Настоящее изобретение относится к антителам человека, гуманизированным и моноклональным антителам со сниженной иммуногенностью, их применению и способам получения. Изобретение, в частности, относится к моноклональным антителам-антагонистам против CD40 человека.
Молекула CD40 представляет собой мембранный гликопротеин типа I с молекулярной массой 50 кДа и экспрессируется на В-клетках, моноцитах/макрофагах, дендритных клетках (DC) и активированных эндотелиальных клетках.1-6 При определенных состояниях CD40 можно также обнаружить на фибробластах, эпителиальных клетках и кератиноцитах.7 Лиганд CD40 (CD40L, CD154), интегральный мембранный гликопротеин типа II с молекулярной массой 32 кДа, транзиторно экспрессируется на активированных Т-клетках CD4+ и небольшой популяции активированных Т-клеток CD8+.8,9 Кроме того, CD40L обнаружен на ряде других типов клеток после активации, включая тучные клетки, базофилы, В-клетки, эозинофилы, DC и тромбоциты.10,11
Исследования на мышиных моделях ясно продемонстрировали вовлеченность взаимодействия CD40L-CD40 в патофизиологию различных аутоиммунных заболеваний (для обзора см. ссылку12). Данные, полученные на трансгенных по CD40L мышах, которые приобретают смертельное воспалительное заболевание кишечника, предоставили первое доказательство того, что взаимодействия CD40-CD40L могут также играть роль в патогенезе воспалительных заболеваний кишечника.13 Моноклональное антитело (мАт) против CD40 мыши эффективно предупреждает воспаление слизистой оболочки и продукцию интерферона-γ Т-клетками CD4+ собственного слоя (lamina propria) при TNBS-индуцированном колите.14 На мышиной модели воспалительного заболевания кишечника с тяжелым комбинированным иммунодефицитом (SCID) показано, что лечение анти-CD40L со дня восстановления Т-клеток полностью предотвращало клиническое и гистологическое проявление экспериментального колита.15 Более того, при введении анти-CD40L с 5 недели после восстановления Т-клеток сохранялась возможность предотвратить прогрессирование заболевания, и получавшие лечение животные демонстрировали улучшение симптомов заболевания и гистологии по сравнению с контрольными животными.15 Кроме того, восстановление Т-клеток мышей с SCID Т-клетками от мышей с выключенным CD40L далее продемонстрировало существенную роль экспрессирующих CD40L Т-клеток в развитии заболевания и продукции интерлейкина-12.16
Взаимодействие CD40-CD40L можно предупредить с помощью моноклональных антител (мАт) против или CD40L, или CD40. Экспрессия CD40L на активированных тромбоцитах привела к тромбоэмболическим событиям во время лечения людей мАт против CD40L человека в виде IgG1 при высших уровнях дозы и прекращению разработки этих мАт.17,19 Поэтому антагонизм в отношении CD40 представляется более привлекательным подходом. Нестимуляторная антагонистическая активность мАт 5D12 (против CD40 человека) была продемонстрирована в различных in vitro исследованиях с использованием различных типов несущих CD40 клеток20,22, и антагонистическая активность химерных 5D12 (ch5D12) была подтверждена in vivo, используя модели заболевания на различных не являющихся людьми приматах.23,27 ch5D12 представляет собой молекулярно сконструированное антитело в виде IgG4 человека, содержащее вариабельные области тяжелой и легкой цепей 5D12 мышы, и было сконструировано для снижения способности к иммуногенности и увеличения полупериода существования in vivo мышиного мАт 5D12 при использовании для людей.
Больные болезнью Крона страдают от вызывающего слабость воспалительного расстройства желудочно-кишечного тракта, точная этиология и патогенез которого остаются неизвестными.28,29 Заболевание характеризуется поступлением в больную слизистую оболочку активированных Т-клеток, В-клеток и макрофагов,30,31 локальной продукцией растворимых медиаторов воспаления и повреждением вовлеченных тканей.28,29 Показано, что Т-клетки CD4+ и макрофаги в слизистой оболочке и цитокины, такие как фактор некроза опухолей (TNF)-α и IL-12, играют центральную роль в инициировании воспалительного цикла при болезни Крона.32,38 Т-клетки воспаленной слизистой оболочки проявляют наивысшую пролиферативную способность28,29 и секретируют увеличенные количества IFN-γ и IL-2. Увеличенные уровни транскриптов мРНК для цитокинов, связанных с Т-клетками, обнаружены в образцах биопсии слизистых оболочек больных болезнью Крона.33 При исследовании экспрессии CD40/CD40L в местах поражений при болезни Крона, проведенном авторами настоящего изобретения, было высказано предположение о том, что CD40L играет основную роль на активированных Т-клетках CD4+.39 CD40L может опосредовать сильную активацию несущих CD40 клеток, главным образом В-клеток и макрофагов, приводя тем самым к увеличению продукции TNF-α и IL-12 в местах поражений. При использовании иммуногистохимии увеличение окрашивания 5D12 было обнаружено во всех образцах больных областей больных болезнью Крона по сравнению с непораженными областями. Двойное окрашивание на CD40 и CD20 (В-клетки) или CD68 (макрофаги) показало, что в срезах образцов больных болезнью Крона клетки CD40+ были, главным образом, В-клетками в лимфоидных фолликулах и макрофагами в собственном слое. Т-клетки собственного слоя воспаленной слизистой оболочки больных болезнью Крона индуцировали продукцию моноцитами значительных количеств IL-12 и TNF-α через 48 ч совместного культивирования. Добавление 5D12 приводило к снижению продукции IL-12 и TNF-α; уровни продукции снижались до уровней, наблюдаемых при использовании контрольных Т-клеток собственного слоя как в отсутствие, так в присутствии IFN-γ.39
Целью настоящего изобретения является получение альтернативных молекул, имеющих по крайней мере такую же безопасность и/или эффективность, что и 5D12 in vivo, но не обязательно такую же степень указанной безопасности и/или эффективности. Антитело 5D12, или по крайней мере его вариабельные области, имеет мышиное происхождение. Настоящее изобретение относится к вариантам вариабельных областей тяжелой и легкой цепей 5D12. В результате, настоящее изобретение относится к полипептиду, включающему аминокислотную последовательность формулы (I)
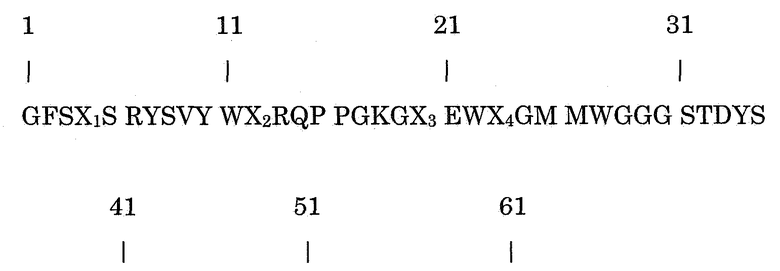
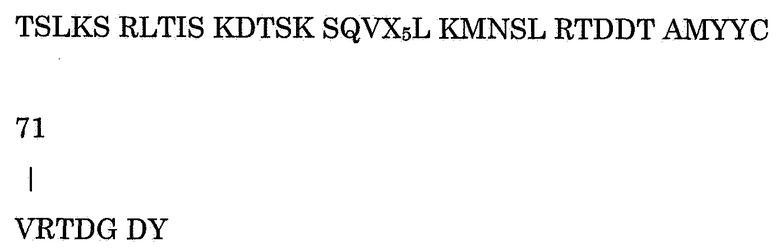
В которой
Х1 представляет собой G, A, V, L, I, P, F, M, W, C, N, Q, S, T, Y, D, E, K, R или H;
Х2 представляет собой G, A, V, L, I, P, F, M, W, C, N, Q, S, T, Y, D, E, K, R или H;
Х3 представляет собой G, A, V, L, I, P, F, M, W, C, N, Q, S, T, Y, D, E, K, R или H;
Х4 представляет собой G, A, V, L, I, P, F, M, W, C, N, Q, S, T, Y, D, E, K, R или H; и
Х5 представляет собой G, A, V, L, I, P, F, M, W, C, N, Q, S, T, Y, D, E, K, R или H.
Указанный полипептид содержит последовательность, которая идентична последовательности вариабельной области тяжелой цепи антитела 5D12, однако полипептид менее иммуногенен для человека, которому его вводят сам по себе или как часть антитела, содержащего указанный полипептид. В указанных положениях Х1-Х5 некоторые аминокислоты могут присутствовать, как указано. Связывающая молекула содержит полипептид по настоящему изобретению, обладает удовлетворительными свойствами связывания CD40. Отмечено, что продукция антитела в клетке применяется до некоторой степени в зависимости от вида аминокислоты в указанных положениях. Это будет подробно описано далее.
Настоящее изобретение, кроме того, относится к полипептиду по настоящему изобретению, содержащему аминокислотную последовательность


Указанный выше полипептид по существу перекрывает вариабельную область тяжелой цепи антитела 5D12 мыши. Аминокислотная последовательность модифицирована в нескольких положениях относительно последовательности мыши. Модифицированный полипептид обладает как хорошими связывающими свойствами, так и хорошей переносимостью людьми, которые получают этот полипептид. В указанные положения могут быть встроены различные аминокислоты без значительного снижения и/или изменения по крайней мере иммунологических свойств полипептида по сравнению с исходным полипептидом мыши. Предпочтительно, полипептид по настоящему изобретению содержит G, A, V, L, P, F, M, W, C, N, Q, S, T, Y, D, E, K, R или H в положении Х1. Таким образом, не снижается и/или значительно не изменяется по крайней мере способность продуцировать полипептид в клетке млекопитающего по сравнению с исходным полипептидом мыши или химерным полипептидом. В особенно предпочтительном варианте осуществления по настоящему изобретению
Х1 представляет собой G, A, V, L, P, F или M;
Х2 представляет собой G, A, V, L, I, P, F или M;
Х3 представляет собой G, A, V, L, I, P, F, M;
Х4 представляет собой G, A, V, L, I, P, F, M; и
Х5 представляет собой G, A, V, L, I, P, F, M, W, C, N, Q, S, T или Y.
Такие полипептиды предпочтительны для продукции на высоком уровне в клетке млекопитающего, в частности, в составе антитела.
Кроме того, настоящее изобретение относится к полипептиду по настоящему изобретению, в котором
Х1 представляет собой G, A, V, L или M;
Х2 представляет собой G, A, V, L, I или M;
Х3 представляет собой G, A, V, L, I, P, F, M;
Х4 представляет собой G, A, V, L, I или M; и
Х5 представляет собой P, F, W, N, Q, S, T или Y.
Такие полипептиды более предпочтительны для продукции антител на высоком уровне и при этом хорошо переносятся человеком. Кроме того, настоящее изобретение относится к полипептиду формулы (I), в котором Х1 представляет собой L; Х2 представляет собой I; Х3 представляет собой P; Х4 представляет собой M; и/или Х5 представляет собой S. Такой полипептид является особенно предпочтительным благодаря способности эффективно продуцироваться в составе антитела, которое воспроизводит связывающие и фармакологические свойства антитела ch5D12, при этом иммунологические свойства людей улучшены по сравнению с антителом мыши или антителом ch5D12, и при этом по крайней мере значительно не снижена продукция полипептида по сравнению с мышиным или химерным аналогом.
Кроме того, настоящее изобретение относится к полипептиду формулы (I), в котором Х1 представляет собой I; Х2 представляет собой V; Х3 представляет собой P; Х4 представляет собой M; и/или Х5 представляет собой S. Этот полипептид является особенно предпочтительным из-за его способности эффективно продуцироваться в составе антитела, которое воспроизводит связывающие и фармакологические свойства антитела ch5D12, при этом обладая улучшенными иммунологическими свойствами у людей по сравнению с антителом мыши или антителом ch5D12, и при этом по крайней мере значительно не снижена продукция полипептида по сравнению с мышиным или химерным аналогом.
Антитело, которое отличается по крайней мере одним положением Х1-Х5 от химерного 5D12 в соответствующих положениях, обладает лучшими иммунологическими свойствами по сравнению с химерным антителом 5D12 у людей. В предпочтительном варианте осуществления таких антител настоящее изобретение, предпочтительно, относится к полипептиду по настоящему изобретению, в котором Х1 представляет собой I, а Х2 представляет собой V; Х1 представляет собой I, а Х2 представляет собой I; Х1 представляет собой L, а Х2 представляет собой I; или Х1 представляет собой L, а Х2 представляет собой V. Особенно предпочтительны такие полипептиды в комбинации с Х3, которая представляет собой Р; Х4, которая представляет собой M; и Х5, которая представляет собой F или S. В одном из вариантов осуществления настоящее изобретение относится к полипептиду по настоящему изобретению, в котором Х1 представляет собой L, Х2 представляет собой V; Х3 представляет собой L; Х4 представляет собой L; и Х5 представляет собой F. Продукция антител, содержащих указанный полипептид, является крайне эффективной при одновременном наличии улучшенных иммунологических свойств у людей по сравнению с ch5D12. Кроме того, настоящее изобретение относится к полипептиду, содержащему аминокислотную последовательность GFSX1SRYSVYWX2R, в которой Х1 представляет собой L, а Х2 представляет собой I; или Х1 представляет собой I, а Х2 представляет собой V. Этот полипептид включает модифицированный CDR1 вариабельной области тяжелой цепи 5D12. Этот CDR1 содержит по крайней мере одну отличную аминокислоту по сравнению с CDR1 вариабельной области тяжелой цепи антитела 5D12. Эта аминокислотная замена приводит к улучшению иммунологических свойств модифицированного антитела 5D12 или ch5D12, причем модификация включает по крайней мере замену соответствующей последовательности указанного полипептида в 5D12 или ch5D12 полипептидом по настоящему изобретению, но при этом обеспечивает высокую продукцию антитела в клетке млекопитающего.
Кроме того, настоящее изобретение относится к вариабельной области тяжелой цепи, содержащей полипептид формулы (I). Указанная вариабельная область содержит, предпочтительно, 90-130, более предпочтительно, 100-120, более предпочтительно, 105-115, наиболее предпочтительно, 113 аминокислот. Полипептид может быть получен синтетически или продуцироваться клеткой. Предпочтительно, указанная вариабельная область тяжелой цепи продуцируется клеткой. В природе существует по крайней мере пять типов тяжелых цепей: γ, δ, α, µ и ε, причем каждый тип определяет класс иммуноглобулинов. Полипептид по настоящему изобретению можно использовать в качестве связывающего вещества непосредственно, или его можно ввести в состав антитела. При введении в состав антитела полипептид, предпочтительно, объединяют с константной областью тяжелой цепи антитела. Таким образом, настоящее изобретение, кроме того, относится к тяжелой цепи антитела, содержащей полипептид формулы (I). В данной области техники известно много производных и аналогов антител с вариабельными областями. При этом, в настоящее время многие различные части, производные и/или аналоги антител используются. Неограничивающими примерами таких частей, производных и/или аналогов являются одноцепочечные фрагменты Fv, монотела, VHH, фрагменты Fab, искусственные связывающие белки, такие как, например, авимеры, и т.п. Общей чертой таких специфических связывающих веществ является наличие вариабельной области тяжелой цепи. Таким образом, настоящее изобретение, кроме того, относится к связывающей конструкции, содержащей полипептид формулы (I).
Предпочтительной связывающей конструкцией по настоящему изобретению является антитело, поскольку антитело содержит природную структуру. Следовательно, настоящее изобретение в предпочтительном варианте осуществления относится к антителу, содержащему полипептид по настоящему изобретению.
Связывающая конструкция по настоящему изобретению, предпочтительно, представляет собой связывающую конструкцию, которая хорошо переносится животным. Толерантность животного в отношении полипептида регулируется многими различными аспектами. Иммунитет, опосредуемый Т-клетками, опосредуемый В-клетками или другими, является одним из параметров, которые включены в толерантность животного в отношении полипептида. Как отмечено выше, антитело 5D12 имеет мышиное происхождение. Полипептид формулы (I) обладает пониженной иммуногенностью для человека. Поэтому он иногда указан как деиммунизированный вариант вариабельной области тяжелой 5D12. Таким образом, в одном из аспектов настоящее изобретение относится к антителу, обладающему эпитопной специфичностью антитела 5D12, причем тяжелая цепь указанного антитела представляет собой полипептид формулы (I). Как используется в настоящем описании, термин «деиммунизированный» определяется как менее иммуногенный для животного, чем исходное антитело. Полипептид формулы (I) является деиммунизированным по сравнению с тяжелой цепью в 5D12 благодаря удалению известных Т-клеточных эпитопов человека. Т-клеточные эпитопы являются аминокислотными последовательностями внутри белков, способными связывать молекулы МНС (главного комплекса гистосовместимости) класса II. После удаления Т-клеточных эпитопов антитело становится менее иммуногенным. Предпочтительно, вариабельная область по настоящему изобретению является, кроме того, гуманизированной, например, венированной. При использовании методов венирования остатки внешней части, которые в изобилии представлены для иммунной системы, избирательно заменяют остатками человека для обеспечения гибридной молекулы, которая обладает либо слабо иммуногенной, либо по существу неиммуногенной венированной поверхностью. Используемым в настоящем изобретении животным, предпочтительно, является млекопитающее, более предпочтительно, примат, наиболее предпочтительно, человек.
Антитело по настоящему изобретению, предпочтительно, содержит константную область антитела человека. В соответствии с различиями в константных областях их тяжелых цепей антитела группируют в пять классов или изотипов: IgG, IgA, IgM, IgD и IgE. Эти классы или изотипы включают по крайней мере одну из указанных тяжелых цепей, которую обозначают соответствующей греческой буквой. В предпочтительном варианте осуществления настоящее изобретение относится к антителу по настоящему изобретению, в котором указанная константная область выбрана из группы константных областей IgG, IgA, IgM, IgD и IgE, более предпочтительно, указанная константная область включает константную область IgG, более предпочтительно, константную область IgG1, предпочтительно, мутированную константную область IgG1, наиболее предпочтительно, указанная константная область представляет собой константную область IgG4. Более того, указанной константной областью IgG4 является, предпочтительно, константная область IgG4 человека. Предпочтительно, антитело IgG4 по настоящему изобретению содержит константные области аминокислотной последовательности тяжелой и легкой цепи, изображенной на фиг.18. Предпочтительно, антитело IgG4 по настоящему изобретению содержит аминокислотную последовательность тяжелой и легкой цепи, изображенную на фиг.18. Некоторая вариация константной области IgG4 происходит в природе и/или допускается без изменения иммунологических свойств получаемого в результате антитела. Как правило, в константной области допускаются приблизительно 1-5 аминокислотных замен. Антитело с константной областью IgG4 или мутированной константной областью IgG1 обладает по крайней мере большей частью фармакологических свойств антитела, но не связывает комплемент и не будет, следовательно, вызывать истощение клеток, с которыми оно связывается in vivo. Предпочтительно, указанной константной областью является константная область антитела человека.
В одном из вариантов осуществления настоящее изобретение относится к нуклеиновой кислоте, кодирующей полипептид по настоящему изобретению и/или связывающую конструкцию по настоящему изобретению, и/или антитело по настоящему изобретению. Используемой в настоящем изобретении нуклеиновой кислотой является, как правило, но не исключительно, рибонуклеиновая кислота (РНК) или дезоксирибонуклеиновая кислота (ДНК). Для квалифицированного в данной области техники специалиста доступны альтернативные нуклеиновые кислоты, такие как, например, пептидо-нуклеиновые кислоты (ПНК). Нуклеиновую кислоту по настоящему изобретению, например, вводят в клетку. Когда указанная нуклеиновая кислота экспрессируется в указанной клетке, то указанная клетка продуцирует полипептид и/или связывающую конструкцию, и/или антитело по настоящему изобретению. Следовательно, настоящее изобретение в одном из вариантов осуществления относится к клетке, содержащей полипептид по настоящему изобретению, связывающую конструкцию по настоящему изобретению, антитело по настоящему изобретению и/или нуклеиновую кислоту по настоящему изобретению. Указанной клеткой, предпочтительно, является клетка животного, более предпочтительно, клетка млекопитающего, более предпочтительно, клетка примата, наиболее предпочтительно, клетка человека. Для целей настоящего изобретения подходящей клеткой является любая клетка, способная включать и, предпочтительно, продуцировать полипептид по настоящему изобретению, связывающую конструкцию по настоящему изобретению, антитело по настоящему изобретению и/или нуклеиновую кислоту по настоящему изобретению.
Кроме того, настоящее изобретение относится к клетке, содержащей антитело по настоящему изобретению. Предпочтительно, указанная клетка продуцирует указанное антитело. В предпочтительном варианте осуществления указанной клеткой является гибридомная клетка, клетка яичника китайского хомячка (СНО), клетка NSO или клетка PER-C6TM. В особенно предпочтительном варианте осуществления указанной клеткой является клетка СНО. Кроме того, изобретение относится к культуре клеток, содержащей клетку по настоящему изобретению. Различными институтами и компаниями были разработаны линии клеток для высокомасштабной продукции антител, например, для клинического применения. Неограничивающими примерами таких линий клеток являются клетки СНО, клетки NSO или клетки PER.C6TM. Эти клетки также можно использовать для других целей, например, для продукции белков. Линии клеток, разработанные для продукции в промышленном масштабе белков и антител, указаны в настоящем описании как промышленные линии клеток. Таким образом, в предпочтительном варианте осуществления настоящее изобретение относится к применению линии клеток, разработанной для высокомасштабной продукции белка и/или антитела, для продукции антитела по настоящему изобретению.
Кроме того, настоящее изобретение относится к способу продуцирования антитела, включающему культивирование клетки по настоящему изобретению и сбор указанного антитела из указанной культуры. Предпочтительно, указанную клетку культивируют в бессывороточной среде. Предпочтительно, указанная клетка адаптирована для роста в суспензии. Кроме того, изобретение относится к антителу, получаемому способом продуцирования антитела по настоящему изобретению. Антитело, предпочтительно, очищают из среды культуры. Предпочтительно, указанное антитело подвергают аффинной очистке.
Клетка по настоящему изобретению представляет собой, например, линию гибридомных клеток, клетку СНО, клетку NSO или другой тип клеток, которые подходят для продукции антител для клинических целей. В особенно предпочтительном варианте осуществления указанной клеткой является клетка человека, предпочтительно, клетка, трансформированная областью Е1 аденовируса или его функциональным эквивалентом. Предпочтительным примером такой линии клеток является линия клеток PER.C6TM или ее эквивалент. В особенно предпочтительном варианте осуществления указанной клеткой является клетка СНО или ее вариант, предпочтительно, вариант, который использует глутаминсинтетазную (GS) векторную систему для экспрессии антитела.
Отмечено, что некоторые аминокислоты в упоминаемых положениях Х1-Х5 меньше подходят для продукции на высоком уровне антитела, включающего полипептид формулы (I), в продуцирующей антитело клетке. В предпочтительном варианте осуществления полипептид формулы (I) в указанном антителе содержит Х1-Х5, причем
Х1 представляет собой G, A, V, L, I, P, F, M, W, C, N, Q, S, T, Y, D, E, K, R или H. В особенно предпочтительном варианте осуществления Х1 представляет собой G, A, V, L, I, P, F или M; Х2 представляет собой G, A, V, L, I, P, F или M; Х3 представляет собой G, A, V, L, I, P, F, M; Х4 представляет собой G, A, V, L, I, P, F, M; и Х5 представляет собой G, A, V, L, I, P, F, M, W, C, N, Q, S, T или Y. Более предпочтительно, указанное антитело включает полипептид формулы (I), где Х1 представляет собой G, A, V, L, I или M; Х2 представляет собой G, A, V, L, I или M; Х3 представляет собой G, A, V, L, I, P, F, M; Х4 представляет собой G, A, V, L, I или M; и Х5 представляет собой P, F, W, N, Q, S, T или Y. Более предпочтительно, Х1 представляет собой L; Х2 представляет собой I; Х3 представляет собой P; Х4 представляет собой M; и/или Х5 представляет собой S. Особенно предпочтительно, когда Х1 представляет собой I, а Х2 представляет собой V; Х1 представляет собой I, а Х2 представляет собой I; Х1 представляет собой L, а Х2 представляет собой I; Х1 представляет собой L, а Х2 представляет собой L; Х1 представляет собой V, а Х2 представляет собой I; Х1 представляет собой V, а Х2 представляет собой V; Х1 представляет собой L, а Х2 представляет собой L; Х1 представляет собой V, а Х2 представляет собой L; или Х1 представляет собой L, а Х2 представляет собой V. Эти последние полипептиды особенно предпочтительны в комбинации с Х3, которая представляет собой P; Х4, которая представляет собой M; и Х5, которая представляет собой или F, или S, предпочтительно, S. В одном из вариантов осуществления настоящее изобретение относится к полипептиду по настоящему изобретению, в котором Х1 представляет собой L; Х2 представляет собой V; Х3 представляет собой L; Х4 представляет собой L; и Х5 представляет собой F. Продукция антител, включающих указанный полипептид, является хорошей при параллельном обеспечении улучшенных по сравнению с ch5D12 иммунологических свойств у людей.
В другом предпочтительном варианте осуществления настоящее изобретение относится к полипептиду по настоящему изобретению, в котором по крайней мере одна из Х1, Х2, Х3, Х4 или Х5 является такой же аминокислотой, что и аминокислота в соответствующем положении последовательности, которая, как продемонстрировано в настоящем изобретении, дает действительно значительные уровни экспрессии, и где, кроме того, по крайней мере одна из Х1, Х2, Х3, Х4 или Х5 является такой же аминокислотой, что и аминокислота в соответствующем положении аминокислотной последовательности 5D12. Преимущество полипептида по настоящему изобретению, в котором по крайней мере одна из Х1, Х2, Х3, Х4 или Х5 является такой же, что и аминокислота в соответствующем положении аминокислотной последовательности 5D12, состоит в том, что связывающая конструкция по настоящему изобретению, включающая такой полипептид, обладает лучшим уровнем экспрессии, чем полипептид по настоящему изобретению, где ни одна из Х1, Х2, Х3, Х4 или Х5 не является такой же аминокислотой, что и аминокислота в соответствующем положении аминокислотной последовательности 5D12. Было высказано не связанное с какой-либо теорией предположение, что такой высокий уровень экспрессии обусловлен тем фактом, что аминокислота в положении Х1, Х2, Х3, Х4 и/или Х5 способствует правильной сборке связывающей конструкции по настоящему изобретению, если указанная аминокислота является такой же аминокислотой, что и аминокислота в соответствующем положении аминокислотной последовательности 5D12.
Антитело, содержащее полипептид формулы (I), обладает нестимуляторной антагонистической активностью. Поскольку взаимодействие CD40L-CD40 вовлечено в патофизиологию различных воспалительных заболеваний, таких как аутоиммунные заболевания и отторжение трансплантата, то полипептид по настоящему изобретению, следовательно, подходит, в частности, для уменьшения интенсивности симптома воспалительного заболевания. В одном из вариантов осуществления антитело содержит связывающую конструкцию по настоящему изобретению. В предпочтительном варианте осуществления указанным антителом является моноклональное антитело. Технология моноклональных антител позволяет продуцировать большие количества по существу чистых антител, получая, таким образом, предсказанные продукты. Следовательно, настоящее изобретение в одном из вариантов осуществления относится к моноклональному антителу-антагонисту против CD40 человека, содержащему полипептид по настоящему изобретению. Связывающая конструкция по настоящему изобретению больше подходит для этой цели, так как она в одном из вариантов осуществления является деиммунизированной по сравнению с мышиным 5D12 и/или химерным 5D12. Следовательно, связывающая конструкция по настоящему изобретению обладает сниженной иммуногенностью и повышенным полупериодом существования у людей по сравнению с мышиным 5D12 и/или химерным 5D12. В результате этого связывающая конструкция по настоящему изобретению имеет устойчивую фармацевтическую эффективность против различных воспалительных заболеваний. Таким образом, в предпочтительном варианте осуществления настоящее изобретение относится к деиммунизированному моноклональному антителу-антагонисту против CD40 по настоящему изобретению.
Как ранее было указано, настоящее изобретение относится к подобным 5D12 молекулам, которые содержат модификации аминокислот по сравнению с аминокислотной последовательностью 5D12, причем модификации присутствуют по крайней мере в вариабельной области тяжелой цепи и, предпочтительно, также в вариабельной области легкой цепи. В этом контексте настоящее изобретение, кроме того, относится к связывающей конструкции по настоящему изобретению, включающей аминокислотную последовательность формулы (II)

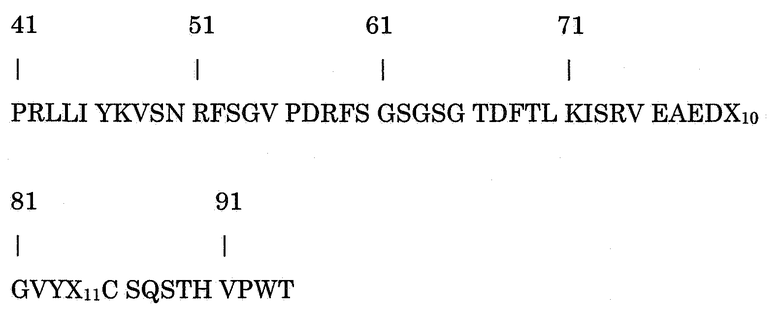
В которой
Х6 представляет собой G, A, V, L, I, P, F, M, W, C, N, Q, S, T, Y, D, E, K, R или H;
Х7 представляет собой G, A, V, L, I, P, F, M, W, C, N, Q, S, T, Y, D, E, K, R или H;
Х8 представляет собой G, A, V, L, I, P, F, M, W, C, N, Q, S, T, Y, D, E, K, R или H;
Х9 представляет собой G, A, V, L, I, P, F, M, W, C, N, Q, S, T, Y, D, E, K, R или H; и
Х10 представляет собой G, A, V, L, I, P, F, M, W, C, N, Q, S, T, Y, D, E, K, R или H, и
Х11 представляет собой G, A, V, L, I, P, F, M, W, C, N, Q, S, T, Y, D, E, K, R или H.
Указанная связывающая композиция, предпочтительно, представляет собой моноклональное антитело-антагонист против CD40 человека по настоящему изобретению. В предпочтительном варианте осуществления Х, который выбирают из Х6, Х7, Х8, Х9, Х10 или Х11, выбран из группы, аналогичной аминокислоте в соответствующем положении в аминокислотной последовательности 5D12 и/или аналогичной аминокислоте в соответствующем положении в последовательности, которая, как показано в настоящий заявке, дает высокие уровни экспрессии, как показано в примере 2. Поэтому связывающая конструкция по настоящему изобретению, предпочтительно, содержит аминокислотную последовательность формулы (II), в которой
Х6 представляет собой N, Q, S, T, Y, W или C;
Х7 представляет собой D, E, N, Q, S, T, Y, W или C;
Х8 представляет собой N, Q, S, T, Y, G, A, V, L, I, P, F, M, W или C;
Х9 представляет собой G, A, V, L, I, P, F, M;
Х10 представляет собой G, A, V, L, I, P, F, M; и
Х11 представляет собой N, Q, S, T, Y, G, A, V, L, I, P, F, M, W или C.
В другом варианте осуществления настоящее изобретение относится к моноклональному антителу-антагонисту против CD40 человека по настоящему изобретению, содержащему полипептид формулы (II) по настоящему изобретению, содержащему следующую аминокислотную последовательность:

В предпочтительном варианте осуществления настоящее изобретение относится к полипептиду формулы (II) по настоящему изобретению, в котором Х6 представляет собой Т или S, Х7 представляет собой D или Q, Х8 представляет собой Q или P, Х9 представляет собой V или А, Х10 представляет собой V или L, и Х11 представляет собой F или Y. Более предпочтительно, Х6 представляет собой Т, Х7 представляет собой Q, Х8 представляет собой P, Х9 представляет собой А, Х10 представляет собой V, и Х11 представляет собой Y. Подобные 5D12 антитела, содержащие полипептид формулы (I) и предпочтительный полипептид формулы (II), сочетают высокие уровни экспрессии в продуцирующей клетке и хорошую переносимость, а также хорошие фармакодинамические свойства у человека.
Антитело-антагонист против CD40 человека по настоящему изобретению, предпочтительно, содержит аминокислотную последовательность вариабельной области тяжелой цепи формулы (I) и аминокислотную последовательность вариабельной области легкой цепи формулы (II). Такое антитело имеет хорошие характеристики. Безусловно, можно создать варианты такого исходного антитела путем модифицирования одной или нескольких аминокислот в нем. Активность многих из таких вариантов будет более или менее аналогично указанному исходному антителу. Такие варианты также входят в объем настоящего изобретения. Существует множество способов модификации антитела по настоящему изобретению. Неограничивающим примером такой модификации является антитело, содержащее пироглутамат вместо глутамата. Другими неограничивающими примерами таких модификаций являются вставка, делеция, инверсия и/или замена одной или нескольких аминокислот по сравнению с указанным исходным антителом. Настоящее изобретение относится к способам и методам создания такого варианта. Также изобретение относится к тестам для определения характеристик указанного варианта. В предпочтительном варианте осуществления настоящее изобретение относится к варианту исходного антитела по настоящему изобретению, содержащему вставку, делецию, инверсию и/или замену приблизительно 1-10 аминокислот по сравнению с аминокислотной последовательностью указанного исходного антитела. Предпочтительно, указанные вставка, делеция, инверсия и/или замена не включают аминокислоты в положении Х1 и положении Х2 вариабельной области тяжелой цепи формулы (I) исходного антитела.
В предпочтительном варианте осуществления настоящее изобретение относится к способу отбора антитела-антагониста против CD40 человека, включающему создание первой линии клеток, которая продуцирует исходное антитело-антагонист против CD40 человека, и определение количества исходного антитела, которое продуцируется указанной первой линией клеток, при этом указанное исходное антитело содержит аминокислотную последовательность вариабельной области тяжелой цепи


в которой Х1 и Х2 попарно выбирают из группы, состоящей из Х1 = I и Х2 = V; Х1 = L и Х2 = I; Х1 = V и Х2 = V; Х1 = L и Х2 = L; или Х1 = L и Х2 = V,
при этом указанный способ, кроме того, включает
создание по крайней мере одной дополнительной линии клеток, которая продуцирует вариант указанного исходного антитела, причем указанный вариант антитела представляет собой модифицированное исходное антитело, содержащее вставку, делецию, инверсию и/или замену приблизительно 1-5 аминокислот по сравнению с указанным исходным антителом, причем указанная модификация не состоит из модификации аминокислот в положении(ях), определяемых Х1 и Х2, и определение количества варианта антитела, который продуцирует указанная по крайней мере одна из дополнительных линий клеток, при этом указанный способ, кроме того, включает отбор варианта антитела, который был продуцирован в количестве, которое составляло по крайней мере 50% количества исходного антитела. Предпочтительно, указанное исходное антитело содержит аминокислотную последовательность легкой цепи

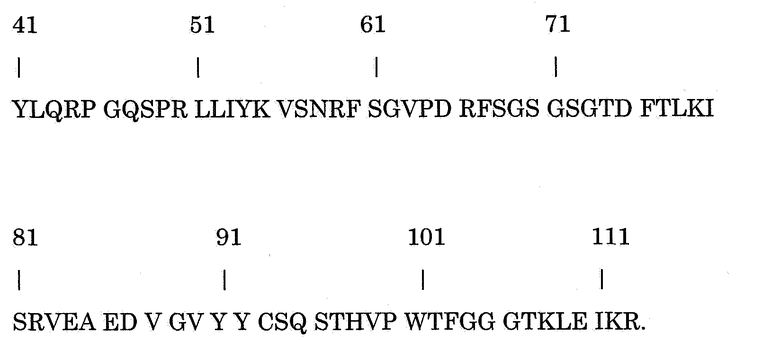
Указанные вставка, делеция, инверсия и/или замена приблизительно 1-5 аминокислот могут быть в любой части антитела, которая не является или не включает аминокислоты в положениях Х1 и Х2. Предпочтительно, указанные вставка, делеция, инверсия и/или замена приблизительно 1-5 аминокислот находятся в указанной аминокислотной последовательности тяжелой цепи или указанной аминокислотной последовательности легкой цепи по сравнению с аминокислотной последовательностью соответствующей цепи в указанном исходном антителе. Предпочтительно, указанные вставка, делеция, инверсия и/или замена приблизительно 1-5 аминокислот находятся в указанной аминокислотной последовательности тяжелой цепи по сравнению с указанной последовательностью тяжелой цепи указанного исходного антитела. Предпочтительно, указанный способ, кроме того, включает создание линии клеток - продуцента антитела, которая продуцирует указанное отобранное антитело. Такая линия клеток - продуцент антитела может быть указанной дополнительной линией клеток или даже другой линией клеток, которая продуцирует указанное отобранное антитело. Предпочтительно, указанный способ, кроме того, включает сбор указанного отобранного антитела. Кроме того, настоящее изобретение относится к выделенному и/или рекомбинантному антителу-антагонисту против CD40 человека, получаемому способом по настоящему изобретению. В предпочтительном варианте осуществления указанное антитело-антагонист против CD40 человека содержит модификацию аминокислотной последовательности тяжелой цепи

В которой Х1 и Х2 попарно выбирают из группы, состоящей из Х1 = I и Х2 = V; Х1 = L и Х2 = I; Х1 = V и Х2 = V; Х1 = L и Х2 = L; или Х1 = L и Х2 = V,
при этом указанная модификация включает вставку, делецию, инверсию и/или замену приблизительно 1-5 аминокислот по сравнению с указанной аминокислотной последовательностью тяжелой цепи, причем указанная модификация не состоит из модификации аминокислот в положении(ях), определяемом Х1 и Х2.
В одном из вариантов осуществления настоящее изобретение относится к фармацевтической композиции, содержащей полипептид по настоящему изобретению, связывающее вещество по настоящему изобретению, антитело по настоящему изобретению, нуклеиновую кислоту по настоящему изобретению и/или клетку по настоящему изобретению. Также настоящее изобретение относится к полипептиду по настоящему изобретению, связывающему веществу по настоящему изобретению, антителу по настоящему изобретению, нуклеиновой кислоте по настоящему изобретению и/или клетке по настоящему изобретению для применения в качестве лекарственного средства, предпочтительно, лекарственного средства для уменьшения интенсивности симптома аутоиммунного заболевания и/или воспалительного заболевания и/или уменьшения отторжения трансплантата, и/или для лечения CD40-положительных раков. В предпочтительном варианте осуществления аутоиммунное и/или воспалительное заболевание выбрано из группы, состоящей из воспалительного заболевания кишечника, ревматоидного артрита, системной красной волчанки, рассеянного склероза, псориаза, буллезного пемфигоида и атопического дерматита.
Поскольку полипептид по настоящему изобретению, в частности, подходит для уменьшения интенсивности симптома воспалительного заболевания из-за его нестимулирующих антагонистических в отношении CD40 свойств, то полипептид по настоящему изобретению подходит для уменьшения интенсивности симптома нескольких нарушений. Используемый в настоящем изобретении термин «воспалительное заболевание» определяется как любое заболевание, в которое вовлечен воспалительный компонент. В интересах настоящего изобретения воспалительное заболевание, в частности, включает аутоиммунное заболевание и/или отторжение трансплантата. Центральную роль во взаимодействии CD40-CD40L для инициации, усиления и пролонгации иммунных ответов играет полипептид по настоящему изобретению, в частности, он подходит для иммуномодуляции аутоиммунного заболевания.
Далее приведена информация о CD40 и CD40L для иллюстрации роли CD40 и его лиганда в воспалительных заболеваниях. Молекула CD40 представляет собой мембранный гликопротеин типа I с молекулярной массой 50 кДа и экспрессируется на В-клетках, моноцитах/макрофагах и дендритных клетках (DC).45-50 Кроме того, при патологических состояниях CD40 можно обнаружить на эндотелиальных клетках (ЕС), фибробластах, эпителиальных клетках и кератиноцитах.51 Лиганд CD40 (gp39, TBAM, TRAP, CD40L, CD154), интегральный мембранный гликопротеин типа II с молекулярной массой 32 кДа, транзиторно экспрессируется на активированных Т-клетках CD4+ и небольшой популяции активированных Т-клеток CD8+.52,53 Кроме того, CD40L обнаружен на ряде других типов клеток после активации, включая тучные клетки, базофилы, В-клетки, эозинофилы, DC и тромбоциты.54,55
Контактирование CD40 и CD40L запускает ряд биологических событий в В-клетках, в том числе пролиферацию, экспрессию маркеров активации, продукцию иммуноглобулинов (Ig), переключение изотипов, гомологичную адгезию и спасение от апоптоза.56,57 Однако, как описано выше, распространение молекулы CD40 не ограничивается В-клетками, как первоначально утверждалось. Свежевыделенные моноциты человека экспрессируют низкие уровни молекулы CD40, которые можно повысить при культивировании в присутствии IFN-γ.47-49,58 Связывание CD40 моноцитов/макрофагов индуцирует секрецию больших количеств провоспалительных медиаторов, таких как IL-1, TNF-α и IL-12, которые индуцируют воспалительные реакции и обладают уничтожающей опухолевые клетки активностью,47-49,58, а также спасают их от апоптоза.48 Связывание CD40 также вызывает усиление дифференциации и активации DC, увеличение экспрессии костимулирующих молекул, таких как CD86, CD80 и CD58, увеличение продукции цитокинов и ингибирование апоптоза.50,59 Кроме того, при экспрессии в условиях воспаления передача сигнала от CD40 может индуцировать экспрессию молекулы 1 межклеточной адгезии (ICAM-1), молекулы 1 адгезии к клеткам сосудов (VCAM-1) и Е-селектина на ЕС.55 Эти результаты говорят о том, что передача сигнала через CD40 во время взаимодействий Т-клетка-ЕС может быть важным этапом при регуляции активации ЕС и привлечения лейкоцитов в нелимфоидные ткани. In vivo исследования показали важность взаимодействий CD40-CD40L в выработке гуморальных иммунных ответов,60,61 в примировании и активации антиген-специфических Т-клеток,62 во временной активации макрофагов,63 а также в защитных клеточно-опосредуемых иммунных ответах через опосредуемую Т-клетками активацию макрофагов против внутриклеточного инфицирования паразитирующими организмами, такими как Pneumocystis, Cryptosporidium и Leishmania.64-66
Специфическая роль CD40 и его лиганда показана на мышиных моделях аутоиммунного заболевания. Исследования на животных моделях ясно показали вовлеченность взаимодействия CD40L-CD40 в патофизиологию различных аутоиммунных заболеваний. В этих исследованиях, используя мышей, страдающих естественными или экспериментальными аутоиммунными заболеваниями, препятствование взаимодействиям CD40L-CD40 оказывало явно благотворные эффекты. Показано, что мАт против CD40L мыши предупреждает или уменьшает симптомы заболевания при индуцированном коллагеном артрите, экспериментальном аллергическом энцефаломиелите (ЕАЕ; в модели MS на животном), у мышей (SWR x NZB)F1 с обыкновенной волчанкой и у мышей с диабетом без ожирения (NOD), у которых спонтанно развились зависимые от Т-клеток аутоиммунные диабеты. Данные указывают на то, что взаимодействия CD40-CD40L также играют роль в патогенезе воспалительных заболеваний кишечника, которые включают болезнь Крона и язвенный колит. Показано, что трансгенные в отношении CD40L мыши с большим числом копий трансгена приобретают смертельное воспалительное заболевание кишечника, отмеченное инфильтрацией клеток CD40+ и Т-клеток CD40L+ в больные ткани.67 мАт против CD40L эффективно предотвращают воспаление слизистых оболочек и продукцию IFN-γ Т-клетками CD4+ собственного слоя у животных с колитом, который был индуцирован 2,4,6-тринитробензолсульфоновой кислотой.44 Совсем недавно провели прямое сравнение между лечением анти-TNF-α и помехой пути CD40-CD40L, используя мАт против CD40L, на модели экспериментального воспалительного заболевания кишечника на мышах с SCID. В этой модели сингенные клетки CD45RBhigh CD4+ вводят мышам с SCID, у которых впоследствии развивается диарея или жидкий стул и у которых отмечается прогрессивное снижение веса, начиная с 3-5 недели после восстановления Т-клеток, в качестве симптомов экспериментального воспалительного заболевания кишечника. Лечение или анти-TNF-α, или анти-CD40L со дня восстановления Т-клеток полностью предотвращало клиническое и гистологическое проявление экспериментального воспалительного заболевания кишечника. Кроме того, введение анти-CD40L с 5 недели после восстановления Т-клеток могло предотвратить прогрессирование заболевания, и у получавших лечение животных наблюдалось улучшение симптомов заболевания и гистологии по сравнению с контрольными животными (неопубликованные наблюдения).
В недавней работе было продемонстрировано, что препятствование пути CD40-CD40L приводит к сильной иммуносупрессии в моделях трансплантации. Комбинированное лечение аллогенными небольшими лимфоцитами или истощенными в отношении Т-клеток небольшими лимфоцитами плюс антителом против CD40L мыши делало возможным выживание аллотрансплантата в виде неопределенных островков Лангерганса у 37 из 40 реципиентов, отличающихся по основным и неосновным локусам гистосовместимости.68 На основании этих экспериментов было сделано заключение, что эффективное препятствование взаимодействию CD40L-CD40, наиболее вероятно, приводило к предотвращению индукции костимулирующих молекул на небольших покоящихся лимфоцитах аллореактивными Т-клетками хозяина. В другом недавнем исследовании продемонстрировано, что введение мАт против CD40L мыши во время трансплантации заметно пролонгировало выживание полностью различных мышиных сердечных аллотрансплантатов как у несенсибилизированных, так и у сенсибилизированных хозяев. Однако если анти-CD40L терапию откладывали до 5 дня после операции, то анти-CD40L не пролонгировало выживание трансплантата. Из этого исследования было сделано заключение, что анти-CD40L терапия ингибировала отторжение аллотрансплантатов, главным образом, путем препятствования помощи Т-клеток для эффекторных функций. Кроме того, было показано, что препятствование путям CD80/CD86-CD28 и CD40-CD40L одновременно эффективно прекращает Т-клеточное клональное увеличение in vitro и in vivo, повышает долгосрочное выживание полностью аллогенных трансплантатов кожи и ингибирует развитие хронического васкулярного отторжения исходно васкуляризированных сердечных аллотрансплантатов. Кроме того, препятствование пути CD40-CD40L, необязательно в комбинации с препятствованием пути CD80/CD86-CD28, предотвращает отторжение почечного аллотрансплантата на модели почечного аллотрансплантата на макаке резус.69,70 Дополнительная информация в отношении эффектов 5D12 на воспалительные заболевания приведена, например, в ссылках71-81.
Рассеянный склероз представляет собой аутоиммунное заболевание центральной (цереброспинальной) нервной системы. При этом нарушении белое вещество, окружающее нервные волокна, затвердевает. Термин «рассеянный склероз» (MS) буквально означает «множество рубцов». Затвердевшие области нервной ткани называются бляшками. Симптомы, тяжесть и течение рассеянного склероза являются в высокой степени непостоянными и зависят отчасти от мест нахождения бляшек и степени поражения белого вещества. Поражение белого вещества нервной системы замедляет нервные импульсы, что приводит к несогласованности нервной системы.
Экспериментальный аутоиммунный энцефаломиелит (ЕАЕ) у обыкновенной игрунки (Callithriz jacchus) представляет собой полезную преклиническую модель рассеянного склероза (обзор которой приведен в ссылках86,87,102,104). Поражения белого вещества центральной нервной системы (ЦНС), которые развиваются при различных вариантах этой модели ЕАЕ, имеют общие с MS патоморфологические, радиологические и иммунологические признаки.95,98,101 Следовательно, модель ЕАЕ на игрунке может служит мостиком в широком иммунологическом зазоре между людьми и грызунами, который затрудняет отбор перспективных лекарственных средств в конвейере их разработок на преклинической стадии.96,99,103
У игрунок, иммунизированных rhMOG, рекомбинантным белком, представляющим собой внеклеточный фрагмент MOG человека (аминокислоты 1-125), развивается ЕАЕ в 100% случаев, что обусловлено наличием мономорфного элемента чувствительности МНС класса II Caja-DRB∗W1201 в репертуаре каждой игрунки.82,84,85,107 Особенно эффективным аспектом этой модели для разработки терапии является то, что поражения, развивающиеся в белом веществе головного мозга, можно визуализировать и в порядке эксперимента охарактеризовать с помощью клинически релевантных методов отображения магнитного резонанса.92,105 Продольный анализ поражений белого вещества головного мозга с помощью отображения магнитного резонанса (MRI) продемонстрировал прогрессирующее увеличение объема и хронической воспалительной активности в большинстве поражений. Более того, с помощью характеристики патологии ЦНС с использованием MRI и ранее описанных гистологических критерией101 обнаружено, что большинство поражений находятся на ранней активной стадии.106
Модель rhMOG-индуцированного ЕАЕ использовали для проверки того, являются ли антитела, нацеленные на костимулирующие молекулы антигенпрезентирующих клеток (АРС) и Т-клеток, возможным лечением для MS. Взаимодействие CD40 с его лигандом CD154 играет важную роль при различных иммунопатогенных процессах, которые функционируют при ЕАЕ, включающих активацию В-клеток, активацию антигенпрезентирующих клеток (АРС), возникновение ответов в виде антиген-специфических Т-клеток и индукцию эффекторных функций макрофагов.90,93,97,7 Исследование, проведенное в 1996 г., подтвердило, что мыши, получавшие лечение антителом против CD154, защищены от ЕАЕ.88. Однако клиническое испытание на больных MS с использованием антитела против CD154 было прекращено из-за неожиданных побочных эффектов, которые не наблюдались в экспериментах на животных.17
Моноклональное антитело мыши (мАт) 5D12 (mu5D12) было индуцировано против CD40 человека. Антитело 5D12 оказалось сильным ингибитором опосредуемой CD40-CD40L активации в нескольких типах клеток и, в отличие от большинства других мАт против CD40, не проявляет CD40-стимулирующей активности.21,22,39 Как мышиное антитело против CD40 человека mu5D12, так и химерный вариант этого антитела ch5D12 продемонстрировали сильные супрессивные эффекты на развитие поражений белого вещества ЦНС и неврологические расстройства в модели ЕАЕ на игрунке и не продемонстрировали отмечаемых побочных эффектов.23,24 Те же исследования показали, что внутривенно вводимое мАт против CD40 пораженным ЕАЕ обыкновенным игрункам может проходить к поражениям белого вещества головного мозга, где молекулы CD40 отчетливо экспрессированы на проникших макрофагах и активированной микроглии,95 что было обнаружено ранее при MS.88 Это подняло вопрос, касающийся того, оказывает ли ch5D12 также терапевтический эффект на уже существующие поражения.
В статье 't Hart и др. описан контроль развития поражения головного мозга у 7 иммунизированных rhMOG мартышек с помощью последовательного отображения магнитного резонанса (MRI) с интервалом в 2 недели.76 Результаты этого исследования продемонстрировали супрессию воспаления в месте поражения у всех 3 подвергнутых лечению ch5D12 мартышек, тогда как увеличение поражения было уменьшено у 2 из 3 подвергнутых лечению ch5D12 мартышек.
Предотвращение контактирования CD40 со своим лигандом CD154 на активированных Т-клетках рано в процессе заболевания оказывает значительное влияние на клиническое и нейропатологическое выражение ЕАЕ у грызунов.88,89,91,94,100 и в моделях на не являющихся людьми приматах.23,24 CD40 отчетливо экспрессируется в поражениях белого вещества в ЦНС больных MS, а также пораженных ЕАЕ грызунов88 и не являющихся людьми приматов.95 То, что несущие CD40 АРС в пределах ЦНС, такие как проникшие макрофаги, а также периваскулярные и паренхимные глиоциты, вносят значительный вклад в патогенез ЕАЕ, было изящно продемонстрировано на химерных в отношении костного мозга мышах.83
Результаты, показанные 't Hart и др., Laman и др. и Boon и др.76,79,81, указывают на то, что блокирование CD40 антителами является потенциально эффективным лечением MS. Важно, что мАт ch5D12 не имеет видимых побочных эффектов в модели ЕАЕ на игрунках, ни у других видов приматов.80 Благоприятный клинический эффект антитела против CD40 был продемонстрирован в контролируемых плацебо экспериментах на двух моделях ЕАЕ на игрунках, а именно, индуцированного миелином человека79 или rhMOG81. Кроме того, в статье 't Hart и др. указан ингибиторный эффект лечения антителом против CD40 на уже существующие поражения.76
Псориаз представляет собой воспалительное заболевание кожи, поражающее 1-2% популяции. При этом заболевании Т-клетки и кератиноциты в местах поражений активированы и экспрессируют маркеры активации и костимулирующие молекулы. Полагают, что некоторые костимулирующие молекулы, экспрессируемые на кератиноцитах и Т-клетках, взаимодействуют друг с другом, и что эти взаимодействия вносят вклад в активность заболевания.108-110 Одной такой совокупностью молекул может быть CD40, который экспрессируется на активированных кератиноцитах, и CD154 (лиганд CD40), который транзиторно экспрессируется на активированных Т-клетках CD4+. Связывание CD40-CD154 между Т-клетками и кератиноцитами может приводить к высвобождению из этих клеток медиаторов воспаления, которые наблюдаются в избытке в местах поражения псориазом. Недавно был представлен обзор CD40, CD154 и взаимодействий CD40-CD154.111
Культивируемые кератиноциты также экспрессируют CD40; экспрессия усиливается при обработке IFN. Связывание CD40, экспрессируемого в высокой степени на обработанных IFN-гамма кератиноцитах (упоминаемых на протяжении этой работы как кератиноциты CD40++), с CD154 приводит к увеличению ICAM и увеличению продукции цитокинов.112-114
До сих пор появилось только одно сообщение, которое показывает, что это связывание вовлечено в патогенез псориаза.113 Типы клеток, которые экспрессируют CD40, в местах поражений псориазом не были идентифицированы в последнем исследовании, и их частота и состояние поражения также не описаны. Также не исследовано присутствие Т-клеток CD154+ в местах поражения псориазом. Таким образом, остается неизвестным, действует ли CD154 в качестве одного из сигналов для продукции кератиноцитами хемокинов и комплемента, которые в избытке обнаруживаются в местах поражения псориазом.115
Недавно Pasch и др. продемонстрировали присутствие и локализацию клеток CD40+ и CD154+ с помощью иммуногистохимии в пораженной и непораженной коже, взятой у десяти больных псориазом.116 Увеличение позитивности в отношении CD40 наблюдалось на скоплениях кератиноцитов, и высокая экспрессия CD40 демонстрировалась на почти всех клетках Лангерганса CD1a+ в нормальном, непораженном и пораженном эпидермисе. Кроме того, высокая экспрессия CD40 была представлена на почти всех клетках CD83a+ в местах поражения псориазом; они редко наблюдались в непораженной и нормальной коже.116 Также небольшая часть Т-клеток продемонстрировала экспрессию CD154 у большинства пациентов в соединении с клетками CD40+. Эти результаты увеличивает возможность того, что Т-клетки CD154+ могут подвергаться связыванию с кератиноцитами CD40+, клетками Лангерганса и дендритными клетками CD83+ и высвобождать из них медиаторы в месте поражения. Кроме того, они продемонстрировали, что связывание CD40 индуцирует высвобождение хемокинов (IL-8, RANTES и MCP-1).116 В той же публикации Pasch и др. продемонстрировали, что связанное с CD40 высвобождение хемокинов IL-8, MCP-1 и в меньшей степени RANTES ингибировалось антителом-антагонистом против CD40 5D12.116 Эти данные говорят о том, что антагонистическое мАт против CD40 5D12 может по крайней мере частично оказывать эффект на воспаление, наблюдаемое в местах поражения псориазом.
В заявке US2003/0165499 сообщается об умеренном антипсориатическом эффекте 5D12 и других антител-антагонистов против CD40 в модельной системе ксеногенного трансплантата на мышах с SCID, которую используют в качестве модели для лечения псориаза, демонстрируя, что антитела-антагонисты против CD40 можно использовать для лечения псориаза. В этой in vivo системе был продемонстрирован терапевтический эффект 5D12.
В особенно предпочтительном варианте осуществления указанное аутоиммунное и/или воспалительное заболевание включает воспалительное заболевание кишечника. Другой предпочтительный вариант осуществления относится к применению настоящего изобретения, при котором указанное воспалительное заболевание кишечника включает язвенный колит (UC) или болезнь Крона (CD).
Как проиллюстрировано выше, полипептид по настоящему изобретению подходит для лечения разнообразных воспалительных заболеваний, включая аутоиммунные заболевания и отторжение трансплантата. Следовательно, настоящее изобретение в одном из вариантов осуществления относится к способу уменьшения интенсивности симптома аутоиммунного заболевания и/или воспалительного заболевания и/или уменьшения отторжения трансплантата. В дополнительном варианте осуществления настоящее изобретение относится к применению полипептида по настоящему изобретению, связывающей конструкции по настоящему изобретению, антитела по настоящему изобретению, нуклеиновой кислоты по настоящему изобретению и/или клетки по настоящему изобретению для получения лекарственного средства, уменьшающего интенсивность симптома аутоиммунного заболевания и/или воспалительного заболевания и/или уменьшающего отторжение трансплантата. Как используется в настоящем описании, уменьшение интенсивности симптома определяется как уменьшение интенсивности по крайней мере одного симптома заболевания по крайней мере частично. Настоящее изобретение представляет особый интерес для лечения аутоиммунных и/или воспалительных заболеваний, для которых в настоящее время не существует эффективного лечения. Примерами таких заболеваний являются ревматоидный артрит, системная красная волчанка, рассеянный склероз, псориаз, воспалительное заболевание кишечника, буллезный пемфигоид и атопический дерматит. Полипептид по настоящему изобретению, необязательно входящий в состав связывающей конструкции или клетку по настоящему изобретению и/или кодируемый нуклеиновой кислотой по настоящему изобретению, подходит, в частности, для уменьшения симптома аутоиммунного и/или воспалительного заболевания, поскольку определенные свойства полипептида по настоящему изобретению позволяют препятствовать пути CD40-CD40L специфическим образом. Кроме того, поскольку связывающая конструкция по настоящему изобретению, предпочтительно, является деиммунизированной, связывающая конструкция по настоящему изобретению присутствует в течение длительного периода времени и, таким образом, проявляет свою антагонистическую активность у пациента в течение значительного периода времени. Следовательно, настоящее изобретение в одном из вариантов осуществления относится к применению полипептида по настоящему изобретению, связывающей конструкции по настоящему изобретению, антитела по настоящему изобретению, нуклеиновой кислоты по настоящему изобретению и/или клетки по настоящему изобретению для получения лекарственного средства, уменьшающего интенсивность симптома аутоиммунного заболевания и/или воспалительного заболевания и/или уменьшающего отторжение трансплантата, причем указанное аутоиммунное и/или воспалительное заболевание выбрано из группы, состоящей из ревматоидного артрита, системной красной волчанки, рассеянного склероза, псориаза, воспалительного заболевания кишечника, буллезного пемфигоида и аллергического дерматита.
Краткое описание фигур
Фиг.1. Концентрация ch5D12 в сыворотке после однократного введения в день 0 четырех уровней доз, указанных символами. Значения приведены в виде мкг ch5D12/мл сыворотки и определялись с помощью иммуноферментного твердофазного анализа, детали которого приведены в ссылке27.
Фиг.2. Изменения показателей активности болезни Крона (CDAI) во время 28-дневного периода после инфузии ch5D12. Заштрихованная область указывает период, в который уровни ch5D12 превышают концентрацию 10 мкг/мл (см. также фиг.1), которая, как обнаружено, является функциональным антагонистическим уровнем в сыворотке у приматов.24-27 В 0,3 мг/кг-группе уровни в сыворотке никогда не превышали уровня 10 мкг/мл.
Фиг.3. Показатели гистологической активности заболевания в день 0 и день 28. Максимальный показатель активности составляет 16, и определение показателя активности проводили в соответствии со ссылкой40. В день 0 и день 28 получали образцы подвздошной кишки (ромбы) девяти субъектов и образцы ободочной кишки (квадраты) 11 субъектов, и результаты представляли для каждого уровня дозы ch5D12 [(а) 0,3 мг/кг; (b) 1,0 мг/кг; (c) 3,0 мг/кг и (d) 10,0 мг/кг.
Фиг.4. Образцы биопсии ободочной кишки пациента 012 (3,0 мг/кг) и биопсии подвздошной кишки пациента 011 (3,0 мг/кг) представлены в качестве примеров уменьшения воспалительной реакции. Образцы окрашивали антителами, узнающими все Т-лимфоциты (CD3), В-клетки (CD19), макрофаги (CD68) и клетки CD40+, до (а, с) и в день 28 после введения ch5D12. НЕ, гематоксилин-эозин.
Фиг.5. Консенсусная последовательность ДНК и выведенная аминокислотная последовательность областей VH и VL 5D12 мыши.
Фиг.6. Сравнение аминокислот мышиных V-областей с венированными и деиммунизированными V-областями 5D12.
Фиг.7. Выравнивание 12 проверяемых вариантов аминокислот (Q5E, K13A, E16Q, T17S, I29L, I37V, P45L, M48L, STS60NSA, T68S, S79F и T108S) деиммунизированной VH 5D12 с исходной мышиной последовательностью (ch5D12) и полностью деиммунизированной последовательностью (DI5D12).
Фиг.8. Анализ FACS с использованием клеток JY. Супернатант клеток PER.C6, транзиторно экспрессирующих варианты (Q5E, K13A, E16Q, T17S, I29L, I37V, P45L, M48L, STS60NSA, T68S, S79F и T108S) 5D12, собирали через 48 ч. В качестве контроля также собирали супернатант клеток, трансфицированных или ch5D12, или DI5D12, вместе с супернатантом от ложно (без плазмиды) трансфицированных клеток. Связывание экспрессированных антител исследовали с помощью FACS с использованием клеток JY вместе с античеловеческим, меченным FITS вторым антителом (разведенным 1/100). В качестве контроля FACS клетки JY инкубировали только со вторым антителом, меченным FITS.
Фиг.9. Выравнивание дополнительных вариантов V-L-I в положениях 29 и 37 (29I-37V, 29V-37I, 29I-37L, 29V-37V, 29L-37L, 29V-37L, 29L-37V) деиммунизированной VH 5D12 с исходной химерной последовательностью (ch5D12), полностью деиммунизированной последовательностью (DI5D12; 29I-37I) и PG102 (29L-37I).
Фиг.10. Распределение антительной продуктивности линий клеток GS-CHO, продуцирующих антитело PG102, и «переросших» культур в 24-луночных планшетах. Число линий клеток отложено по оси Х, диапазоны концентраций антител отложены по оси Y (диапазоны концентраций антител распределены в возрастающие группы, начиная с 0-25 мкг/мл).
Фиг.11. Профили роста и аккумуляции антитела для линий клеток: А) L107 (DC1; верхняя панель), В) L25 (DC2; средняя панель) и С) М95 (DC3; верхняя панель) в подпитываемой культуре CDACF во встряхиваемой колбе.
Фиг.12. Сравнение связывания PG102 (красные) и ch5D12 (голубые) при 10 мкг/мл с CD40, иммобилизованным на 3 различных уровнях.
Фиг.13. Ингибирование антиидиотипическим мАт 173-36-1 связывания мАт против CD40 с клетками JY, экспрессирующими CD40 человека. мАт против CD40 исследовали при концентрации 1 мкг/мл. Данные показывают среднее арифметическое ± значение стандартной ошибки 4 отдельных определений для каждого антитела.
Фиг.14. ELISA CD40-Fc. Зависящее от концентрации связывание ch5D12 (PG100) и PG102 с CD40-Fc человека. Планшеты в течение ночи покрывали 250 нг/мл CD40-Fc человека. мАт контрольного изотипа, chFUN-1, направлено против CD86 человека. Данные показывают среднее арифметическое ± значение стандартной ошибки (все n=3).
Фиг.15. Количественное определение с помощью FACS связывания мАт против CD40 с клетками JY. ch5D12 (PG100) и PG102 продемонстрировали сравнимые концентрации для полумаксимального связывания. мАт против CD86, chFUN-1, также продемонстрировало связывание с клетками JY в этих экспериментах вследствие экспрессии на поверхности этой линии клеток CD86 человека. Данные показывают среднее арифметическое двойных определений для каждого антитела.
Фиг.16. Ингибирование связывания ch5D12 (PG100)-РЕ и PG102-РЕ с клетками JY немечеными антителами ch5D12 (PG100) и PG102. Меченые ch5D12 (PG100) (А) и PG102 (В) инкубировали при 1 мкг/мл в присутствии увеличивающихся концентраций конкурирующего немеченого антитела. Связывание меченых антител определяли в виде средней интенсивности флуоресценции (MFI) с помощью проточной цитометрии. Максимальная MFI для связывания ch5D12 (PG100)-РЕ и PG102-РЕ составляла 369 единиц флуоресценции (А) и 305 единиц флуоресценции (В), соответственно. Данные показывают среднее арифметическое ± значение стандартной ошибки 4 отдельных экспериментов.
Фиг.17. Ингибирование высвобождения IL-8 клетками ТНР-1 после сокультивирования с клетками Jurkat. Клетки ТНР-1, стимулированные IFN-γ, сокультивируют с клетками Jurkat в присутствии увеличивающихся концентраций ch5D12 (PG100) или PG102. Высвобождение IL-8 из клеток ТНР-1, индуцированное связыванием клеток Jurkat с клетками ТНР-1 через CD40-CD40L, определяли с помощью ELISA. Данные показывают результат одного эксперимента.
Фиг.18. Последовательность нуклеиновой кислоты и аминокислотная последовательность тяжелой и легкой цепи IgG4 PG102.
ПРИМЕРЫ
Пример 1
Материалы и методы
Для включения в исследование отобрали восемнадцать взрослых субъектов (возрастом 18-60 лет) с клиническим диагнозом - болезнь Крона, подтвержденным рентгенологическими, эндоскопическими или гистологическими данными, и с показателем активности болезни Крона (CDAI), составляющим по крайней мере 220, но не больше 450 (отмечаемым на протяжении 7 дней до введения исследуемого лекарственного средства). Субъектам позволяли подвергаться следующим лечениям до и во время исследования: лечению мезаламином на протяжении 8 или более недель в дозе, остающейся постоянной на протяжении 4 недель до скринирования; максимум 30 мг кортикостероидов в день (или 9 мг будесонида в день) на протяжении 8 или более недель в дозе, остающейся постоянной на протяжении 2 недель до скринирования; меркаптопурином или азатиоприном на протяжении 4 или более месяцев в дозе, остающейся постоянной на протяжении 8 недель до скринирования. Субъекты не могли получать лечение циклоспорином А или метотрексатом в пределах 3 месяцев до скринирования, и не позволялось, чтобы они предварительно подвергались лечению мАт. Средний возраст всех зарегистрированных субъектов составлял 35,8 лет, из них семь субъектов были мужчинами, и 11 - женщинами. Все субъекты были кавказской этничности. Не наблюдалось видимых различий между пациентами в различных группах при включении (таблица 1) за исключением того, что в трех группах низких доз большинство субъектов были женщинами, в то время как группа наибольшей дозы (10,0 мг/кг) состояла только из являющихся мужчинами субъектов. Не было выдающихся различий между четырьмя группами в отношении электрокардиограммы (ECG), признаков жизни, физических обследований, истории признаков и симптомов болезни Крона базовой линии и лабораторных величин базовой линии. Не было значительных различий в отношении параметров базовой линии среди четырех групп доз. Однако группа наибольшей дозы также продемонстрировала наибольший показатель CDAI на базовой линии. Это исследование было разрешено комитетами по медицинской этике University Hospitals Leuven, Бельгия, Leiden University Medical Center, Нидерланды, Medizinischе Klinik, Kiel, Германия и Hadassah Medical Center, Jerusalem, Израиль. Все пациенты дали информированное согласие на исследование.
Схема исследования и протокол лечения
ch5D12 представляет собой молекулярно сконструированное антитело - IgG4 человека, содержащее вариабельные области тяжелой и легкой цепей исходного варианта мАт 5D12. Показано, что это мАт связывается с несущими CD40 клетками и препятствует опосредуемой CD40 активации различных клеток.20-27 ch5D12 вводили в открытом, проводимом во множестве центров испытании однократно вводимой дозы, в котором исследовались четыре уровня доз. В каждой группе лечения было пять субъектов за исключением группы конечной дозы, в которой были зарегистрированы только три субъекта. Однократное введение на уровнях доз, составляющих 0,3, 1,0, 3,0 и 10,0 мг/кг ch5D12, осуществляли внутривенно. После завершения набора в одну группу дозы, набор в следующую группу начинали только после получения доказательства безопасности на текущем уровне дозы. Клиническую активность заболевания оценивали при каждом визите еженедельно в течение периода первых 28 дней и последующем последнем визите в день 56. Двух субъектов (группа дозы 3,0 мг/кг) исключили из исследования после оценок в 28 день, но их данные также включены в анализ. Субъекты должны были оставаться на одной и той же дозе их текущего лекарственного средства на протяжении исследования. Во время исследования не использовалось неразрешенное сопутствующее лекарственное средство. Ответ на лечение ch5D12 определяли в виде снижения CDAI, составляющего ≥100 единиц, а клиническую ремиссию заболевания определяли как CDAI ≤ 150 (общий показатель) и снижение CDAI, составляющее по крайней мере 100. Всех субъектов, которые подвергались эндоскопии при скрининге и в день 28 (n=11), анализировали на снижение эндоскопического индекса тяжести болезни Крона (CDEIS). Для гистопатологии и иммуногистохимии биопсии брали от этих 11 пациентов до и в день 28 после введения ch5D12. Оценки безопасности включали физические обследования, признаки жизни, ECG и лабораторные данные (биохимия, гематология и анализ мочи), включая оценки анти-dsDNA, pANCA и антитела человека против химерного антитела (НАСА). НАСА определяли с помощью иммуноферментного твердофазного анализа (ELISA), как объясняется в ссылке (27).
Фармакокинетики
Концентрации ch5D12 в сыворотке определяли с помощью (ELISA), как ранее описано.27 Для определения покрытия CD40 инъецированным ch5D12 кровь собирали в пробирки с гепарином и разводили в два раза в PBS. Мононуклеарные клетки периферической крови (PBMC) выделяли центрифугированием в градиенте (Lymphoprep, Nycomed, Roskilde, Дания), и 500000 клеток окрашивали FITC-меченным ch5D12 и PerCP-меченным антителом против CD20 (Becton-Dickinson, Mountain View, CA, США) и инкубировали на льду в течение 30 мин. В качестве фонового контроля для связывания 5D12-FITC отдельную пробирку PBMC окрашивали только PerCP-меченным антителом против CD20. Не связавшиеся клетки смывали, и клетки анализировали с использованием проточной цитометрии (FACSort; Becton-Dickinson). Сбор событий осуществляли с помощью установки дискриминационного окна и получения экспрессирующих CD20 клеток для в целом 5000 событий. Если число экспрессирующих CD20 клеток было низким для набора образцов, заботились о получении того же числа событий как в неокрашенных, так и в окрашенных препаратах.
Гистология и иммуногистохимия
Биопсии слизистой оболочки подвздошной кишки и ободочной кишки, полученные во время илеоколоноскопии до лечения в день 0 и день 28 с помощью стандартных щипцов, фиксировали в 6% формалине для обычного анализа. Дополнительные образцы мгновенно замораживали в соединении с оптимальной температурой резания Tissue-Tek (Miles Laboratories Inc, Naperville, IL, США) в охлажденном в жидком азоте изопентане. Образцы хранили при -80°С до дальнейшего использования. Фиксированные в формалине образцы обычно обрабатывали парафином. Срезы толщиной пять микрометров готовили и окрашивали гематоксилином и эозином. Срезы анализировали, используя микроскоп Leitz Wetzlar (Wetzler, Германия). В целом анализировали четыре полупоследовательных среза для каждого образца. Замороженные образцы использовали для иммуногистохимического анализа, который проводили с использованием набора мАт для оценки присутствия различных субпопуляций лимфоцитов и моноцитов/макрофагов. Набор комплектовали мАт, направленными против CD40 и CD40L. Иммуногистохимическое окрашивание проводили на срезах криостата, сушили в течение ночи при комнатной температуре и фиксировали в абсолютном ацетоне в течение 10 мин. Регидратированные препараты инкубировали в течение 30 мин со следующими мАт: CD3 (клон UCHT1, разведение 1/10) (Dako, Glostrup, Дания), CD4 (клон МТ310, разведение 1/10) (Dako), CD8 (клон 144В, разведение 1/20) (Dako), CD19 (клон HD37, разведение 1/30) (Dako), CD40 (клон 5D12, разведение 1/100) (PanGenetics, Amsterdam, Нидерланды), CD68 (клон Кр1, разведение 1/50) (Dako), CD154 (клон М90, разведение 1/10) (Serotec, Oxford, Соединенное Королевство). Вторым мАт было меченное биотином антитело против иммуноглобулинов мыши (разведение 1/400; Dako), используемое в течение 30 мин. Для эффективного блокирования эндогенной пероксидазы срезы также инкубировали в растворе метанола, содержащем 0,3% (об./об.) Н2О2, в течение 20 мин. После трех промывок PBS добавляли меченный пероксидазой комплекс авидин/биотин (Dako). Между инкубациями срезы промывали в забуференном фосфатом солевом растворе при рН 7 в течение 15 мин. Продукт реакции визуализировали инкубацией среза в течение 10 мин в 0,05 М ацетатном буфере при рН 4,9, содержащем 0,05% 3-амино-9-этилкарбазола (Janssen, Beerse, Бельгия) и 0,01% Н2О2, что приводило к появлению ярко-красного иммунореактивного сайта. Впоследствии срезы подвергали слабому контрастирующему окрашиванию гематоксилином Харриса, ополаскивали дистиллированной водой и покрывали с использованием глицерина покровным стеклом. Контроли, которые были отрицательными, состояли из отсутствия первого или второго антитела, использования отдельно хромогена и использования нерелевантного совпадающего по изотипу мышиного IgG (виментина; Dako).
Все обработанные парафином срезы, окрашенные гематоксилином и эозином, анализировались одним и тем же патологоанатомом (K.G.), не знающем о происхождении образцов. Активность заболевания оценивали с использованием гистологических численных показателей40, из которых максимальная активность соответствовала 16. Также анализировали вслепую имуногистохимически окрашенные среды. Для 15 наборов биопсий клетки с положительным окрашиванием подсчитывали в полях большого увеличения (увеличение х40). Поля выбирали в соответствии с наибольшей плотностью стромальных клеток. Число выражали в виде процента общего числа мононуклеарных клеток в строме. Эти подсчеты использовали для создания системы количественной оценки, посредством чего интенсивность клеток с положительным окрашиванием подразделяли на четыре категории: -, +/-, + и ++. Нормальная величина для CD3+, CD4+, CD8+, CD19+ и CD68+ для подвздошной кишки и ободочной кишки является +. Нормальная величина для клеток CD40+ и CD154+ является отрицательной (-).
Иммунологические параметры безопасности
Для оценки любой неспецифической иммуносупрессии, вызванной ch5D12, оценивали пролиферацию Т-клеток периферической крови в ответ на митоген фитогемагглютинин (РНА). Стимуляцию РНА (1 мкг/мл) осуществляли в отношении выделенных PBMC, как сообщалось ранее.41 Кроме того, для исключения истощения циркулирующих клеток определяли проценты циркулирующих клеток CD3+, CD4+, CD8+ и CD20+. Для проточной цитометрии собирали в указанные моменты времени в пробирки с гепарином кровь, из которой 100 мкл цельной крови в каждой проверяемой пробирке окрашивали следующим образом (все мАт покупали у Becton-Dickinson): анти-CD3-FITC, анти-CD19-PE, анти-CD45-PerCP, анти-CD45-FITC, анти-CD4-PE, анти-CD8-PE и анти-CD14-PE. Окрашенные клетки затем анализировали в проточном цитометре FACScan (Becton-Dickinson). Этот протокол проводили для расчета отношения клеток CD4+ к CD8+ и определения доли В-клеток в популяции лимфоцитов. Сообщаемые для CD3 и CD19 проценты были процентами Т-клеток и В-клеток в популяции лимфоцитов, в то время как о клетках CD4 и CD8 сообщалось в виде процентов клеток CD3. Только CD14 (популяция моноцитов) представлены в виде процента общей популяции CD45.
Результаты
фармакокинетики
Средние максимальные уровни ch5D12 после однократной внутривенной инъекции были зависимы от дозы и пропорциональны дозе (фиг.1). Через 24 ч приблизительно 50% введенного ch5D12 можно было обнаружить в сыворотке в группах наибольших трех доз. В 0,3 мг/кг-группе только 15% присутствовало в сыворотке через 24 ч. Из исследований на не являющихся людьми приматах был рассчитан показатель t1/2β 8-10 дней27, который подтвержден текущими данными на людях. Полного покрытия CD40 на В-клетках периферической крови не добивались при инфузии 0,3 мг/кг, как определено с помощью ex vivo конкурентного анализа с использованием FITC-меченного ch5D12 (не показано). В 1,0 мг/кг-группе покрытие CD40 на циркулирующих В-клетках можно было наблюдать на протяжении приблизительно 1 недели, в то время в двух группах наибольших доз этот период увеличивался до 2-3 недель. Это указывает на то, что в группе самой низкой дозы антагонизм в отношении CD40 был неполным, в то время как полного антагонизма добивались в 1,0 мг/кг-группе в течение 1 недели и даже дольше в группах более высоких доз.
Клинические ответы
Анализ эволюции CDAI показывает, что у 13 из 18 субъектов (72%) наблюдался благоприятный ответ после инфузии ch5D12. Аналогично, у четырех из 18 субъектов (22%) наблюдалась ремиссия во время этого периода. При оценке в день 21 благоприятный ответ был зарегистрирован у 10 из 18 субъектов. Группы 2 и 4 продемонстрировали наибольшее среднее снижение CDAI, и отчетливой связи доза-эффект не наблюдалось (фиг.2; таблица 2). После этого дисперсионный анализ повторных измерений показал статистически значимое снижение CDAI на протяжении 56-дневного периода наблюдений (Р<0,001). Различие между группами не было статистически значимым. Только в группах 3 и 4 мог быть оценен CDEIS (поскольку эндоскопию проводили только у трех субъектов в группе 1 и 2). В группе 3 два из пяти субъектов имели снижение CDEIS, в то время как в группе 4 два из трех субъектов имели снижение. Остальные субъекты в этих двух группах не продемонстрировали изменения (не показано).
Гистологические изменения
Для пациентов, у которых были взяты биопсии (n=11), один исследователь проводил оценку изменений гистопатологии и иммуногистохимии. Оценки, проведенные на материале биопсии, продемонстрировали явное влияние лечения на микроскопическую активность заболевания, а также на интенсивность инфильтрата мононуклеарных клеток в собственном слое.
Изменение от базовой линии до показателя гистопатологической активности в день 28 показано на фиг.3. Имелись образцы подвздошной кишки девяти пациентов и образцы ободочной кишки 11 пациентов в день 0. В четырех случаях подвздошную кишку не включали в скринирование, несмотря на то, что пять пациентов имели активное заболевание подвздошной кишки. Средний гистологический показатель в день 0 для пациентов с активным заболеванием составлял 4,6 (диапазон 3-7). Средний гистологический показатель для пяти пациентов снизился до 1,0 (диапазон 0-3) в день 28. Образцы ободочной кишки от четырех из 11 пациентов не включали в день 0, и образцы ободочной кишки от этих пациентов оставались нормальными в день 28. У семи пациентов ободочная кишка была явно поражена, при этом средний гистологический показатель в день 0 составлял 4,5 (диапазон 2-12). Он снизился в день 28 до среднего показателя, составляющего 1,7 (диапазон 0-7). У пяти из семи пациентов показатель в день 28 был равен или 0, что является индикатором нормальной биопсии, или 1, что является индикатором только архитектурной анормальности. В группе самой низкой дозы (панель а) показатель для ободочной кишки одного субъекта снизился, но показатели для других ободочных кишок в этой группе оставались высокими, а показатель для подвздошной кишки даже возрос. В группах более высоких доз (панели b-d) показатель активности для или подвздошной кишки, или ободочной кишки, или для обеих этих кишок снизился в день 28 после лечения ch5D12. В заключение, положительный ответ [как определено в ссылке (41)] наблюдался у 81% (9/11) субъектов. Кроме того, у семи субъектов с активным заболеванием в день 0 продемонстрировано снижение активности нейтрофилов по существу до того, что нейтрофилы не присутствовали в их биопсиях в день 28.
Для определения клеток, на которые нацелено лечение ch5D12, в каждом протоколе проводили иммуногистохимическую оценку на имеющихся биопсиях, используя маркеры Т-клеток (CD3, CD4 и CD8) и В-клеток (CD19) и маркер макрофагов (CD68). Результаты перечислены в таблице 3. В группе самой низкой дозы уменьшения не наблюдалось в день 28, в то время как во всех других группах наблюдалось уменьшение инфильтрации. В образцах подвздошной кишки на базовой линии было увеличенное количество Т-клеток CD3+ в шести из девяти случаев. В ободочной кишке увеличенное количество Т-клеток CD3+ присутствовало у четырех из 11 пациентов, все из которых также имели активное заболевание по результатам обычной гистологии. В день 28 количество Т-клеток CD3+ нормализовалось у всех, за исключением одного пациента. У этого пациента (пациент 001; группа самой низкой дозы; 0,3 мг/кг) наблюдалось постоянное увеличение клеток CD3+ в подвздошной и ободочной кишке параллельно с хронической воспалительной активностью и пролиферативным воспалением по результатам обычной гистологии. Клетки CD4+ и CD8+ подчинялись схожей с CD3+ картине.
Картина для В-клеток CD19+ была сравнима с картиной, наблюдаемой для Т-клеток. В день 0 было отмечено повышение В-клеток у шести из девяти пациентов, от которых имелись биопсии подвздошной кишки, и в четырех из 11 образцов ободочной кишки. В день 28 количество В-клеток нормализовалось во всех случаях, за исключением одного пациента в группе низкой дозы.
Схожее уменьшение в день 28 наблюдалось для клеток в виде моноцитов/макрофагов CD68+. В день 0 было отмечено увеличенное количество клеток в виде моноцитов/макрофагов CD68+ у пяти из девяти пациентов, от которых имелись биопсии подвздошной кишки, и в четырех из 11 образцов ободочной кишки. Снова все увеличения нормализовалось в день 28, за исключением одного пациента в группе низкой дозы, в образце ободочной кишки которого количество клеток в виде моноцитов/макрофагов CD68+ оставалось высоким, а в биопсии подвздошной кишки наблюдалось даже увеличение, что указывает на то, что пациент все еще имел активное заболевание в день 28.
На фиг.4 представлены характерные примеры уменьшения всех трех основных типов клеток (Т-клеток, В-клеток и клеток в виде моноцитов/макрофагов) в биопсиях подвздошной кишки пациента 011 (показатель активности: 7) и биопсиях ободочной кишки пациента 012 (показатель активности: 12) до введения ch5D12 и в день 28 после введения ch5D12.
Безопасность
Субъектов не исключали из исследования из-за побочных эффектов (АЕ), при этом тяжесть большинства этих АЕ была от легкой до умеренной. С помощью оценки АЕ, происходящих в ≥5% от общего числа АЕ, обнаружено, что наиболее частыми АЕ были гипертермия, артралгия, миалгия и головная боль. Частота артралгии, часто имеющей место как на базовой линии, так и во время периода исследования, уменьшалась в ходе периода наблюдения. Подобные гриппу симптомы и зудящие сыпи, имеющие место у четырех субъектов, из которых только у одного субъекта наблюдалась умеренная сыпь, начавшаяся вскоре после введения исследуемого лекарственного средства (3,0 мг/кг). В целом нет явной реакции на дозу для какого-либо отдельного АЕ, возможно, за исключением головной боли/мигрени.
У шести субъектов были отмечены шесть серьезных побочных эффектов (SAE); в 0,3 мг/кг-группе такие эффекты не происходили, три таких эффекта отмечены в 1,0 мг/кг-группе, два таких эффекта отмечены в 3,0 мг/кг-группе, и один такой эффект отмечен в 10,0 мг/кг-группе. Пять из шести SAE считали, возможно, связанными с исследуемым лекарственным средством, одно SAE таковым не считали. Все за исключением одного такого эффекта относились к желудочно-кишечному тракту (два - к подвздошной кишке у двух 1,0 мг/кг-субъектов; два - к обострению болезни Крона у 1,0 мг/кг-субъекта и 3,0 мг/кг-субъекта; и одно - к усилению абдоминальной боли у 3,0 мг/кг-субъекта). Однако два из таких эффектов происходили в такой момент времени, что причинная связь с исследуемым лекарственным средством является очень невероятной (у одного субъекта в день инфузии, а у другого субъекта после подвергания эндоскопии через 29 дней после введения исследуемого лекарственного средства). Ни одно из наблюдаемых отклонений не представляется связанным с исследуемым лекарственным средством или дозой.
Параметры иммунологической безопасности
Поскольку ch5D12 ингибирует активацию В-клеток in vitro, определяли уровни иммуноглобулинов в целом, и, как обнаружено, они были постоянными в ходе периода наблюдения. Для исключения возможности того, что ch5D12 может истощать В-клетки из кровообращения, авторы настоящего изобретения провели анализ субпопуляции PBMS в ходе исследования. Введение ch5D12 не оказывало влияния на проценты и абсолютные количества CD3-, CD4-, CD8- и CD14-положительных клеток. Абсолютные количества циркулирующих В-клеток CD19+ транзиторно уменьшались вскоре после введения ch5D12 и восстанавливались до своих исходных значений после того, как ch5D12 выводился из кровообращения (не показано).
Для исследования того, вызывало ли лечение ch5D12 общий иммуносупрессивный эффект, PBMS, выделенные в различные моменты времени до и после лечения, стимулировали поликлональным Т-клеточным активатором РНА. К сожалению, клетки от группы дозы 10,0 мг/кг, в том числе клетки, полученные до введения ch5D12, не были заморожены соответствующим образом и поэтому были не способны отвечать на РНА. Индекс стимуляции, индуцируемой инкубацией с РНА, был в высокой степени вариабельным, но тенденции к снижению после лечения ch5D12 не обнаруживалось (данные не показаны).
На протяжении всего исследования антитела против ch5D12 не обнаруживались в двух группах низких доз. В 3,0 мг/кг-группе обнаружено, что у одного субъекта установился ответ против ch5D12 ответ, хотя сигнал был очень низким, и его можно было обнаружить только в неразведенном образце этого субъекта в день 28. В группе наибольшей дозы другой субъект имел антитела против ch5D12 в день 28 и день 56 исследования. Их можно было обнаружить при разведении 1:64. Следовательно, только два субъекта имели положительный иммунный ответ на введение ch5D12.
Обсуждение
В открытом, проводимом во множестве центров испытании четыре уровня доз ch5D12 вводились субъектам с болезнью Крона от умеренной до тяжелой степени. Было зарегистрировано восемнадцать субъектов, при этом в каждой группе лечения было пять субъектов, за исключением группы конечной дозы, в которой были зарегистрированы только три субъекта. Уровни ch5D12 в сыворотке увеличивались в зависимости от дозы, и продолжительный полупериод существования в сыворотке, наблюдаемый у яванских макак,27 был подтвержден у людей. Это исследование показывает возможность препятствования CD40-опосредуемой активации клеток с использованием ch5D12 в отсутствие основных побочных эффектов. Анализ CDAI показывает, что у 72% субъектов наблюдался клинический ответ, а у 22% наблюдалась ремиссия во время исследования. Эти результаты являются многообещающими; однако открытая схема исследования и отсутствие контрольной группы делают невозможным определенно отнести наблюдаемые изменения к введению исследуемого лекарственного средства. Однако у авторов настоящего изобретения в действительности есть несколько доказательств в пользу противовоспалительного эффекта ch5D12. Во-первых, эффективность лечения подтверждает наблюдаемое снижение CDEIS среди субъектов в двух группах наибольших доз. Во-вторых, результаты гистологических и иммуногистохимических анализов, проведенных на биопсиях, показывают, что ch5D12 снижает воспалительную активность ободочной кишке или в подвздошной кишке. В группе низкой дозы явного эффекта не наблюдалось. Субъекты в 10,0 мг/кг-группе имели самые низкие показатели гистологической активности в день 0, и поэтому снижение показателя активности ограничено в этой группе. Наибольшие показатели на базовой линии были обнаружены в 1,0 мг/кг- и 3,0 мг/кг-группах, и у этих пациентов было явное снижение гистологических показателей в день 28.
Хотя у двух пациентов были идентифицированы антитела против ch5D12 в настоящем исследовании, уровни антител НАСА оставались очень низкими и обнаруживались только при очень низких разведениях сыворотки. Остается установить, будут ли ответы против ch5D12 обнаруживаться в последующих исследованиях.
Это испытание представляет первое введение людям мАт против CD40, в общем, и ch5D12, в частности. Однако доказательство биологической активности мАт 5D12 было получено ранее in vivo на модели экспериментального аутоиммунного энцефаломиелита (ЕАЕ) на игрунках. Введение 5D12, начиная со 2-4 недели от индукции заболевания, оказывало явно благотворные эффекты.23 Это in vivo доказательство концепции стимулировало рекомбинантную гуманизации и дальнейшую разработку 5D12. Биологическую активность ch5D12 также определяли на модели ЕАЕ на игрунках. В день 50 после MOG-иммунизации частота заболевания в группе плацебо составляла 100% (50% животных убивали из-за тяжести ЕАЕ), в то время как в группе, подвергнутой лечению ch5D12, признаков заболевания не наблюдалось.24 Более того, ch5D12 показало функциональную активность в модели почечной трансплантации на макаке резус25 и при использовании в качестве иммуносупрессирующей стратегии для предотвращения иммунных ответов против аденовирусных частиц и их продуктов.26 Исследование тканевой перекрестной реактивности на ткани людей и яванских макак показало, что ch5D12 связывается с клеточной поверхностью В-клеток и DC в лимфоидных органах. Неожиданной перекрестной реактивности не наблюдалось ни на ткани человека, ни на ткани яванских макак. Оценку безопасности и переносимости для ch5D12 проводили на яванских макаках, для которых, как было показано, еженедельное введение ch5D12 в течение 4 недель является безопасным и без каких-либо побочных эффектов для всех макак. В этом исследовании было получено функциональное доказательство того, что ch5D12 может предотвращать активацию и пролиферацию В-клеток.27 Вместе взятые эти исследования показывают, что антагонистическое мАт против CD40 ch5D12 не обладает неожиданной перекрестной реактивностью, безопасно, ограничивает иммунореактивность у яванских макак и ингибирует прототипное хроническое воспалительное заболевание центральной нервной системы у игрунок. Следовательно, преклиническая оценка поддерживала дальнейшую разработку для применения для пациентов.
С использованием моделей на животных был оценен костимуляторный путь CD40-CD154 (CD40L) в качестве многообещающей клинической мишени для лечения аутоиммунного заболевания и отторжения трансплантанта.15 Потенциально взаимодействие CD40-CD154 можно ингибировать с помощью нацеливания или на CD40, или на CD154. Все предшествующие исследования фокусировались на осуществлении антагонизма в отношении CD154, который избирательно экспрессируется на активированных Т-клетках и тромбоцитах.8-11 Клинические исследования на людях с использованием мАт против CD154 человека были остановлены из-за тромбоэмболических событий.12-14 Поскольку это мАт против CD154 разработано в виде гуманизированного IgG1, который связывается чрезвычайно хорошо с Fc-рецепторами, мАт против CD154 может перекрестно связывать CD154 с Fc-рецепторами, приводя к образованию сгустков крови. Недавно сообщалось, что CD40 конститутивно экспрессируется на тромбоцитах и, как обнаружено, является функционально важным при использовании растворимого CD154 в качестве стимула для активации тромбоцитов. Эти стимулирующие эффекты, приводящие к активации тромбоцитов, были аннулированы при добавлении мАт против CD154 или CD40.42 Авторы настоящего изобретения подтвердили нестимулирующие свойства ch5D12 также в экспериментах с тромбоцитами человека, поскольку добавление ch5D12 к предварительно стимулированным с использованием субоптимальной концентрации ADP или коллагена тромбоцитов не сопровождалось каким-либо эффектом на состояние активации тромбоцитов (M.F.Hoylaerts, неопубликованные наблюдения, Leuven, Бельгия). Кроме того, Fc-часть IgG4 - основы мАт ch5D12 имеет сниженную способность связываться с Fc-рецепторами, экспрессируемыми на тромбоцитах, по сравнению с основой IgG1. Дальнейшее различие между двумя подходами происходит из того факта, что ch5D12 нацелено на В-клетки, моноциты, макрофаги и DC, в то время как мАт против CD154 нацелено, главным образом, на активированные Т-клетки, что приводит к заметно различным клиническим стратегиям. Признаки тромбоэмболии не наблюдались ни у одного из пациентов, подвергнутых лечению, в этом исследовании.
Эффект ch5D12 в большой мере основан на препятствовании опосредуемой CD40 активации клеток. Стимуляция CD40 индуцирует секрецию цитокинов и хемокинов макрофагами и DC и увеличивает антигенпрезентирующую способность последних. ch5D12 DC проявляет антагонистическое действие в отношении рецептора CD40, благодаря которому индуцируется продукция TNF-α,39 он мог бы также препятствовать секреции TNF-α, который, без сомнения, является важным цитокином при болезни Крона.43 Однако при стимуляции CD40 также индуцирует ряд других воспалительных цитокинов, таких как IL-8 и IL-12,39 и, следовательно, ch5D12 могло бы иметь противовоспалительные свойства помимо предотвращения продукции TNF-α. Выяснение того, зависят ли наблюдаемые эффекты, главным образом, от уменьшения активации Т-клеток, или уменьшения высвобождения цитокинов и хемокинов, ждет дальнейших исследований.
Это первое исследование на людях с использованием ch5D12 доказывает многообещающую клиническую пользу в отсутствие серьезных побочных эффектов. Потребуются дополнительные клинические исследования для установления роли ch5D12 в поддержании ремиссии и в контролировании безопасности множественных инфузий и оптимальной схемы введения доз ch5D12 при этом. Очевидно, что терапевтическая польза антагонизма в отношении CD40 не будет ограничиваться болезнью Крона, и ее потенциально можно расширить на много других воспалительных заболеваний.
Пример 2
Материалы и методы
Создание химерного варианта 5D12
Начиная с гибридомной клеточной линии 5D12, клонировали мышиные вариабельные области 5D12. Вкратце, из лизированных клеток с использованием Qiagen выделяли общую РНК. С помощью ОТ-ПЦР с использованием праймеров 5' CAG GTS MAR CTS SAG SAG TC W GG 3' и 5'GCA TGT ACT AGT AAT TTT TVT TGT CСA CYT TGG TGC T 3' амплифицировали область VH и с использованием праймеров 5' CGA TAC GAS MTY CAG CTG ACC CAG TCT CCA 3' и 5' GAC TCA TCT AGA TAC ACT CAT TCC TGT TGA AGC TCT TG 3' амплифицировали область VL mu5D12. После анализа последовательности определяли консенсусные последовательности для V-областей. Впоследствии V-области реклонировали в pcDNA3002 (Marissen et al., J. of Virol., 2005: 79: 4672-4678) вместе с геномными последовательностями, кодирующими IgG4 и каппа человека, для конструирования экспрессирующей химерный иммуноглобулин IgG4 плазмиды.
Создание деиммунизированного варианта 5D12
В сотрудничестве с Biovation, используя их запатентованную технологию деиммунизации, в пределах V-областей были идентифицированы потенциальные Т-клеточные эпитопы. Эпитопы Т-клеток-хелперов включают короткие аминокислотные последовательности в белках со способностью связываться с молекулами МНС класса II. После удаления Т-клеточных эпитопов антитело больше не может инициировать помощь Т-клетки и последующую иммуногенность против нее. Идентификацию осуществляли с использованием организации поточной обработки пептидов. Подход организации поточной обработки пептидов представляет собой основанный на компьютере способ предсказания связывания пептидов (небольших аминокислотных последовательностей) с 18 различными молекулами МНС класса II человека, основанными на известных структурах, разрешенных с помощью рентгеновской кристаллографии. Более того, был осуществлен анализ с поиском V-областей на присутствие известных связывающих Т-клетки человека пептидов в базе данных (http://www.wehil.wehi.edu.au).
На основе исходных мышиных аминокислотных последовательностях V-областей создавали поверхностные гуманизированные (венированные) V-области. Были идентифицированы 8 потенциальных Т-клеточных эпитопов в пределах этой венированной последовательности VH, и 4 Т-клеточных эпитопа были идентифицированы в пределах этой венированной области VL. Идентифицированные Т-клеточные эпитопы удаляли с помощью аминокислотной замены. На основе этого анализа последовательности ДНК, кодирующие деиммунизированные области VH и VL, синтезировали и реклонировали в экспрессирующую плазмиду pcDNA3002-hIgG4-каппа.
Сайт-направленный мутагенез
Для восстановления мышиных последовательностей внутри области VH подвергнутой деиммунизациии экспрессирующей плазмиды pcDNA3002 использовали в соответствии с протоколом производителя набор Stratagene для сайт-направленного мутагенеза Quick-Change. В таблице 4 представлены использованные олигонуклеотиды. Вкратце, плазмиду денатурировали с последующим отжигом смыслового и антисмыслового праймеров, содержащих желаемую мутацию. С помощью циклирования (30 сек 95°С; 30 сек 55°С; 600 сек 68°С) 16 раз с использованием устройства для ПЦР Biometra T gradient включали мутагенные последовательности. Немутированную исходную плазмиду расщепляли DpnI, которая только разрезает метилированную ДНК. Впоследствии ПЦР-смесь, содержащую мутированную плазмиду, трансформировали в компетентные клетки XL1-Blue. Колонии отбирали и анализировали по последовательностям ДНК на правильную плазмиду. Второй набор экспериментов фокусировался на создании дополнительных вариантов положений 29 и 37, в частности, с помощью замены I на структурно родственный V или L. Сходным с описанным выше образом создавали плазмиды с использованием олигонуклеотидов, представленных в таблице 6. Для создания двойных мутантов потребовалось два раунда мутагенеза (таблица 7). Плазмиды использовали для трансформации компетентных клеток XL1-Blue.
Экспрессия в PER.C6TM
Экспрессию различных вариантов 5D12 осуществляли транзиторно в PER.C6TM (Crucell). Вкратце, за день до трансфекции PER.C6TM подвергали трипсинизации и подсчитывали. Клетки помещали в DMEM, замещенную 10% FCS, при плотности 4×105 клеток на лунку в луночной планшете Т24. На следующий день среду заменяли 0,5 мл свежей среды. Для каждой лунки 1 мкг плазмидной ДНК растворяли в 50 мкл OPTI-MEM I и объединяли с равным объемом OPTI-MEM I, содержащей 3 мкл реагента LipoFectAMINE 2000 (LF2000) (Invitrogen). Смесь инкубировали в течение 20 мин при комнатной температуре для предоставления возможности образования комплексов ДНК-реагент LF2000. Впоследствии комплексы ДНК-реагент LF2000 (100 мкл) непосредственно добавляли в каждую лунку. Трансфицированные клетки инкубировали при 37οС в инкубаторе с СО2. Через 48 ч супернатанты анализировали на экспрессию антител.
ELISA
Количество экспрессированного антитела определяли с помощью сэндвич-ELISA. Вкратце, 96-луночные планшеты EIA/RIA (Corning 3590) покрывали в течение ночи при 4°С поликлональным антителом против IgG человека (Jackson 109-005-088), разведенным 1/100 PBS (100 мкл/лунку). После блокирования PBS, содержащим 0,05% Tween и 1% BSA, планшеты трижды промывали PBS-0,05% Tween. Впоследствии в качестве стандарта использовали очищенное химерное 5D12 при титровании, начинающемся при 400 нг/мл до нуля в трех повторах. Образцы (транзиторные супернатанты) также использовали при титровании. Все разведения осуществляли в PBS-0,05% Tween. Планшеты инкубировали в течение 1 часа при 37°С. После трехкратных промывок в каждую лунку вносили разведенное 1/5000 антитело для обнаружения (меченное щелочной фосфатазой против каппа человека (Southern Biotech Associated 2060-04)). Планшеты инкубировали в течение 1 часа при 37°С. После удаления инкубационной смеси планшеты промывали 5 раз. Окрашивание осуществляли с использованием субстрата PNP (Sigma N-2765) (1 мг PNP/мл субстратного буфера - 100 мМ Tris/HCl, 100 мМ NaCl, 5 мМ MgCl2·6H2О, pH 9,5). ОП405 нм измеряли на BioRad 550 Titertek, используя 655 нм в качестве точки отсчета.
FACS
С помощью анализа FACS анализировали антигенную специфичность экспрессированных антител. Клетки JY являются трансформированными EBV В-клетками человека, которые экспрессируют CD-40. Вкратце, 100000 клеток JY инкубировали со 100 мкл супернатанта из эксперимента транзиторной экспрессии при 4°С в течение 20 минут. Для удаления несвязавшегося антитела клетки промывали PBS, содержащим 0,05% азида Na и 1% BSA. Использовали второе антитело, разведенное 1/100, меченное FITC, против IgG человека (Jackson 109-095-127). После инкубации в течение 20 минут при 4°С клетки промывали. В конце концов, образцы измеряли на FACScan (BD).
Результаты
После анализа нескольких клонов определили консенсусную последовательность для мышиных V-областей 5D12 (фиг.5).
С помощью человеческого поверхностного моделирования V-области были реконструированы, сохраняя критические мышиные аминокислоты. С помощью способа организации поточной обработки пептидов и поиска в базе данных были определены потенциальные Т-клеточные эпитопы в пределах этих реконструированных последовательностей. Были идентифицированы 8 потенциальных эпитопов (первое основание потенциального эпитопа: положение 27, 30, 43, 46, 57, 61, 77 и 83) в пределах области VH и 4 потенциальных эпитопа (положение 7, 13, 28 и 86) в пределах области VL. На основе картирования Т-клеточных эпитопов и процесса венирования были определены оптимальные деиммунизированные аминокислотные последовательности VH и VL (фиг.6). Для удаления Т-клеточного эпитопа в положении 61 в области VH особое внимание уделялось не введению потенциального сайта N-гликозилирования (N-X-T/S): NSA заменяли STS. В этом конечном конструировании были изменены 12 положений в области VH (считая положения 60-62 в качестве 1 замены). Для области VL были изменены 8 положений.
В предварительных экспериментах, в которых сравнивались уровни экспрессии полностью деиммунизированного 5D12 (DI5D12) с ch5D12 (переименованным в PG100), обнаружено, что из-за введенных мутаций DI5D12 экспрессировалось на более низких уровнях, чем исходная последовательность (данные не представлены).
В принципе, каждая замена может влиять на свойства экспрессированного антитела. Поэтому для исследования влияния каждого замененного положения области VH на свойства антитела были сконструированы 12 дополнительных вариантов (фиг.7). В каждом варианте 1 положение меняли обратно на исходную мышиную аминокислотную последовательность.
Для проверки уровней экспрессии и функциональных возможностей различных вариантов были сконструированы 14 экспрессирующих плазмид pcDNA3002. Каждая плазмида содержит основу IgG4 человека, с которой связаны области VH, и область каппа человека, с которой связаны области VL. Одна плазмида кодировала сh5D12, в которой использовались мышиные V-области. Другие 13 плазмид были деиммунизированными вариантами, в которых деиммунизированная область VL используется в комбинации с деиммунизированной областью VH или одним из 12 вариантов VH, представленных на фиг.7. Впоследствии клетки PER.C6TM трансфицировали 14 плазмидами с использованием LipoFectamine 2000. Также были включены отрицательные ложные трансфекции (без плазмиды). Через 48 часов транзиторные супернатанты от каждой трансфекции собирали и анализировали на экспрессию антител. На фиг.8 представлены данные FACS. Ясно, что все проверенные конструкции связываются с клетками JY, что указывает на то, что специфичность связывания CD40 5D12 сохраняется.
Кроме того, данные FACS указывают на то, что уровни продуцируемых антител различаются. Явно различие в уровне экспрессии между DI5D12 и ch5D12 (PG100). Варианты Q5E, K13A, E16Q, T17S, STS60NSA и T68S демонстрируют одинаковые с DI5D12 характеры экспрессии. Другие варианты, по-видимому, увеличивают уровни экспрессии. Для точного определения различных уровней экспрессии проанализировали все супернатанты с помощью количественного ELISA, с помощью которого определяются уровни антител. Как показано в таблице 5, проверенные варианты можно разделить на 3 группы: нет или слабое улучшение уровня экспрессии (Q5E, K13A, E16Q, T17S и STS60NSA), варианты P45L, M48L, S79F и T108S восстанавливают уровни экспрессии по сравнению с ch5D12 (PG100) до 40%, в то время как варианты I29L и I37V восстанавливают уровень выше 50%. Особенно замена в положении 29 (I на V), по-видимому, оказывает наибольшее влияние на уровни экспрессии. Впоследствии вариант I29L переименовали в антитело PG102.
Наблюдение, что замена в положении 29 (I на L) или 37 (I на V) оказывает наибольшее влияние на уровни экспрессии, находилось среди других неожиданных наблюдений, поскольку аминокислоты I, L и V являются структурно близкородственными, и все 3 образуют группу аминокислот с разветвленными цепями. В последующем исследовании было далее установлено, каким является эффект комбинированных замен аминокислот I в положениях 29 и 37 структурно родственными аминокислотами V или L.
На фиг.9 демонстрируются варианты V-L-I, которые были сконструированы соосно с ch5D12 (PG100), DI5D12 и PG102 (вариантом I29L).
Для проверки уровней экспрессии и функциональных возможностей этих вариантов V-L-I были сконструированы дополнительные экспрессирующие плазмиды pcDNA3002, которые содержат основу IgG4 человека, с которой связана область VH одного из вариантов, и область каппа человека, с которой связаны области VL, как описано выше. Впоследствии клетки PER.C6TM трансфицировали плазмидами с использованием LipoFectamine 2000. Через 48 часов транзиторные супернатанты от каждой трансфекции собирали и анализировали на экспрессию антител. Для точного определения различных уровней экспрессии проанализировали все супернатанты с помощью количественного ELISA, с помощью которого определяются уровни антител. Как показано в таблице 8, различные комбинации V, L или I в положениях 29 и 37 сопровождаются различными уровнями экспрессии. Это далее показывает, что как раз не все замены V, L или I приводят в итоге к более высоким уровням продукции.
Обсуждение
Свойства можно изменить с помощью уменьшения иммуногенности терапевтического антитела. В этом исследовании показано, что схема деиммунизации приводит к антителу, которое все еще обладает способностью связываться с антигеном, однако антитело может только продуцироваться на низких уровнях. Наиболее вероятно, что при экспрессии существует проблема сборки деиммунизированных областей VH и VL. С помощью изменения каждого замещенного положения обратно на исходную мышиную последовательность в пределах области VH обнаружено, что некоторые положения являются существенными для надлежащей экспрессии антитела. Особенно сайты, основывающиеся на подходе организации поточной обработки пептидов, оказывали большое влияние на уровень экспрессии. N-концевые замены, все введенные на основе венирования, оказывали небольшое влияние. Замечательно, что замена I на L в положении 29 оказывала наибольшее влияние, что было неожиданным, поскольку обе аминокислоты являются структурно близкородственными. Впоследствии вариант I29L переименовали в антитело PG102.
Последующие исследования продемонстрировали, что различные комбинации V, L или I в положениях 29 и 37 приводят к вариациям в уровнях продукция, демонстрируя, что хотя 3 аминокислоты с разветвленными цепями V, L или I являются структурно близкородственными, замена каждой из них в положениях 29 и 37 оказывает влияние на уровни продукции полных антител.
Пример 3
Материалы и методы
Конструирование, отбор и разработка линии клеток GS-CHO для продукции PG102 (варианта I29L)
PanGenetics BV попросила Lonza Biologics (Lonza) выполнить конструирование, отбор и оценку линии клеток GS-CHO, экспрессирующей человеческое рекомбинантное IgG4/каппа антитело PG102 (вариант I29L).
Линии клеток конструировали трансфекцией клеток-хозяев CHOK1SV векторами, созданными с использованием экспрессующей ген глутаминсинтетазы (GS) системы Lonza (WO2003054172). Последовательности генов поставлялись PanGenetics. Линия клеток CHOK1SV представляет собой суспензионный вариант линии клеток яичника китайского хомячка СНОК1, адаптированный к химически определенной, не содержащей животных компонентов (CDACF) среде (Bebbington et al. (1992) Biotechnology 10: 169-75; de la Cruz Edmonds MC et al. (2006) Mol. Biotechnol. 34: 179-90).
Все реагенты, компоненты сред и материалы получали от аккредитованных поставщиков. Препарат питательных веществ SF40 и SF41, среду CM42 и дополнение к среде SPE получали от Lonza. Среду CD-CHO (Invitrogen) также использовали с добавлением селективного агента L-метионинсульфоксимина (MSX) для обычной субкультуры адаптированных к CDACF линий клеток и с фенолом красным для трансфекции. Селективный агент MSX поставлялся Sigma. Эти среды и питательные вещества не содержат антибиотиков. Конструирование GS-экспрессирующего вектора, кодирующего гены для PG102, также выполнялось Lonza. Клетки-хозяева CHOK1SV получали из пробирки криоконсервированного рабочего банка клеток 269-W Lonza.
Клеточная культура, концентрация и жизнеспособность
Клетки восстанавливали из пробирок криоконсервированных запасов с помощью быстрого нагревания до 37°С и разведения соответствующим образом в 50 мл среды для роста. Криозащитные вещества удаляли центрифугированием клеток, отбросом супернатанта и ресуспендированием клеток в свежей среде для роста. Для статической культуры клетки выращивали в колбах Т25 с объемом культуры, составляющем 5-15 мл. Эти статические культуры помещали в инкубатор при 35,5-37,0°С, содержащий атмосферу 10% (об./об.) СО2 в воздухе.
Для суспензионной культуры культуры наращивали в подходящем объеме среды для роста и субкультивировали каждые 4 дня для роста в среде CDACF. Используемыми сосудами были 125 мл встряхиваемые колбы (для объема культуры, составляющего 10-30 мл), 250 мл встряхиваемые колбы (для объема культуры, составляющего 30-50 мл), 500 мл встряхиваемые колбы (для объема культуры, составляющего 50-100 мл), 1 л вращающиеся бутыли (для объема культуры, составляющего 100-200 мл) или 2 л вращающиеся бутыли (для объема культуры, составляющего 200-400 мл). Свободное пространство каждой культуры газировали 5% объем/объем СО2 в воздухе, и колбы герметизировали. Затем культуры инкубировали при 35,5-37,0°С на платформе для встряхивания, вращающейся при 140±5 об/мин.
Концентрации клеток в целом и жизнеспособных клеток получали с помощью асептического отбора образца культуры, окрашивания нежизнеспособных клеток трипановым синим и микроскопической проверки с использованием модифицированного гемоцитометра Fuchs-Rosenthal. Когда уместно, образцы разводили до подсчета.
В альтернативном случае концентрации клеток в целом и жизнеспособных клеток определяли, используя счетчик частиц CASY-1 или автоматический счетчик клеток Vi-CELL XR. Для счетчика CASY-1 образец культуры асептически отбирали из каждой колбы, разводили буфером Casyton, и определяли число жизнеспособных клеток и клеток в целом. Жизнеспособные и нежизнеспособные клетки отличались по размеру. Процент жизнеспособности рассчитывали в виде отношения жизнеспособных клеток к клеткам в целом, умноженного на 100. Для счетчика Vi-CELL XR 0,7 мл клеточной культуры асептически отбирали, и проводили автоматический подсчет с использованием трипанового синего. Для идентификации и подсчета жизнеспособных и нежизнеспособных клеток используют ряд фотоснимков и алгоритм компьютерного анализа. Для всех способов подсчета процент жизнеспособности рассчитывали в виде отношения жизнеспособных клеток к клеткам в целом, умноженного на 100.
Трансфекция клеток-хозяев CHOK1SV
В день трансфекции клетки, растущие в неселективной среде, собирали центрифугированием и ресуспендировали при концентрации 1,43×107 жизнеспособных клеток/мл. Для каждой трансфекции в кювету для электропорации добавляли приблизительно 0,7 мл клеточной суспензии и 40 мкг плазмидной ДНК. Кювету для электропорации затем помещали в аппарат для электропорации, и посылали одиночный импульс 300 вольт, 900 мкФ. После электропорации клетки из кюветы распредели в 96-луночный планшет при приблизительно 5000 клеток-хозяев/лунку, используя среду CD-CHO/феноловый красный. Планшеты инкубировали при 35,5-37,0°С в атмосфере 10% об./об. СО2 в воздухе. Через день после трансфекции в каждую лунку добавляли 150 мкл селективной среды CD-CHO/феноловый красный/66,6 мкМ MSX. Конечная концентрация MSX в каждой лунке составляла 50 мкМ. Планшеты контролировали для определения момента, когда нетрансфицированные клетки погибали с оставлением фокусов трансфицированных клеток. Фокусы трансфицированных клеток становились видимыми через приблизительно 3-4 недели после трансфекции. Все трансфектанты, проверенные и совершенствуемые далее, происходили из лунок, содержащих только одну колонию, что определялось с помощью визуальной оценки.
Оценка продуктивности линий клеток в статической культуре
96-луночные планшеты инкубировали в течение приблизительно 3-4 недель после трансфекции для того, чтобы сделать возможным образование колоний. Колонии проверяли микроскопически для подтверждения того, что они подходящего размера (покрывая более 60% дна лунки) и что только одна колония присутствует в каждой лунке. Супернатант культуры затем исследовали на продукцию антител, используя анализ ELISA Lonza для собранного антитела, описываемого под названием «ELISA сборки». Процент конфлюэнтности лунки оценивали во время взятия образца. Величину, полученную делением результата анализа на процент конфлюэнтности, использовали для распределения в ряд линий клеток.
Культуры линий клеток, занимающих верхние места в ряду, наращивали в 24-луночных планшетах в среде CD-CHO/феноловый красный/25 мкМ MSX и оставляли для достижения конфлюэнтности. При достижении конфлюэнтности эту культуру использовали для инокуляции колб Т25 со средой CD-CHO/феноловый красный/25 мкМ MSX, в то время как в остальную культуру вновь подавалась свежая среда CD-CHO/феноловый красный/25 мкМ MSX, и осуществлялась инкубация в течение дополнительных 14 дней. В этот момент времени супернатант культуры собирали из 24-луночных планшетов, и определяли концентрацию антител с использованием анализа ELISA Lonza для собранного антитела. Осуществляли отбор наиболее продуктивных линий клеток. При достижении конфлюэнтности в колбах Т25 отобранным линиям клеток предоставляли возможность образовывать много слоев и затем вновь подавали среду CD-CHO/25 мкМ MSX. После подачи среды культуры возвращали в инкубатор на дополнительные четыре-семь дней, пока среда не становилась снова оранжевой/желтой, и клетки извлекали из колб.
Наращивание линий клеток в суспензионной культуре CDACF
Суспензионные культуры инициировали из конфлюэнтных культур в колбах Т25. Суспензионные культуры инокулировали при концентрации 0,05-0,2×106 жизнеспособных клеток/мл в 125 мл встряхиваемые колбы, содержащие среду CD-CHO/25 мкМ MSX. Если концентрация жизнеспособных клеток не достигала 0,05×106 жизнеспособных клеток/мл после максимум 14 дней в колбах Т25, 15 мл каждой культуры автоматически переносили в 15 мл среды CD-CHO/25 мкМ MSX в 125 мл встряхиваемых колбах. Линии клеток затем последовательно субкультивировали в свежей среде CD-CHO/25 мкМ MSX, при концентрации при инокуляции, составляющей 0,2×106 жизнеспособных клеток/мл, в режиме 4-дневной субкультуры, до получения приемлемых и воспроизводимых характеристик роста.
Оценка роста и продуктивности линий клеток в суспензионной культуре
Подпитываемая культура во встряхиваемой колбе
Синглетные культуры каждой отобранной линии клеток готовили в 500 мл встряхиваемых колбах со 100 мл клеточной суспензии с использованием среды CM42/4×SPE. Культуры инокулировали при 0,2×106 жизнеспособных клеток/мл, и свободное пространство каждой культуры уравновешивали 5% об./об. СО2 в воздухе. Культуры инкубировали при 35,5-37,0°С на платформе для встряхивания, вращающейся при 140±5 об/мин, до достижения концентрации жизнеспособных клеток, после пика, равной или меньше 0,6×106 жизнеспособных клеток/мл, или достижения дня 15 («перероста»). В этот момент времени культуры собирали. Концентрацию клеток определяли ежедневно с использованием автоматического счетчика клеток Vi-CELL. При концентрации клеток, составляющей 1,4-2,2×106 жизнеспособных клеток/мл, осуществляли болюсное добавление питательного вещества SF40. Дальнейшие болюсные добавления осуществляли через приблизительно 24, 48 и 72 часа после первоначального добавления. Второе питательное вещество (SF41) использовали только при получении концентраций клеток, превышающих 4,0×106 жизнеспособных клеток/мл. Образцы культур отбирали с соответствующими интервалами, центрифугировали, и супернатанты хранили при -20±5°С до анализа на концентрацию антител с помощью HPLC с использованием белка А. Супернатанты, собранные от каждой переросшей культуры, хранили при -20±5°С. Десять переросших культур с наибольшими концентрациями антител далее оценивали по качеству продуцируемых антител. Образцы оценивали на уровни агрегатов с использованием способа гельпроникающей жидкостной хроматографии высокого разрешения (GP-HPLC) до очистки с использованием белка А и характеристики антител с помощью гель-электрофореза в полиакриламидном геле с додецилсульфатом натрия (SDS-PAGE) в восстанавливающих или невосстанавливающих условиях и анализа с использованием изоэлектрического фокусирования (IEF) и анализа олигосахаридов по времени полета при масс-спектрометрии с ионизацией лазерной десорбцией при содействии матрицы (MALDI TOF-MS).
Криоконсервирование
Клетки подвергали криоконсервированию в ключевых моментах времени на протяжении рабочей программы. Клетки в растущих культурах выделяли центрифугированием и ресуспендировали в смеси для криоконсервирования. Она состояла из соответствующей среды для роста (92,5% об./об.) и диметилсульфоксида (7,5% об./об.).
Аликвоты клеток затем распределяли по маркированным пробиркам, при этом каждая аликвота содержала приблизительно 1,5 мл среды для криоконсервирования и приблизительно 0,5-1,5×107 жизнеспособных клеток. Пробирки затем замораживали в автоматическом программируемом устройстве для заморозки клеток и впоследствии переносили в криостаты с жидким азотом.
Номер поколения
Для каждой отобранной линии клеток номер поколения клеток определяли как нуль в тот момент времени, когда культуру переносили в суспензию. Впоследствии номер поколения рассчитывали в каждой субкультуре, используя процедуру, происходящую из следующего уравнения (округляемый в меньшую сторону до ближайшего 0,5 поколения).
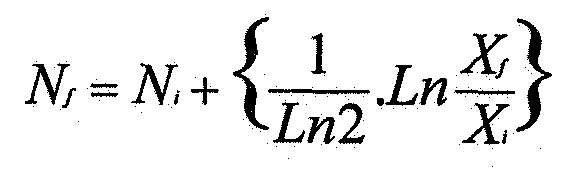
где Nf = номер поколения в момент времени f
Ni = номер поколения в момент времени i
Xf = концентрация жизнеспособных клеток (клеток/мл) в момент времени f
Xi = концентрация жизнеспособных клеток (клеток/мл) в момент времени i
и f>i.
Анализ данных
Интеграл по времени площади под кривой роста (интеграл по времени концентрации жизнеспособных клеток (IVC); 109 клеток.ч/л) рассчитывали суммированием площади (аппроксимированной до прямоугольной трапеции) между смежными величинами концентрации клеток. Площади рассчитывали умножением среднего арифметического двух концентраций жизнеспособных клеток (клеток/мл) на время, прошедшее между двумя определения (ч). Этот способ основан на способе, описанном Renard и др. (1988) в Biotechnology Letters 10(2) pages 91-96.
Когда уместно, рассчитывали общую скорость специфической продукции (qp общая: мг/109 клеток/ч) с помощью анализа линейной регрессии концентрации антитела (мг/л) против интеграла по времени концентрации жизнеспособных клеток. Наклон этой линии равен qp общей. Скорость специфической продукции во время сбора (qp сбора) рассчитывали делением концентрации антител во время сбора на величину IVC во время сбора.
ELISA сборки
Концентрацию антител в образцах супернатантов определяли с использованием ELISA, с помощью которого определяется собранный IgG человека. Оно включало захват собранного антитела в образцах и стандарте 96-луночным планшетом, покрытым козьим анти-Fc Ig человека. Связанное антитело обнаруживали с помощью козьего антитела против легкой цепи человека, связанного с пероксидазой хрена, и хромогенного субстрата тетраметилбензидина. Развитие окраски было пропорционально массе антитела, присутствующего в образце. Концентрацию определяли с использованием стандартной кривой, построенной с использованием стандарта IgG (номер партии L07387/5/2).
HPLC с белком А
Концентрацию продукта определяли с помощью жидкостной хроматографии высокого разрешения с белком А (HPLC) на Agilent 1100 HPLC. Антитело селективно связывалось с колонкой для иммунодетекции с белком А POROS. Несвязавшийся материал элюировали с колонки, и оставшееся связанное антитело высвобождали, понижая рН растворителя. Элюцию контролировали по оптической плотности при 280 нм, используя детектор множества длин волн. Количество антитела определяли (используя программное обеспечение Chemstation) против стандарта антитела (номер партии L07385/4/10) и вводили поправку на коэффициент экстинкции Е280 нм 0,1%=1,52.
Аффинная очистка с использованием белка А
Готовили колонки (5 мл) Sepharose-белок А rmp с подачей самотеком. Перед использованием колонки чистили 6М гуанидина HCl и уравновешивали буфером 50 мМ глицин/глицинат - 250 мМ NaCl, pH 8,0. рН наносимого на колонку материала доводили до рН 8,0±0,1 перед нанесением на колонку. Аффинную колонку с белком А rmp промывали буфером для уравновешивания и элюировали 0,1М глициновым буфером, рН 3,5. Элюаты с колонки нейтрализовали до рН приблизительно 7,0 с помощью 2М Tris-основания и подвергали диализу против забуференного фосфатом солевого раствора Дульбекко в препарате для анализа IEF и электрофорезом в SDS-ПААГ.
Анализ электрофорезом в SDS-ПААГ антитела, подвергнутого аффинной очистке с использованием белка А
Образцы клеточных культур готовили для анализа с помощью очистки с использованием белка А. Электрофорез проводили с использованием 4-20% полиакриламидных гелей изготовления Novex. Для электрофореза в SDS-ПААГ в восстанавливающих условиях образцы восстанавливали, используя 2-меркаптоэтанол, и денатурировали с помощью SDS при рН 8,0. Образцы нагревали в течение 2,0±0,5 минут до загрузки в гель (10 мкг на линию). Электрофорез в SDS-ПААГ в невосстанавливающих условиях проводили с использованием тех же гелей и невосстанавливающего буфера для образцов Novex (2-меркаптоэтанол не добавляется). Образцы нагревали в течение стандартного времени 1,0±0,5 минут до загрузки в гель (4 мкг на линию). После электрофореза белки визуализировали окрашиванием в течение 75 минут кумасси бриллиантовым голубым R250 и обесцвечиванием метанолом/уксусной кислотой.
Анализ IEF антитела, подвергнутого аффинной очистке с использованием белка А
Образцы клеточных культур готовили для анализа с помощью очистки с использованием белка А. Анализ IFA проводили с использованием агарозных гелей для IFA производственного изготовления, рН 3-10. Приблизительно 10 мкг белка в образце на линию загружали в гель и затем подвергали предварительной фокусировке при 70 вольт-часах (В.ч). После извлечения пластинки для загрузки образцов электрофорез продолжали при 1500 В.ч. (1500 В, 20 мА, 25 Вт на гель). IFA-гели фиксировали в растворе трихлоруксусной кислоты: сульфосалициловой кислоты: метанола и затем окрашивали кумасси бриллиантовым голубым R250 (номер партии кумасси голубого 003/L11905/118 и 003/L11905/131). Гели затем обесцвечивали и высушивали (номер партии средства для обесцвечивания 003/L11905/117 и 003/L11905/121).
GP-HPLC
С помощью способа GP-HPLC белковые компоненты разделяются в соответствии с их размером. Большие компоненты, такие как белковые агрегаты, являются слишком большими для того, чтобы проникать в частицы матрицы в какой-либо большой степени и, следовательно, элюируются с колонки перед более маленькими компонентами, такими как мономер белка. Более маленькие компоненты, такие как фрагменты, проникают в матрицу более легко и элюируются после мономера. Поэтому этот метод использовался для отделения мономера антитела как от агрегатов, так и от фрагментов. Мономерный компонент идентифицировали по его характерному времени удержания и положению, относительно калибровочных маркеров.
Неразведенные образцы вводили с получением 50-250 мкг загрузки, причем концентрация продукта составляла 1-5 мг/мл (оцененный диапазон загрузки анализа Lonza). Диапазон молекулярных масс для колонки GF-250 составлял 4-400 кДа.
Компоненты образца обнаруживали при 280 нм, и пиковые хроматограммы анализировали с использованием программного обеспечения Agilent Chemstation. Интегрирование проводили с использованием способа вертикального падения для агрегатов и соприкасающейся пленки для фрагментов.
Долю компонентов в образце определяли расчетом площадей пиков каждого компонента относительно общей интегрированной пиковой площади.
Анализ MALDI TOF-MS
Анализ олигосахарида антитела PG102, продуцируемого каждой линией клеток, проводили с помощью MALDI TOF-MS с использованием MicromassTM MALDI-LR вместе с задержанной экстракцией. MALDI TOF-MS выполняли в составе оборудования рефлектрона способом положительных ионов. Инструмент также калибровали с использованием стандарта в виде смеси N-гликанов.
Антитело очищали из собранных супернатантов, используя колонку с белком А POROS. Очищенные фракции собирали, и дисульфидные связи восстанавливали дитиотрейтолом с последующим алкилированием восстановленных тиолов, используя иодоацетат. Олигосахариды высвобождали, используя фермент N-гликаназу (PNGaseF) и помещали между двумя слоями матрицы в виде супер-дигидроксибензойной кислоты (super-DHB) на установочной пластине для анализа MALDI TOF-MS.
Результаты
Трансфекция клеток-хозяев CHOK1ST
Последовательность гена PG102 была использована Lonza для конструирования вектора с двумя генами pPG102/DGV (не представлено), который использовали для трансфекции линии клеток-хозяев CHOK1ST с помощью электропорации. Используя вектор pPG102/DGV, выполнили шесть рядов трансфекций, и супернатанты 255 трансфектантов скринировали на продукцию антител в 96-луночных планшетах с помощью ELISA сборки. Определяемые уровни антител вырабатывались 99,6% проскринированных трансфектантов. Сорок три трансфектанта, занимающих верхние места в ряду, были отобраны для дальнейшей оценки в 24-луночных планшетах.
Наращивание статических культур и адаптация к суспензии
Культуры отобранных линий клеток GS-CHO сначала наращивали в 24-луночных планшетах, а затем наращивали в статических колбах Т25. Остальным клеткам в 24-луночных планшетах подавалась свежая среда, и их «подвергали переросту» до оценки антительной продуктивности. Отобранные линии клеток демонстрировали диапазон концентраций антител, составляющий от 8,3 до 120 мг/л (фиг.10).
Культуры 23 линий клеток с наибольшими концентрациями продуктов, определенными с помощью ELISA сборки, были отобраны для дальнейшей оценки. Отобранные культуры переносили в среду CDACF в суспензионной культуре. Все линии клеток были успешно адаптированы к суспензионной культуре и были отобраны для дальнейшей оценки в подпитываемых культурах.
Оценка роста и характеристики продуктивности отобранных линий клеток в подпитываемой культуре во встряхиваемой колбе
Рост и характеристики продуктивности 23 отобранных линий клеток оценивали в подпитываемой культуре во встряхиваемой колбе. Для каждой линии клеток получали синглетные культуры. Два питательных вещества, SF40 и SF41, добавляли к культурам, когда концентрация жизнеспособных клеток соответствовала критерию подпитки. Протокол, использованный для режима подпитки, воспроизводит насколько технически возможно обобщенный процесс ферментации GS-CHO Lonza. Данные, касающиеся роста и продуктивности 23 отобранных линий клеток, представлены в таблице 9. Анализ с помощью HPLC с использованием белка А собранных супернатантов подпитываемых культур во встряхиваемых колбах показал, что четыре линии клеток продуцировали антитела в концентрациях, превышающих 1000 мг/л. Линия клеток L107 продуцировала антитела в наибольшей концентрации, 1674 мг/л.
Характеристика антитела, очищенного из подпитываемой культуры во встряхиваемой колбе отобранных линий клеток СНО
Собранные образцы десяти линий клеток с наибольшей концентрацией антител очищали, используя хроматографию на Sepharose-белок А rmp, до анализа. Очищенные с использованием белка А антитела затем анализировали, используя электрофорез в SDS-ПААГ и IEF. Визуальный анализ очищенных с использованием белка А образцов, проанализированных с помощью электрофореза в SDS-ПААГ в невосстанавливающих условиях, показал, что все образцы были сравнимы друг с другом, демонстрируя полосу интактного антитела с молекулярной массой приблизительно 200 кДа (данные не показаны). Были видны дополнительные неосновные полосы, хотя не все из них можно было видеть на изображении геля. Наблюдались также три полосы с молекулярными массами между 116 и 200 кДа, одна полоса с молекулярной массой между 66 и 97 кДа, одна полоса с молекулярной массой между 37 и 55 кДа и две полосы с молекулярными массами между 22 и 31 кДа. Дополнительная полоса, наблюдаемая между 66 и 97 кДа, является половиной антитела, обычной для антител IgG4 в невосстанавливающих условиях. Та же самая половина антитела также наблюдается в контроле между анализами IgG4 (IAC).
Очищенные с использованием белка А образцы, проанализированные с помощью электрофорез в SDS-ПААГ в восстанавливающих условиях, также сравнимы друг с другом, демонстрируя полосу тяжелой цепи с молекулярной массой приблизительно 50 кДа и полосу легкой цепи с молекулярной массой приблизительно 25 кДа (данные не показаны). Анализ IEF показал, что целостность образцов очищенных с использованием белка А антител из отобранных линий клеток демонстрирует сравнимые профили, с шестью главными полосами (3 основными и 3 неосновными полосами) в диапазоне pI 8,15-9,30 для всех линий клеток за исключением L97, M92 и M59. Эти три линии клеток демонстрировали три основные и две неосновные полосы. В образцах J3 и J4 наблюдались дополнительные полосы, хотя не все из них можно видеть на изображении геля.
Олигосахаридный анализ антитела
Олигосахаридный анализ антитела, продуцируемого каждой из десяти линий клеток, отобранных для дальнейшей оценки в подпитываемых культурах во встряхиваемых колбах, проводили с помощью MALDI TOF-MS. Преобладающими олигосахаридными структурами в антителах, полученных из десяти линий клеток, были G0F и G1F, которые являются типичными N-связанными олигосахаридными структурами, наблюдаемыми в антителах. Были обнаружены относительно низкие уровни структур в виде олигоманноз, при этом самый высокий уровень, составляющий 6,7% от гликанов в целом, был определен для антитела, происходящего из линии клеток L52.
Во всех образцах наблюдали структуры олигоманнозы-5 (man-5) в диапазоне 1,1-4,6% от гликанов в целом. Все проанализированные образцы содержали сравнимые уровни олигосахаридов и относительно низкие концентрации олигоманнозы-5 (данные не показаны).
GP-HPLC
Когда антитело, продуцируемое каждой из десяти линий клеток, отобранных для дальнейшей оценки, проанализировали с помощью GP-HPLC, компоненты с низкими молекулярными массами (LMWC) были, по крайней мере, критическими, но предпочтительным был уровень ниже 25%. Все образцы демонстрировали уровни ниже 25%, за исключением L52, который продемонстрировал уровень 27,1% (таблица 10).
Два различных расчета доли агрегатов обычно не использовались, но, как полагали, необходимы, поскольку уровни LMWC варьировали слишком сильно между образцами.
Дальнейший анализ данных, исключающий LMWC и включающий только мономер и более большие пики, продемонстрировал, что площади пиков агрегатов составляют менее 2,2% для всех образцов (таблица 11). Уровни ниже 5% считаются приемлемыми.
Отбор линий клеток
Три линии клеток отобрали для приготовления предварительных источников-запасов (PSS). Используемыми критериями отбора были комбинация высокой концентрации собираемого антитела, приемлемых характеристик роста и приемлемых характеристик качества продукта.
Линия клеток L107 была отобрана в качестве ведущей линии клеток, поскольку она проявляла наибольшую концентрацию собираемого антитела из 23 оцененных линий клеток, вместе с приемлемыми характеристиками роста и качества продукта (фиг.11А).
Линия клеток L25 демонстрировала вторую наибольшую концентрацию собираемого антитела из двадцати трех оцененных линий клеток, вместе с приемлемыми характеристиками роста и качества продукта (фиг.11В). Поэтому линия клеток L25 была отобрана в качестве первой компенсирующей линии клеток.
Линия клеток М95 демонстрировала третью наибольшую концентрацию собираемого антитела из двадцати трех оцененных линий клеток и имела приемлемые характеристиками роста и качества продукта (фиг.11С). На этом основании линия клеток М95 была отобрана в качестве второй компенсирующей линии клеток.
Криоконсервирование предварительных источников-запасов
Предварительные источники-запасы (20 пробирок на каждую линию) подвергали криоконсервированию для каждой из трех отобранных линий клеток. Эти запасы хранили в криостатах с жидким азотом паровой фазы. Линии клеток L107, L25 и М95 были переименованы в DC1, DC2 и DC3, соответственно.
Обсуждение
Lonza Biologics попросили выполнить конструирование, отбор и оценку линии клеток GS-CHO, экспрессирующей человеческое рекомбинантное IgG4/каппа антитело PG102.
Последовательность гена поставлялась PanGenetics, и векторы были созданы с использованием экспрессирующей ген глутаминсинтетазы (GS) системы Lonza. Линию клеток CHOK1SV, суспензионный вариант линии клеток яичника китайского хомячка (СНО) СНОК1, адаптированный к химически определенной, не содержащей животных компонентов среде, подвергли трансфекции электропорацией.
Из трансфектантов CHOK1SV были получены 254 линии клеток, которые секретировали антитело. Сорок три линии клеток были оценены на рост и продуктивность в 24-луночных планшетах. Были определены концентрации антител, составляющие до 120 мкг/мл. Двадцать три линии клеток были оценены в синглетных подпитываемых культурах во встряхиваемых колбах. Были получены концентрации собираемых антител в диапазоне, составляющем 121-1674 мкг/мл. Для четырех линий клеток были получены концентрации собираемых антител, превышающие 1000 мг/л. Десять линий клеток с наибольшими концентрациями антител были отобраны для анализа качества продукта. Качество продукта - очищенного антитела из каждой из отобранных линий клеток было сравнимо друг с другом при анализе электрофорезом в полиакриламидном геле с додецилсульфатом натрия в восстанавливающих или невосстанавливающих условиях и анализе с использованием изоэлектрического фокусирования. Олигосахаридный анализ очищенного антитела из каждой из десяти отобранных линий клеток показал относительно низкие (<4,6%) уровни олигоманнозы-5.
Анализ GP-HPLC показал, что уровни агрегатов были ниже 1% для каждой из трех верхних линий клеток.
Три линии клеток (L107, L25 и М95) были отобраны для дальнейшей оценки на основе данных, касающихся роста, продуктивности и качества продукта. Предварительные источники-запасы из 20 пробирок для каждой линии клеток подвергли криоконсервированию. Три линии клеток были переименованы в DC1, DC2 и DC3, соответственно.
Пример 4
Определение кинетик связывания ch5D12 (PG100) и PG102 (варианта I29L) с CD40-Fc человека с помощью резонанса поверхностных плазмонов (Biacore)
Материалы и методы
Кинетики связывания ch5D12 (PG100) и PG102 (варианта I29L) с CD40 человека сравнивали с помощью резонанса поверхностных плазмонов при постоянной температуре, составляющей 25,0°С, используя прибор Biacore 300. Рекомбинантный человеческий внеклеточный домен CD40, слитый с доменом Fc Ig человека (huCD40-Fc; номер в каталоге R&D Systems - 149-CD-50) использовали в качестве целевого антигена в этих экспериментах. HuCD40-Fc (1 мкг/мл в 10 мМ ацетатном буфере, рН 5,0) связывали через аминогруппы остатков Lys c активированными карбодиимидом чипами СМ5 Biacore, получая уровень иммобилизации антигена, составляющий 127,4 RU (условных единиц). Для кинетических анализов были приготовлены серийные разведения ch5D12 (PG100) и PG102 (в диапазоне 10,0-0,313 мкг/мл) в эксплуатационном буфере для Biacore (забуференном HEPES солевом растворе, содержащем EDTA и поверхностно-активное вещество; HBS-EP). Кинетические анализы проводили при скорости потока 30 мкл/мин. Образцам предоставляли возможность связываться в течение 4 мин и диссоциировать в течение 5 мин при одинаковой скорости потока. Поверхность чипов затем регенерировали в течение 30 секунд, используя 30 мМ NaOH, и базовую линию восстанавливали в HBS-EP в течение 2,5 мин до инъекции следующего образца. Инъецировали чередующиеся образцы PG102 и ch5D12 (PG100) с увеличением концентрации для первого набора дубликатов для каждого разведения и затем со снижением для второго набора дубликатов.
Для сбора данных использовали программное обеспечение для контроля BIAcore (версии 3.2), а для анализа кинетических данных использовали программное обеспечение для оценки BIAevaluation (версии 4.1). Параметры связывания рассчитывали, используя две различные модели молекулярного взаимодействия: модель двухвалентного вещества, определяемого при анализе, и модель связывания в соотношении 1:1 Ленгмюра.
Результаты
Максимальными уровнями наблюдаемого связывания для антител ch5D12 (PG100) и PG102 были 152,5 RU и 140,2 RU, соответственно.
Рассчитанные кинетические константы скорости в соответствии с моделями Ленгмюра и двухвалентного связывания (ka и kd или ka1 и kd1, соответственно) слегка отличались для двух антител. Таким образом, PG102 демонстрировало более высокие скорости ассоциации и диссоциации, чем ch5D12 (PG100) (фиг.12), хотя аффинность в целом (KD) двух антител не отличалась заметно в этих моделях. Кинетические параметры этих экспериментов суммированы ниже.
Модель Ленгмюра (1:1)
Модель двухвалентного связывания (1:2)
Анализ степени согласия для двух способов показал, что данные лучше описываются моделью двухвалентного связывания.
Рассчитанные аффинности двух вариантов антител отличаются между моделями, однако оценки констант диссоциации (KD), полученные для каждого антитела с использованием одной и той же модели, были сравнимыми; 2,71×10-10 М и 2,37×10-10 М для ch5D12 (PG100) и PG102, соответственно, используя модель Ленгмюра, и 5,25×10-9 М и 5,41×10-9 М, используя модель двухвалентного связывания.
Пример 5
Ингибирование связывания ch5D12 (PG100) и PG102 с клетками JY, используя антиидиотипическое антитело
Для демонстрации того, что PG102 (вариант I29L) связывается с тем же эпитопом CD40, что и исходная молекула ch5D12 (PG100), использовали антиидиотипическое антитело для ингибирования связывания PG102 и ch5D12 с экспрессирующими CD40 клетками JY, измеряемого с помощью FACS.
Материалы и методы
Клетки JY, экспрессирующие CD40, блокировали 5% сыворотки человека в течение 30 мин при комнатной температуре. Антиидиотипическое антитело против мышиного мАт 5D12 (клон 173-36-1) предварительно инкубировали при различных концентрациях (0-10 мкг/мл) или с ch5D12 (PG100) (1 мкг/мл), PG102 (1 мкг/мл), или с химерным антителом против CD86 человека (chFUN-1, 1 мкг/мл) в качестве отрицательного контроля в общем объеме 50 мкл/пробирку и инкубировали в течение 15 мин. Клетки JY промывали буфером для FACS (1×PBS, 1% BSA и 0,05% азида натрия), и супернатант отбрасывали. Клетки JY ресуспендировали в буфере для FACS до концентрации 2×106 клеток/мл. 50 мкл суспензии клеток JY добавляли к предварительно проинкубированным смесям антител с получением конечного объема 100 мкл/пробирку. Клетки инкубировали в течение 30 мин при 4°С до промывки 4 мл буфера для FACS для каждой пробирки, и супернатант отбрасывали. Клеточный осадок ресуспендировали в 100 мкл разведенного козьего антитела против IgG человека, конъюгированного с FITC (Jackson Immunoresearch Labs, каталожный номер 109-095-127) в буфере для FACS (1:100) и инкубировали в течение 30 мин при 4°С. Клетки промывали 4 мл буфера для FACS, и супернатант отбрасывали. Осадок ресуспендировали в 200 мкл буфера для FACS, и связанное антитело определяли, используя проточный цитометр FACScan (Becton Dickinson).
Результаты
Антиидиотипическое антитело мАт 173-36-1 ингибировало связывание PG102 и ch5D12 (PG100) с huCD40, экспрессируемым на клетках JY, зависимым от концентрации образом (фиг.13). мАт 173-36-1 не оказывало влияния на связывание chFUN-1 с CD86, который также экспрессируется на клетках JY. Не было значительного различия в блокирующем эффекте антиидиотипического мАт 173-36-1 на связывание специфических в отношении CD40 антител с CD40 на клетках JY. Например, рассчитанными значениями log IC50 для опосредуемого мАт 173-36-1 ингибирования ch5D12 (PG100) (партия 1) и PG102 (партия 2) были -5,74±0,14 и -5,76±0,11, соответственно (р>0,05; оба n=4). Эти значения соответствуют концентрациям антиидиотипического антитела, составляющим приблизительно 1,8 мкг/мл (~12 нМ). В таблице 12 представлена сводка рассчитанных значений -log IC50 для опосредуемого антиидиотипическим антителом ингибирования связывания ch5D12 (PG100) и PG102 с клетками JY.
Пример 6
Связывание ch5D12 (PG100) и PG102 с CD-40Fс, определяемое с помощью ELISA
Материалы и методы
Планшеты для ELISA (планшет Costar EIA/RIA, номер в каталоге Corning - 3590) покрывали 100 мкл/лунку huCD40-muIg (Ancell, каталожный номер 504-020), разведенного до 250 нг/мл в PBS, и инкубировали в течение ночи при 4°С в обстановке влажности. Планшеты промывали 3 раза буфером для промывок (0,05% Tween-20 в PBS), 200 мкл/лунку. Впоследствии планшеты блокировали буфером для блокирования (5% фракции V BSA [Roche, каталожный номер 735094] в буфере для промывок), 200 мкл/лунку, и инкубировали в течение 1 ч при 37°С (в обстановке влажности). Планшеты промывали 3 раза буфером для промывок. Проверяемые антитела (ch5D12 (PG100), PG102 и химерное антитело против CD86 человека chFUN-1) разводили в диапазоне 0-1200 нг/мл в буфере для блокирования и переносили в планшеты для анализа в конечном объеме 100 мкл/лунку с последующей инкубацией в течение 1 ч при 37°С (в обстановке влажности). Планшеты промывали 3 раза буфером для промывок с последующим добавлением 100 мкл/лунку козьего антитела против каппа человека, конъюгированного с АР, для детекции (Southern Biotech Associates, каталожный номер 2060-0), разведенного 1:1000 в буфере для блокирования. Планшеты инкубировали в течение 1 ч при 37°С в обстановке влажности до промывки 3 раза буфером для промывок и дополнительной одной промывки PBS. Субстрат PNP готовили добавлением 1 таблетки пара-нитрофенилфосфата (Sigma, каталожный номер N-2765) в 15 мл буфера для субстрата PNP (12,1 г Tris, 5,84 г NaCl, 1,02 г MgCl2 .6H2O в 1 л H2O (доведенного до рН 9,6 с помощью HCl). В планшеты для анализа добавляли 100 мкл/лунку субстрата PNP. Планшеты инкубировали в течение нескольких минут (максимум 30 мин) при 37°С, чтобы сделать возможным развитие окрашивания до остановки реакции добавлением 3М NaOH. Интенсивность окрашивания определяли при 405 нм, используя считывающее устройство для микропланшетов (Biorad, модель 550).
Результаты
В ряде независимых экспериментов обнаружено, что серийно разведенные ch5D12 (PG100) и PG102 (вариант I29L) сравнимым образом связывались с иммобилизованным CD40-Fc (фиг.14), в то время как контрольное антитело против CD86 не демонстрировало заметного связывания с CD40-Fc. Рассчитанные концентрации для полумаксимального связывания для всех экспериментов суммированы в таблице 13. Значительного различия не наблюдалось в концентрации для полумаксимального связывания для любой из оцененных партий антител против CD40.
Пример 7
Связывание ch5D12 (PG100) и PG102 с huCD40, экспрессируемым на клетках JY, определяемое с помощью FACS
Материалы и методы
Экспрессирующие CD40 клетки JY промывали буфером для FACS (1×PBS, 1% BSA и 0,05% азида натрия). Супернатант отбрасывали, и клетки JY ресуспендировали в буфере для FACS до концентрации 2×106 клеток/мл. Впоследствии 50 мкл суспензии клеток JY добавляли к 50 мкл каждого из проверяемых антител (ch5D12 (PG100), PG102 и химерного антитела против CD86 человека chFUN-1 в качестве отрицательного контроля), приготовленных в диапазоне концентраций 0-900 нг/мл, и инкубировали в течение 30 мин при 4°С. Клетки промывали 4 мл буфера для FACS для каждой пробирки, и супернатант отбрасывали. Клеточный осадок ресуспендировали в 100 мкл козьего антитела против IgG человека, конъюгированного с FITC (Jackson Immunoresearch Labs, каталожный номер 109-095-127), разведенного 1:100 в буфере для FACS, и инкубировали в течение 30 мин при 4°С. Клетки промывали 4 мл буфера для FACS, и супернатант отбрасывали. Осадок ресуспендировали в 200 мкл буфера для FACS, и связанное антитело определяли, используя проточный цитометр FACScan (Becton Dickinson).
Результаты
Все проверенные партии ch5D12 (PG100) и PG102 продемонстрировали схожие свойства связываться с CD40, экспрессируемым на клетках JY, (фиг.15) со сравнимыми концентрациями для полумаксимального связывания антитела (таблица 14). chFUN-1 также связывалось с клетками JY, хотя с более низкой средней интенсивностью флуоресценции (MFI), чем ch5D12 (PG100) и PG102. Это происходило, поскольку клетки JY также экспрессируют CD86 человека.
Обсуждение
Свойства связываться с CD40 различных партий PG102 (варианта I29L) и ch5D12 (PG100) определяли, используя in vitro анализы на основе клеток и ELISA. Связывание PG102 и ch5D12 (PG100) с клетками JY, экспрессирующими CD40, ингибировалось с равной эффективностью антиидиотипическим мАт 173-36-1. Таким образом, диапазон рассчитанных средних значений -logIC50 для партий PG102 и ch5D12 (PG100) составлял 5,51-6,11 (n=8) и 5,43-6,04 (n=10), соответственно (р>0,05). Антитело 173-36-1 направлено против вариабельных областей предшественника ch5D12 (PG100), мышиного мАт 5D12, и не ингибирует связывание антитела контрольного изотипа против CD86. В дальнейших экспериментах была определена концентрация PG102 и ch5D12 (PG100) для полумаксимального связывания с помощью анализа ELISA связывания антитела с CD40-Fc человека и с помощью анализа FACS связывания с клетками JY. В анализах ELISA не было значительной разницы между концентрациями PG102 и ch5D12 (PG100) для полумаксимального связывания для любой проверенной партии. Например, рассчитанные средние значения -log-концентраций для полумаксимального связывания для партий PG102 и ch5D12 (PG100) составляли 7,47±0,04 и 7,54±0,03, соответственно (P>0,05, оба n=4), соответствуя приблизительно 30 нг/мл. Схожие концентрации для полумаксимального связывания наблюдались в FACS-экспериментах по связыванию PG102 и ch5D12 (PG100) с CD40, экспрессируемым на клетках JY. Эти результаты указывают на то, что PG102 и ch5D12 (PG100) имеют один и тот же связывающий CD40 паратоп, что демонстрируется сравнимыми характеристиками активности антиидиотипического антитела в отношении ингибирования связывания PG102 и ch5D12 (PG100) с CD40. Более того, оба антитела проявляют схожие in vitro активности в отношении связывания с CD40 в анализах на основе ELISA и клеток. Вместе представленные данные говорят о том, что способности PG102 и ch5D12 (PG100) связывать CD40 человека являются одинаковыми.
Пример 8
Конкурентное ингибирование связывания ch5D12 (PG100)-РЕ и PG102-РЕ с клетками JY, экспрессирующими CD40, с использованием немеченого антитела против CD40
Материалы и методы
Клетки JY (трансформированную вирусом Эпштейна-Барра линию человеческих лимфобластоидных В-клеток) выращивали в модифицированной по способу Исков среде Дульбекко (IMDM), содержащей 10% инактивированной нагреванием фетальной телячьей сыворотки (FCS), 2 мМ L-глютамина и 50 мкг/мл гентамицина, в атмосфере увлажненного воздуха, содержащей 5% СО2, при 37°С. Клетки собирали в день измерений с использованием проточной цитометрии. В этих экспериментах использовали немеченые антитела ch5D12 (PG100) (партия 4) и PG102 (партия 2). Оба антитела также использовали в меченной РЕ форме для прямого определения связывания с клетками JY с помощью проточной цитометрии. Мечение антител РЕ по заказу осуществлялось AbD Serotec (Oxford, Соединенное Королевство). Для каждого анализа FACS использовали следующий протокол. Клетки JY собирали из клеточной культуры и подсчитывали. Впоследствии 1×105 CD40-экспрессирующий клеток JY/200 мкл буфера для инкубации (PBS, 1% BSA, 0,05% азида натрия) инкубировали с 1 мкг/мл меченых антител ch5D12 (PG100) или PG102 в присутствии варьирующей концентрации (0-10 мкг/мл) немеченого конкурирующего ch5D12 (PG100) или PG102 в течение 30 мин при 4-8°С. Клетки промывали, а затем подвергали анализу проточной цитометрией, при котором определяли MFI при каждой концентрации, используя проточный цитометр FACScan (Becton & Dickinson). Пять тысяч событий на каждый образец измеряли и анализировали с использованием программного обеспечения CellQuest® (Becton & Dickinson).
Результаты
Немеченые антитела ch5D12 (PG100) и PG102 проявляли схожие ингибиторные свойства в этих исследованиях (фиг.16). Средние значения -logIC50 для ингибирования связывания 1 мкг/мл ch5D12 (PG100) немечеными ch5D12 (PG100) и PG102 составляли 7,50±0,03 и 7,63±0,03, соответственно (оба n=4), в то время как средние значения -logIC50 для ингибирования связывания 1 мкг/мл PG102 немечеными ch5D12 (PG100) и PG102 составляли 7,54±0,01 и 7,65±0,02, соответственно (оба n=4). Эти значения -logIC50 соответствуют концентрациям PG102 и ch5D12 (PG100), составляющим приблизительно 20-30 нг/мл (т.е. ~130-200 пМ).
Пример 9
Ингибирование высвобождения IL-8 из клеток ТНР-1
Материалы и методы
Функциональные возможности PG102 определяли, используя функциональный биоанализ на основе клеток. Вкратце, в день 1 клетки человека ТНР-1 и Jurkat 39,8/50 культивировали в модифицированной по способу Исков среде Дульбекко (IMDM, BioWhittaker, каталожный номер - BE12-722F), дополненной 10% фетальной телячьей сыворотки (Gibco, каталожный номер - 10270-106) и 50 мкг/мл гентамицина (Invitrogen, каталожный номер - 15750-045). Клетки ТНР-1 стимулировали rhuIFN-γ при 1000 единиц/мл клеточной культуры.
В день 3 биоанализа клетки ТНР-1 и Jurkat 39,8/50 требовались в концентрации 4×105 клеток/мл. Клетки ТНР-1 и J39,8/50 подсчитывали, и определяли их жизнеспособность. Клетки затем разводили до концентрации, составляющей приблизительно 1×106 клеток/мл. Суспензии клеток смешили в соотношении 1:1 и инкубировали при 37°С в увлажненной атмосфере с 5% СО2 в течение 48 ч.
В день 3 клетки ТНР-1 и J39,8/50 промывали следующим образом (в случае клеток ТНР-1 стадия промывки предназначена для удаления IFN-γ): 5 мл суспензии клеток ТНР-1, простимулированных IFN-γ, и J39,8/50 переносили в 50 мл пробирку Фэлкона. Добавляли 40 мл сбалансированного солевого раствора Хэнка (HBSS), и клетки центрифугировали при 1500 об/мин (используя центрифугу IEC) в течение 6 мин. Супернатант отбрасывали. Осадок из клеток ресуспендировали в предварительно нагретой IMDM, дополненной 10% FCS. Клетки ТНР-1 и J39,8/50 доводили до концентрации 4×105 клеток/мл.
Для анализа ингибирования проверяемый образец ch5D12 (PG100) или PG102 (вариант I29L) серийно разводили в нагретой среде IMDM, дополненной 10% фетальной бычьей сыворотки, для получения конечных исследуемых концентраций в диапазоне 0-160 нг/мл. ТНР-1, J39,8/50 и проверяемый образец (образцы) добавляли в трех повторах в планшеты для культуры клеток с круглым дном (NunclonTM) в следующем порядке: 50 мкл клеток ТНР-1 (эквивалентные 2×104 клеток на каждую лунку), 50 мкл проверяемого образца и 50 мкл клеток J39,8/50. Общий объем составлял 150 мкл на каждую лунку. Планшеты для клеточной культуры упаковывали в пористую липкую пленку и инкубировали при 37°С в увлажненной атмосфере с 5% СО2 в течение 48 ч.
В день 5, после периода культивирования, составляющего 48 ч, 70 мкл супернатантов клеточных культур аспирировали и переносили в микротитрационный планшет с круглым дном. Собранные супернатанты клеточных культур анализировали на содержание IL-8, используя коммерческий ELISA (Biosourse, цитонабор для IL-8 человека, каталожный номер - СНС1304) в соответствии с инструкциями производителя.
Результаты
PG102 (вариант I29L) и ch5D12 (PG100) продемонстрировали схожие ингибиторные эффекты на индуцированное высвобождение IL-8 клетками ТНР-1 (фиг.17). Рассчитанные значения -log IC50 для двух антител составляли 8,42 и 8,28, соответственно. Эти значения соответствуют концентрациях IC50, составляющим приблизительно 30 пМ.
Пример 10
Предшествующее исследование тканевой перекрестной реактивности на ткани людей и яванских макак показало, что ch5D12 (PG100) связывается с клеточной поверхностью В-клеток и DC в лимфоидных органах. Неожиданной перекрестной реактивности не наблюдалось ни на ткани человека, ни на ткани яванских макак. Это исследование повторили для PG102, и были получены схожие результаты, что указывает на то, что ch5D12 (PG100) и PG102 связываются с различными срезами тканей схожим образом.
Также предшествующую оценку безопасности и переносимости для ch5D12 проводили на яванских макаках, для которых, как было показано, еженедельное введение ch5D12 в течение 4 недель является безопасным и без каких-либо побочных эффектов для всех макак. В этом исследовании было получено функциональное доказательство того, что ch5D12 может предотвращать активацию и пролиферацию В-клеток.27 Исследования безопасности повторили для PG102, используя более длительный протокол. Вкратце, было разработано исследование токсичности при внутривенном введении в течение 13 недель на яванских макаках с периодом восстановления в течение 14 недель, используя 3 уровня доз (0, 25 и 100 мг/кг, внутривенно) и 26 макак (6 макак (3 самца, 3 самки) на каждую группу обработки, плюс 4 макаки (2 самца, 2 самки) в качестве групп восстановления в каждой ветви активной обработки). Проводили следующие измерения: ТК, анти-PG102 ответ, проточную цитометрию (в том числе покрытие PBMC CD40), биопсию лимфатических узлов и стандартные токсикологические оценки, такие как гематология и иммуногистохимия.
Результаты этих исследований показали, что для любого из проверенных уровней доз токсичность не наблюдается. PG102 было безопасным и хорошо переносилось.
Вместе взятые эти исследования показывают, что антагонистическое мАт против CD40 PG102 человека, как и его исходное антитело ch5D12 (PG100), не обладает неожиданной перекрестной реактивностью, безопасно и хорошо переносится in vivo.
Таблица 6
Мутагенные олигонуклеотиды. Олигонуклеотиды, используемые для введения мутаций в положения аминокислот 29 и 37. Перечислены смысловые и антисмысловые (rev) олигонуклеотиды.

Таблица 7
Создание двойных мутантов, для которого требуются 2 раунда мутагенеза. Ниже перечислено, какие наборы праймеров использовались на стадии 1 и стадии 2, соответственно, для того, чтобы равняться дополнительным вариантам DI5D12.

Таблица 8
Данные, касающиеся экспрессии, измеряемой с помощью количественного ELISA. Для каждого дополнительного варианта 5D12 (29I-37V, 29V-37I, 29I-37L, 29V-37V, 29L-37L, 29V-37L и 29L-37V) вместе с ch5D12 (PG100), DI5D12 и PG102 (вариантом I29L) показана концентрация антитела в собранном супернатанте.
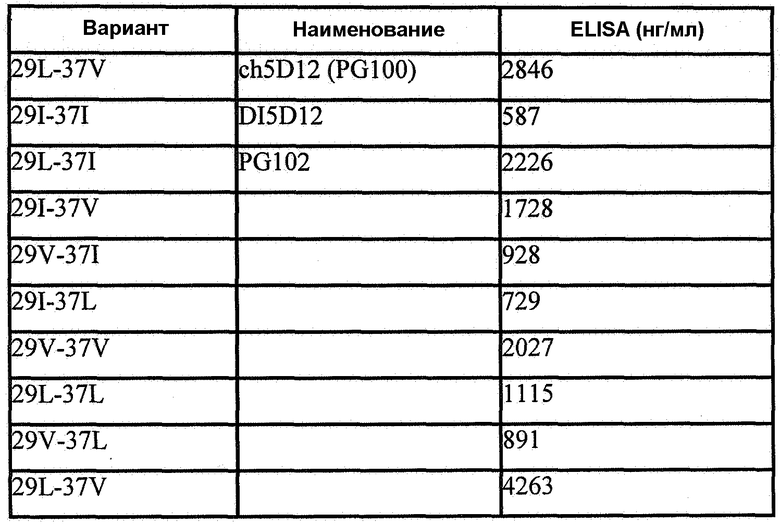
Таблица 9
Сводка данных, касающихся роста и продуктивности линий клеток СНО в подпитываемых культурах CDACF во встряхиваемых колбах
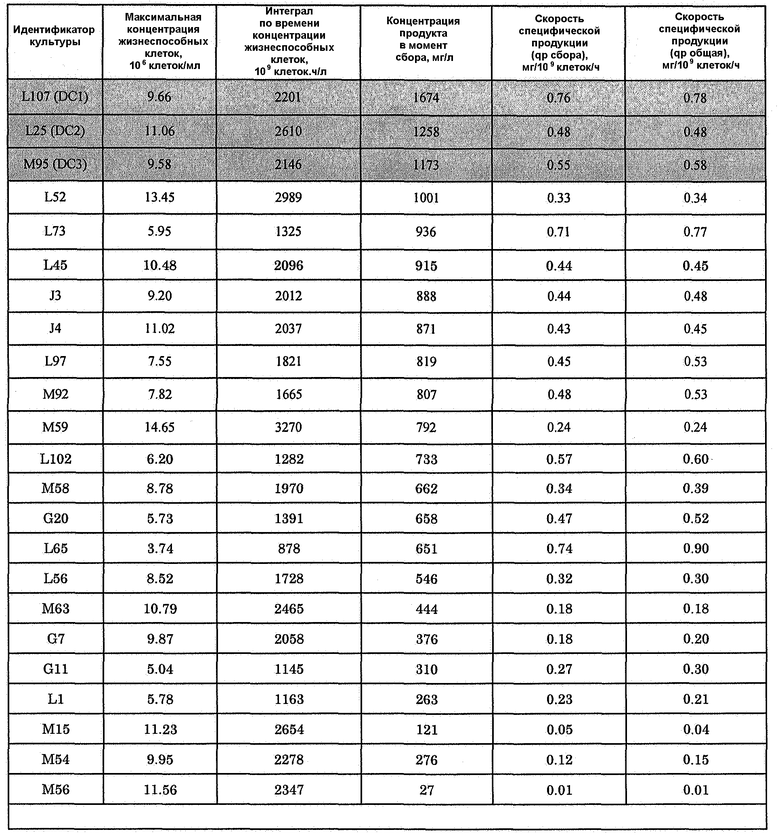
Таблица 10
Анализ GP-HPCL (включающий компоненты с низкими молекулярными массами (LMWC))
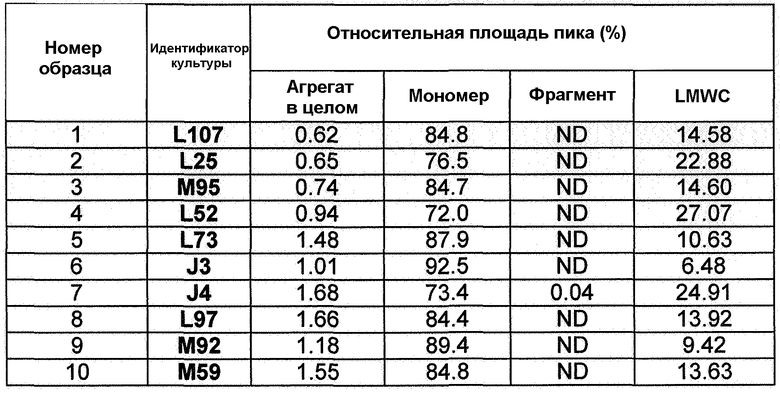
ND = не обнаружен.
Таблица 11
Анализ GP-HPCL (только продукты IgG)
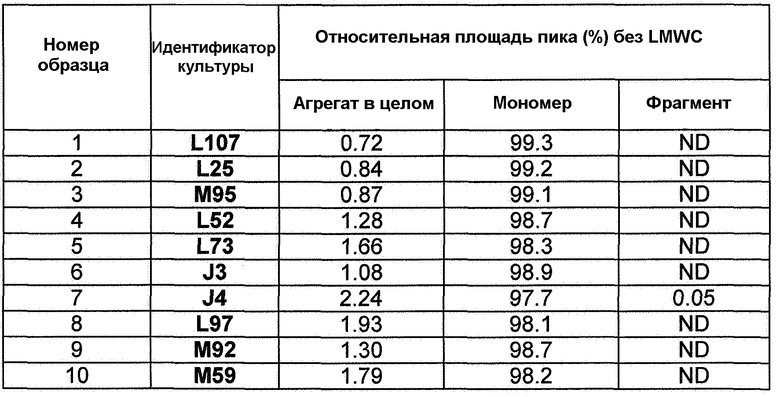
ND = не обнаружен.
Таблица 12
Сводка рассчитанных значений -logIC50 для опосредуемого антиидиотипическим антителом ингибирования связывания ch5D12 (PG100) и PG102 с клетками JY. Данные показывают среднее арифметическое ± значение стандартной ошибки n определений. Не было значительной разницы в ингибиторной активности антиидиотипического антитела с любой из проверенных партий антител (р>0,05, дисперсионный анализ (ANOVA) с последующим анализом Таки для множественных сравнений).
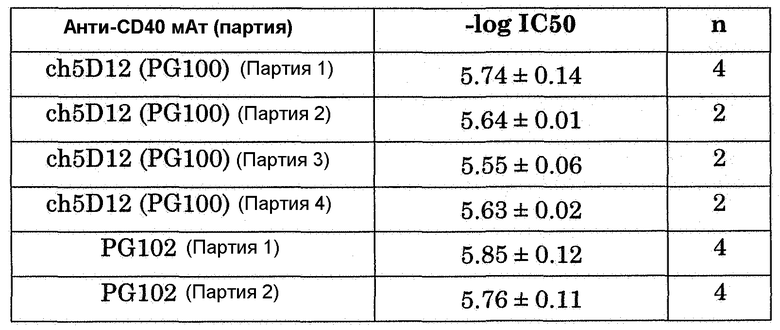
Таблица 13
Сводка рассчитанных значений -log концентрации для полумаксимального связывания для связывания ch5D12 (PG100) и PG102 с CD40-Fc человека, как определено с помощью ELISA. Данные показывают среднее арифметическое ± значение стандартной ошибки n определений. Не было значительной разницы в связывании любой из проверенных партий антител (р>0,05, дисперсионный анализ (ANOVA) с последующим анализом Таки для множественных сравнений). Для ссылки, значение -log концентрации для полумаксимального связывания, составляющее 7,45, соответствует концентрации, составляющей приблизительно 35 нг/мл (~200 пмоль).
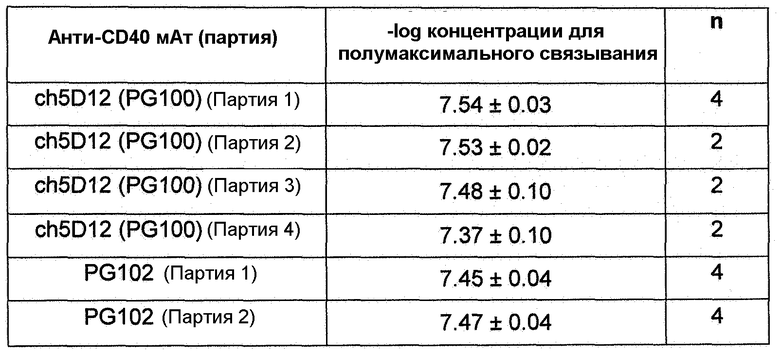
Таблица 14
Сводка рассчитанных значений -log концентрации для полумаксимального связывания для связывания ch5D12 (PG100) и PG102 с клетками JY, экспрессирующими CD40 человека, как определено с помощью ELISA. Данные показывают среднее арифметическое двойных определений. Для ссылки, значение -log концентрации для полумаксимального связывания, составляющее 7,45, соответствует концентрации, составляющей приблизительно 35 нг/мл (~200 пмоль).
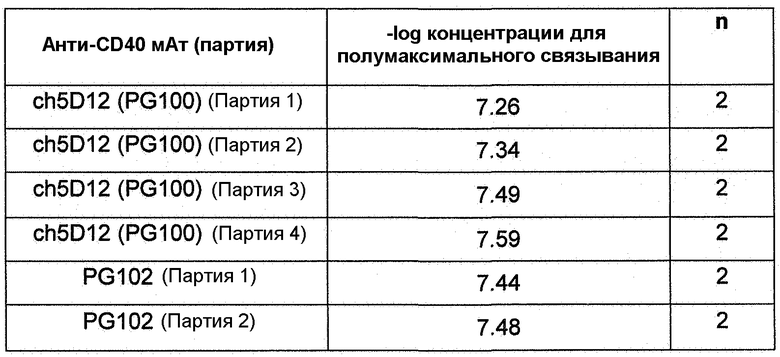
Ссылки
1. Ranheim EA, Kipps TJ. Activated T cells induce expression of B7/BB1 on normal or leukemic B cells through a CD40-dependent signal. J Exp Med (1993); 177: 925-35.
2. Hasbold J, Johnson-Leger C, Atkins CJ, Clark EA, Klaus GGB. Properties of mouse CD40: cellular distribution of CD40 and B cell activation by monoclonal anti-mouse CD40 antibodies. Eur J Immunol (1994); 24: 1835-42.
3. Alderson MR, Armitage RJ, Tough TW, Strockbine L, Fanslow WC, Spriggs MK. CD40 expression by monocytes: regulation by cytokines and activation of monocytes by the ligand for CD40. J Exp Med (1993); 178: 669-74.
4. Kiener PA, Moran-Davis P, Rankin BM, Wahl AF, Aruffo A, Hollenbaugh D. Stimulation of CD40 with purified soluble gp39 induces proinflammatory responses in human monocytes. J Immunol (1995); 155: 4917-25.
5. Shu U, Kiniwa M, Wu CY, et al. Activated T cells induce interleukin-12 production by monocytes via CD40-CD40 ligand interaction. Eur J Immunol (1995); 25: 1125-8.
6. Cella M, Scheidegger D, Palmer-Lehmann K, Lane P, Lanzavecchia A, Alber G. Ligation of CD40 on dendritic cells triggers production of high levels of interleukin-12 and enhances T cell stimulatory capacity: T-T help via APC activation. J Exp Med (1996); 184: 747-52.
7. van Kooten C, Banchereau J. Functions of CD40 on B cells, dendritic cells and other cells. Curr Opin Immunol (1997); 9: 330-7.
8. Cayabyab M, Phillips JH, Larder LL. CD40 preferentially costimulates activation of CD4+ T lymphocytes. J Immunol (1994); 152: 1523-31.
9. Hermann P, Van-Kooten C, Gaillard C, Banchereau J, Blanchard D. CD40 ligand-positive CD8+ T cell clones allow B cell growth and differentiation. Eur J Immunol (1995); 25: 2972-7.
10. Grewal IS, Flavell RA. CD40 and CD154 in cell-mediated immunity. Annu Rev Immunol (1998); 16: 111-35.
11. Henn V, Slupsky JR, Grafe M, et al. CD40 ligand on activated platelets triggers an inflammatory reaction of endothelial cells. Nature (1998); 391: 591-4.
12. van Kooten C, Banchereau J. CD40-CD40 ligand. J Leukoc Biol (2000); 67: 2-17.
13. Clegg CH, Rulffes JT, Haugen HS, et al. Thymus dysfunction and chronic inflammatory disease in gp39 transgenic mice. Int Immunol (1997); 9: 1111-22.
14. Stüber E, Strober W, Neurath M. Blocking the CD40L-CD40 interaction in vivo specifically prevents the priming of T helper 1 cells through the inhibition of interleukin 12 secretion. J Exp Med (1996); 183: 693-8.
15. Liu Z, Geboes K, Colpaert S, et al. Prevention of experimental colitis in SCID mice reconstituted with CD45RBhigh CD4+ T cells by blocking the CD40-CD154 interactions. J Immunol (2000); 164: 6005-14.
16. De Jong YP, Comiskey M, Кailed SL, et al. Chronic murine colitis is dependent on the CD154/CD40 pathway and can be attenuated by anti-CD154 administration. Gastroenterology (2000); 119: 715-23.
17. Kawai T, Andrews D, Colvin RB, Sachs DH, Cosimi AB. Thromboembolic complications after treatment with monoclonal antibody against CD40 ligand. Nat Med (2000); 6: 114.
18. Buhler L, Alwayn IP, Appel JZ III, Robson SC, Cooper DK. Anti-CD154 monoclonal antibody and thromboembolism. Transplantation (2001); 71: 491.
19. Knosalla C, Gollackner B, Cooper DK. Anti-CD154 monoclonal antibody and thromboembolism revisited. Transplantation (2002); 74: 416-17.
20. de Boer M, Conroy J, Min HY, Kwekkeboom J. Generation of monoclonal antibodies to human lymphocyte cell surface antigens using insect cells expressing recombinant proteins. J Immunol Meth (1992); 152: 15-23.
21. Kwekkeboom J, de Boer M, Tager JM, de Groot C. CD40 plays an essential role in the activation of human B cells by murine EL4B5 cells. Immunology (1993); 79: 439-44.
22. Kwekkeboom J, de Rijk D, Kasran A, Barcy S, de Groot C, de Boer M. Helper effector function of human T cells stimulated by anti-CD3 Mab can be enhanced by co-stimulatory signals and is partially dependent on CD40-CD40 ligand interaction. Eur J Immunol (1994); 24: 508-17.
23. Laman JD, 't Hart BA, Brok, HPM, et al. Protection of marmoset monkeys against EAE by treatment with a murine antibody blocking CD40 (mu5D12). Eur J Immunol (2002); 32: 2218-28.
24. Boon L, Brok HPM, Bauer J, et al. Prevention of experimental autoimmune encephalomyelitis in the common marmoset (Callithrix jacchus) using a chimeric antagonist Mab against human CD40 is associated with altered B-cell responses. J Immunol (2001); 167: 2942-9.
25. Haanstra KG, Ringers J, Sick EA, et al. Prevention of kidney allograft rejection using anti-CD40 and anti-CD86 in primates. Transplantation (2003); 75: 637-43.
26. Haegel-Kronenberger H, Haanstra K, Ziller-Remy C, et al. Inhibition of costimulation allows for repeated systemic administration of adenoviral vector in rhesus monkeys. Gene Ther (2004); 11: 241-52.
27. Boon L, Laman JD, Ortiz-Buijsse A, et al. Preclinical assessment of anti-CD40 Mab 5D12 in cynomolgus monkeys. Toxicology (2002); 174: 53-65.
28. Sartor RB. Pathogenesis and immune mechanisms of chronic inflammatory bowel disease. Am J Gastroenterol (1997); 92: 5S-11S.
29. Fiocchi C. Inflammatory bowel disease: etiology and pathogenesis. Gastroenterology (1998); 15: 182-205.
30. Burgio VL, Fais S, Boirivant M, Perrone A, Pallone F. Peripheral monocyte and naive T-cell recruitment and activation in Crohn's disease. Gastroenterology (1995); 109: 1029-38.
31. Rugtveit J, Brandtzaeg P, Halstensen TS, Fausa O, Scott H. Increased macrophage subset in inflammatory bowel disease: apparent recruitment from peripheral blood monocytes. Gut (1994); 35: 669-74.
32. Fuss IJ, Neurath M, Boirivant M, et al. Disparate CD4+ lamina propria (LP) lymphokine secretion profiles in inflammatory bowel disease. J Immunol (1996); 157: 1261-70.
33. Mullin GE, Maycon ZR, Braun-Elwert L, et al. Inflammatory bowel disease mucosal biopsies have specialized lymphokine mRNA profiles. Inflam Bowel Dis (1996); 2: 16-26.
34. Pospai, D, Rene E, Fiasse R, et al. Crohn's disease stable remission after human immunodeficient virus infection. Dig Dis Sci (1998); 43: 412-9.
35. Stronkhorst A, Radema S, Yong SL, et al. CD4 antibody treatment in patients with active Crohn's disease: a phase 1 dose finding study. Gut (1997); 40: 320-7.
36. Sadlack B, Merz H, Schorle H, Schimpl A, Feller AC, Horak I. Ulcerative colitis-like disease in mice with a disrupted interleukin-2 gene. Cell (1993); 75: 253-61.
37. Kuhn R, Lohler J, Rennick D, Rajewsky K, Muller K. Interleukin-10-deficient mice develop chronic enterocolitis. Cell (1993); 75: 263-74.
38. Powrie F, Leach MM, Mauze S, Menon S, Caddle LB, Coffman RA. Inhibition of Th1 responses prevents inflammatory bowel disease in SCID mice reconstituted with CD45RBhi CD4+ T cells. Immunity (1994); 1: 553-62.
39. Liu Z, Colpaert S, D'Haens GR, et al. Hyperexpression of CD40 Ligand (CD154) in inflammatory bowel disease and its role in pathogenic cytokine production. J Immunol (1999); 163: 4049-57.
40. D'Haens G, Geboes K, Peeters M, Baert F, Penninckx F, Rutgeerts P. Early lesions caused by infusion of intestinal contents in excluded ileum in Crohn's disease. Gastroenterology (1998); 114: 262-7.
41. Cornillie F, Shealy D, D'Haens G, et al. Infliximab induces potent anti-inflammatory and local immunomodulatory activity but no systemic immune suppression in patients with Crohn's disease. Aliment Pharmacol Ther (2001); 15: 2041-2.
42. Inwald DP, McDowall A, Peters MJ, Callard RE, Klein NJ. CD40 is constitutively expressed on platelets and provides a novel mechanism for platelet activation. Circ Res (2003); 92: 1041-8.
43. Van Assche G, Rutgeerts P. Anti-TNF agents in Crohn's disease. Expert Opin Investig Drugs (2000); 9: 103-11.
44. Stüber E, Strober W, Neurath M. Blocking the CD40L-CD40 interaction in vivo specifically prevents the priming of T helper 1 cells through the inhibition of interleukin 12 secretion. J Exp Med (1996); 183: 693-698.
45. Ranheim EA, Kipps TJ. Activated T cells induce expression of B7/BB1 on normal or leukemic B cells through a CD40-dependent signal. J Exp Med (1993); 177:925-935.
46. Hasbold J, Johnson-Leger C, Atkins CJ, Clark EA, Klaus GGB. Properties of mouse CD40: cellular distribution of CD40 and B cell activation by monoclonal anti-mouse CD40 antibodies. Eur J Immunol (1994); 24: 1835-1842.
47. Alderson MR, Armitage RJ, Tough TW, Strockbine L, Fanslow WC, Spriggs MK. CD40 expression by monocytes: regulation by cytokines and activation of monocytes by the ligand for CD40. J Exp Med (1993); 178: 669-674.
48. Kiener PA, Moran-Davis P, Rankin BM, Wahl AF, Aruffo A, Hollenbaugh D. Stimulation of CD40 with purified soluble gp39 induces proinflammatory responses in human monocytes. J Immunol (1995); 155: 4917-4925.
49. Shu U, Kiniwa M, Wu CY, Maliszewski C, Vezzio N, Hakimi J, Gately M, Delespesse G. Activated T cells induce interleukin-12 production by monocytes via CD40-CD40 ligand interaction. Eur J Immunol (1995); 25: 1125-1128.
50. Cella M, Scheidegger D, Palmer-Lehmann K, Lane P, Lanzavecchia A, Alber G. Ligation of CD40 on dendritic cells triggers production of high levels of interleukin-12 and enhances T cell stimulatory capacity: T-T help via APC activation. J Exp Med (1996); 184: 747-752.
51. van Kooten C, Banchereau J. Functions of CD40 on B cells, dendritic cells and other cells. Curr Opin Immunol (1997); 9: 330-337.
52. Cayabyab M, Phillips JH, Lanier LL. CD40 preferentially costimulates activation of CD4+ T lymphocytes. J Immunol (1994); 152: 1523-1531.
53. Hermann P, Van-Kooten C, Gaillard C, Banchereau J, Blanchard D. CD40 ligand-positive CD8+ T cell clones allow B cell growth and differentiation. Eur J Immunol (1995); 25: 2972-2977.
54. Grewal IS, Flavell RA. CD40 and CD154 in cell-mediated immunity. Annu Rev Immunol (1998); 16: 111-135.
55. Henn V, Slupsky JR, Grafe M, Anagnostopoulos I, Forster R, Muller-Berghaus G, Kroczek RA. CD40 ligand on activated platelets triggers an inflammatory reaction of endothelial cells. Nature (1998); 391: 591-594.
56. Banchereau J, Bazan F, Blanchard D, Briere F, Galizzi JP, van Kooten C, Liu YJ, Saeland S. The CD40 antigen and its ligand. Annu Rev Immunol (1994); 12: 881-922.
57. Foy TM, Aruffo A, Bajorath J, Buhlmann JE, Noelle RJ. Immune regulation by CD40 and its ligand gp39. Annu Rev Immunol (1996); 14: 591-617.
58. Kato T, Hakamada R, Yamane H, Nariuchi H. Induction of IL-12 p40 messenger RNA expression and IL-12 production of macrophages via CD40-CD40 ligand interaction. J Immunol (1996); 156: 3932-3938.
59. Ludewig B, Graf D, Gelderblom HR, Becker Y, Kroczek RA, Pauli G. Spontaneous apoptosis of dendritic cells is efficiently inhibited by TRAP (CD40-ligand) and TNF-a, but strongly enhanced by interleukin-10. Eur J Immunol (1995); 25: 1943-1950.
60. Allen RC, Armitage RJ, Conley ME, Rosenblatt H, Jenkins NA, Copeland NG, Bedell MA, Edelhoff S, Disteche CM, Simoneaux DK, Fanshow WC, Belmont J, Spriggs MK. CD40 ligand gene defects responsible for X-linked hyper-IgM syndrome. Science (1993); 259: 990-993.
61. Xu J, Foy TM, Laman JD, Elliott EA, Dunn JJ, Waldschmidt TJ, Elsemore J, Noelle RJ, Flavell RA. Mice deficient for the CD40 ligand. Immunity (1994); 1: 423-431.
62. Grewal IS, Xu J, Flavell RA. Impairment of antigen-specific T-cell priming in mice lacking CD40 ligand. Nature (1995); 378: 617-620.
63. Stout RD, Suttles J, Xu J, Grewal IS, Flavell RA. Impaired T cell-mediated macrophage activation in CD40 ligand-deficient mice. J Immunol (1996); 156: 8-11.
64. Levy J, Espanol-Boren T, Thomas C, Fischer A, Tovo P, Bordigoni P, Resnick I, Fasth A, Baer M, Gomez L, Sanders EA, Tabone MD, Plantaz D, Etzioni A, Monafo V, Abinun M, Hammarstrom L, Abrabamsen T, Jones A, Finn A, Klemola T, DeVries E, Sanal O, Peitsch MC, Notarangelo LD. Clinical spectrum of X-linked hyper-IgM syndrome. J Pediatr (1997);.131: 47-54.
65. Soong L, Xu JC, Grewal IS, Kima P, Sun J, Longley BJ Jr, Ruddle NH, McMahon-Pratt D, Flavell RA. Disruption of CD40-CD40 ligand interactions results in an enhanced susceptibility to Leishmania amazonensis infection. Immunity (1996); 4:263-273.
66. Campbell KA, Ovendale PJ, Kennedy MK, Fanslow WC, Reed SG, Maliszewski CR. CD40 ligand is required for protective cell-mediated immunity to Leishmania major. Immunity (1996); 4: 283-289.
67. Clegg CH, Rulffes JT, Haugen HS, Hoggatt IH, Aruffo A, Durham SK, Farr AG, Hollenbaugh D. Thymus dysfunction and chronic inflammatory disease in gp39 transgenic mice. Int Immunol (1997); 9: 1111-1122.
68. Parker et al., Proc. Nat. Acad. Sci. USA (1995) 92: 9560.
69. Haanstra KG, Ringers J, Sick EA, Ramdien-Murli S, Kuhn EM, Boon L, Jonker M. Prevention of kidney allograft rejection using anti-CD40 and anti-CD86 in primates. Transplantation. (2003); 75(5): 637-43.
70. Haanstra KG, Sick EA, Ringers J, Wubben JAM, Kuhn EM, Boon L, Jonker M. Costimulation blockade followed by a 12-week period of cyclosporine A facilitates prolonged drug-free survival of Rhesus Monkey kidney allografts. Transplantation. (2005); 79: 1623-1626.
71. Boon L, Brok HP, Bauer J, Ortiz-Buijsse A, Schellekens MM, Ramdien-Murli S et al. Prevention of experimental autoimmune encephalomyelitis in the common marmoset (Callithrix jacchus) using a chimeric antagonist monoclonal antibody against human CD40 is associated with altered B cell responses. J Immunol (2001); 167: 2942-2949.
72. Laman JD, 't Hart BA, Brok H, Meurs M, Schellekens MM, Kasran A et al. Protection of marmoset monkeys against EAE by treatment with a murine antibody blocking CD40 (mu5D12). Eur J Immunol (2002); 32: 2218-2228.
73. Boon L, Laman JD, Ortiz-Buijsse A, den Hartog MT, Hoffenberg S, Liu P et al. Preclinical assessment of anti-CD40 Mab 5D12 in cynomolgus monkeys. Toxicology (2002); 174: 53-65.
74. Kasran A, Boon L, Wortel CH, Hogezand RA, Schreiber S, Goldin E, Boer M, Geboes K, Rutgeerts P, Ceuppens JL. Safety and tolerability of antagonist anti-human CD40 Mab ch5D12 in patients with moderate to severe Crohn's disease. Aliment Pharmacol Ther. (2005); 22: lll-22.
75. Haanstra KG, Sick EA, Ringers J, Wubben JA, Kuhn EM, Boon L, Jonker M. Costimulation blockade followed by a 12-week period of cyclosporine A facilitates prolonged drug-free survival of rhesus monkey kidney allografts. Transplantation. (2005); 79: 1623-6.
76. 't Hart BA, Blezer EL, Brok HP, Boon L, de Boer M, Bauer J, Laman JD. Treatment with chimeric anti-human CD40 antibody suppresses MRI-detectable inflammation and enlargement of pre-existing brain lesions in common marmosets affected by MOG-induced EAE. J Neuroimmunol. (2005); 163: 31-9.
77. Koenen HJ, den Hartog MT, Heerkens S, Fasse E, Ortiz-Buijsse A, van Neerven RJ, Simons PJ, Joosten I, Boon L. A novel bispecific antihuman CD40/CD86 fusion protein with t-cell tolerizing potential. Transplantation. (2004); 78: 1429-38.
78. de Vos AF, Melief MJ, van Riel D, Boon L, van Eijk M, de Boer M, Laman JD. Antagonist anti-human CD40 antibody inhibits germinal center formation in cynomolgus monkeys. Eur J Immunol. (2004); 34: 3446-55.
79. Laman JD, 't Hart BA, Brok H, Meurs M, Schellekens MM, Kasran A, Boon L, Bauer J, Boer M, Ceuppens J. Protection of marmoset monkeys against EAE by treatment with a murine antibody blocking CD40 (mu5D12). Eur J Immunol. (2002); 32: 2218-28.
80. Boon L, Laman JD, Ortiz-Buijsse A, den Hartog MT, Hoffenberg S, Liu P, Shiau F, de Boer M. Preclinical assessment of anti-CD40 Mab 5D12 in cynomolgus monkeys. Toxicology. (2002); 174: 53-65.
81. Boon L, Brok HP, Bauer J, Ortiz-Buijsse A, Schellekens MM, Ramdien-Murli S, Blezer E, van Meurs M, Ceuppens J, de Boer M, 't Hart BA, Laman JD. Prevention of experimental autoimmune encephalomyelitis in the common marmoset (Callithrix jacchus) using a chimeric antagonist monoclonal antibody against human CD40 is associated with altered B cell responses. J Immunol. (2001); 167: 2942-9.
82. S.G. Antunes, N.G. de Groot, H. Brok, G. Doxiadis, A.A. Menezes, N. Otting and R.E. Bontrop, The common marmoset: a new world primate species with limited Mhc class II variability, Proc. Natl. Acad. Sci. U. S. A. (1998); 95: 11745-11750.
83. B. Becher, B.G. Durell, A.V. Miga, W.F. Hickey and R.J. Noelle, The clinical course of experimental autoimmune encephalomyelitis and inflammation is controlled by the expression of CD40 within the central nervous system, J. Exp. Med. (2001); 193: 967-974.
84. R.E. Bontrop, N. Otting, N.G. de Groot and G.G. Doxiadis, Major histocompatibility complex class II polymorphisms in primates, Immunol. Rev. (1999); 167: 339-350.
85. H.P. Brok, A. Uccelli, N. Kerlero De Rosbo, R.E. Bontrop, L. Roccatagliata, N.G. de Groot, E. Capello, J.D. Laman, K. Nicolay, G.L. Mancardi, A. Ben-Nun and B.A. 't Hart, Myelin/oligodendrocyte glycoprotein-induced autoimmune encephalomyelitis in common marmosets: the encephalitogenic T cell epitope pMOG24-36 is presented by a monomorphic MHC class II molecule, J. Immunol. (2000); 165: 1093-1101.
86. H.P. Brok, J. Bauer, M. Jonker, E. Blezer, S. Amor, R.E. Bontrop, J.D. Laman and B.A. 't Hart, Non-human primate models of multiple sclerosis, Immunol. Rev. (2001); 183: 173-185.
87. C.P. Genain and S.L. Hauser, Experimental allergic encephalomyelitis in the New World monkey Callithrix jacchus, Immunol. Rev. (2001); 183: 159-172.
88. K. Gerritse, J.D. Laman, R.J. Noelle, A. Aruffo, J.A. Ledbetter, W.J. Boersma and E. Claassen, CD40-CD40 ligand interactions in experimental allergic encephalomyelitis and multiple sclerosis, Proc. Natl. Acad. Sci. U.S.A. (1996); 93: 2499-2504.
89. A.M. Girvin, M.C. Dal Canto and S.D. Miller, CD40/CD40L interaction is essential for the induction of EAE in the absence of CD28-mediated co-stimulation, J. Autoimmun.(2002); 18: 83-94.
90. IS. Grewal and R.A. Flavell, A central role of CD40 ligand in the regulation of CD4+ T-cell responses, Immunol. Today (1996); 17: 410-414.
91. L.M. Howard, A.J. Miga, C.L. Vanderlugt, M.C. Dal Canto, J.D. Laman, R.J. Noelle and S.D. Miller, Mechanisms of immunotherapeutic intervention by anti-CD40L (CD154) antibody in an animal model of multiple sclerosis, J. Clin. Invest. (1999); 103: 281-290.
92. E.K. Jordan, H.I. McFarland, B.K. Lewis, N. Tresser, M.A. Gates, M. Johnson, M. Lenardo, L.A. Matis, H.F. McFarland and J.A. Frank, Serial MR imaging of experimental autoimmune encephalomyelitis induced by human white matter or by chimeric myelin-basic and proteolipid protein in the common marmoset, AJNR Am. J. Neuroradiol. (1999); 20: 965-976.
93. J.D. Laman, E. Claassen and R.J. Noelle, Functions of CD40 and its ligand, gp39 (CD40L), Crit. Rev. Immunol.(1996); 16: 59-108.
94. J.D. Laman, C.B. Maassen, M.M. Schellekens, L. Visser, M. Kap, E. de Jong, M. van Puijenbroek, M.J. van Stipdonk, M. van Meurs, C. Schwarzler and U. Gunthert, Therapy with antibodies against CD40L (CD154) and CD44-variant isoforms reduces experimental autoimmune encephalomyelitis induced by a proteolipid protein peptide, Mult. Scler. (1998); 4: 147-153.
95. J.D. Laman, M. van Meurs, M.M. Schellekens, M. de Boer, B. Melchers, L. Massacesi, H. Lassmann, E. Claassen and B.A. 't Hart, Expression of accessory molecules and cytokines in acute EAE in marmoset monkeys (Callithrix jacchus), J. Neuroimmunol. (1998); 86: 30-45.
96. J. Mestas and C.C. Hughes, Of mice and not men: differences between mouse and human immunology, J. Immunol. (2004); 172: 2731-2738.
97. S.A. Quezada, L.Z. Jarvinen, E.F. Lind and R.J. Noelle, CD40/CD154 interactions at the interface of tolerance and immunity, Annu. Rev. Immunol. (2004); 22: 307-328.
98. C.S. Raine, B. Cannella, S.L. Hauser and C.P. Genain, Demyelination in primate autoimmune encephalomyelitis and acute multiple sclerosis lesions: a case for antigen-specific antibody mediation, Ann. Neurol. (1999); 46: 144-160.
99. D.H. Sachs, Tolerance: of mice and men, J. Clin. Invest. (2003); 111: 1819-1821.
100. E.B. Samoilova, J.L. Horton, H. Zhang and Y. Chen, CD40L blockade prevents autoimmune encephalomyelitis and hampers TH1 but not TH2 pathway of T cell differentiation, J. Mol. Med. (1997); 75: 603-608.
101. B.A. 't Hart, J. Bauer, H.J. Muller, B. Melchers, K. Nicolay, H. Brok, R.E. Bontrop, H. Lassmann and L. Massacesi, Histopathological characterization of magnetic resonance imaging-detectable brain white matter lesions in a primate model of multiple sclerosis: a correlative study in the experimental autoimmune encephalomyelitis model in common marmosets (Callithrix jacchus), Am. J. Pathol. (1998); 153: 649-663.
102. B.A. 't Hart, M. van Meurs, H.P. Brok, L. Massacesi, J. Bauer, L. Boon, R.E. Bontrop and J.D. Laman, A new primate model for multiple sclerosis in the common marmoset, Immunol. Today (2000); 21: 290-297.
103. B. 't Hart, S. Amor and M. Jonker, Evaluating the validity of animal models for research into therapies for immune-based disorders, Drug Discov. Today (2004); 9: 517-524.
104. 't Hart et al, 2004b B.A. 't Hart, J.D. Laman, J. Bauer, E.D. Blezer, Y. van Kooyk and R.Q. Hintzen, Modelling of multiple sclerosis: lessons learned in a non-human primate, Lancet Neurol (2004); 3: 589-597.
105. B.A. 't Hart, J. Vogels, J. Bauer, H.P. Brok and E. Blezer, Non-invasive measurement of brain damage in a primate model of multiple sclerosis, Trends Mol. Med. (2004); 10: 85-91.
106. B.A. 't Hart, J.T. Vogels, J. Bauer, H.P.M. Brok and E. Blezer, Non-invasive measurement of brain damage in a primate model of multiple sclerosis, Trends Mol. Med. (2004); 10: 85-91.
107. P. Villoslada, K. Abel, N. Heald, R. Goertsches, S.L. Hauser and C.P. Genain, Frequency, heterogeneity and encephalitogenicity of T cells specific for myelin oligodendrocyte glycoprotein in naive outbred primates, Eur. J. Immunol. (2001); 31: 2942-2950.
108. Bata-Csorgo Z, Hammerberg C, Voorhees JJ, Cooper KD. Intralesional T-lymphocyte activation as a mediator of psoriatic epidermal hyperplasia. J Invest Dermatol (1995); 105(1 Suppl): 89S-94S.
109. Bos JD, De Rie MA. The pathogenesis of psoriasis: immunological facts and speculations. Immunol Today (1999); 20: 40-46.
110. Lebwohl M. Psoriasis. Lancet (2003); 361: 1197-1204.
111. van Kooten C, Banchereau J. CD40-CD40 ligand. J Leukoc Biol (2000); 67: 2-17.
112. Peguet-Navarro J, Dalbiez-Gauthier C, Moulon C, et al. CD40 ligation of human keratinocytes inhibits their proliferation and induces their differentiation. J Immunol (1997); 158: 144-152.
113. Denfeld RW, Hollenbaugh D, Fehrenbach A, et al. CD40 is functionally expressed on human keratinocytes. Eur J Immunol (1996); 26: 2329-2334.
114. Gaspari AA, Sempowski GD, Chess P, Gish J, Phipps RP. Human epidermal keratinocytes are induced to secrete interleukin-6 and co-stimulate T lymphocyte proliferation by a CD40-dependent mechanism. Eur J Immunol (1996); 26: 1371-1377.
115. Pasch MC, Bos JD, Asghar SS. Activation of complement from psoriasis. Clin Exp Dermatol (1998); 23: 189-190.
116. Pasch MC, Timar, K, van Meus M, Heyendaal VMR, Bos JD, Laman JD, Ashgar SS. In situ demonstration of CD40- and CD154-positive cells in psoriatic lesions and keratinocyte production of chemokines by CD40 ligation in vitro. J. Pathol (2004); 203:839-848.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИИ И СПОСОБЫ ИММУНОТЕРАПИИ | 2014 |
|
RU2680010C2 |
| ПРИМЕНЕНИЕ АНТИ-CD40-АНТИТЕЛ | 2006 |
|
RU2442606C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ИММУНОТЕРАПИИ | 2014 |
|
RU2798348C2 |
| ЛЕЧЕНИЕ В-КЛЕТОЧНЫХ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ С ИСПОЛЬЗОВАНИЕМ АНТИТЕЛ ПРОТИВ CD40L В КОМБИНАЦИИ С АНТИТЕЛАМИ ПРОТИВ CD20 И/ИЛИ ХИМИОТЕРАПИЕЙ И ЛУЧЕВОЙ ТЕРАПИЕЙ | 2000 |
|
RU2305561C2 |
| АНТИТЕЛА К CD40 ДЛЯ ПРИМЕНЕНИЯ В ЛЕЧЕНИИ СИНДРОМА ШЕГРЕНА | 2018 |
|
RU2790558C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ АУТОИММУННЫХ ЗАБОЛЕВАНИЙ С ИСПОЛЬЗОВАНИЕМ КОМБИНАЦИИ ЭЛИМИНИРУЮЩЕГО В-КЛЕТКИ/ИММУНОРЕГУЛЯТОРНОГО АНТИТЕЛ | 2001 |
|
RU2285541C2 |
| АНТИ-CD40-АНТИТЕЛА И СПОСОБЫ ПРИМЕНЕНИЯ | 2012 |
|
RU2609647C2 |
| ОДНОЦЕПОЧЕЧНЫЕ БЕЛКИ-АГОНИСТЫ РЕЦЕПТОРА CD40 | 2016 |
|
RU2745801C2 |
| ПРИМЕНЕНИЯ АНТИ-CD40-АНТИТЕЛ | 2008 |
|
RU2491095C2 |
| АНТАГОНИСТ CD40L И ПУТИ ЕГО ПРИМЕНЕНИЯ | 2019 |
|
RU2812919C2 |
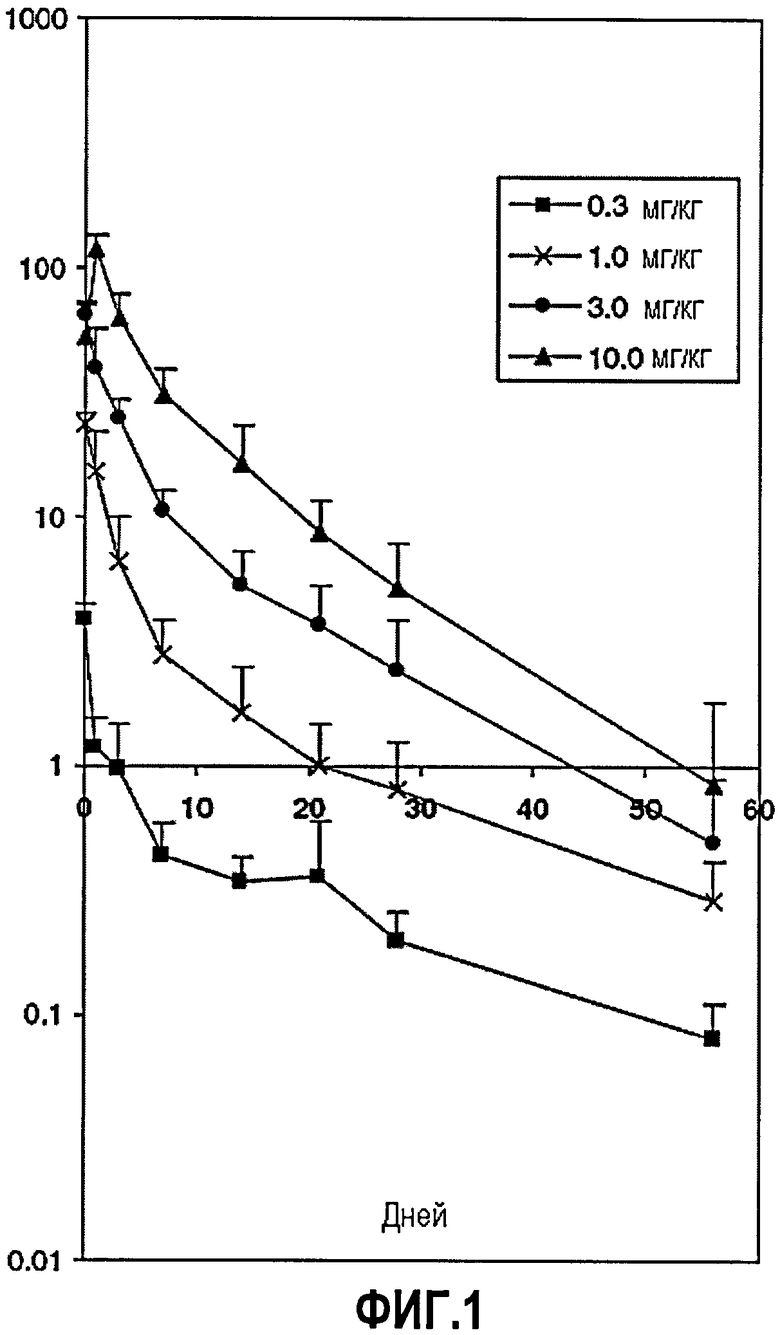
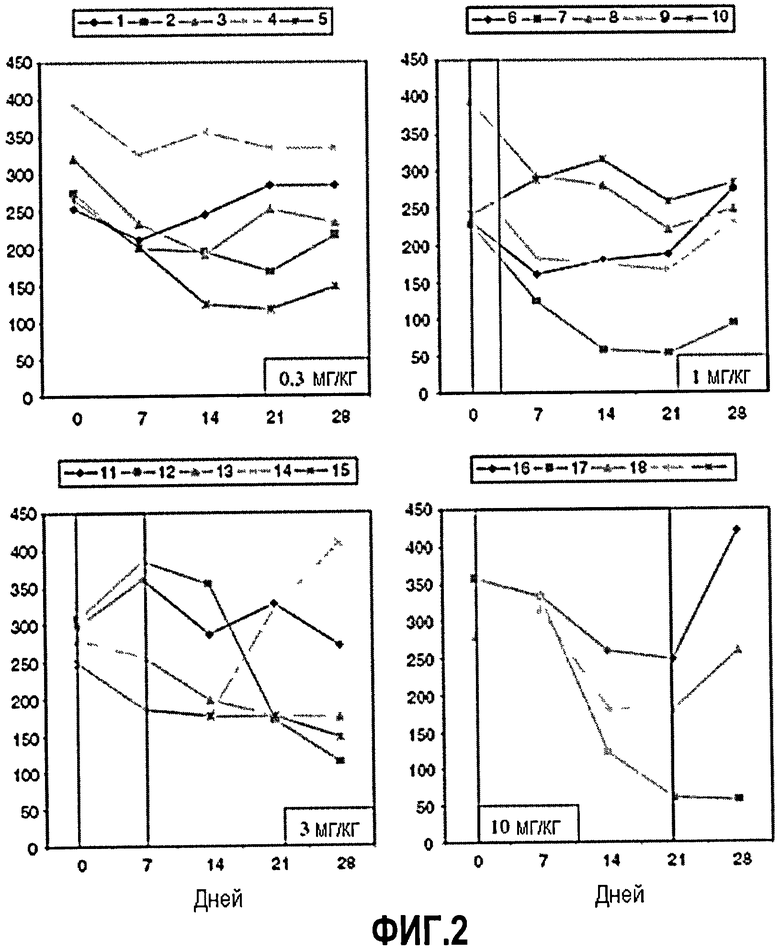
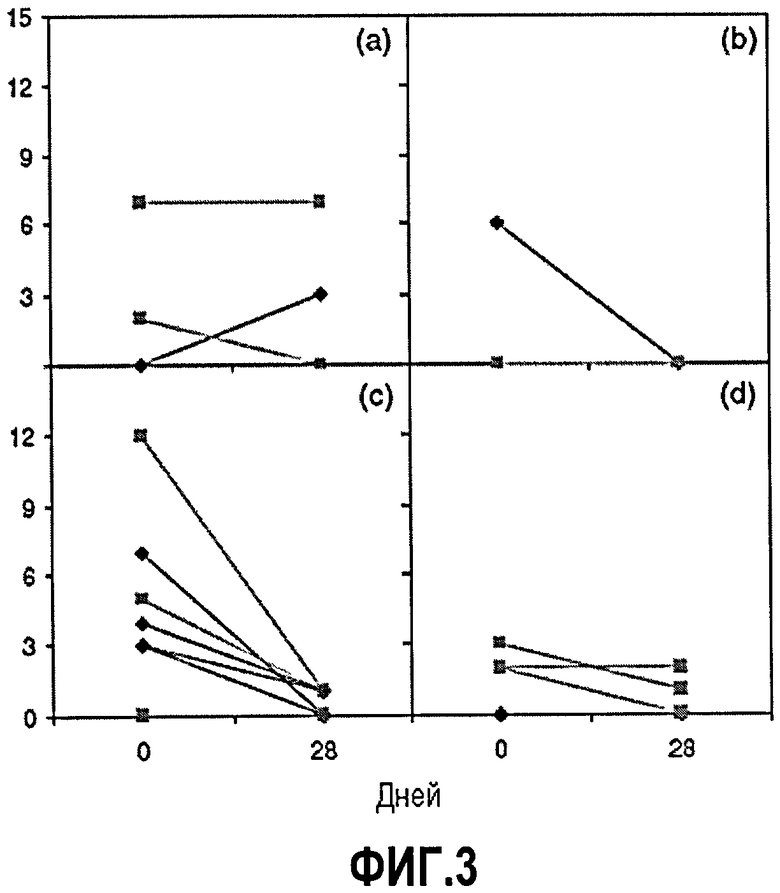
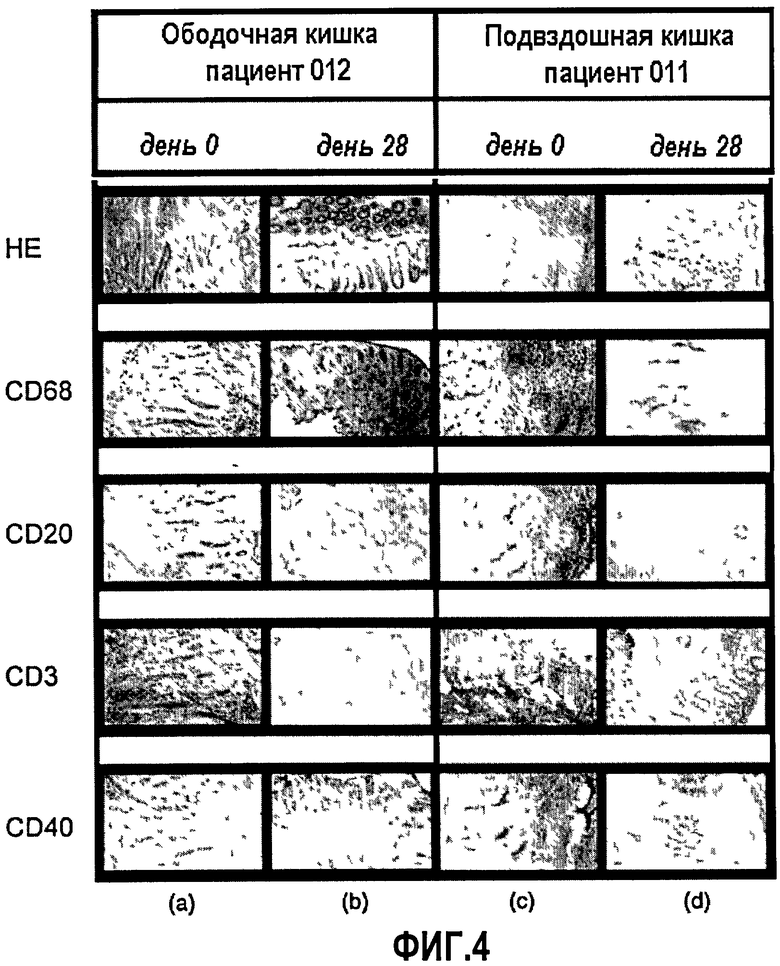
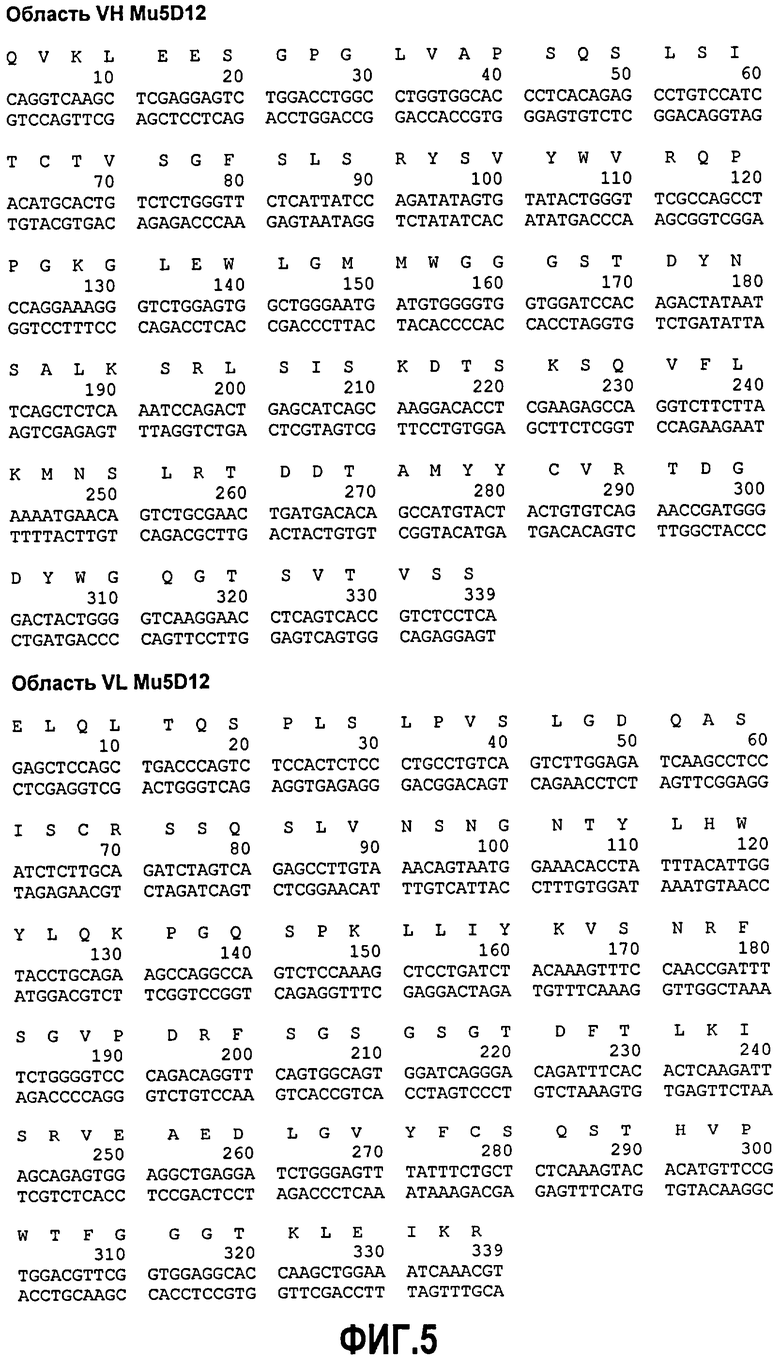
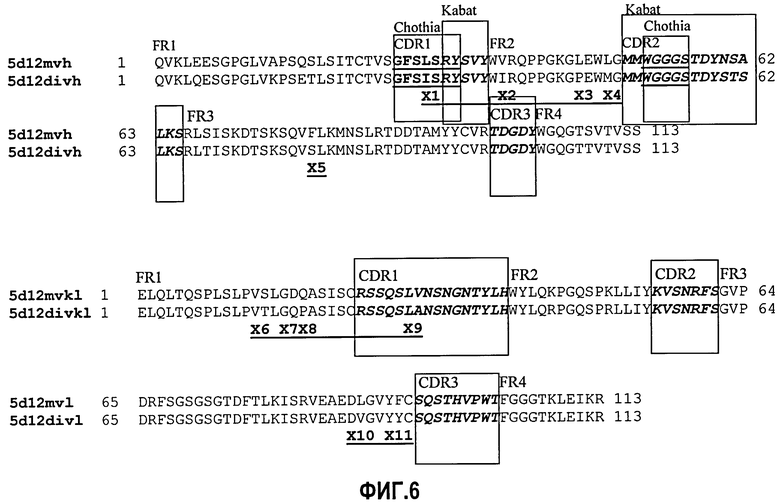
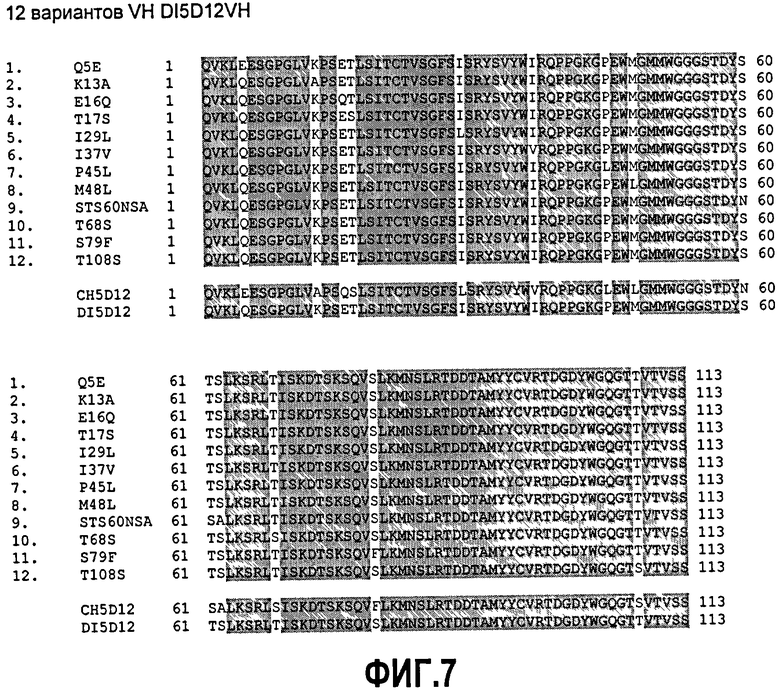
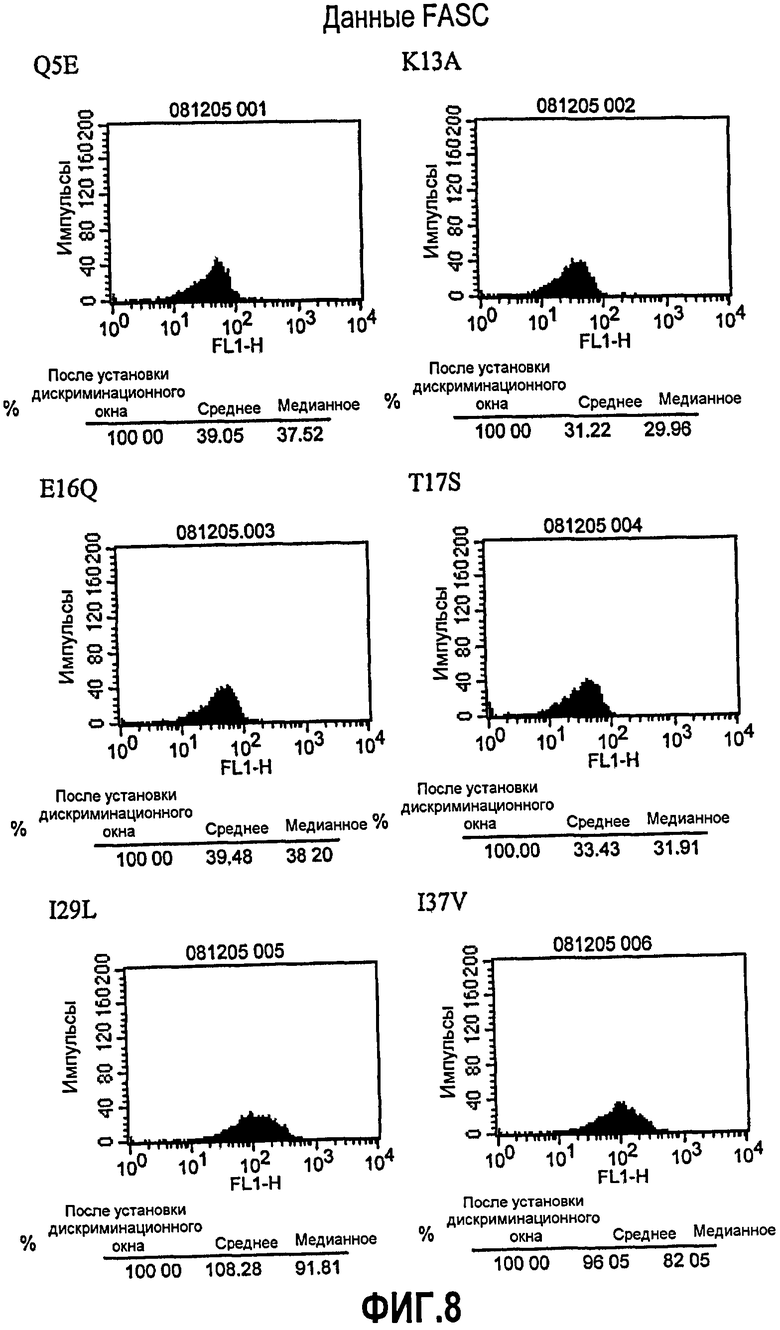
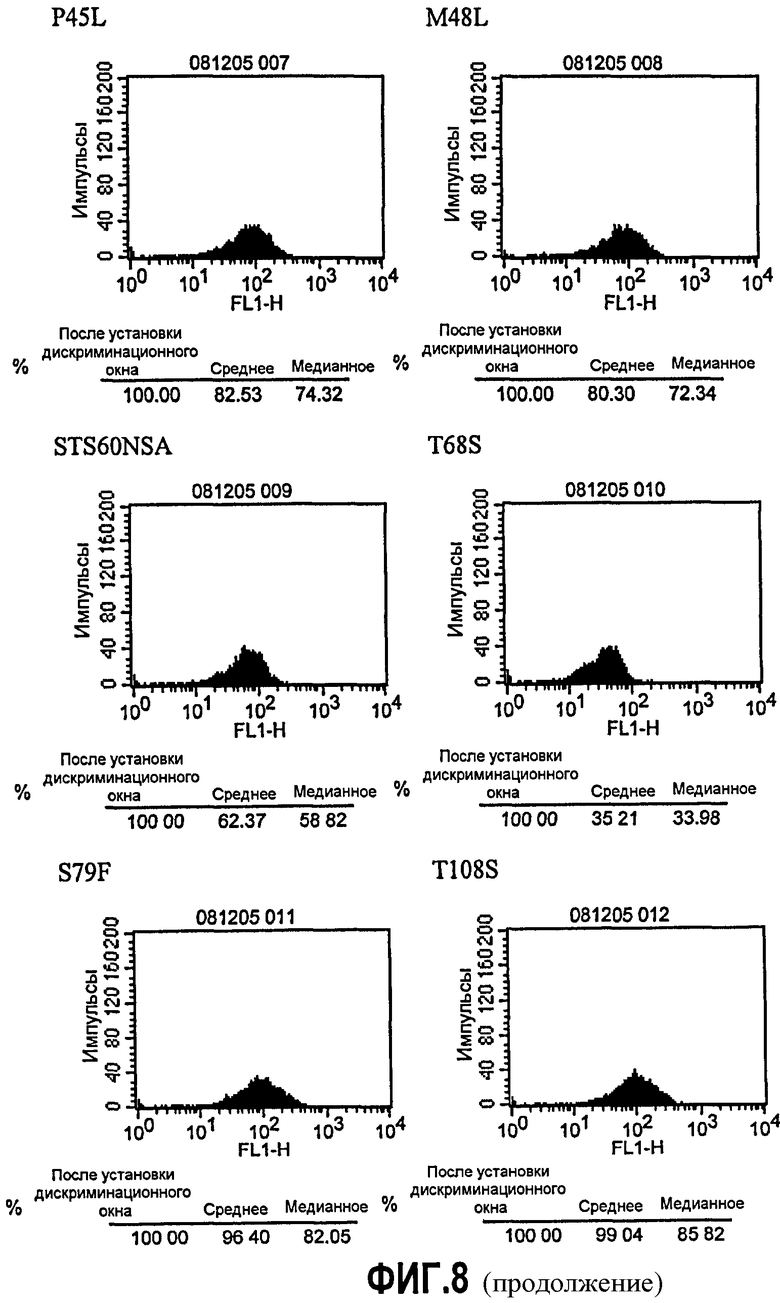
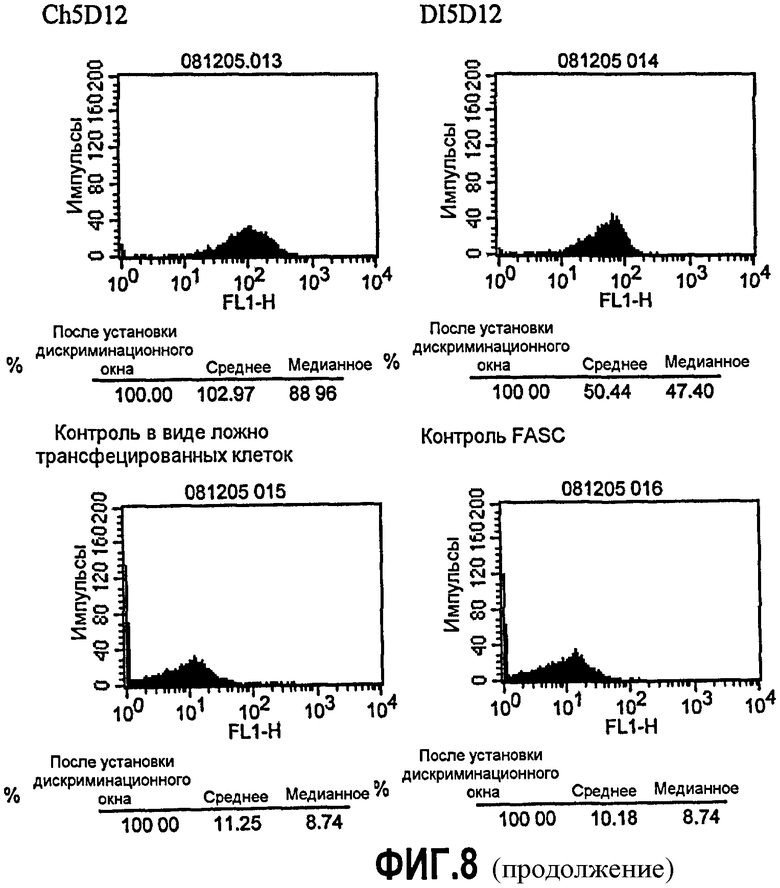
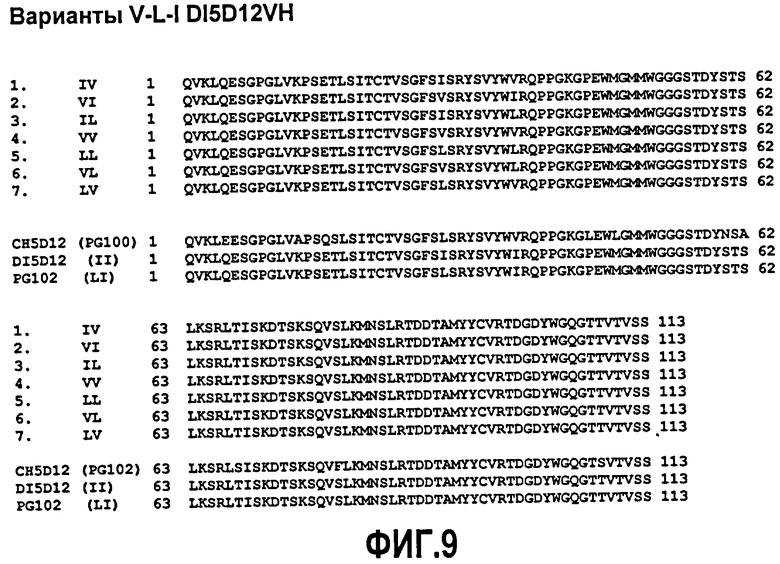
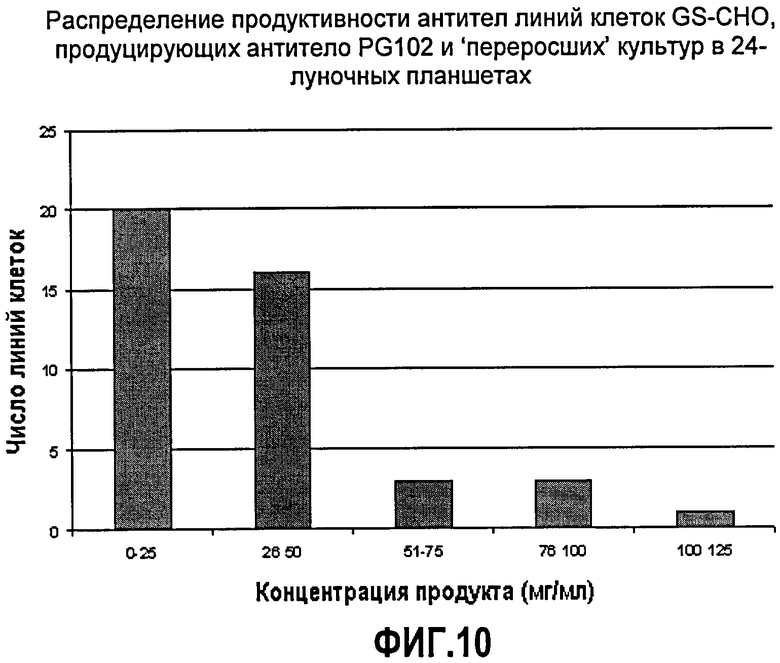
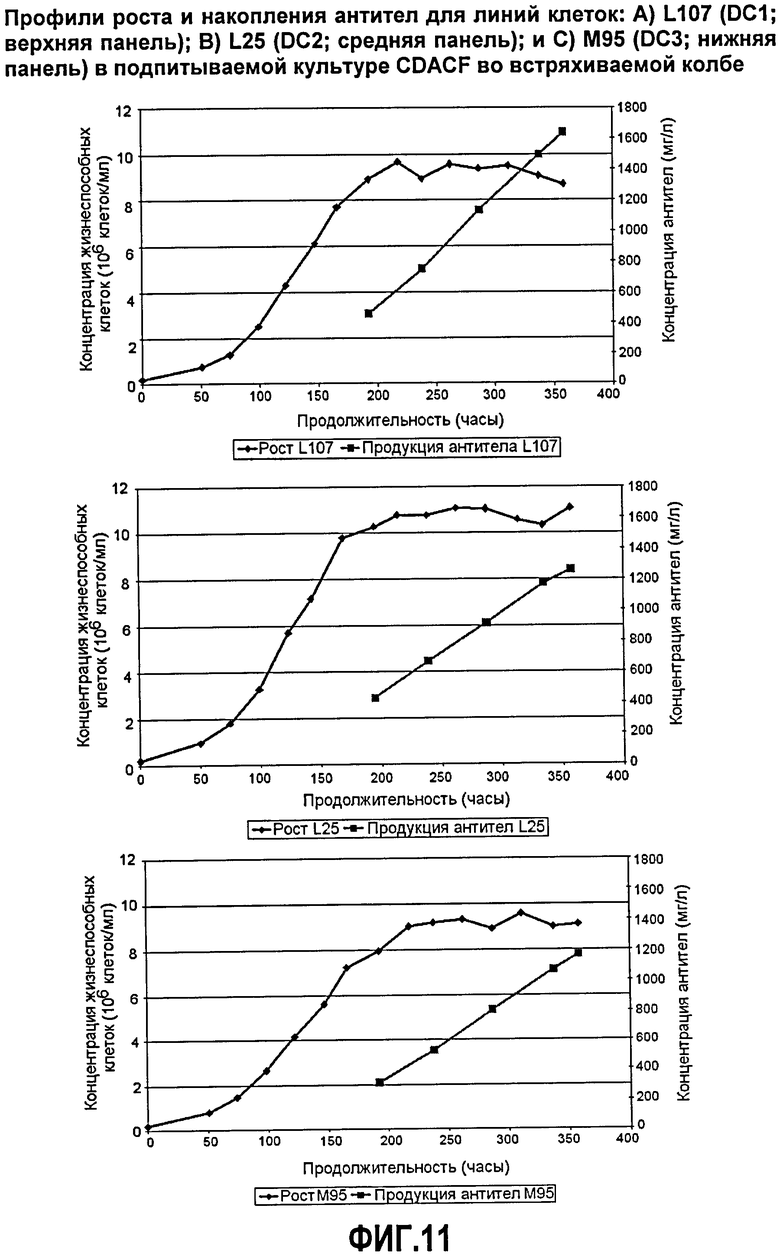
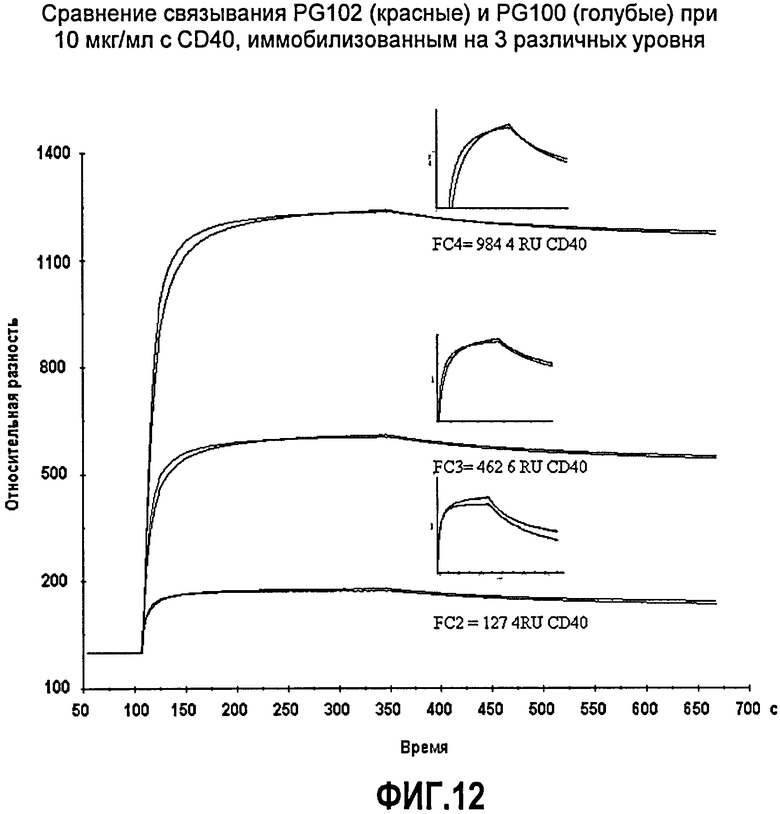
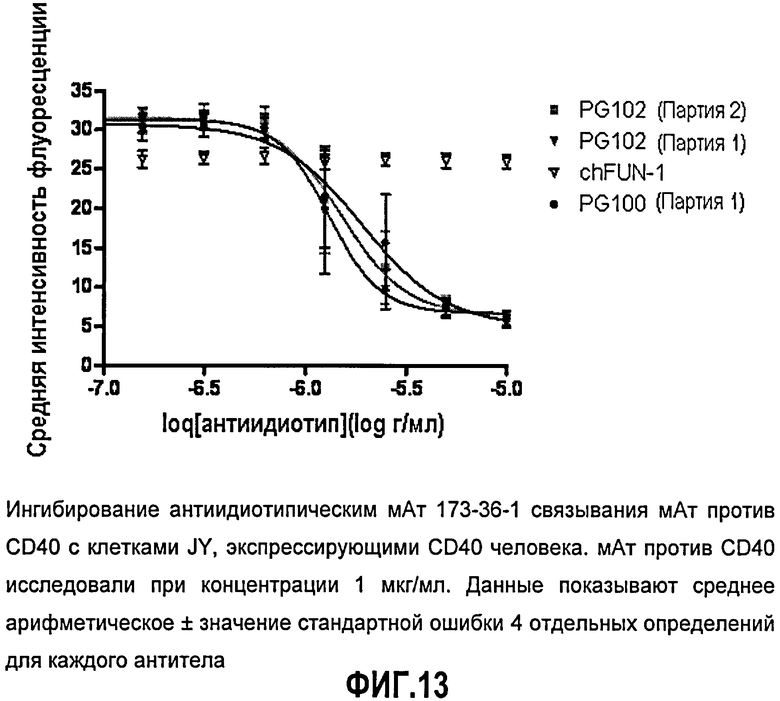
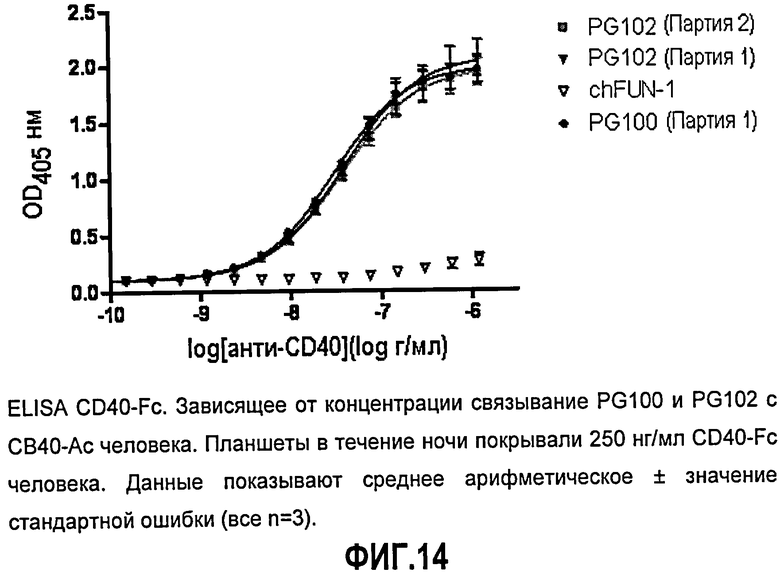
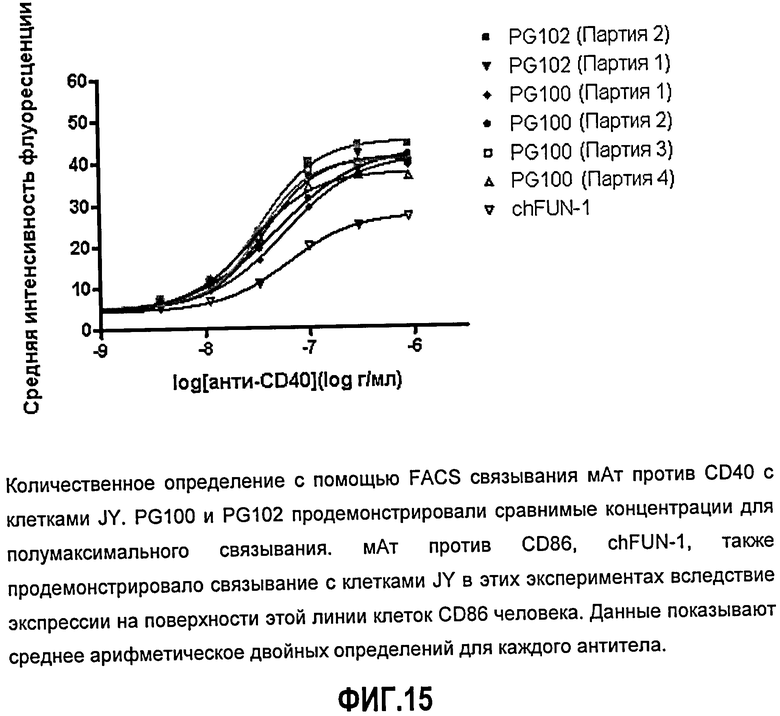
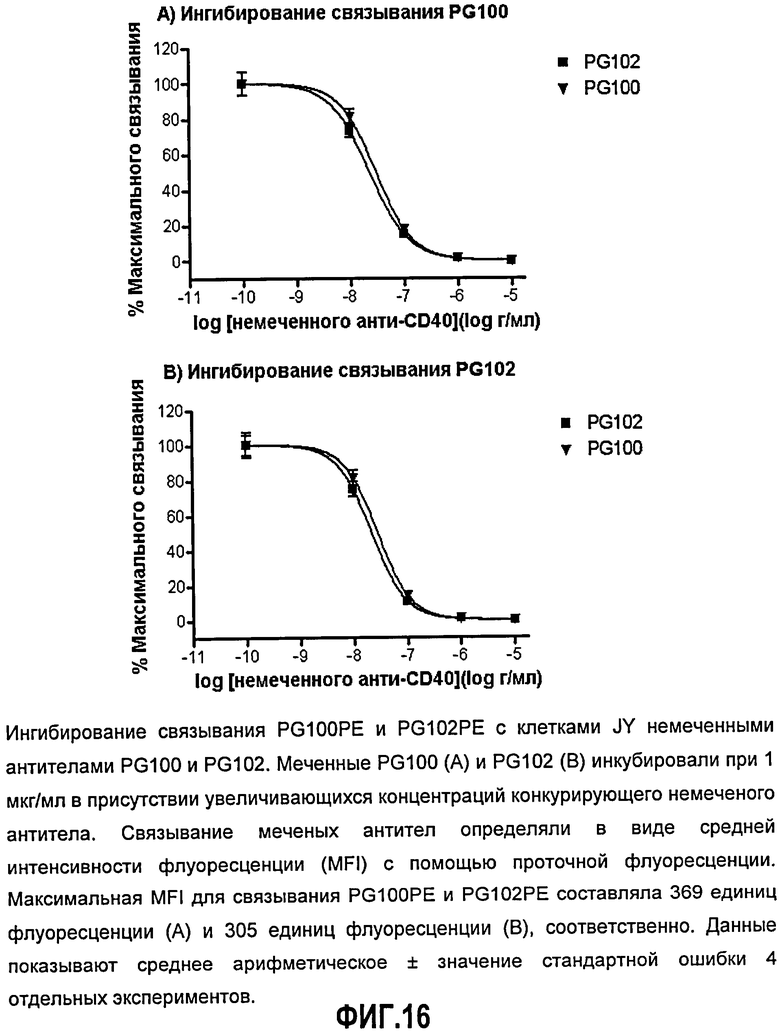
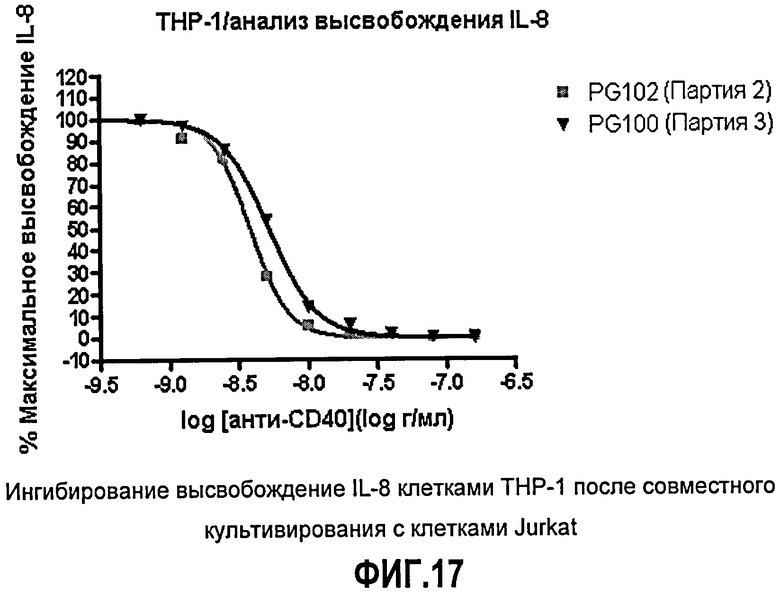

Настоящее изобретение относится к области иммунологии. Предложено моноклональное антитело против CD40 с пониженной иммуногенностью, полученное из химерного антитела 5D12 (ch5D12). Также описаны: нуклеиновая кислота, клетка и культура клеток для получения антитела по изобретению, способ его получения, фармацевтическая композиция, применение антитела для получения лекарственного средства и способ введения индивидууму антитела по изобретению для облегчения симптомов аутоиммунного заболевания, воспалительного заболевания, для уменьшения реакции отторжения трансплантата и/или для лечения CD40-позитивного рака. Рассмотрен способ отбора антител против CD40 человека с повышенной экспрессией, содержащих вставку, делецию, инверсию и/или замену от 1 до 5 аминокислот по сравнению с антителом по изобретению, и антитело, полученное таким способом отбора. 10 н. и 21 з.п. ф-лы, 18 ил., 14 табл., 10 пр.
1. Антитело против CD40, содержащее вариабельную область легкой цепи с аминокислотной последовательностью

где X6 представляет собой N, Q, S, T, Y, W или C;
X7 представляет собой D, E, N, Q, S, T, Y, W или C;
X8 представляет собой N, Q, S, T, Y, G, A, V, L, I, P, F, М, W или C;
X9 представляет собой A, V;
X10 представляет собой G, A, V, L, I, P, F, M; и
X11 представляет собой N, Q, S, T, Y, G, A, V, L, I, P, F, M, W или C,
и вариабельную область тяжелой цепи с аминокислотной последовательностью

где X1 представляет собой I, а X2 представляет собой V;
X1 представляет собой L, а X2 представляет собой I;
X1 представляет собой V, а X2 представляет собой V;
X1 представляет собой L, а X2 представляет собой L; или
X1 представляет собой L, а X2 представляет собой V; а
X3 представляет собой G, A, V, L, I, P, F, M, W, C, N, Q, S, T, Y, D, E, K, R или H;
X4 представляет собой G, A, V, L, I, P, F, M, W, C, N, Q, S, T, Y, D, E, K, R или H; и
X5 представляет собой G, A, V, L, I, P, F, M, W, C, N, Q, S, T, Y, D, E, K, R или H.
2. Антитело по п.1,
где X3 представляет собой G, A, V, L, I, P, F, M;
X4 представляет собой G, A, V, L, I, P, F, M; и
X5 представляет собой G, A, V, L, I, P, F, M, W, C, N, Q, S, T или Y.
3. Антитело по любому из пп.1 или 2,
где X3 представляет собой G, A, V, L, I, P, F, M;
X4 представляет собой G, A, V, L, I или M; и
X5 представляет собой P, F, W, N, Q, S, T или Y.
4. Антитело по п.3,
в котором X1 представляет собой L;
X2 представляет собой I;
X3 представляет собой P; и
X4 представляет собой M; или X5 представляет собой S.
5. Антитело по п.1,
в котором X3 представляет собой P;
X4 представляет собой M; и
X5 представляет собой F или X5 представляет собой S, предпочтительно S.
6. Антитело по п.1,
в котором X1 представляет собой L; X2 представляет собой V; X3 представляет собой L; X4 представляет собой L; и X5 представляет собой F.
7. Антитело по любому из пп.1-6, содержащее константную область антитела человека, предпочтительно, константную область IgG.
8. Антитело по п.7, в котором указанная константная область представляет собой область с недостатком в активации комплемента, предпочтительно константную область IgG4 человека или мутированную константную область IgG1 человека.
9. Нуклеиновая кислота, кодирующая антитело по любому из пп.1-8.
10. Клетка, содержащая нуклеиновую кислоту по п.9, которая экспрессирует антитело по любому из пп.1-8.
11. Антитело по любому из пп.1-8, где антитело представляет собой моноклональное антитело против CD40 человека.
12. Моноклональное антитело против CD40 человека по п.11, где антитело является деиммунизированным.
13. Антитело по п.1, в котором X6 представляет собой T или S, X7 представляет собой D или Q, X8 представляет собой Q или P, X9 представляет собой V или A, X10 представляет собой V или L и X11 представляет собой F или Y.
14. Антитело по п.13, в котором X6 представляет собой T, X7 представляет собой Q, X8 представляет собой P, X9 представляет собой A, X10 представляет собой V и X11 представляет собой Y.
15. Клетка по п.10, являющаяся гибридомной клеткой, клеткой яичника китайского хомячка, клеткой NSO или клеткой PER-C6тм.
16. Культура клеток по п.15 для получения антитела по любому из пп.1-8 и 11-14.
17. Способ получения антитела, включающий культивирование клетки по любому из пп.10 или 15 и сбор указанного антитела из указанной культуры.
18. Фармацевтическая композиция для уменьшения интенсивности симптомов аутоиммунного заболевания, и/или воспалительного заболевания, и/или уменьшения отторжения трансплантата, и/или для лечения CD40-положительного рака, содержащая терапевтически эффективное количество антитела против CD40 по любому из пп.1-8.
19. Антитело по любому из пп.1-8 и 11-14 для применения в качестве лекарственного средства.
20. Применение антитела по любому из пп.1-8 и 11-14 для получения лекарственного средства, уменьшающего интенсивность симптома аутоиммунного заболевания, и/или воспалительного заболевания, и/или уменьшения отторжения трансплантата, и/или для лечения CD40-положительных раков.
21. Применение по п.20, при котором указанное аутоиммунное и/или воспалительное заболевание выбрано из группы, состоящей из ревматоидного артрита, системной красной волчанки, рассеянного склероза, псориаза, буллезного пемфигоида и аллергического дерматита.
22. Применение по п.21, при котором указанным аутоиммунным и/или воспалительным заболеванием является воспалительное заболевание кишечника.
23. Применение по п.22, при котором указанное воспалительное заболевание кишечника включает язвенный колит (UC) или болезнь Крона (CD).
24. Способ введения индивидууму антитела против CD40, включающий введение указанному индивидууму терапевтически эффективного количества антитела по любому из пп.11-14 или 19 для облегчения симптомов аутоиммунного заболевания и/или воспалительного заболевания, и/или для уменьшения реакции отторжения трансплантата, и/или для лечения рака, позитивного по CD40.
25. Способ отбора антитела против CD40 человека с повышенной экспрессией, включающий получение первой линии клеток, которая продуцирует исходное антитело против CD40 человека, и определение количества исходного антитела, которое продуцируется указанной первой линией клеток, при этом указанное исходное антитело содержит вариабельную область легкой цепи, содержащую последовательность

и вариабельной области тяжелой цепи, содержащую последовательность

в которой X1 и X2 попарно выбирают из группы, состоящей из X1=I и X2=V; X1=L и X2=I; X1=V и X2=V; X1=L и X2=L; или X1=L и X2=V, при этом указанный способ дополнительно включает создание по крайней мере одной дополнительной линии клеток, которая продуцирует вариант указанного исходного антитела, причем указанный вариант антитела представляет собой модифицированное исходное антитело, включающее вставку, делецию, инверсию и/или замену приблизительно 1-5 аминокислот по сравнению с указанным исходным антителом, причем указанная модификация не состоит из модификации аминокислот в положении(ях), определяемых X1 и X2, и определение количества варианта антитела, который продуцируется указанной по крайней мере одной дополнительной линией клеток,
при этом указанный способ включает отбор варианта антитела против CD40 человека с повышенной экспрессией.
26. Способ по п.25, в котором указанные вставка, делеция, инверсия и/или замена приблизительно 1-5 аминокислот находятся в указанной аминокислотной последовательности тяжелой цепи или указанной аминокислотной последовательности легкой цепи по сравнению с аминокислотной последовательностью соответствующей цепи в указанном исходном антителе.
27. Способ по п.26, в котором указанные вставка, делеция, инверсия и/или замена приблизительно 1-5 аминокислот находятся в указанной аминокислотной последовательности тяжелой цепи по сравнению с указанной последовательностью тяжелой цепи указанного исходного антитела.
28. Способ по любому из пп.25-27, где указанная вставка, делеция, инверсия и/или замена представляет собой аминокислоту в вариабельной последовательности тяжелой цепи или вариабельной последовательности легкой цепи исходного антитела с соответствующими аминокислотами последовательности вариабельной последовательности тяжелой или легкой цепи химерного 5D12 соответственно.
29. Способ по любому из пп.25-28, дополнительно включающий получение линии клеток, продуцирующей антитело, которая продуцирует указанное отобранное антитело.
30. Способ по п.29, дополнительно включающий сбор указанного отобранного антитела.
31. Антитело против CD40 человека, которое собирают способом по п.30, где антитело содержит вариабельный домен легкой цепи, содержащий последовательность

и вариабельный домен тяжелой цепи, содержащий последовательность

где X1 и X2 представляют собой пару, выбранную из группы, состоящей из X1=I и X2=V; X1=L и X2=I; X1=V и X2=V; или X1=L и X2=L,
где антитело содержит вставку, делецию, инверсию и/или замену между приблизительно 1-5 аминокислотами, где указанная модификация не состоит из модификации аминокислотного положения(ий), определенных как X1 и X2.
| BOON L | |||
| et al | |||
| Приспособление с иглой для прочистки кухонь типа "Примус" | 1923 |
|
SU40A1 |
| Секция механизированной крепи | 1979 |
|
SU945465A1 |
| Дорожная спиртовая кухня | 1918 |
|
SU98A1 |
| Бесколесный шариковый ход для железнодорожных вагонов | 1917 |
|
SU97A1 |
| KASRAN A | |||
| et al | |||
| Приспособление с иглой для прочистки кухонь типа "Примус" | 1923 |
|
SU40A1 |
| Alimentary | |||

