СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[000] Настоящая заявка содержит список последовательностей, представленный в электронном виде в формате ASCII, который полностью включен в данный документ посредством ссылки. Указанная копия ASCII, созданная 23 августа 2016 года, называется PAT057035-WO-PCT_SL.txt и имеет размер 46696 байт.
ОБЛАСТЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
[001] Настоящее изобретение относится к молекулам, например, к антителам или фрагментам антител, способным специфически связываться с тимусным стромальным лимфопоэтином (TSLP), к композициям, содержащим такие молекулы, и к способам применения и получения таких молекул.
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
[002] Тимусный стромальный лимфопоэтин (TSLP) представляет собой цитокин, который передает сигнал через гетеродимерный рецептор, состоящий из субъединицы IL-7Rα и TSLP-R, уникального компонента, гомологичного общей рецепторной γ-цепи (Pandey et al., Nat. Immunol. 2000, 1(1):59-64). TSLP экспрессируется эпителиальными клетками в тимусе, легких, коже, кишечнике и миндалинах, а также клетками гладкой мускулатуры дыхательных путей, фибробластами легких и стромальными клетками (Edwards, 2008, Drug news & perspectives 21, 312-316; Geha, 2010, Annals of New York Academy of Sciences 1183, 13-24; Reche et al., 2001, Journal of immunology 167, 336-343). Эти клетки продуцируют TSLP в ответ на провоспалительные стимулы, а TSLP стимулирует аллергические воспалительные реакции путем воздействия на ряд клеток врожденной иммунной системы, включающих в себя дендритные клетки (Soumelis et al., 2002, Nature immunology 3, 673-680), моноциты (Reche et al., 2001, Journal of immunology 167, 336-343) и тучные клетки (Allahverdi et al., 2007, Journal of Experimental Medicine, 204, 253-258). Клеточные популяции, характеризующиеся максимальной известной экспрессией как TSLP-R, так и IL-7Ra, представляют собой популяции миелоидных дендритных клеток (Reche et al, 2001, Journal of immunology 167, 336-343).
[003] TSLP может стимулировать пролиферацию наивных Т-клеток и способствовать их дифференцировке в клетки Th2, экспрессирующие высокие уровни IL-4, IL-5 и IL-13 (Omori and Ziegler, 2007, Journal of immunology 178, 1396-1404). Высокий уровень экспрессии TSLP обнаружен в эпителиальных клетках астматических легких и в хронических повреждениях, присутствующих при атопическом дерматите, что свидетельствует о роли TSLP в аллергическом воспалении (Ziegler and Artis, 2010, Nature immunology 11, 289-293). Более поздние данные свидетельствуют об участии TSLP в дифференцировке клеток Th17 и в воспалительных процессах, управляемых Th17 (Hartgring et al, 2011, Arthritis and rheumatism 63, 1878-1887; Tanaka et al, 2009, Clinical and experimental allergy: Journal of the British Society for Allergy and Clinical Immunology 39, 89-100, Wu et al, 2014, Journal of molecular and cellular cardiology 76, 33-45). Хроническая аллергическая (атопическая) астма часто характеризуется воспалением Th2-типа, тогда как неаллергическое астматическое воспаление преимущественно характеризуется наличием нейтрофилов со смешанным цитокиновым окружением Th1 и Th17. Последствия хронического воспаления при астме включают гиперреактивность бронхов (BHR), чрезмерную продукцию слизи, ремоделирование стенки дыхательных путей и сужение дыхательных путей (Lambrecht and Hammad, 2014, Nature immunology 16, 45-56). Показано, что TSLP участвует в инициации и поддержании/усилении аллергического астматического ответа (Wang et al., 2006, Immunity 24, 827-838). Совсем недавно было обнаружено, что опосредованная TSLP передача сигнала необходима для вторичного иммунного ответа с участием Т-клеток памяти на местное введение антигена (Wang et al., 2015, The Journal of allergy and Cliniology immunology 135, 781-791 e783).
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[004] В одном аспекте изобретение предлагает молекулы, например моноклональные антитела или их фрагменты, такие как Fab, Fab', F(ab')2, scFv, миниантитела или диатела, способные специфически связывать тимусный стромальный лимфопоэтин (TSLP). В некоторых вариантах осуществления TSLP-связывающие молекулы могут содержать: определяющий комплементарность участок тяжелой цепи 1 (HCDR1), содержащий аминокислотную последовательность SEQ ID NO: 4; определяющий комплементарность участок тяжелой цепи 2 (HCDR2), содержащий аминокислотную последовательность SEQ ID NO: 2; определяющий комплементарность участок тяжелой цепи 3 (HCDR3), содержащий аминокислотную последовательность SEQ ID NO: 3; определяющий комплементарность участок легкой цепи 1 (LCDR1), содержащий аминокислотную последовательность SEQ ID NO: 11; определяющий комплементарность участок легкой цепи 2 (LCDR2), содержащий аминокислотную последовательность SEQ ID NO: 12; и определяющий комплементарность участок легкой цепи 3 (LCDR3), содержащий аминокислотную последовательность SEQ ID NO: 13. В некоторых вариантах осуществления TSLP-связывающие молекулы могут включать в себя: молекулу, которая содержит: HCDR1, содержащий аминокислотную последовательность SEQ ID NO: 5; HCDR2, содержащий аминокислотную последовательность SEQ ID NO: 6; HCDR3, содержащий аминокислотную последовательность SEQ ID NO: 3; LCDR1, содержащий аминокислотную последовательность SEQ ID NO: 14; LCDR2, содержащий аминокислотную последовательность SEQ ID NO: 15; и LCDR3, содержащий аминокислотную последовательность SEQ ID NO: 16.
[005] В некоторых конкретных вариантах осуществления молекула содержит фрагмент антитела, который связывает человеческий TSLP и содержит HCDR1, содержащий аминокислотную последовательность SEQ ID NO: 4; HCDR2, содержащий аминокислотную последовательность SEQ ID NO: 2; HCDR3, содержащий аминокислотную последовательность SEQ ID NO: 3; LCDR1, содержащий аминокислотную последовательность SEQ ID NO: 11; LCDR2, содержащий аминокислотную последовательность SEQ ID NO: 12; и LCDR3, содержащий аминокислотную последовательность SEQ ID NO: 13. В других конкретных вариантах осуществления молекула содержит фрагмент антитела, который связывает человеческий TSLP и содержит HCDR1, содержащий аминокислотную последовательность SEQ ID NO: 5; HCDR2, содержащий аминокислотную последовательность SEQ ID NO: 6; HCDR3, содержащий аминокислотную последовательность SEQ ID NO: 3; LCDR1, содержащий аминокислотную последовательность SEQ ID NO: 14; LCDR2, содержащий аминокислотную последовательность SEQ ID NO: 15; и LCDR3, содержащий аминокислотную последовательность SEQ ID NO: 16.
[006] В некоторых вариантах осуществления TSLP-связывающие молекулы могут содержать: вариабельный участок тяжелой цепи, содержащий аминокислотную последовательность SEQ ID NO: 7, и вариабельный участок легкой цепи, содержащий аминокислотную последовательность SEQ ID NO: 17.
[007] В некоторых вариантах осуществления TSLP-связывающие молекулы могут содержать: тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 22, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 25. В некоторых вариантах осуществления TSLP-связывающие молекулы могут содержать: тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 9, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 19.
[008] В некоторых вариантах осуществления TSLP-связывающие молекулы могут содержать паратоп, содержащий по меньшей мере один, по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть, по меньшей мере семь, по меньшей мере восемь, по меньшей мере девять, по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, или все из следующих остатков: Thr28, Asp31, Tyr32, Trp33, Asp56, Glu101, Ile102, Tyr103, Tyr104, Tyr105 последовательности тяжелой цепи SEQ ID NO: 22, и Gly28, Ser29, Lys30, Tyr31, Tyr48, Asp50, Asn51, Glu52, Asn65 и Trp92 последовательности легкой цепи SEQ ID NO: 25.
[009] В некоторых вариантах осуществления изобретение предлагает молекулы, способные специфически связываться с эпитопом человеческого TSLP, где эпитоп содержит по меньшей мере один, по меньшей мере, два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть, по меньшей мере семь, при не менее восемь, по меньшей мере 9, по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, или все из следующих остатков: Lys38, Ala41, Leu44, Ser45, Thr46, Ser48, Lys49, Ile52, Thr53, Ser56, Gly57, Thr58, Lys59, Lys101, Gln145 и Arg149 последовательности SEQ ID NO: 38. В некоторых вариантах осуществления такие молекулы связывают эпитоп, содержащий по меньшей мере один из следующих наборов остатков SEQ ID NO: 38: (a) Lys49 и Ile52, (b) Gly57 и Lys59, (c) Lys101 или (d) Gln145 и Arg149.
[0010] В некоторых вариантах осуществления TSLP-связывающие молекулы представляют собой человеческие иммуноглобулины, способные специфически связывать человеческий TSLP. В некоторых вариантах осуществления TSLP-связывающие молекулы представляют собой моноклональные антитела или фрагменты антител, выбранные из Fab, Fab', F(ab')2, scFv, миниантитела или диатела. В некоторых вариантах осуществления TSLP-связывающие молекулы представляют собой Fab, например человеческие или гуманизированные Fab, способные специфически связывать человеческий TSLP.
[0011] В некоторых вариантах осуществления описанные здесь молекулы связывают человеческий TSLP с константой диссоциации (KD) менее 100 пМ. В некоторых вариантах осуществления описанные здесь молекулы связывают человеческий TSLP с константой диссоциации (KD) менее 10 пМ.
[0012] В другом аспекте изобретение предлагает фармацевтические композиции, содержащие по меньшей мере одну описанную здесь TSLP-связывающую молекулу и по меньшей мере одно фармацевтически приемлемое вспомогательное вещество. В некоторых вариантах осуществления отношение массы вспомогательного вещества к массе TSLP-связывающей молекулы больше 0,5. В некоторых вариантах осуществления масса TSLP-связывающей молекулы составляет примерно от 5% до 95%, или примерно от 10% до 90%, или примерно от 15% до 85%, или примерно от 20% до 80%, или примерно от 25% до 75%, или примерно от 30% до 70%, или примерно от 40% до 60%, или примерно 40-50% от массы фармацевтической композиции. В некоторых вариантах осуществления фармацевтические композиции содержат средство, образующее оболочку, такое как трилейцин или лейцин. В некоторых вариантах осуществления масса трилейцина или лейцина составляет примерно 10-75% от массы композиции. В некоторых вариантах осуществления масса трилейцина составляет примерно 10-30% от массы композиции. В другом варианте осуществления масса лейцина составляет примерно 50-75% от массы композиции. В некоторых вариантах осуществления фармацевтические композиции содержат по меньшей мере одно стеклообразующее вспомогательное вещество, выбранное из гистидина, трегалозы, маннита, сахарозы или цитрата натрия. В некоторых вариантах осуществления по меньшей мере одно стеклообразующее вспомогательное вещество представляет собой трегалозу или смесь трегалозы и маннита. В некоторых вариантах осуществления масса стеклообразующего вспомогательного вещества составляет примерно 15-35% от массы композиции. В некоторых вариантах осуществления фармацевтические композиции содержат буфер, такой как гистидиновый, глициновый, ацетатный или фосфатный буфер. В некоторых вариантах осуществления содержание буфера в композиции составляет примерно 5-13%.
[0013] В некоторых вариантах осуществления описанные здесь фармацевтические композиции получают в виде сухого порошкообразного состава, например сухого порошкообразного состава, пригодного для вдыхания.
[0014] В некоторых вариантах осуществления описанные здесь фармацевтические композиции содержат высушенные распылением частицы, содержащие оболочку и ядро, причем оболочка содержит трилейцин или лейцин, а ядро содержит: (i) TSLP-связывающую молекулу, трегалозу, маннит и буфер; или (ii) TSLP-связывающую молекулу, трегалозу, буфер и HCl. Буфер может представлять собой гистидиновый, глициновый, ацетатный или фосфатный буфер.
[0015] В некоторых вариантах осуществления описанные здесь фармацевтические композиции содержат высушенные распылением частицы, содержащие: (i) оболочку, содержащую трилейцин или лейцин; и (ii) ядро, содержащее трегалозу, маннит, гистидин и TSLP-связывающую молекулу, или ядро, содержащее трегалозу, гистидин, HCl и TSLP-связывающую молекулу, где TSLP-связывающая молекула представляет собой фрагмент антитела Fab, содержащий: либо (а) HCDR1, содержащий аминокислотную последовательность SEQ ID NO: 4; HCDR2, содержащий аминокислотную последовательность SEQ ID NO: 2; HCDR3, содержащий аминокислотную последовательность SEQ ID NO: 3; LCDR1, содержащий аминокислотную последовательность SEQ ID NO: 11; LCDR2, содержащий аминокислотную последовательность SEQ ID NO: 12; и LCDR3, содержащий аминокислотную последовательность SEQ ID NO: 13; либо (b) HCDR1, содержащий аминокислотную последовательность SEQ ID NO: 5; HCDR2, содержащий аминокислотную последовательность SEQ ID NO: 6; HCDR3, содержащий аминокислотную последовательность SEQ ID NO: 3; LCDR1, содержащий аминокислотную последовательность SEQ ID NO: 14; LCDR2, содержащий аминокислотную последовательность SEQ ID NO: 15; и LCDR3, содержащий аминокислотную последовательность SEQ ID NO: 16.
[0016] В некоторых вариантах осуществления описанные здесь фармацевтические композиции содержат:
(a) 40% (масс./масс.) TSLP-связывающей молекулы, 25% (масс./масс.) трилейцина, 30% (масс./масс.) в сумме трегалозы и маннита и 5% (масс./масс.) гистидина;
(b) 50% (масс./масс.) TSLP-связывающей молекулы, 15% (масс./масс.) трилейцина, 2,6% (масс./масс.) HCl, 5,6% (масс./масс.) гистидина и 26,8% (масс./масс.) в сумме трегалозы и основания; или
(c) 50% (масс./масс.) TSLP-связывающей молекулы, 15% (масс./масс.) трилейцина, 19,4% (масс./масс.) трегалозы, 13,04% (масс./масс.) гистидина и 2,56% (масс./масс.) НС1.
[0017] Изобретение также предлагает нуклеиновые кислоты, кодирующие описанные здесь TSLP-связывающие молекулы, векторы, содержащие такие нуклеиновые кислоты, и клетки-хозяева, содержащие нуклеиновые кислоты или векторы.
[0018] Изобретение также предлагает способы получения описанной здесь TSLP-связывающей молекулы. Такие способы могут включать в себя (a) культивирование клетки-хозяина, экспрессирующей нуклеиновую кислоту, кодирующую молекулу; и (b) сбор молекулы из культуральной среды.
[0019] В другом аспекте изобретение предлагает наборы, содержащие по меньшей мере одну TSLP-связывающую молекулу или описанную здесь фармацевтическую композицию и устройство для доставки молекулы или фармацевтической композиции индивидууму. В некоторых вариантах осуществления устройство может доставлять молекулу или фармацевтическую композицию в аэрозольной форме. В некоторых вариантах осуществления устройство представляет собой ингалятор сухого порошка.
[0020] В другом аспекте изобретение предлагает способы лечения связанного с TSLP состояния у индивидуума, нуждающегося в этом, например, у человека, путем введения индивидууму терапевтически эффективного количества любой описанной здесь TSLP-связывающей молекулы или фармацевтической композиции. Кроме того, изобретение предлагает применение описанных здесь молекул или фармацевтических композиций для лечения связанного с TSLP состояния у индивидуума, нуждающегося в этом. Применение описанных здесь TSLP-связывающих молекул или фармацевтических композиций для лечения связанного с TSLP состояния у индивидуума, нуждающегося в этом, также входит в объем изобретения. Настоящее изобретение также включает в себя применение описанной здесь молекулы в способе промышленного получения лекарственного средства для лечения связанного с TSLP состояния у индивидуума, нуждающегося в этом.
[0021] Связанное с TSLP воспалительное состояние может представлять собой астму, хроническую обструктивную болезнь легких, аллергический ринит, аллергический риносинусит, аллергический конъюнктивит, эозинофильный эзофагит или атопический дерматит. В некоторых вариантах осуществления связанное с TSLP воспалительное состояние представляет собой астму. В некоторых вариантах осуществления TSLP-связывающая молекула входит в состав сухого порошкообразного состава, пригодного для ингаляции. В некоторых вариантах осуществления TSLP-связывающую молекулу вводят индивидууму перорально или интраназально, например, в составе аэрозольной формы. В некоторых вариантах осуществления TSLP-связывающую молекулу вводят индивидууму с помощью ингалятора сухого порошка.
[0022] В некоторых вариантах осуществления способы лечения связанного с TSLP состояния или способы применения TSLP-связывающей молекулы дополнительно включают в себя введение второго средства индивидууму, нуждающемуся в лечении. Второе средство может представлять собой кортикостероид, бронхолитическое средство, антигистаминное средство, антилейкотриеновое средство или ингибитор PDE-4.
[0023] В другом аспекте изобретение предлагает способы получения сухого порошкообразного состава, содержащего описанную здесь TSLP-связывающую молекулу. Такие способы могут включать в себя одну или несколько из следующих стадий: (a) получение водного раствора, содержащего описанную здесь TSLP-связывающую молекулу, трилейцин или лейцин, стеклообразующее вспомогательное вещество и буфер; (b) распылительная сушка водного раствора, полученного на стадии (a), при температуре в диапазоне примерно от 120°С до 200°С (на входе) и примерно от 55°С до 75°С (на выходе) с получением частиц сухого порошка; и (c) сбор частиц сухого порошка. В некоторых вариантах осуществления буфер выбирают из гистидинового, глицинового, ацетатного или фосфатного буфера. В некоторых вариантах осуществления стеклообразующее вспомогательное вещество выбирают из гистидина, гистидина HCl, трегалозы, маннита, сахарозы или цитрата натрия.
[0024] Подробное описание одного или нескольких вариантов осуществления настоящего изобретения можно найти в прилагаемых чертежах и в приведенном ниже описании. Другие признаки, объекты и преимущества изобретения будут очевидны из описания и чертежей, а также из формулы изобретения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0025] На фиг. 1А показана аминокислотная последовательность тяжелой цепи Fab1 против человеческого TSLP (SEQ ID NO: 22), в которой CDR (определенные по системе Kabat) подчеркнуты, а остатки, расположенные на поверхности контакта антитело-антиген, помечены *. На фиг. 1B показана аминокислотная последовательность легкой цепи Fab1 против человеческого TSLP (SEQ ID NO: 25), в которой CDR (определенные по системе Kabat) подчеркнуты, а остатки, расположенные на поверхности контакта антитело-антиген, помечены *.
[0026] На фиг. 2 показана аминокислотная последовательность рекомбинантного человеческого TSLP, используемого в кристаллографических методах исследования (SEQ ID NO: 38), а также элементы вторичной структуры, изображенные ниже аминокислотной последовательности. В рамки заключены α-спиральные участки αA, αB, αC и αD, а жирными линиями обозначены петлевые области. Зрелый человеческий TSLP начинается с Tyr29. Используемая здесь конструкция содержит на N-конце гексагистидиновый маркер (SEQ ID NO: 40) (остатки 15-20), затем участок распознавания протеазой HRV-3C (PreScission) (остатки 21-28) и остатки 11-14, полученные в результате клонирования. Asn64 и Asn119 являются потенциальными участками N-гликозилирования; а остатки 127-130 составляют участок расщепления фурином.
[0027] На фиг. 3 приведена гистограмма, демонстрирующая влияние нейтрализации TSLP на воспаление легких у мышей, сенсибилизированных овальбумином, которым был введен антиген. Мышам, сенсибилизированным овальбумином (OVA) или физиологическим раствором с квасцами, за 1 ч до сенсибилизации внутривенно вводят либо антитело против мышиного TSLP, либо изотипическое контрольное антитело. Всем мышам вводят OVA на 21 день и через 24 часа их умерщвляют. Значения представляют собой средние значения±SEM (стандартная ошибка среднего значения) общего числа клеток и числа отдельных форменных элементов в BAL. Статистический анализ проводят с использованием t-критерия Стьюдента для одной выборки. Значимые различия между мышами, обработанными изотипическим антителом, сенсибилизированными солевым раствором и сенсибилизированными OVA при p<0,05 обозначены (*), а при p<0,01 обозначены (**). Различия между OVA-сенсибилизированными мышами, обработанными изотипическим антителом и антителом против TSLP, при p<0,05 обозначены (#). [PMN: полиморфноядерные клетки (нейтрофилы); Eos: эозинофилы; МО: моноциты; Lymph: лимфоциты; TCC: общее число клеток].
[0028] На фиг. 4A-4C приведен ряд гистограмм, демонстрирующих, что нейтрализация TSLP значительно снижает уровни IL-13 (фиг.4A), эотаксина-2 (CCL24, фиг.4B) и хемокина, регулируемого тимусом и активацией (TARC, CCL17, Фиг.4С), в легких сенсибилизированных овальбумином мышей, которым был введен антиген. Мышам, сенсибилизированным OVA (или физиологическим раствором) и квасцами, за 1 ч до сенсибилизации внутривенно вводят либо антитело против мышиного TSLP, либо изотипическое контрольное антитело. Всем мышам вводят OVA на 21 день и через 24 часа их умерщвляют. Значения представляют собой средние значения±SEM уровней медиаторов, измеренных в BAL методом специфического ELISA. Статистический анализ проводят с использованием t-критерия Стьюдента для одной выборки. Значимые различия между мышами, обработанными изотипическим антителом, сенсибилизированными солевым раствором и сенсибилизированными OVA при p<0,05 обозначены (*), а при p<0,01 обозначены (**). Различия между OVA-сенсибилизированными мышами, обработанными изотипическим антителом и антителом против TSLP, при p<0,05 обозначены (#).
[0029] На фиг. 5 приведен линейный график, демонстрирующий зависимость средней концентрации общего Fab1 против TSLP в сыворотке обезьян от времени.
[0030] На фиг. 6A и 6B приведены гистограммы, демонстрирующие средние концентрации общего Fab1 против TSLP в BAL (6A) или гомогенате легкого (6B) обезьян через 1 час (группы, получающие ингаляции 1, 10, 20 мг/кг/день) или через 6 дней (группа, получающая 1 мг/кг в.в.+ингаляцию 20 мг/кг/день) после последней вдыхаемой дозы.
[0031] На фиг. 7 приведено общее изображение человеческого TSLP в комплексе с Fab1 против TSLP. Спирали TSLP помечены A-D в направлении от N-конца к C-концу.
[0032] На фиг. 8 показан эпитоп TSLP, на который направлен Fab1 против TSLP. В верхней части чертежа показано число непосредственных межмолекулярных контактов между отличными от водорода атомами на расстоянии 4,0 Å, а в нижней части показано уменьшение доступной для растворителя поверхности при образовании комплекса. Аминокислотная последовательность TSLP (SEQ ID NO: 41) показана на горизонтальной оси.
[0033] На фиг. 9 показаны антитело и эпитоп TSLP. TSLP изображен в виде ленты. Все аминокислотные остатки, участвующие в непосредственном контакте с Fab1 (отсечение по расстоянию 4,0 Å), показаны в виде шариков и стержней.
[0034] На фиг. 10А и 10В показан паратоп тяжелой цепи (SEQ ID NO: 42) (А) и легкой цепи (SEQ ID NO: 43) (В) Fab1 против TSLP. В верхней части чертежа показано число непосредственных межмолекулярных контактов (<4,0 Å) между отличными от водорода атомами, а в нижней части показано уменьшение доступной для растворителя поверхности при образовании комплекса. Аминокислотная последовательность вариабельного домена тяжелой или легкой цепи изображена на горизонтальной оси.
[0035] На фиг. 11А-11С показан иеханизм действия Fab1 против TSLP. На фиг. 11А изображен мышиный внеклеточный сигнальный комплекс, где IL-7Rα отмечен черным цветом, а TSLPR - светло-серым. На фиг. 11B изображен комплекс человеческий TSLP-Fab1 в такой же ориентации, что и на фиг. 11А. На фиг. 11С изображено структурное наложение двух комплексов с использованием в качестве основы Сα-атомов цитокина. Мышиный сигнальный комплекс отмечен светло-серым цветом, а комплекс человеческий TSLP-Fab1 - черным цветом.
[0036] На фиг. 12 приведена диаграмма разброса, демонстрирующая, что композиции с более высоким отношением вспомогательное вещество:белок характеризуются улучшенной физико-химической стабильностью Fab1 против TSLP, что иллюстрируется уменьшением скорости агрегации белка.
ПОДРОБНОЕ ОПИСАНИЕ
Определения
[0037] Если контекст явно не указывает иное, единственная форма, используемая в описании и формуле изобретения, включает в себя ссылки на множественную форму. Например, термин "ячейка" включает в себя множество клеток, том числе их смеси.
[0038] Все численные значения, например, значения рН, температуры, времени, концентрации и молекулярной массы, включая диапазоны, являются приближенными и могут варьировать (+) или (-) с шагом 0,1. Следует понимать, хотя это не всегда в явной форме указано, что всем численным значениям предшествует термин "примерно". Кроме того, следует понимать, хотя это не всегда в явной форме указано, что описанные здесь реагенты являются только примерами реагентов, и что можно использовать их эквиваленты, известные в данной области.
[0039] Используемый здесь термин "TSLP" (также известный как "тимусный стромальный лимфопоэтин") относится к цитокину, продуцируемому клетками, отличными от гемопоэтических, в ответ на провоспалительные стимулы. Человеческий ген TSLP относят к хромосомному расположению 5q22.1, а геномную последовательность гена TSLP можно найти в GenBank под номером NC_000005.10. Вследствие альтернативного сплайсинга у людей присутствуют две изоформы TSLP. Белковые последовательности и последовательности мРНК двух изоформ человеческого TSLP приведены в таблице 1.
ТАБЛИЦА 1. Аминокислотные последовательности и последовательности мРНК TSLP
tgcccgtcac tccccgagaa agcgttcgct ttgaaaacga aagcggccct cattaactac tgcccaggct actctgaaac tgagagaaat ggtactctgg aaatgacacg agaaatcaga
aacatctgcc tgaatcaaac ctcacaaatt ctaggattgt ggctttcctg cattcaatct tgaagaaaaa attagctttt ggattatatt atgaaaatat atatcttgtt tttagtagat ataatactaa gggtgtgaca cttaaaagaa cactaatgtt tattctttaa ttatagaagg gattcttaac ttatttttgg catatcgttg tttagtgtag gcgctttaaa tggaaaatga gcattacccc tttaatggaa ataaccgtgc tgttaatgat tggcttcggc ttctgagcag
tctttctcac ctcacctgag acactttact ctagggcaaa cctaactgta gtaggaagaa aatcaaaagt agaaaaacag ttgaaaccaa tgacaggatc tatactccat ttaaaaggca agaatttttg tacctgtaat gattcttcta acattcctac gctaagattt tactaaagaa gaaaataaca gcagaggaaa gtgttcaggc agtcactgcc atgatgaagc tgtcagaatc tgagagctac tgctgcaact gatcgtgtag taaatccagc tgtaaagggg atcttaaccc accacagtgg gatgcacagg cagatcccca agggcattgt gcagctgtga gatatatatt taaagttgta tataatgatt ttctaattta ttccgtgagc acctttgaaa atatacatgt cgctgtgtaa caaatacact tctgaataaa gtaattctca agttc (SEQ ID NO: 37)
[0040] Более длинная изоформа 1 TSLP связана с развитием воспалительного заболевания дыхательных путей (Headley et al, 2009, Journal of immunology 182, 1641-1647, Ying et al., 2005, Journal of immunology 174, 8183-8190). Используемый здесь термин "TSLP" относится к изоформе 1 TSLP. В настоящем описании человеческий белок TSLP также включает в себя белки, последовательности которых по всей длине идентичны аминокислотной последовательности с номером доступа GenBank NP_149024.1 по меньшей мере примерно на 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96% 97%, 98%, 99% или 100%. Нуклеотидная последовательность человеческого TSLP по всей длине идентична нуклеотидной последовательности с номером доступа GenBank NM_033035.4 по меньшей мере примерно на 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81% 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100%. Последовательности белков TSLP мышей, яванских макак и других животных известны в данной области (см., например, таблицу 1).
[0041] Используемый здесь термин "антитело" относится к белковой или полипептидной последовательности, полученной из молекулы иммуноглобулина и способной специфически связываться с антигеном. Антитела могут представлять собой поликлональные или моноклональные, многоцепочечные или одноцепочечные, или интактные иммуноглобулины и могут быть получены из природных источников или с помощью рекомбинантных методов. Природное "антителом" представляет собой гликопротеин, содержащий по меньшей мере две тяжелые (H) цепи и две легкие (L) цепи, соединенные между собой дисульфидными связями. Каждая тяжелая цепь состоит из вариабельного участка тяжелой цепи (сокращенно обозначаемого здесь VH) и константного участка тяжелой цепи. Константный участок тяжелой цепи состоит из трех доменов: CH1, CH2 и CH3. Каждая легкая цепь состоит из вариабельного участка легкой цепи (сокращенно обозначаемого здесь VL) и константного участка легкой цепи. Константный участок легкой цепи состоит из одного домена CL. Участки VH и VL можно далее подразделить на участки гипервариабельности, называемые участками, определяющими комплементарность (CDR), которые чередуются с более консервативными участками, называемыми каркасными (FR). Каждый VH и VL состоит из трех CDR и четырех FR, расположенных в направлении от аминоконца к карбоксиконцу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные участки тяжелой и легкой цепей содержат связывающий домен, который взаимодействует с антигеном. Константные участки антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, в том числе с разными клетками иммунной системы (такими как эффекторные клетки) и первым компонентом (C1q) классической системы комплемента. Антитело может представлять собой моноклональное антитело, человеческое антитело, гуманизированное антитело, верблюжье антитело или химерное антитело. Антитела могут принадлежать к любому изотипу (например, IgG, IgE, IgM, IgD, IgA и IgY), классу (например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2) или подклассу.
[0042] Термины "фрагмент антитела", "антигенсвязывающий фрагмент", "его антигенсвязывающий фрагмент", "антигенсвязывающий домен" антитела и т.п. в настоящем описании относятся к одному или нескольким фрагментам интактного антитела, которые сохраняют способность специфически связываться с конкретным антигеном (например, TSLP). Антигенсвязывающие функции антитела могут выполнять фрагменты интактного антитела. Примеры связывающих фрагментов, охватываемых термином "антигенсвязывающий домен" антитела, включают в себя фрагмент Fab, одновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; фрагмент F(ab)2, двухвалентный фрагмент, содержащий два фрагмента Fab, связанные дисульфидным мостиком в шарнирной области; фрагмент Fd, состоящий из доменов VH и CH1; фрагмент Fv, состоящий из доменов VL и VH одного плеча антитела; однодоменный фрагмент антитела (dAb) (Ward et al, 1989 Nature 341: 544-546), который состоит из домена VH; и выделенный участок определения комплементарности (CDR). Кроме того, хотя два домена фрагмента Fv, VL и VH, кодируются разными генами, их можно объединить с помощью рекомбинантных методов с использованием искусственного пептидного линкера, который позволяет им находиться в одной белковой цепи, где участки VL и VH спариваются, образуя одновалентные молекулы (известные как одноцепочечный Fv (scFv), см., например, Bird et al., 1988 Science 242: 423-426 и Huston et al., 1988 Proc. Natl. Acad. Sci. 85:5879-5883). Такие одноцепочечные антитела содержат один или несколько "антигенсвязывающих доменов" антитела. Указанные фрагменты антител получают с помощью традиционных способов, известных специалистам в данной области, и подвергают скринингу на полезные свойства подобно интактным антителам. Антигенсвязывающие домены также могут быть включены в однодоменные антитела, макситела, минитела, интратела, диатела, триатела, тетратела, v-NAR и бис-scFv (см., например, Hollinger and Hudson, 2005, Nature Biotechnology, 23, 9, 1126-1136). Антигенсвязывающие домены антител можно прививать на каркасы из полипептидов, такие как фибронектин типа III (Fn3) (см. патент США № 6703199, в котором описаны монотела на основе полипептида фибронектина). Антигенсвязывающие домены можно ввести в состав одноцепочечных молекул, содержащих пару тандемных сегментов Fv (VH-CH1-VH-CH1), которые вместе с комплементарными полипептидами легкой цепи образуют пару антигенсвязывающих участков (Zapata et al., 1995 Protein 8 (10): 1057-1062 и патент США 5641870).
[0043] Термин "эпитоп" относится к любой белковой детерминанте, способной специфически связываться с иммуноглобулином, или иным образом взаимодействовать с молекулой. Эпитопные детерминанты обычно состоят из химически активных поверхностных совокупностей фрагментов молекул, таких как боковые цепи аминокислот, или углеводов, или сахаров, и могут иметь специфические трехмерные структурные характеристики, а также специфические характеристики заряда. Эпитоп может быть "линейным" или "конформационным". Конформационные и линейные эпитопы отличаются тем, что связывание с первым, но не с последним, исчезает в присутствии денатурирующих растворителей.
[0044] Термин "паратоп" определяют как и приведенный выше термин "эпитоп", но с обращением концепции. Так, используемый здесь термин "паратоп" относится к области или участку антитела или фрагмента антитела, с которым специфически связывается антиген, т.е. по которому антитело или фрагмент антитела осуществляет физический контакт с антигеном.
[0045] В контексте кристаллической структуры, установленной рентгенографическим методом путем определения пространственных координат комплекса антитела, например фрагмента Fab, с его антигеном, если не указано иное, или если это не противоречит контексту, термин "паратоп" в настоящем описании конкретно определяют как остатки антитела, характеризующиеся наличием тяжелого атома (то есть атома, отличного от водорода), находящегося на определенном расстоянии, например в пределах 4 ангстрем, от тяжелого атома, входящего в состав антигена-мишени.
[0046] Используемые здесь термины "участки, определяющие комплементарность" и "CDR" относятся к аминокислотным остаткам антитела или антигенсвязывающего фрагмента, которые отвечают за связывание антигена.
[0047] Используемый здесь термин "моновалентное антитело" относится к антителу, которое связывается с одним эпитопом на молекуле-мишени.
[0048] Используемый здесь термин "бивалентное антитело" относится к антителу, которое связывается с двумя эпитопами по меньшей мере на двух идентичных молекулах-мишенях. Двухвалентное антитело может также сшивать молекулы-мишени друг с другом. Термин "двухвалентное антитело" также относится к антителу, которое связывается с двумя разными эпитопами по меньшей мере на двух идентичных молекулах-мишенях.
[0049] Термин "поливалентное антитело" относится к одной связывающей молекуле, обладающей несколькими валентностями, где "валентность" определяют как количество антигенсвязывающих фрагментов в молекуле конструкции антитела. Как таковая, одна связывающая молекула может связываться с несколькими участками связывания на молекуле-мишени. Примеры поливалентных антител включают в себя, без ограничения, двухвалентные антитела, трехвалентные антитела, четырехвалентные антитела, пятивалентные антитела и т.п., а также биспецифические антитела и бипаратопические антитела. Например, в случае TSLP, поливалентное антитело, такое как бипаратопическое антитело против TSLP, может содержать связывающий фрагмент, который распознает два разных домена TSLP, соответственно.
[0050] Термин "поливалентное антитело" также относится к одной связывающей молекуле, которая содержит несколько антигенсвязывающих фрагментов, специфичных к двум разным молекулам-мишеням. Примером может служить антитело, которое связывается с TSLP и второй молекулой-мишенью, отличной от TSLP. В одном варианте осуществления поливалентное антитело представляет собой четырехвалентное антитело, которое содержит четыре эпитоп-связывающих домена. Четырехвалентная молекула может быть биспецифичной и двухвалентной по каждому связывающему участку на молекуле-мишени.
[0051] Используемый здесь термин "бипаратопическое антитело" относится к антителу, способному связываться с двумя разными эпитопами на одной молекуле-мишени. Термин также относится к антителу, способному связываться с двумя доменами, присутствующими по меньшей мере на двух молекулах-мишенях, например, к четырехвалентному бипаратопическому антителу.
[0052] Используемый здесь термин "биспецифическое антитело" относится к антителу, способному связываться с двумя или несколькими разными эпитопами по меньшей мере на двух разных мишенях.
[0053] Используемые здесь фразы "моноклональное антитело" или "композиция моноклонального антитела" относятся к полипептидам, включающим в себя антитела, биспецифические антитела и т.д., которые имеют практически идентичные аминокислотные последовательности, или получены из одного и того же генетического источника. Данный термин также относится к препаратам молекул антител, имеющих одинаковый молекулярный состав. Композиция моноклонального антитела характеризуется одной специфичностью связывания и сродством к конкретному эпитопу.
[0054] Используемая здесь фраза "человеческое антитело" относится к антителам, содержащим вариабельные участки, в которых и каркасные участки, и CDR получены из последовательностей человеческого происхождения. Кроме того, если антитело содержит константный участок, он также получен из таких человеческих последовательностей, например, из человеческих зародышевых последовательностей, или из мутантных версий человеческих зародышевых последовательностей, или из антител, содержащих консенсусные каркасные последовательности, полученные в результате анализа человеческих каркасных последовательностей, например, как описано в Knappik, et al. (2000. J Mol Biol 296, 57-86). Структуру и расположение вариабельных доменов иммуноглобулина, например CDR, можно определить с использованием хорошо известных систем нумерации, например, системы нумерации Кабат, системы нумерации Чотиа или сочетания систем Кабат и Чотиа (см., например, Sequences of Proteins of Immunological Interest, U.S. Department of Health and Human Services (1991), eds. Kabat et al.; Al Lazikani et al., (1997) J. Mol. Bio. 273:927 948); Kabat et al., (1991) Sequences of Proteins of Immunological Interest, 5th edit., NIH Publication no. 91-3242 U.S. Department of Health and Human Services; Chothia et al., (1987) J. Mol. Biol. 196:901-917; Chothia et al., (1989) Nature 342:877-883; and Al-Lazikani et al., (1997) J. Mal. Biol. 273:927-948).
[0055] Человеческие антитела настоящего изобретения могут содержать аминокислотные остатки, не кодируемые человеческими последовательностями (например, в результате мутаций, введенных путем неспецифического или сайт-специфического мутагенеза in vitro, или соматической мутации in vivo, или консервативной замены, способствующей стабильности или облегчающей производственный процесс). Однако используемый здесь термин "человеческое антитело" не относится к антителам, в которых последовательности CDR, полученные из зародышей млекопитающих другого вида, таких как мыши, привиты на человеческие каркасные последовательности.
[0056] Используемая здесь фраза "рекомбинантное человеческое антитело" относится ко всем человеческим антителам, которые получают, экспрессируют, создают или выделяют рекомбинантными средствами, таким как антитела, выделенные из животного (например, мыши), которое является трансгенным или трансхромосомным по генам человеческих иммуноглобулинов, или из полученной из него гибридомы, антитела, выделенные из клетки-хозяина, трансформированной с целью обеспечения экспрессии человеческого антитела, например, из трансфектомы, антитела, выделенные из библиотеки рекомбинантных комбинаторных человеческих антител, и антитела, полученные, экспрессированные, созданные или выделенные любыми другими способами, включая сплайсинг последовательностей полноразмерного гена человеческого иммуноглобулина или его части, с получением других последовательностей ДНК. Такие рекомбинантные человеческие антитела содержат вариабельные участки, в которых каркасные участки и участки CDR получены из последовательностей зародышевых человеческих иммуноглобулинов. Однако в некоторых вариантах осуществления такие рекомбинантные человеческие антитела могут подвергаться мутагенезу in vitro (или, если используют животное, трансгенное по человеческим последовательностям Ig, соматическому мутагенезу in vivo), в результате чего аминокислотные последовательности VH- и VL-участков рекомбинантных антител могут представлять собой последовательности, которые, будучи полученными из зародышевых человеческих последовательностей VH и VL и родственными им, могут не существовать в природе в зародышевом наборе человеческих антител in vivo.
[0057] Используемый здесь термин "Fc-участок" относится к полипептиду, содержащему CH3, CH2 и по меньшей мере часть шарнирного участка константного домена антитела. Необязательно Fc-участок может содержать домен CH4, присутствующий в некоторых классах антител. Fc-участок может содержать весь шарнирный участок константного домена антитела. В одном варианте осуществления изобретение включает Fc-участок и участок CH1 антитела. В одном варианте осуществления изобретение включает Fc-участок и участок CH3 антитела. В другом варианте осуществления изобретение включает Fc-участок, участок CH1 и участок Cкаппа/лямбда константного домена антитела. В одном варианте осуществления связывающая молекула настоящего изобретения содержит константный участок, например, константный участок тяжелой цепи. В одном варианте осуществления такая константный участок является модифицированным по сравнению с константным участком дикого типа. То есть, описанные здесь полипептиды настоящего изобретения могут содержать изменения или модификации в одном или нескольких из трех константных доменов тяжелой цепи (CH1, CH2 или CH3), и/или в домене константного участка легкой цепи (CL). Примеры модификаций включают в себя добавления, делеции или замены одной или нескольких аминокислот в одном или нескольких доменах. Такие изменения могут быть введены для оптимизации эффекторной функции, периода полужизни и т.д.
[0058] Используемый здесь термин "сродство" относится к силе взаимодействия между антителом и антигеном в отдельных антигенных сайтах. В каждом антигенном сайте вариабельный участок "плеча" антитела взаимодействует с антигеном посредством слабых нековалентных сил в множестве точек; причем чем больше таких взаимодействий, тем сильнее сродство. Используемый здесь термин "высокое сродство" в применении к антителу IgG или его фрагменту (например, фрагменту Fab) относится к антителу, способному связывать антиген-мишень в концентрации 10-8 М или менее, 10-9 М или менее, или 10-10 М, или 10-11 М или менее, или 10-12 М или менее, или 10-13 М или менее. Однако высокое сродство связывания может варьировать среди антител разных изотипов. Например, высокое сродство связывания в случае изотипа IgM относится к способности антитела связывать антиген в концентрации 10-7 М или менее, или 10-8 М или менее.
[0059] Используемый здесь термин "авидность" относится к информативному показателю общей стабильности или прочности комплекса антитело-антиген. Он определяется тремя основными факторами: сродством антитела к эпитопу; валентностью как антигена, так и антитела; и структурным расположением взаимодействующих частей. В конечном счете эти факторы определяют специфичность антитела, то есть вероятность того, что конкретное антитело связывается с определенным антигенным эпитопом.
[0060] Используемый здесь термин "специфичность связывания" относится к способности антигенсвязывающего участка отдельного антитела взаимодействовать с одной антигенной детерминантой, но не с другой антигенной детерминантой. Антигенсвязывающий участок антитела расположен в Fab-фрагменте молекулы, его составляют гипервариабельные участки тяжелой и легкой цепей. Сродство связывания антитела представляет собой силу взаимодействия между одной антигенной детерминантой и одним антигенсвязывающим участком антитела. Сила взаимодействия определяется суммой сил притяжения и отталкивания, действующих между антигенной детерминантой и антигенсвязывающим участком антитела.
[0061] Термин "лечить" и "лечение" относится как к терапевтическому лечению, так и к профилактическим или предупредительным мерам, целью которых является предотвращение или замедление нежелательного физиологического изменения или расстройства. В целях настоящего изобретения полезные или желательные клинические результаты включают в себя, без ограничения, облегчение симптомов, уменьшение степени заболевания, стабилизацию (то есть отсутствие ухудшения) состояния заболевания, задержку или замедление прогрессирования заболевания, улучшение или облегчение состояния заболевания и ремиссию (как частичную, так и полнойую), независимо от того, являются ли указанные результаты детектируемыми или недетектируемыми. Термин "лечение" также может относиться к увеличению продолжительности жизни по сравнению с ожидаемой продолжительностью жизни в отсутствии лечения.
[0062] Термин "индивидуум" относится к животному, человеку или отличному от человека животному, которому предоставляется лечение в соответствии со способами настоящего изобретения. Предусматриваются ветеринарные и не ветеринарные способы применения. Данный термин включает в себя, без ограничения, млекопитающих, например людей, других приматов, свиней, грызунов, таких как мыши и крысы, кроликов, морских свинок, хомяков, коров, лошадей, кошек, собак, овец и коз. Типичные индивидуумы включают в себя людей, сельскохозяйственных животных и домашних животных, таких как кошки и собаки.
[0063] Термин "эффективное количество" относится к количеству, достаточному для достижения полезных или желательных результатов. Например, терапевтическое количество представляет собой количество, достаточное для достижения желательного терапевтического эффекта. Это количество может быть таким же, как профилактически эффективное количество, или отличаться от профилактически эффективного количества, которое представляет собой количество, необходимое для предотвращения возникновения заболевания или симптомов заболевания. Эффективное количество можно вводить за один или несколько раз, в один или несколько приемов, в одной или нескольких дозах. "Терапевтически эффективное количество" терапевтического соединения (то есть эффективная доза) зависит от выбранного терапевтического соединения. Композиции можно вводить, например, от одного или нескольких раз в день до одного или нескольких раз в неделю, одного или нескольких раз в месяц, или одного или нескольких раз в год. Специалистам в данной области известно, что дозы и режим введения, необходимые для эффективного лечения индивидуума, могут зависеть от некоторых факторов, включающих в себя, без ограничения, тяжесть заболевания или расстройства, предшествующие процедуры, общее состояние здоровья и/или возраст индивидуума и наличие других заболеваний. Кроме того, лечение индивидуума терапевтически эффективным количеством описанных здесь терапевтических соединений может включать в себя однократное или многократное введение.
[0064] Термин "нуклеиновая кислота" или "полинуклеотид" относится к дезоксирибонуклеиновым кислотам (ДНК) или рибонуклеиновым кислотам (РНК) и их полимерам, которые могут находиться в виде одно- или двухцепочечных форм. Если конкретные ограничения отсутствуют, данный термин охватывает нуклеиновые кислоты, содержащие известные аналоги природных нуклеотидов, которые обладают связывающими свойствами, подобными связывающим свойствам исходной нуклеиновой кислоты, и метаболизируются аналогично природным нуклеотидам. Если не указано иное, конкретная нуклеотидная последовательность в неявной форме охватывает ее консервативно модифицированные варианты (например, полученные путем замен на вырожденные кодоны), аллели, ортологи, SNP и комплементарные последовательности, а также последовательность, указанную в явной форме. В частности, замены на вырожденные кодоны можно осуществить путем получения последовательностей, в которых по третьему положению одного или нескольких выбранных (или всех) кодонов осуществлена замена на остатки смешанного основания и/или дезоксиинозина (Batzer et al., Nucleic Acid Res. 19:5081 (1991); Ohtsuka et al., J. Biol. Chem. 260:2605-2608 (1985); и Rossolini et al., Mol. Cell. Probes 8:91-98 (1994)).
[0065] Термины "пептид", "полипептид" и "белок" используются как взаимозаменяемые и относятся к соединению, состоящему из аминокислотных остатков, ковалентно связанных пептидными связями. Белок или пептид должен содержать по меньшей мере две аминокислоты, а ограничения на максимальное число аминокислот, которое может присутствовать в последовательности белка или пептида, отсутствуют. Полипептиды включают в себя любые пептиды или белки, содержащие две или более аминокислот, соединенных друг с другом пептидными связями. В соответствии с настоящим описанием данный термин относится как к короткоцепочечным соединениям, в данной области обычно называемым пептиды, олигопептиды и олигомеры, так и к длинноцепочечным соединениям, в данной области обычно называемым белки, которых существует много типов. "Полипептиды" включают в себя, например, биологически активные фрагменты, по существу гомологичные полипептиды, олигопептиды, гомодимеры, гетеродимеры, варианты полипептидов, модифицированные полипептиды, производные, аналоги, гибридные белки и другие. Полипептид может представлять собой природный пептид, рекомбинантный пептид или их сочетание.
[0066] Термин "консервативные модификации последовательности" относится к аминокислотным модификациям, которые не оказывают существенного влияния на связывающие характеристики, или не изменяют связывающие характеристики антитела или фрагмента антитела, содержащего аминокислотную последовательность. Такие консервативные модификации включают в себя аминокислотные замены, добавления и делеции. Модификации можно вводить в антитело или фрагмент антитела настоящего изобретения с помощью стандартных методов, известных в данной области, таких как сайт-направленный мутагенез и ПЦР-опосредованный мутагенез. Консервативные аминокислотные замены представляют собой замены аминокислотных остатков на аминокислотные остатки, содержащие подобные боковые цепи. В данной области определены семейства аминокислотных остатков, содержащих подобные боковые цепи. Указанные семейства включают в себя аминокислоты, содержащие основные боковые цепи (например, лизин, аргинин, гистидин), кислотные боковые цепи (например, аспарагиновая кислота, глутаминовая кислота), незаряженные полярные боковые цепи (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин, триптофан), неполярные боковые цепи (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин), бета-разветвленные боковые цепи (например, треонин, валин, изолейцин) и ароматические боковые цепи (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, один или несколько аминокислотных остатков в молекуле, такой как антитело или фрагмент антитела настоящего изобретения, можно заменить другими аминокислотными остатками из того же семейства боковых цепей, и измененную молекулу можно подвергнуть тестированию с использованием описанных здесь функциональных анализов.
[0067] Термин "гомологичный" или "идентичный" относится к субъединичной идентичности последовательностей двух полимерных молекул, например двух молекул нуклеиновых кислот, таких как две молекулы ДНК или две молекулы РНК, или двух полипептидных молекул. Если субъединичное положение в обеих молекулах занято одинаковыми мономерными субъединицами; например, если положение в каждой из двух молекул ДНК занято аденином, то они являются гомологичными или идентичными по данному положению. Гомология двух последовательностей является прямой функцией от числа совпадающих или гомологичных положений; например, если половина (например, пять положений в полимере длиной 10 субъединиц) положений в двух последовательностях являются гомологичными, две последовательности гомологичны на 50%; если 90% положений (например, 9 из 10) совпадают или гомологичны, две последовательности являются гомологичными на 90%. Процент "идентичности последовательностей" можно определить путем сравнения двух оптимально выровненных последовательностей в окне сравнения, где фрагмент аминокислотной последовательности в окне сравнения может содержать добавления или делеции (например, гэпы или выступы) по сравнению с исходной последовательностью (которая не содержит добавления или делеции), обеспечивающие оптимальное выравнивание двух последовательностей. Чтобы рассчитать процент, определяют число положений, в которых обе последовательности содержат одинаковые аминокислотные остатки, с получением числа совпадающих положений, делят число совпадающих положений на общее число положений в окне сравнения и умножают результат на 100, получая процент идентичности последовательностей. Результат представляет собой процент идентичности исследуемой последовательности и запрашиваемой последовательности.
[0068] Термин "выделенный" означает изменение природного состояния или удаление из природного состояния. Например, нуклеиновая кислота или пептид, присутствующие в природе в живом животном, не являются "выделенными", но та же нуклеиновая кислота, или тот же пептид, частично или полностью отделенные от веществ, сосуществующих с ними в природе, являются "выделенными". Выделенная нуклеиновая кислота или выделенный белок может существовать в практически очищенной форме, или он может находиться в неприродной среде, такой как, например, клетка-хозяин. Выделенное антитело практически не содержит других антител, имеющих другие антигенные специфичности (например, выделенное антитело, которое специфически связывается с TSLP, практически не содержит антител, специфически связывающихся с антигенами, отличными от TSLP). Однако выделенное антитело, способное специфически связывать молекулу-мишень, может обладать перекрестной реакционноспособностью по отношению к тем же антигенам из других видов, например, выделенное антитело, способное специфически связывать человеческий TSLP, может связывать молекулы TSLP, полученные из других видов. Кроме того, выделенное антитело может практически не содержать другой клеточный материал и/или другие химические вещества.
[0069] В некоторых вариантах сухой порошкообразный состав настоящего изобретения состоит из частиц ядро-оболочка, которые содержат: образующее оболочку вспомогательное вещество и ядро, содержащее API, стеклообразующие вспомогательные вещества и буфер, и иногда также упоминается здесь как платформенный состав, или платформенный состав с ядром и оболочкой.
[0070] В данном описании термины "активный ингредиент", "терапевтически активный ингредиент", "активное средство", "лекарственное средство" или "лекарственное вещество" относятся к активному ингредиенту фармацевтического препарата, также известному как активный фармацевтический ингредиент (API).
[0071] Используемый здесь термин "массовый медианный диаметр", или "MMD", или "×50", относится к медианному диаметру совокупности частиц, обычно из полидисперсной популяции частиц, то есть популяции, характеризующейся определенным диапазоном размеров частиц. Указанные здесь значения MMD определяют методом лазерной дифракции (Sympatec Helos, Clausthal-Zellerfeld, Germany), если контекст не указывает иначе. В отличие от MMD dg обозначает геометрический диаметр одной частицы.
[0072] Термин "насыпная плотность", или ρtapped, в соответствии с данным описанием относится к плотности частиц, измеряемой по способу I, описанному, например, в www.usp.org/sites/default/files/usp_pdf/EN USPNF/revisions/m99375 - bulk_density_and_tapped_density_of_powders.pdf. Значения насыпной плотности соответствуют максимальному приближению плотности частиц, причем измеренные значения примерно на 20% меньше фактической плотности частиц.
[0073] В данном описании термин "шероховатый" относится к наличию многочисленных морщинок или складок, то есть к изборожденной или морщинистой поверхности.
[0074] Используемый здесь термин "шероховатость" является мерой неровности поверхности рекомбинантной частицы. В целях настоящего изобретения шероховатость рассчитывают исходя из удельной площади поверхности, полученной путем измерений BET, истинной плотности, определенной методом гелиевой пикнометрии, путем вычисления отношения поверхности к объему, определенному методом лазерной дифракции (Sympatec), а именно:
Шероховатость=(SSA-ρtrue)/Sv
где Sv=6/D32, где D32 - средний диаметр, определенный на основании удельной площади поверхности. Полагают, что увеличение шероховатости поверхности может приводить к уменьшению сил сцепления между частицами и улучшению направленности аэрозоля на легкие. Ожидается, что улучшение направленности на легкие приведет к снижению вариабельности среди пациентов и уменьшению уровней лекарственного средства в ротоглотке и системном кровотоке. В одном или нескольких вариантах осуществления шероховатость Sv составляет от 3 до 20, например от 5 до 10.
[0075] Используемый здесь термин "средний аэродинамический диаметр первичных частиц", или Da, рассчитываемый исходя из первичного геометрического размера частиц, определяемого с помощью лазерной дифракции (×50), и их насыпной плотности, а именно:
Da=×50 (ρtapped)1/2.
[0076] Используемый здесь термин "доставляемая доза", или "DD", относится к показанию подачи сухого порошка из ингалятора после приведения в действие порошкового устройства или распыления из порошкового устройства. DD определяют как отношение дозы, доставляемой ингалятором, к номинальной или отмеренной дозе. DD представляет собой экспериментально определяемый параметр, который можно определить с использованием настроек устройства in vitro, которые имитируют введение дозы пациенту.
[0077] Используемый здесь термин "массовый медианный аэродинамический диаметр", или "MMAD", относится к среднему аэродинамическому размеру совокупности частиц, обычно из полидисперсной популяции. "Аэродинамический диаметр" представляет собой диаметр сферы единичной плотности, характеризующейся такой же скоростью осаждения, обычно в воздухе, как и порошок, и, следовательно, является полезной характеристикой аэрозольного порошка, или других дисперсных частиц, или композиции, состоящей из частиц, описывающей характер осаждения. Распределение частиц по аэродинамическим размерам (APSD) и MMAD определяют здесь методом каскадного импактора с использованием NEXT GENERATION IMPACTORTM. Как правило, если частицы имеют слишком большие аэродинамические размеры, то глубоких отделов легкого будет достигать незначительное число частиц. Если частицы слишком малы, большой процент частиц может выдыхаться. В отличие от MMAD da представляет собой аэродинамический диаметр одной частицы.
[0078] Используемый здесь термин "суммарная доза, попадающая в легкое" (TLD) относится к проценту активного ингредиента (ингредиентов), который не осаждается в модели идеализированного рта-горла Альберта после ингаляции порошка из сухого порошкового ингалятора при перепаде давления 4 кПа. Данные можно выразить в процентах от номинальной дозы или доставляемой дозы. AIT представляет собой идеализированную версию верхних дыхательных путей среднего взрослого индивидуума. Если не указано иное, TLD измеряют на модели идеализированного горла Альберта. Информацию о AIT и подробное описание экспериментальной установки можно найти в: www.copleyscientific.com.
[0079] Используемый здесь термин "инерционный параметр" относится к параметру, который характеризует инерционное воздействие на верхние дыхательные пути. Параметр определяют на основании закона Стокса по формуле da2Q, где da обозначает аэродинамический диаметр, а Q - объемную скорость потока.
[0080] Используемый здесь термин "содержание твердых веществ" относится к концентрации активного ингредиента (ингредиентов) и вспомогательных веществ, растворенных или диспергированных в жидком растворе или дисперсии, предназначенных для сушки распылением.
[0081] Используемый здесь термин "ALR" относится к параметру процесса, определяющему отношение воздуха к жидкости, используемому в распылителе. Меньшие значения ALR обычно соответствуют более крупным распыленным каплям.
[0082] Используемый здесь термин "плотность популяции частиц" (PPD) относится к безразмерному числу, рассчитанному по содержанию твердых веществ и скорости потока распыляемой жидкости, деленному на общий расход газа осушителя. Обнаружено, что PPD коррелирует с первичным геометрическим размером частиц.
TSLP-связывающие молекулы
[0083] Настоящее изобретение предлагает молекулы, например, антитела или фрагменты антител, включающие в себя фрагменты Fab, Fab', F(ab')2, Fd, Fv и dAb, scFv, однодоменные антитела, макситела, минитела, интратела, диатела, триатела, тетратела, v-NAR и бис-SCFv, способные специфически связывать TSLP и ингибировать активность TSLP. Указанные молекулы можно использовать для лечения воспалительных состояний, связанных с TSLP, таких как астма и хроническая обструктивная болезнь легких. Поскольку TSLP является ключевым узловым цитокином, вышестоящим по отношению к эффекторным цитокинам Th2, ингибирование TSLP может одновременно блокировать несколько нижестоящих эффекторов Th2 (например, IL-4, IL-5, IL-13) и, кроме того, может влиять на пути, не опосредованные Th2 (например, IL-17, IFN-γ).
Антитела против TSLP и TSLP-связывающие фрагменты антител
[0084] В некоторых вариантах осуществления настоящее изобретение предлагает антитела и фрагменты антител, способные специфически связываться с человеческим TSLP. Антитела против TSLP и фрагменты таких антител включают в себя, без ограничения, человеческие и гуманизированные моноклональные антитела и фрагменты антител, полученные с помощью способов, описанных в данном документе, в том числе в примерах. В некоторых вариантах осуществления настоящее изобретение относится к выделенному антителу или его антигенсвязывающему фрагменту, способному связывать человеческий TSLP с константой диссоциации (KD) менее 100 пМ, например, с KD менее 90 пМ, менее 80 пМ, менее чем 70 пМ, менее 60 пМ, менее 50 пМ, менее 40 пМ, менее 30 пМ, менее 20 пМ, менее 10 пМ. В некоторых вариантах осуществления описанные здесь выделенные антитела или антигенсвязывающие фрагменты связывают человеческий TSLP с константой диссоциации (KD) менее 10 пМ.
[0085] В некоторых вариантах осуществления описанные здесь TSLP-связывающие молекулы содержат CDR1 тяжелой цепи, CDR2 тяжелой цепи, CDR3 тяжелой цепи и CDR1 легкой цепи, CDR2 легкой цепи и CDR3 легкой цепи. В некоторых вариантах осуществления описанные здесь TSLP-связывающие молекулы содержат вариабельный участок тяжелой цепи, содержащий CDR1, CDR2 и CDR3, и вариабельный участок легкой цепи, содержащий CDR1, CDR2 и CDR3. В некоторых вариантах осуществления описанные здесь TSLP-связывающие молекулы содержат полноразмерную последовательность тяжелой цепи и полноразмерную последовательность легкой цепи. В некоторых вариантах осуществления молекула представляет собой TSLP-связывающий Fab.
[0086] В таблице 2 приведены последовательности примеров TSLP-связывающих антител и Fab, которые все связываются с человеческим TSLP с высоким сродством. Например, Fab1 против TSLP связывается с рекомбинантным человеческим TSLP с константой диссоциации (KD) 6 пМ. В некоторых вариантах осуществления Fab1 против TSLP связывается с белками TLSP человека и яванского макака со значениями KD 5,0±2,0 пМ и 1,4±0,6 пМ соответственно.
ТАБЛИЦА 2. Аминокислотные последовательности Fab и антител против TSLP
[0087] В некоторых вариантах осуществления антитела содержат CDR VH, имеющий аминокислотную последовательность любого из CDR VH, перечисленных в таблице 2. В частности, изобретение предлагает антитела, способные специфически связываться с белком TSLP, причем указанные антитела содержат (или, альтернативно, состоят из) один, два, три, четыре, пять или шесть CDR VH, имеющих аминокислотную последовательность любого из CDR VH, перечисленных в таблице 2. Настоящее изобретение также относится к антителам, способным специфически связываться с белком TSLP, где указанные антитела содержат CDR VL, имеющий аминокислотную последовательность любого из CDR VL, перечисленных в таблице 2. В частности, изобретение относится к антителам, способным специфически связываться с белком TSLP, где указанные антитела содержат (или, альтернативно, состоят из) один, два, три, четыре, пять или шесть CDR VL, имеющих аминокислотную последовательность любого из CDL VL, перечисленных в таблице 2.
[0088] Изобретение также относится к антителам и их антигенсвязывающим фрагментам, содержащим (или альтернативно, состоящим из) аминокислотную последовательность VH, приведенную в таблице 2, где не более, чем примерно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 аминокислот в каркасной последовательности (например, последовательности, которая не является CDR) подвергнуты мутациям (где мутации, без ограничения, могут представлять собой добавления, замены или удаления).
[0089] Изобретение также относится к антителам и их антигенсвязывающим фрагментам, способным специфически связываться с TSLP, где указанные антитела или их антигенсвязывающие фрагменты содержат (или, альтернативно, состоят из) аминокислотную последовательность VL, приведенную в таблице 2, где не более, чем примерно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 аминокислот в каркасной последовательности (например, последовательности, которая не является CDR) подвергнуты мутациям (где мутации, без ограничения, могут представлять собой добавления, замены или удаления).
[0090] Другие антитела настоящего изобретения и их антигенсвязывающие фрагменты содержат аминокислоты, подвергшиеся мутациям, но обладают по меньшей мере 60, 70, 80, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99 процентами идентичности по участкам CDR, по сравнению с участками CDR, последовательности которых приведены в таблице 2, и могут связываться с TSLP. В одном аспекте другие антитела настоящего изобретения и их антигенсвязывающие фрагменты содержат мутантные аминокислотные последовательности, причем в участках CDR мутациям подвергают не более 1, 2, 3, 4 или 5 аминокислот по сравнению с участками CDR, последовательности которых приведены в таблице 2.
[0091] Настоящее изобретение также относится к нуклеотидным последовательностям, которые кодируют VH, VL, полноразмерную тяжелую цепь и полноразмерную легкую цепь антител и их антигенсвязывающих фрагментов, способных специфически связываться с белком TSLP. Такие нуклеотидные последовательности можно оптимизировать для экспрессии в клетках млекопитающих.
[0092] Другие антитела против TSLP и их антигенсвязывающие фрагменты включают в себя молекулы, в которых аминокислоты или нуклеиновые кислоты, кодирующие аминокислоты, были подвергнуты мутациям, но которые обладают по меньшей мере 60, 70, 80, 90 или 95 процентами идентичности в отношении последовательностей, описанных в таблице 2. В одном варианте осуществления антитела и их антигенсвязывающие фрагменты содержат мутантные аминокислотные последовательности, в вариабельных участках которых мутациям подвергнуты не более 1, 2, 3, 4 или 5 аминокислот по сравнению с вариабельными участками, последовательности которых описаны в таблице 2, и сохраняют при этом практически такую же терапевтическую активность.
[0093] Поскольку каждое из описанных здесь антител может связываться с TSLP, последовательности VH, VL, полноразмерной легкой цепи и полноразмерной тяжелой цепи (аминокислотные последовательности и нуклеотидные последовательности, кодирующие аминокислотные последовательности) можно "смешивать и спаривать" с получением других TSLP-связывающих антител настоящего изобретения и их антигенсвязывающих фрагментов. Полученные в результате "смешивания и спаривания" TSLP-связывающие антитела можно тестировать с использованием анализов связывания, известных в данной области (таких как ELISA и другие анализы, описанные в разделе "Примеры"). Когда указанные цепи смешивают и спаривают, последовательность VH из конкретной пары VH/VL заменяют на структурно подобную последовательностью VH. Подобным образом, полноразмерную последовательность тяжелой цепи из конкретной пары полноразмерная тяжелая цепь/полноразмерная легкая цепь заменяют на структурно подобную полноразмерную тяжелую цепь. Подобным образом, последовательность VL из конкретной пары VH/VL заменяют структурно подобную последовательностью VL. Аналогично полноразмерную последовательность легкой цепи из конкретной пары полноразмерная тяжелая цепь/полноразмерная легкая цепь заменяют на структурно подобную полноразмерную легкую цепь.
[0094] В другом аспекте настоящее изобретение относится к TSLP-связывающим антителам, которые содержат CDR1, CDR2 и CDR3 тяжелой цепи и легкой цепи, описанные в таблице 2, или их сочетания. Участки CDR определяют по системе Кабат (Kabat et al. 1991 Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242) или по системе Чотиа (Chothia et al. al., 1987 J. Mol. Biol. 196: 901-917; и Al-Lazikani et al. 1997 J. Mol. Biol. 273: 927-948). В качестве альтернативы можно использовать другие способы определения участков CDR. Например, участки CDR можно определять по объединенной системе Кабат и Чотиа.
[0095] Поскольку каждое из указанных антител может связываться с TSLP, а антигенсвязывающая специфичность обеспечивается в основном участками CDR1, 2 и 3, последовательности CDR1, 2 и 3 VH и последовательности CDR1, 2 и 3 VL можно "смешивать и спаривать" (т.е. можно смешивать и спаривать CDR из разных антител, но каждое антитело должно содержать CDR1, 2 и 3 VH и CDR1, 2 и 3 VL, чтобы обеспечить создание других TSLP-связывающих молекул настоящего изобретения). Полученные в результате "смешивания и спаривания" TSLP-связывающие антитела можно тестировать с использованием анализов связывания, известных в данной области и описанных в разделе "Примеры" (таких как ELISA). Когда смешивают и спаривают последовательности CDR VH, последовательности CDR1, CDR2 и/или CDR3 из конкретной последовательности VH заменяют на структурно подобные последовательности CDR. Аналогично, когда смешивают и спаривают последовательности CDR VL, последовательность CDR1, CDR2 и/или CDR3 из конкретной последовательности VL заменяют на структурно подобные последовательности CDR. Для рядового специалиста в данной области должно быть очевидно, что новые последовательности VH и VL можно получить путем изменения последовательностей одного или нескольких участков CDR VH и/или VL на структурно подобные последовательности по сравнению с последовательностями CDR, описанными здесь для моноклональных антител настоящего изобретения.
[0096] Соответственно, настоящее изобретение относится к выделенному моноклональному антителу или его антигенсвязывающему фрагменту, содержащему CDR1 вариабельного участка тяжелой цепи (HCDR1), аминокислотная последовательность которого выбрана из последовательностей SEQ ID NO: 1, 4 или 5; CDR2 вариабельного участка тяжелой цепи (HCDR2), аминокислотная последовательность которого выбрана из последовательностей SEQ ID NO: 2 или 6; CDR3 вариабельного участка тяжелой цепи (HCDR3) с аминокислотной последовательностью SEQ ID NO: 3; CDR1 вариабельного участка легкой цепи (LCDR1), аминокислотная последовательность которого выбрана из последовательностей SEQ ID NO: 11 или 14; CDR2 вариабельного участка легкой цепи (LCDR2), аминокислотная последовательность которого выбрана из последовательностей SEQ ID NO: 12 или 15; и CDR3 вариабельного участка легкой цепи (LCDR3), аминокислотная последовательность которого выбрана из последовательностей SEQ ID NO: 13 или 16; где антитело или фрагмент антитела специфически связывают TSLP.
[0097] В некоторых вариантах осуществления антитело или фрагмент антитела, способный специфически связываться с TSLP, представляет собой антитело или фрагмент антитела, описанный в таблице 2.
[0098] В некоторых вариантах осуществления настоящее изобретение относится к выделенному антителу или его антигенсвязывающему фрагменту, способному связывать человеческий TSLP и содержащему последовательности HCDR1, HCDR2 и HCDR3, описанные в SEQ ID NO: 4, 2 и 3 соответственно, и последовательности LCDR1, LCDR2 и LCDR3, описанные в SEQ ID NO: 11, 12 и 13 соответственно.
[0099] В некоторых вариантах осуществления настоящее изобретение относится к выделенному антителу или его антигенсвязывающему фрагменту, способному связывать человеческий TSLP и содержащему последовательности HCDR1, HCDR2 и HCDR3, описанные в SEQ ID NO: 5, 6 и 3 соответственно, и последовательности LCDR1, LCDR2 и LCDR3, описанные в SEQ ID NO: 14, 15 и 16 соответственно.
[00100] В некоторых вариантах осуществления настоящее изобретение относится к выделенному антителу или его антигенсвязывающему фрагменту, способному связывать человеческий TSLP и содержащему последовательности HCDR1, HCDR2 и HCDR3, описанные в SEQ ID NO: 1, 2 и 3 соответственно, и последовательности LCDR1, LCDR2 и LCDR3, описанные в SEQ ID NO: 11, 12 и 13 соответственно.
[00101] В некоторых вариантах осуществления настоящее изобретение относится к выделенному антителу или его антигенсвязывающему фрагменту, способному связывать человеческий TSLP и содержащему VH с аминокислотной последовательностью SEQ ID NO: 7 и VL с аминокислотной последовательностью SEQ ID NO: 17.
[00102] В некоторых вариантах осуществления настоящее изобретение относится к выделенному антителу или его антигенсвязывающему фрагменту, способному связывать человеческий TSLP и содержащему тяжелую цепь с аминокислотной последовательностью SEQ ID NO: 22 и легкую цепь с аминокислотной последовательностью SEQ ID NO: 25.
[00103] В некоторых вариантах осуществления настоящее изобретение относится к выделенному антителу или его антигенсвязывающему фрагменту, способному связывать человеческий TSLP и содержащему тяжелую цепь с аминокислотной последовательностью SEQ ID NO: 9 и легкую цепь с аминокислотной последовательностью SEQ ID NO: 19.
[00104] В соответствии с настоящим описанием, человеческое антитело содержит вариабельные участки тяжелых или легких цепей или полноразмерные тяжелые или легкие цепи, которые являются "продуктом", или которые "получены из" конкретной зародышевой последовательности, если вариабельные участки или полноразмерные цепи антитела получены с использованием системы, в которой применяются гены человеческих зародышевых иммуноглобулинов. Такие системы включают в себя иммунизацию трансгенной мыши, несущей гены человеческих иммуноглобулинов, представляющим интерес антигеном, или скрининг отображенной на фаге библиотеки генов человеческих иммуноглобулинов с использованием представляющего интерес антигена. Человеческое антитело, которое являются "продуктом", или которое "получено из" последовательности человеческого зародышевого иммуноглобулина, можно идентифицировать как таковое путем сравнения аминокислотной последовательности человеческого антитела с аминокислотными последовательностями человеческих зародышевых иммуноглобулинов и выбора последовательности человеческого зародышевого иммуноглобулина, наиболее близкой (то есть, обладающей наибольшим % идентичности) к последовательности человеческого антитела. Человеческое антитело, которое является "продуктом", или которое "получено из" конкретной последовательности человеческого зародышевого иммуноглобулина, может содержать аминокислотные отличия по сравнению с зародышевой последовательностью, например, вследствие естественных соматических мутаций или преднамеренного введения сайт-направленных мутаций. Однако аминокислотные последовательности каркасных участков VH или VL выбранного человеческого антитела обычно по меньшей мере на 90% идентичны аминокислотной последовательности, кодируемой геном человеческого иммуноглобулина, и содержат аминокислотные остатки, которые позволяют идентифицировать антитело как человеческое по сравнению с аминокислотными последовательностями зародышевых иммуноглобулинов других видов (таких как мышиные зародышевые последовательности). В некоторых случаях аминокислотная последовательность человеческого антитела может быть по меньшей мере на 60%, 70%, 80%, 90%, или по меньшей мере на 95%, или даже по меньшей мере на 96%, 97%, 98% или 99% идентичной аминокислотной последовательности, кодируемой геном зародышевого иммуноглобулина. Как правило, рекомбинантное человеческое антитело в каркасных участках VH или VL имеет не более 10 аминокислотных отличий от аминокислотной последовательности, кодируемой геном человеческого зародышевого иммуноглобулина. В некоторых случаях человеческое антитело может отличаться от аминокислотной последовательности, кодируемой геном зародышевого иммуноглобулина, не более, чем по 5, или даже не более, чем по 4, 3, 2 или 1 аминокислотам.
Гомологичные антитела
[00105] В следующем варианте осуществления настоящее изобретение относится к антителу или его антигенсвязывающему фрагменту, содержащему аминокислотные последовательности, которые являются гомологичными последовательностям, описанным в таблице 2, причем указанное антитело связывается с TSLP и сохраняет желательные функциональные свойства антител, описанных в таблице 2.
[00106] Например, изобретение предлагает выделенное моноклональное антитело (или его антигенсвязывающий фрагмент), содержащее вариабельный участок тяжелой цепи (VH) и вариабельный участок легкой цепи (VL), где VH содержит аминокислотную последовательность, которая по меньшей мере на 80%, по меньшей мере на 90% или по меньшей мере на 95% идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 7; VL содержит аминокислотную последовательность, которая по меньшей мере на 80%, по меньшей мере на 90%, или по меньшей мере на 95% идентична аминокислотной последовательности, выбранной из группы, состоящей из SEQ ID NO: 17; антитело специфически связывается с белком TSLP и ингибирует TSLP.
[00107] В одном варианте осуществления аминокислотные последовательности VH и/или VL могут быть на 50%, 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентичны последовательностям, приведенным в таблице 2. В одном варианте осуществления аминокислотные последовательности VH и/или VL могут быть идентичны за исключением аминокислотных замен не более чем в 1, 2, 3, 4 или 5 аминокислотных положениях. Антитело, содержащее участки VH и VL, обладающие высокой степенью (т.е. 80% или более) идентичности по отношению к участкам VH и VL, описанным в таблице 2, можно получить путем мутагенеза (такого как сайт-направленный или ПЦР-опосредованный мутагенез) молекулы нуклеиновой кислоты, кодирующей SEQ ID NO: 8 или 21, или SEQ ID NO: 18 или 24, соответственно, с последующим тестированием кодируемого измененного антитела на сохранение функции с помощью описанных здесь функциональных анализов.
[00108] В одном варианте осуществления аминокислотные последовательности полноразмерной тяжелой цепи и/или полноразмерной легкой цепи могут быть на 50%, 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентичными последовательностям, указанным в таблице 2. Антитело, содержащее полноразмерную тяжелую цепь и полноразмерную легкую цепь, обладающие высокой степенью (т.е. 80% или более) идентичности по отношению к полноразмерной тяжелой цепи SEQ ID NO: 9; и полноразмерной легкой цепи SEQ ID NO: 19, можно получить путем мутагенеза (такого как сайт-направленный или ПЦР-опосредованный мутагенез) молекул нуклеиновой кислоты, кодирующих указанные полипептиды соответственно, с последующим тестированием кодируемого измененного антитела на сохранение функции с помощью описанных здесь функциональных анализов.
[00109] В одном варианте осуществления целые нуклеотидные последовательности полноразмерной тяжелой цепи и/или полноразмерной легкой цепи могут быть на 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентичными последовательностям, описанным в таблице 2.
[00110] В одном варианте осуществления нуклеотидные последовательности вариабельных участков тяжелой цепи и/или легкой цепи могут быть на 60, 70, 80, 90, 95, 96, 97, 98 или 99% идентичными последовательностям, описанным в таблице 2.
[00111] В данном описании процент идентичности двух последовательностей является функцией от числа идентичных положений в последовательностях (то есть, % идентичности равен числу идентичных положений/общее число положений×100) с учетом числа гэпов и длины каждого гэпа, введение которого необходимо для оптимального выравнивания двух последовательностей. Сравнение последовательностей и определение процента идентичности двух последовательностей можно осуществить с помощью математического алгоритма, как описано в приведенных ниже неограничивающих примерах.
[00112] Кроме того, или альтернативно, белковые последовательности настоящего изобретения также можно использовать в качестве "запрашиваемой последовательности" при проведении поиска по открытым базам данных, например, для идентификации родственных последовательностей. Например, такой поиск можно проводить с помощью программы BLAST (версия 2.0) Altschul et al., 1990 J. Mol. Biol. 215:403-10.
Антитела, содержащие консервативными модификации
[00113] В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент настоящего изобретения содержит вариабельный участок тяжелой цепи, содержащий последовательности CDR1, CDR2 и CDR3, и вариабельный участок легкой цепи, содержащий последовательности CDR1, CDR2 и CDR3, где одна или несколько из указанных последовательностей CDR имеют конкретные аминокислотные последовательности, входящие в состав описанных здесь антител, или их консервативные модификации, и где антитела сохраняют желательные функциональные свойства TSLP-связывающих антител настоящего изобретения и их антигенсвязывающих фрагментов. Соответственно, изобретение предлагает выделенное моноклональное антитело или его антигенсвязывающий фрагмент, состоящий из вариабельного участка тяжелой цепи, содержащего последовательности CDR1, CDR2 и CDR3, и вариабельного участка легкой цепи, содержащего последовательности CDR1, CDR2 и CDR3, где: CDR1 вариабельного участка тяжелой цепи содержит аминокислотную последовательность, выбранную из последовательностей SEQ ID NO: 1, 4 или 5, или их консервативных вариантов; CDR2 вариабельного участка тяжелой цепи содержит аминокислотную последовательность, выбранную из последовательностей SEQ ID NO: 2 или 6, или их консервативных вариантов; CDR3 вариабельного участка тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 3 или ее консервативный вариант; CDR1 вариабельного участка легкой цепи содержит аминокислотную последовательность, выбранную из последовательностей SEQ ID NO: 11 или 14, или их консервативных вариантов; CDR2 вариабельного участка легкой цепи содержит аминокислотную последовательность, выбранную из последовательностей SEQ ID NO: 12 или 15, или их консервативных вариантов; и CDR3 вариабельного участка легкой цепи содержит аминокислотную последовательность, выбранную из последовательностей SEQ ID NO: 13 или 16, или их консервативных вариантов; антитело или его антигенсвязывающий фрагмент специфически связываются с TSLP и ингибируют TSLP.
[00114] В некоторых вариантах осуществления антитело настоящего изобретения или его антигенсвязывающий фрагмент содержит вариабельный участок тяжелой цепи и вариабельный участок легкой цепи, где вариабельные участки тяжелой и легкой цепей имеют определенные аминокислотные последовательности, входящие в состав описанных здесь антител, или их консервативные модификации, где антитела сохраняют желательные функциональные свойства TSLP-связывающих антител настоящего изобретения и их антигенсвязывающих фрагментов. Соответственно, изобретение предлагает выделенное моноклональное антитело, или его антигенсвязывающий фрагмент, состоящие из вариабельного участка тяжелой цепи и вариабельного участка легкой цепи, где: вариабельный участок тяжелой цепи содержит аминокислотную последовательность SEQ ID NO: 7, или ее консервативный вариант; вариабельный участок легкой цепи содержит аминокислотную последовательность SEQ ID NO: 17, или ее консервативный вариант; антитело или его антигенсвязывающий фрагмент специфически связываются с TSLP и ингибируют TSLP.
Антитела, которые связываются с одним эпитопом
[00115] Настоящее изобретение относится к антителам, которые связываются с тем же эпитопом, что и TSLP-связывающие антитела или фрагменты антител, перечисленные в таблице 2. Следовательно, можно идентифицировать другие антитела по их способности конкурировать (например, конкурентно ингибировать связывание статистически значимым образом) с другими антителами и антигенсвязывающими фрагментами настоящего изобретения в анализах связывания TSLP. Способность тестируемого антитела ингибировать связывание антител настоящего изобретения и их антигенсвязывающих фрагментов с белком TSLP демонстрирует, что тестируемое антитело может конкурировать с этим антителом за связывание с TSLP; такое антитело может в соответствии с неограничивающей теорией связываться с тем же или родственным (например, структурно подобным или пространственно приближенным) эпитопом на TSLP, что и антитело, с которым оно конкурирует. В некоторых вариантах осуществления антитело, которое связывается с тем же эпитопом на TSLP, что и описанное здесь антитело или его антигенсвязывающий фрагмент, представляет собой человеческое моноклональное антитело. Такие человеческие моноклональные антитела можно получить и выделить с помощью описанных здесь способов. В некоторых вариантах осуществления антитело, которое связывается с тем же эпитопом на TSLP, что и антитело настоящего изобретения, или его антигенсвязывающий фрагмент, представляет собой моноклональное мышиное антитело. В некоторых вариантах осуществления антитело, которое связывается с тем же эпитопом на TSLP, что и описанное здесь антитело, или его антигенсвязывающий фрагмент, представляет собой гуманизированное моноклональное антитело, полученное из моноклонального мышиного антитела. В конкретном варианте осуществления антитело, которое связывается с тем же эпитопом на TSLP, что и описанное здесь антитело, или его антигенсвязывающий фрагмент, представляет собой гуманизированное моноклональное антитело. Такое гуманизированное моноклональное антитело можно получить и выделить с помощью описанных здесь способов.
[00116] В некоторых вариантах осуществления описанное здесь моноклональное антитело, или его антигенсвязывающий фрагмент, специфически связывает эпитоп в человеческом TSLP, где эпитоп содержит один или несколько из следующих остатков: Lys38, Ala41, Leu44, Ser45, Thr46, Ser48, Lys49, Ile52, Thr53, Ser56, Gly57, Thr58, Lys59, Lys101, Gln145 и Arg149 последовательности SEQ ID NO: 38. В некоторых вариантах осуществления описанное здесь моноклональное антитело, или его антигенсвязывающий фрагмент, специфически связывает эпитоп в человеческом TSLP, где эпитоп содержит по меньшей мере один, по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть, по меньшей мере семь, по меньшей мере восемь, по меньшей мере девять, по меньшей мере десять, по меньшей мере одиннадцать, по меньшей мере двенадцать, по меньшей мере тринадцать, по меньшей мере четырнадцать, по меньшей мере пятнадцать, или все из следующих остатков: Lys38, Ala41, Leu44, Ser45, Thr46, Ser48, Lys49, Ile52, Thr53, Ser56, Gly57, Thr58, Lys59, Lys101, Gln145 и Arg149 последовательности SEQ ID NO: 38.
[00117] В некоторых вариантах осуществления описанное здесь моноклональное антитело, или его антигенсвязывающий фрагмент, специфически связывает эпитоп в человеческом TSLP, где эпитоп содержит по меньшей мере один, по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть, по меньшей мере семь, по меньшей мере восемь, или все из следующих остатков: Lys38, Ala41, Leu44, Ser45, Thr46, Ser48, Lys49, Ile52 и Thr53 последовательности SEQ ID NO: 38. Эпитоп такого моноклонального антитела, или его антигенсвязывающего фрагмента, может также содержать один или несколько из следующих остатков: Ser56, Gly57, Thr58, Lys59, Lys101, Gln145 и Arg149 последовательности SEQ ID NO: 38.
[00118] В некоторых вариантах описанное здесь осуществления моноклональное антитело, или его антигенсвязывающий фрагмент, специфически связывает эпитоп в человеческом TSLP, где эпитоп содержит по меньшей мере один, по меньшей мере два, по меньшей мере три, или все из следующих остатков: Ser56, Gly57, Thr58 и Lys59 последовательности SEQ ID NO: 38. Эпитоп такого моноклонального антитела, или его антигенсвязывающего фрагмента, может также содержать один или несколько из следующих остатков: Lys38, Ala41, Leu44, Ser45, Thr46, Ser48, Lys49, Ile52, Thr53, Lys101, Gln145 и Arg149 последовательности SEQ ID NO: 38.
[00119] В некоторых вариантах осуществления описанное здесь моноклональное антитело, или его антигенсвязывающий фрагмент, специфически связывает эпитоп в человеческом TSLP, где эпитоп содержит Lys101 последовательности SEQ ID NO: 38. Эпитоп такого моноклонального антитела, или его антигенсвязывающего фрагмента, может также содержать один или несколько из следующих остатков: Lys38, Ala41, Leu44, Ser45, Thr46, Ser48, Lys49, Ile52, Thr53, Ser56, Gly57, Thr58, Lys59, Gln145 и Arg149 последовательности SEQ ID NO: 38.
[00120] В некоторых вариантах осуществления описанное здесь моноклональное антитело, или его антигенсвязывающий фрагмент, специфически связывает эпитоп в человеческом TSLP, где эпитоп содержит Gln145 или Arg149 последовательности SEQ ID NO: 38. В некоторых вариантах осуществления описанное здесь моноклональное антитело, или его антигенсвязывающий фрагмент, специфически связывает эпитоп в человеческом TSLP, где эпитоп содержит Gln145 и Arg149 последовательности SEQ ID NO: 38. Эпитоп такого моноклонального антитела, или его антигенсвязывающего фрагмента, может также содержать один или несколько из следующих остатков: Lys38, Ala41, Leu44, Ser45, Thr46, Ser48, Lys49, Ile52, Thr53, Ser56, Gly57, Thr58, Lys59, и Lys101 последовательности SEQ ID NO: 38.
[00121] В некоторых вариантах осуществления описанное здесь моноклональное антитело, или его антигенсвязывающий фрагмент, специфически связывает эпитоп в человеческом TSLP, где эпитоп содержит по меньшей мере один, по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть, или все из следующих остатков: Lys49, Ile52, Gly57, Lys59, Lys101, Gln145 и Arg149 последовательности SEQ ID NO: 38. В некоторых вариантах осуществления описанное здесь моноклональное антитело, или его антигенсвязывающий фрагмент, специфически связывает эпитоп в человеческом TSLP, где эпитоп содержит все следующие остатки: Lys49, Ile52, Gly57, Lys59, Lys101, Gln145 и Arg149 последовательности SEQ ID NO: 38.
[00122] В некоторых вариантах осуществления описанное здесь моноклональное антитело, или его антигенсвязывающий фрагмент, специфически связывает эпитоп в человеческом TSLP, где эпитоп содержит, по меньшей мере, один из следующих наборов остатков SEQ ID NO: 38: (a) Lys49 и Ile52, (b) Gly57 и Lys59, (c) Lys101, (d) Gln145 и Arg149. В некоторых вариантах осуществления описанное здесь моноклональное антитело, или его антигенсвязывающий фрагмент, специфически связывает эпитоп в человеческом TSLP, где эпитоп содержит Lys49 и Ile52 последовательности SEQ ID NO: 38. В некоторых вариантах осуществления описанное здесь моноклональное антитело, или его антигенсвязывающий фрагмент, специфически связывает эпитоп в человеческом TSLP, где эпитоп содержит Gly57 и Lys59 последовательности SEQ ID NO: 38. В некоторых вариантах осуществления описанное здесь моноклональное антитело, или его антигенсвязывающий фрагмент, специфически связывает эпитоп в человеческом TSLP, где эпитоп содержит Lys101 последовательности SEQ ID NO: 38. В некоторых вариантах осуществления описанное здесь моноклональное антитело, или его антигенсвязывающий фрагмент, специфически связывает эпитоп в человеческом TSLP, где эпитоп содержит Gln145 и Arg149 последовательности SEQ ID NO: 38.
[00123] В некоторых вариантах осуществления TSLP-связывающие молекулы могут содержать паратоп, содержащий по меньшей мере один, по меньшей мере два, по меньшей мере три, по меньшей мере четыре, по меньшей мере пять, по меньшей мере шесть, по меньшей мере семь, по меньшей мере восемь, по меньшей мере девять, по меньшей мере 10, по меньшей мере 11, по меньшей мере 12, по меньшей мере 13, по меньшей мере 14, по меньшей мере 15, по меньшей мере 16, по меньшей мере 17, по меньшей мере 18, по меньшей мере 19, или все из следующих остатков: Thr28, Asp31, Tyr32, Trp33, Asp56, Glu101, Ile102, Tyr103, Tyr104, Tyr105 последовательности тяжелой цепи SEQ ID NO: 22, или Gly28, Ser29, Lys30, Tyr31, Tyr48, Asp50, Asn51, Glu52, Asn65 и Trp92 последовательности легкой цепи SEQ ID NO: 25.
[00124] После определения целевого эпитопа на антигене можно получить антитела к этому эпитопу, например, с помощью способов, описанных в настоящем изобретении. Альтернативно в процессе получения и характеризации антител может появиться информация о целевых эпитопах. Исходя из этой информации, антитела можно подвергнуть конкурентному скринингу на связывание с тем же эпитопом. С этой целью проводят конкурентные анализы с выявлением антител, способных конкурировать друг с другом за связывание, например, за связывание с антигеном. Способ определения "связывания" антител на основании их способности к конкуренции, характеризующийся высокой пропускной способностью, описан в международной патентной заявке № WO 2003/48731. Как известно специалистам в данной области, практически все, с чем может специфически связываться антитело, может представлять собой эпитоп. Эпитоп может содержать остатки, с которыми связывается антитело.
[00125] Как правило, антитела, специфичные к конкретному антигену-мишени, предпочтительно распознают эпитоп на антигене-мишени в сложной смеси белков и/или макромолекул.
[00126] Участки конкретного полипептида, которые содержат эпитоп, можно идентифицировать с помощью ряда методов картирования эпитопов, хорошо известных в данной области. См., например, Epitope Mapping Protocols in Methods in Molecular Biology, Vol. 66 (Glenn E.Morris, Ed., 1996) Humana Press, Totowa, New Jersey. Например, чтобы определить линейные эпитопы, одновременно синтезируют большое число пептидов на твердых носителях, где пептиды соответствуют фрагментам белковой молекулы, после чего прикрепленные к носителям пептиды подвергают взаимодействию с антителами. Такие способы известны в данной области и описаны, например, в патенте США № 4708871; Geysen et al., (1984) Proc. Natl. Acad. Sci. USA 8:3998-4002; Geysen et al., (1985) Proc. Natl. Acad. Sci. USA 82:78-182; Geysen et al., (1986) Mol. Immunol. 23:709-715. Подобным образом, конформационные эпитопы можно легко идентифицировать путем определения пространственной конформации аминокислот TSLP с помощью таких методов, как, например, водород/дейтериевый обмен, рентгеновская кристаллография и двумерный ядерный магнитный резонанс. См., например, Epitope Mapping Protocols, выше. Антигенные участки белков также можно идентифицировать с помощью стандартных диаграмм антигенности и гидрофобности, испольуя для расчетов, например, программное обеспечение Omiga, версия 1.0, которое можно получить в Oxford Molecular Group. Эта компьютерная программа использует метод Хоппа/Вудса, Hopp et al, (1981) Proc. Natl. Акад. Sci USA 78: 3824-3828; для определения профилей антигенности и метод Кита-Дулитла, Kyte et al, (1982) J. MoI. Biol. 157: 105-132; для построения диаграмм гидрофобности.
Рекомбинантные и модифицированные антитела
[00127] Антитело настоящего изобретения также можно получить путем модификации исходного антитела, содержащего одну или несколько последовательностей VH и/или VL, причем модифицированное антитело может иметь измененные свойства по сравнению с исходным антителом. Рекомбинантное антитело можно получить путем модификации одного или нескольких остатков в одном или обоих вариабельных участках (то есть VH и/или VL), например, в одном или нескольких участках CDR и/или в одном или нескольких каркасных участках. Кроме того, или альтернативно, рекомбинантное антитело можно получить путем модификации остатков в константном участке (константных участках), например, с целью изменения эффекторной функции (эффекторных функций) антитела.
[00128] Одним из рекомбинантных способов получения вариабельных участков является прививка CDR. Антитела взаимодействуют с антигенами-мишенями преимущественно посредством аминокислотных остатков, расположенных в шести определяющих комплементарность участках (CDR) тяжелой и легкой цепей. По этой причине аминокислотные последовательности CDR отдельных антител в большей степени отличаются друг от друга, чем последовательности, находящиеся вне CDR. Поскольку последовательности CDR отвечают за большую часть взаимодействий между антителом и антигеном, можно экспрессировать рекомбинантные антитела, которые имитируют свойства конкретных природных антител, путем конструирования векторов, обеспечивающих экспрессию последовательностей CDR конкретного природного антитела, привитых на каркасные последовательности другого антитела, обладающего другими свойствами (см., например, Riechmann, L. et al., 1998 Nature 332:323-327; Jones, P. et al., 1986 Nature 321:522-525; Queen, C. et al., 1989 Proc. Natl. Acad., U.S.A. 86:10029-10033; патент США № 5225539, Winter, и патенты США №№ 5530101; 5585089; 5693762 и 6180370, Queen et al.)
[00129] Такие каркасные последовательности можно получить из общедоступных баз данных ДНК или опубликованных ссылок, которые содержат последовательности генов зародышевых антител или последовательности перегруппированных антител. Например, зародышевые ДНК-последовательности генов, кодирующих вариабельные участки человеческих тяжелых и легких цепей, можно найти в базе данных человеческих зародышевых последовательностей "VBase" (доступной в Интернете на сайте: www.mrc-cpe.cam.ac.uk/vbase), а также в Kabat, E. A., et al., 1991 Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242; Tomlinson, I. M., et al., 1992 J. fol. Biol. 227: 776-798; and Cox, J. P. L. et al., 1994 Eur. J Immunol. 24: 827-836; содержание каждого из которых в явной форме включено в настоящий документ посредством ссылки. Например, зародышевые ДНК-последовательности генов вариабельных участков человеческих тяжелых и легких цепей, и последовательности перегруппированных антител можно найти в базе данных "IMGT" (доступной в Интернете на сайте www.imgt.org, см. Lefranc, M.P. et al., 1999 Nucleic Acids Res. 27: 209-212; содержание каждого из которых в явной форме включено в настоящий документ посредством ссылки).
[00130] Примеры каркасных последовательностей, подходящих для получения антител настоящего изобретения и их антигенсвязывающих фрагментов, включают в себя последовательности, структурно подобные каркасным последовательностям, используемым выбранными антителами настоящего изобретения и их антигенсвязывающими фрагментами, например, консенсусные последовательности и/или каркасные последовательности, используемые моноклональными антителами настоящего изобретения. Последовательности CDR1, 2 и 3 VH и последовательности CDR1, 2 и 3 VL можно прививать на каркасные участки, последовательности которых идентичны последовательностям зародышевых генов иммуноглобулинов, из которых получают каркасные последовательности, или последовательности CDR можно прививать на каркасные участки, которые содержат одну или несколько мутаций по сравнению с зародышевыми последовательностями. Например, обнаружено, что в некоторых случаях полезно подвергать мутациям остатки в каркасных участках, поскольку такие мутации могут обеспечивать поддержание или усиление антигенсвязывающей способности антитела (см., например, патенты США №№ 5530101; 5585089; 5693762 и 680370, Queen et al).
[00131] Другим типом модификации вариабельного участка является изменение аминокислотных остатков в участках CDR1, CDR2 и/или CDR3 VH и/или VL с целью улучшения одного или нескольких параметров связывания (например, сродства) представляющего интерес антитела, известное как "аффинное созревание". Сайт-направленный мутагенез или ПЦР-опосредованный мутагенез можно использовать для введения мутации (мутаций), а влияние такого мутагенеза на связывание или другое представляющее интерес функциональное свойство антитела можно оценить с помощью анализов in vitro или in vivo, описанных в данном документе и приведенных в разделе Примеры. Можно вводить консервативные модификации (как описано выше). Мутации могут представлять собой аминокислотными замены, добавления или делеции. Кроме того, в участке CDR обычно изменяют не более одного, двух, трех, четырех или пяти остатков.
[00132] Можно использовать широкий ряд каркасных участков или каркасов антител/иммуноглобулинов при условии, что полученный полипептид будет содержать по меньшей мере один связывающий участок, способный специфически связываться с TSLP. Такие каркасные участки или каркасы включают в себя участки человеческих иммуноглобулинов 5 основных идиотипов, их антигенсвязывающих фрагментов, а также иммуноглобулинов животных других видов, предпочтительно имеющих гуманизированные аспекты. Особый интерес в этом отношении представляют антитела, содержащие одну тяжелую цепь, такие как антитела, обнаруженные у верблюдов. Специалисты в данной области продолжают открывать и разрабатывать новые каркасные участки, каркасы и фрагменты.
[00133] В одном аспекте изобретение относится к способу получения антител, не относящихся к иммуноглобулинам, с использованием неиммуноглобулиновых каркасов, на которые можно привить CDR настоящего изобретения. Можно использовать известные или еще не известные неиммуноглобулиновые каркасные участки и каркасы, при условии, что они содержат связывающий участок, специфичный к целевому белку TSLP. Известные неиммуноглобулиновые каркасные участки или каркасы включают в себя, без ограничения, фибронектин (Compound Therapeutics, Inc., Waltham, Mass.), анкирин (Molecular Partners AG, Zurich, Switzerland), доменные антитела (Domantis, Ltd., Cambridge, UK, and Ablynx nv, Zwijnaarde, Belgium), липокалин (Pieris Proteolab AG, Freising, Germany), небольшие модульные иммунофармацевтические препараты (Trubion Pharmaceuticals Inc., Seattle, Wash.), макситела (Avidia, Inc., Mountain View, Calif.), белок A (Affibody AG, Sweden) и аффилин (гамма-кристаллин или убиквитин) (SciI Proteins GmbH, Halle, Germany).
[00134] Фибронектиновые каркасы основаны на домене фибронектина III типа (например, десятый модуль фибронектина типа III (домен 10 Fn3)). Домен фибронектина III типа содержит 7 или 8 бета-цепей, которые распределяются между двумя бета-складками и упаковываются друг против друга, образуя ядро белка, а кроме того он содержат петли (аналогичные CDR), которые соединяют бета-цепи друг с другом и подвергаются воздействию растворителя. По меньшей мере три такие петли присутствуют на каждом краю бета-складчатого сэндвича, где край представляет собой границу белка, перпендикулярную направлению бета-цепей (см. патент США № 6818418). Указанные каркасы на основе фибронектина не являются иммуноглобулиновыми структурами, хотя общая укладка подобна укладке наименьшего функционального фрагмента антитела, вариабельного участка тяжелой цепи, который содержит весь антиген-распознающий элемент в IgG верблюда и ламы. Благодаря такой структуре антитело, не являющееся иммуноглобулином, имитирует антигенсвязывающие свойства, которые сходны по своей природе и сродству с антигенсвязывающими свойствами антител. Указанные каркасы можно использовать в стратегии рандомизации и перетасовки петель in vitro, которая аналогична процессу созревания аффинности антител in vivo. Такие молекулы на основе фибронектина можно использовать в качестве каркасов, в которых петлевые участки молекулы можно заменить на CDR настоящего изобретения с помощью стандартных методов клонирования.
[00135] Анкириновая технология основана на применении белков, содержащих анкириновые повторяющиеся модули, в качестве каркасов, которые могут нести вариабельные участки, способные связываться с разными мишенями. Анкириновый повторяющийся модуль представляет собой полипептид размером 33 аминокислоты, состоящий из двух антипараллельных альфа-спиралей и бета-складки. Связывание вариабельных участков в основном оптимизируют с использованием рибосомного дисплея.
[00136] Авимеры получают из природного A-домен-содержащего белка, такого как LRP-1. Эти домены используются в природе для белок-белковых взаимодействий, а в организме человека A-домены составляют основу структуры более 250 белков. Авимеры состоят из ряда разных "A-доменных" мономеров (2-10), связанных посредством аминокислотных линкеров. Авимеры, способные связываться с антигеном-мишенью, можно получить с использованием методологии, описанной, например, в публикациях патентных заявок США №№ 20040175756; 20050053973; 20050048512; и 20060008844.
[00137] Аффинные лиганды аффител представляют собой небольшие простые белки в виде пучка из трех спиралей, содержащего в качестве каркаса один из IgG-связывающих доменов белка A. Белок A представляет собой поверхностный белок бактерий Staphylococcus aureus. Указанный каркасный домен состоит из 58 аминокислот, 13 из которых рандомизированы с целью получения библиотеки аффител с большим числом вариантов лигандов (см., например, патент США № 583012). Молекулы аффител, имитирующие антитела, имеют молекулярную массу 6 кДа, тогда как молекулярная масса антител составляет 150 кДа. Несмотря на небольшие размеры, связывающий участок молекулы аффитела подобен связывающему участку антитела.
[00138] Антикалины представляют собой продукты, разработанные компанией Pieris ProteoLab AG. Их получают из липокалинов, широко распространенной группы небольших и устойчивых белков, которые обычно участвуют в физиологическом транспорте или хранении химически чувствительных или нерастворимых соединений. Несколько природных липокалинов встречаются в тканях или жидкостях организма человека. Белковая архитектура напоминает иммуноглобулины и характеризуется наличием гипервариабельных петель и жесткого каркаса. Однако, в отличие от антител и их рекомбинантных фрагментов, липокалины состоят из одной полипептидной цепи, содержащей от 160 до 180 аминокислотных остатков, которая лишь немного превышает по размеру один иммуноглобулиновый домен. Набор из четырех петель, который составляет связывающий карман, демонстрирует выраженную структурную пластичность и допускает множество боковых цепей. Следовательно, связывающий участок можно изменить с помощью собственного способа так, чтобы он распознавал нужные молекулы-мишени разной формы с высоким сродством и высокой специфичностью. Один из белков семейства липокалинов, билин-связывающий белок (BBP) Pieris Brassicae используют для разработки антикалинов путем мутагенеза набора из четырех петель. Примером патентных заявок, описывающих антикалины, является публикация PCT № WO 199916873.
[00139] Молекулы аффилина представляют собой небольшие белки, не являющиеся иммуноглобулинами, которые конструируют так, чтобы они обладали специфическим сродством к белкам и небольшим молекулам. Новые молекулы аффилина можно очень быстро выбрать из двух библиотек, каждую из которых получают на основе отдельного человеческого каркасного белка. Молекулы аффилина не обладают структурной гомологией в отношении иммуноглобулиновых белков. В настоящее время используют два аффилиновых каркаса, один из которых представляет собой гамма-кристаллин, человеческий структурный белок хрусталика, а другой представляет собой белок суперсемейства "убиквитинов". Оба человеческих каркаса имеют очень маленькие размеры, обладают высокой температурной стабильностью и являются почти полностью устойчивыми к изменениям рН и денатурирующим средствам. Указанная высокая стабильность в основном обусловлена рястянутой бета-складчатой структурой белков. Примеры белков, полученных на основе гамма-кристаллина, описаны в WO200104144, а примеры "убиквитин-подобных" белков описаны в WO2004106368.
[00140] Миметики белковых эпитопов (PEM) представляют собой циклические, пептидоподобные молекулы среднего размера (МВ 1-2 кДа), имитирующие бета-шпилечную вторичную структуру белков, основную вторичную структуру, участвующую в белок-белковых взаимодействиях.
[00141] Человеческие TSLP-связывающие антитела можно получить с помощью способов, известных в данной области. Например, для превращения нечеловеческих антител в рекомбинантные человеческие антитела, используют технологию "гуманиринг". В патентной публикации США № 20050008625 описан способ, который можно использовать in vivo для замены вариабельного участка нечеловеческого антитела на человеческий вариабельный участок с сохранением таких же характеристик связывания, как у нечеловеческого антитела, или с улучшением характеристик связывания по сравнению с нечеловеческим антителом. Способ основан на эпитоп-определяемой замене вариабельных участков нечеловеческого исходного антитела на вариабельные участки полностью человеческого антитела. Полученное человеческое антитело, как правило, не является структурно родственным исходному нечеловеческому антителу, но связывается с тем же эпитопом на том же антигене, что и исходное антитело. Коротко говоря, подход с серийной эпитоп-определяемой заменой комплементарности осуществляют путем анализа в клетках конкуренции "конкурирующего средства" и библиотеки разных гибридов исходного антитела ("тестируемые антитела") за связывание с лимитирующими количествами антигена в присутствии репортерной системы, которая реагирует на связывание тестируемого антитела с антигеном. Конкурирующее средство может представлять собой исходное антитело или его производное, такое как одноцепочечный фрагмент Fv. Конкурирующее средство также может представлять собой природный или искусственный лиганд антигена, который связывается с тем же эпитопом, что и контрольное антитело. Единственным требованием в отношении конкурирующего средства является наличие способности связываться с тем же эпитопом, что и исходное антитело, и конкурировать с исходным антителом за связывание с антигеном. Тестируемые антитела содержат один антигенсвязывающий V-участок, общий с нечеловеческим исходным антителом, и другой V-участок, произвольно выбранный из источника, содержащего разнообразные элементы, такого как библиотека репертуара человеческих антител. V-участок, общий с исходным антителом, служит для расположения тестируемых антител на том же эпитопе антигена и в той же ориентации, что позволяет проводить отбор по максимальному соответствию антигенсвязывающей способности исходного антитела.
[00142] Для детекции целевых взаимодействий между тестируемыми антителами и антигеном можно использовать репортерные системы разных типов. Например, дополняющие репортерные фрагменты могут связываться с антигеном и тестируемым антителом соответственно, так что активация репортера при дополнения фрагментов происходит только в том случае, если тестируемое антитело связывается с антигеном. Если гибриды тестируемое антитело- и антиген-репортерный фрагмент экспрессируются совместно с конкурирующим средством, активация репортера становится зависимой от способности тестируемого антитела конкурировать с конкурирующим средством, которая пропорциональна сродству тестируемого антитела к антигену. Можно использовать и другие репортерные системы, такие как система, включающая в себя реактиватор автоингибируемой системы реактивации репортера (RAIR), описанная в патентной заявке США № 10/208730 (публикация № 20030198971), или система конкурентной активации, раскрытая в патентной заявке США № 10/076845 (публикация № 20030157579).
[00143] При использовании системы серийной эпитоп-определяемой замены комплементарности в процессе селекции идентифицируют клетки, экспрессирующие одно тестируемое антитело наряду с такими компонентами, как конкурирующее средство, антиген и репортер. В таких клетках каждое тестируемое антитело конкурирует индивидуально с конкурирующим средством за связывание с лимитирующим количеством антигена. Активность репортера пропорциональна количеству антигена, связанного с тестируемым антителом, которое, в свою очередь, пропорционально сродству тестируемого антитела к антигену и стабильности тестируемого антитела. Тестируемые антитела изначально выбирают исходя из их активности, определяемой в сравнении с активностью исходного антитела, экспрессируемого таким же способом, как и тестируемое антитело. Результатом первого цикла отбора является набор "гибридных" антител, каждое из которых состоит из нечеловеческого V-участка исходного антитела и человеческого V-участка, полученного из библиотеки, и связывается с тем же эпитопом на антигене, что и исходное антитело. Антитело, выбранное в первом цикле из нескольких гибридных антител, должно иметь сродство к антигену, сравнимое со сродством исходного антитела, или превышающее его.
[00144] На второй стадии замены V-участков человеческие V-участки, выбранные на первой стадии, используют для отбора продуктов замены оставшихся нечеловеческих V-участков исходного антитела родственными человеческими V-участками из библиотеки, содержащей множество элементов. Гибридные антитела, выбранные в первом цикле, также можно использовать в качестве конкурирующих средств во втором цикле отбора. Результатом второго цикла отбора является выбор полностью человеческих антител, которые структурно отличаются от исходного антитела, но конкурируют с исходным антителом за связывание с тем же антигеном. Некоторые из выбранных человеческих антител связываются с тем же эпитопом на том же антигене, что и контрольное антитело. Среди указанных выбранных человеческих антител одно или несколько связываются с тем же эпитопом со сродством, сопоставимым со сродством исходного антитела, или превышающим его.
Верблюжьи антитела
[00145] Белки антител, полученные от членов семейств верблюжьи и дромедар (Camelus bactrianus и Calelus dromaderius), включая членов Нового Света, таких как виды ламы (Lama paccos, Lama glama и Lama vicugna), были охарактеризованы в отношении размера, сложности структуры и антигенности для людей. Некоторые обнаруженные в природе антитела IgG указанного семейства млекопитающих не содержат легких цепей и, следовательно, структурно отличаются от типичной четырехцепочечной четвертичной структуры, содержащей две тяжелые и две легкие цепи, свойственной для антител других животных. См. PCT/EP93/02214 (WO 94/04678, опубликован 3 марта 1994).
[00146] Участок верблюжьего антитела, который представляет собой небольшой единичный вариабельный домен, идентифицированный как VHH, можно получить рекомбинантным методом в виде небольшого белка, обладающего высоким сродством к мишени, который представляет собой низкомолекулярный полученный из антитела белок, известный как "верблюжья наночастица", "см. патент США № 5759808, выданный 2 июня 1998 г, см. также Stijlemans, B. et al, 2004 J Biol Chem 279: 1256-1261; Dumoulin M. et al., 2003 Nature 424: 783-788; Pleschberger, M. et al., 2003 Bioconjugate Chem 14: 440-448, Cortez-Retamozo, V. et al., 2002 Int J Cancer 89: 456-62 и Lauwereys, M. et al., 1998 EMBO J 17: 3512-3520. Рекомбинантные библиотеки верблюжьих антител и фрагментов антител можно получить на коммерческой основе, например, от Ablynx, Ghent, Belgium. Как и в случае других антител и антигенсвязывающих фрагментов нечеловеческого происхождения, аминокислотную последовательность верблюжьего антитела можно изменить рекомбинантным методом с получением последовательности, более напоминающей человеческую последовательность, то есть нанотело можно "гуманизировать". Таким образом, природно низкую антигенность верблюжьих антител в отношении человека можно дополнительно уменьшить.
[00147] Молекулярная масса верблюжьего нанотела составляет примерно одну десятую от массы молекулы человеческого IgG, а физический диаметр белка составляет всего несколько нанометров. Вследствие небольшого размера верблюжьи нанотела обладают способностью связываться с антигенными участками, функционально невидимыми для более крупных белков-антител, то есть верблюжьи нанотела можно использовать в качестве реагентов для детекции антигенов, которые нельзя обнаружить с помощью классических иммунологических методов, и, возможно, в качестве терапевтических средств. Так, еще одним следствием небольшого размера является то, что верблюжьи нанотела могут ингибировать целевой белок в результате связывания с определенным участком в канавке или узкой щели и, следовательно, их можно использовать в качестве, более близком к функции классического низкомолекулярного средства, чем классического антитела.
[00148] Низкая молекулярная масса и компактные размеры также приводят к тому, что верблюжьи нанотела являются чрезвычайно термостабильными, устойчивыми к экстремальным значениям рН и к протеолитическому расщеплению, и обладают низкой антигенностью. Другим следствием является то, что верблюжьи нанотела легко перемещаются из системы кровообращения в ткани и даже пересекают гематоэнцефалический барьер, что позволяет использовать их для лечения расстройств, поражающих нервную ткань. Нанотела также могут облегчать транспортировку лекарственных средств через гематоэнцефалический барьер. См. патентную заявку США 20040161738, опубликованную 19 августа 2004. Эти особенности в сочетании с низкой антигенностью в отношении людей свидетельствуют о большом терапевтическом потенциале. Кроме того, такие молекулы могут экспрессироваться целиком в прокариотических клетках, таких как E. coli, они могут экспрессироваться в виде гибридных белков бактериофагом и являются функциональными.
[00149] Соответственно, признаком настоящего изобретения является верблюжье антитело или нанотело, обладающее высоким сродством к TSLP. В одном из вариантов осуществления настоящего изобретения верблюжье антитело или нанотело продуцируется естественным образом в организме животного семейства жьих, то есть продуцируется верблюжьим после иммунизации TSLP или его пептидным фрагментом с помощью способов, описанных здесь для других антител. Альтернативно TSLP-связывающее верблюжье нанотело можно получить рекомбинантным способом, то есть путем выбора, например, из библиотеки фага, отображающего соответствующие мутантные белки верблюжьего нанотела, с помощью процедур пэннинга, используя TSLP в качестве мишени, как описано в приведенных здесь примерах. Рекомбинантные нанотела можно дополнительно оптимизировать рекомбинантным методом с достижением периода полужизни в организме индивидуума-реципиента от 45 минут до двух недель. В конкретном варианте осуществления верблюжье антитело или нанотело получают путем прививки последовательностей CDR тяжелой или легкой цепи человеческого антитела настоящего изобретения на каркасные последовательности нанотела или однодоменного антитела, как описано, например, в PCT/EP93/02214.
Биспецифичные молекулы и поливалентные антитела
[00150] В другом аспекте настоящее изобретение относится к биспецифичным или мультиспецифичным молекулам, содержащим TSLP-связывающее антитело настоящего изобретения или его фрагмент. Антитело настоящего изобретения или его антигенсвязывающие фрагменты можно дериватизировать или присоединить к другой функциональной молекуле, например, к другому пептиду или белку (такому как другое антитело или лиганд рецептора), с получением биспецифичной молекулы, способной связываться, по меньшей мере, с двумя разными участками связывания или молекулами-мишенями. Антитело настоящего изобретения фактически можно дериватизировать или присоединить к нескольким другим функциональным молекулам с получением мультиспецифичных молекул, способных связываться с более чем двумя разными участками связывания и/или молекулами-мишенями; такие мультиспецифичные молекулы также охватываются используемым здесь термином "биспецифичная молекула". Чтобы получить биспецифичную молекулу настоящего изобретения, антитело настоящего изобретения можно функционально присоединить (например, путем химического присоединения, генетической гибридизации, нековалентной ассоциации или иным способом) к одной или нескольким другим связывающим молекулам, таким как другое антитело, фрагмент антитела, пептид или связывающий миметик, с получением биспецифичной молекулы.
[00151] Соответственно, настоящее изобретение относится к биспецифическим молекулам, содержащим по меньшей мере одну первую специфичность связывания в отношении TSLP и вторую специфичность связывания в отношении второго эпитопа-мишени. Например, второй эпитоп-мишень может представлять собой другой эпитоп TSLP, отличный от первого эпитопа-мишени. В других вариантах осуществления второй эпитоп может находиться на мишени, не связанной с TSLP, но способной обеспечивать терапевтическое улучшение в сочетании с TSLP.
[00152] Кроме того, если биспецифичная молекула настоящего изобретения является мультиспецифичной, молекула может дополнительно содержать третью связывающую специфичность в дополнение к первому и второму эпитопам-мишеням.
[00153] В одном варианте осуществления биспецифические молекулы настоящего изобретения содержат в качестве специфичности связывания по меньшей мере одно антитело, или фрагмент антитела, включая, например, Fab, Fab', F(ab')2, Fv или одноцепочечный Fv. Антитело также может представлять собой димером легкой цепи или тяжелой цепи, или любой его минимальный фрагмент, такой как Fv или одноцепочечная конструкция, описанная в Ladner et al., патент США № 4946778.
[00154] Диатела представляют собой двухвалентные, биспецифические молекулы, в которых домены VH и VL экспрессируются в одной полипептидной цепи и соединены линкером, который является слишком коротким, чтобы обеспечить спаривание между двумя доменами одной цепи. Домены VH и VL спариваются с комплементарными доменами другой цепи, создавая два антигенсвязывающих участка (см., например, Holliger et al., 1993 Proc. Natl. Acad. Sci. USA 90: 6444-6448; Poijak et al, 1994 Structure 2: 1121-1123). Диатела можно получить путем экспрессии двух полипептидных цепей либо со структурой VHA-VLB и VHB-VLA (конфигурация VH-VL), либо со структурой VLA-VHB и VLB-VHA (конфигурация VL-VH), в одной клетке. Большинство из них могут экспрессироваться в растворимой форме в бактериях. Одноцепочечные диатела (scDb) получают путем соединения двух диатело-образующих полипептидных цепей с линкером, содержащим примерно 15 аминокислотных остатков (см. Holliger and Winter, 1997 Cancer Immunol. Immunother., 45 (3-4): 128-30; Wu et al., 1996 Immunotechnology, 2 (1): 21-36). scDb могут экспрессироваться в бактериях в виде растворимой, активной мономерной формы (см. Holliger and Winter, 1997 Cancer Immunol. Immunother., 45 (34): 128-30; Wu et al., 1996 Immunotechnology, 2 (l): 21-36; Pluckthun and Pack, 1997 Immunotechnology, 3 (2): 83-105; Ridgway et al., 1996 Protein Eng., 9 (7):617-21). Диатело можно гибридизовать с Fc с получением "ди-диатело" (см. Lu et al, 2004 J. Biol. Chem., 279 (4): 2856-65).
[00155] Другие антитела, которые можно использовать для получения биспецифических молекул настоящего изобретения, включают в себя мышиные, химерные и гуманизированные моноклональные антитела.
[00156] Биспецифические молекулы настоящего изобретения можно получить путем конъюгации составляющих связывающих специфичных элементов с помощью способов, известных в данной области. Например, каждый связывающий специфичный элемент биспецифичной молекулы можно получить отдельно и затем конъюгировать их друг с другом. Если связывающие специфичные элементы представляют собой белки или пептиды, для ковалентной конъюгации можно использовать разные связующие или сшивающие средства. Примеры сшивающих средств включают в себя белок А, карбодиимид, N-сукцинимидил-5-ацетилтиоацетат (SATA), 5,5'-дитиобис (2-нитробензойную кислоту) (DTNB), о-фенилендималеимид (oPDM), N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP) и сульфосукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (сульфо-SMCC) (см., например, Karpovsky et al., 1984 J. Exp. Med. 160: 1686; Liu, M A et al., 1985 Proc. Natl. Acad. Sci. USA, 82: 8648). Другие способы описаны в Paulus, 1985 Behring Ins. Mitt. № 78, 118-132; Brennan et al., 1985 Science 229: 81-83) и Glennie et al., 1987 J. Immunol. 139: 2367-2375). В качестве конъюгирующих средств можно использовать SATA и сульфо-SMCC, которые можно приобрести в Pierce Chemical Co. (Rockford, 111.).
[00157] Если связывающие специфичные элементы представляют собой антитела, их можно конъюгировать путем соединения посредством сульфгидрильных групп С-концевых шарнирных участков двух тяжелых цепей. В конкретном варианте осуществления шарнирный участок модифицируют так, чтобы он перед конъюгацией содержал нечетное число сульфгидрильных остатков, например один.
[00158] Альтернативно, оба связывающих специфичных элемента можно закодировать в одном векторе и обеспечить их экспрессию и сборку в одной клетке-хозяине. Данный способ предпочтительно использовать в тех случаях, когда биспецифическая молекула представляет собой гибридный белок mAb X mAb, mAb X Fab, Fab X F(ab')2 или лиганд X Fab. Биспецифическая молекула настоящего изобретения может представлять собой одноцепочечную молекулу, содержащую одноцепочечное антитело и связывающую детерминанту, или одноцепочечную биспецифическую молекулу, содержащую две связывающие детерминанты. Биспецифические молекулы могут содержать по меньшей мере две одноцепочечные молекулы. Способы получения биспецифических молекул описаны, например, в патенте США № 5260203; патенте США № 5455030; патенте США № 4881175; патенте США № 512405; патенте США № 5091513; патенте США № 5476786; патенте США № 5013653; патенте США № 5258498; и патенте США № 5482858.
[00159] Связывание биспецифических молекул с их специфическими мишенями можно подтвердить, например, с помощью иммуноферментного анализа (ELISA), радиоиммуноанализа (REA), анализа FACS, биоанализа (например, путем ингибирования роста) или анализа методом вестерн-блоттинга. Каждый из этих анализов, как правило, позволяет детектировать присутствие комплексов белок-антитело, представляющих особый интерес, с использованием меченого реагента (например, антитела), специфичного для представляющего интерес комплекса.
[00160] В другом аспекте настоящее изобретение относится к поливалентным соединениям, содержащим по меньшей мере два идентичных или разных антигенсвязывающих участков антител и антигенсвязывающих фрагментов настоящего изобретения, связывающихся с TSLP. Антигенсвязывающие участки можно соединить посредством гибридизации белков или ковалентной или нековалентной связи. Альтернативные способы соединения описаны для биспецифических молекул. Тетравалентные соединения можно получить, например, путем сшивки антител и антигенсвязывающих фрагментов настоящего изобретения с антителом или антигенсвязывающим фрагментом, который связывается с константными доменами антител и антигенсвязывающих фрагментов настоящего изобретения, такими как Fc или шарнирный участок.
[00161] Тримеризация доменов описана, например, в патенте Borean Pharma EP 1012280B1. Пентамеризация модулей описана, например, в РСТ/ЕР97/05897.
Антитела с увеличенным периодом полужизни
[00162] Настоящее изобретение относится к антителам, способным специфически связываться с TSLP и имеющим увеличенный период полужизни in vivo.
[00163] Многие факторы могут влиять на период полужизни белка in vivo. Например, почечная фильтрация, метаболизм в печени, деградация протеолитическими ферментами (протеазами) и иммуногенные реакции (например, нейтрализация белка антителами и поглощение его макрофагами и дендритными клетками). Для увеличения периода полужизни антител и антигенсвязывающих фрагментов настоящего изобретения можно использовать множество стратегий. Например, путем химического присоединия к полиэтиленгликолю (PEG), reCODE PEG, каркасу антитела, полисиаловой кислоте (PSA), гидроксиэтилкрахмалу (HES), альбуминсвязывающим лигандам и углеводным оболочкам; путем генетической гибридизации с белками, способными связываться с сывороточными белками, такими как альбумин, IgG, FcRn и переносящие белки; путем присоединения (генетическими или химическими методами) к другим связывающим фрагментам, способным связываться с сывороточными белками, таким как нанотела, Fab, DARPin, авимеры, аффитела и антикалины; путем генетической гибридизации с rPEG, альбумином, доменом альбумина, альбуминсвязывающими белками и Fc; или путем включения в наноносители, композиции с замедленным высвобождением или медицинские устройства.
[00164] Чтобы продлить время пребывания антител в сыворотке in vivo, к антителам или их фрагментам можно присоединить молекулы инертного полимера, такого как PEG с высокой молекулярной массой, с использованием или без использования многофункционального линкера, либо посредством сайт-специфической конъюгации PEG с N- или С-концом антитела, либо через эпсилон-аминогруппы, присутствующие на остатках лизина. Чтобы осуществить пегилирование, антитело, или его антигенсвязывающий фрагмент, обычно подвергают взаимодействию с полиэтиленгликолем (PEG), например, с реакционноспособным сложным эфиром или альдегидным производным PEG, в условиях, в котрых одна или несколько групп PEG присоединяются к антителу или фрагменту антитела. Для проведения пэгирования можно использовать реакцию ацилирования или реакцию алкилирования реакционноспособной молекулой ПЭГ (или аналогичным реакционноспособным водорастворимым полимером). Используемый здесь термин "полиэтиленгликоль" охватывает все формы PEG, используемые для дериватизации других белков, такие как моно(C1-C10)алкокси- или арилоксиполиэтиленгликоль или полиэтиленгликольмалеимид. В одном варианте осуществления антитело, подлежащее пегилированию, представляет собой агликозилированное антитело. Для дериватизации используют линейный или разветвленный полимер, так, чтобы обеспечить минимальную потерю биологической активности. Степень конъюгации тщательно отслеживают методами SDS-PAGE и масс-спектрометрии, чтобы гарантировать надлежащую конъюгацию молекул ПЭГ с антителами. Непрореагировавший ПЭГ можно отделить от конъюгатов антитело-ПЭГ методом гель-хроматографии или ионообменной хроматографии. ПЭГ-дериватизированные антитела можно протестировать на связывающую активность, а также на эффективность in vivo с помощью методов, хорошо известных специалистам в данной области, например, с помощью описанных здесь иммуноанализов. В применении к антителам настоящего изобретения и их антигенсвязывающим фрагментам можно использовать способы пегилирования белков, известные в данной области. См., например, ЕР 0154316, Nishimura et al., и EP 0401384, Ishikawa et al.
[00165] Другие модифицированные технологии пегилирования включают в себя перераспределяющую химически ортогонально направленную рекомбинантную технологию (ReCODE PEG), которая позволяет вставлять химически определенные боковые цепи в биосинтетические белки через систему перераспределения, которая содержит синтетазу тРНК и тРНК. Данная технология позволяет включать более 30 новых аминокислот в биосинтетические белки в клетках E.coli, дрожжей и млекопитающих. тРНК включает нормальную аминокислоту в любом месте, где расположен янтарный кодон, превращая янтарный кодон из стоп-кодона в кодон, который сигнализирует о включении химически определенной аминокислоты.
[00166] Для увеличения периода полужизни в сыворотке также можно использовать рекомбинантную технологию пэгилирования (rPEG). Эта технология включает в себя генетическую гибридизацию неструктурированного белкового хвоста длиной 300-600 аминокислот с существующим фармацевтическим белком. Поскольку кажущаяся молекулярная масса такой неструктурированной белковой цепи примерно в 15 раз больше, чем ее фактическая молекулярная масса, период полужизни белка в сыворотке значительно увеличивается. В отличие от традиционного пегилирования, которое включает в себя химическую конъюгацию и повторную очистку, данный способ позволяет значительно упростить производственный процесс и получить однородный продукт.
[00167] Полисиалирование представляет собой другой способ увеличения активного существования и улучшения стабильности терапевтических пептидов и белков, в котором используют природную полимерную полисиаловую кислоту (PSA). PSA представляет собой полимер сиаловой кислоты (сахар). Используемая для доставки белков и терапевтических пептидных средств полисиаловая кислота обеспечивает защитное микроокружение при конъюгации. Это увеличивает активное существование терапевтического белка в кровотоке и предотвращает его распознавание иммунной системой. Полимер PSA в природных условиях присутствует в организме человека. Он используется некоторыми бактериями, которые эволюционировали в течение миллионов лет, для покрытия клеточных стенок. Такие естественным образом полисиалилированные бактерии способны в силу молекулярной мимикрии преодолеть защитную систему организма. PSA, обеспечивающую природную технологию малой заметности, можно легко получить из таких бактерий в больших количествах и с заданными физическими характеристиками. Бактериальная PSA является полностью неиммуногенной, даже если она связана с белками, поскольку она химически идентична PSA, присутствующей в организме человека.
[00168] Другой способ включает в себя применение производных гидроксиэтилкрахмала ("HES"), связанных с антителами. HES представляет собой модифицированный природный полимер, полученный из воскового кукурузного крахмала и способный метаболизироваться ферментами организма. Обычно растворы HES вводят для пополнения недостающего объема крови и улучшения реологических свойств крови. Хезилирование антител позволяет увеличить период полужизни в кровотоке в результате повышения стабильности молекулы и уменьшения почечного клиренс, что приводит к увеличению биологической активности. Изменяя разные параметры, такие как молекулярная масса HES, можно получить широкий спектр оптимизированных конъюгатов HES с антителами.
[00169] Антитела, характеризующиеся повышенным периодом полужизни in vivo, также можно получить путем введения одной или нескольких модификаций аминокислот (то есть замен, вставок или делеций) в константный домен IgG или его FcRn-связывающий фрагмент (предпочтительно Fc или шарнирный участок домена Fc). См., например, международную публикацию WO 98/23289; международную публикацию № WO 97/34631; и патент США № 6277375.
[00170] Кроме того, чтобы получить антитела или фрагменты антител, обладающие повышенной стабильностью in vivo или имеющие более длительный период полужизни in vivo, антитела можно конъюгировать с альбумином. Такие способы хорошо известны в данной области, см., например, международные публикации №№ WO 93/15199, WO 93/15200 и WO 01/77137; и европейский патент № EP 413622.
[00171] Стратегии увеличения периода полужизни особенно используют при получении нанотел, связующих средств на основе фибронектина и других антител или белков, требующих увеличения периода полужизни in vivo.
Конъюгаты антител
[00172] Настоящее изобретение относится к антителам или их антигенсвязывающим фрагментам, способным специфически связываться с внеклеточным доменом TSLP и гибридизованным рекомбинантными методами или химически конъюгированным (включая как ковалентным, так и нековалентныи способом) с гетерологичным белком или полипептидом (или его антигенсвязывающим фрагментом, предпочтительно с полипептидом, содержащим по меньшей мере 10, по меньшей мере 20, по меньшей мере 30, по меньшей мере 40, по меньшей мере 50, по меньшей мере 60, по меньшей мере 70, по меньшей мере 80, по меньшей мере 90 или по меньшей мере 100 аминокислот) с получением гибридных белков. В частности, изобретение относится к гибридным белкам, содержащим антигенсвязывающий фрагмент описанного здесь антитела (например, фрагмент Fab, фрагмент Fd, фрагмент Fv, фрагмент F(ab)2, домен VH, CDR VH, домен VL или CDR VL) и гетерологичный белок, полипептид или пептид. Способы гибридизации или конъюгирования белков, полипептидов или пептидов с антителом или фрагментом антитела известны в данной области. См., например, патенты США №№ 5336603, 5622929, 5359046, 5349053, 5447851, и 5112946; европейские патенты ЕР 307434 и ЕР 367166; международные публикации №№ WO 96/04388 и WO 91/06570; Ashkenazi et al., 1991, Proc. Natl. Acad. Sci. USA 88: 10535-10539; Zheng et al., 1995, J. Immunol. 154: 5590-5600; и Vil et al., 1992, Proc. Natl. Acad. Sci. USA 89: 11337-11341.
[00173] Другие гибридные белки можно получить с помощью методов перетасовки генов, перетасовки мотивов, перетасовки экзонов и/или перетасовки кодонов (в совокупности называемых "ДНК-перетасовкой"). ДНК-перетасовку можно использовать для изменения активности антител настоящего изобретения и их антигенсвязывающих фрагментов (например, с получением антител и их антигенсвязывающих фрагментов, характеризующихся более высоким сродством и более низкой скоростью диссоциации). Общую информацию можно найти в патентах США №№ 5605793, 5811238, 5830721, 5834252 и 5837458; Patten et al., 1997, Curr. Opinion Biotechnol. 8:724-33; Harayama, 1998, Trends Biotechnol. 16 (2): 76-82; Hansson, et al, 1999, J. Mol. Biol. 287: 265-76; и Lorenzo and Blasco, 1998, Biotechniques 24 (2):308-313 (все указанные патенты и публикации полностью включены в настоящее описание посредством ссылки). Антитела и их антигенсвязывающие фрагменты, или кодируемые антитела и их антигенсвязывающие фрагменты, перед рекомбинацией можно изменить путем неспецифического мутагенеза методом ПЦР с внесением ошибок, случайной вставки нуклеотидов или другими способами. Полинуклеотид, кодирующий антитело или его антигенсвязывающий фрагмент, способный специфически связываться с участком стержня TSLP, можно подвергнуть рекомбинации с одним или несколькими компонентами, мотивами, участками, частями, доменами, фрагментами и т.д. одной или нескольких гетерологичных молекул.
[00174] Кроме того, антитела и их антигенсвязывающие фрагменты можно гибридизовать с маркерными последовательностями, такими как пептид, для облегчения очистки. В одном варианте осуществления маркерная аминокислотная последовательность может включать в себя гексагистидиновый пептид (SEQ ID NO: 40), например, входящий в состав вектора pQE (QIAGEN, Inc., 9259 Eton Avenue, Chatsworth, CA, 91311), а также другие маркеры, многие из которых являются коммерчески доступными. Как описано в Gentz et al., 1989, Proc. Natl. Acad. Sci. USA 86: 821-824, например, гексагистидин (SEQ ID NO: 40) обеспечивает удобную очистку гибридного белка. Другие пептидные маркеры, используемые для очистки, включают в себя, без ограничения, гемагглютининовый ("HA") маркер, который соответствует эпитопу, полученному из белка гемагглютинина гриппа (Wilson et al, 1984, Cell 37: 767), и маркер "FLAG".
[00175] В одном варианте осуществления антитела настоящего изобретения и их антигенсвязывающие фрагменты конъюгируют с диагностическим или детектируемым средством. Такие антитела можно использовать для мониторинга или прогнозирования возникновения, развития, прогрессирования и/или тяжести заболевания или расстройства в процедуре клинических испытаний, таких как определение эффективности конкретной терапии. Диагностику и детекцию можно проводить с использованием антитела, присоединенного к детектируемому веществу, включающему в себя, без ограничения, разные ферменты, такие как, без ограничения, пероксидаза хрена, щелочная фосфатаза, бета-галактозидаза или ацетилхолинэстераза; простетические группы, такие как, без ограничения, стрептавидин/биотин и авидин/биотин; флуоресцентные вещества, такие как, без ограничения, умбеллиферон, флуоресцеин, изотиотионат флуоресцеина, родамин, дихлортриазиниламин флуоресцеин, дансилхлорид или фикоэритрин; люминесцентные вещества, такие как, без ограничения, люминол; биолюминесцентные вещества, такие как, без ограничения, люцифераза, люциферин и аэкорин; радиоактивные вещества, такие как, без ограничения, йод (131I, 125I, 123I и 121I), углерод (14С), сера (35S), тритий (3H), индий (115In, 113In, 112In и 11lIn), технеций (99Tc), таллий (201Ti), галлий (68Ga, 67Ga), палладий (103Pd), молибден (99Mo), ксенон (133Xe), фтор (18F), 153Sm, 177Lu, 159Gd, 149 Pm, 140La, 175Yb, 166Ho, 90Y, 47Sc, 186Re, 188Re, 142Pr, 105Rh, 97Ru, 68Ge, 57Co, 65Zn, 85Sr, 32P, 153Gd, 169Yb, 51Cr, 54Mn, 75Se, 113Sn и 117Tin; а также позитронно-активные металлы, с использованием разных видов позитронно-эмиссионной томографии, и нерадиоактивных парамагнитных ионов металлов.
[00176] Кроме того, антитело или его антигенсвязывающий фрагмент можно конъюгировать с терапевтическим фрагментом или лекарственным фрагментом. Терапевтические фрагменты или лекарственные фрагменты не должны истолковываться как ограниченные классическими химическими терапевтическими средствами. Например, лекарственный фрагмент может представлять собой белок, пептид или полипептид, обладающий желательной биологической активностью. Такие белки могут включать в себя, например, токсин, такой как абрин, рицин А, экзотоксин синегнойной палочки, холерный токсин или дифтерийный токсин; белок, такой как фактор некроза опухоли, альфа-интерферон, бета-интерферон, фактор роста нервов, фактор роста тромбоцитов, активатор тканевого плазминогена, апоптотическое средство, антиангиогенное средство; или, модификатор биологического ответа, такой как, например, лимфокин.
[00177] Кроме того, антитело можно конъюгировать с такими терапевтическими фрагментами, как ионы радиоактивных металлов, например альфа-излучателей, таких как 213Bi, или макроциклические хелаторы, пригодные для конъюгирования ионов радиометаллов, включая, без ограничения, 131In, 131LU, 131Y, 131Ho, 131Sm, с полипептидами. В одном варианте осуществления макроциклический хелатор представляет собой 1,4,7,10-тетраазациклододекан-N,N',N",N'''-тетрауксусную кислоту (DOTA), которую можно присоединить к антителу через молекулу линкера. Такие молекулы-линкеры широко известны в данной области и описаны в Denardo et al., 1998, Clin Cancer Res. 4 (10): 2483-90; Peterson et al., 1999, Bioconjug. Chem. 10 (4): 553-7; и Zimmerman et al., 1999, Nucl. Med. Biol. 26 (8): 943-50, каждый из которых включен в качестве ссылки во всей своей полноте.
[00178] Способы конъюгирования терапевтических фрагментов с антителами хорошо известны, см., например, Amon et al., "Monoclonal Antibodies For Immunotargeting Of Drugs In Cancer Therapy", in Monoclonal Antibodies And Cancer Therapy, Reisfeld et al. (eds.), pp. 243-56 (Alan R. Liss, Inc. 1985); Hellstrom et al., "Antibodies For Drug Delivery", in Controlled Drug Delivery (2nd Ed.), Robinson et al. (eds.), pp. 623-53 (Marcel Dekker, Inc. 1987); Thorpe, "Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review", in Monoclonal Antibodies 84: Biological And Clinical Applications, Pinchera et al. (eds.), pp. 475-506 (1985); "Analysis, Results, And Future Prospective Of The Therapeutic Use Of Radiolabeled Antibody In Cancer Therapy", in Monoclonal Antibodies For Cancer Detection And Therapy, Baldwin et al. (eds.), pp. 303-16 (Academic Press 1985), and Thorpe et al., 1982, Immunol. Rev. 62:119-58.
[00179] Антитела также можно присоединить к твердым носителям, после чего их можно использовать для иммунологических анализов или очистки антигена-мишени. Такие твердые носители включают в себя, без ограничения, стекло, целлюлозу, полиакриламид, найлон, полистирол, поливинилхлорид или полипропилен.
Нуклеиновые кислоты, кодирующие антитела
[00180] Изобретение предлагает по существу очищенные молекулы нуклеиновых кислот, кодирующих полипептиды, содержащие сегменты или домены описанных выше антител против TSLP. Такие полинуклеотиды могут кодировать по меньшей мере один участок CDR, как правило, все три участка CDR тяжелой или легкой цепи описанных здесь антител против TSLP. Такие полинуклеотиды также могут кодировать всю или по существу всю последовательность вариабельного участка тяжелой цепи и/или легкой цепи описанных здесь антител против TSLP. Такие полинуклеотиды также могут кодировать как вариабельный участок, так и константный участок антитела. Вследствие вырожденности кода каждую из аминокислотных последовательностей иммуноглобулина кодируют несколько нуклеотидных последовательностей.
[00181] Полинуклеотидные последовательности можно получить путем твердофазного синтеза ДНК de novo или путем ПЦР-мутагенеза существующей последовательности (например, последовательности, описанной ниже в примерах), кодирующей TSLP-связывающее антитело или его связывающий фрагмент. Прямой химический синтез нуклеиновых кислот можно проводить с помощью способов, известных в данной области, таких как фосфотриэфирный метод Narang et al., 1979, Meth. Enzymol. 68: 90; фосфодиэфирный метод Brown et al., Meth. Enzymol. 68: 109, 1979; диэтилфосфорамидитный метод Beaucage et al., Tetra. Lett., 22: 1859, 1981; и твердофазный метод, патент США № 4458066. Введение мутаций в полинуклеотидную последовательность с помощью метода ПЦР можно осуществлять по способу, описанному, например, в PCR Technology: Principles and Applications for DNA Amplification, H. A. Erlich (Ed.), Freeman Press, NY, N.Y., 1992; PCR Protocols: A Guide to Methods and Applications, Innis et al. (Ed.), Academic Press, San Diego, Calif., 1990; Mattila et al., Nucleic Acids Res. 19:967, 1991; и Eckert et al., PCR Methods and Applications 1:17, 1991.
[00182] Изобретение также относится к векторам экспрессии и клеткам-хозяевам, обеспечивающим продукцию описанных выше TSLP-связывающих антител. Для экспрессии полинуклеотидов, кодирующих цепи или связывающие фрагменты TSLP-связывающих антител, можно использовать разные векторы экспрессии. Для получения антител в клетке-хозяине млекопитающих можно использовать как вирусные, так и невирусные векторы экспрессии. Невирусные векторы и системы включают в себя плазмиды, эписомальные векторы, обычно содержащие экспрессионную кассету для экспрессии белка или РНК, и человеческие искусственные хромосомы (см., например, Harrington et al. Nat Genet. 15:345, 1997). Например, невирусные векторы, подходящие для экспрессии TSLP-связывающих полинуклеотидов и полипептидов в клетках млекопитающих (например, человека), включают в себя pThioHis A, B & C, pcDNA3.1/His, pEBVHis A, B & C, (Invitrogen, San Diego, CA), векторы MPSV и множество других векторов, известных в данной области и используемых для экспрессии других белков. Подходящие вирусные векторы включают в себя векторы на основе ретровирусов, аденовирусов, аденоассоциированных вирусов, вирусов герпеса, векторы на основе SV40, вируса папилломы, HBP вируса Эпштейна-Барра, векторы на основе вируса осповакцины и вируса леса Семлики (SFV). См. Brent et al., выше; Smith, Annu. Rev. Microbiol. 49:807, 1995; и Rosenfeld et al., Cell 68:143, 1992.
[00183] Выбор вектора экспрессии зависит от предполагаемых клеток-хозяев, в которых будет экспрессироваться вектор. Как правило, векторы экспрессии содержат промотор и другие регуляторные последовательности (например, энхансеры), функционально связанные с полинуклеотидами, кодирующими антигенсвязывающий фрагмент цепи TSLP-связывающего антитела. В одном варианте осуществления используют индуцируемый промотор, чтобы предотвратить экспрессию вставленных последовательностей в условиях, отличных от индуцирующих. Индуцируемые промоторы включают в себя, например, промотор арабинозы, lacZ, металлотионеина или промотор теплового шока. Культуры трансформированных организмов можно выращивать в неиндуцирующих условиях без смещения популяции к кодирующим последовательностям, продукты экспрессии которых лучше переносятся клетками-хозяевами. Для эффективной экспрессии цепи или антигенсвязывающего фрагмента TSLP-связывающего антитела помимо промоторов также могут потребоваться, или требуются, другие регуляторные элементы. К таким элементам обычно относятся кодон инициации ATG и соседний участок связывания рибосомы, или другие последовательности. Кроме того, эффективность экспрессии можно повысить путем применения энхансеров, соответствующих используемой клеточной системе (см., например, Scharf et al., Results Probl. Cell Differ. 20:125, 1994; и Bittner et al., Meth. Enzymol., 153:516, 1987). Например, для увеличения экспрессии в клетках-хозяевах млекопитающих можно использовать энхансер SV40 или энхансер CMV.
[00184] Векторы экспрессии также могут содержать последовательность сигнала секреции в таком положении, чтобы обеспечить образование гибридного белка с полипептидами, кодируемыми вставленными последовательностями TSLP-связывающих антител. Чаще всего вставленные последовательности TSLP-связывающих антител присоединяют к сигнальным последовательностям перед включением в вектор. Векторы, которые предполагается использовать для вставки последовательностей, кодирующих вариабельные домены легких и тяжелых цепей TSLP-связывающих антител, иногда также кодируют константные участки или их фрагменты. Такие векторы позволяют экспрессировать вариабельные участки в виде гибридных белков с константными участками, что приводит к образованию интактных антител и их антигенсвязывающих фрагментов. Как правило, такие константные участки являются человеческими.
[00185] Клетки-хозяева, несущие и экспрессирующие цепи TSLP-связывающих антител, могут быть либо прокариотическими, либо эукариотическими. В качестве прокариотического хозяина для клонирования и экспрессии полинуклеотидов настоящего изобретения можно использовать E. coli. Другие подходящие для использования микробные хозяева включают в себя бациллы, такие как Bacillus subtilis и другие энтеробактерии, такие как Salmonella, Serratia и разные виды Pseudomonas. Можно получить векторы экспрессии, подходящие для применения в таких прокариотических хозяевах, которые обычно содержат последовательности, регулирующие экспрессию, совместимые с клеткой-хозяином (такие как точка начала репликации). Кроме того, может присутствовать любое число разных известных промоторов, таких как промоторная система лактозы, промоторная система триптофана (trp), промоторная система бета-лактамазы или промоторная система фага лямбда. Промоторы обычно регулируют экспрессию, необязательно с последовательностью оператора, и содержат последовательности участков связывания с рибосомами и т.п., обеспечивающие инициацию и завершение транскрипции и трансляции. Другие микробы, такие как дрожжи, также можно использовать для экспрессии TSLP-связывающих полипептидов настоящего изобретения. Кроме того, можно использовать клетки насекомых в сочетании с бакуловирусными векторами.
[00186] В одном варианте осуществления для экспрессии и продукции TSLP-связывающих полипептидов настоящего изобретения используют клетки-хозяева млекопитающих. Например, такие клетки могут представлять собой либо гибридомную клеточную линию, экспрессирующую эндогенные гены иммуноглобулинов (например, клон миеломной гибридомы 1D6.C9, описанный в примерах), либо клеточную линию млекопитающих, несущую экзогенный вектор экспрессии (например, клетки миеломы SP2/0, описанные в качестве примера ниже). К ним относятся любые нормальные мортальные или нормальные или аномальные иммортализованные клетки животных или человека. Например, разработано несколько подходящих линий клеток-хозяев, способных секретировать интактные иммуноглобулины, в том числе клеточные линии СНО, разные клеточные линии Cos, клетки HeLa, миеломные клеточные линии, трансформированные B-клетки и гибридомы. Применение культуры клеток тканей млекопитающих для экспрессии полипептидов описано, в целом, например, в Winnacker, FROM GENES TO CLONES, VCH Publishers, N.Y., N.Y., 1987. Векторы экспрессии, предназначенные для использования в клетках-хозяевах млекопитающих, могут содержать последовательности, регулирующие экспрессию, такие как точка начала репликации, промотор и энхансер (см., например, Queen, et al. Immunol. Rev. 89: 49-68, 1986), и необходимые участки процессинга информации, такие как участки связывания рибосом, участки сплайсинга РНК, участки полиаденилирования и последовательности терминатора транскрипции. Такие векторы экспрессии обычно содержат промоторы, полученные из генов млекопитающих или из вирусов млекопитающих. Подходящие промоторы могут быть конститутивными, специфичными для конкретного типа клеток, специфичными для стадии и/или модулируемыми или регулируемыми. Подходящие промоторы включают в себя, без ограничения, промотор металлотионеина, конститутивный основной поздний промотор аденовируса, дексаметазон-индуцируемый промотор MMTV, промотор SV40, промотор MRP poIIII, конститутивный промотор MPSV, тетрациклин-индуцируемый промотор CMV (такой как человеческий немедленно-ранний промотор CMV), конститутивный промотор CMV и сочетания промотор-энхансер, известные в данной области.
[00187] Способы введения векторов экспрессии, содержащих представляющие интерес полинуклеотидные последовательности, варьируют в зависимости от типа клетки-хозяина. Например, трансфекцию прокариотических клеток обычно проводят с использованием хлорида кальция, тогда как трансфекцию других клеток-хозяев проводят путем обработки фосфатом кальция или электропорации. (См., в общих чертах, Sambrook, et al., см. выше). Другие способы включают в себя, например, электропорацию, обработку фосфатом кальция, трансформацию, опосредованную липосомами, инъекцию и микроинъекцию, баллистические методы, использованием виросом, иммунолипосом, конъюгатов поликатион:нуклеиновая кислота, голой ДНК, искусственных вирионов, гибридов со структурным белком вируса герпеса VP22 (Elliot and O'Hare, Cell 88:223, 1997), усиленное средством поглощение ДНК и трансдукцию ex vivo. Чтобы обеспечить продукцию рекомбинантных белков в течение длительного времени и с высоким выходом, часто требуется стабильная экспрессия. Например, клеточные линии, которые стабильно экспрессируют цепи или связывающие фрагменты TSLP-связывающих антител, можно получить с использованием векторов экспрессии настоящего изобретения, которые содержат вирусные точки начала репликации или эндогенные экспрессирующие элементы и селектируемый маркерный ген. После введения вектора клетки оставляют расти в течение 1-2 дней в обогащенной среде и затем среду меняют на селективную. Селектируемый маркер обеспечивает устойчивость к селекции и его присутствие позволяет выращивать клетки, которые успешно экспрессируют введенные последовательности в селективных средах. Устойчивые, стабильно трансфицированные клетки выращивают, используя методы культивирования тканей, соответствующие типу клеток.
Получение антител и фрагментов антител
[00188] Моноклональные антитела (mAb) можно получить разными способами, включающими в себя традиционную методологию моноклональных антител, например, стандартный метод гибридизации соматических клеток Kohler and Milstein, 1975 Nature 256: 495. Можно использовать множество способов получения моноклонального антитела, например, путем трансформации В-лимфоцитов вирусом или онкогеном.
[00189] В качестве животной системы для получения гибридом используют мышиную систему. Получение гибридом в организме мыши представляет собой хорошо разработанную процедуру. Схемы и способы иммунизации с целью выделения иммунизированных спленоцитов, используемых для гибридизации, известны в данной области. Также известны партнеры по гибридизации (например, клетки мышиной миеломы) и способы гибридизации.
[00190] В некоторых вариантах осуществления антитела настоящего изобретения представляют собой гуманизированные моноклональные антитела. Химерные или гуманизированные антитела и антигенсвязывающие фрагменты настоящего изобретения можно получить на основе последовательности мышиного моноклонального антитела, полученного описанным выше способом. ДНК, кодирующие тяжелые и легкие цепи иммуноглобулинов, можно получить из представляющей интерес мышиной гибридомы с последующим введением немышиных (например, человеческих) последовательностей иммуноглобулинов с помощью стандартных методов молекулярной биологии. Например, для получения химерного антитела мышиные вариабельные участки можно соединить с человеческими константными участками с использованием способов, известных в данной области (см., например, патент США № 4816567, Cabilly et al.). Для получения гуманизированного антитела мышиные участки CDR можно вставить в человеческий каркас с использованием способов, известных в данной области. См., например, патент США № 5225539, Winter, и патенты США №№ 5530101; 5585089; 5693762 и 6180370, Queen et al.
[00191] В некоторых вариантах осуществления антитела настоящего изобретения представляют собой человеческие моноклональные антитела. Такие человеческие моноклональные антитела, направленные против TSLP, можно получить с использованием трансгенных или трансхромосомных мышей, несущих части иммунной системы человека, вместо системы мыши. Эти трансгенные и трансхромосомные мыши включают в себя мышей, называемых здесь мыши HuMAb и мыши KM, соответственно, и в совокупности упоминаются здесь как "мыши, продуцирующие человеческие Ig".
[00192] HuMAb Mouse® (Medarex, Inc.) содержит минилокус человеческого гена иммуноглобулина, который кодирует нереаранжированные человеческие последовательности тяжелой (мю и гамма) и легкой цепи каппа иммуноглобулина, вместе с направленными мутациями, которые дезактивируют эндогенные локусы цепей мю и каппа (см., например, Lonberg, et al., 1994 Nature 368 (6474): 856-859). Соответственно, у мышей наблюдается уменьшение экспрессии мышиного IgM или K, и в ответ на иммунизацию вводимые трансгены человеческих тяжелой и легкой цепей подвергаются переключению классов и соматической мутации с образованием моноклонального человеческого IgG-каппа, обладающего высоким сродством (Lonberg, N. et al., 1994 supra; reviewed in Lonberg, N., 1994 Handbook of Experimental Pharmacology 113:49-101; Lonberg, N. and Huszar, D., 1995 Intern. Rev. Immunol. 13: 65-93, and Harding, F. and Lonberg, N., 1995 Ann. N.Y. Acad. Sci. 764:536-546). Получение и использование мышей HuMAb и геномных модификаций, которые несут такие мыши, также описано в Taylor, L. et al., 1992 Nucleic Acids Research 20:6287-6295; Chen, J. et al., 1993 International Immunology 5: 647-656; Tuaillon et al., 1993 Proc. Natl. Acad. Sci. USA 94:3720-3724; Choi et al., 1993 Nature Genetics 4:117-123; Chen, J. et al., 1993 EMBO J. 12: 821-830; Tuaillon et al., 1994 J. Immunol. 152:2912-2920; Taylor, L. et al., 1994 International Immunology 579-591; и Fishwild, D. et al., 1996 Nature Biotechnology 14: 845-851, содержание которых полностью включено в настоящее описание посредством ссылки во всей их полноте. См. также патенты США №№ 5545806; 5569825; 5625126; 5633425; 5789650; 5877397; 5661016; 5814318; 5874299; и 5770429; Lonberg and Kay; патент США № 5545807, Surani et al; публикации PCT №№ WO 92103918, WO 93/12227, WO 94/25585, WO 97113852, WO 98/24884 и WO 99/45962, Lonberg and Kay; и публикацию РСТ № WO 01/14424, Korman et al.
[00193] В некоторых вариантах осуществления человеческие антитела можно получить с использованием мыши, которая несет человеческие последовательности иммуноглобулинов на трансгенах и трансхомосомах, такой как мышь, которая несет человеческий трансген тяжелой цепи и человеческую трансхромосому легкой цепи. Такие мыши, называемые здесь "мыши KM", подробно описаны в публикации PCT WO 02/43478, Ishida et al.
[00194] Кроме того, в данной области существуют альтернативные трансгенные животные системы, экспрессирующие гены человеческих иммуноглобулинов, которые можно использовать для получения TSLP-связывающих антител и их антигенсвязывающих фрагментов. Например, можно использовать альтернативную трансгенную систему, называемую Xenomouse (Abgenix, Inc.). Такие мыши описаны, например, в патентах США №№ 5939598; 6075181; 6114598; 6150584 и 6162963, Kucherlapati et al.
[00195] В данной области также существуют альтернативные трансхромосомные животные системы, экспрессирующие гены человеческих иммуноглобулинов, которые можно использовать для получения TSLP-связывающих антител настоящего изобретения. Например, можно использовать мышей, несущих как трансхромосому тяжелой цепи человека, так и трансхромосому легкой цепи человека, называемых "мыши TC"; такие мыши описаны в Tomizuka et al., 2000 Proc. Natl. Acad. Sci. USA 97: 722-772. Кроме того, в данной области описаны коровы, несущие человеческие трансхромосомы тяжелых и легких цепей (Kuroiwa et al, 2002 Nature Biotechnology 20: 889-894), которых можно использовать для получения TSLP-связывающих антител настоящего изобретения.
[00196] Моноклональные человеческие антитела также можно получить с использованием методов фаговых дисплеев для скрининга библиотек генов человеческих иммуноглобулинов. Такие методы фаговых дисплеев, используемые для выделения человеческих антител, хорошо известны в данной области техники или описаны ниже в примерах. См. например: патенты США №№ 5223409; 5403484; и 5571698, Ladner et al; патенты США №№ 5427908 и 5580717, Dower et al; патенты США №№ 5969108 и 61721919, McCafferty et al.; и патенты США №№ 5885793; 6521404; 6544731; 6555313; 6582915 и 6593081, Griffiths et al.
[00197] Моноклональные человеческие антитела настоящего изобретения также можно получить с использованием мышей SCID, в организм которых введены человеческие иммунные клетки таким образом, чтобы при иммунизации получить человеческий гуморальный ответ. Такие мыши описаны, например, в патентах США №№ 5476996 и 5698767, Wilson et al.
[00198] Fab-фрагменты антител, или Fab, можно получить путем расщепления моноклональных антител папаином с последующей очисткой аффинной хроматографией. Fab также можно получить рекомбинантным синтезом с использованием нуклеиновых кислот, кодирующих Fab, как описано выше. Fab-фрагменты могут сохранять специфичность связывания и/или активность полноразмерной молекулы IgG, но имеют меньший размер и более низкую молекулярную массу, что может сделать их более пригодными для разных применений, чем полноразмерные молекулы IgG.
Получение каркасных участков или Fc-участков рекомбинантными способами
[00199] Рекомбинантные антитела настоящего изобретения и их антигенсвязывающие фрагменты можно получит путем модификаций каркасных остатков в домене VH и/или VL, например, с целью улучшения свойств антитела. Как правило, такие каркасные модификации уменьшают иммуногенность антитела. Например, один подход заключается в введении "обратных мутаций" по одному или нескольким каркасным остаткам с получением соответствующей первоначальной зародышевой последовательности. Более конкретно, антитело, которое было подвергнуто соматической мутации, может содержать каркасные остатки, которые отличаются от остатков зародышевой последовательности, из которой получено антитело. Такие остатки можно идентифицировать путем сравнения каркасных последовательностей антитела с зародышевыми последовательностями, из которых получено антитело. Чтобы вернуть последовательности каркасных участков к зародышевой структуре, соматические мутации можно подвергнуть "обратным мутациям" с получением зародышевой последовательности, например, путем сайт-направленного мутагенеза. Такие антитела, полученные с использованием "обратных мутаций", также входят в объем настоящего изобретения.
[00200] Другим типом каркасной модификации является изменение одного или нескольких остатков в каркасном участке, или даже в одном или нескольких участках CDR, с целью удаления Т-клеточных эпитопов и посредством этого уменьшения потенциальной иммуногенности антитела. Данный подход также упоминается как "деиммунизация" и более подробно описывается в публикации патента США № 20030153043, Carr et al.
[00201] В дополнение, или в качестве альтернативы модификациям каркасных участков или участков CDR, антитела настоящего изобретения можно сконструировать так, чтобы они содержали модификации в участке Fc, как правило, с целью изменения одного или нескольких функциональных свойств антитела, таких как период полужизни в сыворотке, способность фиксировать комплемент, способность связываться с рецептором Fc и/или антиген-зависимая клеточная цитотоксичность. Кроме того, антитело настоящего изобретения можно модифицировать химически (например, к антителу можно присоединить один или несколько химических фрагментов), или его можно модифицировать путем изменения гликозилирования, опять же, с целью изменения одного или нескольких функциональных свойств антитела. Каждый из таких вариантов осуществления более подробно описан ниже. Нумерацию остатков Fc-участка осуществляют по системе EU, описанной Кабат.
[00202] В одном варианте осуществления шарнирный участок CH1 модифицируют путем изменения, например увеличения или уменьшения, числа остатков цистеина. Данный подход описан более подробно в патенте США № 5677425, Bodmer et al. Число остатков цистеина в шарнирном участке CH1 изменяют, например, для облегчения сборки легкой и тяжелой цепей, или для увеличения или уменьшения стабильности антитела.
[00203] В другом варианте осуществления шарнирный участок Fc антитела подвергают мутациям, чтобы уменьшить период биологической полужизни антитела. Более конкретно, одну или несколько аминокислотных мутаций вводят в область поверхностного участка домена CH2-CH3 Fc-шарнирного фрагмента, чтобы уменьшить способность антитела связываться с белком A стафилококка (Staphylococcyl protein A) (SpA) по сравнению со способностью нативного Fc-шарнирного домена связывать SpA. Данный подход более подробно описан в патенте США № 6165745, Ward et al.
[00204] В другом варианте осуществления антитело модифицируют с целью увеличения периода его биологической полужизни. Для этого можно использовать разные подходы. Например, можно ввести одну или несколько из следующих мутаций: T252L, T254S, T256F, как описано в патенте США № 6277375, Ward. Альтернативно, чтобы увеличить период биологической полужизни, антитело можно изменить по участкам CH1 или CL, с введением эпитопа, с которым связывается рецептор реутилизации, образованного двумя петлями домена CH2 Fc-участка IgG, как описано в патентах США №№ 5869046 и 6121022, Presta et al.
[00205] В одном варианте осуществления участок Fc изменяют путем замены по меньшей мере одного аминокислотного остатка на другой аминокислотный остаток с целью изменения эффекторных функций антитела. Например, одну или несколько аминокислот можно заменить на другие аминокислотные остатки, так чтобы изменить сродство антитела к эффекторному лиганду, но сохранить антигенсвязывающую способность исходного антитела. Эффекторный лиганд, сродство к которому изменяется, может представлять собой, например, рецептор Fc или компонент C1 комплемента. Данный подход более подробно описан в патентах США №№ 5624821 и 5648260, Winter et al.
[00206] В другом варианте осуществления одну или несколько аминокислот можно заменить на другие аминокислотные остатки, так чтобы изменить способность антитела связываться с Clq и/или уменьшить или устранить комплементзависимую цитотоксичность (CDC). Данный подход более подробно описан в патенте США № 6194551, Idusogie et al.
[00207] В другом варианте осуществления изменяют один или несколько аминокислотных остатков, чтобы изменять способность антитела фиксировать комплемент. Данный подход более подробно описан в публикации PCT WO 94/29351, Bodmer et al.
[00208] В следующем варианте осуществления участок Fc модифицируют путем изменения одной или нескольких аминокислот так, чтобы увеличить способность антитела опосредовать антителозависимую клеточную цитотоксичность (ADCC) и/или увеличить сродство антитела к рецептору Fc-гамма. Данный подход более подробно описан в публикации PCT WO 00/42072, Presta. Кроме того, определены участки, по которым человеческий IgG1 связывается с Fc-гама RI, Fc-гама RII, Fc-гама RIII и FcRn, и описаны варианты с улучшенными параметрами связывания (см. Shields, R. L. et al., 2001 J. Biol. Chen. 276: 6591-6604).
[00209] В следующем варианте осуществления изменяют гликозилирование антитела. Например, можно получить агликозированное антитело (то есть, не гликозилированное антитело). Изменение гликозилирования можно проводить, например, с целью увеличения сродства антитела к антигену. Такие углеводные модификации можно осуществить, например, путем изменения одного или нескольких участков гликозилирования в последовательности антитела. Например, можно провести одну или несколько аминокислотных замен, приводящих к устранению одного или нескольких участков гликозилирования в каркасном участке вариабельного домена, и тем самым устранить гликозилирование в этом месте. Такое агликозилирование может увеличить сродство антитела к антигену. Данный подход более подробно описан в патентах США №№ 5714350 и 6350861, Co et al.
[00210] Дополнительно или альтернативно можно получить антитело, которое имеет измененный тип гликозилирования, такое как гипофукозилированное антитело, содержащее уменьшенное число фукозильных остатков, или антитело, имеющее увеличенное число точек ветвления GlcNac. Показано, что такие измененные типы гликозилирования повышают способность антител опосредовать ADCC. Такие углеводные модификации можно осуществить, например, путем экспрессии антитела в клетке-хозяине с измененным механизмом гликозилирования. Клетки с измененным механизмом гликозилирования описаны в данной области и могут использоваться в качестве клеток-хозяев для экспрессии рекомбинантных антител настоящего изобретения, в результате чего получают антитело с измененным гликозилированием. Например, в EP 1176195, Hang et al., описана клеточная линия с функционально блокированным геном FUT8, кодирующим фукозилтрансферазу, следовательно, в такой клеточной линии экспрессируюся гипофукозилированные антитела. В публикации РСТ WO 03/035835, Presta, описан вариант клеточной линии CHO, клетки LecB, характеризующиеся пониженной способностью прикреплять фукозу к Asn(297)-связанным углеводам, что также приводит к гипофукозилированию антител, экспрессируемых в данной клетке-хозяине (см. также Shields, R. L. et al., 2002 J. Biol. Chem. 277:26733-26740). В публикации РСТ WO 99/54342, Umana et al., описаны клеточные линии, полученные рекомбинантными методами для экспрессии гликопротеин-модифицирующих гликозилтрансфераз (например, бета(1,4)-N-ацетилглюкозаминилтрансферазы III (GnTIII)), в которых могут экспрессироваться антитела с повышенным числом точек ветвления GlcNac, что приводит к увеличению ADCC-активности антител (см. также Umana et al., 1999 Nat. Biotech. 17: 176-180).
Рекомбинантные способы получения измененных антител
[00211] Как указано выше, описанные здесь TSLP-связывающие антитела, содержащие последовательности VH и VL, или полноразмерные последовательности тяжелой и легкой цепей, можно использовать для получения новых TSLP-связывающих антител путем модификации полноразмерных последовательностей тяжелой цепи и/или легкой цепи, последовательностей VH и/или VL, или присоединенных к ним константных участков. Таким образом, в другом аспекте изобретения структурные особенности TSLP-связывающего антитела настоящего изобретения используют для получения структурно родственных TSLP-связывающих антител, которые сохраняют по меньшей мере одно функциональное свойство антител настоящего изобретения и их антигенсвязывающих фрагментов, такое как способность связываться с человеческим TSLP.
[00212] Например, один или несколько участков CDR из антител настоящего изобретения и их антигенсвязывающих фрагментов, или их мутантные формы, можно соединить с помощью рекомбинантных методов с известными каркасными участками, и/или другими CDR, с получением новых рекомбинантных TSLP-связывающих антител и антигенсвязывающих фрагментов настоящего изобретения, как описано выше. Другие типы модификаций включают в себя описанные в предыдущем разделе. В качестве исходного вещества в рекомбинантном способе можно использовать одну или несколько из представленных здесь последовательностей VH и/или VL, или один или несколько участков CDR. Чтобы создать рекомбинантное антитело, нет необходимости фактически получать (то есть экспрессировать в виде белка) антитело, содержащее одну или несколько из представленных здесь последовательностей VH и/или VL, или один или несколько участков CDR. Скорее, для получения последовательности (последовательностей) второго поколения из исходной последовательности (последовательностей), в качестве исходного материала используют информацию, содержащаяся в последовательности (последовательностях), с последующей продукцией последовательности (последовательностей) второго поколения и экспрессией ее в виде белка.
[00213] Измененную последовательность антитела также можно получить путем скрининга библиотек антител, содержащих фиксированные последовательности CDR3 или минимальные необходимые связывающие детерминанты, описанные в US20050255552, и последовательности, различающиеся по CDR1 и CDR2. Скрининг можно проводить с помощью любого метода, подходящего для скрининга библиотек антител, такого как технология фагового дисплея.
[00214] Для получения и экспрессии измененной последовательности антитела можно использовать стандартные методы молекулярной биологии. Антитело, кодируемое измененной последовательностью (последовательностями), представляет собой антитело, которое сохраняет одно функциональное свойство, несколько функциональных свойств, или все функциональные свойства описанных здесь TSLP-связывающих антител, причем функциональные свойства включают в себя, без ограничения, способность специфически связывать и стабилизировать человеческий белок TSLP.
[00215] Функциональные свойства измененных антител можно оценить с помощью стандартных анализов, известных в данной области и/или описанных здесь, таких как анализы, приведенные в примерах (например, ELISA).
[00216] В некоторых вариантах осуществления рекомбинантных способов получения антител настоящего изобретения и их антигенсвязывающих фрагментов мутации можно вводить произвольно или селективно по всей последовательности, кодирующей TSLP-связывающее антитело, или по ее части, после чего полученные модифицированные TSLP-связывающие антитела можно подвергнуть скринингу на связывающую активность и/или другие функциональные свойства, как описано в данном документе. Способы введения мутаций описаны в данной области. Например, в публикации PCT WO 02/092780, Short, описаны способы введения и скрининга мутаций антител с использованием насыщающего мутагенеза, сборки с синтетическим лигированием или их сочетания. Альтернативно, в публикации PCT WO 03/074679, Lazar et al., описаны способы применения компьютерных методов скрининга для оптимизации физико-химических свойств антител.
Характеристика антител настоящего изобретения
[00217] Антитела и антигенсвязывающие фрагменты настоящего изобретения могут быть охарактеризовать с помощью разных функциональных анализов. Например, их можно охарактеризовать по способности связывать TSLP и ингибировать активность TSLP.
[00218] Способность антитела связываться с TSLP можно детектировать путем непосредственного мечения представляющего интерес антитела, или антитело может быть немеченым и тогда связывание детектируют косвенно с использованием разных форматов сэндвич-анализа, известных в данной области.
[00219] В некоторых вариантах осуществления TSLP-связывающие антитела настоящего изобретения и их антигенсвязывающие фрагменты блокируют связывание исходного TSLP-связывающего антитела с полипептидом TSLP, или конкурируют с исходным TSLP-связывающим антителом за связывание с полипептидом TSLP. Они могут представлять собой описанные выше полностью человеческие или гуманизированные TSLP-связывающие антитела. Они также могут представлять собой другие человеческие, мышиные, химерные или гуманизированные TSLP-связывающие антитела, способные связываться с тем же эпитопом, что и исходное антитело. Способность блокировать связывание исходного антитела, или конкурировать с исходным антителом за связывание, указывает на то, что тестируемое TSLP-связывающее антитело связывается с тем же или подобным эпитопом, что и исходное антитело, или с эпитопом, достаточно близким к эпитопу, с которым связывается исходное TSLP-связывающее антитело. Такие антитела с большой вероятностью могут обладать полезными свойствами, идентифицированными у исходного антитела. Способность блокировать исходное антитело, или конкурировать с исходным антителом, может определить, например, путем анализа конкурентного связывания. При анализе конкурентного связывания тестируемое антитело исследуют на способность ингибировать специфическое связывание исходного антитела с общим антигеном, таким как полипептид TSLP. тестируемое антитело конкурирует с исходным антителом за специфическое связывание с антигеном, если избыток тестируемого антитела значительно ингибирует связывание исходного антитела. Значительное ингибирование означает, что тестируемое антитело снижает специфическое связывание исходного антитела, как правило, по меньшей мере на 10%, 25%, 50%, 75% или 90%.
[00220] Существует ряд известных анализов конкурентного связывания, которые можно использовать для оценки конкуренции антитела с исходным антителом за связывание с конкретным белком, в данном случае TSLP. К ним относятся, например, твердофазный, прямой или косвенный радиоиммуноанализ (RIA), твердофазный, прямой или косвенный иммуноферментный анализ (EIA), конкурентный сэндвич-анализ (см. Stahli et al., Methods in Enzymology, 9: 242-253, 1983); твердофазный прямой EIA с использованием биотина-авидина (см. Kirkland et al., J. Immunol., 137: 3614-3619, 1986); твердофазный прямой анализ с использованием метки, твердофазный прямой сэндвич-анализ с использованием метки (см. Harlow & Lane, выше); твердофазный прямой RIA с использованием I-125 в качестве метки (см. Morel et al., Molec. Immunol. 25: 7-15, 1988); твердофазный прямой EIA с использованием биотина-авидина (Cheung et al., Virology 176: 546-552, 1990); и прямой RIA с использованием метки (Moldenhauer et al., Scand. J. Immunol. 32: 77-82, 1990). Как правило, такой анализ включает в себя использование очищенного антигена, связанного с твердой поверхностью, или клеток, несущих антиген, немеченого тестируемого TSLP-связывающего антитела и меченого исходного антитела. Конкурентное ингибирование измеряют путем определения количества метки, связанной с твердой поверхностью или клетками, в присутствии тестируемого антитела. Обычно тестируемое антитело присутствует в избытке. Антитела, идентифицируемые с помощью конкурентного анализа (конкурирующие антитела), включают в себя антитела, связывающиеся с тем же эпитопом, что и исходное антитело, а также антитела, связывающиеся с соседним эпитопом, достаточно близким к эпитопу, с которым связывается исходное антитело, для стерического затруднения.
[00221] Чтобы определить, связываются ли выбранные TSLP-связывающие моноклональные антитела с уникальными эпитопами, каждое антитело можно биотинилировать с использованием коммерчески доступных реагентов (например, реагентов от Pierce, Rockford, 111.). Исследования конкуренции с использованием немеченых моноклональных антител и биотинилированных моноклональных антител можно проводить с использованием планшетов ELISA, покрытых полипептидом TSLP. Связывание биотинилированных MAb можно детектировать с помощью зонда стрептавидин-щелочная фосфатаза. Изотип очищенного TSLP-связывающего антитела можно определить методом ELISA. Например, лунки микротитрационных планшетов можно покрыть антителом против человеческих IgG в концентрации 1 мкг/мл и инкубировать в течение ночи при 4°С. После блокирования 1% BSA планшеты подвергают взаимодействию с моноклональным TSLP-связывающим антителом, или очищенными изотипическими контролями, в концентрации 1 мкг/мл или менее, при температуре окружающей среды, в течение одного-двух часов. Затем лунки можно подвергнуть взаимодействию с зондами, конъюгированными со щелочной фосфатазой, специфичными либо к человеческому IgG1, либо к человеческому IgM. Затем планшеты оставляют для развития окраски и анализируют, чтобы определить изотип очищенного антитела.
[00222] Чтобы продемонстрировать связывание моноклональных TSLP-связывающих антител с живыми клетками, экспрессирующими полипептид TSLP, можно использовать проточную цитометрию. Коротко говоря, клеточные линии, экспрессирующие TSLP (выращенные в стандартных условиях), можно смешать с разными концентрациями TSLP-связывающего антитела в PBS, содержащем 0,1% BSA и 10% фетальной телячьей сыворотки, и инкубировать при 37°C в течение 1 часа. После промывания клетки подвергают взаимодействию с меченым флуоресцеином антителом против человеческого IgG в тех же условиях, что и окрашивание первичным антителом. Образцы можно проанализировать с помощью прибора FACScan с использованием рассеяния света и бокового рассеяния, чтобы пропускать отдельные клетки. При проведении проточной цитометрии можно использовать (помимо или вместо) альтернативный анализ с использованием флуоресцентной микроскопии. Клетки окрашивают точно так, как описано выше, и анализируют с помощью флуоресцентной микроскопии. Этот метод позволяет визуализировать отдельные клетки, но может иметь низкую чувствительность в зависимости от плотности антигена.
[00223] TSLP-связывающие антитела и антигенсвязывающие фрагменты настоящего изобретения можно дополнительно протестировать на способность взаимодействовать с полипептидом или антигенным фрагментом TSLP методом вестерн-блоттинга. Коротко говоря, получают очищенные полипептиды или гибридные белки TSLP, или экстракты клеток, экспрессирующих TSLP, и подвергают их электрофорезу в полиакриламидном геле в присутствии додецилсульфата натрия. После электрофореза разделенные антигены переносят на нитроцеллюлозные мембраны, блокируют 10% фетальной телячьей сывороткой и обрабатывают моноклональными антителами, подлежащими тестированию. Связывание человеческих IgG можно детектировать с использованием антитела против человеческого IgG-щелочной фосфатазы и таблеток субстрата BCIP/NBT (Sigma Chem. Co., St. Louis, Мо.).
[00224] Примеры функциональных анализов также описаны ниже в разделе Примеры.
Фармацевтические композиции и составы
[00225] Изобретение также предлагает композиции, например фармацевтические композиции, содержащие в качестве активного ингредиента одну или несколько молекул, например, антитела, фрагменты антител, такие как Fab, Fab', F(ab')2, scFv, миниантитела или диатела, способные специфически связывать TSLP.
[00226] Фармацевтические композиции обычно содержат фармацевтически приемлемое вспомогательное вещество. Фармацевтически приемлемое вспомогательное вещество может включать в себя физиологический раствор, растворители, дисперсионные среды, покрытия, противобактериальные и противогрибковые средства, изотонические и замедляющие абсорбцию средства и т.п., совместимые с фармацевтическим введением. Состав фармацевтических композиций обычно соответствует предполагаемому способу введения. Например, при введении путем ингаляции соединения могут доставляться в виде аэрозольной струи из находящегося под давлением контейнера или дозатора, который содержит подходящий пропеллент, например газ, такой как диоксид углерода, или из распылителя. Такие способы включают в себя способы, описанные в патенте США № 6468798.
[00227] В некоторых вариантах осуществления предлагаемые здесь фармацевтические композиции получают в виде формы, подходящей для направленной доставки в дыхательные пути индивидуума, особенно в легкие индивидуума. Такие композиции позволяют избежать осаждения активного ингредиента в верхних дыхательных путях индивидуума, тем самым сводя к минимуму проблемы переносимости или безопасности, связанные с осаждением лекарственного средства во рту и горле. В некоторых вариантах осуществления представленные здесь фармацевтические композиции находятся в виде сухого порошкообразного состава. Такой сухой порошкообразный состав может содержать активный ингредиент, оболочкообразующее вспомогательное вещество, стеклообразующее вспомогательное вещество и буфер.
Активный ингредиент
[00228] Активные ингредиенты сухого порошкообразного состава могут включать одно или несколько из описанных здесь антител и фрагментов антител против TSLP.
[00229] Количество активного ингредиента в фармацевтической композиции можно регулировать так, чтобы обеспечить доставку терапевтически эффективного количества активного ингредиента на единицу дозы, необходимого для достижения желательного результата. На практике это количество значительно варьирует в зависимости от конкретного ингредиента, его активности, тяжести состояния, подлежащего лечению, популяции пациента, требований дозирования, желательного терапевтического эффекта и относительного количества добавок, содержащихся в композиции. Композиция обычно содержит примерно от 1 масс.% до 99 масс.% активного ингредиента, например, примерно от 5% до 95%, примерно от 10% до 90%, примерно от 15% до 85%, примерно от 20% до 80%, примерно от 25% до 75%, примерно от 30% до 70%, примерно от 40% до 60%, или примерно 50% по массе активного ингредиента. Композиции настоящего изобретения предпочтительно используют для доставки активных ингредиентов в дозах от 0,001 мг/день до 100 мг/день, предпочтительно в дозах от 0,01 мг/день до 75 мг/день, и более предпочтительно в дозах от 0,10 мг/день до 50 мг/день. Следует понимать, что описанные здесь композиции могут содержать несколько активных ингредиентов, и что термин "активный ингредиент" никоим образом не исключает использования двух или более таких активных ингредиентов.
Вспомогательные вещества
[00230] В некоторых вариантах осуществления описанный здесь сухой порошкообразный состав содержит фармацевтически приемлемое гидрофобное оболочкообразующее вспомогательное вещество. Оболочкообразующие вспомогательные вещества представляют собой поверхностно-активные средства, которые усиливают диспергируемость высушенных распылением порошков. Гидрофобное оболочкообразующее вспомогательное вещество может принимать разные формы, которые зависят, по меньшей мере, отчасти от состава и предполагаемого использования сухого порошкообразного состава. Подходящие фармацевтически приемлемые гидрофобные вспомогательные вещества, как правило, могут быть выбраны из группы, состоящей из длинноцепочечных фосфолипидов, гидрофобных аминокислот и пептидов и мыл, содержащих длинноцепочечные жирные кислоты.
[00231] В некоторых вариантах осуществления оболочкообразующие вспомогательные вещества включают в себя: глицин, аланин, валин, трилейцин, дилейцин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан, дипальмитоилфосфатидилхолин (DPPC), дистеароилфосфатидилхолин (DSPC) и стеарат магния. В некоторых вариантах осуществления описанные здесь сухие порошкообразные составы содержат трилейцин.
[00232] Контролируя состав и процесс, можно достичь того, что поверхность высушенных распылением частиц будет состоять в основном из оболочкообразующего вспомогательного вещества. Поверхностная концентрация может составлять более 70%, например, более 75%, или 80%, или 85%. В некоторых вариантах осуществления поверхность более чем на 90% состоит из оболочкообразующего вспомогательного вещества, или более на 95%, или 98%, или 99% из гидрофобного вспомогательного вещества. В случае высоко активных ингредиентов поверхность нередко содержит более 95% оболочкообразующего вспомогательного вещества.
[00233] В некоторых вариантах осуществления оболочкообразующее вспомогательное вещество содержит более 70% частиц, образующих граничную поверхность, по данным электронной спектроскопии для химического анализа (ESCA, также известной как рентгеновская фотоэлектронная спектроскопия или XPS), предпочтительно более 90% или 95%.
[00234] В некоторых вариантах осуществления оболочкообразующее вспомогательное вещество облегчает развитие шероховатой морфологии частиц. Это означает, что морфология частиц характеризуется как пористая, морщинистая, ребристая или мятая, но не гладкая. Это означает, что внутренняя и/или внешняя поверхность частиц ингаляционного лекарственного средства являются по меньшей мере частично шероховатыми. Шероховатость способствует высокой эффективности доставки, обеспечивает однородность доз и направленность лекарственного средства путем улучшения флюидизации и диспергируемости порошка. Увеличение шероховатости частиц приводит к уменьшению сил сцепления между частицами, поскольку они не могут приблизиться друг к другу на расстояние, обеспечивающее возникновение ван-дер-ваальсовых взаимодействий. Уменьшение сил сцепления является достаточным для того, чтобы резко улучшить флюидизацию и дисперсию порошка, состоящего из множества шероховатых частиц.
[00235] Если присутствует оболочкообразующее вспомогательное вещество, его содержание обычно варьирует примерно от 15 до 50% масс./масс. по отношению к лекарственному средству. Если в состав композиции входит трилейцин, его содержание должно составлять минимум примерно 15%, чтобы обеспечить приемлемую эффективность в качестве оболочкообразующего вещества. Минимальное содержание лейцина составляет более чем примерно 30%.
[00236] Применение гидрофобных оболочкообразующих вспомогательных веществ, таких как трилейцин, может быть ограничено их растворимостью в жидком сырье. Как правило, содержание трилейцина в готовом порошке составляет менее 30% масс./масс., чаще примерно от 10 масс.% до 20 масс.% (примерно 10-30 масс.%). Вследствие ограниченной растворимости в воде и поверхностной активности трилейцин является превосходным обболочкообразующим веществом. Лейцин также можно использовать в качестве оболочкообразующего вспомогательного вещества, причем варианты осуществления изобретения могут включать в себя частицы, содержащие лейцин в концентрации, составляющей примерно от 50% до 75%.
[00237] Жирнокислотные мыла ведут себя аналогично лейцину и трилейцину и, следовательно, являются подходящими модификаторами поверхности.
[00238] Вследствие краткой временной шкалы события сушки активные ингредиенты, растворенные в исходном сырье, обычно присутствуют в виде аморфных твердых веществ в высушенном распылением лекарственном продукте.
[00239] Подвижность молекул в аморфном твердом веществе гораздо выше, чем в его кристаллическом аналоге. Подвижность молекул включает в себя движения дальнего радиуса действия, связанные с молекулярной диффузией, а также локальные движения, такие как вращения вокруг связей. Основным принципом твердофазной стабилизации аморфных веществ является то, что молекулярная подвижность приводит к нежелательным физическим и химическим изменениям. Поэтому стратегии получения композиций аморфных веществ обычно направлены на подавление подвижности молекул.
[00240] Существование взаимосвязи между молекулярной подвижностью и неустойчивостью является интуитивным и хорошо известным. Однако, чтобы молекулярная подвижность была полезной, ее нужно тщательно определить и понять с точки зрения типов присутствующих движений. Движения молекул дальнего радиуса действия возникают вследствие структурной релаксации, известной как α-релаксация. Временные рамки для таких движений заметно возрастают, когда температура падает ниже температуры стеклования (Tg), или наоборот, когда Tg повышается при фиксированной температуре наблюдения. Поскольку стабилизация молекулы в стекле ограничивает молекулярную подвижность дальнего радиуса действия, это стало наиболее распространенной стратегией твердофазной стабилизации аморфных лекарственных средств при получении композиций.
[00241] Подавление молекулярной подвижности в твердом состоянии посредством стабилизации в стекле, например, путем применения стеклообразующих средств, может улучшить физико-химическую стабильность белка в композиции. Если требуется стеклообразующее средство, его выбор будет определять множество факторов. Основная роль стеклообразующего вспомогательного вещества заключается в снижении общей молекулярной подвижности лекарственного средства большого радиуса действия. На практике это достигается повышением температуры стеклования аморфной фазы, содержащей лекарственное средство. Хотя, как правило, желательно использовать вспомогательные вещества с высокими значениями Tg, для некоторых композиций (например, лекарственных средств со средними значениями Tg, или для композиций, содержащих лекарственное средство в низкой концентрации) можно использовать вспомогательные вещества со средними значениями Tg. В качестве руководства для производителя стоит подчеркнуть свойства идеального стеклообразующего средства: биосовместимое вещество с высокой температурой стеклования, которое смешивается с лекарственным средством, образуя единую аморфную фазу, слабо пластифицируемую водой.
[00242] В некоторых вариантах осуществления описанные здесь сухие порошкообразные составы содержат стеклообразующее вспомогательное вещество. Стеклообразующие вспомогательные вещества, которые подавляют движения молекул большого радиуса действия, включают в себя углеводы, аминокислоты и буферы. В некоторых вариантах осуществления стеклообразующие вспомогательные вещества включают в себя гистидин, гистидин HCl, сахарозу, трегалозу, маннит и цитрат натрия. Таким образом, некоторые вспомогательные вещества, такие как гистидин, можно назвать как буфером, так и стеклообразующим вспомогательным веществом. В некоторых вариантах осуществления описанные здесь сухие порошкообразные составы, например, составы, состоящие из ядра-оболочки, содержат трегалозу.
[00243] Важность других типов молекулярных движений становится все более очевидной в фармацевтической литературе. Номенклатура (α, β и т.д.), используемая для обозначения типов молекулярных движений, исходит из широкополосной диэлектрической спектроскопии. Спектры диэлектрической релаксации обычно изображают на частотной шкале. При интерпретации этих спектров пики диэлектрических потерь на самых низких частотах обозначают как α-движения, движения на более высоких частотах как β-движения, затем γ и т.д. Таким образом, β и другие движения, которые происходят на более высоких частотах, называются "быстрыми" или вторичными движениями (а в некоторых случаях релаксациями Джохари-Голдстайна). Хотя указанные вторичные релаксации часто приписывают внутримолекулярным движениям разных молекулярных групп (например, боковых цепей на белке), они существуют даже у жестких молекул. В упрощенной физической картине β-движения иногда описываются как случайное "дрожание" молекул, уловленной среди ближайших соседей. В какой-то момент локальные движения ближайших соседей могут обеспечить свободный объем, достаточный для диффузионного скачка уловленных молекул. Это движение. Таким образом, β-движения приводят к α-движениям.
[00244] Вторичные движения - это область активных исследований как с теоретической, так и с практической точки зрения. И хотя большая часть литературы относится к лиофилизированным или закаленным из расплава стеклам, внимание также уделяется аморфным, рекомбинантным частицам для ингаляции (например, порошкам, полученным с использованием сушки распылением или некоторых других восходящих процессов). Предполагается, что кристаллизация небольших молекул вблизи Tg происходит вследствие β-движений. Специалистам в области белковых композиций известна важность контроля этих β-движений. Подавление β-движений в аморфных композициях обычно осуществляют путем использования небольших органических вспомогательных веществ, таких как глицерин, маннит, сорбит и диметилсульфоксид. Хотя они наиболее часто упоминаются как вспомогательные вещества для подавления β-движений, для этой цели могут служить и другие низкомолекулярные органические молекулы (например, буферные соли или противоионы). Предполагается, что указанные вспомогательные вещества подавляют движения высокоподвижных доменов путем повышения локальной вязкости. Для читателя, знакомого с обширной литературой по стабилизации стеклом, применение таких вспомогательных веществ может показаться нелогичным. Указанные вещества и большинство других низкомолекулярных соединений имеют низкие значения Tg и способствуют уменьшению Tg композиции, явлению, известному как пластификация. Однако эти вспомогательные вещества также могут уменьшать β-движения. Таким образом, они упоминаются как антипластификаторы, а иногда как пластификаторы, в зависимости от точки отсчета; хотя они пластифицируют α-движения, они антипластифицируют β-движения. Следует отметить, что данная терминология является потенциальным источником путаницы в литературе; указание вещества как пластификатора или антипластификатора зависит от того, является ли его точка отсчета α-движениями или вторичными движениями.
[00245] Поскольку для твердофазной стабилизации белков нужна композиция на стеклообразной основе, вклад α- и β-движений представляет особый интерес. Хотя в литературе присутствует множество ссылок на применение стеклообразующих средств для стабилизации белков, до недавнего времени было мало конкретных ссылок на влияние этих средств на локальные движения. Хотя температуру стеклования белков трудно измерить, большинство данных свидетельствуют о том, что Tg >150°C. Таким образом, вспомогательные вещества (например, дисахариды, такие как сахароза или трегалоза), наиболее часто используемые для стабилизации белков, также пластифицируют α-движения в белке (и антипластифицируют вторичные движения). В недавно опубликованной работе показано, что β-движения в значительной степени определяют стабильность белков в сахарных стеклах. Таким образом, дисахариды антипластифицируют β-движения в белковых композициях.
[00246] В некоторых вариантах осуществления описанные здесь сухие порошкообразные составы содержат стеклообразующие вспомогательные вещества с высокой температурой стеклования (>80°C). В некоторых вариантах осуществления описанные здесь сухие порошкообразные составы содержат такие стеклообразующие средства, как сахароза, трегалоза, маннит, фумарилдикетопиперазин и цитрат натрия.
[00247] Для достижения оптимальной стабилизации аморфного твердого вещества можно использовать смеси стеклообразующих средств. В некоторых вариантах осуществления в качестве основы для композиций, содержащих ядро и оболочку, используют смеси трегалозы и маннита.
[00248] Количество стеклообразующего средства, необходимое для подавления молекулярной подвижности и достижения физической и химической стабильности, зависит от природы активного средства. В некоторых вариантах осуществления композиций, содержащих высушенные распылением белки, молярное отношение стеклообразующего средства к белку может находиться в диапазоне от 300 до 900. В случае небольших молекул необходимое количество стеклообразующего средства зависит от Tg активного средства.
[00249] В некоторых вариантах осуществления описанные здесь сухие порошкообразные составы содержат буфер. Буферы хорошо известны как средства, регулирующие рН, однако их также можно использовать в качестве средств доставки лекарственного средства при физиологически совместимых pH (то есть для повышения переносимости), и для обеспечения условий в растворе, благоприятных для химической стабильности лекарственного средства. В некоторых описанных здесь композициях и способах рН среды, содержащей лекарственное средство, можно контролировать путем совместного использования лекарственного средства и буфера в одной частице.
[00250] Хотя значение рН для твердого лекарственного продукта находится под вопросом, ряд исследований демонстрируют важность регуляции рН для химической стабильности в твердом состоянии. Вода присутствует везде, даже в "сухих" порошкообразных составах, находящихся в твердом состоянии. В дополнение к своей роли как пластификатора аморфных веществ вода является реагентом, продуктом разложения и может также служить в качестве среды для растворения и химических реакций. Имеются данные о том, что адсорбция воды на поверхности частиц может привести к образованию насыщенного раствора в поверхностной пленке. Действительно, в некоторых исследованиях используют рН суспензии лекарственного средства (то есть насыщенного раствора) в качестве показателя локального рН, или рН "микроокружения" лекарственного средства, растворенного в поверхностной пленке, присутствующей в "сухом" порошке. Показано, что в некоторых случаях рН микроокружения имеет значение для стабильности лекарственного средства.
[00251] Как и лекарственное средство, вспомогательные вещества также растворяются в поверхностной пленке адсорбированной воды с образованием насыщенного раствора. Это можно использовать для регуляции локального рН в адсорбированном слое влаги. Буферы или модификаторы рН, такие как гистидин или фосфат, обычно используют в лиофилизированных или высушенных распылением композициях для контроля химической деградации белков в растворе и твердом состоянии.
[00252] В некоторых вариантах осуществления буферы для композиции включают в себя: гистидин, глицин, ацетат и фосфат.
[00253] Дополнительные вспомогательные вещества включают соли (например, хлорид натрия, хлорид кальция, цитрат натрия), антиоксиданты (например, метионин), вспомогательные вещества, уменьшающие агрегацию белков в растворе (например, аргинин), средства, корректирующие вкус лекарственного средства, а также средства, улучшающие абсорбцию макромолекул в системном кровотоке (например, фумарилдикетопиперазин).
Композиция
[00254] Настоящее изобретение предлагает сухие порошкообразные составы, содержащие высушенные распылением частицы, которые позволяют избежать осаждения в ротоглотке среднего взрослого индивидуума и обеспечить целенаправленную доставку лекарственного средства в легкие.
[00255] В некоторых вариантах осуществления для описанных здесь частиц сухих порошкообразных составов общая легочная доза (TLD) in vitro составляет от 80 до 95% масс./масс. от номинальной дозы, например, от 85 до 90% масс./масс. для среднего взрослого индивидуума.
[00256] В некоторых вариантах осуществления для описанных здесь частиц сухих порошкообразных составов общая легочная доза (TLD) in vitro составляет от 90 до 100 масс.% от доставляемой дозы, например, от 90 до 95% масс./масс. для среднего взрослого индивидуума.
[00257] В некоторых вариантах осуществления инерционный параметр доставляемой дозы описанной здесь сухого порошкообразного состава предпочтительно составляет от 120 до 400 мкм2 л/мин, например, от 150 до 300 мкм2 л/мин.
[00258] В некоторых вариантах осуществления описанные здесь сухие порошкообразные составы содержат сконструированные частицы, имеющие пористую, ребристую или шероховатую поверхность. Такие частицы характеризуются уменьшением сил сцепления между частицами по сравнению с тонкоизмельченными кристаллами лекарственного средства с сопоставимым размером первичных частиц. Это обеспечивает улучшение флюидизации и диспергируемости порошка по сравнению с упорядоченными смесями тонкоизмельченного лекарственного средства и крупнозернистой лактозы.
[00259] В некоторых вариантах осуществления частицы описанных здесь сухих порошкообразных составов имеют шероховатость более 1,5, например, от 1,5 до 20, от 3 до 15, или от 5 до 10.
[00260] Для некоторых активных фармацевтических ингредиентов, например, многих пептидов или белков (таких как Fab против TSLP), шероховатую поверхность можно получить путем распылительной сушки чистого препарата. В таком случае композиция может содержать чистый препарат, составляющий 100% масс./масс. от активного средства или лекарственного средства.
[00261] В некоторых вариантах осуществления описанные здесь сухие порошкообразные составы содержат лекарственное средство и буфер. Композиция может содержать от 70% до 99% масс./масс. лекарственного средства или активного средства, а остальную часть составляет буфер.
[00262] В некоторых вариантах осуществления описанные здесь композиции могут содержать от 0,1 до 99 масс.% активного средства, или от 0,1 до 70 масс.% активного средства, или от 0,1 до 50 масс.% активного ингредиента (активных ингредиентов), или от 0,1% до 30% масс./масс. активного ингредиента (активных ингредиентов).
[00263] В некоторых вариантах осуществления описанные здесь сухие порошкообразные составы могут содержать вспомогательные вещества, дополнительно повышающие стабильность или биосовместимость композиции. Например, изобретение охватывает разные соли, буферы, антиоксиданты, оболочкообразующие вспомогательные вещества и стеклообразующие вспомогательные вещества.
[00264] В некоторых вариантах осуществления частицы описанных здесь сухих порошкообразных составов имеют геометрический размер, выраженный в виде массового медианного диаметра (x50), от 0,8 до 2,0 мкм, например, от 1,0 до 1,5 мкм.
[00265] В некоторых вариантах осуществления частицы описанных здесь сухих порошкообразных составов имеют геометрический размер, выраженный в виде x90, от 2,0 мкм до 4,0 мкм, например, от 2,5 до 3,5 мкм.
[00266] В некоторых вариантах осуществления частицы описанных здесь сухих порошкообразных составов имеют насыпную плотность (ρtapped) от 0,03 до 0,40 г/см3, например, от 0,07 до 0,30 г/см3.
[00267] В некоторых вариантах осуществления первичные частицы описанных здесь сухих порошкообразных составов имеют рассчитанный средний аэродинамический размер (Da) от 0,1 до 1,0 мкм, например, от 0,5 до 0,8 мкм.
[00268] В некоторых вариантах осуществления частицы описанных здесь сухих порошкообразных составов имеют рассчитанный аэродинамический диаметр от 0,5 до 1,2 мкм, например, от 0,8 до 1,0 мкм.
[00269] В некоторых вариантах осуществления совокупность частиц описанных здесь сухих порошкообразных составов, присутствующих в доставляемой дозе, предпочтительно имеет массовый медианный аэродинамический диаметр (MMAD) от 1,0 до 3,0 мкм, например, от 1,5 до 2,0 мкм.
[00270] В некоторых вариантах осуществления композиция настоящего изобретения содержит частицы, имеющие оболочку и ядро: трилейцин, как оболочкообразующее средство, присутствует на поверхности частицы, а ядро содержит активный ингредиент (например, Fab против TSLP), трегалозу, или сочетание трегалозы и маннита, и буфер.
[00271] В некоторых вариантах осуществления изобретение предлагает композицию, содержащую примерно 40% (масс./масс.) TSLP-связывающей молекулы, например, Fab1 против TSLP, примерно 25% (масс./масс.) трилейцина, примерно 30% (масс./масс.) трегалозы и маннита и примерно 5% (масс./масс.) гистидина. В других вариантах осуществления настоящее изобретение предлагает композицию, содержащую примерно 50% (масс./масс.) TSLP-связывающей молекулы, примерно 15% (масс./масс.) трилецинина, примерно 2,6% (масс./масс.) HCl, примерно 5,6% масс.) гистидина и примерно 26,8% (масс./масс.) в сумме трегалозы и основания; или примерно 50% (масс./масс.) TSLP-связывающей молекулы, примерно 15% (масс./масс.) трилейцина, примерно 19,4% (масс./масс.) трегалозы, примерно 13% (масс./масс.) гистидина и примерно 2,6% (масс./масс.) HCl.
[00272] В других вариантах осуществления настоящая заявка раскрывает фармацевтический порошкообразный состав, не содержащий носителей, частицы которого, доставляемые из ингалятора сухого порошка, содержат раскрытые здесь молекулы против TSLP, где общая легочная доза in vitro превышает 90% доставляемой дозы, и где частицы в доставляемой дозе имеют инерционный параметр от 120 до 400 мкм2 л/мин.
[00273] В другом варианте осуществления настоящая заявка раскрывает фармацевтическую композицию, не содержащую носителей, доставляемую с помощью ингалятора сухого порошка, причем композиция содержит совокупность частиц, состоящих из ядра, содержащего описанную здесь молекулу против TSLP, и, по меньшей мере, одно стеклообразующее вспомогательное вещество, и оболочки, содержащей гидрофобное вспомогательное вещество и буфер; и где общая легочная доза in vitro превышает 90% масс./масс. от доставляемой дозы. В некоторых вариантах осуществления частицы получают путем распылительной сушки. В другом варианте осуществления гидрофобное вспомогательное вещество включает в себя трилейцин.
[00274] В следующем варианте осуществления настоящая заявка раскрывает фармацевтическую композицию, не содержащую носителей, содержащую совокупность первичных частиц и агломератов частиц, доставляемых из ингалятора сухого порошка, где композиция содержит описанную здесь молекулу против TSLP, и где общая легочная доза in vitro (TLD) составляет более 80% от номинальной дозы, а первичные частицы характеризуются: ребристой морфологией; медианным аэродинамическим диаметром (Da) от 0,3 до 1,0 мкм; и где частицы и агломераты частиц, доставляемые из ингалятора сухого порошка, имеют массовый медианный аэродинамический диаметр (MMAD) от 1,5 до 3,0 мкм. В некоторых вариантах осуществления фармацевтическая композиция дополнительно содержит сосуд для первичных частиц, подходящий для хранения частиц до их аэрозолизации в ингаляторе сухого порошка, и где аэрозоль, содержащий вдыхаемые агломераты, образуется в процессе указанной аэрозолизации.
[00275] В следующем варианте осуществления настоящая заявка раскрывает фармацевтический порошкообразный состав для легочной доставки, где частицы порошка содержат от 1 до 100 масс.% описанной здесь молекулы против TSLP, порошок характеризуется следующим распределением частиц по размеру: по меньшей мере 50% от 1 до 1,5 микрон, а плотность порошка составляет от 0,05 до 0,3 г/см3, аэродинамический диаметр составляет менее 2 мкм, шероховатость от 1,5 до 20; и где порошок вводят путем ингаляции, обеспечивая общую легочную дозу in vitro более 80%. В некоторых вариантах осуществления фармацевтический порошкообразный состав не содержит носитель. В других вариантах осуществления порошок упаковывают в сосуд для применения с помощью ингалятора сухого порошка, причем после аэрозолизации с помощью указанного ингалятора сухого порошка порошок характеризуется наличием подходящих для вдыхания агломератов, имеющих средний аэродинамический диаметр менее, чем примерно 2 мкм.
Способ
[00276] Изобретение также относится к способу получения сухих порошкообразных составов для ингаляции, содержащих высушенные распылением частицы, где композиция содержит по меньшей мере один активный ингредиент и характеризуется общей легочной дозой (TLD) in vitro от 80 до 95% масс./масс., например, от 85 до 90% масс./масс. от номинальной дозы для среднего взрослого индивидуума.
[00277] Кроме того, изобретение предлагает способы получения сухих порошкообразных составов для ингаляции, содержащих высушенные распылением частицы, где композиция содержит по меньшей мере один активный ингредиент и характеризуется общей легочной дозой (TLD) in vitro от 90 до 100 масс.%, например от 90 до 95% масс./масс. от доставляемой дозы для среднего взрослого индивидуума.
[00278] В некоторых вариантах осуществления сухие порошкообразные составы содержат по меньшей мере один активный ингредиент, который подходит для лечения обструктивных или воспалительных заболеваний дыхательных путей, в частности, астмы и/или COPD, например, Fab против TSLP. В некоторых вариантах осуществления сухие порошкообразные составы содержат по меньшей мере один активный ингредиент, который подходит для неинвазивного лечения заболеваний в системном кровотоке.
[00279] Сушка распылением, используемая при получении сконструированных частиц для ингаляции, имеет преимущества, поскольку она позволяет быстро получить сухой порошок и контролировать такие характеристики частиц, как размер, морфология, плотность и поверхностный состав. Процесс сушки протекает очень быстро (порядка миллисекунд). В результате большинство активных ингредиентов, растворенных в жидкой фазе, осаждаются в виде аморфных твердых веществ, поскольку для кристаллизации не хватает времени.
[00280] Сушка распылением включает в себя четыре отдельные операции: приготовление исходного сырья, распыление исходного сырья с получением капель размером несколько микрон, сушка капель в горячем газе и сбор высушенных частиц с помощью мешка или циклонного сепаратора.
[00281] В некоторых вариантах осуществления способы получения частиц сухого порошка включают в себя три стадии, однако в некоторых вариантах осуществления две или даже все три из этих стадий можно проводить практически одновременно, поэтому на практике способ фактически можно считать одностадийным. Исключительно в целях описания способа настоящего изобретения три стадии описаны отдельно, но такое описание не предназначено для ограничения трехстадийного способа.
[00282] В некоторых вариантах осуществления способ включает в себя приготовление раствора исходного сырья и распылительную сушку исходного сырья с получением активных частиц сухого порошка. Исходное сырье содержит, по меньшей мере, один активный ингредиент, растворенный в жидкой среде на водной основе. В некоторых вариантах осуществления исходное сырье содержит по меньшей мере один активный ингредиент (например, Fab1 против TSLP), растворенный в исходной среде на водной основе, содержащей добавленный сорастворитель. В некоторых вариантах осуществления исходное сырье содержит по меньшей мере один активный ингредиент, растворенный в исходной среде этанол/вода, где доля этанола составляет от 5 до 30 масс.%, например, от 5 до 20 масс.%.
[00283] В случае аморфных твердых веществ важно контролировать содержание влаги в лекарственном продукте. Для лекарственных средств, которые не являются гидратами, содержание влаги в порошке предпочтительно составляет менее 5%, чаще менее 3%, или даже 2% масс./масс. Однако содержание влаги должно быть достаточно высоким, чтобы гарантировать отсутствие в порошке значительных электростатических сил притяжения. Содержание влаги в высушенных распылением порошках можно определить путем титрования по методу Карла Фишера.
[00284] В некоторых вариантах осуществления сырье распыляют в потоке теплого отфильтрованного воздуха, который испаряет растворитель и передает высушенный продукт в коллектор. Затем отработанный воздух откачивается с растворителем. Рабочие условия распылительной сушки, такие как температура на входе и выходе, скорость подачи, давление распыления, скорость потока осушающего воздуха и конфигурация сопла, можно регулировать для получения требуемого размера частиц, содержания влаги и выхода образующихся в результате сухих частиц. Выбор подходящих устройств и условий обработки относится к компетенции квалифицированного специалиста с учетом изложенных здесь принципов и может быть осуществлен без чрезмерного экспериментирования. В качестве примера можно использовать следующие параметры масштабной сушки NIRO® PSD-1®: температура воздуха на входе примерно от 80°C до 200°C, например, от 110°C до 170°C; температура воздуха на выходе примерно от 40°С до 120°С, например, примерно от 60°С до 100°С; скорость подачи жидкости примерно от 30 г/мин до 120 г/мин, например, примерно от 50 г/мин до 100 г/мин; общий поток воздуха примерно от 140 стандартных кубических футов в минуту (scfm) до 230 scfm, например, примерно от 160 scfm до 210 scfm; и скорость потока распыляемого воздуха примерно от 30 scfm до 90 scfm, например, примерно от 40 scfm до 80 scfm. Содержание твердых веществ в распыляемом исходном сырье обычно находится в диапазоне от 0,5% масс./об. (5 мг/мл) до 10% масс./об. (100 мг/мл), например от 1,0% масс./об. до 5,0% масс./об. Разумеется, параметры зависят от масштаба и типа используемого оборудования, а также от типа используемой системы растворителей. В любом случае использование этих и подобных методов позволяет получать частицы с диаметрами, обеспечивающими осаждение аэрозоля в легкие.
[00285] В некоторых вариантах осуществления все вспомогательные вещества растворяют в исходном сырье, а покрытия ядра-оболочки на диспергированном активном ингредиенте образуются за счет различий в физических свойствах растворенных веществ.
[00286] Как указано ранее для частиц, содержащих аморфный активный ингредиент, природу поверхности частиц и их морфологию можно контролировать путем регулирования растворимости и диффузии компонентов в исходном сырье. Поверхностно-активные гидрофобные вспомогательные вещества (например, трилейцин, фосфолипиды, жирнокислотные мыла) могут концентрироваться на границе раздела, улучшая флюидизацию и диспергируемость порошка, а также повышая шероховатость поверхности частиц.
[00287] Любую стадию распылительной сушки и/или все стадии распылительной сушки можно проводить с использованием оборудования, традиционно используемого для получения высушенных распылением частиц, предназначенных для применения в фармацевтических препаратах, вводимых путем ингаляции. Коммерчески доступные распылительные сушилки включают в себя сушилки, производимые Büchi Ltd. and Niro Corp.
[00288] В некоторых вариантах осуществления исходное сырье распыляют с помощью двойного сопла для жидкостей. Значительное расширение распределения частиц по размерам капель жидкости происходит при загрузке твердых веществ, превышающей примерно 1,5% масс./масс. Капли большего размера в хвосте распределения приводят к увеличению частиц в соответствующем распределении порошка. В результате в некоторых вариантах осуществления с использованием двойных жидкостных сопел ограничивают загрузку твердых веществ до 1,5% масс./масс., или, например, менее 1,0 масс.%, или 0,75 масс.%.
[00289] В некоторых вариантах осуществления узкого распределения капель по размерам при более высоком содержании твердых веществ можно достичь с помощью плоских пленочных распылителей, как описано, например, в патентах США №№ 7967221 и 8616464,. В некоторых вариантах осуществления исходное сырье распыляют при загрузке твердых веществ от 2% до 10% масс./масс., например, от 3% до 5% масс./масс.
[00290] В некоторых вариантах осуществления плотность популяции частиц или PPD составляет от 0,01×10-6 до 1,0×10-6, например, от 0,03×10-6 до 0,2×10-6.
[00291] В некоторых вариантах осуществления соотношение EtOH/твердые вещества составляет от 1,0 до 20,0, например от 3,0 до 10,0.
[00292] В некоторых вариантах осуществления настоящая заявка раскрывает фармацевтический порошкообразный состав для ингаляции, содержащий частицы, полученные по способу, включающему:
a. получение раствора описанных здесь TSLP-связывающих молекул в смеси вода/этанол, где содержание этанола варьирует от 1 до 20%, а отношение этанола к общему количеству твердых веществ составляет от 1 до 20;
b. распылительную сушку раствора с получением частиц, характеризующих плотностью 0,2 г/см3 или ниже, геометрическим диаметром 1-3 мкм и аэродинамическим диаметром от 1 до 2 мкм;
причем порошок при введении путем ингаляции обеспечивает общую легочную дозу in vitro более чем примерно 80%. В некоторых вариантах осуществления фармацевтический порошкообразный состав дополнительно содержит стеклообразующее вспомогательное вещество. В некоторых вариантах осуществления стеклообразующее вспомогательное вещество включает в себя альфа. В других вариантах осуществления стеклообразующее вспомогательное вещество включает в себя бета. В следующем варианте осуществления стеклообразующее вспомогательное вещество включает в себя трегалозу.
[00293] В некоторых вариантах осуществления фармацевтического порошкообразного состава плотность популяции частиц составляет от 0,01×10-6 до 1,0×10-6.
[00294] Настоящая заявка также раскрывает способ доставки в легкие индивидуума частиц сухого порошка, где способ включает в себя:
a. получение раствора описанных здесь TSLP-связывающих молекул в смеси вода/этанол, где содержание этанола варьирует от 5 до 20%,
b. распылительную сушку раствора с получением частиц, характеризующих плотностью примерно от 0,05 до 0,3 г/см3, геометрическим диаметром 1-3 мкм и аэродинамическим диаметром 1-2 мкм;
c. упаковка высушенного распылением порошка в сосуд;
d. применение ингалятора, содержащего средство для извлечения порошка в сосуд и средство для флюидизации и аэрозолизации порошка, где ингалятор работает при усилии вдоха, совершаемого пациентом, примерно от 2 до 6 кПа; ингалятор и порошок вместе обеспечивают инерционный параметр примерно от 120 до 400 мкм2 л/мин, и где при введении порошка путем ингаляции его отложение в легких составляет по меньшей мере 90%.
[00295] Настоящая заявка также раскрывает способ получения сухого порошкообразного лекарственного состава для легочной доставки, где способ включает
a. получение раствора описанных здесь TSLP-связывающих молекул в смеси вода/этанол, где содержание этанола варьирует от 5 до 20%,
b. распылительную сушку раствора с получением частиц, характеризующих плотностью примерно от 0,05 до 0,3 г/см3, геометрическим диаметром 1-3 мкм и аэродинамическим диаметром 1-2 мкм.
[00296] В следующем варианте осуществления настоящая заявка раскрывает фармацевтический порошкообразный состав, доставляемый из ингалятора сухого порошка, частицы которой содержат описанные здесь TSLP-связывающие молекулы, где общая легочная доза in vitro превышает 90% масс./масс. от доставляемой дозы и где композиция имеет по меньшей мере одну характеристику из следующих: отсутствие носителей, плотность частиц от 0,05 до 0,3 г/см3; шероховатость частиц от 3 до 20; содержит частицы, полученные по способу, включающему в себя распылительную сушку из смеси этанол-вода; и содержит частицы, полученные по способу, включающему в себя распылительную сушку из смеси этанол-вода при отношении этанол:твердые вещества от 1 до 20. В некоторых вариантах осуществления фармацевтический порошкообразный состав имеет, по меньшей мере, две из указанных характеристик; в других вариантах осуществления фармацевтический порошкообразный состав имеет, по меньшей мере, три из указанных характеристик.
Дозирование
[00297] Дозировочный режим, токсичность и терапевтическую эффективность описанных здесь молекул против TSLP, включая фармацевтические композиции, содержащие антитела против TSLP или их фрагменты, можно определить с помощью стандартных фармацевтических процедур на культурах клеток или экспериментальных животных, например, путем определения LD50 (летальная доза для 50% популяции) и ED50 (доза, терапевтически эффективная для 50% популяции). Соотношение доз, вызывающих токсические и терапевтические эффекты, называемое терапевтическим индексом, можно выразить как отношение LD50/ED50. Предпочтительными являются соединения с высоким терапевтическим индексом. Соединения, оказывающие токсические побочные эффекты, можно использовать, но следует позаботиться о разработке системы доставки таких соединений непосредственно к участку пораженной ткани, чтобы свести к минимуму потенциальное повреждение неинфицированных клеток и, следовательно, уменьшить побочные эффекты.
[00298] Данные, полученные путем анализов клеточных культур и исследований на животных, можно использовать при определении диапазона доз, подходящего для людей. Дозировка таких соединений предпочтительно находится в диапазоне концентраций в кровотоке, включающем ED50 при небольшой или отсутствующей токсичности. Дозировка может варьировать в пределах указанного диапазона в зависимости от используемой лекарственной формы и используемого способа введения. Терапевтически эффективную дозу любого соединения, используемого в способе настоящего изобретения, можно первоначально определить с помощью анализов клеточных культур. С помощью животных моделей можно определить дозу, позволяющую достичь диапазона концентраций в плазме, который включает IC50 (то есть концентрацию тестируемого соединения, которая обеспечивает полумаксимальное ингибирование симптомов), определенную на культуре клеток. С помощью такой информации можно более точно определить дозы, подходящие для людей. Уровни в плазме можно измерить, например, методом высокоэффективной жидкостной хроматографии.
Наборы
[00299] Изобретение также относится к наборам, содержащим одну или несколько из описанных здесь фармацевтических композиций, устройство для доставки фармацевтической композиции индивидууму и инструкции по применению. В некоторых вариантах осуществления устройство может доставлять фармацевтическую композицию в аэрозольной форме. В некоторых вариантах осуществления устройство представляет собой ингалятор, например, ингалятор сухого порошка (DPI). В других вариантах осуществления устройство может представлять собой дозирующий ингалятор или распылитель.
[00300] Подходящие ингаляторы сухого порошка включают однодозовые ингаляторы, где сухой порошок хранится в капсуле или блистере, и пациент перед применением загружает одну или несколько капсул или блистеров в устройство. Альтернативно используют многодозовые ингаляторы сухого порошка, где доза предварительно упакована в блистеры из фольги, такие как картриджи, полоски или диски.
[00301] Ингаляторы сухого порошка включают многодозовые ингаляторы сухого порошка, такие как DISKUSTM (GSK, описанный в патенте США 6536427), DISKHALERTM (GSK, описанный в публикации патентной заявки WO 97/25086), GEMINITM (GSK, описанный в публикации патентной заявки WO 05/14089), GYROHALERTM (Vectura, описанный в публикации патентной заявки WO 05/37353) и PROHALERTM (Valois, описанный в публикации патентной заявки WO 03/77979).
[00302] Однодозовые ингаляторы сухого порошка включают AEROLIZERTM (Novartis, описанный в US 3991761) и BREEZHALERTM (Novartis, описанный в патенте США № 8479730 (Ziegler et al.)). Другие подходящие однодозовые ингаляторы описаны в патентах США №№ 8069851 и 7559325.
[00303] Однодозовые блистерные ингаляторы, которые некоторые пациенты находят более простыми и удобными для доставки лекарственных средств, требующих введения один раз в день, включают ингалятор, описанный в патенте США № 8573197 (Axford et al.).
[00304] В некоторых вариантах осуществления ингаляторы представляют собой многодозовые ингаляторы сухого порошка, в которых энергия для флюидизации и диспергирования порошка поставляется пациентом (то есть "пассивные" MD-DPI). Порошки настоящего изобретения эффективно флюидизируются и диспергируются при низкой пиковой скорости вдоха (PIF). В результате небольшие изменения в дисперсии порошка, наблюдаемые при PIF, эффективно уравновешивают увеличение инерционного воздействия, которое происходит при увеличении PIF, что приводит к независимому от скорости потока отложению в легких. Отсутствие зависимости от скорости потока, наблюдаемое для порошков настоящего изобретения, приводит к уменьшению общей вариабельности среди пациентов.
[00305] Инструкции по применению могут включать в себя инструкции по диагностике или лечению связанных с TSLP воспалительных состояний. Предлагаемые наборы можно использовать в соответствии с любым из описанных здесь способов. Для специалистов в данной области будут очевидны другие подходящие способы применения предлагаемых здесь наборов и они смогут использовать наборы в таких способах. Предлагаемые здесь наборы также могут содержать конверт (например, почтовый платный конверт или почтовый пакет), который можно использовать для возврата образца для анализа, например, в лабораторию. Набор может содержать один или несколько контейнеров для образца, или образец может находиться в стандартном флаконе для сбора крови. Набор может также содержать одну или несколько форм информированного согласия, форму заявки на тестирование и инструкции по применению набора в описанном здесь способе. Способы применения таких наборов также входят в объем настоящего изобретения. Одна или несколько форм (например, форма заявки на тестирование) и контейнер, содержащий образец, могут быть закодированы, например штрих-кодом, для идентификации индивидуума, предоставившего образец.
Способы лечения
[00306] Изобретение предлагает способы лечения связанного с TSLP состояния у индивидуума, нуждающегося в лечении такого состояния, например человека, путем введения индивидууму терапевтически эффективного количества одной из описанных здесь TSLP-связывающих молекул или ее фармацевтической композиции. В некоторых вариантах осуществления такие способы дополнительно включают в себя идентификацию и выбор индивидуума, нуждающегося в лечении воспалительного состояния, связанного с TSLP. Изобретение также предусматривает применение описанных здесь TSLP-связывающих молекул, или их фармацевтических композиций для лечения или профилактики заболевания у пациента. В некоторых вариантах осуществления изобретение предлагает описанные здесь TSLP-связывающие молекулы, или их фармацевтические композиции, для применения в способах лечения или профилактики заболевания у пациента. В других вариантах осуществления изобретение предлагает применение описанных здесь TSLP-связывающих молекул, или их фармацевтических композиций, в способе промышленного получения лекарственного средства, предназначенного для лечения или профилактики заболевания у пациента.
[00307] В некоторых вариантах осуществления связанные с TSLP воспалительные состояния могут вызываться аллергическими реакциями или раздражающими веществами или стимулами окружающей среды. В некоторых конкретных вариантах осуществления связанные с TSLP воспалительные состояния включают астму, хроническую обструктивную болезнь легких, аллергический ринит, аллергический риносинусит, аллергический конъюнктивит, атопический дерматит, эозинофильный эзофагит.
[00308] В некоторых вариантах осуществления TSLP-связывающие молекулы, или фармацевтические композиции, содержащие TSLP-связывающие молекулы, вводят индивидууму путем ингаляции, например, в аэрозольной форме с помощью ингалятора сухого порошка. В других вариантах осуществления TSLP-связывающие молекулы или их фармацевтические композиции можно вводить с использованием одного или нескольких из множества способов, известных в данной области. Специалистам в данной области известно, что способ и/или режим введения могут варьировать в зависимости от желаемых результатов. Подходящие способы введения включают в себя внутривенный, внутримышечный, внутрикожный, внутрибрюшинный, подкожный, спинальный или другой парентеральный способ введения, например, путем инъекции или инфузии. К парентеральному введению относятся способы введения, отличные от энтерального и местного введения, обычно проводимые путем инъекции и включающие в себя, без ограничения, внутривенные, внутримышечные, внутриартериальные, интратекальные, интракапсулярные, внутриглазничные, внутрисердечные, внутрикожные, внутрибрюшинные, транстрахеальные, подкожные, субкутикулярные, внутрисуставные, субкапсулярные, субарахноидальные, интраспинальные, эпидуральные и внутригрудинные инъекции и инфузии. Альтернативно TSLP-связывающие молекулы или фармацевтические композиции, содержащие TSLP-связывающие молекулы настоящего изобретения, можно вводить с помощью непарентерального способа, такого как местный, эпидермальный или слизистый путь введения, например, интраназально, перорально, вагинально, ректально, сублингвально или местно.
[00309] В некоторых вариантах осуществления воспалительным состоянием, связанным с TSLP, является астма. Астма представляет собой сложное и гетерогенное хроническое воспалительное заболевание дыхательных путей, которое характеризуется обратимой бронхоконстрикцией и связано с чрезмерным ответом дыхательных путей на широкий спектр бронхоконстрикторных стимулов (гиперреактивность дыхательных путей, AHR). Проведенная в последнее время работа, направленная на идентификацию иммунных путей, участвующих в патогенезе астмы, позволила выявить роли как Т-хелперов типа 2 (Th2), так и эффекторных клеток, не опосредуемых Th2 (Lambrecht and Hammad, Nature immunology 2014, 16: 45-56). В случае аллергической астмы, характеризующейся эозинофильным воспалением и наличием атопии, элементы иммунного пути Th2 имеют решающее значение для развития и поддержания воспаления дыхательных путей и AHR. Тимусный стромальный лимфопоэтин (TSLP) является ключевым вышестоящим регулятором пути Th2. TSLP экспрессируется в эпителиальных клетках слизистой оболочки дыхательных путей в ответ на разные стимулы (например, физическое повреждение, частицы окружающей среды, аллергены, провоспалительные или Th2-поляризующие цитокины и продукты микробов). Роль TSLP заключается в модуляции дендритных клеток (DC), индуцировании дифференцировки наивных Т-клеток в воспалительные Th2-клетки и стимулировании секреции цитокинов из тучных клеток, эозинофилов и макрофагов как части врожденного иммунного ответа. Кроме того, TSLP может препятствовать развитию регуляторных Т-клеток, ухудшая баланс между толерантностью и воспалением. В случае неаллергической астмы, характеризующейся нейтрофильным или pauci-гранулоцитарным воспалением, цитокины, участвующие в развитии воспаления, не до конца изучены, однако, как полагают, важную роль играют не опосредуемые Th2 цитокины IL-17 и интерферон-γ (IFN-γ). Интересно, что результаты доклинических исследований свидетельствуют о том, что помимо опосредования ответа Th2 TSLP усиливает не связанные с Th2 ответы и может также иметь важное значение для развития опосредованного IL-17 и IFN-γ хронического воспаления.
[00310] TSLP является необходимым и достаточным для развития связанного с цитокином Th2 воспаления дыхательных путей у грызунов. У трансгенных мышей с конститутивной секрецией TSLP в эпителии легких под контролем промотора поверхностно-активного белка С развиваются следующие признаки, совместимые с астмой: эозинофильное воспаление дыхательных путей; экспрессия Th2-смещенной инфильтрации Т-клеток CD4; системная эозинофилия; повышенный уровень IgE; гиперчувствительность дыхательных путей; а также значительное реструктурирование дыхательных путей, включая гиперплазию бокаловидных клеток и фиброз дыхательных путей и сосудов. Роль TSLP в развитии аллергического воспаления дополнительно подтверждают данные, свидетельствующие о том, что экспрессии и продукция белка TSLP увеличивается при вдыхании аллергена в легких (Zhou et al, 2005, Nature immunology 6, 1047- 1053), а прямая интраназальная доставка TSLP в присутствии антигена приводит к быстрому возникновению тяжелого заболевания (Headley et al., 2009, Journal of immunology 182, 1641-1647). TSLPR-дефицитные мыши устойчивы к развитию Th2-подобного воспаления у классической мышиной модели праймированной овальбумином и квасцами (Al-Shami et al., 2005, Journal of Experimental Medicine 202, 829-839; Zhou et al., 2005, Nature immunology 6, 1047-1053). Уменьшение воспаления дыхательных путей коррелирует с уменьшением IgE в сыворотке и уменьшением цитокинов и хемокинов Th2, таких как IL-4, -5, -13, эотаксин, а также хемокин, регулируемый тимусом и активацией (TARC).
[00311] Повышенная экспрессия TSLP в собственной пластинке слизистой оболочки дыхательных путей наблюдается, главным образом, у пациентов с тяжелой формой астмы (Shikotra et al., 2012, Journal of Allergy and Clinical Immunology 129, 104-11l.el09). Кроме того, в нескольких исследованиях показана взаимосвязь между частотой однонуклеотидного полиморфизма (SNP) в локусе человеческого TSLP и уровнем экспрессии TSLP и склонностью к астме и эозинофильному эзофагиту (Ferreira et al., 2014, The Journal of allergy and clinical immunology 133, 1564-1571; Harada et al., 2011, American journal of respiratory cell and molecular biology 44, 787-793; He et al., 2009, The Journal of allergy and clinical immunology 124, 222-229; Rothenberg et al., 2010, Nature Genetics 42, 289-291). В проведенном недавно исследовании также было показано, что варианты гена TSLP связаны со значительным увеличением риска астмы при детской астме ерез эпистатические ассоциации (Biagini Myers et al., 2014, The Journal of allergy and Clin immunismology 134, 891-899 e893).
Сочетанные способы лечения
[00312] Разные описанные выше способы лечения можно объединить с другими партнерами по способу лечения, такими как существующее в настоящее время стандартное средство лечения связанных с TSLP воспалительных состояний. Соответственно, описанные здесь способы лечения связанного с TSLP воспалительного состояния могут дополнительно включать в себя введение второго средства индивидууму, нуждающемуся в лечении. В некоторых вариантах осуществления второе средство может быть выбрано, без ограничения, из кортикостероидов, бронходилататоров (SABA, LABA, SAMA, LAMA), антигистаминных средств, антилейкотриенов и ингибиторов PDE-4.
[00313] Термин "сочетание" относится либо к фиксированному сочетанию в одной лекарственной форме, либо к сочетанному введению, где соединение настоящего изобретения и партнер по сочетанию (например, другое лекарственное средство, описанное ниже, также называемое "терапевтическое средство" или "со-средство") можно вводить независимо в одно и то же время, или отдельно с интервалами времени, особенно если эти интервалы времени позволяют партнерам по сочетанию проявлять совместный, например, синергетический эффект. Разные компоненты могут быть упакованы в наборе или отдельно. Один компонент или оба компонента (например, порошки или жидкости) можно перерастворить или разбавить до желательной дозы перед введением. Используемые здесь термины "совместное введение", "сочетанное введение" и т.п. относятся к введению выбранного партнера по сочетанию одному индивидууму, нуждающемуся в этом (например, пациенту), и охватывают схемы лечения, в которых средства не обязательно вводят одним и тем же способом или в одно и то же время. Используемый здесь термин "фармацевтическое сочетание" относится к продукту, который получают в результате смешивания или объединения нескольких терапевтических средств, и включает в себя как фиксированные, так и нефиксированные сочетания терапевтических средств. Термин "фиксированное сочетание" означает, что терапевтические средства, например, соединение настоящего изобретения и партнер по сочетанию вводят пациенту одновременно в виде одного объекта или одной дозы. Термин "нефиксированное сочетание" означает, что терапевтические средства, например соединение настоящего изобретения и партнер по сочетанию вводят пациенту в виде отдельных объектов одновременно, параллельно или последовательно без каких-либо конкретных временных ограничений, причем такое введение обеспечивает терапевтически эффективные уровни двух соединений в организме пациента. Последний термин также относится к коктейльной терапии, например, к введению трех или более терапевтических средств. Используемый здесь термин "фармацевтическое сочетание" относится либо к фиксированному сочетанию в одной лекарственной форме, либо к нефиксированному сочетанию, либо к набору частей для сочетанного введения, где два или более терапевтических средства можно вводить независимо в одно и то же время или отдельно с интервалами времени, особенно в тех случаях, когда эти интервалы времени позволяют партнерам по сочетанию проявлять совместный, например, синергетический эффект.
[00314] Термин "сочетанная терапия" относится к введению двух или более терапевтических средств для лечения терапевтического состояния или расстройства, описанного в настоящем документе. Такое введение включает в себя совместное введение указанных терапевтических средств практически одновременно, например, в одной капсуле, имеющей фиксированное отношение активных ингредиентов. Альтернативно такое введение включает в себя совместное введение активных ингредиентов в нескольких или в отдельных контейнерах (например, в виде таблеток, капсул, порошков и жидкостей). Порошки и/или жидкости перед введением можно перерастворить или развести с достижением желательной дозы. Кроме того, такое введение также охватывает последовательное применение терапевтических средств либо примерно в одно и то же время, либо в разное время. В любом случае схема лечения должна обеспечивать благоприятные эффекты сочетания лекарственных средств при лечении описанных здесь состояний или расстройств.
[00315] Если не указано иное, все используемые здесь технические и научные термины имеют традиционные значения, известные рядовым специалистом в области техники, к которой относится настоящее изобретение. Ниже описаны подходящие способы и материалы, однако для осуществления настоящего изобретения можно использовать способы и материалы, аналогичные или эквивалентные описанным здесь. Все упомянутые здесь публикации, патентные заявки, патенты и другие ссылки включены в качестве ссылки в полном объеме. В случае конфликта настоящее описание, включая определения, будет иметь определяющее значение. Кроме того, материалы, способы и примеры являются только иллюстративными и не предназначены для ограничения. Специалистам в данной области техники известны способы и материалы, аналогичные или эквивалентные описанным здесь, которые можно использовать для осуществления настоящего изобретения. В действительности настоящее изобретение никоим образом не ограничивается описанными способами и материалами.
ПРИМЕРЫ
Пример 1: Получение антител против человеческого TSLP и их Fab-фрагментов методом фагового дисплея
[00316] Fab, которые специфически связываются с изоформой 1 человеческого TSLP (SEQ ID NO: 27), получают с использованием технологии фагового дисплея MorphoSys HuCAL PLATINUM®. Библиотеку фагмидов получают на основе концепции HuCAL® (Knappik et al, 2000, J Mol Biol 296, 57-86) с использованием технологии CysDisplayTM для отображения Fab на поверхности фага (Lohning, WO 2001/05950).
Пэннинг
[00317] Проводят три типа пэннинга: твердофазный пэннинг против непосредственного покрытия рекомбинантным человеческим TSLP (rhTSLP), твердофазный пэннинг с улавливанием амилоидным белком-предшественником (APP) и пэннинг против TSLP в растворе.
[00318] Для твердофазного пэннинга против rhTSLP с непосредственным покрытием 96-луночные планшеты Maxisorp™ покрывают 300 мкл полученного из E.coli rhTSLP (R&D Systems) на лунку при 4°C в течение ночи. Для каждого пэннинга к каждой покрытой антигеном лунке добавляют фаговые антитела HuCAL PLATINUM® и инкубируют в течение 2 часов при комнатной температуре на шейкере для микротитрационных планшетов. Затем неспецифически связанные фаги отмывают с использованием нескольких стадий промывки, а специфически связанные фаги элюируют, используя 25 мМ DTT в 10 мМ Tris/HCl, рН 8. Элюат DTT переносят в 14 мл TGI E. coli и инкубируют, чтобы обеспечить фаговую инфекцию.
[00319] Инфицированные бактерии ресуспендируют в среде 2×YT, высевают на чашки с агаром LB/Cam и инкубируют в течение ночи. Колонии соскребают с чашек и используют для спасения фагов, поликлональной амплификации выбранных клонов и продуцирования фагов. С использованием очищенного фага проводят следующий цикл пэннинга. Второй и третий циклы твердофазного пэннинга проводят в соответствии с протоколом первого цикла, за исключением уменьшенных количеств антигена и более жестких условий промывания.
[00320] При твердофазном пэннинге в режиме захвата APP против TSLP яванского макака, антигены, используемые при пэннинге, содержат метку APP6 (амилоидный белок-предшественник), а гибридные белки антиген-APP6 улавливаются мышиными антителами против APP6, иммобилизованными на планшете Maxisorp™. Чтобы предотвратить выбор фага, связывающегося с APP6-меткой антигена, или с улавливающим антителом против APP6, проводят предварительную блокировку фага с использованием улавливающего антитела и нерелевантного APP6-меченого антигена.
[00321] 96-луночные планшеты Maxisorp™ покрывают 300 мкл антител против APP и нерелевантным APP6-меченым антигеном в течение ночи при 4°C. Используемые в качестве антигенов человеческий TSLP_Avi-APP6 или обезьяний TSLP_APP6-Avi иммобилизуют в течение 1 часа при комнатной температуре на шейкере. Параллельно фаги предварительно адсорбируют дважды на антителах против APP и нерелевантном антигене.
[00322] Помимо процедур покрытия антигеном и блокирования фагов, проводят пэннинг с улавливанием подобно описанному выше твердофазному пэннингу.
[00323] Для проведения пэннинга против TSLP в растворе фаги блокируют смесью, содержащей 50% сыворотки человека/0,33× Chemiblocker/0,05% твин 20. На фаговый пул 4 мг стрептавидиновых шариков (Dynabeads® M-280 Streptavidin, Invitrogen) блокируют в l× Chemiblocker. Для удаления фагов, связывающихся со стрептавидином или с фаговыми шариками, предварительную адсорбцию блокированных фаговых частиц проводят дважды, используя в каждом случае блокированные стрептавидиновые шарики. Затем к фаговым частицам добавляют биотинилированный антиген человеческий TSLP_Avi-APP6. После инкубации комплексы фаг-антиген захватывают с использованием стрептавидиновых шариков, и фаговые частицы, связанные со стрептавидиновыми шариками, собирают с использованием магнитного сепаратора. Неспецифически связанные фаги отмывают, используя несколько стадий промывки, с помощью PBS/0,05% твин 20 и PBS. Специфически связанные фаги элюируют из стрептавидиновых шариков с использованием 25 мМ DTT в 10 мМ Tris/HCl, рН 8. Последующая фаговая инфекция и продуцирование фагов выполняют в соответствии с протоколом твердофазного пэннинга, и начинают следующий цикл пэннинга.
Экспрессия
[00324] Чтобы облегчить быструю экспрессию растворимого Fab, Fab-кодирующие вставки выбранного фага HuCAL PLATINUM® субклонируют из вектора дисплея pMORPH®30 в векторе экспрессии pMORPH®x11_FH. После трансформации E.coli, экспрессию одного клона TG1-F и получение периплазматических экстрактов, содержащих фрагменты HuCAL®-Fab, выполняют, как описано ранее (Rauchenberger et al., 2003, J Biol Chem 278: 38194-38205).
[00325] Резистентные к хлорамфениколу одиночные клоны собирают в лунки стерильного 384-луночного микротитрационного планшета, предварительно заполненные средой 2×YT, и выращивают при температуре 37°С. На следующее утро в каждую лунку мастер-планшетов добавляют среду, содержащую глицерин; планшеты герметизируют алюминиевой фольгой и хранят при -80°C.
Скрининг методом ELISA
[00326] Используя скрининг ELISA, одиночные Fab-клоны идентифицируют по результату пэннинга для связывания с целевым антигеном. Fab тестируют с использованием Fab-содержащих неочищенных лизатов E. coli. Для подтверждения экспрессии Fab в подготовленных лизатах E. coli планшеты Maxisorp™ 384 покрывают Fd-фрагментом специфических овечьих антител против человеческого IgG, разведенным 1:1000 в PBS. После блокирования 5% обезжиренным молочным порошком в PBS, содержащем 0,05% твин 20, добавляют Fab, содержащие лизаты E.coli. Впоследствии связанные фрагменты HuCAL®-Fab обнаруживают путем инкубации с F(ab)2 специфических козьих антител против человеческого IgG, конъюгированного с щелочной фосфатазой (разведенной 1:5000) с последующим добавлением флуоресцентного субстрата AttoPhos (Roche, № 11681982001). Эмиссию флуоресценции при 535 нм регистрируют с использованием возбуждения при 430 нм.
[00327] Для проведения скрининга ELISA на антигене с прямым покрытием, 384-луночные планшеты Maxisorp™ покрывают разными TSLP-антигенами при концентрации 2 мкг/мл в PBS. После блокировки планшетов 5% обезжиренным молочным порошком в PBS добавляют Fab-содержащие лизаты E. coli. Связывание Fab обнаруживают посредством F(ab)2-специфических козьих антител против человеческого IgG, конъюгированных с щелочной фосфатазой (разведенной 1:5000) с использованием флуоресцентного субстрата Attophos (Roche, # 11681982001). Эмиссию флуоресценции при 535 нм регистрируют с использованием возбуждения при 430 нм.
[00328] Для проведения скрининга ELISA на APP-уловленном антигене 384-луночные планшеты Maxisorp™ покрывают специфическим антителом против APP при концентрации 2,5 мкг/мл в PBS. После блокировки планшетов 5% обезжиренным молочным порошком в PBS обеспечивают связывание АРР-меченых антигенов TSLP при концентрации 2 мкг/мл в течение 1 часа при комнатной температуре. Затем добавляют Fab-содержащие лизаты E.coli. Связывание Fab обнаруживают посредством F(ab)2-специфических козьих антител против человеческого IgG, конъюгированных с щелочной фосфатазой (разведенной 1:5000) с использованием флуоресцентного субстрата Attophos (Roche, #11681982001). Эмиссию флуоресценции при 535 нм регистрируют с использованием возбуждения при 430 нм.
[00329] Для проведения скрининга ELISA биотинилированного антигена, 384-луночные планшеты Maxisorp™ покрывают специфическими к фрагменту Fd овечьими антителами против человеческого IgG (сайт связывания #PC075), разбавленными 1:1000 в PBS или специфическими мышиными IgG против His (R&D Systems, #MAB050) соответственно. После блокирования 5% обезжиренным молочным порошком в PBS добавляют Fab-содержащие лизаты E. coli. Впоследствии обеспечивают связывание захваченных фрагментов HuCAL®-Fab с биотинилированным человеческим TSLP 0,7-1,5 мкг/мл, человеческим TSLP или обезьяньим TSLP соответственно, которые обнаруживают путем инкубации со стрептавидином, конъюгированным с щелочной фосфатазой, с последующим добавлением флуоресцентного субстрата AttoPhos (Roche, #11681982001). Эмиссию флуоресценции при 535 нм регистрируют с использованием возбуждения при 430 нм.
[00330] Биотинилированные антигены (2,5-5 мкг/мл) также подвергают улавливанию на планшетах, покрытых нейтравидином. После блокирования 5% обезжиренным молочным порошком в PBS добавляют Fab-содержащие лизаты E. coli. Связывание Fab обнаруживают с помощью F(ab)2-специфических козьих антител против человеческого IgG, конъюгированных с щелочной фосфатазой (разведенной 1:5000) с использованием флуоресцентного субстрата AttoPhos (Roche, #11681982001). Эмиссию флуоресценции при 535 нм регистрируют с использованием возбуждения при 430 нм.
[00331] 9984 клона (384 клона/субкод пэннинга) анализируют в первичном скрининге ELISA на биотинилированном человеческом TSLP_Avi-APP6 и биотинилированном обезьяньем TSLP_APP6-Avi, нанесенными в виде покрытия на NA-планшеты (см. 3.4.4). Результаты ELISA анализируют с помощью программы GENios Pro ʺPrimeScreenʺ. Результаты анализируют по сравнению с фоновым сигналом. Для человеческого антигена выбирают только лунки с сигналами, 10-кратно превышающими фон, а для лунок с обезьяньим антигеном с сигналами, 5-кратно превышающими фон. Сигналы ниже 5-кратного фона, вероятно, будут являться результатом низкой экспрессии Fab, Fab с низким сродством, краевыми эффектами микротитрационного планшета или невоспроизводимых значений. Пэннинг в растворе приводит к 3133 лучшим результатам, твердофазный пэннинг приводит к 240 лучшим результатам. 1472 выбранных первичных лучших результатов дополнительно анализируют во вторичном скрининге ELISA.
[00332] Во вторичном скрининге ELISA используют разные режимы представления антигена, в том числе C- или N-концевые антигены, меченные Avi-APP6, антигены с прямым покрытием, биотинилированные антигены, представленные в растворе, антигены HEK, антигены E.coli, дегликозилированные варианты антигенов (обработанные PNGазой). Кроме того, при вторичном скрининге анализируют неспецифическое связывание с противоцелевым IL-7. Чтобы исключить связывание биотина и метки, используют биотинилированный нерелевантный антиген, меченный APP-Avi. Результаты ELISA анализируют с помощью программы GENios Pro ʺPrimeScreenʺ, и результаты анализируют в сравнении с фоновым сигналом. Для нерелевантного антигена и противоцелевого IL-7 выбирают только лучшие результаты с сигналами, 2-кратно превышающими фон.
[00333] Результаты вторичного скрининга показывают, что способ представления антигена имеет решающее значение для перекрестной реактивности. Скрининг на дегликозилированном антигене показывает, что могут быть связующие, нацеленные на глико-эпитоп. Кроме того, расположение меток и состав меток могут влиять на перекрестную активность вследствие конформационных изменений. Клоны группируют в соответствии с их профилями перекрестной реактивности, что приводит к семи разным группам перекрестной реактивности. Группы 1-3 включают все клоны, которые являются перекрестно-реактивными по отношению к huTSLP из E.coli, либо отдельно, либо в сочетании с антигенами, полученными из HEK. Группа 4 включает в себя все клоны, которые по меньшей мере перекрестно реагируют с TSLP_Avi-APP6 человека, представленным в растворе. Напротив, группа 5 включает все клоны, которые исключительно перекрестно реагируют с TSLP_Avi-APP6 человека в растворе. В группе 6 есть все клоны, которые являются перекрестно-реактивными для TSLP_Avi-APP6 человека и для дегликозилированного TSLP_Avi-APP6 человека, а в группе 7 имеются все клоны, которые являются перекрестно-реактивными для всех антигенов, полученных из НЕК, включая дегликозилированные антигены.
Секвенирование и превращение в IgG
[00334] Анализ последовательности проводят на 73 клонах из групп 1-3 перекрестной реактивности (клоны, перекрестно реагирующие с TSLP, полученным из E.coli), и 569 клонах из групп 4-7 (клоны, способные перекрестно взаимодействовать с HEG-продуцированными антигенами). Всего идентифицировано 297 уникальных клонов HCDR3, 222 клона объединены и 124 клона очищены в формате Fab.
[00335] Клоны, полученные из третьего и четвертого анализа последовательности, сразу вводят в конверсию IgG и последовательно клонируют в векторе pMORPH®4_IgGlf для экспрессии в клетках млекопитающих.
Определение сродства
[00336] Определение константы диссоциации (KD) версии клонов HuCAL® Fab и IgG выполняют следующим образом: биотинилированный человеческий TSLP покрывают 0,2 мкг/мл в аналитическом буфере в течение 1 часа при комнатной температуре на планшетах MSD со стрептавидином. Планшеты со стрептавидином блокируют в течение ночи при 4°C PBS с 3% BSA перед покрытием антигеном. Равновесное титрование в растворе (SET) проводят с человеческим TSLP и обезьяньим TSLP в условиях, описанных ниже. Используют мономерные фракции белка антител (по меньшей мере с 90% содержанием мономеров, анализируемое аналитическим SEC, Superdex75 (Amersham Pharmacia) для Fab или Tosoh G3000SWXL (Tosoh Bioscience) для IgG, соответственно).
[00337] Определение аффинности в растворе в основном выполняют, как описано в литературе (Friquet et al, 1985, J Immnunol Meth 77, 305-319). Чтобы улучшить чувствительность и точность метода SET, его перенесят с классического ELISA на технологию, основанную на ECL (Haenel et al, 2005, Anal Biochem 339, 182-184). 1 мг/мл специфических козьих антител против человеческого фрагмента (Fab)2 (Dianova) метят MSD Sulfo-TAGTM NHS-Сложный эфир (Meso Scale Discovery, Gaithersburg, MD, USA) в соответствии с инструкциями производителя.
[00338] Эксперименты проводят в полипропиленовых микротитрационных планшетах, и PBS pH 7,4, содержащий 0,5% BSA и 0,02% твин-20 в качестве буфера используют для анализа. Немеченый антиген подвергают серийным разведениям 2n, начиная с концентрации, по меньшей мере, в 10 раз превышающей ожидаемую KD. Для определения значений Bмакс используют лунки без антигена; лунки, содержащие только буфер для анализа, используют для определения фона. После добавления соответствующего количества связующего (концентрация антитела, аналогичная или ниже ожидаемой KD, конечный объем 60 мкл), смесь инкубируют в течение ночи при комнатной температуре.
[00339] Планшеты MSD покрывают антигеном (30 мкл на лунку). После промывки планшета PBS с использованием 0,02% Tween-20 уравновешенные образцы переносят в эти планшеты (30 мкл на лунку) и инкубируют в течение 20 мин. После промывки 30 мкл на лунку меченого детектирующего антитела MSD-Sulfo-tag (против человеческого (Fab)2, окончательное разведение обычно 1:2000) добавляют в планшет MSD и инкубируют в течение 30 минут при комнатной температуре на шейкере Эппендорфа (700 об/мин).
[00340] После промывки планшета MSD и добавления 30 мкл/лунку буфера для считывания T с поверхностно-активным веществом, сигналы электрохемилюминесценции обнаруживают с использованием Sector Imager 6000 (Meso Scale Discovery, Gaithersburg, MD, USA).
[00341] Данные оценивают с помощью программного обеспечения XLfit (IDBS) с использованием настраиваемых моделей подгонки. Для определения KD молекул Fab используют следующую модель подгонки (согласно Haenel et al., 2005), модифицированной согласно (Abraham et al., 1996):
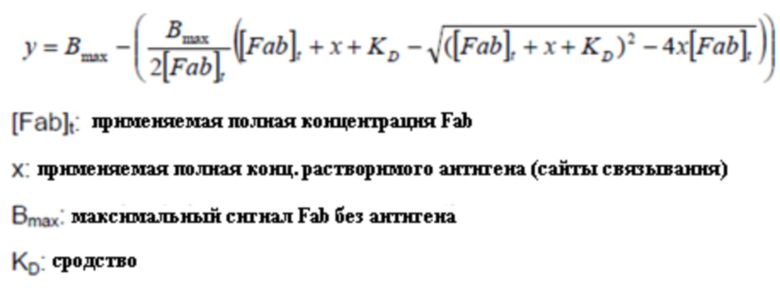
[00342] Для определения KD молекул IgG используют следующую модель подгонки для IgG (модифицированную в соответствии с Piehler et al, 1997):

[00343] Сродство также может быть определено поверхностным плазмонным резонансом на Biacore (SPR) путем определения кинетической константы скорости с использованием прибора Biacore 3000 или T200 (Biacore, GE Healthcare). Определение KD на Biacore с помощью антигена с прямым покрытием, в основном, выполняют следующим образом: 50 RU биотинилированного антигенного человеческого TSRP подвергают улавливанию на чипе SA (Biacore, GE Healthcare). Эталонная проточная ячейка 1 остается пустой. PBS pH 7,2 GIBCO+0,05% Tween 20 используют в качестве рабочего буфера с расходом 30 мкл/мин. Используют концентрации Fab в диапазоне от 3,9 до 500 нМ с объемом инъекции 45 мкл и временем диссоциации 300 с. Регенерацию связанного анализируемого вещества осуществляют с помощью 2-х кратных инъекций 5 мкл 10 мМ глицина, рН 1,5. Необработанные данные аппроксимируют на модели подгонки 1:1, при этом параметр(ы) Rmax устанавливают локально, а RI устанавливают как 0.
Созревание аффинности
[00344] Семь кандидатов из Fab были отобраны для созревания аффинности. Для увеличения сродства и биологической активности выбранных Fab, участки L-CDR3 и HCDR2 были оптимизированы параллельно с помощью кассетного мутагенеза с использованием тринуклеотид-направленного мутагенеза (Virnekas et al., 1994, Nucleic Acids Res 22: 5600-5607), тогда как каркасные участки поддерживают постоянными. Для оптимизации L-CDR3 исходных Fab-фрагментов, LCDR3, каркас 4 и константный участок легкой цепи (405 пар оснований) пула связующих удаляют ферментативным расщеплением и заменяют набором диверсифицированных L-CDR3 вместе с каркасом 4 и константным доменом. Во втором наборе библиотеку диверсифицируют H-CDR2, а соединяющие каркасные участки поддерживают постоянными. Лигирующие смеси электропорируют в 4 мл клетках TOP10F E. coli, получая от 108 до 109 независимых колоний. Этот размер библиотеки обеспечивает покрытие теоретического разнообразия. Амплификацию библиотеки проводят, как описано (Rauchenberger et al., 2003, J Biol Chem 278: 38194-38205). Для контроля качества отдельные клоны случайно выбирают и секвенируют. Для выбора связующих с улучшенной аффинностью фаги, полученные из библиотек созревания, подвергают трем циклам пэннинга в растворе с использованием биотинилированного антигенного человеческого TSLP_Avi-APP6 и обезьяньего TSLP_APP6-Avi. Жесткость условий увеличивают за счет снижения концентрации антигена в каждом цикле пэннинга (Low et al., 1996, J Mol Biol 260, 359-368. 1996.). В дополнение к снижению антигена проводят селекцию по скорости диссоциации (Hawkins et al., 1992, J Mol Biol 226, 889-896). Это объединяют с продолжительными стадиями промывки в течение ночи при комнатной температуре.
[00345] Для дальнейшего увеличения сродства и биологической активности некоторых выбранных фрагментов антитела, участки L-CDR1, L-CDR3, H-CDR2, H-CDR1 оптимизируют параллельно методом кассетного мутагенеза с использованием тринуклеотид-направленного мутагенеза (Virnekas et al., 1994, Nucleic Acids Res 22: 5600-5607), в то время как каркасные участки поддерживают неизменными.
[00346] Посттрансляционные модификации (PTM) в CDR не желательны, поскольку активность таких антител потенциально может быть уменьшена в зависимости от положения PTM, кроме того, ПТМ могут приводить к негомогенному соединению. Перед созреванием аффинности, варианты, лишенные сайтов NG, NS и DG, были сгенерированы и включены в пул с родительским клоном с целью выбора удаленных вариантов PTM во время процесса отбора. Fab, содержащий необработанные лизаты бактериальных клеток сгенерированных вариантов, тестируют на связывание антигена в ELISA на человеческом TSLP. Плазмидную ДНК вариантов смешивают с родительской ДНК для генерации библиотек созревания.
[00347] Для ранжирования созревших связующих веществ методом равновесного титрования в растворе на основе принципов, описанных Haenel et al., 2005, Anal Biochem 339: 182-184, постоянное количество разведенного экстракта BEL уравновешивают в течение ночи с разными концентрациями антигена. Затем смесь переносят в планшеты MSD, которые предварительно покрывают антигеном, и после инкубации и промывки добавляют подходящее меченое детектирующее антитело MSD-Sulfo-метка. Затем концентрацию несвязанного Fab количественно определяют путем детекции ECL с использованием Sector Imager 6000 (Meso Scale Discovery, Gaithersburg, MD, USA). Результаты обрабатывают с использованием программного обеспечения XLfit (IDBS), применяя соответствующую модель подгонки для оценки аффинности и, таким образом, идентификации клонов, наиболее улучшенных посредством созревания.
Продуцирование
[00348] Эукариотические клетки HKB11 трансфицируют с использованием ДНК-вектора экспрессии pMORPH®4, кодирующей как тяжелые, так и легкие цепи Fab или IgG против TSLP. Супернатант клеточной культуры собирают через 3 или 6 дней после трансфекции. После стерильной фильтрации раствор подвергают аффинной хроматографии на белке А (MabSelect SURE, GE Healthcare) с использованием станции обработки жидкости. Если не указано иное, буферный обмен проводят с использованием PBS по Dulbecco (pH 7,2, Invitrogen) и образцы подвергают стерильной фильтрации (размер пор 0,2 мкм). Концентрации белка определяют с помощью УФ-спектрофотометрии, а чистоту IgG анализируют при денатурирующих, восстанавливающих условиях с использованием системы Labchip (Perkin Elmer, USA).
Fab1 против TSLP
[00349] Fab1 против TSLP получают из семейства MOR011086, идентифицированного при первоначальных пэннингах. Созревание аффинности MOR011086 приводит к генерации MOR014051, который содержит мотив посттрансляционной модификации DG в HC-CDR2. Удаление этого мотива DG приводит к получению MOR14701 (DG→DA), который затем превращают в эмбриональную линию для получения MOR014824, то есть Mab1 в таблице 2. Fab1 против TSLP в таблице 2 представляет собой Fab-фрагмент Mab1.
[00350] Аминокислотные последовательности Fab1 CDR тяжелой цепи антитела против TSLP (HCDR), CDR легкой цепи (LCDR), а также аминокислотные последовательности вариабельных областей тяжелой и легкой цепей были определены по методу Кабат, Чотиа или с использованием комбинированных схем нумерации и приведены в таблице 2. Fab1 против TSLP связывается с очень высоким сродством (KD=6 пМ) с рекомбинантным человеческим TSLP, что определяют методом SET. Fab1 против TSLP не связывается со структурно подобным цитокином, IL-7.
Пример 2. Активность Fab1 против TSLP по отношению к рекомбинантному и естественно секретируемому человеческому TSLP в анализах с использованием репортерного гена
[00351] Активность Fab1 против TSLP по отношению к рекомбинантному человеческому TSLP, естественно секретируемому человеческому TSLP и обезьяньему TSLP тестируют в анализе с использованием репортерного гена люциферазы.
Материалы и методы
[00352] Естественно секретируемый человеческий TSLP получают из фибробластных клеток легкого человека путем стимуляции с использованием IL-1β, TNF-α и IL-13 в течение 24 часов.
[00353] Клетки Ba/F3 трансфицируют с использованием hTSLPR, hIL7Rα и конструкции репортера Stat5-люциферазы. Stat5 является нижележащим эффектором сигнального пути TSLP. Клетки выращивают в среде для выращивания клеток: RPMI 1640 (Invitrogen, Grand Island, NY), содержащей 10% FCIII (Fisher Scientific, Pittsburgh, PA), 1% пенициллина/стрептомицина (Invitrogen, Grand Island, NY), 1 мкг/мл пуромицина (Sigma, St. Louis, MO) и 5 нг/мл рекомбинантного человеческого TSLP (rhTSLP, R&D Systems, Minneapolis, MN). Буфер для репортерного анализа получают с использованием RPMI 1640 с 10% FCIII, 1% пенициллина/стрептомицина и 1 мкг/мл пуромицина.
[00354] Клетки Ba/F3 выращивают в суспензии в колбе T162 см2 и разделяют 1:50 дважды в неделю. Клетки Ba/F3 собирают на среднелогарифмической стадии роста, осаждают центрифугированием при 200×g в течение 5 минут и промывают средой для выращивания клеток, не содержащей TSLP. Эту операцию повторяют, а затем инкубируют в течение 18-24 в отсутствии TSLP. На следующий день клетки снова осаждают центрифугированием при 200×g в течение 5 минут и ресуспендируют в буфере для репортерного анализа до концентрации клеток 1 × 106 клеток/мл. 10 мкл клеток Ba/F3 при 1 × 106 клеток/мл объединяют с 70 мкл буфера для репортерного анализа в каждой лунке белого 96-луночного планшета Optiplate (Perkin Elmer, Waltham, Massachusetts). Далее проводят серийное 6-точечное разведение антитела 1:10 по 10 мкл (конечная верхняя концентрация конечного вещества 100 нМ) и инкубируют в течение 30 минут при 37°С/5% СО2 во влажном инкубаторе. Наконец, получают 10 мкл из 0,5 нг/мл человеческого или обезьяньего TSLP или рассчитанную концентрацию естественно секретируемого TSLP с той же относительной активностью, планшет герметизируют для уменьшения испарения и инкубируют в течение 4 часов при 37°C/5% CO2 во влажном инкубаторе. Планшет затем удаляют из инкубатора и уравновешивают при комнатной температуре в течение примерно 15 минут. Затем в каждую лунку добавляют 100 мкл реагента Steady-Glo (Promega, Madison, WI) и инкубируют при комнатной температуре в течение 20 минут. Затем планшеты считывают на приборе Envision, используя программу люминесценции (экспозиция камеры 1 секунда на лунку) и данные анализируют с использованием программ Microsoft Excel и Graphpad Prism.
Результаты
[00355] Fab1 против TSLP обладает превосходной активностью по отношению ко всем трем формам TSLP в анализе с репортерным геном люциферазы, с IC50, равным 15,4 мкМ против рекомбинантного человеческого TSLP (1 нг/мл), IC50, равным 17,1 пМ против естественно секретируемого человеческого TSLP и IC50, равным 10,8 пМ против обезьяньего TSLP. Рассчитывают средние результаты аналитического теста с репортерным геном для нескольких экспериментов (n=3), средние значения IC50 для Fab1 против рекомбинантного человеческого TSLP составляют 15,3 пМ±1,5 пМ SEM. Средние значения IC50 для Fab1 против обезьяньего TSLP составляют 9,5 пМ±0,9 пМ SEM.
[00356] Таким образом, Fab1 против TSLP является мощным ингибитором человеческого и обезьяньего TSLP с пикомолярной активностью. Тот факт, что Fab1 против TSLP демонстрирует превосходную активность против естественно секретируемого TSLP из человеческих легочных фрибробластов, уменьшает вероятность проблем, вызванных дифференциальным гликозилированием активного человеческого TSLP в организме и рекомбинантным человеческий TSLP, используемым для получения Fab против TSLP.
Пример 3. Ингибирование индуцируемой TSLP секреции TARC (хемокина, регулируемого тимусом и активацией) из первичных мононуклеарных клеток периферической крови человека (PBMC) посредством Fab1 против TSLP
[00357] Чтобы определить, способен ли Fab1 против TSLP нейтрализовать TSLP в контексте ответа, управляемого первичными клетками, индуцированную человеческим или обезьяньим TSLP секрецию TARC из человеческих PBMC протестируют в присутствии или отсутствии Fab1 против TSLP.
Материалы и методы
[00358] Венозную кровь, взятую у здоровых доноров, гепаринизируют (Sigma, St. Louis, MO) и собирают в шприцы с объемом 50 мл, а затем разделяют на две стерильные пробирки Falcon, по 25 мл в каждой. Эти пробирки центрифугируют при 1200 об/мин в течение 20 минут с низким ускорением и замедлением перед удалением плазменного слоя с использованием пипетки Пастера. 20 мл крови из каждой пробирки переносят в свежие пробирки Falcon объемом 50 мл и в каждую добавляют 20 мл PBS (1x, Invitrogen, Grand Island, NY) и 10 мл 4% декстрана (м./об., Sigma, St. Louis, MO). Пробирки переворачивают для тщательного перемешивания смеси крови и декстрана, и затем их инкубируют при комнатной температуре в течение 30 минут, чтобы осадить эритроциты. 20 мл супернатанта переносят в свежую пробирку Falcon объемом 50 мл и промывают 30 мл PBS (1400 об/мин в течение 8 минут) перед аспирацией супернатанта и ресуспендированием клеточного осадка в 10 мл PBS.
[00359] Чтобы лизировать красные кровяные клетки, к клеткам добавляют 20 мл стерильной холодной дистиллированной воды (Sigma, St. Louis, MO) и смешивают с использованием пипетки объемом 20 мл в течение 1 минуты до добавления 20 мл стерильного холодного 2×PBS для остановки лизиса. Пробирки несколько раз переворачивают и центрифугируют при 1400 об/мин в течение 8 минут перед объединением в одной пробирке и дважды промывают буфером для анализа (1400 об/мин, 8 минут). Буфер для анализа получают с использованием RPMI 1640 (с GlutaMax, Invitrogen, Grand Island, NY), содержащей 10% человеческой AB-сыворотки (Life Technologies, Grand Island, NY) и 1% пенициллина/стрептомицина (Invitrogen, Grand Island, NY).
[00360] Клетки подсчитывают и ресуспендируют при концентрации 10×106 клеток на мл, из которых 100 мкл добавляют в каждую лунку 96-луночного плоскодонного планшета (1×106 клеток на лунку). 50 мкл/на лунку антител против TSLP добавляют в каждую лунку и оставляют инкубироваться в течение 30 минут при 37°C перед добавлением человеческого или обезьяньего TSLP, получая конечную концентрацию TSLP, равную 1 нг/мл (66 пМ). Клетки инкубируют в течение 24 часов перед центрифугированием планшетов при 1300 об/мин в течение 5 минут и супернатанты собирают для анализа хемокина, регулируемого тимусом и активацией (TARC), с использованием ELISA. Супернатанты хранят при -20°С до тех пор, пока их не оттаивают для анализа TARC методом ELISA (образцы тестируют в чистом виде).
[00361] Анализ TARC методом ELISA проводят, следуя протоколам производителя (R&D Systems, Minneapolis, MN). Вкратце, иммобилизованные антитела разбавляют до рабочей концентрации в PBS в отсутствии белка-носителя. Микропланшеты ImmunomaxiSorp (Fisher Scientific, Pittsburgh, PA) покрывают с использованием 100 мкл на лунку разведенного иммобилизованного антитела, планшеты герметизируют верхними герметизирующими крышечками и инкубируют в течение ночи при комнатной температуре. На следующий день иммобилизованные антитела аспирируют и планшеты промывают промывочным буфером, повторяя процесс два раза на протяжении в общей сложности трех промывок. Лунки промывают, заполняя каждую лунку 300 мкл промывочного буфера, используя многоканальный дозатор или автоматизированное устройство для промывки. После последней промывки оставшийся промывочный буфер удаляют путем переворачивания планшета и промокания его чистыми бумажными полотенцами. Затем планшет блокируют добавлением 300 мкл разбавляющего реагента (1% BSA в PBS) в каждую лунку. Планшеты инкубируют при комнатной температуре в течение как минимум 1 часа. Стадии промывки повторяют и 100 мкл образца или стандартов в разбавителе для реагентов добавляют на лунку. Планшеты покрывают липкой лентой и инкубируют в течение 2 часов при комнатной температуре. Затем повторяют стадии аспирации/промывки и добавляют 100 мкл разбавленного детекторного антитела в каждую лунку, покрывают новой липкой полоской и инкубируют в течение 2 часов при комнатной температуре, прежде чем повторять стадию промывки, как описано ранее. 100 мкл рабочего разведения стрептавидина-HRP добавляют в каждую лунку, после чего планшеты повторно закрывают и инкубируют в течение 20 минут при комнатной температуре, избегая помещения планшета под прямой свет. Затем повторяют стадии аспирации/промывки и 100 мкл раствора субстрата ТМБ добавляют в каждую лунку. Планшеты инкубируют в течение 20 минут при комнатной температуре в темноте с последующим добавлением 50 мкл стоп-раствора. Планшет осторожно встряхивают, чтобы обеспечить смешение лунок, и сразу определяют оптическую плотность в каждой лунке с использованием ридера для микропланшетов, установленного при 450 нм.
Результаты
[00362] Fab1 против TSLP является очень мощным ингибитором индуцированной рекомбинантным человеческим TSLP секреции TARC из человеческих PBMC с IC50, равным 20,3 пМ, и IC90, равным 99,65 пМ, против 1 нг/мл рекомбинантного человеческого TSLP. Было показано, что Fab1 против TSLP является мощным ингибитором индуцированной обезьяньим TSLP секреции TARC из человеческих PBMC с IC50, равной 11,3 мкМ, против 1 нг/мл рекомбинантного обезьяньего TSLP. Когда были рассчитаны средние результаты нескольких экспериментов (n=3) для человеческих РВМС, средние значения IC50 для Fab1 против рекомбинантного человеческого TSLP составляют 19,7 пМ ± 1,9 пмоль SEM. Средние значения IC50 для Fab1 против обезьяньего TSLP составляют 11,1 пМ ± 0,5 пМ SEM.
Пример 4: Ингибирование индуцированной TSLP секреции MDC (макрофагального хемокина) из первичных мононуклеарных клеток периферической крови обезьяны (PBMC) посредством Fab1 против TSLP
Материалы и методы
[00363] Венозную кровь обезьян собирают в вакуумные пробирки, содержащие литиевый гепарин Covance (Dedham, MA). 30 мл крови каждого донора переносят в 50 мл пробирки Falcon и центрифугируют при 1200 об/мин в течение 20 минут с низким ускорением и замедлением перед удалением плазменного слоя с помощью пипетки Пастера, оставляя зазор 0,5 см между слоями. Оставшийся нижний слой клеток ресуспендируют и 10 мл переносят в свежие пробирки Falcon с последующим добавлениеи 10 мл 1× PBS и 5 мл 4% декстрана (мас./об., Sigma, St. Louis, MO) перед переворачиванием пробирок 4-5 раз для тщательного перемешивания. Все пробирки инкубируют при комнатной температуре в вытяжном шкафу в течение 25 минут, чтобы обеспечить осаждение эритроцитов на дне пробирки. 10 мл супернатанта переносят в свежую пробирку Falcon с объемом 50 мл и промывают 40 мл культуральной среды (1400 об/мин в течение 8 минут) перед аспирированием супернатанта и ресуспендированием клеточного осадка в 5 мл 1×PBS.
[00364] Чтобы лизировать красные кровяные клетки, к клеткам добавляют 20 мл стерильной холодной дистиллированной воды (Sigma, St. Louis, MO) и перемешивают с использованием пипетки объемом 20 мл в течение 1 минуты перед добавлением 20 мл стерильного холодного 2×PBS для остановки лизиса. Пробирки несколько раз переворачивают и центрифугируют при 1400 об/мин в течение 8 минут, затем объединяют в одну пробирку и дважды промывают культуральной средой (1400 об/мин, 8 минут, 4°С). Культуральную среду получают с использованием RPMI 1640 (с GlutaMax, Invitrogen, Grand Island, NY) с 10% фетальным клоном III (Fisher Scientific, Pittsburgh, PA) и 1% пенициллина/стрептомицина (Invitrogen, Grand Island, NY).
[00365] Клетки подсчитывают с использованием трипанового синего красителя и ресуспендируют при концентрации 10 × 106 клеток на мл, 100 мкл которых добавляют в каждую лунку 96-луночного плоскодонного планшета (1×106 на лунку). 50 мкл/на лунку антитела против TSLP (верхняя конечная концентрация 100 нМ) добавляют в каждую лунку и оставляют для инкубации в течение 30 минут при 37°C перед добавлением обезьяньего TSLP, получая конечную концентрацию равную 0,5 нг/мл TSLP (33 пМ). Клетки инкубируют в течение 24 часов перед центрифугированием планшетов при 1400 об/мин в течение 8 минут и супернатанты собирают для анализа макрофагального хемокина (MDC, CCL22) с помощью ELISA. Супернатанты хранят при -20°С до их оттаивания для анализа MDC методом ELISA (разведенные 1:2 в буфере для анализа перед добавлением в планшет ELISA). Анализ MDC методом ELISA проводят, следуя протоколам производителя (R&D Systems, Minneapolis, MN).
Результаты
[00366] Показано, что Fab1 против TSLP является мощным ингибитором индуцированной рекомбинантным обезьяньим TSLP секреции MDC из обезьяньих PBMC с IC50, равной 55,5 мкМ, против 0,5 нг/мл рекомбинантного обезьяньего TSLP. Когда были рассчитаны средние результаты нескольких экспериментов (n=3) для обезьяньих PBMC, средние значения IC50 для Fab1 против обезьяньего TSLP составляют 25,1 пМ ± 5,9 пМ SEM.
Пример 5: Межвидовая перекрестная реактивность Fab1 против TSLP
Материалы и методы
[00368] Проводят анализ связывания с использованием поверхностного плазмонного резонанса (SPR) на Biacore, чтобы установить, связывается ли Fab1 против TSLP с человеческим, мышиным или крысиным белком TSLP. Реагенты Biacore, включая сенсорный чип S5 серии S, буфер HBS-EP+, набор иммобилизованных человеческих Fab, EDC (1-этил-3-(3-диметиламинопропил)-карбодиимид), NHS (N-гидроксисукцинимид), этаноламин, BIA-нормализующий раствор, 70% (мас./мас.) глицерина и глицин, были приобретены у GE Healthcare. Подвижный буфер, используемый как для анализа захвата Fab, так и для связывания TSLP, представлял собой I× HBS-EP+, с 10 мМ HEPES (pH 7,4), 150 мМ NaCl, 3 мМ ЭДТА, 0,05% об./об. поверхностно-активного вещества Р20. Рекомбинантные человеческий, обезьяний или мышиный TSLP (ММ 15 кДа) были получены от R&D Systems ((Minneapolis, MN)). Рекомбинантный крысиный TSLP (ММ 15,4 кДа) получают от USCN Life Science Inc. (Ухань, Китай). Подход к захвату используют для подготовки Fab1 против TSLP на чипе CM5 Biacore перед инъекцией человеческого, мышиного или крысиного TSLP. Связующее человеческого Fab было иммобилизовано на всех четырех проточных ячейках чипа CM5 с использованием набора для захвата человеческих Fab, следуя инструкциям производителя. Было установлено время контакта, равное 360 секундам, при скорости потока 10 мкл/мин. Температура отделения для образцов составляет 10°С, а температура анализа составляет 25°С; перед иммобилизацией чип CM5 был примирован HBS-EP+и нормализован с использованием BIA-нормализующего раствора. 375 мкл из 20 мкг/мл связующего человеческого Fab получают посредством объединения 15 мкл 0,5 мг/мл исходного раствора с 360 мкл иммобилизационного буфера с pH 5 (оба предоставлены в наборе для захвата человеческих Fab). Результирующие уровни иммобилизации составляют приблизительно 4000-4400RU связующего человеческого Fab в Fc 1, 2, 3 и 4.
[00369] Пользовательский метод Biacore применяют для разработки кинетического анализа, в котором прибл. 14RU Fab1 против TSLP захватывалось за цикл. Этого достигают путем инжекции 5 нМ Fab1 против TSLP в буфер HBS-EP+ с временем контакта 60 с при скорости потока 10 мкл/мин с последующим периодом стабилизации 30 с. Температура отделения для образцов составляет 10°С, а температура анализа составляет 25°С. Используя этот пользовательский метод Biacore, разрабатывают кинетический анализ для оценки взаимодействия hTSLP, mTSLP и rTSLP с захваченным Fab1 против TSLP. Для каждого антигена получают следующие 10 концентраций в HBS-EP+ и вводят над поверхностью Fab1 против TSLP, включая чистый буфер с 0 nM, 10 нΜ, 5 нМ, 2,5 нМ, 1,25 нМ, 0,625нМ, 0,313 нМ, 0,156 нМ, 0,078 нм, 0,039 нМ, 0,02 нМ. После захвата ~14RU Fab1 против TSLP антиген вводят при 45 мкл/мин на 360 секунд с последующим периодом диссоциации 600 с (для всех тестируемых концентраций) или 1200 с (для концентраций антигена 0 нМ и 2,5 нм). Регенерацию поверхности Fab-связующего проводят после каждого цикла путем инъекции 10 мМ глицина-HCl, pH 2,0 в течение 60 с при 10 мкл/мин с последующей дополнительной промывкой буфером HBS-EP+. Температура отделения для образцов составляет 10°С, а температура анализа составляет 25°С.
[00370] Все эксперименты и анализы SPR выполняют на приборах Biacore T200, управляемых программным обеспечением Biacore T200 Control. Данные обрабатывают с использованием программного обеспечения для оценки Biacore T200. Для качественного анализа межвидовой перекрестной реактивности Fab1 против TSLP на график наносят сенсограммы с вычитанием холостого результата.
Результаты
[00371] Результаты экспериментов по перекрестной реактивности с использованием Biacore SPR показывают плотное связывание Fab1 против TSLP с рекомбинантным человеческим TSLP, тогда как связывание с рекомбинантным крысиным или мышиным TSLP не наблюдают, что согласуется с низкой гомологией человеческого TSLP и TSLP грызунов (примерно 40%).
[00372] Fab1 против TSLP связывается с очень высоким сродством с рекомбинантным TSLP обезьян-макак и является очень мощным ингибитором рекомбинантного обезьяньего TSLP (IC50=10,8 мкМ против 1 нг/мл рекомбинантного TSLP) в анализе с использованием репортерного гена люциферазы. В обоих анализах с использованием первичных PBMC человека и обезьяны, Fab1 против TSLP является очень мощным ингибитором секреции TARC из человеческих PBMC, и секреции MDC из обезьяньих PBMC, индуцированной рекомбинантным обезьяньим TSLP (IC50=11,3 пМ) (IC50=55,5 пМ).
[00373] Таким образом, Fab1 против TSLP показывает ограниченную межвидовую перекрестную реактивность с распознаванием рекомбинантного обезьяньего TSLP, но не крысиного или мышиного TSLP.
Пример 6: Эффективность мышиного антитела против TSLP на мышиных моделях заболевания астмой
Материалы и методы
[00374] Влияние нейтрализации TSLP на аллергические реакции дыхательных путей оценивают на мышиной модели системной сенсибилизации овальбумином (OVA) с последующей локальной антигенной стимуляцией легких. Эта модель характеризуется развитием фенотипа Th2 и ассоциированного эозинофильного воспаления. Поскольку Fab1 против TSLP примера 1 не взаимодействует перекрестно с белками TSLP грызунов, как описано в примере 5, влияние нейтрализации TSLP оценивают с использованием доступного для приобретения суррогатного моноклонального антитела против мышиного TSLP (MAB555, R&D Systems, Minneapolis, MN), причем сообщается о полной нейтрализации биологической активности рекомбинантного мышиного TSLP с IC50 примерно равной 1,3 нМ против 0,5 нМ мышиного TSLP (данные, полученные от R&D Systems). Конкретные наборы ELISA для всех цитокинов и хемокинов также были приобретены у R&D Systems.
[00375] Самок мышей Balb/c иммунизируют OVA (или физиологическим раствором) и квасцами в качестве адъюванта в день 1 и день 14. Вкратце, мышей иммунизируют внутрибрюшинно с использованием 0,2 мл 0,9% мас./об. NaCl (физиологический раствор), содержащего 100 мкг овальбумина (5× кристаллизованного, Sigma, UK), адсорбированного в 1,6 мг гидроксида алюминия (Sigma). На 21-й день мышей стимулируют OVA или физиологическим раствором в виде аэрозоля и сортируют через 24 часа. Воспаление оценивают с помощью дифференциального и общего количества клеток в бронхоальвеолярном лаваже (BAL), в то время как цитокины и хемокины измеряют с помощью специфического ELISA.
[00376] Через двадцать четыре часа после последней интраназальной стимуляции OVA или PBS, мышей анестезируют посредством внутрибрюшинной инъекции 4 мг/кг натрия пентобарбитала (Rhone Merieux, Harlow, UK). Жидкость BAL собирают путем канюлирования трахеи и промывки легких в общей сложности 1,2 мл солевого раствора (3×0,4 мл каждый). Для каждого образца определяют общее количество клеток и проводят подготовку цитоспина (Shandon Scientific Ltd., Cheshire, UK). Клетки окрашивают Diff-Quik (Baxter Dade AG, Dudingen, Швейцария) и дифференциальный подсчет 200 клеток выполняют с использованием стандартных морфологических критериев.
[00377] Чтобы оценить влияние истощения TSLP на фазу сенсибилизации ответа, моноклональное антитело против мышиного TSLP (при 10 мг/кг) или изотипический контроль IgG2a крысы вводят внутривенно за один час до сенсибилизации OVA и снова перед стимуляцией иммунизации на день 14. Чтобы оценить роль TSLP во время стимуляции, некоторым мышам дают антитело только за один час до аэрозолизации OVA на день 21. Никаких побочных эффектов при внутривенном введении этих антител не наблюдают.
[00378] Результаты выражают в виде средних значений ± SEM для указанного количества экспериментов. Один из способов анализа дисперсии (ANOVA) используют для определения значимости среди групп. Если обнаружена значительная дисперсия, для оценки сравнимости между средними значениями используют непарный t-тест Стьюдента. Значение р<0,05 считается значимым.
Результаты
[00379] Сенсибилизация и стимуляция ОВА приводит к увеличению числа клеток в бронхоальвеолярной лаважной жидкости, которая содержит эозинофилы и нейтрофилы по сравнению с контрольными животными (Фиг. 3). Это согласуется с предыдущим опытом ответов после единственной стимуляции антигеном. Кроме того, ряд воспалительных медиаторов также повышается в лаважной жидкости мышей, сенсибилизированных/стимулированных OVA по сравнению с контролями (фиг. 4А-4С).
[00380] Обработка мышиными антителами против TSLP (10 мг/кг) значительно ингибирует общее количество клеток в жидкости BAL примерно на 50%, в то время как количество эозинофилов уменьшается на 80%. Лечение антителом в отсутствии сенсибилизации антигеном не приводит к существенному изменению исходного клеточного состава лаважа. Анализ нисходящих маркеров активности TSLP показывает снижение уровней IL-13 (Фиг. 4A), цитокина, ассоциированного с аллергическим воспалением дыхательных путей, и хемокинов эотаксина-2 и TARC (Фиг. 4B и 4C), оба из которых являются известными хемоаттрактантами Th2-клеток и эозинофилов, которые генерируютсь TSLP-стимулированными дендритными клетками.
Пример 7: Фармакокинетическая характеризация Fab1 против TSLP у крыс
Материалы и методы
[00381] Фармакокинетику (PK) и распределение в легких Fab1 против TSLP исследуют у крыс после внутривенной (ВВ) болюсной инъекции, интратрахеальной инстилляции (ИТИ) или 20-минутной ингаляции только через нос однократной номинальной дозы распыленного Fab1 против TSLP при 1 мг/кг. Концентрации Fab1 против TSLP в разные моменты времени после дозы определяют в плазме, жидкости BAL, а также в образцах гомогената легкого (после BAL и перфузии крови сосудистой системы).
Результаты
[00382] Fab1 против TSLP быстро выводится из системного кровообращения после ВВ инъекции со средним периодом полувыведения около 3 часов. После ITI или ингаляции Fab1 против TSLP медленно всасывался в системное кровообращение, достигая плазменной Cmax примерно за 2 часа для обоих путей введения, а средние конечные периоды полувыведения были длиннее, чем те, которые были определены после ВВ введения (через 7 часов после ITI и 4 ч после ингаляции, по сравнению с 3 ч после ВВ), что указывает на кинетику с ограничением скорости всасывания. Системная биодоступность Fab1 против TSLP составляет в среднем примерно 6% после ITI и 1% после ингаляции, возможно, из-за более высокой фракции отложения в легких после ITI по сравнению с ингаляцией. По сравнению с низкой системной экспозицией, концентрации Fab1 против TSLP в жидкости BAL и гомогенате легкого были намного выше (более чем в 100 раз выше) после ITI или ингаляции, составляя 97-99% от общего количества дозы, извлеченной из всех трех матриц (66-79% для BAL и 20-31% для легких) через 2, 6, 24 или 72 часа после дозирования. Расчетные периоды полужизни Fab1 против TSLP составляют в среднем примерно 7 и 9 часов в BAL и гомогенатах легких соответственно.
Пример 8: Фармакокинетическая характеристика Fab1 против TSLP у обезьян
Материалы и методы
[00383] Токсикокинетику, ФК/ФД и распределение в легких Fab1 против TSLP исследуют у яванских макак после ежедневной 1-часовой ингаляции в течение 14 дней при дозах 1, 10 и 20 мг/кг (группы 3-5) или перекрестного ВВ введения однократной дозы 1 мг/кг, с последующей одной ингаляцией дозы 20 мг/кг после 16-дневного периода вымывания (группа 6). Серийные образцы крови собирают для ФК/ФД, общий TSLP оценивают как маркер ФД и оценки иммуногенности. Кроме того, образцы проб гомогената легкого (в конце) и образцах жидкости БАЛ (терминальные для Групп 3-5 и до введения внутривенной дозы и терминальные для ФК Группы 6) также собирают для оценок ФК, общего TSLP и иммуногенности (только для жидкости БАЛ).
Результаты
[00384] Системное воздействие Fab1 против TSLP в сыворотке было низким после ингаляции с оценкой биодоступности менее 1% при уровне ингалируемой дозы 20 мг/кг. Ингалируемая доза 1 мг/кг не вызывает детектируемых системных эффектов, а ингалируемые дозы, равные 10 и 20 мг/кг, показывают сравнимое системное воздействие Fab1 против TSLP. Cмакс достигалась через 3 часа после ингаляции. Аналогично ФК у крыс, периоды системного полувыведения были более продолжительными после ингаляции (примерно 7 часов) по сравнению с ВВ (примерно 2,3 часа), что указывало на кинетику с ограничением скорости всасывания. Накопление экспозиции в сыворотке наблюдалось после 14 дней дозирования. По сравнению с низкой экспозицией в сыворотке (Фиг. 5) предварительные данные о концентрациях Fab1 против TSLP в терминальной жидкости БАЛ и гомогенатах легких были значительно выше и увеличиваются с увеличением доз (Фиг. 6).
Пример 9: Кристаллография и картирование эпитопа Fab1 против TSLP
[00385] В этом примере Fab1 против TSLP кристаллизуют в свободном состоянии или в комплексе с человеческим TSLP и определяют соответствующие кристаллические структуры. Анализ связывания Fab1 против TSLP с человеческим TSLP на основе данных рентгеновского исследования позволяет получить информацию о эпитопе Fab1 против TSLP на человеческом TSLP.
Материалы и методы
Получение и очистка человеческого TSLP и Fab1 против TSLP
[00386] Fab1 против TSLP получают путем ферментативной обработки mAb1 против TSLP (10,6 мг) 21 мкг папаина в течение 2 часов при комнатной температуре (RT) в 100 мМ Tris (pH 7,0) с 10 мМ DTT. Реакцию останавливают 30 мкл ингибитора папаина E64. Затем Fab1 против TSLP очищали на колонке Lambda Select объемом 5 мл, уравновешенной 20 мМ фосфатом натрия (pH 7,0). Fab элюируют 0,1 М лимонной кислотой, pH 3,0, и рН собранных фракций немедленно регулируют с помощью 1М Tris pH 8,5, разбавленного 1:10. Анализ методом ЖХ-МС показывает наблюдаемую массу 47107,7 Да, которая соответствовала ожидаемой аминокислотной последовательности с тяжелой цепью, расщепленной после Thr228 и имеющей остаток пироглутаминовой кислоты на аминоконце. Для кристаллизации буфер заменяют на 10 мМ Tris-HCl, рН 7,4, 25 мМ NaCl, с помощью повторных стадий разбавления-концентрирования с использованием устройства ультрафильтрации, и образец окончательно концентрируют до 13 мг/мл Fab1 против TSLP.
[00387] Конструкцию человеческого TSLP (вход Uniprot Q969D9, аминокислоты с 29 по 159) с N-концевой гексагистидиновой меткой (SEQ ID NO: 40) с последующим сайтом расщепления PreScission (HRV-3C протеазой) клонируют и экспрессируют в E. coli как тельца включения. Для рефолдинга 89,4 г клеток E. coli лизируют в 715 мл 50 мМ Tris (pH 7,0) с 1 мМ EDTA, 6 мМ MgCl2 и 0,375 М сахарозы с использованием гомогенизатора высокого давления Avestin®. После 30-минутной инкубации с 3,7 кЕ бензоназы лизат центрифугируют в течение 30 минут при 13000 об/мин с помощью ротора с фиксированным углом SS-34. Осадок ресуспендируют в 387 мл 100 мМ Tris (pH 7,0) с 20 мМ ЭДТА, 0,5 М NaCl, 2% Triton Х-100 и затем центрифугируют при 13500 об/мин в течение 50 минут. Осадок снова ресуспендируют в 387 мл 100 мМ Tris pH 7,0 с 20 мМ ЭДТА, центрифугируют при 13500 об/мин в течение 30 минут, и эту процедуру промывки повторяют четыре раза, что приводило к 13 г телец включения. Тельца включения затем солюбилизируют в 65 мл 6 М раствора гуанидина гидрохлорида с 50 мМ ацетата калия (рН 5,0), 5 мМ ЭДТА и 10 мМ TCEP. После 2-часовой инкубации при комнатной температуре образец центрифугируют в течение 30 минут при 20000 об/мин (ротор с фиксированным углом SS-34). Супернатант (70 мл) разбавляют до 100 мл раствором гидрохлорида гуанидиния, описанным выше. Рефолдинг проводят быстрым разбавлением при 4°С 10 л 100 мМ Tris (pH 8,25) с 0,5 М гидрохлоридом аргинина, 5 мМ ЭДТА и 1 мМ GSH. После разбавления добавляют 0,1 М дисульфида глутатиона (GSSG) и смесь рефолдинга инкубируют при медленном перемешивании в течение 7 дней при 4°C. Затем рН доводили до 5,1 с помощью уксусной кислоты и добавляют 0,1 мМ GSSG для разрушения оставшегося TCEP. Немного мутный рефолдинг-раствор фильтруют через фильтрующую капсулу Sartobran 0,65/0,45 мкм и концентрируют с помощью поперечной мембраны Pellicon 10 кДа до 750 мл. Концентрированный раствор диализовали против 10 л 50 мМ ацетата натрия, рН 5,4. Было извлечено примерно 550 мг восстановленного TSLP. Анализ ЖХ-МС конечного очищенного образца подтвердил, что были образованы все дисульфидные мостики и показывает 94% продукта des-Met (ММ=16862,8 Да) и 6% белка с N-концевым метионином. Для кристаллизации с использованием Fab1 против TSLP образец повторно сложенного TSLP используют без расщепления N-концевой метки протеазой PreScission.
[00388] Для получения комплекса TSLP-Fab добавляют двухкратный молярный избыток белка His6-PreSc-TSLP в 25 мМ Tris (рН 7,4) с 50 мМ NaCl, образец концентрируют ультрафильтрацией до примерно 10 мг/мл, загружали на колонку для эксклюзионной хроматографии SPX-75 и элюируют изократически в 10 мМ Tris-HCl, pH 7,4, с 25 мМ NaCl. Пиковую фракцию концентрируют до 9,2 мг/мл путем ультрафильтрации и подвергают кристаллизации.
Кристаллизация и сбор данных рентгеновского исследования
[00389] Кристаллы выращивают в 96-луночных планшетах (планшеты Innovadyne SD2) посредством диффузии паров в сидячей капле. В частности, 0,2 мкл белкового сырья смешивают с 0,2 мкл раствора резервуарного раствора, и каплю уравновешивают против 80 мкл того же резервуарного раствора при 20°С. Эксперименты устанавливают с помощью роботизированной системы Phoenix (Art Robbins Instruments), содержащейся в отделении RockImager (Formulatrix), и визуализируют автоматически.
[00390] Для сбора рентгеновских данных один кристалл непосредственно устанавливают в петле для замораживания и быстро охлаждают в жидком азоте. Наборы данных рентгеновских исследований собирают на источнике света швейцарского типа, с пучком излучения X10SA с детектором пикселей Pilatus, используя рентгеновское излучение 1,00001Å. В обоих случаях на расстоянии от кристалла до детектора, равном 345 мм, регистрируют 720 изображений с частотой колебаний 0,25° для каждого, и их обрабатывают, используя версию XDS Dec. 6, 2010, (Kabsch 1993, J Appl Crystallogr, 26: 795-800), как это выполнялось в APRV.
Определение и анализ структуры
[00391] Структуру Fab1 против TSLP определяют посредством молекулярной замены с использованием программы Phaser (McCoy et al, 2007, J Appl Crystallogr 40: 658-674) с использованием кристаллической структуры фрагмента Fab-фрагмента антитела против CD132 в качестве исходной модели. Fab антитела против CD132 выбирают на основе сходства последовательностей с Fab1 против TSLP. Вариабельные и первые константные участки используют в качестве независимых поисковых моделей для обеспечения изменчивости угла сгибания Fab. Структуру уточняют с использованием итерационных циклов построения модели с последующей автоматизированной кристаллографической обработкой программами Coot 0.8.0 (Crystallographic Object-Oriented Toolkit, Emsley et al, 2010, Acta Crystallogr Sect D: Biol Crystallogr, 66: 486-501) и Autobuster 2.11.5 (Bricogne et al, 2011, версия BUSTER 2.11.2. Кембридж, Великобритания: Global Phasing Ltd.).
[00392] Структуру комплекса TSLP-Fab определяют посредством молекулярной замены с помощью программы Phaser с использованием уточненных структур свободного Fab1 против TSLP и человеческого TSLP, ранее определенных на месте в комплексе с Fab-фрагментом другого антитела. Опять же, вариабельная и первая константные участки Fab1 против TSLP используют в качестве независимых поисковых моделей. Структуру уточняют, как описано выше для свободного Fab, с помощью Coot 0.8.0 и Autobuster 2.11.5.
[00393] Визуальный осмотр кристаллических структур проводят с использованием программ Coot (Emsley et al, 2010, Acta Crystallogr Sect D: Biol Crystallogr, 66: 486-501) и PyMOL (Molecular Graphics System, DeLano Scientific: Palo Alto, CA). Качество окончательных уточненных моделей оценивают с помощью программ Coot и PROCHECK v3.3 (Laskowski et al, 1992, J Appl Crystallogr, 26: 283-291). Остатки человеческого TSLP, которые становятся менее доступными для растворителя при связывании Fab1 против TSLP, были идентифицированы программой AREAIMOL из программного пакета CCP4 (Collaborative Computational Project, Number 4, 1994). Межмолекулярные контакты определяют с использованием расстояния отсечения 4,0 Ǻ и идентифицируют с помощью программы NCOCT CCP4.
Результаты
Кристаллическая структура Fab1 против TSLP
[00394] Свободный Fab1 против TSLP и его комплекс с человеческим TSLP кристаллизуют в 96-луночных планшетах методом диффузии паров в сидячей капле при 19°C. Интересно, что два образца белка кристаллизовались в одних и тех же условиях кристаллизации: 0,17 М (NH4)2SО4, 85 мМ ацетата натрия, рН 5,6, 25,5% PEG MME 2000, 15% глицерина. Кристаллы появляются через 4-5 недель и вырастают до полного размера в течение нескольких дней.
[00395] Свободный Fab-кристалл находится в орторомбической пространственной группе P212121 с одной молекулой Fab на асимметричную единицу. Кристалл комплекса Fab-TSLP находился в пространственной группе 1222 с одним комплексом на асимметричную единицу (Таблица 3). Оба кристалла имели дифракцию с высоким разрешением, и для каждого из них можно было собрать полный набор данных дифракции хорошего качества и с высоким резервированием (Таблица 3).
[00396] Определение структуры путем молекулярной замены проводят с использованием ранее определенной структуры человеческого TSLP. Уточнение с помощью аутобастера приводит к хорошей статистике уточнения и общей геометрии (Таблица 3). Два остатка антител, Asp50L и Asp152L, представляют собой полные маргинальные остатки по карте Рамачандрана в структуре свободного Fab. В дополнение к этим двум остаткам третий остаток антител, Tyr103H, также является полным маргинальным остатком Рамачандрана в структуре комплекса Fab-TSLP. Эти три остатка имеют четко определенную плотность электронов и, таким образом, являются подлинными геометрическими маргинальными остатками. Следует отметить, что Asp50L и Tyrl03H представляют собой остатки CDR, участвующие в связывании TSLP, как описано ниже.
[00397] Аминокислотные последовательности тяжелой и легкой цепей Fab1 против TSLP представлены на Фиг. 1A и 1B, с подчеркнутыми CDR (как определено Kabat, 1991, Sequences of proteins of immunological interest, NIH Publication No. 91-3242), а остатки, расположенные на границе раздела антитело-антиген, помечены *.
ТАБЛИЦА 3 Сбор данных рентгеновского исследования и статистика уточнения
Кристаллическая структура Fab1 против TSLP в комплексе с человеческим TSLP
[00398] Аминокислотная последовательность рекомбинантного человеческого TSLP, используемого в этом примере (SEQ ID NO: 38), представлена на фиг. 2. Зрелый человеческий TSLP начинается с Tyr29. Используемая здесь конструкция содержит N-концевую гексагистидиновую метку (SEQ ID NO: 40) (остатки 15-20) с последующим участком распознавания протеазы HRV-3C (PreScission) (остатки 21-28) и остатки 11-14, являющиеся результатом клонирования. Asn64 и Asn119 являются потенциальными участками N-связанного гликозилирования; а остатки 127-130 составляют потенциальный сайт расщепления фурином (RRKR, SEQ ID NO: 39). Элементы вторичной структуры показаны ниже аминокислотной последовательности: прямоугольники обозначают α-спирали A, B, C и D, а толстые линии обозначают петлеобразные участки.
[00399] TSLP не демонстрирует какого-либо значительного сходства аминокислотной последовательности с другими членами суперсемейства цитокинов IL-2. Тем не менее, TSLP укладывается в виде пучка с четырьмя спиралями с топологией вверх-вверх-вниз-вниз, как IL-2, IL-7 и многие другие цитокины (Фиг.7). Спираль αA обладает сильным изгибом вблизи своего центра, вокруг положения Thr46. Спирали αB and αC являются довольно короткими, только с тремя витками каждая, а C-концевая спираль αD длиннее почти на пять витков (фиг.7). Три дисульфида (Cys34-Cysll0, Cys69-Cys75, Cys90-Cysl37) стабилизируют этот короткоцепочечный четырехспиральный пучок. Однако два перекрестных соединения между αA и αB и αC и αD являются в основном неупорядоченными и не наблюдаются в этой кристаллической структуре. Три аминокислоты из петли αB-αC и последние пять карбоксильных концевых остатков также отсутствуют в окончательной уточненной структуре. Потенциальный участок расщепления фурином и участки N-гликозилирования расположены в пределах отсутствующих соединений.
[00400] Общий вид трехмерной структуры комплекса Fab-TSLP показан на фиг. 7.
[00401] Fab1 против TSLP связывается главным образом со спиралью αA TSLP, которая проходит через мелкую бороздку, выровненную H-CDR1 и H-CDR2 с одной стороны, и H-CDR3 и L-CDR3 с другой стороны. Резкий перегиб спирали αA занимает центральную часть бороздки. Фланкирующие спирали αC and αD и первые четыре остатка петли αA-αB обеспечивают дополнительный контакт с антителом.
[00402] Образование комплекса Fab-TSLP покрывает примерно 1700Ǻ2 комбинированной доступной для растворителя поверхности, с 25 аминокислотными остатками Fab и 25 аминокислотными остатками TSLP, которые испытывают уменьшение поверхности, доступной для растворителей, при образовании комплекса. Среди них 20 остатков Fab и 16 остатков TSLP (см. Табл. 4) участвуют в прямых межмолекулярных контактах при пороговом расстоянии 4,0 Ǻ. Статистика комплементарности формы Sc (Lawrence and Colman, 1993, J Mol Biol, 234: 946-50) составляет 0,72, относительно высокое значение для комплекса антитело-белок (Sundberg and Mariuzza, 2003, Adv Protein Chem, 61: 119- 60). Все шесть CDR Fab1 против TSLP обеспечивают связывание с TSLP. Кроме того, шесть четко определенных молекул воды, расположенных на границе антитело-антиген, опосредуют связывающие взаимодействия.
ТАБЛИЦА 4. Остатки эпитопа и паратопа
[00403] Остатки TSLP, расположенные на границе поверхности связывания, идентифицируют по кристаллографическим координатам путем вычисления (i) межмолекулярных контактов между неводородными атомами менее чем 4,0 Ǻ и (ii) уменьшения доступной для растворителя поверхности при образовании комплекса. Результаты показаны графически на Фиг. 8, вид эпитопа TSLP представлен на Фиг. 9. Как видно из этих двух чертежей, спираль αA вместе с первыми четырьмя остатками петли αA-αB образовала ядро эпитопа и способствовала 82% общего числа межмолекулярных контактов и задействованной поверхности на TSLP, доступной для растворителя. Кроме того, подавляющее большинство ключевых остатков эпитопа было обнаружено в данном участке: Lys49, Ile52, Gly57 и Lys59. Для сравнения, спирали αC and αD вносят очень мало остатков эпитопа: Lys101 (αC), Gin 145 и Argl49 (αD).
[00404] Все шесть определяющих комплементарность участков (CDR) Fab1 против TSLP вносят вклад в граничную поверхность связывания, о чем свидетельствует уменьшение их поверхности, доступной для растворителя, при связывании антигена и их вклад в прямые межмолекулярные контакты (фиг. 10А и 10В). Кроме того, Asn65L третьего каркасного участка легкой цепи (L-FR3) также расположен на антигенсвязывающей граничной поверхности, но он отвечает только за слабое (3,6Ǻ) H-связанное взаимодействие с Lys59 TSLP.
[00405] Особенно важную роль играет петля HCDR3. Glu101H участвует в решающем взаимодействии солевого мостика с Lys49 TSLP, а три последовательных тирозина, расположенных на кончике петли, Tyrl03H, Tyrl04H и Tyrl05H, в совокупности обеспечивают 58% контактов, осуществляемых всей тяжелой цепью. Trp33H H-CDR1 и Asp56H H-CDR2 также являются важными остатками паратопа, которые обеспечивают связывание Lys49 и Lys101 TSLP, соответственно.
[00406] Trp92L является единственным остатком L-CDR3, участвующим в непосредственном контакте с антигеном. Этот остаток расположен на кончике L-CDR3 и не принимает определенной конформации в кристаллической структуре свободного Fab1 против TSLP. Однако в комплексе с TSLP боковая цепь имеет четко выраженную электронную плотность и осуществляет обширные контакты с петлей αA-αB, ее вклад составляет до 42% контактов, создаваемых всей легкой цепью. Asp50L LCDR2 и Tyr31L LCDR1 представляют собой два других важных остатка паратопа, обеспечиваемых легкой цепью. Первый осуществляет очень короткое (2,8 Ǻ) электростатическое взаимодействие с Lys59 TSLP. Последний обеспечивает связывание с Ile52, Lys59 и Gln145 TSLP, а также стабилизирует связанную конформацию петли H-CDR3 через π-π взаимодействия с Tyrl03H и Tyrl04H.
Способ действия Fab1 против TSLP
[00407] Сигнальный путь TSLP требует сборки тройного комплекса, включающего TSLP, родственную цепь TSLPR и общую цепь IL-7Rα. Образование бинарного комплекса TSLP-TSLPR является необходимым условием для мобилизации цепи IL-7Rα. Комплекс человеческий TSLP-Fab1 накладывается на тройной комплекс мышиный TSLP-TSLPR-IL-7Rα, основанный на всех Cα атомах TSLP. Структурное наложение показывает, что Fab1 против TSLP блокирует связывание TSLP как с TSLPR, так и с IL-7Rα. Спираль αA TSLP является центральным элементом эпитопа Fab1 против TSLP (Фиг.11B), и эта спираль также играет главную роль в связывании как с TSLPR, так и с IL-7Rα (Фиг. 11А). Кроме того, спираль αC участвует в связывании IL-7Rα, а спираль αD является частью граничной поверхности связывания TSLPR. Поскольку эти две спирали также образуют эпитоп Fab1 против TSLP, пространственная интерференция между TSLP Fab1 и двумя рецепторными цепями является обширной. Легкая цепь антитела сильно перекрывается с доменом D2 TSLPR, тогда как тяжелая цепь перекрывается с доменом IL-7Ra D2, а также с некоторыми цитокиновыми петлями связывания домена D1 (Фиг. 11С). Данные демонстрируют, что Fab1 против TSLP нейтрализует TSLP, захватывая цитокин и предотвращая его связывание с рецептором TSLPR, тем самым блокируя образование высокоаффинного сигнального комплекса с IL-7Ra.
[00408] Таким образом, с высоким разрешением были определены кристаллические структуры Fab1 против TSLP в свободном состоянии или в комплексе с повторно свернутым рекомбинантным человеческим TSLP. Было обнаружено, что Fab1 против TSLP связывается главным образом со спиралью αA (аминокислотные остатки от Lys 38 до Thr 53) человеческого TSLP, лишь с несколькими, но важными вкладами от спиралей αC (Lys101) и αD (Gln145, Arg149) и от петли αA-αΒ (аминокислотные остатки от Ser56 до Lys 59). Структурное наложение человеческого TSLP в комплексе с Fab1 против TSLP на опубликованный комплекс мышиного TSLP с внеклеточными доменами IL-7RA и TSLPR показывает, что Fab1 против TSLP конкурирует как с IL-7RA, так и с TSLPR за связывание с TSLP. TSLP, связанный с Fab1 против TSLP, не может связываться с TSLPR, и рекрутинг рецептора IL7Rα также ингибируется вследствие обширного стерического затруднения между Fab и рецептором IL-7α.
Пример 10: Процесс распылительной сушки и композиция Fab1 против TSLP
Оборудование и операции распылительной сушки
[00409] Для распылительной сушки сырья используют созданную на заказ распылительную сушилку. Конфигурация распылительной сушилки содержит распылитель с одноканальным соплом и двумя жидкостями, сушильную камеру, циклон, переходник, запорный клапан и 1-литровый коллектор в терморегулируемой рубашке. В описанных в данном документе вариантах осуществления процесс распылительной сушки может включать в себя процесс распыления, процесс сушки и процесс сбора частиц.
[00410] Иллюстративный процесс распыления может включать в себя следующие стадии: (A1) состав жидкого исходного сырья можно подавать через перистальтический насос (Watson Marlow) при контролируемой скорости потока в пневматический распылитель с одноканальным соплом, установленный в распылительной сушилке; (A2) сжатый сухой воздух с регулируемой скоростью потока подают в концентрическое сходящееся газовое сопло; и (A3) расширение воздуха в наконечнике сопла распыляет поток исходного сырья в мелкодисперсный аэрозоль.
[00411] Процесс сушки может включать следующие стадии: (В1) сушильный воздух, нагретый электрическим нагревателем, подают в сушильную камеру с заданной температурой и контролируемой скоростью потока; (B2) горячий осушающий воздух взаимодействует с мелкодисперсным аэрозолем со стадии А3. Растворитель (вода) в капельках испаряется, что приводит к образованию твердых частиц; и (В3) частицы и пар растворителя/воздух выходят из сушильной камеры при заранее определенной температуре.
[00412] Процесс сбора частиц может включать в себя следующие стадии: (C1) частицы и отличный от растворителя пар/воздух со стадии B3 поступают в циклон при высокой тангенциальной скорости; (С2) частицы отделяют от воздушной смеси под действием центробежной силы и собирают на дне циклона в резервуаре для сбора с контролем температуры; и (C3) отработанный отличный от растворителя пар/воздух проходит через фильтр и сбрасывается в атмосферу внутри изолятора.
Процесс получения и композиция вдыхаемых порошков Fab1 против TSLP
[00413] Данная часть представляет композицию и процесс распылительной сушки, используемые для получения подходящих для вдыхания порошков, содержащих частицы Fab1 против TSLP, композиции которых содержат разные вспомогательные вещества. Этот процесс включает распылительную сушку однофазного, водного сырья, содержащего белок (Fab1 против TSLP) и вспомогательные вещества, которые в основном функционируют либо в качестве средств, повышающих диспергируемость (таких как трилейцин), либо в качестве стеклообразующих средств (таких как сахариды, буферные соли). рH сырья контролируют с помощью буфера гистидин-HCl при целевом рН 5,0-рН 5,5. С помощью применения инженерных принципов обработки частиц выбирают вспомогательные вещества и состав для получения порошка, содержащего шероховатые частицы, каждая из которых имеет ядро из белка, стабилизированного в стеклообразной матрице, окруженное оболочкой из гидрофобного вспомогательного вещества, которая улучшает диспергируемость порошка и защищает активное средство.
Таблица 5. Композиции Fab1 против TSLP
[00414] На фиг. 12 показаны композиции с более высоким отношением вспомогательное вещество:белок, характеризующиеся улучшенной физико-химической стабильностью Fab1 против TSLP и сниженной скоростью агрегации Fab1 против TSLP.
Композиция PulmoSol, содержащая Fab1 против TSLP
[00415] Данная часть предоставляет композиции исходного сырья, полученные с использованием разных общих концентраций твердых веществ и разного содержания трилейцина, для увеличения пропускной способности производства и оптимизации образования оболочки. Высокая концентрация твердых веществ увеличивает производительность при получении порошка. В этом примере содержимое TSLP Fab1 фиксируют на уровне 50%, за одним исключением, по сравнению с предыдущим примером.
ТАБЛИЦА 6. Композиции, содержащие Fab1 против TSLP и трилейцин
жание твердых веществ
средство
Композиции, содержащие Fab1 против TSLP и 15% трилейцина
[00416] В данной части представлены несколько композиций, полученные с учетом ограниченной растворимости в воде трилейцина, при сохранении приемлемого рН исходного сырья и полученных частиц. В этом случае трилейцин растворяют в водном HCl и подвергают обратному титрованию с использованием разных основных сред для достижения целевого значения рН исходного раствора (рН 5,0-5,5). Приблизительное молярное отношение HCl к трилейцину 1:1 необходимо для полного растворения трилейцина.
ТАБЛИЦА 7. Композиции, содержащие Fab1 против TSLP и 15% трилейцина
масс/масс
Пример 11: Сродство, с которым Fab1 против TLSP связывается с белками TSLP человека и яванского макака, определяемое путем титрования до конечной точки (SET)
[00417] Измерения методом титрования до конечной точки (SET) проводят, чтобы определить сродство, с которым Fab1 против TLSP связывается с белками TSLP человека и яванского макака. Fab инкубируют при постоянной концентрации с серийными разведениями соответствующих антигенов. Сродство связывания определяют из конкурентной кривой, которую получают путем нанесения на график концентрации несвязанного антитела против концентрации используемого антигена. Fab1 против TSLP связывается со всеми человеческими белками TSLP и белками TSLP яванского макака со сродством в низком пикомолярном (пМ) диапазоне.
Процедура анализа
[00418] Двадцать два серийных разведения 1,6n человеческого TSLP (продуцированного в клетках HEK) и TSLP-антигенов яванского макака готовят в буфере для образцов и добавляют Fab1 в постоянной концентрации. Каждую смесь антиген-Fab в объеме 60 мкл/лунку распределяют с двойными повторами в 384-луночный полипропиленовый микротитрационный планшет (MTP). Буфер для образцов используют в качестве отрицательного контроля, а образец, содержащий только Fab1, в качестве положительного контроля (Bmax). Планшет закрывают и инкубируют в течение ночи (o/n, по меньшей мере, в течение 16 часов) при комнатной температуре (КТ) на шейкере.
[00419] 384-луночный MSD-массив MTP покрывают o/n при 4°C 30 мкл/лунку hsTSLP (продуцированного в E. coli), разведенного PBS до 5 мкг/мл, затем промывают три раза 70 мкл/лунку буфера для промывания и блокируют 50 мкл/лунку блокирующего буфера в течение 1 ч при КТ на шейкере. После промывания уравновешенную смесь антиген-Fab в объеме 30 мкл/лунку переносят из полипропиленового MTP в покрытый планшет MSD и инкубируют в течение 20 минут при КТ.
[00420] После дополнительной стадии промывания в каждую лунку добавляют 30 мкл сульфо-меченого детекционного антитела, разведенного буфером для образцов до концентрации 1,8 мкг/мл, и инкубируют в течение 30 мин при комнатной температуре на шейкере. Планшет MSD промывают и добавляют 35 мкл/лунку 1× MSD-считывающего буфера и инкубируют в течение 5 мин при КТ. Сигналы ECL генерируют и измеряют с помощью устройства для формирования изображения MSD Sector Imager 6000.
[00421] Три независимых эксперимента проводят для каждого антигена и демонстрируют стабильные условия анализа. Из результатов указанных экспериментов рассчитывают средние значения равновесных констант диссоциации KD и стандартных отклонений, приведенные ниже.
ТАБЛИЦА 8. Константы сродства (KD), характеризующие связывание Fab1 с белками TSLP человека и яванского макака
Пример 12: Двухнедельное исследование эффекта введения диапазона доз путем ингаляции на яванских макаках
[00422] Целью данного, не связанного с GLP исследования является определение потенциальной токсичности Fab1, Fab против тимусного стромального лимфопоэтина (TSLP), где введение яванским макакам осуществляют путем ингаляции один раз в день в течение 14 дней, или путем в.в. инъекции однократной дозы на 1 день с последующим 13-дневным периодом отсутствия введения и введением одной дозы путем ингаляции на 15 день. Кроме того, исследуют фармакокинетический/фармакодинамический (PK/PD) профиль и иммуногенность (IG) Fab1.
[00423] Исследование проводят в виде двух отдельных частей одного исследования.
[00424] Только ингаляция: Порошок Fab1 PulmoSol, 39,7% Fab1 в PulmoSol, вводят путем ингаляции 3 группам (3 самца/группу) яванских макак в целевых суточных дозах 1,0, 10,0 и 20,0 мг Fab1/кг/день. Другая группа обезьян (2 самца) получает порошок плацебо PulmoSol и служит контролем. Другой самец получает только воздух и используеся в качестве воздушного контроля. Аэрозоли Fab1 PulmoSol и плацебо PulmoSol получают с помощью вращающегося щеточного генератора (RBG1000). Животных подвергают воздействию аэрозоля порошка Fab1 PulmoSol (группы 3-5) или порошка плацебо PulmoSol (группа 2) в течение 60 минут один раз в день в течение 14 дней с использованием плотно прилегающей маски, охватывающей нос и рот. Одного самца из группы 1 подвергают воздействию только фильтрованного сухого воздуха в течение такого же периода с использованием таких же настроек оборудования. Общие вводимые средние концентрации аэрозоля Fab1 составляют 0,036, 0,31 и 0,66 мг/л для групп 3, 4 и 5 соответственно. Общие средние значения (оцененная сумма) доставляемых доз составляют 1,1, 9,6 и 19,9 мг Fab1/кг/день для групп 3, 4 и 5 соответственно. Массовый медианный аэродинамический диаметр (MMAD) подтверждает, что полученные аэрозоли Fab1 подходят для вдыхания обезьянами и что можно достичь приемлемого осаждения в легких испытуемых видов.
[00425] Внутривенное введение/ингаляция: Fab1 вводят группе из 3 самцов (группа 6) в виде однократной внутривенной болюсной инъекции через подкожную вену на 1 день. Целевая доза составляет 1 мг Fab1/кг. Затем животных оставляют на 13 дней без введения и на 15 день путем ингаляции вводят порошок Fab1 PulmoSol, 39,7% Fabl в PulmoSol. Целевая доза составляет 20 мг Fab1/кг. Аэрозоль Fab1 PulmoSol получают с помощью вращающегося щеточного генератора (RBG1000). Животных подвергают однократному воздействию аэрозоля порошка Fab1 PulmoSol в течение 60 минут с использованием плотно прилегающей маски, охватывающей нос и рот. Затем животных держат в течение 6 дней и на 21 день их подвергают эвтаназии. Общая вводимая средняя концентрация Fab1 в аэрозоле составляет 0,60 мг/л. Общая средняя оцененная суммарная доставляемая доза составляет 16,3 мг Fab1/кг/день. Массовый медианный аэродинамический диаметр (MMAD) подтверждает, что полученный аэрозоль Fab1 подходят для вдыхания обезьянами, и что можно достичь приемлемого осаждения в легких испытуемых видов.
[00426] В данном исследовании оценивают следующие параметры и результаты: клинические симптомы, массу тела, изменение массы тела, параметры клинической патологии (гематология, коагуляция и клиническая химия), биоанализ на концентрации Fab1 и TSLP и токсикокинетические параметры (сыворотка, жидкость бронхоальвеолярного лаважа (BAL), экстракты легочной ткани), антитела против Fab1 (сыворотка и жидкость BAL), результаты макроскопического вскрытия, массы органов и гистопатологические исследования (только группы 1-5).
[00427] Введение Fab1 в течение 14 дней путем ингаляций дозирование приводит к явным изменениям в полости носа (увеличение клеток слизистой оболочки в респираторном эпителии), легких (диффузное накопление альвеолярных макрофагов, увеличение насыщенности лимфатическими клетками бронхиол и альвеол и смешанная альвеолярная инфильтрация воспалительных клеток) и бронхиальном лимфатическом узле (повышенная общая клеточная насыщенность) у самцов яванских макак. Указанные изменения наблюдаются среди животных всех экспериментальных групп. Во всех случаях эффекты имеют тяжесть от минимальной до легкой и наблюдаемые симптомы не считаются неблагоприятными.
[00428] В целом результаты измерения концентрации в сыворотке, бронхоальвеолярной жидкости (BAL) и легочных экстрактах свидетельствуют о том, что Fab1 присутствует у всех животных, получающих Fab1; однако Fab1 не обнаружен ни в одном из образцов, полученных из контрольных животных, получающих воздух или плацебо PulmoSol. Рассчитанная биодоступность составляет примерно 0,2% после введения дозы путем ингаляции на 15 день. Антитела против Fab1 в сыворотке обнаружены только у одного животного из 4 группы после предварительного введения дозы на 1 день и у одного животного из 5 группы на 14-й день, однако считается, что наблюдаемые сигналы не оказывают явного воздействия на активность Fab1 у этих животных. В целом исследование не выявило явной иммуногенности Fab1. Общий TSLP не детектируется в сыворотке, BAL или легочной ткани в большинстве образцов, за исключением некоторых очень низких сигналов, обнаруженных в первых промывных образцах (собранных во время процедуры BAL у трех животных, обработанных Fab1).
[00429] Таким образом, введение Fab1 3 яванским макакам в виде однократной внутривенной болюсной инъекции на 1 день с последующим 13-дневным периодом отсутствия введения и одной ингаляционной дозой на 15 день не вызывает неблагоприятных эффектов. Введение Fab1 яванским макакам путем ингаляции в течение 14 дней связано с явными изменениями в носовой полости, легких и бронхиальном лимфатическом узле животных из всех экспериментальных групп. Во всех случаях эффекты имеют тяжесть от минимальной до легкой и наблюдаемые симптомы не считаются неблагоприятными. Всех животных, получающих Fab1, систематически подвергают тестированию.
Пример 13: Исследование токсичности через тринадцать недель после ингаляции у яванских макаков
[00430] Целью данного исследования является определение потенциальной токсичности Fab1, Fab против тимусного стромального лимфопоэтина (TSLP), после введения его путем ингаляции один раз в день в течение по меньшей мере 92 последовательных дней (13 недель) яванским макакам и оценки потенциальной обратимости результатов после 42-дневного (6-недельного) периода восстановления. Кроме того, определяют токсикокинетические и иммуногенные характеристики Fab1.
[00431] Порошок Fab1 PulmoSol вводят путем ингаляции 3 группам (3/пол/группа) яванских макак в целевых суточных дозах 3, 10 и 22 мг/кг/день. Другая группа обезьян (3/пол) получает в качестве плацебо порошок PulmoSol и служит контрольной группой. Других 2 животных (2/пол) в группах, получающих плацебо и 22 мг/кг/день, используют в исследовании в течение 6-недельного периода восстановления. Аэрозоли Fab1 PulmoSol и плацебо PulmoSol получают с помощью вращающегося щеточного генератора (RBG1000). Животных подвергают воздействию аэрозоля порошка Fab1 PulmoSol (группы 2-4) или порошка плацебо PulmoSol (группа 1) в течение 60 минут один раз в день в течение по меньшей мере 92 последовательных дней с использованием плотно прилегающей маски, охватывающей рот и нос.
[00432] В данном исследовании оценивают следующие параметры и результаты: клинические признаки, массу тела, изменения массы тела, офтальмологические исследования, неврологические исследования (в том числе частота дыхания), электрокардиологические исследования, параметры клинической патологии (гематология, коагуляция, клиническая химия и анализ мочи), иммунофенотипирование лимфоцитов периферической крови (проточная цитометрия), иммунная функция (опосредуемый Т-клетками гуморальный ответ (TDAR) на гемоцианин лимфы улитки (KLH)), биоанализ концентраций Fab1 и токсикокинетических параметров (в сыворотке и экстрактах легочной ткани), антитела против Fab1 (в сыворотке), результаты макроскопического вскрытия, массы органов и гистопатологические анализы.
[00433] Общие вводимые средние концентрации Fab1 в аэрозоле составляют 0,10, 0,33 и 0,72 мг/л для групп 2, 3 и 4 соответственно. Оцененные общие средние дозы, достигаемые при полной доставке (смешанные полы), составляют 3,0, 10,1 и 22,2 мг/кг/день Fab1 для групп 2, 3 и 4 соответственно. Средний массовый медианный аэродинамический диаметр (MMAD) подтверждает, что полученные аэрозоли Fab1 подходят для вдыхания обезьянами, и что достигается приемлемое воздействие на легкие испытуемых видов.
[00434] После введения Fab1 в достигаемых дозах 3, 10 или 22 мг/кг/день отсутствуют явные эффекты, связанные с тестируемым веществом и определяемые по каким-либо из сердечно-сосудистых параметров.
[00435] Результаты исследований иммунной функции (TDAR) демонстрируют тенденцию к снижению уровней антител IgG против KLH при достигнутых дозах 3, 10 или 22 мг/кг/день. Во время последнего анализа (78 день исследования) отмечается снижение уровней антител IgG и IgM против KLH при достигнутых дозах≥3 мг/кг/день. Этот эффект наиболее выражен у самцов во все моменты времени и у самок на 78 день, но менее заметен у самок в другие дни. Тенденция к уменьшению ответа против KLH не считается неблагоприятной. Уровни антител IgG и IgM против KLH у восстанавливающихся животных 4 группы (22,2 мг/кг/день) были выше во всех случаях по сравнению с значениями восстановительного периода параллельного контроля (группа 1) и подобны основным контрольным результатам исследованиям (группа 1), свидетельствуя о восстановлении пониженного гуморального ответа против KLH. Введение Fab1 путем ингаляций в течение по меньшей мере 92 дней приводит к увеличению насыщенности легких клетками лимфоидной ткани при достигнутых дозах 3 мг/кг/день. Обнаруженные эффекты во всех случаях имеют тяжесть от минимальной до легкой и не считаются неблагоприятными. После 6-недельного периода восстановления эффекты отсутствуют у 2/4 (50%) восстанавливающихся животных и наблюдаются с минимальной степенью тяжести у 2/4 (50%) животных, получающих высокую дозу. Такой же эффект также присутствует у одной контрольной восстановливающейся самки (слабый), подтверждая, что такое изменение может иногда случайно встречаться среди контрольных животных. Fab1 не обнаружен ни в одном из образцов сыворотки или легочной ткани животных из контрольной группы, получающих плацебо PulmoSol (группа 1). В целом на протяжении всего периода дозирования у животных, получавших Fab1, наблюдается дозо-зависимое присутствие Fab1 как в сыворотке, так и в ткани легкого. Системное присутствие в сыворотке также обнаруживается во время периода восстановления до 14-28 дней после введения последней дозы на 92 день, тогда как в легочной ткани концентрации не обнаруживаются у всех восстанавливающихся животных до момента вскрытия на 135 день (42 дня после последнего введения дозы). Значительное накопление в сыворотке Fab1 наблюдается с 1 дня исследования до 91 дня исследования после многократного введения суточных доз во всех группах, получающих дозы. Очевидные связанные с полом различия в накоплении отсутствуют. Низкие, но детектируемые сигналы антител против лекарственного средства (ADA) наблюдаются у двух контрольных животных, получавших плацебо PulmoSol, как в исходный момент до введения дозы, так и в моменты времени после введения дозы, вероятно, вследствие существования антител, не специфичных к Fab1. Сигналы ADA не обнаружены ни у каких других контрольных животных. У всех Fab1-обработанных животных развиваются сигналы ADA после введения доз уже после 28 дня исследования. Сильные сигналы ADA у трех животных, по-видимому, связаны с отсутствием Fab1 в сыворотке, но при этом во время вскрытия (через 1-6 часов после введения последней дозы) у этих животных все еще обнаруживается присутствие Fab1 в легких. Можно сделать вывод, что введение Fab1 яванским макакам путем ингаляции в течение 13 недель хорошо переносится при достигаемых при вдыхании уровнях доз до 22,2 мг/кг/день. Показано, что все животные, получающие Fab1, подвергаются воздействию Fab1 как системно, так и в легких. Антитела против Fab1 присутствуют у всех обработанных животных начиная уже с 28 дня и ассоциируются с гораздо меньшим воздействием у 3 особей по сравнению с остальными обработанными животными. Микроскопический анализ подтверждает увеличение насыщенности легких клетками лимфоидной ткани у большинства животных, получающих Fab1, которое наблюдается у 50% животных, восстанавливающихся после высоких доз после 6-недельного периода восстановления. Обнаруженные эффекты во всех случаях имеют тяжесть от минимальной до легкой и не считаются неблагоприятными.
Пример 14: Получение простых высушенных распылением композиций фрагмента моноклонального антитела
[00436] Описанный здесь фрагмент Fab1 моноклонального антитела имеет молекулярную массу 46,6 кДа. Описано применение сухих порошкообразных составов для местной доставки в легкие при лечении астмы. В данном контексте термин "простая" относится к композициям, содержащим только активный фармацевтический ингредиент (Fab1) и буфер.
[00437] Ряд простых композиций антител, содержащих 89,5% активного фармацевтического ингредиента и 10,5% гистидинового буфера, получают из исходного сырья, содержащего разные смеси этанола и воды (таблица 9). Содержание этанола варьирует от 5 до 20% масс./масс. Исходное сырье подвергают распылительной сушке на распылительной сушилке NSD с температурой на входе 105°С, температурой на выходе 70°С, скоростью потока сушильного газа 595 л/мин, скоростью потока распылителя 20 л/мин, скоростью подачи жидкости 8,0 мл/мин и ALR 2,5×103 об./об. Содержание твердых веществ фиксируют при 2% масс./об.
ТАБЛИЦА 9. Влияние параметров процесса на микромеритные свойства простых композиций антител, содержащих 89,5% API в гистидиновом буфере.
(% масс/масс)
масс)
(% масс/масс)
Пример 15: Микромеритные свойства простых высушенных распылением композиций антител
[00438] Микромеритные свойства высушенных распылением композиций антител, описанных в примере 14, приведены в таблице 9. Все простые композиции, содержащие только API и буфер, имеют частицы с гладкой поверхностью (например, с отсутствием морщинистости поверхности). Добавление небольших количеств этанола к водному исходному раствору уменьшает объем и насыпную плотность порошков (как и при получении композиций инсулина). Частицы также являются относительно крупными с точки зрения их распределения по размерам первичных частиц (PPSD).
Пример 16: Аэрозольные характеристики простых высушенных распылением композиций антител
[00439] DD и TLD, определенные для порошков, описанных в примере 15, приведены в таблице 10. Первичные частицы имеют рассчитанный медианный аэродинамический диаметр, Da, от 0,71 до 0,93 мкм (рассчитанный исходя из насыпной плотности и измерений ×50 по уравнению: 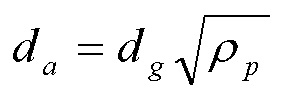 )
)
[00440] Ингалятор сухого порошка Concept1 представляет собой устройство на основе капсул с низкой устойчивостью (R=0,07 см H2O)1/2/(л/мин)).
ТАБЛИЦА 10. Аэрозольные характеристики простых композиций антител. Аэрозольные характеристики оценивают с помощью ингалятора Concept1 (20 мг заполняющей массы) при скорости потока 90 л/мин и общем объеме 2 л (n=5).
(% ND)
(% DD)
[00441] Из приведенных в таблице 10 данных видно, что одного уменьшения плотности недостаточно для получения частиц, которые позволяют эффективно избежать осаждения во рту и в горле. Для этого нужно модифицировать морфологию частиц с увеличением поверхностной шероховатости (морщинистости) и желательно уменьшить размер первичных частиц.
[00442] Интересно отметить, что, хотя пептиды и небольшие белки в природе имеют морщинистую морфологию в отсутствии оболочкообразующего вспомогательного вещества, композиция антитела требует добавления оболочкообразующего вспомогательного вещества, чтобы обеспечить образование морщинистых частиц. В этом отношении добавление оболочкообразующего вспомогательного вещества и добавление этанола выполняют аналогичные функции, изменяя толщину стенки и плотность высушенных распылением частиц. Следовательно, влияние добавления этанола меньше в присутствии оболочкообразующего вещества.
Пример 17: Получение и микромеритные свойства высушенных распылением платформенных составов антител
[00443] В данной серии высушенных распылением порошков условия распылительной сушки поддерживают постоянными и оценивают влияние добавления оболочкообразующего вспомогательного вещества (то есть трилейцина, 0-15% масс./масс.) на составы антител. Указанные составы также содержат трегалозу в качестве стеклообразующего средства (примерно 29-44% масс./масс. в зависимости от содержания трилейцина) и гистидиновый буфер (5,9% масс./масс., РН 5,0).
[00444] Порошки сушат распылением с помощью изготовленной на заказ распылительной сушилки NSD с температурой на входе 105°С, температурой на выходе 70°С, скоростью потока сушильного газа 595 л/мин, скоростью потока распылителя 25 л/мин, скоростью подачи жидкости 10,0 мл/мин и ALR 2,5×103 об./об. Содержание твердых веществ поддерживали постоянным при 2% масс./масс. Все порошки имеют морщинистую морфологию, за исключением партии 761-02-12, которую сушат распылением в отсутствии оболочкообразующего вещества и получают гладкие частицы, подобные, полученным в примере 16. Результаты показаны в таблице 11. В некоторых вариантах осуществления сухой порошкообразный состав настоящего изобретения содержит частицы с ядром и оболочкой, содержащие: оболочкообразующее вспомогательное вещество и ядро, содержащее API, стеклообразующие вспомогательные вещества и буфер, иногда такой состав также упоминается здесь как платформенный состав.
ТАБЛИЦА 11. Влияние параметров процесса на микромеритные свойства "платформенных" составов антител, содержащих 50,0% масс./масс. API, 5,9% гистидинового буфера, трегалозы и трилейцина.
(% масс/масс)
(% масс/масс)
Пример 18: Аэрозольные характеристики "платформенных" высушенных распылением составов антител с разным содержанием трилейцина
[00445] DD и TLD, определенные для порошков, описанных в примере 17, приведены в таблице 12.
ТАБЛИЦА 12. Влияние параметров процесса на микромеритные свойства и аэрозольные характеристики платформенных составов антител. Аэрозольные характеристики оценивают с помощью ингалятора Concept1 (20 мг заполняющей массы) при скорости потока 90 л/мин и общем объеме 2 л (n=5).
(% ND)
(% DD)
[00446] Значительные улучшения DD и TLD наблюдаются в композициях антител с морщинистой морфологией частиц. В вариантах осуществления изобретения желательную морщинистую морфологию достигают благодаря присутствию на поверхности частиц оболочкообразующего вспомогательного вещества трилейцина.
[00447] В некоторых вариантах осуществления изобретения физико-химические свойства вещества, присутствующего на поверхности частиц, влияют на морфологию частиц. В случае больших белков (таких как белки с массой более 20000 Дальтон) для достижения желательной морфологии в качестве оболочкообразующего вспомогательного вещества предпочтительно используют трилейцин. В вариантах осуществления изобретения частицы, образующие композицию, имеют морщинистую морфологию, способствующую уменьшению сил сцепления между частицами, вследствие чего размер агломератов является достаточно маленьким, чтобы агломераты можно было вдыхать.
[00448] При добавлении этанола снижается плотность (в противном случае) морщинистых частиц в результате уменьшения толщины стенки. Это, в свою очередь, способствует уменьшению насыпной плотности, приводя более мелкие первичные частицы в соответствие с желательными аэродинамическими свойствами. В некоторых вариантах осуществления частицы должны иметь пониженную плотность, чтобы первичные частицы и агломераты были пригодными для вдыхания.
[00449] Значительное уменьшение насыпной плотности наблюдается в парных композициях 728-06-04 и 761-02-11, а также 728-06-02 и 761-02-10, когда содержание этанола увеличивается от 0% до 10% масс./масс. В случае конкретных композиций, используемых в данном примере, добавление только 10% этанола не приводит к целевому улучшению аэрозольных характеристик, обеспечиваемому оболочкообразующим вспомогательным веществом. TLD является превосходным (> 80% от DD), но остается ниже желательного значения 90 масс.% DD, в значительной степени вследствие того, что частицы слишком большие и плотные. Расчетный первичный аэродинамический диаметр Da морщинистых частиц составляет от 0,77 до 1,38 мкм.
Пример 19: Влияние модифицированных параметров процесса (содержание твердых веществ и добавление сорастворителя) на микромеритные свойства платформенных составов антител
[00450] Композиции содержат 50,0% масс./масс. API, 5,9% масс./масс. гистидинового буфера (рН 5,0), примерно 14 масс.% или 29 масс.% трегалозы и 15 масс.% или 30 масс.% трилейцина. Порошки сушат распылением с помощью изготовленной на заказ распылительной сушилки NSD с температурой на входе 105°С, температурой на выходе 70°С, скоростью потока сушильного газа 595 л/мин, скоростью потока распылителя 30 л/мин, скоростью подачи жидкости 4,0 мл/мин и ALR 7,5×103 об./об. Содержание твердых веществ уменьшают до 1% масс./масс. Указанные модификации в процессе распылительной сушки осуществляют с целью уменьшения размера первичных частиц. В вариантах осуществления изобретения наблюдается значительное уменьшение распределения первичных частиц по размерам.
ТАБЛИЦА 13. Влияние параметров процесса на микромеритные свойства "платформенных" составов антител, содержащих 50,0% масс./масс. API, 5,9% гистидинового буфера, трегалозы и трилейцина.
(% масс/масс)
масс)
(% масс/
масс)
Пример 20: Влияние модифицированных параметров процесса (содержание твердых веществ и добавление сорастворителя) на аэрозольные характеристики платформенных составов антител
[00451] Влияние уменьшения содержания твердых веществ и увеличения ALR на аэрозольные характеристики платформенных составов антител показано в таблице 14. Наблюдается значительное уменьшение среднего аэродинамического диаметра первичных частиц по сравнению с частицами, полученными по способу примера 18. Это соответствует TLD в диапазоне примерно от 94% до 98% DD, то есть в пределах желательного, предпочтительного или оптимального целевого диапазона характеристики.
ТАБЛИЦА 14. Влияние параметров способа на микромеритные свойства и аэрозольные характеристики платформенных составов антител. Аэрозольные характеристики оценивают с помощью ингалятора Concept1 (20 мг заполняющей массы) при скорости потока 90 л/мин и общем объеме 2 л (n=5).
твердые вещества
(% ND)
(% DD)
[00452] Если не указано иное, используемые здесь технические и научные термины имеют традиционные значения, известные специалистам в области, к которой относится настоящее изобретение.
[00453] Если не указано иное, все способы, стадии, методы и манипуляции, которые конкретно не описаны подробно, можно проводить известными способами, очевидными для специалистов в данной области. Опять же, можно, например, сослаться на стандартные справочники и описанный здесь общий уровнь техники, а также на приведенные в нем другие ссылки.
[00454] Если не указано иное, каждая из приведенных здесь ссылок включена полностью в качестве ссылки.
[00455] Формула изобретения является неограничивающей и приведена ниже.
[00456] Подробное описание конкретных аспектов и пунктов приводится здесь в качестве примера только в целях иллюстрации и не предназначается для ограничения объема прилагаемой формулы изобретения или объема предмета формулы любой соответствующей будущей заявки. В частности, авторы изобретения полагают, что в описание можно внести разные замены, изменения и модификации без отступления от сущности и объема изобретения, определяемых формулой изобретения. Предполагается, что рядовой специалист в данной области может осуществить выбор исходной нуклеиновой кислоты, представляющего интерес клона или типа библиотеки на основе описанных здесь аспектов. Другие аспекты, преимущества и модификации входят в объем приведенной ниже формулы изобретения. В результате не более чем рутинного экспериментирования для специалистов в данной области техники станут очевидны многие эквиваленты описанных здесь конкретных аспектов изобретения. Такие эквиваленты также охватываются нижеследующей формулой изобретения. Пересмотр объема формулы изобретения в последующих поданных соответствующих заявках может быть обусловлен ограничениями патентного законодательства разных стран и не должен толковаться как отказ от предмета формулы изобретения.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> NOVARTIS AG
<120> МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕСЯ С ТИМУСНЫМ СТРОМАЛЬНЫМ ЛИМФОПОЭТИНОМ (TSLP), И СПОСОБЫ ПРИМЕНЕНИЯ ТАКИХ МОЛЕКУЛ
<130> PAT057035-WO-PCT
<140>
<141>
<150> 62/342,511
<151> 2016-05-27
<150> 62/216,050
<151> 2015-09-09
<160> 43
<170> PatentIn version 3.5
<210> 1
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
пептид"
<400> 1
Gly Phe Thr Phe Ser Asp Tyr Trp Met His
1 5 10
<210> 2
<211> 19
<212> Белок
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
пептид"
<400> 2
His Ile Lys Ser Lys Thr Asp Ala Gly Thr Thr Asp Tyr Ala Ala Pro
1 5 10 15
Val Lys Gly
<210> 3
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
пептид"
<400> 3
Glu Ile Tyr Tyr Tyr Ala Phe Asp Ser
1 5
<210> 4
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
пептид"
<400> 4
Asp Tyr Trp Met His
1 5
<210> 5
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
пептид"
<400> 5
Gly Phe Thr Phe Ser Asp Tyr
1 5
<210> 6
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
пептид"
<400> 6
Lys Ser Lys Thr Asp Ala Gly Thr
1 5
<210> 7
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
полипептид"
<400> 7
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly His Ile Lys Ser Lys Thr Asp Ala Gly Thr Thr Asp Tyr Ala Ala
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Ile Tyr Tyr Tyr Ala Phe Asp Ser Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 8
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
полинуклеотид"
<400> 8
gaggttcagc tggtggaatc aggcggcgga ctggttaagc ctggcggмаркер cctмаркерactt 60
agctgcgctg cмаркерtggctt caccttмаркерc gactactgga tgcactgggt маркерacaggcc 120
cctggtaaag gcttggagtg ggtcggacac attaagtcta agaccgacgc cggcactacc 180
gactacgccg ctcccgttaa gggccggttc actatctcta gggacgactc taagaacacc 240
ctctaccttc aaatgaaмаркер ccttaagacc gaggacaccg ccgtctacta ctgcgcмаркерa 300
gaaatctact actacgcctt cgaмаркерctgg ggtcaaggca ccctcgtgac cgtgtcмаркерc 360
<210> 9
<211> 450
<212> Белок
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
полипептид"
<400> 9
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly His Ile Lys Ser Lys Thr Asp Ala Gly Thr Thr Asp Tyr Ala Ala
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Ile Tyr Tyr Tyr Ala Phe Asp Ser Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 10
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
полинуклеотид"
<400> 10
gaggttcagc tggtggaatc aggcggcgga ctggttaagc ctggcggмаркер cctмаркерactt 60
agctgcgctg cмаркерtggctt caccttмаркерc gactactgga tgcactgggt маркерacaggcc 120
cctggtaaag gcttggagtg ggtcggacac attaagtcta agaccgacgc cggcactacc 180
gactacgccg ctcccgttaa gggccggttc actatctcta gggacgactc taagaacacc 240
ctctaccttc aaatgaaмаркер ccttaagacc gaggacaccg ccgtctacta ctgcgcмаркерa 300
gaaatctact actacgcctt cgaмаркерctgg ggtcaaggca ccctcgtgac cgtgtcмаркерc 360
gcмаркерcacta agggcccaag tgtgtttccc ctggccccca gcagcaagtc tacttccggc 420
ggaactgctg ccctgggttg cctggtgaag gactacttcc ccgagcccgt gacagtgtcc 480
tggaactctg gggctctgac ttccggcgtg cacaccttcc ccgccgtgct gcagagcagc 540
ggcctgtaca gcctgagcag cgtggtgaca gtgccctcca gctctctggg aacccagacc 600
tatatctgca acgtgaacca caagcccagc aacaccaagg tggacaagag agtggagccc 660
aagagctgcg acaagaccca cacctgcccc ccctgcccag ctccagaact gctgggaggg 720
ccttccgtgt tcctgttccc ccccaagccc aaggacaccc tgatgatcag caggaccccc 780
gaggtgacct gcgtggtggt ggacgtgtcc cacgaggacc cagaggtgaa gttcaactgg 840
tacgtggacg gcgtggaggt gcacaacgcc aagaccaagc ccagagagga gcagtacaac 900
agcacctaca gggtggtgtc cgtgctgacc gtgctgcacc aggactggct gaacggcaaa 960
gaatacaagt gcaaagtctc caacaaggcc ctgccagccc caatcgaaaa gacaatcagc 1020
aaggccaagg gccagccacg ggagccccag gtgtacaccc tgccccccag ccgggaggag 1080
atgaccaaga accaggtgtc cctgacctgt ctggtgaagg gcttctaccc cagcgatatc 1140
gccgtggagt gggagagcaa cggccagccc gagaacaact acaagaccac ccccccagtg 1200
ctggacagcg acggcagctt cttcctgtac agcaagctga ccgtggacaa gtccaggtgg 1260
cagcagggca acgtgttcag ctgcagcgtg atgcacgagg ccctgcacaa ccactacacc 1320
cagaagtccc tgagcctgag ccccggcaag 1350
<210> 11
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
пептид"
<400> 11
Ser Gly Asp Asn Ile Gly Ser Lys Tyr Val His
1 5 10
<210> 12
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
пептид"
<400> 12
Gly Asp Asn Glu Arg Pro Ser
1 5
<210> 13
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
пептид"
<400> 13
Gln Ala Ala Asp Trp Val Asp Phe Tyr Val
1 5 10
<210> 14
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
пептид"
<400> 14
Asp Asn Ile Gly Ser Lys Tyr
1 5
<210> 15
<211> 3
<212> Белок
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
пептид"
<400> 15
Gly Asp Asn
1
<210> 16
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
пептид"
<400> 16
Ala Asp Trp Val Asp Phe Tyr
1 5
<210> 17
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
полипептид"
<400> 17
Ser Tyr Glu Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Ser Gly Asp Asn Ile Gly Ser Lys Tyr Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Gly Asp Asn Glu Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ala Ala Asp Trp Val Asp Phe Tyr
85 90 95
Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 18
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
полинуклеотид"
<400> 18
agctacgagc tgactcagcc cctмаркерcgtt agcgtggccc tgggtcaaac cgcмаркерaatc 60
acctgмаркерcg gcgataatat cggctctaaa tacgttcact ggtatcagca gaagcccggt 120
caagcccccg tgctcgtgat ctacggcgat aacgagcggc cмаркерcggaat ccccgagcgg 180
ttмаркерcggct ctaaмаркерcgg taacaccgct accctgacta tctcмаркерggc tcaggccggc 240
gacgaggccg actactactg tcaggccgcc gactgggtgg acttctacgt gttcggcgga 300
ggcactaagc tgaccgtgct g 321
<210> 19
<211> 213
<212> Белок
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
полипептид"
<400> 19
Ser Tyr Glu Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Ser Gly Asp Asn Ile Gly Ser Lys Tyr Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Gly Asp Asn Glu Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ala Ala Asp Trp Val Asp Phe Tyr
85 90 95
Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys Ala
100 105 110
Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala
115 120 125
Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala
130 135 140
Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val
145 150 155 160
Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser
165 170 175
Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr
180 185 190
Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala
195 200 205
Pro Thr Glu Cys Ser
210
<210> 20
<211> 639
<212> ДНК
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
полинуклеотид"
<400> 20
agctacgagc tgactcagcc cctмаркерcgtt agcgtggccc tgggtcaaac cgcмаркерaatc 60
acctgмаркерcg gcgataatat cggctctaaa tacgttcact ggtatcagca gaagcccggt 120
caagcccccg tgctcgtgat ctacggcgat aacgagcggc cмаркерcggaat ccccgagcgg 180
ttмаркерcggct ctaaмаркерcgg taacaccgct accctgacta tctcмаркерggc tcaggccggc 240
gacgaggccg actactactg tcaggccgcc gactgggtgg acttctacgt gttcggcgga 300
ggcactaagc tgaccgtgct gggtcaacct aaggctgccc ccagcgtgac cctgttcccc 360
cccagcagcg aggagctgca ggccaacaag gccaccctgg tgtgcctgat cagcgacttc 420
tacccaggcg ccgtgaccgt ggcctggaag gccgacagca gccccgtgaa ggccggcgtg 480
gagaccacca cccccagcaa gcagagcaac aacaagtacg ccgccagcag ctacctgagc 540
ctgacccccg agcagtggaa gagccacagg tcctacagct gccaggtgac ccacgagggc 600
agcaccgtgg aaaagaccgt ggccccaacc gagtgcagc 639
<210> 21
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
полинуклеотид"
<400> 21
gaggtgcagc tggtggaatc aggcggcgga ctggtcaagc ctggcggмаркер cctgagactg 60
agctgcgctg cмаркерtggctt caccttмаркерc gactactgga tgcactgggt cagacaggcc 120
cctggtaaag gcctggagtg ggtcggacac attaagtcta agaccgacgc cggcactacc 180
gactacgccg ctcctgtgaa gggccggttc actatctcta gggacgactc taagaacacc 240
ctgtacctgc agatgaaмаркер cctgaaaacc gaggacaccg ccgtctacta ctgcgcмаркерa 300
gagatctact actacgcctt cgaмаркерctgg ggtcagggca ccctggtcac cgtgtcмаркерc 360
<210> 22
<211> 223
<212> Белок
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
полипептид"
<400> 22
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly His Ile Lys Ser Lys Thr Asp Ala Gly Thr Thr Asp Tyr Ala Ala
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Ile Tyr Tyr Tyr Ala Phe Asp Ser Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Pro Lys Ser Cys
210 215 220
<210> 23
<211> 669
<212> ДНК
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
полинуклеотид"
<400> 23
gaggtgcagc tggtggaatc aggcggcgga ctggtcaagc ctggcggмаркер cctgagactg 60
agctgcgctg cмаркерtggctt caccttмаркерc gactactgga tgcactgggt cagacaggcc 120
cctggtaaag gcctggagtg ggtcggacac attaagtcta agaccgacgc cggcactacc 180
gactacgccg ctcctgtgaa gggccggttc actatctcta gggacgactc taagaacacc 240
ctgtacctgc agatgaaмаркер cctgaaaacc gaggacaccg ccgtctacta ctgcgcмаркерa 300
gagatctact actacgcctt cgaмаркерctgg ggtcagggca ccctggtcac cgtgtcмаркерc 360
gcмаркерcacta agggcccctc cgtgttccct ctggcccctt ccagcaagtc tacctctggc 420
ggcaccgctg ctctgggctg cctggtgaag gactacttcc ctgagcctgt gacagtgtcc 480
tggaactctg gcgccctgac ctccggcgtg cacaccttcc ctgccgtgct gcagtcctcc 540
ggcctgtact ccctgtcctc cgtggtgaca gtgccttcct ccagcctggg cacccagacc 600
tatatctgca acgtgaacca caagccttcc aacaccaagg tggacaagcg ggtggagcct 660
aagtcatgc 669
<210> 24
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
полинуклеотид"
<400> 24
agctacgagc tgactcagcc cctgagcgtc agcgtggccc tgggtcagac cgcмаркерaatc 60
acctgмаркерcg gcgataatat cggctctaaa tacgtgcact ggtatcagca gaagcccggt 120
caggcccccg tgctggtgat ctacggcgat aacgagcggc cмаркерcggaat ccccgagcgg 180
ttмаркерcggct ctaaмаркерcgg taacaccgct accctgacta tctcмаркерggc tcaggccggc 240
gacgaggccg actactactg tcaggccgcc gactgggtgg acttctacgt gttcggcgga 300
ggcactaagc tgaccgtgct g 321
<210> 25
<211> 213
<212> Белок
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
полипептид"
<400> 25
Ser Tyr Glu Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Ser Gly Asp Asn Ile Gly Ser Lys Tyr Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Gly Asp Asn Glu Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ala Ala Asp Trp Val Asp Phe Tyr
85 90 95
Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln Pro Lys Ala
100 105 110
Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala
115 120 125
Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala
130 135 140
Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val
145 150 155 160
Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser
165 170 175
Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Arg Ser Tyr
180 185 190
Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala
195 200 205
Pro Thr Glu Cys Ser
210
<210> 26
<211> 639
<212> ДНК
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
полинуклеотид"
<400> 26
agctacgagc tgactcagcc cctgagcgtc agcgtggccc tgggtcagac cgcмаркерaatc 60
acctgмаркерcg gcgataatat cggctctaaa tacgtgcact ggtatcagca gaagcccggt 120
caggcccccg tgctggtgat ctacggcgat aacgagcggc cмаркерcggaat ccccgagcgg 180
ttмаркерcggct ctaaмаркерcgg taacaccgct accctgacta tctcмаркерggc tcaggccggc 240
gacgaggccg actactactg tcaggccgcc gactgggtgg acttctacgt gttcggcgga 300
ggcactaagc tgaccgtgct gggtcagcct aaggctgccc ccagcgtgac cctgttcccc 360
cccagcagcg aggagctgca ggccaacaag gccaccctgg tgtgcctgat cagcgacttc 420
tacccaggcg ccgtgaccgt ggcctggaag gccgacagca gccccgtgaa ggccggcgtg 480
gagaccacca cccccagcaa gcagagcaac aacaagtacg ccgccagcag ctacctgagc 540
ctgacccccg agcagtggaa gagccacagg tcctacagct gccaggtgac ccacgagggc 600
agcaccgtgg aaaagaccgt ggccccaacc gagtgcagc 639
<210> 27
<211> 159
<212> Белок
<213> Homo sapiens
<400> 27
Met Phe Pro Phe Ala Leu Leu Tyr Val Leu Ser Val Ser Phe Arg Lys
1 5 10 15
Ile Phe Ile Leu Gln Leu Val Gly Leu Val Leu Thr Tyr Asp Phe Thr
20 25 30
Asn Cys Asp Phe Glu Lys Ile Lys Ala Ala Tyr Leu Ser Thr Ile Ser
35 40 45
Lys Asp Leu Ile Thr Tyr Met Ser Gly Thr Lys Ser Thr Glu Phe Asn
50 55 60
Asn Thr Val Ser Cys Ser Asn Arg Pro His Cys Leu Thr Glu Ile Gln
65 70 75 80
Ser Leu Thr Phe Asn Pro Thr Ala Gly Cys Ala Ser Leu Ala Lys Glu
85 90 95
Met Phe Ala Met Lys Thr Lys Ala Ala Leu Ala Ile Trp Cys Pro Gly
100 105 110
Tyr Ser Glu Thr Gln Ile Asn Ala Thr Gln Ala Met Lys Lys Arg Arg
115 120 125
Lys Arg Lys Val Thr Thr Asn Lys Cys Leu Glu Gln Val Ser Gln Leu
130 135 140
Gln Gly Leu Trp Arg Arg Phe Asn Arg Pro Leu Leu Lys Gln Gln
145 150 155
<210> 28
<211> 2629
<212> ДНК
<213> Homo sapiens
<400> 28
gcagccagaa agctctggag catcagggag actccaactt aaggcaacag catgggtgaa 60
taagggcttc ctgtggactg gcaatgagag gcaaaacctg gtgcttgagc actggcccct 120
aaggcaggcc ttacagatct cttacactcg tggtgggaag agttмаркерtgt gaaactgggg 180
tggaattggg tgtccacgta tgttcccttt tgccttacta tatgttctgt cagtttcttt 240
caggaaaatc ttcatcttac aacttgмаркерg gctggtgtta acttacgact tcactaactg 300
tgactttgag aagattaaag cagcctatct cagtactatt tctaaagacc tgattacata 360
tatgagtggg accaaaagta ccgagttcaa caacaccgtc tcttgмаркерca atcggccaca 420
ttgccttact gaaatccaga gcctaacctt caatcccacc gccggctgcg cgtcgctcgc 480
caaagaaatg ttcgccatga aaactaaggc tgcctмаркерct atctggtgcc caggctattc 540
ggaaactcag ataaatgcta ctcaggcaat gaagaagagg agaaaaagga aagtcacaac 600
caataaatgt ctggaacaag tgtcacaatt acaaggattg tggcgtcgct tcaatcgacc 660
tttactgaaa caacagtaaa ccatctttat tatggtcata tttcacagca ccaaaataaa 720
tcatctttat taagмаркерatg aaacattaac tctaactgtg acaaagaaga ccacaaaмаркер 780
ttatctttta attacagaag agtttcttaa cttacttttg taagttttta ttgtgtaagt 840
ttataatgca ggggaagtac tactcctcaa atgttgaggg aagcttccat aacattgatg 900
actggcttca tggcagtaat tctcggctgt agttgcataa gcattgctca agaggaaaat 960
ccaaaagtgc agcaggagaa ctcttttccc tgaaaaagga aaaatattga actcaatgat 1020
agcacctaaa cttacattta aaagacagac attccttcta catgtaatga cacttcttgt 1080
gttaaactaa aaatttacaa gagaagaaag tgaaagcaaa tggggtttca caaaмаркерttg 1140
taaataмаркерt gaagcaattt gaaataattt tcaagcaaag tattgtgaaa gtattctaag 1200
ccaagtttta aatattatct aacagacaag agtggtatat acaagмаркерat cctgagaagt 1260
acctttgtta cagctactat aaatatacat ataaattata gaatctactt taatttattt 1320
tgtgaacact tttgaaaatg tacatgttcc tttgtaattg acactatata tttcttaata 1380
aaataattct caaatttgtt tcttatgaat catctctcaa atcмаркерtмаркер acaatttgca 1440
cacatacttt tctaagggac attatcttcc ttcaggtttt tacctccact catcctмаркерa 1500
gcccactgac tgctcccctt tatacctgtt ggccctgcct aмаркерgagaga atatttggag 1560
aмаркерgcagct tcaggatgca ttgcaatcat ccttttctta aattatgtca cмаркерtctttt 1620
attttttccc ctcttgaact ttcctcacac ctggaagaaa caaagмаркерga aaaagtgaac 1680
aggggatgtc aaatcgattc ttgaattccc gctgcaagct agagccgcag gcaccctctc 1740
actcaatttc cactcagaac cctataaaca ccagtgggaa gggcaaccca ctgcacgtgg 1800
gaatgcactg atttttccta ggagмаркерaca tgttcctcta attactccct gagggtмаркерt 1860
tggggctaaa ccatgacaga agtggggaag ttcaatgtcc ttaaatccat cttacttgcc 1920
aacaggtaag aggaagctta cattacatgt ccagtccaca tttaaagagc acttactgtg 1980
gaacaagcct tcagccaaac aatggggata gaaaagмаркерg taagactcag cctttgtcca 2040
gagaagctca gggtaмаркерct gaaмаркерgcag tttcttttgt cctgaggaaa atcaggacat 2100
gcctgctttc taaaaatctt cctctgaaga cctgacccaa gctcttaaat gctattgtaa 2160
gagaaatttc tttgtctatt aactccattt маркермаркерggat tcactgacta gattttactg 2220
aactatgaaa ataaatacac ataatttttc acaaaatttt gggcccaatt cccctaaaag 2280
aattgaggat маркерggagaaa ggagacaact caaagtcatc ccattaagtg cagtttcttt 2340
gaatcttctg ctttatcttt aaaaatttgt ataatttata tattttattc tatgtgttcc 2400
aмаркерatatct taatgtaaaa tмаркерtcattt aaattacact gtcaattaaa agtaatgggc 2460
aagagattgc atcatactaa ttмаркерtaaga acgttcccaa atgttgtaac aatgtggatc 2520
atacatctct ggttttttaa atgtattgag gctttcttgg tggacмаркерta маркерtatacgg 2580
tcagttatgt caatgtttca tggtcaataa aaaggaagtt gcaaattgt 2629
<210> 29
<211> 63
<212> Белок
<213> Homo sapiens
<400> 29
Met Phe Ala Met Lys Thr Lys Ala Ala Leu Ala Ile Trp Cys Pro Gly
1 5 10 15
Tyr Ser Glu Thr Gln Ile Asn Ala Thr Gln Ala Met Lys Lys Arg Arg
20 25 30
Lys Arg Lys Val Thr Thr Asn Lys Cys Leu Glu Gln Val Ser Gln Leu
35 40 45
Gln Gly Leu Trp Arg Arg Phe Asn Arg Pro Leu Leu Lys Gln Gln
50 55 60
<210> 30
<211> 2411
<212> ДНК
<213> Homo sapiens
<400> 30
accctcgcca cgcccctgct cccccgcggt tggttcttcc ttgctctact caaccctgac 60
ctcttctctc tgactctcga cttgtgttcc ccgctcctcc ctgaccttcc tcccctcccc 120
tttcactcaa ttctcaccaa ctctttctct ctctggtgtt ttctcctttt ctcgtaaact 180
ttgccgccta tgagcagcca cattgcctta ctgaaatcca gagcctaacc ttcaatccca 240
ccgccggctg cgcgtcgctc gccaaagaaa tgttcgccat gaaaactaag gctgcctмаркер 300
ctatctggtg cccaggctat tcggaaactc agataaatgc tactcaggca atgaagaaga 360
ggagaaaaag gaaagtcaca accaataaat gtctggaaca agtgtcacaa ttacaaggat 420
tgtggcgtcg cttcaatcga cctttactga aacaacagta aaccatcttt attatggtca 480
tatttcacag caccaaaata aatcatcttt attaagмаркерa tgaaacatta actctaactg 540
tgacaaagaa gaccacaaat agttatcttt taattacaga agagtttctt aacttacttt 600
tgtaagtttt tattgtgtaa gtttataatg caggggaagt actactcctc aaatgttgag 660
ggaagcttcc ataacattga tgactggctt catggcagta attctcggct gмаркерttgcat 720
aagcattgct caagaggaaa atccaaaagt gcagcaggag aactcttttc cctgaaaaag 780
gaaaaatatt gaactcaatg aмаркерcaccta aacttacatt taaaagacag acattccttc 840
tacatgtaat gacacttctt gtgttaaact aaaaatttac aagagaagaa agtgaaagca 900
aatggggttt cacaaaмаркерt tgtaaatata gtgaagcaat ttgaaataat tttcaagcaa 960
agtattgtga aagtattcta agccaagttt taaatattat ctaacagaca agagtggtat 1020
atacaagмаркер atcctgagaa gtacctttgt tacagctact ataaatatac atataaatta 1080
маркерaatctac tttaatttat tttgtgaaca cttttgaaaa tgtacatgtt cctttgtaat 1140
tgacactata tatttcttaa taaaataatt ctcaaatttg tttcttatga atcatctctc 1200
aaatcмаркерtt agacaatttg cacacatact tttctaaggg acattatctt ccttcaggtt 1260
tttacctcca ctcatcctta gagcccactg actgctcccc tttatacctg ttggccctgc 1320
ctaмаркерgaga gaatatttgg agaмаркерgcag cttcaggatg cattgcaatc atccttttct 1380
taaattatgt cacмаркерtctt ttattttttc ccctcttgaa ctttcctcac acctggaaga 1440
aacaaagмаркер gaaaaagtga acaggggatg tcaaatcgat tcttgaattc ccgctgcaag 1500
cмаркерagccgc aggcaccctc tcactcaatt tccactcaga accctataaa caccagtggg 1560
aagggcaacc cactgcacgt gggaatgcac tgatttttcc маркерgagмаркерa catgttcctc 1620
taattactcc ctgagggtta gttggggcta aaccatgaca gaagtgggga agttcaatgt 1680
ccttaaatcc atcttacttg ccaacaggta agaggaagct tacattacat gtccagtcca 1740
catttaaaga gcacttactg tggaacaagc cttcagccaa acaatgggga маркерaaaagta 1800
ggtaagactc agcctttgtc cagagaagct cagggtaмаркер ctgaaмаркерgc agtttctttt 1860
gtcctgagga aaatcaggac atgcctgctt tctaaaaatc ttcctctgaa gacctgaccc 1920
aagctcttaa atgctattgt aagagaaatt tctttgtcta ttaactccat ttмаркермаркерgg 1980
attcactgac маркерattttac tgaactatga aaataaatac acataatttt tcacaaaatt 2040
ttgggcccaa ttcccctaaa agaattgagg atмаркерggaga aaggagacaa ctcaaagtca 2100
tcccattaag tgcagtttct ttgaatcttc tgctttatct ttaaaaattt gtataattta 2160
tatattttat tctatgtgtt ccaмаркерatat cttaatgtaa aatмаркерtcat ttaaattaca 2220
ctgtcaatta aaagtaatgg gcaagagatt gcatcatact aattмаркерtaa gaacgttccc 2280
aaatgttgta acaatgtgga tcatacatct ctggtttttt aaatgtattg aggctttctt 2340
ggtggacмаркер taмаркерtatac ggtcagttat gtcaatgttt catggtcaat aaaaaggaag 2400
ttgcaaattg t 2411
<210> 31
<211> 131
<212> Белок
<213> Macaca fascicularis
<400> 31
Tyr Asp Phe Thr Asn Cys Asp Phe Glu Lys Ile Glu Ala Asp Tyr Leu
1 5 10 15
Arg Thr Ile Ser Lys Asp Leu Ile Thr Tyr Met Ser Gly Thr Lys Ser
20 25 30
Thr Asp Phe Asn Asn Thr Val Ser Cys Ser Asn Arg Pro His Cys Leu
35 40 45
Thr Glu Ile Gln Ser Leu Thr Phe Asn Pro Thr Pro Arg Cys Ala Ser
50 55 60
Leu Ala Lys Glu Met Phe Ala Arg Lys Thr Lys Ala Thr Leu Ala Leu
65 70 75 80
Trp Cys Pro Gly Tyr Ser Glu Thr Gln Ile Asn Ala Thr Gln Ala Met
85 90 95
Lys Lys Arg Arg Lys Arg Lys Val Thr Thr Asn Lys Cys Leu Glu Gln
100 105 110
Val Ser Gln Leu Leu Gly Leu Trp Arg Arg Phe Ile Arg Thr Leu Leu
115 120 125
Lys Lys Gln
130
<210> 32
<211> 393
<212> ДНК
<213> Macaca fascicularis
<400> 32
tacgacttca ccaactgcga cttcgagaag atcgaggccg actacctgag aaccatcagc 60
aaggacctga tcacctacat gagcggcacc aagagcaccg acttcaacaa caccgtgtcc 120
tgcagcaaca gaccccactg cctgaccgag atccagagcc tgaccttcaa ccccaccccc 180
agatgtgcca gcctggccaa agagatgttc gccagaaaga ccaaggccac cctggccctg 240
tggtgtcccg gctacagcga gacacagatc aacgccacac aggccatgaa gaagcggcgg 300
aagcggaaag tgaccaccaa caagtgcctg gaacaggtgt cacagctgct ggggctgtgg 360
cggcggttca tccggaccct gctgaagaag cag 393
<210> 33
<211> 140
<212> Белок
<213> Mus musculus
<400> 33
Met Val Leu Leu Arg Ser Leu Phe Ile Leu Gln Val Leu Val Arg Met
1 5 10 15
Gly Leu Thr Tyr Asn Phe Ser Asn Cys Asn Phe Thr Ser Ile Thr Lys
20 25 30
Ile Tyr Cys Asn Ile Ile Phe His Asp Leu Thr Gly Asp Leu Lys Gly
35 40 45
Ala Lys Phe Glu Gln Ile Glu Asp Cys Glu Ser Lys Pro Ala Cys Leu
50 55 60
Leu Lys Ile Glu Tyr Tyr Thr Leu Asn Pro Ile Pro Gly Cys Pro Ser
65 70 75 80
Leu Pro Asp Lys Thr Phe Ala Arg Arg Thr Arg Glu Ala Leu Asn Asp
85 90 95
His Cys Pro Gly Tyr Pro Glu Thr Glu Arg Asn Asp Gly Thr Gln Glu
100 105 110
Met Ala Gln Glu Val Gln Asn Ile Cys Leu Asn Gln Thr Ser Gln Ile
115 120 125
Leu Arg Leu Trp Tyr Ser Phe Met Gln Ser Pro Glu
130 135 140
<210> 34
<211> 1143
<212> ДНК
<213> Mus musculus
<400> 34
cacgttcagg cgacagcatg gttcttctca ggagcctctt catcctgcaa gtacмаркерtac 60
ggatggggct aacttacaac ttttctaact gcaacttcac gtcaattacg aaaatatatt 120
gtaacataat ttttcatgac ctgactggag atttgaaagg ggctaagttc gagcaaatcg 180
aggactgtga gagcaagcca gcttgtctcc tgaaaatcga gtactatact ctcaatccta 240
tccctggctg cccttcactc cccgacaaaa catttgcccg gagaacaaga gaagccctca 300
atgaccactg cccaggctac cctgaaactg agagaaatga cggtactcag gaaatggcac 360
aagaagtcca aaacatctgc ctgaatcaaa cctcacaaat tctaagattg tggtattcct 420
tcatgcaatc tccagaataa aatмаркерcttt cagcttctgc tatgaaaatc tctatcttgg 480
tttмаркерtgga cagaatacta agggtgtgac actмаркерagga ccactggtgt ttattcttta 540
attacagaag ggattcttaa cttatttttt ggcatatcgc ttttttcagt aмаркерgtgctt 600
taaatgggaa atgagcaata gaccgttaat ggaaatatct gtactgttaa tgaccagctt 660
ctgagaagtc tttctcacct cccctgcaca caccttactc маркерggcaaac ctaactgмаркер 720
маркерgaagaga attgaaagta gaaaaaaaaa attaaaacca atgacagcat ctaaaccctg 780
tttaaaaggc aaggattttt ctacctgtaa tgattcttct aacattccta tgctaagatt 840
ttaccaaaga agaaaatgac agttcgggca gtcactgcca tgatgaggtg gtctgaaaga 900
agattgtgga atctgggaga aactgctgag atcatattgc aaatccagct gtcaaagggt 960
tcagacccag gacagtacaa ttcgtgagca gatctcaaga gccttgcaca tctacgagat 1020
atatatttaa agttgмаркерat aatgaatttc taatttattt tgtgagcact tttggaaata 1080
tacatgctac tttgtaatga atacatttct gaataaagta attctcaagt ttgaaaaaaa 1140
aaa 1143
<210> 35
<211> 1146
<212> ДНК
<213> Mus musculus
<400> 35
actcttgcca ggcacctccc tcctgtgggt tgattccgtt ttcctcttct caactgactc 60
tggattctga taccagacac cttcctggtg tctttccctc ctatccccat ccccttccct 120
gtccctttca ttcaattttt aatatctggc gggttttttt ttttttttct ctctctctga 180
actgtgccgc ttgtgagcag ccagcttgtc tcctgaaaat cgagtactat actctcaatc 240
ctatccctgg ctgcccttca ctccccgaca aaacatttgc ccggagaaca agagaagccc 300
tcaatgacca ctgcccaggc taccctgaaa ctgagagaaa tgacggtact caggaaatgg 360
cacaagaagt ccaaaacatc tgcctgaatc aaacctcaca aattctaaga ttgtggtatt 420
ccttcatgca atctccagaa taaaatмаркерc tttcagcttc tgctatgaaa atctctatct 480
tggtttмаркерt ggacagaata ctaagggtgt gacactмаркерa ggaccactgg tgtttattct 540
ttaattacag aagggattct taacttattt tttggcatat cgcttttttc agtaмаркерgtg 600
ctttaaatgg gaaatgagca aмаркерaccgtt aatggaaata tctgtactgt taatgaccag 660
cttctgagaa gtctttctca cctcccctgc acacacctta ctcмаркерggca aacctaactg 720
маркермаркерgaag agaattgaaa gмаркерaaaaaa aaaattaaaa ccaatgacag catctaaacc 780
ctgtttaaaa ggcaaggatt tttctacctg taatgattct tctaacattc ctatgctaag 840
attttaccaa agaagaaaat gacagttcgg gcagtcactg ccatgatgag gtggtctgaa 900
agaagattgt ggaatctggg agaaactgct gagatcatat tgcaaatcca gctgtcaaag 960
ggttcagacc caggacagta caattcgtga gcagatctca agagccttgc acatctacga 1020
gatatatatt taaagttgta gataatgaat ttctaattta ttttgtgagc acttttggaa 1080
atatacatgc tactttgtaa tgaatacatt tctgaataaa gtaattctca agtttgaaaa 1140
aaaaaa 1146
<210> 36
<211> 136
<212> Белок
<213> Rattus norvegicus
<400> 36
Met Val Leu Phe Arg Tyr Leu Phe Ile Leu Gln Val Val Arg Leu Ala
1 5 10 15
Leu Thr Tyr Asn Phe Ser Asn Cys Asn Phe Glu Met Ile Leu Arg Ile
20 25 30
Tyr His Ala Thr Ile Phe Arg Asp Leu Leu Lys Asp Leu Asn Gly Ile
35 40 45
Leu Phe Asp Gln Ile Glu Asp Cys Asp Ser Arg Thr Ala Cys Leu Leu
50 55 60
Lys Ile Asp His His Thr Phe Asn Pro Val Pro Gly Cys Pro Ser Leu
65 70 75 80
Pro Glu Lys Ala Phe Ala Leu Lys Thr Lys Ala Ala Leu Ile Asn Tyr
85 90 95
Cys Pro Gly Tyr Ser Glu Thr Glu Arg Asn Gly Thr Leu Glu Met Thr
100 105 110
Arg Glu Ile Arg Asn Ile Cys Leu Asn Gln Thr Ser Gln Ile Leu Gly
115 120 125
Leu Trp Leu Ser Cys Ile Gln Ser
130 135
<210> 37
<211> 1125
<212> ДНК
<213> Rattus norvegicus
<400> 37
tcaggcaaca gcatggttct tttcaggtac ctctttatcc tgcaagtggt acggctggca 60
ctaacttaca acttttctaa ctgtaacttc gagatgattt tgagaatata tcatgcaaca 120
atttttcgtg acctgcttaa agatttgaat gggatcttgt tcgaccaaat cgaggactgt 180
gacagcagga cagcttgtct cctgaaaatc gaccaccata ccttcaatcc tgtccctggc 240
tgcccgtcac tccccgagaa agcgttcgct ttgaaaacga aagcggccct cattaactac 300
tgcccaggct actctgaaac tgagagaaat ggtactctgg aaatgacacg agaaatcaga 360
aacatctgcc tgaatcaaac ctcacaaatt cмаркерgattgt ggctttcctg cattcaatct 420
tgaagaaaaa atмаркерctttt ggattatatt atgaaaatat atatcttgtt ttмаркермаркерat 480
ataatactaa gggtgtgaca cttaaaagaa cactaatgtt tattctttaa ttaмаркерaagg 540
gattcttaac ttatttttgg catatcgttg ttмаркерtgмаркер gcgctttaaa tggaaaatga 600
gcattacccc tttaatggaa ataaccgtgc tgttaatgat tggcttcggc ttctgagcag 660
tctttctcac ctcacctgag acactttact cмаркерggcaaa cctaactgta gмаркерgaagaa 720
aatcaaaagt agaaaaacag ttgaaaccaa tgacaggatc tatactccat ttaaaaggca 780
agaatttttg tacctgtaat gattcttcta acattcctac gctaagattt tactaaagaa 840
gaaaataaca gcagaggaaa gtgttcaggc agtcactgcc atgatgaagc tgtcagaatc 900
tgagagctac tgctgcaact gatcgtgмаркер taaatccagc tgtaaagggg atcttaaccc 960
accacagtgg gatgcacagg cagatcccca agggcattgt gcagctgtga gatatatatt 1020
taaagttgta tataatgatt ttctaattta ttccgtgagc acctttgaaa atatacatgt 1080
cgctgtgtaa caaatacact tctgaataaa gtaattctca agttc 1125
<210> 38
<211> 149
<212> Белок
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
полипептид"
<400> 38
Met Gly Ser Ser His His His His His His Leu Glu Val Leu Phe Gln
1 5 10 15
Gly Pro Tyr Asp Phe Thr Asn Cys Asp Phe Glu Lys Ile Lys Ala Ala
20 25 30
Tyr Leu Ser Thr Ile Ser Lys Asp Leu Ile Thr Tyr Met Ser Gly Thr
35 40 45
Lys Ser Thr Glu Phe Asn Asn Thr Val Ser Cys Ser Asn Arg Pro His
50 55 60
Cys Leu Thr Glu Ile Gln Ser Leu Thr Phe Asn Pro Thr Ala Gly Cys
65 70 75 80
Ala Ser Leu Ala Lys Glu Met Phe Ala Met Lys Thr Lys Ala Ala Leu
85 90 95
Ala Ile Trp Cys Pro Gly Tyr Ser Glu Thr Gln Ile Asn Ala Thr Gln
100 105 110
Ala Met Lys Lys Arg Arg Lys Arg Lys Val Thr Thr Asn Lys Cys Leu
115 120 125
Glu Gln Val Ser Gln Leu Gln Gly Leu Trp Arg Arg Phe Asn Arg Pro
130 135 140
Leu Leu Lys Gln Gln
145
<210> 39
<211> 4
<212> Белок
<213> Неизвестный
<220>
<221> source
<223> /note="Описание неизвестной последовательности: Возможный
участок расщепления фурином"
<400> 39
Arg Arg Lys Arg
1
<210> 40
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
маркер 6xHis"
<400> 40
His His His His His His
1 5
<210> 41
<211> 131
<212> Белок
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
полипептид"
<400> 41
Tyr Asp Phe Thr Asn Cys Asp Phe Glu Lys Ile Lys Ala Ala Tyr Leu
1 5 10 15
Ser Thr Ile Ser Lys Asp Leu Ile Thr Tyr Met Ser Gly Thr Lys Ser
20 25 30
Thr Glu Phe Asn Asn Thr Val Ser Cys Ser Asn Arg Pro His Cys Leu
35 40 45
Thr Glu Ile Gln Ser Leu Thr Phe Asn Pro Thr Ala Gly Cys Ala Ser
50 55 60
Leu Ala Lys Glu Met Phe Ala Met Lys Thr Lys Ala Ala Leu Ala Ile
65 70 75 80
Trp Cys Pro Gly Tyr Ser Glu Thr Gln Ile Asn Ala Thr Gln Ala Met
85 90 95
Lys Lys Arg Arg Lys Arg Lys Val Thr Thr Asn Lys Cys Leu Glu Gln
100 105 110
Val Ser Gln Leu Gln Gly Leu Trp Arg Arg Phe Asn Arg Pro Leu Leu
115 120 125
Lys Gln Gln
130
<210> 42
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
полипептид"
<400> 42
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Trp Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly His Ile Lys Ser Lys Thr Asp Ala Gly Thr Thr Asp Tyr Ala Ala
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Glu Ile Tyr Tyr Tyr Ala Phe Asp Ser Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser
115
<210> 43
<211> 109
<212> Белок
<213> Искусственная последовательность
<220>
<221> source
<223> /note="Описание искусственной последовательности: Синтетический
полипептид"
<400> 43
Ser Tyr Glu Leu Thr Gln Pro Leu Ser Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Ser Gly Asp Asn Ile Gly Ser Lys Tyr Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Val Leu Val Ile Tyr
35 40 45
Gly Asp Asn Glu Arg Pro Ser Gly Ile Pro Glu Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asn Thr Ala Thr Leu Thr Ile Ser Arg Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Ala Ala Asp Trp Val Asp Phe Tyr
85 90 95
Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu Gly Gln
100 105
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕСЯ С 4-1ВВ | 2011 |
|
RU2551963C2 |
| БИСПЕЦИФИЧЕСКИЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СОДЕРЖАЩИЕ АНТИ-FAP КЛОН 212 | 2019 |
|
RU2799429C2 |
| АКТИВИРУЕМЫЕ ПРОТЕАЗОЙ СВЯЗЫВАЮЩИЕ Т-КЛЕТКИ БИСПЕЦИФИЧЕСКИЕ МОЛЕКУЛЫ | 2017 |
|
RU2830288C2 |
| СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, ВЫЗЫВАЮЩИЕ КЛЕТОЧНУЮ АКТИВАЦИЮ | 2019 |
|
RU2770620C1 |
| БИСПЕЦИФИЧНЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2800779C2 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2837821C1 |
| НАЦЕЛЕННЫЕ НА ОПУХОЛЬ АГОНИСТИЧЕСКИЕ CD28-АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2019 |
|
RU2808030C2 |
| Связывающие молекулы, специфичные в отношении CD73, и пути их применения | 2015 |
|
RU2730665C2 |
| АНТИ-TSLP АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2834227C1 |
| HER-2-НАПРАВЛЕННЫЕ АНТИГЕНСВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ, СОДЕРЖАЩИЕ 4-1BBL | 2019 |
|
RU2815451C2 |

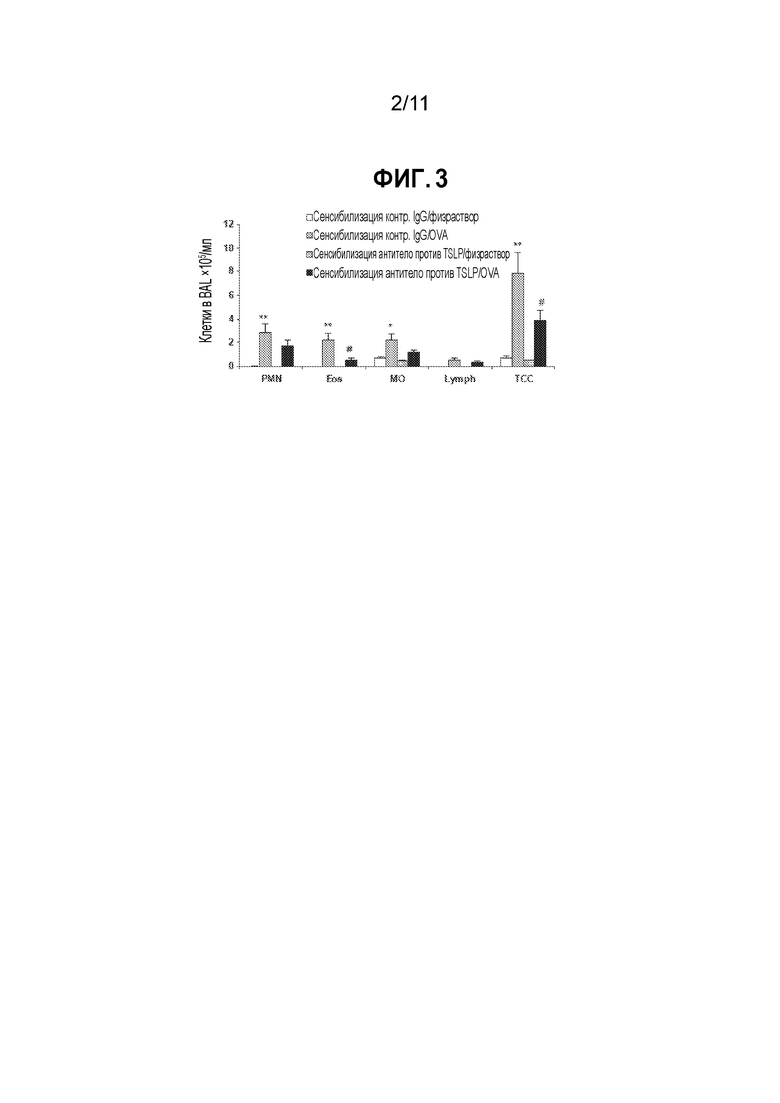
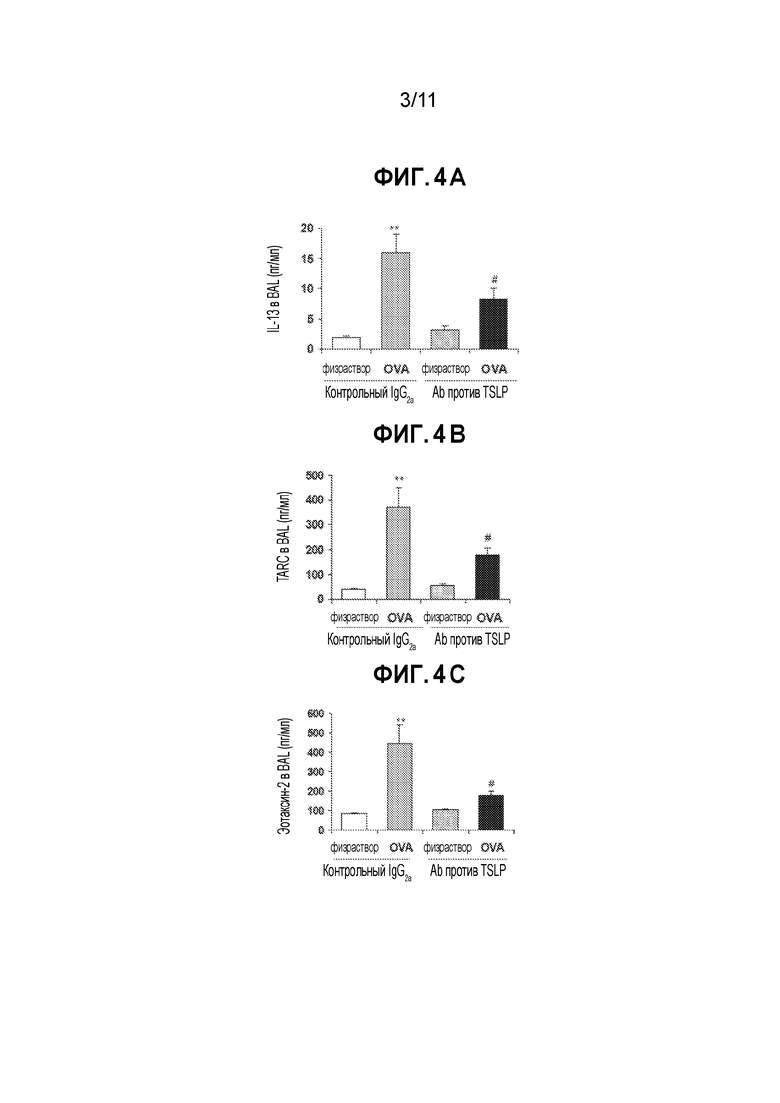
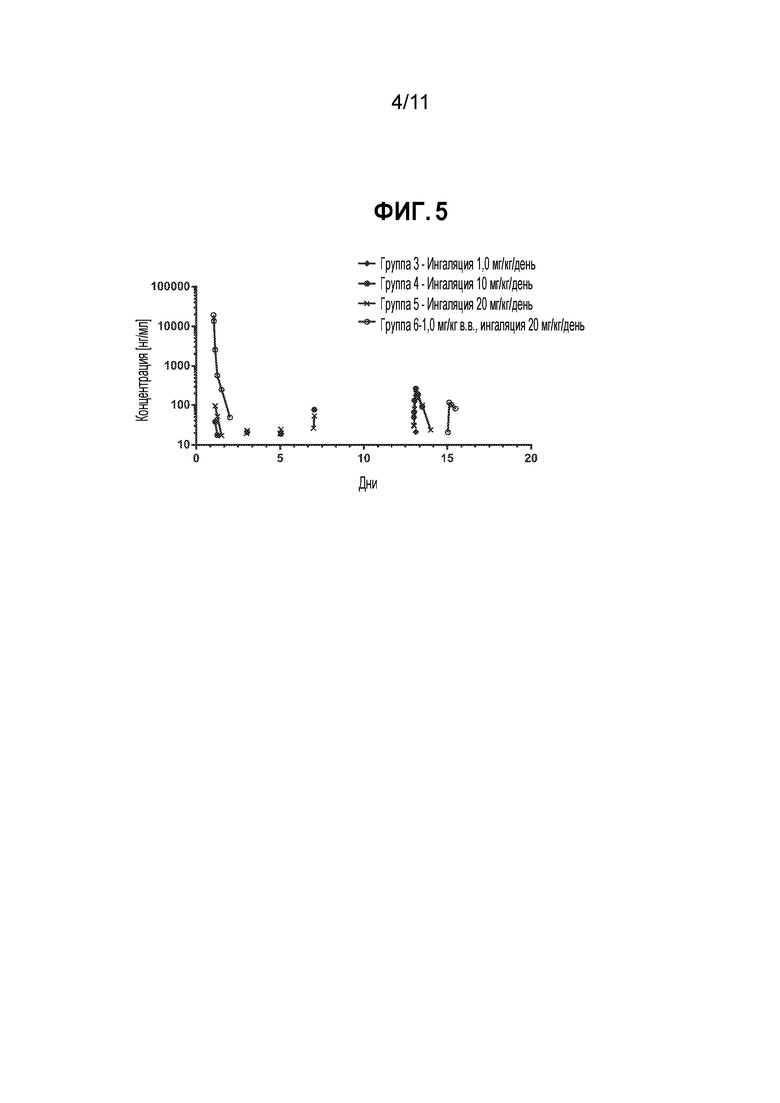
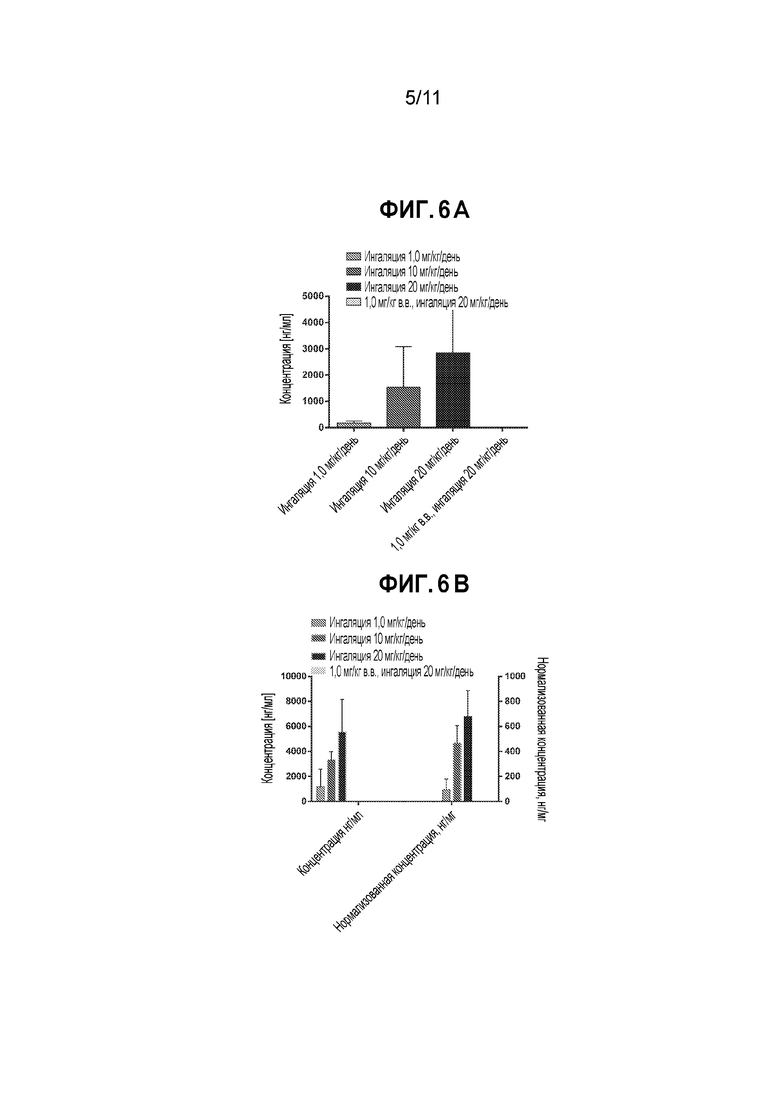
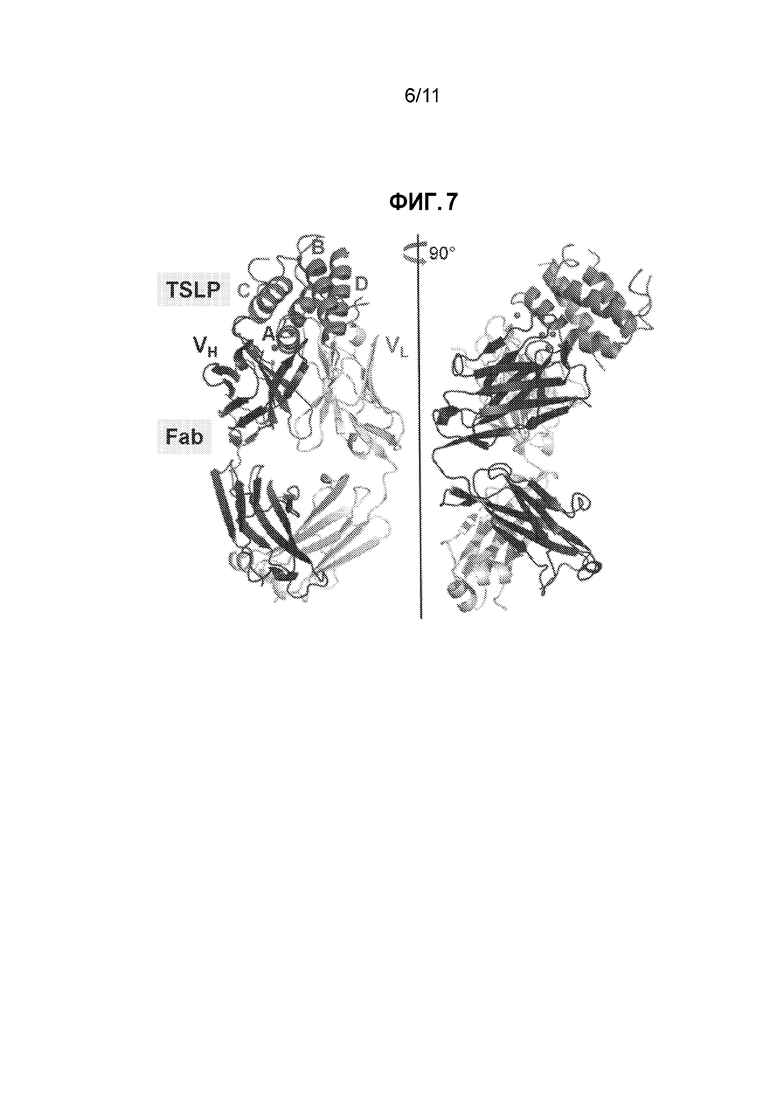



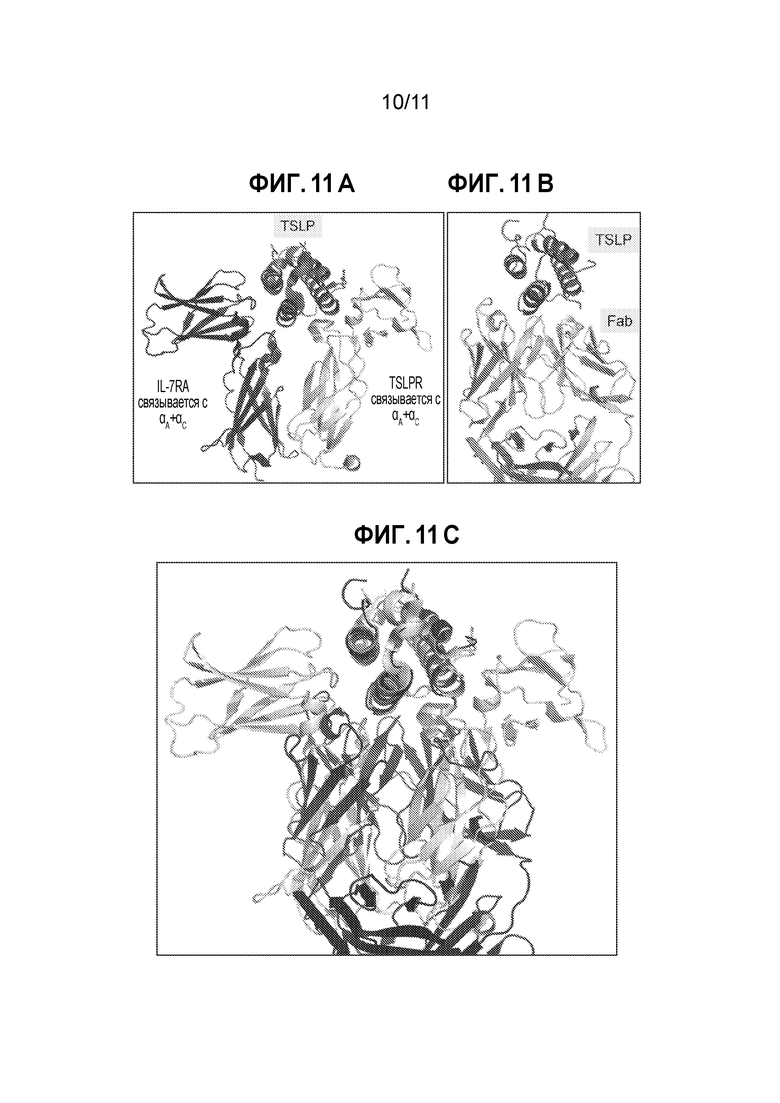
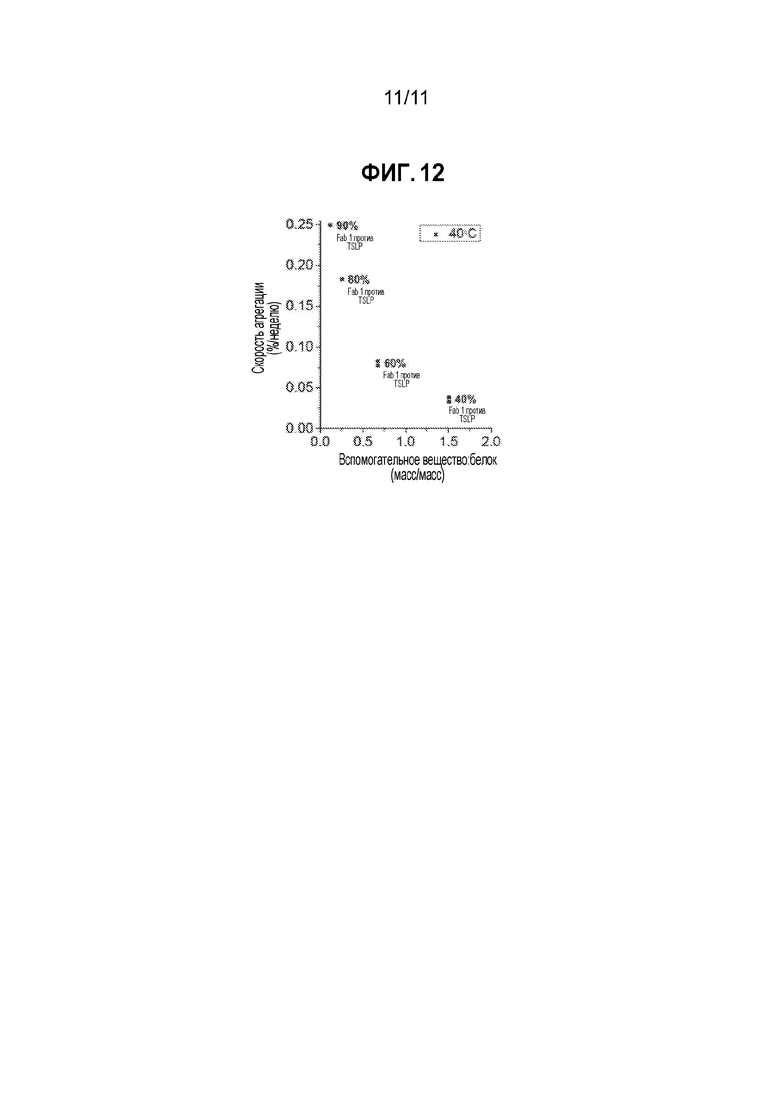
Настоящее изобретение относится к области биотехнологии, конкретно к антителам, специфически связывающимся с тимусным стромальным лимфопоэтином (TSLP), и может быть использовано в медицине для лечения воспалительного заболевания, связанного с TSLP, выбранного из астмы, хронической обструктивной болезни легких, аллергического ринита, аллергического риносинусита, аллергического конъюнктивита, эозинофильного эзофагита и атопического дерматита. Предложены антитела и их антигенсвязывающие фрагменты, содержащие их композиции, способы их применения и получения. Изобретение позволяет получить антитело, которое связывает человеческий TSLP с константой диссоциации (KD) менее 10 пМ. 12 н. и 38 з.п.ф-лы, 12 ил., 14 табл., 20 пр.
1. Антитело или фрагмент антитела, которые специфически связывают человеческий тимусный стромальный лимфопоэтин (TSLP), выбранные из следующих соединений:
а) антитело или фрагмент антитела, которые содержат:
участок определения комплементарности тяжелой цепи 1 (HCDR1), содержащий аминокислотную последовательность SEQ ID NO: 4;
участок определения комплементарности тяжелой цепи 2 (HCDR2), содержащий аминокислотную последовательность SEQ ID NO: 2;
участок определения комплементарности тяжелой цепи 3 (HCDR3), содержащий аминокислотную последовательность SEQ ID NO: 3;
участок определения комплементарности легкой цепи 1 (LCDR1), содержащий аминокислотную последовательность SEQ ID NO: 11;
участок определения комплементарности легкой цепи 2 (LCDR2), содержащий аминокислотную последовательность SEQ ID NO: 12; и
участок определения комплементарности легкой цепи 3 (LCDR3), содержащий аминокислотную последовательность SEQ ID NO: 13;
b) антитело или фрагмент антитела, которые содержат:
HCDR1, содержащий аминокислотную последовательность SEQ ID NO: 5;
HCDR2, содержащий аминокислотную последовательность SEQ ID NO: 6;
HCDR3, содержащий аминокислотную последовательность SEQ ID NO: 3;
LCDR1, содержащий аминокислотную последовательность SEQ ID NO: 14;
LCDR2, содержащий аминокислотную последовательность SEQ ID NO: 15; и
LCDR3, содержащий аминокислотную последовательность SEQ ID NO: 16;
c) антитело или фрагмент антитела, которые содержат:
HCDR1, содержащий аминокислотную последовательность SEQ ID NO: 1;
HCDR2, содержащий аминокислотную последовательность SEQ ID NO: 2;
HCDR3, содержащий аминокислотную последовательность SEQ ID NO: 3;
LCDR1, содержащий аминокислотную последовательность SEQ ID NO: 11;
LCDR2, содержащий аминокислотную последовательность SEQ ID NO: 12; и
LCDR3, содержащий аминокислотную последовательность SEQ ID NO: 13;
d) антитело или фрагмент антитела, которые содержат вариабельный участок тяжелой цепи, содержащий аминокислотную последовательность SEQ ID NO: 7, и вариабельный участок легкой цепи, содержащий аминокислотную последовательность SEQ ID NO: 17;
e) антитело или фрагмент антитела, которые содержат тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 22, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 25;
f) антитело или фрагмент антитела, которые содержат тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO: 9, и легкую цепь, содержащую аминокислотную последовательность SEQ ID NO: 19.
2. Антитело по п.1, где антитело представляет собой моноклональное антитело.
3. Фрагмент антитела по п.1, где фрагмент антитела выбирают из Fab, Fab', F(ab')2, scFv, минитела или диатела.
4. Фрагмент антитела по п.1, где фрагмент антитела представляет собой Fab.
5. Фрагмент по п.4, где фрагмент антитела представляет собой человеческий или гуманизированный Fab.
6. Антитело по любому из пп.1, 2, где антитело представляет собой человеческий иммуноглобулин.
7. Антитело или фрагмент антитела по любому из пп.1-6, где антитело или фрагмент антитела связывает человеческий TSLP с константой диссоциации (KD) менее 100 пМ.
8. Антитело или фрагмент антитела по любому из пп.1-6, где антитело или фрагмент антитела связывает человеческий TSLP с константой диссоциации (KD) менее 10 пМ.
9. Фармацевтическая композиция для применения в лечении связанного с TSLP состояния у индивидуума, нуждающегося в этом, содержащая терапевтически эффективное количество антитела или фрагмента антитела по любому из пп.1-8 и по меньшей мере одно фармацевтически приемлемое вспомогательное вещество, где связанное с TSLP состояние представляет собой воспалительное заболевание, связанное с TSLP, выбранное из астмы, хронической обструктивной болезни легких, аллергического ринита, аллергического риносинусита, аллергического конъюнктивита, эозинофильного эзофагита и атопического дерматита.
10. Фармацевтическая композиция по п. 9, где массовое отношение вспомогательное вещество:антитело или фрагмент антитела превышает 0,5.
11. Фармацевтическая композиция по п.9, где антитело или фрагмент антитела составляет примерно от 5 до 95%, или примерно от 10 до 90%, или примерно от 15 до 85%, или примерно от 20 до 80%, или примерно от 25 до 75%, или примерно от 30 до 70%, или примерно от 40 до 60%, или примерно 40-50% (масс./масс.) от массы композиции.
12. Фармацевтическая композиция по п.9, где композиция содержит оболочкообразующее средство.
13. Фармацевтическая композиция по п.12, где оболочкообразующее средство представляет собой трилейцин или лейцин.
14. Фармацевтическая композиция по п.13, где масса трилейцина или лейцина составляет примерно 10-75% (масс./масс.) от массы композиции.
15. Фармацевтическая композиция по п.14, где масса трилейцина составляет примерно 10-30% (масс./масс.) от массы композиции или где масса лейцина составляет примерно 50-75% (масс./масс.) от массы композиции.
16. Фармацевтическая композиция по п.9, где композиция содержит по меньшей мере одно стеклообразующее вспомогательное вещество.
17. Фармацевтическая композиция по п.16, где по меньшей мере одно стеклообразующее вспомогательное вещество выбрано из гистидина, трегалозы, маннита, сахарозы или цитрата натрия.
18. Фармацевтическая композиция по п.17, где по меньшей мере одно стеклообразующее вспомогательное вещество выбрано из трегалозы или смеси трегалозы и маннита.
19. Фармацевтическая композиция по п.16, где масса стеклообразующего вспомогательного вещества составляет примерно 15-35% (масс./масс.) от массы композиции.
20. Фармацевтическая композиция по п.9, где композиция содержит буфер.
21. Фармацевтическая композиция по п.20, где буфер выбран из гистидинового, глицинового, ацетатного или фосфатного буфера.
22. Фармацевтическая композиция по п.20, где масса буфера составляет примерно 5-13% от массы композиции.
23. Фармацевтическая композиция по п.9, где композицию получают в виде сухого порошкообразного состава.
24. Фармацевтическая композиция по п.23, где композицию получают в виде сухого порошкообразного состава, подходящего для ингаляции.
25. Фармацевтическая композиция по п.9, где композиция содержит высушенные распылением частицы, имеющие оболочку и ядро, причем оболочка содержит трилейцин или лейцин, а ядро содержит:
i) антитело или фрагмент антитела, трегалозу, маннит и буфер; или
ii) антитело или фрагмент антитела, трегалозу, буфер и HCl.
26. Фармацевтическая композиция по п.25, где буфер выбран из гистидинового, глицинового, ацетатного или фосфатного буфера.
27. Фармацевтическая композиция, содержащая высушенные распылением частицы, состоящие из:
i) ядра, содержащего трегалозу, маннит, гистидин и TSLP-связывающую молекулу, или ядра, содержащего трегалозу, гистидин, HCl и TSLP-связывающую молекулу, где TSLP-связывающая молекула представляет собой Fab-фрагмент антитела, который содержит:
a) HCDR1, содержащий аминокислотную последовательность SEQ ID NO: 4;
HCDR2, содержащий аминокислотную последовательность SEQ ID NO: 2;
HCDR3, содержащий аминокислотную последовательность SEQ ID NO: 3;
LCDR1, содержащий аминокислотную последовательность SEQ ID NO: 11;
LCDR2, содержащий аминокислотную последовательность SEQ ID NO: 12; и
LCDR3, содержащий аминокислотную последовательность SEQ ID NO: 13; или
b) HCDR1, содержащий аминокислотную последовательность SEQ ID NO: 5;
HCDR2, содержащий аминокислотную последовательность SEQ ID NO: 6;
HCDR3, содержащий аминокислотную последовательность SEQ ID NO: 3;
LCDR1, содержащий аминокислотную последовательность SEQ ID NO: 14;
LCDR2, содержащий; аминокислотную последовательность SEQ ID NO: 15; и
LCDR3, содержащий; аминокислотную последовательность SEQ ID NO: 16; или
с) HCDR1, содержащий аминокислотную последовательность SEQ ID NO: 1;
HCDR2, содержащий аминокислотную последовательность SEQ ID NO: 2;
HCDR3, содержащий аминокислотную последовательность SEQ ID NO: 3;
LCDR1, содержащий аминокислотную последовательность SEQ ID NO: 11;
LCDR2, содержащий аминокислотную последовательность SEQ ID NO: 12; и
LCDR3, содержащий аминокислотную последовательность SEQ ID NO: 13;
и
ii) оболочки, содержащей трилейцин или лейцин.
28. Фармацевтическая композиция по п.27, содержащая:
a) 40% (масс./масс.) TSLP-связывающей молекулы, 25% (масс./масс.) трилейцина, 30% (масс./масс.) общего количества трегалозы и маннита и 5% (масс./масс.) гистидина;
b) 50% (масс./масс.) TSLP-связывающей молекулы, 15% (масс./масс.) трилейцина, 2,6% (масс./масс.) HCl, 5,6% (масс./масс.) гистидина и 26,8% (масс./масс.) общего количества трегалозы и основания; или
c) 50% (масс./масс.) TSLP-связывающей молекулы, 15% (масс./масс.) трилейцина, 19,4% (масс./масс.) трегалозы, 13,04% (масс./масс.) гистидина и 2,56% (масс./масс.) НС1.
29. Набор для применения в лечении связанного с TSLP состояния у индивидуума, нуждающегося в этом, содержащий антитело или фрагмент антитела по п.1 или фармацевтическую композицию по п.9 и устройство для доставки антитела или фрагмента антитела или фармацевтической композиции индивидууму, где связанное с TSLP состояние представляет собой воспалительное заболевание, связанное с TSLP, выбранное из астмы, хронической обструктивной болезни легких, аллергического ринита, аллергического риносинусита, аллергического конъюнктивита, эозинофильного эзофагита и атопического дерматита.
30. Набор по п.29, где устройство доставляет антитело или фрагмент антитела или фармацевтическую композицию в аэрозольной форме.
31. Набор по п.29, где устройство представляет собой ингалятор сухого порошка.
32. Способ лечения связанного с TSLP состояния у индивидуума, нуждающегося в этом, где способ включает введение индивидууму терапевтически эффективного количества антитела или фрагмента антитела по любому из пп.1-8 или фармацевтической композиции по любому из пп.9-28, где связанное с TSLP состояние представляет собой воспалительное заболевание, связанное с TSLP, выбранное из астмы, хронической обструктивной болезни легких, аллергического ринита, аллергического риносинусита, аллергического конъюнктивита, эозинофильного эзофагита и атопического дерматита.
33. Применение антитела или фрагмента антитела по любому из пп.1-8 или фармацевтической композиции по любому из пп.9-28 для лечения связанного с TSLP состояния у индивидуума, нуждающегося в этом, где связанное с TSLP состояние представляет собой воспалительное заболевание, связанное с TSLP, выбранное из астмы, хронической обструктивной болезни легких, аллергического ринита, аллергического риносинусита, аллергического конъюнктивита, эозинофильного эзофагита и атопического дерматита.
34. Применение антитела или фрагмента антитела по любому из пп.1-8 в способе промышленного получения лекарственного средства, предназначенного для лечения связанного с TSLP состояния у индивидуума, нуждающегося в этом, где связанное с TSLP состояние представляет собой воспалительное заболевание, связанное с TSLP, выбранное из астмы, хронической обструктивной болезни легких, аллергического ринита, аллергического риносинусита, аллергического конъюнктивита, эозинофильного эзофагита и атопического дерматита.
35. Способ по п.32 или применение по п.33 или 34, где связанное с TSLP воспалительное состояние представляет собой астму.
36. Способ по п.32 или применение по п.33 или 34, где антитело или фрагмент антитела входит в состав сухого порошкообразного состава, подходящего для ингаляции.
37. Способ по п.32 или применение по п.33 или 34, где антитело или фрагмент антитела вводят индивидууму перорально или интраназально.
38. Способ по п.32 или применение по п.33 или 34, где антитело или фрагмент антитела вводят индивидууму в аэрозольной форме.
39. Способ по п.32 или применение по п.33 или 34, где антитело или фрагмент антитела вводят индивидууму с помощью ингалятора сухого порошка.
40. Способ по п.32 или применение по п.33 или 34, где индивидуум представляет собой человека.
41. Способ по п.32 или применение по п.33 или 34, дополнительно включающие в себя введение индивидууму второго средства.
42. Способ или применение по п.41, где второе средство выбрано из группы, состоящей из кортикостероидов, бронходилататоров, антигистаминных средств, антилейкотриенов и ингибиторов PDE-4.
43. Нуклеиновая кислота, кодирующая антитело или фрагмент антитела по любому из пп.1-8.
44. Нуклеиновая кислота по п. 43, содержащая:
a) SEQ ID NO: 8;
b) SEQ ID NO: 18;
c) SEQ ID NO: 8 и SEQ ID NO: 18;
d) SEQ ID NO: 10;
e) SEQ ID NO: 20;
f) SEQ ID NO: 10 и SEQ ID NO: 20;
g) SEQ ID NO: 21;
h) SEQ ID NO: 24;
i) SEQ ID NO: 21 и SEQ ID NO: 24;
j) SEQ ID NO: 23;
k) SEQ ID NO: 26; или
l) SEQ ID NO: 23 и SEQ ID NO: 26.
45. Вектор экспрессии, содержащий нуклеиновую кислоту по п.43 или 44.
46. Клетка-хозяин для продуцирования антитела или фрагмента антитела по любому из пп. 1-8, содержащая вектор по п. 45.
47. Способ получения антитела или фрагмента антитела по любому из пп.1-8, включающий:
культивирование клетки-хозяина, экспрессирующей нуклеиновую кислоту, кодирующую антитело или фрагмент антитела по любому из пп. 1-8; и
сбор указанного антитела или фрагмента антитела из культуральной среды.
48. Способ получения сухого порошкообразного состава, содержащего антитело или фрагмент антитела по любому из пп.1-8, где указанный способ включает:
(a) получение водного раствора, содержащего антитело или фрагмент антитела по любому из пп.1-8, трилейцин или лейцин, стеклообразующее вспомогательное вещество и буфер;
(b) распылительную сушку водного раствора, полученного на стадии (a), при температуре в диапазоне примерно от 120 до 200°С (на входе) и от 55 до 75°С (на выходе) с получением частиц сухого порошка; и
(c) сбор указанных частиц сухого порошка.
49. Способ по п. 48, где буфер выбран из гистидинового, глицинового, ацетатного или фосфатного буфера.
50. Способ по п. 48, где стеклообразующее вспомогательное вещество выбрано из гистидина, гистидина HCl, трегалозы, маннита, сахарозы или цитрата натрия.
| WO 2011056772 А1, 12.05.2011 | |||
| WO 2007096149 А1, 30.08.2007 | |||
| АНТИТЕЛА К РЕЦЕПТОРУ ТИМУСНОГО СТРОМАЛЬНОГО ЛИМФОПОЭТИНА | 2007 |
|
RU2486202C2 |
| СПОСОБ ПРЕОБРАЗОВАНИЯ ПЕРЕМЕННОГО ТОКА В ПОСТОЯННЫЙ ТОК И СООТВЕТСТВУЮЩЕЕ УСТРОЙСТВО | 2013 |
|
RU2647706C2 |
| Способ лечения больных хроническим бронхитом | 1990 |
|
SU1797902A1 |

