Цитокины и иммунные клетки опосредуют специфические физиологические механизмы или пути, например, пути, ведущие к различным воспалительным нарушениям. Человеческий тимусный стромальный лимфопоэтин (TSLP) представляет собой ИЛ-7-подобный цитокин, который образуется человеческими эпителиальными клетками. Он стимулирует дифференцировку В-клеток и, кроме того, может костимулировать как тимоциты, так и зрелые Т-клетки. TSLP связывается со специфическим гетеродимерным рецептором на человеческих дендритных клетках CD11c+(DC's). Рецепторный гетеродимер состоит из общей цепи γ-подобного рецептора (рецептор TSLP; TSLPR) и ИЛ-7R-α-цепи. См., например, статьи Tonozuka et al., Cytogenet. Cell Genet. 93:23-25, 2001; Pandey et al., Nat. Immunol. 1:59-64, 2000; L.S.Park et al., J. Exp. Med. 192:659-670, 2000 и Reche et al., J. Immunol. 167:336-343, 2001. Связывание лиганда с рецептором индуцирует секрецию DC's ТН2-привлекающих цитокинов, TARC (тимусного и регулируемого активацией хемокина) и MDC (хемокина, выделенного из макрофагов). Кроме того, TSLP также индуцирует сильную активацию DC, экспансию непримированных (наивных) Т-клеток CD4+ и последующую поляризацию в отношении фенотипа TH2, продуцирующего проаллергические цитокины интерлейкин 4 (ИЛ-4), ИЛ-5, ИЛ-13 и фактор некроза опухоли α.
Обнаружено, что передача сигнала TSLP приводит в результате к активации фактора транскрипции Stat5. Более того, показано, что пациенты, как с острым, так и с хроническим атопическим дерматитом характеризуются сверхэкспрессией TSLP в кожных поражениях, позволяя предположить, что экспрессия TSLP ассоциирована с аллергическим воспалением in vivo. Помимо кожных кератиноцитов высокий уровень экспрессии TSLP обнаружен в эпителиальных клетках бронхов, гладких мышцах и легочных фибробластов, также поддерживая возможную роль TSLP в респираторных аллергических симптомах. Более того, механизмом, который мог бы участвовать в поддержании фенотипа ТН2, является то, что IgE-активированные мастоциты экспрессируют очень высокий уровень TSLP.
Приблизительно 20% населения западных стран страдает от воспалительных нарушений, например, аллергических заболеваний, которые включают астму, ринит, атопический дерматит и лекарственную аллергию. У 50%-80% пациентов с атопическим дерматитом имеется или развивается астма или аллергический ринит. До настоящего времени отсутствует лечение аллергии, вызываемой астмой, атопическим дерматитом и аллергическим ринитом. Современные препараты, такие как антагонисты β-2 адреноцептора для лечения астмы, элидел для лечения атопического дерматита и H1-антигистамин для лечения аллергического ринита, используют для получения направленности на симптомы. Таким образом, в области техники существует повышенная потребность в усовершенствованных лекарственных препаратах для лечения данных воспалительных нарушений, в частности аллергического воспаления. Настоящее изобретения направлено на данную и другие проблемы.
Раскрытие изобретения
Вариант осуществления изобретения в данном контексте предусматривает выделенное человеческое или гуманизированное антитело или его функциональный фрагмент с антигенсвязывающим участком, который специфичен в отношении белка рецептора тимусного стромального лимфопоэтина человека (hTSLPR)-мишени, и антитело или его функциональный фрагмент связывается с hTSLPR. В близком варианте осуществления связывание с hTSLPR определяют по меньшей мере по связыванию рецептора hTSLP клеточной поверхности, препятствующему высвобождение воспалительного медиатора.
В еще одном варианте осуществления изобретение предусматривает выделенный антигенсвязывающий участок антитела или его функциональный фрагмент. В ряде вариантов осуществления выделенный антигенсвязывающий участок включает участок H-CDR1, имеющий последовательность аминокислот TYGMS (SEQ ID NO: 7), и его консервативные варианты. Как описано в данном контексте, консервативные варианты включают остатки аминокислот в любой из идентифицированных последовательностей аминокислот. В близком варианте осуществления выделенный антигенсвязывающий участок представляет собой участок H-CDR2, имеющий последовательность аминокислот WINTYSGVPRYADDFKG (SEQ ID NO: 8), и его консервативные варианты. В другом близком варианте осуществления выделенный антигенсвязывающий участок представляет собой участок H-CDR3, имеющий последовательность аминокислот EGFITTVVGAAGRFVY (SEQ ID NO: 9), и его консервативные варианты.
В другом варианте осуществления выделенный антигенсвязывающий участок представляет собой участок L-CDR1, имеющий последовательность аминокислот KASQDVGTAVA (SEQ ID NO: 10), и его консервативные варианты. В еще одном близком варианте осуществления выделенный антигенсвязывающий участок представляет собой участок L-CDR2, имеющий последовательность аминокислот WASTRHT (SEQ ID NO: 11), и его консервативные варианты. В еще одном близком варианте осуществления выделенный антигенсвязывающий участок представляет собой участок L-CDR3, имеющий последовательность аминокислот QQYSTYPT (SEQ ID NO: 12), и его консервативные варианты.
В другом варианте осуществления выделенный антигенсвязывающий участок представляет собой тяжелую цепь, имеющую последовательность аминокислот вариабельной области SEQ ID NO: 5, и последовательность, имеющую идентичность последовательности на участках CDR по меньшей мере 60, 70, 80, 90 или 95 процентов относительно участка CDR SEQ ID NO: 5. В близком варианте осуществления выделенный антигенсвязывающий участок представляет собой легкую цепь, имеющую последовательность аминокислот вариабельной области SEQ ID NO: 6 и последовательность, имеющую идентичность последовательности на участках CDR по меньшей мере 60, 70, 80, 90 или 95 процентов относительно участка CDR SEQ ID NO: 6.
В другом аспекте настоящее изобретение представляет моноклональные антагонистические антитела к hTSLPR. Некоторые из антител к TSLPR, представленных в изобретении, обладают такой же специфичностью связывания, как у эталонного антитела, которое включает последовательность вариабельной области тяжелой цепи SEQ ID NO: 5 и последовательность вариабельной области легкой цепи SEQ ID NO: 6. Некоторые из данных антител являются полностью человеческими антителами, которые проявляют такую же специфичность связывания, как у эталонного антитела. Некоторые из антител имеют последовательность участка, определяющего комплементарность, тяжелой цепи (CDR) TYGMS (SEQ ID NO: 7), WINTYSGVPRYADDFKG (SEQ ID NO: 8) или EGFITTVVGAAGRFVY (SEQ ID NO: 9); или последовательность CDR легкой цепи KASQDVGTAVA (SEQ ID NO: 10), WASTRHT (SEQ ID NO: 11) или QQYSTYPT (SEQ ID NO: 12).
Некоторые антитела к hTSLPR имеют последовательности CDR1, CDR2 и CDR3 тяжелой цепи TYGMS (SEQ ID NO: 7), WINTYSGVPRYADDFKG (SEQ ID NO: 8) и EGFITTVVGAAGRFVY (SEQ ID NO: 9), соответственно, и последовательности CDR1, CDR2 и CDR3 легкой цепи KASQDVGTAVA (SEQ ID NO: 10). WASTRHT (SEQ ID NO: 11) и QQYSTYPT (SEQ ID NO: 12), соответственно. Некоторые другие антитела, представленные в изобретении, включают последовательность вариабельной области тяжелой цепи аминокислот, которая по меньшей мере на 85% идентична SEQ ID NO: 5, и последовательность вариабельной области легкой цепи аминокислот, которая по меньшей мере на 85% идентична SEQ ID NO: 6. Некоторые другие антитела к hTSLPR, представленные в изобретении, имеют последовательность вариабельной области тяжелой цепи аминокислот, которая идентична SEQ ID NO: 5, и последовательность вариабельной области легкой цепи аминокислот, которая идентична SEQ ID NO: 6.
Некоторые антитела к hTSLPR, представленные в изобретении, представляют собой мышиные антитела. Ряд других представляют собой химерные антитела. Некоторые из химерных антител имеют константную область тяжелой цепи человека и константную область легкой цепи человека. Некоторые другие антитела к hTSLPR, представленные в изобретении, являются гуманизированными антителами. Некоторые другие антитела к hTSLPR, представленные в изобретении, являются полностью человеческими антителами, которые проявляют такую же специфичность связывания, как антитело, которое включает последовательность вариабельной области тяжелой цепи SEQ ID NO: 5 и последовательность вариабельной области легкой цепи SEQ ID NO: 6. В изобретении также представлены одноцепочечные антитела, например, фрагмент Fab. Некоторые из антител к hTSLPR относятся к изотипу IgG1. Некоторые другие антитела относятся к изотипу IgG4.
В другом аспекте изобретение представляет выделенные или рекомбинантные полинуклеотиды (например, ДНК), которые кодируют полипептид, включающий вариабельную область тяжелой цепи или вариабельную область легкой цепи антитела к hTSLPR, представленного в изобретении. Например, полинуклеотиды могут кодировать тяжелую цепь антитела, которая включает последовательности CDR1, CDR2 и CDR3 тяжелой цепи TYGMS (SEQ ID NO: 7), WINTYSGVPRYADDFKG (SEQ ID NO: 8) и EGFITTVVGAAGRFVY (SEQ ID NO: 9), соответственно. Полинуклеотиды могут также кодировать легкую цепь антитела, которая включает последовательности CDR1, CDR2 и CDR3, KASQDVGTAVA (SEQ ID NO: 10), WASTRHT (SEQ ID NO: 11) и QQYSTYPT (SEQ ID NO: 12), соответственно. Некоторые полинуклеотиды, представленные в изобретении, кодируют зрелую последовательность вариабельной области тяжелой цепи, которая по меньшей мере на 90% идентична зрелой области SEQ ID NO: 5. Некоторые другие полинуклеотиды кодируют зрелую последовательность вариабельной области легкой цепи, которая по меньшей мере на 90% идентична зрелой области SEQ ID NO: 6. Некоторые из данных полинуклеотидов кодируют зрелую последовательность вариабельной области тяжелой цепи, которая идентична зрелой области SEQ ID NO: 5 или зрелой последовательности вариабельной области легкой цепи, которая идентична зрелой области SEQ ID NO: 6.
В другом аспекте изобретение представляет выделенные клетки-хозяева, которые несут (1) сегмент рекомбинантной ДНК, кодирующий тяжелую цепь антитела к hTSLPR, соответствующего изобретению, и (2) второй сегмент рекомбинантной ДНК, кодирующий легкую цепь антитела. В некоторых клетках-хозяевах сегменты рекомбинантной ДНК соответственно функционально связаны с первым и вторым промотором и способны экспрессироваться в клетках-хозяевах. Некоторые из данных клеток-хозяев экспрессируют моноклональное антитело, которое имеет последовательности CDR1, CDR2 и CDR3 тяжелой цепи TYGMS (SEQ ID NO: 7), WINTYSGVPRYADDFKG (SEQ ID NO: 8) и EGFITTVVGAAGRFVY (SEQ ID NO: 9), соответственно, и последовательности CDR1, CDR2 и CDR3 легкой цепи KASQDVGTAVA (SEQ ID NO: 10), WASTRHT (SEQ ID NO: 11) и QQYSTYPT (SEQ ID NO: 12), соответственно. Некоторые другие клетки-хозяева экспрессируют антитело к hTSLPR, которое включает зрелую последовательность вариабельной области тяжелой цепи, которая по меньшей мере на 90% идентична зрелому участку SEQ ID NO: 5, и зрелую последовательность вариабельной области легкой цепи, которая по меньшей мере на 90% идентична зрелому участку SEQ ID NO: 6. Некоторые из данных клеток-хозяев экспрессируют антитело к hTSLPR, которое включает зрелую последовательность вариабельной области тяжелой цепи, которая идентична зрелому участку SEQ ID NO: 5, и зрелую последовательность вариабельной области легкой цепи, которая идентична зрелому участку SEQ ID NO: 6. Некоторые из клеток-хозяев представляют собой клетки млекопитающего, отличные от человеческих.
В другом аспекте изобретение представляет способы лечения воспалительного нарушения у субъекта, например, больного человека. Данные способы предусматривают введение пациенту фармацевтической композиции, которая включает эффективное количество антитела к hTSLPR. Как правило, антитело к hTSLPR имеет такую же специфичность связывания, как у антитела к hTSLPR, которое включает последовательность вариабельной области тяжелой цепи SEQ ID NO: 5 и последовательность вариабельной области легкой цепи SEQ ID NO: 6. В ряде способов лечения используют полностью человеческое антитело. В ряде способов, антитело к hTSLPR несет последовательности тяжелой цепи CDR1, CDR2 и CDR3 TYGMS (SEQ ID NO: 7), WINTYSGVPRYADDFKG (SEQ ID NO: 8) и EGFITTVVGAAGRFVY (SEQ ID NO: 9), соответственно, и последовательности легкой цепи CDR1, CDR2 и CDR3 KASQDVGTAVA (SEQ ID NO: 10), WASTRHT (SEQ ID NO: 11) и QQYSTYPT (SEQ ID NO: 12), соответственно. В ряде способов используемое антитело к hTSLPR включает зрелую последовательность вариабельной области тяжелой цепи, которая идентична зрелому участку SEQ ID NO: 5, и зрелую последовательность вариабельной области легкой цепи, которая идентична зрелому участку SEQ ID NO: 6. Некоторые способы направлены на лечение пациентов, которые страдают от аллергического воспалительного заболевания. Примеры аллергических воспалительных заболеваний, которые поддаются лечению, включают атопический дерматит, астму или аллергический ринит.
В еще одном варианте осуществления изобретение представляет иммуноконъюгат, состоящий из первого компонента, который представляет собой антитело или его фрагмент, и второй компонент, имеющий вторую последовательность аминокислот. Например, иммуноконъюгат представляет собой цитотоксин, или иммуноконъюгат представляет собой связывающий белок или антитело, имеющие специфичность связывания с мишенью, которая отлична от hTSLPR.
В другом варианте осуществления изобретение представляет набор, включающий антитело или фрагмент антитела. В некоторых вариантах осуществления набор далее включает фармацевтически приемлемый носитель или, соответственно, наполнитель. В других близких вариантах осуществления антитело в наборе находится в унифицированной дозе. В еще одном близком варианте осуществления набор включает инструкции по применению при введении пациенту.
Дальнейшее понимание природы и преимуществ настоящего изобретения можно получить посредством ссылки на остальные разделы описания и формулу изобретения.
Краткое описание чертежей
На фиг.1 представлен скрининг антагонистических антител к TSLPR с использованием анализа hTSLP-зависимой пролиферации клеток в клетках BaF3/hTSLPR/hIL7Rα.
На фиг.2А-2С представлена очистка мышиных и химерных моноклональных антител к hTSLPR. А: химерное антитело IgGI; В: химерное антитело IgG4 и С: мышиное антитело IgGI.
На фиг.3А-3С показана антагонистическая активность очищенного мышиного антитела к hTSLPR (клон 1D6.C9) с помощью анализа пролиферации клеток и анализа сигнала люциферазы. А: пролиферация клеток Ba/F3-hTSLPR-hIL7Rα; В: пролиферация клеток BaF3/hTSLPR/hIL7Rα/Stat5-Luc и С: активность люциферазы клеток BaF3/hTSLPR/hIL7Rα/Stat5-Luc.
На фиг.4 представлены нуклеотидные последовательности вариабельных областей клона мышиного моноклонального антитела к hTSLPR 1D6.C9.
На фиг.5 представлены последовательности аминокислот вариабельной области клона мышиного антитело к hTSLPR 1D6.C9. Участки, определяющие комплементарность (CDRs), и скелетные области (FRs) показаны подчеркнутыми остатками или остатками, выделенными курсивом.
На фиг.6 показаны результаты анализов сигнала люциферазы, сравнивающие антагонистическую активность очищенного мышиного и химерного антител к hTSLPR в клетках Ba/F3, проявляющих сверхэкспрессию hTSLPR, hIL7Rα и Stat5-Luc.
На фиг.7 показано ингибирование TSLP-опосредованной секреции TARC из человеческих моноцитов посредством мышиного и химерного антител к hTSLPR.
На фиг.8 показана идентификация антитело-связывающего домена - антитело TSLPR связывается с прерывистым эпитопом.
Детальное описание изобретение
Настоящее изобретение основано отчасти на разработке авторами антагонистических антител к человеческому TSLPR. Антитела к hTSLPR, генерированные в мыши, или химерные антитела к hTSLPR, созданные in vitro, как обнаружено, способны ингибировать активности, опосредованные передачей сигнала TSLP, например, TSLP-опосредованная пролиферация клеток. Таким образом данные антитела используют в качестве терапевтических или профилактических агентов против ряда заболеваний или нарушений, опосредованных или ассоциированных с активностями передачи сигнала TSLP, например, аллергических воспалительных заболеваний, таких атопический дерматит и астма. Следующие разделы представляют руководство по получению и использованию композиций, представленных в изобретении, и осуществлению способов, представленных в изобретении.
I. Определения
Пока не определено иначе, все технические и научные термины, используемые в данном контексте, имеют такое же значение, какое, как правило, имеет в виду обычный специалист в области техники, к которой относится данное изобретение. Следующие ссылки представляют специалисту общее определение многих терминов, используемых в данном изобретении: Oxford Dictionary of Biochemistry and Molecular Biology под ред. Smith et al., Oxford University Press (исправленное издание, 2000); Dictionary of Microbiology and Molecular Biology под ред. Singleton, John Wiley & Sons (3 испр. изд., 2002) и А Dictionary of Biology (Oxford Paperback Reference) под ред. Martin and Hine, Oxford University Press (4 испр. изд., 2000). Кроме того, следующие определения представлены, чтобы помочь читателю в практической реализации изобретения.
Для того чтобы настоящее изобретение было легче понять, для ряда терминов сначала даны определения. Дополнительные определения приводят на протяжении детального описания.
Термин "иммунный ответ" относится к действию, например, лимфоцитов, антигенпрезентирующих клеток, фагоцитарных клеток, гранулоцитов и растворимых макромолекул, продуцируемых вышеуказанными клетками или печенью (включая антитела, цитокины и комплемент), которое приводит в результате к избирательному повреждению, разрушению или удалению из организма человека поражающих патогенов, клеток или тканей, инфицированных патогенами, раковых клетки или, в случаях аутоиммунитета или патологического воспаления, нормальных человеческих клеток или тканей.
Термин "путь сигнальной трансдукции" относится к биохимическим взаимоотношениям между рядом молекул сигнальной трансдукции, которые играют роль в передаче сигнала от одной части клетки к другой части клетки.
Термин "антитело" относительно данного контекста включает целые антитела и любые его антигенсвязывающий фрагмент (т.е. "антигенсвязывающую часть") или одиночные цепи. Природное "антитело" представляет собой гликопротеин, включающий по меньшей мере две тяжелых (Н) цепи и две легких (L) цепи, связанные между собой дисульфидными связями. Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (в данном контексте сокращенно VH) и константной области тяжелой цепи. Константная область тяжелой цепи состоит из трех доменов, СН1, СН2 и СН3. Каждая легкая цепь состоит из вариабельной области легкой цепи (в данном контексте сокращенно VL) и константной области легкой цепи. Константная область легкой цепи состоит из одного домена CL. Области VH и VL можно далее подразделить на гипервариабельные участки, называемые участками, определяющими комплементарность (CDR), расположенные между участками, которые являются более консервативными, называемыми скелетными участками (FR). Каждый VH и VL состоит из трех CDR и четырех FR, расположенных от аминоконца к карбоксиконцу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области тяжелой и легкой цепей включают связывающий домен, который взаимодействует с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (Clq) классической системы комплемента.
Термин "антигенсвязывающая часть" антитела (или просто "антигенная часть"), как используют в данном контексте, относится к антителу полной длины или одному или более фрагментов антитела, которые сохраняют способность специфически связываться с антигеном (например, TSLPR). Показано, что антигенсвязывающая функция антитела может быть осуществлена фрагментами антитела полной длины. Примеры связывающих фрагментов, охватываемых термином "антигенсвязывающая часть" антитела, включают фрагмент Fab, одновалентный фрагмент, состоящий из доменов VL, VH, CL и СН1; фрагмент F(ab)2, двухвалентный фрагмент, включающий два фрагмента Fab, связанных дисульфидным мостиком в шарнирной области; фрагмент, состоящий из доменов VH и СН1; фрагмент Fv, состоящий из доменов VL и VH одного плеча антитела; фрагмент dAb (см. статью Ward et al., 1989 Nature 341:544-546), который состоит из домена VH, и выделенный участок, определяющий комплементарность (CDR).
Кроме того, хотя два домена фрагмента Fv, VL и VH, кодируются отдельными генами, они могут быть связаны при использовании рекомбинантных методов синтетическим линкером, который дает возможность получить их в виде одной белковой цепи, в которой участки VL и VH спарены с образованием одновалентных молекул (известных как одноцепочечный Fv (scFv); см., например, статьи Bird et al., 1988 Science 242:423-426 и Huston et al., 1988 Proc. Natl. Acad. Sci. 85:5879-5883). Предусматривают также, что данные одноцепочечные антитела охватываются термином "антигенсвязывающая часть" антитела. Данные фрагменты антитела получают, используя принятые методы, известные компетентным специалистам в области техники, и проводят скрининг фрагментов на применение таким же образом, как интактных антител.
Термин "выделенное антитело", как используют в данном контексте, относится к антителу, которое в существенной степени очищено от других антител, имеющих другие антигенные специфичности (например, выделенное антитело, которое специфически связывает TSLPR, в существенной степени очищено от антител, которые специфически связывают антигены, отличные от TSLPR). Однако выделенное антитело, которое специфически связывает TSLPR, может обладать перекрестной реактивностью с другими антигенами, такими как молекулы TSLPR других видов. Более того, выделенное антитело может быть в существенной степени очищено от другого клеточного материала и/или химических веществ.
Термины "моноклональное антитело" или "композиция моноклональных антител", как используют в данном контексте, относятся к препарату молекул антитела одной молекулярной композиции. Композиция моноклонального антитела проявляет одну специфичность связывания и аффинность в отношении определенного эпитопа.
Термин "человеческое антитело", как используют в данном контексте, предусматривает включение антител, имеющих вариабельные области, в которых как скелетные участки, так и участки CDR выделены из последовательностей человеческий природы. Более того, если антитело включает константную область, то константная область также выделена из данных человеческих последовательностей, например, последовательностей зародышевой линии человека. Человеческие антитело, представленные в изобретении, могут включать остатки аминокислот, не кодируемые человеческими последовательностями (например, мутации, введенные путем неспецифического или сайт-специфического мутагенеза in vitro или путем соматической мутации in vivo).
Термин "человеческое моноклональное антитело" относится к антителам, проявляющим одну специфичность связывания, которые имеют вариабельные области, в которых как скелетные участки, так и участки CDR выделены из человеческих последовательностей. В одном варианте осуществления человеческие моноклональные антитела получают с помощью гибридомы, которая включает В-клетку, полученную от трансгенного животного, отличного от человека, например, трансгенной мыши, имеющей геном, включающий трансген человеческой тяжелой цепи и трансген легкой цепи, слитый с иммортализованной клеткой.
Термин "рекомбинантное человеческое антитело", как используют в данном контексте, включает все человеческие антитела, которые получают, экспрессируют, создают или выделяют рекомбинантными методами, такие как антитела, выделенные у животного (например, мыши), которое является трансгенным или трансхромосомным в отношении генов иммуноглобулина человека, или полученной из него гибридомы, антитела, выделенные из клетки-хозяина, трансформированной с целью получения экспрессии человеческого антитела, например, из трансфектомы, антитела, выделенные из библиотеки рекомбинантных комбинаторных человеческих антител, и антитела, полученные экспрессированные, созданные или выделенные любыми другими средствами, которые включают сплайсинг всей или части последовательностей гена иммуноглобулина человека с другими последовательностями ДНК. Данные рекомбинантные человеческие антитела имеют вариабельные области, в которых скелетные участки и участки CDR выделены из последовательностей иммуноглобулина зародышевой линии человека. Однако в ряде вариантов осуществления данные рекомбинантные человеческие антитела могут быть подвергнуты мутагенезу in vitro (или, когда используют животное, трансгенное по последовательностям Ig человека, соматическому мутагенезу in vivo) и, таким образом, последовательности аминокислот областей VH и VL рекомбинантных антител представляют собой последовательности, которые, хотя и выделены из и относятся к последовательностям VH и VL зародышевой линии человека, могут не существовать в естественных условиях в репертуаре зародышевой линии антител человека in vivo.
"Химерное антитело" представляет собой молекулу антитела, в которой (а) константная область или ее часть изменена, замещена или заменена так, что антигенсвязывающий центр (вариабельная область) связан с константной областью другого или измененного класса, эффекторной функции и/или вида, либо полностью другой молекулой, которая придает химерному антителу новые свойства, например, фермента, токсина, гормона, фактора роста, лекарства и т.п.; или (b) вариабельная область или ее часть изменена, замещена или заменена вариабельной областью, имеющей другую или измененную антигенную специфичность. Например, как показано в примерах ниже, мышиное антитело к hTSLPR можно модифицировать замещением его константной области константной областью из человеческого иммуноглобулина. Вследствие замещения человеческой константной областью химерное антитело может сохранять свою специфичность в плане распознавания человеческого TSLPR, имея при этом пониженную антигенность у человека по сравнению с исходным мышиным антителом.
"Гуманизированное" антитело представляет собой антитело, которое сохраняет реакционность нечеловеческого антитела, являясь при этом менее иммуногенным для человека. Это может достигаться, например, путем сохранения нечеловеческих участков CDR и замещения остальных частей антитела их человеческими аналогами (т.е. константной области, а также скелетных частей вариабельной области). См., например, статьи Morrison et al., Proc. Natl. Acad. Sci. USA, 81:6851-6855, 1984; Morrison and Oi, Adv. ImmunoL, 44:65-92, 1988; Verhoeyen et al., Science, 239:1534-1536, 1988; Padlan, Molec. Immun., 28:489-498, 1991; и Padlan, Molec. Immun., 31:169-217, 1994. Другие примеры технологии человеческой инженерии включают, но без ограничения перечисленным, метод Хота, раскрытый в US 5766886.
Термин "гуманиринг", как используют в данном контексте, относится к способу превращения нечеловеческих антител в инженерные человеческие антитела (См., например, метод KaloBios' Humaneering™).
Как используют в данном контексте, термин "изотип" относится к классу антител (например, IgM, IgE, IgG, такому как IgGI или IgG4), который дают гены константной области тяжелой цепи.
Фразы "антитело, распознающее антиген", и "антитело, специфическое в отношении антигена", используют в данном контексте взаимозаменяемо с термином "антитело, которое связывается специфически с антигеном".
Как используют в данном контексте, антитело, которое "специфически связывается с человеческим TSLPR", относится к антителу, которое связывается с человеческим TSLPR с KD 200×10-12 М или меньше, 150×10-12 М или меньше либо 100×10-12 М или меньше.
Термин "специфичность связывания", как используют в данном контексте, относится к способности паратопа отдельного антитела реагировать только с одной антигенной детерминантой. Паратоп антитела находится в части Fab молекулы и сконструирован из гипервариабельных областей тяжелой и легкой цепей. Аффинность связывания антитела представляет собой силу реакции между одной антигенной детерминантой и одним паратопом на антителе. Она представляет собой сумму сил притяжения и отталкивания, действующих между антигенной детерминантной и паратопом антитела. Аффинность является константой равновесия, которая описывает реакцию антиген-антитело.
Специфическое связывание между двумя элементами означает связывание с константой равновесия (KA) по меньшей мере 1×107 М-1, 108 М-1, 109 М-1 или 1010 М-1. Выражение "специфически (или избирательно) связывается" с антителом (например, антителом к hTSLPR) относится к реакции связывания, которая является определяющей в отношении присутствия родственного антигена (например, человеческий полипептид TSLPR) в гетерогенной популяции белков и других биологических компонентов. В дополнение к вышеупомянутой константе равновесия (KA) антитело к hTSLPR, представленное в изобретении, как правило, имеет константу диссоциации (Kd) приблизительно 1×10-2 сек.-1, 1×10-3 сек.-1, 1×10-4 сек.-1 или ниже и связывается с человеческим TSLPR с аффинностью, которая по меньшей мере в два раза превышает его аффинность связывания с неспецифическим антигеном (например, BSA). Выражения "антитело, распознающее антиген", и "антитело, специфическое в отношении антигена", используют в данном контексте взаимозаменяемо с термином "антитело, которое специфически связывается с антигеном".
Термин "эпитоп" означает белковую детерминанту, способную к специфическому связыванию с антителом. Эпитопы обычно состоят из химически активных поверхностных групп молекул, таких как аминокислотные или сахарные боковые цепи, и, как правило, имеют специфические трехмерные структурные характеристики, а также специфические характеристики заряда. Конформационные и неконформационные эпитопы различаются тем, что связывание с первыми, но не с последними утрачивается в присутствии денатурирующих растворителей.
Термин "нуклеиновая кислота" используют в данном контексте взаимозаменяемо с термином "полинуклеотид" и относится к дезоксирибонуклеотидам или рибонуклеотидам и их полимерам либо в одно-, либо в двухцепочечной форме. Термин охватывает нуклеиновые кислоты, содержащие известные аналоги нуклеотидов или модифицированные скелетные остатки или связи, которые являются синтетическими, природными и неприродными, которые имеют свойства связывания, близкие свойствам эталонной нуклеиновой кислоты, и которые метаболизируются аналогично эталонным нуклеотидам. Примеры данных аналогов включают, но без ограничения перечисленным, фосфоротиоаты, фосфорамидаты, метилфосфонаты, хиральные метилфосфонаты, 2-O-метилрибонуклеотиды, пептид-нуклеиновые кислоты (ПНК).
Пока не указано иначе, термин определенная последовательность нуклеиновой кислоты также подразумеваемым образом охватывает ее консервативно модифицированные варианты (например, замещения вырожденных кодонов) и комплементарные последовательности, а также ясно указываемую последовательность. В частности, как детально описано ниже, замещения вырожденных кодонов могут достигаться посредством генерации последовательностей, в которых третье положение одного или более выбранных (или всех) кодонов замещают смешанным основанием и/или дезоксиинозиновыми остатками (см. статьи Batzer et al,. Nucleic Acid Res. 19:5081, 1991; Ohtsuka et al., J. Biol. Chem. 260:2605-2608, 1985 и Rossolini et al., Mol. Cell. Probes 8:91-98, 1994).
Термин "аминокислота" относится к природным и синтетическим аминокислотам, а также аналогам аминокислот и миметикам аминокислот, которые действуют подобно природным аминокислотам. Природными аминокислотами являются аминокислоты, кодируемые генетическим кодом, а также те аминокислоты, которые модифицируют позднее, например, гидроксипролин, γ-карбоксиглутамат и O-фосфосерин. Аналоги аминокислот относятся к соединениям, которые имеют такую основную химическую структуру, как природная аминокислота, т.е., α-атом углерода, который связан с водородом, карбоксильной группой, аминогруппой и R-группой, например, гомосерин, норлейцин, метионинсульфоксид, метионинметилсульфоний. Данные аналоги имеют модифицированные R-группы (например, норлейцин) или модифицированные пептидные скелеты, но сохраняют такую же основную химическую структуру, как природная аминокислота. Миметики аминокислот относятся к химическим соединениям, которые имеют структуру, отличную от общей химической структуры аминокислоты, но действуют подобно природной аминокислоте.
Термины "полипептид" и "белок" используют в данном контексте взаимозаменяемо в отношении к полимеру из остатков аминокислот. Термины применяют для полимеров аминокислот, в которых один или более остатков аминокислот является искусственным химическим миметиком соответствующей природной аминокислоты, а также для полимеров природных аминокислот и полимера неприродных аминокислот. Пока не указано иначе, термин определенная полипептидная последовательность также подразумеваемым образом охватывает ее консервативно модифицированные варианты.
Термин "консервативно модифицированный вариант" относится к последовательностям, как аминокислот, так и нуклеиновых кислот. Касательно определенных последовательностей нуклеиновых кислот, консервативно модифицированные варианты относятся к тем нуклеиновым кислотам, которые кодируют идентичные или в основном идентичные последовательности аминокислот, или те, в которых нуклеиновая кислота не кодирует последовательность аминокислот в основном идентичных последовательностей. Вследствие вырожденности генетического кода большое число функционально идентичных нуклеиновых кислот кодирует любой заданный белок. Например, все кодоны GCA, GCC, GCG и GCU кодируют аминокислоту аланин. Таким образом в каждом положении, где кодоном определен аланин, кодон может быть изменен на любой из описанных соответствующих кодонов без изменения кодируемого полипептида. Данные варианты нуклеиновой кислоты представляют собой "молчащие варианты", которые представляют собой один из видов консервативно модифицированных вариантов. В данном контексте каждая последовательность нуклеиновой кислоты, которая кодирует полипептид, описывает также каждый возможный вариант нуклеиновой кислоты. Компетентный специалист будет иметь в виду, что каждый кодон в нуклеиновой кислоте (за исключением AUG, который обычно является единственным кодоном для метионина, и TGG, который обычно является единственным кодоном для триптофана), может быть модифицирован с получением функционально идентичной молекулы. Соответственно, каждый молчащий вариант нуклеиновой кислоты, который кодирует полипептид, подразумевается в каждой из описанных последовательностей.
Для полипептидных последовательностей "консервативно модифицированные варианты" включают индивидуальные замены, делеции или вставки в полипептидную последовательность, которые приводят к замене аминокислоты химически близкой аминокислотой. Таблицы консервативных замен, представляющие функционально близкие аминокислоты, хорошо известны в области техники. Данные консервативно модифицированные варианты дополняют и не исключают полиморфные варианты, межвидовые гомологи и аллели, представленные в изобретении. Следующие восемь групп включают аминокислоты, которые представляют собой консервативные замены друг для друга:
1) Аланин (А), Глицин (G);
2) Аспарагиновая кислота (D), Глутаминовая кислота (Е);
3) Аспарагин (N), Глутамин (Q);
4) Аргинин (R), Лизин (K);
5) Изолейцин (I), Лейцин (L), Метионин (М), Валин (V);
6) Фенилаланин (F), Тирозин (Y), Триптофан (W);
7) Серин (S), Треонин (Т) и
8) Цистеин (С), Метионин (М) (см., например, монографию Creighton, Proteins (1984)).
Термины "идентичный" или процент "идентичности" в контексте двух или более последовательностей нуклеиновых кислот или полипептидов относятся к двум или более последовательностей или субпоследовательностей, которые являются одинаковыми. Две последовательности являются "в существенной мере идентичными", если две последовательности имеют определенный процент остатков аминокислот или нуклеотидов, которые являются одинаковыми (т.е., идентичность 60%, необязательно идентичность 65%, 70%, 75%, 80%, 85%, 90%, 95% или 99% относительно заданного участка или, если не определено, относительно целой последовательности) при сравнении и выравнивании для максимального соответствия относительно окна сравнения либо сконструированного участка, как измеряют при использовании одного из следующих алгоритмов сравнения последовательностей или посредством выравнивания вручную и визуального изучения. Необязательно идентичность существует относительно участка, который составляет по меньшей мере приблизительно 50 нуклеотидов (или 10 аминокислот) в длину или, более предпочтительно, относительно участка, который составляет 100-500 или 1000 или более нуклеотидов (либо 20, 50, 200 или более аминокислот) в длину.
При сравнении последовательностей, как правило, одна последовательность действует как эталонная последовательность, с которой сравнивают тест-последовательности. При использовании алгоритма сравнения последовательностей тестируемую и эталонную последовательности вводят в компьютер, при необходимости создают координаты субпоследовательности и задают параметры программы алгоритма последовательности. Можно использовать параметры программы по умолчанию или можно создать альтернативные параметры. Затем алгоритм сравнения последовательностей рассчитывает процент идентичности последовательностей для тест-последовательностей относительно эталонной последовательности, основываясь на параметрах программы.
"Окно сравнения", как используют в данном контексте, включает ссылку на сегмент из любого числа прилегающих друг к другу положений, выбранных из группы, состоящей из 20-600, как правило, от приблизительно 50 до приблизительно 200, чаще от приблизительно 100 до приблизительно 150, в которых последовательность можно сравнить с эталонной последовательностью из того же самого числа прилежащих друг к другу положений после оптимального выравнивания двух последовательностей. Способы выравнивания (элайнмента) последовательностей для сравнения хорошо известны в области техники. Оптимальное выравнивание последовательностей для сравнения можно провести, например, с помощью алгоритма локальной гомологии, см. статью Smith and Waterman (1970) Adv. Appl. Math. 2:482c, с помощью алгоритма выравнивания гомологии, см. статью Needleman and Wunsch, J. Mol. Biol. 48:443, 1970, посредством поиска для метода аналогии, см. статью Pearson and Lipman, Proc. Nat'1. Acad. Sci. USA 85:2444, 1988, с помощью компьютеризированных реализации данных алгоритмов (GAP, BESTFIT, FASTA и TFASTA в пакете программ Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Dr., Madison, WI) или посредством выравнивания вручную и визуального исследования (см., например, раздел Brent et al. в монографии Current Protocols in Molecular Biology, John Wiley & Sons, Inc. (под ред. Ringbou, 2003)).
Двумя примерами алгоритмов, которые подходят для определения процента идентичности последовательности и аналогии последовательности, являются алгоритмы BLAST и BLAST 2.0, которые описаны в статьях Altschul et al., Nuc. Acids Res. 25:3389-3402, 1977 и Altschul et al., J. Mol. Biol. 215:403-410, 1990, соответственно. Пакет программ для проведения анализов BLAST общедоступен через Национальный центр информации по биотехнологии. Данный алгоритм включает, во-первых, идентификацию пар последовательностей с высоким результатом (HSPs) посредством идентификации коротких слов длины W в запрашиваемой последовательности, которые либо соответствуют, либо удовлетворяют некоторому положительно оцениваемому пороговому значению Т при выравнивании со словом той же длины в последовательности базы данных. Т определяют как пороговое значение соседнего слова (см. Altschul et al., выше). Данные исходные наиболее распространенные соседние слова действуют как затравки для начала поисков с целью обнаружения более длинных содержащих их HSPs. Наиболее распространенные слова удлиняют в обоих направлениях вдоль каждой последовательности, пока кумулятивный результат выравнивания может увеличиваться. Кумулятивные результаты рассчитывают, используя для нуклеотидных последовательностей параметры М (премиальный результат для пары соответствующих остатков; всегда >0) и N (штрафной результат для несоответствующих остатков; всегда <0). Для последовательностей аминокислот используют матрицу оценки для расчета общего результата. Удлинение наиболее распространенных слов в каждом направлении прекращают, когда: результат кумулятивного выравнивания падает на величину Х относительно своего максимального достигнутого значения; кумулятивный результат доходит до нуля или меньше вследствие накопления одного или более выравниваний остатков с отрицательным результатом; или достигают конца любой из последовательностей. Параметры алгоритма BLAST W, Т и Х определяют чувствительность и скорость выравнивания. Программа BLASTN (для нуклеотидных последовательностей) использует по умолчанию длину слова (W) 11, ожидание (Е) 10, М=5, N=-4 и сравнение обеих цепей. Для последовательностей аминокислот программа BLASTP использует по умолчанию длину слова 3 и ожидание (Е) 10 и матрицу оценки выравниваний BLOSUM62 (см. статью Henikoff and Henikoff, Proc. Natl. Acad. Sci. USA 89:10915, 1989) (В) 50, ожидание (Е) 10, М=5, N=-4 и сравнение обеих цепей.
Алгоритм BLAST также осуществляет статистический анализ подобия между двумя последовательностями (см., например, статью Karlin and Altschul, Proc. Natl. Acad. Sci. USA 90:5873-5787, 1993). Одним измерением подобия. предусматриваемым алгоритмом BLAST, является наименьшая суммарная вероятность (P(N)), которая дает показатель вероятности, с которой могло бы случайно иметь место соответствие двух последовательностей нуклеотидов или аминокислот. Например, нуклеиновую кислоту считают подобной эталонной последовательности, если наименьшая суммарная вероятность в сравнении тестируемой нуклеиновой кислоты с эталонной нуклеиновой кислотой меньше чем приблизительно 0,2, более предпочтительно меньше чем приблизительно 0,01 и наиболее предпочтительно меньше чем приблизительно 0,001.
Отличным от вышеописанного процента идентичности последовательностей, другим показателем того, что две последовательности нуклеиновых кислот или полипептидов в существенной степени идентичны, является то, что полипептид, кодируемый первой нуклеиновой кислотой, иммунологическим перекрестно реактивен с антителами, образующимися к полипептиду, кодируемому второй нуклеиновой кислотой, как описано ниже. Таким образом, полипептид является типично в существенной степени идентичным второму полипептиду, например, когда два пептида отличаются только консервативными заменами. Другим показателем того, что две последовательности нуклеиновых кислот в существенной степени идентичны, является то, что две молекулы или их комплементы гибридизуются друг с другом в жестких условиях, как описано ниже. Еще одним показателем того, что две последовательности нуклеиновых кислот в существенной степени идентичны, является то, что для амплификации последовательностей могут быть использованы одни и те же праймеры.
Термин "функционально связанный" относится к функциональной связи между двумя или более полинуклеотидными (например, ДНК) сегментами. Как правило, он относится к функциональной связи последовательности регуляции транскрипции с транскрибированной последовательностью. Например, промоторная или энхансерная последовательность функционально связана с кодирующей последовательностью, если она стимулирует или модулирует транскрипцию кодирующей последовательности в соответствующей клетке-хозяине или другой экспрессирующей системе. Как правило, промоторные последовательности регуляции транскрипции, которые функционально связаны с транскрибируемой последовательностью, физически прилегают к транскрибируемой последовательности, т.е. они являются цис-действующими. Однако для некоторых последовательностей регуляции транскрипции, таких как энхансеры, необязательно быть физически прилежащими или находящимися в тесной близости к кодирующим последовательностям, транскрипцию которых они усиливают.
Термин "вектор" предназначен для обозначения полинуклеотидной молекулы, способной транспортировать другой полинуклеотид, к которому он присоединен. Одним типом вектора является "плазмида", которая относится к кольцевой петле двухцепочечной ДНК, в которую можно лигировать дополнительные сегменты ДНК. Другим типом вектора является вирусный вектор, причем дополнительные сегменты ДНК можно лигировать в вирусный геном. Некоторые векторы способны к автономной репликации в клетке-хозяине, в которую их интродуцируют (например, бактериальные векторы, имеющие бактериальный точка начала репликации, и эписомные векторы млекопитающих). Другие векторы (например, неэписомные векторы млекопитающих) можно интегрировать в геном клетки-хозяина путем интродукции в клетку-хозяин, и, таким образом, они реплицируются вместе с геномом хозяина. Кроме того, некоторые векторы способны направлять экспрессию генов, с которыми они функционально связаны. Данные векторы обозначают в данном контексте как "рекомбинантные экспрессирующие векторы" (или просто "экспрессирующие векторы"). Как правило, экспрессирующие векторы, используемые в технологиях рекомбинантной ДНК, часто находятся в форме плазмид. В настоящем описании термины "плазмида" и "вектор" могут быть использованы взаимозаменяемо, поскольку плазмида является наиболее часто используемой формой вектора. Однако изобретение предусматривает включение таких других форм экспрессирующих векторов, как вирусные векторы (например, репликационно-дефектные ретровирусы, аденовирусы и адено-ассоциированные вирусы), которые осуществляют эквивалентные функции.
Термин "рекомбинантная клетка-хозяин" (или просто "клетка-хозяин") относится к клетке, в которую интродуцирован рекомбинантный экспрессирующий вектор. Следует иметь в виду, что данные термины, как предусматривают, относятся не только к определенной конкретной клетке, но к потомству данной клетки. Поскольку некоторые модификации могут иметь место в последующих поколениях вследствие либо мутации, либо воздействий окружающей среды, данное потомство может в действительности не быть идентичным родительской клетке, но еще включено в объем термина "клетка-хозяин", как используют в данном контексте.
Термин "воспалительное заболевание или состояние" относится к любому состоянию, характеризующемуся местным воспалением в области повреждения или инфекции и включает аутоиммунные заболевания, определенные формы инфекционных воспалительных состояний, нежелательную активность нейтрофилов, характерную для трансплантатов органов или других имплантатов и в сущности любого другого состояния, характеризующегося нежелательным накоплением нейтрофилов в локальной области ткани. Данные состояния включают, но без ограничения перечисленным, менингит, отек головного мозга, артрит, нефрит, респираторный дистресс-синдром взрослых, панкреатит, миозит, неврит, заболевания соединительной ткани, флебит, артериит, васкулит, аллергию, анафилаксию, эрлихиоз, подагру, трансплантаты органов и/или язвенный колит.
Термин "субъект" включает человека и животных, отличных от человека. Животные, отличные от человека, включают всех позвоночных, например, млекопитающих и немлекопитающих, таких как приматы, отличные от человека, овца, собака, корова, куры, амфибии и рептилии. За исключением отмеченных случаев, термины "пациент" или "субъект" используют в данном контексте взаимозаменяемо.
Термин "лечение" включает введение соединений или агентов с целью предупреждения или задержки появления симптомов, осложнений или биохимических показателей заболевания (например, аллергического воспалительного заболевания), облегчения симптомов или остановки или подавления развития заболевания, состояния или нарушения. Лечение может быть профилактическим (для предупреждения или задержки появления заболевания или для предупреждения проявлений его клинических или субклинических симптомов) либо терапевтическим подавлением или облегчением симптомов после проявления заболевания.
Выражение "путь сигнальной трансдукции" или "путь передачи сигнала" (например, путь передачи сигнала TSLP) относится к по меньшей мере одной биохимической реакции, но чаще к серии биохимических реакций, которые являются результатом взаимодействия клетки со стимулирующим соединением или агентом. Так, взаимодействие стимулирующего соединения (например, TSLP) с клеткой генерирует "сигнал", который передается через путь сигнальной трансдукции, в конечном счете приводя к клеточному ответу, например, иммунному ответу.
II. Антагонистические антитела к человеческому TSLPR
1. Обзор
Изобретение представляет антитела, которые специфически связываются с человеческим TSLPR. Данные антитела к hTSLPR способны антагонистически воздействовать на TSLP-опосредованные активности передачи сигнала, например, TSLP-опосредованную пролиферацию клеток, как описано ниже в примерах. Общие методы получения моноклональных или поликлональных антител хорошо известны в области техники. См., например, монографию Harlow & Lane, Using Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1998; статьи Kohler & Milstein, Nature 256:495-497, 1975; Kozbor et al.. Immunology Today 4:72, 1983 и раздел Cole et al., стр.77-96 в монографии Monoclonal Antibodies and Cancer Therapy, 1985.
Предпочтительно, когда антитела к hTSLPR, представленные в изобретении, являются моноклональными, как мышиное моноклональное антитело, образующееся к человеческому TSLPR (клон 1D6.C9), как описано в примерах ниже. Моноклональные антитела относятся к антителам, выделенным из одного клона. Любой метод получения моноклонального антитела можно использовать для получения антител к hTSLPR, представленных в изобретении, например, вирусную или онкогенную трансформацию В-лимфоцитов. Одной из систем получения гибридом на животных является система на мышах. Получение гибридомы в мыши является очень хорошо разработанным методом. Как проиллюстрировано в примерах ниже, моноклональные антитела к hTSLPR можно генерировать путем иммунизации животного, отличного от человека (например, мышь) полипептидом hTSLPR или его фрагментом, слитым белком или вариантом. Затем В-клетки, выделенные у животного, сливают с клетками миеломы для генерации гибридом, продуцирующих антитело. Моноклональные мышиные антитела к hTSLPR можно получить скринингом гибридом в анализе ELISA с использованием полипептида hTSLPR или слитого белка. Протоколы иммунизации и методы выделения иммунизированных спленоцитов для слияния известны в области техники. Слитые партнеры (например, клетки мышиной миеломы) и методы слияния также хорошо известны в области техники, см., например, работу Harlow & Lane, выше.
Последовательности аминокислот тяжелой цепи (SEQ ID NO: 5) и вариабельные области легкой цепи (SEQ ID NO: 6) иллюстративного мышиного антитела к hTSLPR, описанные в нижеприведенных примерах, показаны на фиг.5. Кроме того, как указано на фигуре, последовательности CDR вариабельной области тяжелой цепи данного антитела представляют собой TYGMS (CDR1; SEQ ID NO: 7), WINTYSGVPRYADDFKG (CDR2; SEQ ID NO: 8) и EGFITTVVGAAGRFVY (CDR3; SEQ ID NO: 9). Последовательности CDR вариабельной области легкой цепи представляют собой KASQDVGTAVA (CDR1; SEQ ID NO: 10), WASTRHT (CDR2; SEQ ID NO: 11) и QQYSTYPT (CDR3; SEQ ID NO: 12).
Антитела взаимодействуют с антигенами-мишенями преимущественно через остатки аминокислот, которые находятся на шести участках, определяющих комплементарность, тяжелой и легкой цепи (CDR's). Как правило, антитела к hTSLPR, представленные в изобретении, имеют по меньшей мере одну из последовательностей CDR тяжелой цепи или последовательностей CDR легкой цепи, идентичную соответствующим последовательностям CDR, показанным на фиг.5. Некоторые из данных антител к hTSLPR, представленных в изобретении, имеют такую же специфичность связывания, как у иллюстративного мышиного антитела к hTSLPR (клон 1D6.C9), раскрытого в нижеприведенных примерах. Данные антитела могут конкурировать с мышиным антителом к hTSLPR (клон 1D6.C9) за связывание с hTSLPR. Некоторые антитела к hTSLPR, представленные в изобретении, имеют все последовательности CDR в своих вариабельных областях тяжелой цепи и легкой цепи, соответственно, идентичные соответствующим последовательностям CDR, показанным на фиг.5. Так, данные антитела к hTSLPR имеют три последовательности CDR тяжелой цепи, соответственно, идентичные SEQ ID NO: 7, SEQ ID NO: 8 и SEQ ID NO: 9 и три последовательности CDR легкой цепи, соответственно, идентичные SEQ ID NO: 10, SEQ ID NO: 11 и SEQ ID NO: 12.
Кроме того, что оно имеет последовательности CDR, соответственно, идентичные соответствующим последовательностям CDR мышиного антитела к hTSLPR (клон 1D6.C9), некоторые из антител к hTSLPR, представленных в изобретении, имеют свои полные последовательности вариабельной области тяжелой цепи и легкой цепи, соответственно, идентичные соответствующим последовательностям вариабельной области мышиного антитела, как показано на фиг.5 (т.е., SEQ ID NO: 5 и SEQ ID NO: 6). В ряде других вариантов осуществления, отличных от включающих идентичные последовательности CDR, антитела включают остатки аминокислот в скелетных частях вариабельных областей, которые отличаются от соответствующих остатков аминокислот, показанных на фиг.5 (например, ряд гуманизированных антител к hTSLPR, описанных ниже). Тем не менее, данные антитела, как правило, имеют последовательности вариабельной области, которые в существенной степени идентичны (например, на 75%, 85%, 90%, 95% или 99%) соответствующим последовательностям вариабельной области, показанным на фиг.5.
Антитела к hTSLPR, представленные в изобретении, могут представлять собой интактное антитело, которое включает две тяжелые цепи и две легкие цепи. Они могут также представлять собой антигенсвязывающие фрагменты интактного антитела или одноцепочечные антитела. Антитела к hTSLPR, представленные в изобретении, включают антитела, продуцируемые животным, отличным от человека (например, мышиное антитело к hTSLPR, показанное на фиг.5). Они также включают модифицированные антитела, которые представляют собой модифицированные формы мышиного антитела к hTSLPR, показанного на фиг.5. Часто модифицированные антитела представляют собой рекомбинантные антитела, которые имеют близкие или усовершенствованные свойства относительно свойств иллюстративного мышиного антитела. Например, мышиное антитело к hTSLPR, проиллюстрированное в нижеприведенных примерах, может быть модифицировано делецией константной области и замещением ее другой константной областью, которая может привести к увеличенному полупериоду существования, стабильности или аффинности антитела. Модифицированные антитела можно создать, например, путем конструирования экспрессирующих векторов, которые включают последовательности CDR из мышиного антитела, привитые на скелетные последовательности из другого антитела с другими свойствами (см. статью Jones et al., 1986, Nature 321, 522-525). Данные скелетные последовательности можно получить в общедоступных базах данных ДНК.
Некоторые из модифицированных антител представляют собой химерные антитела, которые включают частичные последовательности человеческого иммуноглобулина (например, константные области) и частичные последовательности нечеловеческого иммуноглобулина (например, последовательности вариабельной области мышиного антитела к hTSLPR, показанные на фиг.5). Некоторые другие модифицированные антитела представляют собой гуманизированные антитела. Как правило, гуманизированное антитело имеет один или более остатков аминокислот, интродуцированных в него из источника, который отличен от человека. Методы гуманизации нечеловеческих антител хорошо известны в области техники, например, см. патенты США №№5585089 и 5693762, статьи Jones et al., Nature 321: 522-25, 1986; Riechmann et al., Nature 332: 323-27, 1988 и Verhoeyen et al., Science 239: 1534-36, 1988. Данные методы легко использовать для генерации гуманизированных антител к hTSLPR, представленных в изобретении, посредством замещения по меньшей мере части CDR из нечеловеческого антитела к hTSLPR соответствующими участками человеческого антитела. В ряде вариантов осуществления гуманизированные антитела к hTSLPR, представленные в изобретении, имеют все три CDR в каждой цепи иммуноглобулина из мышиного антитела к hTSLPR, показанного на фиг.5, привитые на соответствующие человеческие скелетные участки.
Антитела к hTSLPR, описанные выше, могут подвергаться заменам, введениям или делециям неосновных аминокислот в обеих, вариабельной и константной областях без утраты специфичности связывания или эффекторных функций либо недопустимого снижения аффинности связывания. Обычно антитела, включающие данные изменения, проявляют существенную идентичность последовательности относительно эталонного антитела (например, мышиного антитела к hTSLPR, показанного на фиг.5), из которого они получены. Например, зрелые вариабельные области легкой цепи ряда антител к hTSLPR, представленных в изобретении, имеют идентичность последовательности по меньшей мере 75% или по меньшей мере 85% относительно последовательности зрелой вариабельной области легкой цепи антитела к hTSLPR, показанного на фиг.5. Аналогично зрелые вариабельные области тяжелой цепи антител, как правило, проявляют идентичность последовательности по меньшей мере 75% или по меньшей мере 85% относительно последовательности зрелой вариабельной области тяжелой цепи антитела к hTSLPR, показанного на фиг.5. Некоторые из модифицированных антител к hTSLPR имеют такую же специфичность и повышенную аффинность по сравнению с мышиным антителом к hTSLPR (клон 1D6.C9), показанным на фиг.5. Обычно аффинность модифицированных антител к hTSLPR (например гуманизированных антител) представляет собой аффинность связывания, которая является такой же или повышенной относительно исходного мышиного антитела. Аффинность связывания модифицированных антител составляет по меньшей мере 70%, 75%, 80%, 85%, 90%, 95% или 100% исходного мышиного антитела.
2. Химерные и гуманизированные антитела к hTSLPR
Некоторые из антител к hTSLPR, представленных в изобретении, являются (например, мышь/человек) антителами, которые получены из участков, выделенных из антагониста нечеловеческого антитела к hTSLPR вместе с участками человеческих антител. Например, химерная Н-цепь может включать антигенсвязывающий участок вариабельной области тяжелой цепи мышиного антитела к hTSLPR (например, последовательность, показанную в SEQ ID NO: 5), связанный с по меньшей мере частью константной области тяжелой цепи человека. Данная химерная тяжелая цепь может быть скомбинирована с химерной L-цепью, которая включает антигенсвязывающий участок вариабельной области легкой цепи мышиного антитела к hTSLPR (например, последовательностью, показанной в SEQ ID NO: 6), связанной с по меньшей мере частью константной области легкой цепи человека.
Химерные антитела к hTSLPR, представленные в изобретении, можно получить в соответствии с описанием в примерах, представленным ниже, а также методами, известными в области техники. Например, ген, кодирующий тяжелую цепь или легкую цепь молекулы мышиного моноклонального антитела к hTSLPR, можно разрезать рестрикциоными ферментами для того, чтобы удалить мышиный участок Fc, и заместить эквивалентной частью гена, кодирующего человеческую константную область Fc. Экспрессирующие векторы и клетки-хозяева, пригодные для экспрессии рекомбинантных антител и гуманизированных антител, в частности, хорошо известны в области техники, Векторы, экспрессирующие химерные гены, кодирующие цепи иммуноглобулина к hTSLPR, можно сконструировать с использованием стандартных рекомбинантных методов, например, см. монографии Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Press (3 изд., 2001) и Brent et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc. (стереотипное изд., 2003). Человеческие последовательности константной области можно выбрать из различных эталонных источников, включая, но без ограничения, перечисленные в работе Kabat et al., Sequences of Proteins of Immunological Interest, 5 изд., U.S. Department of Health and Human Services, U.S. Government Printing Office, 1991. Более специфические описания получения химерных антител путем рекомбинации ДНК также описаны в области техники, например, см. Robinson et al., Международная патентная публикация PCT/US86/02269; Akira et al., Европейская патентная заявка 184187, Taniguchi, М., Европейская патентная заявка 171496, Morrison et al., Европейская патентная заявка 173494; Neuberger et al., Международная заявка WO 86/01533, Cabilly et al. Патент США No. 4816567; Cabilly et al., Европейская патентная заявка 125,023, статьи Better (1988) Science 240:1041-1043; Liu (1987) PNAS 84:3439-3443; Liu (1987) J. Immunol. 139:3521-3526; Sun (1987) PNAS 84:214-218; Nishimura (1987) Cane. Res. 47:999-1005; Wood (1985) Nature 314:446-449; Shaw (1988) J. Natl. Cancer Inst. 80:1553-1559).
Химерные антитела, которые имеют целые вариабельные области из нечеловеческого антитела, могут быть далее гуманизированными для снижения антигенности антитела у человека. Это, как правило, сопровождается замещением ряда последовательностей или остатков аминокислот в вариабельных областях Fv (на скелетных участках или участках не-CDR) эквивалентными последовательностями или остатками аминокислот из человеческих вариабельных областей Fv. Данные дополнительно замещенные последовательности или остатки аминокислот обычно не включены непосредственно в связывание антигена. Чаще гуманизация нечеловеческого антитела предшествует замещению только CDR нечеловеческого антитела (например, мышиного антитела, показанного на фиг.5) CDR в человеческом антителе. В ряде случаев после этого следует замещение ряда дополнительных остатков на человеческих скелетных участках соответствующими остатками из нечеловеческого донорного антитела. Данное дополнительное прививание часто требуется для улучшения связывания с антигеном. Это обусловлено тем, что гуманизированные антитела, которые имеют только CDRs, привитые из нечеловеческого антитела, могут иметь пониженные относительно идеальных активности связывания по сравнению с активностями нечеловеческого донорного антитела. Таким образом в дополнение к CDR гуманизированные антитела к hTSLPR, представленные в изобретении, часто могут включать ряд остатков аминокислот на человеческом скелетном участке, замещенных соответствующими участками из нечеловеческого донорного антитела (например, мышиного антитела, показанного на фиг.5). Методы генерации гуманизированных антител путем замещения, включая критерии выбора скелетных остатков для замещения, хорошо известны с области техники. Например, кроме вышеупомянутых методов, относящихся к получению химерных антител, дополнительные материалы по получению гуманизированных антител представлены, например, в патентной заявке Соединенного королевства Winter et al. GB 2188638A (1987), патенте США 5225539; статьях Jones (1986) Nature 321:552-525, Verhoeyan et al. 1988 Science 239:1534 и Beidler (1988) J. Immunol. 141:4053-4060. Замещение CDR можно также осуществить при использовании сайт-направленного мутагенеза олигонуклеотидов, как описано, например, в WO 94/10332 под названием Humanized Antibodies to Fc Receptors for Иммуноглобулин G on Human Mononuclear Phagocytes.
Химерные или гуманизированные антитела к hTSLPR, представленные в изобретении, могут быть одновалентными, двухвалентными или поливалентными иммуноглобулинами. Например, одновалентное химерное антитело представляет собой димер (HL), сформированный химерной Н-цепью, связанной посредством дисульфидных мостиков с химерной L-цепью, как отмечено выше. Двухвалентное химерное антитело представляет собой тетрамер (Н2 L2), сформированный двумя димерами HL, связанными посредством по меньшей мере одного дисульфидного мостика. Поливалентное химерное антитело основано на агрегации цепей.
3. Человеческие антитела к hTSLPR
Кроме того, химерные или гуманизированные антитела к hTSLPR также включенные в изобретение, представляют собой полностью человеческие антитела, которые проявляют такую же специфичность связывания и сравнимую или повышенную аффинность связывания. Например, человеческие антитела могут иметь такие же или улучшенные свойства относительно свойств эталонного нечеловеческого антитела, которое включает последовательность вариабельной области тяжелой цепи SEQ ID NO: 5 и последовательность вариабельной области легкой цепи SEQ ID NO: 6. По сравнению с химерными или гуманизированными антителами антигенность при введении человеку человеческих антител к hTSLPR, представленных в изобретении дополнительно понижена.
Человеческие антитела к hTSLPR можно генерировать, используя методы, которые известны в области техники. Например, метод in vivo замещения вариабельной области нечеловеческого антитела человеческой вариабельной областью в антителе при поддержании таких же или получении усовершенствованных свойств относительно свойств нечеловеческого антитела раскрыто в патентной заявке США No. 10/778726 (Публикация No. 20050008625). Метод основан на направляемым эпитопом замещением вариабельных областей нечеловеческого эталонного антитела полностью человеческим антителом. Полученное в результате человеческое антитело, как правило, не является структурно родственным эталонному нечеловеческому антителу, но связывается с тем же эпитопом на том же антигене, что эталонное антитело. Вкратце, подход серийного направляемого эпитопом замещения комплементарности осуществляют путем установления конкуренции в клетках между "конкурирующим агентом" и библиотекой разнообразных гибридов эталонного антитела ("тест-антител") за связывание с ограниченными количествами антигена в присутствии сигнальной системы, которая отвечает на связывание тест-антитела с антигеном. Конкурирующий агент может представлять собой эталонное антитело или его производное, такое как одноцепочечный фрагмент Fv. Конкурирующий агент может также представлять собой природный или искусственный лиганд антигена, который связывается с тем же антигеном, который связывается с тем же эпитопом, что эталонное антитело. Единственными требованиями к конкурирующему агенту является то, что он связывается с тем же эпитопом, что эталонное антитело и то, что он конкурирует с эталонным антителом за связывание антигена. Тест-антитела имеют одну общую антигенсвязывающую V-область из нечеловеческого эталонного антитела и другую V-область, выбранную случайным образом из иного источника, такого библиотека репертуара человеческих антител. Общая V-область из эталонного антитела служит направляющим компонентом, помещающим тест-антитела на том же эпитопе на антигене и в той же ориентации, так что выбор направлен на самую высокую точность связывания антигена с эталонным антителом. Идентификацию TSLPR-связывающего домена осуществляют картированием эпитопа и показывают на фиг.8. Антитело к TSLPR связывается с непрерывным эпитопом.
Можно использовать многие типы сигнальной системы для детекции необходимых взаимодействий между тест-антителами и антигеном. Например, комплементирующие сигнальные фрагменты можно связать с антигеном и тест-антителом, соответственно, так что активация сигнала путем комплементации фрагмента имеет место только, когда тест-антитело связывается с антигеном. Когда слияния тест-антитело- и антиген-сигнальный фрагмент соэкспрессируются с конкурирующим агентом, сигнальная активация становится зависимой от способности тест-антитела конкурировать с конкурирующим агентом, которая пропорциональна аффинности тест-антитела в отношении антигена. Другие сигнальные системы, которые могут быть использованы, включают реактиватор аутоингибируемой системы реактивации сигнала (RAIR), как описано в патентной заявке США No. 10/208730 (публикация No. 20030198971), или систему конкурентной активации, раскрытую в патентной заявке США No. 10/076845 (публикация No. 20030157579).
При использовании системы серийного направляемого эпитопом замещения комплементарности проводят отбор с целью идентификации клеток, экспрессирующих одно тест-антитело наряду с конкурирующим агентом, антигеном и компонентами сигнала. В данных клетках каждое тест-антитело конкурирует один на один с конкурирующим агентом за связывание с ограниченным количеством антигена. Активность сигнала пропорциональна количеству антигена, связанного с тест-антителом, которое, в свою очередь, пропорциональна аффинности тест-антитела в отношении антигена и стабильности тест-антитела. Тест-антитела изначально выбирают на основе их активности относительно активности эталонного антитело при экспрессии в качестве тест-антитела. Результатом первого цикла отбора является набор "гибридных" антител, каждое из которых состоит из одной и той же нечеловеческой V-области из эталонного антитела и человеческой V-области из библиотеки, и каждое из которых связывается с одним и тем же эпитопом на антигене, что эталонное антитело. Одно или более из гибридных антител, отобранных в первом цикле, будет обладать аффинностью к антигену, сравнимой или превышающей активность эталонного антитела.
На второй стадии замещения V-области человеческие V-области, отобранные на первой стадии, используют как направляющие для отбора замещений человеческими элементами оставшейся V-области нечеловеческого эталонного антитела с использованием другой библиотеки соседних человеческих V-областей. Гибридные антитела, отобранные в первом цикле, также могут быть использованы в качестве конкурирующих агентов для второго цикла отбора. Результатом второго цикла отбора является набор полностью человеческих антител, которые структурно отличаются от эталонного антитела, но которые конкурируют с эталонным антителом за связывание с одним и тем же антигеном. Некоторые из отобранных человеческих антител связываются с тем же эпитопом на том же антигене, что эталонное антитело. Среди данных отобранных человеческих антител одно или более связываются с одним и тем же эпитопом с аффинностью, которая сравнима или превышает аффинность эталонного антитела.
С использованием одного из вышеописанных мышиных или химерных антител к hTSLPR в качестве эталонного антитела данный метод легко можно применять для генерации человеческих антител, которые связываются с человеческим TSLPR с той же специфичностью связывания и такой же или повышенной аффинностью. Кроме того, данные человеческие антитела к hTSLPR можно также получить коммерчески от фирм, которые традиционно производят человеческие антитела, например, KaloBios, Inc. (Mountain View, CA).
4. Другие типы антител к hTSLPR
Антитела к hTSLPR, представленные в изобретении, также включают одноцепочечные антитела, биспецифические антитела и мультиспецифические антитела. В ряде вариантов осуществления антитела, представленные в изобретении, представляют собой одноцепочечные антитела. Одноцепочечные антитела включают в одной стабильно сложенной полипептидной цепи антигенсвязывающие участки из обеих, тяжелой цепи и легкой цепи. В таком случае одноцепочечные антитела, как правило, сохраняют специфичность связывания и аффинность моноклональных антител, но имеют размер, значительно меньше, чем у классических иммуноглобулинов. Для ряда применений одноцепочечные антитела к hTSLPR, представленные в изобретении, могут давать множество благоприятных свойств по сравнению с интактным антителом к hTSLPR. Они включают, например, ускоренное выведение из организма, повышенную проницаемость тканей, как для диагностической визуализации, так и для терапии, и существенное снижение иммуногенности по сравнению с антителами на мышиной основе. Другие потенциальные преимущества использования одноцепочечных антител включают улучшенные возможности скрининга в высокоэффективных методах скрининга и возможность непарентерального применения.
Одноцепочечные антитела к hTSLPR, представленные в изобретении, можно получить, используя методы, которые описаны в области техники. Примеры данных методов включают описанные в патентах США NoNo 4946778 и 5258498, статьях Huston et al., Methods in Enzymology 203:46-88, 1991; Shu et al., Proc. Natl. Acad. Sci. USA 90:7995-7999, 1993 и Skerra et al., Science 240:1038-1040, 1988.
В некоторых вариантах осуществления изобретение представляет антитела к hTSLPR, дериватизированные или связанные с другой функциональной молекулой с целью генерации биспецифической или мультиспецифической молекулы, которая связывается с множеством центров связывания или целевых эпитопов. Функциональная молекула включает другой пептид или белок (например, цитокин, цитотоксический агент, иммуностимулирующий или ингибирующий агент, фрагмент Fab' или другой фрагмент, связывающий антитело, как обсуждалось выше). Например, антитело к hTSLPR или его антигенсвязывающая часть могут быть функционально связаны (например, путем химического связывания, генетического слияния, нековалентной ассоциации или иным образом) с одной или более из других связывающих молекул, таких как другое антитело, фрагмент антитела, пептид или связывающий миметик. Так, биспецифические и мультиспецифические антитела к hTSLPR, представленные в изобретении, включают по меньшей мере одно моноклональное антитело к hTSLPR или его антигенсвязывающий фрагмент с первой специфичностью связывания с человеческим TSLPR и второй специфичностью связывания со вторым эпитопом-мишенью. Второй эпитоп-мишень может представлять собой Fc-рецептор, например, человеческий FcγRI или человеческий Fcγ-рецептор. Вследствие этого изобретение включает биспецифические и мультиспецифические молекулы, способные связываться как с FcγRI, FcγR или FcεR-экспрессирующими эффекторными клетками (например, моноцитами, макрофагами или полиморфонуклеарными клетками (PMNs)), так и с клетками-мишенями, экспрессирующими человеческий TSLPR (например, человеческими дендритными клетками CD11c+). Данные мультиспецифические (например, биспецифические или мультиспецифические) молекулы направляют человеческие TSLPR-экспрессирующие клетки на эффекторные клетки и переключают опосредованные Fc-рецептором активности эффекторных клеток, такие как фагоцитоз человеческих TSLPR-экспрессирующих клеток, антитело-зависимая опосредованная клетками цитотоксичность (ADCC), высвобождение цитокинов или генерация супероксидного аниона.
Биспецифические и мультиспецифические молекулы к hTSLPR, представленные в настоящем изобретении, можно получить методами, которые описаны в области техники. Они включают химические методы (см., например, статью Kranz, Proc. Natl. Acad. Sci. USA 78:5807, 1981), полидомные методы (см., например, патент США No. 4474893) или технологии рекомбинантной ДНК. Биспецифические и мультиспецифические молекулы, представленные в настоящем изобретении, можно также получить конъюгированием составляющих специфичностей связывания, например, специфичностей связывания с антителом к FcR и антителом к человеческому TSLPR, с использованием методов, известных в области техники и как описано в данном контексте. Например, каждую специфичность связывания биспецифической и мультиспецифической молекулы можно генерировать отдельно и затем конъюгировать друг с другом. Когда специфичности связывания представляют собой белки или пептиды, можно использовать ряд связывающих или перекрестно связывающих агентов для ковалентного конъюгирования. Примеры перекрестно-связывающих агентов включают белок А, карбодиимид, N-сукцинимидил-S-ацетил-тиоацетат (SATA), N-сукцинимидил-3-(2-пиридилтио)пропионат (SPDP) и сульфосукцинимидил 4-(N-малеимидометил)циклогексан-1-карбоксилат (сульфо-SMCC). Когда специфичности связывания представляют собой антитела (например, два гуманизированных антитела), они могут быть конъюгированы посредством сульфгидрильного связывания С-концевых шарнирных участков двух тяжелых цепей. Шарнирный участок может быть модифицирован так, чтобы он включал перед конъюгированием нечетное число сульфгидрильных остатков, например, один.
Связывание биспецифических и мультиспецифических молекул с их специфическими мишенями можно подтвердить с помощью твердофазного иммуноферментного анализа (ELISA), радиоиммуноанализа (RIA) или анализа вестерн-блот.Каждый из данных анализов в основном определяет присутствие комплексов белок-антитело, представляющих особый интерес, с применением меченого реагента (например, антитела), специфического в отношении комплекса, представляющего интерес.Например, комплексы FcR-антитело можно определить с использованием, например, антитела, связанного с ферментом, или фрагмента антитела, который распознает и специфически связывается с комплексами антитело-FcR. Альтернативно комплексы можно определить, используя любой из множества других иммуноанализов. Например, в антитело можно ввести радиоактивную метку и использовать в радиоимуноанализе (RIA) (см., например, монографию Weintraub, В., Principles ofRadioimmunoassays, Seventh Training Course on Radioligand Assay Techniques, The Endocrine Society, March, 1986). Радиоактивный изотоп можно определить такими средствами, как γ-счетчик или сцинтилляционный счетчик либо с помощью авторадиографии.
III. Полинуклеотиды, векторы и клетки-хозяева для получения антител к hTSLPR
Изобретение представляет в существенной степени очищенные полинуклеотиды (ДНК или РНК), которые кодируют полипептиды, включающие сегменты или домены вышеописанных цепей антитела к hTSLPR. Некоторые полинуклеотиды, представленные в изобретении, включают нуклеотидную последовательность вариабельной области тяжелой цепи, показанной в SEQ ID NO: 13 и/или нуклеотидную последовательность вариабельной области легкой цепи, показанной в SEQ ID NO: 14. Ряд других полинуклеотидов, представленных в изобретении, включает нуклеотидные последовательности, которые в существенной степени идентичны (например, по меньшей мере на 65, 80%, 95% или 99%) нуклеотидным последовательностям SEQ ID NO: 13 или SEQ ID NO: 14. При экспрессии с соответствующих экспрессирующих векторов полинуклеотиды, кодируемые данными полинуклеотидами, способны проявлять способность связывать антиген.
Кроме того, в изобретении представлены полинуклеотиды, которые кодируют по меньшей мере один участок CDR и, как правило, все три участка CDR из тяжелой или легкой цепи антитела к hTSLPR, показанного на фиг.5. Ряд других полинуклеотидов кодирует все или в существенной степени все последовательности вариабельной области тяжелой цепи и/или легкой цепи антитела к hTSLPR, показанного на фиг.5. Например, некоторые из данных полинуклеотидов кодируют последовательность аминокислот вариабельной области тяжелой цепи, показанную в SEQ ID NO: 5 и/или последовательность аминокислот вариабельной области легкой цепи, показанную в SEQ ID NO: 6. Вследствие вырожденности кода множество последовательностей нуклеиновых кислот будет кодировать каждую из последовательностей аминокислот иммуноглобулина.
Полинуклеотиды, представленные в изобретении, могут кодировать только последовательность вариабельной области антитела к hTSLPR. Они могут также кодировать как вариабельную область, так и константную область антитела. Некоторые полинуклеотидные последовательности, представленные в изобретении, представляют собой нуклеиновые кислоты, кодирующие зрелую последовательность вариабельной области тяжелой цепи, которая в существенной степени идентичны (например, по меньшей мере на 80%, 90% или 99%) зрелой последовательности вариабельной области тяжелой цепи мышиного антитело к hTSLPR 1D6.C9, показанной в SEQ ID NO: 5. Некоторые другие полинуклеотидные последовательности кодируют зрелую последовательность вариабельной области легкой цепи, которая в существенной степени идентична зрелой последовательности вариабельной области легкой цепи мышиного антитела 1D6.C9, показанной в SEQ ID NO: 6. Ряд полинуклеотидных последовательностей кодирует полипептид, который включает вариабельные области, как тяжелой цепи, так и легкой цепи мышиного антитела. Некоторые другие полинуклеотиды кодируют два полипептидных сегментов, которые, соответственно, в существенной степени идентичны вариабельным областям тяжелой цепи и легкой цепи мышиного антитела.
Полинуклеотидные последовательности можно получить твердофазным синтезом ДНК de novo или путем ПЦР-мутагенеза имеющейся последовательности (например, последовательностей, как описано в нижеприведенных примерах), кодирующей антитело к hTSLPR или его связывающий фрагмент.Прямой химический синтез нуклеиновых кислот можно осуществить методами, известными в области техники, такими как фосфотриэфирный метод, предложенный в статье Narang et al., 1979, Meth. Enzymol. 68:90; фосфодиэфирный метод, предложенный в статье Brown et al., Meth. Enzymol. 68:109, 1979; диэтилфосфорамидитный метод, предложенный Beaucage et al., Tetra. Lett., 22:1859, 1981, и метод с твердой основой, представленный в патенте США No. 4458066. Интродукцию мутаций в полинуклеотидную последовательность посредством ПЦР можно осуществить, как описано, например, в монографиях PCR Technology: Principles and Applications for DNA Amplification, под ред. H.A. Eriich, Freeman Press, NY, NY, 1992; PCR Protocols: A Guide to Methods and Applications, под ред. Innis et al., Academic Press, San Diego, CA, 1990; статьях Mattila et al., Nucleic Acids Res. 19:967, 1991 и Eckert et al., PCR Methods and Applications 1:17, 1991.
Кроме того, в изобретении представлены вышеописанные экспрессирующие векторы и клетки-хозяева для получения антитела к hTSLPR. Различные экспрессирующие векторы можно использовать для экспрессии полинуклеотидов, кодирующих цепи или связывающие фрагменты антитела к hTSLPR. Экспрессирующие векторы, как на вирусной основе, так и невирусной природы, можно использовать для получения антител в клетке млекопитающего-хозяина. Невирусные векторы и системы включают плазмиды, эписомные векторы, как правило, с экспрессирующей кассетой для экспрессии белка или РНК, и человеческие искусственные хромосомы (см., например, статью Harrington et al., Nat Genet 15:345, 1997). Например, невирусные векторы, используемые для экспрессии полинуклеотидов и полипептидов антител к hTSLPR в клетках млекопитающих (например, человека), включают pThioHis A, В & С, pcDNA3. I/His, pEBVHis А, В & С, (Invitrogen, San Diego, CA), векторы MPSV и множество других векторов, известных в области техники, как используемые для экспрессии других белков. Используемые вирусные векторы включают векторы на основе ретровирусов, аденовирусов, аденоассоциированных вирусов, векторы на основе SV40, вируса папилломы, вируса Эпштейна-Барр НВР, векторы на основе вируса вакцины и вируса Semliki Forest (SFV). См. статьи Brent et al., выше; Smith, Annu. Rev. Microbiol. 49:807, 1995 и Rosenfeld et al., Cell 68:143, 1992.
Выбор экспрессирующего вектора зависит от предусматриваемых клеток-хозяев, в которых должен экспрессироваться вектор. Как правило, экспрессирующие векторы включают промотор и другие регуляторные последовательности (например, энхансеры), которые функционально связаны с полинуклеотидами, кодирующими цепь или фрагмент антитела к hTSLPR. В ряде вариантов осуществления используют индуцируемый промотор для предупреждения экспрессии введенных последовательностей в случаях кроме индуцирующих условий. Индуцируемые промоторы включают, например, промотор арабинозы, lacZ, металотионеина или промотор теплового шока. Культуры трансформированных организмов можно размножить в неиндуцирующих условиях без сдвига популяции в сторону кодирующих последовательностей, продукты экспрессии которых лучше переносятся клетками-хозяевами. В дополнение к промоторам могут также потребоваться или быть желательными другие регуляторные элементы для эффективной экспрессии цепи или фрагмента антитела к hTSLPR. Данные элементы, как правило, включают кодон инициации ATG и прилежащий сайт связывания рибосом или другие последовательности. Кроме того, эффективность экспрессии можно повысить включением энхансеров, подходящих для используемой клеточной системы (см., например, статьи Scharfet al.. Results Probl. Cell Differ. 20:125, 1994; and Bittner et al., Meth. Enzymol., 153:516, 1987). Например, энхансер SV40 или энхансер CMV можно использовать для повышения уровня экспрессии в клетках млекопитающих-хозяевах.
Экспрессирующие векторы могут также представлять положение последовательности сигнала секреции для формирования слитого белка с полипептидами, кодируемыми введенными последовательностями антитела к hTSLPR. Чаще введенные последовательности антитела к hTSLPR связывают с сигнальными последовательностями перед включением в вектор. Векторы, предназначенные для использования для получения последовательностей, кодирующих домены легкой и тяжелой цепей антитела к hTSLPR, иногда кодируют также константные области или их части. Данные векторы обеспечивают экспрессию вариабельных областей в виде слитых белков с константными областями, приводя, таким образом, к продукции интактных антител или их фрагментов. Как правило, данные константные области являются человеческими.
Клетки-хозяева, предназначенные для того, чтобы нести и экспрессировать цепи антитела к hTSLPR, могут быть либо прокариотическими, либо эукариотическими. Е. coli представляет собой одного из прокариотических хозяев, используемых для клонирования и экспрессии полинуклеотидов, представленных в настоящем изобретении. Другие микробные хозяева, подходящие для применения, включают бациллы, такие как Bacillus subtilis, и другие представители энтеробактерий, такие как Salmonella, Serratia и различные виды Pseudomonas. В данных прокариотических хозяевах можно также создать экспрессирующие векторы, которые, как правило, включают последовательности контроля экспрессии, совместимые с клеткой-хозяином (например, ориджин репликации). Кроме того, будет присутствовать какое-либо число из ряда хорошо известных промоторов, таких как система лактозного промотора, система триптофанового (trp) промотора, система промотора β-лактамазы или система промотора из фага λ. Промоторы, как правило, контролируют экспрессию, необязательно с последовательностью оператора, и имеют последовательности сайта связывания рибосом и т.п. для инициации и окончания транскрипции и трансляции. Другие микробы, такие как дрожжи, можно также использовать для экспрессии полипептидов антитела к hTSLPR, представленных в изобретении. Можно также использовать клетки насекомых в комбинации с бакуловирусными векторами.
В ряде предпочтительных вариантов осуществления клетки млекопитающих-хозяева используют для экспрессии и получения полипептидов антитела к hTSLPR, представленного в настоящем изобретении. Например, они могут представлять собой либо гибридомную клеточную линию, экспресирующую эндогенные гены иммуноглобулина (например, клон гибридомы миеломы 1D6.C9, как описано в примерах), либо линию клеток млекопитающих, несущую экзогенный экспрессирующий вектор (например, клетки миеломы SP2/0, показанные ниже). Они включают любую нормальную смертную или нормальную или патологическую иммортализованную клетку животного или человека. Например, создан ряд подходящих линий клеток-хозяев, способных секретировать интактные иммуноглобулины, включающие клеточные линии СНО, различные клеточные линии Cos, клетки HeLa, клеточные линии миеломы, трансформированные В-клетки и гибридомы. Использование клеточной культуры ткани млекопитающего для экспрессии полипептидов обсуждают в общем, например, в монографии Winnacker, From Genes to Clones, VCH Publishers, N.Y., N.Y., 1987. Экспрессирующие векторы для клеток млекопитающих-хозяев могут включать последовательности контроля экспрессии, такие как точка начала репликации, промотор и энхансер (см., например, статью Queen, et al., Immunol. Rev. 89:49-68, 1986), и необходимые сайты процессинга информации, такие как сайты связывания рибосом, сплайс-сайты РНК, сайты полиаденилирования и последовательности терминатора транскрипции. Данные экспрессирующие векторы обычно включают промоторы, выделенные из генов млекопитающих или из вирусов млекопитающих. Подходящие промоторы могут быть конститутивными, специфическими в отношении типа клеток, специфическими для стадии и/или модулируемыми либо регулируемыми. Используемые промоторы включают, но без ограничения перечисленным, промотор металлотионеина, конститутивный главный поздний промотор аденовируса, индуцируемый дексаметазоном промотор MMTV, промотор SV40, промотор MRP polIII, конститутивный промотор MPSV, индуцируемый тетрациклином промотор CMV (такой как человеческий средне-ранний промотор CMV), конститутивный промотор CMV и комбинации промотор-энхансер, известные в области техники.
Методы интродукции экспрессирующих векторов, включающих полинуклеотидные последовательности, представляющие интерес, варьируют в зависимости от типа клеточного хозяина. Например, трансфекцию с помощью хлорида кальция обычно используют для прокариотических клеток, тогда как обработку фосфатом кальция или электропорацию можно использовать для других клеточных хозяев. (См. в основном Sambrook, et al., выше). Другие методы включают, например, электропорацию, обработку фосфатом кальция, трансформацию, опосредованную липосомами, инъекцию и микроинъекцию, баллистические методы, виросомы, иммунолипосомы, конъюгаты поликатион:нуклеиновая кислота, голую ДНК, искусственные вирионы, слияние структурного белка вируса герпеса VP22 (см. статью Elliot and O'Hare, Cell 88:223, 1997), усиленное агентом поглощение ДНК и трансдукцию ex vivo. Для длительного высокопродуктивного получения рекомбинантных белков будет часто требоваться стабильная экспрессия. Например, клеточные линии, которые стабильно экспрессируют цепи или связывающие фрагменты антитела к hTSLPR можно получить, используя экспрессирующие векторы, представленные в изобретении, которые включают вирусные точки начала репликации или эндогенные элементы экспрессии и ген селектируемого маркера. После интродукции вектора клеткам можно дать возможность расти в течение 1-2 дней в обогащенной среде перед тем, как их переведут на селективную среду. Целью селектируемого маркера является придать устойчивость к отбору, и его присутствие обеспечивает рост клеток, которые успешно экспрессируют интродуцированные последовательности с селективных средах. Можно обеспечить пролиферацию устойчивых стабильно трансфицированных клеток с использованием методов тканевых культур, подходящих к типу клеток.
IV. Свойства антител к hTSLPR
После того, как вышеописанное антитело к hTSLPR экспрессируется с экспрессирующего вектора в клетке-хозяине или эндогенно с гибридоме, его легко очистить из культуральной среды и клеток-хозяев. Как правило, цепи антитела экспрессируются с сигнальными последовательностями и, таким образом, высвобождаются в культуральную среду. Однако, если цепи антитела не секретируются естественным образом клетками-хозяевами, цепи антитела можно высвободить обработкой мягким детергентом. Затем цепи антитела можно очистить принятыми методами, в том числе осаждением сульфатом аммония, аффинной хроматографией с иммобилизованной мишенью, колоночной хроматографией, гель-электрофорезом и т.п. Данные методы хорошо известны и рутинно практически используются в области техники, см., например, работы Scopes, Protein Purification, Springer-Verlag, NY, 1982 и Harlow & Lane, выше.
В качестве примера, отобранные гибридомы, экспрессирующие антитела к hTSLPR, представленные в изобретении, можно выращивать во вращающихся колбах объемом два литра с целью очистки моноклональных антител. Супернатанты можно отфильтровать и концентрировать перед проведением аффинной хроматографии с белок А-сефарозой или белок G-сефарозой (Pharmacia, Piscataway, NJ). Элюированный IgG можно проверить гель-электрофорезом и высокоэффективной жидкостной хроматографией для обеспечения чистоты. Буферный раствор можно заменить на PBS и концентрацию можно определить чтением при оптической плотности OD280. Можно взять аликвоты моноклональных антител и хранить при -80°С.
Независимо от способа получения моноклональные антитела к hTSLPR, представленные в настоящем изобретении, специфически связываются с hTSLPR или его антигенным фрагментом. Специфическое связывание имеет место, когда константа диссоциации для связывания антитела с hTSLPR или его антигенным фрагментом составляет ≤1 мкМ, предпочтительно ≤100 нМ, и наиболее предпочтительно ≤1 нМ. Способность антитела связываться с hTSLPR можно определить мечением антитела, представляющего интерес, непосредственно, или антитело может быть немеченым и связывание определяют опосредованно, используя различные форматы сандвичевых анализов. См., например, работу Harlow & Lane, выше. Антитела, имеющие данную специфичность связывания, с большей вероятностью разделяют благоприятные свойства, проявляемые мышиным антителом 1D6.C9 к hTSLPR, обсуждаемым ниже в примерах.
Моноклональные антитела к TSLPR, представленные в изобретении, способны антагонистически воздействовать на активности передачи сигнала, опосредованные TSLP. Данные активности включают, например, секрецию Тн2-привлекающих хемокинов дендритными клетками, такими как TARC и MDC; активацию дендритных клеток, экспансию непримированных (наивных) Т-клеток CD4+ и поляризацию в отношении фенотипа TH2, продукцию проаллергических цитокинов, таких как ИЛ-4, ИЛ-5, ИЛ-13 TNFα. Можно использовать ряд анализов для определения, может ли антитело к hTSLPR ингибировать TSLP-опосредованные клеточные активности. Они включают, например, любой из анализов, описанных в примерах, таких как анализ пролиферации клеток с использованием клеток Ba/F3/hTSLPR/hIL7Rα, анализ сигнала люциферазы с использованием клеток Ba/F3/hTSLPR/IL7Rα/Stat5-Luc и анализ секреции TARC. Дополнительные анализы для измерения активности передачи сигнала TSLP также описаны в области техники. См., например, статьи Reche et al., J. ImmunoL, 167:336-43, 2001 и Isaksen et al., J Immunol. 168:3288-94, 2002.
В ряде вариантов осуществления антитела к hTSLPR, представленные в изобретении, блокируют или конкурируют за связывание с эталонным антителом к hTSLPR, имеющим последовательности вариабельной области, показанные на Fig.5 (например, мышиным антителом 1D6.C9 или его химерным антителом, описанным в примерах ниже) к полипептиду hTSLPR. Они могут представлять собой вышеописанные полностью человеческие антитела к hTSLPR. Они могут также являться другими мышиными, химерными или гуманизированными антителами к hTSLPR, которые связываются с тем же эпитопом, что эталонное антитело. Способность блокировать или конкурировать со связыванием эталонного антитела показывает, что исследуемое антитело к hTSLPR связывается с тем же или близким эпитопом, который определен эталонным антителом, или с эпитопом, который в существенной степени проксимален эпитопу, связанному эталонным антителом к hTSLPR. Данные антитела с особенно высокой вероятностью разделяют благоприятные свойства, идентифицированные у эталонного антитела. Способность блокировать или конкурировать с эталонным антителом можно определить, например, анализом конкурентного связывания. С помощью анализа конкурентного связывания тестируемое антитело исследуют на способность ингибировать специфическое связывание эталонного антитела с общим антигеном, таким как полипептид TSLPR. Тест-антитело конкурирует с эталонным антителом за специфическое связывание с антигеном, если избыток тест-антитела в существенной степени ингибирует связывание эталонного антитела. Существенное ингибирование означает, что тест-антитело снижает специфическое связывание эталонного антитела обычно по меньшей мере на 10%, 25%, 50%, 75% или 90%.
Имеется ряд известных анализов конкурентного связывания, которые могут быть использованы для оценки конкуренции антитела к hTSLPR с эталонным антителом к hTSLPR за связывание с человеческим TSLPR. Они включают, например, твердофазный прямой или непрямой радиоиммуноанализ (RIA), твердофазный прямой или непрямой ферментный иммуноанализ (EIA), сандвичевый конкурентный анализ (см. статью Stahli et al.. Methods in Enzymology 9:242-253, 1983); твердофазный прямой EIA с использованием биотин-авидина (см. статью Kirkland et al., J. Immunol. 137:3614-3619, 1986); твердофазный анализ с прямым введением метки, твердофазный сандвичевый анализ с прямым введением метки (см. работу Harlow & Lane, выше); твердофазный RIA с прямым введением метки с использованием метки 1-125 (см. статью Morel et al., Molec. Immunol. 25:7-15, 1988); твердофазный прямой EIA с использованием авидин-биотина (см. статью Cheung et al., Virology 176:546-552, 1990) и RIA с прямым введением метки (см. статью Moldenhauer et al., Scand. J. Immunol. 32:77-82, 1990). Как правило, данный анализ включает применение очищенного антигена, связанного с твердой поверхностью или клетками, несущими один из следующих компонентов: немеченое тест-антитело к hTSLPR и меченое эталонное антитело. Конкурентное ингибирование измеряют путем определения количества метки, связанной с твердой поверхностью или клетками в присутствии тест-антитела. Как правило, тест-антитело присутствует в избытке. Антитела, идентифицированные конкурентным анализом (конкурентные антитела), включают антитела, связывающие тот же самый эпитоп, что эталонное антитело, и антитела, связывающиеся в соседним эпитопом, в существенной степени проксимальным эпитопу, связанному эталонным антителом, вследствие существования стерического препятствия.
Для того, чтобы определить, связывают ли отобранные моноклональные антитела к TSLPR уникальные эпитопы, каждое антитело можно биотинилировать, используя коммерчески доступные реагенты (например, реагенты фирмы Pierce, Rockford, IL). Исследования конкуренции с использованием немеченых моноклональных антител и биотинилированых моноклональных антител можно провести, используя планшеты для ELISA, покрытые полипептидом TSLPR. Связывание биотинилированных MAb можно определить с помощью зонда со стреп-авидин-щелочной фосфатазой. Для установления изотипа очищенного антитело к hTSLPR проводят ELISA изотипа. Например, лунки планшетов для микротитрования могут быть покрыты 1 мкг/мл IgG к человеку в течение ночи при 4°С. После блокирования 1% BSA (бычьим сывороточным альбумином) проводят реакцию планшетов с 1 мкг/мл или меньше моноклонального антитела к hTSLPR или очищенных контролей изотипов при комнатной температуре в течение одного-двух часов. Затем можно провести реакцию лунок либо с человеческим IgGI, либо с зондами, конъюгированными с человеческим IgM-специфической щелочной фосфатазой. Затем планшеты проявляют и анализируют, чтобы можно было определить изотип очищенного антитела.
Для того, чтобы продемонстрировать связывание моноклональных антител к hTSLPR с живыми клетками, экспрессирующими полипептид hTSLPR, можно использовать проточную цитометрию. Вкратце, клеточные линии, экспрессирующие hTSLPR (выращенные со стандартных условиях роста) можно смешать с различными концентрациями антитела к hTSLPR в PBS (забуференный фосфатом солевой раствор), содержащем 0,1% BSA и 10% сыворотки телячьих эмбрионов, и инкубировать при 37°С в течение 1 часа. После промывания проводят реакцию клеток с меченным флуоресцеином антителом IgG к человеку в тех же условиях, что для окрашивания первичного антитела. Образцы можно анализировать на приборе FACScan, используя свет и характеристики бокового рассеяния для пропускания единичных клеток. В альтернативном анализе с использованием флуоресцентной микроскопии можно использовать дополнительно или вместо) анализ с помощью проточной цитометрии. Клетки можно окрасить точно так же, как описано выше, и исследовать флуоресцентной микроскопией. Данный метод дает возможность визуализации отдельных клеток, но может иметь пониженную чувствительность в зависимости от плотности антигена.
Антитела к hTSLPR, представленные в изобретении, можно далее тестировать на реактивность с помощью полипептида или антигенного фрагмента hTSLPR вестерн-блоттингом. Вкратце, можно получить очищенные полипептиды или слитые белки hTSLPR либо клеточные экстракты, полученные из клеток, экспрессирующих TSLPR, и подвергнуть электрофорезу в полиакриламидном геле с додецилсульфатом натрия. После электрофореза разделенные антигены переносят на нитроцеллюлозные мембраны, блокируют 10% сывороткой телячьих эмбрионов и зондируют тестируемыми моноклональными антителами. Связывание человеческого IgG можно определить, используя IgG к человеку - щелочную фосфатазу и проявляют таблетками субстрата BCIP/NBT (Sigma Chem. Co., St. Louis, МО).
V. Неиммуноглобулиновые основы
Можно использовать широкий круг скелетов антител/иммуноглобулинов до тех пор, пока полученный в результате полипептид включает по меньшей мере один участок связывания, который специфичен в отношении белка-мишени. Данные скелеты или основы включают 5 главных идиотипов человеческих иммуноглобулинов или их фрагменты (такие как описаны в другом месте в данном контексте) и включают иммуноглобулины других видов животных, предпочтительно имеющие гуманизированные аспекты. Антитела с одной тяжелой цепью, такие как идентифицированные у верблюжьих, представляют собой в этом плане особенный интерес. Компетентные специалисты в области техники продолжают обнаруживать и разрабатывать новые скелеты, основы и фрагменты.
В одном аспекте изобретение относится к генерации антител на неиммуноглобулиновой основе с использованием неиммуноглобулиновых основ, на которые можно прививать CDR, представленные в изобретении. Известные или еще не найденные неиммуноглобулиновые скелеты и основы можно использовать до тех пор, пока они включают участок связывания, специфический в отношении белка-мишени SEQ ID NO: X. Данные соединения известны в данном контексте как "полипептиды, включающие мишень-специфический участок связывания". Известные неиммуноглобулиновые скелеты и основы включают, но без ограничения перечисленным, аднектины (фибронектин) (Compound Therapeutics, Inc., Waltham, MA), анкирин (Molecular Partners AG, Zurich, Switzerland), домены антител (Domantis, Ltd (Cambridge, MA) и аблинкс nv (Zwijnaarde, Belgium)), липокаин (антикалин) (Pieris Proteolab AG, Freising, Germany), маленькие модульные иммунофармацевтические агенты (Trubion Фармацевтическая Inc., Seattle, WA), макситела (Avidia, Inc. (Mountain View, CA)), белок A (Affibody AG, Sweden) и аффилин (γ-кристаллин или убиквитин) (Scil Proteins GmbH, Halle, Germany).
(i) Аднектины - терапевтические соединения
Аднектиновые основы базируются на домене фибронектина типа III (например, десятом модуле фибронектина типа III (домене 10 Fn3). Домен фибронектина типа III имеет 7 или 8 β-цепей, которые распределены между двумя β-складками, которые, в свою очередь, упакованы друг против друга с образованием ядра белка, и, кроме того, включают петли (аналогичные CDRs), которые связывают β-цепи друг с другом и доступны для воздействия растворителя. Существует по меньшей мере три таких петли на каждом краю сандвича β-складки, где край представляет собой границу белка, перпендикулярную направлению β-цепей. (см. US 6818418).
Данные основы на базе фибронектина не являются иммуноглобулином, хотя общая складка очень близка складке самого маленького функционального фрагмента антитела, вариабельной области тяжелой цепи, которая включает полный элемент распознавания антигена в IgG верблюда и ламы. Вследствие данной структуры неиммуноглобулиновое антитело имитирует свойства связывания антигена, которые аналогичны по природе и аффинности данным свойствам антител. Данные основы можно использовать в стратегии рандомизации петли и перестановки in vitro, которая подобна процессу созревания аффинности антител in vivo. Данные молекулы на базе фибронектина можно использовать как основы, в которых участки петель молекулы могут быть замещены CDR, представленными в изобретении, с помощью стандартных методов клонирования.
(ii) Анкирин-молекулярные партнеры
Способ основан на использовании белков с выделенными из анкирина модулями повторов в качестве основ для того, чтобы нести вариабельные области, которые могут быть использованы для связывания различных мишеней. Модуль анкиринового повтора представляет собой полипептид из 33 аминокислот, состоящий из двух антипараллельных α-спиралей и β-витка. Связывание вариабельных областей обычно оптимизируют, используя рибосомное представление.
(iii) Макситела/авимеры - Avidia
Авимеры получают из белка, включающего природный А-домен, такого как LRP-1. Данные домены используют по природе для взаимодействий белок-белок, и у человека более 250 белков структурно основаны на А-доменах. Авимеры состоят из ряда различных мономеров "А-домена" (2-10), связанных посредством аминокислотных линкеров. Можно создать авимеры, которые могут связываться с антигеном-мишенью, с помощью метода, описанного, например, в 20040175756, 20050053973, 20050048512 и 20060008844.
(vi) Белок А - Affibody
Аффинные лиганды Affibody® представляют собой аленькие простые белки, состоящие из трехспирального пучка на базе основы одного из IgG-связывающих доменов белка А. Белок А представляет собой поверхностный белок из бактерии Staphylococcus aureus. Данный домен основы состоит из 58 аминокислот, 13 из которых рандомизируют с целью генерации библиотек Affibody® с большим числом вариантов лиганда (см., например, US 5831012). Молекулы Affibody® имитируют антитела, они имеют мол. массу 6 кД сравнительно с молекулярной массой антител, которая составляет 150 кД. Несмотря на маленький размер, центр связывания молекул Affibody® аналогичен центру антитела.
(v) Антикалины - Pieris
Антикалины® представляют собой продукты, разработанные фирмой Pieris ProteoLab AG. Они выделены из липокалинов, широко распространенной группы маленьких и устойчивых белков, которые, как правило, включены в физиологический транспорт или накопление химически чувствительных или нерастворимых соединений. Ряд природных липокалинов находится в тканях или жидкостях тела человека.
Структура белка напоминает иммуноглобулины гипервариабельными петлями в верхней части жесткого скелета. Однако в противоположность антителам или их рекомбинантным фрагментам липокалины состоят из одной полипептидной цепи со 160-180 остатками аминокислот, будучи только незначительно больше чем одни домен иммуноглобулина.
Группа из четырех петель, которая создает связывающий карман, демонстрирует выраженную структурную пластичность и допускает множество боковых цепей. Таким образом центр связывания может быть видоизменен в частном способе для того, чтобы распознавать предназначенные молекулы-мишени различной формы с высокой аффинностью и специфичностью.
Один белок из семейства липокалинов, билин-связывающий белок (ВВР) Pieris Brassicae используют для создания антикалинов путем мутагенизации группы из четырех петель. Одним из примеров патентных заявок, описывающих "антикалины" является РСТ WO 199916873.
(vi) Аффилин - белки Scil
Молекулы Аффилин™ представляют собой маленькие неиммуноглобулиновые белки, которые разрабатывают для получения специфических аффинностей в отношении белков и маленьких молекул. Новые молекулы Аффилин™ можно очень быстро отобрать из двух библиотек, базой каждой из которых является другой человеческий белок основы. Молекулы Аффилин™ не проявляют никакой структурной гомологии с белками иммуноглобулина. В белках Scil используют две основы Аффилин™, одна из который представляет собой γ-кристаллический человеческий структурный белок хрусталика глаза, а другой представляет собой белки из семейства "убиквитина". Обе человеческие основы очень маленькие, проявляют высокую термостабильность и в существенной степени устойчивы к изменениям рН и денатурирующим агентам. Данная высокая стабильность в основном обусловлена расширенной структурой β-складки белков. Примеры γ-кристаллических белков описаны в WO 200104144, и примеры "подобных убиквитину" белков описаны в WO 2004106368.
VI. Терапевтическое применение антител к hTSLPR
Антитела к hTSLPR можно использовать во многих терапевтических или профилактических приложениях посредством ингибирования активностей передачи сигнала TSLP. Они включают лечение заболеваний или состояний, опосредованных передачей сигнала TSLP, таких как воздействие на развитие В-клеток, развитие Т-клеток, реаранжировка гена рецептора Т-клеток или регуляция фактора транскрипции Stat5. Например, антагонистические антитела к hTSLPR можно использовать для подавления или снижения уровня нежелательного иммунного ответа, опосредованного клеткой ТН2. В частности, они подходят для лечения больных людей, страдающих от аллергических воспалительных нарушений, ассоциированных или опосредованных передачей сигнала TSLP. Аллергические воспалительные заболевания, поддающиеся лечению антителами к hTSLPR, представленными в изобретении, включают, например, (1) астму, хроническое воспалительное заболевание дыхательных путей, ассоциированное с затруднением прохождения воздуха и бронхиальной гиперчувствительностью; (2) атопический дерматит, хроническое обостряющееся воспалительное кожное заболевание, требующее длительного периодического лечения, и (3) аллергический ринит, воспалительное нарушение слизистой носа, опосредованное лимфоцитами ТН2, которое связано с атопией. Ожидают, что в США и ряде крупных европейских стран диагностированная частота астмы, атопического дерматита и аллергического ринита повысится с 46 миллионов на настоящий момент до 53 миллионов, с 31,7 миллионов на настоящий момент до 37,2 миллионов и с 55,9 миллионов на настоящий момент до 64,5 миллионов в 2013 г., соответственно. Приблизительно у 50-80% пациентов с атопическим дерматитом имеется или будет развиваться астма или аллергический ринит.
Большинство имеющихся в настоящее время лекарственных препаратов для лечения аллергий направлены на получение симптоматического облегчения, тогда как в области иммуномодуляции, делается относительно мало попыток, вероятно обеспечивающих длительную модификацию заболевания. Антитела к hTSLPR, представленные в изобретении, могут обеспечить новое и эффективное лечение пациентов (особенно больных людей), страдающих от любого из данных аллергических заболеваний. TSLP, предупреждая активацию пути сигнальной трансдукции рецептора TSLP, могут блокировать ответ ТН2 и продукцию цитокинов, ответственных как за инициацию, так и за поддержание аллергического воспаления. Следовательно данный подход имеет потенциальную возможность вызывать длительный терапевтический эффект и благоприятное модифицирующее воздействие на заболевание у пациентов с атопическим дерматитом, астмой и аллергическими ринитом.
В другом варианте осуществления изобретение предусматривает фармацевтическую композицию, включающую по меньшей мере одно из каких-либо вышеуказанных антител или функциональных фрагментов либо консервативных вариантов и их фармацевтически приемлемый носитель или наполнитель.
В ряде вариантов осуществления изобретение представляет способ лечения нарушения или состояния, ассоциированных с присутствием клетки, имеющей hTSLP-мишень рецептора. Способ включает введение нуждающемуся в этом пациенту эффективного количества любой из вышеуказанных композиций. В близком варианте осуществления нарушение или состояние, которые предусматривают лечить, представляет собой респираторное нарушение.
В другом варианте осуществления нарушение или состояние, которые предусматривают лечить, представляет бронхиальную астму, которая является распространенным постоянным воспалительным заболеванием легкого, характеризующимся гиперчувствительностью дыхательных путей (AHR), сверхпродукцией слизи, фиброзом и повышенными уровнями сывороточного IgE.
В другом варианте осуществления нарушение или состояние, которые предусматривают лечить, представляет собой атонический (аллергический) дерматит, который является наиболее распространенным кожным заболеванием у детей и характеризуется сильным зудом и хроническими экзематозные бляшки.
В другом варианте осуществления нарушение или состояние, которые предусматривают лечить, выбрано из других воспалительных или обструктивных заболеваний и состояний дыхательных путей, таких как COPD, острое повреждение легкого (ALI), острый респираторный дистресс-синдром/респираторный дистресс-синдром взрослых (ARDS), одышка, аллергическое воспаление дыхательных путей, заболевание мелких дыхательных путей, карцинома легкого, острый грудной синдром у пациентов с серповидно-клеточным заболеванием и легочная гипертензия, а также обострение гиперактивности дыхательных путей в результате другой лекарственной терапии, в частности, лечения другими ингаляционными лекарственными препаратами.
В другом варианте осуществления нарушение или состояние, которые предусматривают лечить, представляет собой бронхит любого типа или генеза, включая, например, острый, арахидический, катаральный, крупозный, хронический или гнойный туберкулезный бронхит.
В другом варианте осуществления нарушение или состояние, которые предусматривают лечить, включает пневмокониоз (воспалительное, как правило, профессиональное, заболевание легких, часто сопровождающееся закупоркой дыхательных путей, хронической или острой, и вызывается повторяющимся вдыханием пыли) любого типа происхождения, включая, например, алюминоз, антракоз, асбестоз, халикоз, птилоз, сидероз, силикоз, табакоз и биссиноз.
В другом варианте осуществления нарушение или состояние, которые предусматривают лечить, выбрано из атопического ринита (сенной лихорадки) и хронического синусита.
В другом варианте осуществления нарушение или состояние, которые предусматривают лечить, выбрано из других воспалительных состояний кожи, например, псориаза или красной волчанки.
В другом варианте осуществления нарушение или состояние, которые предусматривают лечить, представляет собой воспалительную болезнь кишки, такую как язвенный колит или болезнь Крона.
В другом варианте осуществления нарушение или состояние, которые предусматривают лечить, выбрано из других фибротических состояний, таких как системный склероз, фиброз печени, легочный фиброз, идиопатический легочный фиброз или фиброид легкого.
В другом варианте осуществления нарушение или состояние, которые предусматривают лечить, представляет собой рецидив или метастаз опухоли. Ингибирование цитокинов Th2, как показано, усиливает антивирусные вакцины в моделях на животных и может быть благоприятным при лечении ВИЧ и других инфекционных заболеваний [см. статью Ahlers, J.D., et al. Proc Natl Acad Sci USA, 2002].
В другом варианте осуществления, нарушение или состояние, которые предусматривают лечить, представляет собой респираторную вирусную инфекцию, которая обостряет основные хронические состояния, такие как астма, хронический бронхит, COPD, средний отит и синусит.Респираторная вирусная инфекция, которую лечат, может быть связана с вторичной бактериальной инфекцией, такой как средний отит, синусит или пневмония.
В другом варианте осуществления нарушение или состояние, которые предусматривают лечить, выбрано из других заболеваний или состояний, в частности, заболеваний или состояний, имеющих воспалительный компонент, например, заболеваний костей и связок, включающих ревматоидный артрит, псориатический артрит и другие заболевания, такие как атеросклероз, рассеянный склероз и острое и хроническое отторжение трансплантата, например, после трансплантации сердца, почки, печени, легкого или костного мозга.
В другом варианте осуществления нарушение или состояние, которые предусматривают лечить, представляет собой эндотоксический шок, гломерулонефрит, церебральная и сердечная ишемия, болезнь Альцгеймера, кистозный фиброз, вирусные инфекции и связанные с ними осложнения, синдром приобретенного иммунодефицита (СПИД), рассеянный склероз (MS), гастрит, ассоциированный с Helicobacter pylori, и рак, в частности, рост рака яичников.
В другом варианте осуществления нарушение или состояние, которые предусматривают лечить, представляет собой симптомы, вызываемые вирусной инфекцией у человека, которая вызывается риновирусом человека, другим энтеровирусом, коронавирусом, вирусами герпеса, вирусом гриппа, вирусом парагриппа, респираторным синцитиальным вирусом или аденовирусом.
Лечение в соответствии с настоящим изобретением может быть симптоматическим или профилактическим.
Эффективность агента, представленного в изобретении, в ингибировании воспалительных состояний, например при воспалительных заболеваниях дыхательных путей, можно продемонстрировать на модели на животных, например на модели на мышах, крысах или кроликах, воспаления дыхательных путей или других воспалительных состояний, например, как описано в статьях Wada et al, J. Exp. Med (1994) 180:1135-40; Sekido et al. Nature (1993) 365:654-57; Modelska et al., Am. J. Respir. Crit. Care. Med (1999) 160:1450-56 и Laffon et al (1999) Am. J. Respir. Crit. Care Med. 160:1443-49.
В еще одном варианте осуществления изобретение представляет способ идентификации клетки, имеющей рецептор hTSLP. Данный способ включает контактирование клетки с любым из вышеуказанных антител или фрагментами антител, имеющими, кроме того, определяемую метку. Метка является радиоактивной, флуоресцентной, магнитной, парамагнитной или хемилюминесцентной. Способ, кроме того, может включать любой из вышеописанных вариантов визуализации или выделения меченой клетки.
В другом варианте осуществления любое из вышеуказанных человеческих или гуманизированных антител или фрагментов антител является синтетическим.
В другом варианте осуществления изобретение представляет фармацевтическую композицию и дополнительный лечебный фактор.
Дополнительный лечебный фактор может быть выбран из группы, состоящей из противовоспалительных, бронходилатационных, антигистаминовых или противокашлевых лекарственных субстанций, в частности, при лечении обструктивных или воспалительных заболеваний дыхательных путей, таких как упомянутые выше в данном контексте, например, в качестве потенциатора терапевтической активности данных лекарственных препаратов или как средство снижения требуемой дозировки или возможных побочных эффектов данных лекарственных препаратов. Лечебный фактор, представленный в изобретении, можно смешать в другой лекарственной субстанцией в фиксированной фармацевтической композиции, или его можно вводить отдельно перед, одновременно или после другой лекарственной субстанции. Соответственно, изобретение включает комбинацию фактора, представленного в изобретении, как описано выше в данном контексте, с противовоспалительной, бронходилатационной, антигистаминовой или противокашлевой лекарственной субстанцией, причем указанный фактор, представленный в изобретении, и указанная лекарственная субстанция представляет собой одну и ту же или другую фармацевтическую композицию.
Подходящие противовоспалительные лекарственные препараты включают стероиды, в частности, глюкокортикостероиды, такие как будесонид, бекламетазон дипропионат, флутиказон пропионат, циклесонид или мометазон фуорат, либо стероиды, описанные в WO 02/88167, WO 02/12266, WO 02/100879, WO 02/00679 (особенно описанные в примерах 3, 11, 14, 17, 19, 26, 34, 37, 39, 51, 60, 67, 72, 73, 90, 99 и 101), WO 03/35668, WO 03/48181, WO 03/62259. WO 03/64445, WO 03/72592, WO 04/39827 и WO 04/66920; нестероидные агонисты глюкокортикоидного рецептора, такие как описаны в DE 10261874, WO 00/00531, WO 02/10143, WO 03/82280, WO 03/82787, WO 03/86294, WO 03/104195, WO 03/101932, WO 04/05229, WO 04/18429, WO 04/19935 и WO 04/26248; антагонисты LTB4, такие как BIIL 284, СР-195543, DPC11870, LTB4 этаноламид, LY 293111, LY 255283, CGS025019C, СР-195543, ONO-4057, SB 209247, SC-53228 и описанные в US 5451700; антагонисты LTD4, которые включают монтелукаст, пранлукаст, зафирлукаст, акколат, SR2640, Wy-48,252, ICI 198615, МК-571, LY-171883, Ro 24-5913 и L-648051; ингибиторы PDE4, такие как силомиласт (Ariflo® GlaxoSmithKline), рофлумиласт (Byk Gulden),V-11294А (Napp), BAY19-8004 (Bayer), SCH-351591 (Schering-Plough), арофиллин (Almirall Prodesfarma), PD189659/PD168787 (Parke-Davis), AWD-12-281 (Asta Medica), CDC-801 (Celgene), SelCID(TM) CC-10004 (Celgene), VM554/UM565 (Vernalis), T-440 (Tanabe), KW-4490 (Kyowa Hakko Kogyo) и раскрытые в WO 92/19594, WO 93/19749, WO 93/19750, WO 93/19751, WO 98/18796, WO 99/16766, WO 01/13953, WO 03/104204, WO 03/104205, WO 03/39544, WO 04/000814, WO 04/000839, WO 04/005258, WO 04/018450, WO 04/018451, WO 04/018457, WO 04/018465, WO 04/018431, WO 04/018449, WO 04/018450, WO 04/018451, WO 04/018457, WO 04/018465, WO 04/019944, WO 04/019945, WO 04/045607 и WO 04/037805; агонисты А2А, такие как описанные в ЕР 1052264, ЕР 1241176, ЕР 409595А2, WO 94/17090, WO 96/02543, WO 96/02553, WO 98/28319, WO 99/24449, WO 99/24450, WO 99/24451, WO 99/38877, WO 99/41267, WO 99/67263. WO 99/67264, WO 99/67265, WO 99/67266, WO 00/23457, WO 00/77018, WO 00/78774, WO 01/23399, WO 01/27130, WO 01/27131, WO 01/60835, WO 01/94368, WO 02/00676, WO 02/22630, WO 02/96462 и WO 03/086408; а антагонисты А2 В, такие как описаны в WO 02/42298.
Подходящие лекарственные препараты бронходилататоров включают антихолинергические или антимускариновые агенты, в частности, ипратропий бромид, окситропий бромид, соли тиотропия и CHF 4226 (Chiesi) и гликопирролат, а также описанные в ЕР 424021, US 3714357, US 5171744, WO 01/04118, WO 02/00652, WO 02/51841, WO 02/53564, WO 03/00840, WO 03/33495, WO 03/53966, WO 03/87094, WO 04/018422 и WO 04/05285; и агонисты β-2 адреноцептора, такие как альбутерол (сальбутамол), метапротеренол, тербуталин, сальметерол, фенотерол, прокатерол и, особенно, формотерол, кармотерол и их фармацевтически приемлемые соли и соединения (в свободной или солевой либо сольватной форме) формулы I, приведенной в документе WO 00/75114, который включен в данном контексте в виде ссылки, предпочтительно соединения из представленных в нем примеров, особенно соединение (5-[(R)-2-(5,б-диэтил-индан-2-иламино)-1-гидрокси-этил]-8-гидрокси-1Н-хинолин-2-он), и их фармацевтически приемлемые соли, а также соединения (в свободной или солевой либо сольватной форме) формулы I, приведенной в WO 04/16601, и, кроме того, соединения, предложенные в ЕР 1440966, JP 05025045, WO 93/18007, WO 99/64035, US 2002/0055651, WO 01/42193, WO 01/83462, WO 02/66422, WO 02/ 70490, WO 02/76933, WO 03/24439, WO 03/42160, WO 03/42164, WO 03/72539, WO 03/91204, WO 03/99764, WO 04/16578, WO 04/22547, WO 04/32921, WO 04/33412, WO 04/37768, WO 04/37773, WO 04/37807, WO 04/39762, WO 04/39766, WO 04/45618 WO 04/46083, WO 04/80964, EP 1460064, WO 04/087142, WO 04/089892, ЕР 01477167, US 2004/0242622, US 2004/0229904, WO 04/108675, WO 04/108676, WO 05/033121, WO 05/040103 и WO 05/044787.
Подходящие противовоспалительные лекарственные препараты и бронходилататоры включают двойные агонисты β-2 адреноцептор/мускариновые антагонисты, такие как описаны в US 2004/0167167, WO 04/74246 и WO 04/74812.
Подходящие антигистаминовые лекарственные субстанции включают цетиризин гидрохлорид, ацетаминофен, клемастин фумарат, прометазин, лоратидин, деслоратидин, дифенгидрамин и фексофенадин гидрохлорид, активастин, астемизол, азеластин, эбастин, эпинастин, мизоластин и тефенадин, а также описанные в JP 2004107299, WO 03/099807 и WO 04/026841.
Можно также использовать комбинации лечебных факторов, представленных в изобретении, и антихолинергических или антимускариновых агентов, стероидов, β-2-агонистов, ингибиторов PDE4, агонистов допаминового рецептора, антагонистов LTD4 или антагонистов LTB4. Другие эффективные комбинации агентов, представленных в изобретении, с противовоспалительными лекарственными препаратами, представляют собой комбинации с другими антагонистами хемокиновых рецепторов, например, CCR-1, CCR-3, CCR-4, CCR-5, CCR-6, CCR-7, CCR-8, CCR-9 and CCR10, CXCR1, CXCR2, CXCR3, CXCR4, CXCR5, особенно антагонистами CCR-5, такими как антагонисты Schering-Plough SC-351125, SCH-55700 и SCH-D, антагонистами Takeda, такими как N-[[4-[[[6,7-дигидро-2-(4-метилфенил)-5Н-бензоциклогептен-8-ил]карбонил]амино]фенил]-метил]-тетрагидро-N,N-диметил-2Н-пиран-4-аминий хлорид (ТАК-770), антагонистами CCR-5, описанными в US 6166037 (особенно в пунктах формулы 18 и 19), WO 0066558 (особенно пункт формулы 8), WO 0066559 (особенно пункт формулы 9), WO 04/018425 и WO 04/026873.
Дополнительный лечебный фактор может быть также выбран из группы, состоящей из других цитокин-связывающих молекул, особенно антител к другим цитокинам, в частности, с антителом к ИЛ-4, таким как описано в РСТ/ЕР2005/00836, антителом к IgE, таким как Xolair®, антителом к ИЛ-31, антителом к ИЛ-31Р, антителом к ИЛ-13, таким как описано в WO 05/007699, антителом к эндо глину, антителом к ИЛ-1b, антителом к TSLP или другим антителом к hTSLPR.
Антагонистические антитела к hTSLPR, представленные в изобретении, можно использовать для лечения пациента как терапевтически, так и профилактически. В терапевтических приложениях композицию, включающую антагонистическое антитело к hTSLPR (например, гуманизированное антитело к hTSLPR), вводят пациенту, уже пораженному аллергическим заболеванием, вызванным или ассоциированным с передачей сигнала TSLP. Композиция включает антитело в количестве, достаточном для излечения, в частности, задержки или определяемого замедления развития состояния и его осложнений. В профилактических приложениях композиции, включающие моноклональные антитела к hTSLPR, вводят пациенту, еще не страдающему от аллергического воспалительного нарушения. Они скорее направлены на пациентов, которые находятся в группе риска или имеют предрасположенность к развитию аллергического воспалительного нарушения. Данные приложения позволяют повысить устойчивость больного или замедлить развитие аллергического воспалительного нарушения, опосредованного передачей сигнала TSLP.
VII. Фармацевтические композиции
Изобретение представляет фармацевтические композиции, включающие моноклональные антитела к hTSLPR (интактные или связывающие фрагменты), изготовленные вместе с фармацевтически приемлемым носителем. Композиции могут дополнительно включать другие лечебные факторы, которые подходят для лечения или предупреждения данного аллергического нарушения, например, известные вышеуказанные противоаллергические агенты. Фармацевтически приемлемые носители усиливают или стабилизируют композицию или облегчают получение композиции. Фармацевтически приемлемые носители включают растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые агенты, изотонические и замедляющие всасывание агенты и подобные компоненты, которые физиологически совместимы.
Фармацевтическую композицию, представленную в настоящем изобретении, можно ввести рядом методов, известных в области техники. Путь и/или способ введения варьирует в зависимости от требуемых результатов. Предпочтительно, чтобы введение было внутривенным, внутримышечным, внутрибрюшинным или подкожным либо введением, проксимальным области мишени. Фармацевтически приемлемый носитель должен подходить для внутривенного, внутримышечного, подкожного, парентерального, спинального или эпидермального введения (например, путем инъекции или инфузии). В зависимости от пути введения активное соединение, т.е. антитело, биспецифическая или мультиспецифическая молекула, могут быть покрытыми материалом для защиты соединения от действия кислот и других естественных условий, которые могут инактивировать соединение.
Композиция должна быть стерильной и жидкой. Надлежащая текучесть может поддерживаться, например, при использовании покрытия, такого как лецитин, путем поддержания требуемого размера частиц в случае дисперсии и путем применения поверхностно-активных веществ. Во многих случаях предпочтительно включение в композицию изотонических агентов, например, Сахаров, полиспиртов, таких как манит или сорбит, и хлорид натрия. Длительное всасывание инъекционных композиций можно осуществить путем включения в композицию агента, который замедляет всасывание, например, моностеарата алюминия или желатина.
Фармацевтические композиции, представленные в изобретении, можно получить в соответствии с методами, хорошо известными и рутинно практически используемыми в области техники. См., например. Remington: The Science and Practice of Pharmacy, Mack Publishing Co., 20 изд, 2000 и Sustained and Controlled Release Drug Delivery Systems, под ред. J.R.Robinson, Marcel Dekker, Inc., New York, 1978. Предпочтительно, когда фармацевтические композиции производят в условиях GMP. Как правило, в фармацевтических композициях, представленных в изобретении, используют терапевтически эффективную дозу или эффективную дозу антитела к hTSLPR. Антитела к hTSLPR получают в фармацевтически приемлемых лекарственных формах принятыми методами, известными компетентным специалистам в области техники. Схемы дозирования регулируют, чтобы получить оптимальный требуемый ответ (например, терапевтический ответ). Например, можно ввести один болюс, можно ввести несколько разделенных доз в течение времени или дозу можно пропорционально уменьшить или увеличить в соответствии с требованиями терапевтической ситуации. Особенно эффективно получение парентеральных композиций в унифицированной лекарственной форме для легкости введения и унификации дозы. Унифицированная лекарственная форма, как используют в данном контексте, относится к физически дискретным единицам, подходящим в качестве единичных дозировок для пациентов, которых предполагают лечить; каждая единица включает предварительное установленное количество активного соединения, рассчитанное так, чтобы получить требуемый терапевтический эффект, в сочетании с необходимым фармацевтическим носителем.
Фактические уровни доз активных ингредиентов в фармацевтических композициях, представленных в настоящем изобретении, могут варьировать так, чтобы получить количество активного ингредиента, которое эффективно для достижения требуемого терапевтического ответа для конкретного пациента, композиции и способа введения без токсического действия на пациента. Выбранный уровень дозы зависит от множества фармакокинетических факторов, включая активность используемых конкретных композиций, представленных в настоящем изобретении, или их сложного эфира, соли или амида, пути введения, времени введения, скорости выведения конкретного используемого соединения, продолжительности лечения, других лекарственных препаратов, соединений и/или материалов, используемых в комбинации с конкретными используемыми композициями, возраста, пола, массы тела, состояния, общего состояния здоровья и предшествующей истории болезни пациента, которого лечат, и подобных факторов.
Лечащий врач или ветеринар могут начать с доз представленных в изобретении антител, используемых в фармацевтической композиции на уровнях ниже необходимых для достижения требуемого терапевтического эффекта, и постепенно повышать дозу, пока не будет достигнут требуемый эффект. Как правило, эффективные дозы представленных в настоящем изобретении композиций, предназначенных для лечения аллергического воспалительного нарушения, описанного в данном контексте, варьируют в зависимости от многих факторов, включая средства введения, размер мишени, физиологическое состояние пациента, является ли пациент человеком или животным, других введенных лекарственных средств и того, является ли лечение профилактическим или терапевтическим. Лечебные дозы необходимо оттитровать, чтобы оптимизировать безопасность и эффективность. Для введения антитела доза лежит в интервале от приблизительно 0,0001 до 100 мг/кг и чаще 0,01-5 мг/кг массы тела хозяина. Например, дозы могут составлять 1 мг/кг массы тела или 10 мг/кг массы тела либо лежать в интервале 1-10 мг/кг. Иллюстративная схема лечения предусматривает введение один раз каждые две недели или один раз в месяц либо один раз каждые 3-6 месяцев.
Антитело обычно вводят во многих случаях. Интервалы между однократными дозами могут быть недельными, месячными или годичными. Интервалы могут быть также неравномерными, как показано измерением уровней антитела к hTSLPR в крови пациента. В ряде способов дозу регулируют так, чтобы достигнуть концентрацию антитела в плазме 1-1000 мкг/мл и в ряде способов 25-300 мкг/мл. Альтернативно антитело можно ввести в виде препарата с замедленным высвобождением, в таком случае требуется менее частое введение. Доза и частота варьируют в зависимости от полупериода существования антитела в организме пациента. Как правило, гуманизированные антитела демонстрируют более длительный полупериод существования, чем у химерных антител и нечеловеческих антител. Доза и частота введения могут варьировать в зависимости от того, является ли лечение профилактическим или терапевтическим. В профилактических приложениях вводят относительно низкую дозу через относительно нечастые интервалы в течение длительного периода времени. Некоторые пациенты продолжают получать лечение в течение последующей жизни. В терапевтических приложениях иногда требуется относительно высокая доза через относительно короткие интервалы до тех пор, пока развитие заболевания не уменьшится или не закончится, и, предпочтительно, пока у пациента проявляется частичное или полное облечение симптомов заболевания. После этого пациента можно лечить по профилактической схеме.
Примеры
Следующие примеры представлены для дальнейшей иллюстрации изобретения, но не для ограничения его объема. Другие варианты изобретения будут легко очевидны обычному специалисту в области техники и охватываются прилагаемой формулой изобретения.
Пример 1. Создание мышиного антагонистического антитела к hTSLPR
Данный пример описывает создание мышиного антагонистического антитела к hTSLPR. Мышь Вс12 Wehi22 mouse (№1770) иммунизируют слитым белком человеческий TSLPR (Met 1 - Lys 231)/человеческий Fc IgGI (Pro 100 - Lys 330), приобретаемым в фирме R&D System (Minneapolis, MN) (No. no каталогу 981-TR, партия No. EDY1312A). Иммунизацию проводят в целом 8 инъекциями в течение 18 дней. В-клетки выделяют из периферических лимфатических узлов и сливают с клетками миеломы FO (ATCC, Manassas, VA). Проводят скрининг продукции антитела с помощью ELISA в отношении белка hTSLPR/Fc. Получают всего 24 клона, которые продуцируют антитела, специфически распознающие hTSLPR. Связывание антител hTSLPR с клеточным поверхностным TSLPR анализируют с помощью FACS (сортинга клеток по возбужденной флуоресценции) (BD Biosciences, San Jose, CA). Вкратце, клетки BaF3/hTSLPR/hIL7Rα инкубируют с супернатантами культуры гибридомы, а затем с FITC-конъюгированным IgG к мыши и анализируют на приборе для FACS. Результаты показывают, что 14 клонов гибридомы из 24 клонов способны к связыванию клеточного поверхностного рецептора.
Затем hTSLP-зависимый анализ пролиферации клеток на клетках BaF3/hTSLPR/hIL7Rα используют для скрининга антагонистических антител hTSLPR. Клетки экспрессируют как человеческий TSLPR, так и IL7Rα и способны к ответу на стимуляцию человеческим TSLP (см. статью Reche et al., J. ImmunoL, 167:336-43, 2001). Анализ пролиферации клеток проводят в основном, как описано в статье Reche et al. Вкратце, клетки BaF3/hTSLPR/hIL7Rα предварительно инкубируют с антителами, выделенными из культуральных супернатантов, в течение 1 часа и индуцируют с помощью hTSLP в указанных концентрациях, как показано на фиг.1. Пролиферацию клеток измеряют, используя краситель Alamar Blue (TREK Diagnositc Systems, Cleveland, ОН). Для каждого родительского клона в анализе используют один из трех субклонов. Результаты, как показано на фиг.1, показывают, что три суб клона из клона 1D6 демонстрируют сильную антагонистическую активность.
Очищают моноклональное антитело (mAb) из одного из субклонов, 1D6.C9 (см. фиг.2С). Вкратце, бессывороточную кондиционированную среду Miniperm culture (Vivascience, Carlsbad, CA) обрабатывают Cleanascite (LigoChem, Fairfield, NJ). Затем антитело очищают на смоле с белком G (Amersham Biosciences, Piscataway, NJ). Очищенное антитело тестируют в анализе пролиферации клеток, как описано выше. Клетки Ba/F3-hTSLPR-hIL7Rα или клетки Baf3/hTSLPR/hIL7Rα/Stat5-Luc обрабатывают различными концентрациями антитела в течение 1 часа перед добавлением 1 нг/мл hTSLP. Результаты показывают, что TSLPR-зависимая пролиферация как клеток BaF3/hTSLPR/hIL7Rα (см. фиг.3А), так и клеток Baf3/hTSLPR/hIL7Rα/Stat5-Luc (см. фиг.3В) ингибируется добавлением мышиного антитела к hTSLPR дозозависимым образом.
Кроме того, анализ сигнала люциферазы используют также для исследования эффекта антитела на активности TSLP-опосредованной передачи сигнала. TSLP индуцирует фосфорилирование Stat3 и Stat5 в клетках Baf3/hTSLPR/hIL7Rα (см. статью Reche et al., выше). Данный анализ разработан для измерения экспрессии сигнального гена под контролем Stat5 в клетках Baf3/hTSLPR/hIL7Rα/Stat5-Luc в ответ на передачу сигнала TSLP. Вкратце, сигнальную конструкцию Stat-Luc генерируют инсерцией консенсусного сайта связывания Stat в вектор основного сигнала Luc pGL2 (Promega, Madison, WI), который не содержит промотор и энхансер. Введенный инсерцией сайт связывания Stat включает 8 копий элемента последовательности GATTTCCCCGAAATG (SEQ ID NO: 15), который несет последовательность связывания ядра Stat TTCCCGGAA (SEQ ID NO: 16) (см. статьи Yan et al., Cell. 84:421-30, 1996 и Saylors et al., Gene Ther. 6:944-6, 1999). Сигнальную конструкцию Stat-Luc интродуцируют в клетки BaF3/hTSLPR/hIL7Rα. Полученные в результате клетки BaF3/hTSLPR/hIL7Rα/Stat5-Luc обрабатывают различными концентрациями мышиного антитела в течение 1 часа перед добавлением 1 нг/мл hTSLP. Активность люциферазы измеряют, используя Bright Glo (Promega, Madison, WI). Результаты, полученные при анализе, показаны на фиг.3С. Аналогично результатам анализа пролиферации клеток данные анализа сигнала Stat5-Luc также демонстрируют дозозависимую антагонистическую активность мышиного антитела к hTSLPR.
Вариабельные области тяжелой цепи (VH) и легкой цепи (VL) mAb 1D6.C9 клонируют с помощью ОТ-ПЦР (полимеразной цепной реакции с участием обратной транскриптазы). Последовательности праймеров, используемых для идентификации вариабельных областей тяжелой и легкой цепей, являются следующими. Праймерами для VH являются: (1) VH9:
GATGGCAGCWGCYCAAAG (SEQ ID NO: 1) и (2) Н-константная:
GCGTCTAGAAYCTCCACACACAGGRRCCAGTGGATAGAC (SEQ ID NO: 2).
Праймерами для VL являются: (1)
LCV3:GGGTCTAGACACCATGGAGWCACAKWCTCAGGTCTTTRTA (SEQ ID NO: 3) и L-константная: СССТСТАСААСТССАТССТСССААСАТСС (SEQ ID NO: 4).
Вкратце, общую РНК выделяют из клона 1D6.C9. ОТ-ПЦР проводят с прямыми Праймерами к сигнальной последовательности вариабельных областей либо тяжелой, либо легкой цепей, и с обратными Праймерами в области СН1 тяжелой цепи или κ-константной области легкой цепи. ПЦР-продукты клонируют в вектор pCR II или pcDNA3.1/V5-His-TPOP-TA для секвенирования. Затем определяют полинуклеотидные последовательности последовательностей вариабельной области тяжелой цепи и легкой цепи данного клона антитела (см. фиг.4). Соответствующие последовательности аминокислот вариабельных областей (SEQ ID NO: 5 и SEQ ID NO: 6) показаны на фиг.5. На фиг.5 также показаны участки CDR и скелетные области рассчитывают в соответствии с системой нумерации Kabat et al., см. выше.
Пример 2. Генерация химерных антител к hTSLPA
Данный пример описывает генерацию и характеризацию химерных антител к hTSLPR. Химерные антитела включают вариабельные области из клона вышеуказанного мышиного антитела к hTSLPR 1D6.C9 и константные области из человеческих иммуноглобулинов.
Для генерации химерных антител вариабельные области клона мышиного антитела к человеческому TSLPR 1D6.C9 получают, используя технологии ПЦР и праймерные последовательности, описанные в примере 1, созданные для клонирования в кассетные векторы. Затем ПЦР-продукты, соответственно, клонируют в кассетные векторы, которые включают слияния в рамке считывания с лидерными последовательностями человеческого иммуноглобулина, J-сегментами и сплайсер-донорными сигналами с использованием последовательностей, показанных в табл.1. Затем последовательности переносят в экспрессирующие векторы млекопитающих, включающие человеческую константную область иммуноглобулина, например, векторы SP2/0.
Отбор клонов, экспрессирующих тяжелую цепь, основан на селекции на фоне неомицина, тогда как клоны, экспрессирующие легкую цепь, отбирают, используя селекцию на фоне dhfr. Амплификацию генов инициируют с использованием метотрексата. Кассету вариабельной области для тяжелой цепи переносят в векторы млекопитающих, экспрессирующие человеческие IgG1 и IgG4, соответственно.
После субклонирования для получения химерных антител IgG1 ДНК-плазмиды тяжелой цепи и κ-легкой цепи химерного IgG1 совместно электропорируют в клетки миеломы SP2/0. Аналогично для получения химерных антител IgG4 тяжелую цепь и κ-легкую цепь химерного IgG4 совместно электропорируют в клетки миеломы SP2/0, Клетки обирают на фоне генетицина и затем культивируют и размножают в среде для роста, включающей генетицин и метотрексат. Клетки адаптируют в бессывороточной среде для очистки антитела. Химерные антитела IgG1 и IgG4, секретируемые из трансфицированных клеток SP2/0, очищают (см. фиг.2А и фиг.2В). Антагонистическую активность очищенных химерных антител IgGI и IgG4 сравнивают с активностью мышиного антитело в анализе сигнала Luc. Химерное антитело IgG1 к рецептору гибели 5 (DR5) используют в качестве отрицательного контроля. Измеренные по экспрессии сигнального гена в клетках Ba/F3, экспрессирующих hTSLPR, hIL7Rα и Stat5-Luc, результаты показывают, что химерные антитела к hTSLPR проявляют антагонистическую активность, близкую активности мышиного антитела IgG1 (см. фиг.6).
Пример 3. Антитело к hTSLPR ингибирует TSLP-опосредованную секрецию TARC
Данный пример описывает ингибирование TSLP-опосредованной секреции TARC из человеческих моноцитов с помощью антагонистических антител к hTSLPR. Оба, мышиное и химерное антитела к hTSLPR, упомянутые выше, исследуют на эффект TSLP-опосредованной секреции TH2-привлекающего и регулирующего активацию хемокина вилочковой железы (TARC) человеческими моноцитами. За исключением добавления антитела к hTSLPR анализы секреции TARC проводят в основном так, как описано в статье Soumelis et al., Nat Immunol. 3:673-80, 2002. Результаты показаны на фиг.7. Две верхние панели на фигуре, соответственно, показывают секрецию TARC из человеческих моноцитов и дендритных клеток CD11+, стимулируемых TSLP. Три нижние панели, соответственно, показывают эффект трех различных антител к hTSLPR на TSLP-опосредованную секрецию TARC из человеческих моноцитов. Как демонстрируют на фигуре, все, мышиное антитело к hTSLPR и химерные антитела к hTSLPR (оба изотипа IgG1 и IgG4), способны ингибировать секрецию TARC из человеческих моноцитов дозозависимым образом. Это еще более подтверждает антагонистическую активность антитела к hTSLPR в отношении передачи сигнала TSLP.
Исследования связывания с целью определить аффинности связывания антител к hTSLPR проводят с использованием стандартного анализа плазменного резонанса Biacore с целью измерить скорости "на" (ka) и "из" (kd). Данные измерения приводят к вычислению константы связывания (KD) для антитела. Дальнейший анализ также проводят с использованием ForteBiop для исследования кинетики связывания в растворе. Кроме того, клеточный анализ используют для определения IC50 для антитела. Результаты данных исследований суммируют в табл.3.
Последовательности антитела Последовательность NV115-3В

Хотя предшествующее изобретение описано достаточно детально путем иллюстрации и примера с целью ясности понимания, обычному специалисту в области техники будет легко увидеть в свете описания данного изобретения, что в нем можно сделать ряд изменений и модификаций, не выходя из сущности или объема прилагаемой формулы изобретения.
Все публикации, базы данных, последовательности Genbank, патенты и патентные заявки, приведенные в данном описании, включены в данном контексте в виде ссылки так, как если бы было указано, что они специально и отдельно включены в виде ссылки.
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИ-TSLP АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2834227C1 |
| Антитела к TSLP человека и их применение | 2021 |
|
RU2825460C1 |
| АНТИТЕЛО, СПОСОБНОЕ СВЯЗЫВАТЬСЯ С ТИМИЧЕСКИМ СТРОМАЛЬНЫМ ЛИМФОПОЭТИНОМ, И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2825304C2 |
| НОВОЕ АНТИТЕЛО К TSLP-РЕЦЕПТОРУ ЧЕЛОВЕКА | 2014 |
|
RU2689528C2 |
| АНТИТЕЛА, НАПРАВЛЕННЫЕ НА CD127 | 2015 |
|
RU2734076C2 |
| МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕСЯ С ТИМУСНЫМ СТРОМАЛЬНЫМ ЛИМФОПОЭТИНОМ (TSLP), И СПОСОБЫ ПРИМЕНЕНИЯ ТАКИХ МОЛЕКУЛ | 2016 |
|
RU2731644C2 |
| АНТИТЕЛА И ПОЛИПЕПТИДЫ, НАПРАВЛЕННЫЕ ПРОТИВ CD127 | 2017 |
|
RU2769352C2 |
| АНТИТЕЛО К CLDN18.2 И ЕГО ПРИМЕНЕНИЯ | 2018 |
|
RU2797709C2 |
| СКОНСТРУИРОВАННОЕ АНТИТЕЛО ПРОТИВ TSLP | 2010 |
|
RU2575039C2 |
| АНТИТЕЛА, СПЕЦИФИЧНЫЕ К CD22 ЧЕЛОВЕКА, И ИХ ПРИМЕНЕНИЯ В ТЕРАПИИ И ДИАГНОСТИКЕ | 2003 |
|
RU2342401C2 |



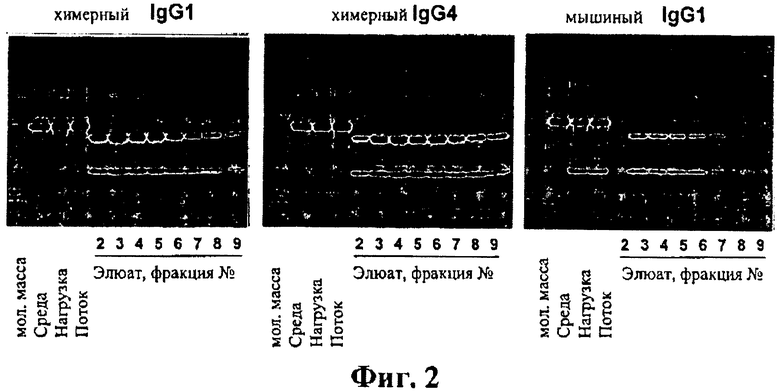
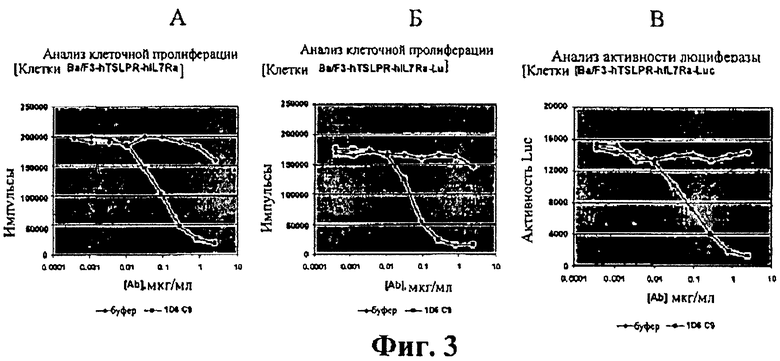


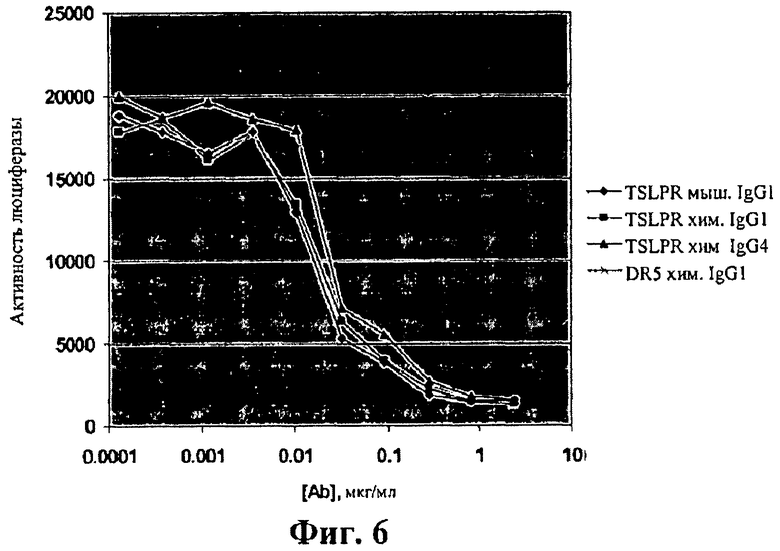
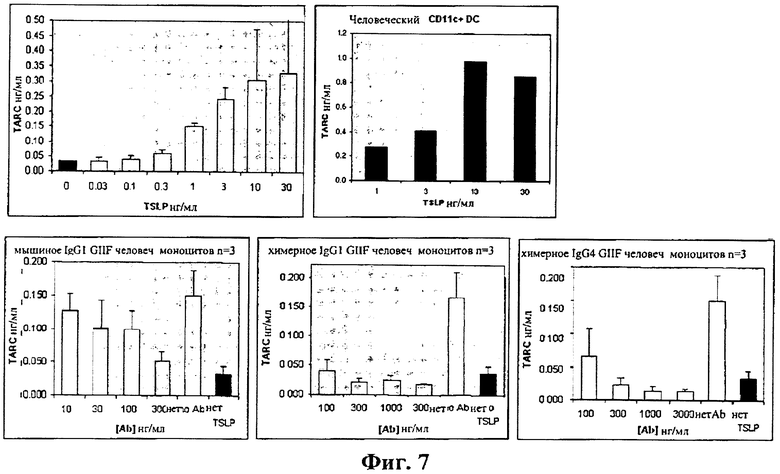
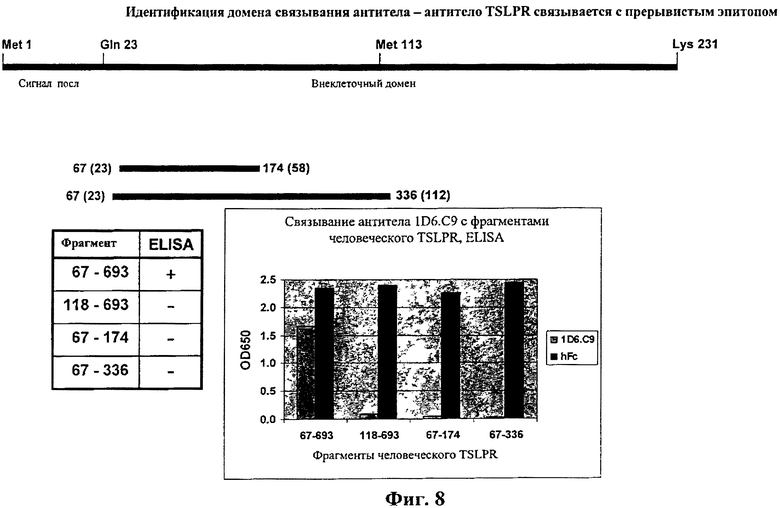
Предложенное изобретение относится к области иммунологии. Предложены варианты антитела к рецептору тимусного стромального лимфопоэтина (hTSLP) человека, характеризующиеся наличием вариабельных областей легкой и тяжелой цепи или 3 CDR легкой цепи и 3 CDR тяжелой цепи соответственно. Описаны варианты выделенного или рекомбинантного кодирующего полинуклеотида, выделенная клетка-хозяина для получения антитела, содержащая кодирующие ДНК. Описан способ лечения воспалительного нарушения, опосредованного hTSLP, включающий введение фармацевтической композиции, включающей эффективное количество антитела. Использование изобретения обеспечивает новые антитела против hTSLP человека, что может найти применение в медицине для лечения воспалительных заболеваний, связанных с активностью hTSLP. 6 н. и 21 з.п. ф-лы, 8 ил., 2 табл., 3 пр.
1. Выделенное антитело к рецептору тимусного стромального лимфопоэтина человека или его антигенсвязывающая часть, включающее последовательность вариабельной области тяжелой цепи аминокислот 10-134 SEQ ID NO: 5 или функционально активную последовательность аминокислот, которая по меньшей мере на 80% идентична аминокислотам 1-134 SEQ ID NO: 5, и последовательность вариабельной области легкой цепи аминокислот 21-128 SEQ ID NO: 6 или функционально активную последовательность аминокислот, которая по меньшей мере на 80% идентична аминокислотам 21-128 SEQ ID NO: 6.
2. Выделенное антитело к рецептору тимусного стромального лимфопоэтина человека или его антигенсвязывающая часть, включающее последовательности CDR1, CDR2 и CDR3 тяжелой цепи TYGMS (SEQ ID NO: 7), WINTYSGVPRYADDFKG (SEQ ID NO: 8) и EGFITTVVGAAGRFVY (SEQ ID NO: 9) соответственно и последовательности CDR1, CDR2 и CDR3 легкой цепи KASQDVGTAVA (SEQ ID NO: 10), WASTRHT (SEQ ID NO: 11) и QQYSTYPT (SEQ ID NO: 12) соответственно.
3. Антитело по п.2, которое представляет собой мышиное антитело.
4. Антитело по п.2, которое представляет собой химерное антитело.
5. Антитело по п.1, которое включает константную область тяжелой цепи человека и константную область легкой цепи человека.
6. Антитело по п.2, которое представляет собой гуманизированное антитело.
7. Антитело по п.1, которое представляет собой человеческое антитело.
8. Антитело по п.2, которое представляет собой одноцепочечное антитело.
9. Антитело по п.2, которое представляет собой фрагмент Fab.
10. Антитело по п.2, которое представляет собой изотип IgG1 или IgG4.
11. Выделенный или рекомбинантный полинуклеотид, который кодирует полипептид вариабельной области тяжелой цепи или легкой цепи антитела по п.1, специфично связывающей hTSLPR.
12. Полинуклеотид по п.11, где антитело представляет собой человеческое антитело.
13. Выделенный или рекомбинантный полинуклеотид, причем полинуклеотид, кодирующий антитело по п.2.
14. Полинуклеотид по п.13, причем антитело включает зрелую последовательность вариабельной области тяжелой цепи, которая идентична аминокислотам 10-134 SEQ ID NO: 5, и зрелую последовательность вариабельной области легкой цепи, которая идентична аминокислотам 21-128 SEQ ID NO: 6.
15. Полинуклеотид по п.13, который представляет собой ДНК.
16. Выделенная клетка-хозяин, предназначенная для получения антитела по п.1 или 2 и включающая (1) сегмент рекомбинантной ДНК, кодирующий тяжелую цепь антитела по п.2, и (2) второй сегмент рекомбинантной ДНК, кодирующей легкую цепь антитела; причем указанные сегменты ДНК соответственно функционально связаны с первым и вторым промотором и способны экспрессироваться в указанной клетке-хозяине.
17. Клетка-хозяин по п.16, причем моноклональное антитело представляет собой гуманизированное антитело.
18. Клетка-хозяин по п.16, причем моноклональное антитело включает последовательности CDR1, CDR2 и CDR3 тяжелой цепи TYGMS (SEQ ID NO: 7), WINTYSGVPRYADDFKG (SEQ ID NO: 8) и EGFITTVVGAAGRFVY (SEQ ID NO: 9) соответственно и последовательности CDR1, CDR2 и CDR3 легкой цепи KASQDVGTAVA (SEQ ID NO: 10), WASTRHT (SEQ ID NO: 11) и QQYSTYPT (SEQ ID NO: 12) соответственно.
19. Клетка-хозяин по п.16, причем моноклональное антитело включает зрелую последовательность вариабельной области тяжелой цепи, которая идентична аминокислотам 10-134 SEQ ID NO: 5, и зрелую последовательность вариабельной области легкой цепи, которая идентична аминокислотам 21-128 SEQ ID NO: 6.
20. Клетка-хозяин по п.16, которая представляет собой клеточную линию млекопитающего, отличного от человека.
21. Способ лечения воспалительного нарушения у пациента, включающий введение пациенту фармацевтической композиции, включающей эффективное количество антитела по п.2.
22. Способ по п.21, в котором антитело представляет собой гуманизированное антитело.
23. Способ по п.21, в котором антитело включает последовательности CDR1, CDR2 и CDR3 тяжелой цепи TYGMS (SEQ ID NO: 7), WINTYSGVPRYADDFKG (SEQ ID NO: 8) и EGFITTVVGAAGRFVY (SEQ ID NO: 9) соответственно и последовательности CDR1, CDR2 и CDR3 легкой цепи KASQDVGTAVA (SEQ ID NO: 10), WASTRHT (SEQ ID NO: 11) и QQYSTYPT (SEQ ID NO: 12).
24. Способ по п.21, в котором антитело включает (i) зрелую последовательность вариабельной области тяжелой цепи, которая идентична зрелому участку SEQ ID NO: 5, и (ii) зрелую последовательность вариабельной области легкой цепи, которая идентична зрелому участку SEQ ID NO: 6.
25. Способ по п.21, в котором пациент является человеком.
26. Способ по п.21, в котором пациент страдает от аллергического воспалительного заболевания.
27. Способ по п.26, в котором аллергическое воспалительное заболевание представляет собой атопический дерматит, астму или аллергический ринит.
| WO 2000029581 А1, 25.05.2000 | |||
| US 2005249712 A1, 10.11.2005 | |||
| SOUMELIS et al., "Human epithelial cells trigger dendritic cell mediated allergic inflammation by producing TSLP" NATURE IMMUNOLOGY, vol.3, no.7, 2002, с.673-680 | |||
| РОЙТ А | |||
| и др | |||
| Иммунология, Пер | |||
| с англ | |||
| - М.: Мир, 2000, с.110-111, 150-151. |

