клеток помещали в каждую 1,5 мл пробирку Ер, добавляли соответствующее мышиное антитело (Антитело 1, Антитело 2, Антитело 3 или Антитело 4) в различных концентрациях и 10 мкг/мл слитого CD112 человека IgG1 Fc протеина, и смесь инкубировали при 4°С в течение 30 мин, затем добавляли 1×PBS и центрифугировали при 3500 об/мин в течение 5 мин для сбора клеток. Пеллет клеток ресуспендировали в 100 мкл 1×PBS, добавляли меченное AF647 мышиное античеловеческое IgG-Fc антитело (доступно от Biolegend), смесь инкубировали при 4°С в течение 30 мин в темноте, затем дважды промывали 1×PBS и собирали клетки центрифугированием при 3500 об/мин в течение 5 мин. Клеточную гранулу ресуспендировали в 200 мл I×PBS и измеряли с помощью проточного цитометра (BD LSR II).
[0194] Как видно из результатов на ФИГ. 3, мышиные антитела из разных клонов были способны эффективно блокировать связывание PVRIG человека с его лигандом CD112.
[0195] Активность мышиных антител из разных клонов в конкурентном связывании с PVRIG человека протеином оценивали с помощью проточной цитометрии. Экспериментальная процедура была следующей:
[0196] Линию клеток 293Т-человека PVRIG собирали, промывали и ресуспендировали в 1×PBS для подсчета клеток. Затем 2×105 клеток помещали в каждую 1,5 мл пробирку Ер, добавляли 10 мкг/мл контрольного антитела мыши IgG1 и мышиного антитела (Антитело 1, Антитело 2, Антитело 3 или Антитело 4) из разных клонов соответственно, смесь инкубировали при 4°С в течение 30 мин, затем промывали 1×PBS и центрифугировали при 3500 об/мин в течение 5 мин для сбора клеток. Пеллет клеток ресуспендировали в 100 мкл 1×PBS, добавляли меченное AF647 Антитело 4, смесь инкубировали при 4°С в течение 30 мин в темноте, затем дважды промывали 1×PBS и собирали клетки центрифугированием при 3500 об/мин в течение 5 мин. Клеточную гранулу ресуспендировали в 200 мкл I×PBS и измеряли с помощью проточного цитометра (BD LSR II), статистические результаты анализа показаны на ФИГ. 4.
[0197] Как видно на ФИГ. 4, Антитело 1, Антитело 2 и Антитело 4 имеют взаимосвязь в конкурентном связывании с PVRIG человека, в то время как Антитело 3 и Антитело 4 связываются с разными эпитопами PVRIG человека.
[0198] Активность мышиных антител в связывании с PVRIG Масаса fascicularis и PVRIG мыши оценивали с помощью проточной цитометрии. Экспериментальная процедура была следующей:
[0199] Клеточные линии 293T Масаса fascicularis PVRIG и 293T мыши PVRIG были собраны, промыты и ресуспендированы в 1×PBS для подсчета клеток. Затем 2×105 клеток помещали в каждую пробирку объемом 1,5 мл Ер, добавляли меченное AF647 антитело 4 (мышиное антитело), смесь инкубировали при 4°С в течение 30 мин в темноте, затем дважды промывали 1×PBS и собирали клетки центрифугированием при 3500 об/мин в течение 5 мин. Клеточную гранулу ресуспендировали в 200 мкл I×PBS и измеряли с помощью проточного цитометра (BD LSR II), статистические результаты анализа показаны на ФИГ. 5.
[0200] Как видно из ФИГ. 5, антитело 4 (мышиное антитело) было способно связываться с белком PVRIG из Масаса fascicularis, но не было способно связываться с белком PVRIG из мыши.
[0201] Активность мышиного антитела в связывании с PVRIG Масаса fascicularis оценивали методом ELISA. Экспериментальная процедура была следующей:
[0202] Рекомбинантный слитый Масаса fascicularis PVRIG человека IgG1 Fc протеин разводили до 1,0 мкг/мл в 1×PBS (фосфатный буфер), разведение помещали в 96-луночный ELISA планшет на ночь при 4°С по 100 мкл/лунку, планшет промывали 3 раза PBST (0,05% Tween20-PBS) и блокировали 1% BSA, и смесь инкубировали в течение 2 ч при 37°С. Планшет промывали 3 раза PBST, добавляли мышиное антитело (Антитело 4), разведенное в нескольких соотношениях, соответственно, для установления 11 концентраций, и смесь инкубировали при 37°С в течение 1 ч, используя мышиный IgG1 в качестве отрицательного контроля при 100 мкл/лунку. Планшет промывали 3 раза PBST, в каждую лунку добавляли 100 мкл меченного пероксидазой хрена (HRP) козьего анти-мышиного IgG антитела (доступно от BOSTER, разведение 1:10000), и смесь инкубировали при 37°С в течение 1 ч. После промывки в каждую лунку добавляли 100 мкл раствора субстрата ТМБ для развития в течение 10-15 мин в темноте, добавляли 100 мкл стоп-раствора (2М⋅H2SO4) в каждую лунку, измеряли раствор с помощью микропланшетного ридера сразу после остановки реакции и считывали абсорбцию при 450 нм (OD450). Результаты показаны на ФИГ. 6. Видно, что антитело 4, мышиное антитело, было способно связываться с PVRIG Масаса fascicularis протеином.
[0203] Активность мышиного антитела в связывании с PVRIG на поверхности мембраны клеточной линии NKG оценивали с помощью проточной цитометрии. Экспериментальная процедура была следующей:
[0204] Линию клеток NKG собирали, промывали и ресуспендировали в 1×PBS для подсчета клеток. Затем 2×105 клеток помещали в каждую 1,5 мл пробирку Ер, добавляли 10 мкл мышиной сыворотки для блокирования при 4°С в течение 30 мин, затем промывали 1×PBS и собирали клетки центрифугированием при 3500 об/мин в течение 5 мин. Клеточную гранулу ресуспендировали в 100 мкл 1×PBS, добавили меченное AF647 Антитело 4, смесь инкубировали при 4°С в течение 30 мин в темноте, затем дважды промывали 1×PBS и собирали клетки центрифугированием при 3500 об/мин в течение 5 мин. Гранулы клеток были ресуспендированы в 200 мкл I×PBS и измерены с помощью проточного цитометра (BD LSR II). Результаты измерений показаны на ФИГ. 7.
[0205] Как видно из ФИГ. 7, мышиное Антитело 4 было способно связываться с PVRIG человека протеином на поверхности клеточной линии NKG.
[0206] Активность мышиного антитела в связывании с PVRIG на поверхности мембраны лимфоцитов оценивали с помощью проточной цитометрии. Экспериментальная процедура была следующей:
[0207] Лимфоциты выделяли из нормальной периферической крови человека путем центрифугирования с использованием Ficoll (Peripheral Blood Lymphocyte Separation Medium, GE Healthcare) при 400g в течение 30 мин, и ресуспендировали в I×PBS для подсчета клеток. Затем 1×106 клеток помещали в каждую пробирку объемом 1,5 мл, добавляли 10 мкл мышиной сыворотки для блокирования при 4°С в течение 30 мин, затем дважды промывали 1×PBS и собирали клетки центрифугированием при 3500 об/мин в течение 5 мин. Гранулы клеток ресуспендировали в 100 мкл 1×PBS и добавили антитело 4, меченное AF647, античеловеческое CD3 антитело мыши, меченное PercP-Су5.5 (доступно от Biolegend), античеловеческое CD8 антитело мыши, меченное РЕ-Су7 (доступно от BD), античеловеческое CD4 антитело мыши, меченное BV421 (доступно от BD) и античеловеческое CD56 антитело мыши, меченное BV605 (доступно от Biolegend). Смесь инкубировали в течение 30 мин при 4°С в темноте, затем дважды промывали 1×PBS и собирали клетки центрифугированием при 3500 об/мин в течение 5 мин. Гранулы клеток были ресуспендированы в 200 мкл I×PBS и измерены с помощью проточного цитометра (BD LSR II). Результаты измерений показаны на ФИГ. 8.
[0208] Как видно из ФИГ. 8, мышиное Антитело 4 было способно связываться с PVRIG протеином на поверхности мембраны лимфоцитов человека.
[0209] Экспрессию CD112 на поверхности линий опухолевых клеток оценивали с помощью проточной цитометрии. Экспериментальная процедура была следующей:
[0210] Линию клеток рака толстой кишки SW620 (Shanghai Cell Bank of Chinese Academy of Sciences), линию клеток меланомы A375 (Shanghai Cell Bank of Chinese Academy of Sciences) и линию клеток рака яичников SK-OV-3 (Shanghai Cell Bank of Chinese Academy of Sciences) собирали, промывали и ресуспендировали в 1×PBS для подсчета клеток. Затем 2×105 клеток помещали в каждую пробирку объемом 1,5 мл, добавляли 10 мкл мышиной сыворотки и блокировали при 4°С в течение 30 мин, затем промывали 1×PBS и собирали клетки центрифугированием при 3500 об/мин в течение 5 мин. Клеточные гранулы ресуспендировали в 100 мкл 1×PBS, добавили АРС-меченное мышиное античеловеческое антитело CD112 (доступно от Biolegend), смесь инкубировали при 4°С в течение 30 мин в темноте, затем дважды промыли 1×PBS и собрали клетки центрифугированием при 3500 об/мин в течение 5 мин. Гранулы клеток были ресуспендированы в 200 мкл I×PBS и измерены с помощью проточного цитометра (BD LSR II). Результаты измерений показаны на ФИГ. 9.
[0211] Как видно из ФИГ. 9, лиганд CD112 PVRIG высоко экспрессировался на поверхности различных линий опухолевых клеток.
[0212] Еще один эксперимент был проведен для оценки влияния мышиных антител на повышение цитотоксичности NK-клеток. Экспериментальная процедура была следующей:
[0213] Лимфоциты были выделены из нормальной периферической крови человека путем центрифугирования с использованием Ficoll (среда для разделения лимфоцитов периферической крови, GE Healthcare) при 400g в течение 30 мин и ресуспендированы в I×PBS; NK-клетки были впоследствии выделены и очищены с помощью набора для выделения NK-клеток (Miltenyi Biotec, Human NK Cell Isolation Kit). Затем очищенные NK-клетки в качестве эффекторных клеток и меченные CFSE (доступные от Thermo Scientific) клетки линии рака толстой кишки SW620 в качестве клеток-мишеней были помещены в 96-луночный круглодонный планшет при соотношении эффекторных и мишеней 1,25:1,2. В культуральную систему добавляли 10 мкг/мл мышиного IgG1 в качестве контрольного антитела и мышиного антитела (антитело 4) соответственно, смесь центрифугировали при 250g в течение 4 мин, а затем инкубировали при 37°С в течение 4 ч. Погибшие опухолевые клетки помечали 7AAD и измеряли с помощью проточного цитометра (BD LSR II), статистические результаты анализа показаны на ФИГ. 10.
[0214] Как видно из ФИГ. 10, мышиное антитело (Антитело 4) было способно значительно усилить эффект NK-клеток по уничтожению опухолевых клеток при различных соотношениях эффекторов к мишеням.
[0215] Далее был оценен эффект мышиных антител на повышение цитотоксичности РВМС человека. Экспериментальная процедура была следующей:
[0216] Лимфоциты выделяли из нормальной периферической крови человека путем центрифугирования с использованием Ficoll (среда для выделения лимфоцитов периферической крови, GE Healthcare) при 400g в течение 30 мин и ресуспендировали в 1×PBS; затем клетки РВМС человека в качестве эффекторных клеток и CFSE (доступный от Thermo Scientific) - меченые клетки рака толстой кишки SW620 или клетки меланомы А375 или клетки рака яичников SK-OV-3 в качестве клеток-мишеней помещали в 96-луночный круглодонный планшет при соотношении эффекторных и мишеней 12. 5:1, 25:1 и 50:1, 10 мкг/мл мышиного IgG1 в качестве контрольного антитела и мышиного антитела (антитело 4) соответственно добавляли в культуральную систему, и смесь центрифугировали при 250g в течение 4 мин, а затем инкубировали при 37°С в течение 4 ч. Погибшие опухолевые клетки помечали 7AAD и измеряли с помощью проточного цитометра (BD LSR II), а статистические результаты анализа представлены на ФИГ. 11.
[0217] Как видно из ФИГ. 11, мышиное антитело Антитело 4 было способно значительно усиливать действие клеток РВМС человека на уничтожение различных опухолевых клеток при различных соотношениях эффекторов к мишеням.
[0218] Проточная цитометрия была далее использована в эксперименте для оценки влияния мышиного антитела на стимулирование экспрессии эффекторных молекул NK-клеток. Экспериментальная процедура была следующей:
[0219] Лимфоциты выделяли из нормальной периферической крови человека путем центрифугирования с использованием Ficoll (среда для выделения лимфоцитов периферической крови, GE Healthcare) при 400g в течение 30 мин и ресуспендировали в 1×PBS; затем клетки РВМС человека в качестве эффекторных клеток и клеточную линию рака толстой кишки SW620 в качестве клеток-мишеней помещали в 96-луночный круглодонный планшет при соотношении эффекторных и мишеней 25: 1, 10 мкг/мл мышиного IgG1 в качестве контрольного антитела и мышиного антитела (антитело 4) соответственно добавляли в культуральную систему, смесь центрифугировали при 250g в течение 4 мин, а затем инкубировали при 37°С в течение 24 ч. 2,5 мкг/мл моненсина (доступен от Sigma) и BV510-меченое мышиное античеловеческое антитело CD107a (доступно от Biolegend) добавляли в течение последних 4 ч инкубации. В конце инкубации клетки собирали, промывали 1×PBS и ресуспендировали в 100 мкл 1×PBS, добавляли 10 мкл мышиной сыворотки для блокирования в течение 30 мин при 4°С. Клетки промывали 1×PBS и ресуспендировали в 100 мкл 1×PBS, добавляли PercP-Су5. 5, меченного античеловеческим CD3 антителом мыши (доступно от Biolegend), РЕ-Су7-меченного античеловеческим CD8 антителом мыши (доступно от BD), BV421-меченного античеловеческим CD4 антителом мыши (доступно от BD) и BV605-меченного античеловеческим CD56 антителом мыши (доступно от Biolegend) добавляли соответственно, смесь инкубировали в течение 30 мин при 4°С в темноте, затем дважды промывали 1×PBS и собирали клетки центрифугированием при 350g в течение 5 мин. Клетки ресуспендировали в Foxp3/Transcription Factor Staining Buffer (доступен от eBioscience), добавили FITC-меченый мышиный античеловеческий IFN-γ (доступен от Biolegend) и РЕ-меченый мышиный античеловеческий TNF-α (доступен от Biolegend) и инкубировали в течение 1 ч при 4°С. Затем клетки дважды промывали 1×PBS и собирали центрифугированием при 500g в течение 5 мин. Клетки ресуспендировали в 200 мкл I×PBS и измеряли с помощью проточного цитометра (BD FACS Celesta), статистические результаты анализа показаны на ФИГ. 12.
[0220] Как видно из ФИГ. 12, мышиное антитело (Антитело 4) было способно усиливать способность NK-клеток к дегрануляции и стимулировать секрецию цитокинов.
[0221] Влияние мышиных антител на стимулирование секреции цитокинов NK-клетками оценивали в экспериментах методом ELISA. Экспериментальная процедура была следующей: лимфоциты выделяли из нормальной периферической крови человека путем центрифугирования с использованием Ficoll (среда для выделения лимфоцитов периферической крови, GE Healthcare) при 400g в течение 30 мин и ресуспендировали в I×PBS. Затем NK-клетки были выделены и очищены с помощью набора для выделения NK-клеток (Miltenyi Biotec, Human NK Cell Isolation Kit). Затем очищенные NK-клетки в качестве эффекторных клеток и клеточная линия рака толстой кишки SW620 в качестве клеток-мишеней были помещены в 96-луночный круглодонный планшет при соотношении эффекторных и мишеней 2. 5:1, 10 мкг/мл мышиного IgG1 в качестве контрольного антитела и мышиного антитела (антитело 4) были соответственно добавлены в систему культуры, смесь центрифугировали при 250g в течение 4 мин, а затем инкубировали при 37°С в течение 18 ч. Затем супернатант культуры клеток был собран, и содержание IFN-γ в супернатанте культуры было измерено с помощью набора ELISA (доступен от Dakewe). Статистические результаты анализа ELISA показаны на ФИГ. 13. Можно видеть, что мышиное антитело (антитело 4) было способно стимулировать секрецию IFN-γ NK-клетками.
[0222] Активность мышиного антитела в ингибировании роста опухоли оценивали в эксперименте. Процедура эксперимента была следующей:
[0223] Клетки рака толстой кишки SW620 (Shanghai Cell Bank of Chinese Academy of Sciences) усиливали in vitro, затем собирали и дважды промывали 1×PBS; клетки собирали центрифугированием при 800 об/мин в течение 5 мин, ресуспендировали в стерильном нормальном физиологическом растворе для подсчета клеток; концентрация клеток была скорректирована до 1×107 клеток/мл; 100 мкл клеток были инокулированы подкожно в правую подмышку 6-8 недельным мышам B-NDG; на 7-й день опухоленосные мыши были случайным образом разделены на группы, и клетки РВМС человека, выделенные с помощью Ficoll, были перелиты через хвостовую вену, с 1×107 клеток на мышь. Внутрибрюшинное введение PBS или контрольного антитела IgG мыши или мышиного антитела Antibody 4 (250 мкг/животное) начинали на следующий день, один раз в 3 дня, в общей сложности 5 раз лечения. Длинную и короткую стороны подкожной опухоли мышей измеряли каждые 3 дня и взвешивали мышей. Размер опухоли рассчитывали по эмпирической формуле (размер опухоли = длинная сторона x короткая сторона x короткая сторона/2). Результаты показаны на ФИГ. 14. Результаты показали, что мышиное антитело (антитело 4) способно ингибировать рост подкожных опухолей in vivo.
[0224] Активность гуманизированного антитела в связывании с PVRIG человека дополнительно оценивали в эксперименте способом ELISA. Экспериментальная процедура была следующей:
[0225] Рекомбинантный слитый PVRIG человека IgG1 Fc протеин разводили до 1,0 мкг/мл в 1×PBS (фосфатный буфер), разведение наносили на 96-луночный ELISA планшет на ночь при 4°С по 100 мкл/лунку, планшет промывали 3 раза PBST (0,05% Tween20-PBS) и блокировали 1% BSA, и смесь инкубировали в течение 2 ч при 37°С. Планшет промывали 3 раза PBST, и гуманизированные антитела, разведенные в нескольких соотношениях, соответственно добавляли для установления 8 концентраций, и смесь инкубировали при 37°С в течение 1 ч, используя человеческий IgG4 в качестве отрицательного контроля в 100 мкл/лунку. Планшет промывали 3 раза PBST, затем в каждую лунку добавляли 100 мкл меченного пероксидазой хрена (HRP) мышиного античеловеческого IgG4-Fc антитела (доступно от Southern Biotech, разведение 1:5000), и смесь инкубировали при 37°С в течение 1 часа. После промывки в каждую лунку добавляли 100 мкл раствора субстрата ТМБ для развития в течение 10-15 мин в темноте, добавляли 100 мкл стоп-раствора (2М⋅H2SO4) в каждую лунку, измеряли раствор с помощью микропланшетного ридера сразу после остановки реакции и считывали абсорбцию при 450 нм (OD450). Результаты показаны на ФИГ. 15.
[0226] Как видно из ФИГ. 15, все гуманизированные антитела были способны специфически связываться с PVRIG человека протеином.
[0227] Активность гуманизированных антител в связывании с PVRIG человека оценивали с помощью проточной цитометрии в эксперименте.
Клеточную линию 293Т-человеческий PVRIG собирали, промывали и ресуспендировали в I×PBS для подсчета клеток. Затем 2×105 клеток помещали в каждую пробирку объемом 1,5 мл, добавляли соответствующие гуманизированные антитела в различных концентрациях, смесь инкубировали при 4°С в течение 30 мин, затем промывали 1×PBS и центрифугировали при 3500 об/мин в течение 5 мин для сбора клеток. Клеточные гранулы ресуспендировали в 100 мкл 1×PBS, добавляли меченное AF647 мышиное античеловеческое IgG-Fc антитело (доступно от Biolegend), смесь инкубировали при 4°С в течение 30 мин в темноте, затем дважды промывали 1×PBS и собирали клетки центрифугированием при 3500 об/мин в течение 5 мин. Пеллету клеток ресуспендировали в 200 мкл I×PBS и измерили с помощью проточного цитометра (BD LSR II), а статистические результаты анализа представлены на ФИГ. 16. Видно, что все гуманизированные антитела специфически связывались с PVRIG человека протеином на поверхности клеточной мембраны.
[0229] Активность гуманизированного антитела в связывании с PVRIG Масаса fascicularis оценивали в эксперименте с помощью проточной цитометрии. Экспериментальная процедура была следующей:
[0230] Клеточную линию 293Т-Масаса fascicularis PVRIG собирали, промывали и ресуспендировали в 1×PBS для подсчета клеток. Затем 2×105 клеток помещали в каждую 1,5 мл пробирку Ер, добавляли соответствующее гуманизированное антитело в различных концентрациях, смесь инкубировали при 4°С в течение 30 мин, затем промывали 1×PBS и центрифугировали при 3500 об/мин в течение 5 мин для сбора клеток. Клеточные гранулы ресуспендировали в 100 мкл 1×PBS, добавляли меченное AF647 мышиное античеловеческое IgG-Fc антитело (доступно от Biolegend), смесь инкубировали при 4°С в течение 30 мин в темноте, затем дважды промывали 1×PBS и собирали клетки центрифугированием при 3500 об/мин в течение 5 мин. Клеточную гранулу ресуспендировали в 200 мкл I×PBS и измеряли с помощью проточного цитометра (BD LSR II), а статистические результаты анализа показаны на ФИГ. 17.
[0231] Как видно из ФИГ. 17, все гуманизированные антитела были способны специфически связываться с PVRIG Масаса fascicularis протеином на поверхности клеточной мембраны.
[0232] Активность гуманизированных антител в блокировании связывания PVRIG человека с лигандом оценивали с помощью проточной цитометрии в эксперименте. Экспериментальная процедура была следующей:
[0233] Линию клеток 293T-human CD112 собирали, промывали и ресуспендировали в 1×PBS для подсчета клеток. Затем 2×105 клеток помещали в каждую пробирку Ер объемом 1,5 мл, добавляли гуманизированное антитело и 10 мкг/мл слитого белка Fc человека PVRIG-human IgG1, и смесь инкубировали при 4°С в течение 30 мин, затем промывали 1×PBS и центрифугировали при 3500 об/мин в течение 5 мин для сбора клеток. Клеточные гранулы ресуспендировали в 100 мкл 1×PBS, добавляли меченное AF647 мышиное античеловеческое IgG-Fc антитело (доступно от Biolegend), смесь инкубировали при 4°С в течение 30 мин в темноте, затем дважды промывали 1×PBS и собирали клетки центрифугированием при 3500 об/мин в течение 5 мин. Гранулы клеток были ресуспендированы в 200 мкл I×PBS и измерены с помощью проточного цитометра (BD LSR II). Как показано на ФИГ. 18, статистические результаты анализа показывают, что все гуманизированные антитела были способны эффективно блокировать связывание человеческого PVRIG с его лигандом CD112.
[0234] Влияние гуманизированных антител на повышение цитотоксичности РВМС человека далее оценивали в эксперименте. Экспериментальная процедура была следующей:
[0235] Лимфоциты выделяли из нормальной периферической крови человека путем центрифугирования с использованием Ficoll (среда для выделения лимфоцитов периферической крови, GE Healthcare) при 400g в течение 30 мин и ресуспендировали в I×PBS; затем клетки РВМС человека в качестве эффекторных клеток и меченную CFSE (доступную от Thermo Scientific) линию клеток рака толстой кишки SW620 в качестве клеток-мишеней помещали в 96-луночный круглодонный планшет при соотношении эффекторных и мишеней 25: 1, 10 мкг/мл человеческого IgG4 в качестве контрольного антитела, мышиное антитело (антитело 4) и гуманизированное антитело соответственно добавляли в культуральную систему, смесь центрифугировали при 250g в течение 4 мин, а затем инкубировали при 37°С в течение 4 ч. Погибшие опухолевые клетки были помечены 7AAD и измерены с помощью проточного цитометра (BD LSR II), статистические результаты анализа показаны на ФИГ. 19. Можно видеть, что все гуманизированные антитела были способны в значительной степени способствовать уничтожению опухолевых клеток клетками РВМС человека.
[0236] Константы сродства были определены с помощью поверхностного плазмонного резонанса (SPR):
[0237] Кинетику связывания и константу равновесия диссоциации (KD) антител настоящего раскрытия к PVRIG человека определяли с помощью анализа поверхностного плазмонного резонанса (Biacore). Вкратце, чип СМ5 покрывали слитым белком Fc человека PVRIG-человеческого IgG1, и после блокирования растворы антител различных концентраций пропускали через чип СМ5 с фиксированной скоростью потока и связывали с антигеном в течение 2 мин, после чего переносили в буфер для диссоциации на 6 мин для определения скорости диссоциации. Кинетику анализировали с использованием модели связывания 1:1.
[0238] В экспериментах, в основном описанных выше, определенная аффинность мышиных антител была такой, как показано в таблице 1, а определенная аффинность гуманизированных антител была такой, как показано в Таблице 2.
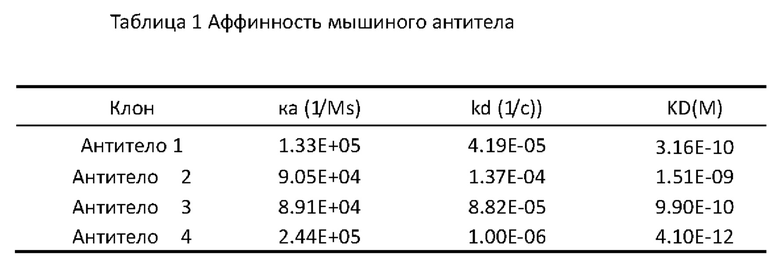
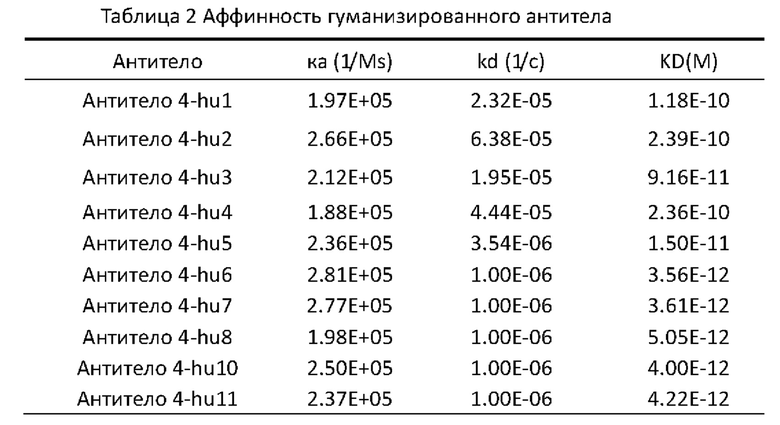
[0241] Из вышеизложенного видно, что полученные Антитела с 1 по 4 и соответствующие гуманизированные антитела Антитела 4 демонстрировали превосходное сродство к PVRIG и проявляли хороший противоопухолевый эффект.
[0242] Кроме того, описание со ссылкой на термин "воплощение", "некоторые воплощения", "пример", "конкретный пример", "некоторые примеры" или тому подобное означает, что конкретный признак, структура, материал или характеристика, описанные в сочетании с воплощением (воплощениями) или примером (примерами), включены по меньшей мере в одно воплощение или пример настоящего раскрытия. В данной спецификации иллюстративные выражения этих терминов не обязательно относятся к одному и тому же варианту воплощения или примеру. Более того, описанные конкретные признаки, структуры, материалы или характеристики могут быть объединены любым подходящим образом в любом одном или нескольких вариантах реализации или примерах. Кроме того, без взаимного противоречия, специалисты в данной области техники могут включать и комбинировать различные варианты реализации или примеры и признаки различных вариантов реализации или примеров, описанных в данной спецификации.
[0243] Хотя варианты реализации настоящего раскрытия были проиллюстрированы и описаны, следует понимать, что приведенные выше варианты являются примерными и не должны толковаться как ограничивающие настоящее раскрытие, и специалисты в данной области могут вносить различные изменения, модификации, замены и вариации в приведенные выше варианты реализации без отхода от объема настоящего раскрытия.
--->
Перечень последовательностей
<110> ХЕФЕИ ТГ ИММУНОФРМА КО., ЛТД.
<120> АНТИТЕЛО АНТИ-PVRIG ПРОТЕИН ИЛИ ФРАГМЕНТ АНТИТЕЛА И
ИХ ИСПОЛЬЗОВАНИЕ
<130> HI5210020P
<160> 51
<170> PatentIn version 3.5
<210> 1
<211> 114
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь вариабельный участок
<400> 1
QVQLQQSGAE LVKPGASVKI SCKATGYTFS SFSIEWVKQR 40
PGHGLAWIGE ILPGSNSTNY NEKFKGKATF TADTSSNTAY 80
MQLSSLTSED SAVYYCSSYW FAYWGQGTLV TVSA 114
<210> 2
<211> 113
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> лёгкая цепь вариабельный участок
<400> 2
DIVMTQSPSS LSVSAGEKVT MSCKSSQSLL NSGNQKNYLA 40
WYQQKPGQPP KLLIYGASTR ESGVPDRFTG SGSGTDFTLT 80
ISSVQAEDLA VYYCQNAHSY PPTFAAGTKL ELK 113
<210> 3
<211> 114
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь вариабельный участок
<400> 3
EVQLVQSGAE VKKPGSSVKV SCKASGGTFS SFSIEWVRQA 40
PGQGLEWMGE ILPGSNSTNY NEKFKGRVTI TADESTSTAY 80
MELSSLRSED TAVYYCARYW FAYWGQGTLV TVSS 114
<210> 4
<211> 441
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь
<400> 4
EVQLVQSGAE VKKPGSSVKV SCKASGGTFS SFSIEWVRQA 40
PGQGLEWMGE ILPGSNSTNY NEKFKGRVTI TADESTSTAY 80
MELSSLRSED TAVYYCARYW FAYWGQGTLV TVSSASTKGP 120
SVFPLAPCSR STSESTAALG CLVKDYFPEP VTVSWNSGAL 160
TSGVHTFPAV LQSSGLYSLS SVVTVPSSSL GTKTYTCNVD 200
HKPSNTKVDK RVESKYGPPC PPCPAPEFLG GPSVFLFPPK 240
PKDTLMISRT PEVTCVVVDV SQEDPEVQFN WYVDGVEVHN 280
AKTKPREEQF NSTYRVVSVL TVLHQDWLNG KEYKCKVSNK 320
GLPSSIEKTI SKAKGQPREP QVYTLPPSQE EMTKNQVSLT 360
CLVKGFYPSD IAVEWESNGQ PENNYKTTPP VLDSDGSFFL 400
YSRLTVDKSR WQEGNVFSCS VMHEALHNHY TQKSLSLSLG 440
K 441
<210> 5
<211> 1323
<212> DNA
<213> Искусственная последовательность
<220>
<221>
<222>
<223> Нуклеотидная последовательность, кодирующая тяжёлую цепь
<400> 5
gaggtgcagc tggtgcagtc cggcgctgag gtgaagaagc ccggctcctc cgtgaaggtg 60
agctgcaagg cttccggcgg caccttttcc agcttctcca tcgagtgggt gaggcaggcc 120
cccggccagg gattggagtg gatgggcgag atcctgcctg gctccaacag caccaactac 180
aatgagaagt ttaagggcag ggtgaccatc accgctgacg agtccacctc caccgcctac 240
atggagctgt cctccctgcg gtccgaggat accgctgtgt actattgtgc caggtattgg 300
ttcgcttatt ggggccaggg caccctggtg accgtgtcct ccgcctccac caagggcccc 360
tccgtgttcc ccctggctcc ctgtagccgg tccaccagcg agtccaccgc tgccctgggc 420
tgtctggtga aggactattt ccccgagccc gtgaccgtga gctggaatag cggcgccctg 480
acctccggcg tgcacacatt ccctgctgtg ctgcagagct ccggcctgta tagcctgtcc 540
tccgtggtga ccgtgccctc ctcctccctg ggcaccaaga cctatacctg caatgtggac 600
cacaagccca gcaacaccaa ggtggacaag agggtggaga gcaagtacgg ccctccttgc 660
cctccttgcc ccgctccaga gttcctgggc ggcccaagcg tgtttctgtt tcctcccaag 720
cctaaggaca ccctgatgat ctccaggacc cctgaggtga cctgcgtggt ggtggatgtg 780
tcccaggagg accctgaggt gcagttcaac tggtacgtgg atggcgtgga ggtgcacaac 840
gccaagacca agccccggga ggagcagttc aacagcacct accgggtggt gagcgtgctg 900
accgtgctgc accaggattg gctgaatggc aaggagtaca agtgcaaggt gtccaacaag 960
ggcctgccta gcagcatcga gaagaccatc tccaaggcca agggccagcc cagggagcct 1020
caggtgtata ccctgccccc tagccaggag gagatgacca agaaccaggt gagcctgacc 1080
tgcctggtga agggctttta tcctagcgat atcgctgtgg agtgggagtc caacggccag 1140
cccgagaaca actacaagac cacccctccc gtgctggata gcgacggctc cttctttctg 1200
tactcccggc tgaccgtgga taagtccagg tggcaggagg gcaatgtgtt cagctgctcc 1260
gtgatgcacg aggccctgca caatcactat acccagaagt ccctgtccct gtccctcggc 1320
aag 1323
<210> 6
<211> 114
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь вариабельный участок
<400> 6
EVQLVQSGAE VKKPGSSVKV SCKASGGTFS SFSIEWVRQA 40
PGQGLEWMGE ILPGSNSTNY NEKFKGRVTI TADESTSTAY 80
MELSSLRSED TAVYYCSSYW FAYWGQGTLV TVSS 114
<210> 7
<211> 441
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь
<400> 7
EVQLVQSGAE VKKPGSSVKV SCKASGGTFS SFSIEWVRQA 40
PGQGLEWMGE ILPGSNSTNY NEKFKGRVTI TADESTSTAY 80
MELSSLRSED TAVYYCSSYW FAYWGQGTLV TVSSASTKGP 120
SVFPLAPCSR STSESTAALG CLVKDYFPEP VTVSWNSGAL 160
TSGVHTFPAV LQSSGLYSLS SVVTVPSSSL GTKTYTCNVD 200
HKPSNTKVDK RVESKYGPPC PPCPAPEFLG GPSVFLFPPK 240
PKDTLMISRT PEVTCVVVDV SQEDPEVQFN WYVDGVEVHN 280
AKTKPREEQF NSTYRVVSVL TVLHQDWLNG KEYKCKVSNK 320
GLPSSIEKTI SKAKGQPREP QVYTLPPSQE EMTKNQVSLT 360
CLVKGFYPSD IAVEWESNGQ PENNYKTTPP VLDSDGSFFL 400
YSRLTVDKSR WQEGNVFSCS VMHEALHNHY TQKSLSLSLG 440
K 441
<210> 8
<211> 1323
<212> DNA
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь кодирующая нуклеиновая кислота
<400> 8
gaggtgcagc tggtgcagag cggcgctgag gtgaagaagc ctggcagctc cgtgaaggtg 60
agctgcaagg cctccggcgg caccttctcc agcttttcca tcgagtgggt gaggcaggcc 120
cctggccagg gactggagtg gatgggcgag atcctgcctg gcagcaattc caccaactat 180
aatgagaagt tcaagggccg ggtgaccatc accgctgacg agagcacctc caccgcttac 240
atggagctgt ccagcctgag gtccgaggac accgctgtgt actactgtag ctcctattgg 300
ttcgcttact ggggccaggg caccctggtg accgtgtcct ccgccagcac caagggccct 360
agcgtgttcc ccctggctcc ctgcagccgg agcacctctg agtccaccgc cgccctgggc 420
tgtctggtga aggattactt tcctgagccc gtgaccgtga gctggaatag cggcgctctg 480
accagcggcg tgcacacctt tcctgccgtg ctgcagagca gcggcctgta cagcctgtcc 540
tccgtggtga ccgtgcctag ctccagcctg ggcaccaaga cctatacctg taatgtggat 600
cacaagccta gcaataccaa ggtggacaag cgggtggagt ccaagtacgg ccccccctgc 660
cctccctgcc cagctcctga atttctgggc ggccccagcg tgtttctgtt tcctcccaag 720
cctaaggata ccctgatgat ctccaggacc cccgaggtga cctgtgtggt ggtggacgtg 780
tcccaggagg atcccgaggt gcagtttaac tggtacgtgg acggcgtgga ggtgcacaat 840
gccaagacca agcctaggga ggagcagttc aatagcacct acagggtggt gagcgtgctg 900
accgtgctgc accaggattg gctgaatggc aaggagtata agtgcaaggt gtccaacaag 960
ggcctgccct ccagcatcga gaagaccatc agcaaggcca agggccagcc ccgggagcct 1020
caggtttata ccctgccccc tagccaggag gagatgacca agaaccaggt gtccctgacc 1080
tgtctggtga aaggcttcta tccttccgat atcgccgtgg agtgggagtc caatggccag 1140
cccgagaaca attataagac cacccctccc gtgctggatt ccgacggcag ctttttcctg 1200
tactcccggc tgaccgtgga taagagccgg tggcaggagg gcaatgtgtt tagctgcagc 1260
gtgatgcacg aggctctgca caaccactac acccagaagt ccctgagcct gtccctgggc 1320
aag 1323
<210> 9
<211> 114
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь вариабельный участок
<400> 9
EVQLVQSGAE VKKPGSSVKV SCKASGGTFS SFSIEWVRQA 40
PGQGLEWMGE ILPGSNSTNY NEKFKGRVTF TADESTSTAY 80
MELSSLRSED TAVYYCSSYW FAYWGQGTLV TVSS 114
<210> 10
<211> 441
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь
<400> 10
EVQLVQSGAE VKKPGSSVKV SCKASGGTFS SFSIEWVRQA 40
PGQGLEWMGE ILPGSNSTNY NEKFKGRVTF TADESTSTAY 80
MELSSLRSED TAVYYCSSYW FAYWGQGTLV TVSSASTKGP 120
SVFPLAPCSR STSESTAALG CLVKDYFPEP VTVSWNSGAL 160
TSGVHTFPAV LQSSGLYSLS SVVTVPSSSL GTKTYTCNVD 200
HKPSNTKVDK RVESKYGPPC PPCPAPEFLG GPSVFLFPPK 240
PKDTLMISRT PEVTCVVVDV SQEDPEVQFN WYVDGVEVHN 280
AKTKPREEQF NSTYRVVSVL TVLHQDWLNG KEYKCKVSNK 320
GLPSSIEKTI SKAKGQPREP QVYTLPPSQE EMTKNQVSLT 360
CLVKGFYPSD IAVEWESNGQ PENNYKTTPP VLDSDGSFFL 400
YSRLTVDKSR WQEGNVFSCS VMHEALHNHY TQKSLSLSLG 440
K 441
<210> 11
<211> 1323
<212> DNA
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь кодирующая нуклеиновая кислота
<400> 11
gaggtgcagc tggtgcagag cggcgctgag gtgaagaagc ctggctccag cgtgaaggtg 60
agctgtaagg ccagcggcgg caccttctcc tccttttcca tcgagtgggt gaggcaggcc 120
cccggccagg gattggagtg gatgggcgag atcctgcccg gcagcaattc caccaattat 180
aatgagaagt ttaagggccg ggtgaccttt accgccgacg agtccacctc caccgcctat 240
atggagctga gctccctgag gagcgaggac accgccgtgt actattgcag ctcctattgg 300
tttgcttatt ggggccaggg caccctggtg accgtgtcca gcgccagcac caagggcccc 360
tccgtgtttc ccctggctcc ctgtagccgg agcaccagcg agtccaccgc tgctctgggc 420
tgtctggtga aggattactt tcccgagccc gtgaccgtgt cttggaatag cggcgccctg 480
accagcggcg tgcacacatt tcccgctgtg ctgcagagct ccggcctgta ctccctgtcc 540
tccgtggtga ccgtgccttc cagctccctg ggcaccaaga cctatacctg caacgtggat 600
cacaagccta gcaacaccaa ggtggacaag cgggtggaga gcaagtacgg ccccccctgc 660
cccccttgtc ctgctcctga gttcctgggc ggcccttccg tgtttctgtt tcctcccaag 720
cccaaggata ccctgatgat cagcaggacc cctgaggtga cctgtgtggt ggtggacgtg 780
tcccaggagg atcctgaggt gcagttcaat tggtatgtgg acggcgtgga ggtgcacaac 840
gccaagacca agccccggga ggagcagttc aattccacct atcgggtggt gagcgtgctg 900
accgtgctgc accaggactg gctgaacggc aaggagtaca agtgcaaggt gagcaataag 960
ggcctgccta gctccatcga gaagaccatc tccaaggcca agggccagcc tcgggagccc 1020
caggtttata ccctgccccc ctcccaggag gagatgacca agaaccaggt gagcctgacc 1080
tgtctggtga aaggcttcta ccccagcgat atcgctgtgg agtgggagtc caacggccag 1140
cccgagaata actataagac caccccccct gtgctggata gcgatggcag cttcttcctg 1200
tactccaggc tgaccgtgga caagtccagg tggcaggagg gcaatgtgtt ctcctgcagc 1260
gtgatgcacg aggccctgca caatcactat acccagaagt ccctgtccct gagcctgggc 1320
aag 1323
<210> 12
<211> 114
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь вариабельный участок
<400> 12
EVQLVQSGAE VKKPGSSVKV SCKASGGTFS SFSIEWVRQA 40
PGQGLEWIGE ILPGSNSTNY NEKFKGRVTF TADESTSTAY 80
MELSSLRSED TAVYYCSSYW FAYWGQGTLV TVSS 114
<210> 13
<211> 441
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь
<400> 13
EVQLVQSGAE VKKPGSSVKV SCKASGGTFS SFSIEWVRQA 40
PGQGLEWIGE ILPGSNSTNY NEKFKGRVTF TADESTSTAY 80
MELSSLRSED TAVYYCSSYW FAYWGQGTLV TVSSASTKGP 120
SVFPLAPCSR STSESTAALG CLVKDYFPEP VTVSWNSGAL 160
TSGVHTFPAV LQSSGLYSLS SVVTVPSSSL GTKTYTCNVD 200
HKPSNTKVDK RVESKYGPPC PPCPAPEFLG GPSVFLFPPK 240
PKDTLMISRT PEVTCVVVDV SQEDPEVQFN WYVDGVEVHN 280
AKTKPREEQF NSTYRVVSVL TVLHQDWLNG KEYKCKVSNK 320
GLPSSIEKTI SKAKGQPREP QVYTLPPSQE EMTKNQVSLT 360
CLVKGFYPSD IAVEWESNGQ PENNYKTTPP VLDSDGSFFL 400
YSRLTVDKSR WQEGNVFSCS VMHEALHNHY TQKSLSLSLG 440
K 441
<210> 14
<211> 1323
<212> DNA
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь кодирующая нуклеиновая кислота
<400> 14
gaggtgcagc tggtgcagtc cggcgccgag gtgaagaagc ccggctcctc cgtgaaggtg 60
agctgtaagg ccagcggcgg caccttctcc tccttttcca tcgagtgggt gaggcaggcc 120
cctggccagg gactggagtg gatcggcgag atcctgcccg gctccaatag caccaattac 180
aacgagaagt ttaagggccg ggtgaccttt accgccgacg agtccacctc caccgcctat 240
atggagctga gctccctgag gagcgaggac accgctgtgt attactgttc ctcctactgg 300
tttgcttact ggggccaggg caccctggtg accgtgtcct ccgcttccac caagggcccc 360
agcgtgtttc ccctggctcc ttgcagccgg agcacctccg agtccaccgc tgctctgggc 420
tgcctggtga aggattattt ccctgagcct gtgaccgtga gctggaacag cggcgctctg 480
acctccggcg tgcacacctt ccctgccgtg ctgcagtcca gcggcctgta tagcctgtcc 540
tccgtggtga ccgtgcccag ctccagcctg ggcaccaaga cctatacctg caacgtggat 600
cacaagcctt ccaataccaa ggtggacaag cgggtggaga gcaagtatgg ccccccctgt 660
cctccttgcc ctgctcctga gttcctgggc ggccccagcg tcttcctgtt tccccctaag 720
cccaaggaca ccctgatgat cagcaggacc cctgaggtga cctgcgtggt ggtggatgtg 780
tcccaggagg atcctgaggt gcagtttaac tggtatgtgg atggcgtgga ggtgcacaac 840
gccaagacca agccccggga ggagcagttt aacagcacct atagggtggt gagcgtgctg 900
accgtgctgc accaggattg gctgaacggc aaggagtata agtgtaaggt gagcaataag 960
ggcctgccta gctccatcga gaagaccatc agcaaggcca agggccagcc tagggagcct 1020
caggtgtaca ccctgcctcc ctcccaggag gagatgacca agaatcaggt gagcctgacc 1080
tgtctggtga agggctttta cccctccgat atcgccgtgg agtgggagtc caatggccag 1140
cctgagaaca attataagac cacccccccc gtgctggact ccgatggcag ctttttcctg 1200
tacagcaggc tgaccgtgga taagagccgg tggcaggagg gcaatgtgtt tagctgcagc 1260
gtgatgcacg aggccctgca caaccactat acccagaaga gcctgagcct gagcctcggc 1320
aag 1323
<210> 15
<211> 114
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь вариабельный участок
<400> 15
EVQLVQSGAE VKKPGSSVKV SCKASGGTFS SFSIEWVRQA 40
PGQGLEWIGE ILPGSNSTNY NEKFKGRVTF TADTSTNTAY 80
MELSSLRSED TAVYYCSSYW FAYWGQGTLV TVSS 114
<210> 16
<211> 441
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь
<400> 16
EVQLVQSGAE VKKPGSSVKV SCKASGGTFS SFSIEWVRQA 40
PGQGLEWIGE ILPGSNSTNY NEKFKGRVTF TADTSTNTAY 80
MELSSLRSED TAVYYCSSYW FAYWGQGTLV TVSSASTKGP 120
SVFPLAPCSR STSESTAALG CLVKDYFPEP VTVSWNSGAL 160
TSGVHTFPAV LQSSGLYSLS SVVTVPSSSL GTKTYTCNVD 200
HKPSNTKVDK RVESKYGPPC PPCPAPEFLG GPSVFLFPPK 240
PKDTLMISRT PEVTCVVVDV SQEDPEVQFN WYVDGVEVHN 280
AKTKPREEQF NSTYRVVSVL TVLHQDWLNG KEYKCKVSNK 320
GLPSSIEKTI SKAKGQPREP QVYTLPPSQE EMTKNQVSLT 360
CLVKGFYPSD IAVEWESNGQ PENNYKTTPP VLDSDGSFFL 400
YSRLTVDKSR WQEGNVFSCS VMHEALHNHY TQKSLSLSLG 440
K 441
<210> 17
<211> 1323
<212> DNA
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь кодирующая нуклеиновая кислота
<400> 17
gaggtgcagc tggtgcagtc cggcgctgag gtgaagaagc ctggctcctc cgtgaaggtg 60
agctgcaagg cttccggcgg caccttttcc agctttagca tcgagtgggt gaggcaggct 120
cctggccagg gcctggagtg gatcggcgag atcctgcctg gctccaacag caccaactac 180
aatgagaagt ttaagggccg ggtgaccttc accgctgaca cctccaccaa caccgcttat 240
atggagctga gctccctgcg gagcgaggat accgctgtgt actattgcag cagctattgg 300
ttcgcttact ggggccaggg caccctggtg accgtgtcct ccgccagcac caagggcccc 360
tccgtgttcc ccctggcccc ttgtagccgg tccacctccg agagcaccgc cgctctgggc 420
tgtctggtga aggattactt ccccgagcct gtgaccgtgt cttggaactc cggcgccctg 480
acctccggcg tgcacacatt ccctgctgtg ctgcagagct ccggcctgta ctccctgagc 540
agcgtggtga ccgtgccctc ctccagcctg ggcaccaaga cctacacctg taatgtggac 600
cacaagccct ccaataccaa ggtggacaag agggtggagt ccaagtacgg ccctccctgt 660
cctccttgcc ctgcccctga gtttctgggc ggccctagcg tgttcctgtt ccctcctaag 720
cctaaggaca ccctgatgat cagccggacc cccgaggtga cctgtgtggt ggtggatgtg 780
tcccaggagg atcccgaggt gcagttcaat tggtacgtgg atggcgtgga ggtgcacaac 840
gccaagacca agcctaggga ggagcagttt aactccacct acagggtggt gtccgtgctg 900
accgtgctgc accaggactg gctgaatggc aaggagtaca agtgtaaggt gagcaacaag 960
ggcctgcctt ccagcatcga gaagaccatc tccaaggcta agggccagcc tcgggagcct 1020
caggtgtaca ccctgccccc tagccaggag gagatgacca agaaccaggt gtccctgacc 1080
tgcctggtga agggctttta tccctccgac atcgccgtgg agtgggagag caatggccag 1140
cctgagaaca attataagac cacccctcct gtgctggaca gcgatggctc cttctttctg 1200
tactcccggc tgaccgtgga caagagcagg tggcaggagg gcaatgtgtt ctcctgtagc 1260
gtgatgcacg aggccctgca caaccactac acccagaagt ccctgtccct gtccctcggc 1320
aag 1323
<210> 18
<211> 114
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь вариабельный участок
<400> 18
EVQLVQSGAE VKKPGSSVKV SCKASGGTFS SFSIEWVRQA 40
PGQGLEWIGE ILPGSNSTNY NEKFKGKATF TADTSTNTAY 80
MELSSLRSED TAVYYCSSYW FAYWGQGTLV TVSS 114
<210> 19
<211> 441
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь
<400> 19
EVQLVQSGAE VKKPGSSVKV SCKASGGTFS SFSIEWVRQA 40
PGQGLEWIGE ILPGSNSTNY NEKFKGKATF TADTSTNTAY 80
MELSSLRSED TAVYYCSSYW FAYWGQGTLV TVSSASTKGP 120
SVFPLAPCSR STSESTAALG CLVKDYFPEP VTVSWNSGAL 160
TSGVHTFPAV LQSSGLYSLS SVVTVPSSSL GTKTYTCNVD 200
HKPSNTKVDK RVESKYGPPC PPCPAPEFLG GPSVFLFPPK 240
PKDTLMISRT PEVTCVVVDV SQEDPEVQFN WYVDGVEVHN 280
AKTKPREEQF NSTYRVVSVL TVLHQDWLNG KEYKCKVSNK 320
GLPSSIEKTI SKAKGQPREP QVYTLPPSQE EMTKNQVSLT 360
CLVKGFYPSD IAVEWESNGQ PENNYKTTPP VLDSDGSFFL 400
YSRLTVDKSR WQEGNVFSCS VMHEALHNHY TQKSLSLSLG 440
K 441
<210> 20
<211> 1323
<212> DNA
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь кодирующая нуклеиновая кислота
<400> 20
gaggtgcagc tggtgcagtc cggcgccgag gtgaagaagc ctggcagctc cgtgaaggtg 60
tcctgtaagg cttccggcgg caccttctcc tccttttcca tcgagtgggt gaggcaggcc 120
cccggccagg gattggagtg gatcggcgag atcctgcctg gcagcaactc caccaattat 180
aatgagaagt ttaagggcaa ggctaccttc accgctgaca ccagcaccaa taccgcctat 240
atggagctgt cctccctgcg gagcgaggac accgccgtgt attattgtag cagctattgg 300
ttcgcctact ggggccaggg caccctggtg accgtgagca gcgctagcac caagggccct 360
agcgtgtttc ctctggctcc ttgttccagg tccaccagcg agagcaccgc tgccctgggc 420
tgcctggtga aggattactt tcctgagcct gtgaccgtgt cttggaacag cggcgctctg 480
accagcggcg tgcacacctt tcccgctgtg ctgcagtcca gcggcctgta ttccctgtcc 540
agcgtggtga ccgtgcccag ctcctccctg ggcaccaaga cctatacctg caacgtggat 600
cacaagccct ccaataccaa ggtggacaag agggtggaga gcaagtacgg ccctccttgc 660
cctccctgtc ctgctcctga gttcctgggc ggcccttccg tgtttctgtt tccccctaag 720
cccaaggata ccctgatgat cagccggacc cccgaggtga cctgcgtggt ggtggatgtg 780
tcccaggagg accccgaggt gcagtttaat tggtatgtgg atggcgtgga ggtgcacaac 840
gctaagacca agcctaggga ggagcagttt aacagcacct atcgggtggt gagcgtgctg 900
accgtgctgc accaggactg gctgaacggc aaggagtaca agtgcaaggt gtccaataag 960
ggcctgccta gctccatcga gaagaccatc agcaaggcca agggccagcc tagggagccc 1020
caggtgtata ccctgccccc ttcccaggag gagatgacca agaatcaggt gagcctgacc 1080
tgtctggtga agggctttta tcctagcgac atcgctgtgg agtgggagag caatggccag 1140
cctgagaata attacaagac caccccccct gtgctggatt ccgatggcag ctttttcctg 1200
tattcccggc tgaccgtgga taagtcccgg tggcaggagg gcaacgtgtt tagctgctcc 1260
gtgatgcacg aggctctgca caatcactac acccagaagt ccctgagcct gtccctgggc 1320
aag 1323
<210> 21
<211> 114
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь вариабельный участок
<400> 21
EVQLVQSGAE VKKPGSSVKV SCKASGYTFS SFSIEWVRQA 40
PGQGLEWMGE ILPGSNSTNY NEKFKGRVTI TADESTSTAY 80
MELSSLRSED TAVYYCSSYW FAYWGQGTLV TVSS 114
<210> 22
<211> 441
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь
<400> 22
EVQLVQSGAE VKKPGSSVKV SCKASGYTFS SFSIEWVRQA 40
PGQGLEWMGE ILPGSNSTNY NEKFKGRVTI TADESTSTAY 80
MELSSLRSED TAVYYCSSYW FAYWGQGTLV TVSSASTKGP 120
SVFPLAPCSR STSESTAALG CLVKDYFPEP VTVSWNSGAL 160
TSGVHTFPAV LQSSGLYSLS SVVTVPSSSL GTKTYTCNVD 200
HKPSNTKVDK RVESKYGPPC PPCPAPEFLG GPSVFLFPPK 240
PKDTLMISRT PEVTCVVVDV SQEDPEVQFN WYVDGVEVHN 280
AKTKPREEQF NSTYRVVSVL TVLHQDWLNG KEYKCKVSNK 320
GLPSSIEKTI SKAKGQPREP QVYTLPPSQE EMTKNQVSLT 360
CLVKGFYPSD IAVEWESNGQ PENNYKTTPP VLDSDGSFFL 400
YSRLTVDKSR WQEGNVFSCS VMHEALHNHY TQKSLSLSLG 440
K 441
<210> 23
<211> 1323
<212> DNA
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь кодирующая нуклеиновая кислота
<400> 23
gaggtgcagc tggtgcagtc cggcgctgag gtgaagaagc ctggcagcag cgtgaaggtg 60
agctgcaagg cttccggcta caccttttcc agcttcagca tcgagtgggt gcggcaggct 120
cccggccagg gattggagtg gatgggcgag atcctgcctg gcagcaactc caccaattac 180
aatgagaagt tcaagggcag ggtgaccatc accgctgacg agagcacctc caccgcttac 240
atggagctga gcagcctgag gtccgaggat accgccgtgt attactgcag cagctattgg 300
tttgcctact ggggccaggg caccctggtg accgtgagct ccgccagcac caagggcccc 360
agcgtgtttc ccctggctcc ttgcagcagg tccaccagcg agagcaccgc cgctctgggc 420
tgtctggtga aggactactt ccccgagcct gtgaccgtgt cttggaatag cggcgctctg 480
accagcggcg tgcacacctt tcctgctgtg ctgcagtcct ccggcctgta ctccctgtcc 540
agcgtggtga ccgtgccctc ctcctccctg ggcaccaaga cctatacctg caatgtggat 600
cacaagccta gcaataccaa ggtggacaag agggtggaga gcaagtacgg ccccccttgc 660
cctccttgcc ccgctcctga gttcctgggc ggcccttccg tgttcctgtt tccccctaag 720
cccaaggaca ccctgatgat ctcccggacc cccgaggtga cctgtgtggt ggtggatgtg 780
agccaggagg accctgaggt gcagttcaat tggtacgtgg atggcgtgga ggtgcacaac 840
gccaagacca agcctaggga ggagcagttc aacagcacct accgggtggt gtccgtgctg 900
accgtgctgc accaggactg gctgaatggc aaggagtaca agtgcaaggt gtccaataag 960
ggcctgcctt cctccatcga gaagaccatc agcaaggcca agggccagcc tcgggagcct 1020
caggtgtata ccctgcctcc cagccaggag gagatgacca agaaccaggt gtccctgacc 1080
tgcctggtga agggctttta cccttccgat atcgccgtgg agtgggagag caatggccag 1140
cctgagaata attacaagac cacccctcct gtgctggata gcgacggcag ctttttcctg 1200
tacagccggc tgaccgtgga taagagccgg tggcaggagg gcaacgtgtt cagctgcagc 1260
gtgatgcacg aggctctgca caaccactat acccagaaga gcctgtccct gagcctgggc 1320
aag 1323
<210> 24
<211> 114
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь вариабельный участок
<400> 24
EVQLVQSGAE VKKPGSSVKV SCKASGYTFS SFSIEWVRQA 40
PGQGLEWMGE ILPGSNSTNY NEKFKGRVTF TADESTSTAY 80
MELSSLRSED TAVYYCSSYW FAYWGQGTLV TVSS 114
<210> 25
<211> 441
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь
<400> 25
EVQLVQSGAE VKKPGSSVKV SCKASGYTFS SFSIEWVRQA 40
PGQGLEWMGE ILPGSNSTNY NEKFKGRVTF TADESTSTAY 80
MELSSLRSED TAVYYCSSYW FAYWGQGTLV TVSSASTKGP 120
SVFPLAPCSR STSESTAALG CLVKDYFPEP VTVSWNSGAL 160
TSGVHTFPAV LQSSGLYSLS SVVTVPSSSL GTKTYTCNVD 200
HKPSNTKVDK RVESKYGPPC PPCPAPEFLG GPSVFLFPPK 240
PKDTLMISRT PEVTCVVVDV SQEDPEVQFN WYVDGVEVHN 280
AKTKPREEQF NSTYRVVSVL TVLHQDWLNG KEYKCKVSNK 320
GLPSSIEKTI SKAKGQPREP QVYTLPPSQE EMTKNQVSLT 360
CLVKGFYPSD IAVEWESNGQ PENNYKTTPP VLDSDGSFFL 400
YSRLTVDKSR WQEGNVFSCS VMHEALHNHY TQKSLSLSLG 440
K 441
<210> 26
<211> 1323
<212> DNA
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь кодирующая нуклеиновая кислота
<400> 26
gaggtgcagc tggtgcagag cggcgctgag gtgaagaagc ccggctccag cgtgaaggtg 60
tcctgcaagg cttccggcta taccttctcc agctttagca tcgagtgggt gaggcaggct 120
cctggccagg gcctggagtg gatgggcgag atcctgcctg gcagcaattc caccaactat 180
aacgagaagt tcaagggccg ggtgaccttc accgccgatg agtccacctc caccgcctac 240
atggagctga gctccctgcg gtccgaggat accgccgtgt actattgcag cagctattgg 300
ttcgcttatt ggggccaggg caccctggtg accgtgagct ccgcttccac caagggccct 360
tccgtgtttc ccctggcccc ctgctcccgg tccacctccg aatccaccgc cgccctgggc 420
tgcctggtga aggactactt ccctgagccc gtgaccgtgt cctggaactc cggcgccctg 480
accagcggcg tgcacacatt tcccgctgtg ctgcagtcct ccggcctgta tagcctgtcc 540
agcgtggtga ccgtgcctag ctccagcctg ggcaccaaga cctacacctg taatgtggat 600
cacaagccca gcaataccaa ggtggataag agggtggagt ccaagtacgg ccccccttgt 660
cctccttgtc ccgccccaga gttcctgggc ggcccatctg tgtttctgtt tccccccaag 720
cctaaggaca ccctgatgat ctcccggacc cctgaggtga cctgtgtggt ggtggacgtg 780
tcccaggagg atcctgaggt gcagttcaat tggtatgtgg acggcgtgga ggtgcacaac 840
gccaagacca agcccaggga ggagcagttt aacagcacct atcgggtggt gtccgtgctg 900
accgtgctgc accaggactg gctgaatggc aaggagtata agtgcaaggt gagcaataag 960
ggcctgccct ccagcatcga gaagaccatc tccaaggcta agggccagcc ccgggagcct 1020
caggtgtaca ccctgcctcc cagccaggag gagatgacca agaaccaggt gtccctgacc 1080
tgtctggtga agggctttta tcccagcgac atcgccgtgg agtgggagtc caacggccag 1140
cctgagaaca actacaagac cacccctccc gtgctggact ccgatggctc ctttttcctg 1200
tattccaggc tgaccgtgga taagagcagg tggcaggagg gcaacgtgtt tagctgctcc 1260
gtgatgcacg aggccctgca caatcactat acccagaagt ccctgagcct gagcctgggc 1320
aag 1323
<210> 27
<211> 114
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь вариабельный участок
<400> 27
EVQLVQSGAE VKKPGSSVKV SCKASGYTFS SFSIEWVRQA 40
PGQGLEWIGE ILPGSNSTNY NEKFKGRVTF TADESTSTAY 80
MELSSLRSED TAVYYCSSYW FAYWGQGTLV TVSS 114
<210> 28
<211> 441
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь
<400> 28
EVQLVQSGAE VKKPGSSVKV SCKASGYTFS SFSIEWVRQA 40
PGQGLEWIGE ILPGSNSTNY NEKFKGRVTF TADESTSTAY 80
MELSSLRSED TAVYYCSSYW FAYWGQGTLV TVSSASTKGP 120
SVFPLAPCSR STSESTAALG CLVKDYFPEP VTVSWNSGAL 160
TSGVHTFPAV LQSSGLYSLS SVVTVPSSSL GTKTYTCNVD 200
HKPSNTKVDK RVESKYGPPC PPCPAPEFLG GPSVFLFPPK 240
PKDTLMISRT PEVTCVVVDV SQEDPEVQFN WYVDGVEVHN 280
AKTKPREEQF NSTYRVVSVL TVLHQDWLNG KEYKCKVSNK 320
GLPSSIEKTI SKAKGQPREP QVYTLPPSQE EMTKNQVSLT 360
CLVKGFYPSD IAVEWESNGQ PENNYKTTPP VLDSDGSFFL 400
YSRLTVDKSR WQEGNVFSCS VMHEALHNHY TQKSLSLSLG 440
K 441
<210> 29
<211> 1323
<212> DNA
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь кодирующая нуклеиновая кислота
<400> 29
gaggtgcagc tggtgcagag cggcgccgag gtgaagaagc ccggctccag cgtgaaggtg 60
tcctgcaagg cttccggcta taccttctcc agcttcagca tcgagtgggt gaggcaggcc 120
cccggccagg gattggagtg gatcggcgag atcctgcctg gcagcaatag caccaactat 180
aatgagaagt tcaagggcag ggtgaccttc accgccgatg agagcacctc caccgcttac 240
atggagctga gcagcctgag gagcgaggat accgctgtgt attattgtag cagctattgg 300
tttgcctact ggggccaggg caccctggtg accgtgtcca gcgccagcac caagggcccc 360
agcgtgttcc ctctggcccc ttgttcccgg tccacctccg agagcaccgc cgctctgggc 420
tgcctggtga aggactattt ccctgagccc gtgaccgtga gctggaactc cggcgctctg 480
accagcggcg tgcacacctt ccctgccgtg ctgcagtcca gcggcctgta ctccctgtcc 540
agcgtggtga ccgtgcccag ctccagcctg ggcaccaaga cctacacctg caacgtggat 600
cacaagccct ccaataccaa ggtggacaag agggtggaga gcaagtatgg ccctccctgt 660
cccccctgcc ctgcccctga atttctgggc ggcccctccg tgtttctgtt tcctcctaag 720
cctaaggata ccctgatgat ctcccggacc cctgaggtga cctgtgtggt ggtggatgtg 780
agccaggagg accccgaggt gcagtttaac tggtacgtgg acggcgtgga ggtgcacaat 840
gctaagacca agccccggga ggagcagttc aatagcacct atagggtggt gagcgtgctg 900
accgtgctgc accaggactg gctgaacggc aaggagtata agtgcaaggt gagcaataag 960
ggcctgccct cctccatcga gaagaccatc tccaaggcta agggccagcc ccgggagcct 1020
caggtgtaca ccctgccccc tagccaggag gagatgacca agaatcaggt gtccctgacc 1080
tgcctggtga aaggctttta cccctccgat atcgccgtgg agtgggagtc caatggccag 1140
cctgagaaca attacaagac cacccctccc gtgctggatt ccgacggcag cttctttctg 1200
tattccaggc tgaccgtgga taagtcccgg tggcaggagg gcaacgtgtt tagctgcagc 1260
gtgatgcacg aggccctgca caatcactac acccagaagt ccctgagcct gtccctgggc 1320
aag 1323
<210> 30
<211> 114
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь вариабельный участок
<400> 30
EVQLVQSGAE VKKPGSSVKV SCKASGYTFS SFSIEWVRQA 40
PGQGLEWIGE ILPGSNSTNY NEKFKGRVTF TADTSTNTAY 80
MELSSLRSED TAVYYCSSYW FAYWGQGTLV TVSS 114
<210> 31
<211> 441
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь
<400> 31
EVQLVQSGAE VKKPGSSVKV SCKASGYTFS SFSIEWVRQA 40
PGQGLEWIGE ILPGSNSTNY NEKFKGRVTF TADTSTNTAY 80
MELSSLRSED TAVYYCSSYW FAYWGQGTLV TVSSASTKGP 120
SVFPLAPCSR STSESTAALG CLVKDYFPEP VTVSWNSGAL 160
TSGVHTFPAV LQSSGLYSLS SVVTVPSSSL GTKTYTCNVD 200
HKPSNTKVDK RVESKYGPPC PPCPAPEFLG GPSVFLFPPK 240
PKDTLMISRT PEVTCVVVDV SQEDPEVQFN WYVDGVEVHN 280
AKTKPREEQF NSTYRVVSVL TVLHQDWLNG KEYKCKVSNK 320
GLPSSIEKTI SKAKGQPREP QVYTLPPSQE EMTKNQVSLT 360
CLVKGFYPSD IAVEWESNGQ PENNYKTTPP VLDSDGSFFL 400
YSRLTVDKSR WQEGNVFSCS VMHEALHNHY TQKSLSLSLG 440
K 441
<210> 32
<211> 1323
<212> DNA
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь кодирующая нуклеиновая кислота
<400> 32
gaggtgcagc tggtgcagag cggcgctgag gtgaagaagc ctggcagcag cgtgaaggtg 60
agctgcaagg cttccggcta caccttttcc agcttcagca tcgagtgggt gaggcaggcc 120
cctggccagg gactggagtg gatcggcgag atcctgcctg gcagcaatag caccaattac 180
aacgagaagt tcaagggccg ggtgaccttc accgccgaca ccagcaccaa taccgcctat 240
atggagctga gctccctgag gtccgaggac accgctgtgt attattgctc cagctattgg 300
ttcgcctact ggggccaggg caccctggtg accgtgtcct ccgccagcac caagggcccc 360
agcgtgtttc ctctggcccc ttgcagcagg tccaccagcg agagcaccgc cgctctgggc 420
tgcctggtga aggattactt ccctgagcct gtgaccgtgt cttggaacag cggcgccctg 480
acctccggcg tgcacacatt tcctgctgtg ctgcagagca gcggcctgta tagcctgagc 540
agcgtggtga ccgtgcctag cagcagcctg ggcaccaaga cctacacctg caatgtggat 600
cacaagccct ccaacaccaa ggtggacaag cgggtggaga gcaagtacgg ccctccctgc 660
cccccctgtc ctgctcctga gtttctgggc ggccctagcg tgttcctgtt ccctcctaag 720
cccaaggaca ccctgatgat cagcaggacc cctgaggtga cctgcgtggt ggtggacgtg 780
tcccaggagg atcccgaggt gcagttcaac tggtacgtgg acggcgtgga ggtgcacaat 840
gccaagacca agcctaggga ggagcagttt aacagcacct atagggtggt gagcgtgctg 900
accgtgctgc accaggactg gctgaacggc aaggagtata agtgtaaggt gtccaacaag 960
ggcctgcctt cctccatcga gaagaccatc tccaaggcta agggccagcc tagggagcct 1020
caggtgtata ccctgcctcc ctcccaggag gagatgacca agaatcaggt gtccctgacc 1080
tgcctggtga aaggctttta ccccagcgac atcgctgtgg agtgggagag caacggccag 1140
cccgagaata attacaagac caccccccct gtgctggatt ccgacggcag cttctttctg 1200
tactccaggc tgaccgtgga caagtccagg tggcaggagg gcaacgtgtt cagctgcagc 1260
gtgatgcacg aggctctgca caaccactac acccagaagt ccctgtccct gagcctcggc 1320
aag 1323
<210> 33
<211> 114
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь вариабельный участок
<400> 33
EVQLVQSGAE VKKPGSSVKV SCKASGYTFS SFSIEWVRQA 40
PGQGLEWIGE ILPGSNSTNY NEKFKGKATF TADTSTNTAY 80
MELSSLRSED TAVYYCSSYW FAYWGQGTLV TVSS 114
<210> 34
<211> 441
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь
<400> 34
EVQLVQSGAE VKKPGSSVKV SCKASGYTFS SFSIEWVRQA 40
PGQGLEWIGE ILPGSNSTNY NEKFKGKATF TADTSTNTAY 80
MELSSLRSED TAVYYCSSYW FAYWGQGTLV TVSSASTKGP 120
SVFPLAPCSR STSESTAALG CLVKDYFPEP VTVSWNSGAL 160
TSGVHTFPAV LQSSGLYSLS SVVTVPSSSL GTKTYTCNVD 200
HKPSNTKVDK RVESKYGPPC PPCPAPEFLG GPSVFLFPPK 240
PKDTLMISRT PEVTCVVVDV SQEDPEVQFN WYVDGVEVHN 280
AKTKPREEQF NSTYRVVSVL TVLHQDWLNG KEYKCKVSNK 320
GLPSSIEKTI SKAKGQPREP QVYTLPPSQE EMTKNQVSLT 360
CLVKGFYPSD IAVEWESNGQ PENNYKTTPP VLDSDGSFFL 400
YSRLTVDKSR WQEGNVFSCS VMHEALHNHY TQKSLSLSLG 440
K 441
<210> 35
<211> 1323
<212> DNA
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь кодирующая нуклеиновая кислота
<400> 35
gaggtgcagc tggtgcagtc cggcgccgag gtgaagaagc ccggcagcag cgtgaaggtg 60
agctgcaagg cttccggcta taccttcagc tccttcagca tcgagtgggt gcggcaggct 120
cctggccagg gactggagtg gatcggcgag atcctgcccg gcagcaattc caccaactat 180
aatgagaagt tcaagggcaa ggctaccttc accgccgaca cctccaccaa taccgcctat 240
atggagctgt ccagcctgag gagcgaggac accgctgtgt actactgctc cagctactgg 300
tttgcctatt ggggccaggg caccctggtg accgtgagca gcgcttccac caagggcccc 360
tccgtgttcc ctctggcccc ctgtagcagg agcacctccg agtccaccgc tgccctgggc 420
tgcctggtga aggactattt ccctgagcct gtgaccgtgt cctggaacag cggcgctctg 480
acctccggcg tgcacacctt ccccgctgtg ctgcagtcct ccggcctgta cagcctgagc 540
tccgtggtga ccgtgcctag cagcagcctg ggcaccaaga cctatacctg caacgtggat 600
cacaagccta gcaacaccaa ggtggacaag cgggtggagt ccaagtacgg ccctccctgt 660
cccccttgcc ccgctcctga gtttctgggc ggcccttccg tgtttctgtt cccccctaag 720
cccaaggata ccctgatgat cagccggacc cccgaggtga cctgcgtggt ggtggatgtg 780
tcccaggagg accccgaggt gcagttcaac tggtacgtgg acggcgtgga ggtgcacaat 840
gccaagacca agcctcggga ggagcagttt aacagcacct atcgggtggt gagcgtgctg 900
accgtgctgc accaggattg gctgaacggc aaggagtata agtgcaaggt gtccaataag 960
ggcctgccta gcagcatcga gaagaccatc agcaaggcta agggccagcc ccgggagccc 1020
caggtttaca ccctgccccc tagccaggag gagatgacca agaatcaggt gtccctgacc 1080
tgcctggtga aaggcttcta tccctccgac atcgctgtgg agtgggagtc caatggccag 1140
cccgagaaca actacaagac cacccctccc gtgctggata gcgacggcag cttctttctg 1200
tactccaggc tgaccgtgga caagagccgg tggcaggagg gcaatgtgtt ctcctgttcc 1260
gtgatgcacg aggccctgca caatcactac acccagaagt ccctgtccct gagcctcggc 1320
aag 1323
<210> 36
<211> 113
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> лёгкая цепь вариабельный участок
<400> 36
DIVMTQSPDS LAVSLGERAT INCKSSQSLL NSGNQKNYLA 40
WYQQKPGQPP KLLIYGASTR ESGVPDRFSG SGSGTDFTLT 80
ISSLQAEDVA VYYCQNAHSY PPTFGQGTKL EIK 113
<210> 37
<211> 220
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> лёгкая цепь
<400> 37
DIVMTQSPDS LAVSLGERAT INCKSSQSLL NSGNQKNYLA 40
WYQQKPGQPP KLLIYGASTR ESGVPDRFSG SGSGTDFTLT 80
ISSLQAEDVA VYYCQNAHSY PPTFGQGTKL EIKRTVAAPS 120
VFIFPPSDEQ LKSGTASVVC LLNNFYPREA KVQWKVDNAL 160
QSGNSQESVT EQDSKDSTYS LSSTLTLSKA DYEKHKVYAC 200
EVTHQGLSSP VTKSFNRGEC 220
<210> 38
<211> 660
<212> DNA
<213> Искусственная последовательность
<220>
<221>
<222>
<223> лёгкая цепь кодирующая нуклеиновая кислота
<400> 38
gacatcgtga tgacccagag ccccgattcc ctggccgtgt ccctgggcga gagggctacc 60
atcaattgca agtcctccca gagcctgctg aatagcggca accagaagaa ctacctggcc 120
tggtaccagc agaagcccgg ccagcctcct aagctgctga tctacggcgc ctccacccgg 180
gagagcggcg ttcctgacag gtttagcggc tccggcagcg gcaccgattt caccctgacc 240
atctccagcc tgcaggccga ggacgtggcc gtgtattatt gccagaatgc ccactcctat 300
ccccccacct ttggccaggg caccaagctg gagatcaagc ggaccgtggc cgccccttcc 360
gtgtttatct tccccccctc cgacgagcag ctgaagtccg gcaccgccag cgtggtgtgt 420
ctgctgaaca acttctaccc cagggaggct aaggtgcagt ggaaggtgga taacgccctg 480
cagtccggca acagccagga gagcgtgacc gagcaggact ccaaggactc cacctactcc 540
ctgtcctcca ccctgaccct gtccaaggcc gattatgaga agcacaaggt gtacgcctgc 600
gaggtgaccc accagggcct gagctccccc gtgaccaagt ccttcaatag gggcgagtgc 660
<210> 39
<211> 114
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь вариабельный участок
<400> 39
QVQLQQSGAE LMKPGASVKI SCKATGYTFS TFAIDWIKQR 40
PGPGLAWIGE ILPGINNTNY NEKFKGKATF TADTSSNTAY 80
MHLSSLTSED SAVYYCSTYW FAYWGQGTLV TVSA 114
<210> 40
<211> 113
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> лёгкая цепь вариабельный участок
<400> 40
DIVMTQSPSS LSVSAGEKVT MTCKSSQSLL NSGNQKNYLA 40
WYQQKPGQPP KLLMYGASTR ESGVPDRFTG SGSGTDFTLT 80
ISSVQAEDLA VYYCQNAHSY PPTFGAGTKL ELK 113
<210> 41
<211> 114
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь вариабельный участок
<400> 41
QVQLQQSGAE LTKPGASVKI SCKATGYTFS TFAIDWIKQR 40
PGHGLAWIGE ILPGGNNTNY NEKFKGKATF TADTSSNTAY 80
MHLSSLTSED SAVYYCSTYW FAYWGQGTLV TVSA 114
<210> 42
<211> 113
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> лёгкая цепь вариабельный участок
<400> 42
DIVMTQSPSS LSVSAGEKVT MSCKSSQSLL NSGNQKNYLA 40
WYQQKPGQPP KLLMYGASTR ESGVPDRFTG SGSGTDFTLT 80
ISSVQAEDLA VYYCQNAHSY PPAFGAGTKL ELK 113
<210> 43
<211> 122
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> тяжёлая цепь вариабельный участок
<400> 43
EVQLQQSGPE LVKPGASMKI SCKASGYSFT AYTMNWVKQS 40
HGKNLEWIGL INPYNGGTSY NQKFKGKATL TGDKSSSTAY 80
MELLSVTSED SAVYFCAREG NYYGSRGDFD YWGQGTTLTV 120
SS 122
<210> 44
<211> 107
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> лёгкая цепь вариабельный участок
<400> 44
DILLTQSPAI LSVSPGERVS FSCRASQTIV TNILWYQQRT 40
NGSPRLLIRY ASESISGIPS RFSGSGSGTD FTLSINSVES 80
EDIAHYYCQQ SHSWPFTFGS GTKLEIK 107
<210> 45
<211> 326
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> human PVRIG full length amino acid
<400> 45
MRTEAQVPAL QPPEPGLEGA MGHRTLVLPW VLLTLCVTAG 40
TPEVWVQVRM EATELSSFTI RCGFLGSGSI SLVTVSWGGP 80
NGAGGTTLAV LHPERGIRQW APARQARWET QSSISLILEG 120
SGASSPCANT TFCCKFASFP EGSWEACGSL PPSSDPGLSA 160
PPTPAPILRA DLAGILGVSG VLLFGCVYLL HLLRRHKHRP 200
APRLQPSRTS PQAPRARAWA PSQASQAALH VPYATINTSC 240
RPATLDTAHP HGGPSWWASL PTHAAHRPQG PAAWASTPIP 280
ARGSFVSVEN GLYAQAGERP PHTGPGLTLF PDPRGPRAME 320
GPLGVR 326
<210> 46
<211> 132
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> human PVRIG extracellular segment amino acid
<400> 46
TPEVWVQVRM EATELSSFTI RCGFLGSGSI SLVTVSWGGP 40
NGAGGTTLAV LHPERGIRQW APARQARWET QSSISLILEG 80
SGASSPCANT TFCCKFASFP EGSWEACGSL PPSSDPGLSA 120
PPTPAPILRA DL 132
<210> 47
<211> 330
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> human IgG1 constant region amino acid
<400> 47
ASTKGPSVFP LAPSSKSTSG GTAALGCLVK DYFPEPVTVS 40
WNSGALTSGV HTFPAVLQSS GLYSLSSVVT VPSSSLGTQT 80
YICNVNHKPS NTKVDKKVEP KSCDKTHTCP PCPAPELLGG 120
PSVFLFPPKP KDTLMISRTP EVTCVVVDVS HEDPEVKFNW 160
YVDGVEVHNA KTKPREEQYN STYRVVSVLT VLHQDWLNGK 200
EYKCKVSNKA LPAPIEKTIS KAKGQPREPQ VYTLPPSRDE 240
LTKNQVSLTC LVKGFYPSDI AVEWESNGQP ENNYKTTPPV 280
LDSDGSFFLY SKLTVDKSRW QQGNVFSCSV MHEALHNHYT 320
QKSLSLSPGK 330
<210> 48
<211> 538
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> human CD112 full length amino acid
<400> 48
MARAAALLPS RSPPTPLLWP LLLLLLLETG AQDVRVQVLP 40
EVRGQLGGTV ELPCHLLPPV PGLYISLVTW QRPDAPANHQ 80
NVAAFHPKMG PSFPSPKPGS ERLSFVSAKQ STGQDTEAEL 120
QDATLALHGL TVEDEGNYTC EFATFPKGSV RGMTWLRVIA 160
KPKNQAEAQK VTFSQDPTTV ALCISKEGRP PARISWLSSL 200
DWEAKETQVS GTLAGTVTVT SRFTLVPSGR ADGVTVTCKV 240
EHESFEEPAL IPVTLSVRYP PEVSISGYDD NWYLGRTDAT 280
LSCDVRSNPE PTGYDWSTTS GTFPTSAVAQ GSQLVIHAVD 320
SLFNTTFVCT VTNAVGMGRA EQVIFVRETP NTAGAGATGG 360
IIGGIIAAII ATAVAATGIL ICRQQRKEQT LQGAEEDEDL 400
EGPPSYKPPT PKAKLEAQEM PSQLFTLGAS EHSPLKTPYF 440
DAGASCTEQE MPRYHELPTL EERSGPLHPG ATSLGSPIPV 480
PPGPPAVEDV SLDLEDEEGE EEEEYLDKIN PIYDALSYSS 520
PSDSYQGKGF VMSRAMYV 538
<210> 49
<211> 329
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> human CD112 extracellular segment amino acid
<400> 49
QDVRVQVLPE VRGQLGGTVE LPCHLLPPVP GLYISLVTWQ 40
RPDAPANHQN VAAFHPKMGP SFPSPKPGSE RLSFVSAKQS 80
TGQDTEAELQ DATLALHGLT VEDEGNYTCE FATFPKGSVR 120
GMTWLRVIAK PKNQAEAQKV TFSQDPTTVA LCISKEGRPP 160
ARISWLSSLD WEAKETQVSG TLAGTVTVTS RFTLVPSGRA 200
DGVTVTCKVE HESFEEPALI PVTLSVRYPP EVSISGYDDN 240
WYLGRTDATL SCDVRSNPEP TGYDWSTTSG TFPTSAVAQG 280
SQLVIHAVDS LFNTTFVCTV TNAVGMGRAE QVIFVRETPN 320
TAGAGATGG 329
<210> 50
<211> 322
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> Macaca fascicularis PVRIG amino acid
<400> 50
MRTEAQVLAL QSPEPGLEGA MGRRTLALPW VLLTLCVTAG 40
TPEVWVQVQM EATELSSFTV HCGFLGPGSI SLVTVSWGGP 80
DGAGGTKLAV LHPELGTRQW APARQARWET QSSISLALED 120
SGASSPFANT TFCCKFASFP EGSWESCGSL PPSSDPGLSA 160
PPTPVPILRA DLAGILGLSG VLLFGCGYLL HLLRRQKHRP 200
TPRLQPSHTN SQALTAQAWA PSQASQAALH DPYATINTSF 240
CPATLDTAHP NGWASLPTHA AHQPQGPAAS ASTPILARGS 280
FVSVENGLYT QAGERPPHTG PGLTLFPDCR GPRALEGRFG 320
VR 322
<210> 51
<211> 133
<212> PRT
<213> Искусственная последовательность
<220>
<221>
<222>
<223> Macaca fascicularis PVRIG extracellular segment
<400> 51
AGTPEVWVQV QMEATELSSF TVHCGFLGPG SISLVTVSWG 40
GPDGAGGTKL AVLHPELGTR QWAPARQARW ETQSSISLAL 80
EDSGASSPFA NTTFCCKFAS FPEGSWESCG SLPPSSDPGL 120
SAPPTPVPIL RAD 133
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| НУКЛЕИНОВЫЕ КИСЛОТЫ, КОДИРУЮЩИЕ АНТИТЕЛА ПРОТИВ СИАЛИРОВАННОГО АНТИГЕНА ЛЬЮИСА ЧЕЛОВЕКА | 2014 |
|
RU2791967C2 |
| АНТИТЕЛО К Myl9 | 2017 |
|
RU2741802C2 |
| АНТИТЕЛА К TIGIT И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2827445C2 |
| АНТИТЕЛО, НАЦЕЛЕННОЕ НА CD47, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2832773C1 |
| АНТИТЕЛО B7H6 И ЕГО ПРИМЕНЕНИЕ | 2022 |
|
RU2834783C2 |
| НУКЛЕИНОВЫЕ КИСЛОТЫ, КОДИРУЮЩИЕ АНТИТЕЛА ПРОТИВ СИАЛИРОВАННОГО АНТИГЕНА ЛЬЮИСА ЧЕЛОВЕКА | 2014 |
|
RU2699289C2 |
| АНТИТЕЛО ПРОТИВ LAG-3, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2017 |
|
RU2757813C2 |
| АНТИТЕЛА ПРОТИВ VISTA И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2750723C1 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА ПРОТИВ БАЗИГИНА И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2755150C2 |
| Антитело против PD-1 и его применение | 2017 |
|
RU2739610C1 |
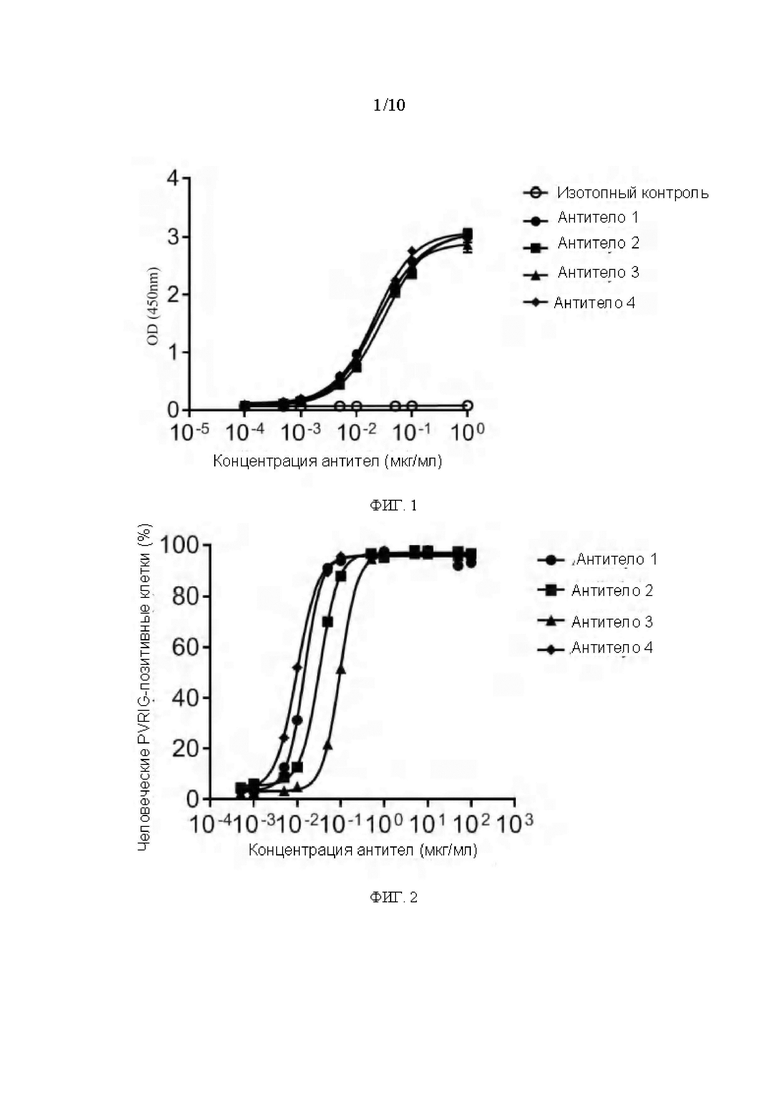
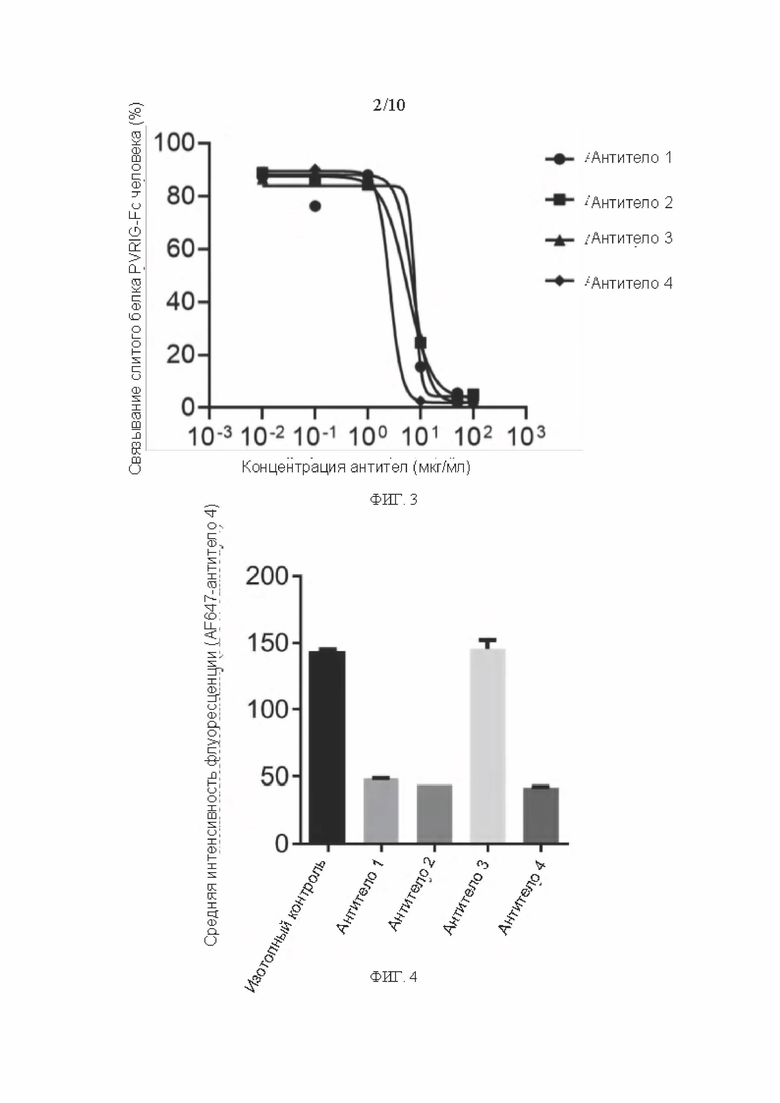
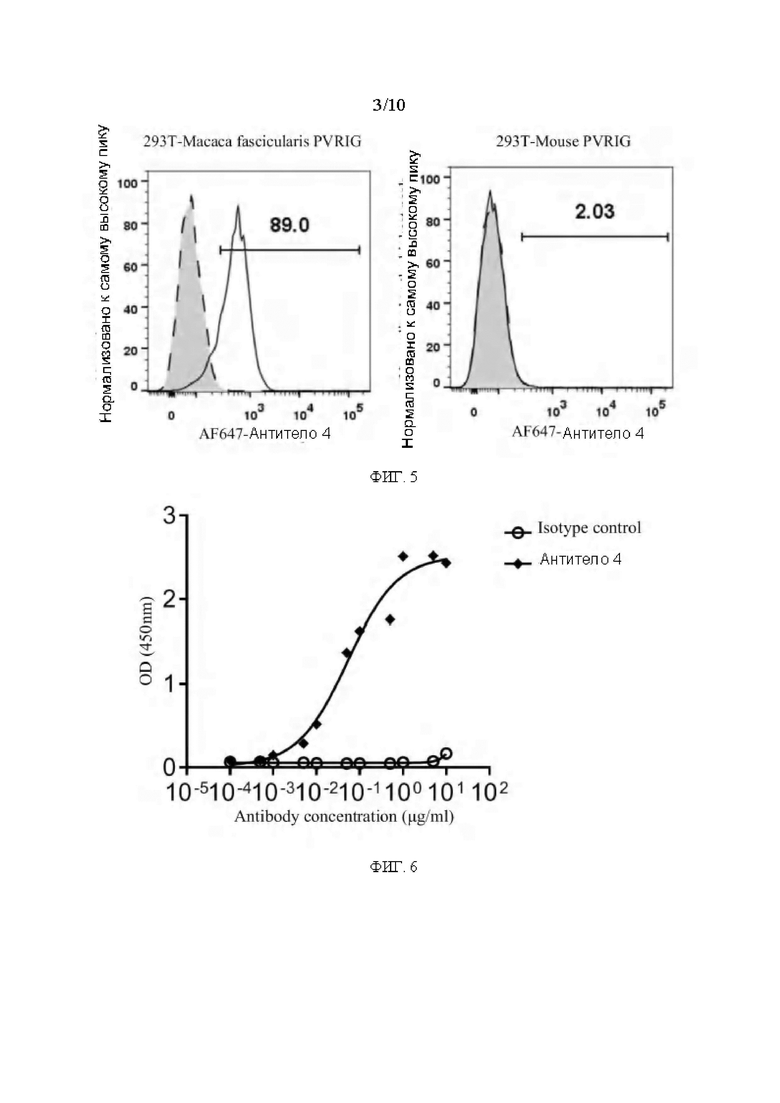
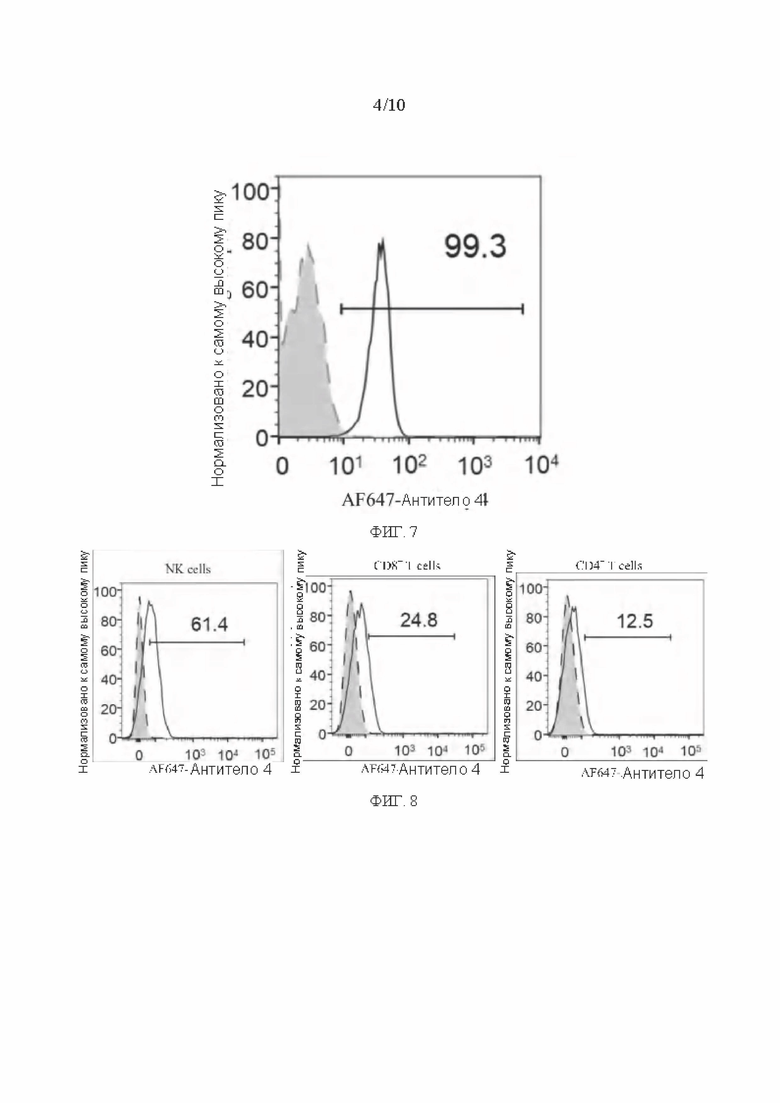

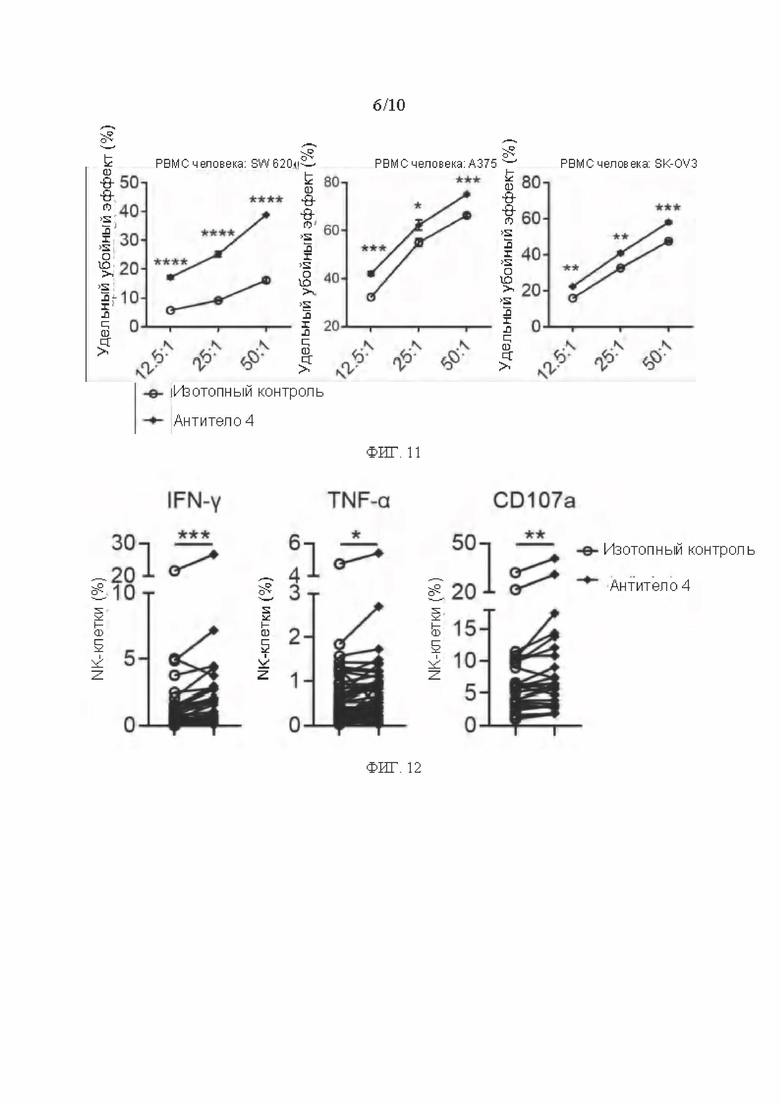
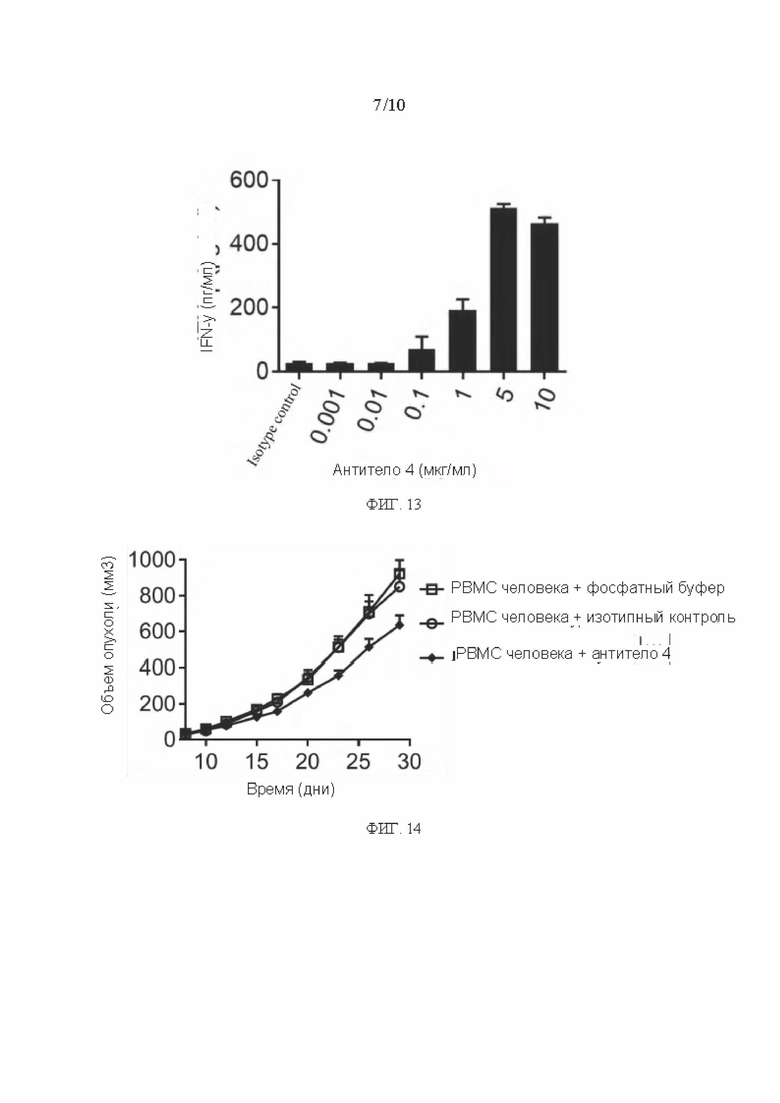
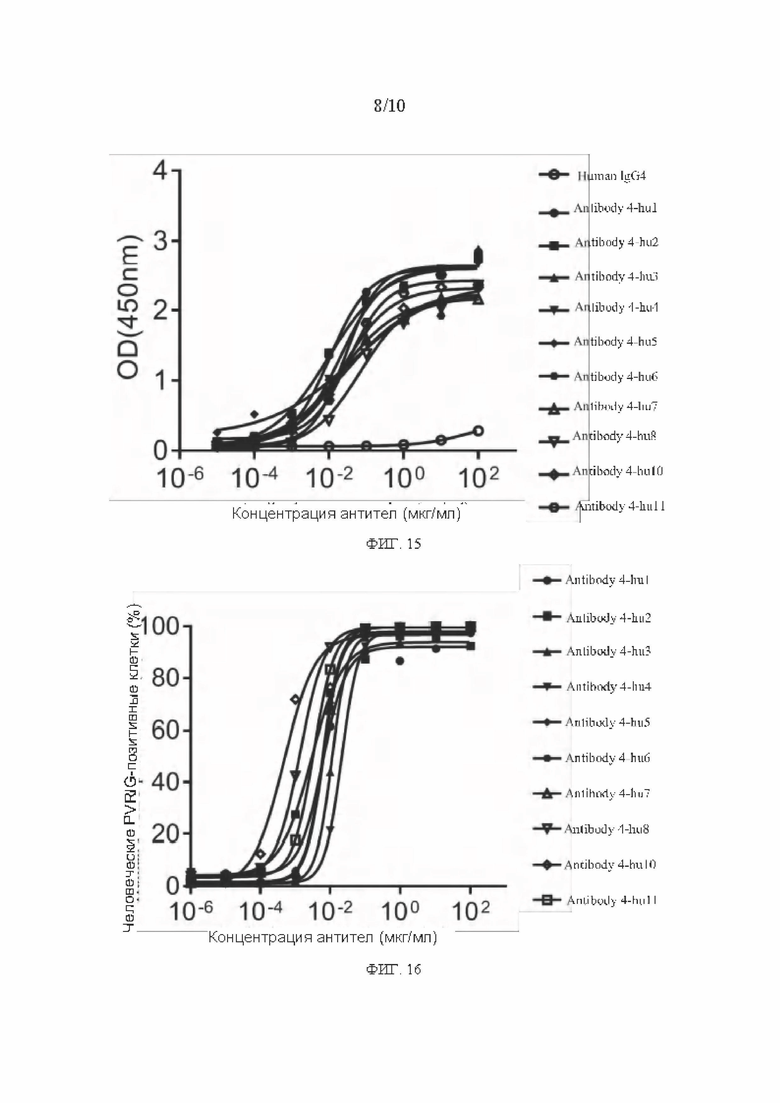
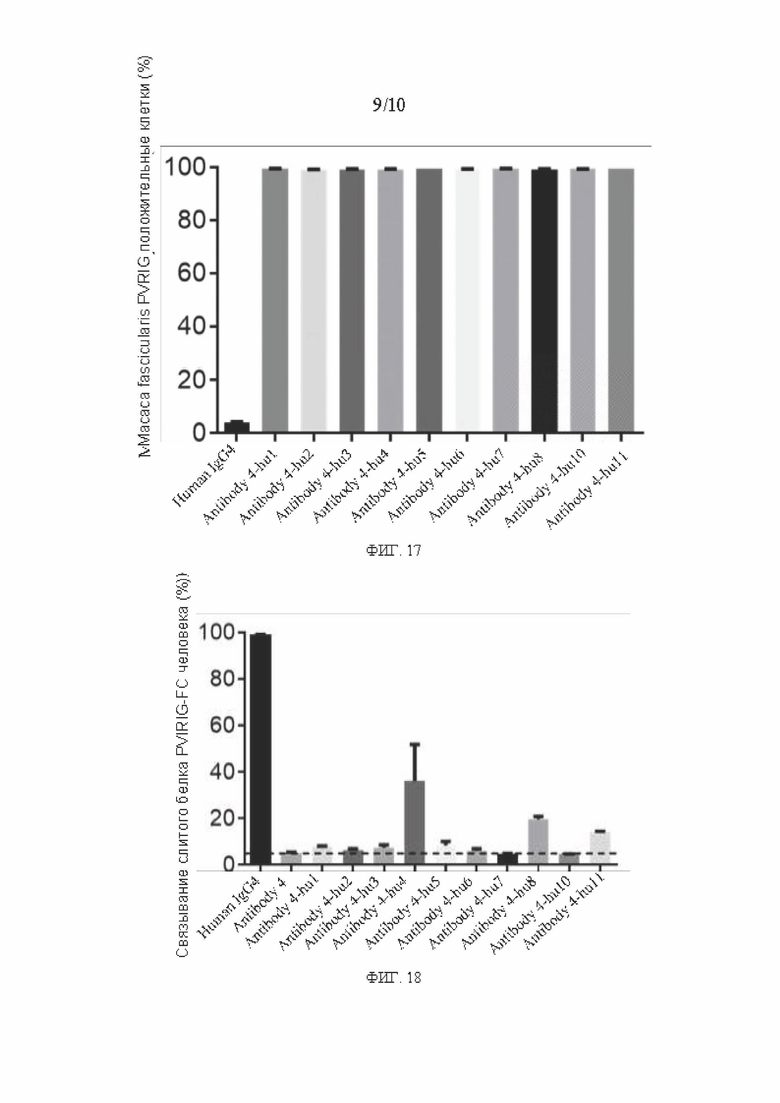
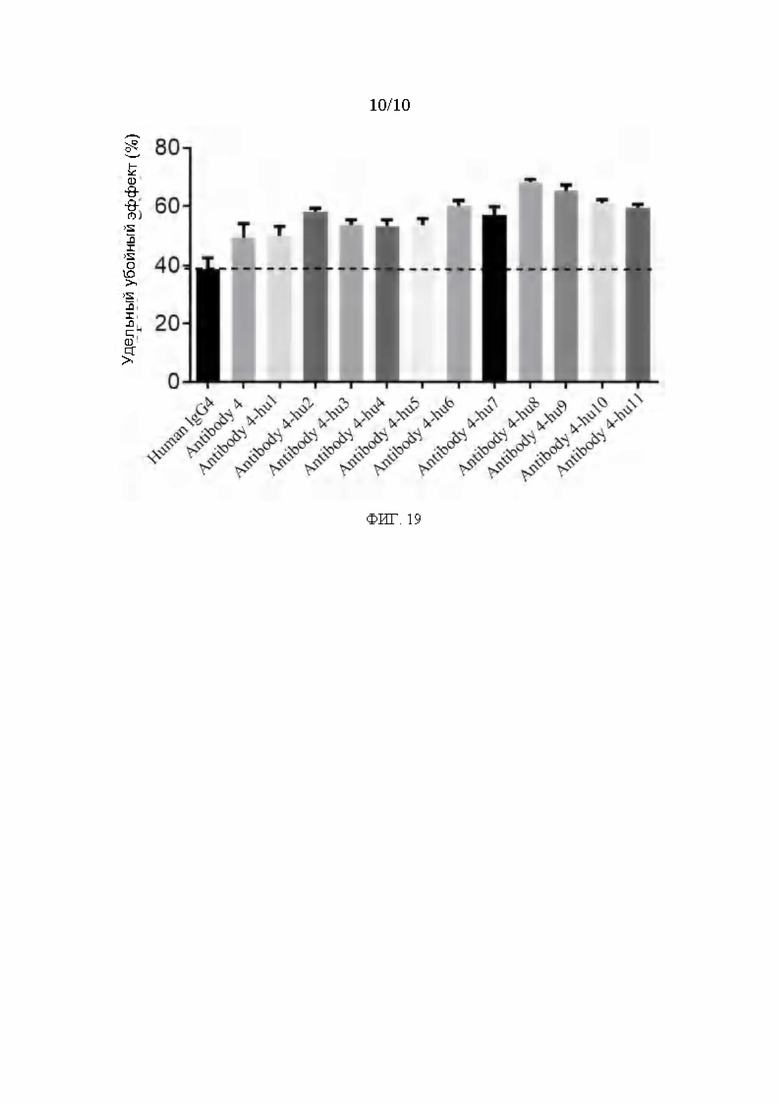
Изобретение относится к области биохимии, в частности к выделенному антителу или фрагменту антитела, которое(ый) специфически связывается с человеческим PVRIG. Также раскрыты нуклеиновая кислота, кодирующая указанное антитело; экспрессионный вектор, содержащий указанную нуклеиновую кислоту; клетка-хозяин, содержащая указанный вектор. Раскрыты способ ингибирования активности PVRIG протеина в образце in vitro с помощью указанного антитела. Изобретение обладает способностью эффективно ингибировать PVRIG. 6 н. и 1 з.п. ф-лы, 19 ил., 2 табл., 2 пр.
1. Антитело или фрагмент антитела, которое(ый) специфически связывается с человеческим PVRIG, выбранное(ый) из одного из следующих:
(1) антитело или фрагмент антитела, включающее(ий) комплементарные определяющие области тяжёлой цепи и комплементарные определяющие области легкой цепи, причём комплементарные определяющие области тяжёлой цепи включают GYTFSSFS, ILPGSNST и SSYWFAY, а комплементарные определяющие области легкой цепи включают QSLLNSGNQKNY, GAS и QNAHSYPPT;
(2) антитело или фрагмент антитела, включающее(ий) комплементарные определяющие области тяжёлой цепи и комплементарные определяющие области легкой цепи, комплементарные определяющие области тяжёлой цепи включают GYTFSTFA, ILPGINNT и STYWFAY, а комплементарные определяющие области лёгкой цепи включают QSLLNSGNQKNY, GAS и QNAHSYPPT;
(3) антитело или фрагмент антитела, включающее(ий) комплементарные определяющие области тяжёлой цепи и комплементарные определяющие области легкой цепи, комплементарные определяющие области тяжёлой цепи включают GYTFSTFA, ILPGGNNT и STYWFAY, а комплементарные определяющие области легкой цепи включают QSLLNSGNQKNY, GAS и QNAHSYPPA;
(4) антитело или фрагмент антитела, включающее(ий) комплементарные определяющие области тяжёлой цепи и комплементарные определяющие области лёгкой цепи, причём комплементарные определяющие области тяжёлой цепи включают GYSFTAYT, INPYNGGT и AREGNYYGSRGDFDY, а комплементарные определяющие области лёгкой цепи включают QTIVTN, YAS и QQSHSWPFT.
2. Антитело или фрагмент антитела, которое(ый) специфически связывается с человеческим PVRIG, выбранное(ый) по меньшей мере из одного из следующих:
(1) антитело или фрагмент антитела, включающее(ий) вариабельную область тяжёлой цепи и вариабельную область лёгкой цепи, причём вариабельная область тяжёлой цепи представляет собой последовательность, представленную в SEQ ID NO: 1, а вариабельная область лёгкой цепи представляет собой последовательность, представленную в SEQ ID NO: 2;
(2) антитело или фрагмент антитела, включающее(ий) вариабельную область тяжёлой цепи и вариабельную область лёгкой цепи, причём вариабельная область тяжёлой цепи выбрана из группы, состоящей из последовательностей, представленных в SEQ ID NO: 3, SEQ ID NO: 6, SEQ ID NO: 9, SEQ ID NO: 12, SEQ ID NO: 15, SEQ ID NO: 18, SEQ ID NO: 21, SEQ ID NO: 24, SEQ ID NO: 27, SEQ ID NO: 30 и SEQ ID NO: 33, и вариабельная область лёгкой цепи представляет собой последовательность, представленную в SEQ ID NO: 36;
(3) антитело или фрагмент антитела, включающее(ий) вариабельную область тяжёлой цепи и вариабельную область лёгкой цепи, причём вариабельная область тяжёлой цепи представляет собой последовательность, представленную в SEQ ID NO: 39, а вариабельная область лёгкой цепи представляет собой последовательность, представленную в SEQ ID NO: 40;
(4) антитело или фрагмент антитела, включающее(ий) вариабельную область тяжёлой цепи и вариабельную область лёгкой цепи, причём вариабельная область тяжёлой цепи представляет собой последовательность, представленную в SEQ ID NO: 41, а вариабельная область лёгкой цепи представляет собой последовательность, представленную в SEQ ID NO: 42;
(5) антитело или фрагмент антитела, включающее(ий) вариабельную область тяжёлой цепи и вариабельную область лёгкой цепи, причём вариабельная область тяжёлой цепи представляет собой последовательность, представленную в SEQ ID NO: 43, а вариабельная область лёгкой цепи представляет собой последовательность, представленную в SEQ ID NO: 44.
3. Антитело или фрагмент антитела по п. 2, включающее(ий) тяжёлую цепь и лёгкую цепь,
где аминокислотная последовательность тяжёлой цепи представляет собой последовательность, представленную в SEQ ID NO: 4, SEQ ID NO: 10, SEQ ID NO: 13, SEQ ID NO: 16, SEQ ID NO: 19, SEQ ID NO: 22, SEQ ID NO: 25, SEQ ID NO: 28, SEQ ID NO:31 или SEQ ID NO: 34; и
где аминокислотная последовательность лёгкой цепи представляет собой последовательность, представленную в SEQ ID NO: 37.
4. Изолируемая нуклеиновая кислота, кодирующая антитело или фрагмент антитела по любому из пп. 1-3.
5. Экспрессионный вектор, включающий нуклеиновую кислоту по п. 4.
6. Клетка-хозяин, включающая вектор экспрессии по п. 5, которая экспресирует антитело по п. 1.
7. Способ ингибирования активности PVRIG протеина в образце in vitro, включающий контакт образца с антителом или фрагментом антитела по любому из пп. 1-3.
| CN 107580500 A, 12.01.2018 | |||
| WO 2016134333 A1, 25.08.2016 | |||
| US 10550173 B2, 04.02.2020 | |||
| RU 2017132214 A, 19.03.2019. |

