Настоящее изобретение относится к применению в технологии белковой инженерии для улучшения свойств ферментов. В частности, настоящее изобретение раскрывает способ определения аминокислот, замещение которых вызывает изменение региоспецифичности. Этот способ применяется к α-D-галактозидазам, относящимся к классу гликозидгидролаз.
В другом аспекте изобретение относится к α-D-галактозидазам с измененной региоспецифичностью.
Альфа-D-галактозидазы (К. Ф.3.2.1.22) - гликозидгидролазы, способные отщеплять концевые остатки a-D-галактозы с невосстанавливающего конца α-галактоолигосахаридов (Encyclopedia of Plant Physiology, 1982, pp 418-471) [1] с сохранением аномерной конфигурации атома С1 субстрата в продукте. В ряде случаев у α-D-галактозидаз обнаружена трансгликозилирующая активность, выражающаяся в способности фермента не только гидролизовать, но и синтезировать галактозосодержащие соединения, в частности, глоботриозу и ее производные.
Глоботриоза (αGal-(1,4)-β-Gal-(1,4)Glc) - это галактоолигосахарид, у которого концевой остаток галактозы присоединен к следующему через α-1,4-O-гликозидную связь. Глоботриоза присутствует на поверхности многих клеток и способна связывать и обеспечивать проникновение в организм человека так называемых шига-токсинов, вырабатываемых Shigella dysenteriae и патогенными штаммами Е. coli, часто обнаруживаемые при пищевых отравлениях: Clin Microbiol Rev 11: 450-479 (1998) [2]. Желудочно-кишечные заболевания, вызываемые шига-токсином, нередко приводят к летально опасным системным осложнениям в организме, включающим острые случаи энцефалопатии и гемолитическому уремическому синдрому: Microbiol Rev 51: 206-220 (1987) [3]. Попадая через эпителий кишечника в кровеносную систему, шига-токсин способен поражать ткани мозга и печени. Одной из эффективных стратегий борьбы с указанными бактериальными болезнями является адсорбция токсинов и патогенных бактерий и последующее их выведение из организма. В качестве сорбента применяют искусственно синтезированную глоботриозу и ее производные: J Infect Dis 189: 360-368 (2004) [4]. Кроме того, на основе глоботриозы, содержащей наночастицы золота, можно проводить экспресс анализ для выявления данных заболеваний на ранних стадиях: Antimicrob Agents Chemother 49: 3842-3846 (2005) [5], либо для обнаружения присутствия бактерий в пищевых продуктах: Microbiol Immunol (2007) 51(6): 581-592. [6]. Таким образом, синтез α-1,4-галактоолигосахаридов (где остатки галактозы соединены между собой или другими сахарными остатками через α-1,4-гликозидную связь) и, в частности, глоботриозы, крайне необходим для удовлетворения возрастающего спроса на такие соединения для фармацевтической промышленности.
Однако традиционные химические методы синтеза α-1,4-галактоолигосахаридов (олигосахаридов) состоят из ряда трудоемких стадий, включающих блокирование и деблокирование реакционноспособных химических групп. В результате такого синтеза помимо основного продукта появляются вредные с экологической точки зрения отходы.
Альтернативой дорогостоящему химическому синтезу олигосахаридов является ферментативный синтез с применением двух классов ферментов, а именно, гликозилтрансфераз и гликозидгидралаз. Однако достаточно высокая цена нуклеотид-активированных Сахаров, являющихся субстратами при направленном синтезе олигосахаридов с помощью гликозилтрансфераз, уменьшает привлекательность использования этого класса ферментов.
В свою очередь гликозидгидролазы, способные осуществлять реакции трансгликозилирования (реакции так называемого обращения гидролиза, перенося связавшийся с активным центром остаток сахарного субстрата (донора) на гидроксил другого сахара или спирта (акцептора)), являются сейчас перспективным инструментом для получения олигосахаридов в биотехнологии: Industrial Enzymes, 2007:141-1, [7].
Одним из существенных недостатков использования гликозидгидролаз в синтезе олигосахаридов является их широкая региоспецифичность, т.е. способность синтезировать широкий спектр региоизомеров (олигосахаридов одинаковой структуры, но с разными положениями присоединения сахаридных остатков друг к другу).
Известны α-D-галактозидазы, способные гидролизовать и синтезировать галактоолигосахариды. Патенты WO 2007071987, US 7514254, US 6740509 (WO 9844145) [8-10] описывают ферментативные способы синтеза различных галактосодержащих олигосахаридов и галактоконьюгатов с помощью трансгликолизирующей активности альфа-галактозидаз из различных бактериальных и растительных источников. Все описанные α-D-галактозидазы не отличаются способностью гидролизовать и синтезировать галактоолигосахариды с заданными типами связями, т.е. обладают широкой региоспецифичностью.
Наиболее близким к заявляемому объекту является α-D-галактозидаза дикого типа из Talaromyces flavus. В патенте CZ291629 [11] описано использование α-D-галактозидазы для ферментативного синтеза изо-глоботриозы (αGal-(1,3)-α-Gal-(1,4)Glc), заключающийся в присоединении галактозного остатка к лактозе, которая предварительно была ацетилирована в 6-ом положении галактозного кольца с помощью дополнительного фермента липаза. Введение стадии ацетилирования при синтезе изо-глоботриозы α-D-галактозидазой из Talaromyces flavus было необходимо в данном случае, поскольку их фермент не обладал узкой региоспецифичностью по отношению ни к α1,4-галактозидным связям, ни к какому-либо другому типу связи.
Таким образом, все рассмотренные α-D-галактозидазы не обладают способностью синтезировать преимущественно α1,4-галактоолигосахариды, в частности, глоботриозу и ее производные, в силу их широкой природной региоспецифичности.
Задачей изобретения является получение α-1,4-специфической D-галактозидазы и создание способа, обеспечивающего изменение природной региоспецифичности α-D-галактозидазы.
Техническим результатом, получаемым при осуществлении данного изобретения, является получение α-1,4-специфической D-галактозидазы, обеспечивающей синтез олигосахаридов с преимущественно α-1,4-галактозидными связями, т.е. синтез галактоолигосахаридов с α-1,4-связями в большем количестве, чем олигосахаридов с другими типами связи.
При этом полученные мутантные формы α1,4-специфической галактозидазы должны удовлетворять следующим требованиям:
1. обладать повышенной трансгликозилирующей активностью по сравнению с ферментом дикого типа;
2. синтезировать преимущественно α1,4-галактозсо держащие соединения;
3. обладать способностью синтезировать глоботриозу преимущественно. Поставленная задача достигается тем, что предлагается α-D-галактозидаза, отличающаяся тем, что она содержит по крайней мере, одну мутацию, а именно, либо Pro402Asp, либо Phe328Ala, обеспечивающую аминокислотную замену в соответствующем положении аминокислотной последовательности исходного фермента дикого типа, выделенного из штамма Thermotoga maritima MSB8, приводящую к изменению региоспецифичности по отношению к α1,4-связи.
Поставленная задача достигается также тем, что предлагается способ получения α-D-галактозидаза, заключающийся в том, что осуществляют выбор позиций для сайт-направленного мутагенеза аминокислотной последовательности исходного фермента дикого типа следующим образом:
а) проводят пространственное наложение активных центров ферментов, обладающих высокой гомологией их аминокислотных последовательностей и сходством трехмерных структур, и кристаллическая структура которых содержит остаток D-галактозы, на основании проведенного наложения структур установливают возможное расположение D-галактозы в активном центре α-галактозидазы из Thermotoga maritima;
б) уточняют методами молекулярной динамики возможное расположение D-галактозы в активном центре α-галактозидазы, определяют общие аминокислотные остатки ферментативного центра, образующие водородные связи с D-галактозой, и выявляют возможные положения D-галактозы относительно пары каталитических аминокислотных остатков фермента;
в) проводят in silico мутации α-галактозидазы дикого типа, при этом отбирают позиции аминокислотных остатков для сайт-направленного мутагенеза по следующим критериям: ослабление взаимодействий и препятствование связыванию лиганда в активных конформациях, выгодных для образования связей 1→3 и 1→6; усиление взаимодействий, способствующих связыванию в активных конформациях, выгодных для образования связи 1→4.
После выбора позиций, по крайней мере, один из отобранных на этапе в) аминокислотных остатков путем введения соответствующей мутации в ген природной α-D-галактозидазы замещают другим аминокислотным остатком, а именно, либо Pro402Asp.либо Phe328Ala, обеспечивающим преимущественное образование α1,4-галактозидной связи и подавление образование других связей.
Совокупность данные признаков основана на проведении предварительных экспериментов.
Экспериментально было обнаружено, что природная α-D-галактозидаза (К.Ф. 3.2.1.22) из термофильной бактерии Thermotoga maritima (ТмГалА) обладает широкой региоспецифичностью, в результате чего концевой остаток α-галактозы может отщепляться от трех различных галактоконъюгатов: αGall-3βGall-pNP, αGall-4βGall-pNP, и αGall-6βGall-pNP (паранитрофенил (ПНФ) замещенные дисахариды), при этом гидролиз α-1,4-связанных галактоконьюгатов происходит с наименьшей скоростью. Поскольку реакции гидролиза и трансгликозилирования взаимосвязаны, следует ожидать, что и в процессе синтеза подобных соединений (с α-1,4-связью между галактозными остатками) будут образовываться два побочных сахарида в количествах, превышающих выход целевой продукта. Кроме того, предварительные эксперименты о наличии у этого фермента трансгликозилирующей активности позволили нам предположить, что ТмГалА может быть использована для синтеза биологически значимого и фармакологически активного галактоконъюгата, глоботриозы αGall-4βGall-4Glc.
Был использован метод компьютерного моделирования α-D-галактозидазы с лигандами для выявления позиций мутирования гена исходного фермента для использования его в синтезе преимущественно α1,4-галактозидных связей.
На фигуре 1 показана структура активированного комплекса фермента ТмГалА, в котором остаток α-D-галактозы ковалентно связан с каталитической аминокислотой Asp327.
На фигуре 2 показано расположение молекулы пара-нитрофенил галактозида вблизи галактозил-α-галактозидазного комплекса мутантной формы Phe328Ala. Показаны каталитические аминокислотные остатки Asp327 и Asp387, мутированый аминокислотный остаток Phe328Ala и галактозил.
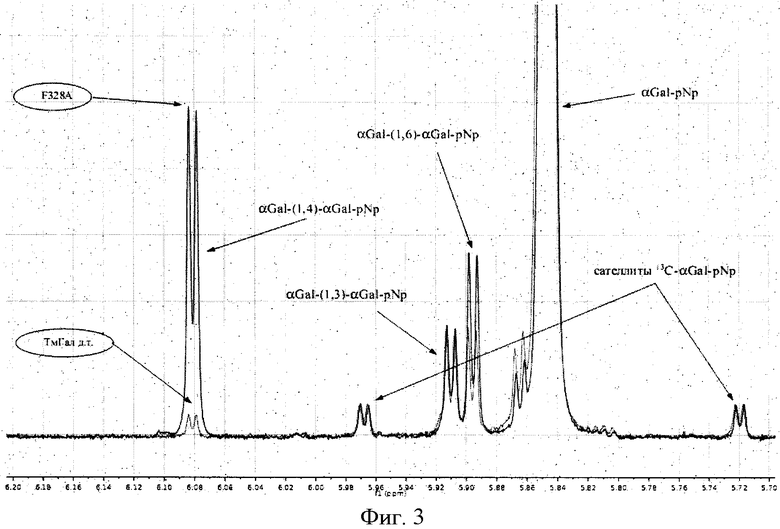
На фигуре 3 представлен сравнительная ЯМР-спектрограмма продуктов, полученных с помощью мутантной формы и природной α-галактозидаз. Обозначены пики, характерные для продуктов, полученных с помощью мутанта Phe328Ala, и природной ТмГалА. Высота пиков характеризует количество полученных продуктов.
Способ получения α-D-галактозидаз с измененной региоспецифичностью заключается в следующем:
Методами компьютерного моделирования были созданы и оптимизированы структуры исследуемых лигандов: четырех олигосахаридов: αGal1,3-Gal1-pNP, αGal1,4-Gal1-pNP, αGal1,6-Gal1-pNP (паранитрофенил (ПНФ) замещенные дисахариды) и Gal1-4Glc (лактоза). Выбор лигандов обусловлен следующими факторами. Нитрофенил является хорошей уходящей группой и его удобно отслеживать спектроскопическими методами в ходе биохимических реакций. Поэтому ПНФ-замещенные химические соединения часто используются при изучении кинетики ферментативных реакций.
Для каждой из трех молекул ПНФ-дисахаридов был проведен докинг в активном центре фермента. Были отобраны структуры ПНФ-дисахаридов с наименьшей конформационной энергией, и эти конформации были взяты в качестве исходных для проведения молекулярной динамики комплекса в явно заданном растворителе с периодическими граничными условиями. Используя уравнение Ярзинского (Physical Review Letters, 1997, 78(14) 2690. [12]) для неравновесных термодинамических процессов, рассчитаны абсолютные величины свободной энергии связывания каждого из pNP-дисахаридов в активном центре ТмГалА. По результатам молекулярно динамического эксперимента установлены ключевые аминокислотные остатки ТмГалА, расположенные в сайтах -1 и +1, и участвующие в связывании агликоновой части изучаемых лигандов и изучены их динамические свойства.
Промоделирована структура активированного комплекса фермента ТмГалА, в котором остаток α-D-галактозы ковалентно связан с каталитической аминокислотой Asp327 (см. фигуру 1). После проведения докинга лактозы с активированным комплексом и анализа полученных результатов выделены аминокислоты, влияющие на ориентацию лактозы вблизи активированного комплекса, и предложены возможные мутации этих аминокислот, выделяющие ориентации лактозы, выгодные для синтеза глоботриозы. Вычисления проводились в следующих пакетах программ: квантово-химических - HyperChem 8.0 или GAMESS 4, в молекулярно-механическом - ICM Pro 3.6, а также в молекулярно-динамическом - GROMACS 4.0.7.
Для синтеза преимущественно 1→4 гликозидной связи была выбрана следующая стратегия: 1) ослабление взаимодействий и препятствование связыванию лиганда в активных конформациях, выгодных для образования связей 1→3 и 1→6; 2) усиление взаимодействий, способствующих связыванию в активных конформациях, выгодных для образования связи 1→4. Согласно выбранной стратегии по первому направлению заслуживают внимания аминокислоты: Trp85, Pro402, Phe391, Pro390, Ala389 и Phe194; по второму направлению: Leul95, Phe391, Pro402. Следует отметить, что более плотному приближению гидроксильных групп остатка галактозы к аномерному центру препятствует боковая цепь Phe328. Можно предположить, что замена ее на меньшую по величине может привести к расширению входа в активный центр в области аномерного углерода стабильного интермедиата. Более плотный контакт гидроксилов акцептора с аномерным центром, вероятно, облегчит реакцию гликозилирования. Исходя из этих предположений, были исследованы следующие замены АК: Phe194Lys, Phe194Met, Phe194Ser, Phe194Tyr, Leu195Ala, Leu195Cys, Leu195Thr, Asn290Ser, Phe328Ala, Phe328Asp, Phe328Gly, Phe328His, Trp85Ala, Trp85Phe, Trp85Tyr, Pro402Asp, Pro402Asn, Pro402Ser.
Для всех замен in silico методом «гибкого докинга» были рассчитаны варианты связывания акцептора вблизи активного центра галактозил-ферментного интермедиата.
После проведения докинга и минимизации системы методами молекулярной динамики (пакет программ ICM-Pro) сравнивались свободные энергии системы и расстояния от 4-го кислорода галактозного кольца в акцепторе до аномерного углерода в интермедиате, Кроме того, для увеличения общей трансгликозилирующей активности представляется целесообразным увеличить кислотность АК, выполняющей роль кислотно-основного катализатора (в нашем случае - остаток Asp387). Это возможно сделать, используя имеющийся в активном центре остаток Arg383. АК Arg383 является консервативным для активных центров нескольких гликозидгидролаз и выполняет роль «активатора» кислотно-основного Asp387. Его положение и конфигурация очень жестко задается цепочками водородных связей Asp387 OD1 - Arg383 NH2 - Gly385 О и Arg383 Nε - Н20565 - Gly385 N, что подтверждается значениями В-факторов в кристаллической структуре ТмГал. Очевидно, обе эти цепочки замыкаются на Gly385. Следовательно, если заменить Gly385 на гидрофобный АК (Pro, Leu или Ile), то гидрофобное взаимодействие с Trp65 вызовет поворот связи СО в остатке 385 и, возможно, изменение угла при N остатка 385, что освободит/ослабит связи с Arg383 NH2 и Arg383 Nε и вызовет его смещение в сторону гидроксила Asp387.
Таким образом, по данным эксперимента in silico предложены следующие мутации фермента, способные вызвать изменение региоспецифичности α-галактозидазы в реакции трансгликозилирования: Phe328Ala (см. фигуру 2), Leu195Cys, Trp85Tyr, Pro402Asp, Phe194Lys. После чего был осуществлен сайт-направленный мутагенез.
Плазмида с геном α-галактозидазы (ТмГалА) дикого типа была любезно предоставлена проф. Робертом Келли из Государственного университета Северной Каролины (США). Ген а-галактозидазы (galA) был клонирован из термофильной бактерии Thermotoga maritima MSB8 (ORF TM1192): Biochemistry, 2007, 46(11): 3319-3330 [13]. Вектор pET24D (Novagen, San Diego, CA), содержащий ген α-галактозидазы, был выделен с помощью QIAGEN plasmid midi kit (кат. №12143). Сайт-направленный мутагенез был осуществлен компанией "Евроген" (http://www.evrogen.com/). Было синтезировано семь плазмид, со следующими точечными мутациями в гене α-галактозидазы: Pro402Asp; Pro402Ser; Gly385Leu; Trp85Tyr; Phe328Ala; Phel94Lys; Leul95Cys. Плазмиды с геном α-галактозидазы дикого типа и мутантных форм были трансформированы в штамм Escherichia coli BL-21 (DE3) по методу, описанному в «Молекулярное клонирование». 1984. С.240-241 [14].
Культуры, выращенные на чашке с агаризованной средой Луриа-Бертони (LB), с добавлением 50 мг/л канамицина, инокулировали в 5 мл жидкой среды LB и растили в течение ночи при 37°C. Далее 1 мл ночной культуры инокулировали в 100 мл среды LB, растили при 37°С на шейкере до ОП600 0,4-0,6 и добавляли ИПТГ до конечной концентрации 1 ммоль. Далее колбу оставляли на шейкере при 37°C в течение ночи.
Все стадии очистки рекомбинантных α-галактозидаз выполняли при 4°С. Клетки центрифугировали (Beckman Coulter, 8000 об/мин, 40 мин), дважды промывали 50 ммоль/л калий-фосфатным буфером, pH 7.0 (буфер А), с добавлением 1 ммоль/л ЭДТА. Клетки ресуспендировали в этот же буфер (объем 5 мл), содержащего 1 мг/мл лизоцима (Sigma Chemical Co., USA) и оставляли на 1 час при 37°C. Полученную суспензию, предварительно выдержанную при +4°C в течение 45 минут, обрабатывали ультразвуком (6×20 сек) и центрифугировали (10000 об/мин, 30 мин.). Полученный супернатант выдерживали 20 мин при температуре +60°C для денатурации нерекомбинантных белков и лизоцима. Осадок центрифугировали (10000 об/мин, 30 мин.). Супернатант диализовали против 10 ммоль/л калий-фосфатного буфера, pH 7.0 (буфер Б). Полученную фракцию α-галактозидазы наносили на колонку DEAE-Toypearl 650M (10×100 мм), уравновешенную буфером Б. Белок элюировали линейным градиентом (0.01-0.5 М) калий-фосфатного буфера, pH 7.0, (скорость протекания 1 мл/мин, в течение 60 мин). Фракции, содержащие α-галактозидазную активность, концентрировали на мембране Millipore (Centriocon YM-30, кат. №4209) до 2 мл. Белок дочищали при помощи гель-фильтрации на колонке Sephacryl S-200 (22×960 мм) (Pharmacia), уравновешенную буфером А (объем фракций 5 мл, скорость протекания 1.5 мл/мин). Чистоту полученных ферментов устанавливали с помощью электрофореза в 10%-ном полиакриламидном геле с использованием додецилсульфата натрия по методу Лаэммли: Nature (1970) 227: 680-685 [15] с использованием стандартного набора белков с молекулярными массами 14400-94000 Да в качестве реперов (LMW, Pharmacia Biotech, Швеция).
Концентрацию фермента измеряли методом Лоури с использованием бычьего сывороточный альбумина (БСА) в качестве стандарта: J. Biol. Chem. (1951)193(1): 265-275 [16].
Ферментативную активность каждого из полученных определяли следующим образом. Реакционную смесь (общий объем - 30 мкл), содержащую 10 мкл субстрата с конечной концентрацией 5 мг/мл и 10 мкл фермента в растворе 50 ммоль/л калий-фосфатного буфера (pH 7.0), инкубировали в течение 10 мин при 37 DC. Реакцию останавливали добавлением 1 мл 5%-го раствора Na2CO3. Удельную активность фермента рассчитывали по количеству освобожденного p-нитрофенола (λ=400 нм, ε=18300 М/см). Одну единицу α-галактозидазной активности определяли как количество фермента, необходимого для освобождения 1 мкмоль галактозы в результате гидролиза ПНФГал за 1 минуту при рН 7.0 и температуре 37°C.
Значения констант Михаэлиса (КМ) и констант скоростей ферментативного гидролиза (kcat) определяли для ПНФГал варьированием концентраций субстратов (6-15 точек) в диапазоне от 0.1 до 4-8 КМ. Обработку данных осуществляли при помощи нелинейного регрессионного анализа уравнений Михаэлиса-Ментен. Условия проведения реакции: реакционную смесь (общий объем - 200 мкл), содержащую субстрат и 0.076 ед. фермента (ТмГал д.т.или ее мутантные формы) в растворе 50 ммоль/л натрий-ацетатного буфера (рН 5.0), инкубировали при 37 ПС. Реакцию останавливали добавлением 1 мл 5%-го раствора Na2CO3.
В качестве субстрата для исследования трансгликозилирующей активности ТмГал и ее мутантных форм использовали ПНФГал. Реакционную смесь (250 мкл), содержащую субстрат в концентрации 100 ммоль/л и 0.057 ед. фермента (ТмГал дикого типа или ее мутантные формы) в растворе 50 ммоль/л калий-фосфатного буфера (pH 7.0), инкубировали при температуре 50°C в течение 30 мин. Реакцию останавливали замораживанием в жидком азоте с последующей лиофилизацией для исследования продуктов реакции.
Продукты трансгликозилирования анализировали при помощи тонкослойной хроматографии на Kieselgel 60 пластинах (Е. Merck) с подвижной фазой бутан-1-ол-уксусная кислота-вода (в соотношении 3:1:1 об./об.). Аликвоты реакционной смеси по 5 мкл наносили на мкл наносили на пластинки, далее высушивали. После окончания хроматографии для проявления хроматограф ических зон пластину обрабатывали 5% H2SO4 в 2-пропаноле и прогревали при 120°C в течение 8 мин. В качестве реперов использовались дисахариды, аутентичные получаемым в реакции. Интегральная интенсивность пятна соответствующего дисахарида служила оценочной мерой трансгликозилирующей активности.
Все 1Н и 13С ЯМР спектры регистрировали на ЯМР спектрометре АМХ-500 Bruker (1Н 500.13 МГц, 13С 125 МГц,) в D2O при комнатной температуре, с использованием ацетона в качестве внутреннего стандарта. Химические сдвиги даны относительно внутреннего стандарта: 2.225 ррм для 1Н и 31.5 ррм для 13С.
Одномерные 1Н, одномерные спектры эффекта Оверхаузера, 13С и фазово-чувствительные двумерные спектры (COSY-DQF, NOESY и протон-углеродные корреляционные спектры) регистрировали с использованием стандартных импульсных программ при 20°C и 50°C. Результаты анализировали с использованием пакета программ XWINNMR (Bruker). Соотнесение сигналов индивидуальных сахарных остатков основывалось на двумерной фазово-чувствительной спектроскопии COSY. Определение последовательности остатков основывалось на кросс-пиках эффекта Оверхаузера в двумерном NOESY эксперименте, (время смешения 180 мсек), и наблюдении эффекта Оверхаузера между аномерными протонами и протоном при замещенном положении. Соотнесение 13С сигналов основывалось на протон-углеродных корреляциях, наблюдаемых в 1Н - 13С корреляционных спектрах. Определение аномерной конфигурации было основано на химических сдвигах и константе спин-спинового взаимодействия J1,2, измеряемых в одномерных 1Н спектрах.
Для исследования реакции трансгликозилирования и количественного определения скоростей реакции использовали метод ЯМР. Для этого реакцию, как описано выше, проводили в стандартной 5-мм ампуле в магните ЯМР спектрометра, общий объем реакционной смеси составлял 600 мкл. Реакционную смесь без фермента подогревали до 50°C и помещали в магнит для настройки однородности магнитного поля и условий подавления сигналов растворителя и буфера. После этого добавляли 40 мкл фермента (0.057 ед.) для инициации реакции и начинали регистрацию ЯМР-сигнала. Для наблюдения 1Н ЯМР спектра использовали стандартную одно-импульсную последовательность с углом поворота 45°, временем наблюдения спада свободной индукции aq=1.5 сек и временем релаксации dl=1 сек. Число точек в спектре составляло 32К и число повторений nt=32, так чтобы общее время регистрации одного спектра составило 80 сек. Для подавления остаточного сигнала растворителя (Н2О) использовали последовательность с насыщением воды во время релаксации dl или последовательность WET. Концентрация компонентов реакционной смеси определялась интегрированием следующих неперекрывающихся сигналов с химическими сдвигами 5:
5.91 для aGal-(1,3)-αGal-pNp (H2 протон),
6.08 для aGal-(1,4)-αGal-pNp (Н1' протон),
5.90 для aGal-(1,6)-αGal-pNp (Н1' протон),
5.26 и 4.58 для α- и β-галактозы (H1 протон),
5.85 для ПНФГал
6.81 для ПНФ
Относительная скорость реакции трансгликозилирования определялась как отношение количества каждого продукта трансгликозилирования к общему количеству образующихся продуктов в единицу времени по формуле:
а1=C1/(C1+C2+C3+CαGal+CαGal),
где ai - относительная скорость образования i-продукта реакции трансгликозилирования;
Ci - концентрация i-соединения, ммоль/л; CαGal и CβGal - концентрации α- и β-галактозы, ммоль/л.
Апробирование региоспецифичности полученных мутантных форм α-галактозидазы
Ферменты ТмГалА дикого типа и ее Pro402Asp, Pro402Ser, Gly385Leu, Trp85Tyr, Phe328Ala, Phe194Lys, Leu195Cys мутанты были выделены в результате двух хроматографических стадий согласно протоколу, описанному выше. На конечной стадии очистки были получены гомогенные по данным электрофореза ферменты. Согласно данным электрофореза в присутствии ДСН, каждый белок мигрировал в виде единичной зоны, молекулярная масса фермента соответствовала 60±1 кДа.
Природная α-D-галактозидаза и ее мутантные варианты были получены в виде водных растворов.
Максимальная гидролитическая активность мутантов Pro402Asp и Phe328Ala была обнаружена при pH 5.0, как и для ТмГал д.т. Трансгликозилируюная активность обоих мутантов отличалась от гидролитической. Так, для мутанта Phe328Ala максимальный выход 1,4-региоизомера был обнаружен при рН 3.0, а мутант Pro402Asp при этом значении pH (3.0) был неактивен.
С помощью процедур ЯМР-спектроскопии были проанализированы реакции трансгликозилирования для всех мутантов. На фигуре 3 проиллюстрировано изменение "региоспецифичности мутанта Phe328Ala по сравнению с ТмГалА дикого типа посредством ЯМРспектроскопии. По данным этих экспериментов были рассчитаны скорости образования продуктов трансгликозилирования для каждого региоизомера отдельно (Табл. 1). Данные дали возможность определить, в каком из полученных мутантов региоспецифичность в отношении альфа-1,4-галактозодной связи была изменена по сравнению с нативной альфа-Д-галактозидазой.
Из таблицы 1 видно, что два мутанта, а именно Pro402Asp и Phe328Ala, существенно изменили свою региоспецифичность в отношении альфа-1,4-галактозодной связи по сравнению с нативной альфа-Д-галактозидазой.
Так, скорость образования альфа 1,4-связи у Pro402Asp и Phe328Ala мутантов увеличилась по сравнению с ТмГал д.т.в 7 и 36 раз, соответственно. При этом скорости образования двух других региоизомеров в среднем в 2 раза ниже для каждого из них. У мутанта Gly385Leu также наблюдается увеличение скорости образования альфа-1.4-дигалактозидов, однако скорости двух других изомеров не изменились.
Кроме того, для всех мутантов были рассчитаны выходы продуктов трансгликозилирования (Табл. 2).
Оказалось, что у мутанта Phe328Ala увеличился как общий выход продуктов трансгликозилирования в почти в 3 раза, так и выход искомых альфа-1,4-дигалактозидов (в 16 раз); для мутанта Pro402Asp общий выход продуктов трансгликозилирования увеличился в 1,4 раза, тогда как выход альфа-1,4-галактоолигосахаридов увеличился в 7 раз по сравнению с нативным ферментом.
Таким образом, из приведенных в табл.1 и табл.2 данных, у мутантов Phe328Ala и Pro402Asp реализованы такие изменения аминокислотных замен в соответствующем положении аминокислотной последовательности исходного фермента дикого типа, выделенного из штамма Thermotoga maritima MSB8, которые привели к изменению региоспецифичности по отношению к образованию α1,4-связи.
Область применения: заявляемые α-1,4-галактозидазы позволяют осуществлять целенаправленный ферментативный синтез α1,4-галактоолигосахаридов, применяемых в качестве пищевых добавок, в частности, глоботриозы, используемой как сорбент токсинов и патогенных бактерий для обнаружения присутствия бактерий в пищевых продуктах.
Литература
1. Н. Meier and J. S. G. Reid (1982) Reverse polysaccharides other than starch in higher plants, in Encyclopedia of Plant Physiology (F.A. Loewusand W. Tanner) pp 418-471, Springer-Verlag, New York).
2. J.C. Paton and A.W. Paton (1998). Pathogenesis and diagnosis of Shiga toxin-producing Escherichia coli infections. Clin Microbiol Rev 11: 450-479.
3. A.D. O'Brien and R.K. Holmes (1987). Shiga and Shiga-like toxins. Microbiol Rev 51: 206-220.
4. M. Watanabe, K. Matsuoka, E. Kita, K. Igai, N. Higashi, A. Miyagawa, T. Watanabe, R. Yanoshita, et al. (2004). Oral therapeutic agents with highly clustered globotriose for treatment of Shiga toxigenic Escherichia coli infections. J Infect Dis 189: 360-368.
5. J.L. Leach, S.A. Garber, A.A. Marcon, and P.A. Prieto (2005). In vitro and in vivo effects of soluble, monovalent globotriose on bacterial attachment and colonization. Antimicrob Agents Chemother 49: 3842-3846.
6. Neri, P., Nagano, S.I., Yokoyama, S. и др. Neutralizing activity of polyvalent Gb3, Gb2 and galacto-trehalose models against Shiga toxins / Neri, P., Nagano, S.I., Yokoyama, S. [и др.]// Microbiol Immunol - 2007. - V.51. - No.6 - P.581-592.
7. Plou, F.J., de Segura, A.G. and Ballesteros, A.: Application of Glycosidases and Transglycosidases in the Synthesis of Oligosaccharides In Industrial Enzymes. Edited by J. PolainaandA. P. MacCabe: Springer Netherlands; 2007:141-157.
8. WO 2007071987 (МКИ: C12N 9/40).
9. US 7514254 (МКИ: C12N 9/40).
10. US 6740509 (WO 9844145) (МКИ: C12N 9/24).
11. CZ 291629 (прототип) (МКИ: С07Н 3/06, С12Р 19/00).
12. Jarzynski, С.Nonequilibrium Equality for Free Energy Differences / Jarzynski, С. // Physical Review Letters - 1997. - V.78. - No.14 - P.2690.
13. Comfort, D.A., Bobrov, K.S., Ivanen, D.R. [и др.]. Biochemical analysis of Thermotoga maritima GH36 alpha-galactosidase (TmGalA) confirms the mechanistic commonality of clan GH-D glycoside hydrolases / Comfort, D.A., Bobrov, K.S., Ivanen, D.R. [и др.] // Biochemistry - 2007. - V.46. - No. 11 - P.3319-3330.
14. Маниатис, Т., и др. //Молекулярное клонирование. / Мир. - 1984, - С.240-241.
15.Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. // Nature. - 1970. - V.227. - 680-685.
16. Lowry, O.H., Rosebrough, N.J., Fair A.L., Randall, R.J. Protein measurement with the Folin phenol reagent. // J. Biol. Chem. - 1951. - V.193. - No.1. - P.265-275.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ ГЛОБОТРИОЗЫ | 2011 |
|
RU2514661C2 |
| ПРИМЕНЕНИЕ РЕКОМБИНАНТНОЙ БЕТА-ГАЛАКТОЗИДАЗЫ AsBgl 1390 ИЗ АРХЕИ SACCHAROVORANS В КАЧЕСТВЕ БЕТА-ГЛЮКОЗИДАЗЫ, БЕТА-КСИЛОЗИДАЗЫ И БЕТА-МАННОЗИДАЗЫ | 2012 |
|
RU2523581C2 |
| ОЛИГОСАХАРИДЫ, МОДИФИЦИРОВАННЫЕ ФЛЮОРОФОРНЫМ МАРКЕРОМ, ДЛЯ ОПРЕДЕЛЕНИЯ ГЛИКАНАЗНЫХ АКТИВНОСТЕЙ И ФЕРМЕНТАТИВНЫЙ СПОСОБ ИХ ПОЛУЧЕНИЯ | 2004 |
|
RU2333915C2 |
| СПОСОБ ПОЛУЧЕНИЯ СИАЛИЛИРОВАННЫХ ОЛИГОСАХАРИДОВ | 2007 |
|
RU2473695C2 |
| ПРОДУКТ И СПОСОБ | 2006 |
|
RU2385931C1 |
| СПОСОБ ПОЛУЧЕНИЯ ГЛИКОЗИДОВ АКРИЛАТНЫХ ПРОИЗВОДНЫХ С ИСПОЛЬЗОВАНИЕМ ПОЛИСАХАРИДОВ И ГЛИКОЗИДАЗ ИЛИ ГЛИКОЗИЛТРАНСФЕРАЗ | 2011 |
|
RU2570556C2 |
| Способ ферментативного синтеза α-L-дифукопиранозидов, модифицированных неуглеводными группами | 2020 |
|
RU2756319C1 |
| БЕТА-ГАЛАКТОЗИДАЗА С ТРАНСГАЛАКТОЗИЛИРУЮЩЕЙ АКТИВНОСТЬЮ | 2007 |
|
RU2435856C2 |
| ГАЛАКТОЗИДАЗА С АКТИВНОСТЬЮ АЛЬФА-ГАЛАКТОЗИЛТРАНСФЕРАЗЫ | 2007 |
|
RU2441913C2 |
| ФЕРМЕНТАТИВНЫЙ СИНТЕЗ 4’-ЭТИНИЛНУКЛЕОЗИДНЫХ АНАЛОГОВ | 2019 |
|
RU2816846C2 |


Изобретение относится к области биохимии. Описана альфа-D-галактозидаза, обладающая измененной региоспецифичностью по отношению к α1,4-связи и содержащая мутацию либо Pro402Asp, либо Phe328Ala в соответствующем положении аминокислотной последовательности исходного фермента дикого типа, выделенного из штамма Thermotoga maritima MSB8. Предложен способ получения указанной альфа-D-галактозидазы, заключающийся в том, что осуществляют выбор позиций для сайт-направленного мутагенеза аминокислотной последовательности фермента дикого типа следующим образом: а) проводят пространственное наложение активных центров ферментов, обладающих высокой гомологией их аминокислотных последовательностей и сходством трехмерных структур, и кристаллическая структура которых содержит остаток D-галактозы, на основании проведенного наложения структур устанавливают возможное расположение D-галактозы в активном центре α-галактозидазы из Thermotoga maritima MSB8, б) уточняют методами молекулярной динамики возможное расположение D-галактозы в активном центре α-галактозидазы, определяют общие аминокислотные остатки ферментативного центра, образующие водородные связи с D-галактозой, и выявляют возможные положения D-галактозы относительно пары каталитических аминокислотных остатков фермента, в) проводят in silico мутации α-галактозидазы дикого типа, при этом отбирают позиции аминокислотных остатков для сайт-направленного мутагенеза, после выбора позиций один из отобранных на этапе в) аминокислотных остатков замещают другим аминокислотным остатком, а именно либо Pro402Asp, либо Phe328Ala. Изобретение позволяет получить региоспецифичные по отношению к α1,4-связи альфа-D-галактозидазы. 2 н.п. ф-лы, 3 ил., 2 табл., 1 пр.
1. Альфа-D-галактозидаза, обладающая измененной региоспецифичностью по отношению к α1,4-связи и содержащая, по крайней мере, одну мутацию, а именно, либо Pro402Asp, либо Phe328Ala, обеспечивающую аминокислотную замену в соответствующем положении аминокислотной последовательности исходного фермента дикого типа, выделенного из штамма Thermotoga maritima MSB8.
2. Способ получения α-D-галактозидаз по п.1, заключающийся в том, что осуществляют выбор позиций для сайт-направленного мутагенеза аминокислотной последовательности исходного фермента дикого типа следующим образом: а) проводят пространственное наложение активных центров ферментов, обладающих высокой гомологией их аминокислотных последовательностей и сходством трехмерных структур, и кристаллическая структура которых содержит остаток D-галактозы, на основании проведенного наложения структур устанавливают возможное расположение D-галактозы в активном центре α-галактозидазы из Thermotoga maritima MSB8, б) уточняют методами молекулярной динамики возможное расположение D-галактозы в активном центре α-галактозидазы, определяют общие аминокислотные остатки ферментативного центра, образующие водородные связи с D-галактозой, и выявляют возможные положения D-галактозы относительно пары каталитических аминокислотных остатков фермента, в) проводят in silico мутации α-галактозидазы дикого типа, при этом отбирают позиции аминокислотных остатков для сайт-направленного мутагенеза по следующим критериям: ослабление взаимодействий и препятствование связыванию лиганда в активных конформациях, выгодных для образования связей 1→3 и 1→6; усиление взаимодействий, способствующих связыванию в активных конформациях, выгодных для образования связи 1→4, после выбора позиций, по крайней мере, один из отобранных на этапе в) аминокислотных остатков путем введения соответствующей мутации в ген природной α-D-галактозидазы замещают другим аминокислотным остатком, а именно, либо Pro402Asp, либо Phe328Аlа, обеспечивающим преимущественное образование α1,4-галактозидной связи и подавление образования других связей.
| RYTCHKOV GN, et аl., "Modification of regiospecificity of the alpha-galactosidase from Thermotoga maritima towards pharmaceutically important globotriose", The second Sant-Petersburg International Conference on NanoBioTechnologies, Book of Abstracts, St.-Petersburg, Russia June 16-18, 2008 | |||
| Джино-прядильная машина | 1922 |
|
SU173A1 |
| JHIMLI DASGUPTA et аl., "In silico mutations | |||

